ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
В настоящем изобретении раскрыты полинуклеотидные последовательности генов, кодирующих сахарозосинтазу (SUS) из Nicotiana tabacum, и их варианты, гомологи и фрагменты. Также раскрыты кодируемые ими полипептидные последовательности и их варианты, гомологи и фрагменты. Также раскрыта модуляция экспрессии одного или более генов NtSUS или функции или активности полипептида(полипептидов) NtSUS, кодируемого(кодируемых) ими, для модулирования уровня редуцирующих сахаров в растении или его части.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
В данной области техники существует постоянная потребность в получении табачного материала, в котором изменено высвобождение вкусоароматических соединений в аэрозоль и дым. В данной области техники также существует постоянная потребность в получении табачного материала, в котором изменено высвобождение акриламида в аэрозоль и дым.
Настоящее изобретение направлено на удовлетворение данных и других потребностей.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Сахарозосинтаза (SUS) представляет собой ключевой фермент, задействованный в синтезе сахарозы, и ключевой фермент, запускающий накопление редуцирующих сахаров, таких как глюкоза, фруктоза, лактоза и мальтоза, в подвергнутых сушке листьях табака. Путем модулирования экспрессии SUS можно модулировать пул редуцирующих сахаров во время и после сушки листа табака. Измененное содержание редуцирующего сахара в подвергнутом сушке табаке может приводить к различному восприятию табачного вкуса и аромата, получаемых при нагревании. Как описано в данном документе, определенные гены NtSUS сверхэкспрессируются во время сушки, тогда как другие не экспрессируются и не будут вносить вклад в модулирование уровней редуцирующих сахаров. Однако данные гены, вероятно, будут участвовать в других метаболических путях, и изменения их экспрессии могут в результате привести к формированию фенотипа, который является неблагоприятным с агрономической точки зрения (например, медленный рост). Знание того, какие гены NtSUS сверхэкспрессируются во время сушки, преимущественно позволяет отобрать растения с изменениями только в соответствующих генах и уменьшает потенциальные негативные эффекты в отношении других метаболических процессов.
В данном документе описаны несколько геномных полинуклеотидных последовательностей SUS из Nicotiana tabacum, в том числе NtSUS1-S (SEQ ID NO: 1), NtSUS1-T (SEQ ID NO: 3), NtSUS2-S (SEQ ID NO: 5), NtSUS2-T (SEQ ID NO: 7), NtSUS3-S (SEQ ID NO: 9), NtSUS3-T (SEQ ID NO: 11), NtSUS4-S (SEQ ID NO: 13), NtSUS4-T (SEQ ID NO: 15), NtSUS5-S (SEQ ID NO: 17), NtSUS5-T (SEQ ID NO: 19), NtSUS6-S (SEQ ID NO: 21) и NtSUS6-T (SEQ ID NO: 23). Также раскрыты соответствующие предсказанные полипептидные последовательности для NtSUS1-S (SEQ ID NO: 2), NtSUS1-T (SEQ ID NO: 4), NtSUS2-S (SEQ ID NO: 6), NtSUS2-T (SEQ ID NO: 8), NtSUS3-S (SEQ ID NO: 10), NtSUS3-T (SEQ ID NO: 12), NtSUS4-S (SEQ ID NO: 14), NtSUS4-T (SEQ ID NO: 16), NtSUS5-S (SEQ ID NO: 18), NtSUS5-T (SEQ ID NO: 20), NtSUS6-S (SEQ ID NO: 22) и NtSUS6-T (SEQ ID NO: 24). NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T могут играть роль в метаболизме сахаров во время сушки. В частности, показано, что NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S играют роль в метаболизме сахаров во время сушки. Во время сушки табака табак трубоогневой сушки обычно содержит в по меньшей мере восемь раз больше редуцирующих сахаров, чем табак воздушной сушки, что обусловлено, главным образом, высоким уровнем накопления крахмала в табаке Вирджиния. Однако после сбора растений и во время процесса старения большая часть крахмала сначала превращается в сахарозу, а затем в редуцирующие сахара, по-видимому, с участием SUS, а также инвертаз, в качестве ключевого фермента для метаболизма сахарозы. Следовательно, настоящее изобретение особенно применимо в модулировании содержания редуцирующих сахаров в подвергнутом сушке растительном материале.
В одном аспекте описана клетка растения, содержащая: (i) полинуклеотид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 9, или SEQ ID NO: 11, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 21, или SEQ ID NO: 23; или полинуклеотид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5, или SEQ ID NO: 7, или SEQ ID NO: 13, или SEQ ID NO: 15; (ii) полипептид, кодируемый полинуклеотидом, представленным в (i); (iii) полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 94% идентичностью последовательности с SEQ ID NO: 2; или по меньшей мере 93% идентичностью последовательности с SEQ ID NO: 4; или по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 6; или по меньшей мере 96% идентичностью последовательности с SEQ ID NO: 8; или по меньшей мере 93% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12; или по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 14; или по меньшей мере 96% идентичностью последовательности с SEQ ID NO: 16; или по меньшей мере 89% идентичностью последовательности с SEQ ID NO: 18; или по меньшей мере 92% идентичностью последовательности с SEQ ID NO: 20; или по меньшей мере 93% идентичностью последовательности с SEQ ID NO: 22; или по меньшей мере 94% идентичностью последовательности с SEQ ID NO: 24; или (iv) конструкцию, вектор или вектор экспрессии, содержащие выделенный полинуклеотид, представленный в (i), где указанная клетка растения содержит по меньшей мере одну модификацию, которая модулирует экспрессию или активность полинуклеотида или полипептида по сравнению с клеткой контрольного растения, в котором экспрессия или активность полинуклеотида или полипептида не были модифицированы.
Также описан полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 69% идентичностью последовательности с SEQ ID NO: 2 или SEQ ID NO: 4; или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 6 или SEQ ID NO: 8; или по меньшей мере 74% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12; или по меньшей мере 76% идентичностью последовательности с SEQ ID NO: 14 или SEQ ID NO: 16; или по меньшей мере 68% идентичностью последовательности с SEQ ID NO: 18 или SEQ ID NO: 20; или по меньшей мере 69% идентичностью последовательности с SEQ ID NO: 22 или SEQ ID NO: 24.
Предпочтительно указанная клетка растения содержит полинуклеотид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5 или SEQ ID NO: 13 или по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 9 или SEQ ID NO: 11.
Предпочтительно указанная клетка растения содержит полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 81% идентичностью последовательности с SEQ ID NO: 6, или по меньшей мере 72% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12, или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 14.
Предпочтительно указанная клетка растения содержит полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 6, или по меньшей мере 74% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12, или по меньшей мере 76% идентичностью последовательности с SEQ ID NO: 14.
Предпочтительно, по меньшей мере одна модификация представляет собой модификацию генома клетки растения, или модификацию конструкции, вектора или вектора экспрессии, или трансгенную модификацию.
Предпочтительно, модификация генома клетки растения или модификация конструкции, вектора или вектора экспрессии представляет собой мутацию или редактирование.
Предпочтительно модификация увеличивает или уменьшает экспрессию или активность полинуклеотида или полипептида по сравнению с клеткой контрольного растения.
Предпочтительно, клетка растения содержит полинуклеотид для обеспечения интерференции, содержащий последовательность, которая на по меньшей мере 80% комплементарна по меньшей мере 19 нуклеотидам РНК, транскрибируемой с полинуклеотида в соответствии с пунктом формулы изобретения 1(i).
Предпочтительно модулированная экспрессия или активность полинуклеотида или полипептида модулируют уровень редуцирующих сахаров в листе, полученном из клетки растения, по сравнению с уровнем редуцирующего сахара в листе, полученном из контрольного растения, соответственно, где редуцирующий сахар представляет собой глюкозу или фруктозу, или, соответственно, где лист представляет собой лист, подвергнутый ранним стадиям сушки, или подвергнутый сушке лист.
В другом аспекте описано растение или его часть, содержащие клетку растения, описанную в данном документе.
В другом аспекте описан растительный материал, подвергнутый сушке растительный материал или гомогенизированный растительный материал, полученные из растения или его части, описанных в данном документе.
Предпочтительно растительный материал предусматривает биомассу, семя, стебель, цветки или листья растения или его части, описанных в данном документе.
Предпочтительно подвергнутый сушке растительный материал представляет собой растительный материал, подвергнутый трубоогневой сушке, солнечной сушке или воздушной сушке.
В другом аспекте описан табачный продукт, содержащий клетку растения, описанную в данном документе, часть растения, описанную в данном документе, или растительный материал, описанный в данном документе.
В другом аспекте описан способ получения растения, описанного в данном документе, включающий следующие стадии: (a) обеспечение клетки растения, содержащей полинуклеотид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 9, или SEQ ID NO: 11, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 21, или SEQ ID NO: 23; или характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5, или SEQ ID NO: 7, или SEQ ID NO: 13, или SEQ ID NO: 15; (b) модификацию клетки растения для модулирования экспрессии указанного полинуклеотида по сравнению с клеткой контрольного растения; и (c) размножение клетки растения с получением растения.
Предпочтительно, стадия (c) включает культивирование растения из побега или саженца, содержащих клетку растения.
Предпочтительно, стадия модификации клетки растения включает модификацию генома клетки с помощью методов редактирования генома или геномной инженерии.
Предпочтительно, методы редактирования генома или геномной инженерии выбраны из технологии CRISPR/Cas, мутагенеза, опосредованного нуклеазой с «цинковыми пальцами», химического или радиационного мутагенеза, гомологичной рекомбинации, олигонуклеотид-направленного мутагенеза и мутагенеза, опосредованного мегануклеазой.
Предпочтительно стадия модификации клетки растения включает трансфекцию клетки конструкцией, содержащей полинуклеотид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 9, или SEQ ID NO: 11, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 21, или SEQ ID NO: 23; или характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5, или SEQ ID NO: 7, или SEQ ID NO: 13, или SEQ ID NO: 15, функционально связанной с конститутивным промотором.
Предпочтительно, стадия модификации клетки растения включает введение в клетку полинуклеотида для обеспечения интерференции, содержащего последовательность, которая на по меньшей мере 80% комплементарна РНК, транскрибируемой с полинуклеотида в соответствии с пунктом формулы изобретения 1(i).
Предпочтительно клетку растения трансфицируют конструкцией, экспрессирующей полинуклеотид для обеспечения интерференции, содержащий последовательность, которая на по меньшей мере 80% комплементарна по меньшей мере 19 нуклеотидам РНК, транскрибируемой с полинуклеотида, описанного в данном документе.
В другом аспекте, описан способ получения подвергнутого сушке растительного материала с измененным количеством редуцирующих сахаров по сравнению с контрольным растительным материалом, включающий следующие стадии: (a) обеспечение растения или его части или растительного материала, описанных в данном документе; (b) сбор из них растительного материала; и (c) сушку растительного материала.
Предпочтительно, растительный материал включает подвергнутые сушке листья, подвергнутые сушке стебли или подвергнутые сушке цветки или их смесь.
Предпочтительно, способ сушки выбран из группы, состоящей из воздушной сушки, огневой сушки, дымовой сушки и трубоогневой сушки.
НЕКОТОРЫЕ ПРЕИМУЩЕСТВА
Преимущественно полинуклеотидные последовательности NtSUS, описанные в данном документе, экспрессируются во время сушки, в частности, с начала сушки. Модулирование экспрессии одной или более полинуклеотидных последовательностей NtSUS может приводить к модулированным уровням редуцирующих сахаров, таких как глюкоза и фруктоза, в подвергнутых сушке листьях табака. В частности, увеличение или уменьшение экспрессии одного или более полинуклеотидов NtSUS может приводить к увеличению или уменьшению уровней редуцирующих сахаров и различному восприятию табака, полученного из них, при нагревании.
В сигаретном дыме редуцирующие сахара оказывают влияние на высвобождение карбонильных компонентов, таких как формальдегид, ацетальдегид, ацетон, 2-фурфурал и акролеин (см. The Chemical Components of Tobacco and Tobacco Smoke, 2nd Edition, Rodgman and Perfetti, 2016). Следовательно, изменение содержания редуцирующих сахаров может менять высвобождение таких карбонильных соединений. Что касается высвобождения аромата в аэрозоле и дыме, глюкоза и в меньшей степени фруктоза могут образовывать соединения Амадори посредством реакции Майяра при взаимодействии с аминокислотами при нагревании. Хотя свободные аминокислоты рассматриваются как ограничивающие факторы в данной химической реакции, присутствие редуцирующих сахаров является необходимым для обеспечения образования соединений Амадори. Хлебный, ореховый или похожий на попкорн вкус и аромат получаются в результате нагревания пролина и глюкозы, таким образом, образуются продукты, подобные 2-ацетил-1-пирролину. Поскольку пролин и редуцирующие сахара являются особенно распространенными в табаке трубоогневой сушки, ореховые нотки получают при нагревании или курении табака Вирджиния. В случае смеси, как правило, акриламид (канцерогенное соединение, получаемое в результате взаимодействия глюкозы (фруктозы) с аспарагином при нагревании) может получаться в результате комбинации табака Берлей (с высоким содержанием аспарагина) с табаком Вирджиния (с высоким содержанием редуцирующих сахаров). Следовательно, модификация баланса редуцирующего сахара и аминокислоты может оказывать влиять на высвобождение как вкусоароматических соединений, так и акриламида в аэрозоле и дыме.
Преимущественно для надлежащего изготовления формованного листа из рекомбинированного табачного материала для нагреваемого табака зачастую требуются редуцирующие сахара. Модулирование экспрессии генов SUS во время сушки листа может оказывать влияние на содержание и баланс редуцирующих сахаров, тем самым воздействуя на изготовление формованного листа. Преимущественно могут быть созданы генетически не модифицированные растения, которые могут быть более приемлемыми для потребителей.
Преимущественно настоящее изобретение не ограничено применением растений, полученных с помощью EMS-мутагенеза. Растение, полученное с помощью EMS-мутагенеза, может характеризоваться меньшим потенциалом передачи улучшенных свойств сельскохозяйственной культуре после селекции. После начала селекции необходимая характеристика (характеристики) растения, полученного с помощью EMS-мутагенеза, по разным причинам может быть утрачена. Например, может потребоваться несколько мутаций, мутация может являться доминантной или рецессивной, и идентификация точечной мутации в гене-мишени может являться труднодостижимой. Напротив, в настоящем изобретении вводится применение полинуклеотидов NtAAT, которыми можно в частности манипулировать для получения растений с необходимым фенотипом. Настоящее изобретение можно применять к различным разновидностям растений или сельскохозяйственных культур.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1 представляет собой столбчатую диаграмму, на которой показано содержание редуцирующих сахаров после сбора (зрелые листья), после двух дней сушки (48 часов сушки) и в конце сушки для каждого сорта.
ПОДРОБНОЕ ОПИСАНИЕ
Заголовки разделов, используемые в настоящем раскрытии, служат для организационных целей и не предусматриваются как ограничивающие.
1. Определения
Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, какое обычно понимает специалист средней квалификации в данной области техники. В случае противоречий данный документ, включая определения, будет иметь преимущественную силу. Предпочтительные способы и материалы описаны ниже, хотя способы и материалы, сходные или эквивалентные описанным в данном документе, могут быть применены при осуществлении настоящего изобретения на практике или его тестировании. Материалы, способы и примеры, раскрытые в данном документе, являются лишь иллюстративными и не предусматриваются как ограничивающие.
Подразумевается, что термины «включают(включает)», «включают(включает) в себя», «характеризуются», «имеет», «может», «содержат(содержит)» и их варианты, используемые в данном документе, являются открытыми переходными фразами, терминами или словами, которые не исключают возможность наличия дополнительных действий или структур.
Формы единственного числа включают ссылки на множественное число, если из контекста явно не следует иное.
Термин «и/или» означает (а) или (b) или как (а), так и (b).
Настоящим изобретением предусмотрены другие варианты осуществления, «содержащие», «состоящие из» и «по существу состоящие из» вариантов осуществления или элементов, представленных в данном документе, независимо от того, указано это явно или нет.
В случае изложения в данном документе числовых диапазонов каждое промежуточное число в них предусматривается в явной форме с той же степенью точности. Например, в случае диапазона 6-9 в дополнение к 6 и 9 предусматриваются числа 7 и 8, а в случае диапазона 6,0-7,0 в явной форме предусматриваются числа 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 и 7,0.
Следующие термины, используемые во всем данном описании и формуле изобретения, имеют следующие значения.
Термины «кодирующая последовательность» или «полинуклеотид, кодирующий» означают нуклеотиды (молекулы РНК или ДНК), которые составляют полинуклеотид, который кодирует полипептид. Кодирующая последовательность может дополнительно содержать сигналы инициации и терминации, функционально связанные с регуляторными элементами, в том числе c промотором и сигналом полиаденилирования, способными управлять экспрессией в клетках индивидуума или млекопитающего, которому вводят полинуклеотид. Кодирующая последовательность может быть кодон-оптимизированной.
Выражения «комплементарная последовательность» или «комплементарный» могут означать спаривание оснований по Уотсону-Крику (например, A-T/U и C-G) или Хугстину между нуклеотидами или аналогами нуклеотидов. «Комплементарность» относится к свойству, присущему совместно двум полинуклеотидам, заключающемуся в том, что при их антипараллельном выравнивании друг относительно друга нуклеотидные основания в каждом положении комплементарны друг другу.
Термин «конструкция» относится к двунитевому фрагменту рекомбинантного полинуклеотида, предусматривающему один или более полинуклеотидов. Конструкция содержит «матричную нить», основания которой спарены с комплементарной «смысловой или кодирующей нитью». Указанная конструкция может быть вставлена в вектор в двух возможных ориентациях: либо в той же (или смысловой) ориентации, либо в противоположной (или антисмысловой) ориентации по отношению к ориентации промотора, расположенного в векторе, таком как вектор экспрессии.
Термин «контроль» в контексте контрольного растения или контрольных клеток растения означает растение или клетки растения, в которых экспрессия, функция или активность одного или более генов или полипептидов не была модифицирована (например, повышена или понижена), и поэтому они могут обеспечивать возможность сравнения с растением, в котором экспрессия, функция или активность одного или более таких же генов или полипептидов была модифицирована. Используемое в данном документе выражение «контрольное растение» означает растение, которое является практически эквивалентным тестируемому растению или модифицированному растению по всем параметрам, за исключением тестируемых параметров. Например, если речь идет о растении, в которое был введен полинуклеотид, контрольным растением является эквивалентное растение, в которое такой полинуклеотид не был введен. Контрольным растением может являться эквивалентное растение, в которое был введен контрольный полинуклеотид. В таких случаях контрольный полинуклеотид представляет собой полинуклеотид, для которого обуславливаемый им фенотипический эффект в отношении растения, как предполагается, является незначительным или отсутствует. Контрольное растение может содержать пустой вектор. Контрольное растение может соответствовать растению дикого типа. Контрольное растение может быть ноль-сегрегантом, при этом сегрегант T1 больше не содержит трансгена.
Термины «донорная ДНК» или «донорная матрица» относятся к фрагменту или молекуле двунитевой ДНК, которые содержат по меньшей мере часть гена, представляющего интерес. Донорная ДНК может кодировать полностью функциональный полипептид или частично функциональный полипептид.
Термин «эндогенный ген или полипептид» относится к гену или полипептиду, которые происходят из генома организма и не претерпели изменения, такого как потеря, приобретение или замена генетического материала. Эндогенный ген подвергается нормальному переносу гена и экспрессии гена. Эндогенный полипептид подвергается нормальной экспрессии.
Термин «энхансерные последовательности» относится к последовательностям, которые могут обеспечивать повышение экспрессии гена. Эти последовательности могут быть расположены выше, в пределах интронов или ниже транскрибируемого участка. Транскрибируемый участок состоит из экзонов и расположенных между ними интронов, от промотора до участка терминации транскрипции. Усиление экспрессии гена может происходить посредством различных механизмов, включая повышение эффективности транскрипции, стабилизацию зрелой мРНК и усиление трансляции.
Термин «экспрессия» относится к выработке функционального продукта. Например, экспрессия фрагмента полинуклеотида может относиться к транскрипции фрагмента полинуклеотида (например, транскрипции, приводящей к получению мРНК или функциональной РНК) и/или трансляции мРНК с получением полипептида-предшественника или зрелого полипептида. «Сверхэкспрессия» относится к выработке продукта гена в трансгенных организмах на уровнях, которые превышают уровни выработки в ноль-сегрегантном (или нетрансгенном) организме из того же эксперимента.
Термины «функциональный» и «полностью функциональный» описывают полипептид, который обладает биологической функцией или активностью. Термин «функциональный ген» относится к гену, транскрибируемому с образованием мРНК, которая транслируется с образованием функционального или активного полипептида.
Термин «генетическая конструкция» относится к молекулам ДНК или РНК, которые содержат полинуклеотид, который кодирует полипептид. Кодирующая последовательность может содержать сигналы инициации и терминации, функционально связанные с регуляторными элементами, в том числе c промотором и сигналом полиаденилирования, способными управлять экспрессией.
Термин «редактирование генома» относится к такому изменению эндогенного гена, который кодирует эндогенный полипептид, при котором достигают экспрессии полипептида, представляющего собой усеченный эндогенный полипептид или эндогенный полипептид, имеющий аминокислотную замену. Редактирование генома может включать замещение участка эндогенного гена, подлежащего нацеливанию, или замещение всего эндогенного гена копией гена, которая характеризуется наличием усечения или аминокислотной замены, с помощью механизма репарации, такого как HDR. Редактирование генома также может включать создание аминокислотной замены в эндогенном гене посредством создания двунитевого разрыва в эндогенном гене, который затем репарируют с применением NHEJ. С помощью NHEJ можно осуществлять добавление или делецию по меньшей мере одной пары оснований в ходе репарации, вследствие чего можно получить аминокислотную замену. Редактирование генома также может включать удаление сегмента гена с помощью одновременного действия двух нуклеаз на одну и ту же нить ДНК для осуществления усечения между двумя сайтами-мишенями нуклеаз, и репарацию разрыва ДНК с помощью NHEJ.
Термин «гетерологичный» в отношении последовательности означает последовательность, которая происходит из чужеродного вида или, в случае, если она происходит из того же вида, является существенно модифицированной по сравнению со своей нативной формой по составу и/или геномному локусу вследствие преднамеренного вмешательства человека.
Термины «репарация путем гомологичной рекомбинации» или «HDR» относятся к механизму репарации двунитевых повреждений ДНК в клетках, если гомологичный фрагмент ДНК присутствует в ядре, главным образом в G2- и S-фазе клеточного цикла. В ходе HDR применяют донорную ДНК или донорную матрицу для направления репарации, и ее можно применять для создания конкретных изменений последовательностей в геноме, в том числе для нацеленного добавления целых генов. Если донорная матрица предоставляется вместе с сайт-специфичной нуклеазой, то клеточный аппарат будет репарировать разрыв с помощью гомологичной рекомбинации, которая усиливается по величине на несколько порядков при наличии расщепления ДНК. Если гомологичный фрагмент ДНК отсутствует, то вместо этого может происходить NHEJ.
Термины «гомология» или «сходство» относятся к степени сходства последовательностей двух полипептидов или двух молекул полинуклеотида, сравниваемых путем выравнивания последовательностей. Степень гомологии между двумя отдельными сравниваемым полинуклеотидами является функцией числа идентичных или совпадающих нуклеотидов в сопоставляемых положениях.
Термины «идентичный» или «идентичность» в контексте двух или более полинуклеотидов или полипептидов означают, что последовательности характеризуются наличием определенной процентной доли остатков, которые являются одинаковыми в пределах определенного участка. Процентную долю можно рассчитать путем оптимального выравнивания двух последовательностей, сравнения двух последовательностей в пределах определенного участка, определения числа положений, в которых в обеих последовательностях находятся идентичные остатки, с получением числа совпадающих положений, деления числа совпадающих положений на общее число положений в определенном участке и умножения результата на 100 с получением процентной доли идентичности последовательностей. В тех случаях, когда эти две последовательности имеют разную длину или при выравнивании создается один или более несимметрично расположенных концов, и определенный участок сравнения включает только одну последовательность, то при расчете остатки одиночной последовательности включаются в знаменатель, а не в числитель. При сравнении ДНК и РНК тимин (Т) и урацил (U) можно считать эквивалентными. Идентичность можно определять самостоятельно или с помощью компьютерного алгоритма для работы с последовательностями, такого как ClustalW, ClustalX, BLAST, FASTA или алгоритм Смита-Уотермана. Популярная программа множественного выравнивания ClustalW (Nucleic Acids Research (1994) 22, 4673-4680; Nucleic Acids Research (1997), 24, 4876-4882) представляет собой подходящий способ получения множественных выравниваний полипептидов или полинуклеотидов. Подходящие параметры для ClustalW могут являться следующими. Для выравниваний полинуклеотидов: штраф за открытие гэпа=15,0, штраф за продолжение гэпа=6,66 и матрица=идентичность. Для выравниваний полипептидов: штраф за открытие гэпа=10,0, штраф за продолжение гэпа=0,2 и матрица=Gonnet. Для выравниваний ДНК и белка: ENDGAP = -1 и GAPDIST=4. Специалистам в данной области будет понятно, что может быть необходимым изменять эти и другие параметры для оптимального выравнивания последовательностей. Предпочтительно, затем проводят расчет процентной доли идентичностей на основании такого выравнивания в виде (N/T), где N представляет собой число положений, в которых в последовательностях присутствует идентичный остаток, и T представляет собой общее число сравниваемых положений, включая гэпы, но за исключением выступов.
Термины «повышение» или «повышенный» относятся к повышению, составляющему от приблизительно 10% до приблизительно 99%, или повышению, составляющему по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 100%, по меньшей мере 150%, или по меньшей мере 200%, или больше, количества, или функции, или активности, такой как без ограничения функция или активность полипептида, транскрипционная функция или активность и/или экспрессия белка. Термин «повышенный» или фраза «повышенное количество» может относиться к количеству, или функции, или активности модифицированного растения или продукта, полученного из модифицированного растения, которое является большим, чем можно обнаружить в растении или продукте из той же разновидности растения, обработанного таким же образом, которое не было модифицировано. Таким образом, в некоторых случаях растение дикого типа той же разновидности, которое было переработано таким же образом, используется в качестве контроля, с помощью которого измеряют, достигнуто ли повышение количества.
Термин «увеличение» или «увеличенный», используемый в данном документе, относится к снижению, составляющему от приблизительно 10% до приблизительно 99%, или снижению на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99% или по меньшей мере 100%, или по меньшей мере 150%, или по меньшей мере 200% и более от количества или функции, такой как функция полипептида, транскрипционная функция или экспрессия полипептида. Термин «повышенный» или фраза «повышенное количество» может относиться к количеству или функции в модифицированном растении или продукте, полученном из модифицированного растения, которое является меньшим, чем можно обнаружить в растении или продукте из той же разновидности растения, обработанного таким же образом, которое не было модифицировано. Таким образом, в некоторых случаях растение дикого типа той же разновидности, которое было переработано таким же образом, используется в качестве контроля, с помощью которого измеряют, достигнуто ли уменьшение количества.
Термин «ингибировать» или «ингибированный» относится к уменьшению, составляющему от приблизительно 98% до приблизительно 100%, или уменьшению, составляющему по меньшей мере 98%, по меньшей мере 99% и, в частности, 100%, количества, или функции, или активности, как, например, без ограничения, функции или активности полипептида, транскрипционной функции или активности и/или экспрессии полипептида.
Термин «введенный» означает доставку полинуклеотида (например, конструкции) или полипептида в клетку. Выражение «введенный» включает ссылку на встраивание полинуклеотида в эукариотическую клетку, где полинуклеотид может быть встроен в геном клетки, а также включает ссылку на транзиентное внедрение полинуклеотида или полипептида в клетку. Выражение «введенный» включает ссылки на способы стабильной или транзиентной трансформации, а также на скрещивание половым путем. Таким образом, выражение «введенный» применительно к вставке полинуклеотида (например, рекомбинантной конструкции/экспрессионной конструкции) в клетку означает «трансфекцию», или «трансформацию», или «трансдукцию» и включает ссылку на встраивание полинуклеотида в эукариотическую клетку, где полинуклеотид может быть встроен в геном клетки (например, хромосомную, плазмидную, пластидную или митохондриальную ДНК), преобразован в автономный репликон или экспрессироваться транзиентно (например, трансфицированная мРНК).
Термины «выделенный» или «очищенный» относятся к материалу, который практически или по существу не содержит компоненты, которые обычно сопутствуют ему, как встречается в его нативном состоянии. Как правило, чистоту и однородность определяют с помощью методик аналитической химии, таких как электрофорез в полиакриламидном геле или высокоэффективная жидкостная хроматография. Полипептид, который является преобладающей молекулой, присутствующей в препарате, является практически очищенным. В частности, выделенный полинуклеотид отделяют от открытых рамок считывания, которые фланкируют требуемый ген и кодируют полипептиды, отличные от необходимого полипептида. Термин «очищенный», применяемый в данном документе, обозначает, что полинуклеотид или полипептид дают по существу одну полосу в электрофорезном геле. В частности, это означает, что полинуклеотид или полипептид являются на по меньшей мере 85% чистыми, более предпочтительно на по меньшей мере 95% чистыми и наиболее предпочтительно на по меньшей мере 99% чистыми. Выделенные полинуклеотиды могут быть очищены из клетки-хозяина, в которой они встречаются в природе. Для получения выделенных полинуклеотидов можно применять общепринятые способы очистки нуклеиновых кислот, известные специалистам в данной области техники. Данный термин также охватывает рекомбинантные полинуклеотиды и химически синтезируемые полинуклеотиды.
Термины «модулировать» или «модулирование» относятся к обеспечению или облегчению качественного или количественного изменения, корректировки или модификации способа, пути, функции или активности, представляющих интерес. Без ограничения такое изменение, корректировка или модификация может представлять собой повышение или понижение уровня соответствующего процесса, пути, функции или активности, представляющих интерес. Например, можно модулировать экспрессию гена или экспрессию полипептида или функцию или активность полипептида. Как правило, относительное изменение, корректировку или модификацию определяют посредством сравнения с контролем.
Термин «путь негомологичного соединения концов (NHEJ)», используемый в данном документе, относится к пути, посредством которого происходит репарация двунитевых разрывов в ДНК путем прямого лигирования концов разрывов без необходимости в гомологичной матрице. Независимое от матрицы повторное лигирование концов ДНК посредством NHEJ является стохастическим, подверженным ошибкам процессом репарации, в ходе которого в точку разрыва ДНК вводятся случайные микровставки и микроделеции (вставки/делеции). Данный способ можно применять для преднамеренного разрушения, делеции или корректировки рамки считывания в последовательностях генов-мишеней. Как правило, в ходе NHEJ используются короткие гомологичные ДНК-последовательности, называемые микрогомологами, для направления репарации. Эти микрогомологи часто присутствуют в однонитевых выступах на концах двунитевых разрывов. Если выступы полностью совместимы, то в ходе NHEJ обычно происходит точная репарация разрыва, хотя может иметь место также неточная репарация, приводящая к потере нуклеотидов, однако гораздо более распространены случаи, когда выступы несовместимы.
Термин «не встречающийся в природе» описывает объект, такой как полинуклеотид, генетическую мутацию, полипептид, растение, клетку растения и растительный материал, который не образован естественным путем или не существует в природе. Такие не встречающиеся в природе объекты или искусственные объекты можно создать, синтезировать, осуществить их инициацию, модифицировать, подвергнуть вмешательству или манипуляции способами, описанными в данном документе, или которые известны в данной области техники. Такие не встречающиеся в природе объекты или искусственные объекты могут быть созданы, синтезированы, инициированы, модифицированы, подвергнуты вмешательству или манипуляции человеком. Таким образом, в качестве примера, не встречающееся в природе растение, не встречающуюся в природе клетку растения или не встречающийся в природе растительный материал можно создать с применением традиционных методик селекции растений, таких как обратное скрещивание, или с помощью технологий манипуляции с генами, например, с применением антисмысловой РНК, интерферирующей РНК, мегануклеазы и т.п. В качестве дополнительного примера, не встречающееся в природе растение, не встречающуюся в природе клетку растения или не встречающийся в природе растительный материал можно создать посредством интрогрессии или путем переноса одной или более генетических мутаций (например, одного или более полиморфизмов) от первого растения или клетки растения ко второму растению или клетке растения (которые сами по себе могут быть встречающимися в природе), таким образом, что полученное растение, клетка растения или растительный материал или их потомство содержит генетическую структуру (например, геном, хромосому или ее сегмент), которая не образуется естественным путем, или которая не существует в природе. Полученное растение, клетка растения или растительный материал, таким образом, являются искусственными или не встречающимися в природе. Соответственно, искусственные или не встречающиеся в природе растение или клетку растения можно создать путем модификации генетической последовательности в первом встречающемся в природе растении или клетке растения, даже если полученная генетическая последовательность встречается в природе во втором растении или клетке растения, которые содержат генетический фон, отличный от такового у первого растения или клетки растения. В определенных вариантах осуществления мутация не является встречающейся в природе мутацией, которая существует в природе в полинуклеотиде или полипептиде, таких как ген или полипептид. Различия в генетическом фоне можно выявить по фенотипическим различиям или с помощью методик молекулярной биологии, известных из уровня техники, таких как секвенирование полинуклеотида, определение наличия или отсутствия генетических маркеров (например, маркеров, представляющих собой микросателлитные РНК).
Термины «олигонуклеотид» или «полинуклеотид» означают по меньшей мере два нуклеотида, ковалентно связанных вместе. Описание отдельной нити также определяет последовательность комплементарной нити. Таким образом, полинуклеотид также охватывает нить, комплементарную описанной отдельной нити. Многие варианты полинуклеотида могут использоваться для той же цели, что и указанный полинуклеотид. Таким образом, полинуклеотид также охватывает практически идентичные полинуклеотиды и комплементарные им последовательности. Отдельная нить представляет собой зонд, который может гибридизироваться с данной последовательностью в жестких условиях гибридизации. Таким образом, полинуклеотид также охватывает зонд, который гибридизируется в жестких условиях гибридизации. Полинуклеотиды могут быть однонитевыми или двунитевыми или могут содержать части как двунитевой, так и однонитевой последовательности. Полинуклеотид может представлять собой ДНК, как геномную, так и кДНК, РНК или гибридную молекулу, где полинуклеотид может содержать комбинации дезоксирибо- и рибонуклеотидов, а также комбинации оснований, в том числе урацила, аденина, тимина, цитозина, гуанина, инозина, ксантина, гипоксантина, изоцитозина и изогуанина. Полинуклеотиды можно получать с помощью способов химического синтеза или с помощью рекомбинантных способов.
Специфичность однонитевой ДНК в отношении гибридизации с комплементарными фрагментами определяется «жесткостью» условий реакции (Sambrook et al., Molecular Cloning and Laboratory Manual, Second Ed., Cold Spring Harbor (1989)). Жесткость гибридизации повышается по мере понижения склонности к образованию ДНК-дуплексов. При реакциях гибридизации полинуклеотидов жесткость можно выбирать таким образом, чтобы содействовать реакциям гибридизации, характеризующимся специфичностью (высокая жесткость), которые можно применять для идентификации, например, клонов полной длины из библиотеки. Реакции гибридизации, характеризующиеся меньшей специфичностью (низкая жесткость), можно применять для идентификации родственных, но не точно соответствующих (гомологичных, но не идентичных) молекул или сегментов ДНК. ДНК-дуплексы стабилизируют посредством (1) определенного числа комплементарных пар оснований; (2) определенного типа пар оснований; (3) концентрации солей (ионной силы) в реакционной смеси; (4) температуры реакции и (5) присутствия определенных органических растворителей, таких как формамид, которые понижают стабильность ДНК-дуплекса. Обычно чем длиннее зонд, тем выше температура, необходимая для надлежащего отжига. Общепринятый подход заключается в изменении температуры; более высокие относительные температуры приводят к более жестким условиям реакции. Для гибридизации в «жестких условиях» описаны протоколы гибридизации, в которых полинуклеотиды, на по меньшей мере 60% гомологичные друг другу, остаются гибридизированными. Обычно жесткие условия выбирают таким образом, чтобы температура была приблизительно на 5°C ниже, чем температура точки плавления (Tm) для конкретной последовательности при определенных значениях ионной силы и рН. Tm представляет собой температуру (при определенных значениях ионной силы, рН и концентрации полинуклеотида), при которой 50% зондов комплементарно данной последовательности, гибридизируются с данной последовательностью в равновесном состоянии. Поскольку данные последовательности обычно присутствуют в избытке, то при Tm 50% зондов заняты в равновесном состоянии.
«Жесткие условия гибридизации» представляют собой условия, которые позволяют зонду, праймеру или олигонуклеотиду гибридизироваться только со своей конкретной последовательностью. Жесткие условия зависят от последовательности и будут различаться. Жесткие условия как правило включают: (1) низкую ионную силу и промывки при высокой температуре, например 15 мМ хлорида натрия, 1,5 мМ цитрата натрия, 0,1% додецилсульфата натрия при 50°C; (2) присутствие денатурирующего средства во время гибридизации, например 50% (об./об.) формамида, 0,1% бычьего сывороточного альбумина, 0,1% фиколла, 0,1% поливинилпирролидона, 50 мМ натрий-фосфатного буфера (750 мМ хлорида натрия, 75 мМ цитрата натрия; pH 6,5) при 42°C или (3) присутствие 50% формамида. Как правило, промывки также предусматривают 5 x SSC (0,75 M NaCl, 75 мМ цитрата натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5x раствор Денхардта, ДНК из молок лососевых рыб, подвергнутую ультразвуковой обработке (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42°C с промывкой при 42°C в 0,2 x SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C и последующей промывкой в условиях высокой жесткости, предусматривающей 0,1 x SSC, содержащий EDTA, при 55°C. Предпочтительно, условия являются такими, что последовательности, гомологичные друг другу на по меньшей мере приблизительно 65%, 70%, 75%, 85%, 90%, 95%, 98% или 99%, обычно остаются гибридизированными друг с другом.
При «условиях умеренной жесткости» используют растворы для промывки и условия гибридизации, которые являются менее жесткими, такими, что полинуклеотид гибридизируется со всем рассматриваемым полинуклеотидом, его фрагментами, производными или аналогами. Один пример предусматривает гибридизацию в 6x SSC, 5x растворе Денхардта, 0,5% SDS и 100 мкг/мл денатурированной ДНК из молок лососевых рыб при 55°C с последующими одной или более промывками в 1 × SSC, 0,1% SDS при 37°C. Температуру, ионную силу и т. д. можно регулировать для обеспечения соответствия экспериментальным факторам, таким как длина зонда. Были описаны другие условия умеренной жесткости (см. Ausubel et al., Current Protocols in Molecular Biology, Volumes 1-3, John Wiley & Sons, Inc., Hoboken, N.J. (1993); Kriegler, Gene Transfer and Expression: A Laboratory Manual, Stockton Press, New York, N.Y. (1990); Perbal, A Practical Guide to Molecular Cloning, 2nd edition, John Wiley & Sons, New York, N.Y. (1988)).
При «условиях низкой жесткости» используют растворы для промывки и условия гибридизации, которые являются менее жесткими, чем таковые в случае умеренной жесткости, такие, что полинуклеотид гибридизируется со всем рассматриваемым полинуклеотидом, его фрагментами, производными или аналогами. Неограничивающий пример условий гибридизации низкой жесткости предусматривает гибридизацию в 35% формамиде, 5x SSC, 50 мМ Tris-HCl (pH 7,5), 5 мМ EDTA, 0,02% PVP, 0,02% фиколле, 0,2% BSA, 100 мкг/мл денатурированной ДНК из молок лососевых рыб, 10% (вес./об.) сульфате декстрана при 40°C с последующими одной или более промывками в 2x SSC, 25 мМ Tris-HCl (pH 7,4), 5 мМ EDTA и 0,1% SDS при 50°C. Хорошо описаны другие условия низкой жесткости, такие как условия для вариантов межвидовой гибридизации (см. Ausubel et al., 1993; Kriegler, 1990).
Термин «функционально связанный» означает, что экспрессия гена находится под контролем промотора, с которым он пространственно соединен. Промотор может быть расположен в 5'-направлении (выше) или 3'-направлении (ниже) от гена, который находится под его контролем. Расстояние между промотором и геном может быть примерно таким же, как расстояние между этим промотором и геном, который он контролирует, в гене, из которого получен промотор. Как известно из уровня техники, изменение этого расстояния можно согласованно осуществлять без потери функции промотора. Термин «функционально связанный» относится к ассоциации фрагментов полинуклеотида в одном фрагменте таким образом, что функция одного регулируется другим. Например, промотор функционально связан с фрагментом полинуклеотида, если он способен регулировать транскрипцию данного фрагмента полинуклеотида.
Термин «растение» относится к любому растению на любой стадии его жизненного цикла или развития и его потомкам. В одном варианте осуществления растение представляет собой растение табака, которое относится к растению, принадлежащему к роду Nicotiana. Термин включает ссылку на целые растения, органы растения, ткани растения, ростки растения, семена растения и клетки растения и их потомство. Клетки растения включают без ограничения клетки из семян, суспензионных культур, зародышей, меристематических участков, каллюсной ткани, листьев, корней, побегов, гаметофитов, спорофитов, пыльцы и микроспор. Подходящие виды, сорта, гибриды и разновидности растений табака описаны в данном документе.
Термины «полинуклеотид», «полинуклеотидная последовательность» или «полинуклеотидный фрагмент» используются в данном документе взаимозаменяемо и относятся к полимеру из РНК или ДНК, который является одно- или двунитевым и необязательно содержит синтетические, неприродные или скорректированные нуклеотидные основания. Нуклеотиды (обычно находящиеся в форме их 5'-монофосфата) обозначают с помощью их однобуквенных обозначений следующим образом: «А» для аденилата или дезоксиаденилата (соответственно для РНК или ДНК), «C» для цитидилата или дезоксицитидилата, «G» для гуанилата или дезоксигуанилата, «U» для уридилата, «T» для дезокситимидилата, «R» для пуринов (A или G), «Y» для пиримидинов (C или T), «K» для G или T, «H» для A, или С, или Т, «I» для инозина и «N» для любого нуклеотида. Полинуклеотид может представлять собой без ограничения геномную ДНК, комплементарную ДНК (кДНК), мРНК или антисмысловую РНК или их фрагмент(фрагменты). Кроме того, полинуклеотид может быть однонитевым или двунитевым, смесью однонитевых и двунитевых участков, гибридной молекулой, содержащей ДНК и РНК, или гибридной молекулой со смесью однонитевых и двунитевых участков или их фрагмента(фрагментов). Дополнительно полинуклеотид может быть составлен из трехнитевых участков, содержащих ДНК, РНК или обе, или их фрагмент(фрагменты). Полинуклеотид может содержать одно или более модифицированных оснований, таких как фосфотиоаты, и может представлять собой пептидную нуклеиновую кислоту (PNA). Как правило, полинуклеотиды могут быть собраны из выделенных или клонированных фрагментов кДНК, геномной ДНК, олигонуклеотидов или отдельных нуклеотидов или комбинации вышеперечисленного. Хотя полинуклеотиды, описанные в данном документе, представлены в виде ДНК-последовательностей, полинуклеотиды включают их соответствующие РНК-последовательности и их комплементарные (например, полностью комплементарные) ДНК- или РНК-последовательности, в том числе цепи, обратно комплементарные им. Полинуклеотиды по настоящему изобретению представлены в прилагаемом перечне последовательностей.
Термины «полипептид» или «полипептидная последовательность» относятся к полимеру из аминокислот, в котором один или более аминокислотных остатков представляют собой искусственный химический аналог соответствующей встречающейся в природе аминокислоты, а также к встречающимся в природе полимерам из аминокислот. Термины также подразумевают модификации, в том числе без ограничения гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и ADP-рибозилирование. Полипептиды по настоящему изобретению представлены в прилагаемом перечне последовательностей.
Термин «промотор» означает синтетическую или полученную природным способом молекулу, которая способна обеспечивать, активировать или усиливать экспрессию полинуклеотида в клетке. Термин относится к элементу/последовательности полинуклеотида, расположенным, как правило, выше по последовательности и функционально связанным с фрагментом двунитевого полинуклеотида. Промоторы могут быть получены целиком из участков вблизи нативного гена, представляющего интерес, или могут состоять из разных элементов, полученных из разных нативных промоторов или сегментов синтетического полинуклеотида. Промотор может содержать одну или более специфических последовательностей, регулирующих транскрипцию, для дополнительного усиления экспрессии и/или корректировки экспрессии пространственно и/или по времени. Промотор может также содержать дистальные энхансерные или репрессорные элементы, которые могут быть расположены на расстоянии до несколько тысяч пар оснований от сайта начала транскрипции. Промотор может быть получен из источников, включающих вирусы, бактерии, грибы, растения, насекомых и животных. Промотор может регулировать экспрессию компонента гена конститутивно или дифференциально по отношению к клетке, ткани или органу, в которых происходит экспрессия, или по отношению к стадии развития, на которой происходит экспрессия, или в ответ на внешние стимулы, такие как физиологические нагрузки, патогены, ионы металлов или индуцирующие средства.
Выражения «тканеспецифичный промотор» и «промотор, предпочтительный для определенной ткани», используемые в данном документе взаимозаменяемо, относятся к промотору, который экспрессируется преимущественно, но не обязательно исключительно, в одном органе или ткани, но может экспрессироваться также в одной конкретной клетке. Выражение «промотор, регулируемый в процессе развития» относится к промотору, функция которого определяется событиями, связанными с развитием. Выражение «конститутивный промотор» относится к промотору, который вызывает экспрессию гена в большинстве типов клеток в большинстве случаев. «Индуцируемый промотор» обеспечивает избирательную экспрессию функционально связанной ДНК-последовательности в ответ на присутствие эндогенных или экзогенных стимулов, например, с помощью химических соединений (химических индукторов) или в ответ на сигналы окружающей среды, гормональные, химические сигналы и/или сигналы, связанные с развитием. Примеры индуцируемых или регулируемых промоторов включают промоторы, регулируемые светом, теплом, стрессом, наводнением или засухой, патогенами, фитогормонами, ранениями или химическими веществами, такими как этанол, жасмонат, салициловая кислота или антидоты.
Термин «рекомбинантный», используемый в данном документе, относится к искусственной комбинации из двух в иных обстоятельствах разделенных сегментов последовательности, полученной, например, посредством химического синтеза или посредством манипуляции с выделенными сегментами полинуклеотидов с помощью методик генной инженерии. Термин также включает ссылку на клетку или вектор, которые были модифицированы путем введения гетерологичного полинуклеотида, или клетку, полученную из модифицированной таким образом клетки, но не охватывает корректировку клетки или вектора в результате встречающихся в природе событий (например, в результате спонтанной мутации, естественной трансформации, или трансдукции, или транспозиции), таких как те, которые происходят без преднамеренного вмешательства человека.
Выражение «рекомбинантная конструкция» относится к комбинации полинуклеотидов, которые обычно не встречаются в природе вместе. Соответственно, рекомбинантная конструкция может содержать регуляторные последовательности и кодирующие последовательности, полученные из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные иначе, чем это обычно встречается в природе. Рекомбинантная конструкция может представлять собой рекомбинантную ДНК-конструкцию.
Выражения «регуляторные последовательности» и «регуляторные элементы», используемые в данном документе взаимозаменяемо, относятся к полинуклеотидным последовательностям, расположенным выше (5'-некодирующие последовательности), в пределах или ниже (3'-некодирующие последовательности) кодирующей последовательности, которые влияют на транскрипцию, процессинг или стабильность РНК или на трансляцию связанной кодирующей последовательности. Регуляторные последовательности включают промоторы, лидерные последовательности, регулирующие трансляцию, интроны и распознаваемые последовательности полиаденилирования. Термины «регуляторная последовательность» и «регуляторный элемент» используются в данном документе взаимозаменяемо.
Выражение «сайт-специфичная нуклеаза» относится к ферменту, способному к специфичному распознаванию и расщеплению ДНК-последовательностей. Сайт-специфичная нуклеаза может быть сконструированной. Примеры сконструированных сайт-специфичных нуклеаз включают нуклеазы с «цинковыми пальцами» (ZFN), TAL-эффекторные нуклеазы (TALEN), системы на основе CRISPR/Cas9 и мегануклеазы.
Термин «табак» используют в собирательном смысле для обозначения табачных культур (например, множества растений табака, выращиваемых в поле, и табака, выращиваемого не методом гидропоники), растений табака и их частей, в том числе без ограничения корней, стеблей, листьев, цветов и семян, подготовленных и/или полученных, как описано в данном документе. Понятно, что «табак» включает растения Nicotiana tabacum и продукты из них.
Термин «табачные продукты» относится к потребительским табачным продуктам, в том числе, без ограничения, к курительным материалам (например, сигаретам, сигарам и трубочному табаку), нюхательному табаку, жевательному табаку, жевательной резинке и леденцам, а также компонентам, материалам и ингредиентам для производства потребительских табачных продуктов. Предпочтительно, данные табачные продукты производят из листьев и стеблей табака, собранных с табака и нарезанных, высушенных, подвергнутых сушке и/или ферментированных в соответствии с общепринятыми методиками получения табака.
Выражения «терминатор транскрипции», «последовательности терминации» или «терминатор» относятся к ДНК-последовательностям, расположенным ниже кодирующей последовательности, включающим распознаваемые последовательности полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные воздействовать на процессинг мРНК или экспрессию гена. Сигнал полиаденилирования обычно характеризуется осуществлением добавления трактов полиадениловой кислоты на 3'-конец предшественника мРНК.
Термин «трансгенный» относится к любой клетке, линии клеток, каллюсу, ткани, части растения или растению, геном которых был скорректирован в результате присутствия гетерологичного полинуклеотида, такого как рекомбинантная конструкция, в том числе к исходным трансгенным объектам, а также полученным с помощью процедур полового скрещивания или бесполого размножения из исходного трансгенного объекта. Термин не охватывает корректировку генома (хромосомную или внехромосомную) с помощью общепринятых способов селекции растений или в результате встречающихся в природе событий, таких как случайное перекрестное опыление, инфекция, вызванная нерекомбинантным вирусом, трансформация нерекомбинантными бактериями, нерекомбинантная транспозиция или спонтанная мутация.
Выражение «трансгенное растение» относится к растению, которое содержит в своем геноме один или более гетерологичных полинуклеотидов, т.е. растение, которое содержит рекомбинантный генетический материал, обычно не обнаруживаемый в нем, и который был введен в рассматриваемое растение (или в предков растения) посредством манипуляции, осуществляемой человеком. Например, гетерологичный полинуклеотид может быть стабильно интегрирован в геном таким образом, что полинуклеотид передается последующим поколениям. Гетерологичный полинуклеотид может быть интегрирован в геном отдельно или в виде части рекомбинантной конструкции. Коммерческая разработка генетически улучшенной идиоплазмы также продвинулась к стадии введения в культурные растения нескольких признаков, что часто называют подходом на основе пирамидирования генов. В этом подходе в растение можно ввести несколько генов, придающих разные характеристики, представляющие интерес. Пирамидирование генов можно осуществлять многими способами, в том числе без ограничения путем котрансформации, повторной трансформации и скрещивания линий с разными трансгенами. Таким образом, растение, выращиваемое из клетки растения, в которую рекомбинантную ДНК вводят с помощью трансформации, является трансгенным растением, равно как и все потомство данного растения, которое содержит введенный трансген (полученное как половым, так и бесполым путем). Понятно, что термин «трансгенное растение» охватывает все растение или дерево и части растения или дерева, например зерна, семена, цветки, листья, корни, плоды, пыльцу, стебли и т.п. Каждый гетерологичный полинуклеотид может придавать трансгенному растению отдельный признак.
Термин «эффектор, подобный активаторам транскрипции» или «TALE» относится к полипептидной структуре, которая распознает определенную ДНК-последовательность и связывается с ней. Выражение «ДНК-связывающий домен TALE» относится к ДНК-связывающему домену, который содержит массив тандемных повторов из 33-35 аминокислот, также известных как RVD-модули, каждый из которых специфично распознает одну пару оснований ДНК. RVD-модули могут располагаться в любом порядке, собираясь в массив, который распознает определенную последовательность. Специфичность связывания ДНК-связывающего домена TALE определяется массивом RVD, за которым расположен один усеченный повтор из 20 аминокислот. ДНК-связывающий домен TALE может иметь от 12 до 27 RVD-модулей, каждый из которых содержит RVD и распознает одну пару оснований ДНК. Были идентифицированы специфичные RVD, которые распознают каждый из четырех возможных нуклеотидов ДНК (А, Т, С и G). Поскольку ДНК-связывающие домены TALE являются модульными, то повторы, которые распознают четыре разных нуклеотида ДНК, можно соединить друг с другом для распознавания любой конкретной ДНК-последовательности. Эти нацеленные ДНК-связывающие домены можно впоследствии объединить с каталитическими доменами для создания функциональных ферментов, в том числе искусственных факторов транскрипции, метилтрансфераз, интеграз, нуклеаз и рекомбиназ.
Термины «эффекторные нуклеазы, подобные активаторам транскрипции» или «TALEN», используемые в данном документе взаимозаменяемо, относятся к сконструированным слитым полипептидам на основе каталитического домена нуклеазы, такой как эндонуклеаза FokI, и разработанного ДНК-связывающего домена TALE, который может быть нацелен на специально синтезированную ДНК-последовательность.
Выражение «мономер TALEN» относится к сконструированному слитому полипептиду с каталитическим доменом нуклеазы и разработанным ДНК-связывающим доменом TALE. Два мономера TALEN могут быть разработаны таким образом, чтобы они нацеливались на участок, являющийся мишенью для TALEN, и расщепляли его.
Термин «трансген» относится к гену или генетическому материалу, содержащему последовательность гена, которые были выделены из одного организма и введены в другой организм. Этот ненативный сегмент ДНК может сохранять способность к обеспечению выработки РНК или полипептида в трансгенном организме или он может обеспечивать корректировку нормальной функции генетического кода трансгенного организма. Введение трансгена имеет потенциал к изменению фенотипа организма.
Термин «вариант» по отношению к полинуклеотиду означает: (i) часть или фрагмент полинуклеотида; (ii) последовательность, комплементарную полинуклеотиду или его части; (iii) полинуклеотид, практически идентичный упоминаемому полинуклеотиду или комплементарной ему последовательности; или (iv) полинуклеотид, который гибридизируется в жестких условиях с упоминаемым полинуклеотидом, комплементарной ему последовательностью или практически идентичным ей полинуклеотидом.
Термин «вариант» по отношению к пептиду или полипептиду означает пептид или полипептид, которые отличаются по последовательности за счет вставки, делеции или консервативной замены аминокислот, но сохраняют по меньшей мере одну биологическую функцию или активность. Вариант также может означать полипептид, который сохраняет по меньшей мере одну биологическую функцию или активность. Консервативную замену аминокислоты, т.е. замещение аминокислоты другой аминокислотой со сходными свойствами (например, степенью гидрофильности и распределением заряженных участков), понимают в данной области техники как обычно включающую незначительное изменение.
Термин «разновидность» относится к популяции растений, которые обладают постоянными характеристиками, отделяющими их от других растений того же вида. Хотя разновидность обладает одним или более отличительными признаками, она дополнительно характеризуется очень небольшим общим варьированием между особями в пределах этой разновидности. Разновидность часто продается на коммерческой основе.
Термин «вектор» относится к полинуклеотидному средству доставки, которое содержит комбинацию компонентов полинуклеотида для обеспечения транспорта полинуклеотидов, полинуклеотидных конструкций и полинуклеотидных конъюгатов и т.п. Вектор может представлять собой вирусный вектор, бактериофаг, искусственную хромосому бактерий или искусственную хромосому дрожжей. Вектор может представлять собой ДНК- или РНК-вектор. Подходящие векторы включают эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды из двунитевой нуклеотидной последовательности; линеаризованные плазмиды из двунитевой нуклеотидной последовательности и другие векторы любого происхождения. «Вектор экспрессии», как используется в данном документе, представляет собой полинуклеотидное средство доставки, которое содержит комбинацию компонентов полинуклеотида для обеспечения экспрессии полинуклеотида(полинуклеотидов), полинуклеотидных конструкций и полинуклеотидных конъюгатов и т.п. Подходящие векторы экспрессии включают эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды из двунитевой нуклеотидной последовательности; линеаризованные плазмиды из двунитевой нуклеотидной последовательности и другие функционально эквивалентные векторы экспрессии любого происхождения. Вектор экспрессии содержит по меньшей мере промотор, расположенный выше по последовательности и функционально связанный с полинуклеотидом, полинуклеотидными конструкциями или полинуклеотидным конъюгатом, как определено ниже.
Термин «цинковый палец» относится к структуре полипептида, которая распознает ДНК-последовательности и связывается с ними. Домен с «цинковыми пальцами» является наиболее распространенным ДНК-связывающим мотивом в протеоме человека. Один домен с «цинковыми пальцами» содержит примерно 30 аминокислот и обычно функционирует путем связывания с 3 последовательными парами оснований ДНК посредством взаимодействий боковой цепи одной аминокислоты с каждой парой оснований.
Термин «нуклеаза с «цинковыми пальцами»» или «ZFN» относится к химерной молекуле полипептида, содержащей по меньшей мере один ДНК-связывающий домен с «цинковыми пальцами», эффективно связанный с по меньшей мере одной нуклеазой или частью нуклеазы, способной в полностью собранном виде расщеплять ДНК.
Если в данном документе не определено иное, то научные и технические термины, используемые в связи с настоящим раскрытием, будут иметь значения, которые обычно понятны специалистам средней квалификации в данной области техники. Например, любые системы номенклатуры и методики, используемые в связи с культурами клеток или тканей, молекулярной биологией, иммунологией, микробиологией, генетикой и химией полипептида и полинуклеотида, а также гибридизацией, которые описаны в данном документе, хорошо известны и широко применяются в данной области техники. Значение и объем терминов должны быть ясны, однако в случае какой-либо скрытой двусмысленности определения, приведенные в данном документе, имеют преимущественную силу по сравнению с любым словарным или не относящимся к данному документу определением. Кроме того, если иное не требуется по контексту, термины в единственном числе будут включать множественное число, и термины во множественном числе будут включать единственное число.
2. Полинуклеотиды
В одном варианте осуществления представлен выделенный полинуклеотид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 60% идентичностью последовательности с любой из последовательностей, описанных в данном документе, включая любой из полинуклеотидов, показанных в перечне последовательностей. Предпочтительно, выделенный полинуклеотид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% идентичностью последовательности с ними.
Предпочтительно полинуклеотид(полинуклеотиды), описанный(описанные) в данном документе, кодирует(кодируют) активный полипептид, который характеризуется по меньшей мере приблизительно 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99%, 100% или большей функцией или активностью полипептида(полипептидов), показанного(показанных) в перечне последовательностей.
В другом варианте осуществления представлен выделенный полинуклеотид, содержащий, состоящий или по существу состоящий из полинуклеотида, характеризующегося по меньшей мере 60% идентичностью последовательности с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23, или c SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
Предпочтительно выделенный полинуклеотид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23, или с SEQ ID NO: 5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
Предпочтительно выделенный полинуклеотид содержит, состоит или по существу состоит из последовательности, характеризующейся меньшей мере, приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23, или с SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
Предпочтительно выделенный полинуклеотид содержит, состоит или по существу состоит из последовательности, характеризующейся меньшей мере, приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 13 или SEQ ID NO: 15.
Предпочтительно выделенный полинуклеотид содержит, состоит или по существу состоит из последовательности, характеризующейся меньшей мере, приблизительно 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23.
В другом варианте осуществления представлены полинуклеотиды, содержащие, состоящие или по существу состоящие из полинуклеотидов с существенной степенью гомологии (т. е. сходством последовательности) или существенной степенью идентичности с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23, или с SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
В другом варианте осуществления предусмотрены фрагменты SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23 с существенной степенью гомологии (то есть сходством последовательности) или с существенной степенью идентичности с ними, которые характеризуются по меньшей мере, приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с соответствующими фрагментами из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23.
В другом варианте осуществления представлены фрагменты из SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13 с существенной степенью гомологии (то есть сходством последовательности) или существенной степенью идентичности с ними, которые характеризуются по меньшей мере, приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с соответствующими фрагментами из SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
В другом варианте осуществления представлены полинуклеотиды, предусматривающие достаточную или существенную степень идентичности или сходства с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23, или с SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13, которые кодируют полипептид, который выполняет функцию SUS.
В другом варианте осуществления представлен полимер, представляющий собой полинуклеотид, который содержит, состоит или по существу состоит из полинуклеотида, обозначенного в данном документе как SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23, или как SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
Предпочтительно полинуклеотиды, описанные в данном документе, кодируют представителей семейства SUS.
Полинуклеотид, описанный в данном документе, может включать полимер из нуклеотидов, который может представлять собой немодифицированную или модифицированную дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). Соответственно, полинуклеотид может представлять собой без ограничения геномную ДНК, комплементарную ДНК (кДНК), мРНК, или антисмысловую РНК, или их фрагмент(фрагменты). Кроме того, полинуклеотид может представлять собой однонитевую или двунитевую ДНК, ДНК, которая является смесью однонитевых и двунитевых участков, гибридную молекулу, содержащую ДНК и РНК, или гибридную молекулу со смесью однонитевых и двунитевых участков или их фрагмента (фрагментов). Дополнительно полинуклеотид может быть составлен из трехнитевых участков, содержащих ДНК, РНК или обе, или их фрагмент (фрагменты). Полинуклеотид может содержать одно или более модифицированных оснований, таких как фосфоротиоаты, и может представлять собой пептидную нуклеиновую кислоту. Как правило, полинуклеотиды могут быть собраны из выделенных или клонированных фрагментов кДНК, геномной ДНК, олигонуклеотидов или отдельных нуклеотидов или комбинации вышеперечисленного. Хотя полинуклеотиды, описанные в данном документе, представлены в виде ДНК-последовательностей, они включают их соответствующие РНК-последовательности и их комплементарные (например, полностью комплементарные) ДНК- или РНК-последовательности, в том числе обратно комплементарные им последовательности.
Полинуклеотид, описанный в данном документе, обычно будет содержать фосфодиэфирные связи, хотя в некоторых случаях включены полинуклеотидные аналоги, которые могут иметь альтернативные остовы, содержащие, например, фосфороамидатные, фосфоротиоатные, фосфородитиоатные или О-метилфосфороамидитные связи; и пептидные полинуклеотидные остовы и связи. Другие аналоги полинуклеотидов включают полинуклеотиды с положительно заряженными остовами, неионогенными остовами и безрибозными остовами. Модификации рибозофосфатного остова можно осуществлять по целому ряду причин, например, для повышения стабильности и периода полужизни таких молекул в физиологических средах или в качестве зондов на биочипе. Можно получать смеси встречающихся в природе полинуклеотидов и аналогов; в качестве альтернативы, можно получать смеси разных аналогов полинуклеотидов и смеси природных полинуклеотидов и аналогов.
Известно множество аналогов полинуклеотидов, в том числе, например, таковые с фосфороамидатными, фосфоротиоатными, фосфородитиоатными или О-метилфосфороамидитными связями и пептидными остовами полинуклеотидов и связями. Другие аналоги полинуклеотидов включают таковые с положительно заряженными остовами, неионогенными остовами и безрибозными остовами. Полинуклеотиды, содержащие один или более карбоциклических сахаров, также включены.
Другие аналоги включают пептидные полинуклеотиды, которые представляют собой таковые на основе пептидных аналогов полинуклеотидов. Эти остовы являются практически неионогенными в нейтральных условиях в отличие от высокозаряженного фосфодиэфирного остова встречающихся в природе полинуклеотидов. Это может давать преимущества. Во-первых, пептидный остов полинуклеотида может характеризоваться улучшенной кинетикой гибридизации. Пептидные полинуклеотиды характеризуются более значительными изменениями температуры плавления в случае наличия несовпадающих пар оснований по сравнению с идеально совпадающими парами оснований. ДНК и РНК, как правило, характеризуются понижением температуры плавления на 2-4 C при наличии внутреннего несовпадения. В случае неионогенного пептидного остова полинуклеотида падение находится ближе к 7-9°C. Сходным образом, из-за их неионогенной природы, гибридизация оснований, присоединенных к этим остовам, является относительно нечувствительной к концентрации солей. Дополнительно пептидные полинуклеотиды могут не разрушаться или разрушаться в меньшей степени клеточными ферментами, и, таким образом, могут быть более стабильными.
В числе применений раскрытых полинуклеотидов и их фрагментов находится применение фрагментов в качестве зондов в анализах на основе гибридизации или в качестве праймеров для применения в анализах на основе амплификации. Такие фрагменты обычно содержат по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или больше смежных нуклеотидов из ДНК-последовательности. В других вариантах осуществления фрагмент ДНК содержит по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или больше смежных нуклеотидов из ДНК-последовательности. Таким образом, согласно одному аспекту также предусмотрен способ выявления полинуклеотида, включающий применение зондов или праймеров, или того и другого. Иллюстративные праймеры описаны в данном документе.
Основные параметры, влияющие на выбор условий гибридизации, и руководство для разработки подходящих условий описаны в Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). Используя данные о генетическом коде в комбинации с полипептидными последовательностями, описанными в данном документе, можно получить наборы вырожденных олигонуклеотидов. Такие олигонуклеотиды применимы в качестве праймеров, например, в полимеразных цепных реакциях (ПЦР), с помощью которых выделяют и амплифицируют фрагменты ДНК. В определенных вариантах осуществления вырожденные праймеры можно применять в качестве зондов для генетических библиотек. Такие библиотеки включают библиотеки кДНК, геномные библиотеки и даже электронные библиотеки меток экспрессируемых последовательностей или ДНК. Гомологичные последовательности, идентифицированные этим способом, будут затем использованы в качестве зондов для идентификации гомологов последовательностей, указанных в данном документе.
Также, потенциально применимыми являются полинуклеотиды и олигонуклеотиды (например, праймеры или зонды), которые гибридизируются в условиях пониженной жесткости, как правило, в условиях средней жесткости и, обычно, в условиях высокой жесткости с полинуклеотидом(полинуклеотидами), описанным(описанными) в данном документе. Основные параметры, влияющие на выбор условий гибридизации, и руководство для разработки подходящих условий изложены в Sambrook J., E. F. Fritsch and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.) и могут быть легко определены специалистами средней квалификации в данной области техники на основе, например, длины или композиции оснований полинуклеотида.
В данном документе определен один способ достижения условий умеренной и высокой жесткости. Следует понимать, что температуру промывки и концентрацию солей при промывке при потребности можно регулировать для достижения необходимой степени жесткости посредством применения основных принципов, которые управляют реакциями гибридизации и стабильностью дуплексов, как это известно специалистам в данной области техники и дополнительно описано ниже (см., например, Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y). При гибридизации полинуклеотида с полинуклеотидом с неизвестной последовательностью предполагается, что длина гибрида будет такой как у гибридизирующегося полинуклеотида. Когда гибридизируют полинуклеотиды с известными последовательностями, длина гибрида может быть определена путем выравнивания последовательностей полинуклеотидов и идентификации участка или участков с оптимальной комплементарностью последовательности. Температура гибридизации для гибридов, длина которых предположительно составят менее 50 пар оснований, должна быть на 5-10 C меньше, чем температура плавления гибрида, где температуру плавления определяют в соответствии со следующими уравнениями. Для гибридов, длина которых составляет менее 18 пар оснований, температура плавления (°С) = 2(число оснований А+Т)+4(число оснований G+C). Для гибридов, длина которых составляет более 18 пар оснований, температура плавления (°C) = 81,5+16,6(log10 [Na+])+0,41(% G+C)-(600/N), где N представляет собой число оснований в гибриде, и [Na+] представляет собой концентрацию ионов натрия в буфере для гибридизации ([Na+] для 1x стандартного цитрата натрия=0,165 M). Как правило, каждый такой гибридизирующийся полинуклеотид имеет длину, которая составляет по меньшей мере 25% (обычно по меньшей мере 50%, 60% или 70% и наиболее часто по меньшей мере 80%) от длины полинуклеотида, с которым он гибридизируется, и характеризуется по меньшей мере 60% идентичностью последовательности (например, по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%) с полинуклеотидом, с которым он гибридизируется.
Как будет понятно специалисту в данной области техники, линейная ДНК имеет две возможные ориентации: направление 5'-3' и направление 3'-5'. Например, если первая последовательность расположена в направлении 5'-3', и если вторая последовательность расположена в направлении 5'-3' в одной и той же полинуклеотидной молекуле/нити, то первая последовательность и вторая последовательность ориентированы в одном направлении или имеют одинаковую ориентацию. Как правило, последовательность промотора и представляющий интерес ген, находящийся под контролем данного промотора, расположены в одинаковой ориентации. Однако если по отношению к первой последовательности, расположенной в направлении 5'-3', вторая последовательность расположена в направлении 3'-5' в одной и той же полинуклеотидной молекуле/нити, тогда первая последовательность и вторая последовательность ориентированы в антисмысловом направлении или имеют антисмысловую ориентацию. Две последовательности, имеющие антисмысловые ориентации по отношению друг к другу, могут быть альтернативно описаны как имеющие одинаковую ориентацию, если первая последовательность (направление 5'-3') и последовательность, обратно комплементарная первой последовательности (первой последовательности, расположенной в 5'-3'), расположены в пределах одной полинуклеотидной молекулы/нити. Последовательности, изложенные в данном документе, показаны в направлении 5'-3'.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS1-S, NtSUS1-T, NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T. По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS2-T, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T и они не модулируются.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS2-T, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T и они не модулируются.
3. Полипептид
В другом аспекте представлен выделенный полипептид, содержащий, состоящий или по существу состоящий из полипептида, характеризующегося по меньшей мере 60% идентичности последовательности с любым из полипептидов, описанных в данном документе, включая любой из полипептидов, приведенных в перечне последовательностей. Предпочтительно выделенный полипептид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с ними.
В одном варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 2 или SEQ ID NO: 4.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 2.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 4.
В одном варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 80%, 81%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 6 или SEQ ID NO: 8.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 6.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 8.
В одном варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12.
В одном варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 14 или SEQ ID NO: 16.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 14.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 16.
В одном варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 18 или SEQ ID NO: 20.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 18.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 20.
В одном варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 22 или SEQ ID NO: 24.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 22.
В другом варианте осуществления представлен полипептид, содержащий, состоящий или по существу состоящий из последовательности, характеризующейся по меньшей мере 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 24.
В одном варианте осуществления представлен полипептид, кодируемый SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 или SEQ ID NO: 23; или SEQ ID NO:5, SEQ ID NO: 9, SEQ ID NO: 11 или SEQ ID NO: 13.
Полипептид может содержать последовательности, имеющие достаточную или значительную степень идентичности или сходства с SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22 или SEQ ID NO: 24; или SEQ ID NO: 6, SEQ ID NO: 10, SEQ ID NO: 12 или SEQ ID NO: 14 для выполнения функции SUS.
Как правило, фрагменты полипептида (полипептидов) сохраняют некоторые или все функции или активность полноразмерной последовательности.
Как обсуждалось в данном документе, полипептиды также включают мутантные варианты, полученных путем введения любого типа корректировок (например, вставки, делеции или замены аминокислот; изменения состояния гликозилирования; изменения, которые влияют на рефолдинг или изомеризацию, трехмерные структуры или состояния самоассоциации), которые могут быть намеренно сконструированы или выделены естественным образом при условии, что они по-прежнему сохраняют некоторую или всю свою функциональность или активность. Предпочтительно, данную функцию или активность модулируют.
Делеция относится к удалению одной или более аминокислот из полипептида. Вставка относится к одному или более аминокислотным остаткам, вводимым в заранее определенное место в полипептиде. Вставки могут предусматривать вставки внутрь последовательности одной или нескольких аминокислот. Замена относится к замещению аминокислот полипептида другими аминокислотами, имеющими сходные свойства (такие как сходная гидрофобность, гидрофильность, антигенность, склонность к образованию или разрыву α-спиральных структур или β-складчатых структур). Аминокислотные замены, как правило, предусматривают один остаток, но могут быть сгруппированы в зависимости от функциональных ограничений, накладываемых на полипептид, и могут предусматривать диапазон от приблизительно 1 до приблизительно 10 аминокислот. Аминокислотные замены являются предпочтительно консервативными аминокислотными заменами, как описано ниже. Аминокислотные замены, делеции и/или вставки можно выполнить с применением методик пептидного синтеза, таких как твердофазный пептидный синтез, или с помощью манипулирования с рекомбинантной ДНК. Способы манипулирования ДНК-последовательностями с получением вариантов полипептида с заменой, вставкой или делецией хорошо известны в данной области техники. Вариант может характеризоваться наличием корректировок, которые вызывают возникновение «молчащего» изменения и в результате приводят к образованию функционально эквивалентного полипептида. Преднамеренные аминокислотные замены можно осуществлять исходя из сходства остатков в отношении полярности, заряда, растворимости, гидрофобности, гидрофильности и амфипатической природы, при условии, что сохраняется связывание, обуславливающее вторичную структуру вещества. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту, положительно заряженные аминокислоты включают лизин и аргинин, и аминокислоты с незаряженными полярными концевыми группами, характеризующиеся сходными значениями гидрофильности, включают лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин. Консервативные замены можно осуществлять, например, в соответствии с приведенной ниже таблицей. Аминокислоты в одном и том же блоке во втором столбце и предпочтительно в одной и той же строке в третьем столбце можно заменять одна на другую:
Полипептид может представлять собой зрелый полипептид, или незрелый полипептид, или полипептид, полученный из незрелого полипептида. Полипептиды могут находиться в линейной форме или являться циклизированными с применением известных способов. Полипептиды, как правило, содержат по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, или по меньшей мере 40 смежных аминокислот.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS1-S, NtSUS1-T, NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS2-T, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T и они не модулируются.
По меньшей мере одна модификация (например, мутация) может быть включена в одну или более из NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S, при этом модификация(модификации) (например, мутация(мутации)) не включена(включены) в одну или более из NtSUS1-S, NtSUS1-T, NtSUS2-T, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T и они не модулируются.
Модификация растений
Трансформация
Рекомбинантные конструкции можно применять для трансформации растений или клеток растения для того, чтобы модулировать экспрессию, функцию или активность полипептида. Рекомбинантная полинуклеотидная конструкция может содержать полинуклеотид, кодирующий один или более полинуклеотидов, описанных в данном документе, функционально связанных с регуляторным участком, подходящим для экспрессии полипептида. Таким образом, полинуклеотид может содержать кодирующую последовательность, которая кодирует полипептид, описанный в данном документе. Растения или клетки растений, в которых модулируют экспрессию, функцию или активность полипептида, могут включать мутантные, не встречающиеся в природе, трансгенные, созданные человеком или полученные с помощью методик генной инженерии растения или клетки растения. Предпочтительно, трансгенное растение или клетка растения содержит геном, который был скорректирован путем стабильной интеграции рекомбинантной ДНК. Рекомбинантная ДНК содержит ДНК, полученную с помощью методик генной инженерии и сконструированную вне клетки, и содержит ДНК, содержащую встречающуюся в природе ДНК, или кДНК, или синтетическую ДНК. Трансгенное растение может включать растение, регенерированное из первоначально трансформированной клетки растения, и трансгенные растения, являющиеся потомством более поздних поколений или гибридов трансформированного растения. Предпочтительно, трансгенная модификация обеспечивает корректировку экспрессии, или функции, или активности полинуклеотида или полипептида, описанных в данном документе, по сравнению с таковыми в контрольном растении.
Полипептид, кодируемый рекомбинантным полинуклеотидом, может являться нативным полипептидом или может являться гетерологичным по отношению к клетке. В некоторых случаях рекомбинантная конструкция содержит полинуклеотид, который обеспечивает модулирование экспрессии, функционально связанный с регуляторным участком. Примеры подходящих регуляторных участков описаны в данном документе.
Также представлены векторы, содержащие рекомбинантные полинуклеотидные конструкции, такие как описанные в данном документе. Подходящие основы для векторов включают, например, те, которые обычно используют в данной области техники, такие как плазмиды, вирусы, искусственные хромосомы, искусственные хромосомы бактерий, искусственные хромосомы дрожжей или искусственные хромосомы бактериофагов. Подходящие векторы экспрессии включают без ограничения плазмиды и вирусные векторы, полученные из, например, бактериофага, бакуловирусов и ретровирусов. Многочисленные векторы и системы экспрессии являются коммерчески доступными.
Векторы могут содержать, например, точки начала репликации, участки связывания с ядерным матриксом или маркеры. Маркерный ген может придавать клетке растения селектируемый фенотип. Например, маркер может придавать устойчивость к биоцидным средствам, такую как устойчивость к антибиотику (например, к канамицину, G418, блеомицину или гигромицину) или к гербициду (например, к глифосату, хлорсульфурону или фосфинотрицину). Дополнительно вектор экспрессии может содержать последовательность метки, предназначенной для облегчения манипуляций или выявления (например, очистки или локализации) экспрессируемого полипептида. Последовательности меток, такие как последовательности люциферазы, бета-глюкуронидазы, зеленого флуоресцентного полипептида, глутатион-S-трансферазы, полигистидина, c-myc или гемагглютинина, как правило, экспрессируются в виде фрагмента, слитого с кодируемым полипептидом. Такие метки могут быть вставлены в любом месте в пределах полипептида, в том числе на карбоксильном или амино-конце.
Растение или клетку растения можно трансформировать путем интегрирования рекомбинантного полинуклеотида в их геном с обеспечением их стабильной трансформации. Растение или клетка растения, описанные в данном документе, могут являться стабильно трансформированными. Стабильно трансформированные клетки, как правило, сохраняют введенный полинуклеотид с каждым клеточным делением. Растение или клетка растения также могут являться транзиентно трансформированными, так что рекомбинантный полинуклеотид не интегрируется в их геном. Транзиентно трансформированные клетки, как правило, теряют весь введенный рекомбинантный полинуклеотид или некоторую его часть с каждым клеточным делением, так что введенный рекомбинантный полинуклеотид нельзя выявить в дочерних клетках после достаточного числа клеточных делений.
Из уровня техники доступен ряд способов для трансформации клетки растения, включающий биолистику; методики с применением генной пушки; трансформацию, опосредованную Agrobacterium; трансформацию, опосредованную вирусным вектором; способ на основе замораживания-оттаивания; бомбардировку микрочастицами; прямое поглощение ДНК; ультразвуковую обработку; микроинъекцию; перенос, опосредованный вирусом растения; и электропорацию. Система на основе Agrobacterium для интеграции чужеродной ДНК в хромосомы растений в широких масштабах изучалась, модифицировалась и использовалась для генной инженерии растений. Молекулы депротеинизированной рекомбинантной ДНК, содержащие ДНК-последовательности, соответствующие исследуемому очищенному полипептиду, функционально связанные в смысловой или антисмысловой ориентации с регуляторными последовательностями, соединяют с соответствующими последовательностями Т-ДНК с помощью общепринятых способов. Их вводят в протопласты с помощью методики с применением полиэтиленгликоля или методики с применением электропорации, обе из которых являются стандартными. В качестве альтернативы, такие векторы, содержащие молекулы рекомбинантной ДНК, кодирующие исследуемый очищенный полипептид, вводят в живые клетки Agrobacterium, которые затем переносят ДНК в клетки растения. Трансформацию с помощью депротеинизированной ДНК без сопутствующих последовательностей вектора на основе Т-ДНК можно выполнять посредством слияния протопластов с ДНК-содержащими липосомами или посредством электропорации. Депротеинизированную ДНК без сопутствующих последовательностей вектора на основе Т-ДНК также можно применять для трансформации клеток с помощью инертных, высокоскоростных микрочастиц.
Если клетку или культивируемую ткань применяют в качестве реципиентной ткани для трансформации, при необходимости растения можно регенерировать из трансформированных культур с помощью методов, известных специалистам в данной области техники.
Выбор регуляторных участков, подлежащих включению в рекомбинантную конструкцию, зависит от нескольких факторов, в том числе без ограничения от эффективности, селектируемости, индуцируемости, необходимого уровня экспрессии и предпочтительной экспрессии в определенных клетках или тканях. Обычной задачей для специалиста в данной области техники является модулирование экспрессии кодирующей последовательности посредством правильного выбора регуляторных участков и их размещения по отношению к кодирующей последовательности. Транскрипцию полинуклеотида можно модулировать сходным образом. Некоторые подходящие регуляторные участки инициируют транскрипцию исключительно или преимущественно в определенных типах клеток. Способы идентификации и установления характеристик регуляторных участков в геномной ДНК растений хорошо известны в данной области техники.
Подходящие промоторы включают тканеспецифичные промоторы, распознаваемые тканеспецифичными факторами, присутствующими в разных тканях или типах клеток (например, специфичные для корня промоторы, специфичные для побега промоторы, специфичные для ксилемы промоторы), или присутствующими на различных стадиях развития, или присутствующими в ответ на разные условия окружающей среды. Подходящие промоторы включают конститутивные промоторы, которые могут быть активированы в большинстве типов клеток, не требуя специфических индукторов. Примеры подходящих промоторов для управления выработкой RNAi-полинуклеотида включают промотор 35S вируса мозаики цветной капусты (CaMV/35S), SSU, OCS, lib4, usp, STLS1, B33, nos или промоторы гена убиквитина или гена фазеолина. Специалисты в данной области техники могут создавать множество вариантов рекомбинантных промоторов.
Тканеспецифичные промоторы представляют собой элементы управления транскрипцией, которые активны только в определенных клетках или тканях в определенные моменты времени в ходе развития растений, например в вегетативных тканях или репродуктивных тканях. Тканеспецифичная экспрессия может быть преимущественной, например, если предпочтительной является экспрессия полинуклеотидов в определенных тканях. Примеры тканеспецифичных промоторов, находящихся под контролем в ходе развития, включают промоторы, которые могут инициировать транскрипцию только (или в основном только) в определенных тканях, таких как вегетативные ткани, например корни или листья, или репродуктивные ткани, такие как плоды, семяпочки, семена, пыльца, тычинки, цветки или любая эмбриональная ткань. Промоторы, специфичные для репродуктивных тканей, могут являться, например, специфичными для пыльника, специфичными для семяпочки, специфичными для зародыша, специфичными для эндосперма, специфичными для интегумента, специфичными для семени и кожуры семени, специфичными для пыльцы, специфичными для лепестка, специфичными для чашелистика или для комбинаций таковых.
Подходящие специфичные для листа промоторы включают промотор гена пируватортофосфатдикиназы (PPDK) из C4-растения (кукурузы), промотор cab-m1Ca+2 из кукурузы, промотор родственных myb генов из Arabidopsis thaliana (Atmyb5), промоторы рибулозобифосфаткарбоксилазы (RBCS) (например, генов томата RBCS 1, RBCS2 и RBCS3A, экспрессируемых в листьях и выращенных на свету саженцах, RBCS1 и RBCS2, экспрессируемых в развивающихся плодах томата, или промотор гена рибулозобифосфаткарбоксилазы, экспрессируемый на высоких уровнях почти исключительно в мезофильных клетках листовых пластинок и листовых пазух).
Подходящие специфичные для стареющих тканей промоторы включают промотор из томата, активный во время созревания плодов, старения и опадения листьев, промотор гена, кодирующего цистеиновую протеазу маиса, промотор 82E4 и промотор генов SAG. Можно применять подходящие специфичные для пыльника промоторы. Можно выбрать подходящие предпочтительные для корней промоторы, известные специалистам в данной области техники. Подходящие промоторы для экспрессии предпочтительно в семенах включают как специфичные для семян промоторы (промоторы, активные в процессе развития семян, такие как промоторы запасных полипептидов семян), так и промоторы прорастающих семян (промоторы, активные во время прорастания семян). Такие предпочтительные для семян промоторы включают Cim1 (цитокинин-индуцированный сигнал); cZ19B1 (19 кДa зеин кукурузы); milps (миоинозитол-1-фосфатсинтаза); mZE40-2, также известный как Zm-40; nuclc; и celA (целлюлозосинтаза). Промотор гена гамма-зеина представляет собой специфичный для эндосперма промотор. Промотор гена Glob-1 представляет собой специфичный для зародыша промотор. Для двудольных растений специфичные для семян промоторы включают промотор гена бета-фазеолина фасоли, гена напина, генаβ-конглицинина, гена лектина сои, гена круциферина и т.п. Для однодольных растений специфичные для семян промоторы включают промотор гена зеина кукурузы с молекулярной массой 15 кДа, промотор гена зеина с молекулярной массой 22 кДа, промотор гена зеина с молекулярной массой 27 кДа, промотор гена g-зеина, промотор гена гамма-зеина с молекулярной массой 27 кДа (такой как промотор gzw64A, см. номер доступа S78780 в Genbank), промотор гена waxy, промотор гена shrunken 1, промотор гена shrunken 2, промотор гена глобулина 1 (см. номер доступа L22344 в Genbank), промотор гена Itp2, промотор гена cim1, промоторы генов end1 и end2 кукурузы, промотор гена nuc1, промотор гена Zm40, промотор гена eep1 и eep2; промотор гена lec1, промотор гена тиоредоксина H; промотор гена mlip15, промотор гена PCNA2 и промотор гена shrunken-2.
Примеры индуцируемых промоторов включают промоторы, реагирующие на воздействие патогенов, анаэробные условия, повышенную температуру, свет, засуху, низкую температуру или высокую концентрацию солей. Патоген-индуцируемые промоторы включают промоторы связанных с патогенезом полипептидов (PR-полипептидов), индуцирование которых происходит после инфицирования патогеном (например, PR-полипептиды, SAR-полипептиды, бета-1,3-глюканаза, хитиназа).
Дополнительно к промоторам растений другие подходящие промоторы могут иметь бактериальное происхождение, например промотор гена октопинсинтазы, промотор гена нопалинсинтазы и другие промоторы, полученные из Ti-плазмид, или они могут быть получены из вирусных промоторов (например, промоторов 35S и 19S РНК вируса мозаики цветной капусты (CaMV), конститутивных промоторов вируса табачной мозаики, промоторов 19S и 35S вируса мозаики цветной капусты (CaMV) или промотора 35S вируса мозаики норичника).
Подходящие способы введения полинуклеотидов в клетки растений и последующего встраивания в геном растения включают микроинъекцию (Biotechniques (1986) 4:320-334), электропорацию (Proc. Natl. Acad. Sci. USA (1986) 83:5602-5606), трансформацию, опосредованную Agrobacterium (US5981840 и US5563055), прямой перенос генов (EMBO J. (1984) 3:2717-2722) и баллистическое ускорение частиц (US4945050, US5879918, US5886244, US5932782; Plant Cell, Tissue, and Organ Culture: Fundamental Methods, ed. Gamborg and Phillips (Springer-Verlag, Berlin) (1995); и Biotechnology (1988) 6:923-926).
Мутация
Раскрыты растение или клетка растения, содержащие мутацию в одном или более полинуклеотидах или полипептидах, описанных в данном документе, где указанная мутация приводит к модуляции функции или активности NtSUS или полипептида (полипептидов), кодируемого(кодируемых) ими.
Представлен способ модулирования уровня полипептида NtSUS в (подвергнутом сушке) растении или в (подвергнутом сушке) растительном материале, при этом указанный способ включает введение в геном указанного растения одной или более мутаций, которые модулируют экспрессию по меньшей мере одного гена NtSUS, где указанный по меньшей мере один ген выбран из последовательностей в соответствии с настоящим изобретением.
Также представлен способ идентификации растения с модулированными уровнями редуцирующих сахаров, при этом указанный способ включает скрининг образца полинуклеотида из представляющего интерес растения в отношении наличия одной или более мутаций в последовательностях в соответствии с настоящим изобретением, и необязательно сопоставление идентифицированной мутации (мутаций) с мутацией (мутациями), которая(которые), как известно, модулирует(модулируют) уровни редуцирующих сахаров.
Также раскрыты растение или клетка растения, которые являются гетерозиготными или гомозиготными по одной или более мутациям в гене NtSUS в соответствии с настоящим изобретением, при этом указанная мутация приводит к модулированной экспрессии гена, или функции, или активности полипептида NtSUS, кодируемого им.
Для комбинирования мутаций в одном растении можно применять целый ряд подходов, в том числе половое скрещивание. Растение, характеризующееся наличием одной или более благоприятных мутаций в гетерозиготном или гомозиготном состоянии в гене в соответствии с настоящим изобретением, которые модулируют экспрессию гена или функцию или активность полипептида, кодируемого таковым, можно скрещивать с растением, характеризующимся наличием одной или более благоприятных мутаций в гетерозиготном или гомозиготном состоянии в одном или более генах, которые модулируют их экспрессию или функцию или активность полипептида, кодируемого таковыми. В одном варианте осуществления скрещивания проводят для введения одной или более благоприятных мутаций в гетерозиготном или гомозиготном состоянии в ген в соответствии с настоящим изобретением в одном и том же растении.
Функция или активность одного или более полипептидов по настоящему изобретению в растении являются повышенными или пониженными, если функция или активность являются более низкими или более высокими, чем функция или активность того же полипептида(полипептидов) в растении, которое не было модифицировано для подавления функции или активности этого полипептида, и которое культивировали, собирали и подвергали сушке с применением тех же протоколов.
В некоторых вариантах осуществления мутацию(мутации) вводят в растение или клетку растения с применением подхода с обеспечением мутагенеза, и введенную мутацию идентифицируют или подвергают отбору с применением способов, известных специалистам в данной области техники, таких как анализ методом Саузерн-блоттинга, секвенирование ДНК, ПЦР-анализ или фенотипический анализ. Мутации, которые влияют на экспрессию гена или которые нарушают функцию кодируемого полипептида, можно определять с применением способов, хорошо известных из уровня техники. Инсерционные мутации в экзонах гена, как правило, в результате приводят к образованию нефункциональных мутантных вариантов. Мутации по консервативным остаткам могут быть особенно эффективными для подавления метаболической функции кодируемого полипептида. Например, понятно, что мутация в одном или более высококонсервативных участках будет, очевидно, обеспечивать корректировку функции полипептида, в то время как мутация вне этих высококонсервативных участков будет, очевидно, оказывать незначительное влияния на функцию полипептида или не будет влиять на него. Дополнительно мутация в одном нуклеотиде может приводить к возникновению стоп-кодона, наличие которого в результате приведет к получению усеченного полипептида и, в зависимости от степени усечения, потере функции.
Также раскрыты способы получения мутантных полинуклеотидов и полипептидов. Любое представляющее интерес растение, в том числе клетку растения или растительный материал, можно генетически модифицировать различными способами, с помощью которых, как известно, индуцируют мутагенез, в том числе сайт-направленный мутагенез, олигонуклеотид-направленный мутагенез, индуцируемый химическими соединениями мутагенез, индуцируемый ионизирующим излучением мутагенез, мутагенез с применением модифицированных оснований, мутагенез с применением ДНК-дуплекса с гэпом, мутагенез с двунитевыми разрывами, мутагенез с применением линий-хозяев с нарушенной репарацией, мутагенез посредством синтеза полного гена, перетасовка ДНК и другие эквивалентные способы.
Также раскрыты фрагменты полинуклеотидов и полипептидов. Фрагменты полинуклеотида могут кодировать фрагменты полипептида, которые сохраняют биологическую функцию нативного полипептида и, следовательно, задействованы в сети транспортировки метаболитов в растении. В качестве альтернативы, фрагменты полинуклеотида, которые являются применимыми в качестве зондов для гибридизации или ПЦР-праймеров, как правило, не кодируют фрагментов полипептида, сохраняющих биологическую функцию. Кроме того, фрагменты раскрытых полинуклеотидов включают такие фрагменты, которые могут быть собраны в рекомбинантные конструкции, как обсуждалось в данном документе. Фрагменты полинуклеотида могут находиться в диапазоне от по меньшей мере приблизительно 25 нуклеотидов, приблизительно 50 нуклеотидов, приблизительно 75 нуклеотидов, приблизительно 100 нуклеотидов, приблизительно 150 нуклеотидов, приблизительно 200 нуклеотидов, приблизительно 250 нуклеотидов, приблизительно 300 нуклеотидов, приблизительно 400 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 600 нуклеотидов, приблизительно 700 нуклеотидов, приблизительно 800 нуклеотидов, приблизительно 900 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 1100 нуклеотидов, приблизительно 1200 нуклеотидов, приблизительно 1300 нуклеотидов или приблизительно 1400 нуклеотидов и до полноразмерного полинуклеотида, кодирующего полипептиды, описанные в данном документе. Фрагменты полипептида могут находиться в диапазоне от по меньшей мере приблизительно 25 аминокислот, приблизительно 50 аминокислот, приблизительно 75 аминокислот, приблизительно 100 аминокислот, приблизительно 150 аминокислот, приблизительно 200 аминокислот, приблизительно 250 аминокислот, приблизительно 300 аминокислот, приблизительно 400 аминокислот, приблизительно 500 аминокислот и до полноразмерного полипептида, описанного в данном документе. Мутантные варианты полипептида можно применять для создания мутантных, не встречающихся в природе или трансгенных растений (например, мутантных, не встречающихся в природе, трансгенных, созданных человеком или полученных с помощью методик генной инженерии растений) или клеток растения, содержащих один или более мутантных вариантов полипептида. Предпочтительно, мутантные варианты полипептида сохраняют функцию полипептида, не подвергнутого мутированию. Функция мутантного варианта полипептида может быть выше, ниже или приблизительно такой же, как у полипептида, не подвергнутого мутированию.
Мутации в полинуклеотидах и полипептидах, описанных в данном документе, могут включать внесенные человеком мутации, или искусственные мутации, или мутации, созданные с помощью генной инженерии. Мутации в полинуклеотидах и полипептидах, описанных в данном документе, могут представлять собой мутации, которые получены или которые можно получить посредством способа, который включает стадию манипуляции in vitro или in vivo. Мутации в полинуклеотидах и полипептидах, описанных в данном документе, могут представлять собой мутации, которые получены или которые можно получить посредством способа, который предусматривает вмешательство человека.
Способы, с помощью которых вводят мутацию случайным образом в полинуклеотид, могут включать химический мутагенез и радиационный мутагенез. Химический мутагенез предусматривает применение экзогенно добавляемых химических веществ, таких как мутагенные, тератогенные или канцерогенные органические соединения, для индуцирования мутаций. Для создания мутаций можно применять мутагены, которые создают главным образом точечные мутации и короткие делеции, вставки, миссенс-мутации, простые повторяющиеся последовательности, трансверсии и/или транзиции, в том числе химические мутагены и излучение. Мутагены включают этилметансульфонат, метилметансульфонат, N-этил-N-нитрозомочевину, триэтилмеламин, N-метил-N-нитрозомочевину, прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, акриламидный мономер, мельфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитро-нитрозогуанидин, нитрозогуанидин, 2-аминопурин, 7,12-диметил-бенз(a)антрацен, этиленоксид, гексаметилфосфорамид, бисульфан, диэпоксиалканы (диэпоксиоктан, диэпоксибутан и т.п.), 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина дигидрохлорид и формальдегид.
Также предусмотрены спонтанные мутации в локусе, которые могут не быть непосредственно вызванными мутагеном, при условии, что они приводят к возникновению необходимого фенотипа. Подходящие мутагенные средства могут также включать, например, ионизирующее излучение, такое как рентгеновское излучение, гамма-излучение, излучение быстрых нейтронов и ультрафиолетовое излучение. Дозу мутагенного химического вещества или излучения определяют экспериментально для каждого типа ткани растения таким образом, чтобы получить частоту мутаций, которая ниже порогового уровня, характеризующегося летальностью или репродуктивной стерильностью. Любой способ получения полинуклеотида растения, известный специалистам в данной области техники, можно применять для получения полинуклеотида растения для скрининга в отношении мутаций.
Способ мутации может включать одну или более стадий скрещивания растений.
После внесения мутации можно провести скрининг для идентификации мутаций, которые обеспечивают возникновение генов с преждевременными стоп-кодонами или иным образом нефункциональных генов. После внесения мутации можно провести скрининг для идентификации мутаций, которые обеспечивают возникновение функциональных генов, которые способны экспрессироваться на повышенных или пониженных уровнях. Скрининг в отношении мутантных вариантов можно выполнять с помощью секвенирования, или путем применения одного или более зондов или праймеров, специфичных для данного гена или полипептида. В полинуклеотидах можно также обеспечивать возникновение конкретных мутаций, которые могут в результате приводить к модулированию экспрессии гена, модулированию стабильности мРНК или модулированию стабильности полипептида. В данном документе такие растения называются «не встречающимися в природе» или «мутантными» растениями. Как правило, мутантные или не встречающиеся в природе растения содержат по меньшей мере часть чужеродного, или синтетического, или созданного человеком нуклеотида (например, ДНК или РНК), которая не присутствовала в растении до проведения с ним манипуляций. Чужеродный нуклеотид может представлять собой один нуклеотид, два или более нуклеотидов, два или более смежных нуклеотидов или два или более несмежных нуклеотидов, как, например, по меньшей мере 10, 20, 30, 40, 50,100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400 или 1500 или больше смежных или несмежных нуклеотидов.
Трансгеника и редактирование гена
Помимо осуществления мутагенеза композиции, которые могут модулировать экспрессию, или функцию, или активность одного или более из полинуклеотидов NtSUS или полипептидов, кодируемых ими, включают специфичные в отношении последовательности полинуклеотиды, которые могут нарушать транскрипцию одного или более эндогенного(эндогенных) гена(генов); специфичные в отношении последовательности полинуклеотиды, которые могут нарушать трансляцию РНК-транскриптов (например, двухнитевые РНК, siRNA, рибозимы); специфичные в отношении последовательности полинуклеотиды, которые могут нарушать стабильность одного или более полипептидов; специфичные в отношении последовательности полинуклеотиды, которые могут нарушать ферментативную функцию одного или более полипептидов или функцию связывания одного или более полипептидов с субстратами или регуляторными полипептидами; антитела, которые проявляют специфичность в отношении одного или более полипептидов; низкомолекулярные соединения, которые могут нарушать стабильность одного или более полипептидов, или ферментативную функцию одного или более полипептидов, или функцию связывания одного или более полипептидов; полипептиды с «цинковыми пальцами», которые связывают один или более полинуклеотидов; и мегануклеазы, которые оказывают воздействие на один или более полинуклеотидов. Технологии редактирования гена, генетические технологии редактирования и технологии редактирование генома хорошо известны в данной области техники.
Нуклеазы с «цинковыми пальцами»
Полипептиды с «цинковыми пальцами» можно применять для модулирования экспрессии, или функции, или активности одного или более из полинуклеотидов NtSUS, описанных в данном документе. В различных вариантах осуществления последовательность геномной ДНК, содержащую часть или всю кодирующую последовательность полинуклеотида, модифицируют путем мутагенеза, опосредованного нуклеазой с «цинковыми пальцами». В последовательности геномной ДНК осуществляют поиск уникального сайта для связывания полипептида с «цинковыми пальцами». В качестве альтернативы, в последовательности геномной ДНК осуществляют поиск двух уникальных сайтов для связывания полипептида с «цинковыми пальцами», при этом оба сайта находятся на противоположных нитях и близко друг к другу, например на расстоянии 1, 2, 3, 4, 5, 6 или более пар оснований друг от друга. Соответственно, предусмотрены полипептиды с «цинковыми пальцами», которые связываются с полинуклеотидами.
Полипептид с «цинковыми пальцами» можно сконструировать для распознавания выбранного сайта-мишени в гене. Полипептид с «цинковыми пальцами» может содержать любую комбинацию мотивов, полученных из природных ДНК-связывающих доменов с «цинковыми пальцами» и неприродных ДНК-связывающих доменов с «цинковыми пальцами» посредством усечения, или удлинения, или способа сайт-направленного мутагенеза в сочетании со способом отбора, таким как без ограничения отбор с помощью фагового дисплея, отбор с помощью бактериальной двугибридной системы или отбор с помощью бактериальной одногибридной системы. Термин «неприродный ДНК-связывающий домен с «цинковыми пальцами»» относится к ДНК-связывающему домену с «цинковыми пальцами», который связывает последовательность из трех пар оснований в полинуклеотиде-мишени и который не встречается в клетке или организме, содержащих полинуклеотид, который подлежит модификации. Способы разработки полипептида с «цинковыми пальцами», который связывает специфические полинуклеотиды, которые являются уникальными для целевого гена, известны из уровня техники.
В других вариантах осуществления может быть выбран полипептид с «цинковыми пальцами» для связывания с регуляторной последовательностью полинуклеотида. Более конкретно, регуляторная последовательность может содержать сайт инициации транскрипции, стартовый кодон, участок экзона, границу раздела экзон-интрон, терминатор или стоп-кодон. Соответственно, настоящее изобретение предусматривает мутантные, не встречающиеся в природе или трансгенные растение или клетки растения, полученные с помощью мутагенеза, опосредованного нуклеазой с «цинковыми пальцами», осуществленного вблизи или в пределах одного или более полинуклеотидов, описанных в данном документе, и способы получения таких растения или клетки растения с помощью мутагенеза, опосредованного нуклеазой с «цинковыми пальцами». Способы доставки полипептида с «цинковыми пальцами» и нуклеазы с «цинковыми пальцами» в растение подобны способам, описанным ниже для доставки мегануклеазы.
Мегануклеазы
В другом аспекте описаны способы получения мутантных, не встречающихся в природе, или трансгенных, или иным образом генетически модифицированных растений с применением мегануклеаз, таких как I-CreI. Встречающиеся в природе мегануклеазы, а также рекомбинантные мегануклеазы можно использовать для того, чтобы специфическим образом выполнять двухнитевой разрыв в одном сайте или в относительно небольшом числе сайтов в геномной ДНК растения для обеспечения разрушения одного или более полинуклеотидов NtSUS, описанных в данном документе. Мегануклеаза может представлять собой сконструированную мегануклеазу со скорректированными свойствами распознавания ДНК. Полипептиды мегануклеаз можно доставлять в клетки растения с помощью ряда разных механизмов, известных из уровня техники.
Настоящее изобретение охватывает применение мегануклеаз для инактивации полинуклеотида(полинуклеотидов) NtSUS, описанного(описанных) в данном документе (или любой их комбинации, описанной в данном документе), в клетке растения или растении. В частности, в настоящем изобретении представлен способ инактивации полинуклеотида NtSUS в растении с применением мегануклеазы, включающий: (a) обеспечение клетки растения, содержащей полинуклеотид NtSUS, описанный в данном документе; (b) введение мегануклеазы или конструкции, кодирующей мегануклеазу, в указанную клетку растения и (c) обеспечение для мегануклеазы возможности в существенной степени инактивировать полинуклеотид(полинуклеотиды) NtSUS.
Мегануклеазы можно применять для расщепления по сайтам распознавания мегануклеазами в пределах кодирующих участков полинуклеотида. Такое расщепление часто приводит в результате к делеции ДНК в сайте распознавания мегануклеазой с последующей репарацией мутагенной ДНК путем соединения негомологичных концов. Такие мутации в кодирующей последовательности гена являются, как правило, достаточными для инактивации гена. Этот способ модификации клетки растения включает, во-первых, доставку кассеты экспрессии мегануклеазы в клетку растения с применением подходящего способа трансформации. Для достижения максимальной эффективности необходимо связать кассету экспрессии мегануклеазы с селектируемым маркером и отобрать успешно трансформированные клетки в присутствии селективного средства. Этот подход в результате приведет к интеграции кассеты экспрессии мегануклеазы в геном, что, однако, может быть нежелательным, если для растения возможно будет требоваться официальное разрешение. В таких случаях кассету экспрессии мегануклеазы (и связанный селектируемый маркерный ген) можно сегрегировать в последующих поколениях растения с применением общепринятых методик селекции.
После доставки кассеты экспрессии мегануклеазы клетки растения выращивают, изначально, в условиях, которые являются типичными для конкретной процедуры трансформации, которую применяли. Это может означать выращивание трансформированных клеток на среде при температуре ниже 26°C, зачастую в темноте. Такие стандартные условия можно применять в течение некоторого периода времени, предпочтительно 1-4 дней, для обеспечения восстановления клетки растения после процесса трансформации. В любой момент после этого начального периода восстановления температуру роста можно повысить, чтобы стимулировать функцию сконструированной мегануклеазы с осуществлением расщепления и мутирования по сайту распознавания мегануклеазой.
TALEN
Один способ редактирования гена включает применение эффекторных нуклеаз, подобных активатору транскрипции (TALEN), которые индуцируют образование двунитевых разрывов, на которые клетка может отвечать посредством механизмов репарации. NHEJ возобновляет связь ДНК с обеих сторон двунитевого разрыва, где участок перекрывания последовательностей является очень маленьким или вовсе отсутствует, для обеспечения отжига. Этот механизм репарации индуцирует ошибки в геноме посредством вставки, или делеции, или хромосомной перестройки. Любые такие ошибки могут сделать продукты гена, закодированные в этом месте, не функциональными. Для определенных видов применения может потребоваться точное удаление полинуклеотида NtSUS из генома растения. Такие применения возможны с применением пары сконструированных мегануклеаз, каждая из которых расщепляет сайт распознавания мегануклеазой по обе стороны от предполагаемой делеции. Также можно применять TALEN, которые способны распознавать ген и связываться с ним и вводить двухнитевой разрыв в геном. Таким образом, в другом аспекте представлены способы получения мутантных, не встречающихся в природе или трансгенных, или иным образом генетически модифицированных растений, описанных в данном документе, с применением эффекторных нуклеаз TAL.
CRISPR/Cas
Другой способ редактирования гена включает применение бактериальной системы CRISPR/Cas. Бактерии и архебактерии характеризуются наличием хромосомных элементов, называемых короткими палиндромными повторами, регулярно расположенными группами (CRISPR), которые являются частью адаптивной иммунной системы, защищающей от проникновения вирусной и плазмидной ДНК. В случае систем CRISPR типа II, CRISPR-РНК (crRNA) функционируют с трансактивирующей crRNA (tracrRNA) и CRISPR-ассоциированными (Cas) полипептидами с обеспечением внесения двунитевых разрывов в ДНК-мишень. Расщепление мишени с помощью Cas9 требует спаривания оснований crRNA и tracrRNA, а также спаривания оснований crRNA и ДНК-мишени. Распознавание мишени облегчается при наличии короткого мотива, называемого прилегающим к протоспейсеру мотивом (PAM), который соответствует последовательности NGG. Данную систему можно применять для редактирования генома. Cas9 обычно программируется двойной РНК, состоящей из crRNA и tracrRNA. Тем не менее, коровые компоненты этих РНК могут быть объединены в единую гибридную «направляющую РНК» для нацеливания Cas9. Использование некодирующей направляющей РНК для нацеливания на ДНК с целью сайт-специфического расщепления обещает быть значительно более простым, чем существующие технологии, такие как TALEN. При применении стратегии CRISPR/Cas перенацеливание нуклеазного комплекса требует только введения новой последовательности РНК, и нет необходимости в реконструировании специфичности факторов транскрипции на основе полипептида. Технология CRISPR/Cas была реализована у растений в соответствии со способом из международной заявки WO 2015/189693 A1, в которой раскрыта опосредованная вирусом платформа редактирования генома, которая является широко применимой для разных видов растений. Геном RNA2 вируса погремковости табака (TRV) был сконструирован для переноса и доставки направляющей РНК в растения Nicotiana benthamiana со сверхэкспрессией эндонуклеазы Cas9. В контексте настоящего изобретения направляющую РНК можно получать из любой из полинуклеотидных последовательностей NtSUS, раскрытых в данном документе, а также идеи из WO 2015/189693 A1 применяют для редактирования генома клетки растения и получения требуемого мутантного растения. Высокий темп развития технологии породил огромное разнообразие протоколов с широкой применяемостью на представителях царства растения, при этом эти протоколы были хорошо каталогизированы в целом ряде недавних научных обзорных статей (например, Plant Methods (2016) 12:8; и Front Plant Sci. (2016) 7: 506). Обзор систем CRISPR/Cas с особым упором на их применение у растений находится в Biotechnology Advances (2015) 33, 1, p41-52. Bortesi и Fischer также проводят сравнение между технологией CRISPR/Cas, нуклеазами с "цинковыми пальцами" и TALEN. Последние разработки для применения CRISPR/Cas в манипулировании геномами растений описаны в Acta Pharmaceutica Sinica B (2017) 7, 3, p292-302) и Curr. Op. in Plant Biol. (2017) 36, 1-8. Плазмиды на основе CRISPR/Cas9 для применения в растениях перечислены в «addgene», некоммерческом хранилище плазмид (addgene.org), и плазмиды на основе CRISPR/Cas являются коммерчески доступными.
Антисмысловая модификация
Технология применения антисмысловых олигонуклеотидов представляет собой другой хорошо известный способ, который можно использовать для модулирования экспрессии полипептида NtSUS. Полинуклеотид гена NtSUS, который подлежит репрессии, клонируют и функционально связывают с регуляторным участком и последовательностью терминации транскрипции так, чтобы антисмысловая нить РНК транскрибировалась. Затем рекомбинантной конструкцией трансформируют клетку растения и получают антисмысловую нить РНК. Полинуклеотид не обязательно является полной последовательностью гена, подлежащего репрессии, но, как правило, является практически комплементарным по меньшей мере части смысловой нити гена, подлежащего репрессии.
Полинуклеотид может быть транскрибирован в рибозим или каталитическую РНК, которая влияет на экспрессию мРНК. Рибозимы можно разрабатывать таким образом, чтобы они специфически спаривались практически с любой целевой РНК и расщепляли фосфодиэфирный остов в определенном месте, тем самым функционально инактивируя целевую РНК. Гетерологичные полинуклеотиды могут кодировать рибозимы, разработанные таким образом, чтобы они расщепляли конкретные транскрипты мРНК, предотвращая таким образом экспрессию полипептида. Рибозимы типа hammerhead являются применимыми для разрушения конкретных мРНК, хотя можно применять различные рибозимы, которые расщепляют мРНК в последовательностях сайт-специфического распознавания. Рибозимы типа hammerhead расщепляют мРНК в местах, определяемых фланкирующими участками, которые образуют комплементарные пары оснований с мРНК-мишени. Единственным требованием является то, что РНК-мишень должна содержать полинуклеотид 5'-UG-3'. Конструирование и получение рибозимов типа hammerhead известно в данной области техники. Последовательности рибозимов типа hammerhead можно встраивать в стабильную РНК, такую как транспортная РНК (tRNA) для повышения эффективности расщепления in vivo.
В одном варианте осуществления специфичный в отношении последовательности полинуклеотид, который может нарушать трансляцию транскрипта(транскриптов) РНК, представляет собой интерферирующую РНК. РНК-интерференция или РНК-сайленсинг представляют собой эволюционно консервативный процесс, с помощью которого конкретные мРНК могут быть нацелены для ферментативного разрушения. Двунитевую РНК (двунитевую РНК) вводят в клетку или получают в клетке (например, вирус с двунитевой РНК или полинуклеотиды, представляющие собой интерферирующую РНК) для инициации пути интерферирующей РНК. Двунитевую РНК можно преобразовывать в несколько дуплексов малых интерферирующих РНК (siRNA) длиной 21-24 п.о. с помощью РНКаз III, которые представляют собой эндонуклеазы, специфичные к двунитевой РНК. Впоследствии, siRNA могут распознаваться индуцируемыми РНК комплексами сайленсинга, которые способствуют раскручиванию siRNA с помощью АТФ-зависимого процесса. Раскрученная антисмысловая нить siRNA направляет активированные РНК-индуцированные комплексы сайленсинга к мРНК, в отношении которой осуществлено нацеливание, которая содержит последовательность, комплементарную антисмысловой нити siRNA. мРНК, в отношении которой осуществлено нацеливание, и антисмысловая нить могут образовывать А-форму спирали, и большая бороздка А-формы спирали может распознаться активированными РНК-индуцированными комплексами сайленсинга. мРНК-мишень может расщепляться с помощью активированных РНК-индуцированных комплексов сайленсинга в одном сайте, определенном сайтом связывания 5'-конца нити siRNA. Активированные РНК-индуцированные комплексы сайленсинга могут повторно использоваться для катализа еще одного события расщепления.
Векторы экспрессии на основе интерферирующей РНК могут содержать конструкции интерферирующей РНК, кодирующие полинуклеотиды интерферирующей РНК, которые осуществляют РНК-интерференцию путем понижения уровня экспрессии мРНК, пре-мРНК или родственных вариантов РНК. Векторы экспрессии могут содержать промотор, расположенный выше по последовательности и функционально связанный с конструкцией интерферирующей РНК, как дополнительно описано в данном документе. Векторы экспрессии на основе интерферирующей РНК могут содержать подходящий минимальный коровый промотор, представляющую интерес конструкцию интерферирующей РНК, расположенный выше по последовательности (5') регуляторный участок, расположенный ниже по последовательности (3') регуляторный участок, в том числе сигналы терминации транскрипции и полиаденилирования, и другие последовательности, известные специалистам в данной области техники, такие как различные селективные маркеры.
Молекулы двунитевой РНК могут включать молекулы siRNA, собранные из отдельного олигонуклеотида в структуре стебель-петля, где самокомплементарные смысловую и антисмысловую участки молекулы siRNA соединены с помощью основанного на полинуклеотидах или не основанного на полинуклеотидах линкера (линкеров), а также кольцевую однонитевую РНК с двумя или более петлевыми структурами и стеблем, содержащую самокомплементарные смысловую и антисмысловую нити, где кольцевую РНК можно процессировать либо in vivo, либо in vitro с образованием активной молекулы siRNA, способной опосредовать интерферирующую РНК.
Также предусмотрено использование молекул малой шпилечной РНК. Они содержат специфичную антисмысловую последовательность в дополнение к обратно комплементарной (смысловой) последовательности, как правило, отделенной спейсером или последовательностью петли. Расщепление спейсера или петли обеспечивает образование молекулы однонитевой РНК и ее обратно комплементарной нити, так что их можно отжечь с образованием двунитевой молекулы РНК (необязательно с дополнительными стадиями обработки, которые могут в результате привести в результате к добавлению или удалению одного, двух, трех или более нуклеотидов с 3'-конца или 5'-конца одной или обеих нитей). Спейсер может иметь достаточную длину для обеспечения отжига антисмысловой и смысловой последовательностей и образования двунитевой структуры (или стебля) до расщепления спейсера (и необязательно последующих стадий обработки, которые могут в результате привести в результате к добавлению или удалению одного, двух, трех, четырех или более нуклеотидов с 3'-конца или с 5'-конца одной или обеих нитей). Спейсерная последовательность как правило представляет собой неродственный полинуклеотид, который находится между двумя комплементарными участками полинуклеотидов, которые после отжига с получением двунитевого полинуклеотида образуют малую шпилечную РНК. Спейсерная последовательность обычно содержит от приблизительно 3 до приблизительно 100 нуклеотидов.
Любой представляющий интерес РНК-полинуклеотид можно получить путем подбора подходящей композиции последовательности, размера петли и длины стебля для получения шпилечного дуплекса. При осуществлении разработки подходящий диапазон длины стебля шпилечного дуплекса включает длины стебля, составляющие по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов, как, например, приблизительно 14-30 нуклеотидов, приблизительно 30-50 нуклеотидов, приблизительно 50-100 нуклеотидов, приблизительно 100-150 нуклеотидов, приблизительно 150-200 нуклеотидов, приблизительно 200-300 нуклеотидов, приблизительно 300-400 нуклеотидов, приблизительно 400-500 нуклеотидов, приблизительно 500-600 нуклеотидов и приблизительно 600-700 нуклеотидов. При осуществлении разработки подходящий диапазон длин петли шпилечного дуплекса включает длину петли, составляющую приблизительно 4-25 нуклеотидов, приблизительно 25-50 нуклеотидов или больше, если длина стебля шпилечного дуплекса является значительной. В определенных вариантах осуществления длина молекулы двунитевой РНК или ssRNA составляет от приблизительно 15 до приблизительно 40 нуклеотидов. В другом варианте осуществления молекула siRNA представляет собой молекулу двунитевой РНК или ssRNA длиной от приблизительно 15 до приблизительно 35 нуклеотидов. В другом варианте осуществления молекула siRNA представляет собой молекулу двунитевой РНК или ssRNA длиной от приблизительно 17 до приблизительно 30 нуклеотидов. В другом варианте осуществления молекула siRNA представляет собой молекулу двунитевой РНК или ssRNA длиной от приблизительно 19 до приблизительно 25 нуклеотидов. В другом варианте осуществления молекула siRNA представляет собой молекулу двунитевой РНК или ssRNA длиной от приблизительно 21 до приблизительно 23 нуклеотидов. В определенных вариантах осуществления шпилечные структуры с участками в виде дуплекса длиннее 21 нуклеотида могут способствовать эффективному сайленсингу, управляемому siRNA, вне зависимости от последовательности и длины петли. Иллюстративные последовательности для осуществления РНК-интерференции описаны в данном документе.
Длина последовательности мРНК-мишени составляет, как правило, от приблизительно 14 до приблизительно 50 нуклеотидов. Следовательно, мРНК-мишень можно проверить на наличие участков длиной от приблизительно 14 до приблизительно 50 нуклеотидов, которые предпочтительно соответствуют одному или более из следующих критериев: соотношение A+T/G+С составляет от приблизительно 2:1 до приблизительно 1:2; наличие динуклеотида AA или динуклеотида СA на 5'-конце; наличие последовательности из по меньшей мере 10 последовательных нуклеотидов, являющихся уникальными для мРНК-мишени (то есть, последовательность не присутствует в других последовательностях мРНК из этого же растения); и отсутствие «рядов» из более чем трех последовательных нуклеотидов, представляющих собой гуанин (G), или более чем трех последовательных нуклеотидов, представляющих собой цитозин (C). Оценку в отношении данных критериев можно осуществлять с помощью различных методик, известных в данной области техники, например, компьютерные программы, такие как BLAST, можно применять для поиска по общедоступным базам данных, чтобы определить, является ли выбранная последовательность уникальной для мРНК-мишени. В качестве альтернативы, последовательность можно отобрать (и разработать последовательность siRNA) с помощью коммерчески доступного компьютерного программного обеспечения (например, OligoEngine, Target Finder и Design Tool для siRNA, которые являются коммерчески доступными).
В одном варианте осуществления отбирают последовательности мРНК-мишени, длина которых составляет от приблизительно 14 до приблизительно 30 нуклеотидов, которые соответствуют одному или более критериям, указанным выше. В другом варианте осуществления отбирают последовательности, длина которых составляет от приблизительно 16 до приблизительно 30 нуклеотидов, которые соответствуют одному или более критериям, указанным выше. В дополнительном варианте осуществления отбирают последовательности, длина которых составляет от приблизительно 19 до приблизительно 30 нуклеотидов, которые соответствуют одному или более критериям, указанным выше. В другом варианте осуществления отбирают последовательности, длина которых составляет от приблизительно 19 до приблизительно 25 нуклеотидов, которые соответствуют одному или более критериям, указанным выше.
В иллюстративном варианте осуществления молекулы siRNA содержат специфическую антисмысловую последовательность, которая комплементарна по меньшей мере 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более смежным нуклеотидам из любого из полинуклеотидов, описанных в данном документе.
Специфическая антисмысловая последовательность, содержащаяся в молекуле siRNA, может быть идентичной или практически идентичной комплементарной последовательности. В одном варианте осуществления специфическая антисмысловая последовательность, содержащаяся в молекуле siRNA, является на по меньшей мере приблизительно 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, комплементарной последовательности мРНК-мишени. Способы определения идентичности последовательности известны в данной области техники, и ее можно определить, например, с помощью программы BLASTN из программного обеспечения Computer Group (GCG) Университета штата Висконсин, или предоставленной на веб-сайте NCBI.
Один способ индукции сайленсинга двунитевой РНК у растений представляет собой трансформацию с помощью генной конструкции, продуцирующей шпилечную РНК (см. Nature (2000) 407, 319-320). Такие конструкции содержат инвертированные участки последовательности целевого гена, отделенные соответствующим спейсером. Вставка функционального интронного участка растения в качестве спейсерного фрагмента дополнительно повышает эффективность индукции сайленсинга гена благодаря выработке шпилечной РНК на основе сплайсинга интрона (Plant J. (2001), 27, 581-590). Предпочтительно длина стебля составляет от приблизительно 50 нуклеотидов до приблизительно 1 тысячи нуклеотидов в длину. Способы получения шпилечной РНК на основе сплайсинга интрона хорошо описаны в данной области техники (см., например, Bioscience, Biotechnology, and Biochemistry (2008) 72, 2, 615-617).
Молекулы интерферирующей РНК, имеющие структуру в виде дуплекса или двунитевую структуру, например двунитевая РНК или малая шпилечная РНК, могут иметь тупые концы, или могут иметь 3'- или 5'-выступы. Используемый в данном документе термин «выступ» относится к неспаренному нуклеотиду или нуклеотидам, которые выступают из структуры в виде дуплекса, если 3'-конец одной нити РНК выходит за пределы 5'-конца другой нити (3'-выступ), или наоборот (5'-выступ). Нуклеотиды, составляющие выступы, могут быть рибонуклеотидами, дезоксирибонуклеотидами или их модифицированными версиями. В одном варианте осуществления по меньшей мере одна нить молекулы интерферирующей РНК имеет 3'-выступ длиной от приблизительно 1 до приблизительно 6 нуклеотидов. В других вариантах осуществления длина 3'-выступа составляет от приблизительно 1 до приблизительно 5 нуклеотидов, от приблизительно 1 до приблизительно 3 нуклеотидов и от приблизительно 2 до приблизительно 4 нуклеотидов.
Если молекула интерферирующей РНК содержит 3'-выступ на одном конце молекулы, другой конец может быть с тупым концом, или также иметь выступ (5' или 3'). Если молекула интерферирующей РНК содержит выступ с обоих концов молекулы, длина выступов может быть одинаковой или разной. В одном варианте осуществления молекула интерферирующей РНК содержит 3'-выступы, состоящие из от приблизительно 1 до приблизительно 3 нуклеотидов, на обоих концах молекулы. В дополнительном варианте осуществления молекула интерферирующей РНК представляет собой двунитевую РНК, имеющую 3'-выступ из 2 нуклеотидов с обоих концов молекулы. В еще одном варианте осуществления нуклеотиды, составляющие выступ интерферирующей РНК, являются динуклеотидами TT или динуклеотидами UU.
Молекулы интерферирующей РНК могут содержать одну или более 5'- или 3'-кэп-структур. Термин «кэп-структура» относится к химической модификации, включенной с любого конца олигонуклеотида, которая защищает молекулу от разрушения эндонуклеазами и может также облегчать доставку или локализацию внутри клетки.
Другой модификацией, применяемой к молекулам интерферирующей РНК, является химическое связывание с молекулой интерферирующей РНК одного или более фрагментов или конъюгатов, которые усиливают функцию, клеточное распределение, клеточный захват, биодоступность или стабильность молекулы интерферирующей РНК. Полинуклеотиды можно синтезировать или модифицировать с помощью способов, общепринятых в данной области техники. Химические модификации включают 2'-модификации, введение неприродных оснований, ковалентное присоединение лиганда и замещение фосфатных связей тиофосфатными связями. В данном варианте осуществления прочность структуры в виде дуплекса укрепляют с помощью по меньшей мере одной и, как правило, двух химических связей.
Нуклеотиды одной или обеих из двух одинарных нитей можно модифицировать для модулирования активации клеточных ферментов, таких как, например, без ограничения определенные нуклеазы. Методики для понижения уровня или ингибирования активации клеточных ферментов известны в данной области техники и включают без ограничения 2'-аминомодификации, 2'-фтормодификации, 2'-алкилмодификации, модификации незаряженного каркаса, морфолиновые модификации, 2'-О-метилмодификации и фосфорамидат.
Молекулу интерферирующей РНК можно конъюгировать с лигандами, например, для усиления ее абсорбции клеткой. В определенных вариантах осуществления гидрофобный лиганд конъюгируют с молекулой для облегчения прямого проникновения через клеточную мембрану. В определенных случаях конъюгация катионного лиганда с олигонуклеотидами часто в результате приводит к улучшению устойчивости к нуклеазам.
«Нацеленные индуцированные локальные повреждения в геноме» (TILLING) представляет собой еще одну технологию мутагенеза, которую можно применять для создания и/или идентификации полинуклеотидов, кодирующих полипептиды с модифицированной экспрессией, функцией и/или активностью. TILLING также обеспечивает возможность отбора растений, несущих такие мутантные варианты. В TILLING комбинируют мутагенез высокой плотности со способами высокопроизводительного скрининга. Способы осуществления TILLING хорошо известны из уровня техники (см. McCallum et al., (2000) Nat Biotechnol 18: 455-457 и Stemple (2004) Nat Rev Genet 5(2): 145-50).
Различные варианты осуществления направлены на векторы экспрессии, содержащие один или более из полинуклеотидов NtSUS или конструкций интерферирующей РНК, которые содержат один или более полинуклеотидов NtSUS, описанных в данном документе.
Различные варианты осуществления направлены на векторы экспрессии, содержащие один или более из полинуклеотидов NtSUS или одну или более конструкций интерферирующей РНК, описанных в данном документе.
Различные варианты осуществления направлены на векторы экспрессии, содержащие один или более полинуклеотидов NtSUS или одну или более конструкций интерферирующей РНК, кодирующих один или более полинуклеотидов интерферирующей РНК, описанных в данном документе, которые способны к самогибридизации с образованием структуры в виде шпильки, при этом конструкция содержит (a) один или более из полинуклеотидов NtSUS, описанных в данном документе; (b) вторую последовательность, кодирующую спейсерный элемент, который образует петлю структуры в виде шпильки; и (c) третью последовательность, содержащую последовательность, обратно комплементарную первой последовательности, расположенную в той же ориентации, что и первая последовательность, где вторая последовательность расположена между первой последовательностью и третьей последовательностью, и вторая последовательность функционально связана с первой последовательностью и третьей последовательностью.
Раскрытые полинуклеотиды NtSUS можно использовать для конструирования различных полинуклеотидов, которые не образуют структуры в виде шпильки. Например, двунитевая РНК может быть образована посредством (1) транскрибирования первой цепи ДНК путем функционального связывания с первым промотором и (2) транскрибирования последовательности, обратно комплементарной фрагменту ДНК первой цепи путем функционального связывания со вторым промотором. Каждую цепь полинуклеотида можно транскрибировать из одного вектора экспрессии, или из разных векторов экспрессии. РНК-дуплекс, обладающий свойствами РНК-интерференции, можно ферментативно преобразовать в siRNA для модулирования уровней РНК.
Таким образом, различные варианты осуществления направлены на векторы экспрессии, содержащие один или более полинуклеотидов NtSUS или конструкций интерферирующей РНК, описанных в данном документе, кодирующих полинуклеотиды интерферирующей РНК, способные к самогибридизации, при этом конструкция содержит (a) один или более из полинуклеотидов NtSUS, описанных в данном документе; и (b) вторую последовательность, содержащую последовательность, комплементарную (например, обратно комплементарную) первой последовательности, расположенную в той же ориентации, что и первая последовательность.
Представлены различные композиции и способы для модулирования уровней эндогенной экспрессии одного или более из полипептидов NtSUS, описанных в данном документе (или любой их комбинации, описанной в данном документе), путем стимулирования косупрессии экспрессии гена.
Различные композиции и способы предусмотрены для модулирования уровня экспрессии эндогенного гена путем модулирования трансляции мРНК. Клетку растения-хозяина (табака) можно трансформировать с помощью вектора экспрессии, содержащего промотор, функционально связанный с полинуклеотидом NtSUS, расположенным в антисмысловой ориентации по отношению к промотору для обеспечения возможности экспрессии полинуклеотидов РНК с последовательностью, комплементарной части мРНК.
Различные векторы экспрессии для модулирования трансляции мРНК могут содержать промотор, функционально связанный с полинуклеотидом NtSUS, в котором последовательность расположена в антисмысловой ориентации по отношению к промотору. Длина полинуклеотидов антисмысловой РНК может варьировать и может составлять приблизительно 15-20 нуклеотидов, приблизительно 20-30 нуклеотидов, приблизительно 30-50 нуклеотидов, приблизительно 50-75 нуклеотидов, приблизительно 75-100 нуклеотидов, приблизительно 100-150 нуклеотидов, приблизительно 150-200 нуклеотидов и приблизительно 200-300 нуклеотидов.
Мобильные генетические элементы
В качестве альтернативы, нацеливание для инактивации в отношении генов можно осуществлять путем введения транспозонов (например, IS-элементов) в геномы представляющих интерес растений. Данные мобильные генетические элементы можно ввести с помощью полового перекрестного опыления и мутантов со вставками можно подвергнуть скринингу в отношении потери функции полипептида. Разрушенный ген родительского растения можно ввести в другие растения путем скрещивания родительского растения с растением, не подвергнутым индуцированному транспозоном мутагенезу, например, путем полового перекрестного опыления. Можно применять любые стандартные методики селекции, известные специалистам в данной области техники. В одном варианте осуществления один или более генов можно инактивировать путем вставки одного или более транспозонов. Мутации могут привести к гомозиготному разрушению одного или более генов, к гетерозиготному разрушению одного или более генов, или к комбинации гомозиготных и гетерозиготных разрушений, если разрушен более чем один ген. Подходящие мобильные элементы включают ретротранспозоны, ретропозоны и SINE-подобные элементы. Такие способы известны специалистам в данной области техники.
Рибозимы
В качестве альтернативы на полинуклеотиды NtSUS можно целенаправленно воздействовать для инактивации путем введения рибозимов, полученных из ряда малых кольцевых РНК, которые способны к саморасщеплению и репликации в растениях. Данные РНК могут реплицироваться либо самостоятельно (РНК вироида), либо с участием вируса-помощника (сателлитные РНК). Примеры подходящих РНК включают полученные из вироида солнечной пятнистости авокадо и сателлитные РНК, полученные из вируса кольцевой пятнистости табака, вируса временной полосатости люцерны, вируса бархатной пятнистости табака, вируса пятнистости Solanum nodiflorum и вируса пятнистости клевера подземного. Различные специфичные к целевой РНК рибозимы известны специалистам в данной области техники.
4. Растения
Мутантные или не встречающиеся в природе растения или клетки растений могут иметь любую комбинацию одной или более мутаций в одном или более генах NtSUS, которая приводит к модулированной экспрессии, или функции, или активности этих генов или продуктов таких генов. Например, мутантные или не встречающиеся в природе растения или клетки растений могут иметь одну мутацию в одном гене NtSUS; несколько мутаций в одном гене NtSUS; одну мутацию в двух или более, или трех или более, или четырех или более генах NtSUS; или несколько мутаций в двух или более, или трех или более, или четырех или более генах NtSUS. В качестве дополнительного примера мутантные или не встречающиеся в природе растения или клетки растений могут иметь одну или более мутаций в конкретной части гена(генов) NtSUS, например, в участке гена NtSUS, который кодирует активный сайт полипептида NtSUS или его часть. В качестве дополнительного примера мутантные или не встречающиеся в природе растения или клетки растений могут иметь одну или более мутаций в участке за пределами одного или более генов NtSUS, в таком как участок, расположенный выше или ниже гена NtSUS, который он регулирует, при условии, что они модулируют функцию или экспрессию гена(генов) NtSUS. Элементы, расположенные выше по последовательности, могут включать промоторы, энхансеры или факторы транскрипции. Некоторые элементы, такие как энхансеры, могут располагаться выше по последовательности или ниже по последовательности от гена, который они регулируют. Элемент (элементы) не обязательно расположен рядом с геном, который он регулирует, так как было обнаружено, что некоторые элементы расположены на расстоянии в несколько тысяч пар оснований выше по последовательности или ниже по последовательности от гена, который они регулируют. Мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну или более мутаций, расположенных в первых 100 нуклеотидах от гена (генов), в первых 200 нуклеотидах от гена (генов), в первых 300 нуклеотидах от гена (генов), в первых 400 нуклеотидах от гена (генов), в первых 500 нуклеотидах от гена (генов), в первых 600 нуклеотидах от гена (генов), в первых 700 нуклеотидах от гена (генов), в первых 800 нуклеотидах от гена (генов), в первых 900 нуклеотидах от гена (генов), в первой 1000 нуклеотидов от гена (генов), в первых 1100 нуклеотидах от гена (генов), в первых 1200 нуклеотидах от гена (генов), в первых 1300 нуклеотидах от гена (генов), в первых 1400 нуклеотидах от гена (генов) или в первых 1500 нуклеотидах от гена (генов). Мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну или более мутаций, расположенных в первом, втором, третьем, четвертом, пятом, шестом, седьмом, восьмом, девятом, десятом, одиннадцатом, двенадцатом, тринадцатом, четырнадцатом или пятнадцатом наборе из 100 нуклеотидов гена (генов) или их комбинации. Раскрыты мутантные или не встречающиеся в природе растения или клетки растения (например, мутантные, не встречающиеся в природе или трансгенные растения или клетки растения и т.п., как описано в данном документе), содержащие варианты мутантного полипептида.
В одном варианте осуществления семена растений подвергают мутагенезу и затем выращивают из них мутантные растения первого поколения. Затем растениям первого поколения дают самоопылиться и из семян от растений первого поколения выращивают растения второго поколения, которые затем проверяют на наличие мутаций в их локусах. Не смотря на то, что подвергнутый мутации растительный материал можно подвергнуть скринингу на наличие мутаций, преимуществом скрининга растений второго поколения является то, что все соматические мутации соответствуют мутациям в зародышевых линиях. Специалисту в данной области техники будет понятно, что различный растительный материал, в том числе без ограничения семена, пыльцу, ткань растения или клетки растения, можно подвергнуть мутагенезу для создания мутантных растений. Однако тип растительного материала, подвергнутого мутагенезу, может иметь значение, когда полинуклеотид растения подвергают скринингу на наличие мутаций. Например, если пыльцу подвергают мутагенезу до опыления не подвергнутого мутагенезу растения, из семян, полученных при этом опылении, выращивают растения первого поколения. Каждая клетка растений первого поколения будет содержать мутации, возникшие в пыльце; таким образом, эти растения первого поколения можно затем подвергнуть скринингу на наличие мутаций вместо того, чтобы ждать появления второго поколения.
Получение модифицированных растений, скрининг и скрещивание
Полинуклеотиды NtSUS, полученные из отдельных растений, клеток растений или растительного материала, необязательно можно объединять, чтобы ускорить скрининг в отношении мутаций в популяции растений, происходящих из подвергнутых мутагенезу растительной ткани, клеток растений или растительного материала. Можно проверить одно или более следующих поколений растений, клеток растения или растительного материала. Размер необязательно объединенной группы зависит от чувствительности применяемого способа скрининга.
После необязательного объединения образцов, их можно подвергнуть анализу с помощью методик полинуклеотид-специфичной амплификации, таких как ПЦР. Любой один или более праймеров или зондов, специфичных в отношении гена или последовательностей, непосредственно примыкающих к гену, можно применять для амплификации последовательностей в пределах необязательно объединенного образца. Предпочтительно для амплификации участков локуса, в которых с наибольшей вероятностью возникают полезные мутации, разрабатывают один или более праймеров или зондов. Наиболее предпочтительно праймер разрабатывают для выявления мутаций в участках полинуклеотида. Дополнительно, предпочтительным для праймера (праймеров) и зонда (зондов) было бы обеспечить избегание известных полиморфных сайтов для облегчения скрининга точечных мутаций. Для облегчения выявления продуктов амплификации один или более праймеров или зондов можно метить с применением любого общепринятого способа введения метки. Праймер (праймеры) или зонд (зонды) можно разрабатывать на основе последовательностей, описанных в данном документе, с помощью способов, которые хорошо известны в данной области техники.
Для облегчения выявления продуктов амплификации праймер (праймеры) или зонд(зонды) можно метить с применением любого общепринятого способа внесения метки. Их можно разрабатывать на основе последовательностей, описанных в данном документе, с помощью способов, которые хорошо известны в данной области техники.
Полиморфизмы можно идентифицировать с помощью средств, известных в данной области техники, и некоторых из описанных в литературе.
В некоторых вариантах осуществления растение можно регенерировать или вырастить из растения, ткани растения или клетки растения. Можно применять любые подходящие способы регенерации или выращивания растения из клетки растения или ткани растения, такие как без ограничения культивирование тканей или регенерация из протопластов. Предпочтительно растения можно регенерировать путем выращивания трансформированных клеток растения на среде для индукции образования каллюса, среде для индукции образования побегов и/или среде для индукции образования корней. См., например, McCormick et al., Plant Cell Reports 5:81-84 (1986). Затем данные растения можно выращивать и либо опылять их с помощью той же трансформированной линии, либо других линий и идентифицировать получаемый в результате гибрид, характеризующийся экспрессией требуемой фенотипической характеристики. Можно выращивать два или более поколений, чтобы убедиться, что экспрессия требуемой фенотипической характеристики стабильно поддерживается и наследуется, и затем собирать семена, чтобы убедиться, что экспрессия требуемой фенотипической характеристики была достигнута. Таким образом, выражение «трансформированные семена», используемое в данном документе, относится к семенам, которые содержат нуклеотидную конструкцию, стабильно интегрированную в геном растения.
Соответственно, в дополнительном аспекте предусмотрен способ получения мутантного растения. Способ включает получение по меньшей мере одной клетки растения, содержащей ген NtSUS, кодирующий функциональный полинуклеотид NtSUS, описанный в данном документе (или любую их комбинацию, описанную в данном документе). Далее эту по меньшей мере одну клетку растения обрабатывают в условиях, эффективных для модулирования функции полинуклеотида (полинуклеотидов) NtSUS, описанного (описанных) в данном документе. По меньшей мере одну мутантную клетку растения затем размножают с получением мутантного растения, где мутантное растение характеризуется модулированным уровнем описанного (описанных) полипептида (полипептидов) (или любой их комбинации, описанной в данном документе) по сравнению с уровнем у контрольного растения. В одном варианте осуществления данного способа получения мутантного растения стадия обработки включает воздействие на по меньшей мере одну клетку химическим мутагенным средством, описанным выше, и в условиях, эффективных для получения по меньшей мере одной мутантной клетки растения. В другом варианте осуществления данного способа стадия обработки включает подвергание по меньшей мере одной клетки воздействию источника ионизирующего излучения в условиях, эффективных для получения по меньшей мере одной мутантной клетки растения. Термин «мутантное растение» включает мутантные растения, у которых генотип является модифицированным по сравнению с генотипом у контрольного растения, предпочтительно с помощью способов, отличных от способов генной инженерии и генетической модификации.
В определенных вариантах осуществления мутантное растение, клетка мутантного растения или мутантный растительный материал может содержать одну или более мутаций, которые встречаются в природе в другом растении, клетке растения или растительном материале и обеспечивают требуемый признак. Эту мутацию можно встроить (например, путем интрогрессии) в другое растение, клетку растения или растительный материал (например, растение, клетку растения или растительный материал с генетическим фоном, отличающимся от такового у растения, из которого происходит мутация) для обеспечения у них данного признака. Таким образом, в качестве примера, мутацию, которая встречается в природе в первом растении, можно ввести во второе растение, такое как второе растение с генетическим фоном, отличающимся от такового у первого растения. Таким образом, специалист в данной области техники может осуществлять поиск и идентифицировать растение, несущее в естественных условиях в своем геноме один или более мутантных аллелей генов, описанных в данном документе, которые обеспечивают желаемый признак. Мутантный аллель (аллели), который встречается в природе, можно перенести во второе растение различными способами, включая селекцию, обратное скрещивание и интрогрессию с получением линий, разновидностей или гибридов, которые имеют одну или более мутаций в генах, описанных в данном документе. Та же методика также может быть применена для интрогрессии одной или более не встречающихся в природе мутации (мутаций) из первого растения во второе растение. Растения, демонстрирующие желаемый признак, можно отобрать из пула мутантных растений. Предпочтительно, отбор осуществляют с применением данных о полинуклеотиде, описанном в данном документе. Следовательно, можно осуществлять отбор по генетическому признаку по сравнению с контролем. Такой скрининговый подход может включать применение общепринятых методик амплификации и/или гибридизации, как обсуждалось в данном документе. Таким образом, дополнительный аспект настоящего изобретения относится к способу идентификации мутантного растения, включающему стадии: (a) получения образца, содержащего полинуклеотид NtSUS из растения; и (b) определения последовательности полинуклеотида NtSUS, где отличие в последовательности полинуклеотида NtSUS по сравнению с полинуклеотидом NtSUS из контрольного растения свидетельствует о том, что указанное растение является мутантным растением. В другом аспекте предложен способ идентификации мутантного растения, которое накапливает редуцирующий (редуцирующие) сахар(сахара) на повышенных или сниженных уровнях по сравнению с контрольным растением, включающий стадии: (a) получения образца из растения, подлежащего скринингу; (b) определения того, содержит ли указанный образец одну или более мутаций в одном или более из полинуклеотидов NtSUS, описанных в данном документе; и (c) определения уровня по меньшей мере одного редуцирующего сахара в указанном растении. Предпочтительно уровень по меньшей мере одного редуцирующего сахара определяют в зеленых листьях, листьях, подвергнутых ранним стадиям сушки, или подвергнутых сушке листьях. В другом аспекте представлен способ получения мутантного растения, которое характеризуется повышенными или сниженными уровнями по меньшей мере одного редуцирующего сахара по сравнению с контрольным растением, включающий стадии: (a) получения образца из первого растения; (b) определения того, содержит ли указанный образец одну или более мутаций в одном или более полинуклеотидах NtSUS, описанных в данном документе, которые приводят к модулированным уровням по меньшей мере одного редуцирующего сахара; и (c) переноса одной или более мутаций во второе растение. Предпочтительно уровень по меньшей мере одного редуцирующего сахара определяют в зеленых листьях, листьях, подвергнутых ранним стадиям сушки, или подвергнутых сушке листьях. Мутацию (мутации) можно перенести во второе растение с помощью различных способов, которые известны в данной области техники, например с помощью генной инженерии, манипуляции с генами, интрогрессии, селекции растений, обратного скрещивания и т.п. В одном варианте осуществления первое растение является встречающимся в природе растением. В одном варианте осуществления второе растение характеризуется генетическим фоном, отличающимся от такового у первого растения. В другом аспекте представлен способ получения мутантного растения, которое характеризуется повышенными или сниженными уровнями по меньшей мере одного редуцирующего сахара по сравнению с контрольным растением, включающий стадии: (a) получения образца из первого растения; (b) определения того, содержит ли указанный образец одну или более мутаций в одном или более из полинуклеотидов NtSUS, описанных в данном документе, которые приводят к модулированным уровням по меньшей мере одного редуцирующего сахара; и (c) интрогрессии одной или более мутаций во второе растение. Предпочтительно уровень по меньшей мере одного редуцирующего сахара определяют в зеленых листьях, листьях, подвергнутых ранним стадиям сушки, или подвергнутых сушке листьях. В одном варианте осуществления стадия интрогрессии включает селекцию растений, необязательно включая обратное скрещивание и т.п. В одном варианте осуществления первое растение является встречающимся в природе растением. В одном варианте осуществления второе растение характеризуется генетическим фоном, отличающимся от такового у первого растения. В одном варианте осуществления первое растение не является сортом или элитным сортом. В одном варианте осуществления второе растение представляет собой сорт или элитный сорт. Дополнительный аспект относится к мутантному растению (включая мутантное растение, являющееся сортом или элитным сортом), полученному или получаемому с помощью способов, описанных в данном документе. В определенных вариантах осуществления «мутантные растения» могут иметь одну или несколько мутаций, локализованных только в конкретном участке растения, например, в пределах последовательности одного или нескольких полинуклеотидов NtSUS, описанных в данном документе. Согласно данному варианту осуществления остальная геномная последовательность мутантного растения будет такой же или практически такой же, как у растения до мутагенеза.
В определенных вариантах осуществления мутантные растения могут иметь одну или более мутаций, локализованных в более чем одном участке генома растения, например, в пределах последовательности одного или более из полинуклеотидов NtSUS, описанных в данном документе, и в одном или более дополнительных участках генома. Согласно данному варианту осуществления остальная геномная последовательность мутантного растения не будет такой же или не будет практически такой же, как у растения до мутагенеза. В определенных вариантах осуществления мутантные растения могут не иметь одну или более мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более экзонах полинуклеотида (полинуклеотидов) NtSUS, описанного (описанных) в данном документе; или могут не иметь одну или более мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более интронах полинуклеотида(полинуклеотидов) NtSUS, описанного(описанных) в данном документе; или могут не иметь одну или более мутаций в промоторе полинуклеотида(полинуклеотидов)NtSUS, описанного(описанных) в данном документе; или могут не иметь одну или более мутаций в 3'-нетранслируемом участке полинуклеотида(полинуклеотидов) NtSUS, описанного(описанных) в данном документе; или могут не иметь одну или более мутаций в 5'-нетранслируемом участке полинуклеотида(полинуклеотидов) NtSUS, описанного(описанных) в данном документе; или могут не иметь одну или более мутаций в кодирующем участке полинуклеотида(полинуклеотидов) NtSUS, описанного(описанных) в данном документе; или могут не иметь одну или более мутаций в некодирующем участке полинуклеотида(полинуклеотидов) NtSUS, описанного(описанных) в данном документе; или любую комбинацию двух или более, трех или более, четырех или более, пяти или более, или шести или более их частей.
В дополнительном аспекте представлен способ идентификации растения, клетки растения или растительного материала, содержащих мутацию в гене, кодирующем полинуклеотид NtSUS, описанный в данном документе, включающий: (a) осуществление мутагенеза растения, клетки растения или растительного материала; (b) получение образца из указанного растения, клетки растения или растительного материала или их потомков и (c) определение полинуклеотидной последовательности гена NtSUS или его варианта или фрагмента, где отличие в указанной последовательности свидетельствует об одной или более мутациях в ней. Этот способ также обеспечивает отбор растений, имеющих мутацию(мутации), которая(которые) встречается(встречаются) в участках генома, которые влияют на экспрессию гена NtSUS в клетке растения, таких как сайт инициации транскрипции, стартовый кодон, участок интрона, граница экзон-интрон, терминатор или стоп-кодон.
Семейства растений, виды, сорта, семена и культура тканей
Растения, подходящие для применения в их отношении генетической модификации, включают однодольные и двудольные растения и системы клеток растений, в том числе виды из одного из следующих семейств: Acanthaceae, Alliaceae, Alstroemeriaceae, Amaryllidaceae, Apocynaceae, Arecaceae, Asteraceae, Berberidaceae, Bixaceae, Brassicaceae, Bromeliaceae, Cannabaceae, Caryophyllaceae, Cephalotaxaceae, Chenopodiaceae, Colchicaceae, Cucurbitaceae, Dioscoreaceae, Ephedraceae, Erythroxylaceae, Euphorbiaceae, Fabaceae, Lamiaceae, Linaceae, Lycopodiaceae, Malvaceae, Melanthiaceae, Musaceae, Myrtaceae, Nyssaceae, Papaveraceae, Pinaceae, Plantaginaceae, Poaceae, Rosaceae, Rubiaceae, Salicaceae, Sapindaceae, Solanaceae, Taxaceae, Theaceae или Vitaceae.
Подходящие виды могут включать представителей рода Abelmoschus, Abies, Acer, Agrostis, Allium, Alstroemeria, Ananas, Andrographis, Andropogon, Artemisia, Arundo, Atropa, Berberis, Beta, Bixa, Brassica, Calendula, Camellia, Camptotheca, Cannabis, Capsicum, Carthamus, Catharanthus, Cephalotaxus, Chrysanthemum, Cinchona, Citrullus, Coffea, Colchicum, Coleus, Cucumis, Cucurbita, Cynodon, Datura, Dianthus, Digitalis, Dioscorea, Elaeis, Ephedra, Erianthus, Erythroxylum, Eucalyptus, Festuca, Fragaria, Galanthus, Glycine, Gossypium, Helianthus, Hevea, Hordeum, Hyoscyamus, Jatropha, Lactuca, Linum, Lolium, Lupinus, Lycopersicon, Lycopodium, Manihot, Medicago, Mentha, Miscanthus, Musa, Nicotiana, Oryza, Panicum, Papaver, Parthenium, Pennisetum, Petunia, Phalaris, Phleum, Pinus, Poa, Poinsettia, Populus, Rauwolfia, Ricinus, Rosa, Saccharum, Salix, Sanguinaria, Scopolia, Secale, Solanum, Sorghum, Spartina, Spinacea, Tanacetum, Taxus, Theobroma, Triticosecale, Triticum, Uniola, Veratrum, Vinca, Vitis и Zea.
Подходящие виды могут включать Panicum spp., Sorghum spp., Miscanthus spp., Saccharum spp., Erianthus spp., Populus spp., Andropogon gerardii (бородач Жерара), Pennisetum purpureum (слоновая трава), Phalaris arundinacea (двукисточник тростниковидны), Cynodon dactylon (свинорой пальчатый), Festuca arundinacea (овсяница тростниковая), Spartina pectinata (спартина гребешковая), Medicago sativa (люцерна), Arundo donax (арундо тростниковый), Secale cereale (рожь), Salix spp. (ива), Eucalyptus spp. (эвкалипт), Triticosecale (тритикале), бамбук, Helianthus annuus (подсолнечник), Carthamus tinctorius (сафлор красильный), Jatropha curcas (ятрофа), Ricinus communis (клещевина), Elaeis guineensis (масличная пальма), Linum usitatissimum (лен), Brassica juncea, Beta vulgaris (сахарная свекла), Manihot esculenta (маниок), Lycopersicon esculentum (томат), Lactuca sativa (латук), Musyclise alca (банан), Solanum tuberosum (картофель), Brassica oleracea (брокколи, цветная капуста, брюссельская капуста), Camellia sinensis (чай), Fragaria ananassa (земляника), Theobroma cacao (какао), Coffeycliseca (кофе), Vitis vinifera (виноград), Ananas comosus (ананас), Capsicum annum (острый и сладкий перец), Allium cepa (лук), Cucumis melo (дыня), Cucumis sativus (огурец), Cucurbita maxima (тыква гигантская), Cucurbita moschata (тыква мускатная), Spinacea oleracea (шпинат), Citrullus lanatus (арбуз), Abelmoschus esculentus (бамия), Solanum melongena (баклажан), Rosa spp. (роза), Dianthus caryophyllus (гвоздика), Petunia spp. (петуния), Poinsettia pulcherrima (пуансеттия), Lupinus albus (люпин), Uniola paniculata (овес), полевица (Agrostis spp.), Populus tremuloides (тополь осинообразный), Pinus spp. (сосна), Abies spp. (пихта), Acer spp. (клен), Hordeum vulgare (ячмень), Poa pratensis (мятлик), Lolium spp. (плевел) и Phleum pratense (тимофеевка), Panicum virgatum (просо), Sorghuycliseor (сорго, суданская трава), Miscanthus giganteus (мискантус), Saccharum sp. (сахарный тростник), Populus balsamifera (тополь), Zea mays (кукуруза), Glycine max (соя), Brassica napus (канола), Triticum aestivum (пшеница), Gossypium hirsutum (хлопчатник), Oryza sativa (рис), Helianthus annuus (подсолнечник), Medicago sativa (люцерна), Beta vulgaris (сахарная свекла) или Pennisetum glaucum (просо жемчужное).
Различные варианты осуществления направлены на мутантный табак, не встречающийся в природе табак или трансгенные растения или клетки растения табака, модифицированные для модулирования уровней экспрессии гена, в результате чего получают растения или клетки растения, например растение или клетку растения табака, в которых уровень экспрессии полипептида модулирован в тканях, представляющих интерес, по сравнению с контролем. Раскрытые композиции и способы можно применять в отношении любого вида рода Nicotiana, включая N. rustica и N. tabacum (например, LA B21, LN KY171, TI 1406, Basma, Galpao, Perique, Beinhart 1000-1 и Petico). Другие виды включают N. acaulis, N. acuminata, N. africana, N. alata, N. ameghinoi, N. amplexicaulis, N. arentsii, N. attenuata, N. azambujae, N. benavidesii, N. benthamiana, N. bigelovii, N. bonariensis, N. cavicola, N. clevelandii, N. cordifolia, N. corymbosa, N. debneyi, N. excelsior, N. forgetiana, N. fragrans, N. glauca, N. glutinosa, N. goodspeedii, N. gossei, N. hybrid, N. ingulba, N. kawakamii, N. knightiana, N. langsdorffii, N. linearis, N. longiflora, N. maritima, N. megalosiphon, N. miersii, N. noctiflora, N. nudicaulis, N. obtusifolia, N. occidentalis, N. occidentalis subsp. hesperis, N. otophora, N. paniculata, N. pauciflora, N. petunioides, N. plumbaginifolia, N. quadrivalvis, N. raimondii, N. repanda, N. rosulata, N. rosulata subsp. ingulba, N. rotundifolia, N. setchellii, N. simulans, N. solanifolia, N. spegazzinii, N. stocktonii, N. suaveolens, N. sylvestris, N. thyrsiflora, N. tomentosa, N. tomentosiformis, N. trigonophylla, N. umbratica, N. undulata, N. velutina, N. wigandioides и N. x sanderae.В одном варианте осуществления растение представляет собой N. tabacum.
Применение сортов табака и элитных сортов табака также предусмотрено в данном документе. Трансгенное, не встречающееся в природе или мутантное растение, следовательно, может представлять собой разновидность табака или элитный сорт табака, которые содержат один или более трансгенов или одну или более генетических мутаций или их комбинацию. Генетическая мутация (мутации) (например, один или более полиморфизмов) могут представлять собой мутации, которые не существуют в природе в отдельной разновидности табака или сорте табака (например, элитном сорте табака), или может представлять собой генетическую мутацию (мутации), которая (которые) существует(существуют) в природе при условии, что мутация не существует в природе в отдельной разновидности табака или сорте табака (например, в элитном сорте табака).
Особенно применимые разновидности Nicotiana tabacum включают табак типа Берлей, табак темного типа, табак трубоогневой сушки и табак восточного типа. Неограничивающими примерами разновидностей или сортов являются: BD 64, CC 101, CC 200, CC 27, CC 301, CC 400, CC 500, CC 600, CC 700, CC 800, CC 900, Coker 176, Coker 319, Coker 371 Gold, Coker 48, CD 263, DF911, DT 538 LC табак Galpao, GL 26H, GL 350, GL 600, GL 737, GL 939, GL 973, HB 04P, HB 04P LC, HB3307PLC, гибрид 403LC, гибрид 404LC, гибрид 501 LC, K 149, K 326, K 346, K 358, K394, K 399, K 730, KDH 959, KT 200, KT204LC, KY10, KY14, KY 160, KY 17, KY 171, KY 907, KY907LC, KY14xL8 LC, Little Crittenden, McNair 373, McNair 944, msKY 14xL8, Narrow Leaf Madole, Narrow Leaf Madole LC, NBH 98, N-126, N-777LC, N-7371LC, NC 100, NC 102, NC 2000, NC 291, NC 297, NC 299, NC 3, NC 4, NC 5, NC 6, NC7, NC 606, NC 71, NC 72, NC 810, NC BH 129, NC 2002, Neal Smith Madole, OXFORD 207, PD 7302 LC, PD 7309 LC, PD 7312 LC, табак «Перик», PVH03, PVH09, PVH19, PVH50, PVH51, R 610, R 630, R 7-11, R 7-12, RG 17, RG 81, RG H51, RGH 4, RGH 51, RS 1410, Speight 168, Speight 172, Speight 179, Speight 210, Speight 220, Speight 225, Speight 227, Speight 234, Speight G-28, Speight G-70, Speight H-6, Speight H20, Speight NF3, TI 1406, TI 1269, TN 86, TN86LC, TN 90, TN 97, TN97LC, TN D94, TN D950, TR (Tom Rosson) Madole, VA 309, VA359, AA 37-1, B13P, Xanthi (Mitchell-Mor), Bel-W3, 79-615, Samsun Holmes NN, KTRDC номер 2 гибрид 49, Burley 21, KY8959, KY9, MD 609, PG01, PG04, PO1, PO2, PO3, RG11, RG 8, VA509, AS44, Banket A1, Basma Drama B84/31, Basma I Zichna ZP4/B, Basma Xanthi BX 2A, Batek, Besuki Jember, C104, Coker 347, Criollo Misionero, Delcrest, Djebel 81, DVH 405, Galpão Comum, HB04P, Hicks Broadleaf, Kabakulak Elassona, Kutsage E1, LA BU 21, NC 2326, NC 297, PVH 2110, Red Russian, Samsun, Saplak, Simmaba, Talgar 28, Wislica, Yayaldag, Prilep HC-72, Prilep P23, Prilep PB 156/1, Prilep P12-2/1, Yaka JK-48, Yaka JB 125/3, TI-1068, KDH-960, TI-1070, TW136, Basma, TKF 4028, L8, TKF 2002, GR141, Basma xanthi, GR149, GR153, Petit Havana. Также предусмотрены подразновидности вышеуказанного с низким уровнем превращения никотина в норникотин, даже если они специально не указаны в данном документе.
Варианты осуществления также направлены на композиции и способы получения мутантных растений, не встречающихся в природе растений, гибридных растений или трансгенных растений, которые были модифицированы для модулирования экспрессии или функции полинуклеотида (полинуклеотидов) NtSUS, описанного (описанных) в данном документе (или любой их комбинации, описанной в данном документе). Преимущественно, полученные мутантные растения, не встречающиеся в природе растения, гибридные растения или трансгенные растения могут быть сходными по общему внешнему виду с контрольными растениями или практически такими же. Различные фенотипические характеристики, такие как степень зрелости, количество листьев на растении, высоту стебля, угол врастания листьев, размер листа (ширина и длина), расстояние междоузлия и соотношение листовая пластина-главная жилка можно оценить путем полевых наблюдений.
Один аспект относится к семени мутантного растения, не встречающегося в природе растения, гибридного растения или трансгенного растения, описанного в данном документе. Предпочтительно, семя представляют собой семя табака. Дополнительный аспект относится к пыльце или семяпочке мутантного растения, не встречающегося в природе растения, гибридного растения или трансгенного растения, описанного в данном документе. Кроме того, предусмотрено мутантное растение, не встречающееся в природе растение, гибридное растение или трансгенное растение, описанное в данном документе, которое дополнительно содержит полинуклеотид, обеспечивающий развитие мужской стерильности.
Также представлена культура тканей из регенерируемых клеток мутантного растения, не встречающегося в природе растения, гибридного растения или трансгенного растения или его части, как описано в данном документе, при этом из культуры регенерируют растения, способные экспрессировать все морфологические и физиологические характеристики родителя. Регенерируемые клетки включают клетки из листьев, пыльцы, зародышей, семядолей, гипокотилей, корней, кончиков корней, пыльников, цветков и их части, семяпочек, побегов, стеблей, черешков, сердцевины и семенных коробочек, или каллюса, или протопластов, полученных из них.
Растительный материал, описанный в данном документе, может представлять собой подвергнутый сушке табачный материал, такой как табачный материал, подвергнутый трубоогневой сушке, такой как табак типа Вирджиния, или табачный материал, подвергнутый солнечной сушке, такой как табак типа Берлей и темного типа. Растительный материал, описанный в данном документе, может представлять собой табачный материал, подвергнутый воздушной сушке, такой как табак типа Берлей и темного типа.
Рекомендация CORESTA в отношении сушки табака описана в справочнике CORESTA № 17, апрель 2016 г., Sustainability in Leaf Tobacco Production (Принципы устойчивого развития в производстве листового табака).
Модулирование содержания редуцирующих сахаров
Одна цель заключается в получении мутантных, трансгенных или не встречающихся в природе растений или их частей, которые проявляют модулированные уровни по меньшей мере одного редуцирующего сахара в растительном материале, например, в подвергнутых сушке листьях. Соответственно, мутантные, трансгенные или не встречающиеся в природе растения или их части проявляют модулированные уровни по меньшей мере одного редуцирующего сахара по сравнению с контрольным растением. Предпочтительно, мутантные, трансгенные или не встречающиеся в природе растения или их части имеют по существу тот же внешний вид, что и контрольное растение.
Соответственно, в данном документе описаны мутантные, трансгенные или не встречающиеся в природе растения или их части или клетки растений, которые характеризуются модулированными уровнями по меньшей мере одного редуцирующего сахара по сравнению с контрольными клетками или контрольными растениями. Мутантные, трансгенные или не встречающиеся в природе растения или клетки растений были подвергнуты модификации для модулирования синтеза или функции одного или более из полипептидов NtSUS, описанных в данном документе, путем модулирования экспрессии одного или более из соответствующих полинуклеотидов NtSUS, описанных в данном документе. Соответственно, модулированные уровни по меньшей мере одного редуцирующего сахара наблюдаются в по меньшей мере зеленых листьях, соответственно, листьях, подвергнутых ранним стадиям сушки, или подвергнутых сушке листьях. В определенных вариантах осуществления можно модулировать уровень редуцирующих сахаров в растении, например, в зеленых листьях, предпочтительно листьях, подвергнутых ранним стадиям сушки, или подвергнутых сушке листьях, или подвергнутом сушке табаке. В определенных вариантах осуществления можно модулировать уровень редуцирующих сахаров в растении, например, в зеленых листьях, предпочтительно листьях, подвергнутых ранним стадиям сушки, подвергнутых сушке листьях или подвергнутом сушке табаке.
В определенных вариантах осуществления модулируется активность и/или экспрессия одной или более из NtSUS1-S, NtSUS1-T, NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
В определенных вариантах осуществления модулируется активность и/или экспрессия одной или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T.
В определенных вариантах осуществления модулируется активность и/или экспрессия одной или более из NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T и не модулируется активность и/или экспрессия одной или более из NtSUS1-S, NtSUS1-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
В определенных вариантах осуществления модулируется активность и/или экспрессия NtSUS2-S, NtSUS2-T, NtSUS3-S, NtSUS3-T, NtSUS4-S и NtSUS4-T и не модулируется активность и/или экспрессия NtSUS1-S, NtSUS1-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
В определенных вариантах осуществления модулируется активность и/или экспрессия одной или более из NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S и не модулируется активность и/или экспрессия одной или более из NtSUS1-S, NtSUS1-T, NtSUS2-T, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T.
В определенных вариантах осуществления модулируется активность и/или экспрессия NtSUS2-S, NtSUS3-S, NtSUS3-T и NtSUS4-S и не модулируется активность и/или экспрессия NtSUS1-S, NtSUS1-T, NtSUS2-T, NtSUS4-T, NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T. Дополнительный аспект относится к мутантным, не встречающимся в природе или трансгенным растению или клетке, где экспрессия или функция одного или более из полипептидов NtSUS, описанных в данном документе, является модулированной, и часть растения (например, зеленые листья, предпочтительно листья, подвергнутые ранним стадиям сушки, или подвергнутые сушке листья, или подвергнутый сушке табак) характеризуется сниженными уровнями по меньшей мере одного редуцирующего сахара на по меньшей мере 5% по сравнению с контрольным растением, в котором экспрессия или функция указанного (указанных) полипептида(полипептидов) NtSUS не была модулирована. В определенных вариантах осуществления уровень по меньшей мере одного редуцирующего сахара в растении, например, в зеленых листьях, предпочтительно листьях, подвергнутых ранним стадиям сушки, или подвергнутых сушке листьях, или подвергнутом сушке табаке, можно модулировать, например, на по меньшей мере приблизительно 5%.
Еще один аспект относится к подвергнутому сушке растительному материалу, такому как подвергнутый сушке лист или подвергнутый сушке табак, полученному или получаемому из мутантных, не встречающихся в природе или трансгенных растения или клетки, где экспрессия одного или более из полинуклеотидов NtSUS, описанных в данном документе, или функция полипептида NtSUS, кодируемого ими, являются модулироваными и где уровень по меньшей мере одного редуцирующего сахара модулирован на по меньшей мере 5% по сравнению с контрольным растением.
Предпочтительно, внешний вид указанного растения или его части (например, листа) является практически таким же, как у контрольного растения. Предпочтительно, растение представляет собой растение табака или кофейное растение.
Варианты осуществления также направлены на композиции и способы получения мутантных, не встречающихся в природе или трансгенных растений или клеток растений, которые были модифицированы для модулирования экспрессии или функции одного или более из полинуклеотидов NtSUS или полипептидов NtSUS, описанных в данном документе, что может приводить к растениям или компонентам растения (например, листьям, таким как зеленые листья, или листья, подвергнутые ранним стадиям сушки, или подвергнутые сушке листья, или табак) или клеткам растений с модулированным содержанием редуцирующих сахаров.
Мутантные, не встречающиеся в природе или трансгенные растения, полученные в соответствии с настоящим изобретением, по внешнему виду могут быть подобны контрольным растениям или быть по существу такими же как контрольные растения. В одном варианте осуществления вес листьев у мутантного, не встречающегося в природе или трансгенного растения является практически таким же, как у контрольного растения. В одном варианте осуществления количество листьев у мутантного, не встречающегося в природе или трансгенного растения является практически таким же, как у контрольного растения. В одном варианте осуществления вес листьев и количество листьев у мутантного, не встречающегося в природе или трансгенного растения является практически таким же, как у контрольного растения. В одном варианте осуществления высота стебля у мутантных, не встречающихся в природе или трансгенных растений является практически такой же, как у контрольных растений, через, например, один, два или три или более месяцев после пересадки в поле или через 10, 20, 30 или 36 или более дней после обрезания верхушек. Например, высота стебля у мутантных, не встречающихся в природе или трансгенных растений не меньше, чем высота стебля у контрольных растений. В другом варианте осуществления содержание хлорофилла у мутантных, не встречающихся в природе или трансгенных растений является практически таким же, как у контрольных растений. В другом варианте осуществления высота стебля у мутантных, не встречающихся в природе или трансгенных растений является практически такой же, как у контрольных растений, и содержание хлорофилла у мутантных, не встречающихся в природе или трансгенных растений является практически таким же, как у контрольных растений. В других вариантах осуществления размер или форма или количество или окраска листьев у мутантных, не встречающихся в природе или трансгенных растений являются практически такими же, как у контрольных растений. Предпочтительно, растение представляет собой растение табака или кофейное растение.
В другом аспекте представлен способ модулирования количества по меньшей мере одного редуцирующего сахара в по меньшей мере части растения (например, листьях, таких как подвергнутые сушке листья, или в табаке), включающий стадии: (i) модулирования экспрессии или функции одного или более из полипептидов NtSUS, описанных в данном документе (или любой их комбинаций, описанной в данном документе), предпочтительно, где полипептид (полипептиды) NtSUS кодируется (кодируются) соответствующими полинуклеотидами NtSUS, описанными в данном документе; (ii) измерения уровня по меньшей мере одного редуцирующего сахара в по меньшей мере части (например, листьях, таких как подвергнутые сушке листья, или в табаке, или в дыме) мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (i); и (iii) идентификации мутантного, не встречающегося в природе или трансгенного растения, в котором уровень по меньшей мере одного редуцирующего сахара был модулирован по сравнению с контрольным растением. Предпочтительно внешний вид указанного мутантного, не встречающегося в природе или трансгенного растения является практически таким же, как у контрольного растения. Предпочтительно растение представляет собой растение табака.
В другом аспекте представлен способ модулирования количества по меньшей мере одного редуцирующего сахара в по меньшей мере части подвергнутого сушке растительного материала, такого как подвергнутый сушке лист, включающий стадии: (i) модулирования экспрессии или функции одного или более из полипептидов NtSUS (или любой их комбинации, описанной в данном документе), предпочтительно, где полипептид (полипептиды) NtSUS кодируется(кодируются) соответствующими полинуклеотидами NtSUS, описанными в данном документе; (ii) сбора растительного материала, такого как один или более листьев, и сушки в течение определенного периода времени; (iii) измерения уровня по меньшей мере одного редуцирующего сахара в по меньшей мере части подвергнутого сушке растительного материала, полученного на стадии (ii) или во время стадии (ii); и (iv) идентификации подвергнутого сушке растительного материала, в котором уровень по меньшей мере одного редуцирующего сахара был модулирован по сравнению с контрольным растением.
Повышение экспрессии по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или повышение составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% или более, например, 200%, 300%, 500%, 1000% или более, что включает повышение транскрипционной функции, или уровня экспрессии полинуклеотида NtSUS, или уровня экспрессии полипептида NtSUS, или их комбинацию.
Повышение функции или активности по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или повышение составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% или более, как, например, 200%, 300%, 500%, 1000% или более.
Снижение экспрессии по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или снижение составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100%, что включает снижение транскрипционной функции, или экспрессии полинуклеотида NtSUS, или экспрессии полипептида NtSUS, или их комбинацию.
Понижение функции или активности по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или понижение составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100%.
Полинуклеотиды и рекомбинантные конструкции, описанные в данном документе, можно применять для модулирования экспрессии, или функции, или активности полинуклеотидов NtSUS или полипептидов NtSUS, описанных в данном документе, в представляющем интерес виде растений, предпочтительно в табаке.
Целый ряд способов на основе полинуклеотидов можно применять для повышения уровня экспрессии генов в растениях и клетках растений. В качестве примера, можно получать конструкцию, вектор или вектор экспрессии, совместимые с подлежащим трансформации растением, которые содержат представляющий интерес ген вместе с расположенным выше по последовательности промотором, способным к обеспечению сверхэкспрессии гена в растении или клетке растения. Иллюстративные промоторы описаны в данном документе. После трансформации и при выращивании в подходящих условиях промотор может управлять экспрессией для модулирования уровней NtSUS в растении или в его конкретной ткани. В одном иллюстративном варианте осуществления создают вектор, несущий один или более полинуклеотидов NtSUS, описанных в данном документе (или любую их комбинацию, описанную в данном документе), для сверхэкспрессии гена в растении или клетке растения. Вектор несет подходящий промотор, такой как промотор 35S вируса мозаики цветной капусты CaMV, расположенный выше по последовательности от трансгена, который запускает его конститутивную экспрессию во всех тканях растения. Вектор также несет ген устойчивости к антибиотику с целью обеспечения возможности отбора трансформированных каллюсов и линий клеток.
Экспрессию последовательностей с промоторов можно усилить посредством включения последовательностей, обеспечивающих контроль экспрессии, в том числе энхансеров, активирующих хроматин элементов, элементов, чувствительных к факторам транскрипции и т. п. Такие последовательности, обеспечивающие контроль, могут быть конститутивными и повышать уровень транскрипции универсальным образом; или они могут быть факультативными и повышать уровень транскрипции в ответ на конкретные сигналы. Отдельно следует указать сигналы, связанные со старением, и сигналы, которые активны во время процедуры сушки.
Таким образом, различные варианты осуществления направлены на способы модулирования уровня экспрессии одного или более полинуклеотидов NtSUS, описанных в данном документе (или любой их комбинации, описанной в данном документе), путем интеграции нескольких копий полинуклеотида NtSUS в геном растения, включающие: трансформацию клетки растения-хозяина с помощью вектора экспрессии, который содержит промотор, функционально связанный с одним или более полинуклеотидами NtSUS, описанными в данном документе. Полипептид, кодируемый рекомбинантным полинуклеотидом, может являться нативным полипептидом или может являться гетерологичным по отношению к клетке.
В одном варианте осуществления растение для применения по настоящему изобретению представляет собой растение, которое подвергнуто трубоогневой сушке, поскольку такие растения характеризуются высоким содержанием редуцирующих сахаров (более приблизительно 14% от сухого веса при выращивании в поле в конце сушки). Мутантные, трансгенные или не встречающиеся в природе растения или их части, которые подвергнуты трубоогневой сушке, могут характеризоваться содержанием редуцирующих сахаров, которое составляет менее приблизительно 14% сухого веса при выращивании в поле в конце сушки, например, менее приблизительно 10% от сухого веса при выращивании в поле в конце сушки, или менее приблизительно 5% от сухого веса при выращивании в поле в конце сушки, или менее приблизительно 1% от сухого веса при выращивании в поле в конце сушки.
В одном варианте осуществления растение для применения по настоящему изобретению представляет собой растение, которое подвергнуто солнечной сушке, поскольку такие растения характеризуются содержанием редуцирующего сахара, составляющим более приблизительно 6,8% от сухого веса при выращивании в поле в конце сушки. Мутантные, трансгенные или не встречающиеся в природе растения или их части, которые подвергнуты солнечной сушке, могут характеризоваться содержанием редуцирующих сахаров, составляющим менее приблизительно 5% от сухого веса при выращивании в поле в конце сушки, например, менее приблизительно 2,5% от сухого веса при выращивании в поле в конце сушки, или менее приблизительно 1% от сухого веса при выращивании в поле в конце сушки.
В одном варианте осуществления растение для применения по настоящему изобретению представляет собой растение, подвергнутое воздушной сушке. Такие растения характеризуются содержанием редуцирующих сахаров, составляющим более приблизительно 1,7% от сухого веса при выращивании в поле в конце сушки. Мутантные, трансгенные или не встречающиеся в природе растения или их части, которые подвергнуты солнечной сушки, могут характеризоваться содержанием редуцирующих сахаров, составляющим менее приблизительно 1,5% от сухого веса при выращивании в поле в конце сушки, например, менее приблизительно 1% от сухого веса при выращивании в поле в конце сушки, или менее приблизительно 0,5% от сухого веса при выращивании в поле в конце сушки.
В определенных вариантах осуществления предпочтительно применение растения, подвергнутого трубоогневой сушке или солнечной сушке.
Измерение содержания свободных аминокислот
Содержание аминокислот можно измерять с помощью различных способов, известных из уровня техники. Одним из таких способов является способ MP 1471 rev 5 2011, Resana, Italy: Chelab Silliker S.r.l, Mérieux NutriSciences Company. Для выявления аминокислот в листьях растения, подвергнутых сушке, после удаления средней жилки пластинки, подвергнутые сушке, при необходимости сушат при 40°C в течение 2-3 дней. Затем табачный материал измельчают в тонкодисперсный порошок (~100 мкM) перед анализом на содержание аминокислот. Другой способ измерения содержания аминокислот в растительном материале описан в UNI EN ISO 13903:2005.В определенных вариантах осуществления измерение содержания свободных аминокислот проводят в соответствии с UNI EN ISO 13903:2005.
Измерение содержания редуцирующих сахаров
Содержание редуцирующих сахаров можно измерять с применением колориметрического способа с сегментированным потоком, разработанного для анализа образцов табака, адаптированного в Skalar Instrument Co (Западный Честер, Пенсильвания) и описанного в Tobacco Science 20: 139-144 (1976). Измерение содержания редуцирующих сахаров также описано в Coresta Recommended Method 38, CRM38, CRM и ISO 15154: 2003. Для определения редуцирующих сахаров в подвергнутых сушке листьях подвергнутые сушке пластинки после удаления средней жилки сушат при 40°C в течение 2-3 дней, если необходимо. Затем табачный материал измельчают в тонкодисперсный порошок (~100 мкм) перед проведением анализов на редуцирующие сахара. В определенных вариантах осуществления измерение содержания редуцирующих сахаров проводят в соответствии с ISO 15154: 2003.
Селекция
Растение, несущее мутантный аллель одного или более полинуклеотидов NtSUS, описанных в данном документе (или любой их комбинации, описанной в данном документе), можно применять в программе селекции растений для создания применимых линий, сортов и гибридов. В частности, мутантный аллель интрогрессируют в коммерчески важные разновидности, описанные выше. Таким образом, предложены способы селекции растений, которые предусматривают скрещивание мутантного растения, не встречающегося в природе растения или трансгенного растения, как описано в данном документе, с растением, характеризующимся иными генетическими особенностями. Способ может дополнительно включать скрещивание растения-потомка с другим растением и необязательно повторение скрещивания до тех пор, пока не будет получено потомка с необходимыми генетическими признаками или генетическим фоном. Одной из целей, для которой подходят такие способы селекции, является введение желаемого генетического признака в другие разновидности, селекционные линии, гибриды или сорта, особенно те, которые представляют коммерческий интерес. Другой целью является облегчение накопления генетических модификаций различных генов в отдельных разновидностях, линиях, гибридах или сортах растений. Предусмотрены внутривидовые, а также межвидовые скрещивания. Растения-потомки, которые возникают в результате таких скрещиваний, также называемые селекционными линиями, являются примерами не встречающихся в природе растений по настоящему изобретению.
В одном варианте осуществления предусмотрен способ получения не встречающегося в природе растения, включающий: (a) скрещивание мутантного или трансгенного растения со вторым растением с получением семени растения табака-потомка; (b) выращивание семени растения табака-потомка в пригодных для роста растений условиях с получением не встречающегося в природе растения. Способ может дополнительно включать: (c) скрещивание предыдущего поколения не встречающегося в природе растения с самим собою или с другим растением с получением семени растения табака-потомка; (d) выращивание семени растения табака-потомка из стадии (c) в пригодных для роста растений условиях с получением дополнительных не встречающихся в природе растений; и (e) повторение стадий скрещивания и выращивания (c) и (d) несколько раз с получением следующих поколений не встречающихся в природе растений. Способ может необязательно предусматривать перед стадией (a) стадию получения родительского растения, которое содержит охарактеризованные генетические особенности и которое не идентично мутантному или трансгенному растению. В некоторых вариантах осуществления в зависимости от программы селекции стадии скрещивания и выращивания повторяют от 0 до 2 раз, от 0 до 3 раз, от 0 до 4 раз, от 0 до 5 раз, от 0 до 6 раз, от 0 до 7 раз, от 0 до 8 раз, от 0 до 9 раз или от 0 до 10 раз для получения поколений не встречающихся в природе растений. Обратное скрещивание является примером такого способа, в котором потомка скрещивают с одним из его родителей или с другим растением, генетически сходным с его родителем, для получения растения-потомка в следующем поколении, которое имеет генетические особенности, более близкие к одному из родителей. Методики селекции растений, в частности селекции растений, хорошо известны и могут применяться в способах по настоящему изобретению. В настоящем изобретении дополнительно предложены не встречающиеся в природе растения, полученные с помощью данных способов. Определенные варианты осуществления исключают стадию отбора растения.
В некоторых вариантах осуществления способов, описанных в данном документе, линии, полученные в результате селекции и скрининга на наличие вариантных генов, оценивают в поле с применением стандартных полевых процедур. Предусмотрены контрольные генотипы, в том числе генотип исходного не подвергнутого мутагенезу родителя, и места посадки растений расположены в поле согласно рандомизированному полноблочному плану или согласно другому соответствующему планированию поля. Для табака используют стандартные агрономические методики, например, табак собирают, взвешивают и отбирают образцы для химического и другого общепринятого тестирования до и во время сушки. Статистический анализ данных выполняют для подтверждения сходства отобранных линий с родительской линией. Цитогенетические анализы отобранных растений необязательно выполняют для подтверждения взаимосвязей набора хромосом и конъюгации хромосом.
ДНК-фингерпринтинг, однонуклеотидный полиморфизм, микросателлитные маркеры или подобные технологии можно применять в программе селекции с отбором с помощью маркера (MAS) для переноса или разведения мутантных аллелей гена в других растениях табака, как описано в данном документе. Например, селекционер может создать расщепляющиеся популяции при гибридизации генотипа, содержащего мутантный аллель, с необходимым с агрономической точки зрения генотипом. Растения из F2 или поколений, полученных в результате обратного скрещивания, можно подвергнуть скринингу с применением маркера, полученного из геномной последовательности или ее фрагмента, с применением одной из методик, перечисленных в данном документе. Растения, идентифицированные как обладающие мутантным аллелем, можно подвергнуть обратному скрещиванию или самоопылению для создания второй популяции, подлежащей скринингу. В зависимости от предполагаемого способа наследования или применяемой технологии MAS для отобранных растений может быть необходимым самоопыление перед каждым циклом обратного скрещивания для облегчения обнаружения необходимых отдельных растений. Обратное скрещивание или другую процедуру селекции можно повторять до тех пор, пока необходимый фенотип рекуррентного родителя не восстановится.
Согласно настоящему изобретению в программе селекции успешные скрещивания дают растения F1, которые являются фертильными. Отобранные растения F1 можно скрещивать с одним из родителей, и для растений первого поколения, полученного в результате обратного скрещивания, можно обеспечивать самоопыление с получением популяции, которую снова подвергают скринингу в отношении экспрессии вариантного гена (например, нулевой версии гена). Процесс обратного скрещивания, самоопыления и скрининга повторяют, например, по меньшей мере 4 раза до тех пор, пока при окончательном скрининге не получат растение, которое является фертильным и в достаточной степени подобным рекуррентному родителю. Это растение, при необходимости, самоопыляют и затем потомство снова подвергают скринингу, чтобы подтвердить, что растение демонстрирует экспрессию вариантного гена. В некоторых вариантах осуществления популяцию растений в поколении F2 подвергают скринингу на наличие экспрессии вариантного гена, например растение, которое не экспрессирует полипептид из-за отсутствия гена, идентифицируют согласно стандартным способам, например с помощью методики ПЦР с применением праймеров, созданных на основе информации о последовательности полинуклеотидов для полинуклеотида (полинуклеотидов), описанного в данном документе (или любой их комбинации, описанной в данном документе).
Разновидности гибридного табака можно получить путем предотвращения самоопыления женских родительских растений (то есть родительских форм) первой разновидности, позволяя пыльце с мужских родительских растений второй разновидности оплодотворить женские родительские растения и обеспечивая образование гибридных семян F1 на женских растениях. Самоопыление женских растений можно предотвратить путем кастрации цветков на ранней стадии развития цветка. В качестве альтернативы, образование пыльцы на женских родительских растениях можно предотвратить, используя какую-либо форму мужской стерильности. Например, мужскую стерильность можно обеспечить посредством цитоплазматической мужской стерильности (CMS) или трансгенной мужской стерильности, где трансген подавляет микроспорогенез и/или образование пыльцы или вызывает самонесовместимость. Женские родительские растения, содержащие CMS, являются особенно применимыми. В вариантах осуществления, в которых женские родительские растения характеризуются CMS, пыльцу собирают с мужских фертильных растений и наносят вручную на рыльца женских родительских растений с CMS и собирают полученные семена F1.
Разновидности и линии, описанные в данном документе, можно применять для образования простых гибридов F1 табака. В таких вариантах осуществления растения родительских разновидностей можно выращивать в виде практически однородных смежных популяций для облегчения естественного перекрестного опыления женских родительских растений мужскими родительскими растениями. Семена F1, образовавшиеся на женских родительских растениях, выборочно собирают с помощью обычных средств. Можно также вырастить две разновидности родительского растения в массе и собрать смесь гибридных F1 семян, образовавшихся на женской особи, и семян, образовавшихся на мужской особи в результате самоопыления. В качестве альтернативы, можно осуществить трехлинейное скрещивание, где простой гибрид F1 используют в качестве женской особи и скрещивают с другой мужской особью. В качестве другой альтернативы, можно создать гибриды двойного скрещивания, где потомство F1 двух разных простых гибридов скрещивают само с собой.
Популяцию мутантных, не встречающихся в природе или трансгенных растений можно подвергнуть скринингу или отбору в отношении тех представителей популяции, которые имеют необходимый признак или фенотип. Например, популяцию из потомства линии с одной трансформацией можно подвергнуть скринингу в отношении тех растений, которые имеют необходимый уровень экспрессии или функции полипептида (полипептидов), кодируемого с ее помощью. Физические и биохимические способы можно применять для выявления уровней экспрессии или активности. Они включают анализ по Саузерну или ПЦР-амплификацию для выявления полинуклеотида; нозерн-блоттинг, анализ с защитой от РНКазы S1, анализ методом удлинения праймера или RT-PCR-амплификацию для выявления РНК-транскриптов; ферментные анализы для выявления ферментативной или рибозимной функции полипептидов и полинуклеотидов; и гель-электрофорез полипептида, вестерн-блоттинг, иммунопреципитацию и иммуноферментные анализы для выявления полипептидов. Другие методики, такие как гибридизация in situ, ферментное окрашивание и иммуноокрашивание, и ферментные анализы также можно применять для выявления присутствия или экспрессии, функции или активности полипептидов NtSUS или полинуклеотидов NtSUS.
В данном документе описаны мутантные, не встречающиеся в природе или трансгенные клетки растения и растения, содержащие один или более рекомбинантных полинуклеотидов, одну или более полинуклеотидных конструкций, одну или более двунитевых РНК, один или более конъюгатов или один или более векторов/векторов экспрессии.
5. Модификация других генов
В частности, растения и их части, описанные в данном документе, можно модифицировать либо до, либо после модулирования экспрессии, функции или активности одного или более полинуклеотидов NtSUS и/или полипептидов NtSUS в соответствии с настоящим изобретением.
Одна или более следующих дополнительных генетических модификаций могут присутствовать в мутантных, не встречающихся в природе или трансгенных растениях или их частях.
Один или более генов, которые вовлечены в превращение промежуточных продуктов азотного обмена могут быть модифицированы, что в результате приводит к более низким уровням по меньшей мере одного специфичного для табака нитрозамина (TSNA). Неограничивающие примеры таких генов включают гены, кодирующие никотин-деметилазу, такие как CYP82E4, CYP82E5 и CYP82E10, описанные в WO2006/091194, WO2008/070274, WO2009/064771 и WO2011/088180, а также нитратредуктазу, описанные в WO2016/046288.
Один или более генов, которые принимают участие в поглощении тяжелых металлов или транспорте тяжелых металлов, могут быть модифицированы, что в результате приводит к более низкому содержанию тяжелых металлов. Неограничивающие примеры включают семейства полипептидов, ассоциированных с множественной лекарственной устойчивостью, семейства посредников диффузии катионов (CDF), семейство Zrt-, Irt-подобных полипептидов (ZIP), семейство катионообменников (CAX), семейство транспортеров меди (COPT), семейство АТФаз тяжелых металлов (например, HMA, как описано в WO2009/074325 и WO2017/129739), семейство гомологов полипептидов макрофагов, ассоциированных с естественной устойчивостью (NRAMP), и другие члены семейства транспортеров с АТФ-связывающей кассетой (ABC) (например, MRP), как описано в WO2012/028309, которые участвуют в транспорте тяжелых металлов, таких как кадмий.
Другие иллюстративные модификации могут приводить к получению растений с модулированной экспрессией или функцией изопропилмалатсинтазы, что в результате приводит к изменению композиции сложного эфира сахарозы, которая может использоваться для изменения ароматического профиля (см. WO2013/029799).
Другие иллюстративные модификации могут приводить к получению растений с модулированной экспрессией или функцией треонинсинтазы, при этом можно модулировать уровни метионала (см. WO2013/029800).
Другие иллюстративные модификации могут приводить к получению растений с модулированной экспрессией или функцией одного или более из неоксантинсинтазы, ликопен-бета-циклазы и 9-цис-эпоксикаротиноиддиоксигеназы для модуляции содержания бета-дамасценона, чтобы изменить вкусоароматический профиль (см. WO2013/064499).
Другие иллюстративные модификации могут приводить к получению растений с модулированными экспрессией или функцией членов семейства CLC хлоридных каналов для модулирования в них уровней нитратов (см. WO2014/096283 и WO2015/197727).
Другие иллюстративные модификации могут приводить к получению растений с модулированной экспрессией или функцией одной или более аспарагинсинтетаз для модулирования уровней аспарагина в листе и обеспечения модулированных уровней акриламида в аэрозоле, полученном при нагревании или горении листа (см. WO2017/042162).
Примеры других модификаций включают модуляцию переносимости гербицида, например, глифосат является активным ингредиентом множества гербицидов широкого спектра действия. Устойчивые к глифосату трансгенные растения были разработаны путем переноса гена aroA (глифосат-EPSP-синтетаза из Salmonella typhimurium и E.coli). Устойчивые к сульфонилмочевине растения были получены путем трансформации мутантного гена ALS (ацетолактатсинтетаза) из Arabidopsis. Полипептид OB фотосистемы II из мутантного Amaranthus hybridus был перенесен в растения с получением атразин-устойчивых трансгенных растений; и бромоксинил-устойчивые трансгенные растения были получены путем встраивания гена bxn из бактерии Klebsiella pneumoniae.
Другие примеры модификаций приводят в результате к получению растений, которые устойчивы к насекомым. Токсины Bacillus thuringiensis(Bt) могут обеспечить эффективный путь отсрочки появления Bt-устойчивых вредителей, как недавно показано на брокколи, где гены Bt cry1Ac и cry1C в составе «пирамиды» обеспечивали контроль капустной моли, устойчивой к любому отдельному полипептиду, и существенно задерживали эволюционное развитие устойчивых насекомых.
Другая иллюстративная модификация в результате приводит к получению растений, которые устойчивы к заболеваниям, вызванным патогенами (например, вирусами, бактериями, грибами). Были разработаны растения, экспрессирующие ген Xa21 (устойчивость к бактериальному некрозу), с растениями, экспрессирующими как ген слияния Bt, так и ген хитиназы (устойчивость к желтой огневке-травянке и выносливость в отношении ризоктониоза).
Другая иллюстративная модификация в результате приводит к корректированию репродуктивной способности, например к мужской стерильности.
Другая иллюстративная модификация приводит в результате к получению растений, которые выносливы в отношении абиотического стресса (например, засухе, изменению температуры, засоленности), и выносливые трансгенные растения были получены путем переноса фермента ацилглицерол-фосфат-ацилтрансферазы из Arabidopsis; гены, кодирующие маннитол-дегидрогеназу и сорбитолдегидрогеназу, которые принимают участие в синтезе маннита и сорбита, улучшают устойчивость к засухе.
Другая иллюстративная модификация приводит к растениям, в которых модулирована активность одной или нескольких эндогенных гликозилтрансфераз, таких как N-ацетилглюкозаминилтрансфераза, β(1,2)-ксилозилтрансфераза и a(1,3)-фукозилтрансфераза (см. WO2011/117249).
Другая иллюстративная модификация в результате приводит к получению растений, в которых активность одной или более никотин-N-деметилаз модулируют таким образом, что могут модулироваться уровни норникотина и метаболитов норникотина, которые образуются во время сушки (см. WO2015169927).
Другие иллюстративные модификации могут приводить к получению растений с улучшенными характеристиками запасных полипептидов и масел, растений с повышенной эффективностью фотосинтеза, растений с длительным сроком хранения, растений с повышенным содержанием углеводов и растений, устойчивых к грибам. Также представлены трансгенные растения, в которых была подвергнута модулированию экспрессия S-аденозил-L-метионина (SAM) и/или цистатионин-гамма-синтазы (CGS).
Один или более генов, которые вовлечены в путь синтеза никотина, можно модифицировать с получением растений или частей растений, которые при сушке вырабатывают модулированные уровни никотина. Гены синтеза никотина могут быть выбраны из группы, состоящей из: A622, BBLa, BBLb, JRE5L1, JRE5L2, MATE1, MATE 2, MPO1, MPO2, MYC2a, MYC2b, NBB1, nic1, nic2, NUP1, NUP2, PMT1, PMT2, PMT3, PMT4 и QPT или комбинации одного или более из них.
Один или более генов, которые вовлечены в регулирование количества одного или более алкалоидов, можно модифицировать с получением растений или частей растений, которые при сушке вырабатывают модулированные уровни алкалоида. Гены, обеспечивающие контроль уровня алкалоидов, могут быть выбраны из группы, состоящей из BBLa, BBLb, JRE5L1, JRE5L2, MATE1, MATE 2, MYC2a, MYC2b, nic1, nic2, NUP1 и NUP2 или комбинации двух или более из них.
Один или более таких признаков можно интрогрессировать в мутантные, не встречающиеся в природе или трансгенные растения другого сорта или можно непосредственно трансформировать в них.
В различных вариантах осуществления предусмотрены мутантные растения, не встречающиеся в природе растения или трансгенные растения, а также биомасса, в которых уровень экспрессии одного или более полинуклеотидов в соответствии с настоящим изобретением являются модулированными для модулирования таким образом уровня полипептида(полипептидов), кодируемого ими.
6. Продукты потребления
Части растений, описанных в данном документе, в частности листовую пластинку и среднюю жилку данных растений, можно включить в изготовление или применять в изготовлении различных продуктов потребления, включая без ограничения материалы, образующие аэрозоль, устройства, образующие аэрозоль, курительные изделия, изделия для курения, бездымные продукты, медицинские или косметические продукты, препараты для внутривенного введения, таблетки, порошки и табачные продукты. Примеры материалов, образующих аэрозоль, включают табачные композиции, виды табака, табачный экстракт, резаный табак, резаный наполнитель, сушеный табак, взорванный табак, гомогенизированный табак, восстановленный табак и виды трубочного табака. Курительные изделия и изделия для курения являются, как правило, устройствами, образующими аэрозоль. Примеры курительных изделий или изделий для курения включают сигареты, сигариллы и сигары. Примеры бездымных продуктов включают жевательный табак и нюхательный табак. В определенных устройствах, образующих аэрозоль, вместо сгорания, табачная композиция или другой материал, образующий аэрозоль, нагревают с помощью одного или более электрических нагревательных элементов с получением аэрозоля. В другом типе нагреваемого устройства, образующего аэрозоль, аэрозоль получают путем перемещения тепла от горючего топливного элемента или источника тепла к физически отделенному материалу, образующему аэрозоль, который может быть расположен внутри, вокруг или ниже по потоку относительно источника тепла. Бездымные табачные продукты и различные табак-содержащие материалы, образующие аэрозоль, могут содержать табак в любом виде, в том числе в виде высушенных частиц, кусков, гранул, порошков или суспензии, нанесенной на, смешанной с, окруженной или иным образом комбинированной с другими ингредиентами в любом формате, таком как хлопья, пленки, таблетки, пены или шарики. Используемый в данном документе термин «дым» используют для описания типа аэрозоля, который получен с помощью курительных изделий, таких как сигареты, или при сжигании материала, образующего аэрозоль.
В одном варианте осуществления также предусмотрен растительный материал, подвергнутый сушке, из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Способы сушки зеленых табачных листьев известны специалистам в данной области и включают без ограничения воздушную сушку, огневую сушку, трубоогневую сушку и солнечную сушку, как описано в данном документе.
В другом варианте осуществления описаны табачные продукты, в том числе табаксодержащие материалы, образующие аэрозоль, содержащие растительный материал, такой как листья, предпочтительно листья, подвергнутые сушке, из мутантных растений табака, трансгенных растений табака или не встречающихся в природе растений табака, описанных в данном документе. Табачные продукты, описанные в данном документе, могут быть смешанными табачными изделиями, которые могут дополнительно содержать немодифицированный табак.
7. Продукты и способы возделывания культур и сельского хозяйства
Для мутантных, не встречающихся в природе или трансгенных растений могут существовать другие варианты применения в, например, сельском хозяйстве. Например, мутантные, не встречающиеся в природе или трансгенные растения, описанные в данном документе, можно применять для изготовления корма для животных и продуктов питания для человека.
В настоящем изобретении также предусмотрены способы получения семян, включающие культивирование мутантного растения, не встречающегося в природе растения или трансгенного растения, описанных в данном документе, и сбор семян культивируемых растений. Семена растений, описанных в данном документе, можно кондиционировать и упаковать в упаковочный материал с помощью средств, известных в данной области техники, с получением промышленного изделия. Упаковочный материал, такой как бумага или ткань, хорошо известен в данной области техники. Упаковка семян может иметь этикетку, например маркировку или этикетку, прикрепленную к упаковочному материалу, этикетку, напечатанную на упаковке, которая описывает происхождение содержащихся в ней семян.
Композиции, способы и наборы для генотипирования растений с целью идентификации, отбора или селекции могут включать средства для выявления присутствия полинуклеотида NtSUS (или любой их комбинации, описанной в данном документе) в образце полинуклеотида. Соответственно, описана композиция, содержащая один или более праймеров для специфической амплификации по меньшей мере части одного или более из полинуклеотидов NtSUS, и необязательно один или более зондов, и необязательно один или более реагентов для проведения амплификации или выявления.
Соответственно, раскрыты специфичные в отношении гена олигонуклеотидные праймеры или зонды, содержащие приблизительно 10 или более смежных полинуклеотидов, соответствующих полинуклеотиду(полинуклеотидам) NtSUS, описанному(описанным) в данном документе. Указанные праймеры или зонды могут содержать или состоять из приблизительно 15, 20, 25, 30, 40, 45 или 50 или более смежных полинуклеотидов, которые гибридизируются (например, специфически гибридизируются) с полинуклеотидом (полинуклеотидами) NtSUS, описанным (описанными) в данном документе. В некоторых вариантах осуществления праймеры или зонды могут содержать или состоять из приблизительно 10-50 смежных нуклеотидов, приблизительно 10-40 смежных нуклеотидов, приблизительно 10-30 смежных нуклеотидов или приблизительно 15-30 смежных нуклеотидов, которые можно применять в зависимых от последовательности способах идентификации гена (например, Саузерн-гибридизации), или выделения (например, гибридизации in situ бактериальных колоний или бляшек бактериофага), или обнаружения гена (например, в качестве одного или более праймеров амплификации при амплификации или обнаружении). Один или более специфических праймеров или зондов можно разработать и применять для амплификации и обнаружения части или всего полинуклеотида (полинуклеотидов). В качестве конкретного примера, два праймера можно применять в протоколе ПЦР для амплификации полинуклеотидного фрагмента. ПЦР можно также проводить с применением одного праймера, который получен из последовательности полинуклеотида, и второго праймера, который гибридизируется с последовательностью выше или ниже последовательности полинуклеотида, такой как последовательность промотора, 3'-конец мРНК-предшественника или последовательность, полученная из вектора. Примеры методик с повышением температуры и изотермических методик, применимых для амплификации полинуклеотидов in vitro, хорошо известны из уровня техники. Образец может происходить или его можно получить из растения, клетки растения, или растительного материала, или табачного продукта, изготовленных из растения, клетки растения или растительного материала, как описано в данном документе.
В дополнительном аспекте также представлен способ выявления в образце полинуклеотида (полинуклеотидов) NtSUS, описанного(-ых) в данном документе (или любой их комбинации, описанной в данном документе), включающий стадию: (a) получения образца, содержащего или предположительно содержащего полинуклеотид; (b) приведения указанного образца в контакт с одним или более праймерами или одним или более зондами для специфического выявления по меньшей мере части полинуклеотида(-ов) NtSUS; и (c) выявления присутствия продукта амплификации, где присутствие продукта амплификации свидетельствует о присутствии полинуклеотида (полинуклеотидов) NtSUS в образце. В дополнительном аспекте также представлено применение одного или более праймеров или зондов для специфического выявления по меньшей мере части полинуклеотида (полинуклеотидов) NtSUS. Также представлены наборы для выявления по меньшей мере части полинуклеотида(полинуклеотидов) NtSUS, которые содержат один или более праймеров или зондов для специфического выявления по меньшей мере части полинуклеотида (полинуклеотидов) NtSUS. Набор может содержать реагенты для амплификации полинуклеотида, например реагенты для ПЦР, или реагенты для технологии обнаружения с применением гибридизационного зонда, такой как Саузерн-блоттинг, нозерн-блоттинг, гибридизация in situ или микрочип. Набор может содержать реагенты для технологии обнаружения связывания антител, такой как вестерн-блоттинг, ELISA, SELDI-масс-спектрометрия или тест-полоски. Набор может содержать реагенты для секвенирования ДНК. Набор может содержать реагенты и инструкции по применению набора.
В некоторых вариантах осуществления набор может содержать инструкции для одного или более описанных способов. Описанные наборы могут быть применимы для определения генетической идентичности, филогенетических исследований, генотипирования, гаплотипирования, анализа родословной или селекции растений, в частности, с количественной оценкой кодоминантных признаков.
В настоящем изобретении также представлен способ генотипирования растения, клетки растения или растительного материала, содержащих полинуклеотид NtSUS, описанный в данном документе. Генотипирование обеспечивает средства различения гомологов пары хромосом и может быть использовано для различения сегрегантов в популяции растений. Способы на основе применения молекулярных маркеров можно применять для филогенетических исследований, характеризующих генетические связи между разновидностями сельскохозяйственных культур, выявления гибридов или соматических гибридов, локализации хромосомных сегментов, влияющих на моногенные признаки, клонирования на основе генетических карт и изучения количественного наследования. В определенном способе генотипирования может использоваться любое количество аналитических методик с молекулярным маркером, включая такие на основе полиморфизма длин амплифицированных фрагментов (AFLP). AFLP является результатом аллельных различий между амплифицированными фрагментами, вызванных изменчивостью полинуклеотида. Таким образом, в настоящем изобретении дополнительно предложены способы отслеживания сегрегации одного или более генов или полинуклеотидов, а также хромосомных последовательностей, генетически связанных с этими генами или полинуклеотидами, с применением таких методик, как анализ AFLP.
Настоящее изобретение дополнительно описано в примерах ниже, которые представлены для более подробного описания настоящего изобретения. Эти примеры, в которых изложен предпочтительный режим, предусмотренный в настоящее время для осуществления настоящего изобретения, предназначены для иллюстрации, а не для ограничения настоящего изобретения.
ПРИМЕРЫ
Пример 1. Идентификация ключевых генов метаболизма сахарозы после сушки в листе табака Берлей, Вирджиния и восточного типа
Для идентификации ключевых функций, вносящих вклад в метаболизм сахарозы во время ранних стадий сушки листа табака Берлей, Вирджиния и восточного типа, проводили анализ преобладания в отношении функции генов, экспрессия которых повышена в подвергнутых сушке листьях после 48 часов сушки в сравнении со спелыми листьями в момент сбора урожая (log2-кратное изменение >2, скорректированное p-значение <0,05) табака Берлей, Вирджиния и восточного типа. Идентифицировали гены, задействованные в выработке редуцирующих сахаров и являющиеся активными после 48 часов сушки независимо от типов сушки и сортов табака. Идентифицировали гены табака, задействованные в выработке редуцирующих сахаров.
Ключевые гены, непосредственно задействованные в выработке редуцирующих сахаров во время ранних стадий сушки в листьях, относятся к семейству генов SUS. SUS, вероятно, представляет собой ключевой фермент для запуска накопления редуцирующих сахаров в подвергнутых сушке отделенных листьях.
Обнаружено, что геном табака имеет 12 продуктов гена NtSUS, распределенных по 6 семействам, с одной копией S и одной копией T от каждого предка: NtSUS1-S (SEQ ID NO: 1), NtSUS1-T (SEQ ID NO: 3), NtSUS2-S (SEQ ID NO: 5), NtSUS2-T (SEQ ID NO: 7), NtSUS3-S (SEQ ID NO: 9), NtSUS3-T (SEQ ID NO: 11), NtSUS4-S (SEQ ID NO: 13), NtSUS4-T (SEQ ID NO: 15), NtSUS5-S (SEQ ID NO: 17), NtSUS5-T (SEQ ID NO: 19), NtSUS6-S (SEQ ID NO: 21) и NtSUS6-T (SEQ ID NO: 23).
Транскрипты SUS происходили из геномных последовательностей NtSUS2-S (SEQ ID NO: 5), NtSUS3-S (SEQ ID NO: 9), NtSUS3-T (SEQ ID NO: 11) и NtSUS4-S (SEQ ID NO: 13). Экспрессия данных генов повышалась во время сушки листа (старения), как показано в таблице 1. Это подтверждает, что копии S, в частности, задействованы в химической модификации листьев, подвергаемых ранним стадиям сушки, и в данном конкретном случае, в повышении уровня глюкозы и фруктозы.
Хотя в подвергнутых сушке листьях табака Берлей обнаружили низкие значения уровней редуцирующих сахаров по сравнению с табаком Вирджиния и восточного типа (см. фигуру 1), тем не менее гены NtSUS являются активированными в табаке Берлей (см. таблицу 1), вероятно, как конститутивный ответ для обеспечения, в свою очередь, доступного источника углерода для синтеза аминокислот во время ранней фазы сушки.
Как в табаке Берлей (BU), так и в табаке Вирджиния (FC) NtSUS1-S и NtSUS1-T, которые не экспрессируются во время ранних стадий сушки (см. таблицу 1), в частности, экспрессируются в корне и стебле, что свидетельствует о возможной специфической функции в этих тканях, заключающейся в доставки углеводов для синтеза клеточных стенок или снабжения ресурсами углерода в условиях аноксии. С другой стороны, NtSUS3-S, NtSUS3-T, NtSUS4-S, которые индуцируются во время ранних стадий сушки листа, также экспрессируются во всех органах, тогда как NtSUS2-S и NtSUS2-T экспрессируются преимущественно в незрелых цветках и лепестках. NtSUS5-S, NtSUS5-T, NtSUS6-S и NtSUS6-T экспрессируются на низких уровнях во всех проанализированных тканях растений (см. таблицу 2).
Для увеличения пула редуцирующих сахаров в подвергнутых сушке листьях могла бы рассматриваться сверхэкспрессия NtSUS2-S, NtSUS3-S, NtSUS3-T и/или NtSUS4-S с применением индуцируемого старением промотора, такого как SAG12 или E4 (применение конститутивного промотора может существенно изменить метаболизм растения). С другой стороны, осуществление нокаута NtSUS2-S, NtSUS3-S, NtSUS3-T и/или NtSUS4-S может внести вклад в снижение содержания редуцирующих сахаров в подвергнутых сушке листьях.
Пример 2 - Сайленсинг экспрессии NtSUS в листе табака Вирджиния
Сайленсинг NtSUS исследовали для определения того, вносят ли эти гены вклад в снижение содержания редуцирующих сахаров в подвергнутых сушке листьях табака Вирджиния. Специфический фрагмент ДНК в пределах кодирующей последовательности обоих NtSUS клонировали с сильным промотором вируса мозаики ночной красавицы (MMV) в вектор GATEWAY. Фрагмент гена NtSUS фланкирован между MMV и 3'-терминаторной последовательностью nos гена нопалинсинтазы Agrobacterium tumefaciens.
Для обеспечения отбора растений с низким содержанием редуцирующих сахаров листья отдельных растений T0 и соответствующих контрольных линий анализировали после 60 ч сушки для определения влияния на уменьшение содержания редуцирующих сахаров. Отбирали самые лучшие линии T0, демонстрирующие самый низкий уровень редуцирующих сахаров. Семена собирали от этих самых лучших линий T0. Потомство T1 анализировали с помощью qPCR для определения эффективности событий сайленсинга NtSUS в аспекте снижения содержания редуцирующих сахаров.
Манипулирование с генами NtSUS (например, либо с применением конститутивного промотора, либо со специфического индуцируемого старением промотора, такого как SAG12 или E4) может изменять химию подвергнутых сушке листьев табака. Аналогичным образом осуществление нокаута генов NtSUS с применением стратегии редактирования гена, такой как CRISPR-Cas или отбор мутантов, может изменять химический состав аминокислот в листьях основных сортов коммерческого табака.
Любая публикация, цитируемая или описанная в данном документе, предоставляет соответствующую информацию, раскрытую до даты подачи настоящей заявки. Заявления, сделанные в данном документе, не должны истолковываться как признание того, что авторы настоящего изобретения не имеют оснований для его противопоставления как более ранним таким раскрытиям. Все публикации, упомянутые в вышеприведенном описании, включены в данный документ с помощью ссылки. Различные модификации и варианты настоящего изобретения будут очевидны специалистам в данной области без отступления от объема и сути настоящего изобретения. Несмотря на то, что настоящее изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что настоящее изобретение, как заявлено, не должно быть неправомерно ограничено такими конкретными вариантами осуществления. В действительности различные модификации описанных способов осуществления изобретения, которые очевидны специалистам в клеточной, молекулярной биологии и биологии растений или в смежных областях, предназначены находиться в пределах объема прилагаемой формулы изобретения.
ТАБЛИЦА 1
Экспрессия генов NtSUS во время ранних стадий сушки для табака Берлей (BU), Вирджиния (FC) и восточного типа (OR)
ТАБЛИЦА 2
Экспрессия генов NtSUS в корне, стебле, листе, расположенном в средней части стебля, незрелом цветке (незр. цветок), чашелистике и лепестке растений табака Берлей (BU) и Вирджиния (FC), выращенных в поле
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1: Полинуклеотидная последовательность NtSUS1-S
atggcagctagtggtcttagcattaagaaaagtttggaggaatccattttggctcatccagatgaaattttggctctcaagtcaaggtacattactacatataatgatattaagaactagaggcttatccaaggttttgttacatttttgaaattataagtttagaacctaatagtacttggtagcacttgtttccttattatctagctgttgttactgcttgttgctactgctttctgttcatctttccttgagcccggtctatcggaaacaacctctctattctcaaagtataaggtttgcgtacatactacctccccagactctacttgtggaatttactgtttttgttgtgttgttgtaatctaatatttattagaattttactgatttttcacatatatatatctatgtcccctgtcgaaaattctatagctcatgttagctaaatacattagtaccattgtttttaattgttttggttttggcacaggattgaaactgaagggaaaggggtaatgaaaccacttgatctcttgaaccatttggtttctgttactagtaagacaaatggagtaaatattgtacctagtgcacttgtggaagttctcagttgcagccaagaagctgtgattgtaccaccaaaactagcactagctgtacgtccgaggcccggtgtatgggagtacttgtcactgaatcttaagacaaagaaagtggctgaattaagcattcctgaataccttcaattgaaagagaacactgttgatgaaaggtaaagtattagtctgcgatttcgctttgtgaaattgaagtttttgttttgattcataatgttttgtgtatcaattatgttaccagtggaaacatattggagttggattttgagccatttacaacagttacaccaccaaaaacactttctgactctattggcaatggtttggagtttcttaatcgccacattgcttcgaaaatgtttcatgataaggagatttccagatgcctccttgacttcctcagaaaccataactacaaaggaaaggtaataaaaaaaagtgtttctttaaacaagttgtatgattatgtgtatatttctaagtatgttaacttgaaaacagtcattgatggtgaaagaaagcattcaaagcctagagagtttccaacttgttctgaaaaaagcagaggaacatttgtgcacattgaatccagaaactccatactccaattttgaatcaaagtttgaagagattggcttggaaagagggtggggaaacaccgctgaacgcgtgcaagacactatcagtcatcttttgcatctccttgaggctcctaacgcgtcttctttggaaaatttccttggtagaatcccattggttttcaatgttgtgattctaactccacatggttattttgctcaagataatgtcttgggctatcctgacactggtggccaggtttgtgtccaatattttgcattcttgatcaagttctttataccatttgaaccaacaatcttnaacattctttttttggttgtgaaatgttgaataggttgtttacattcttgatcaagttccagctatggagcgtgagatgcttcatcgtatgaagcttcaaggactcgatgatatcatccctcgcatccttgttgtaagtggccttaattttcctagtttcatttacacctctaaatgaaattgatcttttttgttgttttatatcaggtaacaaggctgctgcctgatgcagtaggaaccacctgtggcgagcggatggagaaagtatatggggcagaacattctcatataattcgtgttccatttagaactgagaagggaatgttgcgcaaatggatctcacgattcgaagtctggccatacatggaaactttcactgaggttggaacataaaaacaaataaaatccattggaatgttccttctgcaattgaaaatgtcttgctaactgaagacccatttttaaattgatcatcaggatgttgcagaagaacttgtcaaagaattgcaagctaaaccagacttgatcattggaaactacagtgagggaaatcttgctgcctctttgcttgcgaagaaatttggggctactcagtgtactattgctcatgccttggaaaaaactaagtatccaaactctgaccttaattggaagaagtttgatgacaagtatcatttctcaagtcagttcactgctgatctctttgccatgaatcacactgatttcatcatcaccagcactttccaagaaattgctggaaggtaaaagcaaatgcacaccatcatagtatttcatatttttacccttgtttatactatttccattcaccgaccccgacttgtttaggattgagccatagttgttgttgttgtttgtttatactatttccatttgccgaccacaacttgtttaggactgaggtatagttgttgttgttggtttgttcatattattttcattcgctaaccctaacttgtttgggactgaggcatagtagtagtagtagttgttgctattagtttatactatttccatttgccaaccccaacttgtttggtactgagacatagttgttgttgttgttgtttgtttatactatttccatttgccgaccccaacttgtttaggactgaggtatagttgttgttgttggtttgttcatattattttcattcgctaaccccaacttgtttgggactgaggcatagtagtagtagtagtagttgttgctattagtttatactatttccatttgccaaccccaacttgtttggtactgagacatagttgttgttgttgtttgtttatactatttcaatttgtcgaccccaatttgtttgggaccaaggcatggttgttgttgttgtttgtttgtttttactgtttccattgatattggaacatttgttatttgcagcaaaaacactgtaggacagtatgagagtcatactgcttttaccatgcctggattgtaccgagtagtccatggaatcgattcgtttgatccaaagttcaacattgtctcccctggggctgatatgtcaatctacttcccttacactgagaaggagaaaaggctaaccaacttccacccggaaattgaagaactcctctacagtcctgttgagaataaggaccacttgttagtctccttaatttgcttttatttcatcccatttatgatcgcttttatcccaacagatcgattaatcatttgttatcaacataaacagatgtgtgttgaaggaccggaacaagccaattctctttaccatggcaaggctagatcgcgtgaagaatctaacagggctcgtggaatggtatgctaagaatgcaaggctgagggagcttgttaaccttgtggttgtaggcggagacagaaggaaagaatccaaagatttagaagagcaagcagagatgaagaagatgtatgatcttatcgaaacctataacctgaacggccaattcaggtggatttcttcccaaatgaatcgtgtgaggaacggagaactctatcgttacattgcagacacgaggggtgctttcgttcaaccagcattctacgaggcttttggtttgacagttgtagagtctatgacttgtggtttgccaacttttgctacttgtaatggtggaccatttgagattatagtgaatggaaaatctggtttccatattgatcctaatcaaggtgacaaggctgctgatatgttggtaaatttctttgaaaaatctaaagaagatccaagttattgggatgctatttccaagggaggtctgcaacgtattcttgaaaagtaagcttttgcatttgattagcacaagtgcacaaccaagatttaacttttgaacaaactaaaactaacccttttttgtattttcttttgctaggtatacatggcaaatttattcacagaaagtgatcacactatctgggatttatggattctggaagtatgcaaccaagaatgataaagttgctagtgcaaagaagcgctatcttgagatgttttatgaacttggatttaagaaatcagtaagtgtcaattttaaaggggaaccttggatcaacggttaagttgtctttgtgcaacctataggtcaggggtttgagccgtagaagtagccactaatatttacattagggtagactgtgtacatatcacaccccttggggtacggccctttcctggatcctgtatgaacgcgggatgccttgtgcaccgggctgtatttttttttttagtgtcacttctgtattttgtttgagcttgtttataaagtttggaaatctgctgctaatttgtatatttgttggttgtgtatttcaggctgagaaagttccattggctattgatgaatag
SEQ ID NO: 2: Полипептидная последовательность NtSUS1-S
MAASGLSIKKSLEESILAHPDEILALKSRIETEGKGVMKPLDLLNHLVSVTSKTNGVNIVPSALVEVLSCSQEAVIVPPKLALAVRPRPGVWEYLSLNLKTKKVAELSIPEYLQLKENTVDESGNILELDFEPFTTVTPPKTLSDSIGNGLEFLNRHIASKMFHDKEISRCLLDFLRNHNYKGKSLMVKESIQSLESFQLVLKKAEEHLCTLNPETPYSNFESKFEEIGLERGWGNTAERVQDTISHLLHLLEAPNASSLENFLGRIPLVFNVVILTPHGYFAQDNVLGYPDTGGQVVYILDQVPAMEREMLHRMKLQGLDDIIPRILVVTRLLPDAVGTTCGERMEKVYGAEHSHIIRVPFRTEKGMLRKWISRFEVWPYMETFTEDVAEELVKELQAKPDLIIGNYSEGNLAASLLAKKFGATQCTIAHALEKTKYPNSDLNWKKFDDKYHFSSQFTADLFAMNHTDFIITSTFQEIAGSKNTVGQYESHTAFTMPGLYRVVHGIDSFDPKFNIVSPGADMSIYFPYTEKEKRLTNFHPEIEELLYSPVENKDHLCVLKDRNKPILFTMARLDRVKNLTGLVEWYAKNARLRELVNLVVVGGDRRKESKDLEEQAEMKKMYDLIETYNLNGQFRWISSQMNRVRNGELYRYIADTRGAFVQPAFYEAFGLTVVESMTCGLPTFATCNGGPFEIIVNGKSGFHIDPNQGDKAADMLVNFFEKSKEDPSYWDAISKGGLQRILEKYTWQIYSQKVITLSGIYGFWKYATKNDKVASAKKRYLEMFYELGFKKSAEKVPLAIDE
SEQ ID NO: 3: Полинуклеотидная последовательность NtSUS1-T
atggcaggcagtggtcttagcattaaggaaagtttggaggaatccattttggctcatccagatgaaattttggctctcaagtcaaggtacattactgcataatgatattaagacctagaagcggatccaagattttgttacatttttgaaattataagtttagaatctaatatttgttatcgcttgtttccttattatcttgctgttgttactgcctgttgctactagtttctgttcatccttccttgagctgagtttctatcggaaacaacctctctactctcaaagtaggaataagttatgcgtacacactaccctccccagactccacttgtgtaatttactgagtttgttgttgttgttgttgtaatctaatacttgttagaattttactgatttttcacatatatatctatgacccatgtcgaaaatactatagctcatgtgctaaatacattagtaccattgttttgtaattgttttggttttggaacaggattgaaactgaagggaaaggggtaatgaaaccagttgatctcttgaaccatttggtttctgttactagtaaaacaaatggagtaaatgttgtacctagtgcacttgtggaagttctcagttgcagccaagaagctgtgattgtaccaccaaaactagcactagctgtacgtccgaggcccggtgtatgggagtacttgtcactgaatcttaagacaaagaaagtggctgaattgagcattcctgagtaccttcaattgaaagagaatactgttgatgaaaggtaaagtaatagtctgcgatttcgctttgtgaaattgaagttttttgtttgattcttaatgttttgtgtatcaattatgttaccagtggaaacatcttggagttggattttgagccatttacaactgttacaacaccaaaaacactttctgactctattggcaatggtttggagtttcttaatcgccacattgcttcgaaaatgtttcttgataaggagattgccaagtgcctccttgactttctcagaaaccataactacaaaggaaaggtagtaaaaaaagtgtttctttaaacaagttgtatgattatgtgtgtatttctaaatatgtcaatttgaaaacagtcattgatggtgaaagaaagcattcaaagcctggagagtttccaacttgttctgaaaaaagcagaggaatatttgcacacactgaatccagaaactccatactccaaatttgaatccaagtttgaagagattggcttggaaagagggtggggaaacaccgctgaacgcgtgcaagacaccattagtcatcttttgcatctccttgaggctcctaacgcgtcttccttggaaaatttccttggtagaatcccattggttttcaatgttgtgattctcaccccacatggttattttgctcaagataatgtcttgggctatcctgacactggtggccaggtttgtgtccgatataacatatcaagaaattttgcattcttgatcatgttctttataccatttgaaccaacattctttttttggttgtgaaatgttgaataggttgtttacattcttgatcaagttccagctatggagcgtgagatgcttcatcgtatgaagcttcaaggactcgacgatatcatccctcgcatccttgttgtaagtgcccttaattttcctggtttggtttacctctaaatgaaattgattttctggctttctaacttttttggattgatctttttgttgttttatatcaggtaactaggctgctgcctgatgctgtaggaaccacttgtggcgagtggatggagaaagtatatggggcagaacattctcatataattcgtgttccatttagaactgagaaaggaatgttgcgcaaatggatctcacgattcgaagtctggccatacatggaaactttcactgaggttggaacataaaaacaaataaaaatcattggaatgttcttctgcatttgaaaatgtcttgctaactaaagactcatttttaaattaatcatcaggatgttgcagaagaacttgtcaaagaattgcaagctaaaccagacttgataattggaaactacagtgagggaaatcttgctgcctcattgcttgctaagaaatttggggctactcagtgtactattgctcatgccttggaaaaaactaagtatccaaactctgaccttaattggaagaagtttgatgacaagtatcatttctcaagtcagttcactgctgatctttttgccatgaatcacactgatttcattatcaccagcactttccaagaaattgctggaaggtaaaagcaaatgcacaccatcatagtatttcatatttttaccctagtttatactatttccatttgtcaactccaacttgtttgggattgaaccatagttgttgtttgtttatactatttccattcgccgaccccaacttatttgggactgagacataattgttgttattattgtttgtttgtttatactatttccattctcagaccccaacttctttgggactgagccgtagattgttgttgttgttgttgttgttgtttgtttatgctatttccgttcaccgaccccaacttatttgggactgaggtgtagaagtagtcgttgttgtttgtttatacgacttccaattgatattcgaatgtttttatttttgcagcaagaacactgtaggacagtatgagagtcatactgcttttaccatgcctggattgtatcgagtagtccatggaatcaattcgtttgatccaaagttcaacattgtctcccctggggctgatatgtcaatctacttcccttacactgagaaggagaaaagactaaccaacttccacccggaaattgaagaactcctctacagtcctgttgagaataaggaccacttgttagtcttctttatttcattcatttttctacaccttttttttcaacagattgattgattggttcttatcaacgtaaacagatgtgtgttgaaggaccagaacaagccaattctctttaccatggcaaggctagatcgcgtgaagaatctaacagggctcgtggaatggtatgcaaagaatgcaaggctaagggagctcgttaaccttgtggttgtaggcggagacagaaggaaagaatccaaagatttagaagagcaagcagagatgaagaagatgtatgatcttatcgaaacatacaacctgaatggccaattcaggtggatttcttcccaaatgaatcgtgtgaggaacggagaactttatcgatacattgcagacacgaggggtgctttcgttcaaccagcattttatgaggcatttggtttgacagttgttgagtctatgacttgtggtttgccaacttttgctacttgtaatggtggaccatttgagattatagtgaatggaaaatctggtttccatattgatcctaatcaaggtgacaaggctgctgatatgttggttaatttcttcgaaaaatctaaagaagatccaagttattgggatactatttccaagggtggtctgcagcgtattcttgaaaagtaagcttttgcatttgattagcacaagtgtacaaccaagatttaacttatgaacaaactaaaactaaccctttttttattttcttttgctaggtatacatggcaaatttattcacagaaagtgatcacattatctgggatttatggattctggaaatatgcaaccaagaatgacaaagttgctagtgcgaagaagcgctatcttgaaatgttttatgaatttgggtttaagaaatcagtaagtgtcacttctgtattttgtttgagcttgtttgtaaagtttggcaatcttctgctaatttgtactatatttgttgacttgtgcatttcaggctgagaaagttccattggctattgatgaatag
SEQ ID NO: 4: Полипептидная последовательность NtSUS1-T
MAGSGLSIKESLEESILAHPDEILALKSRIETEGKGVMKPVDLLNHLVSVTSKTNGVNVVPSALVEVLSCSQEAVIVPPKLALAVRPRPGVWEYLSLNLKTKKVAELSIPEYLQLKENTVDESGNILELDFEPFTTVTTPKTLSDSIGNGLEFLNRHIASKMFLDKEIAKCLLDFLRNHNYKGKSLMVKESIQSLESFQLVLKKAEEYLHTLNPETPYSKFESKFEEIGLERGWGNTAERVQDTISHLLHLLEAPNASSLENFLGRIPLVFNVVILTPHGYFAQDNVLGYPDTGGQVVYILDQVPAMEREMLHRMKLQGLDDIIPRILVVTRLLPDAVGTTCGEWMEKVYGAEHSHIIRVPFRTEKGMLRKWISRFEVWPYMETFTEDVAEELVKELQAKPDLIIGNYSEGNLAASLLAKKFGATQCTIAHALEKTKYPNSDLNWKKFDDKYHFSSQFTADLFAMNHTDFIITSTFQEIAGSKNTVGQYESHTAFTMPGLYRVVHGINSFDPKFNIVSPGADMSIYFPYTEKEKRLTNFHPEIEELLYSPVENKDHLCVLKDQNKPILFTMARLDRVKNLTGLVEWYAKNARLRELVNLVVVGGDRRKESKDLEEQAEMKKMYDLIETYNLNGQFRWISSQMNRVRNGELYRYIADTRGAFVQPAFYEAFGLTVVESMTCGLPTFATCNGGPFEIIVNGKSGFHIDPNQGDKAADMLVNFFEKSKEDPSYWDTISKGGLQRILEKYTWQIYSQKVITLSGIYGFWKYATKNDKVASAKKRYLEMFYEFGFKKSAEKVPLAIDE
SEQ ID NO: 5: Полинуклеотидная последовательность NtSUS2-2
atggctgaacgtgctctgactcgtgttcacagccttcgtgaacgtcttgatgccactttggctgcacatcgcaatgagatattgctgtttctttcaaggtattgcctaagtagtgttcttgtttcctacaaaagattcagttggtgttcaaaaaacgatatgtgatttgatttatctgcctaagtcttggtagtcataattatccggtacctgtgctggtgcgagttagctggttcggaaactactcttatgaaaacgagagatttagttggtgttgtctgcaattctgtagtatggactattaagcagatagatcatgtttgatatcgaaaaggaatgtatatgtgatgttacttgaactggttttggttattacaggattgaaagccatggaaaagggatcttgaaacctcaccagctattggctgagttcgatgcaattcgccaagatgacaaaaagaagctgaatgatcatgcatttgaagaactcctgaaatctactcaggtaattttgattttggctaaatgtgttaccaagctgaatgatcatgcatttgagtttgtgtccgactactacaatgatatgttataccaggaagcgattgttctgccaccttgggttgcacttgccattcgtttgaggcctggtgtgtgggaatatgtccgtgtgaatgttaatgctctagtcgttgaggagctgaccgtccctgagtatttgcattttaaggaagaacttgttgatggaacgtaagttttagtctcttatttgatactatgttagagaataggcagtggattcaatttatcagtgttgttttttacctaatgcagctccaatggaaatttcgttctcgagttggattttgagcccttcactgcatcctttcctaaaccgaccctcaccaaatctattgggaatggagttgaattcctcaataggcacctttctgcgaaaatgttccatgacaaggaaagcatgaccccgcttcttgaatttcttcgggttcacaattataagggcaaggtaactttgttattcccattcatatatatgttcagtttgtgcttatcatgcgcccaatgatgtatgaatatgtactaaaggatagatgtacgatttcgtttgcagacaatgatgctgaatgacagaatacagaatttaaccactctgcaaaatgtcctaaggaaggcagaggaataccttattatgcttccccctgaaactccattttccgaattcgaacacaagttccaagaaattggattggagaagggatggggcgacactgcggagcgcgtgctagagatgatatgcatgcttcttgatctacttgaggctcccgactcctgtactcttgagaagttcctagggagaattcctatggtgttcaacgtggttatcctttccccccatggatatttcgcccaggaaaatgtcttgggttatcccgacactggtggccaggtgcattactttagtctttgtccgtgagtctatgttgctcagatcctctacaatgccactgtacccgtgtaggatactccaaatataatgcatttttggaggatctgtcaccggtgcaatggcattttggaggtcggagcaacaaacaactgctagtatgcttctaaagcttgcttccataaatgctaaggtccttcacccgtaatgtgcaggttgtctacatattagatcaagttccagccttggagcgtgaaatgcttaaacgcctaaaggagcaaggacttgatataacaccgcgtattcttattgttagtatttcttgtacttgtaattgctgcggattacacaaaattttctctttattggcaacttatcttgatattattcccaggttactcgtctgctgcctgatgcagttggaacaacttgtggtcagcggcttgagaaggtgtatggagccgagcactcacatattcttagggtcccctttaggaccgagaagggcattgttcgcaaatggatatctcgctttgaagtgtggccatacatggagactttcactgaggtgacactaagcttccttgtatttgtctatcttctaattggtattaggaacaatttgctaattattaacgctttggcttttcgtacatcaggatgttgcaaaagaacttgctgcagaactgcaggccaagccagatttgataattggcaactatagcgagggaaatcttgtggcttcattgctggctcacaagttaggcgtaacgcaggtctgtgttatttttcacctcttataaatctgattgtatttccattagtctggaactaaaagtactaaaattttcttttcttcgctgtgttatttgccttctgcagtgcaccattgcccatgcattggagaaaacaaagtatcctgattctgacatctactggaaaaaatttgacgaaaaataccatttctcgtcccagtttaccgctgatcttattgcaatgaatcacaccgattttatcatcaccagcactttccaggagatagcaggaaggtataacatcaattgctaattcggttgcagtaacattttgttcgatttcttccccttatgcttaacctaataccctaatgaattttccagcaaggacactgtcggacagtacgagagtcaccaggcattcacaatgcctggattgtacagagtcgttcacggcattgatgtgttcgatcccaaattcaacattgtctcacctggagctgatataaacctgtatttcccatattccgagaaggaaaagagattgacagcacttcacccagaaattgaggagcttctgtacagtgatgttgagaacgaggaacatctgtaagtttctaacttactcgtaccgtcagtggcagagccagaattttcattaaaatggggtcaaaatataaagacataaattcacaaagaagccaaggggtgtcaatatgtagtataaatatattaaaaaaattacctagctacacaatgtaattttccgacaaaggggtatcggttgcacttcttgaatacatgtggctctgccactgggtacagttacaaagtcctgttacctatgtagatgagcttgtgctgaacatgttgtgattttggtaggtgtgtgctaaaggacaggaataagccaatcttattcacaatggcgagattggatcgtgtgaagaacttaaccggacttgttgagtggtacgccaagaacgcacggctaagggagttggttaaccttgttgtcgttggtggagaccgaaggaaggaatccaaagatttggaagagcaagcagagatgaagaagatgtatgagctaataaagactcacaacttaaatggccaattcagatggatttcttcacagatgaaccgagtaaggaacggcgaactctaccgatacattgccgacactaggggagctttcgtgcagcctgcattctatgaggctttcggtttgactgttgttgaggccatgacctgtggtttgcctacatttgcaactaatcatggcggtccagctgagatcatcgttaacggaaaatccggcttccatatcgatccatatcacggtgagcaagctgctgatctgctagctgatttctttgagaaatgtaagacggaaccttctcattgggaaactatttcaaccggtggcctgaagcgcatccaagagaagtaagcaactctttcttgactctagtcattcaaattaacttgggatttgaggcatagttgattgataatttatcgcgtctctactactatatacaggtacacgtggcaaatctactcggagagattattgacgttggctgctgtttacggtttctggaaacatgtttctaagcttgatcgtctagaaatccgtcgatatctagaaatgttttatgctctcaaataccggaagatggtgagttcttctgcttcctgctcttctcatagtgtttaatatacacttgattgattgcattcacttagactaagttgctcggacacgggtgtggatgtccgacacgagtgcggatctagagttcagatccttcaagatgtaaattataagattcggggatatggatcctagtacggatacgggtgcgagaatccggctaaaaataattttaaaaaaaattatctctaaattatgagatattatgtggaatacttacgtataacttgtaaagtgtagattttttttaattctcaagttgtagattagtaaatgattgatttcctagataagtatgctattttcttcaaatttactcttctgatttcgaaaatcaaattgtatctcgtctcgaatttttccgtccgttatggtcaaagtacccaaaatcgtttgaccaaatcggtacggatcccatacccacacccacactagtgtcgtattgacacgggtgccgcacctaaactgctatgtcggagcaacttagcacttagagaatcattgatgttaaattttcttaattcttgaatctgctaatgaagattttatcttggtttttgtttaggctgaagctgttccattggctgctgaatga
SEQ ID NO: 6: Полипептидная последовательность NtSUS2-2
MAERALTRVHSLRERLDATLAAHRNEILLFLSRIESHGKGILKPHQLLAEFDAIRQDDKKKLNDHAFEELLKSTQEAIVLPPWVALAIRLRPGVWEYVRVNVNALVVEELTVPEYLHFKEELVDGTSNGNFVLELDFEPFTASFPKPTLTKSIGNGVEFLNRHLSAKMFHDKESMTPLLEFLRVHNYKGKTMMLNDRIQNLTTLQNVLRKAEEYLIMLPPETPFSEFEHKFQEIGLEKGWGDTAERVLEMICMLLDLLEAPDSCTLEKFLGRIPMVFNVVILSPHGYFAQENVLGYPDTGGQVVYILDQVPALEREMLKRLKEQGLDITPRILIVTRLLPDAVGTTCGQRLEKVYGAEHSHILRVPFRTEKGIVRKWISRFEVWPYMETFTEDVAKELAAELQAKPDLIIGNYSEGNLVASLLAHKLGVTQCTIAHALEKTKYPDSDIYWKKFDEKYHFSSQFTADLIAMNHTDFIITSTFQEIAGSKDTVGQYESHQAFTMPGLYRVVHGIDVFDPKFNIVSPGADINLYFPYSEKEKRLTALHPEIEELLYSDVENEEHLCVLKDRNKPILFTMARLDRVKNLTGLVEWYAKNARLRELVNLVVVGGDRRKESKDLEEQAEMKKMYELIKTHNLNGQFRWISSQMNRVRNGELYRYIADTRGAFVQPAFYEAFGLTVVEAMTCGLPTFATNHGGPAEIIVNGKSGFHIDPYHGEQAADLLADFFEKCKTEPSHWETISTGGLKRIQEKYTWQIYSERLLTLAAVYGFWKHVSKLDRLEIRRYLEMFYALKYRKMAEAVPLAAE
SEQ ID NO: 7: Полинуклеотидная последовательность NtSUS2-T
atgctttttatgggagtaaattttatggccggtcattcaactttgtgttcattacgcaaaagtcatttttcttggtgtttattacgcaagtcatttttcttttttttttgttacgtaaaaatcattcaactatgtgtttattatctaaaattcaattttttttttccttttgttacacaaaaatcattttactttactctatttatcacaaaagtcaccttggccagattttataataggcttttatcttttgttacacaaaaattattttactttactctatttatcacaaaagtcaccttggccagattttataataggcttttatcttttgttacacaaaaattattttactttactctatttatcacaaaagtcaccttggccagattttacaatacttttaccttaaaagactattatgcccttgacattataaatcctctcatttatataataccttctatatgatacactatataatatatttttacctaggtattttacttataattaaaataatattaaattattttatttatctattttataatatattcatacatttaattttttcatggcaaatcactttgtttaatcatatttaaacatgaacaaattttaaatatcaaaaaaataaaaaaataaaaaaaatatttatttgaaataataacaaacagatttgtttaacaaatgatagtttttttttatagtcaataaaatttttaaaaaaattcaaagatatttgtttttaatattaatatttttaaagctttatctgttaatattatttatttgaaagtattaatctgatgtgtcattgtgttaaatgtgagtattttatttattggattaatgagtatggcttggctgataaaaagctttgattttataattttcattaaaaatattttattaagctagtacctgacaaatttaatatcttgaaaattaacgttaagaaaaaattaaatataaaaatatattataaaaataataaataaataatatcaagttattttaattataaataaaatacatggttaaaaatatattatatagcatataatatagaaggtattacataaatgagatgatttaaagggcataatagacttttcaggtgaatgatttgtaaaatatggttaaagtgattattgtgataattagagcatagtaaaataatttttatgtaacaaaagaaaaaaaaaatgacttttgggtaatgaacataaatttgaataacttttacgtaacaaaagaataaaataaattttggataataaacataaaattgaatgaccacctataaaatttattatttttttgggctcttcttgatttgattttttagtttagcctttgcagtaatcttggttgtcacgcgtagcgttgtgctttcgccacataagtatttagtagacttaattaatgtcattatatcggttggtgtggttttaattacttaactgtactattatattaggtggaaggtttgaaaatttatagtagtaacattctagatcattgaaaatattggtgtttcagtgactttttagtatgtcattttcattttctaagtggttgtactaatatagtatattaaaattttgattggttgagaaacaatctctctcacctacacggtacgggtaaggtatgcgtatacgcttatcctccctacactccatttgtgggactattgttgttattttggataagctgaggtatccatcttctactaactgcactagtttattttttttgctgtttacagttgaaacaattgtctgaggatttctcacctgctgaatcaactgcaatggctgaacgtgtgctgactcgtgttcacagccttcgtgaacgtcttgatgctactttggctgctcatcgcaatgagatattactgtttctttcaaggtatagccaaagatagtattcttgttaactaaaaaagattcagttggtgttcaaaaaacgatacgtttatctgcctaagtcttggtagtcagaattatccggtacctatgctggtgtgagttagctggctaggaaaccactcttatgaaaacaagagatttagttagagttgtctgtaattctgtagtatggactatgtatgtgatgctatttgaactggttttggttattataggattgaaagccatggaaaagggatcttgaaaccgcatcagctattggctgagtttgatgcaattcgccaagatgacaaaaagaaactgaatgatcatgcatttgaagaactcctgaagtccactcaggtaatatggttttggctatatttgtcgccaacgccaagctcatatttttatattattttgagcttgtgtctgaatacgacgatgatatgttatactaggaagcaattgttctgccaccttgggttgcacttgcgattcgtttgaggcctggtgtgtgggaatatgtccgtgtgaatgtcaatgcgctagtcgttgaggagctgactgtccctgagtatttgcatttcaaggaagaacttgtcgatggaacgtaagtgttagtcttcaatttgatgctatgttagagaataggctgtggaatttattgatcaatgctgtgctttgtcctgatacagctccaatggaaatttcgttctcgagttggattttgagcccttcaccgcatcctttcctaaaccaaccctcaccaaatctatcggaaatggagttgaattcctcaataggcacctctctgcgaaaatgttccatgacaaggaaagcatgaccccgcttcttgaatttcttcgggttcacaattataagggcaaggtgacttgctatttccatttatctataggttcggtttgtgcttatcatgcgcccaatgacatatgaatatgcgctaaaggatagatatatgatttcctttgcagacaatgatgctgaacgacagaatacagaatttaaccacactgcaaaatgtcctaaggaaggcagaggaatacctcattatgcttccccctgaaactccattttccgaattcgaacacaagttccaagaaattggattggagaagggatggggcgacactgcagagcgcgtgctggagatgatatgcatgcttcttgatctcctcgaggctcccgattcctgtactcttgagaagttcttggggagaattcctatggtgttcaatgtggttatcctttccccccacggatatttcgcccaggaaaatgtcttgggttatcccgacactggtggccaggtgcattactttaatctttatccgtgagtctatgtttgttcgaatcctctagaaatgtcactgtacctatgtaggatactccaaatataatgcattttggggggatctgttatgggtgcgatggcatttttggaggtcggagcaacaaacaattgctatgtattcttctaaagcttgctttcataaatgctaaggtccttcacccttaatgtgcaggttgtctatatattagatcaagttccagccttggagcgtgaaatgcttaagcgcctaaaggagcaaggacttgatatcacaccgcgtattcttattgttagtatttcctgtacttgtaattactgcggattacacaaaatttcctttttatcttcttaacaacttatcttgatggtattcccaggttactcgtctgctacctgatgcagttggaacgacttgtggtcagcggcttgagaaggtgtatggagccgagcactcacatattctgagggtcccctttaggactgagaagggcattgttcgtaaatggatctctcgctttgaagtgtggccatatatggagactttcactgaggtgacactaaaacttccttatatttgtctatcttctaattggtattaggaataatttgttaattgttaactctttgtcttttcgtacatcaggatgtcgcaaaagaacttgctgcagaattgcaggccaagccagatttgataataggcaactatagcgagggaaatcttgtggcttcattgctcgctcataagttaggcgtaacacaggtctgtgttgtttttcactctcttaaagatctgattgcatttccattagtctggaactagaagtactaaaaagttcttttcttcactgtgttatttgccgtcggcagtgcaccatagctcatgcattggagaaaacaaagtatcctgattctgacatctactggaaaaaattcgatgaaaaataccatttctcgtcccagtttaccgctgatcttattgcaatgaatcacaccgattttatcatcaccagcactttccaggagatagcaggaaggtataacatcaatttgctacttcgactgcaacagcattgtgttcccatttctttcccttatgcttaacctaataccgtcatgaattttccagcaaggacactgtcggacagtacgagagtcatcaggcattcacaatgcccggattgtacagagttgttcacggcattgatgtgttcgaccccaaattcaacattgtctcacctggagctgacataaacctctatttcccatattccgagaaggaaaagagactgacagcacttcaccctgaaatcgaggagctgctgtacagtgacattgagaacgaggaacatctgtaagtttctaccttactcgtacagtcagtggcggagccagaattttcactaaaataaggtcaaaatataaagacataaatccacaaagaagccaagggtgtcaatatatagtataaatacattaaaaaaattacctatctacacagtgtaattttccgacaaaggggtgtcggttgacactccttgaatacatgtggctctgccactgggtacagttacaaagttctgttacctatgtagatgagcttgtgctgaacatgttgtgattttggcaggtgtgtgctaaaggacaggaataagccaatcttattcacaatggcgagattggatcgtgtgaagaatttaaccggacttgttgagtggtatgccaagaacgcacggctaagggagttggttaaccttgttgtggttggtggagatcgaaggaaagaatccaaagatttggaagagcaaacagaaatgaaaaagatgtatgagctaataaagactcacaatttaaatggccaattcagatggatttcttcacagatgaaccgagtgaggaacggtgaactctaccgatacattgctgacactagaggagctttcgtgcagcctgcattctacgaggctttcggtttgactgttgttgaggccatgacctgtggtttgcctacatttgcaactaatcatggcggtccagctgagatcatcgttaacggaaaatctggcttccacatcgatccatatcacggtgagcaagctgctgatctgctagctgatttctttgagaaatgtaagacagaaccttctcattgggaaaccatttcaacgggtggcctgaagcgcatccaagagaagtaagcaactctttcttgactctagtcattgaaattaactttcttgactctagtcattgaaattaactcgggatttgaggcgtagttgattgatattttatcgcgtctctactactgatatatacaggtacacgtggcaaatctactcggagaggctattgacattggctgctgtttacgggttctggaaacatgtttctaagcttgatcgtctagaaatccgtcgatatcttgaaatgttttatgctctcaaataccgcaagatggtgagttcctcttcttccttgcccttctcctagtgtttaagatacaatataattgattgcattatcttagagaatcattaatgttaaattttcttaattcttgaatctgttaatgaagtttttctcttggtttttgtttaggctgaagctgttccattggctgctgagtga
SEQ ID NO: 8: Полипептидная последовательность NtSUS2-T
MLFMGLKQLSEDFSPAESTAMAERVLTRVHSLRERLDATLAAHRNEILLFLSRIESHGKGILKPHQLLAEFDAIRQDDKKKLNDHAFEELLKSTQEAIVLPPWVALAIRLRPGVWEYVRVNVNALVVEELTVPEYLHFKEELVDGTSNGNFVLELDFEPFTASFPKPTLTKSIGNGVEFLNRHLSAKMFHDKESMTPLLEFLRVHNYKGKTMMLNDRIQNLTTLQNVLRKAEEYLIMLPPETPFSEFEHKFQEIGLEKGWGDTAERVLEMICMLLDLLEAPDSCTLEKFLGRIPMVFNVVILSPHGYFAQENVLGYPDTGGQVVYILDQVPALEREMLKRLKEQGLDITPRILIVTRLLPDAVGTTCGQRLEKVYGAEHSHILRVPFRTEKGIVRKWISRFEVWPYMETFTEDVAKELAAELQAKPDLIIGNYSEGNLVASLLAHKLGVTQCTIAHALEKTKYPDSDIYWKKFDEKYHFSSQFTADLIAMNHTDFIITSTFQEIAGSKDTVGQYESHQAFTMPGLYRVVHGIDVFDPKFNIVSPGADINLYFPYSEKEKRLTALHPEIEELLYSDIENEEHLCVLKDRNKPILFTMARLDRVKNLTGLVEWYAKNARLRELVNLVVVGGDRRKESKDLEEQTEMKKMYELIKTHNLNGQFRWISSQMNRVRNGELYRYIADTRGAFVQPAFYEAFGLTVVEAMTCGLPTFATNHGGPAEIIVNGKSGFHIDPYHGEQAADLLADFFEKCKTEPSHWETISTGGLKRIQEKYTWQIYSERLLTLAAVYGFWKHVSKLDRLEIRRYLEMFYALKYRKMAEAVPLAAE
SEQ ID NO: 9: Полинуклеотидная последовательность NtSUS3-S
atggcgaatccaaagttcacaagagtacctagcatgagggagagagttgaggatactctctctgctcaccgtaaccagcttgttgctctcctctccaggtatattaataaactctatatacttgttattttctttatttttttgtctttactgataaatttaactgttttcttctttaaatcttgctttcgatgcatgatttctgttgtgttaaattgcgtaaccattttatctaaaagtttatgctgataaacacttttaaattttaatatgtaaattatattatgtctcaacatcaacatgtggatggccaaaaatataaagcttaattttcgttattttgaatgatttttctctgcgagtgttacggtttgcgtacacattacctaaacctcctccctagtccccacttgtgggaatttaattttttttttctttgtttttttttgttgttgttgttgtctgagttcaattcctaccatgttagcttggcaaaaataagttggtaaagcttgaccccaactagttttagttgatcgatttatttggtgatttatagttcaataataataattactattagagaaagttccagcagcttttctgtttgtttttccagttttagtgattgatatatgtgtatatatattctttgtttcttttaagatacgtggcgcaggggaaggggatattgcaacctcaccacttgatcgatgagttcaacaacgctgtatgtgatgacactgcttgtgagaagctcaaagatggtccctttagtgaagtcttgaaagctactcaggtatattcactaatccatgggaatcaagatgatactgtatatctttattatggtgtctttcagaaatttgacgatgatgaaatgcaacttttctctgtttgtcaccttatccagactgtttttttattttttatttttcattttttaacttgaaatgctcttaatttcctttgtttatcgataagaccggatttacaatgtatgaacggagcatcttaagaaccttctggaatgaagatataagatataaaacatggtgtccgttttctcctttgtggaatcagtgtacatatagactgttattttggtcccactttctggatcttctgatcacaccttctcatgcagaggcgagcttgatggtttcaacctttaaattcttactattgaatccatttcactttcgaaattatgagttcgaaatctaatatttgttgaaatttttgcaaatgttcacatataagtttaagctttgtgtcaagaatactgggctcaatggattccaatagaccaggctgtatccgcctctgtctccactctccctgcatccacttctttcgtgtgactaataatgcttaatgagctagaactcgttttaatgtttgaataagttgcttatatcagagcagcttttgatgtttcaatctttaacgggttatgcagtaccagcattctgcggctgaaaaacaggaatctgagatttacttgtctctggctgaatttcttgttcattttgctaacaagtactttggagttaatgcttgctctctgttgtcaaaataggaagccattgtgctgccaccatttgttgccatagcagttcgtccaaggccaggtgtttgggagtatgttcgtgttaatgtatatgatttgagcgttgaacaattgactgttcctgaatatcttcatttcaaggaagaacttgtggatggagagtaagctctttcttatttcaatacgaaacataaaaatttacagaagttgaataattaacaaatttgttgatttttaatgtatgccaggggtaataatcactttgtgcttgagctggattttgagccatttaatgcatcagttcctcgtccatctcgatcgtcatccattggcaatggagtccaattcctcaatcgtcatctttcctcaattatgtttcgcagcaaagactctctggaccccttacttgatttccttagaggacactgtcataaagggaatgtaagtaccaaaagcagttttccctttgtaaatgtctgcttgtccctgattatctactaaatctttcaacacgcgcaaccattataagaaatgtacaatacttctagttagaatttcatcatcgacaaactatctgctttactttttatttttcccatttgatggatgatagtttagtttatataacagatgatattttggttgaagggtaccatgaactttttcacaaccacttaatggatacatagttgtaatagttgacattttggaataatattgtctcacttggaaatgtttaagaagtattactacttctatttgtaagatggattgtttatctatgcaggtcttgatgttgaatgatcgtatacagcgaatctccaggctggagtctgctctttctaaagcagaggattatctctccaagctatcaccagatacatcctataatgagttcgaatacgcgtgagcttgtacacatttgttttgttttctttcaagcatatgtaatttctcaagaaaagggaaatctataggagttgaaacattctttatggaaccatgtgcatgcagattgcaagaaatgggctttgagagaggttggggtgatactgccagacgtgttttggagacgatgcatcttctttctgacattcttcaggctccggatccatcaaccttggagacatttcttggtagactacctatggtgttcaatgtcgtcatattatcccctcatggatattttggccaagcaaatgtcttgggtttgcccgacactggtggccaggtaataacaaggagaatgaggtcttgtattatgtactccctccgttccaatctatatgaacctatttgactgggtatggaaagaaatgaagacttgtaaaacttgtggttctttagaaattccaaacattacatttggttttttccctcttcctggaaattatactactgaatcatctctagatgttccagtttaacttgagacgtaagggtaaataacggaccattactctgtcctttcttgcagtaggcttggtacaatgaatatagttcgcatagttgccggaagctagagctgtgttagaaaactcaggaacattaatttggcgatgctaatcactgctaatgttactgaagcatccatggttttccttgatgttattctccttttggttgcttcacaggttgtctatatactggatcaagtgcgtgccttggaggccgaaatgcttcttagaataaagcaacaaggacttaacttcaagcctagaatccttgtcgtgagtacatatatattatgcaagctcttatttggtttgtgggattgcagttgacatcaatttgcttactctgattactaaaggtcacacggctgatacctgatgctaaaggaaccatgtgcaaccagaggttggagaggattagtggaactgaatactcgcatattttacgtgtcccttttaggacagagaagggaatccttcataaatggatatctaggtttgatgtatggccttacctggagaagttcactgaggtaacctctttgtcccttggaaattgccttttgttgctgatgtttctgctagtgtgcttaaatgacggatgttaactagtcacttgctagcgtttgcaatagcaacgggaaaagaaaggatttttgctagtttgaagtctgcctccaagaaaaattatattaaaagtttatggctagtggaaacatcagtcattcatgtaccttatttctatgcccaagttgtttaagttgaaagtaatttggccaactatgcaaattgggagaacgtgtagccaactattgtgtttgccgacatgttgatatactttttggtcctgatttatatttgttggtttgtcatactggatgaagcaattctcatgtttttctgcttatatatattggaagaagagatacttgtcgtttcatcatttttctcgacctctctattaccaacactttgccaatttaatgtttggaaatgtcttcttgaccaggatgtggcaagtgaaatgaccgctgagctccagggaaagccagatctgattattggcaactacagtgatggaaatttagttgcctcccttttggcatataaaatgggtgtcacacaggtaggaaatacatgattctttatcttgctagcactaagtcttgaggttatgtatctgcaatagaaattttacgctttgccttcatttctttttaattatttttccagtgtaccattgctcatgccttggaaaaaacaaagtatcctgattctgacatctactggaaaaagtttgaggagaaatatcatttttcatgtcagtttactgctgatctactggcaatgaataattcagatttcattatcaccagtacttatcaagagattgcaggaacgtaagtcattttaatctggtcgtttaaatctgatatttcttccctagtagtctattcaatccgaatttcagttcagtatatgatgtcatcggttgaggaactgtgattggtaaccttatcaaatccgtagctgctctataattttatttcgtaattggagaaacaattttttattattgagcttgtagtctgagctagaatttggttctttatctatcaagtagcataatactacaactattttttatgtgtggcaatttgcaatttcaattttctatttctataagttgcagcttttcttcctgttctgatcatatttacatggctgaaactcaatagaaaactaggctagttgatcaaaagtagttggatgctttaaaattagtagacgttttgctaaatgagtgaccaatgttattaaaaaaacgttcatgttttcaacccttttggcatacatttgaccactgcccaagattttggataagtacatgcagtgcttataattataaagcattttatcccaccttgtttttcattatgaaaattaagtaatttacgagtatttgtataagttacttcataaattagaagtaaatctggattgtgtaaagttattcgccccgtatatactgaaagctacttgaacaagcaaaaaaacagacaaacgtaacattctccatggattaatgagacttgtatatatatatatatatatatgtaaagagagagagagagagatttggcttgtaaccacatgtatattatgccatatggatgtgacattgatgtgactagacctaaatgttttgtttcaatgtccacgggagttttacgtagagttaagaggagaagagagtgaggaatactaatgtttgatggtaccccttggcttcttgacctggatactcagtgttcttattcatgcctatactttggtccttgatttcattctcccttttctagcttgagctgcatcaaagaaattccactgtaaaaaaaataatgctcaccatattggtgcaacatggcaaacatgtatcctatttgatgatcaatcaactttatttttctcctgttaattgacctcagtgtgtaactctctatgtatgatagcattgtaacttgtgtcatgattcataaatagggtactagaattggatggttgacatagtaaatggtcaattgatgatccacaaaatatgcacctactgattaaaatgtgatagggcaggtttatttttgtttgtggttaacacagtacttaaccctatatttaatacaatttggcttatctacaatcttttcttcagtgtttatgcgaattccttattgcacaacaatattgtctttctgagttctattctgttgttgcttacacttttattattccagtaacatagatgtgaagacattagattggttgcttgcaaattgatagccacttgtttcaggaagaatactgttggtcagtacgagagccatactgcattcaccctcccgggactatatcgcgtcgttcatggcattgatgttttcgatcccaaattcaatatagtgtctcctggagctgacatgacaatttatttcccatattctgacaaggaaaaaagactaacgtctttgcatggctcgattgaaaagttgttatttgatcctgcgcagaatgaagagcatatgtaagtggcatccgtttgtacttaatttttttggaatagatgacatattatttgcatgaatatgaaaaggagggtctgatatgattttctatagataaactaccaatgatattatttaaaaactcctggatactgtattaggagaagaagagaaccaggggtagatggcattagaatcccttaaatcttgaagagtcgtcactaacgctcccaacacttctgcctcagaccctcaactaaatactattattgttgatttctttggagaagctataagaatctctctctccttatggtgaaaattttacttggctttatacttaacttccaaggctccctcttataaaatgcaaaaactgtctgtattcactctcttggttaacaattgatccaatcaaatgcatatggaacatctttctttacgtttcttctaaagttcgtttgaggataaggagtagaatctgagaagatagactagtaggtaaccttagggacggatgtggaaattaacatatgggctcagcttttctgccgagtgcagaccatgtatatgcgttaaaaaattcactaaacaagtaaatgtttgattttgaacccagtaaatcaaatgagttgtggtagaatctcgaactcgaaccgataaagttcaaatccaggatccgcttttaggtaaactctaccttgggaagtgttatatatatgtccctgattatttctttttccgtttcctttctattttaatttttaaagttatttttagatggttttattttttgataagtggtaagttgttaatattccaaattaaatgccattgtcataactatatacatttataaagaatgattgatcctagtttctcattcctaagatccaaataaggcaataaacaatgtcttagtaattggacctgcttctggtgatcaacgcttgatcgcgtagttagttatagatgactgtaaaaactttaaccattttaatggttttgtcaaagaacaaatatcggacatattatagagaatggactattgtactttgcttctgattggtcattttattgtgatccgtaaattggctgtgactgatgtcatatctttgcttacagaggtaatctgaatgataaatcaaaacccataattttttcaatggcaaggctagaccatgttaagaacattacgggactagttgagtgctatgctaaaaatgccacattgagggaattggcgaaccttgttgtagtagctggatacaacgatgtaaagaaatccagtgatagagaagaaataacagaaattgagaagatgcatgctcttattaaggagcataaattggatgggcaattcagatgggtatcagcccaaacaaaccgggcacgtaatggtgagctctatcgctatatagctgaccagagaggtatatttgttcaggtatgctatttgtattgtattagtccaatttcattttttgcaccaaaagaaaggttgttattgtgacgtatatgtttgttttagcctgcattttatgaagcatttggactaacggtggttgaagctatgacttgtggtcttccaacatttgcaacttgccatggtggtcctaatgagatcattgaacccggtgtatctgggttccatattgatccttatcatcccgataaagctgctgaactcatgtcagaattctttcaacgctgcaaacaagatcctactcactgggaaaaaatatctgcatctggtctccgaaggattcttgagaggtctgtagttgtgtacatgtatagaagattaaagaatgctaccttgatatttatttgaatcaaaaataacaggaacatctcttttttgaacatcactcaagttcttatattaaataatttttaggtatacgtggaagatttactccgagaggctgatgactttatctggcgtatatggtttctggaagcttgtttcaaaacttgagaggcgtgaaactagacgataccttgagatgttctacattctcaaattccgcgagttggtgagtgccttttagctccttttcagttccaataaactatatatgtggtttaagtaagtattaagcataaacatgtccgtgcttggggctgtcgaaaatgctatggacatatcctgagctaaggatttttcaagaaaattgatgttagctttactctatttacaggcaaaatctgtacctctagcaattgatgacaagtga
SEQ ID NO: 10: Полипептидная последовательность NtSUS3-S
MANPKFTRVPSMRERVEDTLSAHRNQLVALLSRYVAQGKGILQPHHLIDEFNNAVCDDTACEKLKDGPFSEVLKATQEAIVLPPFVAIAVRPRPGVWEYVRVNVYDLSVEQLTVPEYLHFKEELVDGEGNNHFVLELDFEPFNASVPRPSRSSSIGNGVQFLNRHLSSIMFRSKDSLDPLLDFLRGHCHKGNVLMLNDRIQRISRLESALSKAEDYLSKLSPDTSYNEFEYALQEMGFERGWGDTARRVLETMHLLSDILQAPDPSTLETFLGRLPMVFNVVILSPHGYFGQANVLGLPDTGGQVVYILDQVRALEAEMLLRIKQQGLNFKPRILVVTRLIPDAKGTMCNQRLERISGTEYSHILRVPFRTEKGILHKWISRFDVWPYLEKFTEDVASEMTAELQGKPDLIIGNYSDGNLVASLLAYKMGVTQCTIAHALEKTKYPDSDIYWKKFEEKYHFSCQFTADLLAMNNSDFIITSTYQEIAGTKNTVGQYESHTAFTLPGLYRVVHGIDVFDPKFNIVSPGADMTIYFPYSDKEKRLTSLHGSIEKLLFDPAQNEEHIGNLNDKSKPIIFSMARLDHVKNITGLVECYAKNATLRELANLVVVAGYNDVKKSSDREEITEIEKMHALIKEHKLDGQFRWVSAQTNRARNGELYRYIADQRGIFVQPAFYEAFGLTVVEAMTCGLPTFATCHGGPNEIIEPGVSGFHIDPYHPDKAAELMSEFFQRCKQDPTHWEKISASGLRRILERYTWKIYSERLMTLSGVYGFWKLVSKLERRETRRYLEMFYILKFRELAKSVPLAIDDK
SEQ ID NO: 11: Полинуклеотидная последовательность NtSUS3-T
atgtttacatggctgaaactcaatataaaaaacaagggtaggtgatcaaaaatcgttggatgcttaaaatcagtagacgttttgctaaatgagcgaccaatgttattgaaaacgttcatgttttcaacccttttggcatacatttgagcattgcccaagattttggataagtagatgcagtgcttataattttaaagcattgtatcctgccttgtttttcattgtcaaaattaattaacttacaagtatttctataagttgcttcataaattagaagtaaatctggattgtgtaatgttattcgcctcgtaaatactgaaagctgcttgaacaagtgaaaaaacacagacaaacgtaacattctccatggattgatgagacttgtaaaatacatatatagaaatttggcttgtaaccacatgtatattatgccatatggatgtgacattgatgtgactagacctaaatgttttgtttccatgtccactggagttttacgtatagttaagaggagaaaagactgaggaatactaatgtatgatggtacccctttgcttcttgacctggatacccagtgttcctattcatgcctatactttggtccttgatttcactctcccttttctaacttgagctgcatcaaagaaatttccactgtaaaaaaataaataatgctcaccatatctctgcaacattgcaaacatgtatcccatatgattgatattggtgcgacatggcaaacatgtatcctatttgatgatcaatcaaatttatttttcccctgtcaaaatgacctcagtgtgtaattccctatgtatttgatagcattgtaactcgtgtcatgattcatgaatagggtactagaattgcatggttgacaaatattaactggtcgattgatgatccacaaaacatgcacttactgactaaaatgtgatgggacagatttatttttgtttgtgattaacacagtacttaaccctatacttaatacaatttggcctagctacaatcttttcttcagtgcaaattccttgttacacgaccaatattgtctttctgagttctattctgttgttacttacacttttattattcgaataagacattagattgcttgcatgcaaattgatagccacttgtttcaggaagaatactgttggtcagtacgagagccatactgcattcaccctcccaggactatatcgcgtcgttcatggcattgatgttttcgatcccaaattcaatatagtgtctcctggagctgacatgacaatttacttcccatattctgacaaggaaaaaagactaacgtctttgcatggctcgattgagaagttgttatttgatcctgcgcagaatgaagagcatatgtaagtgacatccatttgtacttattttaatttggaatagatgacatacttatttgcatgaatataaactgacaacccagagatttcctacattagaaaaggagggtctgatatgattttctacaaataaattcccagtgatattgttcaaaaagtcctggatactttattatgagagaaccagggatagatggcactagaatcccttaatcttgagaagtcgccacttatcgctcccaacactttctgagaccctcaagtaactactattattgtttgatatcttggagaagctataagaatctttttctccttattgtaattttttttacgtgactttaaacttaacttccaagctccttctgataaaatgcaaaaactgtctgtattcactgtcttggtttattaacaattgatccaatcaaatgcatatggaacatctttctttttgtttcttcaaaagttcgtttgaggataaggagtagaatctgagaagatagactagtaggtaaccttaggggcggatgtagaaatcaacgtatgggttcagctttgttgcagaccctgtatatgcattaaaaaaatcactaaataagtaaataattgattttgaacccagtaaatcaaaatgagttgtagtagaatcctgaactcgaaccgataaagttggatccactaccgggtaaactctaccttgagaagtgtttatatatgtccctaattatttcttttctgtttcctttctattttaattttttaagttcctttttagatggttttattttttgacaagtggtaagttgttagtattccaaattaaatgccattgccataactatatacatttataaagattgattgaccctagtttctcattcctaagatccaaataaggcaataaacaatatgtcttagtacttgaacctgcttctggtggtcaacacttgatcgcgtagttagttatagatgactgtaaaaaccttaatcattttaatggttttgtcaaagaacaaatatcggacatattatagcgaatggactattgtacttttcttctgattggtcattttattgtgatccgtaagttggctgagactgatgtcatatctttgcttacagaggtaatctgaatgataaatcaaaacccataattttttcaatggcaaggctagaccatgttaagaacattacgggactagttgagtgctatgctaaaaatgccacattgagggaattggctaaccttgttgttgtagctggatacaacgatgtaaagaaatccagtgatagagaagaaatagcagaaattgagaagatgcatgctcttattaaggagcataaattggatgggcaattcagatggatagcagcccaaacaaaccgggcacgtaatggtgagctctatcgctatatagctgacaagagaggtatatttgttcaggtacgctgtttgtattgtatttgtccacattcctttttttgcaccgaaagaaaggttgttattgtgacaaatatgtttgttttagcctgcattttatgaagcatttggactcacggtggttgaagctatgacttgtggtcttccaacatttgcaacttgccatggtggtccgaacgagatcattgaacacggtgtatctgggttccatattgatccttatcatcccgataaagctgctgaactcatggcagaattctttcaacgctgcaaacaagatcctactcactgggaaaaaatatctgcatctggtctccgaaggattcttgagaggtttgtagttgtgtacatatatagaagattaaagattgttcccttgatattatttgaatgaaaaataacagtaacatctctttttgaacatcgctcaagttcttgtgttaaataattgttaggtatacgtggaaaatttactccgagaggctgatgactttgtctggtgtatatggtttctggaagcttgtttcaaaacttgagaggcgcgaaactagacgataccttgagatgttctacattctcaaattccgcgagttggtgagtgcctttttgctcattttcagttacaatcaactatatatgtggtttaaatacgtattaagcataaacatgtccgtgattgcggctgtcgaaaatgctatggacatatcctgagctaaggagttttcaagagaattgatttggcttactctgtttacaggcaaaatctgttcctctggcaattgatgacaagtga
SEQ ID NO: 12: Полипептидная последовательность NtSUS3-T
MFTWLKLNIKNKGRKNTVGQYESHTAFTLPGLYRVVHGIDVFDPKFNIVSPGADMTIYFPYSDKEKRLTSLHGSIEKLLFDPAQNEEHIGNLNDKSKPIIFSMARLDHVKNITGLVECYAKNATLRELANLVVVAGYNDVKKSSDREEIAEIEKMHALIKEHKLDGQFRWIAAQTNRARNGELYRYIADKRGIFVQPAFYEAFGLTVVEAMTCGLPTFATCHGGPNEIIEHGVSGFHIDPYHPDKAAELMAEFFQRCKQDPTHWEKISASGLRRILERYTWKIYSERLMTLSGVYGFWKLVSKLERRETRRYLEMFYILKFRELAKSVPLAIDDK
SEQ ID NO: 13: Полинуклеотидная последовательность NtSUS4-S
atggcggaacgtgtgctgactcgtgttcatagccttcgtgaacgtcttgatgctactttggctgctcatcgcaatgagattttgctgtttctttcaaggtatagtcttagcagattgttctttgatttagttgttattgccagttctaatgtatgggcttatatataaacaaagtgttgaagtatgcaaccatataaactgacagcttaaaatgcttgagagaacacacttttatttatttaattatgccttcagcacaagaagtggaacttgacgcaatggaaccataggtcacgggttcaagtcttggaacagcctgcaatctaaggctgcgtgtagtagaccctagtggtccggcccttccacatatctcgcttagtgtaccgggcccattgagtacgggttcggccgaacccagtcgctttggtccaatccatatatttgtcttaaaaatatattgaatatatacaaattgttaatttagtttaaatatgtgtatcatgggttattcatgctggttttggctgttgcaggattgaaagccatggaaaagggatactgaaacctcaccagttgctggctgaatttgattcaattcacaaagaagacaaaaacaaactgaatgatcatgcttttgaagaagtcctgaaatccactcaggtatttgtggttttagtgttaggtgatggatagcatttattgttttactaagatcacatatgtgtcagtttgtggctagtatttaaaatctggtgtattttgtcatactaggaagcaattgttttgtccccttgggttgcgcttgccattcgtctgaggcctggtgtgtgggaatacgttcgtgtgaatgtcaacgctcttgttgttgaggagcttaccgtgcctgagtatttgcaattcaaggaagaacttgttaatggaacgtaagttttaggttcgaatttgttgatttgttagataacatgttctgaactttttgattaaagttgtgtttttgactgatgcagctcgcacgataactttgttcttgagttggattttgagcccttcactgcatcatttccaaaaccaaccctcaccaaatcaattggaaatggagttgaattccttaaccgacacctctctgccaaaatgttccatgacaaggaaagcatgacccctcttctcgagtttcttcgagttcaccactacaagggcaaggtaaacttgtttttcctgtttgtctatgaatttagtttagttgttttgctccgcgaaaatttcagtggaaactgatttatgcaaccactgagtgattaatatgttcaaacttaccgacttctggttttctgtgtagacaatgatgctgaatgacagaattcaggacttaaatactctccaaaatgtcctaaggaaagctgaggaatacctcactaccctttcccctgaaacttcatactcggcatttgagcacaagttccaagaaattggcttggagaggggttggggtgacactgcggagcgtgttctagagatgatctgcatgctcctggatctcctcgaggctcctgactcgtgcacgcttgagaagttccttggtagaattccaatggtttttaatgtggtcatactttcaccccatggttatttcgcccaggaaaatgtcttgggttaccccgacactggtggccaggtgcactgcttatctgtgttcggtcttattatctctttaaaccctactgccacaagtgctgagatgaacctcctttaatttgcaggttgtctatattttggatcaagttcctgctttggagcgtgagatgctcaagcgcataaaggagcaaggacttgacatcaaaccgcgtattcttattgttcgtattcccagtaattgtgtttaaacttatgattatgcaggattttatctgttctaatacagcactcttgcttaaattctcaggttactcggctgctgcctgatgcggttggtaccacttgtggtcagaggcttgagaaagtgtttggaacagagcactcacacattcttagggtcccctttaggaccgagaagggcattgttcgcaaatggatctctcgctttgaagtctggccatacatggagacattcactgaggtgaagcaagctttctctattcatttttcaatcttccaattggttttggcagcaattttctgcttgctttgacttccgctaaaacttcggattttattgcattaggatgtggcgaaagaaattgctgcagaattgcaggctaagccagatcttatcattggcaattatagtgagggcaaccttgctgcctccttgttggctcacaaattaggtgtaacacaggtcggcaatgtttgtgacatgtaatttcatctttgcatttcctttcgtttgcaactaaaagatttaagagttctctctctcttttttttttccgtctactttgccttatgcagtgcacgatagctcatgctttggagaaaacaaaatatcctgattctgatatctacttgaagaaatttgatgaaaaataccatttctcagcccagtttactgccgatcttattgcaatgaatcacaccgatttcatcatcaccagcactttccaggagatagcgggaaggtatttttacatcagtttcccactctgattaaattacaatgtatttccctatatgattaaatactgtgtttgatcctaaatcatttctaaattttccagcaaggacactgttggacagtacgagagccacatggcgttcacaatgcctggactgtatagagttgttcacggcattgatgtgtttgaccccaaatttaacattgtgtcaccaggagctgatatgaatctctatttcccatactacgagaaggaaaagagattgacagcatatcaccctgaaattgaggagctgctgtttagtgatgttgagaatgacgaacacatgtatgttactaaactagcaatcctgctgcaaaattatggctaattatgtaaacaagtttgtactgaatagatttgttattcgatcaggtgtgtgctgaagaacaggaataagcctatcatattcactatggctagattggatcgagtgaagaacttaactggacttgtcgagctgtacgccaagaacccacggctaagggagttggttaaccttgtcgtggttggaggagaccgaaggaaagaatccaaagacttggaagaacaggcagagatgaagaagatgtacgaacttataaagactcacaatttgaacggccaattccgatggatttcttcccagatgaaccgcgtgaggaatggcgaactctacaggtacattgccgatactaggggagctttcgtgcagcctgcattttacgaggcttttggtttgactgttgttgaggccatgacctgtggtttgcctacatttgcaactaatcacggtggtccagctgagatcatcgttcacgggaaatctggtttccacattgatccataccacggggatcaggcagctgaacttctcgctgatttctttgagaaatgtaagaaagaaccttcgcactgggaagccatttccgagggcggccttaagcgtatacaggagaagtaagcaaactgctactcttttcatttttgcaaaacctactatgatcattattaagctcatttttgcaaaacctacttgctgttgttattgtttgttgcttccttttcactgttctttgagctgaaggtctatcagaaacagtctctctaccttcacaaggtaggggtaagatctgcgtgcacgttaccctcctcaaactctacttaattgtgagattacactaggtttgttgttgttgattctttgctaattaattaaaaggtacacatggcaaatatactcggatcggttgttgacactggctgctgtatatggattctggaagcatgtttccaagcttgatcgtcttgaaattcgccgttatcttgaaatgttctatgctctcaaattccgcaagctggtgagtttcattgctttctgcactcctgcaattgtatag
SEQ ID NO: 14: Полипептидная последовательность NtSUS4-S
MAERVLTRVHSLRERLDATLAAHRNEILLFLSRIESHGKGILKPHQLLAEFDSIHKEDKNKLNDHAFEEVLKSTQEAIVLSPWVALAIRLRPGVWEYVRVNVNALVVEELTVPEYLQFKEELVNGTSHDNFVLELDFEPFTASFPKPTLTKSIGNGVEFLNRHLSAKMFHDKESMTPLLEFLRVHHYKGKTMMLNDRIQDLNTLQNVLRKAEEYLTTLSPETSYSAFEHKFQEIGLERGWGDTAERVLEMICMLLDLLEAPDSCTLEKFLGRIPMVFNVVILSPHGYFAQENVLGYPDTGGQVVYILDQVPALEREMLKRIKEQGLDIKPRILIVTRLLPDAVGTTCGQRLEKVFGTEHSHILRVPFRTEKGIVRKWISRFEVWPYMETFTEDVAKEIAAELQAKPDLIIGNYSEGNLAASLLAHKLGVTQCTIAHALEKTKYPDSDIYLKKFDEKYHFSAQFTADLIAMNHTDFIITSTFQEIAGSKDTVGQYESHMAFTMPGLYRVVHGIDVFDPKFNIVSPGADMNLYFPYYEKEKRLTAYHPEIEELLFSDVENDEHMCVLKNRNKPIIFTMARLDRVKNLTGLVELYAKNPRLRELVNLVVVGGDRRKESKDLEEQAEMKKMYELIKTHNLNGQFRWISSQMNRVRNGELYRYIADTRGAFVQPAFYEAFGLTVVEAMTCGLPTFATNHGGPAEIIVHGKSGFHIDPYHGDQAAELLADFFEKCKKEPSHWEAISEGGLKRIQEKYTWQIYSDRLLTLAAVYGFWKHVSKLDRLEIRRYLEMFYALKFRKLVSFIAFCTPAIV
SEQ ID NO: 15: Полинуклеотидная последовательность NtSUS4-T
atggccgaacgtgtgctaactcgtgttcacagccttcgcgaacgtcttgatgctactttggctgctcatcgcaatgagattttgctgtttctttcaaggtatagtcttagcagattgttctttgatttagttggtgttatttgccagttctaatgtatggactaatatatgaacaaagtgcgaccatttcaactgacaacttaaaatgtttgagagaatacacgtttatttacttaattatggcttgagcataggaagtgtatcttggcgtaactcgtaaagttgacctcatgtgacaaggaggtcacggtttcgagccgtggaaacagcctcttgcagaaatgcaggtaaggctgcgtgcaatagatcgcccttccacggacccgcgcatagcgggaacttagtgcaccggttgggctgtccttttttatgtcttcagcacaaaaatttagtttaaacatgtgtatcatggattattcatgctggttttgccggttgcaggattgaaagccacggaaaagggatattgaaacctcaccagttgctggctgagtttgaatcaattcacaaagaagacaaaaacaaactgaatgatcatgcttttgaagaagtcctgaaatctactcaggtaatttgtggttttagtgttaggtgatggatagcatttattgtcttactaagatcatatatgtgtcagtttgtggctagtatttgaaaagtctggtgtggtttgtcatactaggaagcaattgtcttgtccccttgggttgcgcttgccattcgtctgcggcctggtgtgtgggaatatgttcgtgtgaatgtcaatgcacttattgtcgaggagctgactgtgcctgaatatttgcaattcaaggaagaacttgttaatggaacgtaagttttaggttcgaaatgatgatttgttaaataatatgttctgaactttttgattaatgttgtgttttcccctgatgcagctcgaacgataactttgttcttgagctggattttgagcccttcactgcatcatttcccaaaccaaccctcaccaaatcaattggaaatggagttgaattcctcaaccgacacctctctgccaaaatgttccatgacaaggaaagcatgacccctcttctcgagtttcttcgagttcatcactacaagggcaaggtaaacttgtttttcctgtttgtctatgaatttagtttctgaaagttgctttgcttcgtgaattttttagtggcaactgatttatgattttctgtgcagacaatgatgctgaatgacagagttcaggacttaaacactctccaaaatgtcctaaggaaggctgaggaatatctcactaccctttcccctgaaacttcatactcggtatttgagcacaagttccaagaaattggcctagagaggggctggggtgacaatgctgagcgtgttctagagatgatctgcatgctcctggatctcctcgaggctccagactcatgcactcttgagaagttccttggtagaattcctatggtttttaatgtggtcattctttcacctcacggatatttcgcccaggaaaatgtcttgggttaccccgatactggtggccaggtgcactgcttatttgtaacaccttacgcttttccctctgaaacttatttgcggcaagttctaaggtcctccttccttaatttgcaggttgtctatattttggatcaagttccggccttggagcgtgagatgctcaagcgcataaaggagcaaggacttgatatcaaaccgcgtattcttattgttcgtatctccaataattgcgtttaaacttatgattgtgcaggatttgatctgttcaaatctaatgactgattttcttttttttttttttttccctcaggttactcggctgctgcctgatgcggttggtaccacttgtggtcagcggcttgagaaagtgtttggaacagagcattcacatattcttagggtcccctttaggaccgagaagggcatcgttcgcaaatggatctctcgctttgaagtctggccttacatggagacattcactgaggtgaagcaagctttctctattcatttttcaatcttccaatctgttttggcagcaatttttcacttactaacactttggctttcgctaaaacttcggattttattacattaggatgtggcaaaagaaattgctgcagaactgcaggcaaagccagatcttataatcggcaactacagcgagggcaaccttgctgcctccttgttggctcacaagttaggtgtaactcaggtctgtaatgtttgtcacctgttatttcaactttgcatttcctttcatttgcaactagaagttaagagttctctctcttttatcttttccgtctattttgccttctgcagtgcaccatagctcatgcgttggagaaaacaaaatatcctgattctgatatctacttgaagaaatttgatgaaaaataccatttctcagcccagtttactgccgatcttattgcaatgaatcacaccgatttcataatcaccagcactttccaggagatagcgggaaggtattacatcacaatggatttccgatatgattaaattagttaatttaatcctacttcattgtgtttgatcctaaaacttttctaaatttcccagcaaggacactgttggacagtacgagagccacatggctttcacgatgcctggattgtatagagttgttcacggcattgatgtgttcgatcccaaattcaacattgtgtcaccaggagctgatatgaatctctatttcccctacttcgagaaggaaaagcgattgacagcatatcaccctgaaattgaggagctgctgtttagcgatgttgagaatgacgaacacatgtatgttactaaactagcaatcctgctgcaaaattgtggctaattatgtaaaaaagtttttactgaatagatttgtgcttctatcaggtgtgtgctgaaggacaggaataagccaattatattcaccatggctagattggatcgagtgaagaacttaactggacttgtggagttgtacgccaagaacccacggctaagggagttggttaaccttgtcgtggttggtggagaccgaaggaaggaatccaaagatttggaagaacaggcagagatgaagaagatgtatgaacttataaagacgcacaatttaaacggccaattccgatggatttcttcccagatgaaccgcgtgaggaatggcgaactctacaggtacattgccgatactaggggagcttttgtgcagcctgcattttacgaggcttttggtttgactgttgttgaggccatgacctgtggtttgcctacgtttgcaactaatcacggtggtccagctgagatcatcgttcacgggaagtctggttttcacattgatccataccacggcgagcaggcagctgaacttctagctgatttctttgagagatgtaagaaagaaccttcacactgggaagccatttccgagggcggccttaagcgtatacaggagaagtaagcaagctgctactcttttcatttttgcaaaacctaccatgatcattattaagctcatttttgcaaaacctacttgttattctttgttgcttccttttccctgttttttgagccgaggttttatcgaaaacatgctttctaccttcacaaggtaggggtaaggtctgcgtttgttattattgttgttgttgattctctgcgaattaattaaaaggtacacatggcaaatctactcggatcggttgttgacactggctgctgtttatggattctggaagcatgtttccaaacttgatcgtcttgaaattcgtcgttatcttgaaatgttctatgctctaaaattccgcaaactggtgagtttcactgctttctgcactcttccaattgttagttgagtgcactcatttaaactgtagctaaagctgttgtaaatcttcagttaagcagctgctaatgaagtttttatcttttgtttttggttcaggctgaagctgtcccgttggctgttgagtaa
SEQ ID NO: 16: Полипептидная последовательность NtSUS4-T
MAERVLTRVHSLRERLDATLAAHRNEILLFLSRIESHGKGILKPHQLLAEFESIHKEDKNKLNDHAFEEVLKSTQEAIVLSPWVALAIRLRPGVWEYVRVNVNALIVEELTVPEYLQFKEELVNGTSNDNFVLELDFEPFTASFPKPTLTKSIGNGVEFLNRHLSAKMFHDKESMTPLLEFLRVHHYKGKTMMLNDRVQDLNTLQNVLRKAEEYLTTLSPETSYSVFEHKFQEIGLERGWGDNAERVLEMICMLLDLLEAPDSCTLEKFLGRIPMVFNVVILSPHGYFAQENVLGYPDTGGQVVYILDQVPALEREMLKRIKEQGLDIKPRILIVTRLLPDAVGTTCGQRLEKVFGTEHSHILRVPFRTEKGIVRKWISRFEVWPYMETFTEDVAKEIAAELQAKPDLIIGNYSEGNLAASLLAHKLGVTQCTIAHALEKTKYPDSDIYLKKFDEKYHFSAQFTADLIAMNHTDFIITSTFQEIAGSKDTVGQYESHMAFTMPGLYRVVHGIDVFDPKFNIVSPGADMNLYFPYFEKEKRLTAYHPEIEELLFSDVENDEHMCVLKDRNKPIIFTMARLDRVKNLTGLVELYAKNPRLRELVNLVVVGGDRRKESKDLEEQAEMKKMYELIKTHNLNGQFRWISSQMNRVRNGELYRYIADTRGAFVQPAFYEAFGLTVVEAMTCGLPTFATNHGGPAEIIVHGKSGFHIDPYHGEQAAELLADFFERCKKEPSHWEAISEGGLKRIQEKYTWQIYSDRLLTLAAVYGFWKHVSKLDRLEIRRYLEMFYALKFRKLAEAVPLAVE
SEQ ID NO: 17: Полинуклеотидная последовательность NtSUS5-S
atggcctcaacagttgctgatagcatgcctgatgctttgaaacaaagccggtatcatatgaagagatgcttcgctaggtgaacacccttcttttatgttttttcccctctacgtgtttatgtcaaatttccatgcataatgctaactacttttcttctttttgacttcaaaattggatgtgaaaggttcattgcaatgggaaggaggctaatgaagttgaaacatttaacagaagaaatagaagaaactattgaagacaaggcagaaagaaccaggattttggagggttcacttggaaaaattatgagttccacacaggtcagcaccatttaaccaacttagttgaacaggaaaaaaagaaaaagcaaaagagttattgcaaggcgtaacgattttctttgaaattttcaggaggcagctgttgttccaccttatgttgcttttgcagtaaggcacaatcctggcttctgggattatgtcaaagttaacgctgaaactctctctgtggaagctatttcagccagggaatatctcaaattcaaagagatgatctttgacgaagactggtaagtggaaaattgtatcattttaaagagaaacaattttgtaacatacaagaatagttttgatggttgaatgtgcaagcagggcaaaggatgataatgcactggaagtagattttggtgcttttgactactctaatcctcggttagccctttcctcttctgtcggaaatgggctcaactttatctcaaaagttctgtcttcaaagtttggtggaaagccagaggacgcccagcctttgcttgattacttactagctcttaatcatcaaggagaggtatgaaaatggactacctttgtttcttaaaggtattatataatgatgcgcgttataaagttcctttttaaattgaaactttgcagaatctaatgatcaatgagaatctgaatggtgttgctaagcttcaagcagcattgatagtagctgaagtttttgtatcttcctttcccaaagacacaccttataaagactttgagcataagtaagcttctcatatgcttccattgtcatatgcagtataccaatgacatgctaccgaaaagttgtttatgtttgtgacttgattatgaaaactctaggctcaaagaatggggctttgataaagggtggggtcacaatgcaggaagagtaagagagacaatgagactgctttccgagataatccaagcaccagatcccataaatatggagtcctttttcagcaagcttcctactacattcaacattgttatcttctccattcatggttactttggccaagcagatgtccttggtctgcccgatactggaggccaggtctacatatacagcaatttatctccttttgcctcatattgcttattagcgacacttgcatcattgaaatcagacttttacttcacaggttgtttatattctggatcaagtaagggctttagaggaggaaatgttacaaagaatcaagcagcaagggctaaacgtgaagcccaagattcttgtggtgagttttgcaaaaatatgcttagacaggttttgagattgatcggagaagggattaagatgatcaagatctttgtttcctgctttcatgatgtaaacaggtatctcgtctcataccagatgctcgagggacaacatgcaatcaggagatggaacctattcttaactcatcccattctcacatcctgagaattccattcaggactgagaaaggagttcttcgccaatgggtttctcggtttgatatctatccttacttggagaactatgccaaggcaagtcttctaacaaaattaccacctattcatacactttatttactttcttgaactaatcgtttggtttgtgacgtatatcattaggatgcttctgctaagatacttgagctcatggaaggtaaaccagacctcataattgggaactacactgatggaaatttagtggcatctctattggccaacaaacttggagttactcaggttccgtagctgatcatatgatcatattttctacattgtttcttgataattaaatggaaatcttattggatgataacattttagggaaccattgctcatgcattagagaaaactaagtatgaagattctgatgtgaagtggaagcagtttgatcccaagtaccacttttcttgccaatttactgccgatttattggcaatgaatgctgctgattttatcattaccagcacatatcaagaaatcgctggaaggttagcactgactctctcagtatatttggcaacttaatgaatttactgcagtggccaacactaaaagctatcattcgtccttcagcgaaactaggcctggacaatatgaaagtcacacagcatttaccatgccggggctttatagagctgtttcaggcatcaatgtatttgatccaaagttcaacattgctgctcctggggctgaacagtctacctatttccctttcactgagaaacagaaacgattcagcacatttcgtcctgctattaacgaattactttacagtaatgaggaaaacaatgagcacatgtaagtctaattgcccattttcctaatctaaccattgcttaaatcgttctgtttttaccggatgtgtggtacttatcagtaacatttttttttggatcagtggatttcttgcagaccggaaaaaaccaattatattttcaatggcgagatttgatacagtgaagaacctgtcaggcttgactgagtggtatgggaagaataagaagttgcggaacttggtaaaccttgttattgttgggggattcttcgatccatcaaaatcaaaagaccgggaggaagcagctgaaatcaagaagatgcatgaattgattgagaaataccagctcaagggacaaatgagatggatagcagctcaaactgataaatatcgaaatagtgagctataccgaactattgctgacactaagggagcttttgtccaaccggctttatatgaagcttttggactaaccgttattgaagcaatggattgtggattgcctacgtttgcaactaatcaaggtggacctgcagaaatcattgttgatggggtttcaggtttccatattgatccttacaatggggacgaatcaagcaagaaaatagctgatttctttgagaagtgtaaggttgattctaaatattggaacaggatatctgagggaggtctcaagcgcattgaagaatggtaacaaactagttccaagtttaaaaaatggaaaaaatgcttatcatgttatattttcgtggttttaagttctgcttcgatgcagttatacgtggaagatttatgcaaacaaagtgttgaatatgggatcaatctatggattttggagacaattcaatgtggggcaaaagcaggctaagcaaagatactttgagatgttttacaatcctctcttcaggaaattggtaggttgtatatgttgaatacaatttactaagatcctcaaaatgaccaagaaatatacattgactatgctacttttgtaatttcacaggccaaaagcgtgccgatcccacatgaagagccattgccacttgcaacatcagactctactcaatcccaagaattaaaactaccactaccagttccagcagcagtagctaaagttctgccattaacaaggcatgcttttaacttaattacttctctacctagagtaactggtaaagtggatgtcaagtga
SEQ ID NO: 18: Полипептидная последовательность NtSUS5-S
MASTVADSMPDALKQSRYHMKRCFARFIAMGRRLMKLKHLTEEIEETIEDKAERTRILEGSLGKIMSSTQEAAVVPPYVAFAVRHNPGFWDYVKVNAETLSVEAISAREYLKFKEMIFDEDWAKDDNALEVDFGAFDYSNPRLALSSSVGNGLNFISKVLSSKFGGKPEDAQPLLDYLLALNHQGENLMINENLNGVAKLQAALIVAEVFVSSFPKDTPYKDFEHKLKEWGFDKGWGHNAGRVRETMRLLSEIIQAPDPINMESFFSKLPTTFNIVIFSIHGYFGQADVLGLPDTGGQVVYILDQVRALEEEMLQRIKQQGLNVKPKILVVSRLIPDARGTTCNQEMEPILNSSHSHILRIPFRTEKGVLRQWDASAKILELMEGKPDLIIGNYTDGNLVASLLANKLGVTQGTIAHALEKTKYEDSDVKWKQFDPKYHFSCQFTADLLAMNAADFIITSTYQEIAGSETRPGQYESHTAFTMPGLYRAVSGINVFDPKFNIAAPGAEQSTYFPFTEKQKRFSTFRPAINELLYSNEENNEHIGFLADRKKPIIFSMARFDTVKNLSGLTEWYGKNKKLRNLVNLVIVGGFFDPSKSKDREEAAEIKKMHELIEKYQLKGQMRWIAAQTDKYRNSELYRTIADTKGAFVQPALYEAFGLTVIEAMDCGLPTFATNQGGPAEIIVDGVSGFHIDPYNGDESSKKIADFFEKCKVDSKYWNRISEGGLKRIEECYTWKIYANKVLNMGSIYGFWRQFNVGQKQAKQRYFEMFYNPLFRKLAKSVPIPHEEPLPLATSDSTQSQELKLPLPVPAAVAKVLPLTRHAFNLITSLPRVTGKVDVK
SEQ ID NO: 19: Полинуклеотидная последовательность NtSUS5-T
atggcctcaactgttgctggtagcatgcctgatgctttgaaacaaagccgatatcatatgaagagatgcttcgctaggtgaacacccttcttgttctttttgttttttccctctaccatttatgtcaaatttcaatgcataatgctaactactttttttctttttgacttcaaaattggacgtgaaaggttcattgcaatgggaaggaggttgatgaagctgaaacatttaacagaagaaatagaaaaaactattgaagacaaggcagaaagaaccaagattttggagggttcacttggaaaaattatgagttccacacaggtcagcaccatttaaccaacttaattgaataggaagaaaaaaaaaagcaaaagagttattgcaaggcgtaacgatttcctttgaaattttcaggaggcagctgttgtcccaccttatgttgcttttgcagtaaggcacaatcctggcttctgggattatgtcaaagttgacgctgaaactctctctgtggaagctatttcagccagggactatctcaaattcaaagagatgatctttgatgaagattggtaactggaagattgtatcattttaaagaaacaattttttaatattcaagattagttttgatggttgaatgtgcaagcagggcaaaggatgaaaatgcactcgaagtagattttggtgcttttgactactctaatcatcggttagccctttcctcttctgtcggaaatgggctaaacttcatctcgaaagttttgtcttcaaagtttggtggaaaggcagaagatgcccagcctttgcttgattacttactagctcttaatcatcaaggagaggtatggaaatggactaccttcctttcttaaggaattatataatgatgtatgttataaagatcctttttaaacattgacactttgcagaatctaatgatcaatgagaatctgaatggcgtctctaagcttcaagcagcattgatagtagctgaagtttttgtatcttcctttcccaaagacacaccttataaagactttgagcataagtaagcttttcaaacgcttctgttatcatatgcaatataccaagaatatgttgccttttgaaaagttgtttatgtttatgacttgataatgaaaatactaggctcaaagaatggggctttgagaaagggtggggtcacaatgcaggaagagtaagagagacaatgagactgctttccgagataatccaagcgccagatcccataaatatggagtcctttttcagcaggcttcctactacattcaacattgttatcttctccattcatggttactttggccaagcagatgtccttggtttgcccgatactggaggccaggtttacatacacagcaatttatctccttttgcctcatatttacttattagcgacacttgcattattgaaatcacatttgtatttaacaggttgtttatattctggatcaagtaagagccttagaggaggaaatgttacaaagaatcaagcagcaagggttaaatgtgaagcccaagattcttgtggtgagttatgcaaaaatatgcgtagccaaggttttgaaattgttcagaggggattaagatgatcgagatatttgtttccttcttccattgatgtgtacaggtcactcgtctcattccagatgctcgagggactacatgcaatcaggagatggaacctatacttaactcgtcccattctcacatcctgagaattccattcaggacagagaaaggagttcttcgccaatgggtttctcggtttgatatctatccttacttggagaactatgccaaggcaagtctcctaccaaaattaccacctattcatacactttattcagttttttgagctaatcattctcatttgtcacgtatgtgattaggatgcttctgctaagatacttgagctcatggaaggtaaaccagacctcattattgggaactacactgatggaaatttagtggcatctctattggccaacaaacttggagttactcaggttctacagctgatcatttatctgatcagattttctacattgttttcttgataattaaacggaaatcttatgagattgtaacattttagggaaccattgctcatgcattagagaaaaccaagtatgaagattctgatgtcaagtggaagcagtttgattccaagtaccacttttcttgccaattcactgccgatttattggcaatgaatgctgctgattttatcattaccagcacatatcaagaaatcgcaggaaggttagcactgactctctcagtatatttggcaacttaatgaatgtactgcttgtggccaacactaaaagctattactcgtccttcagcgaaactaggcctggacaatatgaaagtcacacagcatttaccatgccggggctttatagagctgtttcaggcatcaatgtatttgatccaaagttcaacattgctgctcctggggctgaacagtctgcctatttccccttcactgagaaacagaaacgattcagcgcgtttcgtcctgctattgaggaactactttacagtaatgagcaaaacaacgagcacatgtaagtctaattgccccattttcctaatctaaccattgcttaaatgttctgtttttacttgatatgtggtacttatcagtgatattttttattggaacagtggatttcttgcagaccgtaaaaaaccaattatattttcaatggcaagatttgatacggtgaagaacttgtcaggcttgactgagtggtatgggaagaataagaagttgcggaacttggttaacctcgttatcgttgggggattcttcgatccatcaaaatcaaaagaccgggaggaagcagctgaaatcaagaagatgcatgaattgattgagaaatacaagctcaagggacaaatgagatggatagcagctcaaactgataaatatcaaaacagtgagctatatcgaactattgctgacactaaaggagctttcgtccaaccggctttatatgaagcttttggactaactgttattgaagcaatgaattgtggactgcctacatttgctactaatcaaggcggacctgcagaaatcattgttgatggggtttcaggcttccatattgatccttacaatggggatgaatcgagcaagaaaatagctgatttctttgagaagtgtaaggttgattctaaatattggaacaagatatgtggaggaggtctcaagcgcattgaagaatggtaa
SEQ ID NO: 20: Полипептидная последовательность NtSUS5-T
MASTVAGSMPDALKQSRYHMKRCFARFIAMGRRLMKLKHLTEEIEKTIEDKAERTKILEGSLGKIMSSTQEAAVVPPYVAFAVRHNPGFWDYVKVDAETLSVEAISARDYLKFKEMIFDEDWAKDENALEVDFGAFDYSNHRLALSSSVGNGLNFISKVLSSKFGGKAEDAQPLLDYLLALNHQGENLMINENLNGVSKLQAALIVAEVFVSSFPKDTPYKDFEHKLKEWGFEKGWGHNAGRVRETMRLLSEIIQAPDPINMESFFSRLPTTFNIVIFSIHGYFGQADVLGLPDTGGQVVYILDQVRALEEEMLQRIKQQGLNVKPKILVVTRLIPDARGTTCNQEMEPILNSSHSHILRIPFRTEKGVLRQWDASAKILELMEGKPDLIIGNYTDGNLVASLLANKLGVTQGTIAHALEKTKYEDSDVKWKQFDSKYHFSCQFTADLLAMNAADFIITSTYQEIAGSETRPGQYESHTAFTMPGLYRAVSGINVFDPKFNIAAPGAEQSAYFPFTEKQKRFSAFRPAIEELLYSNEQNNEHIGFLADRKKPIIFSMARFDTVKNLSGLTEWYGKNKKLRNLVNLVIVGGFFDPSKSKDREEAAEIKKMHELIEKYKLKGQMRWIAAQTDKYQNSELYRTIADTKGAFVQPALYEAFGLTVIEAMNCGLPTFATNQGGPAEIIVDGVSGFHIDPYNGDESSKKIADFFEKCKVDSKYWNKICGGGLKRIEEW
SEQ ID NO: 21: Полинуклеотидная последовательность NtSUS6-S
atggctactgcaccagccctaaatagatcagagtccatagctgatagcatgccagaggccttaaggcaaagccggtaccacatgaagaaatgttttgccaagtacatagagcaaggaaagaggatgatgaaacttcataacttgatggatgagttggagaaagtaattgatgatcctgctgaaaggaaccatgttttggaaggcttacttggctacatattatgcactacaatggtatagctagattcatatgtacttatgatgcccttatattgtttcctgatgtattactcttaaaaccttctttgatcaaatttacaggaggctgcagttgttcctccctacattgcctttgccacgagacagaatcctggattctgggaatatgtgaaagtgaatgctaatgatctttctgttgagggtattacagctacagaatacttgaaattcaaggaaatgatagttgatgaatgctggtatagtatacgttgcagcttatcataccttttgtggttttataacttcaatcagaaaactcatcagagttacctttgtgtgaacatgaaatgcagggcaaaagatgaatatgcactggaaattgattttggagcagtagacttctcaacgcctcgactgaccctatcctcttcaattggcaatggtctcagttatgtttccaagtttctaacttcaaagctaaatgctacctccgcgagtgcacagtgtctggttgactacttgctcactttgaatcatcaaggagatgtacgtcaacaaaaatcaaactccataagtaaacttgtcaactctaagaagaaaaaataggaaaagaagattcacgtaacaaattttctttatgttcaactgcagaaactgatgatcaatgagacactcagcactgtctcaaagcttcaggctgcactggttgtagcagaagcatctatttcctctttaccaacagatacaccatatgagagctttgagctaaggtgatttgttttttcctctacttccctccacttgtgccatgctacgtagtactaagtaacttcaattcttgtaaagattcaaacagtggggttttgagaaaggatggggtgatacagctgaaagggtcagcgacaccatgagaacactgtctgaggtgcttcaggcaccagatccattgaacattcagaagttctttggaagggttccaactgttttcaatattgtattgttctctgtccatggatactttggccaagcagatgttcttggcttgccagacactggtggtcaggtaagcatttaatagcttttacatttaacttctatgcattgacaataaaataatttttaacagtttgaccacttctgctcttgttcaacaggtagtttatgttttggatcaagttgtagcttttgaagaagaaatgctacaaagaattaaacagcaggggctcaatattaagcctcaaattcttgtggtgagttcctagacaatcgacgtgactatgcaattatgtagaggctgtttagaaaagttaatatcatatgttgattgcacagttaacccgactgattccggatgcaaaaggaacaaagtgcaaccaggaactagaaccaatcaagaatacaaaacattcacacatcctcagagttccatttaggacagaaaaaggagtgcttaatcaatgggtttcacgatttgatatctatccatatctggagagatatactcaggtatgtatttttatatcaaccttgctcatcaaagatgtgttgtttcctcaattccatttttccccttggcaaaaggatgctgctgacaaaatcgtcgagctaatggaaggcaaacctgatctaatcattggtaactacactgatgggaatctagtggcttcactaatggctagaaaacttgggataactctggtaacttttcttaatcatatttgatgttgcttcttctccaagttagttcttaatctccactgacctagaccatctttgcaacagggaactattgctcatgctttggagaagacaaaatatgaagactctgacataaaattgaaggaactcgatccgaagtaccacttctcttgccaattcacagctgatttgattgcaatgaattcagcagatttcattatcactagcacataccaagaaatagctggaaggtaagaattagagctaataagtaatgcattcatatgtatttcagcatcgctctttcaccatcatcgaatacacaccactactcagtaaatgtatttgctcaaaagtttgcaacttaatggatctcattcttgaatgcttcaacatatgcagcaaagataaaccaggacagtatgagagccatagtgcatttacccttccagggctttacagagttgcttcaggtatcaatgtctttgatccaaaatttaatattgctgcacctggggcagaccagtcggtgtatttcccttacacagaaaagcagaagcgtttgactgctttccgccctgccattgaggaactgctttttagtaaagtggacaatgacgagcacgtgtaagtctaagtgttaaacttcagcttagtgcctagaacatcccactgctctatgtattgatgtttcacttgtttcaaacagtggatatttagaagacagaaagaaacctatcctgtttaccatggcaaggctggacacagtgaagaacacatctggactaacagaatggtatggcaagaacaagaggctcagaagcttagttaaccttgttgtggttggtggttcctttgatcctacaaaatccaaggatagggaagaagcagctgaaataaaaaagatgcacatgctgatagagaaataccagcttaagggtcagattagatggatagcagctcagactgacagatacagaaatagtgaactctaccgcacaatagcagattccaaaggagcttttgtgcagcctgcattgtatgaagcatttggtctaacagtcattgaggcaatgaactgtggattaccaacctttgctaccaaccaaggtggccctgctgagattattgttgatggggtctcaggctttcatattgatccaaataatggggatgaatcaagcaacaaaattgccaactttttccaaaaatgcagggaggatcctgagtattggaacaggatttcagtccagggtctaaaccgtatatatgaatggtaactcacagataagccattcaaattgcaaagaggcacatatcttgcagaaaatttcttaatccttaaatcctaattttttgcagttacacatggaagatctatgcaaacaaggtattgaatatggggtccatctatactttttggaggacattgtacagagatcagaaacaagcaaagcaaagatacatcgagactttctacaatcttgagtttaggaacttggtatagtgctgcatgacattgacagtataccacaaacatctttatgagatgaattacttttaataaaattgtttttaacctttgcttccttaatggcacttattgcaggtaaaaaatgtgcctatcagaaaggacgaaacaccacaaggaccaaaggagagggagaaagttaagccacagatatcacaaaggcatgctctaaagcttttgcctacagtttttcaagagaccctagtatattctagtactaaattagaattatacagcatgcagcttttgctgttcacctttctaaatcaccagttgtgtcaatcaagttgacaaaatcaataaattgggattttccctttcctatgcttgattgttattactcctactttgtttatggtagtcttccttcattgttttctcctgtacttcttttactacaactgtactgacatactaattatttctgtgtaccaggcgctcacaatcaaggttgcagaagtaagattagataaaattgctactgcatga
SEQ ID NO: 22: Полипептидная последовательность NtSUS6-S
MATAPALNRSESIADSMPEALRQSRYHMKKCFAKYIEQGKRMMKLHNLMDELEKVIDDPAERNHVLEGLLGYILCTTMEAAVVPPYIAFATRQNPGFWEYVKVNANDLSVEGITATEYLKFKEMIVDECWAKDEYALEIDFGAVDFSTPRLTLSSSIGNGLSYVSKFLTSKLNATSASAQCLVDYLLTLNHQGDKLMINETLSTVSKLQAALVVAEASISSLPTDTPYESFELRFKQWGFEKGWGDTAERVSDTMRTLSEVLQAPDPLNIQKFFGRVPTVFNIVLFSVHGYFGQADVLGLPDTGGQVVYVLDQVVAFEEEMLQRIKQQGLNIKPQILVLTRLIPDAKGTKCNQELEPIKNTKHSHILRVPFRTEKGVLNQWVSRFDIYPYLERYTQDAADKIVELMEGKPDLIIGNYTDGNLVASLMARKLGITLGTIAHALEKTKYEDSDIKLKELDPKYHFSCQFTADLIAMNSADFIITSTYQEIAGSKDKPGQYESHSAFTLPGLYRVASGINVFDPKFNIAAPGADQSVYFPYTEKQKRLTAFRPAIEELLFSKVDNDEHVGYLEDRKKPILFTMARLDTVKNTSGLTEWYGKNKRLRSLVNLVVVGGSFDPTKSKDREEAAEIKKMHMLIEKYQLKGQIRWIAAQTDRYRNSELYRTIADSKGAFVQPALYEAFGLTVIEAMNCGLPTFATNQGGPAEIIVDGVSGFHIDPNNGDESSNKIANFFQKCREDPEYWNRISVQGLNRIYECYTWKIYANKVLNMGSIYTFWRTLYRDQKQAKQRYIETFYNLEFRNLVKNVPIRKDETPQGPKEREKVKPQISQRHALKLLPTVFQETLALTIKVAEVRLDKIATA
SEQ ID NO: 23: Полинуклеотидная последовательность NtSUS6-T
atggctactgcaccagccctgaaaagatcagagtccatagctgatagcatgccagaggccttaaggcaaagccggtaccacatgaagaaatgttttgccaagtacatagagcaaggcaagaggatgatgaaacttcataacttgatggatgaattggagaaagtaattgatgatcctgctgaaaggaaccatgttttggaaggcttacttggctacatattatgtactacaatggtatagctagattcatatgtacttatgatgtccttatattgtttccggaggcattattcttaaatccttctttgatcaaatttgtaggaggctgcagttgttcctccctatattgccttcgccacgagacagaatcctggattctgggaatatgtgaaagtcaatgctaatgatctttctgttgagggtattacagctacagattacttgaaattcaaggaaatgatagttgatgaaagctggtatagaatactttgcagcttatcataccttttgtggttttataatttcaatcagaaaactcatcagagttacctttgtgtgaacatgacatgcagggcaaaagatgaatatgcactggaaattgattttggagcagtagacttctcaacgcctcgactgaccctatcctcttcaattggaaatggtctcagttatgtttccaagtttctaacttcaaagctaaatgctacctcagcgagtgcacagtgtctggttgactacttgctcactttgaatcaccaaggagatgtacgtcaacaaaaatcaaactccataagtaaacttgtcaactctaagaagtaaaaataggaaaagaagattcatgtaacaaattttctttatgttcaactgtagaaactgatgatcaatgagacactcggcactgtctcaaagcttcaggctgcactggttgtagcagaagcatctatttcctccttaccaacagatacaccataccagagctttgagctaaggtgatttgttttttcctctacttccttccacttttggtgtgctacatagtactaagtaacttcaattcttgtaaagattcaaacagtggggttttgagaaaggatggggtgatacagctgaaagggtccgcgacaccatgagaacactttctgaggtacttcaggcgccagatccattgaacattgagaagttctttgggagggttccaactgttttcaatattgtattgttctctgttcatggatactttggccaagcaaatgttcttggcttgccagacacaggtggtcaggtaagcatctaatagcttttacatttaacttctatgcattgacaataaaataacttctacactaccaaataatttttgaaagtttgaccacttcggctcttgttcaacaggtggtttatgttttggatcaagttgtagcttttgaagaagaaatgctccaaagaattaaacagcaggggctcaatattaagcctcaaattcttgtggtgagctcctagacaatgacgtgactatgcaattaagtagaggctgtttagaaaagttaatatcatatgttgattgcacagttaacccgactgattccggacgccaaaggaacaaagtgcaaccaggaactagaaccaatcaagaatacaaaacattcacacatcctcagagttccatttaggacagaaaaaggagtgcttaatcaatgggtttcacgatttgatatctatccatatctggagagatatactcaggtgtgtatttttatatcaaccctgctcatcaaagatgtgttgtttcctcaattccatttttcgccttgacaaaaggacgctgctgacaaaatcatcgagctaatggaaggcaaacctgatctaatcattggtaactacactgatgggaatctagtggcttctctaatggctagaaagcttgggataactctggtaacttttcttatcatatttgatgttgtttcttctccaagttggttcttaatgtcaactaacccagaccatctttgtaacagggaactattgctcatgctctggagaagacaaaatatgaagactctgacatcaaattgaaggaactcgatccgaagtaccacttttcttgccaattcacagctgatttgattgcaatgaattcagcagatttcattatcacaagcacatatcaagaaatagccggaaggtaagaattggaactacggaagcagagagctaataagtagtgcactcatatatttcagcatcgctctttcgcataatcgaatacacaccactactcagtaaatgtacttgctcaaaagtttacaagtttatggatcttattcttgaatgcttcaacatatgcagcaaagataggccaggacagtatgagagccatagtgcatttacccttccagggctttacagagttgcttcaggcatcaatgtctttgatcctaaatttaatattgctgcacctggggcagaccaatcggtgtatttcccttacacagaaaagcagacgcgtttgactgctttccgccctgccattgaggaactgctttttagtaaagtggacaatgacgagcacatgtaagtcttagtgttaaacttcagctttcagcttagtgcctagaacattccactggctctatgtattaatgtttcacttgtttcaaacacagtggatatttagaagacagaaagaaacctatcctgtttaccatggcaaggctggacacagtgaagaacacatctggactaacagaatggtatggcaagaacaagaggctcagaagcttagttaaccttgttgtggttggtggttcctttgatcctacaaaatccaaggatagagaagaagcagctgaaataaaaaagatgcacatgctgatagagaaataccagcttaagggtcagatcagatggatagcagctcagactgacagatatagaaacagtgaactctaccgcacaatagcagattccaaaggagcttttgtgcagcctgcattatatgaagcatttggtctaacagtcattgaggcaatgaactgtggattaccaacctttgctaccaaccaaggtggccctgctgagattattgttgatggggtctcaggctttcatattgatccaaataatggggatgaatcaagcaacaaagttgccaactttttccaaaaatgcagggaggatcctgagtattggaacaggatttcagtccagggtctaaaccgtatatatgaatggtaactcacagataagccattcaaattgcaaagaggcacatatcttgctgaaaatttcttaatcctttaatcctaaaattttgcagttacacatggaagatctatgcaaacaaggtattgaatatggggtccatctatactttttggaggacattgtacagagatcagaaacaagcaaagcaaagatacatcgagactttctacaatcttgagtttaggaacttggtatagtgctgcatgacattgacagtataccacaaacatctttatgagatgaattacttttaataaaattgtttttaacctttgcctccttaatgacacttattgcaggtaaaaaatgtgcctatcagacaggacgaaacaccacaaggaccaaaggagaggagggagaaagttaagccacagatatcacaaaggcatgctctaaagcttttgcctatagtttttcaggagaccctagtatattctagtactaaattagaattatacagcatgcagcttgcttctgctgttcacctttctaaatcaccagttatgtcaatcaagttgacaaaatcaataaattcggcttttccctttcctatgcttgattgttattactcctacttcgtttatggtagtcttccttcattgttttctcctgtacttcttttactacaactgtactga
SEQ ID NO: 24: Полипептидная последовательность NtSUS6-T
MATAPALKRSESIADSMPEALRQSRYHMKKCFAKYIEQGKRMMKLHNLMDELEKVIDDPAERNHVLEGLLGYILCTTMEAAVVPPYIAFATRQNPGFWEYVKVNANDLSVEGITATDYLKFKEMIVDESWAKDEYALEIDFGAVDFSTPRLTLSSSIGNGLSYVSKFLTSKLNATSASAQCLVDYLLTLNHQGDKLMINETLGTVSKLQAALVVAEASISSLPTDTPYQSFELRFKQWGFEKGWGDTAERVRDTMRTLSEVLQAPDPLNIEKFFGRVPTVFNIVLFSVHGYFGQANVLGLPDTGGQVVYVLDQVVAFEEEMLQRIKQQGLNIKPQILVLTRLIPDAKGTKCNQELEPIKNTKHSHILRVPFRTEKGVLNQWVSRFDIYPYLERYTQDAADKIIELMEGKPDLIIGNYTDGNLVASLMARKLGITLGTIAHALEKTKYEDSDIKLKELDPKYHFSCQFTADLIAMNSADFIITSTYQEIAGSKDRPGQYESHSAFTLPGLYRVASGINVFDPKFNIAAPGADQSVYFPYTEKQTRLTAFRPAIEELLFSKVDNDEHIGYLEDRKKPILFTMARLDTVKNTSGLTEWYGKNKRLRSLVNLVVVGGSFDPTKSKDREEAAEIKKMHMLIEKYQLKGQIRWIAAQTDRYRNSELYRTIADSKGAFVQPALYEAFGLTVIEAMNCGLPTFATNQGGPAEIIVDGVSGFHIDPNNGDESSNKVANFFQKCREDPEYWNRISVQGLNRIYECYTWKIYANKVLNMGSIYTFWRTLYRDQKQAKQRYIETFYNLEFRNLVKNVPIRQDETPQGPKERREKVKPQISQRHALKLLPIVFQETLVYSSTKLELYSMQLASAVHLSKSPVMSIKLTKSINSAFPFPMLDCYYSYFVYGSLPSLFSPVLLLLQLY
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Филип Моррис Продактс С.А.
<120> МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ РЕДУЦИРУЮЩИХ САХАРОВ В РАСТЕНИИ
<130> P10505EP
<140> EP18164769.4
<141> 2018-03-28
<160> 24
<170> PatentIn версия 3.5
<210> 1
<211> 4384
<212> ДНКДНК
<213> Nicotiana tabacum
<220>
<221> иной_признак
<222> (1520)..(1520)
<223> n представляет собой a, c, g, или t
<400> 1
atggcagcta gtggtcttag cattaagaaa agtttggagg aatccatttt ggctcatcca 60
gatgaaattt tggctctcaa gtcaaggtac attactacat ataatgatat taagaactag 120
aggcttatcc aaggttttgt tacatttttg aaattataag tttagaacct aatagtactt 180
ggtagcactt gtttccttat tatctagctg ttgttactgc ttgttgctac tgctttctgt 240
tcatctttcc ttgagcccgg tctatcggaa acaacctctc tattctcaaa gtataaggtt 300
tgcgtacata ctacctcccc agactctact tgtggaattt actgtttttg ttgtgttgtt 360
gtaatctaat atttattaga attttactga tttttcacat atatatatct atgtcccctg 420
tcgaaaattc tatagctcat gttagctaaa tacattagta ccattgtttt taattgtttt 480
ggttttggca caggattgaa actgaaggga aaggggtaat gaaaccactt gatctcttga 540
accatttggt ttctgttact agtaagacaa atggagtaaa tattgtacct agtgcacttg 600
tggaagttct cagttgcagc caagaagctg tgattgtacc accaaaacta gcactagctg 660
tacgtccgag gcccggtgta tgggagtact tgtcactgaa tcttaagaca aagaaagtgg 720
ctgaattaag cattcctgaa taccttcaat tgaaagagaa cactgttgat gaaaggtaaa 780
gtattagtct gcgatttcgc tttgtgaaat tgaagttttt gttttgattc ataatgtttt 840
gtgtatcaat tatgttacca gtggaaacat attggagttg gattttgagc catttacaac 900
agttacacca ccaaaaacac tttctgactc tattggcaat ggtttggagt ttcttaatcg 960
ccacattgct tcgaaaatgt ttcatgataa ggagatttcc agatgcctcc ttgacttcct 1020
cagaaaccat aactacaaag gaaaggtaat aaaaaaaagt gtttctttaa acaagttgta 1080
tgattatgtg tatatttcta agtatgttaa cttgaaaaca gtcattgatg gtgaaagaaa 1140
gcattcaaag cctagagagt ttccaacttg ttctgaaaaa agcagaggaa catttgtgca 1200
cattgaatcc agaaactcca tactccaatt ttgaatcaaa gtttgaagag attggcttgg 1260
aaagagggtg gggaaacacc gctgaacgcg tgcaagacac tatcagtcat cttttgcatc 1320
tccttgaggc tcctaacgcg tcttctttgg aaaatttcct tggtagaatc ccattggttt 1380
tcaatgttgt gattctaact ccacatggtt attttgctca agataatgtc ttgggctatc 1440
ctgacactgg tggccaggtt tgtgtccaat attttgcatt cttgatcaag ttctttatac 1500
catttgaacc aacaatcttn aacattcttt ttttggttgt gaaatgttga ataggttgtt 1560
tacattcttg atcaagttcc agctatggag cgtgagatgc ttcatcgtat gaagcttcaa 1620
ggactcgatg atatcatccc tcgcatcctt gttgtaagtg gccttaattt tcctagtttc 1680
atttacacct ctaaatgaaa ttgatctttt ttgttgtttt atatcaggta acaaggctgc 1740
tgcctgatgc agtaggaacc acctgtggcg agcggatgga gaaagtatat ggggcagaac 1800
attctcatat aattcgtgtt ccatttagaa ctgagaaggg aatgttgcgc aaatggatct 1860
cacgattcga agtctggcca tacatggaaa ctttcactga ggttggaaca taaaaacaaa 1920
taaaatccat tggaatgttc cttctgcaat tgaaaatgtc ttgctaactg aagacccatt 1980
tttaaattga tcatcaggat gttgcagaag aacttgtcaa agaattgcaa gctaaaccag 2040
acttgatcat tggaaactac agtgagggaa atcttgctgc ctctttgctt gcgaagaaat 2100
ttggggctac tcagtgtact attgctcatg ccttggaaaa aactaagtat ccaaactctg 2160
accttaattg gaagaagttt gatgacaagt atcatttctc aagtcagttc actgctgatc 2220
tctttgccat gaatcacact gatttcatca tcaccagcac tttccaagaa attgctggaa 2280
ggtaaaagca aatgcacacc atcatagtat ttcatatttt tacccttgtt tatactattt 2340
ccattcaccg accccgactt gtttaggatt gagccatagt tgttgttgtt gtttgtttat 2400
actatttcca tttgccgacc acaacttgtt taggactgag gtatagttgt tgttgttggt 2460
ttgttcatat tattttcatt cgctaaccct aacttgtttg ggactgaggc atagtagtag 2520
tagtagttgt tgctattagt ttatactatt tccatttgcc aaccccaact tgtttggtac 2580
tgagacatag ttgttgttgt tgttgtttgt ttatactatt tccatttgcc gaccccaact 2640
tgtttaggac tgaggtatag ttgttgttgt tggtttgttc atattatttt cattcgctaa 2700
ccccaacttg tttgggactg aggcatagta gtagtagtag tagttgttgc tattagttta 2760
tactatttcc atttgccaac cccaacttgt ttggtactga gacatagttg ttgttgttgt 2820
ttgtttatac tatttcaatt tgtcgacccc aatttgtttg ggaccaaggc atggttgttg 2880
ttgttgtttg tttgttttta ctgtttccat tgatattgga acatttgtta tttgcagcaa 2940
aaacactgta ggacagtatg agagtcatac tgcttttacc atgcctggat tgtaccgagt 3000
agtccatgga atcgattcgt ttgatccaaa gttcaacatt gtctcccctg gggctgatat 3060
gtcaatctac ttcccttaca ctgagaagga gaaaaggcta accaacttcc acccggaaat 3120
tgaagaactc ctctacagtc ctgttgagaa taaggaccac ttgttagtct ccttaatttg 3180
cttttatttc atcccattta tgatcgcttt tatcccaaca gatcgattaa tcatttgtta 3240
tcaacataaa cagatgtgtg ttgaaggacc ggaacaagcc aattctcttt accatggcaa 3300
ggctagatcg cgtgaagaat ctaacagggc tcgtggaatg gtatgctaag aatgcaaggc 3360
tgagggagct tgttaacctt gtggttgtag gcggagacag aaggaaagaa tccaaagatt 3420
tagaagagca agcagagatg aagaagatgt atgatcttat cgaaacctat aacctgaacg 3480
gccaattcag gtggatttct tcccaaatga atcgtgtgag gaacggagaa ctctatcgtt 3540
acattgcaga cacgaggggt gctttcgttc aaccagcatt ctacgaggct tttggtttga 3600
cagttgtaga gtctatgact tgtggtttgc caacttttgc tacttgtaat ggtggaccat 3660
ttgagattat agtgaatgga aaatctggtt tccatattga tcctaatcaa ggtgacaagg 3720
ctgctgatat gttggtaaat ttctttgaaa aatctaaaga agatccaagt tattgggatg 3780
ctatttccaa gggaggtctg caacgtattc ttgaaaagta agcttttgca tttgattagc 3840
acaagtgcac aaccaagatt taacttttga acaaactaaa actaaccctt ttttgtattt 3900
tcttttgcta ggtatacatg gcaaatttat tcacagaaag tgatcacact atctgggatt 3960
tatggattct ggaagtatgc aaccaagaat gataaagttg ctagtgcaaa gaagcgctat 4020
cttgagatgt tttatgaact tggatttaag aaatcagtaa gtgtcaattt taaaggggaa 4080
ccttggatca acggttaagt tgtctttgtg caacctatag gtcaggggtt tgagccgtag 4140
aagtagccac taatatttac attagggtag actgtgtaca tatcacaccc cttggggtac 4200
ggccctttcc tggatcctgt atgaacgcgg gatgccttgt gcaccgggct gtattttttt 4260
ttttagtgtc acttctgtat tttgtttgag cttgtttata aagtttggaa atctgctgct 4320
aatttgtata tttgttggtt gtgtatttca ggctgagaaa gttccattgg ctattgatga 4380
atag 4384
<210> 2
<211> 803
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 2
Met Ala Ala Ser Gly Leu Ser Ile Lys Lys Ser Leu Glu Glu Ser Ile
1 5 10 15
Leu Ala His Pro Asp Glu Ile Leu Ala Leu Lys Ser Arg Ile Glu Thr
20 25 30
Glu Gly Lys Gly Val Met Lys Pro Leu Asp Leu Leu Asn His Leu Val
35 40 45
Ser Val Thr Ser Lys Thr Asn Gly Val Asn Ile Val Pro Ser Ala Leu
50 55 60
Val Glu Val Leu Ser Cys Ser Gln Glu Ala Val Ile Val Pro Pro Lys
65 70 75 80
Leu Ala Leu Ala Val Arg Pro Arg Pro Gly Val Trp Glu Tyr Leu Ser
85 90 95
Leu Asn Leu Lys Thr Lys Lys Val Ala Glu Leu Ser Ile Pro Glu Tyr
100 105 110
Leu Gln Leu Lys Glu Asn Thr Val Asp Glu Ser Gly Asn Ile Leu Glu
115 120 125
Leu Asp Phe Glu Pro Phe Thr Thr Val Thr Pro Pro Lys Thr Leu Ser
130 135 140
Asp Ser Ile Gly Asn Gly Leu Glu Phe Leu Asn Arg His Ile Ala Ser
145 150 155 160
Lys Met Phe His Asp Lys Glu Ile Ser Arg Cys Leu Leu Asp Phe Leu
165 170 175
Arg Asn His Asn Tyr Lys Gly Lys Ser Leu Met Val Lys Glu Ser Ile
180 185 190
Gln Ser Leu Glu Ser Phe Gln Leu Val Leu Lys Lys Ala Glu Glu His
195 200 205
Leu Cys Thr Leu Asn Pro Glu Thr Pro Tyr Ser Asn Phe Glu Ser Lys
210 215 220
Phe Glu Glu Ile Gly Leu Glu Arg Gly Trp Gly Asn Thr Ala Glu Arg
225 230 235 240
Val Gln Asp Thr Ile Ser His Leu Leu His Leu Leu Glu Ala Pro Asn
245 250 255
Ala Ser Ser Leu Glu Asn Phe Leu Gly Arg Ile Pro Leu Val Phe Asn
260 265 270
Val Val Ile Leu Thr Pro His Gly Tyr Phe Ala Gln Asp Asn Val Leu
275 280 285
Gly Tyr Pro Asp Thr Gly Gly Gln Val Val Tyr Ile Leu Asp Gln Val
290 295 300
Pro Ala Met Glu Arg Glu Met Leu His Arg Met Lys Leu Gln Gly Leu
305 310 315 320
Asp Asp Ile Ile Pro Arg Ile Leu Val Val Thr Arg Leu Leu Pro Asp
325 330 335
Ala Val Gly Thr Thr Cys Gly Glu Arg Met Glu Lys Val Tyr Gly Ala
340 345 350
Glu His Ser His Ile Ile Arg Val Pro Phe Arg Thr Glu Lys Gly Met
355 360 365
Leu Arg Lys Trp Ile Ser Arg Phe Glu Val Trp Pro Tyr Met Glu Thr
370 375 380
Phe Thr Glu Asp Val Ala Glu Glu Leu Val Lys Glu Leu Gln Ala Lys
385 390 395 400
Pro Asp Leu Ile Ile Gly Asn Tyr Ser Glu Gly Asn Leu Ala Ala Ser
405 410 415
Leu Leu Ala Lys Lys Phe Gly Ala Thr Gln Cys Thr Ile Ala His Ala
420 425 430
Leu Glu Lys Thr Lys Tyr Pro Asn Ser Asp Leu Asn Trp Lys Lys Phe
435 440 445
Asp Asp Lys Tyr His Phe Ser Ser Gln Phe Thr Ala Asp Leu Phe Ala
450 455 460
Met Asn His Thr Asp Phe Ile Ile Thr Ser Thr Phe Gln Glu Ile Ala
465 470 475 480
Gly Ser Lys Asn Thr Val Gly Gln Tyr Glu Ser His Thr Ala Phe Thr
485 490 495
Met Pro Gly Leu Tyr Arg Val Val His Gly Ile Asp Ser Phe Asp Pro
500 505 510
Lys Phe Asn Ile Val Ser Pro Gly Ala Asp Met Ser Ile Tyr Phe Pro
515 520 525
Tyr Thr Glu Lys Glu Lys Arg Leu Thr Asn Phe His Pro Glu Ile Glu
530 535 540
Glu Leu Leu Tyr Ser Pro Val Glu Asn Lys Asp His Leu Cys Val Leu
545 550 555 560
Lys Asp Arg Asn Lys Pro Ile Leu Phe Thr Met Ala Arg Leu Asp Arg
565 570 575
Val Lys Asn Leu Thr Gly Leu Val Glu Trp Tyr Ala Lys Asn Ala Arg
580 585 590
Leu Arg Glu Leu Val Asn Leu Val Val Val Gly Gly Asp Arg Arg Lys
595 600 605
Glu Ser Lys Asp Leu Glu Glu Gln Ala Glu Met Lys Lys Met Tyr Asp
610 615 620
Leu Ile Glu Thr Tyr Asn Leu Asn Gly Gln Phe Arg Trp Ile Ser Ser
625 630 635 640
Gln Met Asn Arg Val Arg Asn Gly Glu Leu Tyr Arg Tyr Ile Ala Asp
645 650 655
Thr Arg Gly Ala Phe Val Gln Pro Ala Phe Tyr Glu Ala Phe Gly Leu
660 665 670
Thr Val Val Glu Ser Met Thr Cys Gly Leu Pro Thr Phe Ala Thr Cys
675 680 685
Asn Gly Gly Pro Phe Glu Ile Ile Val Asn Gly Lys Ser Gly Phe His
690 695 700
Ile Asp Pro Asn Gln Gly Asp Lys Ala Ala Asp Met Leu Val Asn Phe
705 710 715 720
Phe Glu Lys Ser Lys Glu Asp Pro Ser Tyr Trp Asp Ala Ile Ser Lys
725 730 735
Gly Gly Leu Gln Arg Ile Leu Glu Lys Tyr Thr Trp Gln Ile Tyr Ser
740 745 750
Gln Lys Val Ile Thr Leu Ser Gly Ile Tyr Gly Phe Trp Lys Tyr Ala
755 760 765
Thr Lys Asn Asp Lys Val Ala Ser Ala Lys Lys Arg Tyr Leu Glu Met
770 775 780
Phe Tyr Glu Leu Gly Phe Lys Lys Ser Ala Glu Lys Val Pro Leu Ala
785 790 795 800
Ile Asp Glu
<210> 3
<211> 3926
<212> ДНК
<213> Nicotiana tabacum
<400> 3
atggcaggca gtggtcttag cattaaggaa agtttggagg aatccatttt ggctcatcca 60
gatgaaattt tggctctcaa gtcaaggtac attactgcat aatgatatta agacctagaa 120
gcggatccaa gattttgtta catttttgaa attataagtt tagaatctaa tatttgttat 180
cgcttgtttc cttattatct tgctgttgtt actgcctgtt gctactagtt tctgttcatc 240
cttccttgag ctgagtttct atcggaaaca acctctctac tctcaaagta ggaataagtt 300
atgcgtacac actaccctcc ccagactcca cttgtgtaat ttactgagtt tgttgttgtt 360
gttgttgtaa tctaatactt gttagaattt tactgatttt tcacatatat atctatgacc 420
catgtcgaaa atactatagc tcatgtgcta aatacattag taccattgtt ttgtaattgt 480
tttggttttg gaacaggatt gaaactgaag ggaaaggggt aatgaaacca gttgatctct 540
tgaaccattt ggtttctgtt actagtaaaa caaatggagt aaatgttgta cctagtgcac 600
ttgtggaagt tctcagttgc agccaagaag ctgtgattgt accaccaaaa ctagcactag 660
ctgtacgtcc gaggcccggt gtatgggagt acttgtcact gaatcttaag acaaagaaag 720
tggctgaatt gagcattcct gagtaccttc aattgaaaga gaatactgtt gatgaaaggt 780
aaagtaatag tctgcgattt cgctttgtga aattgaagtt ttttgtttga ttcttaatgt 840
tttgtgtatc aattatgtta ccagtggaaa catcttggag ttggattttg agccatttac 900
aactgttaca acaccaaaaa cactttctga ctctattggc aatggtttgg agtttcttaa 960
tcgccacatt gcttcgaaaa tgtttcttga taaggagatt gccaagtgcc tccttgactt 1020
tctcagaaac cataactaca aaggaaaggt agtaaaaaaa gtgtttcttt aaacaagttg 1080
tatgattatg tgtgtatttc taaatatgtc aatttgaaaa cagtcattga tggtgaaaga 1140
aagcattcaa agcctggaga gtttccaact tgttctgaaa aaagcagagg aatatttgca 1200
cacactgaat ccagaaactc catactccaa atttgaatcc aagtttgaag agattggctt 1260
ggaaagaggg tggggaaaca ccgctgaacg cgtgcaagac accattagtc atcttttgca 1320
tctccttgag gctcctaacg cgtcttcctt ggaaaatttc cttggtagaa tcccattggt 1380
tttcaatgtt gtgattctca ccccacatgg ttattttgct caagataatg tcttgggcta 1440
tcctgacact ggtggccagg tttgtgtccg atataacata tcaagaaatt ttgcattctt 1500
gatcatgttc tttataccat ttgaaccaac attctttttt tggttgtgaa atgttgaata 1560
ggttgtttac attcttgatc aagttccagc tatggagcgt gagatgcttc atcgtatgaa 1620
gcttcaagga ctcgacgata tcatccctcg catccttgtt gtaagtgccc ttaattttcc 1680
tggtttggtt tacctctaaa tgaaattgat tttctggctt tctaactttt ttggattgat 1740
ctttttgttg ttttatatca ggtaactagg ctgctgcctg atgctgtagg aaccacttgt 1800
ggcgagtgga tggagaaagt atatggggca gaacattctc atataattcg tgttccattt 1860
agaactgaga aaggaatgtt gcgcaaatgg atctcacgat tcgaagtctg gccatacatg 1920
gaaactttca ctgaggttgg aacataaaaa caaataaaaa tcattggaat gttcttctgc 1980
atttgaaaat gtcttgctaa ctaaagactc atttttaaat taatcatcag gatgttgcag 2040
aagaacttgt caaagaattg caagctaaac cagacttgat aattggaaac tacagtgagg 2100
gaaatcttgc tgcctcattg cttgctaaga aatttggggc tactcagtgt actattgctc 2160
atgccttgga aaaaactaag tatccaaact ctgaccttaa ttggaagaag tttgatgaca 2220
agtatcattt ctcaagtcag ttcactgctg atctttttgc catgaatcac actgatttca 2280
ttatcaccag cactttccaa gaaattgctg gaaggtaaaa gcaaatgcac accatcatag 2340
tatttcatat ttttacccta gtttatacta tttccatttg tcaactccaa cttgtttggg 2400
attgaaccat agttgttgtt tgtttatact atttccattc gccgacccca acttatttgg 2460
gactgagaca taattgttgt tattattgtt tgtttgttta tactatttcc attctcagac 2520
cccaacttct ttgggactga gccgtagatt gttgttgttg ttgttgttgt tgtttgttta 2580
tgctatttcc gttcaccgac cccaacttat ttgggactga ggtgtagaag tagtcgttgt 2640
tgtttgttta tacgacttcc aattgatatt cgaatgtttt tatttttgca gcaagaacac 2700
tgtaggacag tatgagagtc atactgcttt taccatgcct ggattgtatc gagtagtcca 2760
tggaatcaat tcgtttgatc caaagttcaa cattgtctcc cctggggctg atatgtcaat 2820
ctacttccct tacactgaga aggagaaaag actaaccaac ttccacccgg aaattgaaga 2880
actcctctac agtcctgttg agaataagga ccacttgtta gtcttcttta tttcattcat 2940
ttttctacac cttttttttc aacagattga ttgattggtt cttatcaacg taaacagatg 3000
tgtgttgaag gaccagaaca agccaattct ctttaccatg gcaaggctag atcgcgtgaa 3060
gaatctaaca gggctcgtgg aatggtatgc aaagaatgca aggctaaggg agctcgttaa 3120
ccttgtggtt gtaggcggag acagaaggaa agaatccaaa gatttagaag agcaagcaga 3180
gatgaagaag atgtatgatc ttatcgaaac atacaacctg aatggccaat tcaggtggat 3240
ttcttcccaa atgaatcgtg tgaggaacgg agaactttat cgatacattg cagacacgag 3300
gggtgctttc gttcaaccag cattttatga ggcatttggt ttgacagttg ttgagtctat 3360
gacttgtggt ttgccaactt ttgctacttg taatggtgga ccatttgaga ttatagtgaa 3420
tggaaaatct ggtttccata ttgatcctaa tcaaggtgac aaggctgctg atatgttggt 3480
taatttcttc gaaaaatcta aagaagatcc aagttattgg gatactattt ccaagggtgg 3540
tctgcagcgt attcttgaaa agtaagcttt tgcatttgat tagcacaagt gtacaaccaa 3600
gatttaactt atgaacaaac taaaactaac ccttttttta ttttcttttg ctaggtatac 3660
atggcaaatt tattcacaga aagtgatcac attatctggg atttatggat tctggaaata 3720
tgcaaccaag aatgacaaag ttgctagtgc gaagaagcgc tatcttgaaa tgttttatga 3780
atttgggttt aagaaatcag taagtgtcac ttctgtattt tgtttgagct tgtttgtaaa 3840
gtttggcaat cttctgctaa tttgtactat atttgttgac ttgtgcattt caggctgaga 3900
aagttccatt ggctattgat gaatag 3926
<210> 4
<211> 803
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 4
Met Ala Gly Ser Gly Leu Ser Ile Lys Glu Ser Leu Glu Glu Ser Ile
1 5 10 15
Leu Ala His Pro Asp Glu Ile Leu Ala Leu Lys Ser Arg Ile Glu Thr
20 25 30
Glu Gly Lys Gly Val Met Lys Pro Val Asp Leu Leu Asn His Leu Val
35 40 45
Ser Val Thr Ser Lys Thr Asn Gly Val Asn Val Val Pro Ser Ala Leu
50 55 60
Val Glu Val Leu Ser Cys Ser Gln Glu Ala Val Ile Val Pro Pro Lys
65 70 75 80
Leu Ala Leu Ala Val Arg Pro Arg Pro Gly Val Trp Glu Tyr Leu Ser
85 90 95
Leu Asn Leu Lys Thr Lys Lys Val Ala Glu Leu Ser Ile Pro Glu Tyr
100 105 110
Leu Gln Leu Lys Glu Asn Thr Val Asp Glu Ser Gly Asn Ile Leu Glu
115 120 125
Leu Asp Phe Glu Pro Phe Thr Thr Val Thr Thr Pro Lys Thr Leu Ser
130 135 140
Asp Ser Ile Gly Asn Gly Leu Glu Phe Leu Asn Arg His Ile Ala Ser
145 150 155 160
Lys Met Phe Leu Asp Lys Glu Ile Ala Lys Cys Leu Leu Asp Phe Leu
165 170 175
Arg Asn His Asn Tyr Lys Gly Lys Ser Leu Met Val Lys Glu Ser Ile
180 185 190
Gln Ser Leu Glu Ser Phe Gln Leu Val Leu Lys Lys Ala Glu Glu Tyr
195 200 205
Leu His Thr Leu Asn Pro Glu Thr Pro Tyr Ser Lys Phe Glu Ser Lys
210 215 220
Phe Glu Glu Ile Gly Leu Glu Arg Gly Trp Gly Asn Thr Ala Glu Arg
225 230 235 240
Val Gln Asp Thr Ile Ser His Leu Leu His Leu Leu Glu Ala Pro Asn
245 250 255
Ala Ser Ser Leu Glu Asn Phe Leu Gly Arg Ile Pro Leu Val Phe Asn
260 265 270
Val Val Ile Leu Thr Pro His Gly Tyr Phe Ala Gln Asp Asn Val Leu
275 280 285
Gly Tyr Pro Asp Thr Gly Gly Gln Val Val Tyr Ile Leu Asp Gln Val
290 295 300
Pro Ala Met Glu Arg Glu Met Leu His Arg Met Lys Leu Gln Gly Leu
305 310 315 320
Asp Asp Ile Ile Pro Arg Ile Leu Val Val Thr Arg Leu Leu Pro Asp
325 330 335
Ala Val Gly Thr Thr Cys Gly Glu Trp Met Glu Lys Val Tyr Gly Ala
340 345 350
Glu His Ser His Ile Ile Arg Val Pro Phe Arg Thr Glu Lys Gly Met
355 360 365
Leu Arg Lys Trp Ile Ser Arg Phe Glu Val Trp Pro Tyr Met Glu Thr
370 375 380
Phe Thr Glu Asp Val Ala Glu Glu Leu Val Lys Glu Leu Gln Ala Lys
385 390 395 400
Pro Asp Leu Ile Ile Gly Asn Tyr Ser Glu Gly Asn Leu Ala Ala Ser
405 410 415
Leu Leu Ala Lys Lys Phe Gly Ala Thr Gln Cys Thr Ile Ala His Ala
420 425 430
Leu Glu Lys Thr Lys Tyr Pro Asn Ser Asp Leu Asn Trp Lys Lys Phe
435 440 445
Asp Asp Lys Tyr His Phe Ser Ser Gln Phe Thr Ala Asp Leu Phe Ala
450 455 460
Met Asn His Thr Asp Phe Ile Ile Thr Ser Thr Phe Gln Glu Ile Ala
465 470 475 480
Gly Ser Lys Asn Thr Val Gly Gln Tyr Glu Ser His Thr Ala Phe Thr
485 490 495
Met Pro Gly Leu Tyr Arg Val Val His Gly Ile Asn Ser Phe Asp Pro
500 505 510
Lys Phe Asn Ile Val Ser Pro Gly Ala Asp Met Ser Ile Tyr Phe Pro
515 520 525
Tyr Thr Glu Lys Glu Lys Arg Leu Thr Asn Phe His Pro Glu Ile Glu
530 535 540
Glu Leu Leu Tyr Ser Pro Val Glu Asn Lys Asp His Leu Cys Val Leu
545 550 555 560
Lys Asp Gln Asn Lys Pro Ile Leu Phe Thr Met Ala Arg Leu Asp Arg
565 570 575
Val Lys Asn Leu Thr Gly Leu Val Glu Trp Tyr Ala Lys Asn Ala Arg
580 585 590
Leu Arg Glu Leu Val Asn Leu Val Val Val Gly Gly Asp Arg Arg Lys
595 600 605
Glu Ser Lys Asp Leu Glu Glu Gln Ala Glu Met Lys Lys Met Tyr Asp
610 615 620
Leu Ile Glu Thr Tyr Asn Leu Asn Gly Gln Phe Arg Trp Ile Ser Ser
625 630 635 640
Gln Met Asn Arg Val Arg Asn Gly Glu Leu Tyr Arg Tyr Ile Ala Asp
645 650 655
Thr Arg Gly Ala Phe Val Gln Pro Ala Phe Tyr Glu Ala Phe Gly Leu
660 665 670
Thr Val Val Glu Ser Met Thr Cys Gly Leu Pro Thr Phe Ala Thr Cys
675 680 685
Asn Gly Gly Pro Phe Glu Ile Ile Val Asn Gly Lys Ser Gly Phe His
690 695 700
Ile Asp Pro Asn Gln Gly Asp Lys Ala Ala Asp Met Leu Val Asn Phe
705 710 715 720
Phe Glu Lys Ser Lys Glu Asp Pro Ser Tyr Trp Asp Thr Ile Ser Lys
725 730 735
Gly Gly Leu Gln Arg Ile Leu Glu Lys Tyr Thr Trp Gln Ile Tyr Ser
740 745 750
Gln Lys Val Ile Thr Leu Ser Gly Ile Tyr Gly Phe Trp Lys Tyr Ala
755 760 765
Thr Lys Asn Asp Lys Val Ala Ser Ala Lys Lys Arg Tyr Leu Glu Met
770 775 780
Phe Tyr Glu Phe Gly Phe Lys Lys Ser Ala Glu Lys Val Pro Leu Ala
785 790 795 800
Ile Asp Glu
<210> 5
<211> 4532
<212> ДНК
<213> Nicotiana tabacum
<400> 5
atggctgaac gtgctctgac tcgtgttcac agccttcgtg aacgtcttga tgccactttg 60
gctgcacatc gcaatgagat attgctgttt ctttcaaggt attgcctaag tagtgttctt 120
gtttcctaca aaagattcag ttggtgttca aaaaacgata tgtgatttga tttatctgcc 180
taagtcttgg tagtcataat tatccggtac ctgtgctggt gcgagttagc tggttcggaa 240
actactctta tgaaaacgag agatttagtt ggtgttgtct gcaattctgt agtatggact 300
attaagcaga tagatcatgt ttgatatcga aaaggaatgt atatgtgatg ttacttgaac 360
tggttttggt tattacagga ttgaaagcca tggaaaaggg atcttgaaac ctcaccagct 420
attggctgag ttcgatgcaa ttcgccaaga tgacaaaaag aagctgaatg atcatgcatt 480
tgaagaactc ctgaaatcta ctcaggtaat tttgattttg gctaaatgtg ttaccaagct 540
gaatgatcat gcatttgagt ttgtgtccga ctactacaat gatatgttat accaggaagc 600
gattgttctg ccaccttggg ttgcacttgc cattcgtttg aggcctggtg tgtgggaata 660
tgtccgtgtg aatgttaatg ctctagtcgt tgaggagctg accgtccctg agtatttgca 720
ttttaaggaa gaacttgttg atggaacgta agttttagtc tcttatttga tactatgtta 780
gagaataggc agtggattca atttatcagt gttgtttttt acctaatgca gctccaatgg 840
aaatttcgtt ctcgagttgg attttgagcc cttcactgca tcctttccta aaccgaccct 900
caccaaatct attgggaatg gagttgaatt cctcaatagg cacctttctg cgaaaatgtt 960
ccatgacaag gaaagcatga ccccgcttct tgaatttctt cgggttcaca attataaggg 1020
caaggtaact ttgttattcc cattcatata tatgttcagt ttgtgcttat catgcgccca 1080
atgatgtatg aatatgtact aaaggataga tgtacgattt cgtttgcaga caatgatgct 1140
gaatgacaga atacagaatt taaccactct gcaaaatgtc ctaaggaagg cagaggaata 1200
ccttattatg cttccccctg aaactccatt ttccgaattc gaacacaagt tccaagaaat 1260
tggattggag aagggatggg gcgacactgc ggagcgcgtg ctagagatga tatgcatgct 1320
tcttgatcta cttgaggctc ccgactcctg tactcttgag aagttcctag ggagaattcc 1380
tatggtgttc aacgtggtta tcctttcccc ccatggatat ttcgcccagg aaaatgtctt 1440
gggttatccc gacactggtg gccaggtgca ttactttagt ctttgtccgt gagtctatgt 1500
tgctcagatc ctctacaatg ccactgtacc cgtgtaggat actccaaata taatgcattt 1560
ttggaggatc tgtcaccggt gcaatggcat tttggaggtc ggagcaacaa acaactgcta 1620
gtatgcttct aaagcttgct tccataaatg ctaaggtcct tcacccgtaa tgtgcaggtt 1680
gtctacatat tagatcaagt tccagccttg gagcgtgaaa tgcttaaacg cctaaaggag 1740
caaggacttg atataacacc gcgtattctt attgttagta tttcttgtac ttgtaattgc 1800
tgcggattac acaaaatttt ctctttattg gcaacttatc ttgatattat tcccaggtta 1860
ctcgtctgct gcctgatgca gttggaacaa cttgtggtca gcggcttgag aaggtgtatg 1920
gagccgagca ctcacatatt cttagggtcc cctttaggac cgagaagggc attgttcgca 1980
aatggatatc tcgctttgaa gtgtggccat acatggagac tttcactgag gtgacactaa 2040
gcttccttgt atttgtctat cttctaattg gtattaggaa caatttgcta attattaacg 2100
ctttggcttt tcgtacatca ggatgttgca aaagaacttg ctgcagaact gcaggccaag 2160
ccagatttga taattggcaa ctatagcgag ggaaatcttg tggcttcatt gctggctcac 2220
aagttaggcg taacgcaggt ctgtgttatt tttcacctct tataaatctg attgtatttc 2280
cattagtctg gaactaaaag tactaaaatt ttcttttctt cgctgtgtta tttgccttct 2340
gcagtgcacc attgcccatg cattggagaa aacaaagtat cctgattctg acatctactg 2400
gaaaaaattt gacgaaaaat accatttctc gtcccagttt accgctgatc ttattgcaat 2460
gaatcacacc gattttatca tcaccagcac tttccaggag atagcaggaa ggtataacat 2520
caattgctaa ttcggttgca gtaacatttt gttcgatttc ttccccttat gcttaaccta 2580
ataccctaat gaattttcca gcaaggacac tgtcggacag tacgagagtc accaggcatt 2640
cacaatgcct ggattgtaca gagtcgttca cggcattgat gtgttcgatc ccaaattcaa 2700
cattgtctca cctggagctg atataaacct gtatttccca tattccgaga aggaaaagag 2760
attgacagca cttcacccag aaattgagga gcttctgtac agtgatgttg agaacgagga 2820
acatctgtaa gtttctaact tactcgtacc gtcagtggca gagccagaat tttcattaaa 2880
atggggtcaa aatataaaga cataaattca caaagaagcc aaggggtgtc aatatgtagt 2940
ataaatatat taaaaaaatt acctagctac acaatgtaat tttccgacaa aggggtatcg 3000
gttgcacttc ttgaatacat gtggctctgc cactgggtac agttacaaag tcctgttacc 3060
tatgtagatg agcttgtgct gaacatgttg tgattttggt aggtgtgtgc taaaggacag 3120
gaataagcca atcttattca caatggcgag attggatcgt gtgaagaact taaccggact 3180
tgttgagtgg tacgccaaga acgcacggct aagggagttg gttaaccttg ttgtcgttgg 3240
tggagaccga aggaaggaat ccaaagattt ggaagagcaa gcagagatga agaagatgta 3300
tgagctaata aagactcaca acttaaatgg ccaattcaga tggatttctt cacagatgaa 3360
ccgagtaagg aacggcgaac tctaccgata cattgccgac actaggggag ctttcgtgca 3420
gcctgcattc tatgaggctt tcggtttgac tgttgttgag gccatgacct gtggtttgcc 3480
tacatttgca actaatcatg gcggtccagc tgagatcatc gttaacggaa aatccggctt 3540
ccatatcgat ccatatcacg gtgagcaagc tgctgatctg ctagctgatt tctttgagaa 3600
atgtaagacg gaaccttctc attgggaaac tatttcaacc ggtggcctga agcgcatcca 3660
agagaagtaa gcaactcttt cttgactcta gtcattcaaa ttaacttggg atttgaggca 3720
tagttgattg ataatttatc gcgtctctac tactatatac aggtacacgt ggcaaatcta 3780
ctcggagaga ttattgacgt tggctgctgt ttacggtttc tggaaacatg tttctaagct 3840
tgatcgtcta gaaatccgtc gatatctaga aatgttttat gctctcaaat accggaagat 3900
ggtgagttct tctgcttcct gctcttctca tagtgtttaa tatacacttg attgattgca 3960
ttcacttaga ctaagttgct cggacacggg tgtggatgtc cgacacgagt gcggatctag 4020
agttcagatc cttcaagatg taaattataa gattcgggga tatggatcct agtacggata 4080
cgggtgcgag aatccggcta aaaataattt taaaaaaaat tatctctaaa ttatgagata 4140
ttatgtggaa tacttacgta taacttgtaa agtgtagatt ttttttaatt ctcaagttgt 4200
agattagtaa atgattgatt tcctagataa gtatgctatt ttcttcaaat ttactcttct 4260
gatttcgaaa atcaaattgt atctcgtctc gaatttttcc gtccgttatg gtcaaagtac 4320
ccaaaatcgt ttgaccaaat cggtacggat cccataccca cacccacact agtgtcgtat 4380
tgacacgggt gccgcaccta aactgctatg tcggagcaac ttagcactta gagaatcatt 4440
gatgttaaat tttcttaatt cttgaatctg ctaatgaaga ttttatcttg gtttttgttt 4500
aggctgaagc tgttccattg gctgctgaat ga 4532
<210> 6
<211> 805
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 6
Met Ala Glu Arg Ala Leu Thr Arg Val His Ser Leu Arg Glu Arg Leu
1 5 10 15
Asp Ala Thr Leu Ala Ala His Arg Asn Glu Ile Leu Leu Phe Leu Ser
20 25 30
Arg Ile Glu Ser His Gly Lys Gly Ile Leu Lys Pro His Gln Leu Leu
35 40 45
Ala Glu Phe Asp Ala Ile Arg Gln Asp Asp Lys Lys Lys Leu Asn Asp
50 55 60
His Ala Phe Glu Glu Leu Leu Lys Ser Thr Gln Glu Ala Ile Val Leu
65 70 75 80
Pro Pro Trp Val Ala Leu Ala Ile Arg Leu Arg Pro Gly Val Trp Glu
85 90 95
Tyr Val Arg Val Asn Val Asn Ala Leu Val Val Glu Glu Leu Thr Val
100 105 110
Pro Glu Tyr Leu His Phe Lys Glu Glu Leu Val Asp Gly Thr Ser Asn
115 120 125
Gly Asn Phe Val Leu Glu Leu Asp Phe Glu Pro Phe Thr Ala Ser Phe
130 135 140
Pro Lys Pro Thr Leu Thr Lys Ser Ile Gly Asn Gly Val Glu Phe Leu
145 150 155 160
Asn Arg His Leu Ser Ala Lys Met Phe His Asp Lys Glu Ser Met Thr
165 170 175
Pro Leu Leu Glu Phe Leu Arg Val His Asn Tyr Lys Gly Lys Thr Met
180 185 190
Met Leu Asn Asp Arg Ile Gln Asn Leu Thr Thr Leu Gln Asn Val Leu
195 200 205
Arg Lys Ala Glu Glu Tyr Leu Ile Met Leu Pro Pro Glu Thr Pro Phe
210 215 220
Ser Glu Phe Glu His Lys Phe Gln Glu Ile Gly Leu Glu Lys Gly Trp
225 230 235 240
Gly Asp Thr Ala Glu Arg Val Leu Glu Met Ile Cys Met Leu Leu Asp
245 250 255
Leu Leu Glu Ala Pro Asp Ser Cys Thr Leu Glu Lys Phe Leu Gly Arg
260 265 270
Ile Pro Met Val Phe Asn Val Val Ile Leu Ser Pro His Gly Tyr Phe
275 280 285
Ala Gln Glu Asn Val Leu Gly Tyr Pro Asp Thr Gly Gly Gln Val Val
290 295 300
Tyr Ile Leu Asp Gln Val Pro Ala Leu Glu Arg Glu Met Leu Lys Arg
305 310 315 320
Leu Lys Glu Gln Gly Leu Asp Ile Thr Pro Arg Ile Leu Ile Val Thr
325 330 335
Arg Leu Leu Pro Asp Ala Val Gly Thr Thr Cys Gly Gln Arg Leu Glu
340 345 350
Lys Val Tyr Gly Ala Glu His Ser His Ile Leu Arg Val Pro Phe Arg
355 360 365
Thr Glu Lys Gly Ile Val Arg Lys Trp Ile Ser Arg Phe Glu Val Trp
370 375 380
Pro Tyr Met Glu Thr Phe Thr Glu Asp Val Ala Lys Glu Leu Ala Ala
385 390 395 400
Glu Leu Gln Ala Lys Pro Asp Leu Ile Ile Gly Asn Tyr Ser Glu Gly
405 410 415
Asn Leu Val Ala Ser Leu Leu Ala His Lys Leu Gly Val Thr Gln Cys
420 425 430
Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Pro Asp Ser Asp Ile
435 440 445
Tyr Trp Lys Lys Phe Asp Glu Lys Tyr His Phe Ser Ser Gln Phe Thr
450 455 460
Ala Asp Leu Ile Ala Met Asn His Thr Asp Phe Ile Ile Thr Ser Thr
465 470 475 480
Phe Gln Glu Ile Ala Gly Ser Lys Asp Thr Val Gly Gln Tyr Glu Ser
485 490 495
His Gln Ala Phe Thr Met Pro Gly Leu Tyr Arg Val Val His Gly Ile
500 505 510
Asp Val Phe Asp Pro Lys Phe Asn Ile Val Ser Pro Gly Ala Asp Ile
515 520 525
Asn Leu Tyr Phe Pro Tyr Ser Glu Lys Glu Lys Arg Leu Thr Ala Leu
530 535 540
His Pro Glu Ile Glu Glu Leu Leu Tyr Ser Asp Val Glu Asn Glu Glu
545 550 555 560
His Leu Cys Val Leu Lys Asp Arg Asn Lys Pro Ile Leu Phe Thr Met
565 570 575
Ala Arg Leu Asp Arg Val Lys Asn Leu Thr Gly Leu Val Glu Trp Tyr
580 585 590
Ala Lys Asn Ala Arg Leu Arg Glu Leu Val Asn Leu Val Val Val Gly
595 600 605
Gly Asp Arg Arg Lys Glu Ser Lys Asp Leu Glu Glu Gln Ala Glu Met
610 615 620
Lys Lys Met Tyr Glu Leu Ile Lys Thr His Asn Leu Asn Gly Gln Phe
625 630 635 640
Arg Trp Ile Ser Ser Gln Met Asn Arg Val Arg Asn Gly Glu Leu Tyr
645 650 655
Arg Tyr Ile Ala Asp Thr Arg Gly Ala Phe Val Gln Pro Ala Phe Tyr
660 665 670
Glu Ala Phe Gly Leu Thr Val Val Glu Ala Met Thr Cys Gly Leu Pro
675 680 685
Thr Phe Ala Thr Asn His Gly Gly Pro Ala Glu Ile Ile Val Asn Gly
690 695 700
Lys Ser Gly Phe His Ile Asp Pro Tyr His Gly Glu Gln Ala Ala Asp
705 710 715 720
Leu Leu Ala Asp Phe Phe Glu Lys Cys Lys Thr Glu Pro Ser His Trp
725 730 735
Glu Thr Ile Ser Thr Gly Gly Leu Lys Arg Ile Gln Glu Lys Tyr Thr
740 745 750
Trp Gln Ile Tyr Ser Glu Arg Leu Leu Thr Leu Ala Ala Val Tyr Gly
755 760 765
Phe Trp Lys His Val Ser Lys Leu Asp Arg Leu Glu Ile Arg Arg Tyr
770 775 780
Leu Glu Met Phe Tyr Ala Leu Lys Tyr Arg Lys Met Ala Glu Ala Val
785 790 795 800
Pro Leu Ala Ala Glu
805
<210> 7
<211> 5901
<212> ДНК
<213> Nicotiana tabacum
<400> 7
atgcttttta tgggagtaaa ttttatggcc ggtcattcaa ctttgtgttc attacgcaaa 60
agtcattttt cttggtgttt attacgcaag tcatttttct tttttttttg ttacgtaaaa 120
atcattcaac tatgtgttta ttatctaaaa ttcaattttt tttttccttt tgttacacaa 180
aaatcatttt actttactct atttatcaca aaagtcacct tggccagatt ttataatagg 240
cttttatctt ttgttacaca aaaattattt tactttactc tatttatcac aaaagtcacc 300
ttggccagat tttataatag gcttttatct tttgttacac aaaaattatt ttactttact 360
ctatttatca caaaagtcac cttggccaga ttttacaata cttttacctt aaaagactat 420
tatgcccttg acattataaa tcctctcatt tatataatac cttctatatg atacactata 480
taatatattt ttacctaggt attttactta taattaaaat aatattaaat tattttattt 540
atctatttta taatatattc atacatttaa ttttttcatg gcaaatcact ttgtttaatc 600
atatttaaac atgaacaaat tttaaatatc aaaaaaataa aaaaataaaa aaaatattta 660
tttgaaataa taacaaacag atttgtttaa caaatgatag ttttttttta tagtcaataa 720
aatttttaaa aaaattcaaa gatatttgtt tttaatatta atatttttaa agctttatct 780
gttaatatta tttatttgaa agtattaatc tgatgtgtca ttgtgttaaa tgtgagtatt 840
ttatttattg gattaatgag tatggcttgg ctgataaaaa gctttgattt tataattttc 900
attaaaaata ttttattaag ctagtacctg acaaatttaa tatcttgaaa attaacgtta 960
agaaaaaatt aaatataaaa atatattata aaaataataa ataaataata tcaagttatt 1020
ttaattataa ataaaataca tggttaaaaa tatattatat agcatataat atagaaggta 1080
ttacataaat gagatgattt aaagggcata atagactttt caggtgaatg atttgtaaaa 1140
tatggttaaa gtgattattg tgataattag agcatagtaa aataattttt atgtaacaaa 1200
agaaaaaaaa aatgactttt gggtaatgaa cataaatttg aataactttt acgtaacaaa 1260
agaataaaat aaattttgga taataaacat aaaattgaat gaccacctat aaaatttatt 1320
atttttttgg gctcttcttg atttgatttt ttagtttagc ctttgcagta atcttggttg 1380
tcacgcgtag cgttgtgctt tcgccacata agtatttagt agacttaatt aatgtcatta 1440
tatcggttgg tgtggtttta attacttaac tgtactatta tattaggtgg aaggtttgaa 1500
aatttatagt agtaacattc tagatcattg aaaatattgg tgtttcagtg actttttagt 1560
atgtcatttt cattttctaa gtggttgtac taatatagta tattaaaatt ttgattggtt 1620
gagaaacaat ctctctcacc tacacggtac gggtaaggta tgcgtatacg cttatcctcc 1680
ctacactcca tttgtgggac tattgttgtt attttggata agctgaggta tccatcttct 1740
actaactgca ctagtttatt ttttttgctg tttacagttg aaacaattgt ctgaggattt 1800
ctcacctgct gaatcaactg caatggctga acgtgtgctg actcgtgttc acagccttcg 1860
tgaacgtctt gatgctactt tggctgctca tcgcaatgag atattactgt ttctttcaag 1920
gtatagccaa agatagtatt cttgttaact aaaaaagatt cagttggtgt tcaaaaaacg 1980
atacgtttat ctgcctaagt cttggtagtc agaattatcc ggtacctatg ctggtgtgag 2040
ttagctggct aggaaaccac tcttatgaaa acaagagatt tagttagagt tgtctgtaat 2100
tctgtagtat ggactatgta tgtgatgcta tttgaactgg ttttggttat tataggattg 2160
aaagccatgg aaaagggatc ttgaaaccgc atcagctatt ggctgagttt gatgcaattc 2220
gccaagatga caaaaagaaa ctgaatgatc atgcatttga agaactcctg aagtccactc 2280
aggtaatatg gttttggcta tatttgtcgc caacgccaag ctcatatttt tatattattt 2340
tgagcttgtg tctgaatacg acgatgatat gttatactag gaagcaattg ttctgccacc 2400
ttgggttgca cttgcgattc gtttgaggcc tggtgtgtgg gaatatgtcc gtgtgaatgt 2460
caatgcgcta gtcgttgagg agctgactgt ccctgagtat ttgcatttca aggaagaact 2520
tgtcgatgga acgtaagtgt tagtcttcaa tttgatgcta tgttagagaa taggctgtgg 2580
aatttattga tcaatgctgt gctttgtcct gatacagctc caatggaaat ttcgttctcg 2640
agttggattt tgagcccttc accgcatcct ttcctaaacc aaccctcacc aaatctatcg 2700
gaaatggagt tgaattcctc aataggcacc tctctgcgaa aatgttccat gacaaggaaa 2760
gcatgacccc gcttcttgaa tttcttcggg ttcacaatta taagggcaag gtgacttgct 2820
atttccattt atctataggt tcggtttgtg cttatcatgc gcccaatgac atatgaatat 2880
gcgctaaagg atagatatat gatttccttt gcagacaatg atgctgaacg acagaataca 2940
gaatttaacc acactgcaaa atgtcctaag gaaggcagag gaatacctca ttatgcttcc 3000
ccctgaaact ccattttccg aattcgaaca caagttccaa gaaattggat tggagaaggg 3060
atggggcgac actgcagagc gcgtgctgga gatgatatgc atgcttcttg atctcctcga 3120
ggctcccgat tcctgtactc ttgagaagtt cttggggaga attcctatgg tgttcaatgt 3180
ggttatcctt tccccccacg gatatttcgc ccaggaaaat gtcttgggtt atcccgacac 3240
tggtggccag gtgcattact ttaatcttta tccgtgagtc tatgtttgtt cgaatcctct 3300
agaaatgtca ctgtacctat gtaggatact ccaaatataa tgcattttgg ggggatctgt 3360
tatgggtgcg atggcatttt tggaggtcgg agcaacaaac aattgctatg tattcttcta 3420
aagcttgctt tcataaatgc taaggtcctt cacccttaat gtgcaggttg tctatatatt 3480
agatcaagtt ccagccttgg agcgtgaaat gcttaagcgc ctaaaggagc aaggacttga 3540
tatcacaccg cgtattctta ttgttagtat ttcctgtact tgtaattact gcggattaca 3600
caaaatttcc tttttatctt cttaacaact tatcttgatg gtattcccag gttactcgtc 3660
tgctacctga tgcagttgga acgacttgtg gtcagcggct tgagaaggtg tatggagccg 3720
agcactcaca tattctgagg gtccccttta ggactgagaa gggcattgtt cgtaaatgga 3780
tctctcgctt tgaagtgtgg ccatatatgg agactttcac tgaggtgaca ctaaaacttc 3840
cttatatttg tctatcttct aattggtatt aggaataatt tgttaattgt taactctttg 3900
tcttttcgta catcaggatg tcgcaaaaga acttgctgca gaattgcagg ccaagccaga 3960
tttgataata ggcaactata gcgagggaaa tcttgtggct tcattgctcg ctcataagtt 4020
aggcgtaaca caggtctgtg ttgtttttca ctctcttaaa gatctgattg catttccatt 4080
agtctggaac tagaagtact aaaaagttct tttcttcact gtgttatttg ccgtcggcag 4140
tgcaccatag ctcatgcatt ggagaaaaca aagtatcctg attctgacat ctactggaaa 4200
aaattcgatg aaaaatacca tttctcgtcc cagtttaccg ctgatcttat tgcaatgaat 4260
cacaccgatt ttatcatcac cagcactttc caggagatag caggaaggta taacatcaat 4320
ttgctacttc gactgcaaca gcattgtgtt cccatttctt tcccttatgc ttaacctaat 4380
accgtcatga attttccagc aaggacactg tcggacagta cgagagtcat caggcattca 4440
caatgcccgg attgtacaga gttgttcacg gcattgatgt gttcgacccc aaattcaaca 4500
ttgtctcacc tggagctgac ataaacctct atttcccata ttccgagaag gaaaagagac 4560
tgacagcact tcaccctgaa atcgaggagc tgctgtacag tgacattgag aacgaggaac 4620
atctgtaagt ttctacctta ctcgtacagt cagtggcgga gccagaattt tcactaaaat 4680
aaggtcaaaa tataaagaca taaatccaca aagaagccaa gggtgtcaat atatagtata 4740
aatacattaa aaaaattacc tatctacaca gtgtaatttt ccgacaaagg ggtgtcggtt 4800
gacactcctt gaatacatgt ggctctgcca ctgggtacag ttacaaagtt ctgttaccta 4860
tgtagatgag cttgtgctga acatgttgtg attttggcag gtgtgtgcta aaggacagga 4920
ataagccaat cttattcaca atggcgagat tggatcgtgt gaagaattta accggacttg 4980
ttgagtggta tgccaagaac gcacggctaa gggagttggt taaccttgtt gtggttggtg 5040
gagatcgaag gaaagaatcc aaagatttgg aagagcaaac agaaatgaaa aagatgtatg 5100
agctaataaa gactcacaat ttaaatggcc aattcagatg gatttcttca cagatgaacc 5160
gagtgaggaa cggtgaactc taccgataca ttgctgacac tagaggagct ttcgtgcagc 5220
ctgcattcta cgaggctttc ggtttgactg ttgttgaggc catgacctgt ggtttgccta 5280
catttgcaac taatcatggc ggtccagctg agatcatcgt taacggaaaa tctggcttcc 5340
acatcgatcc atatcacggt gagcaagctg ctgatctgct agctgatttc tttgagaaat 5400
gtaagacaga accttctcat tgggaaacca tttcaacggg tggcctgaag cgcatccaag 5460
agaagtaagc aactctttct tgactctagt cattgaaatt aactttcttg actctagtca 5520
ttgaaattaa ctcgggattt gaggcgtagt tgattgatat tttatcgcgt ctctactact 5580
gatatataca ggtacacgtg gcaaatctac tcggagaggc tattgacatt ggctgctgtt 5640
tacgggttct ggaaacatgt ttctaagctt gatcgtctag aaatccgtcg atatcttgaa 5700
atgttttatg ctctcaaata ccgcaagatg gtgagttcct cttcttcctt gcccttctcc 5760
tagtgtttaa gatacaatat aattgattgc attatcttag agaatcatta atgttaaatt 5820
ttcttaattc ttgaatctgt taatgaagtt tttctcttgg tttttgttta ggctgaagct 5880
gttccattgg ctgctgagtg a 5901
<210> 8
<211> 825
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 8
Met Leu Phe Met Gly Leu Lys Gln Leu Ser Glu Asp Phe Ser Pro Ala
1 5 10 15
Glu Ser Thr Ala Met Ala Glu Arg Val Leu Thr Arg Val His Ser Leu
20 25 30
Arg Glu Arg Leu Asp Ala Thr Leu Ala Ala His Arg Asn Glu Ile Leu
35 40 45
Leu Phe Leu Ser Arg Ile Glu Ser His Gly Lys Gly Ile Leu Lys Pro
50 55 60
His Gln Leu Leu Ala Glu Phe Asp Ala Ile Arg Gln Asp Asp Lys Lys
65 70 75 80
Lys Leu Asn Asp His Ala Phe Glu Glu Leu Leu Lys Ser Thr Gln Glu
85 90 95
Ala Ile Val Leu Pro Pro Trp Val Ala Leu Ala Ile Arg Leu Arg Pro
100 105 110
Gly Val Trp Glu Tyr Val Arg Val Asn Val Asn Ala Leu Val Val Glu
115 120 125
Glu Leu Thr Val Pro Glu Tyr Leu His Phe Lys Glu Glu Leu Val Asp
130 135 140
Gly Thr Ser Asn Gly Asn Phe Val Leu Glu Leu Asp Phe Glu Pro Phe
145 150 155 160
Thr Ala Ser Phe Pro Lys Pro Thr Leu Thr Lys Ser Ile Gly Asn Gly
165 170 175
Val Glu Phe Leu Asn Arg His Leu Ser Ala Lys Met Phe His Asp Lys
180 185 190
Glu Ser Met Thr Pro Leu Leu Glu Phe Leu Arg Val His Asn Tyr Lys
195 200 205
Gly Lys Thr Met Met Leu Asn Asp Arg Ile Gln Asn Leu Thr Thr Leu
210 215 220
Gln Asn Val Leu Arg Lys Ala Glu Glu Tyr Leu Ile Met Leu Pro Pro
225 230 235 240
Glu Thr Pro Phe Ser Glu Phe Glu His Lys Phe Gln Glu Ile Gly Leu
245 250 255
Glu Lys Gly Trp Gly Asp Thr Ala Glu Arg Val Leu Glu Met Ile Cys
260 265 270
Met Leu Leu Asp Leu Leu Glu Ala Pro Asp Ser Cys Thr Leu Glu Lys
275 280 285
Phe Leu Gly Arg Ile Pro Met Val Phe Asn Val Val Ile Leu Ser Pro
290 295 300
His Gly Tyr Phe Ala Gln Glu Asn Val Leu Gly Tyr Pro Asp Thr Gly
305 310 315 320
Gly Gln Val Val Tyr Ile Leu Asp Gln Val Pro Ala Leu Glu Arg Glu
325 330 335
Met Leu Lys Arg Leu Lys Glu Gln Gly Leu Asp Ile Thr Pro Arg Ile
340 345 350
Leu Ile Val Thr Arg Leu Leu Pro Asp Ala Val Gly Thr Thr Cys Gly
355 360 365
Gln Arg Leu Glu Lys Val Tyr Gly Ala Glu His Ser His Ile Leu Arg
370 375 380
Val Pro Phe Arg Thr Glu Lys Gly Ile Val Arg Lys Trp Ile Ser Arg
385 390 395 400
Phe Glu Val Trp Pro Tyr Met Glu Thr Phe Thr Glu Asp Val Ala Lys
405 410 415
Glu Leu Ala Ala Glu Leu Gln Ala Lys Pro Asp Leu Ile Ile Gly Asn
420 425 430
Tyr Ser Glu Gly Asn Leu Val Ala Ser Leu Leu Ala His Lys Leu Gly
435 440 445
Val Thr Gln Cys Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Pro
450 455 460
Asp Ser Asp Ile Tyr Trp Lys Lys Phe Asp Glu Lys Tyr His Phe Ser
465 470 475 480
Ser Gln Phe Thr Ala Asp Leu Ile Ala Met Asn His Thr Asp Phe Ile
485 490 495
Ile Thr Ser Thr Phe Gln Glu Ile Ala Gly Ser Lys Asp Thr Val Gly
500 505 510
Gln Tyr Glu Ser His Gln Ala Phe Thr Met Pro Gly Leu Tyr Arg Val
515 520 525
Val His Gly Ile Asp Val Phe Asp Pro Lys Phe Asn Ile Val Ser Pro
530 535 540
Gly Ala Asp Ile Asn Leu Tyr Phe Pro Tyr Ser Glu Lys Glu Lys Arg
545 550 555 560
Leu Thr Ala Leu His Pro Glu Ile Glu Glu Leu Leu Tyr Ser Asp Ile
565 570 575
Glu Asn Glu Glu His Leu Cys Val Leu Lys Asp Arg Asn Lys Pro Ile
580 585 590
Leu Phe Thr Met Ala Arg Leu Asp Arg Val Lys Asn Leu Thr Gly Leu
595 600 605
Val Glu Trp Tyr Ala Lys Asn Ala Arg Leu Arg Glu Leu Val Asn Leu
610 615 620
Val Val Val Gly Gly Asp Arg Arg Lys Glu Ser Lys Asp Leu Glu Glu
625 630 635 640
Gln Thr Glu Met Lys Lys Met Tyr Glu Leu Ile Lys Thr His Asn Leu
645 650 655
Asn Gly Gln Phe Arg Trp Ile Ser Ser Gln Met Asn Arg Val Arg Asn
660 665 670
Gly Glu Leu Tyr Arg Tyr Ile Ala Asp Thr Arg Gly Ala Phe Val Gln
675 680 685
Pro Ala Phe Tyr Glu Ala Phe Gly Leu Thr Val Val Glu Ala Met Thr
690 695 700
Cys Gly Leu Pro Thr Phe Ala Thr Asn His Gly Gly Pro Ala Glu Ile
705 710 715 720
Ile Val Asn Gly Lys Ser Gly Phe His Ile Asp Pro Tyr His Gly Glu
725 730 735
Gln Ala Ala Asp Leu Leu Ala Asp Phe Phe Glu Lys Cys Lys Thr Glu
740 745 750
Pro Ser His Trp Glu Thr Ile Ser Thr Gly Gly Leu Lys Arg Ile Gln
755 760 765
Glu Lys Tyr Thr Trp Gln Ile Tyr Ser Glu Arg Leu Leu Thr Leu Ala
770 775 780
Ala Val Tyr Gly Phe Trp Lys His Val Ser Lys Leu Asp Arg Leu Glu
785 790 795 800
Ile Arg Arg Tyr Leu Glu Met Phe Tyr Ala Leu Lys Tyr Arg Lys Met
805 810 815
Ala Glu Ala Val Pro Leu Ala Ala Glu
820 825
<210> 9
<211> 8323
<212> ДНК
<213> Nicotiana tabacum
<400> 9
atggcgaatc caaagttcac aagagtacct agcatgaggg agagagttga ggatactctc 60
tctgctcacc gtaaccagct tgttgctctc ctctccaggt atattaataa actctatata 120
cttgttattt tctttatttt tttgtcttta ctgataaatt taactgtttt cttctttaaa 180
tcttgctttc gatgcatgat ttctgttgtg ttaaattgcg taaccatttt atctaaaagt 240
ttatgctgat aaacactttt aaattttaat atgtaaatta tattatgtct caacatcaac 300
atgtggatgg ccaaaaatat aaagcttaat tttcgttatt ttgaatgatt tttctctgcg 360
agtgttacgg tttgcgtaca cattacctaa acctcctccc tagtccccac ttgtgggaat 420
ttaatttttt ttttctttgt ttttttttgt tgttgttgtt gtctgagttc aattcctacc 480
atgttagctt ggcaaaaata agttggtaaa gcttgacccc aactagtttt agttgatcga 540
tttatttggt gatttatagt tcaataataa taattactat tagagaaagt tccagcagct 600
tttctgtttg tttttccagt tttagtgatt gatatatgtg tatatatatt ctttgtttct 660
tttaagatac gtggcgcagg ggaaggggat attgcaacct caccacttga tcgatgagtt 720
caacaacgct gtatgtgatg acactgcttg tgagaagctc aaagatggtc cctttagtga 780
agtcttgaaa gctactcagg tatattcact aatccatggg aatcaagatg atactgtata 840
tctttattat ggtgtctttc agaaatttga cgatgatgaa atgcaacttt tctctgtttg 900
tcaccttatc cagactgttt ttttattttt tatttttcat tttttaactt gaaatgctct 960
taatttcctt tgtttatcga taagaccgga tttacaatgt atgaacggag catcttaaga 1020
accttctgga atgaagatat aagatataaa acatggtgtc cgttttctcc tttgtggaat 1080
cagtgtacat atagactgtt attttggtcc cactttctgg atcttctgat cacaccttct 1140
catgcagagg cgagcttgat ggtttcaacc tttaaattct tactattgaa tccatttcac 1200
tttcgaaatt atgagttcga aatctaatat ttgttgaaat ttttgcaaat gttcacatat 1260
aagtttaagc tttgtgtcaa gaatactggg ctcaatggat tccaatagac caggctgtat 1320
ccgcctctgt ctccactctc cctgcatcca cttctttcgt gtgactaata atgcttaatg 1380
agctagaact cgttttaatg tttgaataag ttgcttatat cagagcagct tttgatgttt 1440
caatctttaa cgggttatgc agtaccagca ttctgcggct gaaaaacagg aatctgagat 1500
ttacttgtct ctggctgaat ttcttgttca ttttgctaac aagtactttg gagttaatgc 1560
ttgctctctg ttgtcaaaat aggaagccat tgtgctgcca ccatttgttg ccatagcagt 1620
tcgtccaagg ccaggtgttt gggagtatgt tcgtgttaat gtatatgatt tgagcgttga 1680
acaattgact gttcctgaat atcttcattt caaggaagaa cttgtggatg gagagtaagc 1740
tctttcttat ttcaatacga aacataaaaa tttacagaag ttgaataatt aacaaatttg 1800
ttgattttta atgtatgcca ggggtaataa tcactttgtg cttgagctgg attttgagcc 1860
atttaatgca tcagttcctc gtccatctcg atcgtcatcc attggcaatg gagtccaatt 1920
cctcaatcgt catctttcct caattatgtt tcgcagcaaa gactctctgg accccttact 1980
tgatttcctt agaggacact gtcataaagg gaatgtaagt accaaaagca gttttccctt 2040
tgtaaatgtc tgcttgtccc tgattatcta ctaaatcttt caacacgcgc aaccattata 2100
agaaatgtac aatacttcta gttagaattt catcatcgac aaactatctg ctttactttt 2160
tatttttccc atttgatgga tgatagttta gtttatataa cagatgatat tttggttgaa 2220
gggtaccatg aactttttca caaccactta atggatacat agttgtaata gttgacattt 2280
tggaataata ttgtctcact tggaaatgtt taagaagtat tactacttct atttgtaaga 2340
tggattgttt atctatgcag gtcttgatgt tgaatgatcg tatacagcga atctccaggc 2400
tggagtctgc tctttctaaa gcagaggatt atctctccaa gctatcacca gatacatcct 2460
ataatgagtt cgaatacgcg tgagcttgta cacatttgtt ttgttttctt tcaagcatat 2520
gtaatttctc aagaaaaggg aaatctatag gagttgaaac attctttatg gaaccatgtg 2580
catgcagatt gcaagaaatg ggctttgaga gaggttgggg tgatactgcc agacgtgttt 2640
tggagacgat gcatcttctt tctgacattc ttcaggctcc ggatccatca accttggaga 2700
catttcttgg tagactacct atggtgttca atgtcgtcat attatcccct catggatatt 2760
ttggccaagc aaatgtcttg ggtttgcccg acactggtgg ccaggtaata acaaggagaa 2820
tgaggtcttg tattatgtac tccctccgtt ccaatctata tgaacctatt tgactgggta 2880
tggaaagaaa tgaagacttg taaaacttgt ggttctttag aaattccaaa cattacattt 2940
ggttttttcc ctcttcctgg aaattatact actgaatcat ctctagatgt tccagtttaa 3000
cttgagacgt aagggtaaat aacggaccat tactctgtcc tttcttgcag taggcttggt 3060
acaatgaata tagttcgcat agttgccgga agctagagct gtgttagaaa actcaggaac 3120
attaatttgg cgatgctaat cactgctaat gttactgaag catccatggt tttccttgat 3180
gttattctcc ttttggttgc ttcacaggtt gtctatatac tggatcaagt gcgtgccttg 3240
gaggccgaaa tgcttcttag aataaagcaa caaggactta acttcaagcc tagaatcctt 3300
gtcgtgagta catatatatt atgcaagctc ttatttggtt tgtgggattg cagttgacat 3360
caatttgctt actctgatta ctaaaggtca cacggctgat acctgatgct aaaggaacca 3420
tgtgcaacca gaggttggag aggattagtg gaactgaata ctcgcatatt ttacgtgtcc 3480
cttttaggac agagaaggga atccttcata aatggatatc taggtttgat gtatggcctt 3540
acctggagaa gttcactgag gtaacctctt tgtcccttgg aaattgcctt ttgttgctga 3600
tgtttctgct agtgtgctta aatgacggat gttaactagt cacttgctag cgtttgcaat 3660
agcaacggga aaagaaagga tttttgctag tttgaagtct gcctccaaga aaaattatat 3720
taaaagttta tggctagtgg aaacatcagt cattcatgta ccttatttct atgcccaagt 3780
tgtttaagtt gaaagtaatt tggccaacta tgcaaattgg gagaacgtgt agccaactat 3840
tgtgtttgcc gacatgttga tatacttttt ggtcctgatt tatatttgtt ggtttgtcat 3900
actggatgaa gcaattctca tgtttttctg cttatatata ttggaagaag agatacttgt 3960
cgtttcatca tttttctcga cctctctatt accaacactt tgccaattta atgtttggaa 4020
atgtcttctt gaccaggatg tggcaagtga aatgaccgct gagctccagg gaaagccaga 4080
tctgattatt ggcaactaca gtgatggaaa tttagttgcc tcccttttgg catataaaat 4140
gggtgtcaca caggtaggaa atacatgatt ctttatcttg ctagcactaa gtcttgaggt 4200
tatgtatctg caatagaaat tttacgcttt gccttcattt ctttttaatt atttttccag 4260
tgtaccattg ctcatgcctt ggaaaaaaca aagtatcctg attctgacat ctactggaaa 4320
aagtttgagg agaaatatca tttttcatgt cagtttactg ctgatctact ggcaatgaat 4380
aattcagatt tcattatcac cagtacttat caagagattg caggaacgta agtcatttta 4440
atctggtcgt ttaaatctga tatttcttcc ctagtagtct attcaatccg aatttcagtt 4500
cagtatatga tgtcatcggt tgaggaactg tgattggtaa ccttatcaaa tccgtagctg 4560
ctctataatt ttatttcgta attggagaaa caatttttta ttattgagct tgtagtctga 4620
gctagaattt ggttctttat ctatcaagta gcataatact acaactattt tttatgtgtg 4680
gcaatttgca atttcaattt tctatttcta taagttgcag cttttcttcc tgttctgatc 4740
atatttacat ggctgaaact caatagaaaa ctaggctagt tgatcaaaag tagttggatg 4800
ctttaaaatt agtagacgtt ttgctaaatg agtgaccaat gttattaaaa aaacgttcat 4860
gttttcaacc cttttggcat acatttgacc actgcccaag attttggata agtacatgca 4920
gtgcttataa ttataaagca ttttatccca ccttgttttt cattatgaaa attaagtaat 4980
ttacgagtat ttgtataagt tacttcataa attagaagta aatctggatt gtgtaaagtt 5040
attcgccccg tatatactga aagctacttg aacaagcaaa aaaacagaca aacgtaacat 5100
tctccatgga ttaatgagac ttgtatatat atatatatat atatgtaaag agagagagag 5160
agagatttgg cttgtaacca catgtatatt atgccatatg gatgtgacat tgatgtgact 5220
agacctaaat gttttgtttc aatgtccacg ggagttttac gtagagttaa gaggagaaga 5280
gagtgaggaa tactaatgtt tgatggtacc ccttggcttc ttgacctgga tactcagtgt 5340
tcttattcat gcctatactt tggtccttga tttcattctc ccttttctag cttgagctgc 5400
atcaaagaaa ttccactgta aaaaaaataa tgctcaccat attggtgcaa catggcaaac 5460
atgtatccta tttgatgatc aatcaacttt atttttctcc tgttaattga cctcagtgtg 5520
taactctcta tgtatgatag cattgtaact tgtgtcatga ttcataaata gggtactaga 5580
attggatggt tgacatagta aatggtcaat tgatgatcca caaaatatgc acctactgat 5640
taaaatgtga tagggcaggt ttatttttgt ttgtggttaa cacagtactt aaccctatat 5700
ttaatacaat ttggcttatc tacaatcttt tcttcagtgt ttatgcgaat tccttattgc 5760
acaacaatat tgtctttctg agttctattc tgttgttgct tacactttta ttattccagt 5820
aacatagatg tgaagacatt agattggttg cttgcaaatt gatagccact tgtttcagga 5880
agaatactgt tggtcagtac gagagccata ctgcattcac cctcccggga ctatatcgcg 5940
tcgttcatgg cattgatgtt ttcgatccca aattcaatat agtgtctcct ggagctgaca 6000
tgacaattta tttcccatat tctgacaagg aaaaaagact aacgtctttg catggctcga 6060
ttgaaaagtt gttatttgat cctgcgcaga atgaagagca tatgtaagtg gcatccgttt 6120
gtacttaatt tttttggaat agatgacata ttatttgcat gaatatgaaa aggagggtct 6180
gatatgattt tctatagata aactaccaat gatattattt aaaaactcct ggatactgta 6240
ttaggagaag aagagaacca ggggtagatg gcattagaat cccttaaatc ttgaagagtc 6300
gtcactaacg ctcccaacac ttctgcctca gaccctcaac taaatactat tattgttgat 6360
ttctttggag aagctataag aatctctctc tccttatggt gaaaatttta cttggcttta 6420
tacttaactt ccaaggctcc ctcttataaa atgcaaaaac tgtctgtatt cactctcttg 6480
gttaacaatt gatccaatca aatgcatatg gaacatcttt ctttacgttt cttctaaagt 6540
tcgtttgagg ataaggagta gaatctgaga agatagacta gtaggtaacc ttagggacgg 6600
atgtggaaat taacatatgg gctcagcttt tctgccgagt gcagaccatg tatatgcgtt 6660
aaaaaattca ctaaacaagt aaatgtttga ttttgaaccc agtaaatcaa atgagttgtg 6720
gtagaatctc gaactcgaac cgataaagtt caaatccagg atccgctttt aggtaaactc 6780
taccttggga agtgttatat atatgtccct gattatttct ttttccgttt cctttctatt 6840
ttaattttta aagttatttt tagatggttt tattttttga taagtggtaa gttgttaata 6900
ttccaaatta aatgccattg tcataactat atacatttat aaagaatgat tgatcctagt 6960
ttctcattcc taagatccaa ataaggcaat aaacaatgtc ttagtaattg gacctgcttc 7020
tggtgatcaa cgcttgatcg cgtagttagt tatagatgac tgtaaaaact ttaaccattt 7080
taatggtttt gtcaaagaac aaatatcgga catattatag agaatggact attgtacttt 7140
gcttctgatt ggtcatttta ttgtgatccg taaattggct gtgactgatg tcatatcttt 7200
gcttacagag gtaatctgaa tgataaatca aaacccataa ttttttcaat ggcaaggcta 7260
gaccatgtta agaacattac gggactagtt gagtgctatg ctaaaaatgc cacattgagg 7320
gaattggcga accttgttgt agtagctgga tacaacgatg taaagaaatc cagtgataga 7380
gaagaaataa cagaaattga gaagatgcat gctcttatta aggagcataa attggatggg 7440
caattcagat gggtatcagc ccaaacaaac cgggcacgta atggtgagct ctatcgctat 7500
atagctgacc agagaggtat atttgttcag gtatgctatt tgtattgtat tagtccaatt 7560
tcattttttg caccaaaaga aaggttgtta ttgtgacgta tatgtttgtt ttagcctgca 7620
ttttatgaag catttggact aacggtggtt gaagctatga cttgtggtct tccaacattt 7680
gcaacttgcc atggtggtcc taatgagatc attgaacccg gtgtatctgg gttccatatt 7740
gatccttatc atcccgataa agctgctgaa ctcatgtcag aattctttca acgctgcaaa 7800
caagatccta ctcactggga aaaaatatct gcatctggtc tccgaaggat tcttgagagg 7860
tctgtagttg tgtacatgta tagaagatta aagaatgcta ccttgatatt tatttgaatc 7920
aaaaataaca ggaacatctc ttttttgaac atcactcaag ttcttatatt aaataatttt 7980
taggtatacg tggaagattt actccgagag gctgatgact ttatctggcg tatatggttt 8040
ctggaagctt gtttcaaaac ttgagaggcg tgaaactaga cgataccttg agatgttcta 8100
cattctcaaa ttccgcgagt tggtgagtgc cttttagctc cttttcagtt ccaataaact 8160
atatatgtgg tttaagtaag tattaagcat aaacatgtcc gtgcttgggg ctgtcgaaaa 8220
tgctatggac atatcctgag ctaaggattt ttcaagaaaa ttgatgttag ctttactcta 8280
tttacaggca aaatctgtac ctctagcaat tgatgacaag tga 8323
<210> 10
<211> 810
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 10
Met Ala Asn Pro Lys Phe Thr Arg Val Pro Ser Met Arg Glu Arg Val
1 5 10 15
Glu Asp Thr Leu Ser Ala His Arg Asn Gln Leu Val Ala Leu Leu Ser
20 25 30
Arg Tyr Val Ala Gln Gly Lys Gly Ile Leu Gln Pro His His Leu Ile
35 40 45
Asp Glu Phe Asn Asn Ala Val Cys Asp Asp Thr Ala Cys Glu Lys Leu
50 55 60
Lys Asp Gly Pro Phe Ser Glu Val Leu Lys Ala Thr Gln Glu Ala Ile
65 70 75 80
Val Leu Pro Pro Phe Val Ala Ile Ala Val Arg Pro Arg Pro Gly Val
85 90 95
Trp Glu Tyr Val Arg Val Asn Val Tyr Asp Leu Ser Val Glu Gln Leu
100 105 110
Thr Val Pro Glu Tyr Leu His Phe Lys Glu Glu Leu Val Asp Gly Glu
115 120 125
Gly Asn Asn His Phe Val Leu Glu Leu Asp Phe Glu Pro Phe Asn Ala
130 135 140
Ser Val Pro Arg Pro Ser Arg Ser Ser Ser Ile Gly Asn Gly Val Gln
145 150 155 160
Phe Leu Asn Arg His Leu Ser Ser Ile Met Phe Arg Ser Lys Asp Ser
165 170 175
Leu Asp Pro Leu Leu Asp Phe Leu Arg Gly His Cys His Lys Gly Asn
180 185 190
Val Leu Met Leu Asn Asp Arg Ile Gln Arg Ile Ser Arg Leu Glu Ser
195 200 205
Ala Leu Ser Lys Ala Glu Asp Tyr Leu Ser Lys Leu Ser Pro Asp Thr
210 215 220
Ser Tyr Asn Glu Phe Glu Tyr Ala Leu Gln Glu Met Gly Phe Glu Arg
225 230 235 240
Gly Trp Gly Asp Thr Ala Arg Arg Val Leu Glu Thr Met His Leu Leu
245 250 255
Ser Asp Ile Leu Gln Ala Pro Asp Pro Ser Thr Leu Glu Thr Phe Leu
260 265 270
Gly Arg Leu Pro Met Val Phe Asn Val Val Ile Leu Ser Pro His Gly
275 280 285
Tyr Phe Gly Gln Ala Asn Val Leu Gly Leu Pro Asp Thr Gly Gly Gln
290 295 300
Val Val Tyr Ile Leu Asp Gln Val Arg Ala Leu Glu Ala Glu Met Leu
305 310 315 320
Leu Arg Ile Lys Gln Gln Gly Leu Asn Phe Lys Pro Arg Ile Leu Val
325 330 335
Val Thr Arg Leu Ile Pro Asp Ala Lys Gly Thr Met Cys Asn Gln Arg
340 345 350
Leu Glu Arg Ile Ser Gly Thr Glu Tyr Ser His Ile Leu Arg Val Pro
355 360 365
Phe Arg Thr Glu Lys Gly Ile Leu His Lys Trp Ile Ser Arg Phe Asp
370 375 380
Val Trp Pro Tyr Leu Glu Lys Phe Thr Glu Asp Val Ala Ser Glu Met
385 390 395 400
Thr Ala Glu Leu Gln Gly Lys Pro Asp Leu Ile Ile Gly Asn Tyr Ser
405 410 415
Asp Gly Asn Leu Val Ala Ser Leu Leu Ala Tyr Lys Met Gly Val Thr
420 425 430
Gln Cys Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Pro Asp Ser
435 440 445
Asp Ile Tyr Trp Lys Lys Phe Glu Glu Lys Tyr His Phe Ser Cys Gln
450 455 460
Phe Thr Ala Asp Leu Leu Ala Met Asn Asn Ser Asp Phe Ile Ile Thr
465 470 475 480
Ser Thr Tyr Gln Glu Ile Ala Gly Thr Lys Asn Thr Val Gly Gln Tyr
485 490 495
Glu Ser His Thr Ala Phe Thr Leu Pro Gly Leu Tyr Arg Val Val His
500 505 510
Gly Ile Asp Val Phe Asp Pro Lys Phe Asn Ile Val Ser Pro Gly Ala
515 520 525
Asp Met Thr Ile Tyr Phe Pro Tyr Ser Asp Lys Glu Lys Arg Leu Thr
530 535 540
Ser Leu His Gly Ser Ile Glu Lys Leu Leu Phe Asp Pro Ala Gln Asn
545 550 555 560
Glu Glu His Ile Gly Asn Leu Asn Asp Lys Ser Lys Pro Ile Ile Phe
565 570 575
Ser Met Ala Arg Leu Asp His Val Lys Asn Ile Thr Gly Leu Val Glu
580 585 590
Cys Tyr Ala Lys Asn Ala Thr Leu Arg Glu Leu Ala Asn Leu Val Val
595 600 605
Val Ala Gly Tyr Asn Asp Val Lys Lys Ser Ser Asp Arg Glu Glu Ile
610 615 620
Thr Glu Ile Glu Lys Met His Ala Leu Ile Lys Glu His Lys Leu Asp
625 630 635 640
Gly Gln Phe Arg Trp Val Ser Ala Gln Thr Asn Arg Ala Arg Asn Gly
645 650 655
Glu Leu Tyr Arg Tyr Ile Ala Asp Gln Arg Gly Ile Phe Val Gln Pro
660 665 670
Ala Phe Tyr Glu Ala Phe Gly Leu Thr Val Val Glu Ala Met Thr Cys
675 680 685
Gly Leu Pro Thr Phe Ala Thr Cys His Gly Gly Pro Asn Glu Ile Ile
690 695 700
Glu Pro Gly Val Ser Gly Phe His Ile Asp Pro Tyr His Pro Asp Lys
705 710 715 720
Ala Ala Glu Leu Met Ser Glu Phe Phe Gln Arg Cys Lys Gln Asp Pro
725 730 735
Thr His Trp Glu Lys Ile Ser Ala Ser Gly Leu Arg Arg Ile Leu Glu
740 745 750
Arg Tyr Thr Trp Lys Ile Tyr Ser Glu Arg Leu Met Thr Leu Ser Gly
755 760 765
Val Tyr Gly Phe Trp Lys Leu Val Ser Lys Leu Glu Arg Arg Glu Thr
770 775 780
Arg Arg Tyr Leu Glu Met Phe Tyr Ile Leu Lys Phe Arg Glu Leu Ala
785 790 795 800
Lys Ser Val Pro Leu Ala Ile Asp Asp Lys
805 810
<210> 11
<211> 3604
<212> ДНК
<213> Nicotiana tabacum
<400> 11
atgtttacat ggctgaaact caatataaaa aacaagggta ggtgatcaaa aatcgttgga 60
tgcttaaaat cagtagacgt tttgctaaat gagcgaccaa tgttattgaa aacgttcatg 120
ttttcaaccc ttttggcata catttgagca ttgcccaaga ttttggataa gtagatgcag 180
tgcttataat tttaaagcat tgtatcctgc cttgtttttc attgtcaaaa ttaattaact 240
tacaagtatt tctataagtt gcttcataaa ttagaagtaa atctggattg tgtaatgtta 300
ttcgcctcgt aaatactgaa agctgcttga acaagtgaaa aaacacagac aaacgtaaca 360
ttctccatgg attgatgaga cttgtaaaat acatatatag aaatttggct tgtaaccaca 420
tgtatattat gccatatgga tgtgacattg atgtgactag acctaaatgt tttgtttcca 480
tgtccactgg agttttacgt atagttaaga ggagaaaaga ctgaggaata ctaatgtatg 540
atggtacccc tttgcttctt gacctggata cccagtgttc ctattcatgc ctatactttg 600
gtccttgatt tcactctccc ttttctaact tgagctgcat caaagaaatt tccactgtaa 660
aaaaataaat aatgctcacc atatctctgc aacattgcaa acatgtatcc catatgattg 720
atattggtgc gacatggcaa acatgtatcc tatttgatga tcaatcaaat ttatttttcc 780
cctgtcaaaa tgacctcagt gtgtaattcc ctatgtattt gatagcattg taactcgtgt 840
catgattcat gaatagggta ctagaattgc atggttgaca aatattaact ggtcgattga 900
tgatccacaa aacatgcact tactgactaa aatgtgatgg gacagattta tttttgtttg 960
tgattaacac agtacttaac cctatactta atacaatttg gcctagctac aatcttttct 1020
tcagtgcaaa ttccttgtta cacgaccaat attgtctttc tgagttctat tctgttgtta 1080
cttacacttt tattattcga ataagacatt agattgcttg catgcaaatt gatagccact 1140
tgtttcagga agaatactgt tggtcagtac gagagccata ctgcattcac cctcccagga 1200
ctatatcgcg tcgttcatgg cattgatgtt ttcgatccca aattcaatat agtgtctcct 1260
ggagctgaca tgacaattta cttcccatat tctgacaagg aaaaaagact aacgtctttg 1320
catggctcga ttgagaagtt gttatttgat cctgcgcaga atgaagagca tatgtaagtg 1380
acatccattt gtacttattt taatttggaa tagatgacat acttatttgc atgaatataa 1440
actgacaacc cagagatttc ctacattaga aaaggagggt ctgatatgat tttctacaaa 1500
taaattccca gtgatattgt tcaaaaagtc ctggatactt tattatgaga gaaccaggga 1560
tagatggcac tagaatccct taatcttgag aagtcgccac ttatcgctcc caacactttc 1620
tgagaccctc aagtaactac tattattgtt tgatatcttg gagaagctat aagaatcttt 1680
ttctccttat tgtaattttt tttacgtgac tttaaactta acttccaagc tccttctgat 1740
aaaatgcaaa aactgtctgt attcactgtc ttggtttatt aacaattgat ccaatcaaat 1800
gcatatggaa catctttctt tttgtttctt caaaagttcg tttgaggata aggagtagaa 1860
tctgagaaga tagactagta ggtaacctta ggggcggatg tagaaatcaa cgtatgggtt 1920
cagctttgtt gcagaccctg tatatgcatt aaaaaaatca ctaaataagt aaataattga 1980
ttttgaaccc agtaaatcaa aatgagttgt agtagaatcc tgaactcgaa ccgataaagt 2040
tggatccact accgggtaaa ctctaccttg agaagtgttt atatatgtcc ctaattattt 2100
cttttctgtt tcctttctat tttaattttt taagttcctt tttagatggt tttatttttt 2160
gacaagtggt aagttgttag tattccaaat taaatgccat tgccataact atatacattt 2220
ataaagattg attgacccta gtttctcatt cctaagatcc aaataaggca ataaacaata 2280
tgtcttagta cttgaacctg cttctggtgg tcaacacttg atcgcgtagt tagttataga 2340
tgactgtaaa aaccttaatc attttaatgg ttttgtcaaa gaacaaatat cggacatatt 2400
atagcgaatg gactattgta cttttcttct gattggtcat tttattgtga tccgtaagtt 2460
ggctgagact gatgtcatat ctttgcttac agaggtaatc tgaatgataa atcaaaaccc 2520
ataatttttt caatggcaag gctagaccat gttaagaaca ttacgggact agttgagtgc 2580
tatgctaaaa atgccacatt gagggaattg gctaaccttg ttgttgtagc tggatacaac 2640
gatgtaaaga aatccagtga tagagaagaa atagcagaaa ttgagaagat gcatgctctt 2700
attaaggagc ataaattgga tgggcaattc agatggatag cagcccaaac aaaccgggca 2760
cgtaatggtg agctctatcg ctatatagct gacaagagag gtatatttgt tcaggtacgc 2820
tgtttgtatt gtatttgtcc acattccttt ttttgcaccg aaagaaaggt tgttattgtg 2880
acaaatatgt ttgttttagc ctgcatttta tgaagcattt ggactcacgg tggttgaagc 2940
tatgacttgt ggtcttccaa catttgcaac ttgccatggt ggtccgaacg agatcattga 3000
acacggtgta tctgggttcc atattgatcc ttatcatccc gataaagctg ctgaactcat 3060
ggcagaattc tttcaacgct gcaaacaaga tcctactcac tgggaaaaaa tatctgcatc 3120
tggtctccga aggattcttg agaggtttgt agttgtgtac atatatagaa gattaaagat 3180
tgttcccttg atattatttg aatgaaaaat aacagtaaca tctctttttg aacatcgctc 3240
aagttcttgt gttaaataat tgttaggtat acgtggaaaa tttactccga gaggctgatg 3300
actttgtctg gtgtatatgg tttctggaag cttgtttcaa aacttgagag gcgcgaaact 3360
agacgatacc ttgagatgtt ctacattctc aaattccgcg agttggtgag tgcctttttg 3420
ctcattttca gttacaatca actatatatg tggtttaaat acgtattaag cataaacatg 3480
tccgtgattg cggctgtcga aaatgctatg gacatatcct gagctaagga gttttcaaga 3540
gaattgattt ggcttactct gtttacaggc aaaatctgtt cctctggcaa ttgatgacaa 3600
gtga 3604
<210> 12
<211> 335
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 12
Met Phe Thr Trp Leu Lys Leu Asn Ile Lys Asn Lys Gly Arg Lys Asn
1 5 10 15
Thr Val Gly Gln Tyr Glu Ser His Thr Ala Phe Thr Leu Pro Gly Leu
20 25 30
Tyr Arg Val Val His Gly Ile Asp Val Phe Asp Pro Lys Phe Asn Ile
35 40 45
Val Ser Pro Gly Ala Asp Met Thr Ile Tyr Phe Pro Tyr Ser Asp Lys
50 55 60
Glu Lys Arg Leu Thr Ser Leu His Gly Ser Ile Glu Lys Leu Leu Phe
65 70 75 80
Asp Pro Ala Gln Asn Glu Glu His Ile Gly Asn Leu Asn Asp Lys Ser
85 90 95
Lys Pro Ile Ile Phe Ser Met Ala Arg Leu Asp His Val Lys Asn Ile
100 105 110
Thr Gly Leu Val Glu Cys Tyr Ala Lys Asn Ala Thr Leu Arg Glu Leu
115 120 125
Ala Asn Leu Val Val Val Ala Gly Tyr Asn Asp Val Lys Lys Ser Ser
130 135 140
Asp Arg Glu Glu Ile Ala Glu Ile Glu Lys Met His Ala Leu Ile Lys
145 150 155 160
Glu His Lys Leu Asp Gly Gln Phe Arg Trp Ile Ala Ala Gln Thr Asn
165 170 175
Arg Ala Arg Asn Gly Glu Leu Tyr Arg Tyr Ile Ala Asp Lys Arg Gly
180 185 190
Ile Phe Val Gln Pro Ala Phe Tyr Glu Ala Phe Gly Leu Thr Val Val
195 200 205
Glu Ala Met Thr Cys Gly Leu Pro Thr Phe Ala Thr Cys His Gly Gly
210 215 220
Pro Asn Glu Ile Ile Glu His Gly Val Ser Gly Phe His Ile Asp Pro
225 230 235 240
Tyr His Pro Asp Lys Ala Ala Glu Leu Met Ala Glu Phe Phe Gln Arg
245 250 255
Cys Lys Gln Asp Pro Thr His Trp Glu Lys Ile Ser Ala Ser Gly Leu
260 265 270
Arg Arg Ile Leu Glu Arg Tyr Thr Trp Lys Ile Tyr Ser Glu Arg Leu
275 280 285
Met Thr Leu Ser Gly Val Tyr Gly Phe Trp Lys Leu Val Ser Lys Leu
290 295 300
Glu Arg Arg Glu Thr Arg Arg Tyr Leu Glu Met Phe Tyr Ile Leu Lys
305 310 315 320
Phe Arg Glu Leu Ala Lys Ser Val Pro Leu Ala Ile Asp Asp Lys
325 330 335
<210> 13
<211> 4030
<212> ДНК
<213> Nicotiana tabacum
<400> 13
atggcggaac gtgtgctgac tcgtgttcat agccttcgtg aacgtcttga tgctactttg 60
gctgctcatc gcaatgagat tttgctgttt ctttcaaggt atagtcttag cagattgttc 120
tttgatttag ttgttattgc cagttctaat gtatgggctt atatataaac aaagtgttga 180
agtatgcaac catataaact gacagcttaa aatgcttgag agaacacact tttatttatt 240
taattatgcc ttcagcacaa gaagtggaac ttgacgcaat ggaaccatag gtcacgggtt 300
caagtcttgg aacagcctgc aatctaaggc tgcgtgtagt agaccctagt ggtccggccc 360
ttccacatat ctcgcttagt gtaccgggcc cattgagtac gggttcggcc gaacccagtc 420
gctttggtcc aatccatata tttgtcttaa aaatatattg aatatataca aattgttaat 480
ttagtttaaa tatgtgtatc atgggttatt catgctggtt ttggctgttg caggattgaa 540
agccatggaa aagggatact gaaacctcac cagttgctgg ctgaatttga ttcaattcac 600
aaagaagaca aaaacaaact gaatgatcat gcttttgaag aagtcctgaa atccactcag 660
gtatttgtgg ttttagtgtt aggtgatgga tagcatttat tgttttacta agatcacata 720
tgtgtcagtt tgtggctagt atttaaaatc tggtgtattt tgtcatacta ggaagcaatt 780
gttttgtccc cttgggttgc gcttgccatt cgtctgaggc ctggtgtgtg ggaatacgtt 840
cgtgtgaatg tcaacgctct tgttgttgag gagcttaccg tgcctgagta tttgcaattc 900
aaggaagaac ttgttaatgg aacgtaagtt ttaggttcga atttgttgat ttgttagata 960
acatgttctg aactttttga ttaaagttgt gtttttgact gatgcagctc gcacgataac 1020
tttgttcttg agttggattt tgagcccttc actgcatcat ttccaaaacc aaccctcacc 1080
aaatcaattg gaaatggagt tgaattcctt aaccgacacc tctctgccaa aatgttccat 1140
gacaaggaaa gcatgacccc tcttctcgag tttcttcgag ttcaccacta caagggcaag 1200
gtaaacttgt ttttcctgtt tgtctatgaa tttagtttag ttgttttgct ccgcgaaaat 1260
ttcagtggaa actgatttat gcaaccactg agtgattaat atgttcaaac ttaccgactt 1320
ctggttttct gtgtagacaa tgatgctgaa tgacagaatt caggacttaa atactctcca 1380
aaatgtccta aggaaagctg aggaatacct cactaccctt tcccctgaaa cttcatactc 1440
ggcatttgag cacaagttcc aagaaattgg cttggagagg ggttggggtg acactgcgga 1500
gcgtgttcta gagatgatct gcatgctcct ggatctcctc gaggctcctg actcgtgcac 1560
gcttgagaag ttccttggta gaattccaat ggtttttaat gtggtcatac tttcacccca 1620
tggttatttc gcccaggaaa atgtcttggg ttaccccgac actggtggcc aggtgcactg 1680
cttatctgtg ttcggtctta ttatctcttt aaaccctact gccacaagtg ctgagatgaa 1740
cctcctttaa tttgcaggtt gtctatattt tggatcaagt tcctgctttg gagcgtgaga 1800
tgctcaagcg cataaaggag caaggacttg acatcaaacc gcgtattctt attgttcgta 1860
ttcccagtaa ttgtgtttaa acttatgatt atgcaggatt ttatctgttc taatacagca 1920
ctcttgctta aattctcagg ttactcggct gctgcctgat gcggttggta ccacttgtgg 1980
tcagaggctt gagaaagtgt ttggaacaga gcactcacac attcttaggg tcccctttag 2040
gaccgagaag ggcattgttc gcaaatggat ctctcgcttt gaagtctggc catacatgga 2100
gacattcact gaggtgaagc aagctttctc tattcatttt tcaatcttcc aattggtttt 2160
ggcagcaatt ttctgcttgc tttgacttcc gctaaaactt cggattttat tgcattagga 2220
tgtggcgaaa gaaattgctg cagaattgca ggctaagcca gatcttatca ttggcaatta 2280
tagtgagggc aaccttgctg cctccttgtt ggctcacaaa ttaggtgtaa cacaggtcgg 2340
caatgtttgt gacatgtaat ttcatctttg catttccttt cgtttgcaac taaaagattt 2400
aagagttctc tctctctttt ttttttccgt ctactttgcc ttatgcagtg cacgatagct 2460
catgctttgg agaaaacaaa atatcctgat tctgatatct acttgaagaa atttgatgaa 2520
aaataccatt tctcagccca gtttactgcc gatcttattg caatgaatca caccgatttc 2580
atcatcacca gcactttcca ggagatagcg ggaaggtatt tttacatcag tttcccactc 2640
tgattaaatt acaatgtatt tccctatatg attaaatact gtgtttgatc ctaaatcatt 2700
tctaaatttt ccagcaagga cactgttgga cagtacgaga gccacatggc gttcacaatg 2760
cctggactgt atagagttgt tcacggcatt gatgtgtttg accccaaatt taacattgtg 2820
tcaccaggag ctgatatgaa tctctatttc ccatactacg agaaggaaaa gagattgaca 2880
gcatatcacc ctgaaattga ggagctgctg tttagtgatg ttgagaatga cgaacacatg 2940
tatgttacta aactagcaat cctgctgcaa aattatggct aattatgtaa acaagtttgt 3000
actgaataga tttgttattc gatcaggtgt gtgctgaaga acaggaataa gcctatcata 3060
ttcactatgg ctagattgga tcgagtgaag aacttaactg gacttgtcga gctgtacgcc 3120
aagaacccac ggctaaggga gttggttaac cttgtcgtgg ttggaggaga ccgaaggaaa 3180
gaatccaaag acttggaaga acaggcagag atgaagaaga tgtacgaact tataaagact 3240
cacaatttga acggccaatt ccgatggatt tcttcccaga tgaaccgcgt gaggaatggc 3300
gaactctaca ggtacattgc cgatactagg ggagctttcg tgcagcctgc attttacgag 3360
gcttttggtt tgactgttgt tgaggccatg acctgtggtt tgcctacatt tgcaactaat 3420
cacggtggtc cagctgagat catcgttcac gggaaatctg gtttccacat tgatccatac 3480
cacggggatc aggcagctga acttctcgct gatttctttg agaaatgtaa gaaagaacct 3540
tcgcactggg aagccatttc cgagggcggc cttaagcgta tacaggagaa gtaagcaaac 3600
tgctactctt ttcatttttg caaaacctac tatgatcatt attaagctca tttttgcaaa 3660
acctacttgc tgttgttatt gtttgttgct tccttttcac tgttctttga gctgaaggtc 3720
tatcagaaac agtctctcta ccttcacaag gtaggggtaa gatctgcgtg cacgttaccc 3780
tcctcaaact ctacttaatt gtgagattac actaggtttg ttgttgttga ttctttgcta 3840
attaattaaa aggtacacat ggcaaatata ctcggatcgg ttgttgacac tggctgctgt 3900
atatggattc tggaagcatg tttccaagct tgatcgtctt gaaattcgcc gttatcttga 3960
aatgttctat gctctcaaat tccgcaagct ggtgagtttc attgctttct gcactcctgc 4020
aattgtatag 4030
<210> 14
<211> 808
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 14
Met Ala Glu Arg Val Leu Thr Arg Val His Ser Leu Arg Glu Arg Leu
1 5 10 15
Asp Ala Thr Leu Ala Ala His Arg Asn Glu Ile Leu Leu Phe Leu Ser
20 25 30
Arg Ile Glu Ser His Gly Lys Gly Ile Leu Lys Pro His Gln Leu Leu
35 40 45
Ala Glu Phe Asp Ser Ile His Lys Glu Asp Lys Asn Lys Leu Asn Asp
50 55 60
His Ala Phe Glu Glu Val Leu Lys Ser Thr Gln Glu Ala Ile Val Leu
65 70 75 80
Ser Pro Trp Val Ala Leu Ala Ile Arg Leu Arg Pro Gly Val Trp Glu
85 90 95
Tyr Val Arg Val Asn Val Asn Ala Leu Val Val Glu Glu Leu Thr Val
100 105 110
Pro Glu Tyr Leu Gln Phe Lys Glu Glu Leu Val Asn Gly Thr Ser His
115 120 125
Asp Asn Phe Val Leu Glu Leu Asp Phe Glu Pro Phe Thr Ala Ser Phe
130 135 140
Pro Lys Pro Thr Leu Thr Lys Ser Ile Gly Asn Gly Val Glu Phe Leu
145 150 155 160
Asn Arg His Leu Ser Ala Lys Met Phe His Asp Lys Glu Ser Met Thr
165 170 175
Pro Leu Leu Glu Phe Leu Arg Val His His Tyr Lys Gly Lys Thr Met
180 185 190
Met Leu Asn Asp Arg Ile Gln Asp Leu Asn Thr Leu Gln Asn Val Leu
195 200 205
Arg Lys Ala Glu Glu Tyr Leu Thr Thr Leu Ser Pro Glu Thr Ser Tyr
210 215 220
Ser Ala Phe Glu His Lys Phe Gln Glu Ile Gly Leu Glu Arg Gly Trp
225 230 235 240
Gly Asp Thr Ala Glu Arg Val Leu Glu Met Ile Cys Met Leu Leu Asp
245 250 255
Leu Leu Glu Ala Pro Asp Ser Cys Thr Leu Glu Lys Phe Leu Gly Arg
260 265 270
Ile Pro Met Val Phe Asn Val Val Ile Leu Ser Pro His Gly Tyr Phe
275 280 285
Ala Gln Glu Asn Val Leu Gly Tyr Pro Asp Thr Gly Gly Gln Val Val
290 295 300
Tyr Ile Leu Asp Gln Val Pro Ala Leu Glu Arg Glu Met Leu Lys Arg
305 310 315 320
Ile Lys Glu Gln Gly Leu Asp Ile Lys Pro Arg Ile Leu Ile Val Thr
325 330 335
Arg Leu Leu Pro Asp Ala Val Gly Thr Thr Cys Gly Gln Arg Leu Glu
340 345 350
Lys Val Phe Gly Thr Glu His Ser His Ile Leu Arg Val Pro Phe Arg
355 360 365
Thr Glu Lys Gly Ile Val Arg Lys Trp Ile Ser Arg Phe Glu Val Trp
370 375 380
Pro Tyr Met Glu Thr Phe Thr Glu Asp Val Ala Lys Glu Ile Ala Ala
385 390 395 400
Glu Leu Gln Ala Lys Pro Asp Leu Ile Ile Gly Asn Tyr Ser Glu Gly
405 410 415
Asn Leu Ala Ala Ser Leu Leu Ala His Lys Leu Gly Val Thr Gln Cys
420 425 430
Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Pro Asp Ser Asp Ile
435 440 445
Tyr Leu Lys Lys Phe Asp Glu Lys Tyr His Phe Ser Ala Gln Phe Thr
450 455 460
Ala Asp Leu Ile Ala Met Asn His Thr Asp Phe Ile Ile Thr Ser Thr
465 470 475 480
Phe Gln Glu Ile Ala Gly Ser Lys Asp Thr Val Gly Gln Tyr Glu Ser
485 490 495
His Met Ala Phe Thr Met Pro Gly Leu Tyr Arg Val Val His Gly Ile
500 505 510
Asp Val Phe Asp Pro Lys Phe Asn Ile Val Ser Pro Gly Ala Asp Met
515 520 525
Asn Leu Tyr Phe Pro Tyr Tyr Glu Lys Glu Lys Arg Leu Thr Ala Tyr
530 535 540
His Pro Glu Ile Glu Glu Leu Leu Phe Ser Asp Val Glu Asn Asp Glu
545 550 555 560
His Met Cys Val Leu Lys Asn Arg Asn Lys Pro Ile Ile Phe Thr Met
565 570 575
Ala Arg Leu Asp Arg Val Lys Asn Leu Thr Gly Leu Val Glu Leu Tyr
580 585 590
Ala Lys Asn Pro Arg Leu Arg Glu Leu Val Asn Leu Val Val Val Gly
595 600 605
Gly Asp Arg Arg Lys Glu Ser Lys Asp Leu Glu Glu Gln Ala Glu Met
610 615 620
Lys Lys Met Tyr Glu Leu Ile Lys Thr His Asn Leu Asn Gly Gln Phe
625 630 635 640
Arg Trp Ile Ser Ser Gln Met Asn Arg Val Arg Asn Gly Glu Leu Tyr
645 650 655
Arg Tyr Ile Ala Asp Thr Arg Gly Ala Phe Val Gln Pro Ala Phe Tyr
660 665 670
Glu Ala Phe Gly Leu Thr Val Val Glu Ala Met Thr Cys Gly Leu Pro
675 680 685
Thr Phe Ala Thr Asn His Gly Gly Pro Ala Glu Ile Ile Val His Gly
690 695 700
Lys Ser Gly Phe His Ile Asp Pro Tyr His Gly Asp Gln Ala Ala Glu
705 710 715 720
Leu Leu Ala Asp Phe Phe Glu Lys Cys Lys Lys Glu Pro Ser His Trp
725 730 735
Glu Ala Ile Ser Glu Gly Gly Leu Lys Arg Ile Gln Glu Lys Tyr Thr
740 745 750
Trp Gln Ile Tyr Ser Asp Arg Leu Leu Thr Leu Ala Ala Val Tyr Gly
755 760 765
Phe Trp Lys His Val Ser Lys Leu Asp Arg Leu Glu Ile Arg Arg Tyr
770 775 780
Leu Glu Met Phe Tyr Ala Leu Lys Phe Arg Lys Leu Val Ser Phe Ile
785 790 795 800
Ala Phe Cys Thr Pro Ala Ile Val
805
<210> 15
<211> 4054
<212> ДНК
<213> Nicotiana tabacum
<400> 15
atggccgaac gtgtgctaac tcgtgttcac agccttcgcg aacgtcttga tgctactttg 60
gctgctcatc gcaatgagat tttgctgttt ctttcaaggt atagtcttag cagattgttc 120
tttgatttag ttggtgttat ttgccagttc taatgtatgg actaatatat gaacaaagtg 180
cgaccatttc aactgacaac ttaaaatgtt tgagagaata cacgtttatt tacttaatta 240
tggcttgagc ataggaagtg tatcttggcg taactcgtaa agttgacctc atgtgacaag 300
gaggtcacgg tttcgagccg tggaaacagc ctcttgcaga aatgcaggta aggctgcgtg 360
caatagatcg cccttccacg gacccgcgca tagcgggaac ttagtgcacc ggttgggctg 420
tcctttttta tgtcttcagc acaaaaattt agtttaaaca tgtgtatcat ggattattca 480
tgctggtttt gccggttgca ggattgaaag ccacggaaaa gggatattga aacctcacca 540
gttgctggct gagtttgaat caattcacaa agaagacaaa aacaaactga atgatcatgc 600
ttttgaagaa gtcctgaaat ctactcaggt aatttgtggt tttagtgtta ggtgatggat 660
agcatttatt gtcttactaa gatcatatat gtgtcagttt gtggctagta tttgaaaagt 720
ctggtgtggt ttgtcatact aggaagcaat tgtcttgtcc ccttgggttg cgcttgccat 780
tcgtctgcgg cctggtgtgt gggaatatgt tcgtgtgaat gtcaatgcac ttattgtcga 840
ggagctgact gtgcctgaat atttgcaatt caaggaagaa cttgttaatg gaacgtaagt 900
tttaggttcg aaatgatgat ttgttaaata atatgttctg aactttttga ttaatgttgt 960
gttttcccct gatgcagctc gaacgataac tttgttcttg agctggattt tgagcccttc 1020
actgcatcat ttcccaaacc aaccctcacc aaatcaattg gaaatggagt tgaattcctc 1080
aaccgacacc tctctgccaa aatgttccat gacaaggaaa gcatgacccc tcttctcgag 1140
tttcttcgag ttcatcacta caagggcaag gtaaacttgt ttttcctgtt tgtctatgaa 1200
tttagtttct gaaagttgct ttgcttcgtg aattttttag tggcaactga tttatgattt 1260
tctgtgcaga caatgatgct gaatgacaga gttcaggact taaacactct ccaaaatgtc 1320
ctaaggaagg ctgaggaata tctcactacc ctttcccctg aaacttcata ctcggtattt 1380
gagcacaagt tccaagaaat tggcctagag aggggctggg gtgacaatgc tgagcgtgtt 1440
ctagagatga tctgcatgct cctggatctc ctcgaggctc cagactcatg cactcttgag 1500
aagttccttg gtagaattcc tatggttttt aatgtggtca ttctttcacc tcacggatat 1560
ttcgcccagg aaaatgtctt gggttacccc gatactggtg gccaggtgca ctgcttattt 1620
gtaacacctt acgcttttcc ctctgaaact tatttgcggc aagttctaag gtcctccttc 1680
cttaatttgc aggttgtcta tattttggat caagttccgg ccttggagcg tgagatgctc 1740
aagcgcataa aggagcaagg acttgatatc aaaccgcgta ttcttattgt tcgtatctcc 1800
aataattgcg tttaaactta tgattgtgca ggatttgatc tgttcaaatc taatgactga 1860
ttttcttttt tttttttttt tccctcaggt tactcggctg ctgcctgatg cggttggtac 1920
cacttgtggt cagcggcttg agaaagtgtt tggaacagag cattcacata ttcttagggt 1980
cccctttagg accgagaagg gcatcgttcg caaatggatc tctcgctttg aagtctggcc 2040
ttacatggag acattcactg aggtgaagca agctttctct attcattttt caatcttcca 2100
atctgttttg gcagcaattt ttcacttact aacactttgg ctttcgctaa aacttcggat 2160
tttattacat taggatgtgg caaaagaaat tgctgcagaa ctgcaggcaa agccagatct 2220
tataatcggc aactacagcg agggcaacct tgctgcctcc ttgttggctc acaagttagg 2280
tgtaactcag gtctgtaatg tttgtcacct gttatttcaa ctttgcattt cctttcattt 2340
gcaactagaa gttaagagtt ctctctcttt tatcttttcc gtctattttg ccttctgcag 2400
tgcaccatag ctcatgcgtt ggagaaaaca aaatatcctg attctgatat ctacttgaag 2460
aaatttgatg aaaaatacca tttctcagcc cagtttactg ccgatcttat tgcaatgaat 2520
cacaccgatt tcataatcac cagcactttc caggagatag cgggaaggta ttacatcaca 2580
atggatttcc gatatgatta aattagttaa tttaatccta cttcattgtg tttgatccta 2640
aaacttttct aaatttccca gcaaggacac tgttggacag tacgagagcc acatggcttt 2700
cacgatgcct ggattgtata gagttgttca cggcattgat gtgttcgatc ccaaattcaa 2760
cattgtgtca ccaggagctg atatgaatct ctatttcccc tacttcgaga aggaaaagcg 2820
attgacagca tatcaccctg aaattgagga gctgctgttt agcgatgttg agaatgacga 2880
acacatgtat gttactaaac tagcaatcct gctgcaaaat tgtggctaat tatgtaaaaa 2940
agtttttact gaatagattt gtgcttctat caggtgtgtg ctgaaggaca ggaataagcc 3000
aattatattc accatggcta gattggatcg agtgaagaac ttaactggac ttgtggagtt 3060
gtacgccaag aacccacggc taagggagtt ggttaacctt gtcgtggttg gtggagaccg 3120
aaggaaggaa tccaaagatt tggaagaaca ggcagagatg aagaagatgt atgaacttat 3180
aaagacgcac aatttaaacg gccaattccg atggatttct tcccagatga accgcgtgag 3240
gaatggcgaa ctctacaggt acattgccga tactagggga gcttttgtgc agcctgcatt 3300
ttacgaggct tttggtttga ctgttgttga ggccatgacc tgtggtttgc ctacgtttgc 3360
aactaatcac ggtggtccag ctgagatcat cgttcacggg aagtctggtt ttcacattga 3420
tccataccac ggcgagcagg cagctgaact tctagctgat ttctttgaga gatgtaagaa 3480
agaaccttca cactgggaag ccatttccga gggcggcctt aagcgtatac aggagaagta 3540
agcaagctgc tactcttttc atttttgcaa aacctaccat gatcattatt aagctcattt 3600
ttgcaaaacc tacttgttat tctttgttgc ttccttttcc ctgttttttg agccgaggtt 3660
ttatcgaaaa catgctttct accttcacaa ggtaggggta aggtctgcgt ttgttattat 3720
tgttgttgtt gattctctgc gaattaatta aaaggtacac atggcaaatc tactcggatc 3780
ggttgttgac actggctgct gtttatggat tctggaagca tgtttccaaa cttgatcgtc 3840
ttgaaattcg tcgttatctt gaaatgttct atgctctaaa attccgcaaa ctggtgagtt 3900
tcactgcttt ctgcactctt ccaattgtta gttgagtgca ctcatttaaa ctgtagctaa 3960
agctgttgta aatcttcagt taagcagctg ctaatgaagt ttttatcttt tgtttttggt 4020
tcaggctgaa gctgtcccgt tggctgttga gtaa 4054
<210> 16
<211> 805
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 16
Met Ala Glu Arg Val Leu Thr Arg Val His Ser Leu Arg Glu Arg Leu
1 5 10 15
Asp Ala Thr Leu Ala Ala His Arg Asn Glu Ile Leu Leu Phe Leu Ser
20 25 30
Arg Ile Glu Ser His Gly Lys Gly Ile Leu Lys Pro His Gln Leu Leu
35 40 45
Ala Glu Phe Glu Ser Ile His Lys Glu Asp Lys Asn Lys Leu Asn Asp
50 55 60
His Ala Phe Glu Glu Val Leu Lys Ser Thr Gln Glu Ala Ile Val Leu
65 70 75 80
Ser Pro Trp Val Ala Leu Ala Ile Arg Leu Arg Pro Gly Val Trp Glu
85 90 95
Tyr Val Arg Val Asn Val Asn Ala Leu Ile Val Glu Glu Leu Thr Val
100 105 110
Pro Glu Tyr Leu Gln Phe Lys Glu Glu Leu Val Asn Gly Thr Ser Asn
115 120 125
Asp Asn Phe Val Leu Glu Leu Asp Phe Glu Pro Phe Thr Ala Ser Phe
130 135 140
Pro Lys Pro Thr Leu Thr Lys Ser Ile Gly Asn Gly Val Glu Phe Leu
145 150 155 160
Asn Arg His Leu Ser Ala Lys Met Phe His Asp Lys Glu Ser Met Thr
165 170 175
Pro Leu Leu Glu Phe Leu Arg Val His His Tyr Lys Gly Lys Thr Met
180 185 190
Met Leu Asn Asp Arg Val Gln Asp Leu Asn Thr Leu Gln Asn Val Leu
195 200 205
Arg Lys Ala Glu Glu Tyr Leu Thr Thr Leu Ser Pro Glu Thr Ser Tyr
210 215 220
Ser Val Phe Glu His Lys Phe Gln Glu Ile Gly Leu Glu Arg Gly Trp
225 230 235 240
Gly Asp Asn Ala Glu Arg Val Leu Glu Met Ile Cys Met Leu Leu Asp
245 250 255
Leu Leu Glu Ala Pro Asp Ser Cys Thr Leu Glu Lys Phe Leu Gly Arg
260 265 270
Ile Pro Met Val Phe Asn Val Val Ile Leu Ser Pro His Gly Tyr Phe
275 280 285
Ala Gln Glu Asn Val Leu Gly Tyr Pro Asp Thr Gly Gly Gln Val Val
290 295 300
Tyr Ile Leu Asp Gln Val Pro Ala Leu Glu Arg Glu Met Leu Lys Arg
305 310 315 320
Ile Lys Glu Gln Gly Leu Asp Ile Lys Pro Arg Ile Leu Ile Val Thr
325 330 335
Arg Leu Leu Pro Asp Ala Val Gly Thr Thr Cys Gly Gln Arg Leu Glu
340 345 350
Lys Val Phe Gly Thr Glu His Ser His Ile Leu Arg Val Pro Phe Arg
355 360 365
Thr Glu Lys Gly Ile Val Arg Lys Trp Ile Ser Arg Phe Glu Val Trp
370 375 380
Pro Tyr Met Glu Thr Phe Thr Glu Asp Val Ala Lys Glu Ile Ala Ala
385 390 395 400
Glu Leu Gln Ala Lys Pro Asp Leu Ile Ile Gly Asn Tyr Ser Glu Gly
405 410 415
Asn Leu Ala Ala Ser Leu Leu Ala His Lys Leu Gly Val Thr Gln Cys
420 425 430
Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Pro Asp Ser Asp Ile
435 440 445
Tyr Leu Lys Lys Phe Asp Glu Lys Tyr His Phe Ser Ala Gln Phe Thr
450 455 460
Ala Asp Leu Ile Ala Met Asn His Thr Asp Phe Ile Ile Thr Ser Thr
465 470 475 480
Phe Gln Glu Ile Ala Gly Ser Lys Asp Thr Val Gly Gln Tyr Glu Ser
485 490 495
His Met Ala Phe Thr Met Pro Gly Leu Tyr Arg Val Val His Gly Ile
500 505 510
Asp Val Phe Asp Pro Lys Phe Asn Ile Val Ser Pro Gly Ala Asp Met
515 520 525
Asn Leu Tyr Phe Pro Tyr Phe Glu Lys Glu Lys Arg Leu Thr Ala Tyr
530 535 540
His Pro Glu Ile Glu Glu Leu Leu Phe Ser Asp Val Glu Asn Asp Glu
545 550 555 560
His Met Cys Val Leu Lys Asp Arg Asn Lys Pro Ile Ile Phe Thr Met
565 570 575
Ala Arg Leu Asp Arg Val Lys Asn Leu Thr Gly Leu Val Glu Leu Tyr
580 585 590
Ala Lys Asn Pro Arg Leu Arg Glu Leu Val Asn Leu Val Val Val Gly
595 600 605
Gly Asp Arg Arg Lys Glu Ser Lys Asp Leu Glu Glu Gln Ala Glu Met
610 615 620
Lys Lys Met Tyr Glu Leu Ile Lys Thr His Asn Leu Asn Gly Gln Phe
625 630 635 640
Arg Trp Ile Ser Ser Gln Met Asn Arg Val Arg Asn Gly Glu Leu Tyr
645 650 655
Arg Tyr Ile Ala Asp Thr Arg Gly Ala Phe Val Gln Pro Ala Phe Tyr
660 665 670
Glu Ala Phe Gly Leu Thr Val Val Glu Ala Met Thr Cys Gly Leu Pro
675 680 685
Thr Phe Ala Thr Asn His Gly Gly Pro Ala Glu Ile Ile Val His Gly
690 695 700
Lys Ser Gly Phe His Ile Asp Pro Tyr His Gly Glu Gln Ala Ala Glu
705 710 715 720
Leu Leu Ala Asp Phe Phe Glu Arg Cys Lys Lys Glu Pro Ser His Trp
725 730 735
Glu Ala Ile Ser Glu Gly Gly Leu Lys Arg Ile Gln Glu Lys Tyr Thr
740 745 750
Trp Gln Ile Tyr Ser Asp Arg Leu Leu Thr Leu Ala Ala Val Tyr Gly
755 760 765
Phe Trp Lys His Val Ser Lys Leu Asp Arg Leu Glu Ile Arg Arg Tyr
770 775 780
Leu Glu Met Phe Tyr Ala Leu Lys Phe Arg Lys Leu Ala Glu Ala Val
785 790 795 800
Pro Leu Ala Val Glu
805
<210> 17
<211> 3752
<212> ДНК
<213> Nicotiana tabacum
<400> 17
atggcctcaa cagttgctga tagcatgcct gatgctttga aacaaagccg gtatcatatg 60
aagagatgct tcgctaggtg aacacccttc ttttatgttt tttcccctct acgtgtttat 120
gtcaaatttc catgcataat gctaactact tttcttcttt ttgacttcaa aattggatgt 180
gaaaggttca ttgcaatggg aaggaggcta atgaagttga aacatttaac agaagaaata 240
gaagaaacta ttgaagacaa ggcagaaaga accaggattt tggagggttc acttggaaaa 300
attatgagtt ccacacaggt cagcaccatt taaccaactt agttgaacag gaaaaaaaga 360
aaaagcaaaa gagttattgc aaggcgtaac gattttcttt gaaattttca ggaggcagct 420
gttgttccac cttatgttgc ttttgcagta aggcacaatc ctggcttctg ggattatgtc 480
aaagttaacg ctgaaactct ctctgtggaa gctatttcag ccagggaata tctcaaattc 540
aaagagatga tctttgacga agactggtaa gtggaaaatt gtatcatttt aaagagaaac 600
aattttgtaa catacaagaa tagttttgat ggttgaatgt gcaagcaggg caaaggatga 660
taatgcactg gaagtagatt ttggtgcttt tgactactct aatcctcggt tagccctttc 720
ctcttctgtc ggaaatgggc tcaactttat ctcaaaagtt ctgtcttcaa agtttggtgg 780
aaagccagag gacgcccagc ctttgcttga ttacttacta gctcttaatc atcaaggaga 840
ggtatgaaaa tggactacct ttgtttctta aaggtattat ataatgatgc gcgttataaa 900
gttccttttt aaattgaaac tttgcagaat ctaatgatca atgagaatct gaatggtgtt 960
gctaagcttc aagcagcatt gatagtagct gaagtttttg tatcttcctt tcccaaagac 1020
acaccttata aagactttga gcataagtaa gcttctcata tgcttccatt gtcatatgca 1080
gtataccaat gacatgctac cgaaaagttg tttatgtttg tgacttgatt atgaaaactc 1140
taggctcaaa gaatggggct ttgataaagg gtggggtcac aatgcaggaa gagtaagaga 1200
gacaatgaga ctgctttccg agataatcca agcaccagat cccataaata tggagtcctt 1260
tttcagcaag cttcctacta cattcaacat tgttatcttc tccattcatg gttactttgg 1320
ccaagcagat gtccttggtc tgcccgatac tggaggccag gtctacatat acagcaattt 1380
atctcctttt gcctcatatt gcttattagc gacacttgca tcattgaaat cagactttta 1440
cttcacaggt tgtttatatt ctggatcaag taagggcttt agaggaggaa atgttacaaa 1500
gaatcaagca gcaagggcta aacgtgaagc ccaagattct tgtggtgagt tttgcaaaaa 1560
tatgcttaga caggttttga gattgatcgg agaagggatt aagatgatca agatctttgt 1620
ttcctgcttt catgatgtaa acaggtatct cgtctcatac cagatgctcg agggacaaca 1680
tgcaatcagg agatggaacc tattcttaac tcatcccatt ctcacatcct gagaattcca 1740
ttcaggactg agaaaggagt tcttcgccaa tgggtttctc ggtttgatat ctatccttac 1800
ttggagaact atgccaaggc aagtcttcta acaaaattac cacctattca tacactttat 1860
ttactttctt gaactaatcg tttggtttgt gacgtatatc attaggatgc ttctgctaag 1920
atacttgagc tcatggaagg taaaccagac ctcataattg ggaactacac tgatggaaat 1980
ttagtggcat ctctattggc caacaaactt ggagttactc aggttccgta gctgatcata 2040
tgatcatatt ttctacattg tttcttgata attaaatgga aatcttattg gatgataaca 2100
ttttagggaa ccattgctca tgcattagag aaaactaagt atgaagattc tgatgtgaag 2160
tggaagcagt ttgatcccaa gtaccacttt tcttgccaat ttactgccga tttattggca 2220
atgaatgctg ctgattttat cattaccagc acatatcaag aaatcgctgg aaggttagca 2280
ctgactctct cagtatattt ggcaacttaa tgaatttact gcagtggcca acactaaaag 2340
ctatcattcg tccttcagcg aaactaggcc tggacaatat gaaagtcaca cagcatttac 2400
catgccgggg ctttatagag ctgtttcagg catcaatgta tttgatccaa agttcaacat 2460
tgctgctcct ggggctgaac agtctaccta tttccctttc actgagaaac agaaacgatt 2520
cagcacattt cgtcctgcta ttaacgaatt actttacagt aatgaggaaa acaatgagca 2580
catgtaagtc taattgccca ttttcctaat ctaaccattg cttaaatcgt tctgttttta 2640
ccggatgtgt ggtacttatc agtaacattt ttttttggat cagtggattt cttgcagacc 2700
ggaaaaaacc aattatattt tcaatggcga gatttgatac agtgaagaac ctgtcaggct 2760
tgactgagtg gtatgggaag aataagaagt tgcggaactt ggtaaacctt gttattgttg 2820
ggggattctt cgatccatca aaatcaaaag accgggagga agcagctgaa atcaagaaga 2880
tgcatgaatt gattgagaaa taccagctca agggacaaat gagatggata gcagctcaaa 2940
ctgataaata tcgaaatagt gagctatacc gaactattgc tgacactaag ggagcttttg 3000
tccaaccggc tttatatgaa gcttttggac taaccgttat tgaagcaatg gattgtggat 3060
tgcctacgtt tgcaactaat caaggtggac ctgcagaaat cattgttgat ggggtttcag 3120
gtttccatat tgatccttac aatggggacg aatcaagcaa gaaaatagct gatttctttg 3180
agaagtgtaa ggttgattct aaatattgga acaggatatc tgagggaggt ctcaagcgca 3240
ttgaagaatg gtaacaaact agttccaagt ttaaaaaatg gaaaaaatgc ttatcatgtt 3300
atattttcgt ggttttaagt tctgcttcga tgcagttata cgtggaagat ttatgcaaac 3360
aaagtgttga atatgggatc aatctatgga ttttggagac aattcaatgt ggggcaaaag 3420
caggctaagc aaagatactt tgagatgttt tacaatcctc tcttcaggaa attggtaggt 3480
tgtatatgtt gaatacaatt tactaagatc ctcaaaatga ccaagaaata tacattgact 3540
atgctacttt tgtaatttca caggccaaaa gcgtgccgat cccacatgaa gagccattgc 3600
cacttgcaac atcagactct actcaatccc aagaattaaa actaccacta ccagttccag 3660
cagcagtagc taaagttctg ccattaacaa ggcatgcttt taacttaatt acttctctac 3720
ctagagtaac tggtaaagtg gatgtcaagt ga 3752
<210> 18
<211> 840
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 18
Met Ala Ser Thr Val Ala Asp Ser Met Pro Asp Ala Leu Lys Gln Ser
1 5 10 15
Arg Tyr His Met Lys Arg Cys Phe Ala Arg Phe Ile Ala Met Gly Arg
20 25 30
Arg Leu Met Lys Leu Lys His Leu Thr Glu Glu Ile Glu Glu Thr Ile
35 40 45
Glu Asp Lys Ala Glu Arg Thr Arg Ile Leu Glu Gly Ser Leu Gly Lys
50 55 60
Ile Met Ser Ser Thr Gln Glu Ala Ala Val Val Pro Pro Tyr Val Ala
65 70 75 80
Phe Ala Val Arg His Asn Pro Gly Phe Trp Asp Tyr Val Lys Val Asn
85 90 95
Ala Glu Thr Leu Ser Val Glu Ala Ile Ser Ala Arg Glu Tyr Leu Lys
100 105 110
Phe Lys Glu Met Ile Phe Asp Glu Asp Trp Ala Lys Asp Asp Asn Ala
115 120 125
Leu Glu Val Asp Phe Gly Ala Phe Asp Tyr Ser Asn Pro Arg Leu Ala
130 135 140
Leu Ser Ser Ser Val Gly Asn Gly Leu Asn Phe Ile Ser Lys Val Leu
145 150 155 160
Ser Ser Lys Phe Gly Gly Lys Pro Glu Asp Ala Gln Pro Leu Leu Asp
165 170 175
Tyr Leu Leu Ala Leu Asn His Gln Gly Glu Asn Leu Met Ile Asn Glu
180 185 190
Asn Leu Asn Gly Val Ala Lys Leu Gln Ala Ala Leu Ile Val Ala Glu
195 200 205
Val Phe Val Ser Ser Phe Pro Lys Asp Thr Pro Tyr Lys Asp Phe Glu
210 215 220
His Lys Leu Lys Glu Trp Gly Phe Asp Lys Gly Trp Gly His Asn Ala
225 230 235 240
Gly Arg Val Arg Glu Thr Met Arg Leu Leu Ser Glu Ile Ile Gln Ala
245 250 255
Pro Asp Pro Ile Asn Met Glu Ser Phe Phe Ser Lys Leu Pro Thr Thr
260 265 270
Phe Asn Ile Val Ile Phe Ser Ile His Gly Tyr Phe Gly Gln Ala Asp
275 280 285
Val Leu Gly Leu Pro Asp Thr Gly Gly Gln Val Val Tyr Ile Leu Asp
290 295 300
Gln Val Arg Ala Leu Glu Glu Glu Met Leu Gln Arg Ile Lys Gln Gln
305 310 315 320
Gly Leu Asn Val Lys Pro Lys Ile Leu Val Val Ser Arg Leu Ile Pro
325 330 335
Asp Ala Arg Gly Thr Thr Cys Asn Gln Glu Met Glu Pro Ile Leu Asn
340 345 350
Ser Ser His Ser His Ile Leu Arg Ile Pro Phe Arg Thr Glu Lys Gly
355 360 365
Val Leu Arg Gln Trp Asp Ala Ser Ala Lys Ile Leu Glu Leu Met Glu
370 375 380
Gly Lys Pro Asp Leu Ile Ile Gly Asn Tyr Thr Asp Gly Asn Leu Val
385 390 395 400
Ala Ser Leu Leu Ala Asn Lys Leu Gly Val Thr Gln Gly Thr Ile Ala
405 410 415
His Ala Leu Glu Lys Thr Lys Tyr Glu Asp Ser Asp Val Lys Trp Lys
420 425 430
Gln Phe Asp Pro Lys Tyr His Phe Ser Cys Gln Phe Thr Ala Asp Leu
435 440 445
Leu Ala Met Asn Ala Ala Asp Phe Ile Ile Thr Ser Thr Tyr Gln Glu
450 455 460
Ile Ala Gly Ser Glu Thr Arg Pro Gly Gln Tyr Glu Ser His Thr Ala
465 470 475 480
Phe Thr Met Pro Gly Leu Tyr Arg Ala Val Ser Gly Ile Asn Val Phe
485 490 495
Asp Pro Lys Phe Asn Ile Ala Ala Pro Gly Ala Glu Gln Ser Thr Tyr
500 505 510
Phe Pro Phe Thr Glu Lys Gln Lys Arg Phe Ser Thr Phe Arg Pro Ala
515 520 525
Ile Asn Glu Leu Leu Tyr Ser Asn Glu Glu Asn Asn Glu His Ile Gly
530 535 540
Phe Leu Ala Asp Arg Lys Lys Pro Ile Ile Phe Ser Met Ala Arg Phe
545 550 555 560
Asp Thr Val Lys Asn Leu Ser Gly Leu Thr Glu Trp Tyr Gly Lys Asn
565 570 575
Lys Lys Leu Arg Asn Leu Val Asn Leu Val Ile Val Gly Gly Phe Phe
580 585 590
Asp Pro Ser Lys Ser Lys Asp Arg Glu Glu Ala Ala Glu Ile Lys Lys
595 600 605
Met His Glu Leu Ile Glu Lys Tyr Gln Leu Lys Gly Gln Met Arg Trp
610 615 620
Ile Ala Ala Gln Thr Asp Lys Tyr Arg Asn Ser Glu Leu Tyr Arg Thr
625 630 635 640
Ile Ala Asp Thr Lys Gly Ala Phe Val Gln Pro Ala Leu Tyr Glu Ala
645 650 655
Phe Gly Leu Thr Val Ile Glu Ala Met Asp Cys Gly Leu Pro Thr Phe
660 665 670
Ala Thr Asn Gln Gly Gly Pro Ala Glu Ile Ile Val Asp Gly Val Ser
675 680 685
Gly Phe His Ile Asp Pro Tyr Asn Gly Asp Glu Ser Ser Lys Lys Ile
690 695 700
Ala Asp Phe Phe Glu Lys Cys Lys Val Asp Ser Lys Tyr Trp Asn Arg
705 710 715 720
Ile Ser Glu Gly Gly Leu Lys Arg Ile Glu Glu Cys Tyr Thr Trp Lys
725 730 735
Ile Tyr Ala Asn Lys Val Leu Asn Met Gly Ser Ile Tyr Gly Phe Trp
740 745 750
Arg Gln Phe Asn Val Gly Gln Lys Gln Ala Lys Gln Arg Tyr Phe Glu
755 760 765
Met Phe Tyr Asn Pro Leu Phe Arg Lys Leu Ala Lys Ser Val Pro Ile
770 775 780
Pro His Glu Glu Pro Leu Pro Leu Ala Thr Ser Asp Ser Thr Gln Ser
785 790 795 800
Gln Glu Leu Lys Leu Pro Leu Pro Val Pro Ala Ala Val Ala Lys Val
805 810 815
Leu Pro Leu Thr Arg His Ala Phe Asn Leu Ile Thr Ser Leu Pro Arg
820 825 830
Val Thr Gly Lys Val Asp Val Lys
835 840
<210> 19
<211> 3268
<212> ДНК
<213> Nicotiana tabacum
<400> 19
atggcctcaa ctgttgctgg tagcatgcct gatgctttga aacaaagccg atatcatatg 60
aagagatgct tcgctaggtg aacacccttc ttgttctttt tgttttttcc ctctaccatt 120
tatgtcaaat ttcaatgcat aatgctaact actttttttc tttttgactt caaaattgga 180
cgtgaaaggt tcattgcaat gggaaggagg ttgatgaagc tgaaacattt aacagaagaa 240
atagaaaaaa ctattgaaga caaggcagaa agaaccaaga ttttggaggg ttcacttgga 300
aaaattatga gttccacaca ggtcagcacc atttaaccaa cttaattgaa taggaagaaa 360
aaaaaaagca aaagagttat tgcaaggcgt aacgatttcc tttgaaattt tcaggaggca 420
gctgttgtcc caccttatgt tgcttttgca gtaaggcaca atcctggctt ctgggattat 480
gtcaaagttg acgctgaaac tctctctgtg gaagctattt cagccaggga ctatctcaaa 540
ttcaaagaga tgatctttga tgaagattgg taactggaag attgtatcat tttaaagaaa 600
caatttttta atattcaaga ttagttttga tggttgaatg tgcaagcagg gcaaaggatg 660
aaaatgcact cgaagtagat tttggtgctt ttgactactc taatcatcgg ttagcccttt 720
cctcttctgt cggaaatggg ctaaacttca tctcgaaagt tttgtcttca aagtttggtg 780
gaaaggcaga agatgcccag cctttgcttg attacttact agctcttaat catcaaggag 840
aggtatggaa atggactacc ttcctttctt aaggaattat ataatgatgt atgttataaa 900
gatccttttt aaacattgac actttgcaga atctaatgat caatgagaat ctgaatggcg 960
tctctaagct tcaagcagca ttgatagtag ctgaagtttt tgtatcttcc tttcccaaag 1020
acacacctta taaagacttt gagcataagt aagcttttca aacgcttctg ttatcatatg 1080
caatatacca agaatatgtt gccttttgaa aagttgttta tgtttatgac ttgataatga 1140
aaatactagg ctcaaagaat ggggctttga gaaagggtgg ggtcacaatg caggaagagt 1200
aagagagaca atgagactgc tttccgagat aatccaagcg ccagatccca taaatatgga 1260
gtcctttttc agcaggcttc ctactacatt caacattgtt atcttctcca ttcatggtta 1320
ctttggccaa gcagatgtcc ttggtttgcc cgatactgga ggccaggttt acatacacag 1380
caatttatct ccttttgcct catatttact tattagcgac acttgcatta ttgaaatcac 1440
atttgtattt aacaggttgt ttatattctg gatcaagtaa gagccttaga ggaggaaatg 1500
ttacaaagaa tcaagcagca agggttaaat gtgaagccca agattcttgt ggtgagttat 1560
gcaaaaatat gcgtagccaa ggttttgaaa ttgttcagag gggattaaga tgatcgagat 1620
atttgtttcc ttcttccatt gatgtgtaca ggtcactcgt ctcattccag atgctcgagg 1680
gactacatgc aatcaggaga tggaacctat acttaactcg tcccattctc acatcctgag 1740
aattccattc aggacagaga aaggagttct tcgccaatgg gtttctcggt ttgatatcta 1800
tccttacttg gagaactatg ccaaggcaag tctcctacca aaattaccac ctattcatac 1860
actttattca gttttttgag ctaatcattc tcatttgtca cgtatgtgat taggatgctt 1920
ctgctaagat acttgagctc atggaaggta aaccagacct cattattggg aactacactg 1980
atggaaattt agtggcatct ctattggcca acaaacttgg agttactcag gttctacagc 2040
tgatcattta tctgatcaga ttttctacat tgttttcttg ataattaaac ggaaatctta 2100
tgagattgta acattttagg gaaccattgc tcatgcatta gagaaaacca agtatgaaga 2160
ttctgatgtc aagtggaagc agtttgattc caagtaccac ttttcttgcc aattcactgc 2220
cgatttattg gcaatgaatg ctgctgattt tatcattacc agcacatatc aagaaatcgc 2280
aggaaggtta gcactgactc tctcagtata tttggcaact taatgaatgt actgcttgtg 2340
gccaacacta aaagctatta ctcgtccttc agcgaaacta ggcctggaca atatgaaagt 2400
cacacagcat ttaccatgcc ggggctttat agagctgttt caggcatcaa tgtatttgat 2460
ccaaagttca acattgctgc tcctggggct gaacagtctg cctatttccc cttcactgag 2520
aaacagaaac gattcagcgc gtttcgtcct gctattgagg aactacttta cagtaatgag 2580
caaaacaacg agcacatgta agtctaattg ccccattttc ctaatctaac cattgcttaa 2640
atgttctgtt tttacttgat atgtggtact tatcagtgat attttttatt ggaacagtgg 2700
atttcttgca gaccgtaaaa aaccaattat attttcaatg gcaagatttg atacggtgaa 2760
gaacttgtca ggcttgactg agtggtatgg gaagaataag aagttgcgga acttggttaa 2820
cctcgttatc gttgggggat tcttcgatcc atcaaaatca aaagaccggg aggaagcagc 2880
tgaaatcaag aagatgcatg aattgattga gaaatacaag ctcaagggac aaatgagatg 2940
gatagcagct caaactgata aatatcaaaa cagtgagcta tatcgaacta ttgctgacac 3000
taaaggagct ttcgtccaac cggctttata tgaagctttt ggactaactg ttattgaagc 3060
aatgaattgt ggactgccta catttgctac taatcaaggc ggacctgcag aaatcattgt 3120
tgatggggtt tcaggcttcc atattgatcc ttacaatggg gatgaatcga gcaagaaaat 3180
agctgatttc tttgagaagt gtaaggttga ttctaaatat tggaacaaga tatgtggagg 3240
aggtctcaag cgcattgaag aatggtaa 3268
<210> 20
<211> 732
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 20
Met Ala Ser Thr Val Ala Gly Ser Met Pro Asp Ala Leu Lys Gln Ser
1 5 10 15
Arg Tyr His Met Lys Arg Cys Phe Ala Arg Phe Ile Ala Met Gly Arg
20 25 30
Arg Leu Met Lys Leu Lys His Leu Thr Glu Glu Ile Glu Lys Thr Ile
35 40 45
Glu Asp Lys Ala Glu Arg Thr Lys Ile Leu Glu Gly Ser Leu Gly Lys
50 55 60
Ile Met Ser Ser Thr Gln Glu Ala Ala Val Val Pro Pro Tyr Val Ala
65 70 75 80
Phe Ala Val Arg His Asn Pro Gly Phe Trp Asp Tyr Val Lys Val Asp
85 90 95
Ala Glu Thr Leu Ser Val Glu Ala Ile Ser Ala Arg Asp Tyr Leu Lys
100 105 110
Phe Lys Glu Met Ile Phe Asp Glu Asp Trp Ala Lys Asp Glu Asn Ala
115 120 125
Leu Glu Val Asp Phe Gly Ala Phe Asp Tyr Ser Asn His Arg Leu Ala
130 135 140
Leu Ser Ser Ser Val Gly Asn Gly Leu Asn Phe Ile Ser Lys Val Leu
145 150 155 160
Ser Ser Lys Phe Gly Gly Lys Ala Glu Asp Ala Gln Pro Leu Leu Asp
165 170 175
Tyr Leu Leu Ala Leu Asn His Gln Gly Glu Asn Leu Met Ile Asn Glu
180 185 190
Asn Leu Asn Gly Val Ser Lys Leu Gln Ala Ala Leu Ile Val Ala Glu
195 200 205
Val Phe Val Ser Ser Phe Pro Lys Asp Thr Pro Tyr Lys Asp Phe Glu
210 215 220
His Lys Leu Lys Glu Trp Gly Phe Glu Lys Gly Trp Gly His Asn Ala
225 230 235 240
Gly Arg Val Arg Glu Thr Met Arg Leu Leu Ser Glu Ile Ile Gln Ala
245 250 255
Pro Asp Pro Ile Asn Met Glu Ser Phe Phe Ser Arg Leu Pro Thr Thr
260 265 270
Phe Asn Ile Val Ile Phe Ser Ile His Gly Tyr Phe Gly Gln Ala Asp
275 280 285
Val Leu Gly Leu Pro Asp Thr Gly Gly Gln Val Val Tyr Ile Leu Asp
290 295 300
Gln Val Arg Ala Leu Glu Glu Glu Met Leu Gln Arg Ile Lys Gln Gln
305 310 315 320
Gly Leu Asn Val Lys Pro Lys Ile Leu Val Val Thr Arg Leu Ile Pro
325 330 335
Asp Ala Arg Gly Thr Thr Cys Asn Gln Glu Met Glu Pro Ile Leu Asn
340 345 350
Ser Ser His Ser His Ile Leu Arg Ile Pro Phe Arg Thr Glu Lys Gly
355 360 365
Val Leu Arg Gln Trp Asp Ala Ser Ala Lys Ile Leu Glu Leu Met Glu
370 375 380
Gly Lys Pro Asp Leu Ile Ile Gly Asn Tyr Thr Asp Gly Asn Leu Val
385 390 395 400
Ala Ser Leu Leu Ala Asn Lys Leu Gly Val Thr Gln Gly Thr Ile Ala
405 410 415
His Ala Leu Glu Lys Thr Lys Tyr Glu Asp Ser Asp Val Lys Trp Lys
420 425 430
Gln Phe Asp Ser Lys Tyr His Phe Ser Cys Gln Phe Thr Ala Asp Leu
435 440 445
Leu Ala Met Asn Ala Ala Asp Phe Ile Ile Thr Ser Thr Tyr Gln Glu
450 455 460
Ile Ala Gly Ser Glu Thr Arg Pro Gly Gln Tyr Glu Ser His Thr Ala
465 470 475 480
Phe Thr Met Pro Gly Leu Tyr Arg Ala Val Ser Gly Ile Asn Val Phe
485 490 495
Asp Pro Lys Phe Asn Ile Ala Ala Pro Gly Ala Glu Gln Ser Ala Tyr
500 505 510
Phe Pro Phe Thr Glu Lys Gln Lys Arg Phe Ser Ala Phe Arg Pro Ala
515 520 525
Ile Glu Glu Leu Leu Tyr Ser Asn Glu Gln Asn Asn Glu His Ile Gly
530 535 540
Phe Leu Ala Asp Arg Lys Lys Pro Ile Ile Phe Ser Met Ala Arg Phe
545 550 555 560
Asp Thr Val Lys Asn Leu Ser Gly Leu Thr Glu Trp Tyr Gly Lys Asn
565 570 575
Lys Lys Leu Arg Asn Leu Val Asn Leu Val Ile Val Gly Gly Phe Phe
580 585 590
Asp Pro Ser Lys Ser Lys Asp Arg Glu Glu Ala Ala Glu Ile Lys Lys
595 600 605
Met His Glu Leu Ile Glu Lys Tyr Lys Leu Lys Gly Gln Met Arg Trp
610 615 620
Ile Ala Ala Gln Thr Asp Lys Tyr Gln Asn Ser Glu Leu Tyr Arg Thr
625 630 635 640
Ile Ala Asp Thr Lys Gly Ala Phe Val Gln Pro Ala Leu Tyr Glu Ala
645 650 655
Phe Gly Leu Thr Val Ile Glu Ala Met Asn Cys Gly Leu Pro Thr Phe
660 665 670
Ala Thr Asn Gln Gly Gly Pro Ala Glu Ile Ile Val Asp Gly Val Ser
675 680 685
Gly Phe His Ile Asp Pro Tyr Asn Gly Asp Glu Ser Ser Lys Lys Ile
690 695 700
Ala Asp Phe Phe Glu Lys Cys Lys Val Asp Ser Lys Tyr Trp Asn Lys
705 710 715 720
Ile Cys Gly Gly Gly Leu Lys Arg Ile Glu Glu Trp
725 730
<210> 21
<211> 3937
<212> ДНК
<213> Nicotiana tabacum
<400> 21
atggctactg caccagccct aaatagatca gagtccatag ctgatagcat gccagaggcc 60
ttaaggcaaa gccggtacca catgaagaaa tgttttgcca agtacataga gcaaggaaag 120
aggatgatga aacttcataa cttgatggat gagttggaga aagtaattga tgatcctgct 180
gaaaggaacc atgttttgga aggcttactt ggctacatat tatgcactac aatggtatag 240
ctagattcat atgtacttat gatgccctta tattgtttcc tgatgtatta ctcttaaaac 300
cttctttgat caaatttaca ggaggctgca gttgttcctc cctacattgc ctttgccacg 360
agacagaatc ctggattctg ggaatatgtg aaagtgaatg ctaatgatct ttctgttgag 420
ggtattacag ctacagaata cttgaaattc aaggaaatga tagttgatga atgctggtat 480
agtatacgtt gcagcttatc ataccttttg tggttttata acttcaatca gaaaactcat 540
cagagttacc tttgtgtgaa catgaaatgc agggcaaaag atgaatatgc actggaaatt 600
gattttggag cagtagactt ctcaacgcct cgactgaccc tatcctcttc aattggcaat 660
ggtctcagtt atgtttccaa gtttctaact tcaaagctaa atgctacctc cgcgagtgca 720
cagtgtctgg ttgactactt gctcactttg aatcatcaag gagatgtacg tcaacaaaaa 780
tcaaactcca taagtaaact tgtcaactct aagaagaaaa aataggaaaa gaagattcac 840
gtaacaaatt ttctttatgt tcaactgcag aaactgatga tcaatgagac actcagcact 900
gtctcaaagc ttcaggctgc actggttgta gcagaagcat ctatttcctc tttaccaaca 960
gatacaccat atgagagctt tgagctaagg tgatttgttt tttcctctac ttccctccac 1020
ttgtgccatg ctacgtagta ctaagtaact tcaattcttg taaagattca aacagtgggg 1080
ttttgagaaa ggatggggtg atacagctga aagggtcagc gacaccatga gaacactgtc 1140
tgaggtgctt caggcaccag atccattgaa cattcagaag ttctttggaa gggttccaac 1200
tgttttcaat attgtattgt tctctgtcca tggatacttt ggccaagcag atgttcttgg 1260
cttgccagac actggtggtc aggtaagcat ttaatagctt ttacatttaa cttctatgca 1320
ttgacaataa aataattttt aacagtttga ccacttctgc tcttgttcaa caggtagttt 1380
atgttttgga tcaagttgta gcttttgaag aagaaatgct acaaagaatt aaacagcagg 1440
ggctcaatat taagcctcaa attcttgtgg tgagttccta gacaatcgac gtgactatgc 1500
aattatgtag aggctgttta gaaaagttaa tatcatatgt tgattgcaca gttaacccga 1560
ctgattccgg atgcaaaagg aacaaagtgc aaccaggaac tagaaccaat caagaataca 1620
aaacattcac acatcctcag agttccattt aggacagaaa aaggagtgct taatcaatgg 1680
gtttcacgat ttgatatcta tccatatctg gagagatata ctcaggtatg tatttttata 1740
tcaaccttgc tcatcaaaga tgtgttgttt cctcaattcc atttttcccc ttggcaaaag 1800
gatgctgctg acaaaatcgt cgagctaatg gaaggcaaac ctgatctaat cattggtaac 1860
tacactgatg ggaatctagt ggcttcacta atggctagaa aacttgggat aactctggta 1920
acttttctta atcatatttg atgttgcttc ttctccaagt tagttcttaa tctccactga 1980
cctagaccat ctttgcaaca gggaactatt gctcatgctt tggagaagac aaaatatgaa 2040
gactctgaca taaaattgaa ggaactcgat ccgaagtacc acttctcttg ccaattcaca 2100
gctgatttga ttgcaatgaa ttcagcagat ttcattatca ctagcacata ccaagaaata 2160
gctggaaggt aagaattaga gctaataagt aatgcattca tatgtatttc agcatcgctc 2220
tttcaccatc atcgaataca caccactact cagtaaatgt atttgctcaa aagtttgcaa 2280
cttaatggat ctcattcttg aatgcttcaa catatgcagc aaagataaac caggacagta 2340
tgagagccat agtgcattta cccttccagg gctttacaga gttgcttcag gtatcaatgt 2400
ctttgatcca aaatttaata ttgctgcacc tggggcagac cagtcggtgt atttccctta 2460
cacagaaaag cagaagcgtt tgactgcttt ccgccctgcc attgaggaac tgctttttag 2520
taaagtggac aatgacgagc acgtgtaagt ctaagtgtta aacttcagct tagtgcctag 2580
aacatcccac tgctctatgt attgatgttt cacttgtttc aaacagtgga tatttagaag 2640
acagaaagaa acctatcctg tttaccatgg caaggctgga cacagtgaag aacacatctg 2700
gactaacaga atggtatggc aagaacaaga ggctcagaag cttagttaac cttgttgtgg 2760
ttggtggttc ctttgatcct acaaaatcca aggataggga agaagcagct gaaataaaaa 2820
agatgcacat gctgatagag aaataccagc ttaagggtca gattagatgg atagcagctc 2880
agactgacag atacagaaat agtgaactct accgcacaat agcagattcc aaaggagctt 2940
ttgtgcagcc tgcattgtat gaagcatttg gtctaacagt cattgaggca atgaactgtg 3000
gattaccaac ctttgctacc aaccaaggtg gccctgctga gattattgtt gatggggtct 3060
caggctttca tattgatcca aataatgggg atgaatcaag caacaaaatt gccaactttt 3120
tccaaaaatg cagggaggat cctgagtatt ggaacaggat ttcagtccag ggtctaaacc 3180
gtatatatga atggtaactc acagataagc cattcaaatt gcaaagaggc acatatcttg 3240
cagaaaattt cttaatcctt aaatcctaat tttttgcagt tacacatgga agatctatgc 3300
aaacaaggta ttgaatatgg ggtccatcta tactttttgg aggacattgt acagagatca 3360
gaaacaagca aagcaaagat acatcgagac tttctacaat cttgagttta ggaacttggt 3420
atagtgctgc atgacattga cagtatacca caaacatctt tatgagatga attactttta 3480
ataaaattgt ttttaacctt tgcttcctta atggcactta ttgcaggtaa aaaatgtgcc 3540
tatcagaaag gacgaaacac cacaaggacc aaaggagagg gagaaagtta agccacagat 3600
atcacaaagg catgctctaa agcttttgcc tacagttttt caagagaccc tagtatattc 3660
tagtactaaa ttagaattat acagcatgca gcttttgctg ttcacctttc taaatcacca 3720
gttgtgtcaa tcaagttgac aaaatcaata aattgggatt ttccctttcc tatgcttgat 3780
tgttattact cctactttgt ttatggtagt cttccttcat tgttttctcc tgtacttctt 3840
ttactacaac tgtactgaca tactaattat ttctgtgtac caggcgctca caatcaaggt 3900
tgcagaagta agattagata aaattgctac tgcatga 3937
<210> 22
<211> 860
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 22
Met Ala Thr Ala Pro Ala Leu Asn Arg Ser Glu Ser Ile Ala Asp Ser
1 5 10 15
Met Pro Glu Ala Leu Arg Gln Ser Arg Tyr His Met Lys Lys Cys Phe
20 25 30
Ala Lys Tyr Ile Glu Gln Gly Lys Arg Met Met Lys Leu His Asn Leu
35 40 45
Met Asp Glu Leu Glu Lys Val Ile Asp Asp Pro Ala Glu Arg Asn His
50 55 60
Val Leu Glu Gly Leu Leu Gly Tyr Ile Leu Cys Thr Thr Met Glu Ala
65 70 75 80
Ala Val Val Pro Pro Tyr Ile Ala Phe Ala Thr Arg Gln Asn Pro Gly
85 90 95
Phe Trp Glu Tyr Val Lys Val Asn Ala Asn Asp Leu Ser Val Glu Gly
100 105 110
Ile Thr Ala Thr Glu Tyr Leu Lys Phe Lys Glu Met Ile Val Asp Glu
115 120 125
Cys Trp Ala Lys Asp Glu Tyr Ala Leu Glu Ile Asp Phe Gly Ala Val
130 135 140
Asp Phe Ser Thr Pro Arg Leu Thr Leu Ser Ser Ser Ile Gly Asn Gly
145 150 155 160
Leu Ser Tyr Val Ser Lys Phe Leu Thr Ser Lys Leu Asn Ala Thr Ser
165 170 175
Ala Ser Ala Gln Cys Leu Val Asp Tyr Leu Leu Thr Leu Asn His Gln
180 185 190
Gly Asp Lys Leu Met Ile Asn Glu Thr Leu Ser Thr Val Ser Lys Leu
195 200 205
Gln Ala Ala Leu Val Val Ala Glu Ala Ser Ile Ser Ser Leu Pro Thr
210 215 220
Asp Thr Pro Tyr Glu Ser Phe Glu Leu Arg Phe Lys Gln Trp Gly Phe
225 230 235 240
Glu Lys Gly Trp Gly Asp Thr Ala Glu Arg Val Ser Asp Thr Met Arg
245 250 255
Thr Leu Ser Glu Val Leu Gln Ala Pro Asp Pro Leu Asn Ile Gln Lys
260 265 270
Phe Phe Gly Arg Val Pro Thr Val Phe Asn Ile Val Leu Phe Ser Val
275 280 285
His Gly Tyr Phe Gly Gln Ala Asp Val Leu Gly Leu Pro Asp Thr Gly
290 295 300
Gly Gln Val Val Tyr Val Leu Asp Gln Val Val Ala Phe Glu Glu Glu
305 310 315 320
Met Leu Gln Arg Ile Lys Gln Gln Gly Leu Asn Ile Lys Pro Gln Ile
325 330 335
Leu Val Leu Thr Arg Leu Ile Pro Asp Ala Lys Gly Thr Lys Cys Asn
340 345 350
Gln Glu Leu Glu Pro Ile Lys Asn Thr Lys His Ser His Ile Leu Arg
355 360 365
Val Pro Phe Arg Thr Glu Lys Gly Val Leu Asn Gln Trp Val Ser Arg
370 375 380
Phe Asp Ile Tyr Pro Tyr Leu Glu Arg Tyr Thr Gln Asp Ala Ala Asp
385 390 395 400
Lys Ile Val Glu Leu Met Glu Gly Lys Pro Asp Leu Ile Ile Gly Asn
405 410 415
Tyr Thr Asp Gly Asn Leu Val Ala Ser Leu Met Ala Arg Lys Leu Gly
420 425 430
Ile Thr Leu Gly Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Glu
435 440 445
Asp Ser Asp Ile Lys Leu Lys Glu Leu Asp Pro Lys Tyr His Phe Ser
450 455 460
Cys Gln Phe Thr Ala Asp Leu Ile Ala Met Asn Ser Ala Asp Phe Ile
465 470 475 480
Ile Thr Ser Thr Tyr Gln Glu Ile Ala Gly Ser Lys Asp Lys Pro Gly
485 490 495
Gln Tyr Glu Ser His Ser Ala Phe Thr Leu Pro Gly Leu Tyr Arg Val
500 505 510
Ala Ser Gly Ile Asn Val Phe Asp Pro Lys Phe Asn Ile Ala Ala Pro
515 520 525
Gly Ala Asp Gln Ser Val Tyr Phe Pro Tyr Thr Glu Lys Gln Lys Arg
530 535 540
Leu Thr Ala Phe Arg Pro Ala Ile Glu Glu Leu Leu Phe Ser Lys Val
545 550 555 560
Asp Asn Asp Glu His Val Gly Tyr Leu Glu Asp Arg Lys Lys Pro Ile
565 570 575
Leu Phe Thr Met Ala Arg Leu Asp Thr Val Lys Asn Thr Ser Gly Leu
580 585 590
Thr Glu Trp Tyr Gly Lys Asn Lys Arg Leu Arg Ser Leu Val Asn Leu
595 600 605
Val Val Val Gly Gly Ser Phe Asp Pro Thr Lys Ser Lys Asp Arg Glu
610 615 620
Glu Ala Ala Glu Ile Lys Lys Met His Met Leu Ile Glu Lys Tyr Gln
625 630 635 640
Leu Lys Gly Gln Ile Arg Trp Ile Ala Ala Gln Thr Asp Arg Tyr Arg
645 650 655
Asn Ser Glu Leu Tyr Arg Thr Ile Ala Asp Ser Lys Gly Ala Phe Val
660 665 670
Gln Pro Ala Leu Tyr Glu Ala Phe Gly Leu Thr Val Ile Glu Ala Met
675 680 685
Asn Cys Gly Leu Pro Thr Phe Ala Thr Asn Gln Gly Gly Pro Ala Glu
690 695 700
Ile Ile Val Asp Gly Val Ser Gly Phe His Ile Asp Pro Asn Asn Gly
705 710 715 720
Asp Glu Ser Ser Asn Lys Ile Ala Asn Phe Phe Gln Lys Cys Arg Glu
725 730 735
Asp Pro Glu Tyr Trp Asn Arg Ile Ser Val Gln Gly Leu Asn Arg Ile
740 745 750
Tyr Glu Cys Tyr Thr Trp Lys Ile Tyr Ala Asn Lys Val Leu Asn Met
755 760 765
Gly Ser Ile Tyr Thr Phe Trp Arg Thr Leu Tyr Arg Asp Gln Lys Gln
770 775 780
Ala Lys Gln Arg Tyr Ile Glu Thr Phe Tyr Asn Leu Glu Phe Arg Asn
785 790 795 800
Leu Val Lys Asn Val Pro Ile Arg Lys Asp Glu Thr Pro Gln Gly Pro
805 810 815
Lys Glu Arg Glu Lys Val Lys Pro Gln Ile Ser Gln Arg His Ala Leu
820 825 830
Lys Leu Leu Pro Thr Val Phe Gln Glu Thr Leu Ala Leu Thr Ile Lys
835 840 845
Val Ala Glu Val Arg Leu Asp Lys Ile Ala Thr Ala
850 855 860
<210> 23
<211> 3905
<212> ДНК
<213> Nicotiana tabacum
<400> 23
atggctactg caccagccct gaaaagatca gagtccatag ctgatagcat gccagaggcc 60
ttaaggcaaa gccggtacca catgaagaaa tgttttgcca agtacataga gcaaggcaag 120
aggatgatga aacttcataa cttgatggat gaattggaga aagtaattga tgatcctgct 180
gaaaggaacc atgttttgga aggcttactt ggctacatat tatgtactac aatggtatag 240
ctagattcat atgtacttat gatgtcctta tattgtttcc ggaggcatta ttcttaaatc 300
cttctttgat caaatttgta ggaggctgca gttgttcctc cctatattgc cttcgccacg 360
agacagaatc ctggattctg ggaatatgtg aaagtcaatg ctaatgatct ttctgttgag 420
ggtattacag ctacagatta cttgaaattc aaggaaatga tagttgatga aagctggtat 480
agaatacttt gcagcttatc ataccttttg tggttttata atttcaatca gaaaactcat 540
cagagttacc tttgtgtgaa catgacatgc agggcaaaag atgaatatgc actggaaatt 600
gattttggag cagtagactt ctcaacgcct cgactgaccc tatcctcttc aattggaaat 660
ggtctcagtt atgtttccaa gtttctaact tcaaagctaa atgctacctc agcgagtgca 720
cagtgtctgg ttgactactt gctcactttg aatcaccaag gagatgtacg tcaacaaaaa 780
tcaaactcca taagtaaact tgtcaactct aagaagtaaa aataggaaaa gaagattcat 840
gtaacaaatt ttctttatgt tcaactgtag aaactgatga tcaatgagac actcggcact 900
gtctcaaagc ttcaggctgc actggttgta gcagaagcat ctatttcctc cttaccaaca 960
gatacaccat accagagctt tgagctaagg tgatttgttt tttcctctac ttccttccac 1020
ttttggtgtg ctacatagta ctaagtaact tcaattcttg taaagattca aacagtgggg 1080
ttttgagaaa ggatggggtg atacagctga aagggtccgc gacaccatga gaacactttc 1140
tgaggtactt caggcgccag atccattgaa cattgagaag ttctttggga gggttccaac 1200
tgttttcaat attgtattgt tctctgttca tggatacttt ggccaagcaa atgttcttgg 1260
cttgccagac acaggtggtc aggtaagcat ctaatagctt ttacatttaa cttctatgca 1320
ttgacaataa aataacttct acactaccaa ataatttttg aaagtttgac cacttcggct 1380
cttgttcaac aggtggttta tgttttggat caagttgtag cttttgaaga agaaatgctc 1440
caaagaatta aacagcaggg gctcaatatt aagcctcaaa ttcttgtggt gagctcctag 1500
acaatgacgt gactatgcaa ttaagtagag gctgtttaga aaagttaata tcatatgttg 1560
attgcacagt taacccgact gattccggac gccaaaggaa caaagtgcaa ccaggaacta 1620
gaaccaatca agaatacaaa acattcacac atcctcagag ttccatttag gacagaaaaa 1680
ggagtgctta atcaatgggt ttcacgattt gatatctatc catatctgga gagatatact 1740
caggtgtgta tttttatatc aaccctgctc atcaaagatg tgttgtttcc tcaattccat 1800
ttttcgcctt gacaaaagga cgctgctgac aaaatcatcg agctaatgga aggcaaacct 1860
gatctaatca ttggtaacta cactgatggg aatctagtgg cttctctaat ggctagaaag 1920
cttgggataa ctctggtaac ttttcttatc atatttgatg ttgtttcttc tccaagttgg 1980
ttcttaatgt caactaaccc agaccatctt tgtaacaggg aactattgct catgctctgg 2040
agaagacaaa atatgaagac tctgacatca aattgaagga actcgatccg aagtaccact 2100
tttcttgcca attcacagct gatttgattg caatgaattc agcagatttc attatcacaa 2160
gcacatatca agaaatagcc ggaaggtaag aattggaact acggaagcag agagctaata 2220
agtagtgcac tcatatattt cagcatcgct ctttcgcata atcgaataca caccactact 2280
cagtaaatgt acttgctcaa aagtttacaa gtttatggat cttattcttg aatgcttcaa 2340
catatgcagc aaagataggc caggacagta tgagagccat agtgcattta cccttccagg 2400
gctttacaga gttgcttcag gcatcaatgt ctttgatcct aaatttaata ttgctgcacc 2460
tggggcagac caatcggtgt atttccctta cacagaaaag cagacgcgtt tgactgcttt 2520
ccgccctgcc attgaggaac tgctttttag taaagtggac aatgacgagc acatgtaagt 2580
cttagtgtta aacttcagct ttcagcttag tgcctagaac attccactgg ctctatgtat 2640
taatgtttca cttgtttcaa acacagtgga tatttagaag acagaaagaa acctatcctg 2700
tttaccatgg caaggctgga cacagtgaag aacacatctg gactaacaga atggtatggc 2760
aagaacaaga ggctcagaag cttagttaac cttgttgtgg ttggtggttc ctttgatcct 2820
acaaaatcca aggatagaga agaagcagct gaaataaaaa agatgcacat gctgatagag 2880
aaataccagc ttaagggtca gatcagatgg atagcagctc agactgacag atatagaaac 2940
agtgaactct accgcacaat agcagattcc aaaggagctt ttgtgcagcc tgcattatat 3000
gaagcatttg gtctaacagt cattgaggca atgaactgtg gattaccaac ctttgctacc 3060
aaccaaggtg gccctgctga gattattgtt gatggggtct caggctttca tattgatcca 3120
aataatgggg atgaatcaag caacaaagtt gccaactttt tccaaaaatg cagggaggat 3180
cctgagtatt ggaacaggat ttcagtccag ggtctaaacc gtatatatga atggtaactc 3240
acagataagc cattcaaatt gcaaagaggc acatatcttg ctgaaaattt cttaatcctt 3300
taatcctaaa attttgcagt tacacatgga agatctatgc aaacaaggta ttgaatatgg 3360
ggtccatcta tactttttgg aggacattgt acagagatca gaaacaagca aagcaaagat 3420
acatcgagac tttctacaat cttgagttta ggaacttggt atagtgctgc atgacattga 3480
cagtatacca caaacatctt tatgagatga attactttta ataaaattgt ttttaacctt 3540
tgcctcctta atgacactta ttgcaggtaa aaaatgtgcc tatcagacag gacgaaacac 3600
cacaaggacc aaaggagagg agggagaaag ttaagccaca gatatcacaa aggcatgctc 3660
taaagctttt gcctatagtt tttcaggaga ccctagtata ttctagtact aaattagaat 3720
tatacagcat gcagcttgct tctgctgttc acctttctaa atcaccagtt atgtcaatca 3780
agttgacaaa atcaataaat tcggcttttc cctttcctat gcttgattgt tattactcct 3840
acttcgttta tggtagtctt ccttcattgt tttctcctgt acttctttta ctacaactgt 3900
actga 3905
<210> 24
<211> 913
<212> БЕЛОКБЕЛОК
<213> Nicotiana tabacum
<400> 24
Met Ala Thr Ala Pro Ala Leu Lys Arg Ser Glu Ser Ile Ala Asp Ser
1 5 10 15
Met Pro Glu Ala Leu Arg Gln Ser Arg Tyr His Met Lys Lys Cys Phe
20 25 30
Ala Lys Tyr Ile Glu Gln Gly Lys Arg Met Met Lys Leu His Asn Leu
35 40 45
Met Asp Glu Leu Glu Lys Val Ile Asp Asp Pro Ala Glu Arg Asn His
50 55 60
Val Leu Glu Gly Leu Leu Gly Tyr Ile Leu Cys Thr Thr Met Glu Ala
65 70 75 80
Ala Val Val Pro Pro Tyr Ile Ala Phe Ala Thr Arg Gln Asn Pro Gly
85 90 95
Phe Trp Glu Tyr Val Lys Val Asn Ala Asn Asp Leu Ser Val Glu Gly
100 105 110
Ile Thr Ala Thr Asp Tyr Leu Lys Phe Lys Glu Met Ile Val Asp Glu
115 120 125
Ser Trp Ala Lys Asp Glu Tyr Ala Leu Glu Ile Asp Phe Gly Ala Val
130 135 140
Asp Phe Ser Thr Pro Arg Leu Thr Leu Ser Ser Ser Ile Gly Asn Gly
145 150 155 160
Leu Ser Tyr Val Ser Lys Phe Leu Thr Ser Lys Leu Asn Ala Thr Ser
165 170 175
Ala Ser Ala Gln Cys Leu Val Asp Tyr Leu Leu Thr Leu Asn His Gln
180 185 190
Gly Asp Lys Leu Met Ile Asn Glu Thr Leu Gly Thr Val Ser Lys Leu
195 200 205
Gln Ala Ala Leu Val Val Ala Glu Ala Ser Ile Ser Ser Leu Pro Thr
210 215 220
Asp Thr Pro Tyr Gln Ser Phe Glu Leu Arg Phe Lys Gln Trp Gly Phe
225 230 235 240
Glu Lys Gly Trp Gly Asp Thr Ala Glu Arg Val Arg Asp Thr Met Arg
245 250 255
Thr Leu Ser Glu Val Leu Gln Ala Pro Asp Pro Leu Asn Ile Glu Lys
260 265 270
Phe Phe Gly Arg Val Pro Thr Val Phe Asn Ile Val Leu Phe Ser Val
275 280 285
His Gly Tyr Phe Gly Gln Ala Asn Val Leu Gly Leu Pro Asp Thr Gly
290 295 300
Gly Gln Val Val Tyr Val Leu Asp Gln Val Val Ala Phe Glu Glu Glu
305 310 315 320
Met Leu Gln Arg Ile Lys Gln Gln Gly Leu Asn Ile Lys Pro Gln Ile
325 330 335
Leu Val Leu Thr Arg Leu Ile Pro Asp Ala Lys Gly Thr Lys Cys Asn
340 345 350
Gln Glu Leu Glu Pro Ile Lys Asn Thr Lys His Ser His Ile Leu Arg
355 360 365
Val Pro Phe Arg Thr Glu Lys Gly Val Leu Asn Gln Trp Val Ser Arg
370 375 380
Phe Asp Ile Tyr Pro Tyr Leu Glu Arg Tyr Thr Gln Asp Ala Ala Asp
385 390 395 400
Lys Ile Ile Glu Leu Met Glu Gly Lys Pro Asp Leu Ile Ile Gly Asn
405 410 415
Tyr Thr Asp Gly Asn Leu Val Ala Ser Leu Met Ala Arg Lys Leu Gly
420 425 430
Ile Thr Leu Gly Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Glu
435 440 445
Asp Ser Asp Ile Lys Leu Lys Glu Leu Asp Pro Lys Tyr His Phe Ser
450 455 460
Cys Gln Phe Thr Ala Asp Leu Ile Ala Met Asn Ser Ala Asp Phe Ile
465 470 475 480
Ile Thr Ser Thr Tyr Gln Glu Ile Ala Gly Ser Lys Asp Arg Pro Gly
485 490 495
Gln Tyr Glu Ser His Ser Ala Phe Thr Leu Pro Gly Leu Tyr Arg Val
500 505 510
Ala Ser Gly Ile Asn Val Phe Asp Pro Lys Phe Asn Ile Ala Ala Pro
515 520 525
Gly Ala Asp Gln Ser Val Tyr Phe Pro Tyr Thr Glu Lys Gln Thr Arg
530 535 540
Leu Thr Ala Phe Arg Pro Ala Ile Glu Glu Leu Leu Phe Ser Lys Val
545 550 555 560
Asp Asn Asp Glu His Ile Gly Tyr Leu Glu Asp Arg Lys Lys Pro Ile
565 570 575
Leu Phe Thr Met Ala Arg Leu Asp Thr Val Lys Asn Thr Ser Gly Leu
580 585 590
Thr Glu Trp Tyr Gly Lys Asn Lys Arg Leu Arg Ser Leu Val Asn Leu
595 600 605
Val Val Val Gly Gly Ser Phe Asp Pro Thr Lys Ser Lys Asp Arg Glu
610 615 620
Glu Ala Ala Glu Ile Lys Lys Met His Met Leu Ile Glu Lys Tyr Gln
625 630 635 640
Leu Lys Gly Gln Ile Arg Trp Ile Ala Ala Gln Thr Asp Arg Tyr Arg
645 650 655
Asn Ser Glu Leu Tyr Arg Thr Ile Ala Asp Ser Lys Gly Ala Phe Val
660 665 670
Gln Pro Ala Leu Tyr Glu Ala Phe Gly Leu Thr Val Ile Glu Ala Met
675 680 685
Asn Cys Gly Leu Pro Thr Phe Ala Thr Asn Gln Gly Gly Pro Ala Glu
690 695 700
Ile Ile Val Asp Gly Val Ser Gly Phe His Ile Asp Pro Asn Asn Gly
705 710 715 720
Asp Glu Ser Ser Asn Lys Val Ala Asn Phe Phe Gln Lys Cys Arg Glu
725 730 735
Asp Pro Glu Tyr Trp Asn Arg Ile Ser Val Gln Gly Leu Asn Arg Ile
740 745 750
Tyr Glu Cys Tyr Thr Trp Lys Ile Tyr Ala Asn Lys Val Leu Asn Met
755 760 765
Gly Ser Ile Tyr Thr Phe Trp Arg Thr Leu Tyr Arg Asp Gln Lys Gln
770 775 780
Ala Lys Gln Arg Tyr Ile Glu Thr Phe Tyr Asn Leu Glu Phe Arg Asn
785 790 795 800
Leu Val Lys Asn Val Pro Ile Arg Gln Asp Glu Thr Pro Gln Gly Pro
805 810 815
Lys Glu Arg Arg Glu Lys Val Lys Pro Gln Ile Ser Gln Arg His Ala
820 825 830
Leu Lys Leu Leu Pro Ile Val Phe Gln Glu Thr Leu Val Tyr Ser Ser
835 840 845
Thr Lys Leu Glu Leu Tyr Ser Met Gln Leu Ala Ser Ala Val His Leu
850 855 860
Ser Lys Ser Pro Val Met Ser Ile Lys Leu Thr Lys Ser Ile Asn Ser
865 870 875 880
Ala Phe Pro Phe Pro Met Leu Asp Cys Tyr Tyr Ser Tyr Phe Val Tyr
885 890 895
Gly Ser Leu Pro Ser Leu Phe Ser Pro Val Leu Leu Leu Leu Gln Leu
900 905 910
Tyr
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ САХАРОВ И АМИНОКИСЛОТ В РАСТЕНИИ (SULTR3) | 2020 |
|
RU2826107C1 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2018 |
|
RU2809244C2 |
| СЛИТЫЙ БЕЛОК ИЗ БЕЛКА DCTN1 С БЕЛКОМ RET | 2018 |
|
RU2813996C2 |
| СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ ОДНОДОЛЬНЫХ РАСТЕНИЙ | 2015 |
|
RU2727428C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2817591C2 |
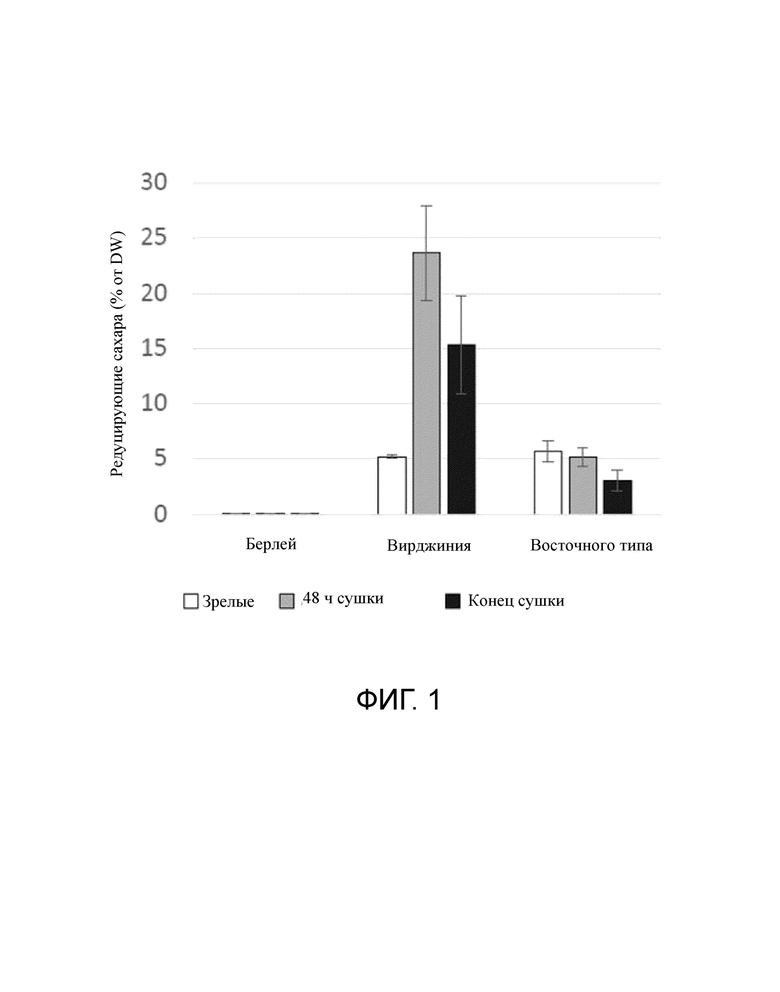
Изобретение относится к области биохимии, в частности к подвергнутому сушке растительному материалу из Nicotiana tabacum. Также раскрыт табачный продукт, содержащий указанный растительный материал из Nicotiana tabacum. Раскрыт способ получения указанного подвергнутого сушке растительного материала из Nicotiana tabacum. Изобретение позволяет эффективно модулировать экспрессию или активность полинуклеотида или полипептида, влияющие на уровни редуцирующих сахаров в подвергнутому сушке растительному материалу из Nicotiana tabacum. 3 н. и 9 з.п. ф-лы, 1 ил., 2 табл., 2 пр.
1. Подвергнутый сушке растительный материал из Nicotiana tabacum для применения в получении табачного продукта, содержащий:
(i) полинуклеотид, состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 9 или SEQ ID NO: 11; или полинуклеотид, состоящий из последовательности, характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5 или SEQ ID NO: 13;
(ii) полипептид, кодируемый полинуклеотидом, указанным в (i);
(iii) полипептид, состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 6 или SEQ ID NO: 14; или по меньшей мере 93% идентичностью последовательности с SEQ ID NO: 10 или SEQ ID NO: 12; или
(iv) конструкцию, вектор или вектор экспрессии, содержащие выделенный полинуклеотид, указанный в (i),
при этом указанный растительный материал содержит по меньшей мере одну модификацию, модулирующую экспрессию или активность полинуклеотида или полипептида по сравнению с таковыми контрольного подвергнутого сушке растительного материала, в котором экспрессия или активность полинуклеотида или полипептида являются немодифицированными, и
при этом модулированная экспрессия или активность полинуклеотида или полипептида модулирует уровень редуцирующих сахаров в подвергнутом сушке растительном материале по сравнению с уровнем редуцирующих сахаров в контрольном подвергнутом сушке растительном материале, и
при этом по меньшей мере одна модификация представляет собой модификацию генома подвергнутого сушке растительного материала или модификацию конструкции, вектора или вектора экспрессии или трансгенную модификацию, и/или
при этом по меньшей мере одна модификация представляет собой полинуклеотид для обеспечения интерференции, содержащий последовательность, которая комплементарна по меньшей мере 19 нуклеотидам РНК, транскрибируемой с полинуклеотида (i).
2. Растительный материал по п. 1, в котором модификация генома подвергнутого сушке растительного материала или модификация конструкции, вектора или вектора экспрессии представляет собой мутацию или редактирование.
3. Растительный материал по любому из предыдущих пунктов, в котором модификация повышает или снижает экспрессию или активность полинуклеотида или полипептида по сравнению с контрольным подвергнутым сушке растительным материалом.
4. Растительный материал по любому из предыдущих пунктов, в котором редуцирующий сахар представляет собой глюкозу или фруктозу.
5. Растительный материал по любому из предыдущих пунктов, содержащий биомассу, семя, стебель, цветки или листья.
6. Растительный материал по любому из предыдущих пунктов, подвергнутый трубоогневой сушке, солнечной сушке или воздушной сушке.
7. Растительный материал по любому из предыдущих пунктов, подвергнутый гомогенизации.
8. Табачный продукт, содержащий подвергнутый сушке растительный материал по любому из предыдущих пунктов.
9. Способ получения подвергнутого сушке растительного материала из Nicotiana tabacum по любому из пп. 1-7, содержащий
(a) обеспечение клетки растения Nicotiana tabacum, содержащей полинуклеотид, состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 9 или SEQ ID NO: 11; или характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5 или SEQ ID NO: 13;
(b) модификация клетки растения для обеспечения модулирования экспрессии полинуклеотида по сравнению с таковой в клетке контрольного растения;
(c) регенерация клетки растения с получением растения;
(d) сбор из него растительного материала; и
(f) сушку растительного материала.
10. Способ по п. 9, в котором стадия модификации клетки растения включает модификацию генома клетки с помощью методов редактирования генома или геномной инженерии; предпочтительно,
при этом методы редактирования генома или геномной инженерии выбраны из технологии CRISPR/Cas, мутагенеза, опосредованного нуклеазой с «цинковыми пальцами», химического или радиационного мутагенеза, гомологичной рекомбинации, олигонуклеотид-направленного мутагенеза и мутагенеза, опосредованного мегануклеазой.
11. Способ по п. 9 или 10, в котором стадия модификации клетки растения включает трансфекцию клетки конструкцией, содержащей полинуклеотид, состоящий из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 9, или SEQ ID NO: 11; или характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5 или SEQ ID NO: 13, функционально связанной с конститутивным промотором; и/или
при этом клетку растения трансфицируют конструкцией, экспрессирующей полинуклеотид для обеспечения интерференции, содержащий последовательность, которая комплементарна по меньшей мере 19 нуклеотидам РНК, транскрибируемым с полинуклеотида, состоящего из последовательности, характеризующейся по меньшей мере 95% идентичностью последовательности с SEQ ID NO: 9 или SEQ ID NO: 11; или полинуклеотида, состоящего из последовательности, характеризующейся по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 5 или SEQ ID NO: 13.
12. Способ по любому из пп. 9-11, в котором растительный материал содержит подвергнутые сушке листья, подвергнутые сушке стебли или подвергнутые сушке цветки или их смесь; и/или
в котором способ сушки выбран из группы, состоящей из воздушной сушки, огневой сушки, дымовой сушки и трубоогневой сушки.
| СПОСОБ ДВУХСТУПЕНЧАТОГО СЖИГАНИЯ ТОПЛИВА И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2006 |
|
RU2334166C2 |
| WO 2013029799 A1, 07.03.2013 | |||
| RU 2015126909 A, 27.01.2017 | |||
| SIERRO N., et al., The tobacco genome sequence and its comparison with those of tomato and potato, NATURE COMMUNICATIONS, 2014, pp.1-9. | |||
