Ссылка на родственные заявки
[0001] Согласно настоящей заявке испрашивается приоритет в соответствии с заявкой на выдачу патента США №62/565980, поданной 29 сентября 2017 года, заявкой на выдачу патента США №62/679142, поданной 1 июня 2018 года, и заявкой на выдачу патента США №62/720292, поданной 21 августа 2018 года, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
Ссылка на перечень последовательностей, представленный в текстовом файле на веб-сайте EFS
[0002] Перечень последовательностей, изложенный в файле 519832SEQLIST.txt размером 139 килобайт, был создан 25 сентября 2018 года и тем самым включен в настоящий документ посредством ссылки.
Предшествующий уровень техники настоящего изобретения
[0003] Транстиретин (TTR) представляет собой белок, обнаруженный в сыворотке и спинномозговой жидкости, который переносит гормон щитовидной железы и ретинол-связывающий белок к ретинолу. Печень секретирует TTR в кровь, а хороидное сплетение секретирует его в спинномозговую жидкость. TTR также продуцируется в пигментированном эпителии сетчатки и секретируется в стекловидное тело. Неправильно свернутый и агрегированный TTR накапливается во многих тканях и органах при амилоидных заболеваниях, старческом системном амилоидозе (SSA), семейной амилоидной полинейропатии (FAP) и семейной амилоидной кардиомиопатии (FAC).
[0004] Одним из многообещающих терапевтических подходов при заболеваниях TTR амилоидозом является снижение нагрузки TTR у больного. Однако по-прежнему сохраняется потребность в подходящих отличных от человека животных, обеспечивающих реальную человеческую мишень или близкое подобие реальной человеческой мишени для нацеленных на человеческий TTR реагентов по эндогенному Ttr локусу, позволяющих тем самым тестировать эффективность и способ действия таких средств на живых животных, а также выполнять фармакокинетические и фармакодинамические исследования в условиях, при которых гуманизированный белок и гуманизированный ген являются единственной версией присутствующего TTR.
Краткое раскрытие настоящего изобретения
[0005] Представлены отличные от человека животные, включающие в себя гуманизированный TTR локус, а также способы применения таких отличных от человека животных. Также представлены геномы отличных от человека животных или клетки, включающие в себя гуманизированный TTR локус.
[0006] Согласно одному аспекту представлены геномы отличных от человека животных, клетки отличного от человека животного или отличные от человека животные, включающие в себя гуманизированный TTR локус. Такие геном отличного от человека животного, клетка отличного от человека животного или отличное от человека животное могут включать в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность. Некоторые такие геномы отличных от человека животных, клетки отличного от человека животного или отличные от человека животные могут включать в себя генетически модифицированный эндогенный Ttr локус, в котором область эндогенного Ttr локуса, включающая в себя как Ttr кодирующую последовательность, так и некодирующую последовательность, была делетирована и заменена соответствующей человеческой TTR последовательностью, включающей в себя как TTR кодирующую последовательность, так и некодирующую последовательность. Необязательно генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор. Необязательно человеческая TTR последовательность функционально связана с эндогенным Ttr промотором. Необязательно по меньшей мере один интрон и по меньшей мере один экзон эндогенного Ttr локуса были делетированы и заменены соответствующей человеческой TTR последовательностью.
[0007] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных вся Ttr кодирующая последовательность эндогенного Ttr локуса была делетирована и заменена соответствующей человеческой TTR последовательностью. Необязательно область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью.
[0008] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область. В некоторых таких отличных от человека животных эндогенная Ttr 5' нетранслируемая область не была делетирована и заменена соответствующей человеческой TTR последовательностью.
[0009] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена человеческой TTR последовательностью, включающей в себя соответствующую человеческую TTR последовательность и человеческую TTR 3' нетранслируемую область, а эндогенная Ttr 5' нетранслируемая область не была делетирована и заменена соответствующей человеческой TTR последовательностью, и эндогенный Ttr промотор не был делетирован и заменен соответствующей человеческой TTR последовательностью. Необязательно человеческая TTR последовательность в генетически модифицированном эндогенном Ttr локусе включает в себя, состоит, по сути, из или состоит из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 18. Необязательно генетически модифицированный эндогенный Ttr локус кодирует белок, включающий в себя, состоящий, по сути, из или состоящий из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 1. Необязательно генетически модифицированный эндогенный Ttr локус включает в себя кодирующую последовательность, включающую в себя, состоящую, по сути, из или состоящую из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 90. Необязательно генетически модифицированный эндогенный Ttr локус включает в себя, состоит, по сути, из или состоит из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 14 или 15.
[0010] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных генетически модифицированный эндогенный Ttr локус кодирует белок-предшественник транстиретина, включающий в себя сигнальный пептид, и область эндогенного Ttr локуса, кодирующая сигнальный пептид, не была делетирована и заменена соответствующей человеческой TTR последовательностью. Необязательно первый экзон эндогенного Ttr локуса не был делетирован и заменен соответствующей человеческой TTR последовательностью. Необязательно первый экзон и первый интрон эндогенного Ttr локуса не были делетированы и заменены соответствующей человеческой TTR последовательностью. Необязательно область эндогенного Ttr локуса от начала второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью. Необязательно генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область.
[0011] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных область эндогенного Ttr локуса от второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена человеческой TTR последовательностью, включающей в себя соответствующую человеческую TTR последовательность и человеческую TTR 3' нетранслируемую область, а эндогенная Ttr 5' нетранслируемая область не была делетирована и заменена соответствующей человеческой TTR последовательностью, и эндогенный Ttr промотор не был делетирован и заменен соответствующей человеческой TTR последовательностью. Необязательно человеческая TTR последовательность в генетически модифицированном эндогенном Ttr локусе включает в себя, состоит, по сути, из или состоит из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 19. Необязательно генетически модифицированный эндогенный Ttr локус кодирует белок, включающий в себя, состоящий, по сути, из или состоящий из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 2. Необязательно генетически модифицированный эндогенный Ttr локус включает в себя кодирующую последовательность, включающую в себя, состоящую, по сути, из или состоящую из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 91. Необязательно генетически модифицированный эндогенный Ttr локус включает в себя, состоит, по сути, из или состоит из последовательности, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, приведенной в SEQ ID NO: 16 или 17.
[0012] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных генетически модифицированный эндогенный Ttr локус не включает в себя кассету селекции или репортерный ген. В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных генетически модифицированный эндогенный Ttr локус включает в себя кассету селекции или репортерный ген. В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных геном отличного от человека животного, клетка отличного от человека животного или отличное от человека животное являются гомозиготными по генетически модифицированному эндогенному Ttr локусу. В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных геном отличного от человека животного, клетка отличного от человека животного или отличное от человека животное являются гетерозиготными по генетически модифицированному эндогенному Ttr локусу.
[0013] В некоторых таких геномах отличных от человека животных, клетках отличного от человека животного или отличных от человека животных отличным от человека животным является млекопитающее. Необязательно млекопитающее представляет собой грызуна. Необязательно грызуном является крыса или мышь. Необязательно отличное от человека животное представляет собой мышь.
[0014] Согласно другому аспекту представлены способы применения отличных от человека животных, включающих в себя гуманизированный TTR локус, для оценивания активности нацеливающихся на человеческий TTR реагентов in vivo. Такие способы могут предусматривать: (а) ведение нацеливающегося на человеческий TTR реагента любому из упомянутых выше отличных от человека животных и (b) оценивание активности нацеливающегося на человеческий TTR реагента у отличного от человека животного.
[0015] В некоторых таких способах ведение включает в себя опосредованную адено-ассоциированным вирусом (AAV) доставку, опосредованную липидной наночастицей (LNP) доставку или гидродинамическую доставку (HDD). Необязательно ведение включает в себя опосредованную LNP доставку, и необязательно доза LNP составляет от приблизительно 0,1 мг/кг до приблизительно 2 мг/кг. Необязательно ведение включает в себя опосредованную AAV8 доставку.
[0016] В некоторых таких способах стадия (b) предусматривает выделение печени из отличного от человека животного и оценивание активности нацеливающегося на человеческий TTR реагента в печени. Необязательно стадия (b) дополнительно предусматривает оценивание активности нацеливающегося на человеческий TTR реагента в органе или ткани, отличных от печени.
[0017] В некоторых таких способах нацеливающийся на человеческий TTR реагент представляет собой редактирующее геном средство, а оценивание предусматривает оценивание модификации генетически модифицированного Ttr локуса. Необязательно оценивание предусматривает измерение частоты вставок или делеций в генетически модифицированном Ttr локусе. В некоторых таких способах оценивание предусматривает измерение экспрессии Ttr информационной РНК, кодируемой генетически модифицированным Ttr локусом. В некоторых таких способах оценивание предусматривает измерение экспрессии TTR белка, кодируемого генетически модифицированным Ttr локусом. Необязательно измерение экспрессии TTR белка включает в себя измерение содержания TTR белка в сыворотке крови у отличного от человека животного. Необязательно активность оценивают в печени отличного от человека животного.
[0018] В некоторых таких способах нацеливающийся на человеческий TTR реагент включает в себя нуклеазное средство, разработанное для нацеливания на область человеческого TTR гена. Необязательно нуклеазное средство включает в себя белок Cas и направляющую РНК, разработанную для нацеливания на целевую последовательность направляющей РНК в человеческом TTR гене. Необязательно белок Cas представляет собой белок Cas9. Необязательно нацеливающийся на человеческий TTR реагент дополнительно включает в себя экзогенную донорную нуклеиновую кислоту, при этом экзогенная донорная нуклеиновая кислота разработана для рекомбинации с человеческим TTR геном. Необязательно экзогенная донорная нуклеиновая кислота представляет собой однонитевый олигодезоксинуклеотид (ssODN).
[0019] Согласно другому аспекту представлены способы оптимизации активности нацеливающегося на человеческий TTR реагента in vivo. Такие способы могут предусматривать: (I) выполнение любого из упомянутых выше способов оценивания активности нацеливающихся на человеческий TTR реагентов in vivo первый раз у первого отличного от человека животного, включающего в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как TTR кодирующую последовательность, так и некодирующую последовательность; (II) изменение переменной и выполнение способа стадии (I) во второй раз с измененной переменной у второго отличного от человека животного, включающего в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как TTR кодирующую последовательность, так и некодирующую последовательность; и (III) сравнение активности нацеливающегося на человеческий TTR реагента на стадии (I) с активностью нацеливающегося на человеческий TTR реагента на стадии (II) и выбор способа, приводящего в результате к более высокой активности. Необязательно стадия (III) может предусматривать выбор способа, приводящего в результате к более высокой эффективности, более высокой точности, более высокой однородности показателей или более высокой специфичности.
[0020] Необязательно измененная переменная на стадии (II) представляет собой способ доставки для введения нацеливающегося на человеческий TTR реагента в отличное от человека животное. Необязательно ведение включает в себя опосредованную LNP доставку, и измененная переменная на стадии (II) представляет собой состав LNP. Необязательно измененная переменная на стадии (II) представляет собой путь введения для введения нацеливающегося на человеческий TTR реагента в отличное от человека животное. Необязательно измененная переменная на стадии (II) представляет собой концентрацию или количество нацеливающегося на человеческий TTR реагента, водимого в отличное от человека животное. Необязательно измененная переменная на стадии (II) представляет собой форму нацеливающегося на человеческий TTR реагента, водимого в отличное от человека животное. Необязательно измененная переменная на стадии (II) представляет собой нацеливающийся на человеческий TTR реагент, вводимый в отличное от человека животное.
[0021] В некоторых таких способах нацеливающийся на человеческий TTR реагент включает в себя белок Cas (например, белок Cas9) и направляющую РНК, разработанную для нацеливания на целевую последовательность направляющей РНК в человеческом TTR гене. Необязательно измененная переменная на стадии (II) представляет собой последовательность направляющей РНК или целевую последовательность направляющей РНК. Необязательно каждые из белка Cas и направляющей РНК вводят в форме РНК, а измененная переменная на стадии (II) представляет собой отношение иРНК Cas к направляющей РНК. Необязательно измененная переменная на стадии (II) представляет собой модификации направляющей РНК.
[0022] Согласно другому аспекту представлены способы создания отличных от человека животных, включающих в себя гуманизированный TTR локус. Некоторые такие способы предусматривают: (а) введение в отличное от человека животное эмбриональной стволовой (ES) клетки: (i) нуклеазного средства, которое нацеливается на целевую последовательность в эндогенном Ttr локусе; и (ii) нацеливающегося вектора, включающего в себя вставку нуклеиновой кислоты, включающую в себя человеческую TTR последовательность, фланкированную 5' плечом гомологии, соответствующим 5' целевой последовательности в эндогенном Ttr локусе, и 3' плечом гомологии, соответствующим 3' целевой последовательности в эндогенном Ttr локусе, при этом нацеливающийся вектор рекомбинируется с эндогенным Ttr локусом с получением генетически модифицированной не являющейся человеческой ES клетки, включающей в свой геном генетически модифицированный эндогенный Ttr локус, включающие в себя человеческую TTR последовательность; (b) введение генетически модифицированной отличной от человеческой ES клетки в эмбрион отличного от человека животного-хозяина и (с) гестацию эмбриона отличного от человека животного-хозяина в суррогатной матери, при этом суррогатная мать производит генетически модифицированное отличное от человека животное-потомка F0, включающее в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность.
[0023] В некоторых таких способах нуклеазное средство включает в себя белок Cas (например, белок Cas9) и направляющую РНК. В некоторых таких способах нацеливающийся вектор представляет собой большой нацеливающийся вектор по меньшей мере 10 т.п.н. в длину, или в котором общая длина 5' и 3' плеч гомологии составляет по меньшей мере 10 т.п.н. В некоторых таких способах отличное от человека животное представляет собой мышь или крысу. В некоторых таких способах отличное от человека животное представляет собой мышь.
Краткое описание графических материалов
[0024] На фиг. 1А показано выравнивание человеческого и мышиного белков-предшественников транстиретина (TTR) (SEQ ID NO: 1 и 6, соответственно). Изображены сигнальный пептид, Т4 связывающий домен, границы экзон/интрон в фазе 0 и границы экзон/интрон в фазе 1/2.
[0025] На фиг. 1В показано выравнивание кодирующих человеческий и мышиный транстиретин (TTR) последовательностей (SEQ ID NO: 90 и 92, соответственно).
[0026] На фиг. 2 показаны схемы (без соблюдения масштаба) мышиного Ttr локуса дикого типа, первой версии гуманизированного мышиного Ttr локуса и второй версии гуманизированного мышиного Ttr локуса. Обозначены экзоны, интроны, 5' нетранслируемые области (UTR), 3' UTR, инициирующие кодоны (ATG), терминирующие кодоны (TGA) и SCAR loxP из кассет селекции. Белые боксы показывают мышиную последовательность; черные боксы показывают человеческую последовательность.
[0027] На фиг. 3 показана схема (без соблюдения масштаба) нацеливания для создания первой версии гуманизированного мышиного Ttr локуса. Показаны мышиный Ttr локус дикого типа, аллель F0 гуманизированного мышиного Ttr локуса с самоудаляющейся кассетой селекции неомицином (SDC-Neo) (MAID 7576) и аллель F1 гуманизированного мышиного Ttr локуса с SCAR loxP из удаленной кассеты селекции SDC-Neo (MAID 7577). Белые боксы показывают мышиную последовательность; черные боксы показывают человеческую последовательность.
[0028] На фиг. 4 показана схема (без соблюдения масштаба) нацеливания для создания второй версии гуманизированного мышиного Ttr локуса. Показаны мышиный Ttr локус дикого типа, аллель F0 гуманизированного мышиного Ttr локуса с кассетой селекции SDC-Neo и аллель F1 гуманизированного мышиного Ttr локуса с SCAR loxP из удаленной кассеты селекции SDC-Neo. Белые боксы показывают мышиную последовательность; черные боксы показывают человеческую последовательность.
[0029] На фиг. 5А показана схема (без соблюдения масштаба) стратегии для скрининга первого целевого мышиного Ttr локуса, в том числе анализов на потерю аллеля (7576mTU, 9090mTM и 9090mTD), анализов на приобретение аллеля (7576hTU, 7576hTD, Neo), анализов удерживания (9090retU, 9090retU2, 9090retU3, 9090retD, 9090retD2, 9090retD3) и анализов CRISPR, разработанных для охватывания области, которая расщепляется направляющими CRISPR (9090mTGU, mGU, 9090mTGD и mGD). Белые боксы показывают мышиную последовательность; черные боксы показывают человеческую последовательность.
[0030] На фиг. 5В показана схема (без соблюдения масштаба) стратегии для скрининга второго целевого мышиного Ttr локуса, в том числе анализов на потерю аллеля (4552mTU, 9212mTU, 9090mTM, 9212mTD), анализов на приобретение аллеля (7655hTU, 7576hTD, Neo), анализов удерживания (9204mretU, 9204mretD) и анализов CRISPR, разработанных для охватывания области, которая расщепляется направляющими CRISPR (mGU, mGD и 9212mTGD). Белые боксы показывают мышиную последовательность; черные боксы показывают человеческую последовательность.
[0031] На фиг. 6 показана экспрессия иРНК бета-актина (Actb), бета-2-микроглобулина (В2М), транстиретина Mus musculus (Mm Ttr) и транстиретина Homo sapiens (Hs TTR) в образцах печени от (1) мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3), (2) в образцах печени от мышей дикого типа, (3) в образцах селезенки от мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса, и (4) в образцах селезенки от мышей дикого типа. Более низкие значения Ct указывают на более высокую экспрессию.
[0032] На фиг. 7А и 7В показаны результаты анализов ELISA на предмет содержания человеческого TTR белка (фиг. 7А) и содержания мышиного TTR белка (фиг. 7В) в сыворотке крове и спинномозговой жидкости (CSF). Тестируемые образцы включают в себя сыворотку крови и CSF от мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3), контроли человеческой сыворотки крови и CSF, а также контроли мышиной (F1H4) сыворотки крови и CSF.
[0033] На фиг. 7С показаны результаты анализов ELISA на предмет содержания в сыворотке крови (1) человеческого TTR и (2) мышиного TTR белка. Тестируемые образцы включают в себя образцы сыворотки крови от мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3), полученных из первого клона (клона 7576C-G7), мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (МАЮ 7576; аллель F0 на фиг.3), полученных из второго клона (клона 7576А-А5), и мышей дикого типа (F1H4). Мышиную сыворотку крови и человеческую сыворотку крови использовали в качестве контролей.
[0034] На фиг. 8 показана экспрессия человеческого TTR белка, определяемая с помощью вестерн-блоттинга в образцах сыворотки крови от мышей дикого типа (F1H4), мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3), полученных из первого клона (клона 7576C-G7), и мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса, полученных из второго клона (7576А-А5). Мышиную сыворотку крови использовали в качестве отрицательного контроля, а человеческую сыворотку крови использовали в качестве положительного контроля. Мышиный IgG использовали в качестве контроля нагрузки.
[0035] На фиг. 9 показана экспрессия человеческого TTR белка, определяемая с помощью вестерн-блоттинга в образцах печени и почки от мышей дикого типа (F1H4), мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3), полученных из первого клона (клона 7576C-G7), и мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса, полученных из второго клона (7576А-А5). Мышиную сыворотку крови использовали в качестве отрицательного контроля, а человеческую сыворотку крови использовали в качестве положительного контроля. GAPDH использовали в качестве контроля нагрузки.
[0036] На фиг. 10 показан процент редактирования генома (суммарное количество вставок или делеций, наблюдаемых в суммарном количестве последовательностей, считанных в ПЦР-реакции из пула лизированных клеток) в гуманизированном мышином Ttr локусе, как определяли с помощью секвенирования следующего поколения (NGS) в первичных гепатоцитах, выделенных от мышей поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3). Тестируемые образцы включали в себя необработанные гепатоциты и гепатоциты, обработанные липидными наночастицами, содержащими иРНК Cas9 и направляющие РНК, разработанные для нацеливания на человеческий TTR.
[0037] На фиг. 11А-11Н показан химический анализ в сыворотке крови аланинаминотрансферазы (ALT) (фиг. 11А), аспартатаминотрансферазы (AST) (фиг. 11В), триглицеридов (фиг. 11С), холестерина (фиг. 11D), липопротеина высокой плотности (HDL) (фиг. 11Е), липопротеина низкой плотности (LDL) (фиг. 11F), неэстерифицированных жирных кислот (NEFA) (фиг. 11G) и альбумина (фиг 11Н) через 14 суток после инъекции липидных наночастиц, включающих в себя иРНК Cas9 и направляющие РНК, разработанные для нацеливания на человеческий TTR в мышах поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3). Ед./л относится к единицам на литр, мг/дл относится к миллиграммам на децилитр, мэкв./л относится к миллиэквивалентам на литр, а г/дл относится к граммам на децилитр.
[0038] На фиг. 12 показан процент редактирования генома (суммарное количество вставок или делеций, наблюдаемых в суммарном количестве последовательностей, считанных в ПЦР-реакции из пула лизированных клеток) в гуманизированном мышином Ttr локусе, как определяли с помощью секвенирования следующего поколения (NGS) в образцах из печени через 14 суток после инъекции буферного контроля или липидных наночастиц, включающих в себя иРНК Cas9 и направляющие РНК, разработанные для нацеливания на человеческий TTR, в мышах поколения F0, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F0 на фиг. 3).
[0039] На фиг. 13 показаны результаты ELISA, анализирующего содержание в сыворотке крови человеческого TTR у мышей дикого типа (F1H4), мышей, которым плазмиды человеческого TTR вводили путем гидродинамической доставки (HDD), и мышей, которым вводили химерную плазмиду мышиного/человеческого TTR (область, кодируемая экзоном 1, является мышиной, область, кодируемая экзонами 2-4, является человеческой) путем HDD. Показаны два отрицательных контроля, и человеческую сыворотку крови использовали в качестве положительного контроля.
[0040] На фиг. 14 показаны результаты ELISA, анализирующего содержание человеческого TTR в лизатах печени через 8 суток после инъекции буферного контроля или липидных наночастиц, включающих в себя иРНК Cas9 и направляющую РНК 1 человеческого TTR, разработанную для нацеливания на человеческий TTR, у мышей поколения F2, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F1 на фиг. 3; получали из клона 7576B-F10).
[0041] На фиг. 15А и 15В показаны результаты ELISA, анализирующего содержание человеческого TTR в образцах сыворотки крови (разбавление 1:5000 на фиг. 15А, и разбавление 1:10000 на фиг. 15В) через 8 суток после инъекции буферного контроля или липидных наночастиц, включающих в себя иРНК Cas9 и направляющую РНК 1 человеческого TTR, разработанную для нацеливания на человеческий TTR, у мышей поколения F2, гомозиготных по первой версии гуманизированного мышиного Ttr локуса (MAID 7576; аллель F1 на фиг. 3; получали из клона 7576B-F10).
[0042] На фиг. 16 показаны результаты ELISA, анализирующего содержание человеческого TTR в образцах плазмы крови мышей hTTR7577/7577, hTTR7655/7655, hTTR7655/7656, hTTR7656/7656 и hTTR7656/WT.
[0043] На фиг. 17А и 17В показаны результаты ELISA, анализирующего содержание человеческого TTR и мышиного TTR в образцах плазмы крови мышей nTTRWT/WT и hTTR7577/7577 (возрастом 3 месяца). Человеческую сыворотку крови использовали в качестве контроля.
[0044] На фиг. 17С показана экспрессия иРНК mTTR (1) и hTTR (2) в образцах печени от 3-месячных мышей hTTRWT/WT и hTTR7577/7577. Более низкие значения Ct указывают на более высокую экспрессию.
[0045] На фиг. 18 показаны результаты ELISA, анализирующего содержание человеческого TTR в образцах плазмы крови мышей дикого типа (F1H4), hTTR7577/7577 (hTTR v1) и hTTR7656/7656 (hTTRv2) (возрастом 2-3 месяца).
[0046] На фиг. 19 показан процент редактирования генома в гуманизированном мышином Ttr локусе, как определяли с помощью секвенирования следующего поколения (NGS) в образцах из печени после инъекции буферного контроля или липидных наночастиц, включающих в себя иРНК Cas9 и направляющие РНК, разработанные для нацеливания на человеческий TTR, в мышах, гомозиготных по первой версии гуманизированного мышиного Ttr локуса.
[0047] На фиг. 20 показаны результаты ELISA, анализирующего содержание человеческого TTR в образцах сыворотки крови после инъекции буферного контроля или липидных наночастиц, включающих в себя иРНК Cas9 и направляющие РНК, разработанные для нацеливания на человеческий TTR, в мышах, гомозиготных по первой версии гуманизированного мышиного Ttr локуса.
Определения
[0048] Используемые в настоящем документе взаимозаменяемо термины «белок», «полипептид» и «пептид» включают в себя полимерные формы аминокислот любой длины, в том числе кодируемые и некодируемые аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают в себя полимеры, которые были модифицированы, такие как полипептиды, имеющие модифицированные пептидные остовы. Термин «домен» относится к любой части белка или полипептида, обладающего определенной функцией или структурой.
[0049] Говорят, что белки имеют «N-конец» и «С-конец». Термин «N-конец» относится к началу белка или полипептида, оканчивающемуся аминокислотой со свободной аминогруппой (-NH2). Термин «С-конец» относится к концу аминокислотной цепи (белка или полипептида), оканчивающемуся свободной карбоксильной группой (-СООН).
[0050] Используемые в настоящем документе взаимозаменяемо термины «нуклеиновая кислота» и «полинуклеотид» включают в себя полимерные формы нуклеотидов любой длины, в том числе рибонуклеотиды, дезоксирибонуклеотиды, или их аналоги, или модифицированные версии. Они включают в себя одно-, двух- и многонитевые ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, включающие в себя пуриновые основания, пиримидиновые основания или другие натуральные, химически модифицированные, биохимически модифицированные, ненатуральные или дериватизированные нуклеотидные основания.
[0051] Говорят, что нуклеиновые кислоты имеют «5'-концы» и «3'-концы», поскольку мононуклеотиды реагируют с получением олигонуклеотидов таким способом, что 5'-фосфат одного мононуклеотидного пентозного кольца присоединяется к 3' кислороду его соседа в одном направлении через фосфодиэфирную связь. Конец олигонуклеотида называют «5'-концом», если его 5' фосфат не связан с 3' кислородом мононуклеотидного пентозного кольца. Конец олигонуклеотида называют «3'-концом», если его 3' фосфат не связан с 5' фосфатом другого мононуклеотидного пентозного кольца. Также можно сказать, что последовательность нуклеиновой кислоты, даже если она находится внутри более крупного олигонуклеотида, имеет 5' - и 3'-концы. Либо в линейной, либо в кольцевой молекуле ДНК дискретные элементы, называют либо элементами «в 3'-5' направлении» или 5', либо элементами «в 5'-3' направлении» или 3'.
[0052] Термин «геномно интегрированная» относится к нуклеиновой кислоте, которая была введена в клетку так, что нуклеотидная последовательность интегрируется в геном клетки. Любой протокол может быть использован для стабильного встраивания нуклеиновой кислоты в геном клетки.
[0053] Термины «вектор экспрессии», или «экспрессионная конструкция», или «экспрессионная кассета» относятся к рекомбинантной нуклеиновой кислоте, включающей в себя желаемую кодирующую последовательность, функционально связанную с соответствующими последовательностями нуклеиновой кислоты, необходимыми для экспрессии функционально связанной кодирующей последовательности в конкретных клетке-хозяине или организме-хозяине. Последовательности нуклеиновой кислоты, необходимые для экспрессии в прокариотах обычно включают в себя промотор, оператор (необязательный) и сайт связывания рибосомы, а также другие последовательности. Известно, что эукариотические клетки, как правило, используют промоторы, энхансеры и сигналы терминации и полиаденилирования, хотя некоторые элементы могут быть делетированы, а другие элементы добавлены с сохранением необходимой экспрессии.
[0054] Термин «нацеливающийся вектор» относится к рекомбинантной нуклеиновой кислоте, которая может быть введена с помощью гомологичной рекомбинации, опосредованного негомологичным соединением концов лигирования или любыми другими средствами рекомбинации в целевое положение в геноме клетки.
[0055] Термин «вирусный вектор» относится к рекомбинантной нуклеиновой кислоте, которая включает в себя по меньшей мере один элемент вирусного происхождения и включает в себя элементы, достаточные для упаковки в частицу вирусного вектора или обеспечивающие такую упаковку. Вектор и/или частица могут быть использованы с целью переноса ДНК, РНК или других нуклеиновых кислот в клетки либо ex vivo, либо in vivo. Известны многочисленные формы вирусных векторов.
[0056] Термин «выделенный» в отношении белков, нуклеиновых кислот и клеток включает в себя белки, нуклеиновые кислоты и клетки, которые являются относительно очищенными в отношении других компонентов клетки или организма, которые обычно могут присутствовать in situ, вплоть до и включительно, по сути, чистого препарата белка, нуклеиновой кислоты или клетки. Термин «выделенный» также включает в себя белки и нуклеиновые кислоты, которые не имеют встречающихся в природе аналога, или белков, или нуклеиновых кислот, которые были химически синтезированы и тем самым, по сути, не загрязнены другими белками или нуклеиновыми кислотами. Термин «выделенный» также включает в себя белки, нуклеиновые кислоты или клетки, которые были отделены или очищены от большей части других клеточных компонентов или компонентов организма, с которыми они встречаются в природе (например, другие клеточные белки, нуклеиновые кислоты, или клеточные, или внеклеточные компоненты).
[0057] Термин «дикий тип» включает в себя объекты, имеющие структуру и/или активность, обнаруживаемую в нормальном состоянии или контексте (в отличие от мутантного, болезненного, измененного и т.д.). Гены и полипептиды дикого типа часто существуют в нескольких разных формах (например, аллелях).
[0058] Термин «эндогенная последовательность» относится к последовательности нуклеиновой кислоты, которая встречается в природе в клетке или в отличном от человека животном. Например, эндогенная Ttr последовательность отличного от человека животного относится к нативной Ttr последовательности, которая встречается в природе в Ttr локусе у отличного от человека животного.
[0059] Термин «экзогенные» молекулы или последовательности включает в себя молекулы или последовательности, которые в норме не присутствуют в клетке в такой форме. Нормальное присутствие включает в себя присутствие в отношении конкретной стадии развития и условий окружающей среды клетки. Например, экзогенная молекула или последовательность может включать в себя мутантную версию соответствующей эндогенной последовательности в клетке, такую как гуманизированная версия эндогенной последовательности, или может включать в себя последовательность, соответствующую эндогенной последовательности в клетке, но в другой форме (т.е. не в пределах хромосомы). Напротив, эндогенные молекулы или последовательности включают в себя молекулы или последовательности, которые в норме присутствуют в такой форме в конкретной клетке на конкретной стадии развития в определенных условиях окружающей среды.
[0060] Термин «гетерологичный» при использовании в контексте нуклеиновой кислоты или белка указывает на то, что нуклеиновая кислота или белок включает в себя по меньшей мере два сегмента, которые не встречаются в природе вместе в одной и той же молекуле. Например, термин «гетерологичный» при использовании в отношении сегментов нуклеиновой кислоты или сегментов белка указывает на то, что нуклеиновая кислота или белок включает в себя две или более субпоследовательности, которые не находятся в одном и том же отношении друг к другу (например, совместном соединении) в природе. В качестве одного примера, «гетерологичная» область вектора нуклеиновой кислоты представляет собой сегмент нуклеиновой кислоты, находящийся в другой молекуле нуклеиновой кислоты или присоединенный к другой молекуле нуклеиновой кислоты, который в природе не находится в соединении с другой молекулой. Например, гетерологичная область вектора нуклеиновой кислоты может включать в себя кодирующую последовательность, фланкированную последовательностями, не встречающимися в природе в соединении с кодирующей последовательностью. Подобным образом, «гетерологичная» область белка представляет собой сегмент аминокислот, находящийся в другой пептидной молекуле или присоединенный к другой пептидной молекуле, который в природе не встречается в соединении с другой пептидной молекулой (например, белок слияния или белок с меткой). Подобным образом, нуклеиновая кислота или белок может предусматривать гетерологичную метку, или гетерологичную секрецию, или последовательность локализации.
[0061] «Оптимизация кодона» использует преимущество вырожденности кодонов, что проявляется в множественности комбинаций кодонов из трех пар оснований, которые определяют аминокислоту, и, как правило, включает в себя процесс модификации последовательности нуклеиновой кислоты для усиления экспрессии в конкретной клетке-хозяине путем замены по меньшей мере одного кодона нативной последовательности на кодон, который чаще или наиболее часто используется в генах клетки-хозяина, при сохранении нативной аминокислотной последовательности. Например, нуклеиновую кислоту, кодирующую белок Cas9, можно модифицировать с заменой кодонов, имеющих более высокую частоту использования в данной прокариотической или эукариотической клетке, в том числе в бактериальной клетке, дрожжевой клетке, человеческой клетке, не являющейся человеческой клетке, клетки млекопитающих, клетке грызунов, мышиной клетке, крысиной клетке, клетки хомяка или любой другой клетке-хозяине, по сравнению со встречающейся в природе последовательностью нуклеиновой кислоты. Таблицы частот использования кодона легко доступны, например, в «Базе данных частот использования кодона». Эти таблицы могут быть адаптированы несколькими способами. См. Nakamura et al. (2000) Nucleic Acids Research 28:292, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Также доступны компьютерные алгоритмы для оптимизации кодона конкретной последовательности для экспрессии в конкретном хозяине (см., например, Gene Forge).
[0062] Термин «локус» относится к определенной локализации гена (или характерной последовательности), последовательности ДНК, кодирующей полипептид последовательности или к положению в хромосоме генома организма. Например, термин «Ttr локус» может относиться к определенной локализации Ttr гена, Ttr последовательности ДНК, кодирующей транстиретин последовательности или к положению Ttr в хромосоме генома организма, который был идентифицирован как содержащий такую последовательность. Термин «Ttr локус» может включать в себя регуляторный элемент Ttr гена, в том числе, например, энхансер, промотор, 5' и/или 3' нетранслируемую область (UTR) или их комбинацию.
[0063] Термин «ген» относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, продукт РНК и/или полипептидный продукт) и включает в себя кодирующую область, прерываемую некодирующими нитронами, и последовательность, расположенную рядом с кодирующей областью и на 5'-, и на 3'-концах, так что ген соответствует полноразмерной иРНК (в том числе 5' и 3' нетранслируемым последовательностям). Термин «ген» также включает в себя другие некодирующие последовательности, в том числе регуляторные последовательности (например, промоторы, энхансеры и сайты связывания фактора транскрипции), сигналы полиаденилирования, внутренние участки посадки рибосомы, сайленсеры, изолирующую последовательность и участки прикрепления к матриксу. Эти последовательности могут быть расположены близко к кодирующей области гена (например, в пределах 10 т.п.н.) или на отдаленных сайтах, и они влияют на уровень или скорость транскрипции и трансляции гена.
[0064] Термин «аллель» относится к вариантной форме гена. Некоторые гены имеют ряд различных форм, которые располагаются в одном и том же положении или генном локусе в хромосоме. Диплоидный организм имеет два аллеля в каждом генном локусе. Каждая пара аллелей представляет генотип определенного генного локуса. Генотипы описывают как гомозиготные, если имеется два идентичных аллеля в определенном локусе, и как гетерозиготные, если два аллеля отличаются.
[0065] «Кодирующая область» или «кодирующая последовательность» гена, состоит из части ДНК или РНК гена, состоящей из экзонов, которая кодирует белок. Область начинается с инициирующего кодона на 5'-конце и заканчивается терминирующим ко доном на 3'-конце.
[0066] «Промотор» представляет собой регуляторную область ДНК, обычно включающую в себя ТАТА-бокс, способный направлять РНК-полимеразу II для инициации синтеза РНК по соответствующему сайту инициации транскрипции для конкретной полинуклеотидной последовательности. Промотор, кроме того, может включать в себя другие области, которые влияют на скорость инициации транскрипции. Промоторные последовательности, раскрываемые в настоящем документе, модулируют транскрипцию функционально связанного полинуклеотида. Промотор может быть активным в одном или нескольких из типов клеток, раскрываемых в настоящем документе (например, в эукариотической клетке, клетке отличных от человека млекопитающих, человеческой клетке, клетке грызуна, плюрипотентной клетке, эмбрионе на стадии одной клетки, дифференцированной клетке или их комбинации). Промотор может быть, например, конститутивно активным промотором, условным промотором, индуцибельным промотором, ограниченным по времени промотором (например, регулируемым развитием промотором) или пространственно ограниченным промотором (например, специфическим по отношению к клетке или специфическим по отношению к ткани промотором). Примеры промоторов можно найти, например, в WO 2013/176772, включенной в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[0067] Термин «функциональное связывание» или «являющийся функционально связанным» включает в себя такое смежное расположение двух или более компонентов (например, промотора и другого элемента последовательности), что оба компонента функционируют нормально, и допускается возможность того, что по меньшей мере один из компонентов может опосредовать функцию, которая влияет по меньшей мере на один из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или нескольких транскрипционных регуляторных факторов. Функциональное связывание может включать в себя такие последовательности, которые являются смежными друг по отношению к другу или действуют в транс-положении (например, регуляторная последовательность может действовать на расстоянии для контроля транскрипции кодирующей последовательности).
[0068] Термин «комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной нити нуклеиновой кислоты из-за ориентации ее групп нуклеиновых оснований образует водородные связи с другой последовательностью на противоположной нити нуклеиновой кислоты. Комплементарными основаниями в ДНК, как правило, являются А с Т и С с G. В РНК они, как правило, представляют собой С с G и U с А. Комплементарность может быть абсолютной или существенной/достаточной. Абсолютная комплементарность между двумя нуклеиновыми кислотами означает, что две нуклеиновых кислоты могут образовывать дуплекс, при этом каждое основание в дуплексе связывается с комплементарным основанием путем спаривания оснований по Уотсону-Крику. Термин «существенно» или «достаточно» комплементарная означает, что последовательность в одной нити не полностью и/или не абсолютно комплементарна последовательности в противоположной нити, но что наблюдается связывание между основаниями на двух нитях, достаточное для образования стабильного гибридного комплекса при наборе условий гибридизации (например, концентрация соли и температура). Такие условия могут быть предсказаны с использованием последовательностей и стандартных математических вычислений для предсказания Tm (температуры плавления) гибридизированных нитей или с помощью эмпирического определения Tm с использованием рутинных способов. Tm включает в себя температуру, при которой популяция комплексов гибридизации, образованных между двумя нитями нуклеиновой кислоты, денатурируются на 50% (т.е. популяция двухнитевых молекул нуклеиновой кислоты наполовину диссоциируется до отдельных нитей). При температуре ниже Tm предпочтительным является образование комплекса гибридизации, тогда как при температуре выше Tm предпочтительным является плавление или разделение нитей в комплексе гибридизации. Tm можно оценить для нуклеиновой кислоты, имеющей известное содержание G+С в водном 1 М растворе NaCl, с использованием, например, Tm=81,5+0,41 (%G+С), хотя другие известные вычисления Tm учитывают структурные характеристики нуклеиновой кислоты.
[0069] Термин «условие гибридизации» включает в себя кумулятивную среду, в которой одна нить нуклеиновой кислоты связывается со второй нитью нуклеиновой кислоты посредством взаимодействий комплементарных нитей и водородной связи с образованием комплекса гибридизации. Такие условия включают в себя химические компоненты и их концентрации (например, соли, хелатирующие средства, формамид) водного или органического раствора, включающего в себя нуклеиновые кислоты, и температуру смеси. Другие факторы, такие как продолжительность инкубации или размеры реакционной камеры, могут влиять на среду. См., например, Sambrook et al., Molecular Cloning, A Laboratory Manual, 2. sup. nd ed., pp. 1,90-1,91, 9,47-9,51, 11,47-11,57 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989), включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[0070] Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны ошибки спаривания между основаниями. Условия, подходящие для гибридизации между двумя нуклеиновыми кислотами, зависят от длины нуклеиновых кислот и степени комплементации, переменных, которые хорошо известны. Чем выше степень комплементации между двумя нуклеотидными последовательностями, тем выше значение температуры плавления (Tm) для гибридов нуклеиновых кислот, имеющих такие последовательности. Для гибридизации между нуклеиновыми кислотами с короткими участками комплементарности (например, комплементарность более 35 или меньше, 30 или меньше, 25 или меньше, 22 или меньше, 20 или меньше, или 18 или меньше нуклеотидов) важным становится положение ошибок спаривания (см. Sambrook et al., supra, 11,7-11,8). Как правило, длина для гибридизируемой нуклеиновой кислоты составляет по меньшей мере приблизительно 10 нуклеотидов. Иллюстративная минимальная длина для гибридизируемой нуклеиновой кислоты включает в себя по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 22 нуклеотида, по меньшей мере приблизительно 25 нуклеотидов и по меньшей мере приблизительно 30 нуклеотидов. Кроме того, температура и концентрация соли в промывочном растворе могут быть отрегулированы при необходимости в соответствии с факторами, такими как длина области комплементации и степени комплементации.
[0071] Последовательность полинуклеотида не должна быть на 100% комплементарна последовательности его целевой нуклеиновой кислоты, подлежащей специфической гибридизации. Кроме того, полинуклеотид может гибридизироваться с одним или несколькими сегментами, так что промежуточные или смежные сегменты не вовлекаются в событие гибридизации (например, структура петли или структура шпильки). Полинуклеотид (например, gRNA) может характеризоваться по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99% или 100% комплементарностью последовательностей в отношении целевой области в целевой последовательности нуклеиновой кислоты, на которую он нацеливается. Например, gRNA, в которой 18 из 20 нуклеотидов комплементарны целевой области, и которая, следовательно, будет специфически гибридизироваться, будет характеризоваться 90% комплементарностью. В данном примере оставшиеся некомплементарные нуклеотиды могут быть кластеризованы или перемежеваны с комплементарными нуклеотидами и не должны быть смежными друг с другом или с комплементарными нуклеотидами.
[0072] Процент комплементарности между конкретными участками последовательностей нуклеиновой кислоты в нуклеиновых кислотах может быть определен рутинным способом с использованием программы BLAST (средства поиска основного локального выравнивания) и программы PowerBLAST (Altschul et al. (1990) J. Mol. Biol. 215:403-410; Zhang and Madden (1997) Genome Res. 7:649-656, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей) или с помощью программы Gap (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, Madison Wis.) с использованием параметров по умолчанию, в которой используется алгоритм Смита и Уотермана (1981) из Adv. Appl. Math. 2:482-489, включенной в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[0073] В способах и композициях, представленных в настоящем документе, используется ряд различных компонентов. Некоторые компоненты в настоящем описании могут иметь активные варианты и фрагменты. Такие компоненты включают в себя, например, белки Cas, РНК CRISPR, tracrRNA и направляющие РНК. Биологическая активность для каждого из этих компонентов описывается в других частях настоящего документа. Термин «функциональный» относится к присущей способности белка или нуклеиновой кислоты (или их фрагмента или варианта) проявлять биологическую активность или функцию. Такие биологические активности или функции могут включать в себя, например, способность белка Cas связываться с направляющей РНК и с целевой последовательностью ДНК. Биологические функции функциональных фрагментов или вариантов могут быть одинаковыми или могут быть фактически измененными (например, в отношении их специфичности, или селективности, или эффективности) по сравнению с оригинальной, но с сохранением основной биологической функции.
[0074] Термин «вариант» относится к нуклеотидной последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, одним нуклеотидом), или к белковой последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, одной аминокислотой).
[0075] Термин «фрагмент» в отношении белка означает белок, который является более коротким или имеет меньше аминокислот, чем полноразмерный белок. Термин «фрагмент» в отношении нуклеиновой кислоты означает нуклеиновую кислоту, которая является более короткой или имеет меньше нуклеотидов, чем полноразмерная нуклеиновая кислота. Фрагмент может быть, например, N-концевым фрагментом (т.е. удаляется часть С-терминального конца белка), С-концевым фрагментом (т.е. удаляется часть N-терминального конца белка) или внутренним фрагментом.
[0076] Термин «идентичность последовательностей» или «идентичность» в контексте двух полинуклеотидов или полипептидных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия в определенном окне сравнения. Если процент идентичности последовательностей используют в отношении белков, то положения остатков, которые не идентичны, часто отличаются консервативными аминокислотными заменами, при этом аминокислотные остатки замещаются другими аминокислотными остатками с подобными химическими свойствами (например, заряд или гидрофобность), и поэтому функциональные свойства молекулы не изменяются. Если последовательности отличаются консервативными заменами, то процент идентичности последовательностей может быть повышен, чтобы скорректировать консервативный характер замены. Говорят, что последовательности, которые отличаются такими консервативными заменами, характеризуются «подобием последовательностей» или «подобием». Средства для такой корректировки хорошо известны. Как правило, они включают в себя оценку консервативной замены как частичной, а не полной ошибки спаривания, что тем самым повышает процент идентичности последовательностей. Таким образом, например, если идентичная аминокислота получает оценку 1, а неконсервативная замена получает оценку ноль, то консервативная замена получает оценку от нуля до 1. Вычисляют оценку консервативных замен, например, как это реализовано в программе PC/GENE (Intelligenetics, Mountain View, California).
[0077] Термин «процент идентичности последовательностей» включает в себя значение, определяемое путем сравнения двух оптимально выровненных последовательностей (наибольшее число абсолютно совпадающих остатков) в окне сравнения, при этом часть полинуклеотидной последовательности в окне сравнения может включать в себя добавления или делеций (т.е. гэпы) по сравнению с эталонной последовательностью (которая не включает в себя добавления или делеций) для оптимального выравнивания двух последовательностей. Процент вычисляют путем определения количества положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях, с получением числа совпадающих положений, деления количества совпадающих положений на суммарное количество положений в окне сравнения и умножения результата на 100 с получением процента идентичности последовательностей. Если не указано иное (например, более короткая последовательность включает в себя связанную гетерологичную последовательность), окно сравнения представляет собой полную длину более короткой из двух последовательностей, подлежащих сравнению.
[0078] Если не указано иное, значения идентичности/подобия последовательностей включают в себя значение, полученное с использованием GAP версии 10 с использованием следующих параметров: % идентичности и % подобия для нуклеотидной последовательности с использованием штрафа за открытие гэпа 50, штрафа за продолжение гэпа 3 и оценочной матрицы nwsgapdna.cmp; % идентичности и % подобия для аминокислотной последовательности с использованием штрафа за открытие гэпа 8, штрафа за продолжение гэпа 2 и оценочной матрицы BLOSUM62; или любой эквивалентной ей программы. Термин «эквивалентная программа» включает в себя любую программу сравнения последовательностей, которая для любых двух рассматриваемых последовательностей генерирует выравнивание, имеющее идентичные совпадения нуклеотидных или аминокислотных остатков и идентичный процент идентичности последовательностей по сравнению с соответствующим выравниванием, сгенерированным с помощью GAP версии 10.
[0079] Термин «консервативная аминокислотная замена» относится к замене аминокислоты, которая обычно присутствует в последовательности другой аминокислоты подобного размера, заряда или полярности. Примеры консервативных замен включают в себя замену неполярного (гидрофобного) остатка, такого как изолейцин, валин или лейцин другим неполярным остатком. Подобным образом, примеры консервативных замен включают в себя замену одного полярного (гидрофильного) остатка другим, например, между аргинином и лизином, между глутамином и аспарагином или между глицином и серином. Кроме того, замена основного остатка, такого как лизин, аргинин или гистидин другим или замена одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, другим кислотным остаткам являются дополнительными примерами консервативных замен. Примеры неконсервативных замен включают в себя замену неполярного (гидрофобного) аминокислотного остатка, такого как изолейцин, валин, лейцин, аланин или метионин, полярным (гидрофильным) остатком, таким как цистеин, глутамин, глутаминовая кислота или лизин, и/или полярного остатка неполярным остатком. Типичные классификации аминокислот кратко описаны в приведенной ниже таблице 1.
[0080]

[0081] Термин «гомологичная» последовательность (например, последовательность нуклеиновой кислоты) включает в себя последовательность, которая либо идентична, либо, по сути, подобна известной эталонной последовательности таким образом, что она, например, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична известной эталонной последовательности. Гомологичные последовательности могут включать в себя, например, ортологическую последовательность и паралогичные последовательности. Гомологичные гены, например, как правило, происходят от общей предковой последовательности ДНК, либо через событие видообразования (ортологические гены), либо через событие генетической дупликации (паралогичные гены). «Ортологические» гены включают в себя гены разных видов, которые произошли от общего предкового гена путем видообразования. Ортологи обычно сохраняют ту же функцию в ходе эволюции. «Паралогичные» гены включают в себя гены, связанные путем дупликации в геноме. Паралоги могут развивать новые функции в ходе эволюции.
[0082] Термин «in vitro» включает в себя искусственные среды и процессы или реакции, которые происходят в искусственной среде (например, в тестовой пробирке). Термин «in vivo» включает в себя природные среды (например, клетку, или организм, или тело) и процессы или реакции, которые происходят в природной среде. Термин «ех vivo» включает в себя клетки, которые были выделены из тела индивидуума, и процессы или реакции, которые происходят в таких клетках.
[0083] Термин «репортерный ген» относится к нуклеиновой кислоте, имеющей последовательность, кодирующую генный продукт (как правило, фермент), который легко и количественно анализировать, если конструкцию, включающую в себя последовательность репортерного гена, функционально связанную с эндогенным или гетерологичным промоторным и/или энхансерным элементом, вводят в клетки, включающие в себя (или которые могут быть выполнены с возможностью включать в себя) факторы, необходимые для активации промоторных и/или энхансерных элементов. Примеры репортерных генов включают в себя без ограничения гены, кодирующие бета-галактозидазу (lacZ), гены бактериальной хлорамфениколацетилтрансферазы (cat), гены люциферазы светлячка, гены, кодирующие бета-глюкуронидазу (GUS), и гены, кодирующие флуоресцентные белки. Термин «репортерный белок» относится к белку, кодируемому репортерным геном.
[0084] Используемый в настоящем документе термин «флуоресцентный репортерный белок» означает репортерный белок, который является выявляемым на основании флуоресценции, при этом флуоресценция может происходить либо от репортерного белка непосредственно, активности репортерного белка на флуорогенном субстрате, либо от белка с аффинностью связывания с флуоресцентно меченным соединением. Примеры флуоресцентных белков включают в себя зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP и ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP и ZsYellowl), синие флуоресцентные белки (например, BFP, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire и T-sapphire), голубые флуоресцентные белки (например, CFP, eCFP, Cerulean, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, мономер DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, DsRed-Monomer, HcRed-Tandem, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine и tdTomato), а также любой другой подходящий флуоресцентный белок, присутствие которого в клетках можно выявить с помощью способов проточной цитометрии.
[0085] Репарация в ответ на двухнитевые разрывы (DSB) происходит главным образом посредством двух консервативных путей репарации ДНК: гомологичная рекомбинация (HR) и негомологичное соединение концов (NHEJ). См. Kasparek & Humphrey (2011) Seminars in Клетка & Dev. Biol. 22:886-897, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Подобным образом, репарация целевой нуклеиновой кислоты, опосредованная экзогенной донорной нуклеиновой кислотой, может включать в себя любой процесс обмена генетической информацией между двумя полинуклеотидами.
[0086] Термин «рекомбинация» включает в себя любой процесс обмена генетической информацией между двумя полинуклеотидами, что может происходить по любому механизму. Рекомбинация может происходить путем направляемой гомологией репарации (HDR) или гомологичной рекомбинации (HR). HDR или HR включает в себя форму репарации нуклеиновой кислоты, при которой может потребоваться гомология нуклеотидных последовательностей, использует «донорную» молекулу в качестве матрицы для репарации «целевой» молекулы (т.е. той, которая испытала двухнитевый разрыв) и ведет к переносу генетической информации от донора к цели. Без углубления в какую-либо конкретную теорию предполагается, что такой перенос может включать в себя коррекцию ошибки спаривания гетеродуплексной ДНК, которая образуется между поврежденной целью и донором, и/или отжиг нитей в зависимости от синтеза, при котором донор используется для повторного синтезирования генетической информации, которая станет частью цели, и/или связанных процессов. В некоторых случаях донорный полинуклеотид, часть донорного полинуклеотида, копия донорного полинуклеотида или часть копии донорного полинуклеотида интегрируется в целевую ДНК. См. Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) PLOS ONE 7:e45768:1-9; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[0087] NHEJ включает в себя репарацию двухнитевых разрывов в нуклеиновой кислоте путем прямого лигирования концов разрывов друг с другом или с экзогенной последовательностью без необходимости в гомологичной матрице. Лигирование несмежных последовательностей с помощью NHEJ часто может приводить к делециям, вставкам или транслокациям вблизи сайта двухнитевого разрыва. Например, NHEJ также может приводить к целевой интеграции экзогенной донорной нуклеиновой кислоты посредством прямого лигирования концов разрыва с концами экзогенной донорной нуклеиновой кислоты (т.е. захват на основе NHEJ). Такая NHEJ-опосредованная целевая интеграция может быть предпочтительной для вставки экзогенной донорной нуклеиновой кислоты, когда пути направляемой гомологией репарации (HDR) сложно использовать (например, в неделящихся клетках, первичных клетках и клетках, которые плохо выполняют репарацию ДНК на основе гомологии). Кроме того, в отличие от направляемой гомологией репарации, информация, касающаяся больших областей идентичности последовательностей, фланкирующих сайт расщепления, не требуется, что может быть полезным при попытке целенаправленной вставки в организмы, у которых есть геномы, информация о геномной последовательности которых ограничена. Интеграция может происходить посредством лигирования тупых концов между экзогенной донорной нуклеиновой кислотой и расщепленной геномной последовательностью или посредством лигирования липких концов (т.е. имеющих 5' или 3' выступающие концы) с использованием экзогенной донорной нуклеиновой кислоты, которая фланкирована выступающими концами, которые совместимы с таковыми, создаваемыми нуклеазным средством в расщепленной геномной последовательности. См., например, US 2011/020722, WO 2014/033644, WO 2014/089290 и Maresca et al. (2013) Genome Res. 23(3):539-546, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Если тупые концы лигированы, может потребоваться резекция цели и/или донора для создания областей микрогомологии, необходимых для соединения фрагментов, что может привести к нежелательным изменениям в целевой последовательности.
[0088] Термин «антигенсвязывающий белок» включает в себя любой белок, который связывается с антигеном. Примеры антигенсвязывающих белков включают в себя антитело, антигенсвязывающий фрагмент антитела, мультиспецифическое антитело (например, биспецифическое антитело), scFV, bis-scFV, диатело, триатело, тетратело, V-NAR, VHH, VL, F(ab), F(ab)2, DVD (антигенсвязывающий белок с двойным вариабельным доменом), SVD (антигенсвязывающий белок с одинарным вариабельным доменом), биспецифической Т-клеточный рекрутер (BiTE) или Davisbody (см. патент США №8586713, включенный в настоящий документ посредством ссылки во всей своей полноте для всех целей).
[0089] Термин «антиген» относится к веществу, либо полной молекуле, либо домену в молекуле, которое способно индуцировать продуцирование антител со специфичностью связывания с этим веществом. Термин «антиген» также включает в себя вещества, которые в организмах-хозяевах дикого типа не будут индуцировать продуцирование антител из-за когнатного распознавания, но могут индуцировать такой ответ у животного-хозяина с соответствующей генной инженерией с нарушением иммунологической толерантности.
[0090] Термин «эпитоп» относится к сайту на антигене, с которым связывается антигенсвязывающий белок (например, антитело). Эпитоп может быть образован из смежных аминокислот или несмежных аминокислот, расположенных рядом за счет третичной укладки одного или нескольких белков. Эпитопы, образованные из смежных аминокислот (также известные как линейные эпитопы), обычно сохраняются при воздействии денатурирующих растворителей, тогда как эпитопы, образованные третичной укладкой (также известные как конформационные эпитопы), обычно теряются при обработке денатурирующими растворителями. Эпитоп, как правило, включает в себя по меньшей мере 3, а чаще по меньшей мере 5 или 8-10 аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов включают в себя, например, рентгеновскую кристаллографию и 2-мерный ядерный магнитный резонанс. См., например, Epitope Mapping Protocols, in Methods in Molecular Biology, Vol. 66, Glenn E. Morris, Ed. (1996), включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[0091] Паратоп антитела, описываемый в настоящем документе, как правило, включает в себя по меньшей мере определяющую комплементарность область (CDR), которая специфически распознает гетерологичный эпитоп (например, CDR3 область вариабельного домена тяжелой и/или легкой цепи).
[0092] Термин «антитело» включает в себя молекулы иммуноглобулина, включающие в себя четыре полипептидных цепи, две тяжелых (Н) цепи и две легких (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь включает в себя вариабельный домен тяжелой цепи и константную область тяжелой цепи (СН). Константная область тяжелой цепи включает в себя три домена: СН1, СН2 и СН3. Каждая легкая цепь включает в себя вариабельный домен легкой цепи и константную область легкой цепи (CL). Вариабельные домены тяжелой цепь и легкой цепи могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), перемежающиеся областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждый вариабельный домен тяжелой цепи и легкой цепи включает в себя три CDR и четыре FR, расположенные от амино-конца до карбокси-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут быть сокращенно названы HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут быть сокращенно названы LCDR1, LCDR2 и LCDR3). Термин «высокоаффинное» антитело относится к антителу, которое характеризуется KD в отношении своего целевого эпитопа приблизительно 10-9 М или ниже (например, приблизительно 1×10-9 М, 1×10-10 М, 1×10-11 М или приблизительно 1×10-12 М). Согласно одному варианту осуществления KD измеряют с помощью поверхностного плазмонного резонанса, например, BIACORE™; согласно другому варианту осуществления KD измеряют с помощью ELISA.
[0093] Специфическое связывание антигенсвязывающего белка с его целевым антигеном включает в себя связывание с аффинностью по меньшей мере 106, 107, 108, 109 или 1010 М-1. Специфическое связывание заметно выше по величине и отличается от неспецифического связывания, происходящего по меньшей мере с одной неродственной целью. Специфическое связывание может быть результатом образования связей между конкретными функциональными группами или определенным пространственным соответствием (например, типом замка и ключа), тогда как неспецифическое связывание обычно является результатом Ван-дер-Ваальсовых сил. Однако специфическое связывание не обязательно означает, что антигенсвязывающий белок связывается с одной и только одной целью.
[0094] Термин «антисмысловая РНК» относится к однонитевой РНК, которая комплементарна нити информационной РНК, транскрибированной в клетке.
[0095] Термин «малая интерферирующая РНК (siRNA)» относится к обычно двухнитевой молекуле РНК, которая индуцирует путь РНК-интерференции (РНКi). Эти молекулы могут варьировать по длине (обычно от 18 до 30 пар оснований) и характеризоваться варьирующей степенью комплементарности своей целевой иРНК в антисмысловой цепи. Некоторые, но не все, siRNA имеют непарные выступающие основания на 5'- или 3'-конце смысловой нити и/или антисмысловой нити. Термин «siRNA» включает дуплексы из двух отдельных нитей, а также одиночные нити, которые могут формировать шпилечные структуры, включающие в себя дуплексную область. Двухнитевая структура может иметь длину, например, менее 20, 25, 30, 35, 40, 45 или 50 нуклеотидов. Например, двухнитевая структура может иметь длину приблизительно 21-23 нуклеотида, длину приблизительно 19-25 нуклеотидов или длину приблизительно 19-23 нуклеотида.
[0096] Термин «короткая шпилечная РНК (shRNA)» относится к одной нити оснований РНК, которая самогибридизируется в структуре шпильки и может индуцировать путь РНК-интерференции (RNAi) после процессинга. Эти молекулы могут варьировать в длину (как правило, приблизительно 50-90 нуклеотидов в длину или в некоторых случаях более чем 250 нуклеотидов в длину, например, для микроРНК-адаптированной shPHK). Молекулы shRNA процессируются внутри клетки с образованием siRNA, которые, в свою очередь, могут приводить к нокдауну экспрессии генов. shRNA могут быть включены в векторы. Термин «shRNA» также относится к молекуле ДНК, из которой может быть транскрибирована короткая шпилечная молекула РНК.
[0097] Композиции или способы, «предусматривающие» или «включающие в себя» один или несколько перечисленных элементов, могут включать в себя и другие элементы, которые конкретно не указаны. Например, композиция, которая «предусматривает» или «включает в себя» белок, может включать в себя белок отдельно или в комбинации с другими ингредиентами. Переходная фраза «состоящий, по сути, из» означает, что объем формулы изобретения должен интерпретироваться как охватывающий указанные элементы, перечисленные в формуле изобретения, и элементы, которые не оказывают существенного влияния на основную(ые) и новую(ые) характеристику(и) заявляемого изобретения. Таким образом, термин «состоящий, по сути, из» при использовании в формуле изобретения не означает, что он эквивалентен термину «включающий в себя».
[0098] Термин «необязательный» или «необязательно» означает, что впоследствии описанное событие или обстоятельство может или не может произойти, и что описание включает в себя случаи, в которых событие или обстоятельство происходит, и случаи, в которых оно не происходит.
[0099] Обозначение диапазона значений включает в себя все целые числа в пределах или определяющие диапазон и все поддиапазоны, определенные целыми числами, в пределах диапазона.
[00100] Если иное не очевидно из контекста, термин «приблизительно» охватывает значения в пределах стандартного предела ошибки измерения (например, SEM) заявленного значения.
[00101] Термин «и/или» означает и охватывает все без исключения возможные комбинации одного или нескольких связанных перечисленных элементов, а также отсутствие комбинаций при интерпретации в альтернативе («или»).
[00102] Термин «или» относится к любому представителю конкретного перечня, а также включает в себя любую комбинацию представителей этого перечня.
[00103] Формы единственного числа предусматривают ссылки во множественном числе, если иное не очевидно из контекста. Например, термин «белок» или «по меньшей мере один белок» может включать в себя множество белков, в том числе их смеси.
[00104] Термин «статистически значимый» означает р ≤ 0,05.
Подробное раскрытие настоящего изобретения
I. Обзор
[00105] В настоящем документе раскрываются геномы отличных от человека животных, клетки отличного от человека животного и отличные от человека животные, включающие в своем геноме гуманизированный TTR локус, а также способы применения таких клеток отличного от человека животного и отличных от человека животных. Клетки отличного от человека животного или отличные от человека животные, включающие в себя гуманизированный TTR локус, экспрессируют человеческий белок транстиретин или химерный белок транстиретин, включающий в себя один или нескольких фрагментов человеческого белка транстиретина. Такие клетки отличного от человека животного и отличные от человека животные могут быть использованы для оценивания доставки или эффективности нацеливающихся на человеческий TTR средств (например, средств редактирования генома CRISPR/Cas9) ex vivo или in vivo и могут быть использованы в способах оптимизации доставки или эффективности таких средств ex vivo или in vivo.
[00106] В некоторых клетках отличного от человека животного и отличных от человека животных, раскрываемых в настоящем документе, большая часть или вся геномная ДНК человеческого TTR вставляется в соответствующий ортологический Ttr локус отличного от человека животного, некоторых клеток отличного от человека животного и отличных от человека животных, раскрываемых в настоящем документе, большая часть или вся геномная ДНК Ttr отличного от человека животного заменяется один к одному соответствующей ортологической геномной ДНК человеческого TTR. По сравнению с отличными от человека животными со вставками cDNA уровни экспрессии должны быть выше, если сохраняется структура интрон-экзон и механизм сплайсинга, поскольку консервативные регуляторные элементы, скорее всего, останутся нетронутыми, и сплайсированные транскрипты, которые подвергаются процессированию РНК, являются более стабильными, чем cDNA. Напротив, вставка cDNA человеческого TTR (например, вместе со вставкой искусственного интрона бета-глобулина в 5' UTR) в Ttr локус отличного от человека животного устранила бы консервативные регуляторные элементы, такие как элементы, содержащиеся в первом экзоне и интроне Ttr отличного от человека животного. Замена геномной последовательности отличного от человека животного соответствующей ортологической человеческой геномной последовательностью или вставка геномной последовательности человеческого TTR в соответствующий ортологический не являющийся человеческим Ttr локус, скорее всего, приведет к правильной экспрессии трансгена из эндогенного Ttr локуса. Подобным образом, трансгенные отличные от человека животные с трансгенной вставкой кодирующих человеческий TTR последовательностей в рандомном геномном локусе, а не в эндогенном Ttr локусе отличного от человека животного, не будут так точно отражать эндогенную регуляцию экспрессии Ttr. Гуманизированный TTR аллель, полученный в результате замены один к одному большей части или всей геномной ДНК отличного от человека животного соответствующей ортологической человеческой геномной ДНК, или вставки человеческой геномной последовательности TTR в соответствующий ортологический, не являющийся человеческим, Ttr локус, обеспечит реальную человеческую мишень или близкое подобие реальной человеческой мишени для нацеливающихся на человеческий TTR реагентов (например, реагентов, разработанную для нацеливания человеческий TTR, CRISPR/Cas9), что позволяет тестировать эффективность и способ действия таких средств на живых животных, а также выполнять фармакокинетические и фармакодинамические исследования в условиях, при которых гуманизированный белок и гуманизированный ген являются единственной версией присутствующего TTR.
II. Отличные от человека животные, включающие в себя гуманизированный TTR локус
[00107] Клетки и отличные от человека животные, раскрываемые в настоящем документе, включают в свой геном гуманизированный TTR локус. Клетки или отличные от человека животные, включающие в себя гуманизированный TTR локус, экспрессируют человеческий белок транстиретин или частично гуманизированный, химерный белок транстиретин, в котором один или несколько фрагментов нативного белка транстиретина были замещены соответствующими фрагментами из человеческого транстиретина.
А. Транстиретин (TTR)
[00108] Клетки и отличные от человека животные, описываемые в настоящем документе, включают в себя гуманизированный транстиретиновый (Ttr) локус. Транстиретин (TTR) представляет собой 55-кДа транспортный белок сыворотки крови и спинномозговой жидкости из 127 аминокислот, главным образом синтезируемый печенью, но также продуцируемый хороидным сплетением. Он также был назван преальбумином, связывающим тироксин преальбумином, ATTR, ТВРА, CTS, CTS1, HEL111, HsT2651 и PALB. В своем нативном состоянии TTR существует в виде тетрамера. В гомозиготах гомотетрамеры включают в себя идентичные богатые бета-листами субъединицы из 127 аминокислот. В гетерозиготах тетрамеры TTR могут быть собраны из вариантных субъединиц и/или субъединиц дикого типа, как правило, объединенных статистическим путем. TTR отвечает за перенос тироксина (Т4) и связанного с ретинолом RBP (связывающегося с ретинолом белка) как в сыворотке крови, так и в спинномозговой жидкости.
[00109] Если иное не очевидно из контекста, упоминание человеческого транстиретина (TTR) или его фрагментов или доменов включает в себя натуральные человеческие аминокислотные последовательности дикого типа, в том числе их изоформы и аллельные варианты. Белок-предшественник транстиретина включает в себя сигнальную последовательность (как правило, 20 аминокислот), тогда как зрелый белок транстиретина нет. Иллюстративные полипептидные последовательности TTR обозначены номерами доступа NP 000362.1 (NCBI) и Р02766.1 (UniProt) (идентичные, каждый описан под SEQ ID NO: 1). Остатки могут быть пронумерованы согласно номеру доступа в UniProt Р02766.1, при этом первая аминокислота зрелого белка (т.е. без сигнальной последовательности из 20 аминокислот) обозначена как остаток 1. В любом другом TTR белке остатки пронумерованы согласно соответствующим остаткам в номере доступа в UniProt Р02766.1 при максимальном выравнивании.
[00110] Человеческий TTR ген располагается на хромосоме 18 и включает в себя четыре экзона и три интрона. Иллюстративный человеческий TTR ген состоит из остатков 5001-12258 в последовательности, обозначенной номером доступа в GenBank NG_009490.1 (SEQ ID NO: 3). Четыре экзона в SEQ ID NO: 3 включают в себя остатки 1-205, 1130-1260, 3354-3489 и 6802-7258, соответственно. TTR кодирующая последовательность в SEQ ID NO: 3 включает в себя остатки 137-205, 1130-1260, 3354-3489 и 6802-6909. Иллюстративная иРНК человеческого TTR обозначена номером доступа в NCBI NM_000371.3 (SEQ ID NO: 4).
[00111] Мышиный Ttr ген располагается на хромосоме 18 и также включает в себя четыре экзона и три интрона. Иллюстративный мышиный Ttr ген состоит из остатков 20665250-20674326 последовательности, обозначенной номером доступа в GenBank NC_000084.6 (SEQ ID NO: 5). Четыре экзона в SEQ ID NO: 5 включают в себя остатки 1-258, 1207-1337, 4730-4865 и 8382-9077, соответственно. Ttr кодирующая последовательность в SEQ ID NO: 5 включает в себя остатки 190-258, 1207-1337, 4730-4865 и 8382-8489. Иллюстративный мышиный TTR белок обозначается номером доступа в UniProt Р07309.1 или номером доступа в NCBI NP 038725.1 (идентичны, каждый описан под SEQ ID NO: 6). Иллюстративная иРНК мышиного Ttr обозначена номером доступа в NCBI NM_013697.5 (SEQ ID NO: 7).
[00112] Иллюстративный TTR белок крысы обозначается номером доступа в UniProt Р02767. Иллюстративный TTR белок свиньи обозначается номером доступа в UniProt Р50390. Иллюстративный TTR белок курицы обозначается номером доступа в UniProt Р27731. Иллюстративный TTR белок коровы обозначается номером доступа в UniProt O46375. Иллюстративный TTR белок овцы обозначается номером доступа в UniProt P12303. Иллюстративный TTR белок шимпанзе обозначается номером доступа в UniProt Q5U7I5. Иллюстративный TTR белок орангутана обозначается номером доступа в UniProt Q5NVS2. Иллюстративный TTR белок кролика обозначается номером доступа в UniProt Р07489. Иллюстративный TTR белок яванского макака обозначается номером доступа в UniProt Q8HXW1.
[00113] Транстиретиновый (TTR) амилоидоз представляет собой системное нарушение, характеризующееся патогенным, неправильно свернутым TTR и внеклеточным отложением амилоидных фибрилл, состоящих из TTR. TTR амилоидоз обычно вызывается дестабилизацией нативной тетрамерной формы TTR (из-за окружающих или генетических условий), что приводит к диссоциации, неправильной укладке и агрегации TTR в амилоидные фибриллы, которые накапливаются в различных органах и тканях, вызывая прогрессирующую дисфункцию. Диссоциированные мономеры имеют склонность к образованию неправильно свернутых белковых агрегатов и амилоидных фибрилл.
[00114] У людей как TTR тетрамеры дикого типа, так и смешанные тетрамеры, состоящие из мутантных субъединиц и субъединиц дикого типа, могут диссоциировать, неправильно укладываться и агрегировать, при этом процесс амилоидогенеза приводит к дегенерации постмитотической ткани. Таким образом, TTR амилоидозы охватывают заболевания, вызванные патогенным неправильно свернутым TTR, возникающим в результате мутаций в TTR или в результате немутантного, неправильно свернутого TTR.
[00115] Старческий системный амилоидоз (SSA) и старческий сердечный амилоидоз (SCA) представляют собой возрастные типы амилоидоза, которые возникают в результате отложения амилоида TTR дикого типа снаружи и внутри кардиомиоцитов сердца. TTR амилоидоз также является наиболее распространенной формой наследственного (семейного) амилоидоза, который вызывается мутациями, дестабилизирующими белок TTR. TTR амилоидозы, связанные с точечными мутациями в TTR гене, включают в себя семейную амилоидную полинейропатию (FAP), семейную амилоидную кардиомиопатию (FAC) и селективный амилоидоз центральной нервной системы (CNSA).
В. Гуманизированные TTR локусы
[00116] Гуманизированный TTR локус, раскрываемый в настоящем документе, может представлять собой Ttr локус, в котором весь Ttr ген замещается соответствующей ортологической человеческой TTR последовательностью, или он может представлять собой Ttr локус, в котором только часть Ttr гена замещается соответствующей ортологической человеческой TTR последовательностью (т.е. гуманизированный). Гуманизированный TTR локус также может включать в себя человеческую TTR последовательность, вставленную в эндогенный Ttr локус без замещения соответствующей ортологической эндогенной последовательности. Человеческая TTR последовательность, соответствующая конкретному сегменту эндогенной Ttr последовательности, относится к области человеческого TTR, которая выравнивается с конкретным сегментом эндогенной Ttr последовательности при оптимальном выравнивании человеческого TTR и эндогенного Ttr. Необязательно человеческая TTR последовательность модифицируется для оптимизации кодонов на основании частоты использования кодонов у отличного от человека животного. Замененные или вставленные (т.е. гуманизированные) области могут включать в себя кодирующие области, такие как экзон, некодирующие области, такие как интрон, нетранслируемые области или регуляторные области (например, промотор, энхансер или связывающийся с транскрипционным репрессором элемент) или любую их комбинацию.
[00117] Гуманизированный TTR локус является локусом, в котором область эндогенного Ttr локуса была делетирована и заменена соответствующей ортологической человеческой TTR последовательностью (например, ортологической человеческой TTR последовательностью дикого типа). В качестве альтернативы, гуманизированный TTR локус является локусом, в котором область человеческого TTR локуса была вставлена в соответствующий эндогенный Ttr локус отличного от человека животного. В качестве одного примера, замещенная или вставленная область эндогенного Ttr локуса могут включать в себя как кодирующую последовательность (т.е. весь экзон или часть экзона), так и некодирующую последовательность (т.е. весь интрон или часть интрона), такую как по меньшей мере один экзон и по меньшей мере один интрон. Например, замещенная или вставленная область может включать в себя по меньшей мере один экзон и по меньшей мере один интрон. Замещенная или вставленная область, включающая в себя как кодирующую последовательность, так и некодирующую последовательность, может представлять собой смежную область эндогенного Ttr локуса, что означает отсутствие промежуточной последовательности между замещенной или вставленной кодирующей последовательностью и замещенной или вставленной некодирующей последовательностью. Например, замещенная или вставленная область может включать в себя по меньшей мере один экзон и по меньшей мере один соседний интрон. Замещенная или вставленная область может включать в себя один экзон, два экзона, три экзона, четыре экзона или все экзоны эндогенного Ttr локуса. Вставленная человеческая TTR последовательность может включать в себя один экзон, два экзона, три экзона, четыре экзона или все экзоны человеческого TTR гена. Подобным образом, замещенная область может включать в себя один интрон, два интрона, три интрона или все интроны эндогенного Ttr локуса. Вставленная человеческая TTR последовательность может включать в себя один интрон, два интрона, три интрона или все интроны человеческого TTR гена. Необязательно один или несколько интронов и/или один или несколько экзонов эндогенного Ttr локуса остаются немодифицированными (т.е. неделетированными и незамещенными). Например, первый экзон эндогенного Ttr локуса может оставаться немодифицированным. Подобным образом, первый экзон и первый интрон эндогенного Ttr локуса может оставаться немодифицированным.
[00118] Согласно одному конкретному примеру вся кодирующую последовательность для белка-предшественника транстиретина может быть делетирована и замещена соответствующей ортологической человеческой TTR последовательностью. Например, область эндогенного Ttr локуса, начинающаяся с инициирующего кодона и заканчивающаяся терминирующим кодоном, может быть делетирована и замещена соответствующей ортологической человеческой TTR последовательностью. Согласно другому конкретному примеру вся кодирующую последовательность для белка-предшественника транстиретина может быть вставлена. Например, область человеческого TTR локуса, начинающаяся с инициирующего кодона и заканчивающаяся терминирующим кодоном, может быть вставлена.
[00119] Фланкирующие нетранслируемые области, включающие в себя регуляторные последовательности, также могут быть гуманизированными. Первый экзон Ttr локуса, как правило, включает в себя 5' нетранслируемую область в 3'-5' направлении инициирующего кодона. Подобным образом, последний экзон Ttr локуса, как правило, включает в себя 3' нетранслируемую область в 5'-3' направлении терминирующего кодона. Области в 3'-5' направлении инициирующего кодона Ttr и в 5'-3' направлении терминирующего кодона Ttr либо могут быть немодифицированными, либо могут быть делетированы и замещены соответствующей ортологической человеческой TTR последовательностью. Например, 5' нетранслируемая область (UTR), 3' UTR или как 5' UTR, так и 3' UTR могут быть гуманизированными, или 5' UTR, 3' UTR или как 5' UTR, так и 3' UTR могут оставаться эндогенными. Согласно одному конкретному примеру 5' UTR остается эндогенной. Согласно другому конкретному примеру 3' UTR является гуманизированной, но 5' UTR остается эндогенной. Согласно другому конкретному примеру 5' UTR остается эндогенной, а человеческая 3' UTR TTR вставляется в эндогенный Ttr локус. Например, человеческая 3' UTR TTR может заменять эндогенную 3' UTR или может быть вставлена без замены эндогенной 3' UTR (например, она может быть вставлена в 3'-5' направлении эндогенной 3' UTR).
[00120] Одна или несколько областей эндогенного Ttr локуса, кодирующего один или несколько доменов белка-предшественника транстиретина, могут быть гуманизированными. Подобным образом, одна или несколько областей эндогенного Ttr локуса, кодирующего один или несколько доменов белка-предшественника транстиретина, могут оставаться немодифицированными (т.е. неделетированными и незамещенными). Например, белки-предшественники транстиретина, как правило, имеют сигнальный пептид на N-конце. Сигнальный пептид может составлять, например, приблизительно 20 аминокислот в длину. Область эндогенного Ttr локуса, кодирующая сигнальный пептид, может оставаться немодифицированной (т.е. неделетированной и незамещенной) или может быть делетирована и замещена соответствующей ортологической человеческой TTR последовательностью. Подобным образом, область эндогенного Ttr локуса, кодирующая эпитоп, распознаваемый антигенсвязывающим белком против человеческого TTR, может быть гуманизирована.
[00121] В зависимости от степени замещения соответствующими ортологическими последовательностями или от степени вставки человеческих TTR последовательностей регуляторные последовательности, такие как промотор, могут быть эндогенными или обеспеченными замещающей или вставленной человеческой ортологической последовательностью. Например, гуманизированный TTR локус может включать в себя эндогенный Ttr промотор отличного от человека животного. Кодирующая последовательность для белка-предшественника транстиретина в генетически модифицированном эндогенном Ttr локусе может быть функционально связана с эндогенным Ttr промотором. Например, человеческая TTR последовательность может быть функционально связана с эндогенным Ttr промотором.
[00122] В качестве конкретного примера, гуманизированный TTR локус может представлять собой локус, в котором область эндогенного Ttr локуса, которая делетирована и замещена соответствующей ортологической человеческой TTR последовательностью, или область человеческого TTR локуса, которая вставлена, включает в себя, состоит, по сути, из или состоит из области от инициирующего кодона до терминирующего кодона Ttr. Человеческая TTR последовательность, которая вставлена, может дополнительно включать в себя человеческую 3' UTR TTR. Например, человеческая TTR последовательность в гуманизированном TTR локусе может включать в себя, состоять, по сути, из или состоять из области от инициирующего кодона до конца 3' UTR TTR. Необязательно Ttr кодирующая последовательность в модифицированном эндогенном Ttr локусе функционально связана с эндогенным Ttr промотором. Человеческая TTR последовательность в гуманизированном TTR локусе может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 18. Гуманизированный TTR локус может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 14 или 15. Кодирующая последовательность (CDS) в гуманизированном TTR локусе может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 90 (или ее вырожденных последовательностей, которые кодируют тот же белок). Полученный в результате человеческой белок-предшественник транстиретина, кодированный гуманизированным TTR локусом, может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 1.
[00123] В качестве другого конкретного примера, гуманизированный TTR локус может представлять собой локус, в котором область эндогенного Ttr локуса, которая делетирована и замещена соответствующей ортологической человеческой TTR последовательностью, или область человеческого TTR локуса, которая вставлена, включает в себя, состоит, по сути, из или состоит из области от начала второго экзона до терминирующего кодона Ttr. Человеческая TTR последовательность, которая вставлена, может дополнительно включать в себя человеческую 3' UTR TTR. Например, человеческая TTR последовательность в гуманизированном TTR локусе может включать в себя, состоять, по сути, из или состоять из области от начала второго экзона до конца человеческой 3' UTR TTR. Необязательно Ttr кодирующая последовательность в модифицированном эндогенном Ttr локусе функционально связана с эндогенным Ttr промотором. Человеческая TTR последовательность в гуманизированном TTR локусе может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 19. Гуманизированный TTR локус может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 16 или 17. Кодирующая последовательность (CDS) в гуманизированном TTR локусе может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 91 (или ее вырожденных последовательностей, которые кодируют тот же белок). Полученный в результате химерный белок-предшественник транстиретина, кодируемый гуманизированным TTR локусом, может включать в себя, состоять, по сути, из или состоять из последовательности, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична по отношению к SEQ ID NO: 2.
[00124] TTR белок, экспрессируемый из гуманизированного TTR локуса, может быть полностью человеческим TTR белком или химерным эндогенным/человеческим TTR белком (например, если отличное от человека животное представляет собой мышь, то химерным мышиным/человеческим TTR белком). Например, сигнальный пептид белка-предшественника транстиретина может быть эндогенным, а остальная часть белка может быть человеческой. В качестве альтернативы, N-конец белка-предшественника транстиретина может быть эндогенным, а остальная часть белка может быть человеческой. Например, N-концевые 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот могут быть эндогенными, а остальная часть может быть человеческой. Согласно конкретному примеру 23 аминокислоты на N-конце являются эндогенными, а остальная часть белка является человеческой.
[00125] Необязательно гуманизированный TTR локус может включать в себя другие элементы. Примеры таких элементов могут включать в себя кассеты селекции, репортерные гены, сайты распознавания рекомбиназой или другие элементы. В качестве одного примера, гуманизированный TTR локус может включать в себя удаляемую кассету селекции (например, самоудаляющуюся кассету селекции), фланкированную последовательностями распознавания рекомбиназой (например, сайтами loxP). В качестве альтернативы, гуманизированный TTR локус может не содержать других элементов (например, может не содержать кассету селекции и/или может не содержать репортерный ген). Примеры подходящих репортерных генов и репортерных белков раскрываются в разных частях настоящего документа. Примеры подходящих селекционных маркеров включают в себя неомицин-фосфотрансферазу (neor), гигромицин-В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин-S-деаминазу (bsrr), ксантин/гуанин-фосфорибозилтрансферазу (gpt) и тримидинкиназу вируса простого герпеса (HSV-k). Примеры рекомбиназ включают в себя Cre-, Flp- и Dre-рекомбиназы. Одним примером гена Cre-рекомбиназы является Crei, в котором два экзона, кодирующие Cre-рекомбиназу, отделяются интроном для предотвращения его экспрессии в прокариотической клетке. Такие рекомбиназы могут дополнительно включать в себя сигнал ядерной локализации для облегчения локализации в ядре (например, NLS-Crei). Сайты распознавания рекомбиназой включают в себя нуклеотидные последовательности, которые распознаются сайт-специфической рекомбиназой и могут служить субстратом для события рекомбинации. Примеры сайтов распознавания рекомбиназой включают в себя FRT, FRT11, FRT71, attr, att, rox и сайты lox, такие как loP, lox511, lox2272, lox66, lox71, loxM2 и lox5171.
[00126] Другими элементами, такими как репортерные гены или кассеты селекции, могут быть самоудаляющиеся кассеты, фланкированные сайтами распознавания рекомбиназой. См., например, US 8697851 и US 2013/0312129, каждый из которых включен в настоящий документ посредством ссылки в полном своем объеме для всех целей. В качестве примера, самоудаляющаяся кассета может включать в себя ген Crei (включающий в себя два экзона, кодирующие Cre-рекомбиназу, которые разделены интроном), функционально связанный с мышиным промотором Prm1, и ген устойчивости к неомицину, функционально связанный с человеческим убиквитиновым промотором. С использованием промотора Prm1 самоудаляющаяся кассета может быть специфически делетирована в мужских зародышевых клетках животных F0. Полинуклеотид, кодирующий селекционный маркер, может быть функционально связан с промотором, активным в клетке, которая является целевой. Примеры промоторов описываются в других частях настоящего документа. В качестве другого конкретного примера, самоудаляющаяся кассета селекции может включать в себя ген устойчивости к гигромицину, кодирующий последовательность, функционально связанную с одним или несколькими промоторами (например, как человеческим убиквитиновым, так и ЕМ7 промоторами), за которым следует сигнал полиаденилирования, а затем кодирующую последовательность Crei, функционально связанную с одним или несколькими промоторами (например, промотором mPrm1), затем другой сигнал полиаденилирования, при этом полная кассета фланкируется сайтами loxP.
[00127] Гуманизированный TTR локус также может представлять собой условный аллель. Например, условный аллель может быть многофункциональным аллелем, описанным в US 2011/0104799, включенной в настоящий документ посредством ссылки в полном своем объеме для всех целей. Например, условный аллель может включать в себя: (а) запускающую последовательность в смысловой ориентации по отношению к транскрипции целевого гена; (b) кассету селекции лекарственными средствами (DSC) в смысловой или антисмысловой ориентации; (с) представляющую интерес нуклеотидную последовательность (NSI) в антисмысловой ориентации и (d) условный по инверсии модуль (COIN, в котором используются разделяющий экзоны интрон и инвертируемый модуль, подобный генной ловушке) в обратной ориентации. См., например, US 2011/0104799. Условный аллель может дополнительно включать в себя рекомбинируемые блоки, которые рекомбинируются при воздействии первой рекомбиназой с образованием условного аллеля, который (i) не имеет запускающей последовательности и DSC; и (ii) содержит NSI в смысловой ориентации и COIN в антисмысловой ориентации. См., например, US 2011/0104799.
С. Геномы отличных от человека животных, клетки отличного от человека животного и отличные от человека животные, включающие в себя гуманизированный TTR локус
[00128] Представлены геномы отличных от человека животных, клетки отличного от человека животного или отличные от человека животные, включающие в себя гуманизированный TTR локус, как описывается в других частях настоящего документа. Геномы, клетки или отличные от человека животные могут быть гетерозиготными или гомозиготными по гуманизированному TTR локусу. Диплоидный организм имеет два аллеля в каждом генном локусе. Каждая пара аллелей представляет генотип определенного генного локуса. Генотипы описывают как гомозиготные, если имеется два идентичных аллеля в определенном локусе, и как гетерозиготные, если два аллеля отличаются.
[00129] Геномы или клетки отличного от человека животного, представленные в настоящем документе, могут быть, например, любой не являющейся человеческой клеткой, включающей в себя Ttr локус или геномный локус, гомологичный или ортологический по отношению к человеческому TTR локусу. Геномы могут быть из эукариотической клетки, или клетки могут быть эукариотическими клетками, которые включают в себя, например, животные клетки, клетки млекопитающих, клетки отличных от человека млекопитающих и человеческие клетки. Термин «животное» включает в себя млекопитающих, рыб и птиц. Клеткой млекопитающего может быть, например, клетка отличного от человека млекопитающего, клетка грызуна, клетка крысы, клетка мыши или клетка хомяка. Другие отличные от человека млекопитающие включают в себя, например, отличных от человека приматов, обезьян, высших обезьян, орангутанов, кошек, собак, кроликов, лошадей, домашний скот (например, виды крупного рогатого скота, такие как коровы, быки и т.д.; виды овечьих, такие как овцы, козы и т.д., и виды свиней, такие как свиньи и кабаны). Также включены домашние животные и сельскохозяйственные животные. Термин «отличный от человека» исключает людей. [00130] Клетки также могут быть любого типа недифференцированного или дифференцированного состояния. Например, клетка может быть тотипотентной клеткой, плюрипотентной клеткой (например, человеческой плюрипотентной клеткой или не являющейся человеческой плюрипотентной клеткой, такой как мышиная эмбриональная стволовая (ES) клетка или крысиная ES клетка) или неплюрипотентной клеткой. Тотипотентные клетки включают в себя недифференцированные клетки, которые могут давать начало любому типу клеток, а плюрипотентные клетки включают в себя недифференцированные клетки, которые обладают способностью развиваться в более чем один тип дифференцированных клеток. Такие плюрипотентные и/или тотипотентные клетки могут представлять собой, например, ES клетки или подобные ES клетки, такие как индуцированные плюрипотентные стволовые (iPS) клетки. ES клетки включают в себя происходящие из эмбриона тотипотентные или плюрипотентные клетки, которые способны быть источником любой ткани развивающегося эмбриона при введении в эмбрион. ES клетки могут быть получены из внутренней клеточной массы бластулы и способны дифференцироваться в клетки любого из трех зародышевых листков позвоночных (эндодермы, эктодермы и мезодермы).
[00131] Клетки, представленные в настоящем документе, также могут быть зародышевыми клетками (например, спермиями или ооцитами). Клетки могут быть митотически компетентными клетками или митотически неактивными клетками, мейотически компетентными клетками или мейотически неактивными клетками. Подобным образом, клетки также могут быть первичными соматическими клетками или клетками, которые не являются первичной соматической клеткой. Соматические клетки включают в себя любую клетку, которая не является гаметой, зародышевой клеткой, гаметоцитом или недифференцированной стволовой клеткой. Например, клетки могут представлять собой клетки печени, такие как гепатобласты или гепатоциты.
[00132] Подходящие клетки, представленные в настоящем документе, также включают в себя первичные клетки. Первичные клетки включают в себя клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают в себя клетки, которые не являются ни трансформированными, ни иммортализованными. Они включают в себя любую клетку, полученную из организма, органа или ткани, которая ранее не высевалась в тканевую культуру или ранее высевалась в тканевую культуру, но не способна быть бесконечно пересеваемой в тканевой культуре. Такие клетки могут быть выделены традиционными методиками и включают в себя, например, гепатоциты.
[00133] Другие подходящие клетки, представленные в настоящем документе, включают в себя иммортализованные клетки. Иммортализованные клетки включают в себя клетки из многоклеточного организма, которые в норме не пролиферировали бы бесконечно долго, но из-за мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать подвергаться делению. Такие мутации или изменения могут происходить естественным путем или могут быть преднамеренно индуцированы. Конкретным примером иммортализованной клеточной линии является клеточная линия рака печени человека HepG2. Многочисленные типы иммортализованных клеток хорошо известны. Иммортализованные или первичные клетки включают в себя клетки, которые обычно используют для культивирования или для экспрессии рекомбинантных генов или белков.
[00134] Клетки, представленные в настоящем документе, также включают в себя эмбрионы на одноклеточной стадии (т.е. оплодотворенные ооциты или зиготы). Такие эмбрионы на одноклеточной стадии могут происходить из любого генетического фона (например, BALB/c, C57BL/6, 129 или их комбинации для мышей), могут быть свежими или замороженными и могут быть получены в результате естественного скрещивания или in vitro оплодотворения.
[00135] Клетки, представленные в настоящем документе, могут быть нормальными, здоровыми клетками или могут быть больными или несущими мутацию клетками.
[00136] Согласно конкретному примеру клетки отличного от человека животного являются эмбриональными стволовыми (ES) клетками или клетками печени, такими как мышиные или крысиные ES клетки или клетки печени.
[00137] Отличные от человека животные, включающие в себя гуманизированный TTR локус, как описывается в настоящем документе, могут быть получены способами, описываемыми в других частях настоящего документа. Термин «животное» включает в себя млекопитающих, рыб и птиц. Отличные от человека млекопитающие включают в себя, например, отличных от человека приматов, обезьян, высших обезьян, орангутанов, кошек, собак, лошадей, кроликов, грызунов (например, мышей, крыс, хомяков и морских свинок) и домашний скот (например, виды крупного рогатого скота, такие как коровы и быки; виды овечьих, такие как овцы и козы, и виды свиней, такие как свиньи и кабаны). Также включены домашние животные и сельскохозяйственные животные. Термин «отличное от человека животное» исключает людей. Предпочтительные отличные от человека животные включают в себя, например, грызунов, таких как мыши и крысы.
[00138] Отличные от человека животные могут быть из любого генетического фона. Например, подходящие мыши могут быть из штамма 129, штамма C57BL/6, смеси штамма 129 и C57BL/6, BALB/c или штамма Swiss Webster. Примеры штаммов 129 включают в себя 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129Sl/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1 и 129Т2. См., например, Festing et al. (1999) Mammalian Genome 10:836, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Примеры штаммов C57BL включают в себя C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal_wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Подходящие мыши также могут быть из смеси упомянутого выше штамма 129 и упомянутого выше штамма C57BL/6 (например, 50% 129 и 50% C57BL/6). Подобным образом, подходящие мыши могут быть из смеси упомянутых выше штаммов 129 или смеси упомянутых выше штаммов BL/6 (например, штаммов 129S6 (129/SvEvTac)).
[00139] Подобным образом, крысы могут быть из любого крысиного штамма, в том числе, например, крысиного штамма ACI, крысиного штамма Dark Agouti (DA), крысиного штамма Wistar, крысиного штамма LEA, крысиного штамма Sprague Dawley (SD) или крысиного штамма Fischer, такого как Fisher F344 или Fisher F6. Крысы также могут быть получены из штамма, полученного из смеси двух или более штаммов, упомянутых выше. Например, подходящая крыса может быть из штамма DA или штамма ACI. Крысиный штамм ACI характеризуется наличием окраски «черный агути» с белым брюхом и лапами и гаплотипом RT1avl. Такие штаммы доступны из различных источников, в том числе из Harlan Laboratories. Крысиный штамм Dark Agouti (DA) характеризуется наличием окраса «агути» и гаплотипом RT1av1. Такие крысы доступны из различных источников, в том числе из Charles River и Harlan Laboratories. Некоторые подходящие крысы могут быть из инбредного крысиного штамма. См., например, US 2014/0235933, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
III. Способы применения отличных от человека животных, включающих в себя гуманизированный TTR локус, для оценивания эффективности нацеливающихся на человеческий TTR реагентов in vivo или ex vivo
[00140] Представлены различные способы применения отличных от человека животных, включающих в себя гуманизированный TTR локус, как описывается в других частях настоящего документа, для оценивания или оптимизации доставки или эффективности нацеливающихся на человеческий TTR реагентов (например, терапевтических молекул или комплексов) in vivo или ex vivo. Поскольку отличные от человека животные включают в себя гуманизированный TTR локус, отличные от человека животные более точно будут отражать эффективность нацеливающегося на человеческий TTR реагента. Такие отличные от человека животные, в частности, применимы для тестирования редактирующих геном реагентов, разработанных для нацеливания на человеческий TTR ген, поскольку отличные от человека животные, раскрываемые в настоящем документе, включают в себя гуманизированные эндогенные Ttr локусы, а не трансгенные вставки человеческой TTR последовательности в рандомных геномных локусах, и гуманизированные эндогенные Ttr локусы включают в себя ортологическую человеческую геномную TTR последовательность как из кодирующих, так и из некодирующих областей, а не искусственную последовательность cDNA.
А. Способы тестирования эффективности нацеливающихся на человеческий TTR реагентов in vivo или ex vivo
[00141] Представлены различные способы оценивания доставки или эффективности нацеливающихся на человеческий TTR реагентов in vivo с использованием отличных от человека животных, включающих в себя гуманизированный TTR локус, как описывается в других частях настоящего документа. Такие способы могут предусматривать: (а) введение в отличное от человека животное нацеливающегося на человеческий TTR реагента (т.е. ведение нацеливающегося на человеческий TTR реагента отличному от человека животному) и (b) оценивание активности нацеливающегося на человеческий TTR реагента.
[00142] Нацеливающийся на человеческий TTR реагент может представлять собой любое биологическое или химическое средство, которое нацеливается на человеческий TTR локус (человеческий TTR ген), иРНК человеческого TTR или человеческий белок транстиретин. Примеры нацеливающихся на человеческий TTR реагентов раскрываются в разных частях настоящего документа и включают в себя, например, редактирующие геном средства.
Например, нацеливающийся на человеческий TTR реагент может представлять собой нацеливающуюся на TTR нуклеиновую кислоту (например, направляющие РНК CRISPR/Cas, короткие шпилечные РНК (shRNA) или малые интерферирующие РНК (siRNA)) или нуклеиновую кислоту, кодирующую нацеливающийся на TTR белок (например, белки Cas, такие как Cas9, ZFN или TALEN). В качестве альтернативы, нацеливающийся на человеческий TTR реагент может представлять собой нацеливающиеся на TTR антитело или антигенсвязывающий белок, или любую другую крупную молекулу или малую молекулу, которая нацеливается на человеческий TTR.
[00143] Такие нацеливающиеся на человеческий TTR реагенты могут быть введены любым способом доставки (например, AAV, LNP или HDD), как раскрывается более подробно в других частях настоящего документа, и любым путем введения. Средства доставки терапевтических комплексов и молекул, а также пути введения раскрывается более подробно в других частях настоящего документа. В конкретных способах реагенты, доставляли посредством AAV-опосредованной доставки. Например, AAV8 могут быть использованы для нацеливания на печень. В других конкретных способах реагенты доставляют опосредованной LNP доставкой. В других конкретных способах реагенты доставляют гидродинамической доставкой (HDD). Дозой может быть любая подходящая доза. Например, в некоторых способах, в которых реагенты (например, иРНК и gRNA Cas9) доставляют опосредованной LNP доставкой, доза может составлять от приблизительно 0,01 до приблизительно 10 мг/кг, от приблизительно 0,01 до приблизительно 5 мг/кг, от приблизительно 0,01 до приблизительно 4 мг/кг, от приблизительно 0,01 до приблизительно 3 мг/кг, от приблизительно 0,01 до приблизительно 2 мг/кг, от приблизительно 0,01 до приблизительно 1 мг/кг, от приблизительно 0,1 до приблизительно 10 мг/кг, от приблизительно 0,1 до приблизительно 6 мг/кг; от приблизительно 0,1 до приблизительно 5 мг/кг, от приблизительно 0,1 до приблизительно 4 мг/кг, от приблизительно 0,1 до приблизительно 3 мг/кг, от приблизительно 0,1 до приблизительно 2 мг/кг, от приблизительно 0,1 до приблизительно 1 мг/кг, от приблизительно 0,3 до приблизительно 10 мг/кг, от приблизительно 0,3 до приблизительно 6 мг/кг; от приблизительно 0,3 до приблизительно 5 мг/кг, от приблизительно 0,3 до приблизительно 4 мг/кг, от приблизительно 0,3 до приблизительно 3 мг/кг, от приблизительно 0,3 до приблизительно 2 мг/кг, от приблизительно 0,3 до приблизительно 1 мг/кг, приблизительно 0,1 мг/кг, приблизительно 0,3 мг/кг, приблизительно 1 мг/кг, приблизительно 2 мг/кг или приблизительно 3 мг/кг. Согласно конкретному примеру доза составляет от приблизительно 0,1 до приблизительно 6 мг/кг; от приблизительно 0,1 до приблизительно 3 мг/кг или от приблизительно 0,1 до приблизительно 2 мг/кг. Согласно конкретному примеру нацеливающийся на человеческий TTR реагент представляет собой редактирующий геном реагент, доза LNP составляет приблизительно 1 мг/кг, а процент редактирования генома по гуманизированному TTR локусу составляет от приблизительно 70% до приблизительно 80%. Согласно другому конкретному примеру нацеливающийся на человеческий TTR реагент представляет собой редактирующий геном реагент, доза LNP составляет приблизительно 0,3 мг/кг, а процент редактирования составляет от приблизительно 50% до приблизительно 80%. Согласно другому конкретному примеру нацеливающийся на человеческий TTR реагент представляет собой редактирующий геном реагент, доза LNP составляет приблизительно 0,1 мг/кг, а процент редактирования составляет от приблизительно 20% до приблизительно 80%. Согласно другому конкретному примеру доза LNP составляет приблизительно 1 мг/кг, и содержание TTR в сыворотке крови снижается на от приблизительно 0% до приблизительно 10% или от приблизительно 0% до приблизительно 35% контрольных уровней. Согласно другому конкретному примеру доза LNP составляет приблизительно 0,3 мг/кг, и содержание TTR в сыворотке крови снижается на от приблизительно 0% до приблизительно 20% или от приблизительно 0% до приблизительно 95% контрольных уровней. Согласно другому конкретному примеру доза LNP составляет приблизительно 0,1 мг/кг, и содержание TTR в сыворотке крови снижается на от приблизительно 0% до приблизительно 60% или от приблизительно 0% до приблизительно 99% контрольных уровней.
[00144] Способы оценивания активности нацеливающегося на человеческий TTR реагента хорошо известны и представлены в других частях настоящего документа. Оценивание активности может осуществляться в любом типе клеток, любом типе тканей или любом типе органов, как раскрывается в других частях настоящего документа. В некоторых способах оценивание активности осуществляют в клетках печени. Если нацеливающийся на TTR реагент представляет собой редактирующий геном реагент (например, нуклеазное средство), то такие способы могут предусматривать оценивание модификации гуманизированного TTR локуса. В качестве одного примера, оценивание может включать в себя измерение активности негомологичного соединения концов (NHEJ) по гуманизированному TTR локусу. Это может включать в себя, например, измерение частоты вставок или делеций в гуманизированном TTR локусе. Например, оценивание может включать в себя секвенирование гуманизированного TTR локуса в одной или нескольких клетках, выделенных из отличного от человека животного (например, секвенирование следующего поколения). Оценивание может включать в себя выделение целевых органа (например, печени) или ткани из отличного от человека животного и оценивание модификации гуманизированного TTR локуса в целевых органе или ткани. Оценивание также может включать в себя оценивание модификации гуманизированного TTR локуса в двух или более разных типах клеток в целевых органе или ткани. Подобным образом, оценивание может включать в себя выделение нецелевых органа или ткани (например, двух или более нецелевых органов или тканей) из отличного от человека животного и оценивание модификации гуманизированного TTR локуса в нецелевых органе или ткани.
[00145] Такие способы также могут предусматривать измерение уровней экспрессии иРНК, продуцируемой гуманизированным TTR локусом, или измерение уровней экспрессии белка, кодируемого гуманизированным TTR локусом. Например, содержание белка может быть измерено в конкретном типе клеток, тканей или органов (например, в печени), или секретируемые содержания могут быть измерены в сыворотке крови. Способы оценивания экспрессии иРНК Ttr или белка, экспрессируемого гуманизированным TTR локусом, представлены в других частях настоящего документа и хорошо известны.
[00146] Различные представленные выше способы оценивания активности in vivo также могут быть использованы для оценивания активности нацеливающихся на человеческий TTR реагентов ex vivo, как описывается в других частях настоящего документа.
[00147] В качестве одного примера, если нацеливающийся на человеческий TTR реагент представляет собой редактирующий геном реагент (например, нуклеазное средство), то может быть оценен процент редактирования по гуманизированному TTR локусу (например, в клетках печени). Например, процент редактирования (например, суммарное количество вставок или делеций, наблюдаемое в суммарном количестве последовательностей, считываемых в ПЦР-реакции из пула лизированных клеток) может составлять по меньшей мере приблизительно 10%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 99% или, например, от приблизительно 1% до приблизительно 99%, от приблизительно 10% до приблизительно 99%, от приблизительно 20% до приблизительно 99%, от приблизительно 30% до приблизительно 99%, от приблизительно 40% до приблизительно 99%, от приблизительно 50% до приблизительно 99%, от приблизительно 60% до приблизительно 99%, от приблизительно 1% до приблизительно 90%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 1% до приблизительно 80%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80% или от приблизительно 60% до приблизительно 80%.
[00148] В качестве другого примера, может быть оценено содержание TTR в сыворотке крови. Например, содержание TTR в сыворотке крови может быть снижено по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 20%, по меньшей мере на приблизительно 30%, по меньшей мере на приблизительно 40%, по меньшей мере на приблизительно 50%, по меньшей мере на приблизительно 60%, по меньшей мере на приблизительно 65%, по меньшей мере на приблизительно 70%, по меньшей мере на приблизительно 80%, по меньшей мере на приблизительно 90%, по меньшей мере на приблизительно 95%, по меньшей мере на приблизительно 99% или, например, на от приблизительно 1% до приблизительно 99%, на от приблизительно 10% до приблизительно 99%, на от приблизительно 20% до приблизительно 99%, на от приблизительно 30% до приблизительно 99%, на от приблизительно 40% до приблизительно 99%, на от приблизительно 50% до приблизительно 99%, на от приблизительно 60% до приблизительно 99%, на от приблизительно 70% до приблизительно 99%, на от приблизительно 80% до приблизительно 99%, на от приблизительно 1% до приблизительно 90%, на от приблизительно 10% до приблизительно 90%, на от приблизительно 20% до приблизительно 90%, на от приблизительно 30% до приблизительно 90%, на от приблизительно 40% до приблизительно 90%, на от приблизительно 50% до приблизительно 90%, на от приблизительно 60% до приблизительно 90%, на от приблизительно 70% до приблизительно 90% или на от приблизительно 80% до приблизительно 90%.
[00149] В некоторых способах нацеливающийся на человеческий TTR реагент представляет собой нуклеазное средство, такое как нуклеазное средство CRISPR/Cas, которое нацеливается на человеческий TTR ген. Такие способы могут предусматривать, например, (а) введение в отличное от человека животное нуклеазного средства, разработанного для расщепления человеческого TTR гена (например, белка Cas, такого как Cas9, и направляющей РНК, разработанной для нацеливания на целевую последовательность направляющей РНК в человеческом TTR гене); и (b) оценивание модификации гуманизированного TTR локуса. [00150] В случае нуклеазы CRISPR/Cas, например, будет индуцироваться модификация гуманизированного TTR локуса, если направляющая РНК формирует комплекс с белком Cas и направляет белок Cas на гуманизированный TTR локус, и комплекс Cas/направляющая РНК расщепляет целевую последовательность направляющей РНК, запуская в клетке репарацию (например, путем негомологичного соединения концов (NHEJ), если донорная последовательность отсутствует).
[00151] Необязательно могут быть введены две или более направляющих РНК, каждая из которых разработана для нацеливания на отдельную целевую последовательность направляющей РНК в человеческом TTR гене. Например, две направляющих РНК могут быть разработаны для вырезания геномной последовательности между двумя целевыми последовательностями направляющей РНК. Модификация гуманизированного TTR локуса будет индуцироваться, если первая направляющая РНК формирует комплекс с белком Cas и направляет белок Cas на гуманизированный TTR локус, вторая направляющая РНК формирует комплекс с белком Cas и направляет белок Cas на гуманизированный TTR локус, первый комплекс Cas/направляющая РНК расщепляет первую целевую последовательность направляющей РНК, и второй комплекс Cas/направляющая РНК расщепляет вторую целевую последовательность направляющей РНК, что приводит в результате к вырезанию промежуточной последовательности.
[00152] Необязательно экзогенную донорную нуклеиновую кислоту, способную рекомбинироваться с человеческим TTR геном и модифицировать его, также вводят в отличное от человека животное. Необязательно нуклеазное средство или белок Cas могут быть привязаны к экзогенной донорной нуклеиновой кислоте, как описывается в других частях настоящего документа. Модификация гуманизированного TTR локуса будет индуцироваться, например, если направляющая РНК формирует комплекс с белком Cas и направляет белок Cas на гуманизированный TTR локус, комплекс Cas/направляющая РНК расщепляет целевую последовательность направляющей РНК, и гуманизированный TTR локус рекомбинируется с экзогенной донорной нуклеиновой кислотой с модификацией гуманизированного TTR локуса. Экзогенная донорная нуклеиновая кислота может рекомбинироваться с гуманизированным TTR локусом, например, путем направляемой гомологией репарации (HDR) или путем NHEJ опосредованной вставки. Может быть использован любой тип экзогенной донорной нуклеиновой кислоты, примеры которого представлены в других частях настоящего документа.
В. Способы оптимизации доставки или эффективности нацеливающегося на человеческий TTR реагента in vivo или ex vivo
[00153] Представлены различные способы оптимизации доставки нацеливающихся на человеческий TTR реагентов в клетку или отличное от человека животное или оптимизации активности или эффективности нацеливающихся на человеческий TTR реагентов in vivo. Такие способы могут предусматривать, например, (а) выполнение способа тестирования эффективности нацеливающегося на человеческий TTR реагента, как описано выше, первый раз у первого отличного от человека животного или в первой клетке, включающих в себя гуманизированный TTR локус; (b) изменение переменной и выполнение способа во второй раз у второго отличного от человека животного (т.е. того же вида) или во второй клетке, включающих в себя гуманизированный TTR локус с измененной переменной; (с) сравнение активности нацеливающегося на человеческий TTR реагента на стадии (а) с активностью нацеливающегося на человеческий TTR реагента на стадии (b) и выбор способа, приводящего в результате к более высокой активности.
[00154] Способы измерения доставки, эффективности или активности нацеливающихся на человеческий TTR реагентов раскрываются в разных частях настоящего документа. Например, такие способы могут предусматривать измерение модификации гуманизированного TTR локуса. Более эффективная модификация гуманизированного TTR локуса может означать разные вещи в зависимости от желаемого эффекта в отличном от человека животном или клетке. Например, более эффективная модификация гуманизированного TTR локуса может означать одно, или несколько, или все из более высоких уровней модификации, более высокой точности, более высокой однородности показателей или более высокой специфичности. Более высокие уровни модификации (т.е. более высокая эффективность) гуманизированного TTR локуса относится к более высокому проценту клеток-мишеней в конкретном целевом типе клеток, в конкретной целевой ткани или в конкретном целевом органе (например, печени). Более высокая точность относится к более точной модификации гуманизированного TTR локуса (например, к более высокому проценту целевых клеток, имеющих одну и ту же модификацию или имеющих желаемую модификацию без дополнительных непредусмотренных вставок и делеций (например, инсерционно-делеционных мутаций NHEJ)). Более высокая однородность показателей относится к более соответствующей модификации гуманизированного TTR локуса среди других типов целевых клеток, тканей или органов, если нацеливанию подлежит более чем один тип клетки, ткани или органа (например, модификация большего числа клеточных типов в печени). Если нацеливанию подлежит конкретный орган, то более высокая однородность показателей также может относиться к более соответствующей модификации во всех локализациях внутри органа (например, печени). Более высокая специфичность может относиться к более высокой специфичности в отношении целевых геномного локуса или локусов, к более высокой специфичности в отношении целевого типа клеток к более высокой специфичности в отношении целевого типа тканей или к более высокой специфичности в отношении целевого органа. Например, повышенная специфичность в отношении геномного локуса относится к меньшей модификации нецелевых геномных локусов (например, к меньшему проценту целевых клеток, имеющих модификации в непредусмотренных, нецелевых геномных локусов вместо модификации целевого геномного локуса или в дополнение к таковой). Подобным образом, повышенная специфичность в отношении типа клеток, ткани или типа органа относится к меньшей модификации нецелевых типов клеток, типов тканей или типов органов, если нацеливанию подлежит конкретный тип клеток, тип тканей или тип органов (например, при нацеливании на конкретный орган (например, печень) наблюдается меньшая модификация клеток в органах или тканях, которые не предназначены для нацеливания).
[00155] Переменная, которая изменяется, может представлять собой любой параметр. В качестве одного примера, измененная переменная может представлять собой упаковку или способ доставки, с помощью которых нацеливающийся на человеческий TTR реагент или реагенты вводят в клетку или отличное от человека животное. Примеры способов доставки, таких как LNP, HDD и AAV, раскрываются в разных частях настоящего документа. Например, измененная переменная может представлять собой серотип AAV. Подобным образом, ведение может включать в себя опосредованную LNP доставку, и измененная переменная может представлять собой состав LNP. В качестве другого примера, измененная переменная может представлять собой путь введения для введения нацеливающегося на человеческий TTR реагента или реагентов в клетку или отличное от человека животное. Примеры путей введения, такие как внутривенная, интравитреальная, интрапаренхимальная и назальная инсталляция, раскрываются в разных частях настоящего документа.
[00156] В качестве другого примера, измененная переменная может представлять собой концентрацию или количество введенного нацеливающегося на человеческий TTR реагента или реагентов. В качестве другого примера, измененная переменная может представлять собой концентрацию или количество одного введенного нацеливающегося на человеческий TTR реагента (например, направляющей РНК, белка Cas или экзогенной донорной нуклеиновой кислоты) относительно концентрации или количества другого введенного нацеливающегося на человеческий TTR реагента (например, направляющей РНК, белка Cas или экзогенной донорной нуклеиновой кислоты).
[00157] В качестве другого примера, измененная переменная может представлять собой время введения нацеливающегося на человеческий TTR реагента или реагентов относительно времени оценивания активности или эффективности реагентов. В качестве другого примера, измененная переменная может представлять собой число раз или частоту, с которой вводят нацеливающийся на человеческий TTR реагент или реагенты. В качестве другого примера, измененная переменная может представлять собой время введения одного водимого нацеливающегося на человеческий TTR реагента (например, направляющей РНК, белка Cas или экзогенной донорной нуклеиновой кислоты) относительно времени введения другого вводимого нацеливающегося на человеческий TTR реагента (например, направляющей РНК, белка Cas или экзогенной донорной нуклеиновой кислоты).
[00158] В качестве другого примера, измененная переменная может представлять собой форму, в которой вводят нацеливающийся на человеческий TTR реагент или реагенты. Например, направляющая РНК может быть введена в форме ДНК или в форме РНК. Белок Cas (например, Cas9) может быть введен в форме ДНК, в форме РНК или в форме белка (например, в комплексе с направляющей РНК). Экзогенная донорная нуклеиновая кислота может представлять собой ДНК, РНК, однонитевую, двухнитевую, линейную, кольцевую и т.д. Подобным образом, каждый из компонентов может включать в себя различные комбинации модификаций для стабильности, для снижения нецелевых эффектов, для облегчения доставки и т.д. В качестве другого примера, измененная переменная может представлять собой нацеливающийся на человеческий TTR реагент или реагенты, которые вводят (например, введение другой направляющей РНК с другой последовательностью, введение другого белка Cas (например, введение другого белка Cas с другой последовательностью или нуклеиновой кислоты с другой последовательностью, но кодирующей ту же аминокислотную последовательность белка Cas) или введение другой экзогенной донорной нуклеиновой кислоты с другой последовательностью).
[00159] Согласно конкретному примеру нацеливающийся на человеческий TTR реагент включает в себя белок Cas и направляющую РНК, разработанные для нацеливания целевой последовательности направляющей РНК на человеческий TTR ген. В таких способах измененная переменная может представлять собой последовательность направляющей РНК и/или целевую последовательность направляющей РНК. В некоторых таких способах каждые из белка Cas и направляющей РНК могут быть введены в форме РНК, и измененная переменная может представлять собой отношение иРНК Cas к направляющей РНК (например, в составе LNP). В некоторых таких способах измененная переменная может представлять собой модификации направляющей РНК (например, направляющую РНК с модификацией сравнивают с направляющей РНК без модификации).
С. Нацеливающиеся на человеческий TTR реагенты
[00160] Нацеливающийся на человеческий TTR реагент может представлять собой любой реагент, который нацеливается на человеческий TTR ген, иРНК человеческого TTR или человеческий TTR белок. Например, это может быть редактирующий геном реагент, такой как нуклеазное средство, которое расщепляет целевую последовательность в человеческом TTR гене, это может быть антисмысловой олигонуклеотид, нацеливающийся на иРНК человеческого TTR, это может быть антигенсвязывающий белок, нацеливающийся на эпитоп человеческого TTR белка, или это может быть малая молекула, нацеливающаяся на человеческий TTR. Нацеливающиеся на человеческий TTR реагенты в способах, раскрываемых в настоящем документе, могут быть известными нацеливающимися на человеческий TTR реагентами, могут быть предполагаемыми нацеливающимися на TTR реагентами (например, кандидатными реагентами, разработанными для нацеливания на человеческий TTR) или могут быть реагентами, подлежащими скринингу по активности нацеливания на человеческий TTR.
(1) Нуклеазные средства, нацеливающиеся на человеческий TTR ген
[00161] Нацеливающийся на человеческий TTR реагент может представлять собой редактирующий геном реагент, такой как нуклеазное средство, которое расщепляет целевую последовательность в человеческом TTR гене. Целевая последовательность нуклеазы включает в себя последовательность ДНК, в которой нуклеазным средством индуцируется однонитевый разрыв или двухнитевый разрыв. Целевая последовательность для нуклеазного средства может быть эндогенной (или нативной) по отношению к клетке, или целевая последовательность может быть экзогенной по отношению к клетке. Целевая последовательность, которая является экзогенной по отношению к клетке, не встречается в природе в геноме клетки. Целевая последовательность также может быть экзогенной по отношению к представляющим интерес полинуклеотидам, которые желательно расположить в целевом локусе. В некоторых случаях целевая последовательность представлена только один раз в геноме клетки-хозяина.
[00162] Длина целевой последовательности может варьировать и включает в себя, например, целевые последовательности, которые составляют приблизительно 30-36 пар оснований для пары нуклеаз с «цинковыми пальцами» (ZFN) (т.е. приблизительно 15-18 пар оснований для каждой ZFN), приблизительно 36 пар оснований для подобной активаторам транскрипции эффекторной нуклеазы (TALEN) или приблизительно 20 пар оснований для направляющей РНК CRISPR/Cas9.
[00163] Любое нуклеазное средство, которое индуцирует однонитевый разрыв или двухнитевый разрыв в желаемой целевой последовательности, может быть использовано в способах и композициях, раскрываемых в настоящем документе. Может быть использовано встречающееся в природе или нативное нуклеазное средство при условии, что нуклеазное средство индуцирует однонитевый разрыв или двухнитевый разрыв в желаемой целевой последовательности. В качестве альтернативы, может быть использовано модифицированное или созданное методами генной инженерии нуклеазное средство. Термин «созданное методами генной инженерии нуклеазное средство» включает в себя нуклеазу, которая создана методами генной инженерии (модифицирована или получена) из своей нативной формы для специфического распознавания и индуцирования однонитевого разрыва или двухнитевого разрыва в желаемой целевой последовательности. Таким образом, созданное методами генной инженерии нуклеазное средство может быть получено из нативного, встречающегося в природе нуклеазного средства, или оно может быть искусственно создано или синтезировано. Созданная методами генной инженерии нуклеаза может индуцировать однонитевый разрыв или двухнитевый разрыв в целевой последовательности, например, при этом целевая последовательность не является последовательностью, которая была бы распознана нативным (не созданным методами генной инженерии или немодифицированным) нуклеазным средством. Модификация нуклеазного средства может заключаться всего лишь в одной аминокислоте в расщепляющем белок средстве или в одном нуклеотиде в расщепляющем нуклеиновую кислоту средстве. Получение однонитевого разрыва или двухнитевого разрыва в целевой последовательности или другой ДНК может упоминаться в настоящем документе как «разрезание» или «расщепление» целевой последовательности или другой ДНК.
[00164] Также представлены активные варианты и фрагменты иллюстративных целевых последовательностей. Такие активные варианты могут характеризоваться по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичностью последовательностей по отношению к данной целевой последовательности, при этом активные варианты сохраняют биологическую активность и, следовательно, могут распознаваться и расщепляться нуклеазным средством специфическим в отношении последовательности образом. Анализы для измерения двухнитевого разрыва целевой последовательности нуклеазным средством хорошо известны. См., например, Frendewey et al. (2010) Methods in Enzymology 476:295-307, которая включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00165] Целевая последовательность нуклеазного средства может быть расположена в любом месте локуса Ttr или вблизи него. Целевая последовательность может быть расположена в пределах кодирующей области Ttr гена или в пределах регуляторных областей, которые влияют на экспрессию гена. Целевая последовательность нуклеазного средства может быть расположена в интроне, экзоне, промоторе, энхансере, регуляторной области или любой не кодирующей белок области.
[00166] Одним типом нуклеазного средства является подобная активаторам транскрипции эффекторная нуклеаза (TALEN). TAL эффекторные нуклеазы представляют собой класс специфических в отношении последовательности нуклеаз, которые могут быть использованы для создания двухнитевых разрывов в специфических целевых последовательностях в геноме прокариотического или эукариотического организма. TAL эффекторные нуклеазы создают путем слияния нативного или созданного методами генной инженерии подобного активатору транскрипции (TAL) эффектора или его функциональной части с каталитическим доменом эндонуклеазы, таким как, например, FokI. Уникальный модульный TAL-эффекторный ДНК-связывающий домен обеспечивает конструирование белков с потенциально любой заданной специфичностью распознавания ДНК. Таким образом, для распознавания целевых сайтов со специфической ДНК могут быть созданы методами генной инженерии связывающие ДНК домены TAL-эффекторных нуклеаз, и, таким образом, их можно использовать для создания двухнитевых разрывов в желаемых целевых последовательностях. См. WO 2010/079430; Morbitzer et al. (2010) PNAS 10.1073/pnas.1013133107; Scholze & Boch (2010) Virulence 1:428-432; Christian et al. Genetics (2010) 186:757-761; Li et al. (2010) Nuc. Acids Res. (2010) doi:10.1093/nar/gkq704; и Miller et al. (2011) Nature Biotechnology 29:143-148, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме.
[00167] Примеры подходящих TAL нуклеаз и способов получения подходящих TAL нуклеаз раскрываются, например, в US 2011/0239315 A1, US 2011/0269234 A1, US 2011/0145940 A1, US 2003/0232410 A1, US 2005/0208489 A1, US 2005/0026157 A1, US 2005/0064474 A1, US 2006/0188987 A1 и US 2006/0063231 A1, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме. Согласно различным вариантам осуществления методами генной инженерии создают TAL эффекторные нуклеазы, которые выполняют разрез в целевой последовательности нуклеиновой кислоты или возле нее, например, в представляющем интерес локусе или представляющем интерес геномном локусе, в котором целевая последовательность нуклеиновой кислоты находится в последовательности, подлежащей модификации посредством нацеливающего вектора, или возле нее. TAL нуклеазы, подходящие для применения с различными способами и композициями, представленные в настоящем документе, включают в себя те, которые специфически разработаны для связывания на целевых последовательностях нуклеиновой кислоты, подлежащей модификации посредством нацеливающих векторов, или возле них, как описывается в настоящем документе.
[00168] В некоторых TALEN каждый мономер TALEN включает в себя 33-35 TAL-повторов, которые распознают одну пару нуклеотидов посредством двух гипервариабельных остатков. В некоторых TALEN нуклеазное средство представляет собой химерный белок, включающий в себя ДНК-связывающий домен на основе TAL-повторов, функционально связанный с независимой нуклеазой, такой как эндонуклеаза FokI. Например, нуклеазное средство может включать в себя первый ДНК-связывающий домен на основе TAL-повторов и второй ДНК-связывающий домен на основе TAL-повторов, при этом каждый из первого и второго ДНК-связывающих доменов на основе TAL-повторов функционально связан с нуклеазой FokI, при этом первый и второй ДНК-связывающие домены на основе TAL-повторов распознают две непрерывные целевые последовательности ДНК в каждой нити целевой последовательности ДНК, разделенные спейсерной последовательностью различной длины (12-20 пар оснований), и при этом субъединицы нуклеазы FokI димеризуются с образованием активной нуклеазы, которая выполняет двухнитевый разрыв в целевой последовательности.
[00169] Нуклеазное средство, применяемое в различных способах и композициях, раскрываемых в настоящем документе, может дополнительно включать в себя нуклеазу с «цинковыми пальцами» (ZFN). В некоторых ZFN каждый мономер ZFN включает в себя 3 или более ДНК-связывающих доменов на основе «цинковых пальцев», при этом каждый ДНК-связывающий домен на основе «цинковых пальцев» связывается с субсайтом из 3 пар оснований. В некоторых ZFN ZFN представляет собой химерный белок, включающий в себя ДНК-связывающий домен на основе «цинковых пальцев», функционально связанный с независимой нуклеазой, такой как эндонуклеаза FokI. Например, нуклеазное средство может включать в себя первую ZFN и вторую ZFN, при этом каждая из первой ZFN и второй ZFN функционально связана с субъединицей нуклеазы FokI, при этом первая и вторая ZFN распознают две непрерывные целевые последовательности ДНК в каждой нити целевой последовательности ДНК, разделенные спейсером из приблизительно 5-7 пар оснований, и при этом субъединицы нуклеазы FokI димеризуются с созданием активной нуклеазы, которая выполняет двухнитевый разрыв. См., например, US 20060246567; US 20080182332; US 20020081614; US 20030021776; WO/2002/057308 A2; US 20130123484; US 20100291048; WO/2011/017293 А2; и Gaj et al. (2013) Trends in Biotechnology, 31(7):397-405, каждая из которых включена в настоящий документ посредством ссылки.
[00170] Другой тип нуклеазного средства представляет собой мегануклеазу. Мегануклеазы были разделены на четыре семейства на основании консервативных мотивов последовательностей, семейства представляют собой LAGLIDADG, GIY-YIG, H-N-H и His-Cys box. Эти мотивы участвуют в координации ионов металлов и гидролизе фосфодиэфирных связей. Мегануклеазы отличаются своими длинными целевыми последовательностями и допуском некоторых видов полиморфизма последовательностей в их ДНК-субстратах. Домены, структура и функции мегануклеаз известны; см., например, Guhan and Muniyappa (2003) Crit Rev BiochemMol Biol 38:199-248; Lucas et al., (2001) Nucleic Acids Res 29:960-9; Jurica and Stoddard, (1999) Cell Mol Life Sci 55:1304-26; Stoddard, (2006) Q Rev Biophys 38:49-95; и Moure et al., (2002) Nat Struct Biol 9:764. В некоторых примерах используют встречающийся в природе вариант и/или созданное методами генной инженерии производное мегануклеазы. Известны способы модификации кинетических параметров, взаимодействия кофакторов, экспрессии, оптимальных условий и/или специфичности целевой последовательности и скрининга по активности. См., например, Epinat et al., (2003) Nucleic Acids Res 31:2952-62; Chevalier et al., (2002) Mol Cell 10:895-905; Gimble et al., (2003) Mol Biol 334:993-1008; Seligman et al., (2002) Nucleic Acids Res 30:3870-9; Sussman et al., (2004) J Mol Biol 342:31-41; Rosen et al., (2006) Nucleic Acids Res 34:4791-800; Chames et al., (2005) Nucleic Acids Res 33:e178; Smith et al., (2006) Nucleic Acids Res 34:e149; Gruenet al., (2002) Nucleic Acids Res 30: e29; Chen and Zhao, (2005) Nucleic Acids Res 33:e154; WO 2005105989; WO 2003078619; WO 2006097854; WO 2006097853; WO 2006097784 и WO 2004031346, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме.
[00171] Может быть использована любая мегануклеаза, в том числе, например, I-SceI, I-SceII, I-SceIII, I-SceIV, I-SceV, I-SceVI, I-SceVII, I-CeuI, I-CeuAIIP, I-CreI, I-CrepsbIP, I-CrepsbIIP, I-CrepsbIIIP, I-CrepsbIVP, I-TliI, I-PpoI, PI-PspI, F-SceI, F-SceII, F-SuvI, F-TevI, F-TevII, I-AmaI, I-AniI, I-ChuI, I-CmoeI, I-CpaI, I-CpaII, I-CsmI, I-CvuI, I-CvuAIP, I-DdiI, I-DdiII, I-DirI, I-DmoI, I-HmuI, I-HmuII, I-HsNIP, I-LlaI, I-MsoI, I-NaaI, I-NanI, I-NcIIP, I-NgrIP, I-NitI, I-NjaI, I-Nsp236IP, I-PakI, I-PboIP, I-PcuIP, I-PcuAI, I-PcuVI, I-PgrIP, I-PobIP, I-PorI, I-PorIIP, I-PbpIP, I-SpBetaIP, I-ScaI, I-SexIP, I-SneIP, I-SpomI, I-SpomCP, I-SpomIP, I-SpomIIP, I-SquIP, I-Ssp6803I, I-SthPhiJP, I-SthPhiST3P, I-SthPhiSTe3bP, I-TdeIP, I-TevI, I-TevII, I-TevIII, I-UarAP, I-UarHGPAIP, I-UarHGPA13P, I-VinIP, I-ZbiIP, PI-MtuI, PI-MtuHIP PI-MtuHIIP, PI-PfuI, PI-PfuII, PI-PkoI, PI-PkoII, PI-Rma43812IP, PI-SpBetaIP, PI-SceI, PI-TfuI, PI-TfuII, PI-ThyI, PI-TliI, PI-TliII или любые ее активные варианты или фрагменты.
[00172] Мегануклеазы могут распознавать, например, двухнитевые последовательности ДНК из 12-40 пар оснований. В некоторых случаях мегануклеаза распознает одну идеально соответствующую целевую последовательность в геноме.
[00173] Некоторые мегануклеазы являются хоуминг-нуклеазами. Один тип хоуминг-нуклеазы представляет собой семейство LAGLIDADG хоуминг-нуклеаз, включающее в себя, например, I-SceI, I-CreI и I-DmoI.
[00174] Нуклеазные средства, кроме того, могут включать в себя системы CRISPR/Cas, как описано более подробно ниже.
[00175] Также представлены активные варианты и фрагменты нуклеазных средств (т.е. созданное методами генной инженерии нуклеазное средство). Такие активные варианты могут характеризоваться по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше идентичностью последовательностей в отношении нативного нуклеазного средства, при этом активные варианты сохраняют способность разрезать желаемую целевую последовательность и следовательно сохранять индуцирующую однонитевый разрыв или двухнитевый разрыв активность. Например, любое из нуклеазных средств, описываемых в настоящем документе, может быть модифицировано из нативной эндонуклеазной последовательности и разработано для распознавания и индуцирования однонитевого разрыва или двухнитевого разрыва в целевой последовательности, которая не распознается нативным нуклеазным средством. Таким образом, некоторые созданные методами генной инженерии нуклеазы обладают специфичностью индуцирования однонитевого разрыва или двухнитевого разрыва в целевой последовательности, которая отличается от целевой последовательности соответствующего нативного нуклеазного средства. Анализы на предмет активности индуцирования однонитевого разрыва или двухнитевого разрыва известны и, как правило, измеряют общую активность и специфичность эндонуклеазы на ДНК-субстратах, включающих в себя целевую последовательность.
[00176] Нуклеазное средство может быть введено в клетку или отличное от человека животное известными средствами. Полипептид, кодирующий нуклеазное средство, может быть непосредственно введен в клетку или отличное от человека животное. В качестве альтернативы, полинуклеотид, кодирующий нуклеазное средство, может быть введен в клетку или отличное от человека животное. При введении полинуклеотида, кодирующего нуклеазное средство, нуклеазное средство может быть временно, условно или конститутивно экспрессировано в клетке. Полинуклеотид, кодирующий нуклеазное средство, может содержаться в экспрессионной кассете и может быть функционально связан с условным промотором, индуцибельным промотором, конститутивным промотором или специфическим по отношению к ткани промотором. Примеры промоторов обсуждаются более подробно в других частях настоящего документа. В качестве альтернативы, нуклеазное средство может быть введено в клетку в виде иРНК, кодирующей нуклеазное средство.
[00177] Полинуклеотид, кодирующий нуклеазное средство, может быть стабильно интегрирован в геном клетки и функционально связан с промотором, активным в клетке. В качестве альтернативы, полинуклеотид, кодирующий нуклеазное средство, может находиться в нацеливающемся векторе.
[00178] При обеспечении нуклеазного средства в клетке посредством введения полинуклеотида, кодирующего нуклеазное средство, такой полинуклеотид, кодирующий нуклеазное средство, может быть модифицирован с замещением кодонов, характеризующихся более высокой частотой использования в представляющей интерес клетке, по сравнению со встречающейся в природе полинуклеотидной последовательностью, кодирующей нуклеазное средство. Например, полинуклеотид, кодирующий нуклеазное средство, может быть модифицирован с замещением кодонов, характеризующихся более высокой частотой использования в данной представляющей интерес эукариотической клетке, в том числе человеческой клетке, не являющейся человеческой клетке, клетке млекопитающего, клетке грызуна, клетке мыши, клетке крысы или любой другой представляющей интерес клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью.
(2) Системы CRISPR/Cas, нацеливающиеся на человеческий TTR ген
[00179] Конкретным типом нацеливающегося на человеческий TTR реагента может быть система CRISPR/Cas, которая нацеливается на человеческий TTR ген. Системы CRISPR/Cas включают в себя транскрипты и другие элементы, участвующие в экспрессии Cas генов или управляющие активностью Cas генов. Система CRISPR/Cas может представлять собой, например, систему типа I, типа II или типа III. В качестве альтернативы, система CRISPR/Cas может представлять собой систему типа V (например, подтипа V-A или подтипа V-B). Системы CRISPR/Cas, используемые в композициях и способах, раскрываемых в настоящем документе, могут не встречаться в природе. «Невстречающаяся в природе» система включает в себя что-либо, что указывает на вмешательство человека, например, один или несколько компонентов системы изменены или мутированы по сравнению с их встречающимся в природе состоянием, при этом они по меньшей мере, по сути, не содержат по меньшей мере один другой компонент, с которым они естественно связаны в природе, или они связаны по меньшей мере с другим компонентом, с которым они не связаны в природе. Например, в невстречающихся в природе системах CRISPR/Cas могут использоваться комплексы CRISPR, включающие в себя gRNA и белок Cas, которые вместе не встречаются в природе, белок Cas, который не встречается в природе, или gRNA, которая не встречается в природе.
[00180] Белки Cas и полинуклеотиды, кодирующие белки Cas. Белки Cas, как правило, включают в себя по меньшей мере один домен распознавания или связывания РНК, который может взаимодействовать с направляющими РНК (gRNA, описанными более подробно ниже). Белки Cas могут также включать в себя нуклеазные домены (например, домены ДНКазы или РНКазы), ДНК-связывающие домены, геликазные домены, домены межбелковых взаимодействий, домены димеризации и другие домены. Некоторые такие домены (например, домены ДНКазы) могут быть из нативного белка Cas. Могут быть добавлены другие такие домены для получения модифицированного белка Cas. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновых кислот, которая включает в себя разрыв ковалентных связей молекулы нуклеиновой кислоты. В результате расщепления могут образовываться тупые концы или ступенчатые концы, и расщепление может быть однонитевым или двухнитевым. Например, белок Cas9 дикого типа, как правило, будет создавать продукт расщепления с тупыми концами. В качестве альтернативы, белок Cpf1 дикого типа (например, FnCpf1) может давать в результате продукт расщепления с 5' выступающим концом из 5 нуклеотидов, при этом расщепление происходит после 18-й пары оснований от последовательности РАМ на нецелевой нити и после 23-го основания на целевой нити. Белок Cas может обладать полной активностью расщепления с созданием двухнитевого разрыва в целевом геномном локусе (например, двухнитевого разрыва с тупыми концами), или он может представлять собой никазу, которая создает однонитевый разрыв в целевом геномном локусе.
[00181] Примеры белков Cas включают в себя Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cul966, а также их гомологи или модифицированные версии.
[00182] Иллюстративный белок Cas представляет собой белок Cas9 или белок, полученный из белка Cas9. Белки Cas9 относятся к системе CRISPR/Cas типа II и, как правило, имеют общих четыре ключевых мотива с консервативной архитектурой. Мотивы 1, 2 и 4 являются подобными RuvC мотивами, а мотив 3 является мотивом HNH. Иллюстративные белки Cas9 происходят из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, Alicyclobacillus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus, Acaryochloris marina, Neisseria meningitidis или Campylobacter jejuni. Дополнительные примеры представителей семейства Cas9 описаны в WO 2014/131833, включенной в настоящий документ посредством ссылки в полном своем объеме для всех целей. Cas9 из S. pyogenes (SpCas9) (обозначенный номером доступа в SwissProt Q99ZW2) является иллюстративным белком Cas9. Cas9 из S. aureus (SaCas9) (обозначенный номером доступа в UniProt J7RUA5) является другим иллюстративным белком Cas9. Cas9 из Campylobacter jejuni (CjCas9) (обозначенный номером доступа в UniProt Q0P897) является другим иллюстративным белком Cas9. См., например, Kim et al. (2017) Nat. Comm. 8:14500, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. SaCas9 меньше, чем SpCas9, a CjCas9 меньше, чем SaCas9 и SpCas9. Иллюстративная последовательность белка Cas9 может включать в себя, состоять, по сути, из или состоять из SEQ ID NO: 94. Иллюстративная ДНК, кодирующая белок Cas9, может включать в себя, состоять, по сути, из или состоять из SEQ ID NO: 93.
[00183] Другим примером белка Cas является белок Cpf1 (CRISPR из Prevotella и Francisella 1). Cpf1 является крупным белком (приблизительно 1300 аминокислот), который включает в себя домен подобной RuvC нуклеазы, гомологичный соответствующему домену Cas9, вместе с аналогом характерного богатого аргинином кластера Cas9. Однако в Cpf1 отсутствует нуклеазный домен HNH, который присутствует в белке Cas9s, и подобный RuvC домен является смежным в последовательности Cpf1, в отличие от Cas9, где он содержит длинные вставки, включающие в себя домен HNH. См., например, Zetsche et al. (2015) Cell 163(3):759-771, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Иллюстративные белки Cpf1 происходят из Francisella tularensis 1, Francisella tularensis subsp.novicida, Prevotella albensis, Lachnospiraceae bacterium MC2017 1, Butyrivibrio proteoclasticus, Peregrinibacteria bacterium GW2011 GWA2_33_10, Parcubacteria bacterium GW2011 GWC2_44_17, Smithella sp. SCADC, Acidaminococcus sp. BV3L6, Lachnospiraceae bacterium MA2020, CandidatusMethanoplasma termitum, Eubacterium eligens, Moraxella bovoculi 237, Leptospira inadai, Lachnospiraceae bacterium ND2006, Porphyromonas crevioricanis 3, Prevotella disiens и Porphyromonas macacae. Cpf1 из Francisella novicida U112 (FnCpf1; обозначенный номером доступа в UniProt A0Q7Q2) является иллюстративным белком Cpf1.
[00184] Белки Cas могут быть белками дикого типа (т.е. белками, которые встречаются в природе), модифицированными белками Cas (т.е. вариантами белка Cas) или фрагментами белков Cas дикого типа или модифицированных белков Cas. Белки Cas также могут быть активными вариантами или фрагментами в отношении каталитической активности белков Cas дикого типа или модифицированных белков Cas. Активные варианты или фрагменты в отношении каталитической активности могут характеризоваться по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичностью последовательностей в отношении белка Cas дикого типа или модифицированного белка Cas или их части, при этом активные варианты сохраняют способность разрезать по желаемому сайту расщепления и, следовательно, сохраняют индуцирующую однонитевый разрыв или индуцирующую двухнитевый разрыв активность. Анализы на предмет индуцирующей однонитевый разрыв или индуцирующей двухнитевый разрыв активности известны и, как правило, измеряют общую активность и специфичность белка Cas на ДНК-субстратах, включающих в себя сайт расщепления.
[00185] Белки Cas могут быть модифицированы для усиления или уменьшения одного или нескольких из аффинности связывания нуклеиновой кислоты, специфичности связывания нуклеиновой кислоты и ферментативной активности. Белки Cas также могут быть модифицированы с изменением любых других активности или свойства белка, такого как стабильность. Например, один или несколько нуклеазных доменов белка Cas могут быть модифицированы, делетированы или инактивированы, или белок Cas может быть усечен с удалением доменов, которые не являются необходимыми для функции белка или для оптимизации (например, повышения или снижения) активности или свойства белка Cas.
[00186] Одним примером модифицированного белка Cas является модифицированный белок SpCas9-HFl, который является высокоточным вариантом Cas9 Streptococcus pyogenes, содержащим изменения (N497A/R661A/Q695A/Q926A), разработанным для уменьшения неспецифических контактов ДНК. См., например, Kleinstiver et al. (2016) Nature 529(7587):490-495, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Другим примером модифицированного белка Cas является модифицированный вариант eSpCas9 (K848A/K1003A/R1060A), разработанный для уменьшения нецелевых эффектов. См., например, Slaymaker et al. (2016) Science 351(6268):84-88, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Другие варианты SpCas9 включают в себя K855А и K810A/K1003A/R1060A.
[00187] Белки Cas могут включать в себя по меньшей мере один нуклеазный домен, такой как домен ДНКазы. Например, белок Cpf1 дикого типа, как правило, включает в себя подобный RuvC домен, который расщепляет обе нити целевой ДНК, вероятно, в димерной конфигурации. Белки Cas также могут включать в себя по меньшей мере два нуклеазных домена, таких как домены ДНКазы. Например, белок Cas9 дикого типа, как правило, включает в себя подобный RuvC нуклеазный домен и подобный HNH нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать отличную нить двухнитевой ДНК с осуществлением двухнитевого разрыва в ДНК. См., например, Jinek et al. (2012) Science 337:816-821, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00188] Один, или несколько, или все из нуклеазных доменов могут быть делетированы или подвергнуты мутации таким образом, чтобы прекратить их функционирование или снизить нуклеазную активность. Например, при делеции или мутировании одного из нуклеазных доменов в белке Cas9, полученный в результате белок Cas может называться никазой и может осуществлять однонитевый разрыв в целевой последовательности направляющей РНК в пределах двухнитевой ДНК, но не двухнитевый разрыв (т.е. он может расщеплять комплементарную нить или некомплементарную нить, но не обе). При делеции или мутировании обоих нуклеазных доменов полученный в результате белок Cas (например, Cas9) будет обладать сниженной способностью к расщеплению обеих нитей двухнитевой ДНК (например, нуклеаза-пустой или нуклеаза-неактивный белок Cas или каталитически мертвый белок Cas (dCas)). Примером мутации, превращающей Cas9 в никазу, является мутация D10A (замена аспартата на аланин в положении 10 Cas9) в домене RuvC белка Cas9 из S. pyogenes. Аналогичным образом, мутация Н939А (замена гистидина на аланин в положении аминокислоты 839), Н840А (замена гистидина на аланин в положении аминокислоты 840) или N863A (замена аспарагина на аланин в положении аминокислоты N863) в домене HNH Cas9 из S. pyogenes может превращать Cas9 в никазу. Другие примеры мутаций, превращающих Cas9 в никазу, включают в себя соответствующие мутации Cas9 из S. thermophilus. См., например, Sapranauskas et al. (2011) Nucleic Acids Research 39:9275-9282, и WO 2013/141680, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Такие мутации могут быть созданы с использованием таких способов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез или синтез всего гена. Примеры других мутаций, создающих никазы, можно найти, например, в WO 2013/176772 и WO 2013/142578, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Если все из нуклеазных доменов делетированы или подвергнуты мутации в белке Cas (например, оба нуклеазных домена D10A/H840A Cas9), то полученный в результате белок Cas (например, Cas9) будет обладать сниженной способностью к расщеплению обеих нитей двухнитевой ДНК (например, нуклеаза-пустой или нуклеаза-неактивный белок Cas). Одним конкретным примером является двойной мутант Cas9 D10A/H840A S. pyogenes или соответствующий двойной мутант в Cas9 из других видов при оптимальном выравнивании с Cas9 S. pyogenes. Другим конкретным примером является двойной мутант Cas9 D10A/N863A S. pyogenes или соответствующий двойной мутант в Cas9 других видов при оптимальном выравнивании с Cas9 S. pyogenes.
[00189] Примеры инактивирующих мутаций в каталитических доменах белков Cas9 Staphylococcus aureus также известны. Например, фермент Cas9 Staphyloccocus aureus (SaCas9) может включать в себя замену в положении N580 (например, замену N580A) и замену в положении D10 (например, замену D10A) с образованием нуклеаза-неактивного белка Cas. См., например, WO 2016/106236, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00190] Примеры инактивирующих мутаций в каталитических доменах белков Cpf1 также известны. По отношению к белкам Cpf1 из Francisella novicida U112 (FnCpf1), Acidaminococcus sp. BV3L6 (AsCpf1), Lachnospiraceae bacterium ND2006 (LbCpf1) и Moraxella bovoculi 237 (MbCpf1 Cpf1) такие мутации могут включать в себя мутации в положениях 908, 993 или 1263 AsCpf1, или в соответствующих положениях в ортологах Cpf1, или в положениях 832, 925, 947 или 1180 LbCpf1, или в соответствующих положениях в ортологах Cpf1. Такие мутации могут включать в себя, например, одну или несколько из мутаций D908A, Е993А и D1263A AsCpf1, или соответствующих мутаций в ортологах Cpf1, или D832A, Е925А, D947A и D1180A LbCpf1, или соответствующих мутаций в ортологах Cpf1. См., например, US 2016/0208243, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00191] Белки Cas (например, нуклеаза-активные белки Cas или нуклеаза-неактивные белки Cas) также могут быть функционально связаны с гетерологичными полипептидами как белки слияния. Например, белок Cas может быть слит с доменом расщепления или доменом эпигенетической модификации. См. WO 2014/089290, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Белки Cas также могут быть слиты с гетерологичным полипептидом с обеспечением повышенной или пониженной стабильности. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, С-конце или внутри белка Cas.
[00192] В качестве одного примера, белок Cas может быть слит с одним или несколькими гетерологичными полипептидами, которые обеспечивают субклеточную локализацию. Такие гетерологичные полипептиды могут включать в себя, например, один или несколько сигналов ядерной локализации (NLS), таких как однокомпонентный NLS SV40 и/или двухкомпонентный NLS альфа-импортин, для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, ER удерживающий сигнал и т.п. См., например, Lange et al. (2007) J. Biol. Chem. 282:5101-5105, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Такие сигналы субклеточной локализации могут быть локализованы на N-конце, С-конце или в любом месте белка Cas. NLS может включать в себя участок основных аминокислот и может представлять собой однокомпонентную последовательность или двухкомпонентную последовательность. Необязательно белок Cas может включать в себя два или более NLS, в том числе NLS (например, NLS альфа-импортин или однокомпонентный NLS) на N-конце и NLS (например, NLS SV40 или двухкомпонентный NLS) на С-конце. Белок Cas также может включать в себя два более NLS на N-конце и/или два или более NLS на С-конце.
[00193] Белки Cas также могут быть функционально связаны с доменом проникновения в клетку или доменом трансдукции белка. Например, домен проникновения в клетку может быть полученный из белка TAT HIV-1, мотива проникновения в клетку TLM из вируса гепатита В человека, MPG, Рер-1, VP22, проникающего в клетку пептида из вируса Herpes simplex или полиаргининовой пептидной последовательности. См., например, WO 2014/089290 и WO 2013/176772, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Домен проникновения в клетку может быть локализован на N-конце, С-конце или в любом месте белка Cas.
[00194] Белки Cas также могут быть функционально связаны с гетерологичным полипептидом для облегчения отслеживания или очистки, таким как флуоресцентный белок, метка очистки или метка эпитопа. Примеры флуоресцентных белков включают в себя зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP, ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP и ZsYellowl), синие флуоресцентные белки (например, BFP, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire и T-sapphire), голубые флуоресцентные белки (например, CFP, eCFP, Cerulean, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, мономер DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, DsRed-Monomer, HcRed-Tandem, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine и tdTomato) и любой другой подходящий флуоресцентный белок. Примеры меток включают в себя глутатион-S-трансферазу (GST), хитин-связывающий белок (СВР), мальтоза-связывающий белок, тиоредоксин (TRX), поли(NANP), метку тандемной аффинной очистки (ТАР), myc, AcV5, AU1, AU5, Е, ECS, Е2, FLAG, гемагглютинин (НА), nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, S1, T7, V5, VSV-G, гистидин (His), биотин-карбоксил-переносящий белок (В ССР) и кальмодулин.
[00195] Белки Cas также могут быть привязаны к экзогенным донорным нуклеиновым кислотам или меченым нуклеиновым кислотам. Такая привязка (т.е. физическое связывание) может быть достигнута посредством ковалентных взаимодействий или нековалентных взаимодействий, и привязка может быть непосредственной (например, путем непосредственного слияния или химической конъюгации, что может быть достигнуто с помощью модификации остатков цистеина или лизина в белок или модификации интеина) или может быть достигнута посредством одного или нескольких промежуточных линкеров или адаптерных молекул, таких как стрептавидин, или аптамеров. См., например, Pierce et al. (2005) Mini Rev. Med. Chem. 5(1):41-55; Duckworth et al. (2007) Angew. Chem. Int. Ed. Engl. 46(46):8819-8822; Schaeffer и Dixon (2009) Australian J. Chem. 62(10): 1328-1332; Goodman et al. (2009) Chembiochem. 10(9): 1551-1557; и Khatwani et al. (2012) Bioorg. Med. Chem. 20(14):4532-4539, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Нековалентные стратегии для синтеза конъюгатов белок-нуклеиновая кислота включают в себя способы с использованием биотина-стрептавидина и никеля-гистидина. Ковалентные конъюгаты белок-нуклеиновая кислота могут быть синтезированы путем соединения соответствующим образом функционализированных нуклеиновых кислот и белков с использованием широкого ряда химических соединений. Некоторые из таких химических соединений вовлекаются непосредственно в присоединения олигонуклеотида к аминокислотному остатку на поверхности белка (например, к амину лизина или тиолуцистеина), в то время как другие более сложные схемы требуют посттрансляционной модификации белка или участия каталитического или реактивного домена белка. Способы ковалентного присоединения белков к нуклеиновым кислотам могут включать в себя, например, химическое сшивание олигонуклеотидов с остатками лизина или цистеина белка, лигирование экспрессируемого белка, хемоферментативные способы и применение фотоаптамеров. Экзогенная донорная нуклеиновая кислота или меченая нуклеиновая кислота могут быть привязаны к С-концу, N-концу или внутренней области внутри белка Cas. В одном примере экзогенная донорная нуклеиновая кислота или меченая нуклеиновая кислота привязаны к С-концу или N-концу белка Cas. Подобным образом, белок Cas может быть привязан к 5'-концу, 3'-концу или к внутренней области внутри экзогенной донорной нуклеиновой кислоты или меченой нуклеиновой кислоты. То есть экзогенная донорная нуклеиновая кислота или меченая нуклеиновая кислота могут быть привязаны в любой ориентации и полярности. Например, белок Cas может быть привязан к 5'-концу или 3'-концу экзогенной донорной нуклеиновой кислоты или меченой нуклеиновой кислоты.
[00196] Белки Cas могут быть обеспечены в любой форме. Например, белок Cas может быть обеспечен в форме белка, такого как белок Cas, в комплексе с gRNA. В качестве альтернативы, белок Cas может быть обеспечен в форме нуклеиновой кислоты, кодирующей белок Cas, такой как РНК (например, информационная РНК (иРНК)) или ДНК. Необязательно нуклеиновая кислота, кодирующая белок Cas, может быть оптимизирована по кодонам для эффективной трансляции в белок в конкретных клетке или организме. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована с замещением кодонов, имеющих более высокую частоту использования, в бактериальной клетке, дрожжевой клетке, человеческой клетке, не являющейся человеческой клетке, клетке млекопитающего, клетке грызуна, клетке мыши, клетке крысы или любой другой представляющей интерес клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью. При введении нуклеиновой кислоты, кодирующей белок Cas, в клетку белок Cas может быть временно, условно, или конститутивно экспрессироваться в клетке.
[00197] Белки Cas, обеспеченные в виде иРНК, могут быть модифицированы для улучшения свойств стабильности и/или иммуногенности. Модификации могут быть выполнены с одним или несколькими нуклеозидами в иРНК. Примеры химических модификаций в нуклеиновых основаниях иРНК включают в себя псевдоуредин, 1-метил-псевдоуредин и 5-метил-цитидин. Например, может быть использована кэппированная и полиаденилированная иРНК Cas, включающая в себя N1-метил-псевдоуредин. Подобным образом, иРНК Cas могут быть модифицированы с помощью истощения уредина с использованием синонимичных кодонов.
[00198] Нуклеиновые кислоты, кодирующие белки Cas, могут быть стабильно интегрированы в геном клетки и функционально связаны с промотором, активным в клетке. В качестве альтернативы, нуклеиновые кислоты, кодирующие белки Cas, могут быть функционально связаны с промотором в экспрессионной конструкции. Экспрессионные конструкции включают в себя любые конструкции нуклеиновых кислот, способные управлять экспрессией гена или другой представляющей интерес последовательности нуклеиновой кислоты (например, Cas гена), и которые могут переносить такую представляющую интерес последовательность нуклеиновой кислоты в целевую клетку. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в нацеливающемся векторе, включающем в себя вставку нуклеиновой кислоты, и/или в векторе, включающем в себя ДНК, кодирующую gRNA. В качестве альтернативы, она может находиться в векторе или плазмиде, которая отделяется от нацеливающегося вектора, включающего в себя вставку нуклеиновой кислоты, и/или отделяется от вектора, включающего в себя ДНК, кодирующую gRNA. Промоторы, которые могут быть использованы в экспрессионной конструкции, включают в себя промоторы, активные, например, в одной или нескольких из эукариотической клетки, человеческой клетки, не являющейся человеческой клетки, клетки млекопитающего, клетки отличного от человека млекопитающего, клетки грызуна, клетки мыши, клетки крысы, клетки хомяка, клетки кролика, плюрипотентной клетки, эмбриональной стволовой (ES) клетки или зиготе. Такие промоторы могут представлять собой, например, условные промоторы, индуцибельные промоторы, конститутивные промоторы или специфические по отношению к ткани промоторы. Необязательно промотор может быть двунаправленным промотором, управляющим экспрессией как белка Cas в одном направлении, так и направляющей РНК в другом направлении. Такие двунаправленные промоторы могут состоять из (1) полного традиционного однонаправленного промотора Pol III, который содержит 3 внешних контрольных элемента: элемент дистальной последовательности (DSE), элемент проксимальной последовательности (PSE) и ТАТА-бокс; и (2) второго основного промотора Pol III, который включает в себя PSE и ТАТА-бокс, слитый с 5'-концом DSE в обратной ориентации. Например, в промоторе H1 DSE соседствует с PSE и ТАТА-боксом, и промотор можно сделать двунаправленным путем создания гибридного промотора, в котором транскрипция в обратном направлении контролируется путем добавления PSE и ТАТА-бокса, полученного из промотора U6. См., например, US 2016/0074535, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Применение двунаправленного промотора для экспрессии генов, кодирующих белок Cas и направляющую РНК, одновременно позволяет создавать компактные экспрессионные кассеты для облегчения доставки.
[00199] Направляющие РНК. Термин «направляющая РНК» или «gRNA» представляет собой молекулу РНК, которая связывается с белком Cas (например, белком Cas9) и нацеливает белок Cas в определенное местоположение в целевой ДНК. Направляющие РНК могут включать в себя два сегмента: «нацеливающийся на ДНК сегмент» и «связывающийся с белком сегмент». Термин «сегмент» включает в себя сектор или область молекулы, например, смежный участок нуклеотидов в РНК. Некоторые gRNA, такие как gRNA для Cas9, могут включать в себя две отдельных молекулы РНК: «РНК-активатор» (например, tracrRNA) и «РНК-нацеливатель» (например, РНК CRISPR или crRNA). Другие gRNA представляют собой одинарные молекулы РНК (одинарный полинуклеотид РНК), которые также можно назвать «одномолекулярной gRNA», «одиночной направляющей РНК» или «sgRNA». См., например, WO 2013/176772, WO 2014/065596, WO 2014/089290, WO 2014/093622, WO 2014/099750, WO 2013/142578 и WO 2014/131833, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Для Cas9, например, однонаправляющая РНК может включать в себя crRNA, слитую с tracrRNA (например, через линкер). Для Cpf1, например, необходима только crRNA для достижения связывания с целевой последовательностью и/или расщепления целевой последовательности. Термины «направляющая РНК» и «gRNA» включают в себя как двухмолекулярные (т.е. модульные) gRNA, так и одномолекулярные gRNA.
[00200] Иллюстративная двухмолекулярная gRNA включает в себя crRNA-подобную («РНК CRISPR», или «РНК-нацеливатель», или «crRNA», или «повтор crRNA») молекулу и соответствующую tracrRNA-подобную («транс-действующую РНК CRISPR», или «РНК-активатор», или «tracrRNA») молекулу. crRNA включает в себя как нацеливающийся на ДНК сегмент (однонитевый) gRNA, так и участок нуклеотидов (т.е. хвост crRNA), который образует одну половину дуплекса dsRNA связывающегося с белком сегмента gRNA. Пример хвоста crRNA, расположенного в 5'-3' направлении (3') нацеливающегося на ДНК сегмента, включает в себя, состоит, по сути, из или состоит из GUUUUAGAGCUAUGCU (SEQ ID NO: 87). Любой из нацеливающихся на ДНК сегментов, раскрываемых в настоящем документе, может быть соединен с 5'-концом SEQ ID NO: 87 с образованием crRNA.
[00201] Соответствующая tracrRNA (РНК-активатор) включает в себя участок нуклеотидов, который образует другую половину дуплекса dsRNA связывающегося с белком сегмента gRNA. Участок нуклеотидов crRNA является комплементарным и гибридизируется с участком нуклеотидов tracrRNA с образованием дуплекса dsRNA связывающегося с белком домена gRNA. Таким образом, можно сказать, что каждая crRNA имеет соответствующую tracrRNA. Пример последовательности tracrRNA включает в себя, состоит, по сути, из, или состоит из AGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGA GUCGGUGCUUU (SEQ ID NO: 88).
[00202] В системах, в которых необходимы и crRNA, и tracrRNA, crRNA и соответствующая tracrRNA гибридизируются с образованием gRNA. В системах, в которых необходима только crRNA, crRNA может представлять собой gRNA. CrRNA, кроме того, обеспечивает однонитевый нацеливающийся на ДНК сегмент, который нацеливается на целевую последовательность направляющей РНК путем гибридизации с противоположной нитью (т.е. комплементарной нитью). Если используется для модификации внутри клетки, точная последовательность данной молекулы crRNA или tracrRNA может быть разработана так, чтобы она была специфичной для видов, в которых будут использоваться молекулы РНК. См., например, Mali et al. (2013) Science 339:823-826; Jinek et al. (2012) Science 337:816-821; Hwang et al. (2013)Nat. Biotechnol. 31:227-229; Jiang et al. (2013) Nat. Biotechnol. 31:233-239; и Cong et al. (2013) Science 339:819-823, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00203] Нацеливающийся на ДНК сегмент (crRNA) данной gRNA включает в себя нуклеотидную последовательность, которая комплементарна последовательности (т.е. комплементарной нити последовательности распознавания направляющей РНК в нити, противоположной целевой последовательности направляющей РНК) в целевой ДНК. Нацеливающийся на ДНК сегмент gRNA взаимодействует с целевой ДНК специфическим в отношении последовательности образом путем гибридизации (т.е. спаривания оснований). Поэтому, нуклеотидная последовательность нацеливающегося на ДНК сегмента может варьировать и определяет локализацию в целевой ДНК, с которой gRNA и целевая ДНК будут взаимодействовать. Нацеливающийся на ДНК сегмент рассматриваемой gRNA может быть модифицирован для гибридизации с любой желаемой последовательностью в целевой ДНК. Встречающиеся в природе crRNA отличаются в зависимости от системы CRISPR/Cas и организма, но зачастую содержат нацеливающийся сегмент длиной от 21 и 72 нуклеотидов, фланкированный двумя прямыми повторами (DR) длиной от 21 до 46 нуклеотидов (см., например, WO 2014/131833, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей). В случае S. pyogenes DR в длину составляют 36 нуклеотидов, а нацеливающийся сегмент в длину составляет 30 нуклеотидов. 3'-расположенный DR является комплементарным и гибридизируется с соответствующей tracrRNA, которая, в свою очередь, связывается с белком Cas.
[00204] Нацеливающийся на ДНК сегмент может иметь длину по меньшей мере приблизительно 12 нуклеотидов, по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 17 нуклеотидов, по меньшей мере приблизительно 18 нуклеотидов, по меньшей мере приблизительно 19 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 25 нуклеотидов, по меньшей мере приблизительно 30 нуклеотидов, по меньшей мере приблизительно 35 нуклеотидов или по меньшей мере приблизительно 40 нуклеотидов. Такие нацеливающиеся на ДНК сегменты могут иметь длину от приблизительно 12 нуклеотидов до приблизительно 100 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 80 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 50 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 40 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 30 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 25 нуклеотидов или от приблизительно 12 нуклеотидов до приблизительно 20 нуклеотидов. Например, нацеливающийся на ДНК сегмент может составлять от приблизительно 15 нуклеотидов до приблизительно 25 нуклеотидов (например, от приблизительно 17 нуклеотидов до приблизительно 20 нуклеотидов или приблизительно 17 нуклеотидов, приблизительно 18 нуклеотидов, приблизительно 19 нуклеотидов или приблизительно 20 нуклеотидов). См., например, US 2016/0024523, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Для Cas9 из S. pyogenes типичный нацеливающийся на ДНК сегмент составляет от 16 до 20 нуклеотидов в длину или от 17 до 20 нуклеотидов в длину. Для Cas9 из S. aureus типичный нацеливающийся на ДНК сегмент составляет от 21 до 23 нуклеотидов в длину. Для Cpf1 типичный нацеливающийся на ДНК сегмент составляет по меньшей мере 16 нуклеотидов в длину или по меньшей мере 18 нуклеотидов в длину.
[00205] TracrRNA могут иметь любую форму (например, полноразмерные tracrRNA или активные частичные tracrRNA) и варьирующие длины. Они могут включать в себя первичные транскрипты или процессированные формы. Например, tracrRNA (как часть одиночной направляющей РНК, или как отдельная молекула, или как часть двухмолекулярной gRNA) могут включать в себя, состоять, по сути, из или состоять из всей или части последовательности tracrRNA дикого типа (например, приблизительно или более чем приблизительно 20, 26, 32, 45, 48, 54, 63, 67, 85 или больше нуклеотидов последовательности tracrRNA дикого типа). Примеры последовательностей tracrRNA дикого типа из S. pyogenes включают в себя версии из 171 нуклеотида, 89 нуклеотидов, 75 нуклеотидов и 65 нуклеотидов. См., например, Deltcheva et al. (2011) Nature 471:602-607; WO 2014/093661, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Примеры tracrRNA в одиночных направляющих РНК (sgRNA) включают в себя сегменты tracrRNA, обнаруженные в версиях +48, +54, +67 и +85 sgRNA, при этом «+n» указывает на то, что до +n нуклеотидов tracrRNA дикого типа включено в sgRNA. См. патент США №8697359, включенный в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00206] Процент комплементарности между нацеливающимся на ДНК сегментом и комплементарной нитью последовательности распознавания направляющей РНК в целевой ДНК может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%). Процент комплементарности между нацеливающимся на ДНК сегментом и комплементарной нитью последовательности распознавания направляющей РНК в целевой ДНК может составлять по меньшей мере 60% в пределах приблизительно 20 смежных нуклеотидов. В качестве примера, процент комплементарности между нацеливающимся на ДНК сегментом и комплементарной нитью последовательности распознавания направляющей РНК в целевой ДНК составляет 100% в пределах 14 смежных нуклеотидов на 5'-конце комплементарной нити последовательности распознавания направляющей РНК в комплементарной нити целевой ДНК и всего 0% в пределах остальной части. В таком случае нацеливающийся на ДНК сегмент можно считать имеющим в длину 14 нуклеотидов. В качестве другого примера, процент комплементарности между нацеливающимся на ДНК сегментом и комплементарной нитью последовательности распознавания направляющей РНК в целевой ДНК составляет 100% в пределах семи смежных нуклеотидов на 5'-конце комплементарной нити последовательности распознавания направляющей РНК в комплементарной нити целевой ДНК и всего 0% в пределах остальной части. В таком случае нацеливающийся на ДНК сегмент можно считать имеющим в длину 7 нуклеотидов. В некоторых направляющих РНК по меньшей мере 17 нуклеотидов в нацеливающемся на ДНК сегменте комплементарны целевой ДНК. Например, нацеливающийся на ДНК сегмент может составлять 20 нуклеотидов в длину и может включать в себя 1, 2 или 3 ошибки спаривания с комплементарной нитью последовательности распознавания направляющей РНК. Предпочтительно ошибки спаривания не являются смежными с последовательностью прилегающего к протоспейсеру мотива (РАМ) (например, ошибки спаривания находятся на 5'-конце нацеливающегося на ДНК сегмента, или ошибки спаривания находятся на расстоянии по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19 пар оснований от последовательности РАМ).
[00207] Связывающийся с белком сегмент gRNA может включать в себя два участка нуклеотидов, которые комплементарны друг другу. Комплементарные нуклеотиды связывающегося с белком сегмента гибридизируются с образованием дуплекса двухнитевой РНК (dsRNA). Связывающийся с белком сегмент рассматриваемой gRNA взаимодействует с белком Cas, и gRNA направляет связанный белок Cas на определенную нуклеотидную последовательность в целевой ДНК посредством нацеливающегося на ДНК сегмента.
[00208] Одиночные направляющие РНК имеют нацеливающийся на ДНК сегмент и каркасную последовательность (т.е. связывающую белок или связывающую Cas последовательность направляющей РНК). Например, такие направляющие РНК имеют 5' нацеливающийся на ДНК сегмент и 3' каркасную последовательность. Иллюстративные каркасные последовательности включают в себя, состоят, по сути, из или состоят из GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAA AAGUGGCACCGAGUCGGUGCU (версия 1; SEQ ID NO: 89); GUUGGAACCAUUCAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAAC UUGAAAAAGUGGCACCGAGUCGGUGC (версия 2; SEQ ID NO: 8); GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAA AAGUGGCACCGAGUCGGUGC (версия 3; SEQ ID NO: 9) и GUUUAAGAGCUAUGCUGGAAACAGCAUAGCAAGUUUAAAUAAGGCUAGUCCGUUAU CAACUUGAAAAAGUGGCACCGAGUCGGUGC (версия 4; SEQ ID NO: 10). Направляющие РНК, нацеливающиеся на любую целевую последовательность направляющей РНК, могут включать в себя, например, нацеливающийся на ДНК сегмент на 5'-конце направляющей РНК, слитый с любой из иллюстративных каркасных последовательностей направляющей РНК на 3'-конце направляющей РНК. То есть любой из нацеливающихся на ДНК сегментов, раскрываемых в настоящем документе, может быть соединен с 5'-концом любого из SEQ ID NO: 89, 8, 9 или 10 с образованием одиночной направляющей РНК (химерная направляющая РНК). Версии 1, 2, 3 и 4 направляющей РНК, раскрываемые в других частях настоящего документа, относятся к нацеливающимся на ДНК сегментам (т.е. к направляющим последовательностям или направляющим), соединенным с версиями 1, 2, 3 и 4 каркаса, соответственно.
[00209] Направляющие РНК могут включать в себя модификации или последовательности, обеспечивающие дополнительные желаемые свойства (например, модифицированную или регулируемую стабильность; субклеточное нацеливание; отслеживание по флуоресцентной метке; сайт связывания для белка или белкового комплекса и т.п.). К примерам таких модификаций относятся, например, 5'-кэп (например, 7-метилгуанилатный кэп (m7G)); 3'-полиаденилированный хвост (т.е. 3'-поли(А) хвост); последовательность-рибопереключатель (например, для обеспечения регулируемой стабильности и/или регулируемой доступности белками и/или белковыми комплексами); последовательность контроля стабильности; последовательность, образующая дуплекс dsRNA (т.е. шпилька)); модификация или последовательность, нацеливающая РНК в субклеточную локализацию (например, ядро, митохондрии, хлоропласта и т.п.); модификация или последовательность, обеспечивающая отслеживание (например, прямая конъюгация с флуоресцентной молекулой, конъюгация с фрагментом, который способствует выявлению флуоресценции, последовательность, обеспечивающая выявление флуоресценции и т.д.); модификация или последовательность, обеспечивающая сайт связывания для белков (например, белков, действующих на ДНК, в том числе ДНК-метилтрансфераз, ДНК-деметилаз, гистонацетилтрансфераз, гистондеацетилаз и т.п.); и их комбинации. Другие примеры модификаций включают в себя созданные методами генной инженерии дуплексные структуры «стебель-петля», созданные методами генной инженерии области выпетливания, созданные методами генной инженерии шпильки 3' дуплексной структуры «стебель-петля» или любую их комбинацию. См., например, US 2015/0376586, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Выпетливание может представлять собой неспаренную область нуклеотидов в дуплексе, состоящую из crRNA-подобной области и минимальной tracrRNA-подобной области. Выпетливание может включать в себя на одной стороне дуплекса неспаренную 5'-XXXY-3', в которой X представляет собой любой пурин, a Y может представлять собой нуклеотид, который может образовывать неоднозначную пару с нуклеотидом на противоположной нити, и область неспаренных нуклеотидов на другой стороне дуплекса.
[00210] Немодифицированные нуклеиновые кислоты могут быть склонны к разложению. Экзогенные нуклеиновые кислоты также могут индуцировать врожденный иммунный ответ. Модификации могут помочь ввести стабильность и уменьшить иммуногенность. Направляющие РНК могут включать в себя модифицированные нуклеозиды и модифицированные нуклеотиды, в том числе, например, один или несколько из следующих: (1) изменение или замена одного или обоих не связывающих атомов кислорода фосфата и/или одного или нескольких связывающих атомов кислорода фосфата в фосфодиэфирной связи остова; (2) изменение или замена компонента рибозного сахара, например, изменение или замена 2'-гидроксила в рибозном сахаре; (3) замена фосфатного фрагмента на линкеры дефосфо; (4) модификация или замена встречающегося в природе нуклеинового основания; (5) замена или модификация рибозофосфатного остова; (6) модификация 3'-конца или 5'-конца олигонуклеотида (например, удаление, модификация или замена концевой фосфатной группы или конъюгация фрагмента) и (7) модификация сахара. Другие возможные модификации направляющей РНК включают в себя модификации или замену урацилов или полиуретановых путей. См., например, WO 2015/048577 и US 2016/0237455, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Подобные модификации могут быть выполнены в кодирующих Cas нуклеиновых кислотах, таких как иРНК Cas.
[00211] В качестве одного примера, нуклеотиды на 5'- или 3'-конце направляющей РНК могут включать в себя фосфоротиоатные связи (например, основания могут иметь модифицированную фосфатную группу, которая представляет собой фосфоротиоатную группу). Например, направляющая РНК может включать в себя фосфоротиоатные связи между 2, 3 или 4 концевыми нуклеотидами на 5'- или 3'-конце направляющей РНК. В качестве другого примера, нуклеотиды на 5'- и/или 3'-конце направляющей РНК могут иметь 2'-O-метил модификации. Например, направляющая РНК может включать в себя 2'-O-метил-модификации в 2, 3 или 4 концевых нуклеотидах на 5'- и/или 3'-конце направляющей РНК (например, на 5'-конце). См., например, WO 2017/173054 А1 и Finn et al. (2018) Cell Reports 22: 1-9, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Согласно одному конкретному примеру направляющая РНК включает в себя 2'-O-метил-аналоги и 3' фосфоротиоатные межнуклеотидные связи в первых трех 5'- и 3'-концевых остатках РНК. Согласно другому конкретному примеру направляющая РНК модифицирована так, что все группы 2'ОН, которые не взаимодействуют с белком Cas9, заменяются 2'-O-метил-аналогами, и хвостовая область направляющей РНК, которая характеризуется минимальным взаимодействием с Cas9, модифицирована 5' и 3' фосфоротиоатными межнуклеотидными связями. См., например, Yin et al. (2017) Nat. Biotech. 35(12): 1179-1187, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Другие примеры модифицированных направляющих РНК представлены, например, в WO 2018/107028 А1, включенной в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00212] Направляющие РНК могут быть обеспечены в любой форме. Например, gRNA могут быть обеспечены в форме РНК, либо в виде двух молекул (отдельные crRNA и tracrRNA), либо в виде одной молекулы (sgRNA) и необязательно в форме комплекса с белком Cas. gRNA также могут быть обеспечены в форме ДНК, кодирующей gRNA. ДНК, кодирующая gRNA, может кодировать одну молекулу РНК (sgRNA) или отдельные молекулы РНК (например, отдельные crRNA и tracrRNA). В последнем случае ДНК, кодирующая gRNA, может быть обеспечена в виде одной молекулы ДНК или в виде отдельных молекул ДНК, кодирующих crRNA и tracrRNA, соответственно.
[00213] При обеспечении gRNA в форме ДНК gRNA может быть временно, условно или конститутивно экспрессирована в клетке. ДНК, кодирующие gRNA, могут быть стабильно интегрированы в геном клетки и функционально связаны с промотором, активным в клетке. В качестве альтернативы, ДНК, кодирующие gRNA, могут быть функционально связаны с промотором в экспрессионной конструкции. Например, ДНК, кодирующая gRNA, может находиться в векторе, включающем в себя гетерологичную нуклеиновую кислоту, такую как нуклеиновая кислота, кодирующая белок Cas. В качестве альтернативы, она может находиться в векторе или плазмиде, которая отделена от вектора, включающего в себя нуклеиновую кислоту, кодирующую белок Cas. Промоторы, которые могут быть использованы в таких экспрессионных конструкциях, включают в себя промоторы, активные, например, в одной или нескольких из эукариотической клетки, человеческой клетки, не являющаяся человеческой клетки, клетки млекопитающего, клетки отличного от человека млекопитающего, клетки грызуна, клетки мыши, клетки крысы, клетки хомяка, клетки кролика, плюрипотентной клетки, эмбриональной стволовой (ES) клетки, стволовой клетки взрослого человека, онтогенетически ограниченной клетки-предшественника, индуцированной плюрипотентной стволовой (iPS) клетки или эмбриона на стадии одной клетки. Такими промоторами могут быть, например, условные промоторы, индуцибельные промоторы, конститутивные промоторы или специфические по отношению к ткани промоторы. Такими промоторами также могут быть, например, двунаправленные промоторы. Конкретные примеры подходящих промоторов включают в себя промотор РНК-полимеразы III, такой как промотор U6 человека, промотор U6-полимеразы III крысы или промотор U6-полимеразы III мыши.
[00214] В качестве альтернативы, gRNA могут быть получены различными другими способами. Например, gRNA могут быть получены путем in vitro транскрипции с использованием, например, Т7 РНК-полимеразы (см., например, WO 2014/089290 и WO 2014/065596, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей). Направляющие РНК также могут представлять собой полученную синтетическим путем молекулу, полученную с помощью химического синтеза.
[00215] Последовательности распознавания направляющей РНК и целевые последовательности направляющей РНК. Термин «последовательность распознавания направляющей РНК» включает в себя последовательности нуклеиновой кислоты, присутствующие в целевой ДНК, с которой будет связываться нацеливающийся на ДНК сегмент gRNA, при условии наличия достаточных условий для связывания. Используемый в настоящем документе термин «последовательность распознавания направляющей РНК» охватывает обе нити целевой двухнитевой ДНК (т.е. последовательность в комплементарной нити, с которой направляющая РНК гибридизируется, и соответствующую последовательность в некомплементарной нити, смежную с прилегающим к протоспейсеру мотивом (РАМ)). Используемый в настоящем документе термин «целевая последовательность направляющей РНК» относится, в частности, к последовательности в некомплементарной нити, смежной с РАМ (т.е. в 3'-5' направлении или 5' по отношению к РАМ). То есть целевая последовательность направляющей РНК относится к последовательности в некомплементарной нити, соответствующей последовательности, с которой направляющая РНК гибридизируется в комплементарной нити. Целевая последовательность направляющей РНК эквивалентна нацеливающемуся на ДНК сегменту направляющей РНК, но с тиминами вместо урацилов. В качестве одного примера, целевая последовательность направляющей РНК для фермента Cas9 будет относиться к последовательности в некомплементарной нити, смежной с 5'-NGG-3' РАМ. Последовательности распознавания направляющей РНК включают в себя последовательности, по отношению к которым направляющая РНК разрабатывается как комплементарная, при этом гибридизация между комплементарной нитью последовательности распознавания направляющей РНК и нацеливающимся на ДНК сегментом направляющей РНК способствует образованию комплекса CRISPR. Полная комплементарность не является обязательной при условии, что существует комплементарность, достаточная для обеспечения гибридизации и способствования образованию комплекса CRISPR. Последовательности распознавания направляющей РНК или целевые последовательности направляющей РНК также включают в себя сайты расщепления для белков Cas, описанные более подробно ниже. Последовательность распознавания направляющей РНК или целевая последовательность направляющей РНК может включать в себя любой полинуклеотид, который может быть расположен, например, в ядре или цитоплазме клетки или в органелле клетки, такой как митохондрия или хлоропласт.
[00216] На последовательность распознавания направляющей РНК в целевой ДНК могут быть нацелены (т.е. связаны, или гибридизированы, или комплементарны) белок Cas или gRNA. Подходящие условия связывания ДНК/РНК включают в себя физиологические условия, обычно присутствующие в клетке. Известны другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) (см., например, Molecular Cloning: А Laboratory Manual, 3rd Ed. (Sambrook et al., Harbor Laboratory Press 2001), включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей). Нить целевой ДНК, которая комплементарна и гибридизируется с белком Cas или gRNA, можно назвать «комплементарной нитью», а нить целевой ДНК, которая комплементарна «комплементарной нити» (и, следовательно, не комплементарна белку Cas или gRNA), можно назвать «некомплементарной цепью» или «матричной нитью».
[00217] Белок Cas может расщеплять нуклеиновую кислоту по сайту внутри или снаружи последовательности нуклеиновой кислоты, присутствующей в целевой ДНК, с которой будет связываться нацеливающийся на ДНК сегмент gRNA. Термин «сайт расщепления» включает в себя положение нуклеиновой кислоты, по которому белок Cas осуществляет однонитевый разрыв или двухнитевый разрыв. Например, образование комплекса CRISPR (включающего в себя gRNA, гибридизированную с комплементарной нитью последовательности распознавания направляющей РНК и образовавшую комплекс с белком Cas) может привести к расщеплению одной или обеих нитей в пределах или около (например, в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или больше пар оснований из) последовательности нуклеиновой кислоты, присутствующей в целевой ДНК, с которой будет связываться нацеливающийся на ДНК сегмент gRNA. Если сайт расщепления находится за пределами последовательности нуклеиновой кислоты, с которой будет связываться нацеливающийся на ДНК сегмент gRNA, то сайт расщепления по-прежнему считается находящимся в «последовательности распознавания направляющей РНК» или целевой последовательности направляющей РНК. Сайт расщепления может быть только в одной нити или в обеих нитях нуклеиновой кислоты. Сайты расщепления могут находиться в одном и том же положении в обеих нитях нуклеиновой кислоты (образуя тупые концы) или могут находиться в разных сайтах в каждой нити (продуцируя ступенчатые концы (т.е. выступающие концы)). Ступенчатые концы могут быть получены, например, с использованием двух белков Cas, каждый из которых осуществляет однонитевый разрыв в другом сайте расщепления на другой нити с осуществлением тем самым двухнитевого разрыва. Например, первая никаза может создавать однонитевый разрыв в первой нити двухнитевой ДНК (dsDNA), а вторая никаза может создавать однонитевый разрыв во второй нити dsDNA, так что образуются выступающие последовательности. В некоторых случаях последовательность распознавания направляющей РНК или целевая последовательность направляющей РНК никазы в первой нити отделяется от последовательности распознавания направляющей РНК или целевой последовательности направляющей РНК никазы во второй нити по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 парами оснований.
[00218] Сайт-специфическое связывание и/или расщепление целевой ДНК белками Cas может происходить в местах, определяемых как (i) комплементарностью спаривания оснований между gRNA и целевой ДНК, так и (ii) коротким мотивом, называемым прилегающим к протоспейсеру мотивом (РАМ), в целевой ДНК. РАМ может фланкировать целевую последовательность направляющей РНК в некомплементарной нити, противоположной нити, с которой направляющая РНК гибридизируется. Необязательно целевая последовательность направляющей РНК может быть фланкирована на 3'-конце с помощью РАМ. В качестве альтернативы, целевая последовательность направляющей РНК может быть фланкирована на 5' -конце с помощью РАМ. Например, сайт расщепления белков Cas может составлять от приблизительно 1 до приблизительно 10 или от приблизительно 2 до приблизительно 5 пар оснований (например, 3 пар оснований) в 3'-5' направлении или в 5'-3' направлении последовательности РАМ. В некоторых случаях (например, при использовании Cas9 из S. pyogenes или близкородственного Cas9) последовательность РАМ некомплементарной нити может представлять собой 5'-N1GG-3', при этом Ni представляет собой любой нуклеотид ДНК и находится непосредственно в 3' относительно последовательности распознавания направляющей РНК некомплементарной нити целевой ДНК (т.е. непосредственно 3' относительно целевой последовательности направляющей РНК). Таким образом, последовательность РАМ комплементарной нити будет представлять собой 5'-CCN2-3', при этом N2 представляет собой любой нуклеотид ДНК и находится непосредственно в 5' относительно последовательности распознавания направляющей РНК некомплементарной нити целевой ДНК. В некоторых таких случаях N1 и N2 могут быть комплементарными, а пара оснований N1-N2 может быть любой парой оснований (например, N1=C и N2=G; N1=G и N2=C; N1=A и N2=T или N1=T и N2=A). В случае Cas9 из S. aureus РАМ может представлять собой NNGRRT или NNGRR, при этом N может представлять собой A, G, С или Т, a R может представлять собой G или А. В случае Cas9 из С. jejuni РАМ может представлять собой, например, NNNNACAC или NNENNRYAC, при этом N может представлять собой A, G, С или Т, a R может представлять собой G или А. В некоторых случаях (например, для FnCpf1) последовательность РАМ может располагаться в 3'-5' направлении 5'-конца и иметь последовательность 5'-TTN-3'.
[00219] Ниже представлены примеры целевых последовательностей направляющей РНК или целевых последовательностей направляющей РНК в дополнение к последовательности РАМ. Например, целевая последовательность направляющей РНК может быть 20-нуклеотидной последовательностью ДНК, непосредственно предшествующей мотиву NGG, распознаваемому белком Cas9. Примерами таких целевых последовательностей направляющей РНК плюс последовательность РАМ являются GN19NGG (SEQ ID NO: 11) или N20NGG (SEQ ID NO: 12). См., например, WO 2014/165825, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Гуанин на 5'-конце может облегчать транскрипцию РНК-полимеразой в клетках. Другие примеры целевых последовательностей направляющей РНК плюс последовательность РАМ могут включать в себя два гуаниновых нуклеотида на 5'-конце (например, GGN20NGG; SEQ ID NO: 13) для облегчения эффективной транскрипции с помощью Т7-полимеразы in vitro. См., например, WO 2014/065596, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Другие целевые последовательности направляющей РНК плюс последовательность РАМ могут иметь в длину от 4 до 22 нуклеотидов из SEQ ID NO: 11-13, в том числе 5' G или GG и 3' GG или NGG. При этом другие целевые последовательности направляющей РНК могут иметь в длину от 14 до 20 нуклеотидов из SEQ ID NO: 11-13.
[00220] Последовательность распознавания направляющей РНК или целевая последовательность направляющей РНК может быть любой последовательностью нуклеиновой кислоты, эндогенной или экзогенной по отношению к клетке. Последовательность распознавания направляющей РНК или целевая последовательность направляющей РНК может быть последовательностью, кодирующей генный продукт (например, белок), или некодирующей последовательностью (например, регуляторной последовательностью) или может включать в себя и ту, и другую.
(3) Экзогенные донорные нуклеиновые кислоты, нацеливающиеся на человеческий TTR ген
[00221] В способах и композициях, раскрываемых в настоящем документе, можно использовать экзогенные донорные нуклеиновые кислоты для модификации гуманизированного TTR локуса после расщепления гуманизированного TTR локуса нуклеазным средством. В таких способах белок нуклеазного средства расщепляет гуманизированный TTR локус с созданием однонитевого разрыва (ника) или двухнитевого разрыва, и экзогенная донорная нуклеиновая кислота рекомбинируется с гуманизированным TTR локусом путем опосредованного негомологичным соединением концов (NHEJ) лигирования или посредством события направляемой гомологией репарации. Необязательно репарация с использованием экзогенной донорной нуклеиновой кислоты удаляет или нарушает целевую последовательность нуклеазы, так что на аллели, на которые происходило нацеливание, нуклеазное средство не может быть повторно нацелено.
[00222] Экзогенные донорные нуклеиновые кислоты могут включать в себя дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), они могут быть однонитевыми или двухнитевыми, и они могут иметь линейную или кольцевую форму. Например, экзогенная донорная нуклеиновая кислота может представлять собой однонитевый олигодезоксинуклеотид (ssODN). См., например, Yoshimi et al. (2016) Nat. Commun. 7:10431, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Иллюстративная экзогенная донорная нуклеиновая кислота составляет от приблизительно 50 нуклеотидов до приблизительно 5 т.п.н. в длину, составляет от приблизительно 50 нуклеотидов до приблизительно 3 т.п.н. в длину или составляет от приблизительно 50 до приблизительно 1000 нуклеотидов в длину. Другие иллюстративные экзогенные донорные нуклеиновые кислоты составляют от приблизительно 40 до приблизительно 200 нуклеотидов в длину. Например, экзогенная донорная нуклеиновая кислота может составлять приблизительно 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140, 140-150, 150-160, 160-170, 170-180, 180-190 или 190-200 нуклеотидов в длину. В качестве альтернативы, экзогенная донорная нуклеиновая кислота может составлять приблизительно 50-100, 100-200, 200-300, 300-400, 400-500, 500-600, 600-700, 700-800, 800-900 или 900-1000 нуклеотидов в длину. В качестве альтернативы, экзогенная донорная нуклеиновая кислота может составлять приблизительно 1-1,5, 1,5-2, 2-2,5, 2,5-3, 3-3,5, 3,5-4, 4-4,5 или 4,5-5 т.п.н. в длину. В качестве альтернативы, экзогенная донорная нуклеиновая кислота может составлять, например, не более чем 5 т.п.н., 4,5 т.п.н., 4 т.п.н., 3,5 т.п.н., 3 т.п.н., 2,5 т.п.н., 2 т.п.н., 1,5 т.п.н., 1 т.п.н., 900 нуклеотидов, 800 нуклеотидов, 700 нуклеотидов, 600 нуклеотидов, 500 нуклеотидов, 400 нуклеотидов, 300 нуклеотидов, 200 нуклеотидов, 100 нуклеотидов или 50 нуклеотидов в длину. Экзогенные донорные нуклеиновые кислоты (например, нацеливающиеся векторы) также могут быть длиннее.
[00223] Согласно одному примеру экзогенная донорная нуклеиновая кислота представляет собой ssODN, которая составляет от приблизительно 80 нуклеотидов до приблизительно 200 нуклеотидов в длину. Согласно другому примеру экзогенная донорная нуклеиновая кислота представляет собой ssODN, которая составляет от приблизительно 80 нуклеотидов до приблизительно 3 т.п.н. в длину. Такая ssODN может иметь плечи гомологии, например, каждое из которых составляет от приблизительно 40 нуклеотидов до приблизительно 60 нуклеотидов в длину. Такая ssODN также может иметь плечи гомологии, например, каждое из которых составляет от приблизительно 30 нуклеотидов до 100 нуклеотидов в длину. Плечи гомологии могут быть симметричными (например, длина каждого 40 нуклеотидов или длина каждого 60 нуклеотидов), или они могут быть асимметричными (например, длина одного плеча гомологии составляет 36 нуклеотидов, а длина другого плеча гомологии составляет 91 нуклеотид).
[00224] Экзогенные донорные нуклеиновые кислоты могут включать в себя модификации или последовательности, которые обеспечены для дополнительных желаемых признаков (например, модифицированной или регулируемой стабильности; отслеживания или выявления с помощью флуоресцентной метки; сайта связывания для белка или белкового комплекса и т.д.). Экзогенные донорные нуклеиновые кислоты могут включать в себя одну или несколько флуоресцентных меток, меток очистки, меток эпитопа или их комбинацию. Например, экзогенная донорная нуклеиновая кислота может включать в себя одну или несколько флуоресцентных меток (например, флуоресцентные белки или другие флуорофоры или красители), например, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 флуоресцентных меток. Иллюстративные флуоресцентные метки включают в себя флуорофоры, такие как флуоресцеин (например, 6-карбоксифлуоресцеин (6-FAM)), Texas Red, HEX, Су3, Су5, Су5,5, Pacific Blue, 5-(и -6)-карбокситетраметилродамин (TAMRA) и Су7. Широкий ряд флуоресцентных красителей коммерчески доступен для мечения олигонуклеотидов (например, от компании Integrated DNA Technologies). Такие флуоресцентные метки (например, внутренние флуоресцентные метки) могут быть использованы, например, для выявления экзогенной донорной нуклеиновой кислоты, которая была непосредственно встроена в расщепленную целевую нуклеиновую кислоту, имеющую выступающие концы, совместимые с концами экзогенной донорной нуклеиновой кислоты. Метка или маркер может находиться на 5'-конце, 3'-конце или внутри экзогенной донорной нуклеиновой кислоты. Например, экзогенная донорная нуклеиновая кислота может быть конъюгирована на 5'-конце с IR700 флуорофором от компании Integrated DNA Technologies (5'IRDYE®700).
[00225] Экзогенные донорные нуклеиновые кислоты также могут включать в себя вставки нуклеиновой кислоты, в том числе сегменты ДНК, подлежащие интеграции в гуманизированный TTR локус. Интеграция вставки нуклеиновой кислоты в гуманизированный TTR локус может приводить в результате к добавлению представляющей интерес последовательности нуклеиновой кислоты в гуманизированный TTR локус, делеции представляющей интерес последовательности нуклеиновой кислоты в гуманизированный TTR локус или замене представляющей интерес последовательности нуклеиновой кислоты в гуманизированном TTR локусе (т.е. делеция и вставка). Некоторые экзогенные донорные нуклеиновые кислоты разработаны для помещения вставки нуклеиновой кислоты в гуманизированный TTR локус без какой-либо соответствующей делеции в гуманизированном TTR локусе. Другие экзогенные донорные нуклеиновые кислоты разработаны для делеции представляющей интерес последовательности нуклеиновой кислоты в гуманизированном TTR локусе без какого-либо соответствующего помещения вставки нуклеиновой кислоты. Следующие экзогенные донорные нуклеиновые кислоты разработаны для делеции представляющей интерес последовательности нуклеиновой кислоты в гуманизированном TTR локусе и замены ее вставкой нуклеиновой кислоты.
[00226] Вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в гуманизированном TTR локусе, подлежащая делеции и/или замене, может иметь различную длину. Иллюстративная вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в гуманизированном TTR локусе, подлежащая делеции и/или замене, составляет от приблизительно 1 нуклеотида до приблизительно 5 т.п.н. в длину или составляет от приблизительно 1 нуклеотида до приблизительно 1000 нуклеотидов в длину. Например, вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в гуманизированном TTR локусе, подлежащая делеции и/или замене, может составлять приблизительно 1-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140, 140-150, 150-160, 160-170, 170-180, 180-190 или 190-120 нуклеотидов в длину. Подобным образом, вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в гуманизированном TTR локусе, подлежащая делеции и/или замене, может составлять 1-100, 100-200, 200-300, 300-400, 400-500, 500-600, 600-700, 700-800, 800-900 или 900-1000 нуклеотидов в длину. Подобным образом, вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в гуманизированном TTR локусе, подлежащая делеции и/или замене, может в длину составлять приблизительно 1-1,5, 1,5-2, 2-2,5, 2,5-3, 3-3,5, 3,5-4, 4-4,5 или 4,5-5 т.п.н. или больше.
[00227] Вставка нуклеиновой кислоты может включать в себя последовательность, которая является гомологичной или ортологической по отношению ко всей или части целевой последовательности для замены. Например, вставка нуклеиновой кислоты может включать в себя последовательность, которая включает в себя одну или несколько точковых мутаций (например, 1, 2, 3, 4, 5 или больше) по сравнению с целевой последовательностью для замены в гуманизированном TTR локусе. Необязательно такие точковые мутации могут приводить в результате к консервативной аминокислотной замене (например, замене аспарагиновой кислоты [Asp, D] глутаминовой кислотой [Glu, Е]) в кодируемом полипептиде.
[00228] Донорные нуклеиновые кислоты для опосредованной негомологичным соединением концов вставки. Некоторые экзогенные донорные нуклеиновые кислоты имеют короткие однонитевые области на 5'-конце и/или 3'-конце, которые комплементарны одному или нескольким выступающим концам, созданным путем опосредованного нуклеазой расщепления по гуманизированному TTR локусу. Такие выступающие конца также могут быть названы 5' и 3' плечами гомологии. Например, некоторые экзогенные донорные нуклеиновые кислоты имеют короткие однонитевые области на 5'-конце и/или 3'-конце, которые комплементарны одному или нескольким выступающим концам, созданным путем опосредованного нуклеазой расщепления в 5' и/или 3' целевых последовательностях по гуманизированному TTR локусу. Некоторые из таких экзогенных донорных нуклеиновых кислот имеют комплементарную область только на 5'-конце или только на 3'-конце. Например, некоторые из таких экзогенных донорных нуклеиновых кислот имеют комплементарную область только на 5'-конце, комплементарном выступающему концу, созданному в 5' целевой последовательности по гуманизированному TTR локусу или только на 3'-конце, комплементарном выступающему концу, созданному в 3' целевой последовательности по гуманизированному TTR локусу. Другие из таких экзогенных донорных нуклеиновых кислот имеют комплементарные области как на 5'-, так и на 3'-конце. Например, другие из таких экзогенных донорных нуклеиновых кислот имеют комплементарные области как на 5'-, так и на 3'-конце, например, комплементарных первому и второму выступающим концам, соответственно, созданных путем опосредованного нуклеазой расщепления по гуманизированному TTR локусу. Например, если экзогенная донорная нуклеиновая кислота является двухнитевой, то однонитевые комплементарные области, могут начинаться от 5'-конца верхней нити донорной нуклеиновой кислоты и от 5'-конца нижней нити донорной нуклеиновой кислоты с образованием 5' выступающих концов на каждом конце. В качестве альтернативы, однонитевая комплементарная область может начинаться от 3'-конца верхней нити донорной нуклеиновой кислоты и от 3'-конца нижней нити матрицы с образованием 3' выступающих концов.
[00229] Комплементарные области могут иметь любую длину, достаточную для обеспечения лигирования между экзогенной донорной нуклеиновой кислотой и целевой нуклеиновой кислотой. Иллюстративные комплементарные области составляют от приблизительно 1 до приблизительно 5 нуклеотидов в длину, от приблизительно 1 до приблизительно 25 нуклеотидов в длину или от приблизительно 5 до приблизительно 150 нуклеотидов в длину. Например, комплементарная область может составлять по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину. В качестве альтернативы, комплементарная область в длину может составлять приблизительно 5-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140 или 140-150 нуклеотидов или больше.
[00230] Такие комплементарные области могут быть комплементарны выступающим концам, образованным двумя парами никаз. Два двухнитевых разрыва со ступенчатыми концами могут быть созданы с использованием первой и второй никаз, которые расщепляют противоположные нити ДНК с образованием первого двухнитевого разрыва, а также третьей и четвертой никаз, которые расщепляют противоположные нити ДНК с образованием второго двухнитевого разрыва. Например, белок Cas может быть использован для осуществления однонитевого разрыва первой, второй, третьей и четвертой целевых последовательностей направляющей РНК, соответствующих первой, второй, третьей и четвертой направляющим РНК. Первую и вторую целевые последовательности направляющей РНК можно расположить так, чтобы создать первый сайт расщепления таким образом, что однонитевые разрывы, образованные первой и второй никазами в первой и второй нитях ДНК, образуют двухнитевый разрыв (т.е. первый сайт расщепления включает в себя однонитевые разрывы в первой и второй целевых последовательностях направляющей РНК). Подобным образом, третью и четвертую целевые последовательности направляющей РНК можно расположить таким образом, чтобы создать второй сайт расщепления так, что однонитевые разрывы, образованные третьей и четвертой никазами в первой и второй нитях ДНК, образуют двухнитевый разрыв (т.е. второй сайт расщепления включает в себя однонитевые разрывы в третьей и четвертой целевых последовательностях направляющей РНК). Предпочтительно однонитевые разрывы в первой и второй целевых последовательностях направляющей РНК и/или третьей и четвертой целевых последовательностях направляющей РНК могут быть смещенными однонитевыми разрывами, которые образуют выступающие концы. Окно смещения может составлять, например, по меньшей мере приблизительно 5 пар оснований, 10 пар оснований, 20 пар оснований, 30 пар оснований, 40 пар оснований, 50 пар оснований, 60 пар оснований, 70 пар оснований, 80 пар оснований, 90 пар оснований, 100 пар оснований или больше. См. Ran et al. (2013) Cell 154:1380-1389; Mali et al. (2013) Nat. Biotech.31:833-838; и Shen et al. (2014) Nat. Methods 11:399-404, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. В таких случаях двухнитевая экзогенная донорная нуклеиновая кислота может быть разработана с однонитевыми комплементарными областями, которые являются комплементарными по отношению к выступающим концам, образованными однонитевыми разрывами в первой и второй целевых последовательностях направляющей РНК и однонитевыми разрывами в третьей и четвертой целевых последовательностях направляющей РНК. Затем такая экзогенная донорная нуклеиновая кислота может быть вставлена путем опосредованного негомологичным соединением концов лигирования.
[00231] Донорные нуклеиновые кислоты для вставки с помощью направляемой гомологией репарации. Некоторые экзогенные донорные нуклеиновые кислоты включают в себя плечи гомологии. Если экзогенная донорная нуклеиновая кислота также включает в себя вставку нуклеиновой кислоты, то плечи гомологии могут фланкировать вставку нуклеиновой кислоты. Для удобства пользования плечи гомологии называют в настоящем документе как 5' и 3' плечи гомологии (т.е. в 3'-5' направлении и в 5'-3' направлении). Данная терминология относится к относительному положению плеч гомологии по отношению к вставке нуклеиновой кислоты в экзогенной донорной нуклеиновой кислоте. 5' и 3' плечи гомологии соответствуют областям в гуманизированном TTR локусе, которые упоминаются в настоящем документе как «5' целевая последовательность» и «3' целевая последовательность», соответственно.
[00232] Плечо гомологии и целевая последовательность «соответствуют» или являются «соответствующими» друг другу, если две области обладают достаточным уровнем идентичности последовательностей друг другу для действия в качестве субстратов в реакции гомологичной рекомбинации. Термин «гомология» включает в себя последовательности ДНК, которые либо являются идентичными, либо обладают идентичностью последовательностей по отношению к соответствующей последовательности. Идентичность последовательностей между данной целевой последовательностью и соответствующим плечом гомологии, находящимся в экзогенной донорной нуклеиновой кислоте, может иметь любую степень идентичности последовательностей для обеспечения осуществления гомологичной рекомбинации. Например, количественный показатель идентичности последовательностей, которой характеризуются плечо гомологии экзогенной донорной нуклеиновой кислоты (или ее фрагмента) и целевая последовательность (или ее фрагмент), может составлять по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей, так что последовательности подвергаются гомологичной рекомбинации. Более того, соответствующая область гомологии между плечом гомологии и соответствующей целевой последовательностью может иметь любую длину, которая достаточна для обеспечения гомологичной рекомбинации. Иллюстративные плечи гомологии составляют от приблизительно 25 нуклеотидов до приблизительно 2,5 т.п.н. в длину, от приблизительно 25 нуклеотидов до приблизительно 1,5 т.п.н. в длину или от приблизительно 25 до приблизительно 500 нуклеотидов в длину. Например, данное плечо гомологии (или каждое из плеч гомологии) и/или соответствующая целевая последовательность могут включать в себя соответствующие области гомологии, которые составляют приблизительно 25-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-150, 150-200, 200-250, 250-300, 300-350, 350-400, 400-450 или 450-500 нуклеотидов в длину, так что плечи гомологии характеризуются достаточной гомологией, чтобы подвергаться гомологичной рекомбинации с соответствующими целевыми последовательностями в целевой нуклеиновой кислоте. В качестве альтернативы, данное плечо гомологии (или каждое плечо гомологии) и/или соответствующая целевая последовательность могут включать в себя соответствующие области гомологии, которые составляют от приблизительно 0,5 т.п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 1,5 т.п.н., от приблизительно 1,5 т.п.н. до приблизительно 2 т.п.н. или от приблизительно 2 т.п.н. до приблизительно 2,5 т.п.н. в длину. Например, каждое из плеч гомологии может составлять приблизительно 750 нуклеотидов в длину. Плечи гомологии могут быть симметричными (длина каждого приблизительно одинакова), или они могут быть асимметричными (одно длиннее другого).
[00233] При использовании нуклеазного средства в комбинации с экзогенной донорной нуклеиновой кислотой 5' и 3' целевые последовательности предпочтительно располагаются в достаточной близости к сайту расщепления нуклеазой (например, в достаточной близости к целевой последовательности нуклеазы), чтобы способствовать осуществлению события гомологичной рекомбинации между целевыми последовательностями и плечами гомологии при однонитевом разрыве (нике) или двухнитевом разрыве по сайту расщепления нуклеазой. Термин «сайт расщепления нуклеазой» включает в себя последовательность ДНК, в которой образуется однонитевый разрыв или двухнитевый разрыв с помощью нуклеазного средства (например, белка Cas9 в комплексе с направляющей РНК). Целевые последовательности в целевом локусе, которые соответствуют 5' и 3' плечам гомологии экзогенной донорной нуклеиновой кислоты «располагаются в достаточной близости» к сайту расщепления нуклеазой, если расстояние является таким, что способствует осуществлению события гомологичной рекомбинации между 5' и 3' целевыми последовательностями и плечами гомологии при однонитевом разрыве или двухнитевом разрыве в сайте расщепления нуклеазой. Таким образом, целевые последовательности, соответствующие 5' и/или 3' плечам гомологии экзогенной донорной нуклеиновой кислоты, могут находиться, например, в пределах по меньшей мере 1 нуклеотида от данного сайта расщепления нуклеазой или в пределах по меньшей мере от 10 нуклеотидов до приблизительно 1000 нуклеотидов от данного сайта расщепления нуклеазой. В качестве примера, сайт расщепления нуклеазой может непосредственно соседствовать по меньшей мере с одной или обеими целевыми последовательностями.
[00234] Пространственное расположение целевых последовательностей, которые соответствуют плечам гомологии экзогенной донорной нуклеиновой кислоты, и сайта расщепления нуклеазой может варьировать. Например, целевые последовательности могут быть расположены 5' по отношению к сайту расщепления нуклеазой, целевые последовательности могут быть расположены 3' по отношению к сайту расщепления нуклеазой, или целевые последовательности могут фланкировать сайт расщепления нуклеазой.
(4) Другие нацеливающиеся на человеческий TTR реагенты
[00235] Активность любого другого известного или предполагаемого нацеливающегося на человеческий TTR реагента также может быть оценена с использованием отличных от человека животных, раскрываемых в настоящем документе. Подобным образом, любая другая молекула может быть скринирована по нацеливающейся на человеческий TTR активности с использованием отличных от человека животных, раскрываемых в настоящем документе.
[00236] Примеры других нацеливающихся на человеческий TTR реагентов включают в себя антисмысловые олигонуклеотиды (например, siRNA или shRNA), которые действуют посредством РНК-интерференции (RNAi). Антисмысловые олигонуклеотиды (ASO) или антисмысловые РНК представляют собой короткие синтетические нити нуклеотидов, разработанные для предупреждения экспрессии целевого белка путем селективного связывания с РНК, которая кодирует целевой белок и тем самым предупреждает трансляцию. Эти соединения связываются с РНК с высокой аффинностью и селективностью благодаря хорошо охарактеризованному спариванию оснований Уотсона-Крика (гибридизации). РНК-интерференция (RNAi) представляет собой эндогенный клеточный механизм для контроля экспрессии генов, при котором малые интерферирующие РНК (siRNA), которые связаны с РНК-индуцируемым сайленсинг-комплексом (RISC), опосредуют расщепление целевой информационной РНК (иРНК). Примеры нацеливающихся на человеческий TTR антисмысловых олигонуклеотидов известны. См., например, Ackermann et al. (2012) Amyloid Suppl 1:43-44; и Coelho et al. (2013) N. Engl. J. Med. 369(9):819-829, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
Ф[00237] Другие нацеливающиеся на человеческий TTR реагенты включают в себя антитела или антигенсвязывающие белки, разработанные для специфического связывания человеческого TTR эпитопа.
[00238] Другие нацеливающиеся на человеческий TTR реагенты включают в себя низкомолекулярные реагенты. Одним примером такого низкомолекулярного реагента является тафамидис, который функционирует путем кинетической стабилизации корректно свернутой тетрамерной формы транстиретинового (TTR) белка. См., например, Hammarstrom et al. (2003) Science 299:713-716, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
D. Ведение нацеливающихся на человеческий TTR реагентов в отличных от человека животных или клетки
[00239] Способы, раскрываемые в настоящем документе, могут предусматривать введение в отличное от человека животное или клетку различных молекул (например, нацеливающихся на человеческий TTR реагентов, таких как терапевтические молекулы или комплексы), в том числе нуклеиновых кислот, белков, комплексов нуклеиновая кислота-белок или белковых комплексов. Термин «введение» включает в себя обеспечение в клетке или отличном от человека животном молекулы (например, нуклеиновой кислоты или белка) таким образом, что это обеспечивает доступ к внутренней части клетки или к внутренней части клетки в отличном от человека животном. Введение может быть выполнено любыми средствами, и два или более из компонентов (например, два компонента или все компоненты) могут быть введены в клетку или отличное от человека животное одновременно или последовательно в любой комбинации. Например, белок Cas может быть введен в клетку или отличное от человека животное до введения направляющей РНК, или он может быть введен после введения направляющей РНК. В качестве другого примера, экзогенная донорная нуклеиновая кислота может быть введена до введения белка Cas и направляющей РНК, или она может быть введена после введения белка Cas и направляющей РНК (например, экзогенная донорная нуклеиновая кислота может быть введена в пределах приблизительно 1, 2, 3, 4, 8, 12, 24, 36, 48 или 72 часов до или после введения белка Cas и направляющей РНК). См., например, US 2015/0240263 и US 2015/0110762, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Кроме того, два или более из компонентов могут быть введены в клетку или отличное от человека животное одним и тем же способом доставки или разными способами доставки. Подобным образом, два или более из компонентов могут быть введены в отличное от человека животное одним и тем же путем введения или разными путями введения.
[00240] В некоторых способах компоненты системы CRISPR/Cas вводят в отличное от человека животное или клетку. Направляющая РНК может быть введена в отличное от человека животное или клетку в форме РНК (например, in vitro транскрибированной РНК) или в форме ДНК, кодирующей направляющую РНК. При введении в форме ДНК кодирующая направляющую РНК ДНК может быть функционально связана с промотором, активным в клетке у отличного от человека животного. Например, направляющая РНК может быть доставлена посредством AAV и экспрессирована in vivo под промотором U6. Такие ДНК могут находиться в одной или нескольких экспрессионных конструкциях. Например, такие экспрессионные конструкции могут быть компонентами одной молекулы нуклеиновой кислоты. В качестве альтернативы, они могут быть разделены в любой комбинации на две или более молекул нуклеиновой кислоты (т.е. ДНК, кодирующие одну или несколько РНК CRISPR, и ДНК, кодирующие одну или несколько tracrRNA, могут быть компонентами отдельных молекул нуклеиновой кислоты).
[00241] Подобным образом, белки Cas, могут быть обеспечены в любой форме. Например, белок Cas может быть обеспечен в форме белка, такого как белок Cas, образующий комплекс с gRNA. В качестве альтернативы, белок Cas может быть обеспечен в форме нуклеиновой кислоты, кодирующей белок Cas, такой как РНК (например, информационной РНК (иРНК)) или ДНК. Необязательно нуклеиновая кислота, кодирующая белок Cas, может быть оптимизирована по кодону для эффективной трансляции в белок в конкретных клетке или организме. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована с замещением кодонов, характеризующихся более высокой частотой использования в клетке млекопитающего, клетке грызуна, клетке мыши, клетке крысы или в любой другой представляющей интерес клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью. При введении нуклеиновой кислоты, кодирующей белок Cas, в отличное от человека животное, белок Cas может быть временно, условно или конститутивно экспрессирован в клетке у отличного от человека животного.
[00242] Нуклеиновые кислоты, кодирующие белки Cas или направляющие РНК, могут быть функционально связаны с промотором в экспрессионной конструкции. Экспрессионные конструкции включают в себя любые конструкции нуклеиновой кислоты, способные управлять экспрессией гена или другой представляющей интерес последовательностью нуклеиновой кислоты (например, Cas геном), и которые могут переносить такую представляющую интерес последовательность нуклеиновой кислоты в целевую клетку. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в векторе, включающем в себя ДНК, кодирующую одну или несколько gRNA. В качестве альтернативы, она может находиться в векторе или плазмиде, которая отделена от вектора, включающего в себя ДНК, кодирующую одну или несколько gRNA. Подходящие промоторы, которые могут быть использованы в экспрессионной конструкции включают в себя промоторы, активные, например, в одной или нескольких из эукариотической клетки, человеческой клетки, не являющейся человеческой клетки, клетки млекопитающего, клетки отличного от человека млекопитающего, клетки грызуна, клетки мыши, клетки крысы, клетки хомяка, клетки кролика, плюрипотентной клетки, эмбриональной стволовой (ES) клетки, стволовой клетки взрослого человека, онтогенетически ограниченной клетки-предшественника, индуцированной плюрипотентной стволовой (iPS) клетки или эмбриона на стадии одной клетки. Такими промоторами могут быть, например, условные промоторы, индуцибельные промоторы, конститутивные промоторы или специфические по отношению к ткани промоторы. Необязательно промотор может быть двунаправленным промотором, управляющим экспрессией как белка Cas в одном направлении, так и направляющей РНК в другом направлении. Такие двунаправленные промоторы могут состоять из (1) полного традиционного однонаправленного промотора Pol III, который содержит 3 внешних контрольных элемента: элемент дистальной последовательности (DSE), элемент проксимальной последовательности (PSE) и ТАТА-бокс; и (2) второго основного промотора Pol III, который включает в себя PSE и ТАТА-бокс, слитый с 5'-концом DSE в обратной ориентации. Например, в промоторе HI DSE соседствует с PSE и ТАТА-боксом, и промотор можно сделать двунаправленным путем создания гибридного промотора, в котором транскрипция в обратном направлении контролируется путем добавления PSE и ТАТА-бокса, полученного из промотора U6. См., например, US 2016/0074535, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. Применение двунаправленного промотора для экспрессии генов, кодирующих белок Cas и направляющую РНК, одновременно позволяет создавать компактные экспрессионные кассеты для облегчения доставки.
[00243] Молекулы (например, белки Cas или направляющие РНК), вводимые в отличное от человека животное или клетку, могут быть обеспечены в композициях, включающих в себя носитель, повышающий стабильность вводимых молекул (например, пролонгирующие период хранения при данных условиях (например, при -20°С, 4°С или окружающей температуре), в течение которого разложение продуктов остается ниже порога, например, ниже 0,5% по массе исходных нуклеиновой кислоты или белка; или повышающий стабильность in vivo). Неограничивающие примеры таких носителей включают в себя микросферы поли(молочной кислоты) (PLA), микросферы сополимера D,L-молочной и гликолевой кислот (PLGA), липосомы, мицеллы, обращенные мицеллы, липидные кохлеаты и липидные микротрубочки.
[00244] В настоящем документе представлены различные способы и композиции для обеспечения введения нуклеиновой кислоты или белка в клетку или отличное от человека животное. Способы введения нуклеиновых кислот в различные типы клеток известны и включают в себя, например, способы стабильной трансфекции, способы временной трансфекции и опосредованные вирусом способы.
[00245] Протоколы трансфекции, а также протоколы для введения последовательностей нуклеиновой кислоты в клетки могут варьировать. Неограничивающие способы трансфекции включают в себя способы химической трансфекции с использованием липосом, наночастиц, фосфата кальция (Graham et al. (1973) Virology 52 (2): 456-67, Bacchetti et al. (1977) Proc. Natl. Acad. Sci. USA 74 (4): 1590-4, и Kriegler, M (1991). Transfer and Expression: A Laboratory Manual. New York: W. H. Freeman and Company, pp. 96-97); дендромеров или катионных полимеров, таких как DEAE-дектсран или полиэтиленимин. Нехимические способы включают в себя электропорацию, сонопорацию и оптическую трансфекцию. Трансфекция на основе частиц предусматривает применение генной пушки или трансфекции с помощью магнита (Bertram (2006) Current Pharmaceutical Biotechnology 7, 277-28). Вирусные способы также могут быть использованы для трансфекции.
[00246] Введение нуклеиновых кислот или белков в клетку также может быть опосредовано электропорацией, внутрицитоплазматической инъекцией, вирусной инфекцией, аденовирусом, адено-ассоциированным вирусом, лентивирусом, ретровирусом, трансфекцией, липид-опосредованной трансфекцией или нуклеофекцией. Нуклеофекция представляет собой улучшенную технологию электропорации, которая позволяет доставлять субстраты нуклеиновых кислот не только в цитоплазму, но и через ядерную мембрану, а также в ядро. Кроме того, применение нуклеофекции в способах, раскрываемых в настоящем документе, как правило, требует гораздо меньше клеток, чем обычная электропорация (например, только приблизительно 2 миллиона по сравнению с 7 миллионами при обычной электропорации). Согласно одному примеру нуклеофекцию выполняют с использованием системы LONZA® NUCLEOFECTOR™.
[00247] Введение нуклеиновых кислот или белков в клетку (например, зиготу) также может быть выполнено с помощью микроинъекции. В зиготах (т.е. эмбрионах на стадии одной клетки) микроинъекцию можно осуществлять в материнский и/или отцовский пронуклеус или в цитоплазму. Если микроинъекцию осуществляют только в один пронуклеус, то отцовский пронуклеус является предпочтительным из-за его более крупного размера. Микроинъекция иРНК осуществляется предпочтительно в цитоплазму (например, для доставки иРНК непосредственно в механизм трансляции), тогда как микроинъекция белка Cas или полинуклеотида, кодирующего белок Cas или кодирующего РНК, осуществляется предпочтительно в ядро/пронуклеус. В качестве альтернативы, микроинъекция может быть выполнена путем инъекции как в ядро/пронуклеус, так и в цитоплазму: сначала можно ввести иглу в ядро/пронуклеус и можно ввести первое количество, и при удалении иглы из эмбриона на стадии одной клетки второе количество может быть введено в цитоплазму. Если белок Cas инъецируют в цитоплазму, то белок Cas предпочтительно включает в себя сигнал ядерной локализации для обеспечения доставки в ядро/пронуклеус. Способы проведения микроинъекции хорошо известны. См., например, Nagy et al. (Nagy A, Gertsenstein M, Vintersten K, Behringer R., 2003, Manipulating the Mouse Embryo. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); см. также Meyer et al. (2010) Proc. Natl. Acad. Sci. USA 107: 15022-15026; и Meyer et al. (2012) Proc. Natl. Acad. Sci. USA 109: 9354-9359.
[00248] Другие способы введения нуклеиновой кислоты или белков в клетку или отличное от человека животное может предусматривать, например, векторную доставку, опосредованную частицами доставку, опосредованную экзосомой доставку, опосредованную липидными наночастицами доставку, опосредованную проникающим в клетку пептидом доставку или опосредованную имплантируемым устройством доставку. В качестве конкретных примеров, нуклеиновая кислота или белок могут быть введены в клетку или отличное от человека животное в носителе, таком как микросфера поли(молочной кислоты) (PLA), микросфера сополимера D,L-молочной и гликолевой кислот (PLGA), липосома, мицелла, обращенная мицелла, липидный кохлеат и липидная микротрубочка. Некоторые конкретные примеры доставки в отличное от человека животное включают в себя гидродинамическую доставку, опосредованную вирусом доставку (например, опосредованную адено-ассоциированным вирусом (AAV) доставку) и опосредованную липидными наночастицами доставку.
[00249] Введение нуклеиновых кислот и белков в клетки или в отличных от человека животных может быть выполнено путем гидродинамической доставки (HDD). Гидродинамическая доставка появилась как метод внутриклеточной доставки ДНК in vivo. Для доставки генов в паренхиматозные клетки через выбранный кровеносный сосуд необходимо вводить только необходимые последовательности ДНК, что устраняет проблемы безопасности, связанные с современными вирусными и синтетическими векторами. При инъекции в кровоток ДНК способна достигать клеток в различных тканях, доступных для крови. При гидродинамической доставке используется сила, создаваемая быстрой инъекцией большого объема раствора в несжимаемую кровь в кровотоке, чтобы преодолеть физические барьеры эндотелия и клеточных мембран, которые препятствуют проникновению крупных и непроницаемых для мембран соединений в паренхиматозные клетки. В дополнение к доставке ДНК этот способ применим для эффективной внутриклеточной доставки РНК, белков и других небольших соединений in vivo. См., например, Bonamassa et al. (2011) Pharm. Res. 28(4): 694-701, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00250] Введение нуклеиновых кислот также может быть достигнуто путем опосредованной вирусом доставки, такой как опосредованная AAV доставка или опосредованная лентивирусом доставка. Другие иллюстративные вирусы/вирусные векторы включают в себя ретровирусы, аденовирусы, вирусы коровьей оспы, поксвирусы и вирусы простого герпеса. Вирусы могут инфицировать делящиеся клетки, неделящиеся клетки или как делящиеся, так и неделящиеся клетки. Вирусы могут интегрироваться в геном хозяина или, в качестве альтернативы, не интегрироваться в геном хозяина. Такие вирусы также могут быть созданы методами генной инженерии для снижения иммунитета. Вирусы могут быть способными к репликации или могут быть дефектными по репликации (например, дефектными в одном или нескольких генах, необходимых для дополнительных циклов репликации и/или упаковки вириона). Вирусы могут вызывать временную экспрессию, длительную экспрессию (например, по меньшей мере, 1 неделя, 2 недели, 1 месяц, 2 месяца или 3 месяца) или постоянную экспрессию (например, Cas9 и/или рРНК). Иллюстративные вирусные титры (например, титры AAV) включают в себя 1012, 1013, 1014, 1015 и 1016 векторных геномов/мл.
[00251] Геном ssDNA AAV состоит из двух открытых рамок считывания. Rep и Cap, фланкированных двумя инвертированными концевыми повторами, которые позволяют синтезировать нить комплементарной ДНК. При конструировании транспортной плазмиды AAV трансген помещают между двумя ITR, a Rep и Cap могут доставляться in trans. В дополнение к Rep и Cap для AAV может потребоваться хелперная плазмида, включающая в себя гены из аденовируса. Эти гены (Е4, Е2а и VA) опосредуют репликацию AAV. Например, транспортная плазмида Rep/Cap и хелперная плазмида могут быть трансфицированы в клетки HEK293, включающие в себя ген аденовируса Е1+, для продуцирования инфекционных частиц AAV. В качестве альтернативы. Rep, Cap и хелперные гены аденовируса могут быть объединены в одну плазмиду. Подобные упаковывающие клетки и способы могут быть использованы для других вирусов, таких как ретровирусы.
[00252] Идентифицировали множество серотипов AAV. Эти серотипы отличаются по типам клеток, которые они инфицируют (т.е. по их тропизму), что позволяет преимущественную трансдукцию определенных типов клеток. Серотипы для ткани ЦНС включают в себя AAV1, AAV2, AAV4, AAV5, AAV8 и AAV9. Серотипы для сердечной ткани включают AAV1, AAV8 и AAV9. Серотипы для ткани почки включают в себя AAV2. Серотипы для ткани легкого включают в себя AAV4, AAV5, AAV6 и AAV9. Серотипы для ткани поджелудочной железы включают в себя AAV8. Серотипы для фоторецепторных клеток включают в себя AAV2, AAV5 и AAV8. Серотипы для ткани пигментного эпителия сетчатки включают в себя т AAV1, AAV2, AAV4, AAV5 и AAV8. Серотипы для скелетной мышечной ткани включают в себя AAV1, AAV6, AAV7, AAV8 и AAV9. Серотипы для ткани печени включают в себя AAV7, AAV8, AAV9 и, в частности, AAV8.
[00253] Тропизм можно дополнительно уточнить посредством псевдотипирования, которое представляет собой смешивание капсида и генома из различных вирусных серотипов. Например, AAV2/5 указывает на вирус, включающий в себя геном серотипа 2, упакованный в капсид из серотипа 5. Применение псевдотипированных вирусов может улучшать эффективность трансдукции, а также изменять тропизм. Гибридные капсиды, полученные из разных серотипов, также могут быть использованы для изменения вирусного тропизма. Например, AAV-DJ включает в себя гибридный капсид от восьми серотипов и демонстрирует высокую инфекционность в широком диапазоне клеточных типов in vivo. AAV-DJ8 является другим примером, который демонстрирует свойства AAV-DJ, но с усиленным поглощением в головном мозге. Серотипы AAV также могут быть модифицированы посредством мутаций. Примеры мутационных модификаций AAV2 включают в себя Y444F, Y500F, Y730F и S662V. Примеры мутационных модификаций AAV3 включают в себя Y705F, Y731F и T492V. Примеры мутационных модификаций AAV6 включают в себя S663V и T492V. Другие псевдотипированные/модифицированные варианты AAV включают в себя AAV2/1, AAV2/6, AAV2/7, AAV2/8, AAV2/9, AAV2.5, AAV8.2 и AAV/SASTG.
[00254] Для ускорения экспрессии трансгена могут использоваться варианты самокомплементарного AAV (scAAV). Поскольку AAV зависит от механизма репликации ДНК клетки, чтобы синтезировать комплементарную нить генома однонитевой ДНК AAV, экспрессия трансгена может быть отсрочена. Чтобы устранить эту задержку, можно использовать комплементарные последовательности, включающие в себя scAAV, которые способны к спонтанному отжигу при инфекции, что устраняет необходимость синтеза ДНК клетки-хозяина. Однако также можно использовать векторы однонитевого AAV (ssAAV).
[00255] Для повышения емкости упаковки более длинные трансгены могут быть разделены между двумя транспортными плазмидами AAV, первой с 3' донором сплайсинга и второй с 5' акцептором сплайсинга. При совместном инфицировании клетки эти вирусы образуют конкатемеры, сплайсируются вместе, и может быть экспрессирован полноразмерный трансген. Хотя это позволяет экспрессию более длинного трансгена, экспрессия менее эффективна. В подобных способах повышения емкости используют гомологичную рекомбинацию. Например, трансген может быть разделен между двумя транспортными плазмидами, но с существенным перекрыванием последовательностей, так что совместная экспрессия индуцирует гомологичную рекомбинацию и экспрессию полноразмерного трансгена.
[00256] Введение нуклеиновых кислот и белков также может быть выполнено путем опосредованной липидной наночастицей (LNP) доставки. Например, опосредованная LNP доставка может быть использована для доставки комбинации иРНК Cas и направляющей РНК или комбинации белка Cas и направляющей РНК. Доставка с помощью таких способов дает в результате временную экспрессию Cas и улучшенное выведение биоразлагаемых липидов, улучшенную переносимость и пониженную иммуногенность. Липидные составы могут защищать биологические молекулы от разложения с улучшением при этом их поглощения клеткой. Липидные наночастицы представляют собой частицы, включающие в себя множество липидных молекул, физически ассоциированных друг с другом с помощью межмолекулярных связей. Они включают в себя микросферы (в том числе однослойные и многослойные везикулы, например, липосомы), диспергированную фазу в эмульсии, мицеллы или внутреннюю фазу в суспензии. Такие липидные наночастицы могут быть использованы для инкапсулирования одной или нескольких нуклеиновых кислот или белков для доставки. Составы, которые содержат катионные липиды, применимы для доставки полианионов, таких как нуклеиновые кислоты. Другими липидами, которые могут быть включены, являются нейтральные липиды (т.е. незаряженные или цвиттер-ионные липиды), анионные липиды, хелперные липиды, которые усиливают трансфекцию, и липиды-невидимки, которые увеличивают продолжительность времени, на протяжении которого наночастицы могут существовать in vivo. Примеры подходящих катионных липидов, нейтральных липидов, анионных липидов, хелперных липидов и липидов-невидимок могут быть найдены в WO 2016/010840 A1, включенной в настоящий документ посредством ссылки в полном своем объеме для всех целей. Иллюстративная липидная наночастица может включать в себя катионный липид и один или несколько других компонентов. Согласно одному примеру другой компонент может включать в себя хелперный липид, такой как холестерин. Согласно другому примеру другие компоненты могут включать в себя хелперный липид, такой как холестерин, и нейтральный липид, такой как DSPC. Согласно другому примеру другие компоненты могут включать в себя хелперный липид, такой как холестерин, необязательный нейтральный липид, такой как DSPC, и липид-невидимку, такой как S010, S024, S027, S031 или S033.
[00257] LNP может содержать одно, или несколько, или все из следующих: (i) липид для инкапсулирования и для эндосомального высвобождения; (ii) нейтральный липид для стабилизации; (in) хелперный липид для стабилизации и (iv) липид-невидимка. См., например, Finn et al. (2018) Cell Reports 22: 1-9; и WO 2017/173054 А1, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. В некоторых LNP груз может включать в себя направляющую РНК или нуклеиновую кислоту, кодирующую направляющую РНК. В некоторых LNP груз может включать в себя иРНК, кодирующую нуклеазу Cas, такую как Cas9, и направляющую РНК или нуклеиновую кислоту, кодирующую направляющую РНК.
[00258] Липид для инкапсулирования и эндосомального высвобождения может представлять собой катионный липид. Липид также может представлять собой биоразлагаемый липид, такой как биоразлагаемый ионизируемый липид. Одним примером подходящего липида является липид А или LP01, который представляет собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-октадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-(9Z,12Z)-октадека-9,12-диеноатом. См., например Finn et al. (2018) Cell Reports 22: 1-9; и WO 2017/173054 A1, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Другим примером подходящего липида является липид В, который представляет собой ((5-((диметиламино)метил)-1,3-фенилен)бис(окси))бис(октан-8,1-диил)бис(деканоат), также называемый ((5-((диметиламино)метил)-1,3-фенилен)бис(окси))бис(октан-8,1-диил)бис(деканоатом). Другим примером подходящего липида является липид С, который представляет собой 2-((4-(((3-(диметиламино)пропокси)карбонил)окси)гексадеканоил)окси)пропан-1,3-диил(9Z,9'Z,12Z,12'Z)-бис(октадека-9,12-диеноат). Другим примером подходящего липида является липид D, который представляет собой 3-(((3-(диметиламино)пропокси)карбонил)окси)-13-(октаноилокси)тридецил-3-октилундеканоат. Другие подходящие липиды включают в себя гептатриаконта-6,9,28,31-тетраен-19-ил-4-(диметиламино)бутаноат (также известный как Dlin-MC3-DMA (MC3))).
[00259] Некоторые такие липиды, подходящие для применения в LNP, описываемых в настоящем документе, являются биоразлагаемыми in vivo. Например, LNP, включающие в себя такие липиды, включают в себя те, в которых по меньшей мере 75% липида выводится из плазмы за 8, 10, 12, 24 или 48 часов, или за 3, 4, 5, 6, 7 или 10 суток. В качестве другого примера, по меньшей мере 50% LNP выводится из плазмы за 8, 10, 12, 24 или 48 часов, или за 3, 4, 5, 6, 7 или 10 суток.
[00260] Такие липиды могут быть ионизируемыми в зависимости от рН среды, в которой они находятся. Например, в слабокислой среде липиды могут протонироваться и, таким образом, нести положительный заряд. И наоборот, в слабоосновной среде, такой как, например, кровь, где рН составляет приблизительно 7,35, липиды не могут протонироваться и, таким образом, нести заряд. Согласно некоторым вариантам осуществления липиды могут быть протонированы при рН по меньшей мере приблизительно 9, 9,5 или 10. Способность такого липида нести заряд связана с его собственной pKa. Например, липид независимо может характеризоваться pKa в диапазоне от приблизительно 5,8 до приблизительно 6,2.
[00261] Нейтральные липиды функционируют со стабилизацией и улучшением процессинга LNP. Примеры подходящих нейтральных липидов включают в себя ряд нейтральных, незаряженных или цвиттер-ионных липидов. Примеры нейтральных фосфолипидов, подходящих для применения в настоящем раскрытии, включают в себя без ограничения 5-гептадецилбензол-1,3-диол (резорцин), дипальмитоилфосфатидилхолин (DPPC), дистеароилфосфатидилхолин (DSPC), фосфохолин (DOPC), димиристоилфосфатидилхолин (DMPC), фосфатидилхолин (PLPC), 1,2-дистеароил-sn-глицеро-3-фосфохолин (DAPC), фосфатидилэтаноламин (РЕ), яичный фосфатидилхолин (ЕРС), дилаурилоилфосфатидилхолин (DLPC), димиристоилфосфатидилхолин (DMPC), 1-миристоил-2-пальмитоилфосфатидилхолин (МРРС), 1-пальмитоил-2-миристоилфосфатидилхолин (РМРС), 1-пальмитоил-2-стеароилфосфатидилхолин (PSPC), 1,2-диарахидоил-sn-глицеро-3-фосфохолин (DBPC), 1-стеароил-2-пальмитоилфосфатидилхолин (SPPC), 1,2-диэйкозеноил-sn-глицеро-3-фосфохолин (DEPC), пальмитоилолеоилфосфатидилхолин (РОРС), лизофосфатидилхолин, диолеоилфосфатидилэтаноламин (DOPE), дилинолеоилфосфатидилхолиндистеароилфосфатидилэтаноламин (DSPE), димиристоилфосфатидилэтаноламин (DMPE), дипальмитоилфосфатидилэтаноламин (DPPE), пальмитоилолеоилфосфатидилэтаноламин (POPE), лизофосфатидилэтаноламин и их комбинации. Например, нейтральный фосфолипид может быть выбран из группы, состоящей из дистеароилфосфатидилхолина (DSPC) и димиристоилфосфатидилэтаноламина (DMPE).
[00262] Хелперные липиды включают в себя липиды, которые усиливают трансфекцию. Механизм, с помощью которого хелперный липид усиливает трансфекцию, может включать в себя усиление стабильности частиц. В некоторых случаях хелперный липид может усиливать мембранную фузогенность. Хелперные липиды включают в себя стероиды, стеролы и алкилрезорцины. Примеры подходящих хелперных липидов включают в себя холестерин, 5-гептадецилрезорцин и холестерина гемисукцинат. Согласно одному примеру хелперный липид может представлять собой холестерин или холестерина гемисукцинат.
[00263] Липиды-невидимки включают в себя липиды, которые изменяют продолжительность времени существования наночастиц in vivo. Липиды-невидимки могут способствовать процессу составления, например, путем уменьшения агрегации частиц и контроля размера частиц. Липиды-невидимки могут модулировать фармакокинетические свойства LNP. Подходящие липиды-невидимки включают в себя липиды, имеющие гидрофильную концевую группу, связанную с липидным фрагментом.
[00264] Гидрофильные концевые группы липида-невидимки могут включать в себя, например, полимерный фрагмент, выбранный из полимеров на основе PEG (иногда называемых поли(этиленоксидом)), поли(оксазолина), поли(винилового спирта), поли(глицерина), поли(N-винилпирролидона), полиаминокислот и поли-N-(2-гидроксипропил)метакриламида. Термин «PEG» означает любой полиэтиленгликоль или другой полимер полиалкиленового эфира. В некоторых составах LNP PEG представляет собой PEG-2K, также называемый PEG 2000, который характеризуется средней молекулярной массой приблизительно 2000 дальтон. См., например, WO 2017/173054 A1, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00265] Липидный фрагмент липида-невидимки может быть получен, например, из диацилглицерина или диацилгликамида, в том числе из включающих в себя диалкилглицериновую или диалкилгликамидную группу с алкильной цепью, независимо включающей в себя от приблизительно С4 до приблизительно С40 насыщенных или ненасыщенных атомов углерода, при этом цепь может включать в себя одну или несколько функциональных групп, таких как, например, амид или сложный эфир. Диалкилглицериновая или диалкилгликамидная группа может дополнительно включать в себя одну или несколько замещенных алкильных групп.
[00266] В качестве одного примера, липид-невидимка может быть выбран из PEG-дилауроилглицерина, PEG-димиристоилглицерина (PEG-DMG), PEG-дипальмитоилглицерина, PEG-дистеароилглицерина (PEG-DSPE), PEG-дилаурилгликамида, PEG-димиристилгликамида, PEG-дипальмитоилгликамида и PEG-дистеароилгликамида, PEG-холестерина (1-[8'-(холест-5-ен-3[бета]-окси)карбоксамидо-3',6'-диоксаоктанил]карбамоил-[омега]-метил-поли(этиленгликоля), PEG-DMB (3,4-дитетрабекоксилбензил-[омега]-метил-поли(этиленгликоль)эфира), 1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин-N-[метокси(полиэтиленгликоля)-2000] (PEG2k-DMG), 1,2-дистеароил-sn-глицеро-3-фосфоэтаноламин-N-[метокси(полиэтиленгликоля)-2000] (PEG2k-DSPE), 1,2-дистеароил-sn-глицерина, метоксиполиэтиленгликоля (PEG2k-DSG), поли(этиленгликоль)-2000-диметакрилата (PEG2k-DMA) и 1,2-дистеарилоксипропил-3-амин-N-[метокси(полиэтиленгликоля)-2000] (PEG2k-DSA). Согласно одному конкретному примеру липид-невидимка может представлять собой PEG2k-DMG.
[00267] LNP могут характеризоваться различными соответствующими молярными отношениями компонентных липидов в составе. Мол. % CCD липида может составлять, например, от приблизительно 30 мол. % до приблизительно 60 мол. %, от приблизительно 35 мол. % до приблизительно 55 мол. %, от приблизительно 40 мол. % до приблизительно 50 мол. %, от приблизительно 42 мол. % до приблизительно 47 мол. % или приблизительно 45%. Мол. % хелперного липида может составлять, например, от приблизительно 30 мол. % до приблизительно 60 мол. %, от приблизительно 35 мол. % до приблизительно 55 мол. %, от приблизительно 40 мол. % до приблизительно 50 мол. %, от приблизительно 41 мол. % до приблизительно 46 мол. % или приблизительно 44 мол. %. Мол. % нейтрального липида может составлять, например, от приблизительно 1 мол. % до приблизительно 20 мол. %, от приблизительно 5 мол. % до приблизительно 15 мол. %, от приблизительно 7 мол. % до приблизительно 12 мол. % или приблизительно 9 мол. %. Мол. % липида-невидимки может составлять, например, от приблизительно 1 мол. % до приблизительно 10 мол. %, от приблизительно 1 мол. % до приблизительно 5 мол. %, от приблизительно 1 мол. % до приблизительно 3 мол. %, приблизительно 2 мол. % или приблизительно 1 мол. %.
[00268] LNP могут характеризоваться разными отношениями между положительно заряженными аминовыми группами биоразлагаемого липида (N) и отрицательно заряженными фосфатными группами (Р) нуклеинововой кислоты, подлежащей инкапсулированию. Это может быть математически представлено уравнением N/P. Например, отношение N/P может составлять от приблизительно 0,5 до приблизительно 100, от приблизительно 1 до приблизительно 50, от приблизительно 1 до приблизительно 25, от приблизительно 1 до приблизительно 10, от приблизительно 1 до приблизительно 7, от приблизительно 3 до приблизительно 5, от приблизительно 4 до приблизительно 5, приблизительно 4, приблизительно 4,5 или приблизительно 5. Отношение N/P также может составлять от приблизительно 4 до приблизительно 7 или от приблизительно 4,5 до приблизительно 6. Согласно конкретным примерам отношение N/P может составлять 4,5 или может составлять 6. [00269] В некоторых LNP груз может включать в себя иРНК Cas и gRNA. иРНК Cas и gRNA могут находиться в разных отношениях. Например, состав LNP может характеризоваться отношением иРНК Cas к нуклеиновой кислоте gRNA, варьирующим от приблизительно 25:1 до приблизительно 1:25, варьирующим от приблизительно 10:1 до приблизительно 1:10, варьирующим от приблизительно 5:1 до приблизительно 1:5 или приблизительно 1:1. В качестве альтернативы, состав LNP может характеризоваться отношением иРНК Cas к нуклеиновой кислоте gRNA от приблизительно 1:1 до приблизительно 1:5 или приблизительно 10:1. В качестве альтернативы, состав LNP может характеризоваться отношением иРНК Cas к нуклеиновой кислоте gRNA приблизительно 1:10, 25:1, 10:1, 5:1, 3:1, 1:1, 1:3, 1:5, 1:10 или 1:25. В качестве альтернативы, состав LNP может характеризоваться отношением иРНК Cas к нуклеиновой кислоте gRNA от приблизительно 1:1 до приблизительно 1:2. Согласно конкретным примерам отношение иРНК Cas к gRNA может составлять приблизительно 1:1 или приблизительно 1:2.
[00270] В некоторых LNP груз может включать в себя экзогенную донорную нуклеиновую кислоту и gRNA. Экзогенная донорная нуклеиновая кислота и gRNA могут находиться в разных отношениях. Например, состав LNP может характеризоваться отношением экзогенной донорной нуклеиновой кислоты к нуклеиновой кислоте gRNA, варьирующим от приблизительно 25:1 до приблизительно 1:25, варьирующим от приблизительно 10:1 до приблизительно 1:10, варьирующим от приблизительно 5:1 до приблизительно 1:5 или приблизительно 1:1. В качестве альтернативы, состав LNP может характеризоваться отношением экзогенной донорной нуклеиновой кислоты к нуклеиновой кислоте gRNA от приблизительно 1:1 до приблизительно 1:5, приблизительно 5:1 до приблизительно 1:1, приблизительно 10:1 или приблизительно 1:10. В качестве альтернативы, состав LNP может характеризоваться отношением экзогенной донорной нуклеиновой кислоты к нуклеиновой кислоте gRNA приблизительно 1:10,25:1, 10:1, 5:1, 3:1, 1:1, 1:3, 1:5, 1:10 или 1:25.
[00271] Конкретный пример подходящей LNP характеризуется отношением азота к фосфату (N/P) 4,5 и включает в себя биоразлагаемый катионный липид, холестерин, DSPC и PEG2k-DMG в молярном отношении 45:44:9:2. Биоразлагаемый катионный липид может представлять собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилоктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил(9Z,12Z)-октадека-9,12-диеноатом. См., например, Finn et al. (2018) Cell Reports 22: 1-9, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей. иРНК Cas9 может находиться в отношении по массе 1:1 к направляющей РНК. Другой конкретный пример подходящей LNP включает в себя Dlin-MC3-DMA (MC3), холестерин, DSPC и PEG-DMG в молярном отношении 50:38,5:10:1,5.
[00272] Другой конкретный пример подходящей LNP характеризуется отношением азота к фосфату (N/P) 6 и включает в себя биоразлагаемый катионный липид, холестерин, DSPC и PEG2k-DMG в молярном отношении 50:38:9:3. Биоразлагаемый катионный липид может представлять собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилоктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил(9Z,12Z)-октадека-9,12-диеноатом, иРНК Cas9 может находиться в отношении 1:2 по массе к направляющей РНК.
[00273] Способ доставки может быть выбран для снижения иммуногенности. Например, белок Cas и gRNA могут быть доставлены различными способами (например, бимодальной доставкой). Такие разные способы могут обеспечивать различные фармакодинамические параметры или фармакокинетические свойства у рассматриваемой доставляемой молекулы (например, Cas или кодирующей нуклеиновой кислоты, gRNA или кодирующей нуклеиновой кислоты, или экзогенной донорной нуклеиновой кислоты/матрицы для репарации). Например, разные способы могут приводить в результате к разному распределению в ткани, разному период полувыведения или разному распределению во времени. Некоторые способы доставки (например, доставка вектора нуклеиновой кислоты, который сохраняется в клетке посредством автономной репликации или геномной интеграции) приводят в результате к более постоянной экспрессии и присутствию молекулы, тогда как другие способы доставки являются временными и менее постоянными (например, доставка РНК или белка). Доставка белков Cas более кратковременным способом, например, в виде мРНК или белка, может гарантировать, что комплекс Cas/gRNA присутствует и активен только в течение короткого периода времени, и может снизить иммуногенность, вызванную пептидами из бактериального фермента Cas, представленного на поверхности клетки молекулами МНС. Такая временная доставка может также уменьшить вероятность нецелевых модификаций.
[00274] Введение in vivo может быть осуществлено любым подходящим путем, в том числе, например, парентеральным, внутривенным, пероральным, подкожным, внутриартериальным, внутричерепным, внутритекальным, внутриперитонеальным, местным, интраназальным или внутримышечным. Системные способы введения включают в себя, например, пероральный и парентеральный пути. Примеры парентеральных путей включают в себя внутривенные, внутриартериальные, внутрикостные, внутримышечные, внутрикожные, подкожные, внутриназальные и внутриперитонеальные пути. Конкретным примером является внутривенная инфузия. Назальная инстилляция и интравитреальная инъекция являются другими конкретными примерами. Локальные способы введения включают в себя, например, внутритекальный, интрацеребровентрикулярный, интрапаренхимальный (например, локализованную интрапаренхимальную доставку в стриатум (например, в хвостатое ядро или в путамен), кору головного мозга, прецентральную извилину, гиппокамп (например, в зубчатую извилину или область СА3), височную кору, миндалину, лобную кору, таламус, мозжечок, мозговое вещество, гипоталамус, тектум, тегментум или черную субстанцию), интраокулярный, интраорбитальный, субконъюктивальный, интравитреальный, субретинальный и транссклеральный пути. Значительно меньшие количества компонентов (по сравнению с системными подходами) могут оказывать эффект при локальном введении (например, интрапаренхимальном или интравитреальном) по сравнению с системным введением (например, внутривенным). Локальные способы введения могут также уменьшать или исключать частоту потенциально токсических побочных эффектов, которые могут возникнуть, когда терапевтически эффективные количества компонента вводятся системно.
[00275] Введение in vivo может быть осуществлено любым подходящим путем в том числе, например, парентеральным, внутривенным, пероральным, подкожным, внутриартериальным, внутричерепным, внутритекальным, внутриперитонеальным, местным, внутриназальным или внутримышечным. Конкретным примером является внутривенная инфузия. Композиции, включающие в себя направляющие РНК и/или белки Cas (или нуклеиновые кислоты, кодирующие направляющие РНК и/или белки Cas), могут быть составлены с использованием одного или нескольких физиологически и фармацевтически приемлемых носителей, разбавителей, вспомогательных средств или вспомогательных веществ. Состав может зависеть от выбранного пути введения. Термин «фармацевтически приемлемый» означает, что носитель, разбавитель, вспомогательное средство или вспомогательное вещество является совместимым с другими ингредиентами состава и, по сути, не опасным для его реципиента.
[00276] Частота введения и число дозировок могут зависеть от периода полувыведения экзогенных донорных нуклеиновых кислот, направляющих РНК или белков Cas (или нуклеиновых кислот, кодирующих направляющие РНК или белка Cas) и пути введения среди других факторов. Введение нуклеиновых кислот или белков в клетку или в отличное от человека животное может быть выполнено за один раз или несколько раз на протяжении периода времени. Например, введение может быть выполнено по меньшей мере два раза на протяжении периода времени, по меньшей мере три раза на протяжении периода времени, по меньшей мере четыре раза на протяжении периода времени, по меньшей мере пять раз на протяжении периода времени, по меньшей мере шесть раз на протяжении периода времени, по меньшей мере семь раз на протяжении периода времени, по меньшей мере восемь раз на протяжении периода времени, по меньшей мере девять раз на протяжении периодов времени, по меньшей мере десять раз на протяжении периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз на протяжении периода времени, по меньшей мере тринадцать раз на протяжении периода времени, по меньшей мере четырнадцать раз на протяжении периода времени, по меньшей мере пятнадцать раз на протяжении периода времени, по меньшей мере шестнадцать раз на протяжении периода времени, по меньшей мере семнадцать раз на протяжении периода времени, по меньшей мере восемнадцать раз на протяжении периода времени, по меньшей мере девятнадцать раз на протяжении периода времени или по меньшей мере двадцать раз на протяжении периода времени.
Е. Измерение доставки, активности или эффективности нацеливающихся на человеческий TTR реагентов in vivo или ex vivo
[00277] Способы, раскрываемые в настоящем документе, могут дополнительно предусматривать выявление или измерение активности нацеливающихся на человеческий TTR реагентов. Например, если нацеливающийся на человеческий TTR реагент представляет собой редактирующий геном реагент (например, CRISPR/Cas, разработанную для нацеливания на человеческий TTR локус), то измерение может включать в себя оценивание гуманизированного TTR локуса на предмет модификаций.
[00278] Различные способы могут быть использованы для идентификации клеток, имеющих целевую генетическую модификацию. Скрининг может включать в себя количественный анализ на предмет оценивания модификации аллеля (МОА) родительской хромосомы. Например, количественный анализ может быть выполнен посредством количественной ПЦР, такой как ПЦР в режиме реального времени (qPCR). При ПЦР в режиме реального времени можно использовать первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может включать в себя флуоресцентный зонд, который распознает амплифицированную последовательность. Другие примеры подходящих количественных анализов включают в себя опосредованную флуоресценцией in situ гибридизацию (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным зондом(ами), зонды INVADER®, зонды Molecular Beacon TAQMAN® или зондовую технологию ECLIPSE™ (см., например, US 2005/0144655, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей).
[00279] Секвенирование следующего поколения (NGS) также может быть использовано для скрининга. Секвенирование следующего поколения также может быть названо «NGS», или «массивно-параллельным секвенированием», или «секвенированием с высокой пропускной способностью». NGS может быть использовано в качестве инструмента скрининга в дополнение к анализам МОА для определения точной природы целевой генетической модификации и того, соответствует ли она типам клеток, или типам тканей, или типам органов.
[00280] Оценивание модификации гуманизированного TTR локуса у отличного от человека животного может быть осуществлено на любом типе клеток из любых ткани или органа. Например, оценивание может быть осуществлено на нескольких типах клеток из одних и тех же ткани или органа или на клетках из множества местоположений в ткани или органе. Это может предоставить информацию о том, на какие типы клеток в целевых ткани или органе происходит нацеливание, или какие участки ткани или органа достигаются нацеливающимся на человеческий TTR реагентом. В качестве другого примера, оценивание может быть осуществлено на нескольких типах ткани или в нескольких органах. В способах, при которых нацеливанию подлежит конкретный тип ткани, органа или клетки, это может предоставить информацию о том, насколько эффективно осуществляется нацеливание на эти ткань или орган, и возникают ли нецелевые эффекты в других тканях или органах.
[00281] Если реагент разработан для инактивации гуманизированного TTR локуса, воздействия на экспрессию гуманизированного TTR локуса, предотвращения трансляции иРНК гуманизированного TTR или очистки гуманизированного TTR белок, измерение может включать в себя оценивание экспрессии иРНК гуманизированного TTR или белка. Это измерение может быть осуществлено в печени или конкретных типах клеток или в участках печени, или оно может предусматривать измерение содержания в сыворотке крови секретируемого гуманизированного TTR белка.
[00282] Продуцирование и секреция гуманизированного TTR белка могут быть оценены любыми известными средствами. Например, экспрессия может быть оценена путем измерения содержания кодируемой иРНК в печени отличного от человека животного или содержания кодируемого белка в печени отличного от человека животного с использованием известных анализов. Секреция гуманизированного TTR белка может быть оценена путем измерения либо содержания в плазме, либо содержания в сыворотке крови кодируемого гуманизированного TTR белка у отличного от человека животного с использованием известных анализов.
IV. Способы создания отличных от человека животных, включающих в себя гуманизированный TTR локус
[00283] Представлены различные способы создания отличного от человека животного, включающего в себя гуманизированный TTR локус, как раскрывается в других частях настоящего документа. Любой удобный способ или протокол для получения генетически модифицированного организма подходит для получения такого генетически модифицированного отличного от человека животного. См., например, Cho et al. (2009) Current Protocols in Cell Biology 42:19.11:19.11.1-19.11.22; и Gama Sosa et al. (2010) Brain Struct. Funct. 214(2-3): 91-109, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Такие генетически модифицированные отличные от человека животные могут быть созданы, например, посредством нокина гена в целевом Ttr локусе.
[00284] Например, способ получения отличного от человека животного, включающего в себя гуманизированный TTR локус, может предусматривать: (1) модификацию генома плюрипотентной клетки с содержанием гуманизированного TTR локуса; (2) идентификацию или отбор генетически модифицированной плюрипотентной клетки, включающей в себя гуманизированный TTR локус; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион отличного от человека животного-хозяина и (4) имплантацию суррогатной матери и гестацию эмбриона-хозяина в суррогатной матери. Необязательно эмбрион-хозяин, включающий в себя модифицированную плюрипотентную клетку (например, не являющуюся человеческой ES клетку), может быть инкубирован до стадии бластулы перед имплантацией суррогатной матери и гестацией в суррогатной матери для получения отличного от человека животного F0. Суррогатная мать затем может произвести поколение отличного от человека животного F0, включающее в себя гуманизированный TTR локус.
[00285] Способы дополнительно могут предусматривать идентификацию клетки или животного, имеющих модифицированный целевой геномный локус (т.е. гуманизированный TTR локус). Различные способы могут быть использованы для идентификации клеток и животных, имеющих целевую генетическую модификацию.
[00286] Стадия скрининга может включать в себя, например, количественный анализ на предмет оценивания модификации аллеля (МОА) родительской хромосомы. Например, количественный анализ может быть выполнен посредством количественной ПЦР, такой как ПЦР в режиме реального времени (qPCR). В ПЦР в режиме реального времени можно использовать набор первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может включать в себя флуоресцентный зонд, который распознает амплифицированную последовательность.
[00287] Другие примеры подходящих количественных анализов включают в себя опосредованную флуоресценцией in situ гибридизацию (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным зондом(ами), зонды INVADER®, зонды Molecular Beacon TAQMAN® или зондовую технологию ECLIPSE™ (см., например, US 2005/0144655, включенную в настоящий документ посредством ссылки в полном своем объеме для всех целей).
[00288] Примером подходящей плюрипотентной клетки является эмбриональная стволовая (ES) клетка (например, ES клетка мыши или ES клетка крысы). Модифицированная плюрипотентная клетка может быть создана, например, с помощью рекомбинации путем (а) введения в клетку одного или нескольких нацеливающихся векторов, включающих в себя вставку нуклеиновой кислоты, фланкированную 5' и 3' плечами гомологии, соответствующими 5' и 3' целевым сайтам, при этом вставка нуклеиновой кислоты включает в себя гуманизированный TTR локус; и (b) идентификации по меньшей мере одной клетки, включающей в свой геном вставку нуклеиновой кислоты, интегрированную в эндогенный Ttr локус. В качестве альтернативы, модифицированная плюрипотентная клетка может быть создана путем (а) введения в клетку (i) нуклеазного средства, при этом нуклеазное средство индуцирует однонитевый разрыв или двухнитевый разрыв в целевой последовательности в эндогенном Ttr локусе; и (ii) одного или нескольких нацеливающихся векторов, включающих в себя вставку нуклеиновой кислоты, фланкированную 5' и 3' плечами гомологии, соответствующими 5' и 3' целевым сайтам, расположенным в достаточной близости к целевой последовательности, при этом вставка нуклеиновой кислоты включает в себя гуманизированный TTR локус; и (с) идентификации по меньшей мере одной клетки, включающей в себя модификацию (например, интеграцию вставки нуклеиновой кислоты) в эндогенном Ttr локусе. Может быть использовано любое нуклеазное средство, которое индуцирует однонитевый разрыв или двухнитевый разрыв в желаемой целевой последовательности. Примеры подходящих нуклеаз включают в себя подобную активаторам транскрипции эффекторную нуклеазу (TALEN), нуклеазу с «цинковыми пальцами» (ZFN), мегануклеазу и ассоциированные с короткими палиндромными регулярно расположенными группами повторами (CRISPR)/CRISPR (Cas) системы или компоненты таких систем (например, CRISPR/Cas9). См., например, US 2013/0309670 и US 2015/0159175, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00289] Согласно конкретному примеру способ создания отличного от человека животного, включающего в себя гуманизированный TTR локус, может предусматривать: (а) введение в отличное от человека животное эмбриональной стволовой (ES) клетки: (i) нуклеазного средства, которое нацеливается на целевую последовательность в эндогенном Ttr локусе; и (ii) нацеливающегося вектора, включающего в себя вставку нуклеиновой кислоты, включающую в себя человеческую TTR последовательность, фланкированную 5' плечом гомологии, соответствующим 5' целевой последовательности в эндогенном Ttr локусе, и 3' плечом гомологии, соответствующим 3' целевой последовательности в эндогенном Ttr локусе, при этом нацеливающийся вектор рекомбинируется с эндогенным Ttr локусом с получением генетически модифицированной не являющейся человеческой ES клетки, включающей в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность; (b) введение генетически модифицированной не являющейся человеческой ES клетки в эмбрион отличного от человека животного-хозяина; и (с) гестацию эмбриона отличного от человека животного-хозяина в суррогатной матери, при этом суррогатная мать производит генетически модифицированное отличное от человека животное-потомка F0, включающее в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность.
[00290] В некоторых таких способах нуклеазное средство может включать в себя белок Cas (например, белок Cas9) и направляющую РНК, которая нацеливается на целевую последовательность в эндогенном Ttr локусе, но также могут быть использованы другие подходящие нуклеазные средства. Системы CRISPR/Cas и CRISPR/Cas9 описываются более подробно в других частях настоящего документа. Необязательно могут быть использованы два или более (например, три или четыре) нуклеазных средств (например, направляющих РНК). Целевая(ые) последовательность(и) может(могут) представлять собой любую подходящую целевую последовательность в эндогенном Ttr локусе. Например, целевая(ые) последовательность(и) может(могут) находиться в пределах приблизительно 10, приблизительно 20, приблизительно 30, приблизительно 40, приблизительно 50, приблизительно 100, приблизительно 200, приблизительно 300, приблизительно 400, приблизительно 500, приблизительно 1000 нуклеотидов, приблизительно 2000, приблизительно 3000, приблизительно 4000 или приблизительно 5000 нуклеотидов от инициирующего кодона и/или терминирующего кодона (например, одна целевая последовательность в непосредственной близости от инициирующего кодона и одна целевая последовательность в непосредственной близости от терминирующего кодона).
[00291] В некоторых таких способах нацеливающийся вектор представляет собой большой нацеливающийся вектор по меньшей мере 10 т.п.н. в длину, или в котором общая длина 5' и 3' плеч гомологии составляет по меньшей мере 10 т.п.н., но другие типы экзогенных донорных нуклеиновых кислот также могут быть использованы и хорошо известны. 5' и 3' плечи гомологии могут соответствовать 5' и 3' целевым последовательностям, соответственно, которые фланкируют область, подлежащую замене человеческой TTR вставленной нуклеиновой кислотой, или которые фланкируют область, в которой человеческая TTR вставленная нуклеиновая кислота подлежит вставке. Экзогенная донорная нуклеиновая кислота или нацеливающийся вектор могут рекомбинироваться с целевым локусом с помощью направляемой гомологией репарации или могут быть вставлены с помощью NHEJ-опосредованной вставки с образованием гуманизированного TTR локуса.
[00292] Донорная клетка может быть введена в эмбрион-хозяина на любой стадии, такой как стадия бластулы или стадия преморулы (т.е. стадия 4 клеток или стадия 8 клеток). Получают потомство, которое способно передавать генетическую модификацию через зародышевую линию. См., например, патент США №7294754, включенный в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00293] В качестве альтернативы, способ получения отличных от человека животных, описываемых в других частях настоящего документа, может предусматривать: (1) модификацию генома эмбриона на стадии одной клетки с содержанием гуманизированного TTR локуса с использованием описанных выше способов модификации плюрипотентной клетки; (2) отбор генетически модифицированного эмбриона и (3) имплантацию суррогатной матери и гестацию в суррогатной матери генетически модифицированного эмбриона. Получают потомство, которое способно передавать генетическую модификацию через зародышевую линию.
[00294] Для создания отличных от человека млекопитающих животных можно также использовать методики ядерного переноса. В кратком изложении способы ядерного переноса могут предусматривать стадии: (1) энуклеирования ооцита или обеспечения энуклеированного ооцита; (2) выделения или обеспечения донорных клетки или ядра, которые должны быть объединены с энуклеированным ооцитом; (3) вставки клетки или ядра в энуклеированный ооцит с образованием восстановленной клетки; (4) имплантации восстановленной клетки в матку животного для формирования эмбриона и (5) обеспечения развития эмбриона. В таких способах ооциты обычно извлекают из умерщвленных животных, хотя их можно выделять также из яйцеводов и/или яичников живых животных. До энуклеации ооциты могут созревать в различных хорошо известных средах. Энуклеация ооцитов может быть выполнена с помощью ряда хорошо известных способов. Вставку донорных клетки или ядра в энуклеированный ооцит с образованием восстановленной клетки можно проводить перед слиянием посредством микроинъекции донорной клетки под вителлиновый слой. Слияние можно индуцировать с применением электрического импульса постоянного тока в плоскости контакта/слияния (электрослияние), путем воздействия на клетки химическими веществами, стимулирующими слияние, такими как полиэтиленгликоль, или путем использования инактивированного вируса, например вируса Сендай. Восстановленную клетку можно активировать с использованием электрического и/или неэлектрического средства до, во время и/или после слияния ядра донора и ооцита реципиента. Способы активации включают в себя электрические импульсы, химически индуцированный шок, пенетрацию с помощью сперматозоида, повышение содержания двухвалентных катионов в ооцитах, а также снижение фосфорилирования клеточных белков (например, с помощью ингибиторов киназы) в ооците. Активированные восстановленные клетки или эмбрионы можно культивировать в хорошо известной среде, после чего их переносят в матку животного. См., например, US 2008/0092249, WO 1999/005266, US 2004/0177390, WO 2008/017234 и патент США №7612250, каждые из которых включены в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00295] Различные способы, представленные в настоящем документе, обеспечивают создание генетически модифицированного отличного от человека животного F0, при этом клетки генетически модифицированного животного F0 включают в себя гуманизированный TTR локус. В зависимости от используемого способа создания животного F0 число клеток в животном F0, которые имеют гуманизированный TTR локус, будет варьировать. Введение донорных ES клеток в эмбрион на стадии преморулы из соответствующего организма (например, в эмбрион мыши на стадии 8 клеток), посредством, например, способа VELOCIMOUSE®, обеспечивает то, что более высокий процент популяции клеток животного F0 будет включать в себя клетки, имеющие представляющую интерес нуклеотидную последовательность, включающую в себя целевую генетическую модификацию. Например, по меньшей мере 50%, 60%, 65%, 70%, 75%, 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада отличного от человека животного F0 может включать в себя клеточную популяцию, имеющею целевую модификацию.
[00296] Клетки генетически модифицированного животного F0 могут быть гетерозиготными по гуманизированному TTR локусу или могут быть гомозиготными по гуманизированному TTR локусу.
[00297] Все заявки на выдачу патентов, веб-сайты, другие публикации, номера доступа и т.п., упомянутые выше или ниже, включены в настоящий документ посредством ссылки в полном своем объеме для любых целей в равной мере, как если бы было указано, что каждая индивидуальная позиция отдельно и в индивидуальном порядке включена тем самым путем ссылки на нее. Если в различные моменты времени с номером доступа связаны различные версии последовательности, подразумевается версия, связанная с номером доступа на действительную дату подачи настоящей заявки. Под действительной датой подачи подразумевается наступающая ранее из фактической даты подачи или даты подачи приоритетной заявки при приведении ссылки на номер доступа, если применимо. Подобным образом, если в различные моменты времени издаются различные версии публикации, веб-сайта и т.п., то, если не указано иное, подразумевается наиболее поздняя опубликованная версия на действительную дату подачи настоящей заявки. Если специально не указано иное, любые признак, стадия, элемент, вариант осуществления или аспект настоящего изобретения могут использоваться в комбинации с любым другим из перечисленного. Хотя настоящее изобретение было подробно описано посредством иллюстрации и примера в целях четкости и лучшего понимания, будет очевидно, что некоторые изменения и модификации могут быть осуществлены в рамках объема приложенной формулы изобретения.
Краткое описание последовательностей
[00298] Нуклеотидные и аминокислотные последовательности, перечисленные в приложенном перечне последовательностей, показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот. Нуклеотидные последовательности согласно стандартному соглашению начинаются с 5'-конца последовательности и продолжаются (т.е. слева направо в каждой строке) до 3'-конца. Показана только одна нить каждой нуклеотидной последовательности, но считается, что комплементарная нить включена в любую ссылку на отображаемую нить. Когда предоставляется нуклеотидная последовательность, кодирующая аминокислотную последовательность, подразумевается, что также предусматриваются ее кодон-вырожденные варианты, которые кодируют ту же аминокислотную последовательность. Аминокислотные последовательности согласно стандартному соглашению начинаются с амино-конца последовательности и продолжаются (т.е. слева направо в каждой строке) до карбокси-конца.
[00299]
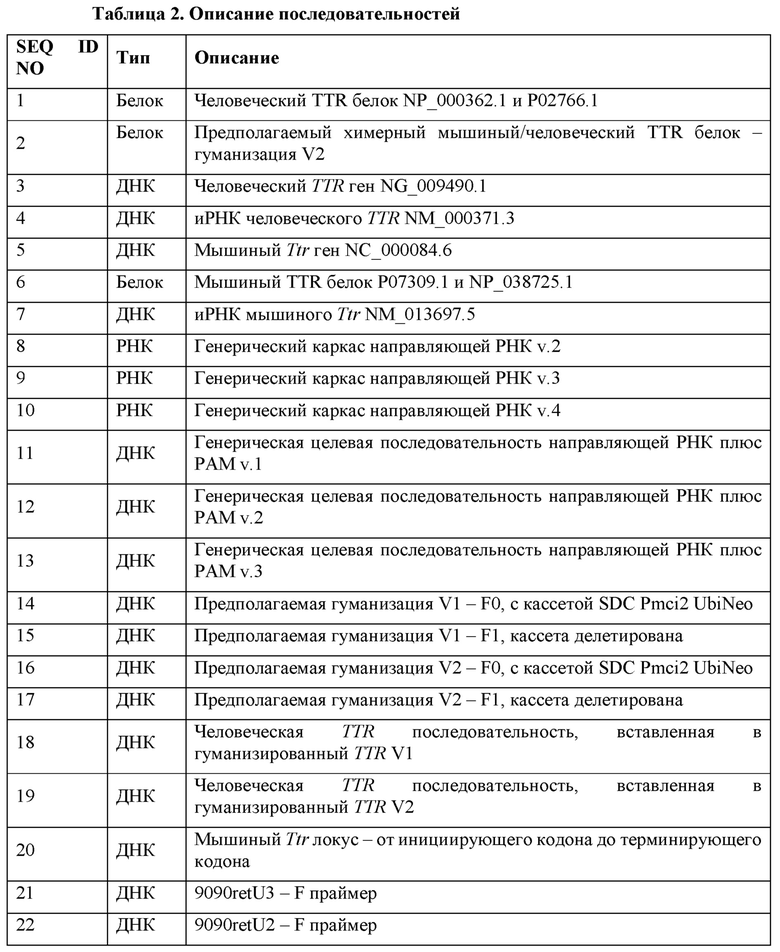
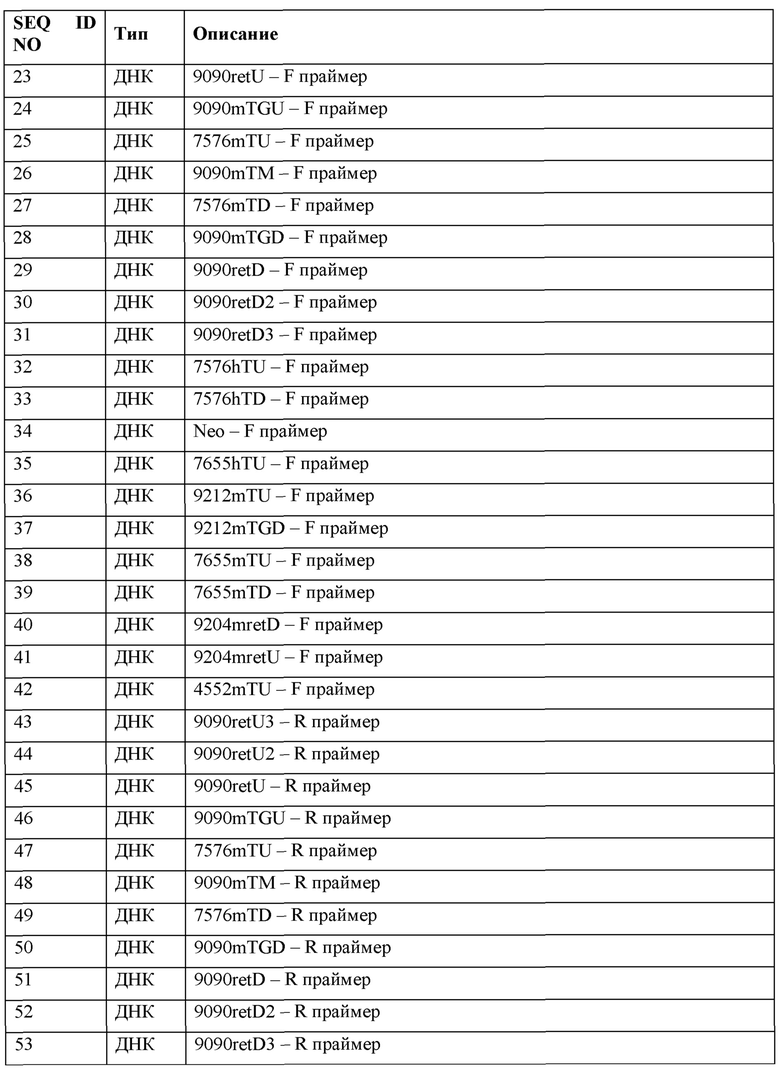
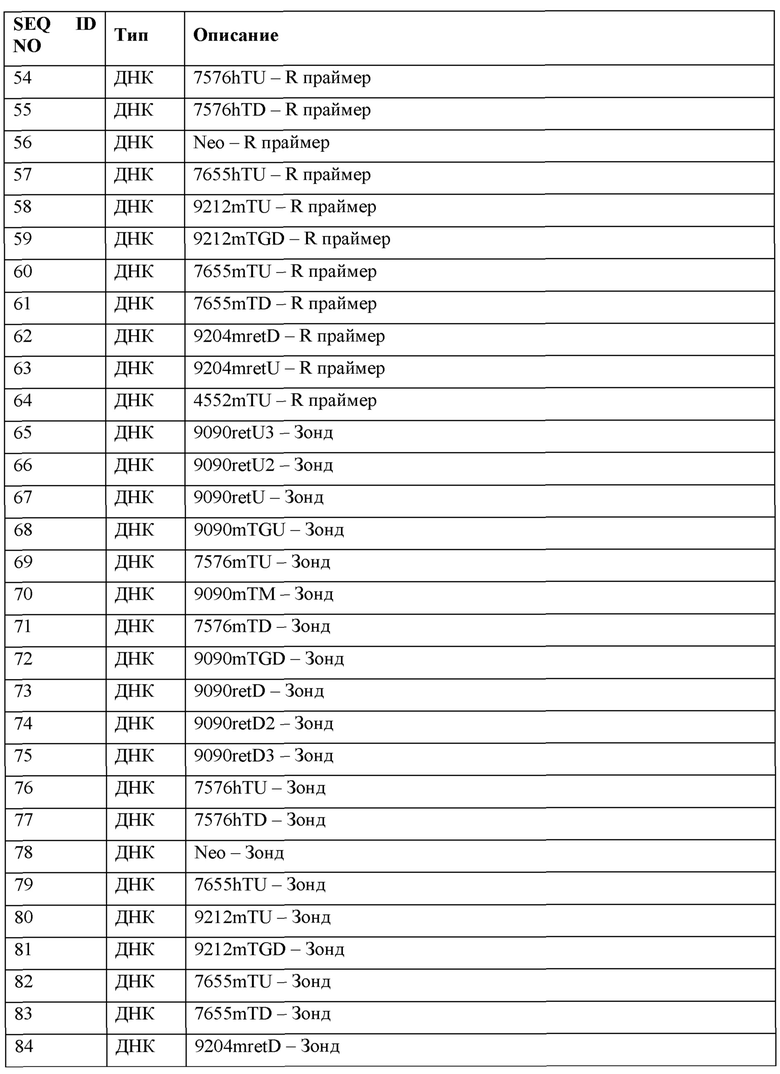

ПРИМЕРЫ
Пример 1. Создание мышей, включающих в себя гуманизированный TTR локус
[00300] Одним из многообещающих терапевтических подходов при заболеваниях TTR амилоидозом является снижение нагрузки TTR у больного путем инактивации гена с помощью технологии редактирования генома, такой как технология CRISPR/Cas9. Для помощи в разработке терапевтических средств на основе CRISPR/Cas9 создавали мышей с целевыми модификациями в Ttr гене.
[00301] Первый созданный Ttr аллель представлял собой полную делецию кодирующей мышиный транстиретин последовательности и ее замену ортологической частью человеческого TTR гена. Создавали большой нацеливающийся вектор, включающий в себя 5' плечо гомологии, содержащее 33,7 т.п.н. последовательности в 3'-5' направлении от инициирующего кодона мышиного Ttr до 34,5 т.п.н. последовательности в 5'-3' направлении терминирующего кодона мышиного Ttr, для замены приблизительно области 8,3 т.п.н. от инициирующего кодона мышиного Ttr до терминирующего кодона мышиного Ttr ортологической человеческой TTR последовательностью приблизительно 7,1 т.п.н. от инициирующего кодона человеческого TTR до конца последнего экзона человеческого TTR (экзона 4, включающего в себя человеческую 3' UTR), и самоудаляющуюся кассету селекции неомицином (SDC Neo), фланкированную сайтами loxP. См. фиг. 3. Кассета SDC Neo включает в себя следующие компоненты от 5' до 3': сайт loxP, мышиный протаминовый (Prml) промотор, Crei (кодирующую последовательность Cre, оптимизированную с включением интрона), полиА, человеческий убиквитиновый промотор, кодирующую последовательность неомицин-фосфотрансферазы (neor), полиА, loxP. Для создания гуманизированного аллеля компоненты CRISPR/Cas9, нацеливающиеся на мышиный Ttr локус, вводили в мышиные эмбриональные стволовые клетки F1H4 вместе с большим нацеливающимся вектором. Анализы на потерю аллеля, анализы на приобретение аллеля, анализы удерживания и анализы CRISPR с использованием праймеров и зондов, изложенных на фиг. 5А и в таблице 3, выполняли для подтверждения гуманизации мышиного Ttr аллеля. Анализы на потерю аллеля, на приобретение аллеля и анализы удерживания описаны, например, в US 2014/0178879; US 2016/0145646; WO 2016/081923; и Frendewey et al. (2010) Methods Enzymol. 476:295-307, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей. Анализы CRISPR представляют собой анализы TAQMAN®, разработанные для схватывания области, которая разрушается с помощью gRNA CRISPR. При разрезании gRNA CRISPR и создании инсерционно-делеционной мутации (вставки или делеции) в анализе TAQMAN® не будет происходить амплификация, что таким образом подтвердит расщепление CRISPR. Версии с кассетой SDC Neo и после удаления кассеты SDC Neo показаны на фиг. 3. Затем создавали мышей F0 с использованием способа VELOCIMOUSE®. См., например, US 7,576,259; US 7,659,442; US 7,294,754; US 2008/007800 и Poueymirou et al. (2007) Nature Biotech. 25(1):91-99, каждая из которых включена в настоящий документ посредством ссылки в полном своем объеме для всех целей.
[00302] Мышей поколения F0 (50% C57BL/6NTac и 50% 129S6/SvEvTac) создавали из нескольких клонов гуманизированных ES клеток, в том числе клонов 7576А-А5, 7576C-G7 и 7576B-F10. Последовательность для предполагаемого гуманизированного мышиного Ttr локуса у мышей поколения F0 приведена в SEQ ID NO: 14 и включает в себя кассету SDC Neo. Затем создавали мышей поколения F1 и F2 (75% C57BL/6NTac и 25% 129S6/SvEvTac) путем скрещивания. Последовательность для предполагаемого гуманизированного мышиного Ttr локуса у мышей поколения F1 и F2 приведена в SEQ ID NO: 15 и не включает в себя кассету SDC Neo.
[00303] Сравнение последовательностей человеческого и мышиного белка-предшественника транстиретина показано на фиг. 1А, сравнение кодирующих последовательностей человеческого и мышиного транстиретина показано на фиг. 1В, а схематическое представление мышиного Ttr локуса дикого типа и конечного гуманизированного мышиного Ttr локуса (гуманизированная TTR версия 1 с делетированной кассетой SDC Neo) показано на фиг. 2. Эндогенная последовательность мышиного Ttr локуса от инициирующего ко дона до терминирующего ко дона представлена под SEQ ID NO: 20. Последовательности для предполагаемого гуманизированного мышиного Ttr локуса с кассетой SDC Neo и без кассеты SDC Neo приведены в SEQ ID NO: 14 и 15, соответственно. Предполагаемый белок-предшественник транстиретина, кодируемый гуманизированным мышиным Ttr локусом, приведен в SEQ ID NO: 1. Этот аллель обеспечивает истинную человеческую мишень для терапевтических средств на основе CRISPR/Cas9 человеческого TTR, тем самым обеспечивая возможность тестирования эффективности и механизма действия терапевтических средств на основе CRISPR/Cas9 на живых животных, а также фармакокинетических и фармакодинамических исследований в условиях, когда белок человека является единственной версией имеющегося TTR.
[00304]

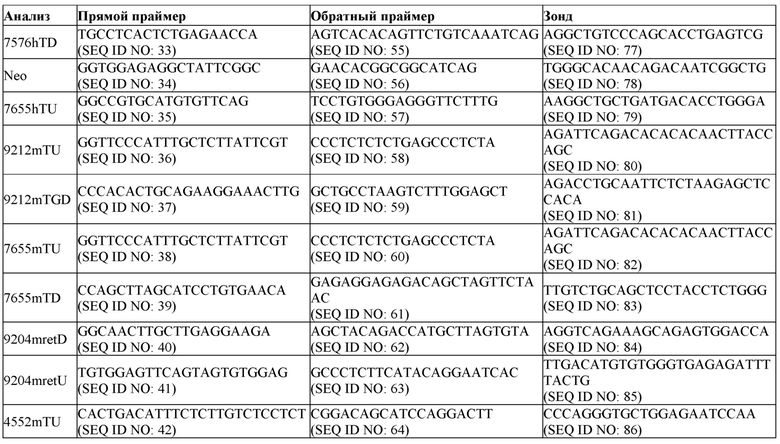
Пример 2. Характеристика мышей, включающих в себя гуманизированный TTR локус
[00305] Затем характеризовали когорту мышей с гуманизированным TTR F0 из клонов 7576А-А5 и 7576C-G7. Как показано на фиг. 6, иРНК гуманизированного TTR надежно экспрессировали в печени 8-недельных, гомозиготных мышей поколения F0 с гуманизированным TTR. Равные массовые количества РНК из каждой ткани анализировали с помощью RT-qPCR. Пять мышей анализировали на предмет генотипа с четырьмя техническими повторностями на образец. Из каждой ткани извлекали РНК. Геномную ДНК разлагали таким образом, чтобы она не учитывалась в qPCR-реакции. РНК подвергали обратной транскрипции, и анализы, специфические для человеческого TTR и мышиного Ttr, использовали для выявления человеческих TTR транскриптов и мышиных Ttr транскриптов, соответственно. Как и предполагали, гомозиготная мышь с гуманизированным TTR демонстрировала значительную экспрессию человеческого TTR в печени (значения Ct ниже 30), тогда как мыши WT демонстрировали значения Ct 30 и выше, что свидетельствует об отсутствии экспрессии человеческого TTR. Напротив, мыши дикого типа демонстрировали значительную экспрессию мышиного Ttr в печени, тогда как гомозиготные мыши с гуманизированным TTR демонстрировали значения Ct 30 и выше, что указывает на отсутствие эндогенной экспрессии мышиного Ttr.
[00306] Выполняли анализ ELISA для оценивания содержания человеческого TTR и мышиного TTR белков в сыворотке крове и спинномозговой жидкости из 8-недельных, гомозиготных гуманизированных по TTR мышей поколения F0. Использовали набор ELISA для человеческого TTR (Aviva Systems Biology; № по каталогу OKIA00081; 1:250 разбавление для сыворотки крови; 1:1000 разбавление для CSF) для оценивания содержания человеческого TTR. Использовали набор ELISA для мышиного TTR (Aviva Systems Biology; № по каталогу OKIA00111; 1:4000 разбавление для сыворотки крови; 1:10000 разбавление для CSF) для оценивания содержания мышиного TTR. Человеческую сыворотку крови и человеческую CSF использовали в качестве положительных контролей для человеческого TTR и отрицательных контролей для мышиного TTR, а сыворотку крови F1H4 и CSF F1H4 использовали в качестве отрицательных контролей для человеческого TTR и положительных контролей для мышиного TTR. Как показано на фиг. 7А, человеческий TTR выявляли в сыворотке крови от гуманизированных по TTR мышей, но не в сыворотке крови от мышей дикого типа (F1H4). Как показано на фиг. 7В, мышиный TTR не выявляли в сыворотке крови от гуманизированных по TTR мышей, но выявляли в сыворотке крови мышей дикого типа. Содержание человеческого и мышиного TTR в сыворотке крови дополнительно оценивали у гуманизированных по TTR мышей, полученных из двух отдельных клонов ES клеток с гуманизированным мышиным Ttr. 7576C-G7 и 7576А-А5. Как показано на фиг. 7С, человеческий TTR выявляли в сыворотке крови гуманизированных по TTR мышей, полученных из клона 7576А-А5.
[00307] Экспрессию человеческого TTR белка также оценивали у 8-недельных, гомозиготных мышей с гуманизированным TTR с помощью вестерн-блоттинга на образцах сыворотки крови, образцах печени и образцах почек. Результаты показаны на фиг. 8-9. Образцы сыворотки крови (суммарный объем 5 мкл на лунку) кипятили в буфере Лэммли (включающем в себя SDS и бета-меркаптоэтанол) и разделяли на 4-20% денатурирующем градиентном геле (антитело против TTR: 1:1000; Abcam; ab75815). Мышиный IgG (антитело против мышиного IgG-HRP: 1:7500, Jackson ImmunoResearch) использовали в качестве контроля нагрузки. Тестировали три мыши на группу на предмет гуманизированных мышиных Ttr клонов 7576C-G7 и 7576А-А5. Тестировали пять мышей на группу на предмет контроля в виде мыши дикого типа (F1H4). Мышиную сыворотку крови и человеческую сыворотку крови использовали в качестве отрицательного и положительного контролей, соответственно. Как показано на фиг. 8, человеческий TTR выявляли с помощью вестерн-блоттинга в сыворотке крови от обоих гуманизированных мышиных Ttr клонов.
[00308] Образцы печени и почек (100 мкг суммарного белка на лунку) кипятили в буфере Лэммли (включающем в себя SDS и бета-меркаптоэтанол) и разделяли на 4-20% денатурирующем градиентном геле (антитело против TTR: 1:1000; Abcam; ab75815). GAPDH (антитело против GAPDH: 1:2000, Santa Cruz) использовали в качестве контроля нагрузки. Тестировали три мыши на группу на предмет гуманизированных мышиных Ttr клонов 7576C-G7 и 7576А-А5. Тестировали пять мышей на группу на предмет контроля в виде мыши дикого типа (F1H4). Мышиную сыворотку крови и человеческую сыворотку крови использовали в качестве отрицательного и положительного контролей, соответственно. Как показано на фиг.9, человеческий TTR выявляли с помощью вестерн-блоттинга в сыворотке крови от обеих гомозиготных гуманизированных по TTR мышей, созданных из клона 7576А-А5.
[00309] Таким образом, обнаружили, что мыши TTR HumIn (TTR7576/7576) F0 имеют выявляемое количество циркулирующего hTTR. Кроме того, мыши клона 7576С-А5 имели выявляемые количества hTTR в печени и плазме.
[00310] Авторы настоящего изобретения выдвинули гипотезу о том, что удаление кассеты селекции лекарственным средством неомицином может усиливать секрецию человеческого TTR. Содержание человеческого TTR измеряли в образцах плазмы из неокончательных подчелюстных кровопусканий у 5-недельных мышей, гомозиготных по полностью гуманизированному мышиному Ttr локусу с кассетой селекции неомицином (TTR7576/7576), мышей, гетерозиготных по полностью гуманизированному мышиному Ttr локусу с кассетой селекции неомицином (TTR7576/WT), мышей, гетерозиготных по полностью гуманизированному мышиному Ttr локусу без кассеты селекции неомицином (TTR7577/WT), и мышей дикого типа (F1H4). Содержание человеческого TTR анализировали с помощью набора для ELISA человеческого TTR (hTTR) AssayPro (№ по каталогу ЕР3010-1; 1:40000 разбавление). Содержание мышиного TTR в сыворотке крови анализировали с помощью набора для ELISA мышиного TTR (mTTR) Aviva Systems Biology (№ по каталогу OK1A00111; 1:4000 разбавление). Предварительно определяли, что набор для ELISA человеческого TTR AssayPro является специфическим для выявления человеческого TTR, но не мышиного TTR, а также предварительно определяли, что набор для ELISA мышиного TTR Aviva Systems Biology является специфическим для выявления мышиного TTR, но не человеческого TTR (данные не показаны). Как показано в таблице 4, удаление кассеты селекции неомицином приводило в результате к статистически значимому увеличению содержания человеческого TTR в сыворотке крови. MAID7576 относится к гуманизированному TTR локусу с кассетой селекции неомицином. MAID7577 относится к гуманизированному TTR локусу с удаленной кассетой селекции неомицином. Повышенную экспрессию иРНК человеческого TTR также наблюдали в печени (данные не показаны). Мыши, гетерозиготные по hTTR и mTTR (TTR-WT7576/WT и TTR-WT7577/WT), характеризовались повышенным циркулирующим hTTR, возможно, из-за повышенной стабильности от взаимодействия гетеромеров (например, между видами TTR).
[00311]
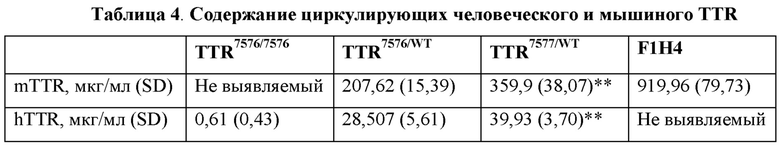
[00312] Также измеряли содержание TTR в сыворотке крови и печени гомозиготных гуманизированных по TTR мышей поколения F2 (по три на группу), которых создавали из другого клона: 7576B-F10. Как показано на фиг. 14 (контрольный образец трис-солевого раствора с сахарозой (TSS)), человеческий TTR выявляли в образцах печени с содержанием более чем 1000 нг/мл. Как показано на фиг. 15А и 15В (образец TSS), человеческий TTR выявляли в образцах сыворотки крови с содержанием 80000 нг/мл или больше.
[00313] В другом эксперименте кровь собирали путем подчелюстных кровопусканий у гомозиготных мышей TTR WT HumIn (v1.0, hTTR7577/7577, клон B-F10) F2 в возрасте 3 месяцев. Определяли содержание hTTR в плазме с использованием специфического в отношении вида набора для ELISA (hTTR, Aviva, № по каталогу OKIA00081; mTTR, Aviva, № по каталогу OKIA00111). Как показано на фиг. 17А и 17В и в таблице 5, hTTR секретировался в кровоток у гомозиготных мышей TTR WT HumIn (v1.0, клон B-F10) F2 при 52,1+/- 10,7 мкг/мл, при этом содержание mTTR не выявляли. Содержание иРНК mTTR и hTTR в образцах печени от контрольных мышей дикого типа (F1H4) и мышей WT HumIn (v1.0, hTTR7577/7577, клон B-F10) показано на фиг. 17С.
[00314]

Пример 3. Применение мышей, включающих в себя гуманизированный TTR локус, для тестирования направляющих РНК, нацеливающихся на человеческий TTR, ex vivo и in vivo
[00315] Когорты гуманизированных по TTR мышей поколения F0 затем использовали для тестирования направляющих РНК, нацеливающихся на человеческий TTR, ex vivo и in vivo. В доказательство концепции направляющие РНК человеческого TTR сначала тестировали в первичных гепатоцитах, выделенных их гуманизированных по TTR мышей поколения F0, полученных из клона 7576C-G7. Печени от мышей huTTRhI/hI перфузировали с 50 мл среды для перфузии печени, включающей в себя IX PenStrep, а затем с 50 мл среды для расщепления печени (HBSS, 100 мМ CaCl2, 500 мМ HEPES, коллагеназа). Как только печень оказывалась переваренной, ее помещали в промывочную среду, включающую в себя IX PenStrep и L-глутамин. Печени иссекали для высвобождения из них гепатоцитов путем осторожного встряхивания. После высвобождения клеток их пропускали через сетчатый фильтр 70 мкм и вращали при 50 g в течение 4 минут при 4°С. Дебрис промывали 2 раза промывочным буфером. Затем дебрис повторно суспендировали в 20 мл 38-40% перколла и вращали при 200 g × 10 минут при 4°С. Дебрис промывали 2 раза и повторно суспендировали в среде для высевания (Williams E Media, IX Penstrep, IX L-глутамин, 5% FBS). Клетки высевали при 300000 клеток на лунку в 24-луночные покрытые коллагеном планшеты для тканевых культур. После того как обеспечивали прикрепление клеток в течение 6-18 часов, среду для высевания заменяли средой без FBS. Используемые реагенты показаны в таблице 6.
[00316]
[00317] Липидные наночастицы (LNP) (включающие в себя иРНК Cas9 плюс gRNA человеческого TTR (версии 1 и 2, каждая нацеливающаяся на экзон 2 человеческого TTR)) добавляли к гепатоцитам через 24 часа после выделения. Вкратце, в каждую лунку LNP добавляли при концентрации 500 нг в 500 мкл среды для поддержания гепатоцитов, включающей в себя 3% мышиной сыворотки крови, и инкубировали в течение 5 минут при 37°С. Высеянные клетки промывали и добавляли 500 мкл инкубированных LNP в среде в каждую лунку. Через 48 часов лизаты ДНК получали из клеток и выполняли секвенирование следующего поколения для каждой тестируемой направляющей РНК.
[00318] Как показано на фиг. 10, редактирование в гуманизированном TTR гене наблюдали с обеими направляющими РНК в первичных гепатоцитах, выделенных из гуманизированных по TTR мышей. Направляющая РНК человеческого TTR 1 характеризовалась эффективностью редактирования 20,4%, а направляющая РНК человеческого TTR 2 характеризовалась эффективностью редактирования 7,53%. Подобные результаты наблюдали для нацеливающейся на экзон 3 направляющей РНК человеческого TTR (эффективность редактирования 17,37%; данные не показаны). Эффективность редактирования относится к суммарному количеству вставок или делеций, наблюдаемых в суммарном количестве последовательностей, считываемых в ПЦР-реакции из пула лизированных клеток, как определяли с помощью секвенирования следующего поколения.
[00319] Затем направляющие РНК человеческого TTR 1 и 2 тестировали in vivo у гуманизированных по TTR мышей. Использовали гуманизированных по TTR мышей (TtrhI/hI) поколения F0 из клонов номер 7576А-А5 и 7576C-G7. На три группы животных воздействовали с помощью свежих LNP, как показано в таблице 7.
[00320]

[00321] LNP составляли с направляющими РНК человеческого TTR и иРНК, кодирующими Cas9, с одним NLS и без метки НА. LNP характеризовались отношением азота к фосфату (N/P) 4,5 и содержали катионный липид, холестерин, DSPC и PEG2k-DMG в молярном отношении 45:44:9:2. Катионный липид, используемый в in vitro и in vivo экспериментах с LNP, описываемых в настоящем документе, представлял собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-октадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-(9Z,12Z)-октадека-9,12-диеноатом. Отношение (направляющая РНК) : (иРНК Cas9) в каждой составляло 1:1 по массе. Дополнительные подробности состава LNP представлены в таблице 8.
[00322]

[00323] Мышей взвешивали перед инъекцией и LNP (включающие в себя иРНК Cas9 плюс gRNA человеческого TTR) водили дозой 2 мг/кг с помощью разбавления в трис-солевом растворе с сахарозой так, что доставляемый объем составлял 200 мкл на мышь. Доставку осуществляли внутривенно путем инъекции в хвостовую вену. Через 8-14 суток мышей умерщвляли и собирали сыворотку крови вместе с тканями печени. Ткани перерабатывали в лизаты ДНК, которые затем анализировали с помощью секвенирования следующего поколения (NGS).
[00324] Химический анализ сыворотки крови на предмет печеночных ферментов ALT, AST, триглицеридов, общего холестерина, HDL, LDL, неэстерифицированных жирных кислот (NEFA) и альбумина не показывал статистической разницы между различными группами обработки. См. фиг. 11А - 11Н. Подобные результаты наблюдали для нацеливающегося экзона 3 направляющей РНК человеческого TTR (данные не показаны).
[00325] NGS показывало значительное редактирование в печени по gRNA 1 человеческого TTR (среднее значение 42,8%) и gRNA 2 человеческого TTR (среднее значение 36,0%). См. фиг. 12. Эффективность редактирования относится к суммарному количеству вставок или делеций, наблюдаемых в суммарном количестве последовательностей, считанных в ПЦР-реакции из пула лизированных клеток. Минимальные или статистически незначимые уровни редактирования гена наблюдали в других тканях (данные не показаны).
[00326] Затем направляющую РНК 1 человеческого TTR тестировали in vivo у гомозиготных гуманизированных по TTR мышей поколения F2 из клона номер 7576B-F10. Животных взвешивали перед введением дозы для вычисления дозы, а затем контролировали состояние здоровья в течение 24 часов после введения дозы. Животным вводили дозу внутривенно при 1 мг/кг, 0,3 мг/кг и 0,1 мг/кг с LNP, составленными с направляющей РНК 1 человеческого TTR и иРНК, кодирующей Cas9, как описано выше. Трис-солевого раствора с сахарозой использовали в качестве контроля. Тестировали три мыши на группу. После вскрытия (через 8 суток после введения дозы) собирали печень и кровь (для сыворотки крови) для анализа. Процент редактирования гуманизированного TTR локуса, наблюдаемый в печени, составлял 50,7% при дозе 1 мг/кг LNP, составленной с направляющей РНК 1 человеческого TTR и иРНК, кодирующей Cas9, 13,0% при дозе 0,3 мг/кг и 2,3% при дозе 0,1 мг/кг, при этом менее чем 0,1% редактирование наблюдали у контрольных мышей. Затем измеряли содержание человеческого TTR в печеночном лизате и в сыворотке крови от мышей, получавших дозу. Печени лизировали в RIPA и ингибиторах протеазы при 100 мг/мл. Для оценивания содержания человеческого TTR использовали набор для ELISA человеческого TTR (Aviva Systems Biology; № по каталогу OKIA00081; 1:100 разбавление для печеночных лизатов; 1:5000 или 1:10000 разбавление для сыворотки крови). Как показано на фиг. 14, содержание более чем 1000 нг/мл человеческого TTR измеряли в лизатах печени от контрольных животных, и это содержание было ниже на более чем 50% у животных, получавших дозу 1 мг/кг LNP, составленной с направляющей РНК 1 человеческого TTR и иРНК, кодирующей Cas9. Как показано на фиг. 15А и 15В, человеческий TTR измеряли с содержанием 80000 нг/мл или больше в сыворотке крови от контрольных животных, и содержание человеческого TTR снижалось на 66% у животных, получавших дозу 1 мг/кг LNP, составленной с направляющей РНК 1 человеческого TTR и иРНК, кодирующей Cas9.
[00327] Затем тестировали три разных направляющих РНК человеческого TTR (направляющих РНК 3, 4 и 5 человеческого TTR) in vivo у гомозиготных гуманизированных по TTR мышей. Составы LNP содержали иРНК Cas9 в отношении 1:2 по массе к направляющей РНК. LNP содержали катионный липид (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-октадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-(9Z,12Z)-октадека-9,12-диеноатом), холестерин, DSPC и PEG2k-DMG в молярном отношении 50:38:9:3, соответственно, и характеризовались отношением N:P 6.
[00328] Сначала оценивали редактирование по гуманизированному TTR локусу. Мышей взвешивали перед инъекцией и LNP (включающие в себя иРНК Cas9 плюс gRNA человеческого TTR) водили дозами 1 мг/кг, 0,3 мг/кг и 0,1 мг/кг (n=5 мышей на группу). Доставку осуществляли внутривенно путем инъекции в хвостовую вену. Как описано выше, мышей позже умерщвляли и собирали сыворотку крови вместе с тканями печени. Ткани перерабатывали в лизаты ДНК, которые затем анализировали с помощью секвенирования следующего поколения (NGS). NGS показывало значительное редактирование в печени по каждой gRNA человеческого TTR при всех тестируемых дозах зависимым от дозы образом. См. фиг. 19. Результаты редактирования в печени определяли с использованием праймеров, разработанных для амплификации представляющей интерес области для анализа NGS.
[00329] Затем оценивали содержание TTR в сыворотке крови. Мышей взвешивали перед введением дозы для вычисления дозы. Мышам вводили дозу внутривенно при 1 мг/кг, 0,3 мг/кг и 0,1 мг/кг (n=5 мышей на группу) с LNP, составленными с направляющей РНК 3, 4 или 5 человеческого TTR и иРНК, кодирующей Cas9, как описано выше. Составы LNP содержали иРНК Cas9 в отношении 1:2 по массе к направляющей РНК. LNP содержали катионный липид (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-октадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-(9Z,12Z)-октадека-9,12-диеноатом), холестерин, DSPC и PEG2k-DMG в молярном отношении 50:38:9:3, соответственно, и характеризовались отношением N:P 6. Трис-солевой раствор с сахарозой использовали в качестве контроля. Как описано выше, позже собирали кровь (для сыворотки крови) для анализа. Затем измеряли содержание человеческого TTR в сыворотке крови от получавших дозу мышей. Содержание человеческого TTR в сыворотке крови оценивали, как описано выше. Как показано на фиг. 20, содержание человеческого TTR значительно снижалось у мышей, получавших дозу каждой направляющей РНК при всех дозах зависимым от дозы образом.
Пример 4. Создание мышей, включающих в себя гуманизированный TTR локус, кодирующий химерный мышиный/человеческий TTR белок с мышиной сигнальной последовательностью
[00330] Авторы настоящего изобретения выдвинули гипотезу о том, что мышиная сигнальная последовательность TTR может усиливать секрецию hTTR до более устойчивых уровней. Конструировали плазмиды для гидродинамической доставки (HDD), включающие в себя вставку cDNA для сигнальной последовательности мышиного Ttr (mTtr) + hTTR («m/hTTR»). Конструкции HDD с использованием вектора pRG977 со вставкой cDNA, приведенные в таблице 9, вводили инъекцией с помощью HDD самцам мышей C57/BL6, каждый возрастом 59 суток. ELISA выполняли на полученной подчелюстным кровопусканием крови в день 4 после HDD. Плазму F1H4 и человеческую сыворотку крови включали в ELISA в качестве отрицательных и положительных контролей, соответственно.
[00331]
[00332] Результаты показаны на фиг. 13. HDD у мышей C57/BL6 дикого типа показала, что использование мышиной сигнальной последовательности TTR действительно усиливало секрецию hTTR в плазме по сравнению с человеческой сигнальной последовательностью TTR + hTTR («hTTR»). Это продемонстрировало, что мыши C57/BL6 могут быть использованы для предсказания конструкций TTR, которые будут приводить в результате к стабильной секреции hTTR
[00333] На основании этих результатов создавали второй гуманизированный TTR аллель, включающий в себя делецию области мышиного Ttr локуса от начала второго экзона до терминирующего кодона и ее замену ортологической частью человеческого TTR гена, но также включающий в себя 3' UTR человеческого TTR гена. Большой нацеливающийся вектор, включающий в себя 5' плечо гомологии, включающее в себя 36 т.п.н. последовательности в 3'-5' направлении от второго экзона мышиного Ttr гена и 34,5 т.п.н. последовательности в 5'-3' направлении терминирующего кодона мышиного Ttr, создавали для замены области приблизительно 7,3 т.п.н. от начала второго экзона в мышином Ttr гене до терминирующего кодона мышиного Ttr ортологической последовательностью человеческого TTR приблизительно 6,1 т.п.н. от начала второго экзона в человеческом TTR гене до конца последнего экзона человеческого TTR (экзона 4, включающего в себя человеческую 3' UTR) и самоудаляющейся кассетой селекции неомицином (SDC Neo), фланкированной сайтами loxP. См. фиг. 4. Для создания гуманизированного аллеля компоненты CRISPR/Cas9, нацеливающиеся на мышиный Ttr локус, вводили в мышиные эмбрионные стволовые клетки F1H4 вместе с большим нацеливающимся вектором. Выполняли анализы на потерю аллеля, анализы на приобретение аллеля и анализы удерживания с использованием праймеров и зондов, изложенных на фиг. 5В и в таблице 3, для подтверждения гуманизации мышиного Ttr аллеля. Версии с кассетой SDC Neo и после вырезания кассеты SDC Neo показаны на фиг. 4. Затем создавали мышей F0 с использованием способа VELOCIMOUSE®.
[00334] Сравнение человеческих и мышиных последовательностей белка-предшественника транстиретина показано на фиг. 1А, сравнение человеческих и мышиных кодирующих транстиретин последовательностей показано на фиг. 1В, и схематическое представление мышиного Ttr локуса дикого типа и конечного гуманизированного мышиного Ttr локуса (версия 2 гуманизированного TTR с делегированной кассетой SDC Neo) показано на фиг. 2. Последовательности для предполагаемого гуманизированного мышиного Ttr локуса с кассетой SDC Neo и без кассеты SDC Neo приведены в SEQ ID NO: 16 и 17, соответственно. MAID7655 относится к гуманизированному TTR локусу (содержащему мышиную сигнальную последовательность) с кассетой селекции неомицином. MAID7656 относится к гуманизированному TTR локусу (содержащему мышиную сигнальную последовательность) с удаленной кассетой селекции неомицином. Предполагаемый белок-предшественник транстиретина, кодируемый гуманизированным мышиным Ttr локусом (химерный мышиный/человеческий TTR белок), приведен в SEQ ID NO: 2.
[00335] Затем использовали набор ELISA для человеческого TTR (Aviva Systems Biology; № по каталогу OKIA00081; 1:2000 разбавление) для оценивания содержания человеческого TTR в плазме крови у разных версий гуманизированных по TTR мышей возрастом от 1 до 3 месяцев. Мыши включали в себя контрольную мышь дикого типа и мышей с различными комбинациями аллелей дикого типа, MAID7577, MAID7655 и MAID7656. MAID7577 относится к гуманизированному TTR локусу с удаленной кассетой селекции неомицином. MAID7655 относится к гуманизированному TTR локусу (содержащему мышиную сигнальную последовательность) с кассетой селекции неомицином. MAID7656 относится к гуманизированному TTR локусу (содержащему мышиную сигнальную последовательность) с удаленной кассетой селекции неомицином. Данные представлены на фиг. 16 и в таблице 10. Как показано на фиг. 16, мыши hTTR7577/7577 (клон B-F10) имели ~55 мкг/мл циркулирующего hTTR, что является существенным, но ниже физиологического содержания у мышей дикого типа или в человеческой сыворотке крови. Гуманизированные по TTR мыши с сигнальной последовательностью мышиного TTR (hTTR7655/7655, hTTR7655/7656 и hTTR7656/7656) не имели повышенного содержания секретируемого TTR по сравнению с гуманизированными по TTR мышами с сигнальной последовательностью человеческого TTR (hTTR7577/7577).
[00336]

[00337] Затем использовали набор ELISA для человеческого TTR (Aviva Systems Biology; № по каталогу OKIA00081; 1:2000 разбавление) для оценивания содержания человеческого TTR в плазме крови у разных версий гуманизированных по TTR мышей возрастом от 2 до 3 месяцев в другом эксперименте. Мыши включали в себя контрольную мышь дикого типа (помеченную F1H4) и мышей, гомозиготных по аллелям MAID7577 или MAID7656. MAID7577 относится к гуманизированному TTR локусу с удаленной кассетой селекции неомицином. MAID7656 относится к гуманизированному TTR локусу (содержащему мышиную сигнальную последовательность) с удаленной кассетой селекции неомицином. Данные представлены на фиг. 18 и в таблице 11. Как показано на фиг 18, мыши hTTR7577/7577 (hTTR v1) имели ~55 мкг/мл циркулирующего hTTR, что является существенным, но ниже физиологического содержания у мышей дикого типа или в человеческой сыворотке крови. Гуманизированные по TTR мыши с сигнальной последовательностью мышиного TTR (hTTR7656/7656; hTTRv2) характеризовались повышенным содержанием секретируемого TTR по сравнению с гуманизированными по TTR мышами с сигнальной последовательностью человеческого TTR (hTTR7577/7577).
[00338]
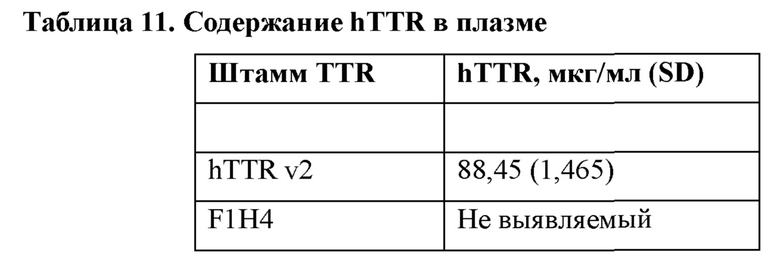
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Редженерон Фармасьютикалс, Инк.
<120> ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ВКЛЮЧАЮЩИЕ В СЕБЯ ГУМАНИЗИРОВАННЫЙ
TTR ЛОКУС, И СПОСОБЫ ПРИМЕНЕНИЯ
<130> 57766-519832
<150> US 62/565,980
<151> 2017-09-29
<150> US 62/679,142
<151> 2018-06-01
<150> US 62/720,292
<151> 2018-08-21
<160> 94
<170> Патентная версия 3.5
<210> 1
<211> 147
<212> PRT
<213> Homo sapiens
<400> 1
Met Ala Ser His Arg Leu Leu Leu Leu Cys Leu Ala Gly Leu Val Phe
1 5 10 15
Val Ser Glu Ala Gly Pro Thr Gly Thr Gly Glu Ser Lys Cys Pro Leu
20 25 30
Met Val Lys Val Leu Asp Ala Val Arg Gly Ser Pro Ala Ile Asn Val
35 40 45
Ala Val His Val Phe Arg Lys Ala Ala Asp Asp Thr Trp Glu Pro Phe
50 55 60
Ala Ser Gly Lys Thr Ser Glu Ser Gly Glu Leu His Gly Leu Thr Thr
65 70 75 80
Glu Glu Glu Phe Val Glu Gly Ile Tyr Lys Val Glu Ile Asp Thr Lys
85 90 95
Ser Tyr Trp Lys Ala Leu Gly Ile Ser Pro Phe His Glu His Ala Glu
100 105 110
Val Val Phe Thr Ala Asn Asp Ser Gly Pro Arg Arg Tyr Thr Ile Ala
115 120 125
Ala Leu Leu Ser Pro Tyr Ser Tyr Ser Thr Thr Ala Val Val Thr Asn
130 135 140
Pro Lys Glu
145
<210> 2
<211> 147
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 2
Met Ala Ser Leu Arg Leu Phe Leu Leu Cys Leu Ala Gly Leu Val Phe
1 5 10 15
Val Ser Glu Ala Gly Pro Ala Gly Thr Gly Glu Ser Lys Cys Pro Leu
20 25 30
Met Val Lys Val Leu Asp Ala Val Arg Gly Ser Pro Ala Ile Asn Val
35 40 45
Ala Val His Val Phe Arg Lys Ala Ala Asp Asp Thr Trp Glu Pro Phe
50 55 60
Ala Ser Gly Lys Thr Ser Glu Ser Gly Glu Leu His Gly Leu Thr Thr
65 70 75 80
Glu Glu Glu Phe Val Glu Gly Ile Tyr Lys Val Glu Ile Asp Thr Lys
85 90 95
Ser Tyr Trp Lys Ala Leu Gly Ile Ser Pro Phe His Glu His Ala Glu
100 105 110
Val Val Phe Thr Ala Asn Asp Ser Gly Pro Arg Arg Tyr Thr Ile Ala
115 120 125
Ala Leu Leu Ser Pro Tyr Ser Tyr Ser Thr Thr Ala Val Val Thr Asn
130 135 140
Pro Lys Glu
145
<210> 3
<211> 7258
<212> ДНК
<213> Homo sapiens
<400> 3
gttgactaag tcaataatca gaatcagcag gtttgcagtc agattggcag ggataagcag 60
cctagctcag gagaagtgag tataaaagcc ccaggctggg agcagccatc acagaagtcc 120
actcattctt ggcaggatgg cttctcatcg tctgctcctc ctctgccttg ctggactggt 180
atttgtgtct gaggctggcc ctacggtgag tgtttctgtg acatcccatt cctacattta 240
agattcacgc taaatgaagt agaagtgact ccttccagct ttgccaacca gcttttatta 300
ctagggcaag ggtacccagc atctattttt aatataatta attcaaactt caaaaagaat 360
gaagttccac tgagcttact gagctgggac ttgaactctg agcattctac ctcattgctt 420
tggtgcatta ggtttgtaat atctggtacc tctgtttcct cagatagatg atagaaataa 480
agatatgata ttaaggaagc tgttaatact gaattttcag aaaagtatcc ctccataaaa 540
tgtatttggg ggacaaactg caggagatta tattctggcc ctatagttat tcaaaacgta 600
tttattgatt aatctttaaa aggcttagtg aacaatattc tagtcagata tctaattctt 660
aaatcctcta gaagaattaa ctaatactat aaaatgggtc tggatgtagt tctgacatta 720
ttttataaca actggtaaga gggagtgact atagcaacaa ctaaaatgat ctcaggaaaa 780
cctgtttggc cctatgtatg gtacattaca tcttttcagt aattccactc aaatggagac 840
ttttaacaaa gcaactgttc tcaggggacc tattttctcc cttaaaattc attatacaca 900
tccctggttg atagcagtgt gtctggaggc agaaaccatt cttgctttgg aaacaattac 960
gtctgtgtta tactgagtag ggaagctcat taattgtcga cacttacgtt cctgataatg 1020
ggatcagtgt gtaattcttg tttcgctcca gatttctaat accacaaaga ataaatcctt 1080
tcactctgat caattttgtt aacttctcac gtgtcttctc tacacccagg gcaccggtga 1140
atccaagtgt cctctgatgg tcaaagttct agatgctgtc cgaggcagtc ctgccatcaa 1200
tgtggccgtg catgtgttca gaaaggctgc tgatgacacc tgggagccat ttgcctctgg 1260
gtaagttgcc aaagaaccct cccacaggac ttggttttat cttcccgttt gcccctcact 1320
tggtagagag aggctcacat catctgctaa agaatttaca agtagattga aaaacgtagg 1380
cagaggtcaa gtatgccctc tgaaggatgc cctctttttg ttttgcttag ctaggaagtg 1440
accaggaacc tgagcatcat ttaggggcag acagtagaga aaagaaggaa tcagaactcc 1500
tctcctctag ctgtggtttg caaccctttt gggtcacaga acactttatg taggtgatga 1560
aaagtaaaca ttctatgccc agaaaaaatg cacagataca cacacataca aaatcatata 1620
tgtgatttta ggagtttcac agattccctg gtgtccctgg gtaacaccaa agctaagtgt 1680
ccttgtctta gaattttagg aaaaggtata atgtgtatta acccattaac aaaaggaaag 1740
gaattcagaa atattattaa ccaggcatct gtctgtagtt aatatggatc acccaaaacc 1800
caaggctttt gcctaatgaa cactttgggg cacctactgt gtgcaaggct gggggctgtc 1860
aagctcagtt aaaaaaaaaa agatagaaga gatggatcca tgaggcaaag tacagcccca 1920
ggctaatccc acgatcaccc gacttcatgt ccaagagtgg cttctcacct tcattagcca 1980
gttcacaatt ttcatggagt ttttctacct gcactagcaa aaacttcaag gaaaatacat 2040
attaataaat ctaagcaaag tgaccagaag acagagcaat caggagaccc tttgcatcca 2100
gcagaagagg aactgctaag tatttacatc tccacagaga agaatttctg ttgggtttta 2160
attgaacccc aagaaccaca tgattcttca accattattg ggaagatcat tttcttaggt 2220
ctggttttaa ctggcttttt atttgggaat tcatttatgt ttatataaaa tgccaagcat 2280
aacatgaaaa gtggttacag gactattcta agggagagac agaatggaca ccaaaaatat 2340
tccaatgttc ttgtgaatct tttccttgca ccaggacaaa aaaaaaaaga agtgaaaaga 2400
agaaaggagg aggggcataa tcagagtcag taaagacaac tgctattttt atctatcgta 2460
gctgttgcag tcaaatggga agcaatttcc aacattcaac tatggagctg gtacttacat 2520
ggaaatagaa gttgcctagt gtttgttgct ggcaaagagt tatcagagag gttaaatata 2580
taaaagggaa aagagtcaga tacaggttct tcttcctact ttaggttttc cactgtgtgt 2640
gcaaatgata ctccctggtg gtgtgcagat gcctcaaagc tatcctcaca ccacaaggga 2700
gaggagcgag atcctgctgt cctggagaag tgcagagtta gaacagctgt ggccacttgc 2760
atccaatcat caatcttgaa tcacagggac tctttcttaa gtaaacatta tacctggccg 2820
ggcacggtgg ctcacgcctg taatcccagc actttgggat gccaaagtgg gcatatcatc 2880
tgaggtcagg agttcaagac cagcctggcc aacatggcaa aactccgtct ttatgaaaaa 2940
tacaaaaatt agccaggcat ggtggcaggc gcctgtaatc ccagctaatt gggaggctga 3000
ggctggagaa tcccttgaat ctaggaggca gaggttgcag tgagctgaga tcgtgccatt 3060
gcactccagc ctgggtgaca agagtaaaac tctgtctcaa aaaaaaaaaa ttatacctac 3120
attctcttct tatcagagaa aaaaatctac agtgagcttt tcaaaaagtt tttacaaact 3180
ttttgccatt taatttcagt taggagtttt ccctacttct gacttagttg aggggaaatg 3240
ttcataacat gtttataaca tgtttatgtg tgttagttgg tgggggtgta ttactttgcc 3300
atgccatttg tttcctccat gcgtaactta atccagactt tcacacctta taggaaaacc 3360
agtgagtctg gagagctgca tgggctcaca actgaggagg aatttgtaga agggatatac 3420
aaagtggaaa tagacaccaa atcttactgg aaggcacttg gcatctcccc attccatgag 3480
catgcagagg tgagtataca gaccttcgag ggttgttttg gttttggttt ttgcttttgg 3540
cattccagga aatgcacagt tttactcagt gtaccacaga aatgtcctaa ggaaggtgat 3600
gaatgaccaa aggttccctt tcctattata caagaaaaaa ttcacaacac tctgagaagc 3660
aaatttcttt ttgactttga tgaaaatcca cttagtaaca tgacttgaac ttacatgaaa 3720
ctactcatag tctattcatt ccactttata tgaatattga tgtatctgct gttgaaataa 3780
tagtttatga ggcagccctc cagaccccac gtagagtgta tgtaacaaga gatgcaccat 3840
tttatttctc gaaaacccgt aacattcttc attccaaaac acatctggct tctcggaggt 3900
ctggacaagt gattcttggc aacacatacc tatagagaca ataaaatcaa agtaataatg 3960
gcaacacaat agataacatt taccaagcat acaccatgtg gcagacacaa ttataagtgt 4020
tttccatatt taacctactt aatcctcagg aataagccac tgaggtcagt cctattatta 4080
tccccatctt atagatgaag aaaatgaggc accaggaagt caaataactt gtcaaaggtc 4140
acaagactag gaaatacaca agtagaaatg tttacaatta aggcccaggc tgggtttgcc 4200
ctcagttctg ctatgcctcg cattatgccc caggaaactt tttcccttgt gaaagccaag 4260
cttaaaaaaa gaaaagccac atttgtaacg tgctctgttc ccctgcctat ggtgaggatc 4320
ttcaaacagt tatacatgga cccagtcccc ctgccttctc cttaatttct taagtcattt 4380
gaaacagatg gctgtcatgg aaatagaatc cagacatgtt ggtcagagtt aaagatcaac 4440
taattccatc aaaaatagct cggcatgaaa gggaactatt ctctggctta gtcatggatg 4500
agactttcaa ttgctataaa gtggttcctt tattagacaa tgttaccagg gaaacaacag 4560
gggtttgttt gacttctggg gcccacaagt caacaagaga gccccatcta ccaaggagca 4620
tgtccctgac tacccctcag ccagcagcaa gacatggacc ccagtcaggg caggagcagg 4680
gtttcggcgg cgcccagcac aagacattgc ccctagagtc tcagccccta ccctcgagta 4740
atagatctgc ctacctgaga ctgttgtttg cccaagagct gggtctcagc ctgatgggaa 4800
ccatataaaa aggttcactg acatactgcc cacatgttgt tctctttcat tagatcttag 4860
cttccttgtc tgctcttcat tcttgcagta ttcattcaac aaacattaaa aaaaaaaaaa 4920
agcattctat gtgtggaaca ctctgctaga tgctgtggat ttagaaatga aaatacatcc 4980
cgacccttgg aatggaaggg aaaggactga agtaagacag attaagcagg accgtcagcc 5040
cagcttgaag cccagataaa tacggagaac aagagagagc gagtagtgag agatgagtcc 5100
caatgcctca ctttggtgac gggtgcgtgg tgggcttcat gcagcttctt ctgataaatg 5160
cctccttcag aactggtcaa ctctaccttg gccagtgacc caggtggtca tagtagattt 5220
accaagggaa aatggaaact tttattagga gctcttaggc ctcttcactt catggatttt 5280
tttttccttt ttttttgaga tggagttttg ccctgtcacc caggctggaa tgcagtggtg 5340
caatctcagc tcactgcaac ctccgcctcc caggttcaag caattctcct gcctcagcct 5400
cccgagtagc tgggactaca ggtgtgcgcc accacaccag gctaattttt gtattttttg 5460
taaagacagg ttttcaccac gttggccagg ctggtctgaa ctccagacct caggtgattc 5520
acctgtctca gcctcccaaa gtgctgggat tacaggtgtg agccaccgtg cccggctact 5580
tcatggattt ttgattacag attatgcctc ttacaatttt taagaagaat caagtgggct 5640
gaaggtcaat gtcaccataa gacaaaagac atttttatta gttgattcta gggaattggc 5700
cttaagggga gccctttctt cctaagagat tcttaggtga ttctcacttc ctcttgcccc 5760
agtattattt ttgtttttgg tatggctcac tcagatcctt ttttcctcct atccctaagt 5820
aatccgggtt tctttttccc atatttagaa caaaatgtat ttatgcagag tgtgtccaaa 5880
cctcaaccca aggcctgtat acaaaataaa tcaaattaaa cacatcttta ctgtcttcta 5940
cctctttcct gacctcaata tatcccaact tgcctcactc tgagaaccaa ggctgtccca 6000
gcacctgagt cgcagatatt ctactgattt gacagaactg tgtgactatc tggaacagca 6060
ttttgatcca caatttgccc agttacaaag cttaaatgag ctctagtgca tgcatatata 6120
tttcaaaatt ccaccatgat cttccacact ctgtattgta aatagagccc tgtaatgctt 6180
ttacttcgta tttcattgct tgttatacat aaaaatatac ttttcttctt catgttagaa 6240
aatgcaaaga ataggagggt gggggaatct ctgggcttgg agacaggaga cttgccttcc 6300
tactatggtt ccatcagaat gtagactggg acaatacaat aattcaagtc tggtttgctc 6360
atctgtaaat tgggaagaat gtttccagct ccagaatgct aaatctctaa gtctgtggtt 6420
ggcagccact attgcagcag ctcttcaatg actcaatgca gttttgcatt ctccctacct 6480
tttttttcta aaaccaataa aatagataca gcctttaggc tttctgggat ttcccttagt 6540
caagctaggg tcatcctgac tttcggcgtg aatttgcaaa acaagacctg actctgtact 6600
cctgctctaa ggactgtgca tggttccaaa ggcttagctt gccagcatat ttgagctttt 6660
tccttctgtt caaactgttc caaaatataa aagaataaaa ttaattaagt tggcactgga 6720
cttccggtgg tcagtcatgt gtgtcatctg tcacgttttt cgggctctgg tggaaatgga 6780
tctgtctgtc ttctctcata ggtggtattc acagccaacg actccggccc ccgccgctac 6840
accattgccg ccctgctgag cccctactcc tattccacca cggctgtcgt caccaatccc 6900
aaggaatgag ggacttctcc tccagtggac ctgaaggacg agggatggga tttcatgtaa 6960
ccaagagtat tccattttta ctaaagcagt gttttcacct catatgctat gttagaagtc 7020
caggcagaga caataaaaca ttcctgtgaa aggcactttt cattccactt taacttgatt 7080
ttttaaattc ccttattgtc ccttccaaaa aaaagagaat caaaatttta caaagaatca 7140
aaggaattct agaaagtatc tgggcagaac gctaggagag atccaaattt ccattgtctt 7200
gcaagcaaag cacgtattaa atatgatctg cagccattaa aaagacacat tctgtaaa 7258
<210> 4
<211> 938
<212> ДНК
<213> Homo sapiens
<400> 4
gttgactaag tcaataatca gaatcagcag gtttgcagtc agattggcag ggataagcag 60
cctagctcag gagaagtgag tataaaagcc ccaggctggg agcagccatc acagaagtcc 120
actcattctt ggcaggatgg cttctcatcg tctgctcctc ctctgccttg ctggactggt 180
atttgtgtct gaggctggcc ctacgggcac cggtgaatcc aagtgtcctc tgatggtcaa 240
agttctagat gctgtccgag gcagtcctgc catcaatgtg gccgtgcatg tgttcagaaa 300
ggctgctgat gacacctggg agccatttgc ctctgggaaa accagtgagt ctggagagct 360
gcatgggctc acaactgagg aggaatttgt agaagggata tacaaagtgg aaatagacac 420
caaatcttac tggaaggcac ttggcatctc cccattccat gagcatgcag aggtggtatt 480
cacagccaac gactccggcc cccgccgcta caccattgcc gccctgctga gcccctactc 540
ctattccacc acggctgtcg tcaccaatcc caaggaatga gggacttctc ctccagtgga 600
cctgaaggac gagggatggg atttcatgta accaagagta ttccattttt actaaagcag 660
tgttttcacc tcatatgcta tgttagaagt ccaggcagag acaataaaac attcctgtga 720
aaggcacttt tcattccact ttaacttgat tttttaaatt cccttattgt cccttccaaa 780
aaaaagagaa tcaaaatttt acaaagaatc aaaggaattc tagaaagtat ctgggcagaa 840
cgctaggaga gatccaaatt tccattgtct tgcaagcaaa gcacgtatta aatatgatct 900
gcagccatta aaaagacaca ttctgtaaaa aaaaaaaa 938
<210> 5
<211> 9077
<212> ДНК
<213> Mus musculus
<400> 5
ctaatctccc taggcaaggt tcatatttgt gtaggttact tattctcctt ttgttgacta 60
agtcaataat cagaatcagc aggtttggag tcagcttggc agggatcagc agcctgggtt 120
ggaaggaggg ggtataaaag ccccttcacc aggagaagcc gtcacacaga tccacaagct 180
cctgacagga tggcttccct tcgactcttc ctcctttgcc tcgctggact ggtatttgtg 240
tctgaagctg gccccgcggt gagtgatcct gtgagcgatc cagacatggc agttagacct 300
tagataaaga agaagtgcct tcttccagat gtgagaacta gagtactcag actctatatt 360
taccattaga ctccaaagag aagagctgga gtgcctctgg ctcttccttc tattgcttta 420
gcgcattggg tctgtagtgc tcagtctctg gtgtccttag ataataaaga tatgagatta 480
acatagaaat aaagatataa aagggctgga tgtatagttt agtggtccag tgtatgccta 540
gtatgtgaaa agccttctgt tcaacctcta gcaatagaaa aacaagatat attctcggtg 600
gggctgttaa tattgaattc tcataaaatc tttaatatat ttagtatgcc tattatgttg 660
ttatatttta gttctttagc taatcaaaat gcattattga tctttctttg tctttttttg 720
gccaacactc tattccagtc tttgaaaaag tcctttaaaa gagttaatca gtataattaa 780
atgagtcagg aagtatgtga gggttatttt acaaccagag ggaattacta tagcaacagc 840
tgattagaat gatctcaaga aaaagcccat tctgtctttt tgcaccatgc acctttcagt 900
ggctccattc agatggagag gcaaacagag caatggctct cagagggcct attttccctt 960
tgaacattca ttatccatat ccctggtgca cagcagtgca tctgggggca gaaactgttc 1020
ttgctttgga aacaatgctg tctatgtcat actggataaa gaagctcatt aattgtcaac 1080
acttatgtta tcataatggg atcagcatgt acttttggtt ttgttccaga gtctatcacc 1140
ggaaagaaca agccggttta ctctgaccca tttcactgac atttctcttg tctcctctgt 1200
gcccagggtg ctggagaatc caaatgtcct ctgatggtca aagtcctgga tgctgtccga 1260
ggcagccctg ctgtagacgt ggctgtaaaa gtgttcaaaa agacctctga gggatcctgg 1320
gagccctttg cctctgggta agcttgtaga aagcccacca tgggaccggt tccaggttcc 1380
catttgctct tattcgtgtt agattcagac acacacaact taccagctag agggctcaga 1440
gagagggctc aggggcgaag ggcacgtatt gctcttgtaa gagacacagg tttaattcct 1500
agcaccagaa tggcagctca taaccatctg aaactcacag tcttaggaga tctgggtatc 1560
tgacattctc ttctacccac catgtgtgtg gtgcacaaat tcacatgcag gcatcaaatc 1620
ttataaacaa caacaaaaaa ccaacaaacc tggtagcaaa agaagattag aaggttaaac 1680
atatgagccg agagcttttg ttttgttttg ttttgttttg ttttgtttac atttcaaatg 1740
ttatcccctt tctcggtccc cctccccaaa ccctctaccc cattctctcc tccccttctt 1800
ctatgagggt gttccccacc aacccactcc caccttcctg ctctcgaatt cccctatact 1860
gggacatcaa gccttcacag aatcaagggc ctctcctccc attgatgccc gacaatgtca 1920
tcctctgcta cctatgtggc tggagccatg ggtcccttca tgtatcctcc ttggttggtg 1980
gtttagtctc tgggaggtct gggggatctg gttgattgat attattgttc ttcctatgag 2040
attgcaaacc ccttcagctc cttcggtcct ttaactcctc cactggggac cccgagctca 2100
gtccaatggt tggctgtgag catccaccag cagaggcctt tttttttttt tttaacaaag 2160
ctgctttatt atgttgctta gagcatgacc aggaaccaga gcacagtcca agactgaagg 2220
gaggaaaagg gggggagtca ataaccccac tgtttcatag tggtttgcaa cccttttata 2280
tcacagccca ctttaggcaa ataatgaaaa ttatagtctc cagggacaga gaagatggtg 2340
caggaagtga agtgcctgct cagaaaatgg gggcttgaat gtgagttccc agactctgtg 2400
taagatgccc agcatcgaag tgcatgctta taacaccagc ctggaggtag aagcttagaa 2460
acaggggtac cctgaagttg cttgttcacc agtgtccctg aatgggtagg tgcatgtttg 2520
gtgagagacc ctgtctcaaa aatcaaggtg taggataatt gaaaatacct agctttgagc 2580
ttagatcatg caaatgtgta cacacactca cacacaccac acacacaaaa aaatgcagag 2640
acagagagat acagagagac agagagatac agagacagag acagagagaa aaggagaaag 2700
taaaaaacaa ataatttaaa gacccatggc cacaaagagg ctcaaagaca agcacgtata 2760
aaaccataca catgtaattt taggagtttt cagattccct ggtacccgtg ggtgatgcac 2820
aagctttgaa tcccagtctt aaaatcttac gaagaacgtg ttcgtgtgtg ctaatttatt 2880
gatgagagga aaggaattga caaagtgccc ttccggagct tcctgcatta cccagactca 2940
gggttttttt aaatgtacac tcagaacaga gtagctctgt gcaagggtag caaccacgaa 3000
gcttaataag aaacatatcg tgagagatct gcaaggcaaa tctaggggct gaccaatctc 3060
acagtcaccc actagcatgt caacacaact tcccacctgt gctagccact tagcaatttt 3120
gtgttgttct gttttgtttt tgtttttaac aaagcaattt caaagagatt tctaattcat 3180
ctaaacaaac aaaccaaaag gaaaacagca aagacgccct gagcacttag cagagcagct 3240
atgcagttat gactcctggg tggagacttt atatcaggct tcaactgaat acctagaacc 3300
tactagtgct cttcatcaat ccttgggaag gtcattttct tttggtgctg ttttgagttt 3360
ctatttgtta atgtcttcat aattatacac gtgttgagca cagcatgcaa agtgattagg 3420
ggaatctagt tggagtggaa tggataccca aatattcaga ctttcttgtg actcttcttt 3480
cttgtaccca catcaaaaaa aaaaaaaatg gagatgagac atggtcagag tcactaaaac 3540
cagctgctac ttttaattac gtggggagca gtttctaaca ttgccattat tgaactgatg 3600
ctgcctgggt ggaaatggaa atcacttagt atttcttgtt ggcaaagaat tactgaatgg 3660
attaaatttc caaagggaga agtcagttac aagtcttttc tttgtttatt aggctttctg 3720
ctatgataaa ttacactact tccagaagtt acccttaggc catgggacac tggactatca 3780
ctctgctgtc acaagagatt acagagttag tcaaggcagc ttgtgacacc ttcagggact 3840
gtcataaact tccagcaagt cattaatcct gaatgcaata ctgtgtgtgt gtgtctatgt 3900
gtgtttgtat gtctgtgtgt gtcttatgtc tgtgtctctg tgtgtgtgtg tgtttgtgtg 3960
tgtgtgtgta tgtatgcctg tgtgtgtctt atgtctgtgt ttgtgtgtct gtgtgtgtct 4020
tatgtctgtg tttgtatgtc tgtgtgtgtc tgtgtgtgtc ttatgtctgt gtctctgtgt 4080
gtgtgtgtgt gtatgtatgt atgtatgtat gtatgtgtat gtgtttgcat ctctctgtgt 4140
gtctgcgctt atatatttgt gtatgtgttt atgtgttcgc ctttgtgcgt tgttggggat 4200
tgaatccagg ggaatacaaa tgttaagaaa gaacgttacc actaagcttc acctgtaggc 4260
cttaaagctt ttctttcttt taaaaattgt aattaattca ttttcagtca ggatctccac 4320
acctcgtccc tgctgctcta gaactcacta tttaaacaca atcgccctca aacctgcagc 4380
aaccctcccg cctctaccct gcgagcacta gaataataac aggtgacccc acacgcctag 4440
attaagacct ttaaggtaaa cattttacta tattttagtc tcataagaca agatgctaca 4500
ataaagctgt acataaagtt ccctcgaatt tcttgctatt ttaactcaaa cataaggatt 4560
tcctcctttt tgattcaggt aacagaaaaa atacacaggt acatacatgt acacacatga 4620
acacacacgc atcacaacca catatgcgca cgcttgtgtg atctatcatt taccatgcca 4680
ctgaactctt ctttccccat aaattcctct ggacttgtgt gccctccagg aagaccgcgg 4740
agtctggaga gctgcacggg ctcaccacag atgagaagtt tgtagaagga gtgtacagag 4800
tagaactgga caccaaatcg tactggaaga cacttggcat ttccccgttc catgaattcg 4860
cggatgtaag tggacacacc aagttgtttg gattttgttt ttagtctcag gaaattccct 4920
tcgctcttgc tgtacgatgg gcatgagtgg aaagtagatt ccacagccag aatccacagt 4980
gctgggaaag caagccttct gaatttttct aaaactcatt tagcaacatg gcctgaacct 5040
gttcacactg cttatggtca gctaactata tttatgtaaa tattcatttc tctgttgagg 5100
aaatgttagt atttgctttt gaggcaacct ccagatacca tggagggcat gtcatagtca 5160
aagagagggc tccctatggt atttctctaa attctggcat ttcctttatt ccaaagcaca 5220
tctagtgtcc ccagaagttt gggtagacaa ttcttggcaa cacagagaat tacaacatgt 5280
tcaaaaccca acagcttaat atctaaatca tcaagcaaac atcacatggc aaagggattt 5340
ctgaatcaaa actgtttcat ccttatgatc aacctatgga ggtctagcct cgacttacac 5400
ccattttacc aataagctaa gagaagctaa gttcctcatc aaggacacaa ggctagcatg 5460
tgtgagcaag tgacagagtt gccctctatg ttggttagtg tgccttagcc agtgtctcag 5520
taagaaatgg agctaaatca aaacccaagg ccaacagcca aaggcacatg agtaaccttt 5580
gcttggcact gggctcagtt tccctggctc ctctcagtcc tcagttcaca gaggcagctg 5640
tcatgcaaat agaatccaag cttgttggtc agacctggag ataacaaatt ccatcaaaaa 5700
tagctcctca tgtgacctag tttgctgtct gttgctatga tacacaccat gaccgaaaag 5760
caaccctggg gagagaaggg tttatttcat cttacagctt acagttcacc atggaggaaa 5820
gccaggtggg aacctggaag tggaaattga agcagagacc agaaaggaat gctgtttact 5880
ggctggctta gctccttttc ttatacagct taggtctatg tgcccagggg atggtactgc 5940
cgagcatagg ctgagcccgc ctacatcaac cattagtcaa aaaaaggtcc atagacttgc 6000
ctacaggcca atctcatgga ggcaataccc cagtggaggg tccctcttcg caggttactc 6060
tagtttgtgt caagttgaca aaacctaacc acaaagcaca aacagggtct gcccttgtgg 6120
cttagccatg gatgacactc tcagatgatg gtgttaccag acaaaccaga ggggctcacc 6180
aagagtctgc cacctaccaa ggtagtactc tactcctcac tgggcaccaa cacccatatt 6240
agctgggcca gtacaggacc cttgctgttt cctgcatgaa ttgtccatag accctgggtc 6300
tcagcctgcc gggagtacct gtaagtagtc gcctcaaaca cattattcct gttggaagac 6360
ttgtctgatt ctcttttaga actcaatcaa caaacgtttt tattttgttt tggctttttg 6420
gagacaagat ctctcatagg ccagcctgac ttgaatgtag ctgaggatga cctgtgctgc 6480
taatcttctc gcctcttcct cccaagtggt aggataatag gcataagaca ccacagcagt 6540
tttactccat accagggctc tgaacccaga ctttaaacac tctatcaact gattcacatt 6600
cccaccccat cattcaacaa acatttgaaa aataaaaccc ttctgccttg agcactctgc 6660
taaatacagc ctttgagtgc ggagtatttc ctcacaacca gggtccaaga tgaccccatc 6720
atacatacca cggaaaatta ggagatgttt ttaggtctct ttgcttgggg taatttttat 6780
gtgtgtgtgt acacagccct gtgcgtgtgt gtgtgtgtgt gtgtgtgtgt gtacaggcac 6840
acacgtgtat gcatgtagag gctacataaa aaccttaggt gtcattctca ggcactctgt 6900
tcaccccttc acacagcccg aacacacaaa atttgaggca ttagcctgga gctcaccagt 6960
taggctagac tgacttgcca gcagacccca ggctgtctcc atctccccag ctctgggatt 7020
acaaactcta tcataccaga catttttata catattctga gcataaaatt catgtcttca 7080
ggctaacaag tcaagagctt aaatgactga gctctcttac gtggtggatt ttttttaaaa 7140
ctacataata tctttttttt ttttttcact tctggggaag aaacaaatga gcctgagtga 7200
caatgcgaca gaaaagaaat tttgaggagt gtgtgtgtct gtgtgtgtgg tggcacatgc 7260
ctctcatcta atgctagagg ctacagtaga atgctcctga attagtggcc agccaaggcc 7320
aagggctagg gttgtaactc agtggcagag ggcttgccta gcattcgcag gatttgatcc 7380
atagcgctat aaataataat aaataaatac aacagtctaa gatgattctc cctttcattt 7440
atctggatgt tatttttgtg ttagttttac tctgtcatcc aatcattgtt tgccctatat 7500
ttggacattt aaaaaaaatc tttattccaa gtgtgttcaa agctgtatcc aaaacctgtc 7560
caccaaatga gtccaatgac atacatcttc tatattacca tctgttccag atttggctga 7620
ctcccggcac ctgggctgtt gctgcaccca tgtctcagat agtctagtga tttgagaagt 7680
gactagtaat tgcaaaatcc agactttgtc cagaaacttc tatgagctcc aaaactttca 7740
tttacatttc tgccagccac aaaccgcttg tgttgtggag agaaccctgt gatgtcttcc 7800
cacagcatct cagccttgtt tcttccctta aaatattcat cttttcacat tagaacatgc 7860
aaagggacag tgggagcgaa acccctggac tgggacgcac gaagccttcc tttctggtca 7920
ggctctcact gtagaaactt aggccggttt cagcatgcag tctgctggag aatggctcct 7980
gccaacattc caggtctgga agtttgtagt ggagttgttg ataaccactg ttcgccacag 8040
gtcttttgtt tgtgggtgtc agtgtttcta ctctcctgac ttttatctga acccaagaaa 8100
gggaacaata gccttcaagc tctctgtgac tctgatctga ccagggccac ccacactgca 8160
gaaggaaact tgcaaagaga gacctgcaat tctctaagag ctccacacag ctccaaagac 8220
ttaggcagca tattttaatc taattattcg tcccccaacc ccaccccaga ggacagttag 8280
acaataaaag gaagattacc agcttagcat cctgtgaaca ctttgtctgc agctcctacc 8340
tctgggctct gttagaacta gctgtctctc ctctctccta ggtggttttc acagccaacg 8400
actctggcca tcgccactac accatcgcag ccctgctcag cccatactcc tacagcacca 8460
cggctgtcgt cagcaacccc cagaattgag agactcagcc caggaggacc aggatcttgc 8520
caaagcagta gcatcccatt tgtaccaaaa cagtgttctt gctctataaa ccgtgttagc 8580
agctcaggaa gatgccgtga agcattctta ttaaaccacc tgctatttca ttcaaactgt 8640
gtttcttttt tatttcctca tttttctccc ctgctcctaa aacccaaaat cttctaaaga 8700
attctagaag gtatgcgatc aaacttttta aagaaagaaa atactttttg actcatggtt 8760
taaaggcatc ctttccatct tggggaggtc atgggtgctc ctggcaactt gcttgaggaa 8820
gataggtcag aaagcagagt ggaccaaccg ttcaatgttt tacaagcaaa acatacacta 8880
agcatggtct gtagctatta aaagcacaca atctgaaggg ctgtagatgc acagtagtgt 8940
tttcccagag catgttcaaa agccctgggt tcaatcacaa tactgaaaag taggccaaaa 9000
aacattctga aaatgaaata tttgggtttt tttttataac ctttagtgac taaataaaga 9060
caaatctaag agactaa 9077
<210> 6
<211> 147
<212> PRT
<213> Mus musculus
<400> 6
Met Ala Ser Leu Arg Leu Phe Leu Leu Cys Leu Ala Gly Leu Val Phe
1 5 10 15
Val Ser Glu Ala Gly Pro Ala Gly Ala Gly Glu Ser Lys Cys Pro Leu
20 25 30
Met Val Lys Val Leu Asp Ala Val Arg Gly Ser Pro Ala Val Asp Val
35 40 45
Ala Val Lys Val Phe Lys Lys Thr Ser Glu Gly Ser Trp Glu Pro Phe
50 55 60
Ala Ser Gly Lys Thr Ala Glu Ser Gly Glu Leu His Gly Leu Thr Thr
65 70 75 80
Asp Glu Lys Phe Val Glu Gly Val Tyr Arg Val Glu Leu Asp Thr Lys
85 90 95
Ser Tyr Trp Lys Thr Leu Gly Ile Ser Pro Phe His Glu Phe Ala Asp
100 105 110
Val Val Phe Thr Ala Asn Asp Ser Gly His Arg His Tyr Thr Ile Ala
115 120 125
Ala Leu Leu Ser Pro Tyr Ser Tyr Ser Thr Thr Ala Val Val Ser Asn
130 135 140
Pro Gln Asn
145
<210> 7
<211> 1237
<212> ДНК
<213> Mus musculus
<400> 7
ctaatctccc taggcaaggt tcatatttgt gtaggttact tattctcctt ttgttgacta 60
agtcaataat cagaatcagc aggtttggag tcagcttggc agggatcagc agcctgggtt 120
ggaaggaggg ggtataaaag ccccttcacc aggagaagcc gtcacacaga tccacaagct 180
cctgacagga tggcttccct tcgactcttc ctcctttgcc tcgctggact ggtatttgtg 240
tctgaagctg gccccgcggg tgctggagaa tccaaatgtc ctctgatggt caaagtcctg 300
gatgctgtcc gaggcagccc tgctgtagac gtggctgtaa aagtgttcaa aaagacctct 360
gagggatcct gggagccctt tgcctctggg aagaccgcgg agtctggaga gctgcacggg 420
ctcaccacag atgagaagtt tgtagaagga gtgtacagag tagaactgga caccaaatcg 480
tactggaaga cacttggcat ttccccgttc catgaattcg cggatgtggt tttcacagcc 540
aacgactctg gccatcgcca ctacaccatc gcagccctgc tcagcccata ctcctacagc 600
accacggctg tcgtcagcaa cccccagaat tgagagactc agcccaggag gaccaggatc 660
ttgccaaagc agtagcatcc catttgtacc aaaacagtgt tcttgctcta taaaccgtgt 720
tagcagctca ggaagatgcc gtgaagcatt cttattaaac cacctgctat ttcattcaaa 780
ctgtgtttct tttttatttc ctcatttttc tcccctgctc ctaaaaccca aaatcttcta 840
aagaattcta gaaggtatgc gatcaaactt tttaaagaaa gaaaatactt tttgactcat 900
ggtttaaagg catcctttcc atcttgggga ggtcatgggt gctcctggca acttgcttga 960
ggaagatagg tcagaaagca gagtggacca accgttcaat gttttacaag caaaacatac 1020
actaagcatg gtctgtagct attaaaagca cacaatctga agggctgtag atgcacagta 1080
gtgttttccc agagcatgtt caaaagccct gggttcaatc acaatactga aaagtaggcc 1140
aaaaaacatt ctgaaaatga aatatttggg ttttttttta taacctttag tgactaaata 1200
aagacaaatc taagagacta aaaaaaaaaa aaaaaaa 1237
<210> 8
<211> 82
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 8
guuggaacca uucaaaacag cauagcaagu uaaaauaagg cuaguccguu aucaacuuga 60
aaaaguggca ccgagucggu gc 82
<210> 9
<211> 76
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 9
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugc 76
<210> 10
<211> 86
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 10
guuuaagagc uaugcuggaa acagcauagc aaguuuaaau aaggcuaguc cguuaucaac 60
uugaaaaagu ggcaccgagu cggugc 86
<210> 11
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (2)..(21)
<223> n = A, T, C или G
<400> 11
gnnnnnnnnn nnnnnnnnnn ngg 23
<210> 12
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (1)..(21)
<223> n = A, T, C или G
<400> 12
nnnnnnnnnn nnnnnnnnnn ngg 23
<210> 13
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (3)..(23)
<223> n = A, T, C или G
<400> 13
ggnnnnnnnn nnnnnnnnnn nnngg 25
<210> 14
<211> 12208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (1)..(97)
<223> Мышиная последовательность
<220>
<221> прочий признак
<222> (98)..(7298)
<223> Человеческая последовательность
<220>
<221> прочий признак
<222> (7299)..(12108)
<223> Кассета SDC Neo
<220>
<221> прочий признак
<222> (12109)..(12208)
<223> Мышиная последовательность
<400> 14
agcttggcag ggatcagcag cctgggttgg aaggaggggg tataaaagcc ccttcaccag 60
gagaagccgt cacacagatc cacaagctcc tgacaggatg gcttctcatc gtctgctcct 120
cctctgcctt gctggactgg tatttgtgtc tgaggctggc cctacggtga gtgtttctgt 180
gacatcccat tcctacattt aagattcacg ctaaatgaag tagaagtgac tccttccagc 240
tttgccaacc agcttttatt actagggcaa gggtacccag catctatttt taatataatt 300
aattcaaact tcaaaaagaa tgaagttcca ctgagcttac tgagctggga cttgaactct 360
gagcattcta cctcattgct ttggtgcatt aggtttgtaa tatctggtac ctctgtttcc 420
tcagatagat gatagaaata aagatatgat attaaggaag ctgttaatac tgaattttca 480
gaaaagtatc cctccataaa atgtatttgg gggacaaact gcaggagatt atattctggc 540
cctatagtta ttcaaaacgt atttattgat taatctttaa aaggcttagt gaacaatatt 600
ctagtcagat atctaattct taaatcctct agaagaatta actaatacta taaaatgggt 660
ctggatgtag ttctgacatt attttataac aactggtaag agggagtgac tatagcaaca 720
actaaaatga tctcaggaaa acctgtttgg ccctatgtat ggtacattac atcttttcag 780
taattccact caaatggaga cttttaacaa agcaactgtt ctcaggggac ctattttctc 840
ccttaaaatt cattatacac atccctggtt gatagcagtg tgtctggagg cagaaaccat 900
tcttgctttg gaaacaatta cgtctgtgtt atactgagta gggaagctca ttaattgtcg 960
acacttacgt tcctgataat gggatcagtg tgtaattctt gtttcgctcc agatttctaa 1020
taccacaaag aataaatcct ttcactctga tcaattttgt taacttctca cgtgtcttct 1080
ctacacccag ggcaccggtg aatccaagtg tcctctgatg gtcaaagttc tagatgctgt 1140
ccgaggcagt cctgccatca atgtggccgt gcatgtgttc agaaaggctg ctgatgacac 1200
ctgggagcca tttgcctctg ggtaagttgc caaagaaccc tcccacagga cttggtttta 1260
tcttcccgtt tgcccctcac ttggtagaga gaggctcaca tcatctgcta aagaatttac 1320
aagtagattg aaaaacgtag gcagaggtca agtatgccct ctgaaggatg ccctcttttt 1380
gttttgctta gctaggaagt gaccaggaac ctgagcatca tttaggggca gacagtagag 1440
aaaagaagga atcagaactc ctctcctcta gctgtggttt gcaacccttt tgggtcacag 1500
aacactttat gtaggtgatg aaaagtaaac attctatgcc cagaaaaaat gcacagatac 1560
acacacatac aaaatcatat atgtgatttt aggagtttca cagattccct ggtgtccctg 1620
ggtaacacca aagctaagtg tccttgtctt agaattttag gaaaaggtat aatgtgtatt 1680
aacccattaa caaaaggaaa ggaattcaga aatattatta accaggcatc tgtctgtagt 1740
taatatggat cacccaaaac ccaaggcttt tgcctaatga acactttggg gcacctactg 1800
tgtgcaaggc tgggggctgt caagctcagt taaaaaaaaa aagatagaag agatggatcc 1860
atgaggcaaa gtacagcccc aggctaatcc cacgatcacc cgacttcatg tccaagagtg 1920
gcttctcacc ttcattagcc agttcacaat tttcatggag tttttctacc tgcactagca 1980
aaaacttcaa ggaaaataca tattaataaa tctaagcaaa gtgaccagaa gacagagcaa 2040
tcaggagacc ctttgcatcc agcagaagag gaactgctaa gtatttacat ctccacagag 2100
aagaatttct gttgggtttt aattgaaccc caagaaccac atgattcttc aaccattatt 2160
gggaagatca ttttcttagg tctggtttta actggctttt tatttgggaa ttcatttatg 2220
tttatataaa atgccaagca taacatgaaa agtggttaca ggactattct aagggagaga 2280
cagaatggac accaaaaata ttccaatgtt cttgtgaatc ttttccttgc accaggacaa 2340
aaaaaaaaag aagtgaaaag aagaaaggag gaggggcata atcagagtca gtaaagacaa 2400
ctgctatttt tatctatcgt agctgttgca gtcaaatggg aagcaatttc caacattcaa 2460
ctatggagct ggtacttaca tggaaataga agttgcctag tgtttgttgc tggcaaagag 2520
ttatcagaga ggttaaatat ataaaaggga aaagagtcag atacaggttc ttcttcctac 2580
tttaggtttt ccactgtgtg tgcaaatgat actccctggt ggtgtgcaga tgcctcaaag 2640
ctatcctcac accacaaggg agaggagcga gatcctgctg tcctggagaa gtgcagagtt 2700
agaacagctg tggccacttg catccaatca tcaatcttga atcacaggga ctctttctta 2760
agtaaacatt atacctggcc gggcacggtg gctcacgcct gtaatcccag cactttggga 2820
tgccaaagtg ggcatatcat ctgaggtcag gagttcaaga ccagcctggc caacatggca 2880
aaactccgtc tttatgaaaa atacaaaaat tagccaggca tggtggcagg cgcctgtaat 2940
cccagctaat tgggaggctg aggctggaga atcccttgaa tctaggaggc agaggttgca 3000
gtgagctgag atcgtgccat tgcactccag cctgggtgac aagagtaaaa ctctgtctca 3060
aaaaaaaaaa attataccta cattctcttc ttatcagaga aaaaaatcta cagtgagctt 3120
ttcaaaaagt ttttacaaac tttttgccat ttaatttcag ttaggagttt tccctacttc 3180
tgacttagtt gaggggaaat gttcataaca tgtttataac atgtttatgt gtgttagttg 3240
gtgggggtgt attactttgc catgccattt gtttcctcca tgcgtaactt aatccagact 3300
ttcacacctt ataggaaaac cagtgagtct ggagagctgc atgggctcac aactgaggag 3360
gaatttgtag aagggatata caaagtggaa atagacacca aatcttactg gaaggcactt 3420
ggcatctccc cattccatga gcatgcagag gtgagtatac agaccttcga gggttgtttt 3480
ggttttggtt tttgcttttg gcattccagg aaatgcacag ttttactcag tgtaccacag 3540
aaatgtccta aggaaggtga tgaatgacca aaggttccct ttcctattat acaagaaaaa 3600
attcacaaca ctctgagaag caaatttctt tttgactttg atgaaaatcc acttagtaac 3660
atgacttgaa cttacatgaa actactcata gtctattcat tccactttat atgaatattg 3720
atgtatctgc tgttgaaata atagtttatg aggcagccct ccagacccca cgtagagtgt 3780
atgtaacaag agatgcacca ttttatttct cgaaaacccg taacattctt cattccaaaa 3840
cacatctggc ttctcggagg tctggacaag tgattcttgg caacacatac ctatagagac 3900
aataaaatca aagtaataat ggcaacacaa tagataacat ttaccaagca tacaccatgt 3960
ggcagacaca attataagtg ttttccatat ttaacctact taatcctcag gaataagcca 4020
ctgaggtcag tcctattatt atccccatct tatagatgaa gaaaatgagg caccaggaag 4080
tcaaataact tgtcaaaggt cacaagacta ggaaatacac aagtagaaat gtttacaatt 4140
aaggcccagg ctgggtttgc cctcagttct gctatgcctc gcattatgcc ccaggaaact 4200
ttttcccttg tgaaagccaa gcttaaaaaa agaaaagcca catttgtaac gtgctctgtt 4260
cccctgccta tggtgaggat cttcaaacag ttatacatgg acccagtccc cctgccttct 4320
ccttaatttc ttaagtcatt tgaaacagat ggctgtcatg gaaatagaat ccagacatgt 4380
tggtcagagt taaagatcaa ctaattccat caaaaatagc tcggcatgaa agggaactat 4440
tctctggctt agtcatggat gagactttca attgctataa agtggttcct ttattagaca 4500
atgttaccag ggaaacaaca ggggtttgtt tgacttctgg ggcccacaag tcaacaagag 4560
agccccatct accaaggagc atgtccctga ctacccctca gccagcagca agacatggac 4620
cccagtcagg gcaggagcag ggtttcggcg gcgcccagca caagacattg cccctagagt 4680
ctcagcccct accctcgagt aatagatctg cctacctgag actgttgttt gcccaagagc 4740
tgggtctcag cctgatggga accatataaa aaggttcact gacatactgc ccacatgttg 4800
ttctctttca ttagatctta gcttccttgt ctgctcttca ttcttgcagt attcattcaa 4860
caaacattaa aaaaaaaaaa aagcattcta tgtgtggaac actctgctag atgctgtgga 4920
tttagaaatg aaaatacatc ccgacccttg gaatggaagg gaaaggactg aagtaagaca 4980
gattaagcag gaccgtcagc ccagcttgaa gcccagataa atacggagaa caagagagag 5040
cgagtagtga gagatgagtc ccaatgcctc actttggtga cgggtgcgtg gtgggcttca 5100
tgcagcttct tctgataaat gcctccttca gaactggtca actctacctt ggccagtgac 5160
ccaggtggtc atagtagatt taccaaggga aaatggaaac ttttattagg agctcttagg 5220
cctcttcact tcatggattt ttttttcctt tttttttgag atggagtttt gccctgtcac 5280
ccaggctgga atgcagtggt gcaatctcag ctcactgcaa cctccgcctc ccaggttcaa 5340
gcaattctcc tgcctcagcc tcccgagtag ctgggactac aggtgtgcgc caccacacca 5400
ggctaatttt tgtatttttt gtaaagacag gttttcacca cgttggccag gctggtctga 5460
actccagacc tcaggtgatt cacctgtctc agcctcccaa agtgctggga ttacaggtgt 5520
gagccaccgt gcccggctac ttcatggatt tttgattaca gattatgcct cttacaattt 5580
ttaagaagaa tcaagtgggc tgaaggtcaa tgtcaccata agacaaaaga catttttatt 5640
agttgattct agggaattgg ccttaagggg agccctttct tcctaagaga ttcttaggtg 5700
attctcactt cctcttgccc cagtattatt tttgtttttg gtatggctca ctcagatcct 5760
tttttcctcc tatccctaag taatccgggt ttctttttcc catatttaga acaaaatgta 5820
tttatgcaga gtgtgtccaa acctcaaccc aaggcctgta tacaaaataa atcaaattaa 5880
acacatcttt actgtcttct acctctttcc tgacctcaat atatcccaac ttgcctcact 5940
ctgagaacca aggctgtccc agcacctgag tcgcagatat tctactgatt tgacagaact 6000
gtgtgactat ctggaacagc attttgatcc acaatttgcc cagttacaaa gcttaaatga 6060
gctctagtgc atgcatatat atttcaaaat tccaccatga tcttccacac tctgtattgt 6120
aaatagagcc ctgtaatgct tttacttcgt atttcattgc ttgttataca taaaaatata 6180
cttttcttct tcatgttaga aaatgcaaag aataggaggg tgggggaatc tctgggcttg 6240
gagacaggag acttgccttc ctactatggt tccatcagaa tgtagactgg gacaatacaa 6300
taattcaagt ctggtttgct catctgtaaa ttgggaagaa tgtttccagc tccagaatgc 6360
taaatctcta agtctgtggt tggcagccac tattgcagca gctcttcaat gactcaatgc 6420
agttttgcat tctccctacc ttttttttct aaaaccaata aaatagatac agcctttagg 6480
ctttctggga tttcccttag tcaagctagg gtcatcctga ctttcggcgt gaatttgcaa 6540
aacaagacct gactctgtac tcctgctcta aggactgtgc atggttccaa aggcttagct 6600
tgccagcata tttgagcttt ttccttctgt tcaaactgtt ccaaaatata aaagaataaa 6660
attaattaag ttggcactgg acttccggtg gtcagtcatg tgtgtcatct gtcacgtttt 6720
tcgggctctg gtggaaatgg atctgtctgt cttctctcat aggtggtatt cacagccaac 6780
gactccggcc cccgccgcta caccattgcc gccctgctga gcccctactc ctattccacc 6840
acggctgtcg tcaccaatcc caaggaatga gggacttctc ctccagtgga cctgaaggac 6900
gagggatggg atttcatgta accaagagta ttccattttt actaaagcag tgttttcacc 6960
tcatatgcta tgttagaagt ccaggcagag acaataaaac attcctgtga aaggcacttt 7020
tcattccact ttaacttgat tttttaaatt cccttattgt cccttccaaa aaaaagagaa 7080
tcaaaatttt acaaagaatc aaaggaattc tagaaagtat ctgggcagaa cgctaggaga 7140
gatccaaatt tccattgtct tgcaagcaaa gcacgtatta aatatgatct gcagccatta 7200
aaaagacaca ttctgtaaat gagagagcct tattttcctg taaccttcag caaatagcaa 7260
aagacacatt ccaagggccc acttctttac tgtgggcact cgagataact tcgtataatg 7320
tatgctatac gaagttatat gcatgccagt agcagcaccc acgtccacct tctgtctagt 7380
aatgtccaac acctccctca gtccaaacac tgctctgcat ccatgtggct cccatttata 7440
cctgaagcac ttgatggggc ctcaatgttt tactagagcc cacccccctg caactctgag 7500
accctctgga tttgtctgtc agtgcctcac tggggcgttg gataatttct taaaaggtca 7560
agttccctca gcagcattct ctgagcagtc tgaagatgtg tgcttttcac agttcaaatc 7620
catgtggctg tttcacccac ctgcctggcc ttgggttatc tatcaggacc tagcctagaa 7680
gcaggtgtgt ggcacttaac acctaagctg agtgactaac tgaacactca agtggatgcc 7740
atctttgtca cttcttgact gtgacacaag caactcctga tgccaaagcc ctgcccaccc 7800
ctctcatgcc catatttgga catggtacag gtcctcactg gccatggtct gtgaggtcct 7860
ggtcctcttt gacttcataa ttcctagggg ccactagtat ctataagagg aagagggtgc 7920
tggctcccag gccacagccc acaaaattcc acctgctcac aggttggctg gctcgaccca 7980
ggtggtgtcc cctgctctga gccagctccc ggccaagcca gcaccatggg aacccccaag 8040
aagaagagga aggtgcgtac cgatttaaat tccaatttac tgaccgtaca ccaaaatttg 8100
cctgcattac cggtcgatgc aacgagtgat gaggttcgca agaacctgat ggacatgttc 8160
agggatcgcc aggcgttttc tgagcatacc tggaaaatgc ttctgtccgt ttgccggtcg 8220
tgggcggcat ggtgcaagtt gaataaccgg aaatggtttc ccgcagaacc tgaagatgtt 8280
cgcgattatc ttctatatct tcaggcgcgc ggtctggcag taaaaactat ccagcaacat 8340
ttgggccagc taaacatgct tcatcgtcgg tccgggctgc cacgaccaag tgacagcaat 8400
gctgtttcac tggttatgcg gcggatccga aaagaaaacg ttgatgccgg tgaacgtgca 8460
aaacaggtaa atataaaatt tttaagtgta taatgatgtt aaactactga ttctaattgt 8520
ttgtgtattt taggctctag cgttcgaacg cactgatttc gaccaggttc gttcactcat 8580
ggaaaatagc gatcgctgcc aggatatacg taatctggca tttctgggga ttgcttataa 8640
caccctgtta cgtatagccg aaattgccag gatcagggtt aaagatatct cacgtactga 8700
cggtgggaga atgttaatcc atattggcag aacgaaaacg ctggttagca ccgcaggtgt 8760
agagaaggca cttagcctgg gggtaactaa actggtcgag cgatggattt ccgtctctgg 8820
tgtagctgat gatccgaata actacctgtt ttgccgggtc agaaaaaatg gtgttgccgc 8880
gccatctgcc accagccagc tatcaactcg cgccctggaa gggatttttg aagcaactca 8940
tcgattgatt tacggcgcta aggatgactc tggtcagaga tacctggcct ggtctggaca 9000
cagtgcccgt gtcggagccg cgcgagatat ggcccgcgct ggagtttcaa taccggagat 9060
catgcaagct ggtggctgga ccaatgtaaa tattgtcatg aactatatcc gtaacctgga 9120
tagtgaaaca ggggcaatgg tgcgcctgct ggaagatggc gattaggcgg ccggccgcta 9180
atcagccata ccacatttgt agaggtttta cttgctttaa aaaacctccc acacctcccc 9240
ctgaacctga aacataaaat gaatgcaatt gttgttgtta acttgtttat tgcagcttat 9300
aatggttaca aataaagcaa tagcatcaca aatttcacaa ataaagcatt tttttcactg 9360
cattctagtt gtggtttgtc caaactcatc aatgtatctt atcatgtctg gatcccccgg 9420
ctagagttta aacactagaa ctagtggatc ccccgggatc atggcctccg cgccgggttt 9480
tggcgcctcc cgcgggcgcc cccctcctca cggcgagcgc tgccacgtca gacgaagggc 9540
gcagcgagcg tcctgatcct tccgcccgga cgctcaggac agcggcccgc tgctcataag 9600
actcggcctt agaaccccag tatcagcaga aggacatttt aggacgggac ttgggtgact 9660
ctagggcact ggttttcttt ccagagagcg gaacaggcga ggaaaagtag tcccttctcg 9720
gcgattctgc ggagggatct ccgtggggcg gtgaacgccg atgattatat aaggacgcgc 9780
cgggtgtggc acagctagtt ccgtcgcagc cgggatttgg gtcgcggttc ttgtttgtgg 9840
atcgctgtga tcgtcacttg gtgagtagcg ggctgctggg ctggccgggg ctttcgtggc 9900
cgccgggccg ctcggtggga cggaagcgtg tggagagacc gccaagggct gtagtctggg 9960
tccgcgagca aggttgccct gaactggggg ttggggggag cgcagcaaaa tggcggctgt 10020
tcccgagtct tgaatggaag acgcttgtga ggcgggctgt gaggtcgttg aaacaaggtg 10080
gggggcatgg tgggcggcaa gaacccaagg tcttgaggcc ttcgctaatg cgggaaagct 10140
cttattcggg tgagatgggc tggggcacca tctggggacc ctgacgtgaa gtttgtcact 10200
gactggagaa ctcggtttgt cgtctgttgc gggggcggca gttatggcgg tgccgttggg 10260
cagtgcaccc gtacctttgg gagcgcgcgc cctcgtcgtg tcgtgacgtc acccgttctg 10320
ttggcttata atgcagggtg gggccacctg ccggtaggtg tgcggtaggc ttttctccgt 10380
cgcaggacgc agggttcggg cctagggtag gctctcctga atcgacaggc gccggacctc 10440
tggtgagggg agggataagt gaggcgtcag tttctttggt cggttttatg tacctatctt 10500
cttaagtagc tgaagctccg gttttgaact atgcgctcgg ggttggcgag tgtgttttgt 10560
gaagtttttt aggcaccttt tgaaatgtaa tcatttgggt caatatgtaa ttttcagtgt 10620
tagactagta aattgtccgc taaattctgg ccgtttttgg cttttttgtt agacgtgttg 10680
acaattaatc atcggcatag tatatcggca tagtataata cgacaaggtg aggaactaaa 10740
ccatgggatc ggccattgaa caagatggat tgcacgcagg ttctccggcc gcttgggtgg 10800
agaggctatt cggctatgac tgggcacaac agacaatcgg ctgctctgat gccgccgtgt 10860
tccggctgtc agcgcagggg cgcccggttc tttttgtcaa gaccgacctg tccggtgccc 10920
tgaatgaact gcaggacgag gcagcgcggc tatcgtggct ggccacgacg ggcgttcctt 10980
gcgcagctgt gctcgacgtt gtcactgaag cgggaaggga ctggctgcta ttgggcgaag 11040
tgccggggca ggatctcctg tcatctcacc ttgctcctgc cgagaaagta tccatcatgg 11100
ctgatgcaat gcggcggctg catacgcttg atccggctac ctgcccattc gaccaccaag 11160
cgaaacatcg catcgagcga gcacgtactc ggatggaagc cggtcttgtc gatcaggatg 11220
atctggacga agagcatcag gggctcgcgc cagccgaact gttcgccagg ctcaaggcgc 11280
gcatgcccga cggcgatgat ctcgtcgtga cccatggcga tgcctgcttg ccgaatatca 11340
tggtggaaaa tggccgcttt tctggattca tcgactgtgg ccggctgggt gtggcggacc 11400
gctatcagga catagcgttg gctacccgtg atattgctga agagcttggc ggcgaatggg 11460
ctgaccgctt cctcgtgctt tacggtatcg ccgctcccga ttcgcagcgc atcgccttct 11520
atcgccttct tgacgagttc ttctgagggg atccgctgta agtctgcaga aattgatgat 11580
ctattaaaca ataaagatgt ccactaaaat ggaagttttt cctgtcatac tttgttaaga 11640
agggtgagaa cagagtacct acattttgaa tggaaggatt ggagctacgg gggtgggggt 11700
ggggtgggat tagataaatg cctgctcttt actgaaggct ctttactatt gctttatgat 11760
aatgtttcat agttggatat cataatttaa acaagcaaaa ccaaattaag ggccagctca 11820
ttcctcccac tcatgatcta tagatctata gatctctcgt gggatcattg tttttctctt 11880
gattcccact ttgtggttct aagtactgtg gtttccaaat gtgtcagttt catagcctga 11940
agaacgagat cagcagcctc tgttccacat acacttcatt ctcagtattg ttttgccaag 12000
ttctaattcc atcagacctc gacctgcagc ccctagataa cttcgtataa tgtatgctat 12060
acgaagttat gctaggtaac tataacggtc ctaaggtagc gagctagcga gactcagccc 12120
aggaggacca ggatcttgcc aaagcagtag catcccattt gtaccaaaac agtgttcttg 12180
ctctataaac cgtgttagca gctcagga 12208
<210> 15
<211> 7476
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (1)..(97)
<223> Мышиная последовательность
<220>
<221> прочий признак
<222> (98)..(7298)
<223> Человеческая последовательность
<220>
<221> прочий признак
<222> (7299)..(7376)
<223> Кассета LoxP Scar
<220>
<221> прочий признак
<222> (7377)..(7476)
<223> Мышиная последовательность
<400> 15
agcttggcag ggatcagcag cctgggttgg aaggaggggg tataaaagcc ccttcaccag 60
gagaagccgt cacacagatc cacaagctcc tgacaggatg gcttctcatc gtctgctcct 120
cctctgcctt gctggactgg tatttgtgtc tgaggctggc cctacggtga gtgtttctgt 180
gacatcccat tcctacattt aagattcacg ctaaatgaag tagaagtgac tccttccagc 240
tttgccaacc agcttttatt actagggcaa gggtacccag catctatttt taatataatt 300
aattcaaact tcaaaaagaa tgaagttcca ctgagcttac tgagctggga cttgaactct 360
gagcattcta cctcattgct ttggtgcatt aggtttgtaa tatctggtac ctctgtttcc 420
tcagatagat gatagaaata aagatatgat attaaggaag ctgttaatac tgaattttca 480
gaaaagtatc cctccataaa atgtatttgg gggacaaact gcaggagatt atattctggc 540
cctatagtta ttcaaaacgt atttattgat taatctttaa aaggcttagt gaacaatatt 600
ctagtcagat atctaattct taaatcctct agaagaatta actaatacta taaaatgggt 660
ctggatgtag ttctgacatt attttataac aactggtaag agggagtgac tatagcaaca 720
actaaaatga tctcaggaaa acctgtttgg ccctatgtat ggtacattac atcttttcag 780
taattccact caaatggaga cttttaacaa agcaactgtt ctcaggggac ctattttctc 840
ccttaaaatt cattatacac atccctggtt gatagcagtg tgtctggagg cagaaaccat 900
tcttgctttg gaaacaatta cgtctgtgtt atactgagta gggaagctca ttaattgtcg 960
acacttacgt tcctgataat gggatcagtg tgtaattctt gtttcgctcc agatttctaa 1020
taccacaaag aataaatcct ttcactctga tcaattttgt taacttctca cgtgtcttct 1080
ctacacccag ggcaccggtg aatccaagtg tcctctgatg gtcaaagttc tagatgctgt 1140
ccgaggcagt cctgccatca atgtggccgt gcatgtgttc agaaaggctg ctgatgacac 1200
ctgggagcca tttgcctctg ggtaagttgc caaagaaccc tcccacagga cttggtttta 1260
tcttcccgtt tgcccctcac ttggtagaga gaggctcaca tcatctgcta aagaatttac 1320
aagtagattg aaaaacgtag gcagaggtca agtatgccct ctgaaggatg ccctcttttt 1380
gttttgctta gctaggaagt gaccaggaac ctgagcatca tttaggggca gacagtagag 1440
aaaagaagga atcagaactc ctctcctcta gctgtggttt gcaacccttt tgggtcacag 1500
aacactttat gtaggtgatg aaaagtaaac attctatgcc cagaaaaaat gcacagatac 1560
acacacatac aaaatcatat atgtgatttt aggagtttca cagattccct ggtgtccctg 1620
ggtaacacca aagctaagtg tccttgtctt agaattttag gaaaaggtat aatgtgtatt 1680
aacccattaa caaaaggaaa ggaattcaga aatattatta accaggcatc tgtctgtagt 1740
taatatggat cacccaaaac ccaaggcttt tgcctaatga acactttggg gcacctactg 1800
tgtgcaaggc tgggggctgt caagctcagt taaaaaaaaa aagatagaag agatggatcc 1860
atgaggcaaa gtacagcccc aggctaatcc cacgatcacc cgacttcatg tccaagagtg 1920
gcttctcacc ttcattagcc agttcacaat tttcatggag tttttctacc tgcactagca 1980
aaaacttcaa ggaaaataca tattaataaa tctaagcaaa gtgaccagaa gacagagcaa 2040
tcaggagacc ctttgcatcc agcagaagag gaactgctaa gtatttacat ctccacagag 2100
aagaatttct gttgggtttt aattgaaccc caagaaccac atgattcttc aaccattatt 2160
gggaagatca ttttcttagg tctggtttta actggctttt tatttgggaa ttcatttatg 2220
tttatataaa atgccaagca taacatgaaa agtggttaca ggactattct aagggagaga 2280
cagaatggac accaaaaata ttccaatgtt cttgtgaatc ttttccttgc accaggacaa 2340
aaaaaaaaag aagtgaaaag aagaaaggag gaggggcata atcagagtca gtaaagacaa 2400
ctgctatttt tatctatcgt agctgttgca gtcaaatggg aagcaatttc caacattcaa 2460
ctatggagct ggtacttaca tggaaataga agttgcctag tgtttgttgc tggcaaagag 2520
ttatcagaga ggttaaatat ataaaaggga aaagagtcag atacaggttc ttcttcctac 2580
tttaggtttt ccactgtgtg tgcaaatgat actccctggt ggtgtgcaga tgcctcaaag 2640
ctatcctcac accacaaggg agaggagcga gatcctgctg tcctggagaa gtgcagagtt 2700
agaacagctg tggccacttg catccaatca tcaatcttga atcacaggga ctctttctta 2760
agtaaacatt atacctggcc gggcacggtg gctcacgcct gtaatcccag cactttggga 2820
tgccaaagtg ggcatatcat ctgaggtcag gagttcaaga ccagcctggc caacatggca 2880
aaactccgtc tttatgaaaa atacaaaaat tagccaggca tggtggcagg cgcctgtaat 2940
cccagctaat tgggaggctg aggctggaga atcccttgaa tctaggaggc agaggttgca 3000
gtgagctgag atcgtgccat tgcactccag cctgggtgac aagagtaaaa ctctgtctca 3060
aaaaaaaaaa attataccta cattctcttc ttatcagaga aaaaaatcta cagtgagctt 3120
ttcaaaaagt ttttacaaac tttttgccat ttaatttcag ttaggagttt tccctacttc 3180
tgacttagtt gaggggaaat gttcataaca tgtttataac atgtttatgt gtgttagttg 3240
gtgggggtgt attactttgc catgccattt gtttcctcca tgcgtaactt aatccagact 3300
ttcacacctt ataggaaaac cagtgagtct ggagagctgc atgggctcac aactgaggag 3360
gaatttgtag aagggatata caaagtggaa atagacacca aatcttactg gaaggcactt 3420
ggcatctccc cattccatga gcatgcagag gtgagtatac agaccttcga gggttgtttt 3480
ggttttggtt tttgcttttg gcattccagg aaatgcacag ttttactcag tgtaccacag 3540
aaatgtccta aggaaggtga tgaatgacca aaggttccct ttcctattat acaagaaaaa 3600
attcacaaca ctctgagaag caaatttctt tttgactttg atgaaaatcc acttagtaac 3660
atgacttgaa cttacatgaa actactcata gtctattcat tccactttat atgaatattg 3720
atgtatctgc tgttgaaata atagtttatg aggcagccct ccagacccca cgtagagtgt 3780
atgtaacaag agatgcacca ttttatttct cgaaaacccg taacattctt cattccaaaa 3840
cacatctggc ttctcggagg tctggacaag tgattcttgg caacacatac ctatagagac 3900
aataaaatca aagtaataat ggcaacacaa tagataacat ttaccaagca tacaccatgt 3960
ggcagacaca attataagtg ttttccatat ttaacctact taatcctcag gaataagcca 4020
ctgaggtcag tcctattatt atccccatct tatagatgaa gaaaatgagg caccaggaag 4080
tcaaataact tgtcaaaggt cacaagacta ggaaatacac aagtagaaat gtttacaatt 4140
aaggcccagg ctgggtttgc cctcagttct gctatgcctc gcattatgcc ccaggaaact 4200
ttttcccttg tgaaagccaa gcttaaaaaa agaaaagcca catttgtaac gtgctctgtt 4260
cccctgccta tggtgaggat cttcaaacag ttatacatgg acccagtccc cctgccttct 4320
ccttaatttc ttaagtcatt tgaaacagat ggctgtcatg gaaatagaat ccagacatgt 4380
tggtcagagt taaagatcaa ctaattccat caaaaatagc tcggcatgaa agggaactat 4440
tctctggctt agtcatggat gagactttca attgctataa agtggttcct ttattagaca 4500
atgttaccag ggaaacaaca ggggtttgtt tgacttctgg ggcccacaag tcaacaagag 4560
agccccatct accaaggagc atgtccctga ctacccctca gccagcagca agacatggac 4620
cccagtcagg gcaggagcag ggtttcggcg gcgcccagca caagacattg cccctagagt 4680
ctcagcccct accctcgagt aatagatctg cctacctgag actgttgttt gcccaagagc 4740
tgggtctcag cctgatggga accatataaa aaggttcact gacatactgc ccacatgttg 4800
ttctctttca ttagatctta gcttccttgt ctgctcttca ttcttgcagt attcattcaa 4860
caaacattaa aaaaaaaaaa aagcattcta tgtgtggaac actctgctag atgctgtgga 4920
tttagaaatg aaaatacatc ccgacccttg gaatggaagg gaaaggactg aagtaagaca 4980
gattaagcag gaccgtcagc ccagcttgaa gcccagataa atacggagaa caagagagag 5040
cgagtagtga gagatgagtc ccaatgcctc actttggtga cgggtgcgtg gtgggcttca 5100
tgcagcttct tctgataaat gcctccttca gaactggtca actctacctt ggccagtgac 5160
ccaggtggtc atagtagatt taccaaggga aaatggaaac ttttattagg agctcttagg 5220
cctcttcact tcatggattt ttttttcctt tttttttgag atggagtttt gccctgtcac 5280
ccaggctgga atgcagtggt gcaatctcag ctcactgcaa cctccgcctc ccaggttcaa 5340
gcaattctcc tgcctcagcc tcccgagtag ctgggactac aggtgtgcgc caccacacca 5400
ggctaatttt tgtatttttt gtaaagacag gttttcacca cgttggccag gctggtctga 5460
actccagacc tcaggtgatt cacctgtctc agcctcccaa agtgctggga ttacaggtgt 5520
gagccaccgt gcccggctac ttcatggatt tttgattaca gattatgcct cttacaattt 5580
ttaagaagaa tcaagtgggc tgaaggtcaa tgtcaccata agacaaaaga catttttatt 5640
agttgattct agggaattgg ccttaagggg agccctttct tcctaagaga ttcttaggtg 5700
attctcactt cctcttgccc cagtattatt tttgtttttg gtatggctca ctcagatcct 5760
tttttcctcc tatccctaag taatccgggt ttctttttcc catatttaga acaaaatgta 5820
tttatgcaga gtgtgtccaa acctcaaccc aaggcctgta tacaaaataa atcaaattaa 5880
acacatcttt actgtcttct acctctttcc tgacctcaat atatcccaac ttgcctcact 5940
ctgagaacca aggctgtccc agcacctgag tcgcagatat tctactgatt tgacagaact 6000
gtgtgactat ctggaacagc attttgatcc acaatttgcc cagttacaaa gcttaaatga 6060
gctctagtgc atgcatatat atttcaaaat tccaccatga tcttccacac tctgtattgt 6120
aaatagagcc ctgtaatgct tttacttcgt atttcattgc ttgttataca taaaaatata 6180
cttttcttct tcatgttaga aaatgcaaag aataggaggg tgggggaatc tctgggcttg 6240
gagacaggag acttgccttc ctactatggt tccatcagaa tgtagactgg gacaatacaa 6300
taattcaagt ctggtttgct catctgtaaa ttgggaagaa tgtttccagc tccagaatgc 6360
taaatctcta agtctgtggt tggcagccac tattgcagca gctcttcaat gactcaatgc 6420
agttttgcat tctccctacc ttttttttct aaaaccaata aaatagatac agcctttagg 6480
ctttctggga tttcccttag tcaagctagg gtcatcctga ctttcggcgt gaatttgcaa 6540
aacaagacct gactctgtac tcctgctcta aggactgtgc atggttccaa aggcttagct 6600
tgccagcata tttgagcttt ttccttctgt tcaaactgtt ccaaaatata aaagaataaa 6660
attaattaag ttggcactgg acttccggtg gtcagtcatg tgtgtcatct gtcacgtttt 6720
tcgggctctg gtggaaatgg atctgtctgt cttctctcat aggtggtatt cacagccaac 6780
gactccggcc cccgccgcta caccattgcc gccctgctga gcccctactc ctattccacc 6840
acggctgtcg tcaccaatcc caaggaatga gggacttctc ctccagtgga cctgaaggac 6900
gagggatggg atttcatgta accaagagta ttccattttt actaaagcag tgttttcacc 6960
tcatatgcta tgttagaagt ccaggcagag acaataaaac attcctgtga aaggcacttt 7020
tcattccact ttaacttgat tttttaaatt cccttattgt cccttccaaa aaaaagagaa 7080
tcaaaatttt acaaagaatc aaaggaattc tagaaagtat ctgggcagaa cgctaggaga 7140
gatccaaatt tccattgtct tgcaagcaaa gcacgtatta aatatgatct gcagccatta 7200
aaaagacaca ttctgtaaat gagagagcct tattttcctg taaccttcag caaatagcaa 7260
aagacacatt ccaagggccc acttctttac tgtgggcact cgagataact tcgtataatg 7320
tatgctatac gaagttatgc taggtaacta taacggtcct aaggtagcga gctagcgaga 7380
ctcagcccag gaggaccagg atcttgccaa agcagtagca tcccatttgt accaaaacag 7440
tgttcttgct ctataaaccg tgttagcagc tcagga 7476
<210> 16
<211> 11218
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (1)..(100)
<223> Мышиная последовательность
<220>
<221> прочий признак
<222> (101)..(6308)
<223> Человеческая последовательность
<220>
<221> прочий признак
<222> (6309)..(11118)
<223> Кассета SDC Neo
<220>
<221> прочий признак
<222> (11119)..(11218)
<223> Мышиная последовательность
<400> 16
atgtactttt ggttttgttc cagagtctat caccggaaag aacaagccgg tttactctga 60
cccatttcac tgacatttct cttgtctcct ctgtgcccag ggcaccggtg aatccaagtg 120
tcctctgatg gtcaaagttc tagatgctgt ccgaggcagt cctgccatca atgtggccgt 180
gcatgtgttc agaaaggctg ctgatgacac ctgggagcca tttgcctctg ggtaagttgc 240
caaagaaccc tcccacagga cttggtttta tcttcccgtt tgcccctcac ttggtagaga 300
gaggctcaca tcatctgcta aagaatttac aagtagattg aaaaacgtag gcagaggtca 360
agtatgccct ctgaaggatg ccctcttttt gttttgctta gctaggaagt gaccaggaac 420
ctgagcatca tttaggggca gacagtagag aaaagaagga atcagaactc ctctcctcta 480
gctgtggttt gcaacccttt tgggtcacag aacactttat gtaggtgatg aaaagtaaac 540
attctatgcc cagaaaaaat gcacagatac acacacatac aaaatcatat atgtgatttt 600
aggagtttca cagattccct ggtgtccctg ggtaacacca aagctaagtg tccttgtctt 660
agaattttag gaaaaggtat aatgtgtatt aacccattaa caaaaggaaa ggaattcaga 720
aatattatta accaggcatc tgtctgtagt taatatggat cacccaaaac ccaaggcttt 780
tgcctaatga acactttggg gcacctactg tgtgcaaggc tgggggctgt caagctcagt 840
taaaaaaaaa aagatagaag agatggatcc atgaggcaaa gtacagcccc aggctaatcc 900
cacgatcacc cgacttcatg tccaagagtg gcttctcacc ttcattagcc agttcacaat 960
tttcatggag tttttctacc tgcactagca aaaacttcaa ggaaaataca tattaataaa 1020
tctaagcaaa gtgaccagaa gacagagcaa tcaggagacc ctttgcatcc agcagaagag 1080
gaactgctaa gtatttacat ctccacagag aagaatttct gttgggtttt aattgaaccc 1140
caagaaccac atgattcttc aaccattatt gggaagatca ttttcttagg tctggtttta 1200
actggctttt tatttgggaa ttcatttatg tttatataaa atgccaagca taacatgaaa 1260
agtggttaca ggactattct aagggagaga cagaatggac accaaaaata ttccaatgtt 1320
cttgtgaatc ttttccttgc accaggacaa aaaaaaaaag aagtgaaaag aagaaaggag 1380
gaggggcata atcagagtca gtaaagacaa ctgctatttt tatctatcgt agctgttgca 1440
gtcaaatggg aagcaatttc caacattcaa ctatggagct ggtacttaca tggaaataga 1500
agttgcctag tgtttgttgc tggcaaagag ttatcagaga ggttaaatat ataaaaggga 1560
aaagagtcag atacaggttc ttcttcctac tttaggtttt ccactgtgtg tgcaaatgat 1620
actccctggt ggtgtgcaga tgcctcaaag ctatcctcac accacaaggg agaggagcga 1680
gatcctgctg tcctggagaa gtgcagagtt agaacagctg tggccacttg catccaatca 1740
tcaatcttga atcacaggga ctctttctta agtaaacatt atacctggcc gggcacggtg 1800
gctcacgcct gtaatcccag cactttggga tgccaaagtg ggcatatcat ctgaggtcag 1860
gagttcaaga ccagcctggc caacatggca aaactccgtc tttatgaaaa atacaaaaat 1920
tagccaggca tggtggcagg cgcctgtaat cccagctaat tgggaggctg aggctggaga 1980
atcccttgaa tctaggaggc agaggttgca gtgagctgag atcgtgccat tgcactccag 2040
cctgggtgac aagagtaaaa ctctgtctca aaaaaaaaaa attataccta cattctcttc 2100
ttatcagaga aaaaaatcta cagtgagctt ttcaaaaagt ttttacaaac tttttgccat 2160
ttaatttcag ttaggagttt tccctacttc tgacttagtt gaggggaaat gttcataaca 2220
tgtttataac atgtttatgt gtgttagttg gtgggggtgt attactttgc catgccattt 2280
gtttcctcca tgcgtaactt aatccagact ttcacacctt ataggaaaac cagtgagtct 2340
ggagagctgc atgggctcac aactgaggag gaatttgtag aagggatata caaagtggaa 2400
atagacacca aatcttactg gaaggcactt ggcatctccc cattccatga gcatgcagag 2460
gtgagtatac agaccttcga gggttgtttt ggttttggtt tttgcttttg gcattccagg 2520
aaatgcacag ttttactcag tgtaccacag aaatgtccta aggaaggtga tgaatgacca 2580
aaggttccct ttcctattat acaagaaaaa attcacaaca ctctgagaag caaatttctt 2640
tttgactttg atgaaaatcc acttagtaac atgacttgaa cttacatgaa actactcata 2700
gtctattcat tccactttat atgaatattg atgtatctgc tgttgaaata atagtttatg 2760
aggcagccct ccagacccca cgtagagtgt atgtaacaag agatgcacca ttttatttct 2820
cgaaaacccg taacattctt cattccaaaa cacatctggc ttctcggagg tctggacaag 2880
tgattcttgg caacacatac ctatagagac aataaaatca aagtaataat ggcaacacaa 2940
tagataacat ttaccaagca tacaccatgt ggcagacaca attataagtg ttttccatat 3000
ttaacctact taatcctcag gaataagcca ctgaggtcag tcctattatt atccccatct 3060
tatagatgaa gaaaatgagg caccaggaag tcaaataact tgtcaaaggt cacaagacta 3120
ggaaatacac aagtagaaat gtttacaatt aaggcccagg ctgggtttgc cctcagttct 3180
gctatgcctc gcattatgcc ccaggaaact ttttcccttg tgaaagccaa gcttaaaaaa 3240
agaaaagcca catttgtaac gtgctctgtt cccctgccta tggtgaggat cttcaaacag 3300
ttatacatgg acccagtccc cctgccttct ccttaatttc ttaagtcatt tgaaacagat 3360
ggctgtcatg gaaatagaat ccagacatgt tggtcagagt taaagatcaa ctaattccat 3420
caaaaatagc tcggcatgaa agggaactat tctctggctt agtcatggat gagactttca 3480
attgctataa agtggttcct ttattagaca atgttaccag ggaaacaaca ggggtttgtt 3540
tgacttctgg ggcccacaag tcaacaagag agccccatct accaaggagc atgtccctga 3600
ctacccctca gccagcagca agacatggac cccagtcagg gcaggagcag ggtttcggcg 3660
gcgcccagca caagacattg cccctagagt ctcagcccct accctcgagt aatagatctg 3720
cctacctgag actgttgttt gcccaagagc tgggtctcag cctgatggga accatataaa 3780
aaggttcact gacatactgc ccacatgttg ttctctttca ttagatctta gcttccttgt 3840
ctgctcttca ttcttgcagt attcattcaa caaacattaa aaaaaaaaaa aagcattcta 3900
tgtgtggaac actctgctag atgctgtgga tttagaaatg aaaatacatc ccgacccttg 3960
gaatggaagg gaaaggactg aagtaagaca gattaagcag gaccgtcagc ccagcttgaa 4020
gcccagataa atacggagaa caagagagag cgagtagtga gagatgagtc ccaatgcctc 4080
actttggtga cgggtgcgtg gtgggcttca tgcagcttct tctgataaat gcctccttca 4140
gaactggtca actctacctt ggccagtgac ccaggtggtc atagtagatt taccaaggga 4200
aaatggaaac ttttattagg agctcttagg cctcttcact tcatggattt ttttttcctt 4260
tttttttgag atggagtttt gccctgtcac ccaggctgga atgcagtggt gcaatctcag 4320
ctcactgcaa cctccgcctc ccaggttcaa gcaattctcc tgcctcagcc tcccgagtag 4380
ctgggactac aggtgtgcgc caccacacca ggctaatttt tgtatttttt gtaaagacag 4440
gttttcacca cgttggccag gctggtctga actccagacc tcaggtgatt cacctgtctc 4500
agcctcccaa agtgctggga ttacaggtgt gagccaccgt gcccggctac ttcatggatt 4560
tttgattaca gattatgcct cttacaattt ttaagaagaa tcaagtgggc tgaaggtcaa 4620
tgtcaccata agacaaaaga catttttatt agttgattct agggaattgg ccttaagggg 4680
agccctttct tcctaagaga ttcttaggtg attctcactt cctcttgccc cagtattatt 4740
tttgtttttg gtatggctca ctcagatcct tttttcctcc tatccctaag taatccgggt 4800
ttctttttcc catatttaga acaaaatgta tttatgcaga gtgtgtccaa acctcaaccc 4860
aaggcctgta tacaaaataa atcaaattaa acacatcttt actgtcttct acctctttcc 4920
tgacctcaat atatcccaac ttgcctcact ctgagaacca aggctgtccc agcacctgag 4980
tcgcagatat tctactgatt tgacagaact gtgtgactat ctggaacagc attttgatcc 5040
acaatttgcc cagttacaaa gcttaaatga gctctagtgc atgcatatat atttcaaaat 5100
tccaccatga tcttccacac tctgtattgt aaatagagcc ctgtaatgct tttacttcgt 5160
atttcattgc ttgttataca taaaaatata cttttcttct tcatgttaga aaatgcaaag 5220
aataggaggg tgggggaatc tctgggcttg gagacaggag acttgccttc ctactatggt 5280
tccatcagaa tgtagactgg gacaatacaa taattcaagt ctggtttgct catctgtaaa 5340
ttgggaagaa tgtttccagc tccagaatgc taaatctcta agtctgtggt tggcagccac 5400
tattgcagca gctcttcaat gactcaatgc agttttgcat tctccctacc ttttttttct 5460
aaaaccaata aaatagatac agcctttagg ctttctggga tttcccttag tcaagctagg 5520
gtcatcctga ctttcggcgt gaatttgcaa aacaagacct gactctgtac tcctgctcta 5580
aggactgtgc atggttccaa aggcttagct tgccagcata tttgagcttt ttccttctgt 5640
tcaaactgtt ccaaaatata aaagaataaa attaattaag ttggcactgg acttccggtg 5700
gtcagtcatg tgtgtcatct gtcacgtttt tcgggctctg gtggaaatgg atctgtctgt 5760
cttctctcat aggtggtatt cacagccaac gactccggcc cccgccgcta caccattgcc 5820
gccctgctga gcccctactc ctattccacc acggctgtcg tcaccaatcc caaggaatga 5880
gggacttctc ctccagtgga cctgaaggac gagggatggg atttcatgta accaagagta 5940
ttccattttt actaaagcag tgttttcacc tcatatgcta tgttagaagt ccaggcagag 6000
acaataaaac attcctgtga aaggcacttt tcattccact ttaacttgat tttttaaatt 6060
cccttattgt cccttccaaa aaaaagagaa tcaaaatttt acaaagaatc aaaggaattc 6120
tagaaagtat ctgggcagaa cgctaggaga gatccaaatt tccattgtct tgcaagcaaa 6180
gcacgtatta aatatgatct gcagccatta aaaagacaca ttctgtaaat gagagagcct 6240
tattttcctg taaccttcag caaatagcaa aagacacatt ccaagggccc acttctttac 6300
tgtgggcact cgagataact tcgtataatg tatgctatac gaagttatat gcatgccagt 6360
agcagcaccc acgtccacct tctgtctagt aatgtccaac acctccctca gtccaaacac 6420
tgctctgcat ccatgtggct cccatttata cctgaagcac ttgatggggc ctcaatgttt 6480
tactagagcc cacccccctg caactctgag accctctgga tttgtctgtc agtgcctcac 6540
tggggcgttg gataatttct taaaaggtca agttccctca gcagcattct ctgagcagtc 6600
tgaagatgtg tgcttttcac agttcaaatc catgtggctg tttcacccac ctgcctggcc 6660
ttgggttatc tatcaggacc tagcctagaa gcaggtgtgt ggcacttaac acctaagctg 6720
agtgactaac tgaacactca agtggatgcc atctttgtca cttcttgact gtgacacaag 6780
caactcctga tgccaaagcc ctgcccaccc ctctcatgcc catatttgga catggtacag 6840
gtcctcactg gccatggtct gtgaggtcct ggtcctcttt gacttcataa ttcctagggg 6900
ccactagtat ctataagagg aagagggtgc tggctcccag gccacagccc acaaaattcc 6960
acctgctcac aggttggctg gctcgaccca ggtggtgtcc cctgctctga gccagctccc 7020
ggccaagcca gcaccatggg aacccccaag aagaagagga aggtgcgtac cgatttaaat 7080
tccaatttac tgaccgtaca ccaaaatttg cctgcattac cggtcgatgc aacgagtgat 7140
gaggttcgca agaacctgat ggacatgttc agggatcgcc aggcgttttc tgagcatacc 7200
tggaaaatgc ttctgtccgt ttgccggtcg tgggcggcat ggtgcaagtt gaataaccgg 7260
aaatggtttc ccgcagaacc tgaagatgtt cgcgattatc ttctatatct tcaggcgcgc 7320
ggtctggcag taaaaactat ccagcaacat ttgggccagc taaacatgct tcatcgtcgg 7380
tccgggctgc cacgaccaag tgacagcaat gctgtttcac tggttatgcg gcggatccga 7440
aaagaaaacg ttgatgccgg tgaacgtgca aaacaggtaa atataaaatt tttaagtgta 7500
taatgatgtt aaactactga ttctaattgt ttgtgtattt taggctctag cgttcgaacg 7560
cactgatttc gaccaggttc gttcactcat ggaaaatagc gatcgctgcc aggatatacg 7620
taatctggca tttctgggga ttgcttataa caccctgtta cgtatagccg aaattgccag 7680
gatcagggtt aaagatatct cacgtactga cggtgggaga atgttaatcc atattggcag 7740
aacgaaaacg ctggttagca ccgcaggtgt agagaaggca cttagcctgg gggtaactaa 7800
actggtcgag cgatggattt ccgtctctgg tgtagctgat gatccgaata actacctgtt 7860
ttgccgggtc agaaaaaatg gtgttgccgc gccatctgcc accagccagc tatcaactcg 7920
cgccctggaa gggatttttg aagcaactca tcgattgatt tacggcgcta aggatgactc 7980
tggtcagaga tacctggcct ggtctggaca cagtgcccgt gtcggagccg cgcgagatat 8040
ggcccgcgct ggagtttcaa taccggagat catgcaagct ggtggctgga ccaatgtaaa 8100
tattgtcatg aactatatcc gtaacctgga tagtgaaaca ggggcaatgg tgcgcctgct 8160
ggaagatggc gattaggcgg ccggccgcta atcagccata ccacatttgt agaggtttta 8220
cttgctttaa aaaacctccc acacctcccc ctgaacctga aacataaaat gaatgcaatt 8280
gttgttgtta acttgtttat tgcagcttat aatggttaca aataaagcaa tagcatcaca 8340
aatttcacaa ataaagcatt tttttcactg cattctagtt gtggtttgtc caaactcatc 8400
aatgtatctt atcatgtctg gatcccccgg ctagagttta aacactagaa ctagtggatc 8460
ccccgggatc atggcctccg cgccgggttt tggcgcctcc cgcgggcgcc cccctcctca 8520
cggcgagcgc tgccacgtca gacgaagggc gcagcgagcg tcctgatcct tccgcccgga 8580
cgctcaggac agcggcccgc tgctcataag actcggcctt agaaccccag tatcagcaga 8640
aggacatttt aggacgggac ttgggtgact ctagggcact ggttttcttt ccagagagcg 8700
gaacaggcga ggaaaagtag tcccttctcg gcgattctgc ggagggatct ccgtggggcg 8760
gtgaacgccg atgattatat aaggacgcgc cgggtgtggc acagctagtt ccgtcgcagc 8820
cgggatttgg gtcgcggttc ttgtttgtgg atcgctgtga tcgtcacttg gtgagtagcg 8880
ggctgctggg ctggccgggg ctttcgtggc cgccgggccg ctcggtggga cggaagcgtg 8940
tggagagacc gccaagggct gtagtctggg tccgcgagca aggttgccct gaactggggg 9000
ttggggggag cgcagcaaaa tggcggctgt tcccgagtct tgaatggaag acgcttgtga 9060
ggcgggctgt gaggtcgttg aaacaaggtg gggggcatgg tgggcggcaa gaacccaagg 9120
tcttgaggcc ttcgctaatg cgggaaagct cttattcggg tgagatgggc tggggcacca 9180
tctggggacc ctgacgtgaa gtttgtcact gactggagaa ctcggtttgt cgtctgttgc 9240
gggggcggca gttatggcgg tgccgttggg cagtgcaccc gtacctttgg gagcgcgcgc 9300
cctcgtcgtg tcgtgacgtc acccgttctg ttggcttata atgcagggtg gggccacctg 9360
ccggtaggtg tgcggtaggc ttttctccgt cgcaggacgc agggttcggg cctagggtag 9420
gctctcctga atcgacaggc gccggacctc tggtgagggg agggataagt gaggcgtcag 9480
tttctttggt cggttttatg tacctatctt cttaagtagc tgaagctccg gttttgaact 9540
atgcgctcgg ggttggcgag tgtgttttgt gaagtttttt aggcaccttt tgaaatgtaa 9600
tcatttgggt caatatgtaa ttttcagtgt tagactagta aattgtccgc taaattctgg 9660
ccgtttttgg cttttttgtt agacgtgttg acaattaatc atcggcatag tatatcggca 9720
tagtataata cgacaaggtg aggaactaaa ccatgggatc ggccattgaa caagatggat 9780
tgcacgcagg ttctccggcc gcttgggtgg agaggctatt cggctatgac tgggcacaac 9840
agacaatcgg ctgctctgat gccgccgtgt tccggctgtc agcgcagggg cgcccggttc 9900
tttttgtcaa gaccgacctg tccggtgccc tgaatgaact gcaggacgag gcagcgcggc 9960
tatcgtggct ggccacgacg ggcgttcctt gcgcagctgt gctcgacgtt gtcactgaag 10020
cgggaaggga ctggctgcta ttgggcgaag tgccggggca ggatctcctg tcatctcacc 10080
ttgctcctgc cgagaaagta tccatcatgg ctgatgcaat gcggcggctg catacgcttg 10140
atccggctac ctgcccattc gaccaccaag cgaaacatcg catcgagcga gcacgtactc 10200
ggatggaagc cggtcttgtc gatcaggatg atctggacga agagcatcag gggctcgcgc 10260
cagccgaact gttcgccagg ctcaaggcgc gcatgcccga cggcgatgat ctcgtcgtga 10320
cccatggcga tgcctgcttg ccgaatatca tggtggaaaa tggccgcttt tctggattca 10380
tcgactgtgg ccggctgggt gtggcggacc gctatcagga catagcgttg gctacccgtg 10440
atattgctga agagcttggc ggcgaatggg ctgaccgctt cctcgtgctt tacggtatcg 10500
ccgctcccga ttcgcagcgc atcgccttct atcgccttct tgacgagttc ttctgagggg 10560
atccgctgta agtctgcaga aattgatgat ctattaaaca ataaagatgt ccactaaaat 10620
ggaagttttt cctgtcatac tttgttaaga agggtgagaa cagagtacct acattttgaa 10680
tggaaggatt ggagctacgg gggtgggggt ggggtgggat tagataaatg cctgctcttt 10740
actgaaggct ctttactatt gctttatgat aatgtttcat agttggatat cataatttaa 10800
acaagcaaaa ccaaattaag ggccagctca ttcctcccac tcatgatcta tagatctata 10860
gatctctcgt gggatcattg tttttctctt gattcccact ttgtggttct aagtactgtg 10920
gtttccaaat gtgtcagttt catagcctga agaacgagat cagcagcctc tgttccacat 10980
acacttcatt ctcagtattg ttttgccaag ttctaattcc atcagacctc gacctgcagc 11040
ccctagataa cttcgtataa tgtatgctat acgaagttat gctaggtaac tataacggtc 11100
ctaaggtagc gagctagcga gactcagccc aggaggacca ggatcttgcc aaagcagtag 11160
catcccattt gtaccaaaac agtgttcttg ctctataaac cgtgttagca gctcagga 11218
<210> 17
<211> 6486
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (1)..(100)
<223> Мышиная последовательность
<220>
<221> прочий признак
<222> (101)..(6308)
<223> Человеческая последовательность
<220>
<221> прочий признак
<222> (6309)..(6386)
<223> Кассета LoxP Scar
<220>
<221> прочий признак
<222> (6387)..(6486)
<223> Мышиная последовательность
<400> 17
atgtactttt ggttttgttc cagagtctat caccggaaag aacaagccgg tttactctga 60
cccatttcac tgacatttct cttgtctcct ctgtgcccag ggcaccggtg aatccaagtg 120
tcctctgatg gtcaaagttc tagatgctgt ccgaggcagt cctgccatca atgtggccgt 180
gcatgtgttc agaaaggctg ctgatgacac ctgggagcca tttgcctctg ggtaagttgc 240
caaagaaccc tcccacagga cttggtttta tcttcccgtt tgcccctcac ttggtagaga 300
gaggctcaca tcatctgcta aagaatttac aagtagattg aaaaacgtag gcagaggtca 360
agtatgccct ctgaaggatg ccctcttttt gttttgctta gctaggaagt gaccaggaac 420
ctgagcatca tttaggggca gacagtagag aaaagaagga atcagaactc ctctcctcta 480
gctgtggttt gcaacccttt tgggtcacag aacactttat gtaggtgatg aaaagtaaac 540
attctatgcc cagaaaaaat gcacagatac acacacatac aaaatcatat atgtgatttt 600
aggagtttca cagattccct ggtgtccctg ggtaacacca aagctaagtg tccttgtctt 660
agaattttag gaaaaggtat aatgtgtatt aacccattaa caaaaggaaa ggaattcaga 720
aatattatta accaggcatc tgtctgtagt taatatggat cacccaaaac ccaaggcttt 780
tgcctaatga acactttggg gcacctactg tgtgcaaggc tgggggctgt caagctcagt 840
taaaaaaaaa aagatagaag agatggatcc atgaggcaaa gtacagcccc aggctaatcc 900
cacgatcacc cgacttcatg tccaagagtg gcttctcacc ttcattagcc agttcacaat 960
tttcatggag tttttctacc tgcactagca aaaacttcaa ggaaaataca tattaataaa 1020
tctaagcaaa gtgaccagaa gacagagcaa tcaggagacc ctttgcatcc agcagaagag 1080
gaactgctaa gtatttacat ctccacagag aagaatttct gttgggtttt aattgaaccc 1140
caagaaccac atgattcttc aaccattatt gggaagatca ttttcttagg tctggtttta 1200
actggctttt tatttgggaa ttcatttatg tttatataaa atgccaagca taacatgaaa 1260
agtggttaca ggactattct aagggagaga cagaatggac accaaaaata ttccaatgtt 1320
cttgtgaatc ttttccttgc accaggacaa aaaaaaaaag aagtgaaaag aagaaaggag 1380
gaggggcata atcagagtca gtaaagacaa ctgctatttt tatctatcgt agctgttgca 1440
gtcaaatggg aagcaatttc caacattcaa ctatggagct ggtacttaca tggaaataga 1500
agttgcctag tgtttgttgc tggcaaagag ttatcagaga ggttaaatat ataaaaggga 1560
aaagagtcag atacaggttc ttcttcctac tttaggtttt ccactgtgtg tgcaaatgat 1620
actccctggt ggtgtgcaga tgcctcaaag ctatcctcac accacaaggg agaggagcga 1680
gatcctgctg tcctggagaa gtgcagagtt agaacagctg tggccacttg catccaatca 1740
tcaatcttga atcacaggga ctctttctta agtaaacatt atacctggcc gggcacggtg 1800
gctcacgcct gtaatcccag cactttggga tgccaaagtg ggcatatcat ctgaggtcag 1860
gagttcaaga ccagcctggc caacatggca aaactccgtc tttatgaaaa atacaaaaat 1920
tagccaggca tggtggcagg cgcctgtaat cccagctaat tgggaggctg aggctggaga 1980
atcccttgaa tctaggaggc agaggttgca gtgagctgag atcgtgccat tgcactccag 2040
cctgggtgac aagagtaaaa ctctgtctca aaaaaaaaaa attataccta cattctcttc 2100
ttatcagaga aaaaaatcta cagtgagctt ttcaaaaagt ttttacaaac tttttgccat 2160
ttaatttcag ttaggagttt tccctacttc tgacttagtt gaggggaaat gttcataaca 2220
tgtttataac atgtttatgt gtgttagttg gtgggggtgt attactttgc catgccattt 2280
gtttcctcca tgcgtaactt aatccagact ttcacacctt ataggaaaac cagtgagtct 2340
ggagagctgc atgggctcac aactgaggag gaatttgtag aagggatata caaagtggaa 2400
atagacacca aatcttactg gaaggcactt ggcatctccc cattccatga gcatgcagag 2460
gtgagtatac agaccttcga gggttgtttt ggttttggtt tttgcttttg gcattccagg 2520
aaatgcacag ttttactcag tgtaccacag aaatgtccta aggaaggtga tgaatgacca 2580
aaggttccct ttcctattat acaagaaaaa attcacaaca ctctgagaag caaatttctt 2640
tttgactttg atgaaaatcc acttagtaac atgacttgaa cttacatgaa actactcata 2700
gtctattcat tccactttat atgaatattg atgtatctgc tgttgaaata atagtttatg 2760
aggcagccct ccagacccca cgtagagtgt atgtaacaag agatgcacca ttttatttct 2820
cgaaaacccg taacattctt cattccaaaa cacatctggc ttctcggagg tctggacaag 2880
tgattcttgg caacacatac ctatagagac aataaaatca aagtaataat ggcaacacaa 2940
tagataacat ttaccaagca tacaccatgt ggcagacaca attataagtg ttttccatat 3000
ttaacctact taatcctcag gaataagcca ctgaggtcag tcctattatt atccccatct 3060
tatagatgaa gaaaatgagg caccaggaag tcaaataact tgtcaaaggt cacaagacta 3120
ggaaatacac aagtagaaat gtttacaatt aaggcccagg ctgggtttgc cctcagttct 3180
gctatgcctc gcattatgcc ccaggaaact ttttcccttg tgaaagccaa gcttaaaaaa 3240
agaaaagcca catttgtaac gtgctctgtt cccctgccta tggtgaggat cttcaaacag 3300
ttatacatgg acccagtccc cctgccttct ccttaatttc ttaagtcatt tgaaacagat 3360
ggctgtcatg gaaatagaat ccagacatgt tggtcagagt taaagatcaa ctaattccat 3420
caaaaatagc tcggcatgaa agggaactat tctctggctt agtcatggat gagactttca 3480
attgctataa agtggttcct ttattagaca atgttaccag ggaaacaaca ggggtttgtt 3540
tgacttctgg ggcccacaag tcaacaagag agccccatct accaaggagc atgtccctga 3600
ctacccctca gccagcagca agacatggac cccagtcagg gcaggagcag ggtttcggcg 3660
gcgcccagca caagacattg cccctagagt ctcagcccct accctcgagt aatagatctg 3720
cctacctgag actgttgttt gcccaagagc tgggtctcag cctgatggga accatataaa 3780
aaggttcact gacatactgc ccacatgttg ttctctttca ttagatctta gcttccttgt 3840
ctgctcttca ttcttgcagt attcattcaa caaacattaa aaaaaaaaaa aagcattcta 3900
tgtgtggaac actctgctag atgctgtgga tttagaaatg aaaatacatc ccgacccttg 3960
gaatggaagg gaaaggactg aagtaagaca gattaagcag gaccgtcagc ccagcttgaa 4020
gcccagataa atacggagaa caagagagag cgagtagtga gagatgagtc ccaatgcctc 4080
actttggtga cgggtgcgtg gtgggcttca tgcagcttct tctgataaat gcctccttca 4140
gaactggtca actctacctt ggccagtgac ccaggtggtc atagtagatt taccaaggga 4200
aaatggaaac ttttattagg agctcttagg cctcttcact tcatggattt ttttttcctt 4260
tttttttgag atggagtttt gccctgtcac ccaggctgga atgcagtggt gcaatctcag 4320
ctcactgcaa cctccgcctc ccaggttcaa gcaattctcc tgcctcagcc tcccgagtag 4380
ctgggactac aggtgtgcgc caccacacca ggctaatttt tgtatttttt gtaaagacag 4440
gttttcacca cgttggccag gctggtctga actccagacc tcaggtgatt cacctgtctc 4500
agcctcccaa agtgctggga ttacaggtgt gagccaccgt gcccggctac ttcatggatt 4560
tttgattaca gattatgcct cttacaattt ttaagaagaa tcaagtgggc tgaaggtcaa 4620
tgtcaccata agacaaaaga catttttatt agttgattct agggaattgg ccttaagggg 4680
agccctttct tcctaagaga ttcttaggtg attctcactt cctcttgccc cagtattatt 4740
tttgtttttg gtatggctca ctcagatcct tttttcctcc tatccctaag taatccgggt 4800
ttctttttcc catatttaga acaaaatgta tttatgcaga gtgtgtccaa acctcaaccc 4860
aaggcctgta tacaaaataa atcaaattaa acacatcttt actgtcttct acctctttcc 4920
tgacctcaat atatcccaac ttgcctcact ctgagaacca aggctgtccc agcacctgag 4980
tcgcagatat tctactgatt tgacagaact gtgtgactat ctggaacagc attttgatcc 5040
acaatttgcc cagttacaaa gcttaaatga gctctagtgc atgcatatat atttcaaaat 5100
tccaccatga tcttccacac tctgtattgt aaatagagcc ctgtaatgct tttacttcgt 5160
atttcattgc ttgttataca taaaaatata cttttcttct tcatgttaga aaatgcaaag 5220
aataggaggg tgggggaatc tctgggcttg gagacaggag acttgccttc ctactatggt 5280
tccatcagaa tgtagactgg gacaatacaa taattcaagt ctggtttgct catctgtaaa 5340
ttgggaagaa tgtttccagc tccagaatgc taaatctcta agtctgtggt tggcagccac 5400
tattgcagca gctcttcaat gactcaatgc agttttgcat tctccctacc ttttttttct 5460
aaaaccaata aaatagatac agcctttagg ctttctggga tttcccttag tcaagctagg 5520
gtcatcctga ctttcggcgt gaatttgcaa aacaagacct gactctgtac tcctgctcta 5580
aggactgtgc atggttccaa aggcttagct tgccagcata tttgagcttt ttccttctgt 5640
tcaaactgtt ccaaaatata aaagaataaa attaattaag ttggcactgg acttccggtg 5700
gtcagtcatg tgtgtcatct gtcacgtttt tcgggctctg gtggaaatgg atctgtctgt 5760
cttctctcat aggtggtatt cacagccaac gactccggcc cccgccgcta caccattgcc 5820
gccctgctga gcccctactc ctattccacc acggctgtcg tcaccaatcc caaggaatga 5880
gggacttctc ctccagtgga cctgaaggac gagggatggg atttcatgta accaagagta 5940
ttccattttt actaaagcag tgttttcacc tcatatgcta tgttagaagt ccaggcagag 6000
acaataaaac attcctgtga aaggcacttt tcattccact ttaacttgat tttttaaatt 6060
cccttattgt cccttccaaa aaaaagagaa tcaaaatttt acaaagaatc aaaggaattc 6120
tagaaagtat ctgggcagaa cgctaggaga gatccaaatt tccattgtct tgcaagcaaa 6180
gcacgtatta aatatgatct gcagccatta aaaagacaca ttctgtaaat gagagagcct 6240
tattttcctg taaccttcag caaatagcaa aagacacatt ccaagggccc acttctttac 6300
tgtgggcact cgagataact tcgtataatg tatgctatac gaagttatgc taggtaacta 6360
taacggtcct aaggtagcga gctagcgaga ctcagcccag gaggaccagg atcttgccaa 6420
agcagtagca tcccatttgt accaaaacag tgttcttgct ctataaaccg tgttagcagc 6480
tcagga 6486
<210> 18
<211> 7201
<212> ДНК
<213> Homo sapiens
<400> 18
atggcttctc atcgtctgct cctcctctgc cttgctggac tggtatttgt gtctgaggct 60
ggccctacgg tgagtgtttc tgtgacatcc cattcctaca tttaagattc acgctaaatg 120
aagtagaagt gactccttcc agctttgcca accagctttt attactaggg caagggtacc 180
cagcatctat ttttaatata attaattcaa acttcaaaaa gaatgaagtt ccactgagct 240
tactgagctg ggacttgaac tctgagcatt ctacctcatt gctttggtgc attaggtttg 300
taatatctgg tacctctgtt tcctcagata gatgatagaa ataaagatat gatattaagg 360
aagctgttaa tactgaattt tcagaaaagt atccctccat aaaatgtatt tgggggacaa 420
actgcaggag attatattct ggccctatag ttattcaaaa cgtatttatt gattaatctt 480
taaaaggctt agtgaacaat attctagtca gatatctaat tcttaaatcc tctagaagaa 540
ttaactaata ctataaaatg ggtctggatg tagttctgac attattttat aacaactggt 600
aagagggagt gactatagca acaactaaaa tgatctcagg aaaacctgtt tggccctatg 660
tatggtacat tacatctttt cagtaattcc actcaaatgg agacttttaa caaagcaact 720
gttctcaggg gacctatttt ctcccttaaa attcattata cacatccctg gttgatagca 780
gtgtgtctgg aggcagaaac cattcttgct ttggaaacaa ttacgtctgt gttatactga 840
gtagggaagc tcattaattg tcgacactta cgttcctgat aatgggatca gtgtgtaatt 900
cttgtttcgc tccagatttc taataccaca aagaataaat cctttcactc tgatcaattt 960
tgttaacttc tcacgtgtct tctctacacc cagggcaccg gtgaatccaa gtgtcctctg 1020
atggtcaaag ttctagatgc tgtccgaggc agtcctgcca tcaatgtggc cgtgcatgtg 1080
ttcagaaagg ctgctgatga cacctgggag ccatttgcct ctgggtaagt tgccaaagaa 1140
ccctcccaca ggacttggtt ttatcttccc gtttgcccct cacttggtag agagaggctc 1200
acatcatctg ctaaagaatt tacaagtaga ttgaaaaacg taggcagagg tcaagtatgc 1260
cctctgaagg atgccctctt tttgttttgc ttagctagga agtgaccagg aacctgagca 1320
tcatttaggg gcagacagta gagaaaagaa ggaatcagaa ctcctctcct ctagctgtgg 1380
tttgcaaccc ttttgggtca cagaacactt tatgtaggtg atgaaaagta aacattctat 1440
gcccagaaaa aatgcacaga tacacacaca tacaaaatca tatatgtgat tttaggagtt 1500
tcacagattc cctggtgtcc ctgggtaaca ccaaagctaa gtgtccttgt cttagaattt 1560
taggaaaagg tataatgtgt attaacccat taacaaaagg aaaggaattc agaaatatta 1620
ttaaccaggc atctgtctgt agttaatatg gatcacccaa aacccaaggc ttttgcctaa 1680
tgaacacttt ggggcaccta ctgtgtgcaa ggctgggggc tgtcaagctc agttaaaaaa 1740
aaaaagatag aagagatgga tccatgaggc aaagtacagc cccaggctaa tcccacgatc 1800
acccgacttc atgtccaaga gtggcttctc accttcatta gccagttcac aattttcatg 1860
gagtttttct acctgcacta gcaaaaactt caaggaaaat acatattaat aaatctaagc 1920
aaagtgacca gaagacagag caatcaggag accctttgca tccagcagaa gaggaactgc 1980
taagtattta catctccaca gagaagaatt tctgttgggt tttaattgaa ccccaagaac 2040
cacatgattc ttcaaccatt attgggaaga tcattttctt aggtctggtt ttaactggct 2100
ttttatttgg gaattcattt atgtttatat aaaatgccaa gcataacatg aaaagtggtt 2160
acaggactat tctaagggag agacagaatg gacaccaaaa atattccaat gttcttgtga 2220
atcttttcct tgcaccagga caaaaaaaaa aagaagtgaa aagaagaaag gaggaggggc 2280
ataatcagag tcagtaaaga caactgctat ttttatctat cgtagctgtt gcagtcaaat 2340
gggaagcaat ttccaacatt caactatgga gctggtactt acatggaaat agaagttgcc 2400
tagtgtttgt tgctggcaaa gagttatcag agaggttaaa tatataaaag ggaaaagagt 2460
cagatacagg ttcttcttcc tactttaggt tttccactgt gtgtgcaaat gatactccct 2520
ggtggtgtgc agatgcctca aagctatcct cacaccacaa gggagaggag cgagatcctg 2580
ctgtcctgga gaagtgcaga gttagaacag ctgtggccac ttgcatccaa tcatcaatct 2640
tgaatcacag ggactctttc ttaagtaaac attatacctg gccgggcacg gtggctcacg 2700
cctgtaatcc cagcactttg ggatgccaaa gtgggcatat catctgaggt caggagttca 2760
agaccagcct ggccaacatg gcaaaactcc gtctttatga aaaatacaaa aattagccag 2820
gcatggtggc aggcgcctgt aatcccagct aattgggagg ctgaggctgg agaatccctt 2880
gaatctagga ggcagaggtt gcagtgagct gagatcgtgc cattgcactc cagcctgggt 2940
gacaagagta aaactctgtc tcaaaaaaaa aaaattatac ctacattctc ttcttatcag 3000
agaaaaaaat ctacagtgag cttttcaaaa agtttttaca aactttttgc catttaattt 3060
cagttaggag ttttccctac ttctgactta gttgagggga aatgttcata acatgtttat 3120
aacatgttta tgtgtgttag ttggtggggg tgtattactt tgccatgcca tttgtttcct 3180
ccatgcgtaa cttaatccag actttcacac cttataggaa aaccagtgag tctggagagc 3240
tgcatgggct cacaactgag gaggaatttg tagaagggat atacaaagtg gaaatagaca 3300
ccaaatctta ctggaaggca cttggcatct ccccattcca tgagcatgca gaggtgagta 3360
tacagacctt cgagggttgt tttggttttg gtttttgctt ttggcattcc aggaaatgca 3420
cagttttact cagtgtacca cagaaatgtc ctaaggaagg tgatgaatga ccaaaggttc 3480
cctttcctat tatacaagaa aaaattcaca acactctgag aagcaaattt ctttttgact 3540
ttgatgaaaa tccacttagt aacatgactt gaacttacat gaaactactc atagtctatt 3600
cattccactt tatatgaata ttgatgtatc tgctgttgaa ataatagttt atgaggcagc 3660
cctccagacc ccacgtagag tgtatgtaac aagagatgca ccattttatt tctcgaaaac 3720
ccgtaacatt cttcattcca aaacacatct ggcttctcgg aggtctggac aagtgattct 3780
tggcaacaca tacctataga gacaataaaa tcaaagtaat aatggcaaca caatagataa 3840
catttaccaa gcatacacca tgtggcagac acaattataa gtgttttcca tatttaacct 3900
acttaatcct caggaataag ccactgaggt cagtcctatt attatcccca tcttatagat 3960
gaagaaaatg aggcaccagg aagtcaaata acttgtcaaa ggtcacaaga ctaggaaata 4020
cacaagtaga aatgtttaca attaaggccc aggctgggtt tgccctcagt tctgctatgc 4080
ctcgcattat gccccaggaa actttttccc ttgtgaaagc caagcttaaa aaaagaaaag 4140
ccacatttgt aacgtgctct gttcccctgc ctatggtgag gatcttcaaa cagttataca 4200
tggacccagt ccccctgcct tctccttaat ttcttaagtc atttgaaaca gatggctgtc 4260
atggaaatag aatccagaca tgttggtcag agttaaagat caactaattc catcaaaaat 4320
agctcggcat gaaagggaac tattctctgg cttagtcatg gatgagactt tcaattgcta 4380
taaagtggtt cctttattag acaatgttac cagggaaaca acaggggttt gtttgacttc 4440
tggggcccac aagtcaacaa gagagcccca tctaccaagg agcatgtccc tgactacccc 4500
tcagccagca gcaagacatg gaccccagtc agggcaggag cagggtttcg gcggcgccca 4560
gcacaagaca ttgcccctag agtctcagcc cctaccctcg agtaatagat ctgcctacct 4620
gagactgttg tttgcccaag agctgggtct cagcctgatg ggaaccatat aaaaaggttc 4680
actgacatac tgcccacatg ttgttctctt tcattagatc ttagcttcct tgtctgctct 4740
tcattcttgc agtattcatt caacaaacat taaaaaaaaa aaaaagcatt ctatgtgtgg 4800
aacactctgc tagatgctgt ggatttagaa atgaaaatac atcccgaccc ttggaatgga 4860
agggaaagga ctgaagtaag acagattaag caggaccgtc agcccagctt gaagcccaga 4920
taaatacgga gaacaagaga gagcgagtag tgagagatga gtcccaatgc ctcactttgg 4980
tgacgggtgc gtggtgggct tcatgcagct tcttctgata aatgcctcct tcagaactgg 5040
tcaactctac cttggccagt gacccaggtg gtcatagtag atttaccaag ggaaaatgga 5100
aacttttatt aggagctctt aggcctcttc acttcatgga tttttttttc cttttttttt 5160
gagatggagt tttgccctgt cacccaggct ggaatgcagt ggtgcaatct cagctcactg 5220
caacctccgc ctcccaggtt caagcaattc tcctgcctca gcctcccgag tagctgggac 5280
tacaggtgtg cgccaccaca ccaggctaat ttttgtattt tttgtaaaga caggttttca 5340
ccacgttggc caggctggtc tgaactccag acctcaggtg attcacctgt ctcagcctcc 5400
caaagtgctg ggattacagg tgtgagccac cgtgcccggc tacttcatgg atttttgatt 5460
acagattatg cctcttacaa tttttaagaa gaatcaagtg ggctgaaggt caatgtcacc 5520
ataagacaaa agacattttt attagttgat tctagggaat tggccttaag gggagccctt 5580
tcttcctaag agattcttag gtgattctca cttcctcttg ccccagtatt atttttgttt 5640
ttggtatggc tcactcagat ccttttttcc tcctatccct aagtaatccg ggtttctttt 5700
tcccatattt agaacaaaat gtatttatgc agagtgtgtc caaacctcaa cccaaggcct 5760
gtatacaaaa taaatcaaat taaacacatc tttactgtct tctacctctt tcctgacctc 5820
aatatatccc aacttgcctc actctgagaa ccaaggctgt cccagcacct gagtcgcaga 5880
tattctactg atttgacaga actgtgtgac tatctggaac agcattttga tccacaattt 5940
gcccagttac aaagcttaaa tgagctctag tgcatgcata tatatttcaa aattccacca 6000
tgatcttcca cactctgtat tgtaaataga gccctgtaat gcttttactt cgtatttcat 6060
tgcttgttat acataaaaat atacttttct tcttcatgtt agaaaatgca aagaatagga 6120
gggtggggga atctctgggc ttggagacag gagacttgcc ttcctactat ggttccatca 6180
gaatgtagac tgggacaata caataattca agtctggttt gctcatctgt aaattgggaa 6240
gaatgtttcc agctccagaa tgctaaatct ctaagtctgt ggttggcagc cactattgca 6300
gcagctcttc aatgactcaa tgcagttttg cattctccct accttttttt tctaaaacca 6360
ataaaataga tacagccttt aggctttctg ggatttccct tagtcaagct agggtcatcc 6420
tgactttcgg cgtgaatttg caaaacaaga cctgactctg tactcctgct ctaaggactg 6480
tgcatggttc caaaggctta gcttgccagc atatttgagc tttttccttc tgttcaaact 6540
gttccaaaat ataaaagaat aaaattaatt aagttggcac tggacttccg gtggtcagtc 6600
atgtgtgtca tctgtcacgt ttttcgggct ctggtggaaa tggatctgtc tgtcttctct 6660
cataggtggt attcacagcc aacgactccg gcccccgccg ctacaccatt gccgccctgc 6720
tgagccccta ctcctattcc accacggctg tcgtcaccaa tcccaaggaa tgagggactt 6780
ctcctccagt ggacctgaag gacgagggat gggatttcat gtaaccaaga gtattccatt 6840
tttactaaag cagtgttttc acctcatatg ctatgttaga agtccaggca gagacaataa 6900
aacattcctg tgaaaggcac ttttcattcc actttaactt gattttttaa attcccttat 6960
tgtcccttcc aaaaaaaaga gaatcaaaat tttacaaaga atcaaaggaa ttctagaaag 7020
tatctgggca gaacgctagg agagatccaa atttccattg tcttgcaagc aaagcacgta 7080
ttaaatatga tctgcagcca ttaaaaagac acattctgta aatgagagag ccttattttc 7140
ctgtaacctt cagcaaatag caaaagacac attccaaggg cccacttctt tactgtgggc 7200
a 7201
<210> 19
<211> 6208
<212> ДНК
<213> Homo sapiens
<400> 19
ggcaccggtg aatccaagtg tcctctgatg gtcaaagttc tagatgctgt ccgaggcagt 60
cctgccatca atgtggccgt gcatgtgttc agaaaggctg ctgatgacac ctgggagcca 120
tttgcctctg ggtaagttgc caaagaaccc tcccacagga cttggtttta tcttcccgtt 180
tgcccctcac ttggtagaga gaggctcaca tcatctgcta aagaatttac aagtagattg 240
aaaaacgtag gcagaggtca agtatgccct ctgaaggatg ccctcttttt gttttgctta 300
gctaggaagt gaccaggaac ctgagcatca tttaggggca gacagtagag aaaagaagga 360
atcagaactc ctctcctcta gctgtggttt gcaacccttt tgggtcacag aacactttat 420
gtaggtgatg aaaagtaaac attctatgcc cagaaaaaat gcacagatac acacacatac 480
aaaatcatat atgtgatttt aggagtttca cagattccct ggtgtccctg ggtaacacca 540
aagctaagtg tccttgtctt agaattttag gaaaaggtat aatgtgtatt aacccattaa 600
caaaaggaaa ggaattcaga aatattatta accaggcatc tgtctgtagt taatatggat 660
cacccaaaac ccaaggcttt tgcctaatga acactttggg gcacctactg tgtgcaaggc 720
tgggggctgt caagctcagt taaaaaaaaa aagatagaag agatggatcc atgaggcaaa 780
gtacagcccc aggctaatcc cacgatcacc cgacttcatg tccaagagtg gcttctcacc 840
ttcattagcc agttcacaat tttcatggag tttttctacc tgcactagca aaaacttcaa 900
ggaaaataca tattaataaa tctaagcaaa gtgaccagaa gacagagcaa tcaggagacc 960
ctttgcatcc agcagaagag gaactgctaa gtatttacat ctccacagag aagaatttct 1020
gttgggtttt aattgaaccc caagaaccac atgattcttc aaccattatt gggaagatca 1080
ttttcttagg tctggtttta actggctttt tatttgggaa ttcatttatg tttatataaa 1140
atgccaagca taacatgaaa agtggttaca ggactattct aagggagaga cagaatggac 1200
accaaaaata ttccaatgtt cttgtgaatc ttttccttgc accaggacaa aaaaaaaaag 1260
aagtgaaaag aagaaaggag gaggggcata atcagagtca gtaaagacaa ctgctatttt 1320
tatctatcgt agctgttgca gtcaaatggg aagcaatttc caacattcaa ctatggagct 1380
ggtacttaca tggaaataga agttgcctag tgtttgttgc tggcaaagag ttatcagaga 1440
ggttaaatat ataaaaggga aaagagtcag atacaggttc ttcttcctac tttaggtttt 1500
ccactgtgtg tgcaaatgat actccctggt ggtgtgcaga tgcctcaaag ctatcctcac 1560
accacaaggg agaggagcga gatcctgctg tcctggagaa gtgcagagtt agaacagctg 1620
tggccacttg catccaatca tcaatcttga atcacaggga ctctttctta agtaaacatt 1680
atacctggcc gggcacggtg gctcacgcct gtaatcccag cactttggga tgccaaagtg 1740
ggcatatcat ctgaggtcag gagttcaaga ccagcctggc caacatggca aaactccgtc 1800
tttatgaaaa atacaaaaat tagccaggca tggtggcagg cgcctgtaat cccagctaat 1860
tgggaggctg aggctggaga atcccttgaa tctaggaggc agaggttgca gtgagctgag 1920
atcgtgccat tgcactccag cctgggtgac aagagtaaaa ctctgtctca aaaaaaaaaa 1980
attataccta cattctcttc ttatcagaga aaaaaatcta cagtgagctt ttcaaaaagt 2040
ttttacaaac tttttgccat ttaatttcag ttaggagttt tccctacttc tgacttagtt 2100
gaggggaaat gttcataaca tgtttataac atgtttatgt gtgttagttg gtgggggtgt 2160
attactttgc catgccattt gtttcctcca tgcgtaactt aatccagact ttcacacctt 2220
ataggaaaac cagtgagtct ggagagctgc atgggctcac aactgaggag gaatttgtag 2280
aagggatata caaagtggaa atagacacca aatcttactg gaaggcactt ggcatctccc 2340
cattccatga gcatgcagag gtgagtatac agaccttcga gggttgtttt ggttttggtt 2400
tttgcttttg gcattccagg aaatgcacag ttttactcag tgtaccacag aaatgtccta 2460
aggaaggtga tgaatgacca aaggttccct ttcctattat acaagaaaaa attcacaaca 2520
ctctgagaag caaatttctt tttgactttg atgaaaatcc acttagtaac atgacttgaa 2580
cttacatgaa actactcata gtctattcat tccactttat atgaatattg atgtatctgc 2640
tgttgaaata atagtttatg aggcagccct ccagacccca cgtagagtgt atgtaacaag 2700
agatgcacca ttttatttct cgaaaacccg taacattctt cattccaaaa cacatctggc 2760
ttctcggagg tctggacaag tgattcttgg caacacatac ctatagagac aataaaatca 2820
aagtaataat ggcaacacaa tagataacat ttaccaagca tacaccatgt ggcagacaca 2880
attataagtg ttttccatat ttaacctact taatcctcag gaataagcca ctgaggtcag 2940
tcctattatt atccccatct tatagatgaa gaaaatgagg caccaggaag tcaaataact 3000
tgtcaaaggt cacaagacta ggaaatacac aagtagaaat gtttacaatt aaggcccagg 3060
ctgggtttgc cctcagttct gctatgcctc gcattatgcc ccaggaaact ttttcccttg 3120
tgaaagccaa gcttaaaaaa agaaaagcca catttgtaac gtgctctgtt cccctgccta 3180
tggtgaggat cttcaaacag ttatacatgg acccagtccc cctgccttct ccttaatttc 3240
ttaagtcatt tgaaacagat ggctgtcatg gaaatagaat ccagacatgt tggtcagagt 3300
taaagatcaa ctaattccat caaaaatagc tcggcatgaa agggaactat tctctggctt 3360
agtcatggat gagactttca attgctataa agtggttcct ttattagaca atgttaccag 3420
ggaaacaaca ggggtttgtt tgacttctgg ggcccacaag tcaacaagag agccccatct 3480
accaaggagc atgtccctga ctacccctca gccagcagca agacatggac cccagtcagg 3540
gcaggagcag ggtttcggcg gcgcccagca caagacattg cccctagagt ctcagcccct 3600
accctcgagt aatagatctg cctacctgag actgttgttt gcccaagagc tgggtctcag 3660
cctgatggga accatataaa aaggttcact gacatactgc ccacatgttg ttctctttca 3720
ttagatctta gcttccttgt ctgctcttca ttcttgcagt attcattcaa caaacattaa 3780
aaaaaaaaaa aagcattcta tgtgtggaac actctgctag atgctgtgga tttagaaatg 3840
aaaatacatc ccgacccttg gaatggaagg gaaaggactg aagtaagaca gattaagcag 3900
gaccgtcagc ccagcttgaa gcccagataa atacggagaa caagagagag cgagtagtga 3960
gagatgagtc ccaatgcctc actttggtga cgggtgcgtg gtgggcttca tgcagcttct 4020
tctgataaat gcctccttca gaactggtca actctacctt ggccagtgac ccaggtggtc 4080
atagtagatt taccaaggga aaatggaaac ttttattagg agctcttagg cctcttcact 4140
tcatggattt ttttttcctt tttttttgag atggagtttt gccctgtcac ccaggctgga 4200
atgcagtggt gcaatctcag ctcactgcaa cctccgcctc ccaggttcaa gcaattctcc 4260
tgcctcagcc tcccgagtag ctgggactac aggtgtgcgc caccacacca ggctaatttt 4320
tgtatttttt gtaaagacag gttttcacca cgttggccag gctggtctga actccagacc 4380
tcaggtgatt cacctgtctc agcctcccaa agtgctggga ttacaggtgt gagccaccgt 4440
gcccggctac ttcatggatt tttgattaca gattatgcct cttacaattt ttaagaagaa 4500
tcaagtgggc tgaaggtcaa tgtcaccata agacaaaaga catttttatt agttgattct 4560
agggaattgg ccttaagggg agccctttct tcctaagaga ttcttaggtg attctcactt 4620
cctcttgccc cagtattatt tttgtttttg gtatggctca ctcagatcct tttttcctcc 4680
tatccctaag taatccgggt ttctttttcc catatttaga acaaaatgta tttatgcaga 4740
gtgtgtccaa acctcaaccc aaggcctgta tacaaaataa atcaaattaa acacatcttt 4800
actgtcttct acctctttcc tgacctcaat atatcccaac ttgcctcact ctgagaacca 4860
aggctgtccc agcacctgag tcgcagatat tctactgatt tgacagaact gtgtgactat 4920
ctggaacagc attttgatcc acaatttgcc cagttacaaa gcttaaatga gctctagtgc 4980
atgcatatat atttcaaaat tccaccatga tcttccacac tctgtattgt aaatagagcc 5040
ctgtaatgct tttacttcgt atttcattgc ttgttataca taaaaatata cttttcttct 5100
tcatgttaga aaatgcaaag aataggaggg tgggggaatc tctgggcttg gagacaggag 5160
acttgccttc ctactatggt tccatcagaa tgtagactgg gacaatacaa taattcaagt 5220
ctggtttgct catctgtaaa ttgggaagaa tgtttccagc tccagaatgc taaatctcta 5280
agtctgtggt tggcagccac tattgcagca gctcttcaat gactcaatgc agttttgcat 5340
tctccctacc ttttttttct aaaaccaata aaatagatac agcctttagg ctttctggga 5400
tttcccttag tcaagctagg gtcatcctga ctttcggcgt gaatttgcaa aacaagacct 5460
gactctgtac tcctgctcta aggactgtgc atggttccaa aggcttagct tgccagcata 5520
tttgagcttt ttccttctgt tcaaactgtt ccaaaatata aaagaataaa attaattaag 5580
ttggcactgg acttccggtg gtcagtcatg tgtgtcatct gtcacgtttt tcgggctctg 5640
gtggaaatgg atctgtctgt cttctctcat aggtggtatt cacagccaac gactccggcc 5700
cccgccgcta caccattgcc gccctgctga gcccctactc ctattccacc acggctgtcg 5760
tcaccaatcc caaggaatga gggacttctc ctccagtgga cctgaaggac gagggatggg 5820
atttcatgta accaagagta ttccattttt actaaagcag tgttttcacc tcatatgcta 5880
tgttagaagt ccaggcagag acaataaaac attcctgtga aaggcacttt tcattccact 5940
ttaacttgat tttttaaatt cccttattgt cccttccaaa aaaaagagaa tcaaaatttt 6000
acaaagaatc aaaggaattc tagaaagtat ctgggcagaa cgctaggaga gatccaaatt 6060
tccattgtct tgcaagcaaa gcacgtatta aatatgatct gcagccatta aaaagacaca 6120
ttctgtaaat gagagagcct tattttcctg taaccttcag caaatagcaa aagacacatt 6180
ccaagggccc acttctttac tgtgggca 6208
<210> 20
<211> 8300
<212> ДНК
<213> Mus musculus
<400> 20
atggcttccc ttcgactctt cctcctttgc ctcgctggac tggtatttgt gtctgaagct 60
ggccccgcgg tgagtgatcc tgtgagcgat ccagacatgg cagttagacc ttagataaag 120
aagaagtgcc ttcttccaga tgtgagaact agagtactca gactctatat ttaccattag 180
actccaaaga gaagagctgg agtgcctctg gctcttcctt ctattgcttt agcgcattgg 240
gtctgtagtg ctcagtctct ggtgtcctta gataataaag atatgagatt aacatagaaa 300
taaagatata aaagggctgg atgtatagtt tagtggtcca gtgtatgcct agtatgtgaa 360
aagccttctg ttcaacctct agcaatagaa aaacaagata tattctcggt ggggctgtta 420
atattgaatt ctcataaaat ctttaatata tttagtatgc ctattatgtt gttatatttt 480
agttctttag ctaatcaaaa tgcattattg atctttcttt gtcttttttt ggccaacact 540
ctattccagt ctttgaaaaa gtcctttaaa agagttaatc agtataatta aatgagtcag 600
gaagtatgtg agggttattt tacaaccaga gggaattact atagcaacag ctgattagaa 660
tgatctcaag aaaaagccca ttctgtcttt ttgcaccatg cacctttcag tggctccatt 720
cagatggaga ggcaaacaga gcaatggctc tcagagggcc tattttccct ttgaacattc 780
attatccata tccctggtgc acagcagtgc atctgggggc agaaactgtt cttgctttgg 840
aaacaatgct gtctatgtca tactggataa agaagctcat taattgtcaa cacttatgtt 900
atcataatgg gatcagcatg tacttttggt tttgttccag agtctatcac cggaaagaac 960
aagccggttt actctgaccc atttcactga catttctctt gtctcctctg tgcccagggt 1020
gctggagaat ccaaatgtcc tctgatggtc aaagtcctgg atgctgtccg aggcagccct 1080
gctgtagacg tggctgtaaa agtgttcaaa aagacctctg agggatcctg ggagcccttt 1140
gcctctgggt aagcttgtag aaagcccacc atgggaccgg ttccaggttc ccatttgctc 1200
ttattcgtgt tagattcaga cacacacaac ttaccagcta gagggctcag agagagggct 1260
caggggcgaa gggcacgtat tgctcttgta agagacacag gtttaattcc tagcaccaga 1320
atggcagctc ataaccatct gaaactcaca gtcttaggag atctgggtat ctgacattct 1380
cttctaccca ccatgtgtgt ggtgcacaaa ttcacatgca ggcatcaaat cttataaaca 1440
acaacaaaaa accaacaaac ctggtagcaa aagaagatta gaaggttaaa catatgagcc 1500
gagagctttt gttttgtttt gttttgtttt gttttgttta catttcaaat gttatcccct 1560
ttctcggtcc ccctccccaa accctctacc ccattctctc ctccccttct tctatgaggg 1620
tgttccccac caacccactc ccaccttcct gctctcgaat tcccctatac tgggacatca 1680
agccttcaca gaatcaaggg cctctcctcc cattgatgcc cgacaatgtc atcctctgct 1740
acctatgtgg ctggagccat gggtcccttc atgtatcctc cttggttggt ggtttagtct 1800
ctgggaggtc tgggggatct ggttgattga tattattgtt cttcctatga gattgcaaac 1860
cccttcagct ccttcggtcc tttaactcct ccactgggga ccccgagctc agtccaatgg 1920
ttggctgtga gcatccacca gcagaggcct tttttttttt ttttaacaaa gctgctttat 1980
tatgttgctt agagcatgac caggaaccag agcacagtcc aagactgaag ggaggaaaag 2040
ggggggagtc aataacccca ctgtttcata gtggtttgca acccttttat atcacagccc 2100
actttaggca aataatgaaa attatagtct ccagggacag agaagatggt gcaggaagtg 2160
aagtgcctgc tcagaaaatg ggggcttgaa tgtgagttcc cagactctgt gtaagatgcc 2220
cagcatcgaa gtgcatgctt ataacaccag cctggaggta gaagcttaga aacaggggta 2280
ccctgaagtt gcttgttcac cagtgtccct gaatgggtag gtgcatgttt ggtgagagac 2340
cctgtctcaa aaatcaaggt gtaggataat tgaaaatacc tagctttgag cttagatcat 2400
gcaaatgtgt acacacactc acacacacca cacacacaaa aaaatgcaga gacagagaga 2460
tacagagaga cagagagata cagagacaga gacagagaga aaaggagaaa gtaaaaaaca 2520
aataatttaa agacccatgg ccacaaagag gctcaaagac aagcacgtat aaaaccatac 2580
acatgtaatt ttaggagttt tcagattccc tggtacccgt gggtgatgca caagctttga 2640
atcccagtct taaaatctta cgaagaacgt gttcgtgtgt gctaatttat tgatgagagg 2700
aaaggaattg acaaagtgcc cttccggagc ttcctgcatt acccagactc agggtttttt 2760
taaatgtaca ctcagaacag agtagctctg tgcaagggta gcaaccacga agcttaataa 2820
gaaacatatc gtgagagatc tgcaaggcaa atctaggggc tgaccaatct cacagtcacc 2880
cactagcatg tcaacacaac ttcccacctg tgctagccac ttagcaattt tgtgttgttc 2940
tgttttgttt ttgtttttaa caaagcaatt tcaaagagat ttctaattca tctaaacaaa 3000
caaaccaaaa ggaaaacagc aaagacgccc tgagcactta gcagagcagc tatgcagtta 3060
tgactcctgg gtggagactt tatatcaggc ttcaactgaa tacctagaac ctactagtgc 3120
tcttcatcaa tccttgggaa ggtcattttc ttttggtgct gttttgagtt tctatttgtt 3180
aatgtcttca taattataca cgtgttgagc acagcatgca aagtgattag gggaatctag 3240
ttggagtgga atggataccc aaatattcag actttcttgt gactcttctt tcttgtaccc 3300
acatcaaaaa aaaaaaaaat ggagatgaga catggtcaga gtcactaaaa ccagctgcta 3360
cttttaatta cgtggggagc agtttctaac attgccatta ttgaactgat gctgcctggg 3420
tggaaatgga aatcacttag tatttcttgt tggcaaagaa ttactgaatg gattaaattt 3480
ccaaagggag aagtcagtta caagtctttt ctttgtttat taggctttct gctatgataa 3540
attacactac ttccagaagt tacccttagg ccatgggaca ctggactatc actctgctgt 3600
cacaagagat tacagagtta gtcaaggcag cttgtgacac cttcagggac tgtcataaac 3660
ttccagcaag tcattaatcc tgaatgcaat actgtgtgtg tgtgtctatg tgtgtttgta 3720
tgtctgtgtg tgtcttatgt ctgtgtctct gtgtgtgtgt gtgtttgtgt gtgtgtgtgt 3780
atgtatgcct gtgtgtgtct tatgtctgtg tttgtgtgtc tgtgtgtgtc ttatgtctgt 3840
gtttgtatgt ctgtgtgtgt ctgtgtgtgt cttatgtctg tgtctctgtg tgtgtgtgtg 3900
tgtatgtatg tatgtatgta tgtatgtgta tgtgtttgca tctctctgtg tgtctgcgct 3960
tatatatttg tgtatgtgtt tatgtgttcg cctttgtgcg ttgttgggga ttgaatccag 4020
gggaatacaa atgttaagaa agaacgttac cactaagctt cacctgtagg ccttaaagct 4080
tttctttctt ttaaaaattg taattaattc attttcagtc aggatctcca cacctcgtcc 4140
ctgctgctct agaactcact atttaaacac aatcgccctc aaacctgcag caaccctccc 4200
gcctctaccc tgcgagcact agaataataa caggtgaccc cacacgccta gattaagacc 4260
tttaaggtaa acattttact atattttagt ctcataagac aagatgctac aataaagctg 4320
tacataaagt tccctcgaat ttcttgctat tttaactcaa acataaggat ttcctccttt 4380
ttgattcagg taacagaaaa aatacacagg tacatacatg tacacacatg aacacacacg 4440
catcacaacc acatatgcgc acgcttgtgt gatctatcat ttaccatgcc actgaactct 4500
tctttcccca taaattcctc tggacttgtg tgccctccag gaagaccgcg gagtctggag 4560
agctgcacgg gctcaccaca gatgagaagt ttgtagaagg agtgtacaga gtagaactgg 4620
acaccaaatc gtactggaag acacttggca tttccccgtt ccatgaattc gcggatgtaa 4680
gtggacacac caagttgttt ggattttgtt tttagtctca ggaaattccc ttcgctcttg 4740
ctgtacgatg ggcatgagtg gaaagtagat tccacagcca gaatccacag tgctgggaaa 4800
gcaagccttc tgaatttttc taaaactcat ttagcaacat ggcctgaacc tgttcacact 4860
gcttatggtc agctaactat atttatgtaa atattcattt ctctgttgag gaaatgttag 4920
tatttgcttt tgaggcaacc tccagatacc atggagggca tgtcatagtc aaagagaggg 4980
ctccctatgg tatttctcta aattctggca tttcctttat tccaaagcac atctagtgtc 5040
cccagaagtt tgggtagaca attcttggca acacagagaa ttacaacatg ttcaaaaccc 5100
aacagcttaa tatctaaatc atcaagcaaa catcacatgg caaagggatt tctgaatcaa 5160
aactgtttca tccttatgat caacctatgg aggtctagcc tcgacttaca cccattttac 5220
caataagcta agagaagcta agttcctcat caaggacaca aggctagcat gtgtgagcaa 5280
gtgacagagt tgccctctat gttggttagt gtgccttagc cagtgtctca gtaagaaatg 5340
gagctaaatc aaaacccaag gccaacagcc aaaggcacat gagtaacctt tgcttggcac 5400
tgggctcagt ttccctggct cctctcagtc ctcagttcac agaggcagct gtcatgcaaa 5460
tagaatccaa gcttgttggt cagacctgga gataacaaat tccatcaaaa atagctcctc 5520
atgtgaccta gtttgctgtc tgttgctatg atacacacca tgaccgaaaa gcaaccctgg 5580
ggagagaagg gtttatttca tcttacagct tacagttcac catggaggaa agccaggtgg 5640
gaacctggaa gtggaaattg aagcagagac cagaaaggaa tgctgtttac tggctggctt 5700
agctcctttt cttatacagc ttaggtctat gtgcccaggg gatggtactg ccgagcatag 5760
gctgagcccg cctacatcaa ccattagtca aaaaaaggtc catagacttg cctacaggcc 5820
aatctcatgg aggcaatacc ccagtggagg gtccctcttc gcaggttact ctagtttgtg 5880
tcaagttgac aaaacctaac cacaaagcac aaacagggtc tgcccttgtg gcttagccat 5940
ggatgacact ctcagatgat ggtgttacca gacaaaccag aggggctcac caagagtctg 6000
ccacctacca aggtagtact ctactcctca ctgggcacca acacccatat tagctgggcc 6060
agtacaggac ccttgctgtt tcctgcatga attgtccata gaccctgggt ctcagcctgc 6120
cgggagtacc tgtaagtagt cgcctcaaac acattattcc tgttggaaga cttgtctgat 6180
tctcttttag aactcaatca acaaacgttt ttattttgtt ttggcttttt ggagacaaga 6240
tctctcatag gccagcctga cttgaatgta gctgaggatg acctgtgctg ctaatcttct 6300
cgcctcttcc tcccaagtgg taggataata ggcataagac accacagcag ttttactcca 6360
taccagggct ctgaacccag actttaaaca ctctatcaac tgattcacat tcccacccca 6420
tcattcaaca aacatttgaa aaataaaacc cttctgcctt gagcactctg ctaaatacag 6480
cctttgagtg cggagtattt cctcacaacc agggtccaag atgaccccat catacatacc 6540
acggaaaatt aggagatgtt tttaggtctc tttgcttggg gtaattttta tgtgtgtgtg 6600
tacacagccc tgtgcgtgtg tgtgtgtgtg tgtgtgtgtg tgtacaggca cacacgtgta 6660
tgcatgtaga ggctacataa aaaccttagg tgtcattctc aggcactctg ttcacccctt 6720
cacacagccc gaacacacaa aatttgaggc attagcctgg agctcaccag ttaggctaga 6780
ctgacttgcc agcagacccc aggctgtctc catctcccca gctctgggat tacaaactct 6840
atcataccag acatttttat acatattctg agcataaaat tcatgtcttc aggctaacaa 6900
gtcaagagct taaatgactg agctctctta cgtggtggat tttttttaaa actacataat 6960
atcttttttt tttttttcac ttctggggaa gaaacaaatg agcctgagtg acaatgcgac 7020
agaaaagaaa ttttgaggag tgtgtgtgtc tgtgtgtgtg gtggcacatg cctctcatct 7080
aatgctagag gctacagtag aatgctcctg aattagtggc cagccaaggc caagggctag 7140
ggttgtaact cagtggcaga gggcttgcct agcattcgca ggatttgatc catagcgcta 7200
taaataataa taaataaata caacagtcta agatgattct ccctttcatt tatctggatg 7260
ttatttttgt gttagtttta ctctgtcatc caatcattgt ttgccctata tttggacatt 7320
taaaaaaaat ctttattcca agtgtgttca aagctgtatc caaaacctgt ccaccaaatg 7380
agtccaatga catacatctt ctatattacc atctgttcca gatttggctg actcccggca 7440
cctgggctgt tgctgcaccc atgtctcaga tagtctagtg atttgagaag tgactagtaa 7500
ttgcaaaatc cagactttgt ccagaaactt ctatgagctc caaaactttc atttacattt 7560
ctgccagcca caaaccgctt gtgttgtgga gagaaccctg tgatgtcttc ccacagcatc 7620
tcagccttgt ttcttccctt aaaatattca tcttttcaca ttagaacatg caaagggaca 7680
gtgggagcga aacccctgga ctgggacgca cgaagccttc ctttctggtc aggctctcac 7740
tgtagaaact taggccggtt tcagcatgca gtctgctgga gaatggctcc tgccaacatt 7800
ccaggtctgg aagtttgtag tggagttgtt gataaccact gttcgccaca ggtcttttgt 7860
ttgtgggtgt cagtgtttct actctcctga cttttatctg aacccaagaa agggaacaat 7920
agccttcaag ctctctgtga ctctgatctg accagggcca cccacactgc agaaggaaac 7980
ttgcaaagag agacctgcaa ttctctaaga gctccacaca gctccaaaga cttaggcagc 8040
atattttaat ctaattattc gtcccccaac cccaccccag aggacagtta gacaataaaa 8100
ggaagattac cagcttagca tcctgtgaac actttgtctg cagctcctac ctctgggctc 8160
tgttagaact agctgtctct cctctctcct aggtggtttt cacagccaac gactctggcc 8220
atcgccacta caccatcgca gccctgctca gcccatactc ctacagcacc acggctgtcg 8280
tcagcaaccc ccagaattga 8300
<210> 21
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 21
cacagacaat cagacgtacc agta 24
<210> 22
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 22
ccagctttgc cagtttacga 20
<210> 23
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 23
ttggacggtt gccctctt 18
<210> 24
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 24
gatggcttcc cttcgactct tc 22
<210> 25
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 25
cactgacatt tctcttgtct cctct 25
<210> 26
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 26
gggctcacca cagatgagaa g 21
<210> 27
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 27
cactgttcgc cacaggtctt 20
<210> 28
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 28
gctcagccca tactcctaca 20
<210> 29
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 29
gcccaggagg accaggat 18
<210> 30
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 30
ggcaacttgc ttgaggaaga 20
<210> 31
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 31
gcagcaaccc agcttcactt 20
<210> 32
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 32
actgagctgg gacttgaac 19
<210> 33
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 33
tgcctcactc tgagaacca 19
<210> 34
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 34
ggtggagagg ctattcggc 19
<210> 35
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 35
ggccgtgcat gtgttcag 18
<210> 36
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 36
ggttcccatt tgctcttatt cgt 23
<210> 37
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 37
cccacactgc agaaggaaac ttg 23
<210> 38
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 38
ggttcccatt tgctcttatt cgt 23
<210> 39
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 39
ccagcttagc atcctgtgaa ca 22
<210> 40
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 40
ggcaacttgc ttgaggaaga 20
<210> 41
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 41
tgtggagttc agtagtgtgg ag 22
<210> 42
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 42
cactgacatt tctcttgtct cctct 25
<210> 43
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 43
gggacatctc ggtttcctga ctt 23
<210> 44
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 44
tccacactac tgaactccac aa 22
<210> 45
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 45
cggaacactc gctctacgaa a 21
<210> 46
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 46
gggccagctt cagacaca 18
<210> 47
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 47
cccagggtgc tggagaatcc aa 22
<210> 48
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 48
gccaagtgtc ttccagtacg at 22
<210> 49
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 49
gttccctttc ttgggttcag a 21
<210> 50
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 50
gatgctactg ctttggcaag atc 23
<210> 51
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 51
cctgagctgc taacacggtt 20
<210> 52
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 52
agctacagac catgcttagt gta 23
<210> 53
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 53
tgccagttta ggaggaatat gttc 24
<210> 54
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 54
ctgaggaaac agaggtacca gatat 25
<210> 55
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 55
agtcacacag ttctgtcaaa tcag 24
<210> 56
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 56
gaacacggcg gcatcag 17
<210> 57
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 57
tcctgtggga gggttctttg 20
<210> 58
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 58
ccctctctct gagccctcta 20
<210> 59
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 59
gctgcctaag tctttggagc t 21
<210> 60
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 60
ccctctctct gagccctcta 20
<210> 61
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 61
gagaggagag acagctagtt ctaac 25
<210> 62
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 62
agctacagac catgcttagt gta 23
<210> 63
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 63
gccctcttca tacaggaatc ac 22
<210> 64
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 64
cggacagcat ccaggactt 19
<210> 65
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 65
tcatgtaatc tggcttcaga gtggga 26
<210> 66
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 66
tgggaggcaa ttcttagttt caatgga 27
<210> 67
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 67
tcccaaaggt gtctgtctgc aca 23
<210> 68
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 68
ctcctttgcc tcgctggact gg 22
<210> 69
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 69
cggacagcat ccaggactt 19
<210> 70
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 70
agaaggagtg tacagagtag aactggaca 29
<210> 71
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 71
tgtttgtggg tgtcagtgtt tctactc 27
<210> 72
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 72
caccacggct gtcgtcagca a 21
<210> 73
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 73
cttgccaaag cagtagcatc cca 23
<210> 74
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 74
aggtcagaaa gcagagtgga cca 23
<210> 75
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 75
cccaggcaat tcctaccttc cca 23
<210> 76
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 76
tctgagcatt ctacctcatt gctttggt 28
<210> 77
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 77
aggctgtccc agcacctgag tcg 23
<210> 78
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 78
tgggcacaac agacaatcgg ctg 23
<210> 79
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 79
aaggctgctg atgacacctg gga 23
<210> 80
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 80
agattcagac acacacaact taccagc 27
<210> 81
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 81
agacctgcaa ttctctaaga gctccaca 28
<210> 82
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 82
agattcagac acacacaact taccagc 27
<210> 83
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 83
ttgtctgcag ctcctacctc tggg 24
<210> 84
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 84
aggtcagaaa gcagagtgga cca 23
<210> 85
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 85
ttgacatgtg tgggtgagag attttactg 29
<210> 86
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 86
cccagggtgc tggagaatcc aa 22
<210> 87
<211> 16
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 87
guuuuagagc uaugcu 16
<210> 88
<211> 67
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 88
agcauagcaa guuaaaauaa ggcuaguccg uuaucaacuu gaaaaagugg caccgagucg 60
gugcuuu 67
<210> 89
<211> 77
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 89
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcu 77
<210> 90
<211> 444
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 90
atggcttctc atcgtctgct cctcctctgc cttgctggac tggtatttgt gtctgaggct 60
ggccctacgg gcaccggtga atccaagtgt cctctgatgg tcaaagttct agatgctgtc 120
cgaggcagtc ctgccatcaa tgtggccgtg catgtgttca gaaaggctgc tgatgacacc 180
tgggagccat ttgcctctgg gaaaaccagt gagtctggag agctgcatgg gctcacaact 240
gaggaggaat ttgtagaagg gatatacaaa gtggaaatag acaccaaatc ttactggaag 300
gcacttggca tctccccatt ccatgagcat gcagaggtgg tattcacagc caacgactcc 360
ggcccccgcc gctacaccat tgccgccctg ctgagcccct actcctattc caccacggct 420
gtcgtcacca atcccaagga atga 444
<210> 91
<211> 444
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 91
atggcttccc ttcgactctt cctcctttgc ctcgctggac tggtatttgt gtctgaagct 60
ggccccgcgg gcaccggtga atccaagtgt cctctgatgg tcaaagttct agatgctgtc 120
cgaggcagtc ctgccatcaa tgtggccgtg catgtgttca gaaaggctgc tgatgacacc 180
tgggagccat ttgcctctgg gaaaaccagt gagtctggag agctgcatgg gctcacaact 240
gaggaggaat ttgtagaagg gatatacaaa gtggaaatag acaccaaatc ttactggaag 300
gcacttggca tctccccatt ccatgagcat gcagaggtgg tattcacagc caacgactcc 360
ggcccccgcc gctacaccat tgccgccctg ctgagcccct actcctattc caccacggct 420
gtcgtcacca atcccaagga atga 444
<210> 92
<211> 444
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 92
atggcttccc ttcgactctt cctcctttgc ctcgctggac tggtatttgt gtctgaagct 60
ggccccgcgg gtgctggaga atccaaatgt cctctgatgg tcaaagtcct ggatgctgtc 120
cgaggcagcc ctgctgtaga cgtggctgta aaagtgttca aaaagacctc tgagggatcc 180
tgggagccct ttgcctctgg gaagaccgcg gagtctggag agctgcacgg gctcaccaca 240
gatgagaagt ttgtagaagg agtgtacaga gtagaactgg acaccaaatc gtactggaag 300
acacttggca tttccccgtt ccatgaattc gcggatgtgg ttttcacagc caacgactct 360
ggccatcgcc actacaccat cgcagccctg ctcagcccat actcctacag caccacggct 420
gtcgtcagca acccccagaa ttga 444
<210> 93
<211> 4176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> прочий признак
<222> (1)..(3)
<223> Инициирующий кодон
<220>
<221> прочий признак
<222> (10)..(30)
<223> 5' NLS
<220>
<221> прочий признак
<222> (4126)..(4173)
<223> 3' NLS
<220>
<221> прочий признак
<222> (4174)..(4176)
<223> Терминирующий кодон
<400> 93
atggacaagc ccaagaaaaa gcggaaagtg aagtacagca tcggcctgga catcggcacc 60
aactctgtgg gctgggccgt gatcaccgac gagtacaagg tgcccagcaa gaaattcaag 120
gtgctgggca acaccgacag gcacagcatc aagaagaacc tgatcggcgc cctgctgttc 180
gacagcggcg aaacagccga ggccaccaga ctgaagagaa ccgccagaag aagatacacc 240
aggcggaaga acaggatctg ctatctgcaa gagatcttca gcaacgagat ggccaaggtg 300
gacgacagct tcttccacag actggaagag tccttcctgg tggaagagga caagaagcac 360
gagagacacc ccatcttcgg caacatcgtg gacgaggtgg cctaccacga gaagtacccc 420
accatctacc acctgagaaa gaaactggtg gacagcaccg acaaggccga cctgagactg 480
atctacctgg ccctggccca catgatcaag ttcagaggcc acttcctgat cgagggcgac 540
ctgaaccccg acaacagcga cgtggacaag ctgttcatcc agctggtgca gacctacaac 600
cagctgttcg aggaaaaccc catcaacgcc agcggcgtgg acgccaaggc tatcctgtct 660
gccagactga gcaagagcag aaggctggaa aatctgatcg cccagctgcc cggcgagaag 720
aagaacggcc tgttcggcaa cctgattgcc ctgagcctgg gcctgacccc caacttcaag 780
agcaacttcg acctggccga ggatgccaaa ctgcagctga gcaaggacac ctacgacgac 840
gacctggaca acctgctggc ccagatcggc gaccagtacg ccgacctgtt cctggccgcc 900
aagaacctgt ctgacgccat cctgctgagc gacatcctga gagtgaacac cgagatcacc 960
aaggcccccc tgagcgcctc tatgatcaag agatacgacg agcaccacca ggacctgacc 1020
ctgctgaaag ctctcgtgcg gcagcagctg cctgagaagt acaaagaaat cttcttcgac 1080
cagagcaaga acggctacgc cggctacatc gatggcggcg ctagccagga agagttctac 1140
aagttcatca agcccatcct ggaaaagatg gacggcaccg aggaactgct cgtgaagctg 1200
aacagagagg acctgctgag aaagcagaga accttcgaca acggcagcat cccccaccag 1260
atccacctgg gagagctgca cgctatcctg agaaggcagg aagattttta cccattcctg 1320
aaggacaacc gggaaaagat cgagaagatc ctgaccttca ggatccccta ctacgtgggc 1380
cccctggcca gaggcaacag cagattcgcc tggatgacca gaaagagcga ggaaaccatc 1440
accccctgga acttcgagga agtggtggac aagggcgcca gcgcccagag cttcatcgag 1500
agaatgacaa acttcgataa gaacctgccc aacgagaagg tgctgcccaa gcacagcctg 1560
ctgtacgagt acttcaccgt gtacaacgag ctgaccaaag tgaaatacgt gaccgaggga 1620
atgagaaagc ccgccttcct gagcggcgag cagaaaaagg ccatcgtgga cctgctgttc 1680
aagaccaaca gaaaagtgac cgtgaagcag ctgaaagagg actacttcaa gaaaatcgag 1740
tgcttcgact ccgtggaaat ctccggcgtg gaagatagat tcaacgcctc cctgggcaca 1800
taccacgatc tgctgaaaat tatcaaggac aaggacttcc tggataacga agagaacgag 1860
gacattctgg aagatatcgt gctgaccctg acactgtttg aggaccgcga gatgatcgag 1920
gaaaggctga aaacctacgc tcacctgttc gacgacaaag tgatgaagca gctgaagaga 1980
aggcggtaca ccggctgggg caggctgagc agaaagctga tcaacggcat cagagacaag 2040
cagagcggca agacaatcct ggatttcctg aagtccgacg gcttcgccaa ccggaacttc 2100
atgcagctga tccacgacga cagcctgaca ttcaaagagg acatccagaa agcccaggtg 2160
tccggccagg gcgactctct gcacgagcat atcgctaacc tggccggcag ccccgctatc 2220
aagaagggca tcctgcagac agtgaaggtg gtggacgagc tcgtgaaagt gatgggcaga 2280
cacaagcccg agaacatcgt gatcgagatg gctagagaga accagaccac ccagaaggga 2340
cagaagaact cccgcgagag gatgaagaga atcgaagagg gcatcaaaga gctgggcagc 2400
cagatcctga aagaacaccc cgtggaaaac acccagctgc agaacgagaa gctgtacctg 2460
tactacctgc agaatggccg ggatatgtac gtggaccagg aactggacat caacagactg 2520
tccgactacg atgtggacca tatcgtgcct cagagctttc tgaaggacga ctccatcgat 2580
aacaaagtgc tgactcggag cgacaagaac agaggcaaga gcgacaacgt gccctccgaa 2640
gaggtcgtga agaagatgaa gaactactgg cgacagctgc tgaacgccaa gctgattacc 2700
cagaggaagt tcgataacct gaccaaggcc gagagaggcg gcctgagcga gctggataag 2760
gccggcttca tcaagaggca gctggtggaa accagacaga tcacaaagca cgtggcacag 2820
atcctggact cccggatgaa cactaagtac gacgaaaacg ataagctgat ccgggaagtg 2880
aaagtgatca ccctgaagtc caagctggtg tccgatttcc ggaaggattt ccagttttac 2940
aaagtgcgcg agatcaacaa ctaccaccac gcccacgacg cctacctgaa cgccgtcgtg 3000
ggaaccgccc tgatcaaaaa gtaccctaag ctggaaagcg agttcgtgta cggcgactac 3060
aaggtgtacg acgtgcggaa gatgatcgcc aagagcgagc aggaaatcgg caaggctacc 3120
gccaagtact tcttctacag caacatcatg aactttttca agaccgaaat caccctggcc 3180
aacggcgaga tcagaaagcg ccctctgatc gagacaaacg gcgaaaccgg ggagatcgtg 3240
tgggataagg gcagagactt cgccacagtg cgaaaggtgc tgagcatgcc ccaagtgaat 3300
atcgtgaaaa agaccgaggt gcagacaggc ggcttcagca aagagtctat cctgcccaag 3360
aggaacagcg acaagctgat cgccagaaag aaggactggg accccaagaa gtacggcggc 3420
ttcgacagcc ctaccgtggc ctactctgtg ctggtggtgg ctaaggtgga aaagggcaag 3480
tccaagaaac tgaagagtgt gaaagagctg ctggggatca ccatcatgga aagaagcagc 3540
tttgagaaga accctatcga ctttctggaa gccaagggct acaaagaagt gaaaaaggac 3600
ctgatcatca agctgcctaa gtactccctg ttcgagctgg aaaacggcag aaagagaatg 3660
ctggcctctg ccggcgaact gcagaaggga aacgagctgg ccctgcctag caaatatgtg 3720
aacttcctgt acctggcctc ccactatgag aagctgaagg gcagccctga ggacaacgaa 3780
cagaaacagc tgtttgtgga acagcataag cactacctgg acgagatcat cgagcagatc 3840
agcgagttct ccaagagagt gatcctggcc gacgccaatc tggacaaggt gctgtctgcc 3900
tacaacaagc acagggacaa gcctatcaga gagcaggccg agaatatcat ccacctgttc 3960
accctgacaa acctgggcgc tcctgccgcc ttcaagtact ttgacaccac catcgaccgg 4020
aagaggtaca ccagcaccaa agaggtgctg gacgccaccc tgatccacca gagcatcacc 4080
ggcctgtacg agacaagaat cgacctgtct cagctgggag gcgacaagag acctgccgcc 4140
actaagaagg ccggacaggc caaaaagaag aagtga 4176
<210> 94
<211> 1391
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> ПРОЧИЙ ПРИЗНАК
<222> (4)..(10)
<223> 5' NLS
<220>
<221> ПРОЧИЙ ПРИЗНАК
<222> (1376)..(1391)
<223> 3' NLS
<400> 94
Met Asp Lys Pro Lys Lys Lys Arg Lys Val Lys Tyr Ser Ile Gly Leu
1 5 10 15
Asp Ile Gly Thr Asn Ser Val Gly Trp Ala Val Ile Thr Asp Glu Tyr
20 25 30
Lys Val Pro Ser Lys Lys Phe Lys Val Leu Gly Asn Thr Asp Arg His
35 40 45
Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu Leu Phe Asp Ser Gly Glu
50 55 60
Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr
65 70 75 80
Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln Glu Ile Phe Ser Asn Glu
85 90 95
Met Ala Lys Val Asp Asp Ser Phe Phe His Arg Leu Glu Glu Ser Phe
100 105 110
Leu Val Glu Glu Asp Lys Lys His Glu Arg His Pro Ile Phe Gly Asn
115 120 125
Ile Val Asp Glu Val Ala Tyr His Glu Lys Tyr Pro Thr Ile Tyr His
130 135 140
Leu Arg Lys Lys Leu Val Asp Ser Thr Asp Lys Ala Asp Leu Arg Leu
145 150 155 160
Ile Tyr Leu Ala Leu Ala His Met Ile Lys Phe Arg Gly His Phe Leu
165 170 175
Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser Asp Val Asp Lys Leu Phe
180 185 190
Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu Phe Glu Glu Asn Pro Ile
195 200 205
Asn Ala Ser Gly Val Asp Ala Lys Ala Ile Leu Ser Ala Arg Leu Ser
210 215 220
Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala Gln Leu Pro Gly Glu Lys
225 230 235 240
Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala Leu Ser Leu Gly Leu Thr
245 250 255
Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala Glu Asp Ala Lys Leu Gln
260 265 270
Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu Asp Asn Leu Leu Ala Gln
275 280 285
Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu Ala Ala Lys Asn Leu Ser
290 295 300
Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg Val Asn Thr Glu Ile Thr
305 310 315 320
Lys Ala Pro Leu Ser Ala Ser Met Ile Lys Arg Tyr Asp Glu His His
325 330 335
Gln Asp Leu Thr Leu Leu Lys Ala Leu Val Arg Gln Gln Leu Pro Glu
340 345 350
Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser Lys Asn Gly Tyr Ala Gly
355 360 365
Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu Phe Tyr Lys Phe Ile Lys
370 375 380
Pro Ile Leu Glu Lys Met Asp Gly Thr Glu Glu Leu Leu Val Lys Leu
385 390 395 400
Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg Thr Phe Asp Asn Gly Ser
405 410 415
Ile Pro His Gln Ile His Leu Gly Glu Leu His Ala Ile Leu Arg Arg
420 425 430
Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp Asn Arg Glu Lys Ile Glu
435 440 445
Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr Val Gly Pro Leu Ala Arg
450 455 460
Gly Asn Ser Arg Phe Ala Trp Met Thr Arg Lys Ser Glu Glu Thr Ile
465 470 475 480
Thr Pro Trp Asn Phe Glu Glu Val Val Asp Lys Gly Ala Ser Ala Gln
485 490 495
Ser Phe Ile Glu Arg Met Thr Asn Phe Asp Lys Asn Leu Pro Asn Glu
500 505 510
Lys Val Leu Pro Lys His Ser Leu Leu Tyr Glu Tyr Phe Thr Val Tyr
515 520 525
Asn Glu Leu Thr Lys Val Lys Tyr Val Thr Glu Gly Met Arg Lys Pro
530 535 540
Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala Ile Val Asp Leu Leu Phe
545 550 555 560
Lys Thr Asn Arg Lys Val Thr Val Lys Gln Leu Lys Glu Asp Tyr Phe
565 570 575
Lys Lys Ile Glu Cys Phe Asp Ser Val Glu Ile Ser Gly Val Glu Asp
580 585 590
Arg Phe Asn Ala Ser Leu Gly Thr Tyr His Asp Leu Leu Lys Ile Ile
595 600 605
Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu Asn Glu Asp Ile Leu Glu
610 615 620
Asp Ile Val Leu Thr Leu Thr Leu Phe Glu Asp Arg Glu Met Ile Glu
625 630 635 640
Glu Arg Leu Lys Thr Tyr Ala His Leu Phe Asp Asp Lys Val Met Lys
645 650 655
Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp Gly Arg Leu Ser Arg Lys
660 665 670
Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser Gly Lys Thr Ile Leu Asp
675 680 685
Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg Asn Phe Met Gln Leu Ile
690 695 700
His Asp Asp Ser Leu Thr Phe Lys Glu Asp Ile Gln Lys Ala Gln Val
705 710 715 720
Ser Gly Gln Gly Asp Ser Leu His Glu His Ile Ala Asn Leu Ala Gly
725 730 735
Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln Thr Val Lys Val Val Asp
740 745 750
Glu Leu Val Lys Val Met Gly Arg His Lys Pro Glu Asn Ile Val Ile
755 760 765
Glu Met Ala Arg Glu Asn Gln Thr Thr Gln Lys Gly Gln Lys Asn Ser
770 775 780
Arg Glu Arg Met Lys Arg Ile Glu Glu Gly Ile Lys Glu Leu Gly Ser
785 790 795 800
Gln Ile Leu Lys Glu His Pro Val Glu Asn Thr Gln Leu Gln Asn Glu
805 810 815
Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly Arg Asp Met Tyr Val Asp
820 825 830
Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp Tyr Asp Val Asp His Ile
835 840 845
Val Pro Gln Ser Phe Leu Lys Asp Asp Ser Ile Asp Asn Lys Val Leu
850 855 860
Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser Asp Asn Val Pro Ser Glu
865 870 875 880
Glu Val Val Lys Lys Met Lys Asn Tyr Trp Arg Gln Leu Leu Asn Ala
885 890 895
Lys Leu Ile Thr Gln Arg Lys Phe Asp Asn Leu Thr Lys Ala Glu Arg
900 905 910
Gly Gly Leu Ser Glu Leu Asp Lys Ala Gly Phe Ile Lys Arg Gln Leu
915 920 925
Val Glu Thr Arg Gln Ile Thr Lys His Val Ala Gln Ile Leu Asp Ser
930 935 940
Arg Met Asn Thr Lys Tyr Asp Glu Asn Asp Lys Leu Ile Arg Glu Val
945 950 955 960
Lys Val Ile Thr Leu Lys Ser Lys Leu Val Ser Asp Phe Arg Lys Asp
965 970 975
Phe Gln Phe Tyr Lys Val Arg Glu Ile Asn Asn Tyr His His Ala His
980 985 990
Asp Ala Tyr Leu Asn Ala Val Val Gly Thr Ala Leu Ile Lys Lys Tyr
995 1000 1005
Pro Lys Leu Glu Ser Glu Phe Val Tyr Gly Asp Tyr Lys Val Tyr
1010 1015 1020
Asp Val Arg Lys Met Ile Ala Lys Ser Glu Gln Glu Ile Gly Lys
1025 1030 1035
Ala Thr Ala Lys Tyr Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe
1040 1045 1050
Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu Ile Arg Lys Arg Pro
1055 1060 1065
Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile Val Trp Asp Lys
1070 1075 1080
Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser Met Pro Gln
1085 1090 1095
Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly Phe Ser
1100 1105 1110
Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala
1115 1120 1125
Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser
1130 1135 1140
Pro Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys
1145 1150 1155
Gly Lys Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile
1160 1165 1170
Thr Ile Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe
1175 1180 1185
Leu Glu Ala Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile
1190 1195 1200
Lys Leu Pro Lys Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys
1205 1210 1215
Arg Met Leu Ala Ser Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu
1220 1225 1230
Ala Leu Pro Ser Lys Tyr Val Asn Phe Leu Tyr Leu Ala Ser His
1235 1240 1245
Tyr Glu Lys Leu Lys Gly Ser Pro Glu Asp Asn Glu Gln Lys Gln
1250 1255 1260
Leu Phe Val Glu Gln His Lys His Tyr Leu Asp Glu Ile Ile Glu
1265 1270 1275
Gln Ile Ser Glu Phe Ser Lys Arg Val Ile Leu Ala Asp Ala Asn
1280 1285 1290
Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys His Arg Asp Lys Pro
1295 1300 1305
Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu Phe Thr Leu Thr
1310 1315 1320
Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp Thr Thr Ile
1325 1330 1335
Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp Ala Thr
1340 1345 1350
Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp
1355 1360 1365
Leu Ser Gln Leu Gly Gly Asp Lys Arg Pro Ala Ala Thr Lys Lys
1370 1375 1380
Ala Gly Gln Ala Lys Lys Lys Lys
1385 1390
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Животные, отличные от человека, содержащие гуманизированный локус альбумина | 2020 |
|
RU2815514C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ХАРАКТЕРИЗУЮЩИЕСЯ ЭКСПАНСИЕЙ ГЕКСАНУКЛЕОТИДНЫХ ПОВТОРОВ В ЛОКУСЕ C9ORF72 | 2017 |
|
RU2760877C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ СКОНСТРУИРОВАННЫЙ ГЕН ANGPTL8 | 2017 |
|
RU2742354C2 |
| ОПОСРЕДОВАННЫЕ ГУМАНИЗИРОВАННЫМИ Т-КЛЕТКАМИ ИММУННЫЕ ОТВЕТЫ У НЕ ОТНОСЯЩИХСЯ К ЧЕЛОВЕКУ ЖИВОТНЫХ | 2016 |
|
RU2732628C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОЦЕНКИ ОПОСРЕДОВАННОГО CRISPR/CAS РАЗРУШЕНИЯ ИЛИ ВЫРЕЗАНИЯ И ИНДУЦИРОВАННОЙ CRISPR/CAS РЕКОМБИНАЦИИ С ЭКЗОГЕННОЙ ДОНОРСКОЙ НУКЛЕИНОВОЙ КИСЛОТОЙ IN VIVO | 2018 |
|
RU2782356C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС ASGR1 | 2018 |
|
RU2796949C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАПРАВЛЕННОЙ МОДИФИКАЦИИ ГЕНОМА | 2014 |
|
RU2725520C2 |
| ЭКСПРЕССИРУЮЩИЕ CAS МЫШИНЫЕ ЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ И МЫШИ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2782358C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАПРАВЛЕННОЙ МОДИФИКАЦИИ ГЕНОМА | 2020 |
|
RU2751237C1 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2799086C2 |

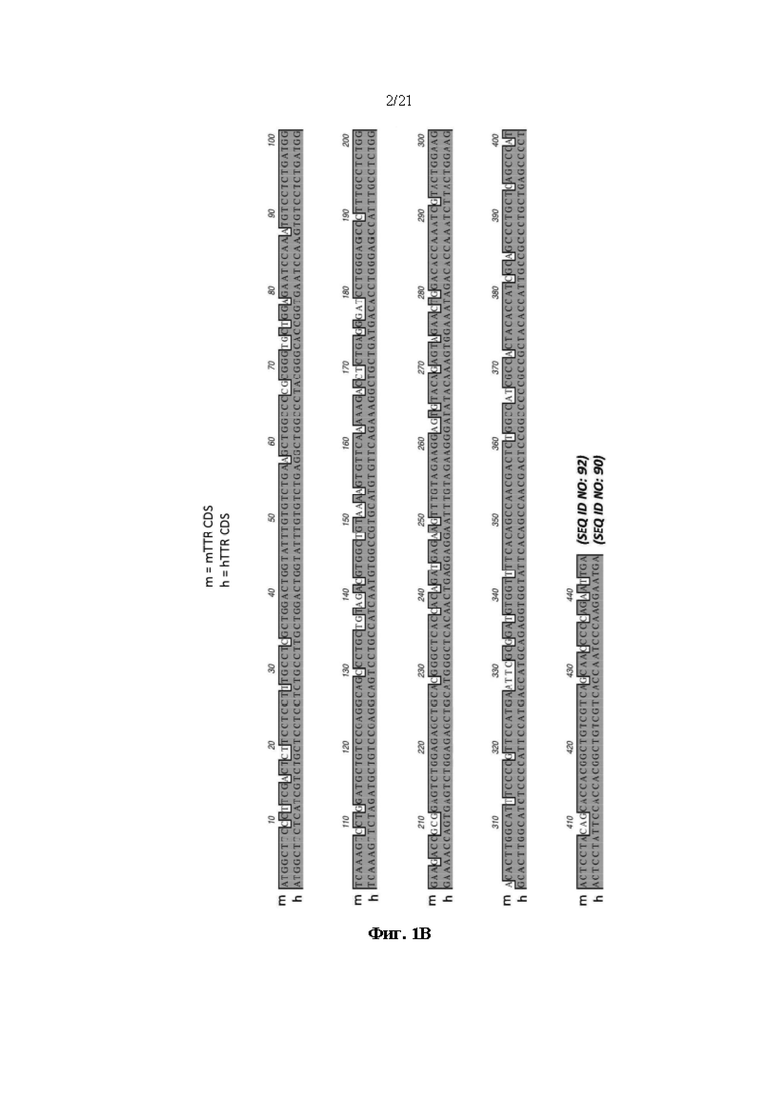

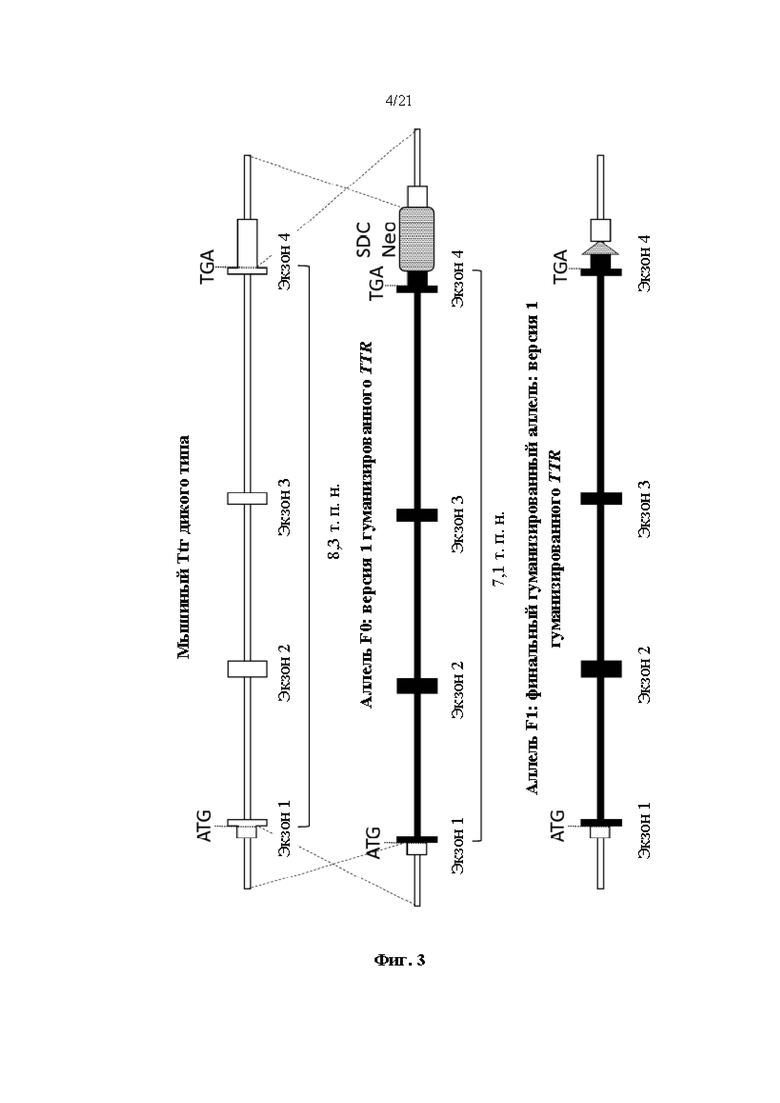
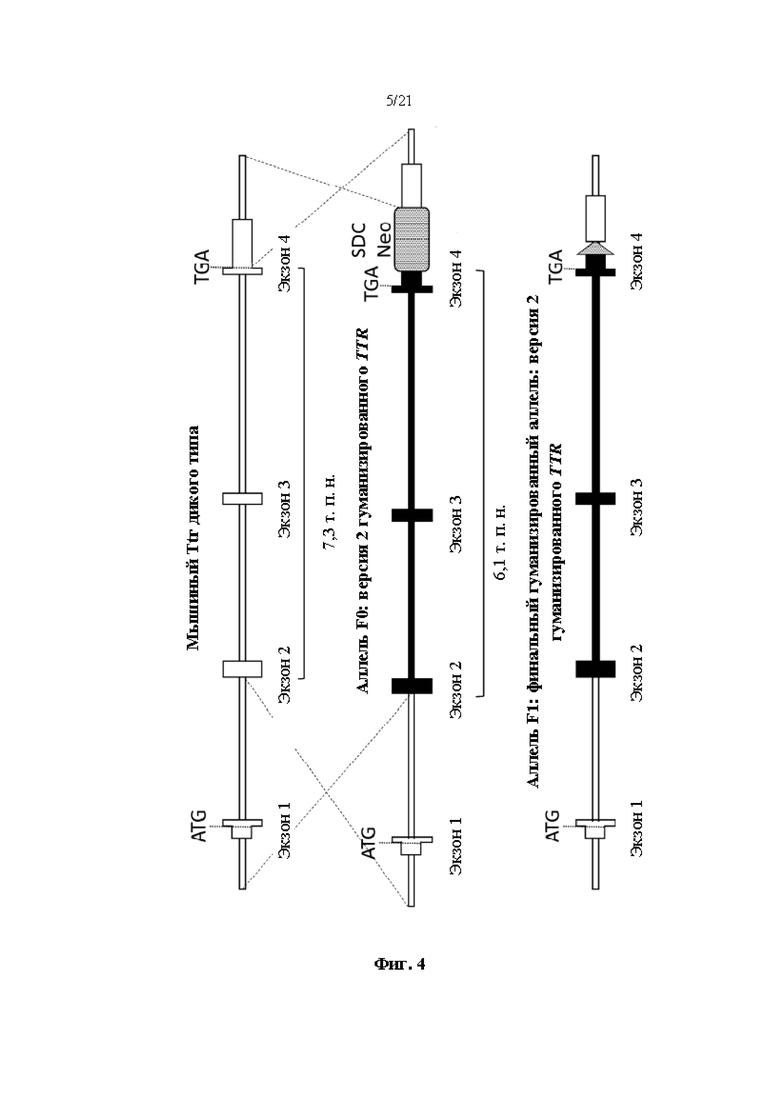
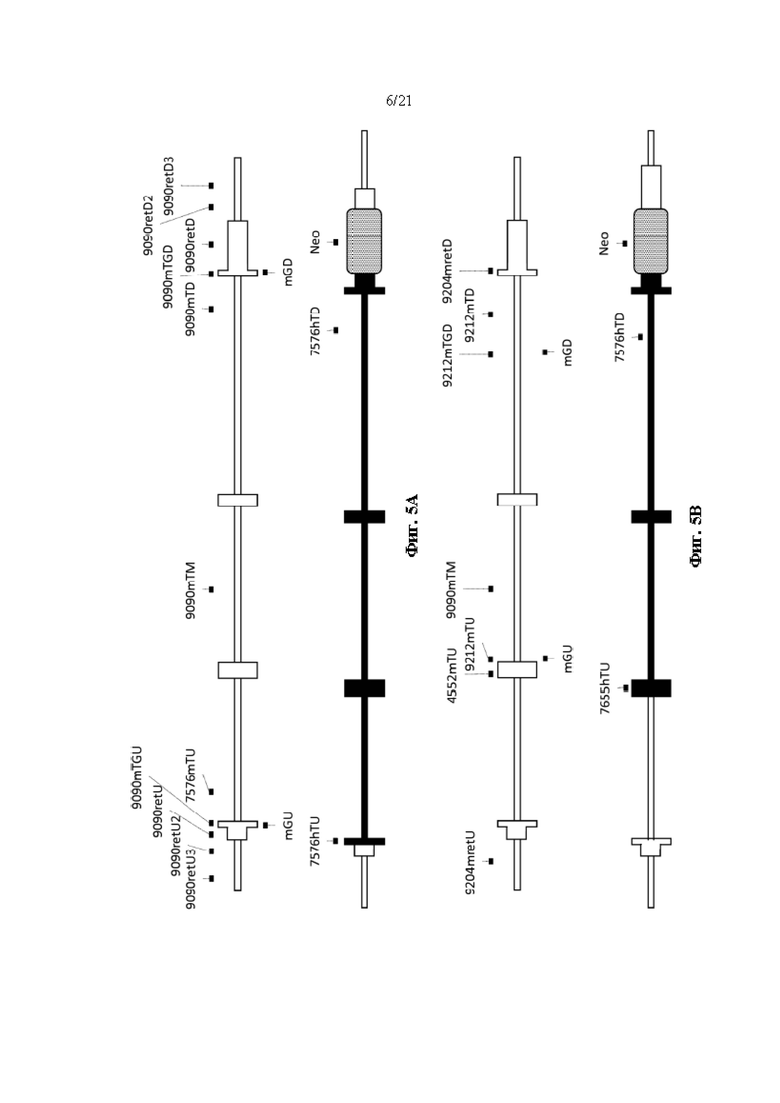
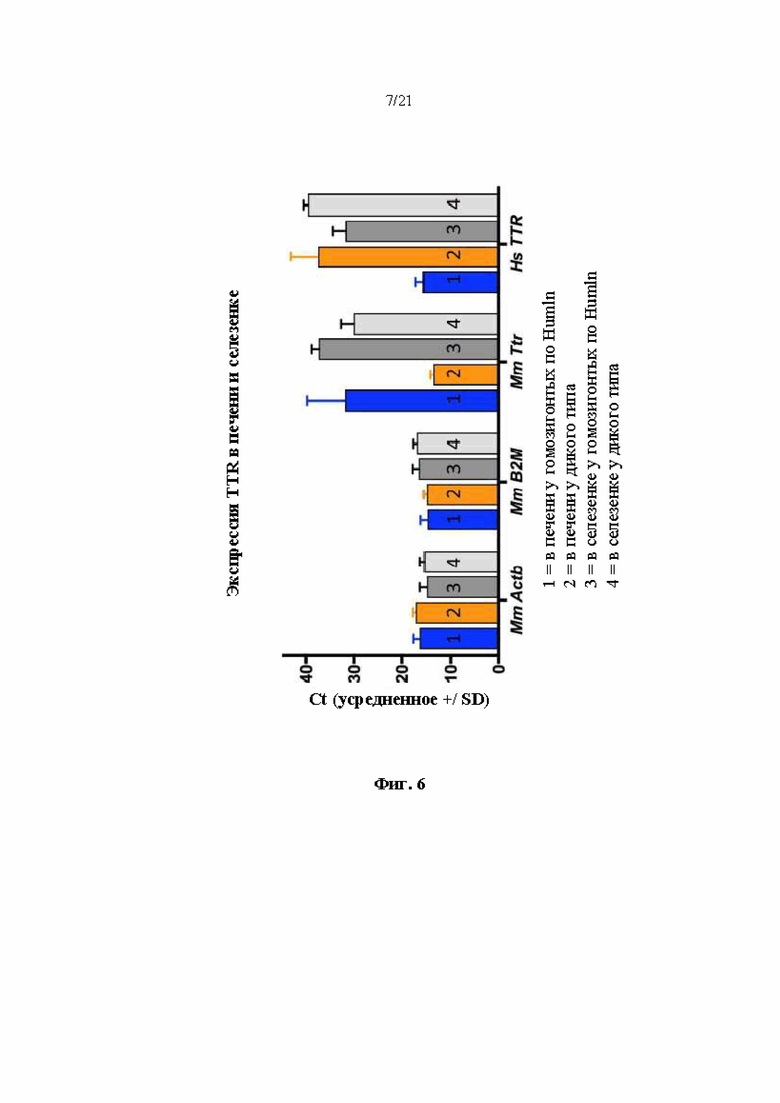
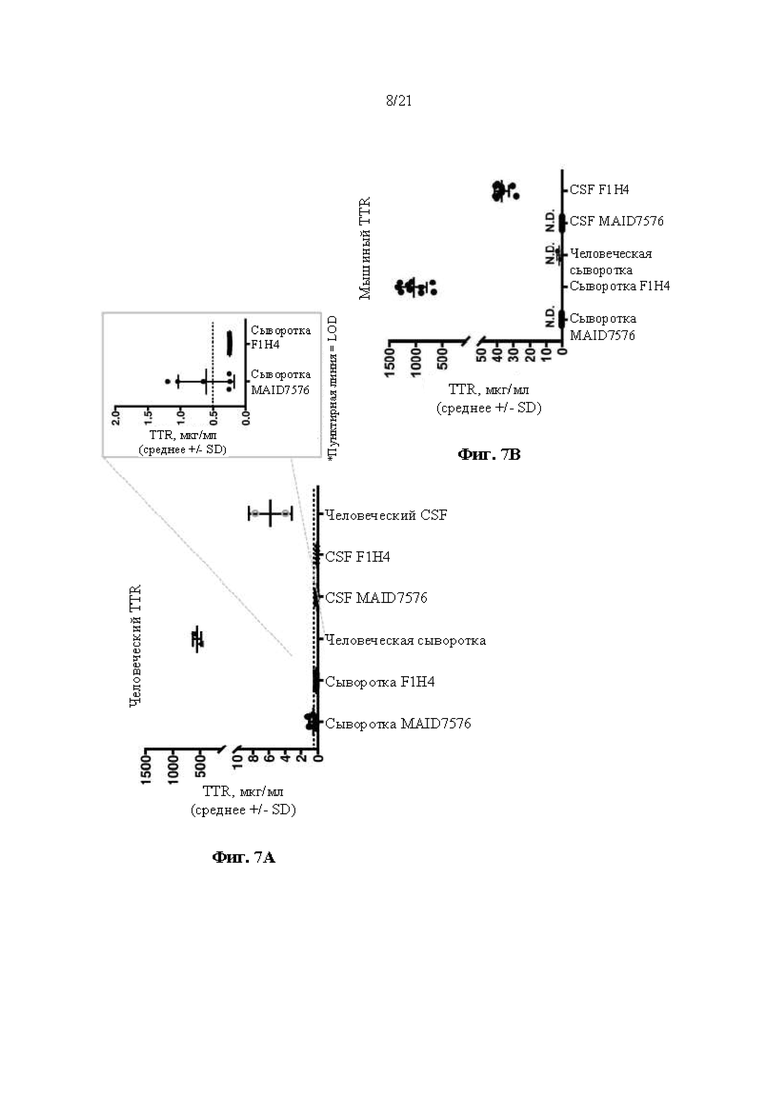
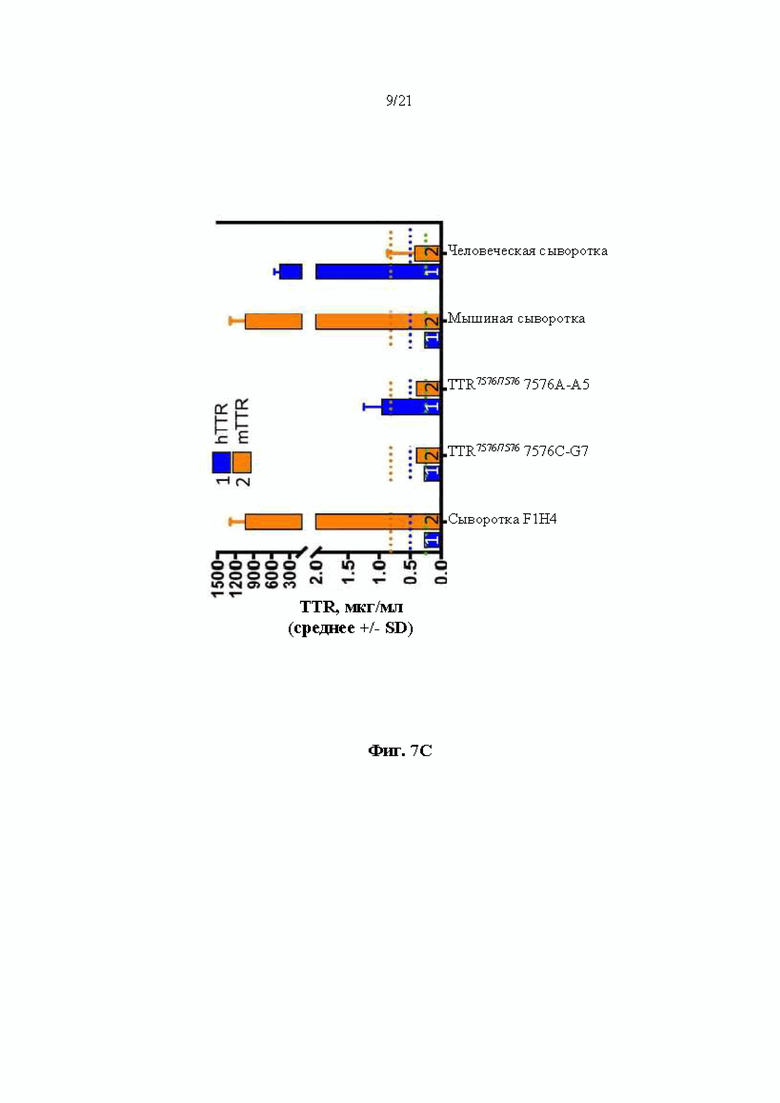
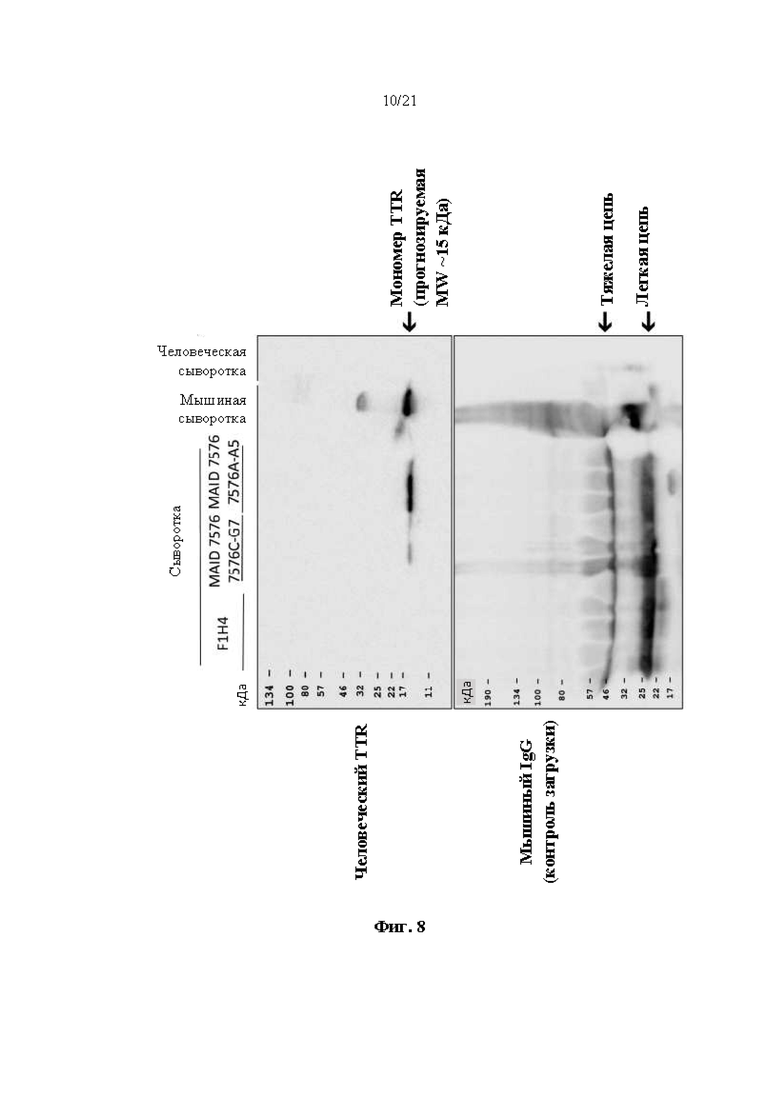
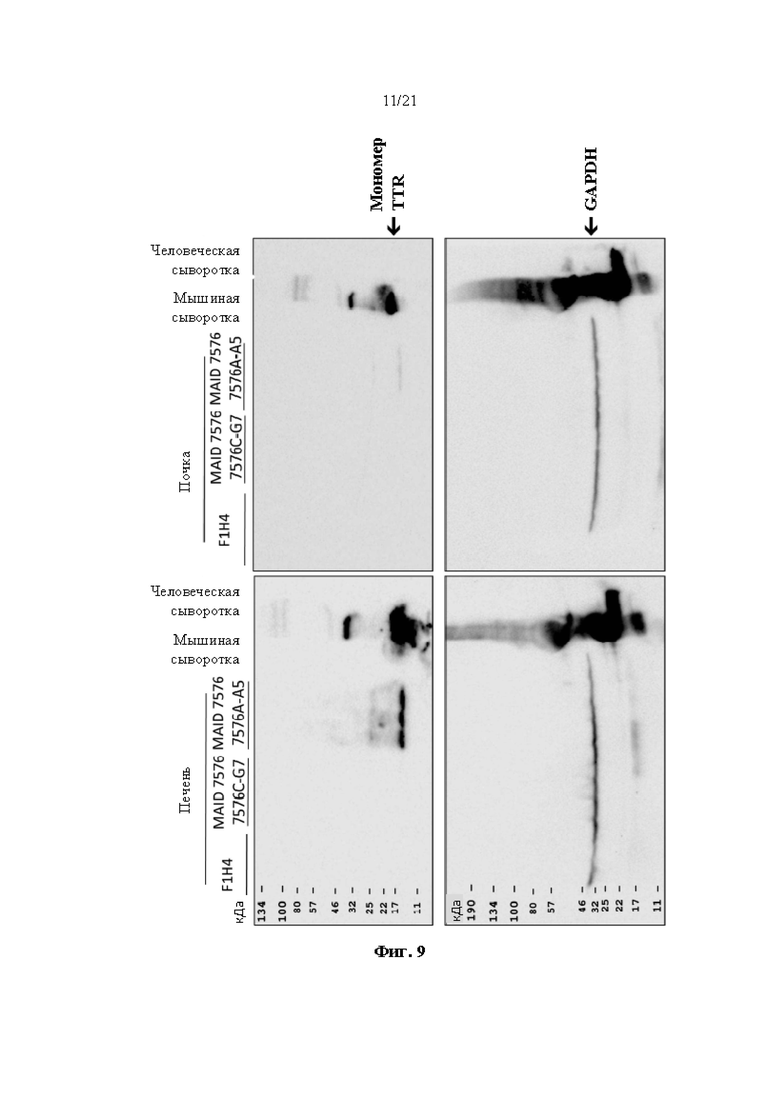

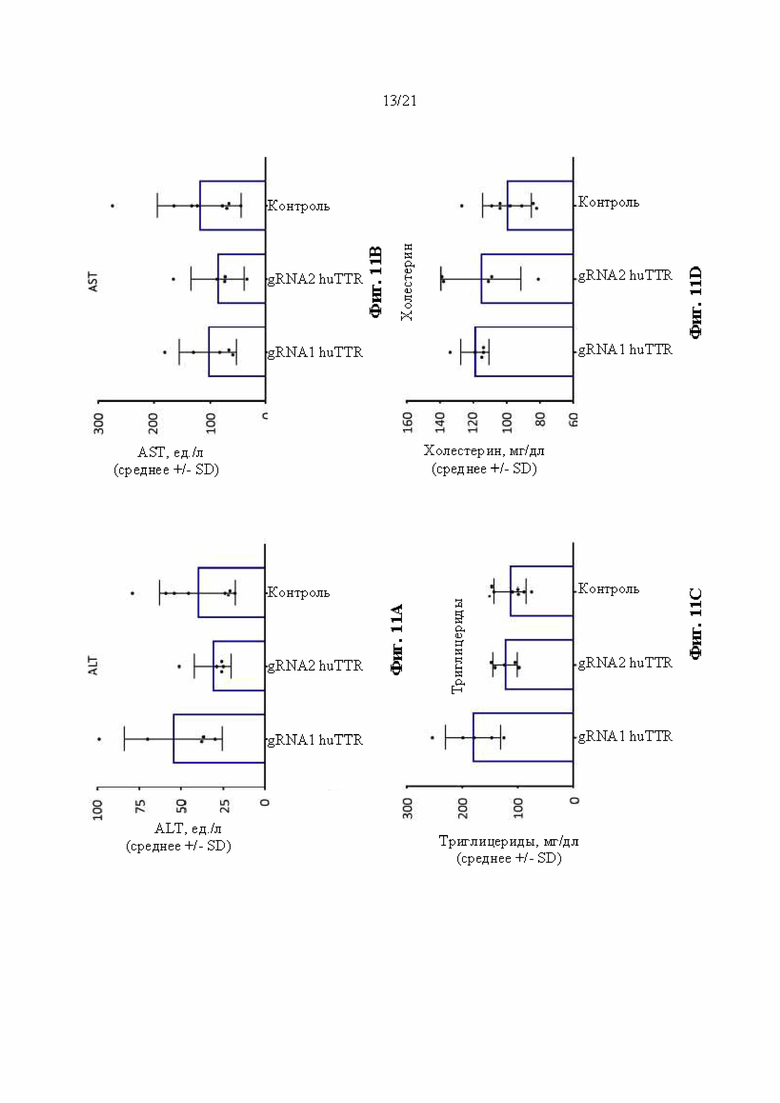
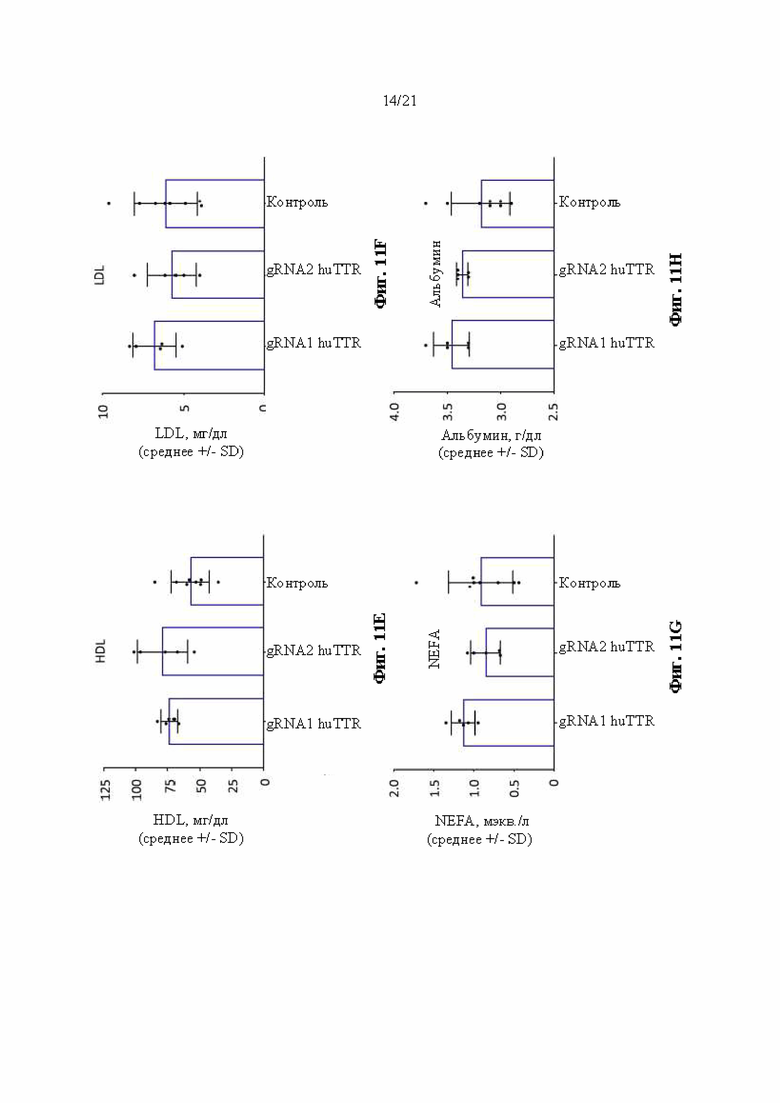
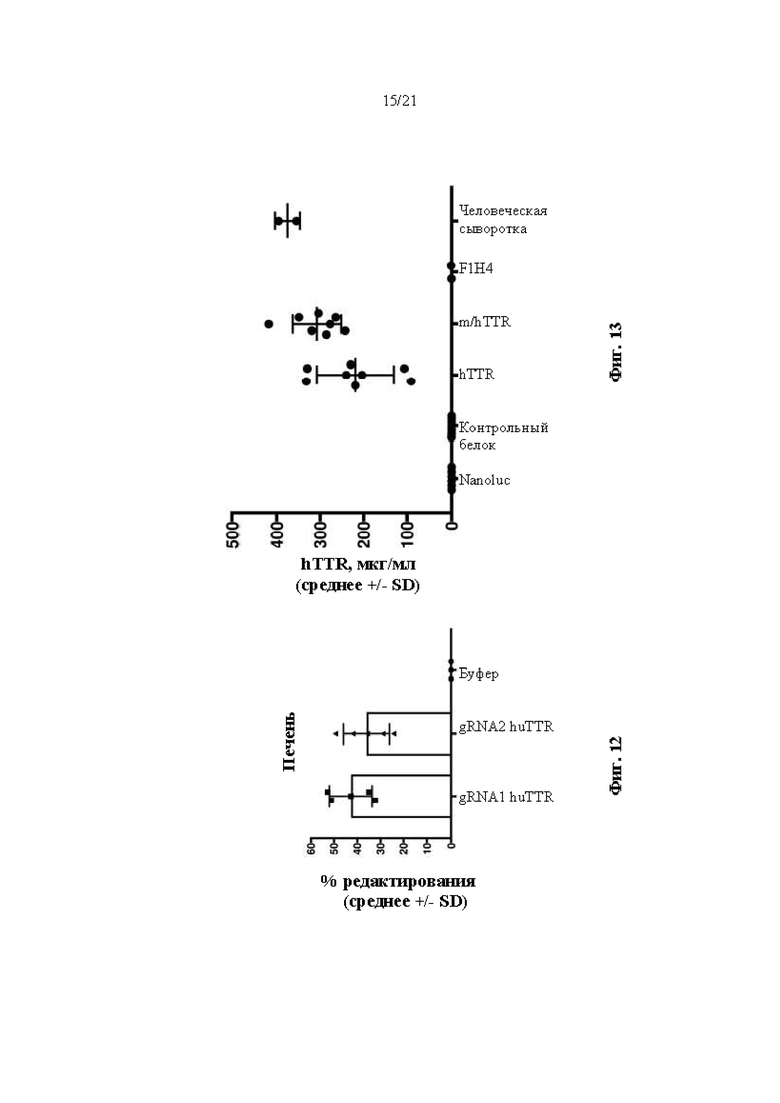
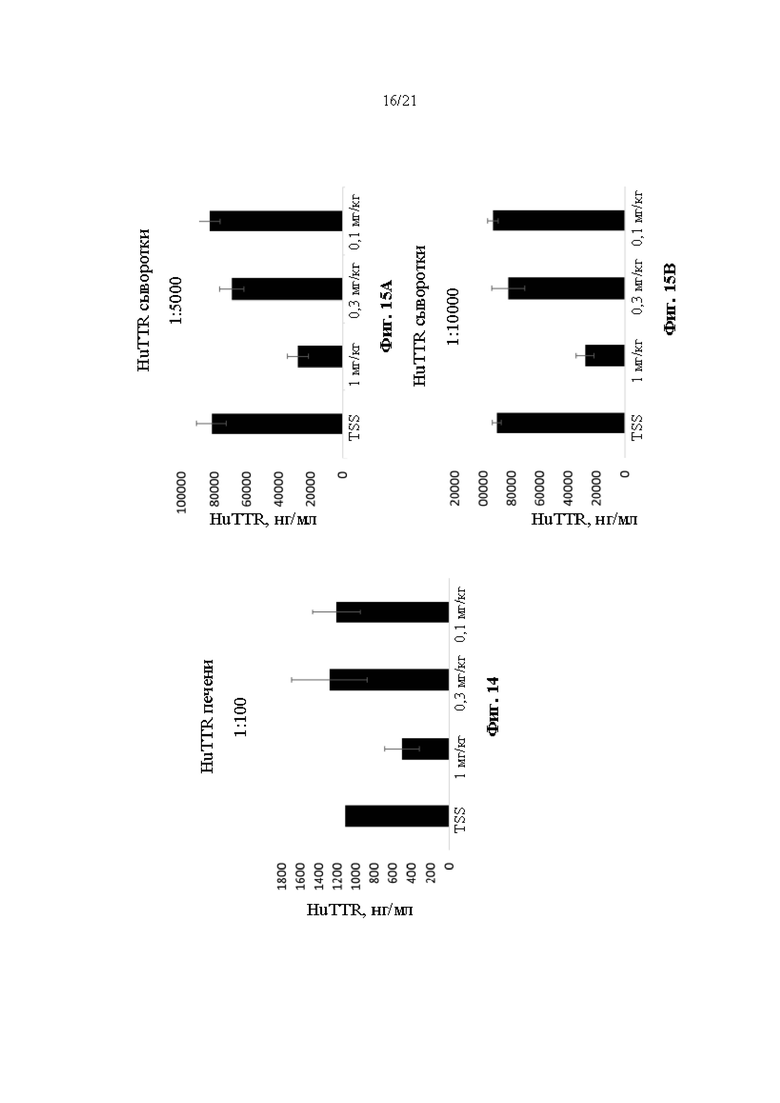
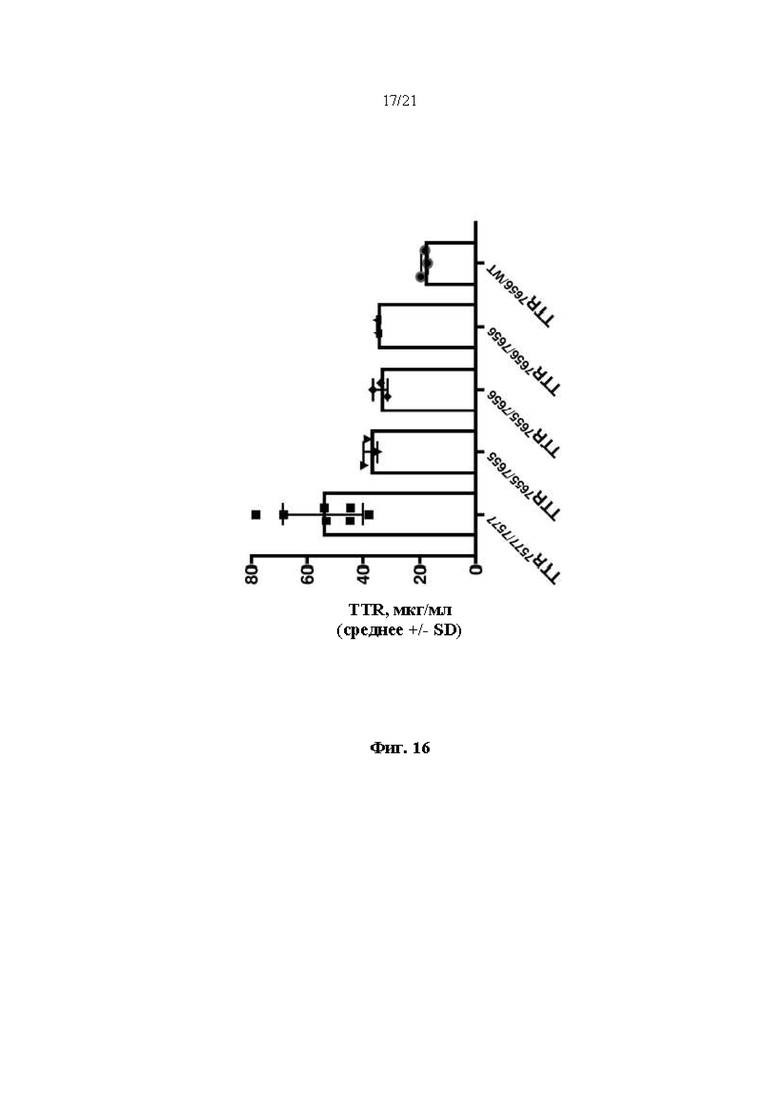
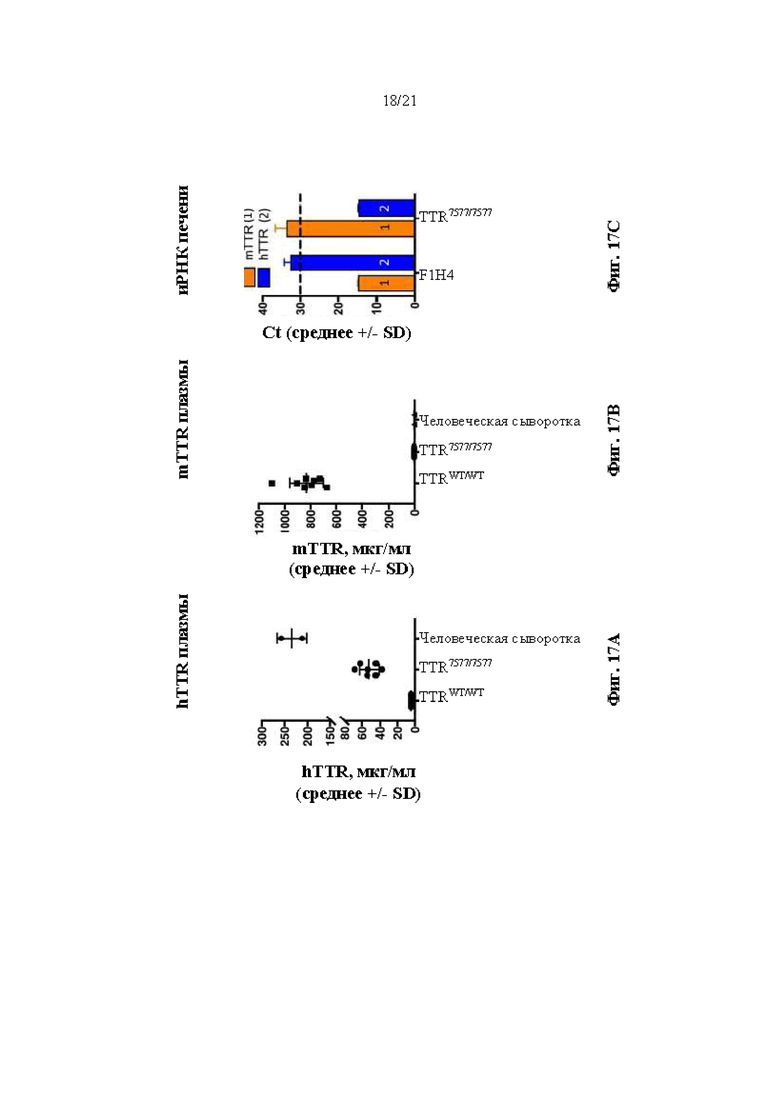

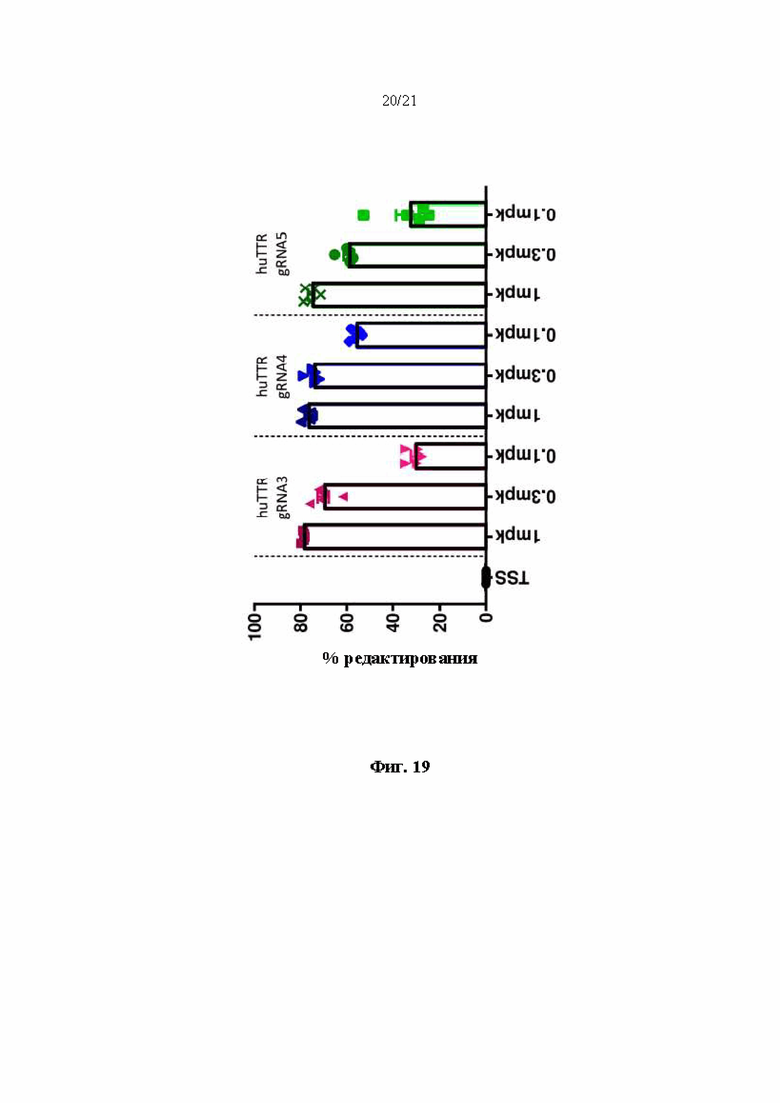

Изобретение относится к области биотехнологии, в частности к отличному от человека животному для экспрессии человеческого белка транстиретина (TTR) или химерного TTR белка, включающему генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность, его клетке, а также к способу его получения. Также раскрыты молекула нуклеиновой кислоты, кодирующая вышеуказанное антитело или его фрагмент, а также клетка и вектор, ее содержащие. Изобретение также относится к способу получения антитела против Абета или его антигенсвязывающего фрагмента. Изобретение эффективно для оптимизации активности нацеливающегося на человеческий TTR реагента in vivo, а также для оценивания активности нацеливающегося на человеческий TTR реагента in vivo. 8 н. и 18 з.п. ф-лы, 20 ил., 11 табл., 4 пр.
1. Отличное от человека животное для экспрессии человеческого белка транстиретина (TTR) или химерного TTR белка, включающего в себя один или несколько фрагментов человеческого TTR белка, причем отличное от человека животное включает в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
где отличное от человека животное представляет собой мышь или крысу,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
при этом вся кодирующая Ttr последовательность эндогенного Ttr локуса была делетирована и заменена соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью.
2. Отличное от человека животное по п. 1, в котором область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена человеческой TTR последовательностью, включающей в себя соответствующую человеческую TTR последовательность и человеческую TTR 3' нетранслируемую область, и
в котором 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
в котором эндогенный Ttr промотор не был делетирован и заменен соответствующей человеческой TTR последовательностью.
3. Отличное от человека животное по п. 2, в котором:
(i) человеческая TTR последовательность в генетически модифицированном эндогенном Ttr локусе включает в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 18;
(ii) генетически модифицированный эндогенный Ttr локус кодирует белок, включающий в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 1;
(iii) генетически модифицированный эндогенный Ttr локус включает в себя кодирующую последовательность, включающую в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 90; или
(iv) генетически модифицированный эндогенный Ttr локус включает в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 14 или 15.
4. Отличное от человека животное по любому из предыдущих пунктов, в котором генетически модифицированный эндогенный Ttr локус не включает в себя кассету селекции или репортерный ген.
5. Отличное от человека животное по любому из предыдущих пунктов, при этом отличное от человека животное является гомозиготным по генетически модифицированному эндогенному Ttr локусу.
6. Отличное от человека животное по любому из предыдущих пунктов, при этом отличное от человека животное представляет собой мышь.
7. Отличное от человека животное для экспрессии человеческого белка транстиретина (TTR) или химерного TTR белка, включающего в себя один или несколько фрагментов человеческого TTR белка, причем отличное от человека животное включает в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
где отличное от человека животное представляет собой мышь или крысу,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и при этом
генетически модифицированный эндогенный Ttr локус кодирует белок-предшественник TTR, включающий в себя сигнальный пептид, область эндогенного Ttr локуса, кодирующая сигнальный пептид, не была делетирована и заменена соответствующей человеческой TTR последовательностью, первый экзон эндогенного Ttr локуса не был делетирован и заменен соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от начала второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью.
8. Отличное от человека животное по п. 7, в котором область эндогенного Ttr локуса от второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена человеческой TTR последовательностью, включающей в себя соответствующую человеческую TTR последовательность и человеческую TTR 3' нетранслируемую область, и
в котором первый экзон и первый интрон эндогенного Ttr локуса не были делетированы и заменены соответствующей человеческой TTR последовательностью, и
в котором 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
в котором эндогенный Ttr промотор не был делетирован и заменен соответствующей человеческой TTR последовательностью.
9. Отличное от человека животное по п. 8, в котором:
(i) человеческая TTR последовательность в генетически модифицированном эндогенном Ttr локусе включает в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 19;
(ii) генетически модифицированный эндогенный Ttr локус кодирует белок, включающий в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 2;
(iii) генетически модифицированный эндогенный Ttr локус включает в себя кодирующую последовательность, включающую в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 91; или
(iv) генетически модифицированный эндогенный Ttr локус включает в себя последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 16 или 17.
10. Отличное от человека животное по любому из пп. 7-9, в котором генетически модифицированный эндогенный Ttr локус не включает в себя кассету селекции или репортерный ген.
11. Отличное от человека животное по любому из пп. 7-10, при этом отличное от человека животное является гомозиготным по генетически модифицированному эндогенному Ttr локусу.
12. Отличное от человека животное по любому из пп. 7-11, при этом отличное от человека животное представляет собой мышь.
13. Способ оценивания активности нацеливающегося на человеческий TTR реагента in vivo, предусматривающий:
(a) ведение нацеливающегося на человеческий TTR реагента в отличное от человека животное по любому из пп. 1-12 и
(b) оценивание активности нацеливающегося на человеческий TTR реагента у отличного от человека животного.
14. Способ по п. 13, при котором ведение включает в себя: (1) опосредованную адено-ассоциированным вирусом (AAV) доставку, при этом введение необязательно включает в себя опосредованную AAV8 доставку; (2) опосредованную липидной наночастицей (LNP) доставку, при этом необязательно доза LNP составляет от приблизительно 0,1 мг/кг до приблизительно 2 мг/кг; или (3) гидродинамическую доставку (HDD).
15. Способ по п. 13 или 14, при котором стадия (b) предусматривает выделение печени у отличного от человека животного и оценивание активности нацеливающегося на человеческий TTR реагента в печени, при этом необязательно стадия (b) дополнительно предусматривает оценивание активности нацеливающегося на человеческий TTR реагента в органе или ткани, отличных от печени.
16. Способ по любому из пп. 13-15, при котором нацеливающийся на человеческий TTR реагент представляет собой редактирующее геном средство и оценивание предусматривает оценивание модификации генетически модифицированного Ttr локуса, при этом оценивание необязательно предусматривает измерение частоты вставок или делеций в генетически модифицированном Ttr локусе.
17. Способ по любому из пп. 13-16, при котором оценивание предусматривает измерение экспрессии информационной РНК Ttr, кодируемой генетически модифицированным Ttr локусом, или измерение экспрессии TTR белка, кодируемого генетически модифицированным Ttr локусом, при этом необязательно измерение экспрессии TTR белка предусматривает измерение содержания в сыворотке крови TTR белка у отличного от человека животного.
18. Способ по любому из пп. 13-17, при котором нацеливающийся на человеческий TTR реагент включает в себя нуклеазное средство, разработанное для нацеливания на область человеческого TTR гена.
19. Способ по п. 18, при котором нуклеазное средство включает в себя белок Cas и направляющую РНК, разработанную для нацеливания целевой последовательности направляющей РНК на человеческий TTR ген, при этом необязательно белок Cas представляет собой белок Cas9.
20. Способ по п. 18 или 19, при котором нацеливающийся на человеческий TTR реагент дополнительно включает в себя экзогенную донорную нуклеиновую кислоту, при этом экзогенная донорная нуклеиновая кислота разработана для рекомбинации с человеческим TTR геном, и при этом необязательно экзогенная донорная нуклеиновая кислота представляет собой однонитевой олигодезоксинуклеотид (ssODN).
21. Способ оптимизации активности нацеливающегося на человеческий TTR реагента in vivo, предусматривающий:
(I) выполнение способа по любому из пп. 13-20 первый раз у первого отличного от человека животного, включающего в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность, причем способ выполняют с неизмененной переменной;
(II) изменение переменной и выполнение способа стадии (I) во второй раз с измененной переменной у второго отличного от человека животного, включающего в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность; и
(III) сравнение активности нацеливающегося на человеческий TTR реагента на стадии (I) с активностью нацеливающегося на человеческий TTR реагента на стадии (II) и выбор способа, приводящего в результате к более высокой активности.
22. Способ по п. 21, при котором:
(i) измененная переменная на стадии (II) представляет собой способ доставки или путь введения для введения нацеливающегося на человеческий TTR реагента в отличное от человека животное, при котором необязательно ведение включает в себя опосредованную LNP доставку, и измененная переменная на стадии (II) представляет собой состав LNP;
(ii) измененная переменная на стадии (II) представляет собой концентрацию или количество нацеливающегося на человеческий TTR реагента, вводимого в отличное от человека животное;
(iii) измененная переменная на стадии (II) представляет собой форму нацеливающегося на человеческий TTR реагента, вводимого в отличное от человека животное;
(iv) измененная переменная на стадии (II) представляет собой нацеливающийся на человеческий TTR реагент, вводимый в отличное от человека животное; или
(v) нацеливающийся на человеческий TTR реагент включает в себя белок Cas и направляющую РНК, разработанную для нацеливания на целевую последовательность направляющей РНК в человеческом TTR гене, и (1) измененная переменная на стадии (II) представляет собой последовательность направляющей РНК или целевую последовательность направляющей РНК, (2) каждые из белка Cas и направляющей РНК вводят в форме РНК и измененная переменная на стадии (II) представляет собой отношение иРНК Cas к направляющей РНК или (3) измененная переменная на стадии (II) представляет собой модификации направляющей РНК.
23. Клетка отличного от человека животного для экспрессии человеческого белка транстиретина (TTR) или химерного TTR белка, включающего в себя один или несколько фрагментов человеческого TTR белка, причем клетка отличного от человека животного включает в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
где отличное от человека животное представляет собой мышь или крысу,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и при этом вся кодирующая Ttr последовательность эндогенного Ttr локуса была делетирована и заменена соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью.
24. Клетка отличного от человека животного для экспрессии человеческого белка транстиретина (TTR) или химерного TTR белка, включающего в себя один или несколько фрагментов человеческого TTR белка, причем клетка отличного от человека животного включает в свой геном генетически модифицированный эндогенный Ttr локус, включающий в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
где отличное от человека животное представляет собой мышь или крысу,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
при этом генетически модифицированный эндогенный Ttr локус кодирует белок-предшественник TTR, включающий в себя сигнальный пептид, область эндогенного Ttr локуса, кодирующая сигнальный пептид, не была делетирована и заменена соответствующей человеческой TTR последовательностью, первый экзон эндогенного Ttr локуса не был делетирован и заменен соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от начала второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью.
25. Способ получения отличного от человека животного по любому из пп. 1-6, предусматривающий:
(А) (а) модификацию генома плюрипотентной клетки мыши или плюрипотентной клетки крысы для содержания генетически модифицированного эндогенного Ttr локуса, включающего в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
причем вся кодирующая Ttr последовательность эндогенного Ttr локуса была делетирована и заменена соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью;
(b) идентификацию или отбор генетически модифицированной плюрипотентной клетки мыши или генетически модифицированной плюрипотентной клетки крысы, содержащей генетически модифицированный эндогенный Ttr локус;
(c) введение генетически модифицированной плюрипотентной клетки мыши в эмбрион мыши-хозяина или введение генетически модифицированной плюрипотентной клетки крысы в эмбрион крысы-хозяина; и
(d) имплантацию и гестацию эмбриона мыши-хозяина в суррогатную мышь-мать или имплантацию и гестацию эмбриона крысы-хозяина в суррогатную крысу-мать; или
(В) (а) модификацию генома эмбриона мыши на стадии одной клетки или эмбриона крысы на стадии одной клетки для содержания генетически модифицированного эндогенного Ttr локуса, включающего в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
причем вся кодирующая Ttr последовательность эндогенного Ttr локуса была делетирована и заменена соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от инициирующего кодона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью;
(b) отбор генетически модифицированного эмбриона мыши на стадии одной клетки или генетически модифицированного эмбриона крысы на стадии одной клетки, содержащего генетически модифицированный эндогенный Ttr локус; и
(c) имплантацию и гестацию генетически модифицированного эмбриона мыши на стадии одной клетки в суррогатную мышь-мать или имплантацию и гестацию генетически модифицированного эмбриона крысы на стадии одной клетки в суррогатную крысу-мать.
26. Способ получения отличного от человека животного по любому из пп. 7-12, предусматривающий:
(A) (а) модификацию генома плюрипотентной клетки мыши или плюрипотентной клетки крысы для содержания генетически модифицированного эндогенного Ttr локуса, включающего в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
причем генетически модифицированный эндогенный Ttr локус кодирует белок-предшественник TTR, включающий в себя сигнальный пептид, область эндогенного Ttr локуса, кодирующая сигнальный пептид, не была делетирована и заменена соответствующей человеческой TTR последовательностью, первый экзон эндогенного Ttr локуса не был делетирован и заменен соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от начала второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью;
(b) идентификацию или отбор генетически модифицированной плюрипотентной клетки мыши или плюрипотентной клетки крысы, содержащей генетически модифицированный эндогенный Ttr локус;
(c) введение генетически модифицированной плюрипотентной клетки мыши в эмбрион мыши-хозяина или введение генетически модифицированной плюрипотентной клетки крысы в эмбрион крысы-хозяина; и
(d) имплантацию и гестацию эмбриона мыши-хозяина в суррогатную мышь-мать или имплантацию и гестацию эмбриона крысы-хозяина в суррогатную крысу-мать; или
(B) (а) модификацию генома эмбриона мыши на стадии одной клетки или крысы на стадии одной клетки для содержания генетически модифицированного эндогенного Ttr локуса, включающего в себя человеческую TTR последовательность, включающую в себя как кодирующую TTR последовательность, так и некодирующую последовательность,
причем генетически модифицированный эндогенный Ttr локус включает в себя эндогенный Ttr промотор, в котором человеческая TTR последовательность функционально связана с эндогенным Ttr промотором,
причем генетически модифицированный эндогенный Ttr локус включает в себя человеческую TTR 3' нетранслируемую область,
причем 5' нетранслируемая область эндогенного Ttr не была делетирована и заменена соответствующей человеческой TTR последовательностью, и
причем генетически модифицированный эндогенный Ttr локус кодирует белок-предшественник TTR, включающий в себя сигнальный пептид, область эндогенного Ttr локуса, кодирующая сигнальный пептид, не была делетирована и заменена соответствующей человеческой TTR последовательностью, первый экзон эндогенного Ttr локуса не был делетирован и заменен соответствующей человеческой TTR последовательностью и область эндогенного Ttr локуса от начала второго экзона Ttr до терминирующего кодона Ttr была делетирована и заменена соответствующей человеческой TTR последовательностью;
(b) отбор генетически модифицированного эмбриона мыши на стадии одной клетки или генетически модифицированного эмбриона крысы на стадии одной клетки, содержащего генетически модифицированный эндогенный Ttr локус; и
(c) имплантацию и гестацию генетически модифицированного эмбриона мыши на стадии одной клетки в суррогатную мышь-мать или имплантацию и гестацию генетически модифицированного эмбриона крысы на стадии одной клетки в суррогатную крысу-мать.
| GANG ZHAO et al., Inconsistency between hepatic expression and serum concentration of transthyretin in mice humanized at the transthyretin locus, Genes to Cells, 2008, Vol | |||
| Насос | 1917 |
|
SU13A1 |
| MEI-HUI TENG et al., Amyloid and Nonfibrillar Deposits in Mice Transgenic for Wild-Type Human Transthyretin: A Possible Model for Senile Systemic Amyloidosis, | |||

