Область техники
Настоящее изобретение относится к способу определения активности ботулотоксина с использованием клеток.
Уровень техники
В настоящее время биологический анализ LD50 на мышах (mLD50) является общепринятым методом обнаружения BoNT/A, остающегося в пищевых, клинических или экологических образцах. В частности, фармацевтическая промышленность использует mLD50 в качестве стандартного теста для измерения активности BoNT/A в эстетических или клинических целях. Однако известно, что активность BoNT/A, оцениваемая с помощью mLD50, в значительной степени варьируется в зависимости от исследовательских институтов/учреждений и исследователей, поэтому точное и воспроизводимое количественное определение биологической активности BoNT/A является сложной задачей.
Встреча ZEBET была проведена 27-28 апреля в Берлине, Германия, для продвижения подходов, альтернативных mLD50, с целью стандартизации измерения BoNT/A, а также для минимизации количества задействованных подопытных животных и причиняемых им страданий (Altern Lab Anim. Август 2010; 38 (4): 315-30). Следовательно, многие научно-исследовательские институты и отрасли в мире приступили к разнообразным исследованиям для разработки способов определения активности с использованием клеток (cell-based potency assay, СВРА) или биоанализа с использованием клеток (cell-based bioassay, СВВ), которые могли бы заменить mLD50 с удовлетворительной специфичностью, чувствительностью и воспроизводимостью. Чтобы успешно разработать СВРА или СВВ, они пытались получить (1) моноклональные или поликлональные антитела, специфичные к SNAP25197, и (2) линии нейрональных клеток, которые проявляют высокую чувствительность к низким уровням BoNT/A (~пМ).
Еще в 2004 году доктор Чепмен и его коллеги изобрели флуоресцентный репортерный анализ, в котором использовали эпигенетически экспрессируемый SNAP25, слитый с двумя флуоресцентными белками (Proc Natl Acad Sci USA. 2004 Oct 12; 101 (41): 14701-6), что потом стало технологической платформой BoCellTM (BioSentinel Inc). Хотя это первый в своем роде метод, позволяющий определять активность эндопептидазы BoNT/A в клетках млекопитающих, растущих в 96-луночном культуральном планшете, анализ BoCellTM примерно на 2-3 порядка менее чувствителен, чем биоанализ на мышах (Appl Environ Microbiol vol. 78, 21 (2012): 7687-97).
Как было сказано выше, во всем мире были предприняты усилия по разработке СВРА для замены mLD50, что в конечном итоге исключило бы испытания на животных. Хотя несколько институтов разработали СРВА, остается несколько недостатков, которые необходимо преодолеть. Изобретение значительно улучшенного СВРА не только будет способствовать разработке разнообразных продуктов, имеющих отношение к BoNT/A, но также повысит конкурентоспособность продуктов, предоставив потребителям более высокий уровень уверенности в отношении контроля качества.
Следовательно, настоящее изобретение призвано преодолеть ограничения обычного способа СРВА и разработать более эффективный способ СВРА. Путем сравнительного анализа 13 различных линий нейронных клеток с последующим обширным клональным отбором в рамках настоящего изобретение была создана новая клеточная линия N2-42F. В дополнение к уровню чувствительности к BoNT/A, который можно сравнить с уровнем чувствительности SiMa, время удвоения популяции (PDT), равное ~24 часам, и стабильное прикрепление к культуральной пластине, покрытой поли-D-лизином (PDL), делают клетки N2-42F очень привлекательными и надежными клетками-хозяевами для способа СВРА.
Настоящее изобретение также относится к антителу для способа СРВА для замены биоанализа LD50 на мышах (mLD50), при этом указанное антитело представляет собой моноклональное антитело, обладающее значительно более высокой аффинностью связывания и специфичностью в отношении SNAP25, которое позволяет преодолеть ограничения обычного способа СРВА и делает возможным разработку более эффективного способа СВРА. Таким образом, ожидается, что указанное антитело будет активно использоваться в фармацевтической и косметической областях.
В дополнение, настоящее изобретение относится к оптимальному способу анализа активности с использованием клеток (СВРА), в котором используют клетки N2-42F и моноклональные антитела, обладающие значительно более высокой аффинностью связывания и специфичностью по отношению к SNAP25, и этот способ СВРА позволяет измерять активность ботулотоксина на уровне 0,5 ЕД или менее. Ожидается, что указанный способ СВРА, использующий клетку и антитело согласно настоящему изобретению, станет высоконадежным и воспроизводимым способом анализа активности ботулотоксина с использованием клеток.
Описание
Техническая задача
Настоящее изобретение было создано для решения вышеописанных проблем, имеющих место в предшествующем уровне техники, и целью настоящего изобретения является предоставление клетки, антител и способа определения активности ботулотоксина.
Однако техническая цель, которая должна быть достигнута настоящим изобретением, не ограничивается вышеупомянутой технической целью, и другие цели, которые не упомянуты выше, могут быть легко поняты специалистами в данной области техники из следующего описания.
Техническое решение
В дальнейшем различные варианты осуществления изобретения, описанные в данном документе, будут описаны со ссылкой на фигуры. В нижеследующем описании изложены многочисленные конкретные детали, такие как конкретные конфигурации, составы, процессы и т.д., чтобы обеспечить полное понимание настоящего изобретения. Однако определенные варианты осуществления изобретения могут быть реализованы без одной или нескольких из этих конкретных деталей или в сочетании с другими известными способами и конфигурациями. В других случаях, известные процессы и способы получения не были описаны подробно, чтобы излишне не затруднять понимание настоящего изобретения. Ссылка в данном описании на «один вариант осуществления» или «вариант осуществления» означает, что конкретный признак, конфигурация, композиция или характеристика, описанные в связи с указанным вариантом осуществления, включены по меньшей мере в один вариант осуществления настоящего изобретения. Таким образом, появление фразы «в одном варианте осуществления» или «варианте осуществления» в различных местах в этом описании не обязательно относится к одному и тому же варианту осуществления настоящего изобретения. Кроме того, конкретные особенности, конфигурации, композиции или характеристики могут быть объединены любым подходящим образом в одном или нескольких вариантах осуществления настоящего изобретения.
Если в описании не указано иное, все научные и технические термины, используемые в настоящем описании, имеют те же значения, которые обычно понимаются специалистами в области техники, к которой относится настоящее изобретение.
Компания Allergan Inc., производитель Botox®, успешно произвела моноклональные антитела, специфичные к SNAP25197, а также идентифицировала нейробластому человека SiMa как идеальную линию клеток-хозяев, высокочувствительных к BoNT/A (PLoS One.2012; 7 (11): е49516). Используя эти реагенты, они разработали новый способ СВРА для оценки стабильности и эффективности BoNT/A, который был впервые одобрен FDA для замены mLD50 в 2010 году (патент США US 8455213 B2 и патентная заявка US 2010/0204559 А1). Компания MERZ Pharma, немецкий производитель ботулотоксина, разработала способ CBA-ELISA в 2014 году (WO 2014/207109 A1) и получила одобрение FDA в 2015 году. Способ CBA-ELISA использует фиксацию дифференцированных нейронных клеток in situ с последующим использованием проницаемой мембраны и иммунологическим обнаружением эндогенных SNAP25 (белок 25, ассоциированный с синаптосомным нервом).
Способы СВРА компании Allergan и способ CBA-ELISA компании MERZ демонстрируют превосходную чувствительность при субпикомолярных концентрациях ЕС50 (т.е. <1,0 Ед/лунку), что эквивалентно биопробам с использованием мышей. Технологические платформы Allergan и MERZ обычно использует коммерческие поликлональные антитела кролика (Sigma S9684) для обнаружения SNAP25 в оптимальных для них условиях. Дифференцированные клетки нейробластомы человека SiMa используют исключительно в способе СВРА компании Allergan, в то время как дифференцированные индуцированные плюрипотентные стволовые клетки человека (iPS) используют в качестве клеток-хозяина в стандартизированном и оптимизированном способе CBA-ELISA компании MERZ. Клетки SiMa растут медленно, время удвоения популяции (PDT) превышает 70 часов. Точно так же создание нейрональных дифференцированных iPS-клеток человека занимает много времени, и, кроме того, их хранение так же сложно, как и создание. Таким образом, способ СВРА станет более надежным, если линия нейронных клеток будет не только высокочувствительна к BoNT/A, но и ее можно будет легко поддерживать и хранить, и она будет иметь более быстрое PDT. Более того, исследовательская группа во главе с доктором Дэвидом Биби из Университета Висконсина поставила вопрос о пригодности линии клеток SiMa, поскольку она не демонстрирует характеристики, подобные характеристикам двигательных нейронов (J Biomol Screen. 2016 Jan; 21(1): 65-73). Они разработали альтернативный способ СВРА, используя линию клеток NG108-15, подобных мотонейронам, которая демонстрировала ЕС50 ~7,9 пМ. Поскольку способ СВРА полагается на анализ вестерн-блоттинга для сравнительного определения эндогенных уровней SNAP25197 и SNAP25FL, его использование в качестве высокопроизводительного анализа в настоящее время представляется сложной задачей.
В одном из вариантов настоящего изобретения «ботулотоксин» представляет собой нейротоксический белок, продуцируемый бактерией Clostridium botulinum. Род Clostridium насчитывает более 127 видов, сгруппированных по морфологии и функциям. Анаэробные грамположительные бактерии Clostridium botulinum продуцируют мощный полипептидный нейротоксин, ботулотоксин, который вызывает нервно-паралитическое заболевание у людей и животных, известное как ботулизм. Споры Clostridium botulinum обнаруживаются в почве и могут расти в неправильно стерилизованных и герметичных пищевых контейнерах на домашних консервных заводах, что является причиной многих случаев ботулизма. Симптомы ботулизма обычно проявляются через 18-36 часов после употребления в пищу продуктов, инфицированных культурой или спорами Clostridium botulinum. Ботулотоксин, по-видимому, может проходить без ослабления через слизистую оболочку кишечника и проявляет высокое сродство к холинергическим мотонейронам. Симптомы отравления ботулотоксином могут прогрессировать от затруднений при ходьбе, глотании и разговоре до паралича дыхательных мышц и смерти.
Ботулотоксин типа А известен как самый ядовитый и смертоносный природный биологический агент для человека. Примерно 50 пикограмм коммерчески доступного ботулотоксина типа А (очищенный комплекс нейротоксинов) составляет LD50 (т.е. 1 единица). Интересно, что в пересчете на молярность, ботулотоксин типа А примерно в 1,8 миллиарда раз более ядовит, чем дифтерия, примерно в 600 миллионов раз более ядовит, чем цианид натрия, примерно в 30 миллионов раз более ядовит, чем токсин кобры, и примерно в 12 миллионов раз более ядовит, чем холера. Одной единицей (ЕД) ботулотоксина считается LD50 при внутрибрюшинной инъекции самкам мышей Swiss Webster массой от 18 до 20 граммов каждая.
В целом были охарактеризованы 7 иммунологически отличных ботулинических нейротоксинов: ботулинические нейротоксины серотипов А, В, C1, D, Е, F и G, каждый из которых отличается нейтрализацией типоспецифическими антителами. Различные серотипы ботулотоксина отличаются в зависимости от вида животных, на которой они действуют, а также они отличаются по тяжести и продолжительности паралича, который они вызывают. Например, было определено, что ботулотоксин типа А в 500 раз более эффективен, если судить по скорости паралича у крыс, чем ботулотоксин типа В. Кроме того, ботулотоксин типа В не токсичен для приматов при дозе 480 Ед/кг, что примерно в 12 раз превышает LD50 для ботулотоксина типа А у приматов. Очевидно, ботулотоксин связывается с высоким сродством с холинергическими мотонейронами, перемещается в нейрон и блокирует высвобождение ацетилхолина. Дополнительное поглощение может происходить через рецепторы с низким сродством, а также за счет фагоцитоза и пиноцитоза.
Независимо от серотипа, молекулярный механизм интоксикации токсином является похожим и включает не менее 3 этапов. На первом этапе процесса токсин связывается с пресинаптической мембраной целевого нейрона посредством специфического взаимодействия между тяжелой цепью (Н-цепью или НС) и рецептором на поверхности клетки. Считается, что рецепторы для каждого типа ботулотоксина и столбнячного токсина различаются. Карбоксильный концевой сегмент НС, по-видимому, важен для присоединения ботулотоксина к поверхности клетки.
На втором этапе ботулотоксин проникает через плазматическую мембрану клетки-мишени. Ботулотоксин сначала поглощается клеткой через рецептор-опосредованный эндоцитоз, и образуется эндосома, содержащая ботулотоксин. Затем токсин выходит из эндосомы в цитоплазму клетки. Считается, что эта стадия опосредуется концевым амино-сегментом тяжелой цепи, HN, который запускает конформационное изменение токсина в ответ на рН около 5,5 или ниже. Известно, что эндосомы обладают протонным насосом, который снижает внутриэндосомный рН. Конформационный сдвиг обнажает гидрофобные остатки в токсине, что позволяет ботулотоксину внедряться в эндосомальную мембрану. Затем ботулотоксин (или, по крайней мере, легкая цепь ботулотоксина) перемещается через эндосомальную мембрану в цитоплазму.
Последний этап механизма действия ботулотоксина, по-видимому, включает восстановление дисульфидной связи, соединяющей тяжелую цепь и легкую цепь. Вся токсическая активность ботулинического и столбнячного токсинов определяется легкой цепью голотоксина; легкая цепь представляет собой эндопептидазу цинка (Zn++), которая избирательно расщепляет белки, необходимые для распознавания и стыковки везикул, содержащих нейротрансмиттер, с цитоплазматической поверхностью плазматической мембраны и слияния везикул с плазматической мембраной. Нейротоксин столбняка, ботулотоксин типов В, D, F и G вызывают деградацию синаптобревина (также называемого мембранным белком, ассоциированным с пузырьками (VAMP)), белком синаптосомной мембраны. Большая часть VAMP, присутствующего на цитоплазматической поверхности синаптического пузырька, удаляется в результате любого из этих событий расщепления. Серотипы А и Е расщепляют SNAP25. Первоначально предполагалось, что серотип С1 расщепляет синтаксин, но было обнаружено, что он расщепляет синтаксин и SNAP25. Каждый из ботулотоксинов специфически расщепляет разные связи, за исключением ботулотоксина типа В (и столбнячного токсина), которые расщепляют одну и ту же связь. Каждое из этих расщеплений блокирует процесс связывания везикул с мембраной, тем самым предотвращая экзоцитоз содержимого везикул.
Ботулотоксины использовались в клинических условиях для лечения нервно-мышечных расстройств, характеризующихся гиперактивностью скелетных мышц (т.е. двигательных расстройств). В 1989 году комплекс ботулотоксина типа А был одобрен Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) для лечения блефароспазма, косоглазия и гемифациального спазма. Впоследствии ботулотоксин типа А был также одобрен FDA для лечения цервикальной дистонии и для лечения глабеллярных линий, а ботулотоксин типа В был одобрен для лечения цервикальной дистонии. Серотипы ботулотоксина, не относящиеся к типу А, по-видимому, имеют более низкую эффективность и/или более короткую продолжительность активности по сравнению с ботулотоксином типа А. Клинические эффекты периферического внутримышечного ботулотоксина типа А обычно наблюдаются в течение одной недели после инъекции. Типичная продолжительность облегчения симптомов после однократной внутримышечной инъекции ботулотоксина типа А составляет в среднем около 3 месяцев, хотя сообщалось о значительно более длительных периодах терапевтической активности.
Хотя все серотипы ботулотоксинов, по-видимому, ингибируют высвобождение нейромедиатора ацетилхолина в нервно-мышечном соединении, они делают это, воздействуя на разные нейросекреторные белки и расщепляя эти белки в разных местах. Например, ботулинические типы А и Е оба расщепляют синаптосомный белок массой 25 кДа (SNAP25), но они нацелены на различные аминокислотные последовательности в этом белке. Ботулотоксин типов В, D, F и G действует на ассоциированный с везикулами мембранный белок (VAMP, также называемый синаптобревином), причем каждый серотип расщепляет белок в разных местах. Наконец, ботулотоксин типа С1, по-видимому, расщепляет синтаксин и SNAP25. Эти различия в механизме действия могут влиять на относительную эффективность и/или продолжительность действия различных серотипов ботулотоксина. В частности, субстрат для ботулотоксина можно найти во множестве различных типов клеток.
Молекулярная масса ботулотоксина для всех семи известных серотипов ботулотоксина составляет около 150 кДа. Интересно, что ботулотоксины высвобождаются клостридиальными бактериями в виде комплексов, содержащих молекулу белка ботулотоксина 150 кДа вместе с ассоциированными белками, не являющимися токсинами. Таким образом, комплекс ботулотоксина типа А может продуцироваться клостридиальными бактериями в формах 900 кДа, 500 кДа или 300 кДа. Ботулотоксин типов В и С1, по-видимому, производится только в виде комплекса 700 или 500 кДа. Ботулотоксин типа D производится в виде комплексов 300 или 500 кДа. Наконец, типы ботулотоксина Е и F производятся только в виде комплексов с молекулярной массой приблизительно 300 кДа. Считается, что указанные комплексы (то есть, комплексы с молекулярной массой более примерно 150 кДа) содержат гемагглютининовые белки, не являющиеся токсинами, и не гемагглютининовые белки, не являющиеся токсинами. Эти два типа белков, не являющихся токсинами, (которые вместе с молекулой ботулотоксина образуют соответствующий нейротоксиновый комплекс) могут обеспечивать стабильность молекулы ботулотоксина в отношении денатурации и защиту от пищеварительных кислот при проглатывании ботулотоксина. Кроме того, возможно, что наличие более крупных комплексов ботулотоксина (молекулярная масса более 150 кДа) приводит к более медленной скорости диффузии ботулотоксина от места внутримышечной инъекции комплекса ботулотоксина.
Исследования in vitro показали, что ботулотоксин ингибирует вызванное катионом калия высвобождение ацетилхолина и норэпинефрина из первичных культур клеток ткани ствола мозга. Кроме того, сообщалось, что ботулотоксин ингибирует индуцированное высвобождение как глицина, так и глутамата в первичных культурах нейронов спинного мозга, а в препаратах синаптосом головного мозга ботулотоксин подавляет высвобождение каждого из нейротрансмиттеров, представленных ацетилхолином, дофамином, норадреналином, CGRP, веществом Р и глутаматом. Таким образом, при использовании адекватных концентраций индуцированное высвобождение большинства нейротрансмиттеров может быть заблокировано ботулотоксином.
Ботулотоксин типа А может быть получен путем создания и выращивания культур Clostridium botulinum в ферментере, а затем сбора и очистки ферментированной смеси в соответствии с известными процедурами. Все серотипы ботулотоксина изначально синтезируются как неактивные одноцепочечные белки, которые должны быть расщеплены или разрезаны протеазами, чтобы стать нейроактивными. Бактериальные штаммы, которые продуцируют ботулотоксин серотипов А и G, обладают эндогенными протеазами, и поэтому серотипы А и G могут быть выделены из бактериальных культур преимущественно в активной форме. Напротив, серотипы ботулотоксина C1, D и Е синтезируются непротеолитическими штаммами и поэтому обычно являются неактивными при извлечении из культуры. Серотипы В и F продуцируются как протеолитическими, так и непротеолитическими штаммами и, таким образом, могут быть выделены как в активной, так и в неактивной форме. Однако даже протеолитические штаммы, которые продуцируют, например, серотип ботулотоксина типа В, расщепляют только часть продуцируемого токсина. Точное соотношение расщепленных и нерасщепленных молекул зависит от продолжительности инкубации и температуры культуры. Следовательно, определенный процент любого препарата, например, токсина ботулотоксина типа В, вероятно, будет неактивным, что, возможно, объясняет известный факт о том, что ботулотоксин типа В имеет значительно более низкую эффективность по сравнению с ботулотоксином типа А. Присутствие неактивных молекул ботулотоксина в клиническом препарате будет вносить вклад в общую белковую нагрузку препарата, которая связана с повышенной антигенностью, не влияя на его клиническую эффективность. Более того, известно, что ботулотоксин типа В при внутримышечной инъекции имеет более короткую продолжительность активности и также менее эффективен, чем ботулотоксин типа А при том же уровне дозировки.
Высококачественный кристаллический ботулотоксин типа А может быть получен из штамма Hall А бактерии Clostridium botulinum с характеристиками ≥3×107 Ед/мг, А260/А278 менее 0,60 и четким рисунком полос при гель-электрофорезе. Известный процесс Шанца можно использовать для получения кристаллического ботулотоксина типа А. Обычно комплекс ботулотоксина типа А может быть выделен и очищен из культуры анаэробной ферментации при культивировании Clostridium botulinum типа А в подходящей среде. Известный процесс также может быть использован после отделения нетоксиновых белков для получения чистых ботулотоксинов, таких как, например, очищенный ботулотоксин типа А с молекулярной массой приблизительно 150 кДа с удельной эффективностью 1-2×108 LD50 Ед/мг или больше; очищенный ботулотоксин типа В с молекулярной массой приблизительно 156 кДа с удельной эффективностью 1-2×108 LD50 Ед/мг или более, и; очищенный ботулотоксин типа F с молекулярной массой приблизительно 155 кДа с удельной эффективностью 1-2×107 LD50 Ед/мг или более.
Ботулотоксины и/или комплексы ботулотоксина являются коммерчески доступными от производителей соединений, известных в данной области, и чистый ботулотоксин также может быть использован для приготовления фармацевтической композиции.
Как и в случае с ферментами в целом, биологическая активность ботулотоксинов (которые являются внутриклеточными пептидазами) зависит, по крайней мере частично, от их трехмерной конформации. Таким образом, ботулотоксин типа А инактивируется при нагревании, при действии различных химических веществ, вызывающих растяжение и высыхание поверхности. Кроме того, известно, что разбавление комплекса ботулотоксина, полученного посредством известного культивирования, ферментации и очистки до очень низких концентраций токсина, используемых для приготовления фармацевтической композиции, приводит к быстрой инактивации токсина, если не присутствует подходящий стабилизирующий агент. Разведение токсина от миллиграммовых количеств до раствора, содержащего нанограммы на миллилитр, представляет значительные трудности из-за быстрой потери специфической токсичности при таком большом разбавлении. Поскольку ботулотоксин можно использовать через месяцы или годы после составления токсинсодержащей фармацевтической композиции, токсин следует стабилизировать подходящим стабилизирующим агентом. Таким образом, как раскрыто в настоящем изобретении, разработка технологии с оптимальным стабилизатором необходима для контроля высвобождением ботулотоксина in vivo для формы с замедленным высвобождением.
Сообщалось, что ботулотоксин типа А использовался в клинических приложениях следующим образом:
Обычная продолжительность действия внутримышечных инъекций ботулотоксина, вводимого in vivo, обычно составляет от 3 до 4 месяцев. Однако в некоторых случаях ботулотоксин подтипа А может иметь эффективность до 12 месяцев (European J. Neurology 6 (Supp 4): S111-S1150: 1999), а в некоторых случаях до 27 месяцев при использовании для лечения желез, например, при лечении гипергидроза.
Помимо фармакологического действия на периферические места, ботулотоксины могут также оказывать тормозящее действие на центральную нервную систему. Работа Weigand и др., Nauny-Schmiedeberg's Arch. Pharmacol. 1976; 292, 161 165, и Habermann, Nauny-Schmiedeberg's Arch. Pharmacol. 1974; 281, 47-56 показали, что ботулотоксин способен попадать в область позвоночника за счет ретроградного транспорта. Таким образом, ботулотоксин, вводимый в периферическое место, например внутримышечно, может ретроградно переноситься в спинной мозг.
Ботулотоксин также был предложен для лечения кожных ран костей и сухожилий или использовался для этого (патент США №6,447,787); интратекальной боли (см. патент США №6,113,915); различных расстройств вегетативной нервной системы, включая заболевания потовых желез (см., например, патент США №5,766,605 и Goldman (2000), Aesthetic Plastic Surgery, июль-август 24 (4): 280-282); тензионной головной боли (патент США №6,458,365); мигрени (патент США №5,714,468); послеоперационной боли и висцеральной боли (патент США №6,464,986); чрезмерного роста волос и сохранения волос (патент США №6,299,893); псориаза и дерматита (патент США №5,670,484); поврежденных мышц (патент США №6,423,319); различных раковых заболеваний (патенты США №6,139,845 и 6,063,768), нарушений гладкой мускулатуры (патент США №5,437,291); синдромов защемления нерва (патентная заявка США 20030224019); угрей (WO 03/011333); нейрогенного воспаления (патент США №6,063,768); оптических расстройств (см. патент США №6,265,379); заболеваний поджелудочной железы (см. патенты США №№6,143,306 и 6,261,572); заболеваний простаты, включая гиперплазию простаты, рак простаты и недержание мочи (см. патенты США №№6,365,164 и 6,667,041 и Doggweiler R. и др. Botulinum toxin type А causes diffuse and highly selective atrophy of rat prostate, Neurourol Urodyn 1998; 17 (4): 363); фибромиалгии (патент США №6,623,742) и синдрома грушевидной мышцы (см. Childers и др. (2002), American Journal of Physical Medicine & Rehabilitation, 81: 751-759).
В патенте США №5,989,545 раскрывается, что модифицированный клостридиальный нейротоксин или его фрагмент, предпочтительно ботулотоксин, химически конъюгированный или рекомбинантно слитый с конкретным нацеливающим фрагментом, можно использовать для лечения боли путем введения агента в спинной мозг. Кроме того, было обнаружено, что целевые ботулотоксины (т.е. с неприродным связывающим фрагментом) можно использовать для лечения различных состояний (см., WO 96/33273; WO 99/17806; WO 98/07864; WO 00/57897; WO 01/21213; WO 00/10598).
Кроме того, ботулотоксин вводили в грудную мышцу для контроля спазма грудной клетки (Senior М., Botox and the management of pectoral spasm after subpectoral implant insertion, Plastic and Recon Surg, июль 2000 г., стр. 224-225). Известны имплантаты токсина с контролируемым высвобождением (см. патенты США №№6,306,423 и 6,312,708), как и трансдермальное введение ботулотоксина (патентная заявка США №10/194,805). Известно, что ботулотоксин можно использовать для ослабления жевательной или кусающей мышцы рта, чтобы можно было вылечить нанесенные самому себе раны и образовавшиеся язвы (Payne М. и др., Botulinum toxin as a novel treatment for self mutilation in Lesch-Nyhan syndrome, Ann Neurol 2002 Sep.; 52 (3 Supp 1): S157); для осуществления заживления доброкачественных кистозных поражений или опухолей (Blugerman G., и др., Multiple eccrine hidrocystomas: A new therapeutic option with botulinum toxin, Dermatol Surg 2003 May; 29 (5): 557-9); для лечения анальной трещины (Jost W., Ten years' experience with botulinum toxin in anal fissure, Int J Colorectal Dis, 2002, сентябрь; 17 (5): 298-302); и для лечения некоторых видов атопического дерматита (Heckmann М., и др., Botulinum toxin type A injection in the treatment of lichen simplex: An open pilot study, J Am Acad Dermatol 2002 April; 46(4):617-9).
Кроме того, ботулотоксин может уменьшать индуцированную воспалительную боль на крысиной модели с формалином (Aoki K., и др., Mechanisms of the antinociceptive effect of subcutaneous Botox: Inhibition of peripheral and central nociceptive processing, Cephalalgia 2003 Sept; 23(7):649). Кроме того, сообщалось, что блокада нервов ботулотоксином может вызывать уменьшение толщины эпидермиса (Li Y, и др., Sensory and motor denervation influences epidermal thickness in rat foot glabrous skin, Exp Neurol 1997; 147:452-462). Наконец, известно введение ботулотоксина в стопу для лечения чрезмерного потоотделения стоп (Katsambas А., и др., Cutaneous diseases of the foot: Unapproved treatments, Clin Dermatol 2002 November-December; 20(6):689-699; Sevim, S., и др., Botulinum toxin-A therapy for palmar and plantar hyperhidrosis, Acta Neurol Belg 2002 Dec; 102(4): 167-70), спастики пальцев ног (Suputtitada, A., Local botulinum toxin type A injections in the treatment of spastic toes, Am J Phys Med Rehabil 2002 October; 81(10):770-5) идиопатической ходьбы на пальцах ног (Tacks, L., и др., Idiopathic toe walking: Treatment with botulinum toxin A injection, Dev Med Child Neurol 2002; 44(Suppl 91):6), и дистонии стоп (Rogers J., и др., Injections of botulinum toxin A in foot dystonia, Neurology 1993 Apr.; 43(4 Suppl 2)).
Столбнячный токсин, а также его производные (т.е. с ненативным нацеливающим фрагментом), его фрагменты, гибриды и химеры также могут иметь терапевтическое применение. Указанный столбнячный токсин во многом похож на ботулотоксины. Таким образом, и столбнячный токсин, и ботулотоксины представляют собой полипептиды, производимые близкородственными видами Clostridium (Clostridium tetani и Clostridium botulinum, соответственно). Кроме того, как столбнячный токсин, так и ботулотоксины представляют собой двухцепочечные белки, состоящие из легкой цепи (молекулярная масса: примерно 50 кДа), ковалентно связанной одной дисульфидной связью с тяжелой цепью (молекулярная масса: примерно 100 кДа). Следовательно, молекулярная масса столбнячного токсина и каждого из 7 ботулотоксинов (не входящих в комплекс) составляет около 150 кДа. Кроме того, как для столбнячного токсина, так и для ботулотоксинов легкая цепь несет домен, который проявляет внутриклеточную биологическую (протеазную) активность, тогда как тяжелая цепь содержит домены связывания рецептора (иммуногенные) и транслокационные домены клеточной мембраны.
Кроме того, как столбнячный токсин, так и ботулотоксины проявляют высокое специфическое сродство к ганглиозидным рецепторам на поверхности пресинаптических холинергических нейронов. Рецептор-опосредованный эндоцитоз столбнячного токсина в периферических холинергических нейронах приводит к ретроградному аксональному транспорту, блокируя высвобождение тормозных нейротрансмиттеров из центральных синапсов и вызывая спастический паралич. Напротив, считалось, что рецептор-опосредованный эндоцитоз ботулотоксина в периферических холинергических нейронах вряд ли приводит к ретроградному транспорту, ингибированию экзоцитоза ацетилхолина из центральных синапсов и вялому параличу. Однако в недавнем сообщении было высказано предположение, что ботулотоксин также может подвергаться ретроградному транспорту по аксонам и, возможно, ингибировать высвобождение ацетилхолина в центральном синапсе (Bomba-Warczak и др., Interneuronal Transfer and Distal Action of Tetanus Toxin and Botulinum Neurotoxins A and D in Central Neurons, Cell Reports, 2016 August; 16, 1974-1987).
Наконец, столбнячный токсин и ботулотоксины похожи друг на друга как по биосинтезу, так и по молекулярной архитектуре. Таким образом, существует общая идентичность 34% между последовательностями белков столбнячного токсина и ботулотоксина типа А, а идентичность последовательностей достигает 62% для некоторых функциональных доменов (Binz Т. и др., The Complete Sequence of Botulinum Neurotoxin Type A and Comparison with Other Clostridial Neurotoxins, J Biological Chemistry 265(16); 9153-9158:1990).
В одном из вариантов осуществления настоящего изобретения «ацетилхолин» представляет собой сложный эфир холина и уксусной кислоты, который является первым известным нейромедиатором. Он распределен по нейронам, имеет химическую формулу C7H16NO2 и молекулярную массу 146,21 кДа.
Как правило, только один тип низкомолекулярных нейромедиаторов высвобождается каждым типом нейронов нервной системы млекопитающих, хотя есть данные, свидетельствующие о том, что один и тот же нейрон может высвобождать несколько нейромодуляторов. Нейромедиатор ацетилхолин секретируется нейронами во многих областях мозга, в частности, большими пирамидными клетками моторной коры, несколькими различными нейронами в базальных ганглиях, моторными нейронами, которые иннервируют скелетные мышцы, преганглионарными нейронами вегетативной нервной системы (как симпатическими, так и парасимпатическими), волокнами мешка 1 волокон мышечного веретена, постганглионарными нейронами парасимпатической нервной системы и некоторыми постганглионарные нейронами симпатической нервной системы. По сути, только постганглионарные симпатические нервные волокна потовых желез, пилореекторные мышцы и несколько кровеносных сосудов являются холинергическими, поскольку большинство постганглионарных нейронов симпатической нервной системы секретируют нейромедиатор норадреналин. В большинстве случаев ацетилхолин оказывает возбуждающее действие. Однако известно, что ацетилхолин оказывает ингибирующее действие на некоторые периферические парасимпатические нервные окончания (например, подавление частоты сердечных сокращений блуждающим нервом).
Эфферентные сигналы вегетативной нервной системы передаются телу либо через симпатическую нервную систему, либо через парасимпатическую нервную систему. Преганглионарные нейроны симпатической нервной системы отходят от тел клеток преганглионарных симпатических нейронов, расположенных в промежуточно-латеральном роге спинного мозга. Преганглионарные симпатические нервные волокна, отходящие от тела клетки, образуют синапс с постганглионарными нейронами, расположенными либо в паравертебральном симпатическом ганглии, либо в превертебральном ганглии. Поскольку преганглионарные нейроны симпатической и парасимпатической нервной системы являются холинергическими, нанесение ацетилхолина на ганглии будет возбуждать как симпатические, так и парасимпатические постганглионарные нейроны.
Ацетилхолин активирует два типа рецепторов, мускариновые и никотиновые рецепторы. Мускариновые рецепторы обнаруживаются во всех эффекторных клетках, стимулируемых постганглионарными нейронами парасимпатической нервной системы, а также в клетках, которые стимулируются постганглионарными холинергическими нейронами симпатической нервной системы. Никотиновые рецепторы находятся в мозговом веществе надпочечников, а также в вегетативных ганглиях, то есть на клеточной поверхности постганглионарного нейрона в синапсе между преганглионарными и постганглионарными нейронами как симпатической, так и парасимпатической систем. Никотиновые рецепторы также обнаруживаются во многих не вегетативных нервных окончаниях, например, в мембранах волокон скелетных мышц в нервно-мышечном соединении.
Ацетилхолин высвобождается из холинергических нейронов, когда маленькие прозрачные внутриклеточные пузырьки сливаются с пресинаптической мембраной нейрональной клетки. Большое разнообразие секреторных клеток, не являющихся нейронами, таких как мозговое вещество надпочечников (а также клеточная линия РС12) и островковые клетки поджелудочной железы, выделяют катехоламины и паратироидный гормон, соответственно, из больших везикул с плотным ядром. Клеточная линия PC12 представляет собой клон клеток феохромоцитомы крысы, широко используемых в качестве модели культуры тканей для изучения развития симпатоадреналовой функции. Ботулотоксин ингибирует высвобождение обоих типов соединений из обоих типов клеток in vitro, когда денервированные клетки проницаемы (например, путем электропорации) или когда непосредственно в них введен указанный токсин. Известно также, что ботулотоксин блокирует высвобождение глутамата нейромедиатора из культур клеток кортикальных синаптосом.
Нервно-мышечное соединение образуется в скелетных мышцах из-за близости аксонов к мышечным клеткам. Сигнал, передаваемый через нервную систему, приводит к возникновению потенциала действия на терминальном аксоне с активацией ионных каналов и, как следствие, высвобождением нейромедиатора ацетилхолина из внутринейрональных синаптических пузырьков, например, на моторной замыкательной пластинке нервно-мышечного соединения. Ацетилхолин пересекает внеклеточное пространство и связывается с белками рецептора ацетилхолина на поверхности концевой пластинки мышцы. Как только произошло достаточное связывание, потенциал действия мышечной клетки вызывает определенные изменения мембранного ионного канала, что приводит к сокращению мышечной клетки. Затем ацетилхолин высвобождается из мышечных клеток и метаболизируется холинэстеразами во внеклеточном пространстве. Метаболиты возвращаются обратно в терминальный аксон для переработки в ацетилхолин.
В одном из вариантов осуществления настоящего изобретения «активность ботулотоксина» означает активность токсина. Одна единица (ЕД) ботулотоксина определяется как количество ботулотоксина, которое убивает 50% группы мышей весом от 18 до 20 г каждая, при измерении с помощью стандартного способа анализа - биоанализа LD50 (mLD50) с использованием мышей. Ботулотоксин, особенно ботулотоксин серотипа А, является наиболее смертоносным природным биологическим агентом, известным человеку. Он в 1,8 миллиарда раз более ядовит, чем дифтерийный токсин, в 600 миллионов раз более ядовит, чем цианид натрия, в 30 миллионов раз более ядовит, чем токсин кобры, и в 12 миллионов раз более ядовит, чем токсин холеры. Таким образом, разница в эффективности ботулотоксина примерно на 20% приводит к значительному различию в действии, например, в 360 миллионов раз сильнее дифтерийного токсина или в 2,4 миллиона раз сильнее токсина холеры.
Препараты ботулотоксина, предназначенные для медицинских или косметических целей, обычно распространяются в виде лиофилизированных составов или жидких составов, и проблема заключается в том, что, поскольку ботулотоксин сам по себе является белком, его активность становится очень нестабильной из-за температуры, рН, света, физического воздействия или газа (воздух, азот, кислород и др.). Когда эффективность ботулотоксина снижается, как описано выше, он вряд ли проявляет ожидаемый эффект, и, следовательно, обязательно требуется точно предсказать эффективность ботулотоксина на стадии приготовления или стадии применения.
В одном варианте осуществления настоящего изобретения «антитело» - это термин, известный в данной области, который относится к конкретной молекуле белка, которая направлена против антигенного сайта. Для целей настоящего изобретения термин «антитело» означает антитело, которое специфически связывается с белком SNAP25. Это антитело можно получить обычным способом. Антитела согласно настоящему изобретению включают частичный пептид, который может быть получен из белка, и указанный частичный пептид согласно настоящему изобретению содержит по меньшей мере 7 аминокислот, предпочтительно по меньшей мере 9 аминокислот, более предпочтительно по меньшей мере 12 аминокислот. Форма антитела согласно настоящему изобретению не ограничивается каким-либо образом, и поликлональные антитела, моноклональные антитела или их части, которые обладают способностью к связыванию антигена, включены в антитела согласно настоящему изобретению, и все иммуноглобулиновые антитела включены в антитела согласно настоящему изобретению. Кроме того, антитела согласно настоящему изобретению также включают специальные антитела, такие как гуманизированные антитела. Антитела согласно настоящему изобретению включают не только полное антитело, имеющее легкие и тяжелые цепи, но также функциональный фрагмент молекулы антитела. Выражение «функциональный фрагмент молекулы антитела» относится к фрагменту, обладающему по меньшей мере антигенсвязывающей способностью, и примеры функционального фрагмента включают Fab, F(ab'), F(ab')2, Fv и подобные им.
В одном варианте осуществления настоящего изобретения термин «набор» означает набор композиций и аксессуаров для определенного назначения. Что касается назначения настоящего изобретения, указанный набор согласно настоящему изобретению включает либо антитело, которое специфически связывается с SNAP25FL или SNAP25197, либо композицию, содержащую антитело, либо чашку для культивирования клеток, покрытую антителом, для измерения активности ботулотоксина.
В одном варианте осуществления настоящего изобретения предоставляется линия клеток, клонально выбранная из Neuro-2a, которая является родительской линией нейрональных клеток.
Авторы настоящего изобретения выбрали клетки Neuro-2a, обладающие чувствительностью к ботулотоксину, которая аналогична чувствительности линии клеток SiMa, из 13 различных линий нейрональных клеток, и в конце концов выбрали линию N2-42F (регистрационный номер: KCTC 13712ВР), которая постоянно показывает высокую чувствительность к ботулотоксину, из клеток Neuro-2a посредством процесса клональной селекции, создав таким образом настоящее изобретение.
Линия клеток согласно настоящему изобретению может использоваться для определения активности ботулотоксина или для обнаружения ботулотоксина.
Линия клеток согласно настоящему изобретению сохраняет свою чувствительность к ботулотоксину, даже если продолжается пассирование, и, таким образом, ее можно с большим удобством использовать в платформе анализа на основе клеток.
Линия клеток для определения активности ботулотоксина в соответствии с настоящим изобретением означает гомогенные одиночные клетки, выделенные из родительской линии нейронных клеток, соответствующей популяции, включающей различные типы клеток, и относится к клеткам, имеющим общие генетические особенности, например, высокую или низкую экспрессию генов, уровни определенного гена и т.п.
Линия клеток для определения активности ботулотоксина может быть выделена из родительской нейрональной клеточной линии с помощью такого метода, как клональная селекция, или получена путем регулирования уровней экспрессии генов. Регуляция уровней экспрессии генов может быть достигнута обычным способом регуляции экспрессии генов, например трансформацией, манипуляциями с промоторами и т.п.
Линия родительских нейронных клеток согласно настоящему изобретению может включать любую иммортализованную линию клеток, полученную из нерва, и может быть предпочтительно клетками Neuro-2a, более предпочтительно клетками Neuro-2a (регистрационный номер: KCTC АС28106), но не ограничивается ими. Клетки Neuro-2a представляют собой нейрональные клетки мыши, которые обычно можно использовать для измерения LD50, при этом время удвоения составляет от 34 до 100 часов для обычных клеток SiMa, которые используются для определения активности ботулотоксина, а для клеток Neuro-2a оно составляет всего 24 часа, что указывает на то, что клетки Neuro-2a очень подходят не только для определения активности ботулотоксина с использованием клеток, но также для обнаружения ботулотоксина. Кроме того, клетки Neuro-2a соответствуют популяции, содержащей различные типы клеток при наблюдении под микроскопом, и, таким образом, имеют то преимущество, что они очень подходят для отбора из них только отдельных клеток, более чувствительных к ботулотоксину.
Линией клеток согласно настоящему изобретению, клонально выбранной из родительской линии нейрональных клеток Neuro-2a, может быть N2-42F (регистрационный номер: KCTC 13712ВР), но не ограничивается ей.
Линия клеток согласно настоящему изобретению для определения активности ботулотоксина может использоваться для обнаружения ботулотоксина или определения его активности. Клеточная линия чувствительна к ботулотоксину и, таким образом, может обнаруживать присутствие или отсутствие ботулотоксина в интересующем образце, а также может измерять степень токсичности ботулотоксина в зависимости от концентрации ботулотоксина.
Ботулотоксин согласно настоящему изобретению представляет собой нейротоксический белок, продуцируемый бактерией Clostridium botulinum, и его можно разделить в общей сложности на семь серотипов: А, В, С (C1, С2), D, Е, F и G. Ботулотоксин влияет на различные нейросекреторные белки в зависимости от серотипа и расщепляет эти белки в разных местах. В частности, оба серотипа ботулотоксина А и Е могут расщеплять SNAP25 (белок 25, связанный с синаптосомным нервом), серотипы ботулотоксина В, D, F и G могут расщеплять VAMP (белок мембран, связанный с пузырьками), а серотип ботулотоксина С1 может расщеплять как синтаксин, так и SNAP25, тем самым вызывая нейротоксичность. Предпочтительно, ботулотоксин может быть ботулотоксином серотипа А или ботулотоксином серотипа В, более предпочтительно ботулотоксином серотипа А, но не ограничивается этим.
Поскольку серотипы ботулотоксина А и В согласно настоящему изобретению очищены и широко используются для лечения дистонии, эстетических применений и т.п., и, таким образом, когда линию клеток согласно настоящему изобретению для определения активности ботулотоксина используют для измерения эффективности ботулотоксина, преимущество заключается в том, что можно определить концентрацию, при которой могут возникать побочные эффекты, тем самым решая проблемы, которые могут возникнуть, когда ботулотоксин используют для вышеописанных применений.
В другом варианте осуществления настоящего изобретения предоставляется способ определения активности ботулотоксина с использованием клеток.
Способ согласно настоящему изобретению включает стадии: культивирования линии клеток согласно настоящему изобретению; обработки культивируемой линии клеток ботулотоксином; и измерение чувствительности обработанной ботулотоксином линии клеток к ботулотоксину.
Способ определения активности ботулотоксина с использованием клеток согласно настоящему изобретению может быть осуществлен путем обработки ботулотоксином линии клеток для определения активности ботулотоксина и измерения чувствительности этой линии клеток к ботулотоксину. Таким образом, описание содержания, относящегося к линии клеток для определения активности ботулотоксина, ботулотоксину, нейросекреторным белкам, расщепляемым ботулотоксином, линии родительских нейрональных клеток и т.п., будет опущено во избежание чрезмерного усложнения описания в связи с повторным изложением указанного содержания.
На стадии культивирования линии клеток согласно настоящему изобретению культивирование указанной линии клеток может быть выполнено в культуральном планшете, покрытом поли-D-лизином. При использовании планшета, покрытого поли-D-лизином, линия клеток согласно настоящему изобретению может быть равномерно распределена, прочно прикреплена и может поддерживаться в здоровом состоянии клеток по сравнению с использованием планшета, который обычно используется для культивирования линии клеток, или планшета, покрытого желатином или коллагеном.
Стадия измерения чувствительности линии клеток к ботулотоксину согласно настоящему изобретению может включать измерение расщепления эндогенного нейросекреторного белка, вызванного ботулотоксином. В частности, в случае ботулотоксина серотипа А и серотипа Е можно измерять расщепление SNAP25, а в случае серотипов ботулотоксина В, D, F и G можно измерять расщепление VAMP, а в случае ботулотоксина серотипа С1 можно измерять расщепление синтаксина и/или SNAP25.
Измерение расщепления согласно настоящему изобретению может быть осуществлено с помощью метода обнаружения белка с использованием антитела, специфичного к расщепленному пептиду эндогенного нейросекреторного белка или тому подобного.
Антитело согласно настоящему изобретению означает белковую молекулу, которая может распознавать целый или расщепленный пептид нейросекреторного белка в качестве антигена и может специфически связываться с нейросекреторным белком, и примеры таких антител включают поликлональные антитела, моноклональные антитела и рекомбинантные антитела.
Метод обнаружения белка, который используется в настоящем изобретении, может представлять собой любой традиционный метод обнаружения белка, и его примеры включают, помимо прочего, вестерн-блоттинг, ELISA (иммуноферментный анализ), RIA (радиоиммуноанализ), радио иммунодиффузию, иммунодиффузию по Оухтерлони, ракетный иммуноэлектрофорез, иммуногистокрашивание, анализ иммунопреципитации, анализ связывания комплемента, использование сортировщика клеток с флуоресцентной активацией (FACS), анализ на белковом чипе и т.п.
В еще одном варианте настоящего изобретения предоставляется способ обнаружения ботулотоксина с использованием клеток.
Способ согласно настоящему изобретению включает этапы: культивирования линии клеток в соответствии с настоящим изобретением; обработка культивируемой линии клеток представляющим интерес образцом; и измерение чувствительности обработанной образцом линии клеток к ботулотоксину.
Способ обнаружения ботулотоксина с использованием клеток в соответствии с настоящим изобретением может быть реализован путем обработки линии клеток исследуемым образцом вместо ботулотоксина, используемого в методе определения активности ботулотоксина с использованием, и измерения чувствительности лини клеток к ботулотоксину. Таким образом, описание содержания, относящегося к линии клеток для определения активности ботулотоксина, нейросекреторных белков, расщепляемых ботулотоксином, исходной нейрональной линии клеток, планшета, покрытого клетками, метода обнаружения белка, антитела и подобного будет опущено, чтобы избежать чрезмерной сложности описания из-за повторного описания.
Образец, представляющий интерес, который используется в настоящем изобретении, представляет собой образец, который, как ожидается, будет содержать ботулотоксин, и его примеры могут включать биологические образцы, включая супернатанты культур клеток, кровь, слюну, мокроту, спинномозговую жидкость, секреты, лимфатические жидкости, диализные жидкости, биологические жидкости, мочу и т.п., а также химические образцы, содержащие различные соединения.
В еще одном варианте осуществления настоящего изобретения обеспечивается антитело, антитело, которое специфически связывается с SNAP25, при этом указанным SNAP25 является SNAP25FL или SNAP25197, и указанное антитело включает: участок CDR1 тяжелой цепи, выбранный из группы, включающей SEQ ID NO: 11-13, 28-33, и 55-56; участок CDR2 тяжелой цепи, выбранный из группы, включающей SEQ ID NO: 14-16, 34-39, и 57-58; участок CDR3 тяжелой цепи, выбранный из группы, включающей SEQ ID NO: 17-19, 40-46, и 59-60; участок CDR1 легкой цепи, выбранный из группы, включающей SEQ ID NO: 20-22, 47-49, и 61-62; участок CDR2 легкой цепи, выбранный из группы, включающей SEQ ID NO: 23-24, 50-51, и 63-64; и участок CDR3 легкой цепи, выбранный из группы, включающей SEQ ID NO: 25-27, 52-54, и 65-66.
Более конкретно, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25FL и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 11; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 14; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 17; участок CDR1 легкой цепи, представленный SEQ ID NO: 20; участок CDR2 легкой цепи, представленный SEQ ID NO: 23; и участок CDR3 легкой цепи, представленный SEQ ID NO: 25. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 83 и 84, но не ограничивается им.
Кроме того, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25FL и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 12; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 15; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 18; участок CDR1 легкой цепи, представленный SEQ ID NO: 21; участок CDR2 легкой цепи, представленный SEQ ID NO: 24; и участок CDR3 легкой цепи, представленный SEQ ID NO: 26. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 87 и 88, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25FL и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 13; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 16; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 19; участок CDR1 легкой цепи, представленный SEQ ID NO: 22; участок CDR2 легкой цепи, представленный SEQ ID NO: 24; и участок CDR3 легкой цепи, представленный SEQ ID NO: 27. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 89 и 90, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 28; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 34; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 40; участок CDR1 легкой цепи, представленный SEQ ID NO: 47; участок CDR2 легкой цепи, представленный SEQ ID NO: 50; и участок CDR3 легкой цепи, представленный SEQ ID NO: 52. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 71 и 72, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 29; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 35; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 41; участок CDR1 легкой цепи, представленный SEQ ID NO: 48; участок CDR2 легкой цепи, представленный SEQ ID NO: 50; и участок CDR3 легкой цепи, представленный SEQ ID NO: 52. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 73 и 74, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 29; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 36; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 42; участок CDR1 легкой цепи, представленный SEQ ID NO: 47; участок CDR2 легкой цепи, представленный SEQ ID NO: 50; и участок CDR3 легкой цепи, представленный SEQ ID NO: 52. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 75 и 76, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 33; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 35; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 43; участок CDR1 легкой цепи, представленный SEQ ID NO: 48; участок CDR2 легкой цепи, представленный SEQ ID NO: 50; и участок CDR3 легкой цепи, представленный SEQ ID NO: 52. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 77 и 78, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 30; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 37; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 44; участок CDR1 легкой цепи, представленный SEQ ID NO: 48; участок CDR2 легкой цепи, представленный SEQ ID NO: 50; и участок CDR3 легкой цепи, представленный SEQ ID NO: 52. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 79 и 80, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 31; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 38; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 45; участок CDR1 легкой цепи, представленный SEQ ID NO: 47; участок CDR2 легкой цепи, представленный SEQ ID NO: 50; и участок CDR3 легкой цепи, представленный SEQ ID NO: 53. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 81 и 82, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 32; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 39; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 46; участок CDR1 легкой цепи, представленный SEQ ID NO: 49; участок CDR2 легкой цепи, представленный SEQ ID NO: 51; и участок CDR3 легкой цепи, представленный SEQ ID NO: 54. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 85 и 86, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25FL и SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 55; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 57; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 59; участок CDR1 легкой цепи, представленный SEQ ID NO: 61; участок CDR2 легкой цепи, представленный SEQ ID NO: 63; и участок CDR3 легкой цепи, представленный SEQ ID NO: 65. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 67 и 68, но не ограничивается им.
В дополнение, указанным антителом предпочтительно является антитело, которое специфически связывается с SNAP25FL и SNAP25197 и содержит: участок CDR1 тяжелой цепи, представленный SEQ ID NO: 56; участок CDR2 тяжелой цепи, представленный SEQ ID NO: 58; участок CDR3 тяжелой цепи, представленный SEQ ID NO: 60; участок CDR1 легкой цепи, представленный SEQ ID NO: 62; участок CDR2 легкой цепи, представленный SEQ ID NO: 64; и участок CDR3 легкой цепи, представленный SEQ ID NO: 66. Более конкретно, указанным антителом может быть антитело, представленное SEQ ID NO: 69 и 70, но не ограничивается им.
В еще одном варианте осуществления настоящего изобретения предоставляется композиция антитела, содержащая указанное антитело, или чашку для культивирования, покрытую антителом, или набор, содержащий композицию антитела, или указанную чашку для культивирования.
В еще одном варианте осуществления настоящего изобретения предоставляется гибридомная клетка, способная продуцировать указанное антитело, при этом указанная гибридомная клетка представляет собой гибрид клетки селезенки и миеломной клетки мыши, инъецированной одним или несколькими пептидами, выбранными из группы, состоящей из SEQ ID NO: 1-10.
В еще одном варианте осуществления настоящего изобретения предоставляется способ определения активности ботулотоксина, включающий стадии: (а) обработки нейрональной клетки ботулотоксином; и (б) измерение SNAP25FL или SNAP25197 в нейрональной клетке с помощью одного или нескольких антител, выбранных из числа антител, представленных SEQ ID NO: 67-90, при этом указанным ботулотоксином является ботулотоксин типа А
В еще одном варианте осуществления настоящего изобретения предоставляется способ обнаружения ботулотоксина, включающий стадии: (а) обработки нейрональной клетки представляющим интерес образцом; (б) измерение SNAP25197 в нейрональной клетке с помощью одного или нескольких антител, выбранных из числа антител, представленных SEQ ID NO: 67-82, SEQ ID NO: 85 или SEQ ID NO: 86; и (в) определение того, что при измерении SNAP25197 ботулотоксин присутствует в образце, при этом указанным ботулотоксином является ботулотоксин типа А.
В еще одном варианте осуществления настоящего изобретения предоставляется способ с использованием клеток для анализа активности нейротоксина, включающий стадии: (а) культивирования нейрональной клеточной линии, полученной из Neuro-2a; (б) обработка нейрональной клеточной линии нейротоксином; (в) обработка линии нейрональных клеток или образца, полученного из линии нейрональных клеток, антителом, которое специфически связывается с SNAP25FL и SNAP25197; и (г) обработка нейрональной клеточной линии стадии (в) антителом, которое специфически связывается с SNAP25197 без связывания с SNAP25FL, при этом указанная клеточная линия, полученная из Neuro-2a, представляет собой линию клеток N2-42F (регистрационный номер: KCTC 13712ВР), указанный нейротоксин представляет собой ботулотоксин, а указанный нейротоксин на стадии (б) разбавляют средой, содержащей GT1b (тринатриевая соль ганглиозида GT1b), и используют для обработки указанной линии клеток.
Указанное антитело, используемое на стадии (в) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 55 или 56; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 57 или 58; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 59 или 60; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 61 или 62; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 63 или 64; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 65 или 66. Более конкретно, указанное антитело, используемое на стадии (с) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 55; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 57; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 59; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 61; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 63; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 65. В дополнение, указанное антитело, используемое на стадии (в) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 56; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 58; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 60; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 62; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 64; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 66.
В дополнение, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из любого участка, выбранного из группы, состоящей из SEQ ID NO: 28-33; участок CDR2 тяжелой цепи, состоящий из любого участка, выбранного из группы, состоящей из SEQ ID NO: 34-39; участок CDR3 тяжелой цепи, состоящий из любого участка, выбранного из группы, состоящей из SEQ ID NO: 40-46; участок CDR1 легкой цепи, состоящий из любого участка, выбранного из группы, состоящей из SEQ ID NO: 47-49; участок CDR2 легкой цепи, состоящий из любого участка, выбранного из группы, состоящей из SEQ ID NO: 50-51; и участок CDR3 легкой цепи, состоящий из любого участка, выбранного из группы, состоящей из SEQ ID NO: 52-54. Более конкретно, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 28; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 34; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 40; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 47; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52. В дополнение, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 29; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 35; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 41; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 48; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52. В дополнение, указанное антитело, используемое на стадии (d) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 29; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 36; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 42; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 47; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52. Кроме того, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 33; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 35; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 43; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 48; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52. Кроме того, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 30; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 37; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 44; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 48; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52. В дополнение, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 31; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 38; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 45; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 47; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 53. В дополнение, указанное антитело, используемое на стадии (г) способа анализа активности нейротоксина с использованием клеток, содержит: участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 32; участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 39; участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 46; участок CDR1 легкой цепи, состоящий из SEQ ID NO: 49; участок CDR2 легкой цепи, состоящий из SEQ ID NO: 51; и участок CDR3 легкой цепи, состоящий из SEQ ID NO: 54.
В еще одном варианте осуществления настоящего изобретения предоставляется среда для культивирования клеток для обработки нейрональной клетки нейротоксином, содержащая GT1b (тринатриевую соль ганглиозида GT1b). Концентрация GT1b в среде для культивирования клеток может составлять от 25 до 75 мкг/мл, а среда для культивирования клеток может дополнительно содержать креатин и аргинин. Кроме того, концентрация креатина в среде для культивирования клеток может составлять от 0,1 до 10 мМ, а концентрация аргинина в среде для культивирования клеток может составлять от 0,5 до 50 мМ. Кроме того, средой для культивирования клеток может быть среда RPMI 1640 (Roswell Park Memorial Institute 1640).
Далее будет подробно описана каждая стадия настоящего изобретения.
Преимущество настоящего изобретения
В последнее время наблюдается быстрый рост спроса на ботулотоксин для медицинских и косметических целей, но нет стабильного и воспроизводимого способа анализа для измерения эффективности ботулотоксина с использованием клеток. Поскольку ботулотоксин является очень мощным белком нейротоксином, разработка высокоспецифичных и чувствительных клеток и антител особенно необходима для точного измерения активности ботулотоксина с использованием клеток.
Настоящее изобретение относится к антителу для определения активности ботулотоксина и композиции антител, содержащей его. Новая линия клеток в соответствии с настоящим изобретением имеет значительно более короткое время удвоения по сравнению с обычными клетками SiMa, которые используются для определения активности ботулотоксина или для обнаружения ботулотоксина, а также имеет значительно более высокую чувствительность к ботулотоксину по сравнению с родительской линией клеток, что указывает на то, что она очень подходит для определения или обнаружения активности ботулотоксина с использованием клеток. Кроме того, линия клеток согласно настоящему изобретению может быть прикреплена и может стабильно культивироваться в культуральной чашке, покрытой поли-d-лизином (PDL), и, таким образом, может очень эффективно использоваться для определения или обнаружения активности ботулотоксина с использованием клеток.
Антитело против ботулотоксина согласно настоящему изобретению представляет собой моноклональное антитело, обладающее специфичностью связывания с  SNAP25FL,
SNAP25FL,  SNAP25197 или
SNAP25197 или  SNAP25FL и SNAP25197, и имеет превосходную специфичность и чувствительность. Таким образом, ожидается его активное использование в фармацевтической и косметической сферах.
SNAP25FL и SNAP25197, и имеет превосходную специфичность и чувствительность. Таким образом, ожидается его активное использование в фармацевтической и косметической сферах.
Кроме того, настоящее изобретение относится к анализу оптимальной активности клеток (СВРА), в котором используются клетки N2-42F и моноклональные антитела, обладающие значительно более высокой аффинностью связывания и специфичностью в отношении SNAP25, и этот анализ СВРА позволяет измерять активность ботулотоксина от 0,5 ед. или менее. Ожидается, что анализ СВРА, использующий клетки и антитело согласно настоящему изобретению, станет высоконадежным и воспроизводимым анализом активности ботулотоксина с использованием клеток.
Описание чертежей
На Фиг. 1 показаны результаты вестерн-блоттинга, проведенного для измерения чувствительности к ботулотоксину A (BoNT/A) в нейрональных клетках согласно одному из примеров согласно настоящему изобретению. Дорожка М представляет собой маркер размера белка; дорожка 1 представляет уровень экспрессии белка SNAP25 в общем клеточном лизате, не обработанном BoNT/A; а дорожка 2 показывает уровень экспрессии белка SNAP25 в общем клеточном лизате, обработанном BoNT/A. Кроме того, N2a обозначает клетки Neuro-2a, а K-ВМ1 обозначает клетки KP-N-RT-BM-1.
На Фиг. 2 показаны результаты вестерн-блоттинга, проведенного для изучения степени расщепления SNAP25, подтвержденной в трехэтапном процессе клональной селекции согласно одному из примеров согласно настоящему изобретению. При 2-м клональном отборе только клон 42, показывающий значительное расщепление SNAP25, вызванное BoNT/A, был выбран из 6 клонов, включающих клон 42, и при 3-м клональном отборе клон 24 (42F) последовательно показывает расщепление SNAP25 под действием BoN/A во множестве одинаковых экспериментов.
На Фиг. 3 представлен график, показывающий результаты измерения времени удвоения клеток N2-42F и их родительских клеток Neuro-2a согласно одному из примеров согласно настоящему изобретению.
На Фиг. 4 показаны изображения в 20-кратном увеличении клеток Neuro-2a (которые являются клетками родительской линии), клеток SiMa и клона N2-42F, полученные с использованием Leica DMi8, при достижении 60% конфлюентности, чтобы подтвердить морфологию клеток в соответствии с одним из примеров согласно настоящему изобретению.
На Фиг. 5 показаны изображения клона N2-42F, культивированного в чашках, покрытых каждым из следующих соединений: коллагеном IV типа, желатином и поли-D-лизином, в соответствии с одним из примеров согласно настоящему изобретению.
На Фиг. 6а и 6б показаны результаты вестерн-блоттинга, проведенного для изучения степени расщепления SNAP25, которое появляется, когда клетки SiMa и N2-42F обрабатывали различными концентрациями BoNT/A, согласно одному из примеров согласно настоящему изобретению, и степень расщепления SANP25 или Vamp2, которое происходит при обработке N2-42F и Neuro-2a различными типами ботулотоксина.
На Фиг. 7а и 7б показаны результаты анализа вестерн-блоттинга, проведенного для изучения стабильности N2-42F при пассировании, полученного в процессе клональной селекции согласно настоящему изобретению, согласно одному из примеров согласно настоящему изобретению.
На Фиг. 8 представлена схема, показывающая положения пептидов антигена SNAP25 для получения моноклонального или поликлонального антитела с использованием синтетических пептидов согласно одному из примеров согласно настоящему изобретению.
На Фиг. 9 представлена схема, показывающая процесс образования клеток гибридомы для получения моноклонального антитела и процесс скрининга клонов согласно одному из примеров согласно настоящему изобретению.
На Фиг. 10а-10в показаны результаты первоначального скрининга гибридомных клеток с использованием ELISA для получения моноклонального антитела согласно одному из примеров согласно настоящему изобретению.
На Фиг. 11а-11в показаны результаты повторного скрининга клеток для получения клонов одной клетки на основе результатов первоначального скрининга клеток гибридомы согласно одному из примеров согласно настоящему изобретению.
На Фиг. 12а-12в показаны результаты второго повторного скрининга для получения клонов одной клетки согласно одному из примеров согласно настоящему изобретению.
На Фиг. 13а-13с показаны результаты третьего повторного скрининга для получения клонов одной клетки согласно одному из примеров согласно настоящему изобретению.
На Фиг. 14а и 14б показан образец IgG, выделенного из белка сыворотки кролика, для получения поликлонального антитела согласно одному из способов согласно настоящему изобретению. На Фиг. 14а дорожка М представляет собой маркер размера белка; полоса 1 представляет собой элюат; дорожка 2 представляет собой пул элюированных IgG при рН 5,5; дорожка 3 представляет собой пул элюированных IgG при рН 4,0; дорожка 4 представляет собой пул элюированных IgG при рН 2,5; и дорожка 5 представляет собой пул элюированных IgG при рН 11,5.
На Фиг. 15а и 15б показаны результаты кинетического анализа моноклональных антител, полученных в соответствии с настоящим изобретением согласно одному из способов согласно настоящему изобретению. На Фиг. 15 IgG, загруженные на биосенсоры АМС, включают С16 IgG (I), С24 IgG (II), С4 IgG (III) и С7 IgG (IV); а) представляет загрузку антитела; 6) представляет собой промывку; в) представляет собой ассоциацию антигена; и г) представляет собой диссоциацию антигена.
На Фиг. 16а и 16б показаны результаты кинетического анализа моноклональных антител, полученных в соответствии с настоящим изобретением, ассоциированных и диссоциированных с серийно разведенным рекомбинантным GST-SNAP25, согласно одному из примеров согласно настоящему изобретению. На Фиг. 16 IgG, загруженные на биосенсоры АМС, включают С14 (I), С24 (II), D2 (III), D6 (IV), Е6 (V) и А15 (VI); а) представляет загрузку антитела; б) представляет собой ассоциацию антигена; и в) представляет собой диссоциацию антигена.
На Фиг. 17 показаны результаты вестерн-блоттинга, проведенного для изучения специфичности связывания антигена моноклональных антител, полученных в соответствии с настоящим изобретением, согласно одному из примеров согласно настоящему изобретению. На Фиг. 17 полоса 1 представляет собой SNAP25FL, а полоса 2 представляет собой GST-SNAP25197.
На Фиг. 18а и 18б показаны результаты вестерн-блоттинга, проведенного для изучения специфичности связывания антигена моноклонального антитела, полученного в соответствии с настоящим изобретением, и конъюгированного с пероксидазой хрена (HRP), согласно одному из примеров согласно настоящему изобретению. На Фиг. 18а дорожка М представляет собой маркер размера белка; полоса 1 представляет собой неконъюгированный С16 IgG (9 мг); дорожка 2 представляет активированную HRP (4 мг); дорожка 3 представляет смесь С16 IgG/HRP (С16 IgG-HRP) перед инкубацией (4,5 мг); полоса 4 представляет собой С16 IgG-HRP после инкубации (4,5 мг); полоса 5 представляет собой С16 IgG-HRP после блокирования (4,3 мг); полоса 6 представляет собой С16 IgG-HRP после удаления свободной HRP диализом (4,3 мг); и а) представляет собой конъюгат С16 IgG-HRP.
На Фиг. 19а-19в показаны результаты электрофореза в ДСН-ПААГ моноклонального антитела, полученного согласно настоящему изобретению и конъюгированного с биотином, согласно одному из примеров согласно настоящему изобретению. На Фиг. 19а и 19б дорожка М представляет маркер размера белка; дорожка 1 представляет собой только А15 IgG; полоса 2 представляет собой IgG А15, конъюгированный с 0,1 мМ биотина; дорожка 3 представляет собой IgG А15, конъюгированный с 0,25 мМ биотина; и полоса 4 представляет собой IgG А15, конъюгированный с 0,5 мМ биотина.
На Фиг. 20а и 20б показаны результаты электрофореза в ДСН-ПААГ поликлонального антитела, полученного согласно настоящему изобретению и конъюгированного с АР, согласно одному из примеров согласно настоящему изобретению. На Фиг. 20а дорожка М представляет маркер размера белка; дорожка 1 представляет собой неконъюгированный IgG; дорожка 2 представляет собой АР; дорожка 3 представляет собой конъюгаты AP-IgG; а) представляет собой концентрирующую часть полиакриамидного геля; и б) представляет собой конъюгаты AP-IgG.
На Фиг. 21а-21в показаны результаты оптимизации времени обработки токсином в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 22 показаны результаты оптимизации среды обработки токсином в способе определения активности ботулотоксина согласно одному из способов согласно настоящему изобретению.
На Фиг. 23а и 23б показаны результаты оптимизации сенсибилизатора в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 24а-24в показаны результаты оптимизации GT1b в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 25а и 25б показаны результаты оптимизации N2/B27 в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 26а и 26б показаны результаты оптимизации обработки связывающими антителами в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 27а и 27б показаны результаты оптимизации обработки детектирующими антителами в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 28 показаны результаты оптимизации способа определения активности конъюгатов HRP в способе определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 29 показана схема, показывающая метод ELISA типа «сэндвич» для определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 30а-30в показаны результаты исследования точности и линейности метода ELISA типа «сэндвич» для определения активности ботулотоксина согласно одному из примеров согласно настоящему изобретению.
На Фиг. 31а-31в показаны результаты измерения биологической активности методом ELISA типа «сэндвич» для определения активности ботулотоксина согласно одному из способов согласно настоящему изобретению.
Наилучший способ осуществления настоящего изобретения
Настоящее изобретение состоит в том, чтобы преодолеть ограничения обычного СРВА и разработать более эффективный СВРА. Путем сравнительного анализа 13 различных линий нейрональных клеток с последующим обширным клональным отбором в настоящем изобретении была создана новая линия клеток N2-42F. В дополнение к чувствительности к BoNT/A, которую можно сравнить с чувствительностью SiMa, время удвоения, равное ~24 часа, и стабильное прикрепление к культуральному планшету, покрытому поли-d-лизином (PDL), делают клетки N2-42F очень привлекательными и надежными хозяевами для СВРА. Антитело согласно настоящему изобретению представляет собой моноклональное антитело, обладающее очень высокой аффинностью связывания и специфичностью в отношении SNAP25. Кроме того, настоящее изобретение относится к оптимальному анализу активности с использованием клеток (СВРА), в котором используются клетки N2-42F и моноклональные антитела, обладающие очень высокой аффинностью связывания и специфичностью в отношении SNAP25, и этот СВРА может измерять активность ботулотоксин от 0,5 Ед или менее.
Примеры
Далее настоящее изобретение будет более подробно со ссылкой на примеры. Для специалистов в данной области техники будет очевидно, что эти примеры предназначены исключительно для иллюстративных целей и не предназначены для ограничения объема настоящего изобретения.
Материалы и методы 1: Реагенты для получения клеток
Материалы, используемые для получения клеток согласно настоящему изобретению, описаны ниже.
0,25% трипсин-ЭДТА (Gibco™ 25200056), GlutaMAX™ (Gibco™ 35050061), MEM (Gibco™ 11095080), заменимые аминокислоты MEM (NEAA) (100х) (Gibco™ 11140050), пируват натрия (Gibco™ 11360070), экспресс-фермент TrypLE™ (1x) (Gibco™ 12604021), раствор антимикотика и антибиотика (АА) (100х) (Sigma А5955), борная кислота (Sigma В6768), коллаген из плаценты человека (Sigma С5533), ДТТ (Sigma D0632), ДМСО (Sigma D2650), раствор желатина (Sigma G1393), гидробромид поли-D-лизина (Sigma Р6407), полисорбат (Sigma Р7949), тетраборат натрия (Sigma 221732), DPBS (Welgene LB001-02), фетальная бычья сыворотка (ФБС) (YI Frontier US-ФБС-500), глицерин (Affymatrix USB 16374), набор ПЦР для определения микоплазмы (Takara Bio 6601), буфер RIPA (10х) (abcam ab156034), 6-луночный планшет (Falcon 353046), 12-луночный планшет (Corning CLS3513), 24-луночный планшет (Falcon 353047), колба Т75 (Falcon BD353136), TaKaRa EX Tag™ (Takara Bio RR001)
Материалы и методы 2. Приготовление исходного раствора ботулотоксина, разбавителя и токсичной среды BoNT/A.
Очищенный ботулотоксин серотипа A(BoNT/А) получали от Hugel (EXBII1501).
Материалы и методы 2-1. Приготовление рабочего исходного раствора BoNT/A
Очищенный BoNT/A получали от Hugel (EXBII1501). BoNT/A разбавляли для получения рабочего исходного раствора (10 нМ) с использованием буфера для разведения токсина, состоящего из 50 мМ фосфата натрия, рН 7,0, 1 мМ ДТТ, 0,05% полисорбата, 20% глицерина и 0,2 мг/мл ацетилированного БСА. Исходный раствор BoNT/A перед использованием хранили в аликвотах при -80°С. Исходный раствор BoNT/A готовили следующим образом.
(1) Основной исходный раствор (200 Ед/мл): ресуспендировали лиофилизированный BoNT/A (200 Ед) в 1 мл интоксикационной среды или физиологического раствора и оставляли при комнатной температуре на 10 мин.
(2) Исходный раствор А (50 Ед/мл): растворяли аликвоту 150 мкл основного исходного раствора в стерильной пробирке для микроцентрифугирования в 450 мкл интоксикационной среды при комнатной температуре (т.е. разводили основной исходный раствор в пропорции 1:4).
(3) Исходный раствор В (5 Ед/мл): растворяли аликвоту 20 мкл исходного раствора А в стерильной пробирке для микроцентрифугирования в 180 мкл интоксикационной среды при комнатной температуре (т.е. разводили исходный раствор А в пропорции 1:10).
(4) Исходный раствор С (0.5 Ед/мл): растворяли аликвоту 20 мкл исходного раствора В в пробирке, содержащей 180 мкл интоксикационной среды при комнатной температуре (т.е. разводили исходный раствор В в пропорции 1:10).
Материалы и методы 2-2. Подготовка образца для построения стандартной кривой
Стандартные эталонные образцы BoNT/А для построения стандартной кривой готовили следующим образом.
1. Стандартный исходный раствор А (50 пМ, 113,6 Ед/мл): ресуспендировали лиофилизированный BoNT/A (100 Ед) в 880 мкл интоксикационной среды или физиологического раствора и оставляли при комнатной температуре на 10 мин.
2. Стандартный исходный раствор В (10 пМ, 22,7 Ед/мл): растворяли аликвоту 50 мкл исходного раствора А в стерильной пробирке для микроцентрифугирования в 200 мкл среды для интоксикации при комнатной температуре (т.е. разводили исходный раствор А в пропорции 1:5).
3. Стандартный исходный раствор С (2 пМ, 4,54 Ед/мл): растворяли аликвоту 50 мкл исходного раствора В в стерильной пробирке для микроцентрифугирования в 200 мкл интоксикационной среды при комнатной температуре (т.е. разводили исходный раствор В в пропорции 1:5).
Материалы и методы 3. Покрытие планшетов для культуры нейрональных клеток
Планшет для культивирования покрывали в течение ночи либо раствором желатина (0,1% в 1x PBS), либо коллагеном типа IV (0,1 мг/мл), либо поли-D-лизином (PDL) (50 мкг/мл). Лиофилизированный коллаген восстанавливали в деионизированной H2O до конечной концентрации 0,1 мг/мл. Раствор PDL готовили растворением 5 мг порошка в 0,1 М боратном буфере, рН 8,5, до рабочей концентрации 50 мкг/мл. Планшет для культивирования дважды промывали 1x DPBS и сушили на воздухе в вытяжном шкафу для культивирования тканей.
Материалы и методы 4. Размножение линий клеток и питательной среды
Тринадцать нейрональных линий клеток были собраны из 5 различных институтов (Таблица 1). Нейрональные клетки были приобретены в Американской коллекции культур тканей (АТСС), Немецкой коллекции микроорганизмов и клеточных культур (DSMZ), Японской коллекции исследовательских биоресурсов (JCRB), Корейской коллекции типовых культур (KCTC) и Корейском банке линий клеток (KCLB). Их поддерживали и размножали в рекомендованной среде. Загрязнение микоплазмой всех клеточных линий контролировали с помощью набора ПЦР для обнаружения микоплазмы (Takara Bio Inc. 6601) через каждые 4 пассажа клеток N2-42F или 10 пассажей клеток SiMa и гибридомы. ПЦР-тест проводили согласно рекомендованной методике. Вкратце, культуральный супернатант собирали и инкубировали в течение 3-4 дней. ПЦР (50 мкл) выполняли с аликвотами (3 мкл) культурального супернатанта, 1х буфером для ПЦР, смесью dNTP, праймерами MCGp F1/R2 и TaKaRa EX Taq™ (Takara Bio RR001). Аликвоты (10 мкл) продуктов ПЦР разделяли на 1% агарозном геле и визуализировали окрашиванием этидием.
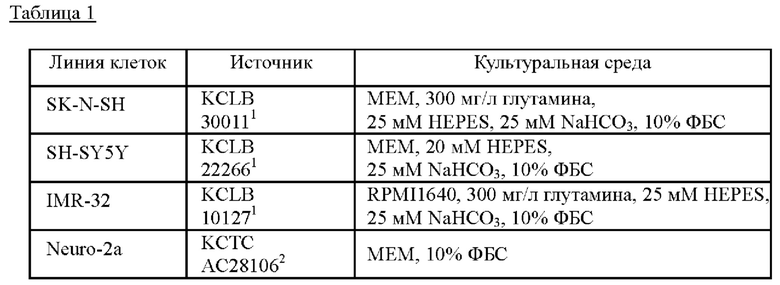
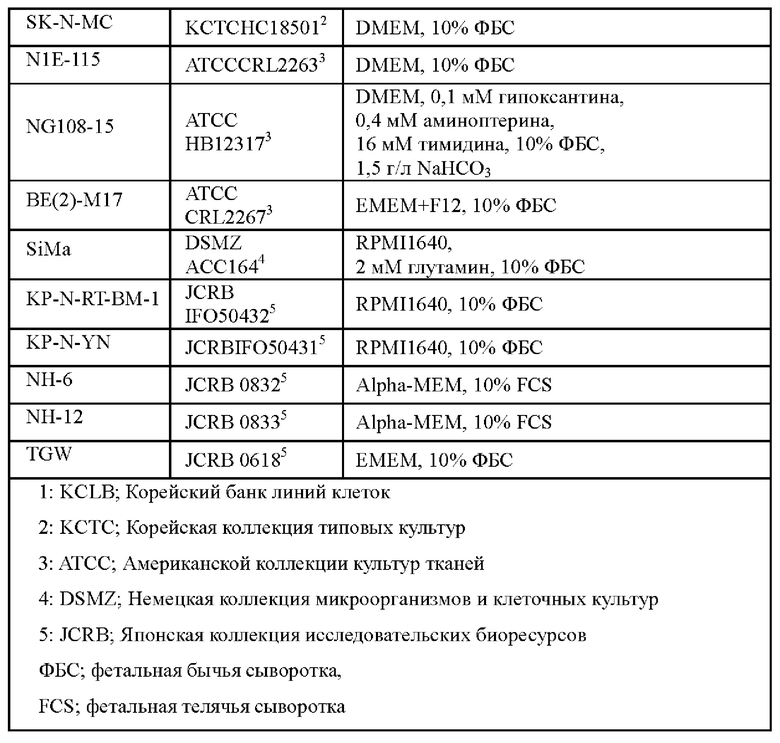
Материалы и методы 5: Реагенты для получения антител
Материалы, использованные для получения антител согласно настоящему изобретении, описаны ниже.
2-меркаптоэтанол (Sigma М3148), 4-йодфенилборная кислота (Sigma 471933), 10х TBS (BIO-RAD 170-6435), буфер 10х Трис/Глицин/ДСН (Bio-Rad 161-0772), 12% Mini-PROTEAN® TGX™ (Bio-Rad 456-1046), уксусная кислота (Merck 100063), щелочная фосфатаза (Sigma P0114), AMICON Ultra-15, Ultracel 30K (Millipore UFC903024), раствор антимикотика и антибиотика (АА) (100х) (Sigma А5955), бромфеноловый синий (Sigma В0126), DMEM (Gibco™ 11995065), ДМСО (Sigma D2650), ДМСО (Sigma 472301), EZ-link NHS-PEG4-Biotin (Thermo Fisher Scientific 21329), этиленгликоль (Sigma 324558), фетальная бычья сыворотка (ФБС) (YI Frontier US-ФБС-500), глицерин (Affymetrix USB 16374), глицин (Bioshop GLN001), раствор глутаральдегида (Sigma G7651), пероксидаза хрена (Sigma Р6782), раствор пероксида водорода (Sigma 216763), люминол (Sigma 123072), хлорид магния (Sigma М8266), заменимые аминокислоты MEM (NEAA) (100х) (Gibco™ 11140050), метанол (Merck 106009), PCR® STRIP TUBE (Axygen PCR-0208-CP-C), поливинилиденфторидная (ПВДФ) мембрана (Millipore ISEQ00010), хлорид калия (Sigma Р9333), 20% (вес/об) раствор ДСН (Bio-Rad 161-0418) стандартов двойного цвета Precision plus protein™ (Bio-Rad 1610374), обезжиренное молоко (BD Difco™ 232-100), ацетат натрия (Sigma W302406), бикарбонат натрия (Sigma S6014), боргидрид натрия (Sigma 452882), хлорид натрия (Merck 106404), (мета)перйодат натрия (Sigma S1878), двухосновный фосфат натрия (Sigma S7907), одноосновный фосфат натрия (Sigma S5011), станнат натрия тригидрат (Sigma 336262), боргидрид тетрабутиламмония (Sigma, 230170), Т175 (SPL 74175), колба Т75 (SPL 70375), Трис (Bioshop TRS001), хлорид цинка (Sigma 229997)
Материалы и методы 6: Разработка метода производства антител
Материалы и методы 6-1. Создание поликлональных и моноклональных антител с использованием синтетических пептидов.
Синтетические пептиды конъюгировали с гемоцианином лимфы улитки (ГЛУ) либо на С-, либо на N-конце, как показано на Фиг. 8. Сначала двух кроликов иммунизировали пептидными антигенами для получения поликлональной сыворотки. После первоначальной инъекции кроликов периодически бустировали в течение 6 недель. Сыворотки кроликов тестировали на их реактивность и специфичность с помощью вестерн-блоттинга и ELISA. Затем сыворотки хранили при -80°С перед использованием. Потом, чтобы получить моноклональное антитело, четырем мышам инъецировали пептидные антигены. Затем собирали продуцирующие антитела спленоциты и объединяли их с клетками миеломы с образованием гибридом, после чего следовали три раунда одноклеточного клонального отбора, как показано на Фиг. 9. Вкратце, гибридомы сначала подвергали скринингу с помощью ELISA с использованием пептидных антигенов, затем путем Вестерн-блоттинг с использованием рекомбинантных белков SNAP25 (GST-SNAP25FL и GST-SNAP25197) или общих клеточных лизатов, полученных из нейрональных клеток (SiMa), и, наконец, сэндвич-ELISA с общими клеточными лизатами. Гибридомы, продуцирующие антитела, размножали и хранили в паровой фазе жидкого азота до тех пор, пока они не были восстановлены для производства антител, как подробно описано ниже.
Материалы и методы 6-2. Получение аффинной колонки с SNAP25
Рекомбинантный SNAP25197 концентрировали до 10 мг/мл с использованием AMICON Ultra-15 путем повторного центрифугирования при 1000х g в течение 10 мин при 4°С, при этом измеряя концентрацию белка с помощью нано-спектрофотометра (Drawell Scientific Instrument Co., Ltd, Шанхай). Аликвоту (3 мл) SNAP25197 в AMICO Ultra-15 смешивали с 12 мл буфера для связывания (0,1 М HEPES, рН 7,5, 0,1 М NaCl) и повторно концентрировали центрифугированием при 1000х g. Эту замену буфера повторяли 6 раз, и концентрат SANP25197 добавляли к 1 мл суспензии Affi-Gel 15 (Bio-Rad) в деионизированной Н2О. После инкубации на шейкере-инкубаторе в течение 1 часа при комнатной температуре Affi-Gel 15 центрифугировали при 1000х g в течение 10 минут при 4°С. Этот конъюгированный с SNAP25197 Affi-Gel 15 (SNAP25-AffiGel) ресуспендировали в 10 мл 10 мМ гидрохлорида этанол амина, рН 8,0, и инкубировали в течение 1 часа при комнатной температуре. После промывания 1x PBS перед использованием SNAP25-AffiGel хранили в 1x PBS, содержащем 0,2% азида натрия.
Материалы и методы 6-3. Очистка поликлонального антитела
Сыворотку кролика разводили в 10 раз 10 мМ Трис-HCl, рН 7,5, и центрифугировали при 10,000х g в течение 10 мин при 4°С. После фильтрации через фильтр на крышке бутылки 0,45 мкм (Nalgene™) прозрачный разбавитель сыворотки трижды пропускали через SNAP25-AffiGel. Затем SNAP25-AffGel промывали 20 объемами колонки 10 мМ Tris-HCl, рН 7,5 и 0,5 М NaCl, и связанные белки последовательно элюировали 12 объемами колонки 0,1 М ацетата натрия, рН 5,5, 0,1 М глицина, рН 4,0, и 0,1 М глицина, рН 2,5 и 0,1 М триэтиламина, рН 11,5. Во время элюирования образцы белка собирали в пробирки, содержащие 0,1 мл 1 М Трис-HCl, рН 8,0. Фракции пиков, содержащие белок, объединяли и концентрировали до 1 мл с помощью AMICON Ultra-15. После диализа с четырьмя заменами 1x PBS и 10% глицерина (0,5 л) каждые 90 мин их анализировали с помощью ДСН-ПААГ и ELISA.
Материалы и методы 6-4. Конъюгация антител с пероксидазой хрена (HRP)
Для конъюгации с HRP очищенный В4 или С16 IgG концентрировали до 10 мг/мл с помощью AMICON Ultra-15 путем повторных центрифугирований при 1000 g в течение ~15 мин на каждое центрифугирование при 4°С, обеспечивая при этом избыточный объем буфера для конъюгации (0,1 М NaHCO3, рН 9,5, 0,9% NaCl) в конце каждого центрифугирования. HRP (5 мг) солюбилизировали в 1,2 мл деионизованной Н2О и смешивали с 0,3 мл 0,1 М периодата натрия в 10 мМ фосфате натрия, рН 7,0. После инкубации в течение 20 минут при комнатной температуре раствор HRP подвергали диализу с четырьмя заменами 1 мМ ацетата натрия, рН 4,0, в течение 6 часов при 4°С. Концентрированное антитело (5 мг) и активированную HRP (5 мг) смешивали вместе в пробирке для микроцентрифугирования и инкубировали в течение 2 часов при комнатной температуре с защитой от света. Реакцию конъюгации останавливали добавлением 0,1 мл боргидрида натрия (4 мг/мл в деионизированной H2O). Конъюгат HRP-антитело подвергали диализу с использованием Pur-A-Lyzer Maxi 50000 с тремя заменами 1x PBS и один раз с 1x PBS/50% глицерин с часовым интервалом при 4°С. Конъюгат HRP-антитело хранили при 4°С или -80°С для длительного хранения перед использованием.
Материалы и методы 6-5. Конъюгация антитела с активированным биотином
А15 IgG (1 мг/мл) переносили в Pur-A-Lyzer Maxi 20000 и подвергали диализу с четырьмя заменами реакционного буфера, состоящего из 0,1 М фосфата, рН 7,2 и 0,15 М NaCl с часовым интервалом при 4°С с защитой от света. Активированный биотин (10 мМ в реакционном буфере) (EZ-Link NHS-PEG4-Biotin) смешивали с 50 мкг аликвоты А15 IgG до конечной концентрации 0,1 мМ, 0,25 мМ или 0,5 мМ. Смесь доводили до 100 мкл реакционным буфером и инкубировали в течение 2 часов при 4°С с защитой от света. После добавления 2 мкл 0,1 М глицина реакционную смесь подвергали диализу с тремя заменами 1x PBS и один раз с 1x PBS/50% глицерин с часовым интервалом при 4°С и хранили в пробирке для микроцентрифугирования при -20°С перед использованием. Степень биотинилирования IgG оценивали с использованием набора Pierce Biotin Quantitation Kit (Thermo Scientific 28005), а реактивность и специфичность биотинилированного IgG исследовали с помощью сэндвич-ELISA.
Материалы и методы 6-6. Сшивание щелочной фосфатазы (ЩФ) с антителами
Конъюгирование щелочной фосфатазы (ЩФ) с антителом инициировали добавлением глутарового альдегида до 0,25% в смеси (20 мкл), состоящей из реакционного буфера (0,1 М фосфат натрия, рН 6,8), 50 мкг ЩФ (2,2 мг/мл) (Sigma-Aldrich P0114-10KU), и 100 мкг очищенного IgG, такого как моноклональное антитело А15, поликлональное антитело rA15 IgG и поликлональный анти-SNAP25 IgG (Sigma-Aldrich S9684). После инкубации на льду в течение 1 часа с защитой от света в реакционную смесь добавляли 1 мкл 1 М этаноламина и инкубировали в течение 1 часа при комнатной температуре с защитой от света. Конъюгат AP-IgG подвергали диализу с тремя заменами 1x PBS и один раз с буфером для хранения (25 мМ Tris-HCl, рН 7,5, 1 мМ MgCl2, 0,1 мМ ZnCl2 и 50% глицерин) с часовым интервалом при 4°С.Специфичность связывания антигена и АР-активность конъюгата AP-IgG исследовали прямым ELISA, описанным ниже.
Материалы и методы 6-7. Измерение KD посредством OCTET RED96
Для анализа кинетики моноклональных антител выполняли анализ биослойной интерферометрии (BLI) при 30°С с использованием прибора  Octet Red96 в соответствии с процедурой, рекомендованной производителем. Вкратце, рекомбинантный GST-SNAP25FL или SNAP25197 разбавляли до 125 или 250 нМ в 1х буфере для измерения кинетики (1x KB)/1х PBS, и аналит, т.е. очищенный IgG, серийно разбавляли до 3,9, 7,8, 15,6, 31,25, 62,5, 125, 250 нМ в 1х буфере для измерения кинетики. После уравновешивания в 1х буфере для измерения кинетики в течение 1 минуты зонды анти-GST (
Octet Red96 в соответствии с процедурой, рекомендованной производителем. Вкратце, рекомбинантный GST-SNAP25FL или SNAP25197 разбавляли до 125 или 250 нМ в 1х буфере для измерения кинетики (1x KB)/1х PBS, и аналит, т.е. очищенный IgG, серийно разбавляли до 3,9, 7,8, 15,6, 31,25, 62,5, 125, 250 нМ в 1х буфере для измерения кинетики. После уравновешивания в 1х буфере для измерения кинетики в течение 1 минуты зонды анти-GST ( 18-5096) загружали GST-SANP25FL или GST-SNAP25197 на 30 минут с последующим погружением в 1х буфер для измерения кинетики на 10 минут. После ассоциации и диссоциации аналита в течение 10 мин для каждой процедуры были получены кинетические кривые, и KD (константу равновесной диссоциации) оценивали с использованием программного обеспечения для анализа
18-5096) загружали GST-SANP25FL или GST-SNAP25197 на 30 минут с последующим погружением в 1х буфер для измерения кинетики на 10 минут. После ассоциации и диссоциации аналита в течение 10 мин для каждой процедуры были получены кинетические кривые, и KD (константу равновесной диссоциации) оценивали с использованием программного обеспечения для анализа  Octet.
Octet.
В качестве альтернативного кинетического анализа биосенсоры захвата Fc IgG мыши (АМС) загружали антителом и подвергали ассоциации/диссоциации с серийно разведенным GST-SNAP25. Вкратце, очищенный IgG разбавляли до 100 или 200 нМ, тогда как GST-SNAP25FL или SNAP25197 серийно разбавляли до 1,56, 3,125, 6,25, 12,5, 25, 50 и 100 нМ. После уравновешивания в 1х буфере для измерения кинетики в течение 1 минуты биосенсоры АМС загружали IgG на 10 минут с последующим погружением в 1х буфер для измерения кинетики на 10 минут. Ассоциацию и диссоциацию аналита проводили по 10 мин каждая и оценивали KD, как описано выше.
Материалы и методы 6-8. Прямой метод ELISA
Иммунопланшет (Thermo Fisher Scientific А71125) покрывали в течение 2 часов при 37°С 1 мкг GST-SNAP25FL или GST-SNAP25197 в 0,1 М карбонатном буфере, рН 9,5. После промывания 1xPBS иммунопланшет инкубировали с 300 мкл блокирующего буфера (5% обезжиренное сухое молоко в 1xPBS) в течение 15 минут при комнатной температуре. После трехкратной промывки 1xPBST (1xPBS/0,05% Твин-20) в микропланшет вносили 100 мкл супернатанта гибридомной культуры (разведение 1:20~10,000) или асцитную жидкость (разведение 1:1,000~312,500) и инкубировали в течение 1 ч при комнатной температуре. Микропланшет трижды промывали 1xPBST, к которому добавляли аликвоты (100 мкл на лунку) конъюгата козьего антимышиного IgG-HRP (разведение 1:1000) (Ab Frontier LF-SA8001). После инкубации в течение 1 часа при комнатной температуре микропланшет трижды промывали 1xPBST и проводили реакцию HRP с 50 мкл 1-StepTMUltra TMB-ELISA (Thermo Fisher Scientific 34028) при комнатной температуре в течение 3-25 минут. Реакцию HRP останавливали добавлением 50 мкл 1 М H2SO4 и сигнал ELISA оценивали при 450 нм с помощью Bio-Tek SynergyNeo2.
Материалы и методы 6-9. Анализ вестерн-блоттинг
Рекомбинантный GST-SNAP25 (20 нг ~1 мкг на лунку) или общий клеточный лизат клеток Neuro-2a (15 мкг на лунку) разделяли вместе со стандартами белка Precision (3 мкл на лунку) с помощью 10% или 12% ДСН-ПААГ. После 5 мин замачивания в буфере для переноса, состоящем из 48 мМ Трис, 38,9 мМ глицина, 20% метанола, 0,05% ДСН, белки переносили на ПВДФ-мембрану с использованием Trans-Blot® Semi-Dry (Bio-Rad 170-3940). в течение 45 минут при 25 В. ПВДФ-мембрану на короткое время промывали 1xTBST (1xTBS/0,05% Твин 20) и инкубировали с блокирующим буфером (5% обезжиренное сухое молоко в 1xTBST) в течение 15 минут при комнатной температуре. Затем мембрану из ПВДФ инкубировали с супернатантом любой культуры гибридомы (разведение 1:100 в блокирующем растворе) в течение 45 мин при комнатной температуре. Поликлональные IgG к SNAP25 (Sigma S9684) (разведение 1:8000) и IgG против SNAP25197 (R&D МС6050) (разведение 1:100) использовали в качестве положительных контролей. После отмывки 1xTBST три раза в течение 15 минут, ПВДФ-мембрану инкубировали либо с конъюгатом козьих антител против кроличьего IgG-HRP (разведение 1:10000), либо с конъюгатом козьих антител против мышиного IgG-HRP (разведение 1:10000) в течение 45 минут при комнатной температуре. После трехкратной промывки с помощью 1xTBST рекомбинантный GST-SNAP25 или эндогенный SNAP25 обнаруживали и количественно определяли с использованием раствора ECL (см. ниже) и системы визуализации Bio-Rad ChemiDoc ТМ MP (Bio-Rad Universal hood III).
Примеры
Пример 1: Скрининг нейрональных клеток, чувствительных к BoNT/A
Пример 1-1. Сравнительный анализ нейрональных линий клеток на их чувствительность к BoNT/A
Нейрональные клетки высевали из расчета 2×105 клеток/лунку в 24-луночный планшет, и через 24 часа их обрабатывали 2 нМ BoNT/А в среде, указанной в Таблице 1, в течение 3 дней. Общие клеточные лизаты (ОКЛ) получали с использованием 1xRIPA, а аликвоты (3,5 мкг) исследовали с помощью 12% ДСН-ПААГ.
SNAP25FL и SNAP25197 анализировали с помощью вестерн-блоттинга с использованием поликлональных IgG кролика (Sigma S9684), специфичных к SNAP25, как описано в разделе «Материалы и методы». Раствор ECL составлен и оптимизирован AbBio Inc. Рабочий раствор ECL получали путем смешивания SolA и SolB в соотношении 1:1 перед использованием. SolA и SolB представлены в Таблице 2. Чувствительность клеточных линий к BoNT/A (2 нМ) представлена в Таблице 3.

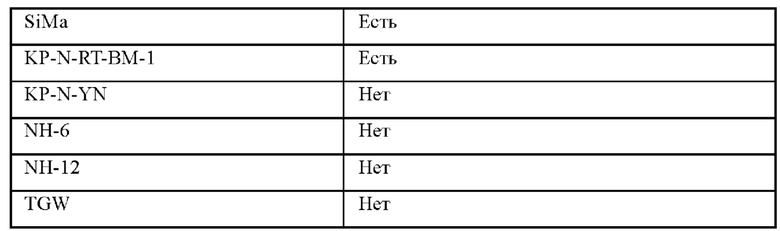
Как суммировано в Таблице 3, за исключением клеток SiMa, только клетки Neuro-2a и KP-N-RT-BM1 (K-ВМ1) давали детектируемые уровни расщепления SNAP25.
Пример 1-2. Скрининг нейрональных клеток, чувствительных к BoNT/A
Клетки Neuro-2a, SiMa и KP-N-RT-BM-1 отдельно культивировали на 24-луночном планшете в тех же условиях, что и в разделе «Материалы и методы», а феномен расщепления белка SNAP25 анализировали с помощью вестерн-блоттинга показано на Фиг. 1. Степень расщепления SNAP25 оценивается примерно в 33%, 66% и 29% для клеток Neuro-2a, SiMa и K-ВМ1, соответственно.
В дальнейшем для клональной селекции использовали Neuro-2a, а не K-ВМ1 по следующей причине. Во-первых, эффективность BoNT/A in vivo, которую обычно измеряют по летальности мышей (то есть, LD50 мыши), можно было бы лучше воспроизвести с помощью линии клеток мыши. Фактически, Neuro-2a - это линия клеток мыши, а K-ВМ1 - линия клеток человека. Во-вторых, K-ВМ1 имеет тенденцию расти гораздо медленнее, чем Neuro-2a (данные не показаны). Следует отметить, что время удвоения популяции клеток SiMa, линии клеток нейробластомы человека, составляет от 34 до 100 часов (DSMZ АСС-164). В-третьих, под микроскопом клетки Neuro-2a представляют собой смешанную популяцию нескольких типов клеток. Таким образом, вероятно, что клетки, чувствительные к BoNT/A, существуют в гетерогенной популяции клеток Neuro-2a.
Пример 2: Отбор клонов нейрональных клеток, чувствительных к BoNT/A
Среди популяций клеток Neuro-2a, выбранных в Примере 1, отбирали клоны, высокочувствительные к BoNT/A.
Пример 2-1. Клепальная культура клеток Neuro-2a
Клетки Neuro-2a размножали в MEM с добавлением 10% фетальной бычьей сыворотки (ФБС), 1х заменимых аминокислот (1x NEAA), 1х пирувата натрия, 1x GlutaMAX™ и 1x антибиотик-антимикотик (1x АА). Когда клетки достигли ~80-90% конфлюэнтности, их обрабатывали 1x TrypLE (Gibco™ 12604021) для получения суспензии отдельных клеток. После определения количества жизнеспособных клеток с помощью гемоцитометра и трипанового синего клетки разводили до плотности 10 клеток на мл среды. Затем аликвоты по 100 мкл (~эквивалент одной клетки) добавляли в 7х 96-луночный культуральный планшет. Рост колоний периодически контролировали под микроскопом. При ~60% слияния клетки переносили в 24-луночный культуральный планшет. Позже их делили поровну, и одну половину хранили в емкости с жидким азотом, а остальную тестировали на чувствительность к BoNT/A.
Клональные клетки Neuro-2a субкультивировали с 24-луночного планшета на 96-луночный микропланшет. На следующий день культуральную среду заменяли на PRMI1640 с добавлением 2 мМ L-аланил-L-глутамина, 1x В27, 1х N2, 1x NEAA (среда для дифференцировки) в течение 2 дней, чтобы вызвать дифференцировку нейронов. На 4-й день добавляли GT1b до конечной концентрации 25 мкг/мл (1х интоксикационная среда). После инкубации в течение 24 часов культуральную среду заменяли средой для интоксикации, содержащей BoNT/A (0,1 нМ), и клетки инкубировали еще 2 дня. После удаления среды путем аспирации клетки обрабатывали 1х ДСН-буфером для образцов, аликвоты исследовали на степень расщепления SNAP25 с помощью 12% ДСН-ПААГ и вестерн-блоттинга. Эту процедуру отбора клонов повторяли три раза, и результаты, подтвержденные в каждой процедуре отбора клонов, показаны на Фиг. 2. Следуя процедуре, оптимизированной для клеток SiMa компанией Allergan с небольшими модификациями, как описано в разделе «Материалы и методы» (PLoS One. 2012; 7 (11): е49516.), 142 клональных клетки перемещали из 24-луночного планшета в 96-луночный культуральный планшет и тестировали на чувствительность к BoNT/A. Как показано на Фиг. 2, всего было идентифицировано 19 положительных клонов, которые проявляли заметно более высокую чувствительность к BoNT/A, чем родительский Neuro-2a. Из положительных клонов, подвергнутых многократным циклам одноклеточной клональной селекции, только клон 42 постоянно проявлял более высокую чувствительность к BoNT/A, чем родительский Neuro-2a. После трех раундов одноклеточного клонального отбора клон 42 был переименован в N2-42F в соответствии с номенклатурой, рекомендованной для клеточных линий (Cell Stem Cell. 2011 Jun 3; 8(6):607-8.; Bioinformatics. 2008 Dec 1; 24(23):2760-6).
Пример 2-2. Восстановление, пролиферация и хранение замороженных клеток
Восстановление замороженных клеток проводили следующим образом..
(1) Быстро размораживали (<2 минут) пробирку с исходным раствором клеток, извлеченную из морозильной камеры с жидким азотом, осторожным встряхиванием в водяной бане при 37°С.
(2) После размораживания обеззараживали пробирку с исходным раствором клеток опрыскиванием 70% этанолом.
(3) Отвинчивали верхнюю часть флакона в вытяжном шкафу с ламинарным потоком для тканевых культур и переносили содержимое в стерильную коническую пробирку объемом 15 мл, содержащую 9 мл предварительно подогретой полной среды.
(4) После осторожного центрифугирования (125х g в течение 10 мин) удаляли супернатант путем аспирации и повторно суспендировали клетки в 2 мл полной среды.
(5) Осторожно пипетировали, чтобы разрыхлить осадок и разбить комки, и переносили суспензию клеток в колбу Т75, содержащую 25 мл полной среды.
(6) Инкубировали колбу при 37°С в СО2-инкубаторе.
Культивирование клеток проводили следующим образом.
(1) Когда клетки достигали ~90% конфлюэнтности, культуральную среду удаляли путем аспирации и промывали колбу для культивирования один раз 1x DPBS.
(2) Добавляли 1 мл 0,25% трипсин-ЭДТА и инкубировали колбу в течение 10 минут при 37°С.
(3) Добавляли 9 мл полной среды и разделяли скопления клеток повторным пипетированием.
(4) Переносили суспензию клеток в коническую пробирку на 15 мл и центрифугировали при 800х g в течение 5 мин.
(5) Удаляли супернатант путем аспирации и повторно суспендировали осадок клеток с 10 мл полной среды.
(6) Определяли плотность жизнеспособных клеток с помощью гемоцитометра.
(7) Переносили аликвоты суспензии клеток по 3 мл в свежие колбы Т75.
(8) Инкубировали в течение 3 дней, пока клетки не достигали ~90% конфлюэнтности.
Замораживание клеток для создания банка клеток выполняли следующим образом.
(1) Пересаживали ~90% конфлюэнтные клетки из двух колб Т75 в восемь колб Т75.
(2) Когда клетки достигали поздней фазы роста (~2,0×107 клеток/флакон), удаляли культуральную среду и готовили суспензию отдельных клеток, обработав их 0,25% трипсин-ЭДТА.
(3) Повторно суспендировали осадок клеток при плотности 5×106 клеток/мл в среде для заморозки.
(4) Помещали аликвоту 1 мл суспензии клеток во флаконы с криогенным исходным раствором.
(5) Помещали пробирки в контейнер для замораживания, заполненный изопропанолом (обязательно закручивая крышку пробирки).
(6) Помещали контейнер для замораживания на ночь при -80°С.
(7) На следующий день немедленно переносили замороженные клетки в хранилище с паровой фазой жидкого азота.
Пример 3: Определение характеристик N2-42F и культуральной среды
Пример 3-1. Подтверждение времени удвоения N2-42F
Определяли времена расщепления клеток Neuro-2a и N2-42F, которые, как было подтверждено, способны обнаруживать укороченные формы белка SNAP25.
Клетки N2-42F и Neuro-2a высевали из расчета 1,5×105 клеток на лунку в 6-луночный культуральный планшет. Ежедневно в течение 6 дней подсчитывали общее количество жизнеспособных клеток с помощью гемоцитометра и окрашивания трипановым синим. Время удвоения рассчитывали с использованием количества жизнеспособных клеток, полученных во время фазы экспоненциального роста, как показано ниже в Таблице 4 и результаты показаны в Таблице 5 и на Фиг. 3. Т - время инкубации, Xb и Хе - количество клеток в начале и в конце времени инкубации в Таблице 4 соответственно.

Как показано в Таблице 5 и на Фиг. 3, время расщепления составляло 24±4,7 часов для Neuro-2a и 24±2,9 часов для N2-42F.
Пример 3-2. Подтверждение морфологии N2-42F
Allergan сообщил о выделении BoNT/А-чувствительного клона (H1) из клеток SiMa (PLoS One. 2012; 7 (11): е49516.). Таким образом, ожидается, что подобно Neuro-2a, клетки SiMa также представляют собой гетерогенную клеточную популяцию смешанных типов клеток, но их субклоны, такие как H1 и N2-42F, являются гомогенными типами клеток. Это подтвердили микроскопические исследования клеток Neuro-2a, N2-42F и SiMa. Как показано на Фиг. 4, и Neuro-2a, и SiMa демонстрируют гетерогенную морфологию под микроскопом, но клетки N2-42F выглядят очень гомогенными.
Из приведенных выше результатов можно видеть, что N2-42F соответствует гомогенному типу клеток в качестве единственного клона среди различных клонов, составляющих Neuro-2a.
Пример 3-3. Подтверждение условий культивирования N2-42F
В целях оптимизации условий интоксикации BoNT/A клетки N2-42F выращивали в культуральных планшетах, покрытых различными матрицами. В отличие от родительских клеток Neuro-2a, которые обычно размножаются в планшетах для культивирования без покрытия, клетки N2-42F явно предпочитают планшеты для культивирования, покрытые поли-D-лизином (PDL) (Фиг. 5). Ни пластины без покрытия, ни пластины, покрытые коллагеном IV типа или желатином, не поддерживали эффективный рост клеток N2-42F. На этих пластинах клетки N2-42F выглядели нездоровыми, образуя сгустки. Кроме того, они были неплотно прикреплены к пластине. Напротив, клетки N2-42F, выращенные на планшетах, покрытых PDL, были прочно прикреплены и равномерно распределены. Таким образом, если не указано иное, в настоящем исследовании культуральные планшеты, покрытые PDL, использовали исключительно для клеток N2-42F.
Пример 4: Подтверждение чувствительности N2-42F к ботулиновым нейротоксинам
Используя клетки SiMa в качестве контроля, тщательно исследовали чувствительность клеток N2-42F к BoNT/A в трех наборах культур 96-луночных планшетов, снабженных 1х средой для интоксикации, содержащей различные концентрации BoNT/А. После инкубации в течение 4 дней клетки обрабатывали 50 мкл 1х ДСН-буфера для образцов и аликвоты (12 мкл) подвергали анализу вестерн-блоттинг с использованием поликлональных IgG против SNAP25 (Sigma S9684), как описано в материалах и методах. Эндогенное расщепление SNAP25 в незначительных количествах было обнаружено в клетках SiMa и N2-42F, обработанных 0,93 пМ или более низкими концентрациями BoNT/A. Обработка 2,78~25 пМ BoNT/A приводила к 25~72% расщеплению SNAP25 в клетках N2-42F и 33~75% в клетках SiMa (Фиг. 6а). Этот результат показывает, что клетки N2-42F так же чувствительны, как и клетки SiMa.
Включая BoNT/A, существует семь серологически различных ботулиновых нейротоксинов от BoNT/A до BoNT/G. Подобно BoNT/A, BoNT/B также был лицензирован для фармацевтического применения в виде Myobloc® или Neurobloc®. Таким образом, было исследовано, можно ли использовать клетки N2-42F в любых анализах активности различных серотипов нейротоксинов ботулина с использованием клеток. С этой целью сравнивали чувствительность дифференцирующихся клеток N2-42F и Neuro-2a к различным комплексам нейротоксинов (Metabiologics C08RA188-RA194).
Как показано на Фиг. 6б, 25 пМ BoNT/A (M-BoNT/A) от Metabiologics приводили к насыщающей степени расщепления SNAP25 в N2-42F. В тех же условиях примерно 44% расщепления SNAP25 наблюдалось в клетках Neuro-2a. Более высокая чувствительность клеток N2-42F к M-BoNT/A согласуется с их чувствительностью к BoNT/A, полученному HUGEL (то есть Botulax). Клетки N2-42F также проявляли значительно более высокую чувствительность к M-BoNT/B. Интоксикация 2 нМ М-BoNT/B приводила к почти полному расщеплению Vamp2 в клетках N2-42F, но только к около 50% расщеплению в Neuro-2a. При этом клетки N2-42F в меньшей степени проявляли высокую чувствительность к M-BoNT/C и M-BoNT/F. Но с M-BoNT/D (5 пМ или 200 пМ) не было заметной разницы в чувствительности между клетками N2-42F и Neuro-2a (данные не показаны). Также ни одни из клеток не показали какой-либо уровень чувствительности к M-BoNT/E (10~400 пМ) или BoNT/G (50~2000 пМ). Наши результаты показывают, что клетки N2-42F могут использоваться в качестве хозяев в клеточном анализе активности для BoNT/A, BoNT/B, BoNT/C и BoNT/F.
Пример 5: Подтверждение стабильности N2-42F
Стабильность при пассировании может быть ключевой особенностью нейронных клеток, которые могут использоваться в качестве хозяина на любых платформах анализа с использованием клеток. Клетки N2-42F непрерывно размножались в течение нескольких пассажей. Клетки в ранних пассажах использовали для создания основного банка клеток, как описано в разделе «Материалы и методы» и через каждые 5 пассажей клетки также хранили в резервуарах с жидким азотом для стабильности пассирования. Вкратце, клетки N2-42F, хранящиеся на пассаже 5 (Р5) и 15 (Р15), были восстановлены из резервуара с жидким азотом и при ~90% конфлюэнтности, их сравнивали по чувствительности к BoNT/A с использованием среды для интоксикации 1х, содержащей 0,1 нМ BoNT/A в соответствии с процедурой, описанной на Фиг. 6. Эксперимент проводили в трех повторах с использованием клеток SiMa в качестве контроля. Как показано на Фиг. 7, 63~68% расщепления SNAP25 было обнаружено в клетках N2-42 на Р5 и 63~71% на Р15. Клетки SiMa показали 70-77% расщепления.
Этот результат указывает на то, что подобно клону H1 клеток SiMa, идентифицированному Allergan, чувствительность к BoNT/A является стабильно наследуемым свойством клеток N2-42F, что делает их подходящим хозяином, вторым в своем роде после клона H1 клеток SiMa, в платформе для анализа с использованием клеток.
Пример 6: Определение активности BoNT/А на основе N2-42F
Пример 6-1. Экспериментальный способ
96-луночный планшет покрывали методом, описанным в разделе «Материалы и методы». Пока сушили на воздухе 96-луночный культуральный планшет, готовили в общей сложности 7 мл суспензии клеток N2-42F с плотностью 5,5×105 клеток/мл. Примерно 90% конфлюэнтных клеток N2-42F в одной колбе Т75 будет достаточно для трех 96-луночных культуральных планшетов. Переносили суспензию клеток в стерильный буферный резервуар, распределяли аликвоты (100 мкл) клеточной суспензии в каждую лунку, используя многоканальную пипетку, и инкубировали 96-луночный культуральный планшет в инкубаторе с СО2. Наружные лунки 96-луночного планшета заполняли аликвотами (100 мкл) раствора 1x АА, чтобы избежать образование сильных краевых эффектов.
На следующий день после распределения клеток всю среду для культивирования клеток удаляли с 96-луночного планшета с помощью многоканальной пипетки и добавляли 100 мкл RPMI 1640 для промывания. Затем интоксикационной средой BoNT/A обрабатывали каждый 96-луночный планшет и инкубировали в течение 4 дней при 37°С, 5% CO2. В дополнение, готовили связывающее антитело, В4 IgG, в количестве 7 мл, которое затем разделяли по 50 мкл в планшетах для ELISA и хранили при 4°С в течение ночи.
Для измерения среду интоксикации BoNT/А удаляли с 96-луночного планшета с помощью многоканальной пипетки и каждую лунку обрабатывали 60 мкл лизирующего буфера (рН 7,5, 20 мМ HEPES, 1% Тритон-200 мМ NaCl, 1 мМ ЭГТА, и 5 мМ ЭДТА, с добавлением ингибитора протеолиза непосредственно перед использованием), и инкубировали при 4°С в течение 20 минут при встряхивании со скоростью 500 об/мин. После этого отбирали лизирующего буфер, содержащийся в каждой лунке, и центрифугировали при 4000 об/мин в течение 20 минут при 4°С.
Планшет для ELISA промывали 3 раза промывочным буфером, добавляли аликвоты по 300 мкл блокирующего буфера в каждую лунку, инкубировали в течение 15 мин при комнатной температуре и дважды промывали промывочным буфером после удаления блокирующего буфера.
Аликвоты по 50 мкл ОКЛ переносили из 96-луночного культурального планшета в планшет для ELISA, покрытого иммобилизованным антителом (В4), и планшет для ELISA инкубировали в течение 4 часов при 4°С на шейкере для микропланшетов при 200 об/мин. Наконец, планшет 3 раза промывали промывочным буфером.
Для обнаружения SNAP25 и SNAP25197 детектирующие антитела добавляли в планшет для ELISA (50 мкл на лунку) и планшет инкубировали в течение 1 часа при комнатной температуре на инкубаторе Thermo shaker (200 об/мин). Затем планшет трижды промывали промывочным буфером, в планшет добавляли аликвоты по 50 мкл 1-StepTMUltra TMB-ELISA, реакцию пероксидазы хрена останавливали добавлением 2 М серной кислоты (50 мкл/лунку) через 5 мин. Реакцию пероксидазы хрена измеряли при 450 нм. Значение А450 может указывать на относительное количество SNAP25197.
Пример 6-2. Построение стандартной кривой и определение активности BoNT/A
Стандартную кривую строили и активность BoNT/A тестировали следующим образом. Рассчитывали среднее значение А450 для контрольных лунок, в которых сэндвич-ELISA проводили без обработки BoNT/A (т.е. 0 пМ). Вычитали среднее контрольное значение А450 из значений теста А450 и вычисляли нормированное среднее значение теста А450. Затем откладывали нормализованные средние значения А450 по оси Y в зависимости от активности BoNT/A по оси X, используя Prism 5.0 (GraphPad Software, La Jolla, CA). Анализировали график, последовательно выбрав «анализ», «нелинейная регрессия (аппроксимация кривой)» и «сигмоидальная доза-реакция», что дало значение ЕС50. Строили стандартную кривую, используя нормализованные значения А450 для тестовых лунок, обработанных 0,1~0,93 пМ BoNT/A в виде стандартного контроля (см. Приложение 2, Раздел Г). Использовали уравнение стандартной кривой со значением R2 0,95 или выше для определения активности BoNT/А в тестируемых образцах.
Пример 7: Получение моноклональных антител, специфичных к SNAP25
Allergan использовал пептид N-CDSNKTRIDEANQ-C из 13 аминокислотных остатков (АА) для повышения уровня антител. Указанный пептид был разработан таким образом, чтобы он был идентичен С-концу SNAP25197, полученному при расщеплении посредством BoNT/A. Поскольку SNAP25FL также имеет идентичную аминокислотную последовательность, было бы разумно предположить, что специфичность моноклональных антител, распознающих SNAP25197, связана с еще не идентифицированной особенностью SNAP25197, а не с его первичной последовательностью аминокислот. SNAP25 состоит из 206 аминокислотных остатков (Фиг. 8). С помощью мотивов SNARE он образует стабильный тройной комплекс с синтаксином 1А и синаптобревином 2 (VAMP2). Менее стабильный и нефункциональный тройной комплекс образуется с SNAP25197, тогда как тройной комплекс вообще не может образоваться с SNAP180 (Peer J. 2015 Jun 30; 3: e1065). Их открытие о том, что 9 С-концевых аминокислот SNAP25 важны для функции in vivo и образования стабильного тройного комплекса, предполагает, что расщепление в позиции 197-й аминокислоты с помощью BoNT/A вызывает еще не идентифицированные структурные изменения, в частности на С-конце и во втором домене SNARE. Таким образом, постулируемое структурное различие между SNAP25FL и SNAP25197 позволяет получить моноклональные антитела, специфичные к SNAP25197.
Всего было создано 10 пептидных антигенов, удовлетворяющих следующим критериям (Таблица 6). Во-первых, были исключены альфа-спиральные области, проявляющие относительно более низкую антигенность. Во-вторых, пептидные последовательности неизбыточны и уникальны, их свойства важны для снижения перекрестной реактивности антител.

Из всех 10 пептидов, описанных в Таблице 6, пептиды М и N использовали для получения моноклональных антител, способных обнаруживать как SNAP25FL, так и SNAP25197. С использованием пептидов С ожидалось получить три различных моноклональных антитела со специфичностью связывания в отношении (1) SNAP25FL, (2) SNAP25197 и (3) обоих SNAP25FL и SNAP25197. На протяжении всего нашего исследования два коммерчески доступных антитела использовали в качестве контроля в анализах ELISA и вестерн-блоттинга. Они включают поликлональные IgG кролика против SNAP25 (Sigma-Aldrich) и МС6050 (R&D). Первое распознает как SNAP25FL, так и SNAP25197, а второе является моноклональным антителом, специфичным к SNAP25197.
Индивидуальный синтетический пептид вводили двум кроликам для получения поликлональной сыворотки и 4 мышам для получения моноклональных антител, как описано в разделе «Материалы и методы». В частности, процедура получения гибридомных клеток, продуцирующих моноклональные антитела, суммирована на Фиг. 9. Вкратце, четыре мыши были иммунизированы пептидным антигеном для индукции продукции антител. После ELISA-скрининга сыворотки мышей на продукцию антител спленоциты выделяли из ELISA-положительных мышей и сливали с клетками миеломы SP2. Впоследствии клетки гибридомы высевали при плотности единичных клеток в 96-луночный планшет на пептидный антиген. Как показано на Фиг. 9, первоначальный скрининг для получения положительных клонов выполняли с использованием прямого ELISA с использованием пептидного антигена, за которым следовал более строгий скрининг с использованием рекомбинантного SNAP25 и общего клеточного лизата.
Типичный результат начального скрининга гибридомы показан на Фиг. 10. Супернатанты культур собирали из клеток гибридомы (клон 4, В4), выращенных в 7х 96-луночном планшете, и тестировали на их активность к пептидному антигену прямым ELISA. Около 25% клонов дали положительные сигналы ELISA (Фиг. 10а, и Фиг. 10б), и 24 относительно сильных положительных клонов отбирали и тестировали на их активность к эндогенному SNAP25 с помощью сэндвич-ELISA с использованием общего клеточного лизата (Фиг. 10в). Были отобраны десять клонов для нескольких раундов отбора клонов из одной клетки.
Для клональной селекции из одной клетки клоны гибридомы высевали с одноклеточной плотностью в один 96-луночный планшет. Четыре дня спустя культуральные супернатанты тестировали на активность по отношению к рекомбинантному SNAP25 прямым ELISA, и для дальнейшего анализа отбирали пять клонов, дающих относительно сильные сигналы ELISA. Для примера, в случае клона 4, более 80% субклонов, выращенных в 96-луночном планшете, дали положительный результат в прямом ELISA (Фиг. 11а). Двенадцать субклонов давали сигналы ELISA ниже, чем отрицательный контроль, включая А8-А10, В11, С10, D4, D8, D9, Е5, E11, F10 и G7 (Фиг. 11а). Пять субклонов также анализировали с помощью сэндвич-ELISA (Фиг. 11б) и вестерн-блоттинга (Фиг. 11в) с использованием общего клеточного лизата (ОКЛ), как описано в материалах и методах. Следует отметить, что сигналы ELISA с субклонами клона 4 хорошо согласуются с сигналами вестерн-блоттинга. Поскольку ОКЛ претерпел денатурацию во время вестерн-блоттинга, этот результат предполагает, что моноклональные антитела, продуцируемые гибридомным клоном 4, одинаково реагируют как с неденатурированным (т.е. ELISA), так и с денатурированным SNAP25.
Одноклеточный клональный отбор повторяли еще два раунда, как показано на Фиг. 12 и 13. После второго раунда отбора два клона 6 и 8 были исключены из дальнейшего скрининга, поскольку все культуральные супернатанты в 96-луночном планшете не дали сигналов ELISA, значительно более высоких, чем отрицательный контроль. Посредством серии одноклеточной клональной селекции было получено всего 12 клонов гибридомы, которые продуцируют моноклональные антитела, специфичные для SNAP25FL, SNAP25197 или обоих.
Пример 8: Получение и очистка моноклональных антител
Клетки гибридомы размножали в колбах Т175 и использовали для создания банка основных клеток. Для продукции моноклональных антител клетки гибридомы выделяли из флакона с исходным раствором, хранящимся в паровой фазе жидкого азота. Когда клетки достигли примерно 90% конфлюэнтности, их субкультивировали в соотношении 1:4~10 в нескольких колбах Т175. После инкубации в течение 3-4 дней клетки собирали центрифугированием и ресуспендировали в бессывороточной среде при концентрации 1,0×106 клеток/мл. После инкубации в течение 4 дней супернатанты культур собирали и использовали для очистки IgG с использованием колонки с протеином G (колонка HiTrap® Protein G HP), как описано в разделе «Материалы и методы». Выход моноклональных антител, очищенных с использованием культуральных супернатантов, варьировался от партии к партии культуры гибридомы, а также для индивидуального клона гибридомы. Более десятка раз очистки с различными партиями культурального супернатанта (165~542 мл) дали примерно 2,45~5,52 мг С16 IgG (Таблица 7).
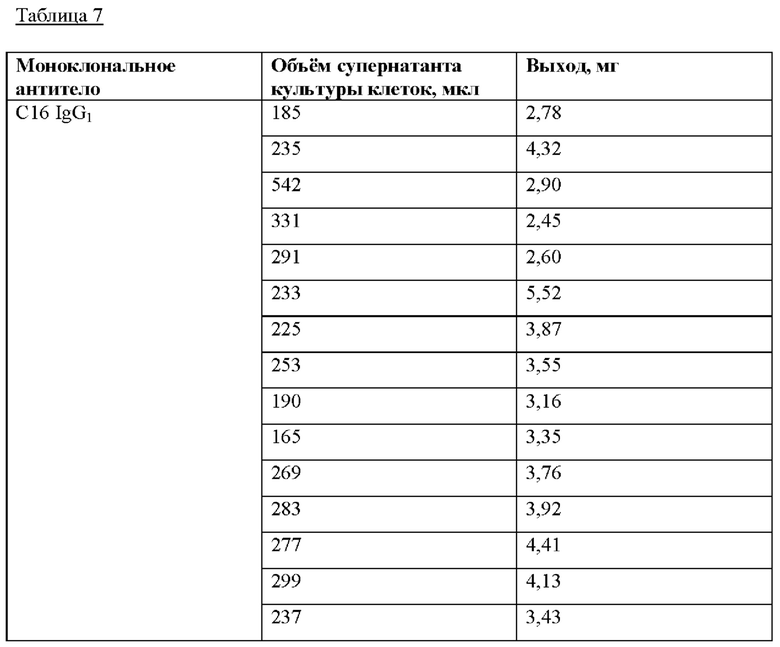
Результаты очистки других моноклональных антител представлена в Таблице 8.
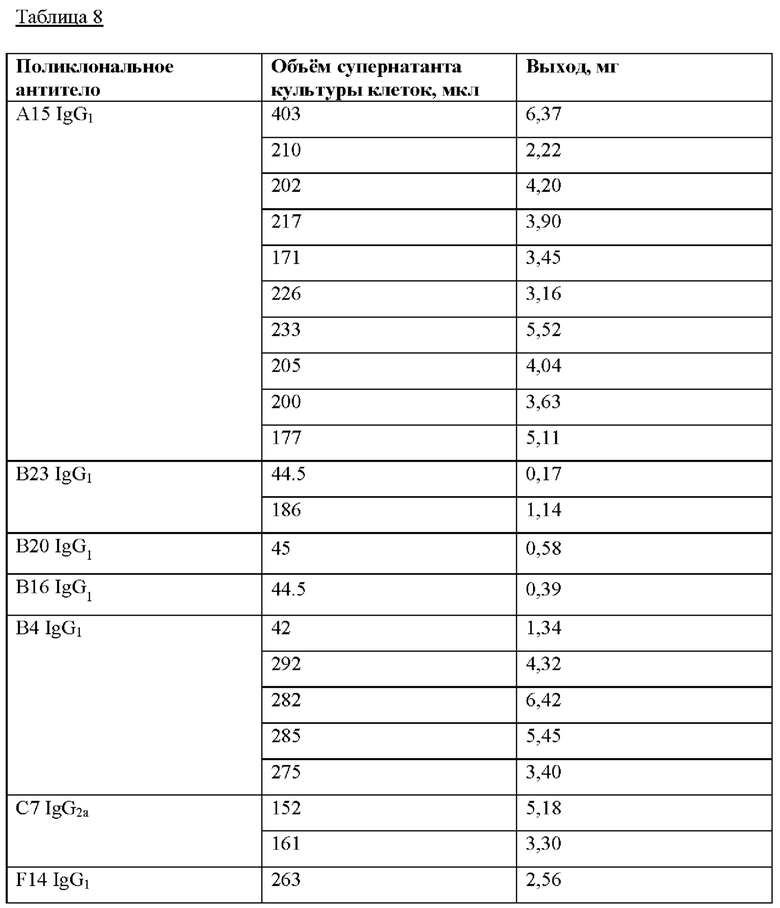
В среднем примерно 1-2 мг IgG было получено из 100 мл супернатанта культуры. Хотя данные не представлены, чистота IgG и их специфичность связывания антигена были подтверждены с помощью ДСН-ПААГ и ELISA.
Пример 9: Очистка поликлональных антител
Поликлональные антитела очищали из сыворотки кролика с использованием SNAP25-AffiGel, как описано в разделе «Материалы и методы». Вкратце, сыворотку кролика разводили в 10 раз 10 мМ Трис-HCl, рН 7,5, и очищали центрифугированием при 10,000х g в течение 10 мин и фильтровали через фильтр 0,45 мкм. Прозрачный разбавитель сыворотки повторно загружали на SNAP25-AffiGel три раза, и связанные белки последовательно элюировали 0,1 М ацетатом натрия, рН 5,5, 0,1 М глицином, рН 4,0, 0,1 М глицином, рН 2,5 и 0,1 М триэтиламином, рН 11.5. Белковые фракции собирали в пробирки, содержащие 0,1 мл 1 М Трис-HCl, рН 8,0, объединяли и концентрировали до 1 мл с помощью AMICON Ultra-15. После диализа с четырьмя заменами 1x PBS и 10% глицерина каждые 90 мин их анализировали с помощью ДСН-ПААГ и ELISA.
В Таблице 9 суммированы результаты относительного распределения белков сыворотки во фракциях SNAP25-AffiGel, полученных из 4 различных партий сывороток rA15.
Несмотря на то, что каждый образец сыворотки демонстрировал разные паттерны распределения белка во фракциях SNAP25-AffiGel (Таблица 9), IgG был обнаружен как основной компонент всех фракций, исследованных с помощью 10% ДСН-ПААГ (Фиг. 14а). Однако при тестировании с помощью сэндвич-ELISA активность в отношении эндогенного SNAP25 была обнаружена только для фракции с рН 4,0. Высокое отношение сигнала к фону (>25) (Фиг. 14б) указывает на то, что IgG во фракции с рН 4,0 является высокоспецифичным для эндогенного SNAP25. Поскольку аналогичные результаты были получены с другими сыворотками кроликов, использовали исключительно фракцию рН 4,0 на протяжении всего нашего исследования с использованием поликлональной сыворотки, если не указано иное.
Пример 10: Анализ последовательности моноклональных антител
Тотальную РНК, экстрагированную из клеток гибридомы, подвергали обратной транскрипции в кДНК с использованием либо антисмыслового праймера oligo-dT, либо антисмыслового праймера, специфичного для генов (мышиный IgG1 СН и каппа CL). Для амплификации кДНК с помощью ПЦР для определения изотипа антитела использовали специфические праймеры константных доменов мыши. Вырожденные VH и VL праймеры использовали для амплификации вариабельных доменов из кДНК. Для 5' RACE к 3' концу кДНК был добавлен гомополимерный [dC] хвост. Затем тяжелую и легкую цепи амплифицировали с помощью олиго [dG] смыслового праймера и ген-специфического (СН/KC) антисмыслового праймера. Продукты ПЦР клонировали в вектор с тупыми концами или в вектор ТА для секвенирования. Результаты секвенирования сопоставляли с цепями VH и VL для определения консенсусных последовательностей.
Последовательности CDR объединены в три разные группы IgG в соответствии с их антигенной специфичностью: (1) SNAP25FL-специфические IgG (Таблица 10), (2) SNAP25197-специфические IgG (Таблица 11), и (3) Би-специфические IgG, взаимодействующие как с SNAP25FL, так и с SNAP25197 (Таблица 12).
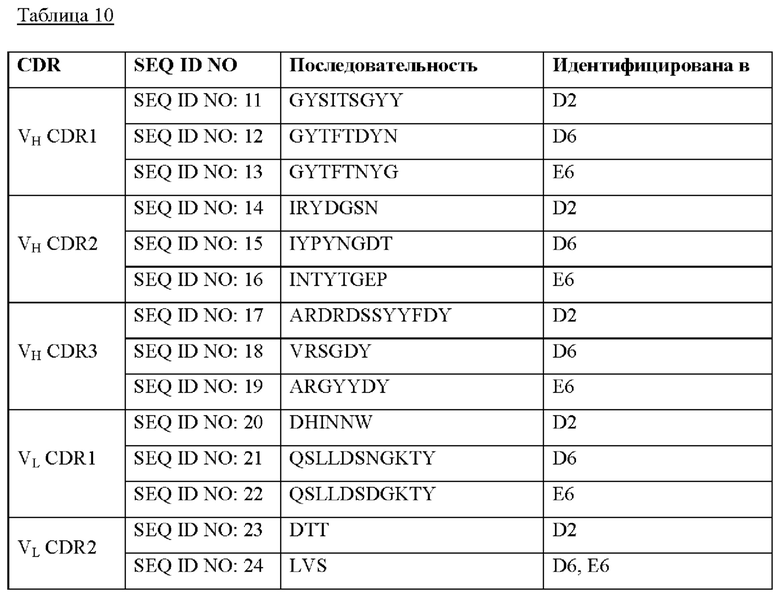



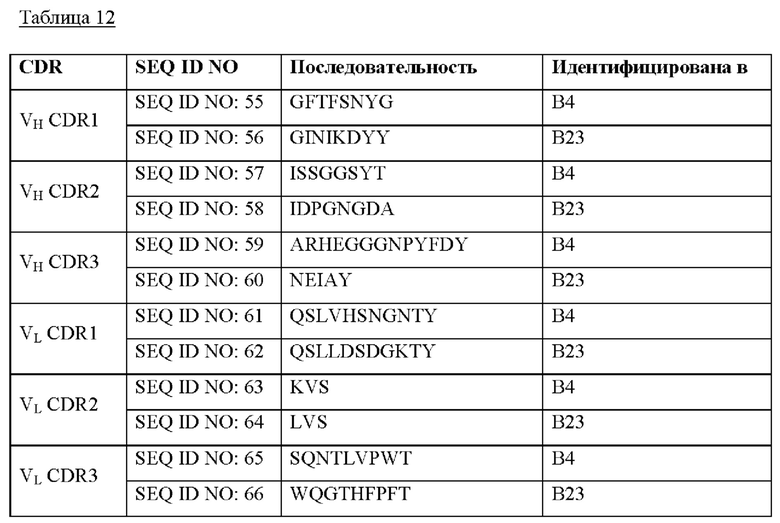
Последовательности доменов VH и VL антител, продуцируемых в соответствии с настоящим изобретением, суммированы в Таблица 13.

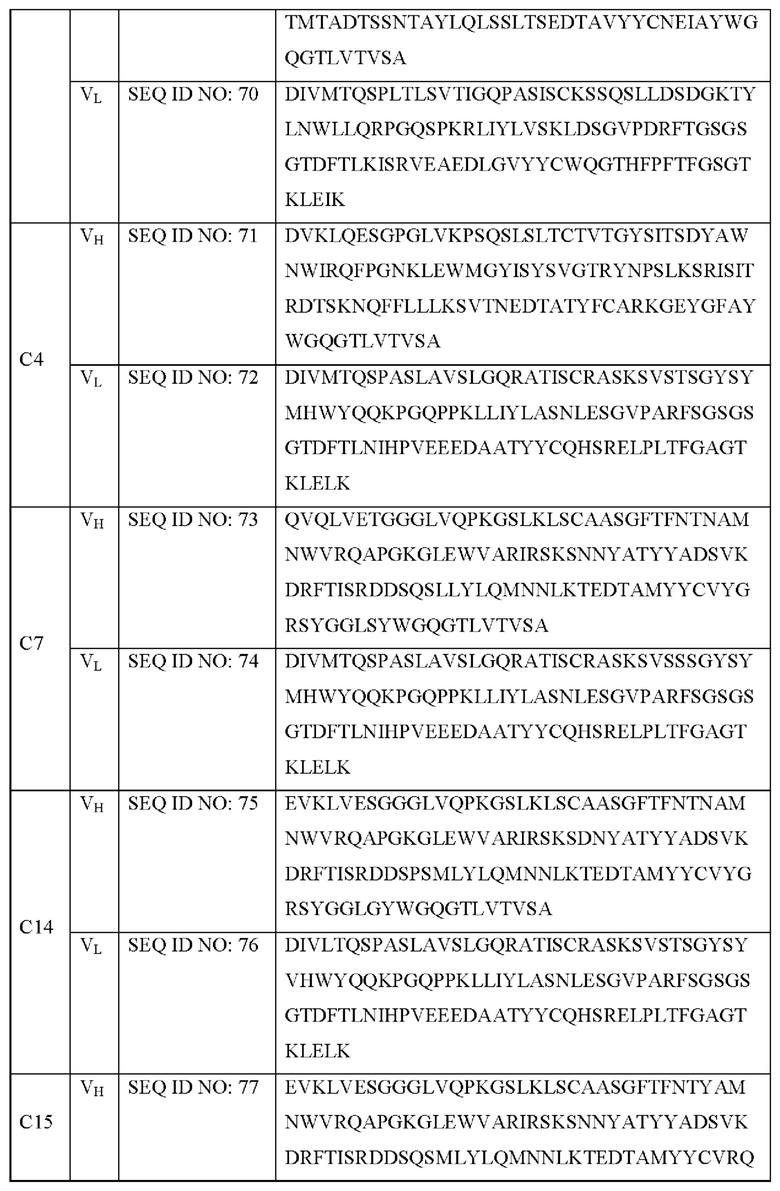
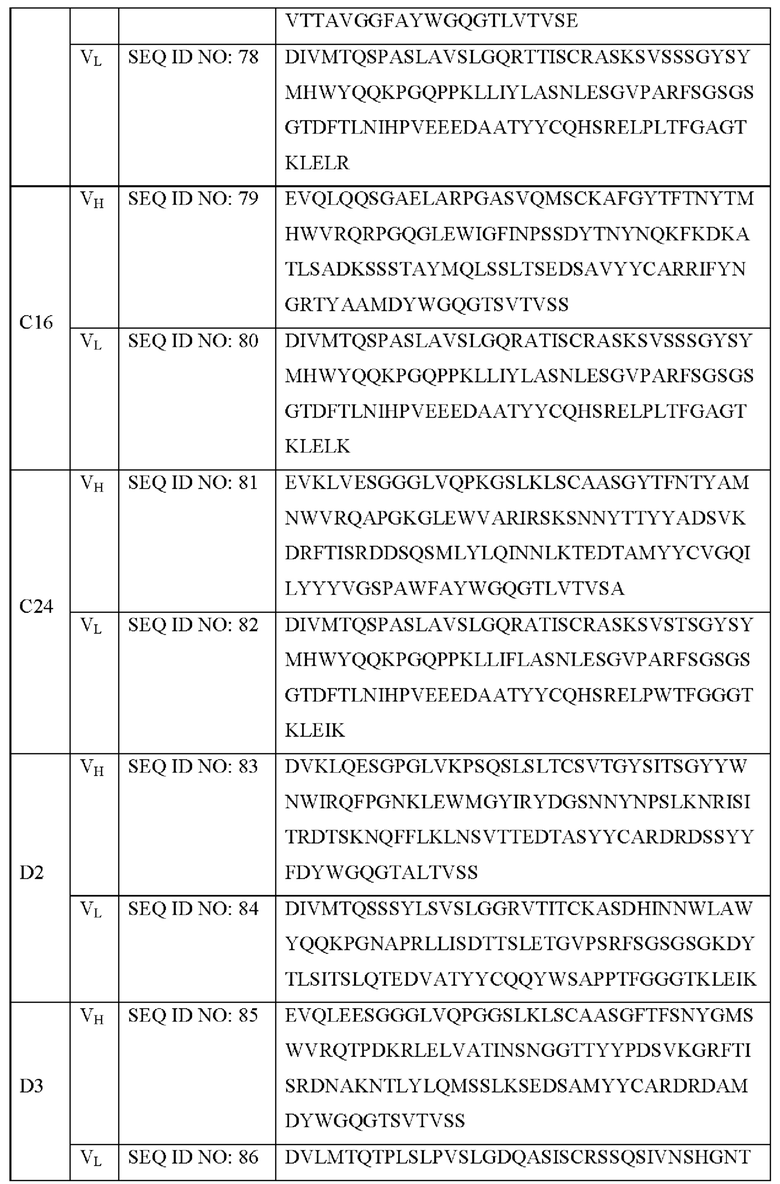
Интересно отметить, что последовательность VL CDR3 является общей для SNP25197-специфичных IgG, таких как С4, С7, С14, С15 и С16 IgG, тогда как их последовательности VH имеют тенденцию быть IgG-специфичными (Таблица 11). Анализ выравнивания последовательностей показал, что ни одна последовательность CDR IgG, перечисленных в Таблицах 10-12, не перекрывается с последовательностями CDR VL и VH ранее описанных IgG (патент США US 8,198,034 B2). Это может быть приписано использованию более строгой стратегии скрининга положительных клеток гибридомы в настоящем исследовании. То есть на протяжении нескольких раундов скрининга только тройные положительные клоны гибридомы были отобраны прямым ELISA с использованием пептидного антигена, сэндвич-ELISA с ОКЛ и вестерн-блоттингом с ОКЛ. Эта строгая стратегия скрининга должна была способствовать значительному снижению значений KD IgG, полученных в настоящем исследовании (см. ниже).
Пример 11: Кинетический анализ моноклональных антител
Как подробно описано в разделе «Материалы и методы», кинетический анализ моноклональных антител проводили исключительно методом BLI с использованием прибора  Octet Red96 в соответствии с процедурой, рекомендованной производителем. Во-первых, был проведен кинетический анализ с использованием IgG, специфичных к SNAP25197, с использованием биосенсоров против GST, нагруженных рекомбинантный GST-SNAP25197 (125 нМ или 250 нМ). На Фиг. 15 показан набор кинетических кривых, полученных с С4, С7, С16 и С24, в качестве примера таких исследований. Вкратце, биосенсоры анти-GST, загруженные GST-SNAP25197, последовательно погружали в 1х буфер для измерения кинетики, серийно разведенные образцы IgG (7,8, 15,6, 31,2, 62,5, 125, 250, 500 нМ) (ассоциация аналитов) и 1х буфер для измерения кинетики (диссоциация аналита). После ассоциации и диссоциации аналита в течение 10 мин каждой из процедур были получены исходные данные кинетических кривых (Фиг. 15а). Затем кинетические кривые выравнивали путем вычитания базового сигнала BLI (Фиг. 15б), из которого с использованием программного обеспечения для анализа Octet получали KD (Таблица 14).
Octet Red96 в соответствии с процедурой, рекомендованной производителем. Во-первых, был проведен кинетический анализ с использованием IgG, специфичных к SNAP25197, с использованием биосенсоров против GST, нагруженных рекомбинантный GST-SNAP25197 (125 нМ или 250 нМ). На Фиг. 15 показан набор кинетических кривых, полученных с С4, С7, С16 и С24, в качестве примера таких исследований. Вкратце, биосенсоры анти-GST, загруженные GST-SNAP25197, последовательно погружали в 1х буфер для измерения кинетики, серийно разведенные образцы IgG (7,8, 15,6, 31,2, 62,5, 125, 250, 500 нМ) (ассоциация аналитов) и 1х буфер для измерения кинетики (диссоциация аналита). После ассоциации и диссоциации аналита в течение 10 мин каждой из процедур были получены исходные данные кинетических кривых (Фиг. 15а). Затем кинетические кривые выравнивали путем вычитания базового сигнала BLI (Фиг. 15б), из которого с использованием программного обеспечения для анализа Octet получали KD (Таблица 14).
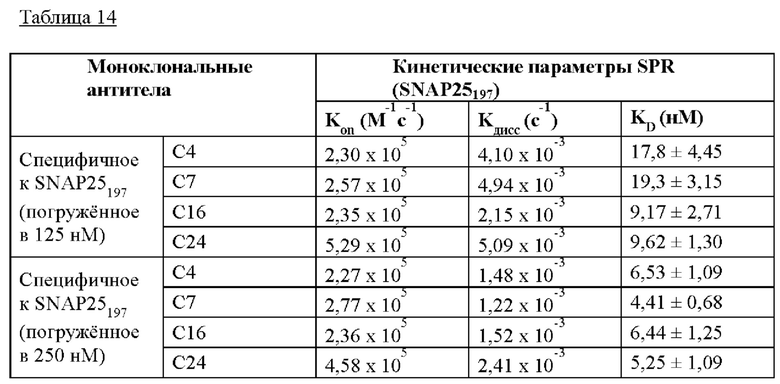
Учитывая диапазон значений KD в нМ, кинетический анализ, как казалось, был проведен нормально. Однако, когда кинетический анализ проводился с биосенсором, погруженным в более низкую концентрацию GST-SNAP25197 (125 нМ), значения KD пропорционально уменьшались, в основном из-за изменений константы скорости диссоциации Kдисс (Таблица 14). Также было странно, что значения Kдисс (1,22~5,09×10-3) были на 1 или 2 порядка ниже, чем ранее сообщалось (3,11×10-4~6,74×10-5) для моноклональных антител с аналогичной антигенной специфичностью (Патент США US 8,198,034 B2).
Можно рассматривать две вероятные причины относительно высоких значений KD и Kдисс. Первый - это использование неподходящего буфера для анализа, а второй - неотъемлемое ограничение всего кинетического анализа, проводимого с биосенсором к GST (примечание по применению 14  анализ кинетики биомолекулярного связывания на платформе Octet). Сравнительное использование различных буферов для анализа в кинетическом анализе показало, что 1х буфер для измерения кинетики был наиболее подходящим среди всех протестированных буферов (данные не показаны). Поэтому, в качестве альтернативного кинетического анализа биосенсоры захвата Fc мышиных IgG (АМС) были непосредственно загружены антителом и подвергнуты ассоциации/диссоциации с серийно разведенным рекомбинантный GST-SNAP25. Вкратце, очищенный IgG разбавляли до 100 или 200 нМ, тогда как GST-SNAP25FL или SNAP25197 серийно разбавляли до 1,56, 3,125, 6,25, 12,5, 25, 50 и 100 нМ. После уравновешивания в 1х буфере для измерения кинетики в течение 1 минуты биосенсоры АМС загружали IgG на 10 минут с последующим погружением в 1х буфер для измерения кинетики на 10 минут. Ассоциацию и диссоциацию аналита проводили по 10 мин для каждой операции, KD оценивали, как описано выше.
анализ кинетики биомолекулярного связывания на платформе Octet). Сравнительное использование различных буферов для анализа в кинетическом анализе показало, что 1х буфер для измерения кинетики был наиболее подходящим среди всех протестированных буферов (данные не показаны). Поэтому, в качестве альтернативного кинетического анализа биосенсоры захвата Fc мышиных IgG (АМС) были непосредственно загружены антителом и подвергнуты ассоциации/диссоциации с серийно разведенным рекомбинантный GST-SNAP25. Вкратце, очищенный IgG разбавляли до 100 или 200 нМ, тогда как GST-SNAP25FL или SNAP25197 серийно разбавляли до 1,56, 3,125, 6,25, 12,5, 25, 50 и 100 нМ. После уравновешивания в 1х буфере для измерения кинетики в течение 1 минуты биосенсоры АМС загружали IgG на 10 минут с последующим погружением в 1х буфер для измерения кинетики на 10 минут. Ассоциацию и диссоциацию аналита проводили по 10 мин для каждой операции, KD оценивали, как описано выше.
Расчетные значения KD с использованием программного обеспечения для анализа Octet приведены в Таблице 15.
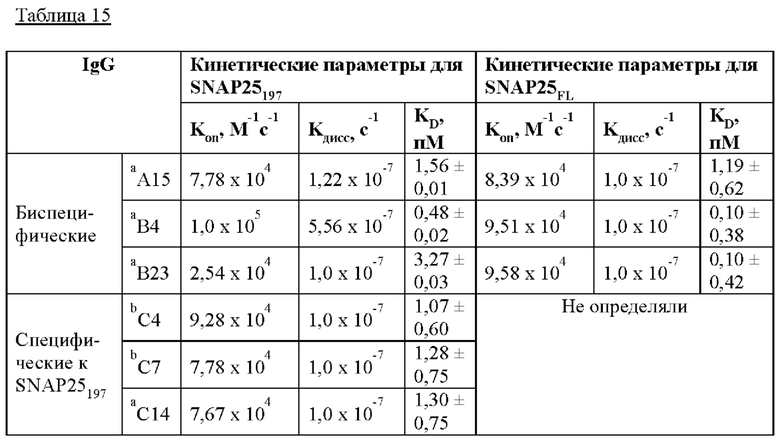

Наиболее заметными являются значения Kдисс (т.е. 2,57×10-6~1,0×10-7) (Таблица 15), что на два-три порядка ниже, чем полученные с биосенсором к GST (Таблица 14). Фактически, скорости диссоциации были слишком низкими, чтобы их можно было точно измерить с помощью Octet, поэтому нижний предел значения Kдисс, 1,0×10-7, был предварительно задан для шести IgG, как описано в Таблице 15.
Оценка значений KD с этими ориентировочными константами скорости диссоциации дала 0,48~3,27 пМ. Следует отметить, что биосенсоры АМС, нагруженные SNAP25197-специфическими IgG, не показали статистически значимой степени ассоциации GST-SNAP25FL до 1 мкМ. Точно так же GST-SNAP25197 не ассоциировался с биосенсорами АМС, загруженными SNAP25FL-специфическими IgG. Эти специфичности связывания IgG хорошо согласуются с их активностью в ELISA.
Пример 12: Антигенсвязывающая специфичность моноклональных антител
Моноклональные антитела исследовали в сравнении на их активность по отношению к SNAP25FL и SNAP25197 с использованием прямого ELISA и вестерн-блоттинга. Сначала проводили прямой ELISA с очищенным IgG (50 нг на лунку) в микропланшете, покрытом GST-SNAP25FL или GST-SNAP25197, как описано в разделе «Материалы и методы». Реакцию пероксидазы хрена проводили в течение 5 мин, и степень активности пероксидазы хрена определяли путем измерения А450 с помощью Bio-Tek SynergyNeo2. Значения А450 получали после вычитания фоновой А450, измеренной без IgG, рассчитывали отношение А450 для SNAP25197 к А450 для SNAP25FL и представляли как отношение 197/206 в Таблице 16. В соответствии с результатом, полученным с помощью анализа BLI, значения отношения 197/206 для биспецифических IgG, таких как А15, В4 и В23, были близки к 1,0. Значения отношения 197/206 для SNAP25197-специфических IgG, таких как С7, С14, С16 и С24, были более 950, а значения отношения 197/206 для SNAP25FL-специфических IgG давали 0,02~0,03.
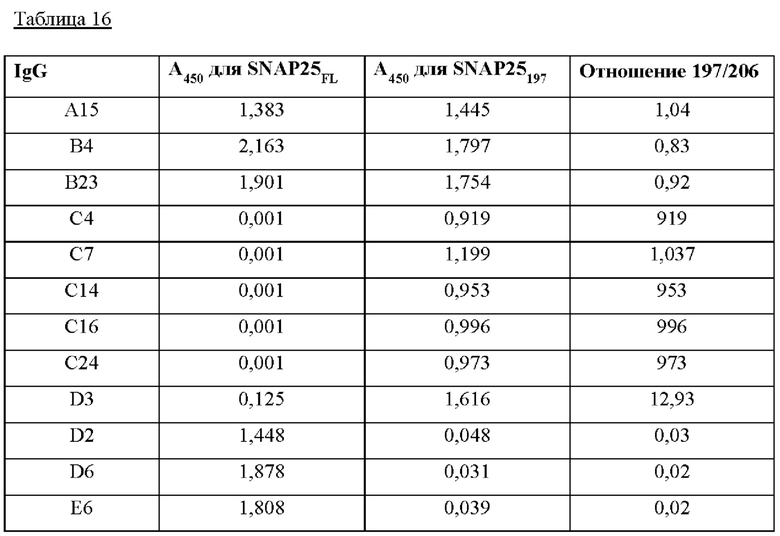
Вестерн-блоттинг проводили с супернатантами культур гибридомы (разведение 1:100) в качестве источника первичных антител. Аликвоты (0,5 мкг) GST-SNAP25FL и GST-SNAP25197, разделенные на денатурирующем геле и впоследствии перенесенные на мембрану ПВДФ, тестировали как антиген. Моноклональные антитела реагировали с денатурированным антигеном SNAP25 с той же специфичностью, что и в анализе ELISA (Фиг. 17). Эти результаты не являются неожиданными, поскольку только дважды положительные клетки гибридомы были отобраны с помощью строгого анализа ELISA и вестерн-блоттинга.
Пример 13: Конъюгирование пероксидазы хрена (HRP) с антителами
Среди протестированных моноклональных антител, С16 IgG продемонстрировал наиболее воспроизводимое сохранение антигенной аффинности и специфичности после конъюгирования с пероксидазой хрена (данные не показаны). В типичной реакции конъюгации HRP С16 IgG (5 мг) инкубировали с активированной пероксидазой хрена (5 мг) в течение 2 часов при комнатной температуре с защитой от света, как описано в разделе «Материалы и методы». После добавления 0,1 мл боргидрида натрия (4 мг/мл) конъюгат HRP-антитело подвергали диализу против 1x PBS и 1xPBS/50% глицерина при 4°С. Как показано на Фиг. 18а, конъюгация активированной пероксидазы хрена с С16 IgG была очень эффективной, так что образование конъюгата С16 IgG-HRP было обнаружено даже без инкубации (сравните дорожки 1-3). Неполная конъюгация С16 IgG, наблюдаемая в виде свободных IgG и HRP (дорожки 4-6), предполагает необходимость относительно высоких концентраций свободных пероксидазы хрена и IgG в реакционной смеси для эффективной конъюгации.
Два разных конъюгата С16 IgG-HRP исследовали на активность по отношению к SNAP25197 в прямом ELISA. 96-луночный микропланшет покрывали 0-200 мкг GST-SNAP25197, имитируя примерно до ~0,8% расщепление эндогенного SNAP25 в клетках, выращенных в лунке микропланшета. Аликвоты (200 нг) конъюгатов С16 IgG-HRP были способны детектировать всего 50 пг GST-SNAP25197 при измерении в течение 30 минут с помощью 50 мкл 1-StepTM Ultra TMB-ELISA (Фиг. 18б). Кроме того, более высокие значения А450 были получены пропорционально увеличению количества GST-SNAP25197. На основании этих результатов конъюгаты C16-HRP использовали исключительно в качестве детектирующих антител в оптимизированном сэндвич-ELISA.
Пример 14: Конъюгация биотина с антителами
Сэндвич-ELISA, разработанный Allergan, использует два антитела: SNAP25197-специфический IgG в качестве захватывающего антитела и поликлональный SNAP25-специфический IgG в качестве детектирующего антитела. Напротив, сэндвич-ELISA, разработанный в настоящем исследовании, использует два моноклональных антитела в качестве связывающих каждое из детектирующих антител, что делает контроль качества более осуществимым и простым. Этот новый сэндвич-ELISA демонстрирует высокий уровень повторяемости, воспроизводимости и точности. Тем не менее, добавление второго детектирующего антитела, такого как IgG, конъюгированного с щелочной фосфатазой или биотином, могло бы дополнительно повысить точность сэндвич-ELISA за счет нормализации SNAP25, связавшегося в каждой лунке.
В качестве первой попытки биспецифическое моноклональное антитело А15 было конъюгировано с биотином, как описано в разделе «Материалы и методы». Как показано на Фиг. 19, чем выше была концентрация биотина, тем больше молекул биотина было конъюгировано на один IgG. Количественное определение конъюгации биотина с использованием HABA/Avidin Premix (Thermo Scientific) показало, что примерно 8 моль биотина конъюгировались на моль IgG в реакции с 0,25 мМ биотина. Соответственно, тяжелая цепь IgG, конъюгированная с 0,5 мМ биотином, демонстрировала заметно более медленную миграцию на ДСН-ПААГ (Фиг. 19а и 19б).
Активность IgG А15, конъюгированного с 0,1 мМ биотином, исследовали с помощью сэндвич-ELISA. Вкратце, ОКЛ (60 мкл) получали из клеток N2-42F, обработанных указанной концентрацией BoNT/А. Микропланшет, покрытый В4 IgG, инкубировали с 50 мкл аликвотами ОКЛ, и эндогенный SNAP25197, связавшийся с IgG В4, выявляли путем инкубации с конъюгатом А15-биотин и конъюгатом стрептавидин-АР (разведение 1:500 в 1x TBS). А15 IgG был охарактеризован в качестве биспецифического моноклонального антитела, связывающего как SNAP25FL, так и SNAP25197 со сравнимой аффинностью и специфичностью (см. Фиг. 17). Эти свойства входят в число критериев выбора IgG А15 в качестве потенциально второго антитела для обнаружения с целью нормализации. Таким образом, ожидалось, что связывание А15 с SNAP25 останется относительно независимым от степени расщепления SNAP25. Однако, напротив, активность АР, отражающая связывание биотинилированного IgG А15 с SNAP25, увеличивалась пропорционально концентрации BoNT/А (Фиг. 19в). Этот результат можно объяснить изменением его антигенной специфичности при биотинилировании как легкой, так и тяжелой цепей IgG (Фиг. 19б).
Пример 15: Прямое перекрестное связывание щелочной фосфатазы (ЩФ) с антителами.
В качестве альтернативного подхода ко второму детектирующему антителу очищенные IgG напрямую перекрестно сшивали с ЩФ с использованием глутаральдегида, поскольку степень перекрестного сшивания можно регулировать концентрацией глутаральдегида. Два поликлональных и три моноклональных антитела были сшиты с ЩФ с использованием глутаральдегида. За исключением С16, все они были биспецифическими антителами, связывающимися как с SNAP25FL так и с SNAP25197, что было ключевым свойством в качестве второго антитела для обнаружения для нормализации. С16 IgG использовали в качестве контроля, поскольку конъюгация пероксидазы хрена была эффективной, но не влияла на антигенную специфичность С16 (Фиг. 18). В поисках оптимальных условий для получения конъюгата ЩФ-IgG, сохраняющего его антигенную специфичность и реактивность, сшивание ЩФ с IgG выполняли с различными соотношениями IgG к ЩФ в различных условиях инкубации (Таблица 17).
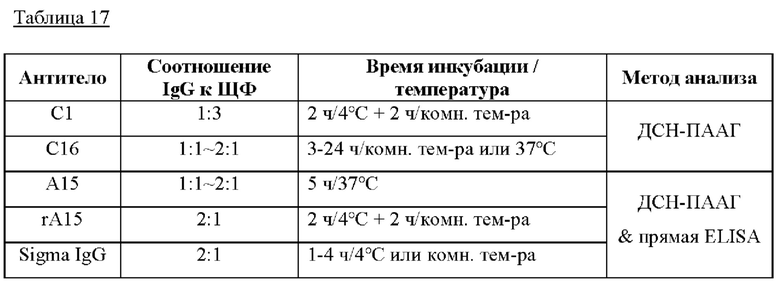
После завершения сшивания с глутаральдегидом полученные конъюгаты ЩФ-IgG анализировали с помощью ДСН-ПААГ. Репрезентативный результат, показывающий электрофоретическое разрешение конъюгатов ЩФ-IgG и последующую визуализацию с помощью окрашивания кумасси синим, показан на Фиг. 20. Вкратце, поликлональный IgG против SNAP25 (Sigma) был перекрестно связан с ЩФ в реакции с 0,2% глутаровым альдегидом. Сшивание ЩФ было высокоэффективным, так как все IgG образовывали высокомолекулярные комплексы с ЩФ. Полученные конъюгаты ЩФ-IgG демонстрировали гораздо более медленную миграцию на геле с ДСН, и некоторые не могли проникнуть в часть концентрирующего геля (Фиг. 20а). Аналогичные картины были получены для всех протестированных IgG (данные не показаны).
После подтверждения сшивания анализом ДСН-ПААГ конъюгаты ЩФ-rA15 IgG и ЩФ-Sigma IgG (100 нг на лунку) были исследованы в сравнении на активность антигена и специфичность в прямом ELISA с использованием микропланшета, покрытого 2 нг смесей GST-SNAP25, содержащие различные количества GST-SNAP25197. Конъюгаты ЩФ-Sigma IgG не давали никакого сигнала ELISA, тогда как конъюгаты ЩФ-rA15 IgG приводили к результату, аналогичному результату с биотинилированным IgG А15 (сравните Фиг. 19 и Фиг. 20б). При увеличении количества GST-SNAP25197 более сильные сигналы ELISA были получены с конъюгатами АР-rA15 IgG.
Результаты, показанные на Фиг. 19 и 20, предполагают, что в случае антител, реагирующих с SNAP25FL, будь то моноклональные или поликлональные антитела, как тяжелая, так и легкая цепи IgG эффективно конъюгированы с активированным биотином или перекрестно связаны с ЩФ с помощью глутаральдегида. Таким образом, второе детектирующее антитело еще предстоит разработать, пока не станет доступна технология конъюгации, более специфичная для Fc.
Пример 16: Оптимизация выращивания N2-42F и обработки BoNT/A
Получив ключевые реагенты, такие как нейрональные клетки, высокочувствительные к BoNT/А, и моноклональные антитела, специфичные к SNAP25, была проведена серия экспериментов для оптимизации всех этапов анализа активности с использованием клеток, включая среду интоксикации, сенсибилизатор, BoNT/А, время обработки и пару антител связывания/обнаружения.
Пример 16-1. Оптимизация времени интоксикации BoNT/A
Во-первых, было оптимизировано время обработки BoNT/A (время токсинизации). Протокол А (Фиг. 21а) представляет собой стандартизированную процедуру СВРА, оптимизированную для SiMa. Для исследования чувствительности к BoNT/A в соответствии с протоколом А клетки N2-42F высевали из расчета 5,6×105 клеток на лунку в 12-луночный культуральный планшет, и на следующий день среду заменяли на 1х интоксикационную среду без GT/1b. Двумя днями позже в среду добавляли GT1b (25 мг/мл), и еще через один день инкубации в культуральную среду добавляли 1х интоксикационную среду, содержащую BoNT/A, и инкубировали в течение дополнительных 2 дней. Протокол В (Фиг. 21б) представляет собой процедуру СВРА, разработанную в этом исследовании. Вкратце, клетки N2-42F высевали из расчета 5,6×105 клеток на лунку в 12-луночном культуральном планшете, и на следующий день среду заменяли 1х средой для интоксикации, содержащей 25 пМ BoNT/A. Общие клеточные лизаты готовили в указанный день путем добавления 1х буфера для образцов ДСН (200 мкл на лунку) и хранили при -20°С перед использованием. Аликвоты (12 мкл) наносили на 12% ДСН-ПААГ, и SNAP25FL и SNAP25197 детектировали вестерн-блоттингом с использованием поликлональных IgG против SNAP25 (Sigma S9684, разведение 1:8000) и козьих антител против кроличьих IgG Fc-HRP (AbFrontier LF - SA8002, разведение 1:8000). Степень расщепления SNAP25 определяли количественно с использованием программного обеспечения Image Lab (Bio-Rad).
Как описано выше, для установления оптимального времени интоксикации для клеток N2-42F протокол А был изменен путем увеличения времени культивирования клеток либо в 1х интоксикационной среде, не содержащей GT1b, либо в 1х интоксикационной среде, содержащей BoNT/A. Эти изменения не улучшили степень расщепления SNAP25. Кроме того, клетки N2-42F, выращенные в среде для интоксикации 1х более 4 дней, выглядели очень нездоровыми под микроскопом (данные не показаны). На основании этого наблюдения был разработан новый протокол В, в котором время культивирования в 1х интоксикационной среде с добавлением как GT1b, так и BoNT/A было сокращено (Фиг. 21б). По прошествии суток, начиная с третьего дня (d4) после посева клеток, степень расщепления SNAP25 в клетках N2-42F сравнивали с помощью вестерн-блоттинга. Как показано на Фиг. 21в, на d4 степень расщепления SNAP25 оценивалась менее 20%, но продолжительное культивирование клеток N2-42F более d4 в 1х интоксикационной среде с добавлением GT1b и BoNT/A привело к значительному увеличению расщепления SNAP25 до 64% на d7. Поскольку клетки N2-42F на d7 выглядели нездоровыми под микроскопом, d6 определяли как день сбора клеток N2-42 и сэндвич-анализа ELISA для измерения активности BoNT/А.
Пример 16-2. Оптимизация времени культивирования
Осмолярность и температура влияли на чувствительность BoCell ТМ к BoNT/A (патент США US 9,526,345 B2). Кроме того, чувствительность клеток NG108-15 к BoNT/A была значительно улучшена за счет оптимизации среды нейральной дифференцировки (J Biomol Screen. 2016 Jan; 21 (1): 65-73). Поскольку чувствительность к BoNT/A отражает степень транспорта BoNT/A внутрь клетки двумя независимыми рецепторами на поверхности клетки, рецептором полисиалоганглиозида (PSG) и рецептором белка (SV2) (J Neurochem. 2009 Jun; 109 (6): 1584-95), повышенная чувствительность к BoNT/A за счет оптимизации культуральной среды или более высокой температуры связана с облегчением транспорта BoNT/A внутрь клетки. С этой целью клетки N2-42F тестировали на чувствительность к BoNT/A в трех различных питательных средах: RPMI1640, Neurobasal™ и MEM, все с добавлением 1х N2, 1x В27 и 1x GT1b. Во время культивирования в указанной среде клетки N2-42F обрабатывали различными концентрациями (0,93~25 пМ) BoNT/A в соответствии с протоколом В. Чувствительность клеток N2-42F к BoNT/А, измеренная с помощью вестерн-блоттинга, была самой высокой в среде RPMI1640 (Фиг. 22). Чувствительность к BoNT/A, измеренная в среде Neurobasal™ или MEM, была на 25% или 50% ниже, чем в RPMI1640. Содержание KCl обычно составляет 5,33 мМ во всех средах, но концентрация NaCl составляет 103 мМ в RPMI1640, 117 мМ в MEM и 52 мМ в среде Neurobasal™. Несмотря на это различие, известно, что осмолярность среды для культивирования клеток для большинства клеток позвоночных находится в узком диапазоне от 260 мОсм/кг до 320 мОсм/кг (АТСС Culture Cell Guide). Таким образом, чувствительность клеток N2-24F в RPMI1640 к BoNT/А, вероятно, обусловлена еще не идентифицированным компонентом среды, кроме осмолярности.
В условиях, описанных на Фиг. 22, около 48% эндогенного SNAP25 расщеплялось в клетках N2-42F под действием 8,33 пМ BoNT/A, что эквивалентно активности примерно 10 единиц/мл. Поскольку объем культуры в 96-луночном планшете составляет 0,1 мл, ЕС50 может быть оценена как биоактивность ~1 Ед. BoNT/A на лунку в микропланшете. Таким образом, чувствительность к BoNT/A, измеренная с клетками N2-42F в соответствии с протоколом В, достаточно чувствительна для измерения биологической активности BoNT/А, определенной с помощью биоанализа LD50 для мышей.
Пример 16-3. Идентификация и оптимизация сенсибилизаторов
Воспользовавшись наличием полностью охарактеризованных моноклональных антител, специфичных к SNAP25 (см. ниже), исследовали чувствительность клеток N2-42F к BoNT/А в 1х интоксикационной среде, содержащей различные концентрации BoNT/A (0,03~5,5 пМ), с помощью сэндвич-анализа ELISA, следуя протоколу В, при исследовании, влияют ли на чувствительность к BoNT/A аргинин или любые соединения с продемонстрированным влиянием на выживаемость или дифференцировку нейронов, включая АТФ, (см. выше) (Trends Neurosci. 2000 Dec; 23 (12): 625-33), креатин (J Neurochem. 2005 Oct; 95 (1): 33-45) и липоевая кислота (J Neurosci Res. 2014 Jan; 92 (1): 86-94). Как показано на Фиг. 23а, добавление 1 мМ креатина или 5 мМ аргинина в среду интоксикации 1х заметно увеличивало чувствительность к BoNT/А, снижая значение ЕС50 с 2,51 пМ до 2,13 или 2,03 пМ, соответственно. Напротив, АТФ и липоевая кислота действовали как очень эффективные ингибиторы, поэтому расщепление SNAP25 в клетках N2-42F не обнаруживалось даже при 25 пМ BoNT/A.
В исследовании оптимизации токсинизированной среды считалось, что чувствительность клеток N2-24F к BoNT/A зависит от факторов, отличных от осмотического давления. Потому что чувствительность была выше в среде RPMI1640, чем в среде Neurobasal™ или MEM. Аргинин присутствовал в составе среды в концентрациях 1,15 мМ в RPMI1640, 0,6 мМ в MEM и 0,4 мМ в Neurobasal™. Поскольку аргинин является аминокислотой-предшественником креатина, аргинин отдельно исследовали на предмет его влияния на чувствительность к BoNT/A с использованием 2-кратной интоксикационной среды, содержащей 2~10 мМ аргинина. Как показано на Фиг. 23б, чувствительность к BoNT/A постепенно повышалась с увеличением концентрации аргинина до 5 мМ, что отражалось в снижении значений ЕС50. Чувствительность к BoNT/A с 10 мМ аргинина (ЕС50=2,34 пМ), хотя и выше, чем в контроле (ЕС50=2,94 пМ), была ниже, чем с 5 мМ аргинином (ЕС50=1,65 пМ). Основываясь на этом результате, хотя механизм его действия еще предстоит понять, в оптимизированном стандартном протоколе СВРА использовали интоксикационную среду, содержащую 5 мМ аргинина.
В 2002 году Шенгрунд и его коллеги впервые предоставили экспериментальные доказательства того, что в клетках Neuro-2a для эффективного расщепления SNAP25 с помощью BoNT/A требуется более 25 мкг/мл GT1b в DMEM (J Biol Chem. 2002, 6 сентября; 277 (36): 32815-9). Поскольку клетки N2-42F происходят из Neuro-2a, потребность в GT1b для активности BoNT/A исследовали с использованием 1х интоксикационной среды, содержащей 25-75 мкг/мл GT1b (1~3х GT1b), методом вестерн-блоттинга. Без добавления GT1b наличие BoNT/A в концентрации 8,3 пМ приводило к примерно 18% расщеплению SNAP25 (Фиг. 24а). Добавление 1х или 3х GT1b приводило к 10% и 18% увеличению расщепления SNAP25 при 8,3 пМ BoNT/A, соответственно, в клетках N2-42F (Фиг. 24а).
В целях оптимизации концентрации GT1b, чувствительность клеток N2-42F к BoNT/A тестировали в 2х интоксикационной среде, содержащей 25 пМ BoNT/А, увеличивая концентрации GT1b от 1х до 5х. При измерении методом вестерн-блоттинга расщепление SNAP25 было значительно усилено добавлением 1х или 2х GT1b, но с более чем 2х GT1b расщепление SNAP25 увеличилось лишь незначительно с 63%, 65%, 68% до 70% (Фиг. 24б). Параллельный тест проводили с использованием оптимизированного сэндвич-ELISA. Принимая во внимание чувствительность сэндвич-ELISA, клетки N2-42F обрабатывали 0,93 пМ BoNT/A в 2х интоксикационной среде согласно протоколу В. Сэндвич-ELISA более четко продемонстрировал потребность в GT1b для активности BoNT/A. Вкратце, когда клетки N2-42F обрабатывали BoNT/A в интоксикационной среде без GT1b, в сэндвич-ELISA были получены относительно низкие значения А450 (Фиг. 24в). Напротив, добавление GT1b к среде интоксикации привело к заметному увеличению значения А450. Результат, показывающий устойчивое увеличение значения А450 до 2х GT1b, может отражать эффективное трехмолекулярное взаимодействие между BoNT/A, GT1b и рецептором полисиалоганглиозида (PSG). Следовательно, значение насыщения А450, полученное с 4х GT1b и даже уменьшенное значение А450 с 5х, может быть объяснено насыщением рецептора PSG и/или эффектом молярного избытка свободного GT1b, который конкурирует с комплексом BoNT/A-GT1b за рецептор PSG. Основываясь на этом рассуждении, в качестве оптимальной концентрации GT1b была выбрана 2х кратная концентрация, то есть 50 мкг/мл, когда активность BoNT/A измеряется с использованием клеток N2-42F.
Было протестировано влияние N2 (добавка N2, Thermo Fisher Scientific 17502048) / В27 (бессывороточная добавка В27ТМ, Thermo Fisher Scientific 17504-044) в культурах нейронов на чувствительность к BoNT/А. Известно, что В27, содержащий весь трансретинол (0,1 мг/л), поддерживает дифференцировку двигательных нейронов нервных клеток-предшественников (J Cell Biochem. 2008 Oct 15; 105 (3): 633-40). N2 содержит подмножество компонентов В27, включая инсулин, и известно, что он способствует (1) дифференцировке эмбриональных стволовых клеток человека и (2) пролиферации / выживанию нервных клеток-предшественников (J Cell Biochem. 15 октября 2008 г.; 105 (3): 633 -40). Чувствительность к BoNT/A клеток N2-42F в RPMI1640 с добавлением 1х GT1b, 8,3 пМ BoNT/A и указанной концентрации N2/B27 в соответствии с протоколом В. Расщепление SNAP25 количественно анализировали с помощью вестерн-блоттинга. Как показано на Фиг. 25а, внутриклеточные уровни всего SNP25 (SNAP25FL+SNAP25197) увеличивались пропорционально концентрации N2/B27, тогда как уровень SNAP25197 оставался относительно неизменным и даже снижался в присутствии 4х или 5х N2/B27. Недавнее исследование показало, что N2 и В27 действуют совместно, защищая нейрон от гибели после истощения глюкозы, ограничивая гликолиз (Front Mol Neurosci. 29 сентября 2017; 10: 305). Соответственно, результат на Фиг. 25а, скорее всего, отражает усиленный синтез эндогенного SNAP25 с увеличением концентраций N2/B27 как часть их функции по содействию пролиферации клеток и выживанию в бессывороточной среде RPMI1640, содержащей относительно низкую концентрацию глюкозы (11,1 мМ).
Используя RPMI1640 с добавлением 3х GT1b и 5 мМ аргинина, дополнительно исследовали влияние N2/B27 на активность BoNT/A. Степень расщепления SNAP25 в клетках N2-42F, обработанных 8,3 пМ BoNT/A, увеличилась с 13% до 56% и 69% при 1х и 2х N2/B27, несмотря на заметное увеличение общего внутриклеточного уровня SNAP25 (Фиг. 25б). На основании этого наблюдения к оптимизированной среде для интоксикации добавляли 2х N2/B27.
Пример 17: Оптимизация сэндвич-ELISA
Пример 17-1. Оптимизация буферов
После оптимизации культуральной среды и добавок для среды для интоксикации клеток N2-42F, систематически оценивали эффективность сэндвич-ELISA в различных условиях, приведенных в Таблице 18, и выбирали оптимизированные условия, сведенные в Таблицу 19.
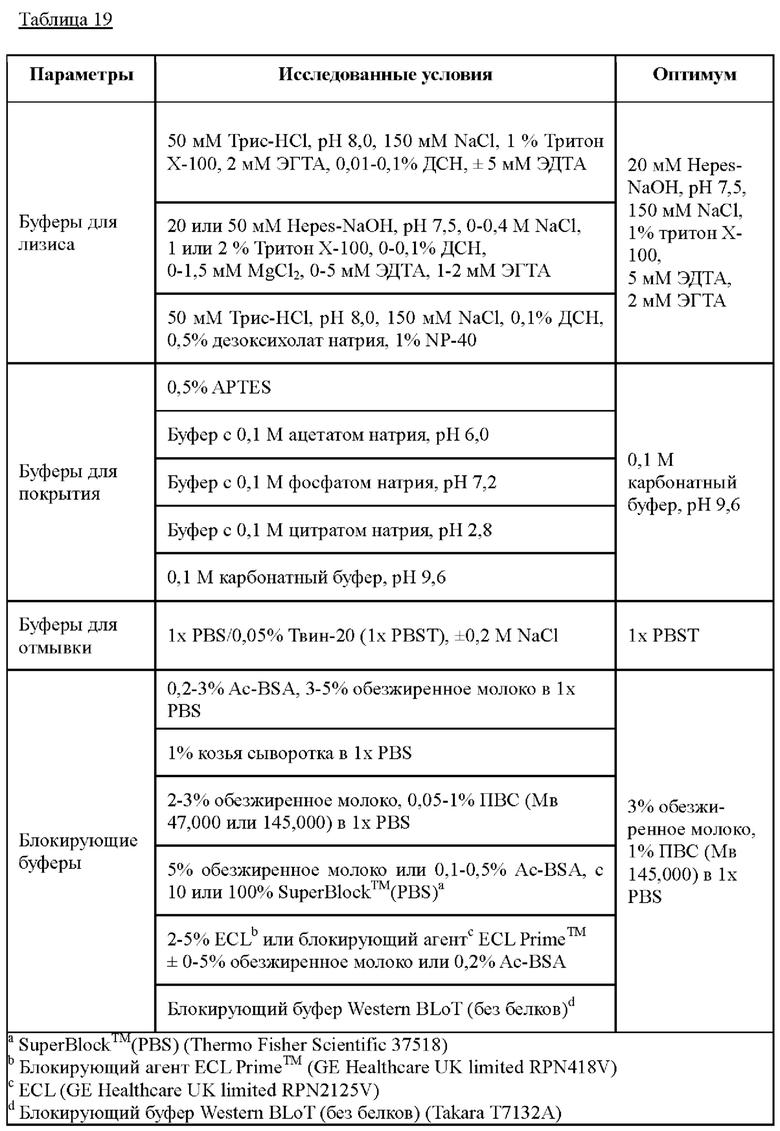
Пример 17-2. Оптимизация связывающих антител
Во вторую очередь, три биспецифических антитела, А15, В4 и В23, сравнивали на предмет их функции в качестве захватывающего антитела в сэндвич-ELISA. ОКЛ готовили из клеток N2-42F, обработанных различными концентрациями (0-25 пМ) BoNT/A в 1х интоксикационной среде. ОКЛ вносили в микропланшет, покрытый указанным антителом (400 нг на лунку). После инкубации в течение 4 часов при 4°С количество SNAP25197 среди общего количества SNAP25, связанного указанным антителом, обнаруживали и количественно определяли с использованием конъюгатов С16 IgG-HRP в соответствии с процедурой, описанной в разделе «Материалы и методы». Единственным исключением было то, что реакцию с пероксидазой хрена проводили в течение 30 минут до тех пор, пока все тестовые группы не дали положительный сигнал ELISA. Как показано на Фиг. 26а, расчетные значения ЕС50 составляли 15 пМ, 0,7 пМ и 5,5 пМ для А15, В4 и В23 IgG соответственно. Этот результат согласуется с их значениями KD для SNAP25FL и SNAP25197.
Оптимальное количество В4 IgG определяли с помощью ОКЛ, приготовленных из клеток N2-42F, инкубированных в 2-кратной интоксикационной среде, содержащей указанную концентрацию BoNT/A, согласно протоколу В. Сэндвич-ELISA осуществляли с микропланшетом, покрытым увеличивающимся количеством (100~400 нг на лунку) В4 IgG. После инкубации в течение 4 часов при 4°С связанный SNAP25197 обнаруживали и количественно определяли с использованием конъюгатов С16 IgG-HRP, как описано выше. Следует отметить, что реакцию с пероксидазой хрена проводили в течение 15 мин. Как показано на Фиг. 26б, значение ЕС50 неуклонно снижалось с увеличением количества В4 IgG со 100 до 300 нг. Дальнейшего статистически значимого улучшения ЕС50 при применении 400 нг В4 IgG не наблюдалось. Принимая во внимание все вышеуказанное, в качестве стандартного количества связывающих антител в оптимизированном сэндвич-ELISA использовали 300 нг В4 IgG.
Пример 17-3. Оптимизация условий инкубации ОКЛ и детектирования антител
ОКЛ, приготовленные из клеток SiMa, обработанных 8,33 или 25 пМ BoNT/A в 1х интоксикационной среде, инкубировали в микропланшете при указанных условиях. Впоследствии связанный SNAP25197 обнаруживали и количественно определяли с использованием конъюгатов С16 IgG-HRP. Как показано на Фиг. 27а, сигнал ELISA для SNAP25197 был самым сильным, когда микропланшет инкубировали в течение 4 часов при 4°С, но был очень слабым после инкубации при 37°С в течение 1 часа. Хотя данные не показаны, аналогичные результаты были получены с ОКЛ клеток N2-42F, и сигнал ELISA существенно не изменился при инкубации более 4 часов. Таким образом, в оптимизированном сэндвич-ELISA ОКЛ инкубировали в течение 4 часов при 4°С.
Оптимальные условия инкубации для обнаружения антител исследовали прямым ELISA. Вкратце, микропланшеты покрывали различными количествами (от 10 пг до 1 нг) рекомбинантного GST-SNAP25197, которые представляют собой эндогенное расщепление SNAP25 от 0,05% до 50% в стандартном СВРА. После добавления конъюгатов С16 IgG-HRP (200 нг на лунку) микропланшеты инкубировали в указанных условиях. В условиях теста инкубация конъюгатов Cg16 IgG-HRP при комнатной температуре в течение 1 часа давала наибольший сигнал ELISA для всех диапазонов исследованных GST-SNAP25197, но в других условиях также генерировались приемлемые уровни сигнала ELISA. Однако при рассмотрении последующей реакции с пероксидазой хрена, проводимой при комнатной температуре, в оптимизированном сэндвич-ELISA конъюгаты C16-HRP инкубировали в течение 1 часа при комнатной температуре.
Пример 17-4. Оптимизация способа обнаружения конъюгатов С16 IgG-HRP
Активность пероксидазы хрена обычно измеряли с использованием традиционного субстрата ТМВ (1-StepTM Ultra TMB-ELISA). Одним из недостатков использования субстрата ТМВ является то, что на значение ЕС50 обычно влияет время реакции с пероксидазой хрена (Фиг. 26 и 29). Чем дольше реакция с пероксидазой хрена, тем ниже ЕС50. В качестве альтернативного способа обнаружения сравнительно оценивали флуориметрическое измерение с помощью сэндвич-ELISA. За исключением предоставленного реакционного субстрата пероксидазы хрена, колориметрические и флуориметрические измерения проводили параллельно, следуя по существу одинаковому способу на всем протяжении сэндвич-ELISA. Флуориметрические и колориметрические измерения дали 0,66 и 0,78 пМ ЕС50 (Фиг. 28), и оба значения ЕС50 в равной степени зависели от времени реакции с пероксидазой хрена (данные не показаны). В заключение, несмотря на то, что для флуориметрического измерения реакции с пероксидазой хрена необходимы черный микропланшет и встроенный считыватель микропланшетов с флуориметром, оба измерения являются точными и достаточно чувствительными, чтобы определить активность конъюгатов С16 IgG-HRP в оптимизированном ELISA.
Пример 18: Подтверждение эффективности и практическое применение СВРА
Пример 18-1. Оптимизированный временной график СВРА
Объединив все оптимизированные условия и реагенты, окончательно отлаженная оптимальная СВРА может быть осуществлена всего за три рабочих дня, как показано на Фиг. 29. Вкратце, клетки высевали на 1 день, а на следующий день 2 среду заменяли на 2х интоксикационную среду, содержащую BoNT/A (0,1~10 Ед/мл или менее 4 пМ). На 6 день клетки обрабатывали буфером для лизиса, и с полученными ОЛК проводят сэндвич-ELISA и определяют биологическую активность BoNT/A. Избегая лабораторной работы в выходные дни, можно выполнять оптимизированную СВРА три раза в неделю, тратя только 1-2 рабочих часа в течение первых двух дней и один полный рабочий день. Таким образом, оптимизированный СВРА подходит для измерения биологической активности нескольких партий фармацевтических или косметических продуктов BoNT/A в неделю. Использование двух моноклональных антител для сэндвич-ELISA, за исключением поликлональных антител кролика, делает СВРА очень надежным, поскольку их получение и последующий контроль качества превосходят таковые для поликлональных антител.
Пример 18-2. Точность и линейность СВРА
Следуя стандартному протоколу В, три оператора провели в общей сложности 18 СВРА.
В общей сложности 18 СВРА были выполнены тремя операторами в соответствии с протоколом В. Клетки N2-42F обрабатывали различными концентрациями (0, 0,03, 0,1, 0,2, 0,31, 0,62, 0,93, 2,78, 5,55, 8,33 пМ) BoNT/A в 2х среде для интоксикации (Фиг. 30а) или 2х среде для интоксикации, содержащей 3х GT1b (Фиг. 30б и Фиг. 30в). Реакцию с пероксидазой хрена проводили в течение 5 минут (Фиг. 30а и 30б) или 9 минут (Фиг. 30в), а ЕС50 определяли с использованием программного обеспечения Gen5. Анализ линейной регрессии был выполнен со значениями А450, полученными с клетками N2-42F, обработанными 0,1-0,93 пМ BoNT/А, и результаты показаны на Фиг. 30.
Первая серия СВРА дала 1,39 пМ ЕС50 с 1,5 фМ (относительная активность 3,4 мЕ/мл) предела обнаружения (ПО) и 4,6 фМ предела количественного определения (ПКО) (Clin Biochem Rev. 2008 Aug; 29 Suppl 1: S49-52; Anal Sci. 2007, Feb; 23 (2): 215-8). Поскольку 1,39 пМ BoNT/A эквивалентно относительной активности ~0,3 Ед на анализ, первый СВРА достаточно чувствителен для измерения биологической активности BoNT/A вместо анализа LD50 с использованием мышей. Немного более низкое значение ЕС50, 1,24 пМ, было получено путем продолжительной реакции с пероксидазой хрена, используемой во второй серии СВРА (Фиг. 30б). Влияние времени реакции с пероксидазой хрена на ЕС50 было замечено ранее (Фиг. 29). Третья серия СВРА, выполненная с использованием интоксикационной среды с 3х GT1b, дала самый низкий уровень ЕС50 1,09 пМ, что согласуется с результатами на Фиг. 27. Этот результат указывает на то, что эффективность количественного определения и предела обнаружения СВРА, разработанные в настоящем исследовании, можно регулировать, регулируя время реакции с пероксидазой хрена или изменяя концентрацию GT1b в среде интоксикации. Линейный регрессионный анализ сигналов ELISA для клеток N2-42F, обработанных 0,03~0,93 пМ BoNT/А, выявил отличную линейную корреляцию между относительной эффективностью BoNT/A и сигналом ELISA во всех трех сериях СВРА (Фиг. 30). Этот результат показывает, что СВРА не только высокочувствителен, но и очень точен при измерении относительной активности BoNT/A между 6,8 мЕд. и 0,2 Ед. на анализ.
Пример 18-3. Измерение биологической активности образцов BOTULAX®
Эффективность BoNT/A в BOTULAX® определяли по LD50 с использованием мышей. Для измерения этой биологической активности с использованием СВРА две разные партии (HUB 18009 и HUB 18011) BOTULAX® (200 Ед на флакон) растворяли либо в интоксикационной среде (матрица а), либо в деионизированной Н2О (матрица б), либо в 5 мМ аргинине, рН 6.0 (матрица в). После 10 мин инкубации при комнатной температуре растворенный в среде BOTULAX® серийно разбавляли до 0,015, 0,05, 0,15, 0,5, 1,5, 5, 15 и 50 Ед/мл, а остальные - до 0,23, 0,48, 0,70, 0,98, 1,41., 2,11, 4,20, 6,32 и 12,6 Ед/мл в оптимизированной среде для интоксикации. Проводили три различных серии СВРА с использованием этих образцов в соответствии со стандартным протоколом В. Были определены значения ЕС50. Как показано на Фиг. 31, значения ЕС50 для двух партий составляли 10,6±0,12 и 10,3±0,52 Ед/мл с матрицей а, 4,6±0,04 и 4,6±0,08 Ед/мл с матрицей 6 и 4,5±0,11 и 4,4±0,22 Ед/мл с матрицей в. Этот результат показывает, что СВРА чувствителен и достаточно точен для измерения биологической активности BOTULAX®, а его чувствительность (0,4-0,5 ЕД на лунку) эквивалентна или превосходит биоанализ с использованием мышей.
Хотя настоящее описание изобретения было подробно раскрыто со ссылкой на конкретные признаки, специалистам в данной области техники будет очевидно, что это описание представляет собой только его предпочтительный вариант осуществления и не ограничивает объем настоящего изобретения. Таким образом, объем настоящего изобретения по существу определяется прилагаемой формулой изобретения и ее эквивалентами.
Регистрационный номер
Орган-депозитарий: Корейский научно-исследовательский институт биологии и биотехнологии;
Регистрационный номер: KCTC13712BP;
Дата депонирования: 13 ноября 2018 г.
Промышленная применимость
Настоящее изобретение призвано преодолеть ограничения обычного СРВА и разработать более эффективный СВРА. В настоящее время существует необходимость в разработке анализа активности с использованием клеток (СВРА) для замены биоанализа LD50 с использованием мышей (mLD50) в области измерения активности ботулотоксина. Ожидается, что СВРА, использующий клетку и антитело согласно настоящему изобретению, станет высоконадежным и воспроизводимым анализом активности ботулотоксина с использованием клеток.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ ГЛИПИКАНА 3 | 2005 |
|
RU2427588C2 |
| АНТИТЕЛО ПРОТИВ ГЛИПИКАНА 3 | 2011 |
|
RU2611751C2 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО, СОДЕРЖАЩЕЕ АНТИТЕЛО К ФОСФОЛИПАЗЕ D4 | 2014 |
|
RU2709741C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ПРОТИВ СПЕЦИФИЧЕСКОГО МЕМБРАННОГО АНТИГЕНА ПРОСТАТЫ (PSMA) И СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК, ЭКСПРЕССИРУЮЩИХ PSMA | 2006 |
|
RU2421466C2 |
| АНТИ-VEGF-АНТИТЕЛО И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ, ДИАГНОСТИКИ ИЛИ ЛЕЧЕНИЯ РАКА ИЛИ СВЯЗАННОГО С АНГИОГЕНЕЗОМ ЗАБОЛЕВАНИЯ | 2014 |
|
RU2644245C2 |
| АНТИТЕЛО ПРОТИВ PD-L1 | 2017 |
|
RU2744862C2 |
| АНТИТЕЛА РЕЦЕПТОРА 1 ИНТЕРФЕРОНА АЛЬФА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2600884C2 |
| ИММУНОЛОГИЧЕСКИЕ ТЕСТЫ НА АКТИВНОСТЬ ЭНДОПЕПТИДАЗ С ИЗМЕНЕННОЙ НАЦЕЛЕННОСТЬЮ | 2010 |
|
RU2543650C2 |
| АНТИТЕЛА РЕЦЕПТОРА 1 ИНТЕРФЕРОНА АЛЬФА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2412202C2 |
| АНТИТЕЛО, СПЕЦИФИЧНО СВЯЗЫВАЮЩЕЕСЯ С ИЗОЛИРОВАННЫМ ПЕПТИДОМ, ОБРАЗОВАННЫМ ИЗ ВИМЕНТИНА, ИЛИ СВЯЗЫВАЮЩИЙСЯ С ЭТИМ ПЕПТИДОМ ФРАГМЕНТ | 2016 |
|
RU2705899C2 |
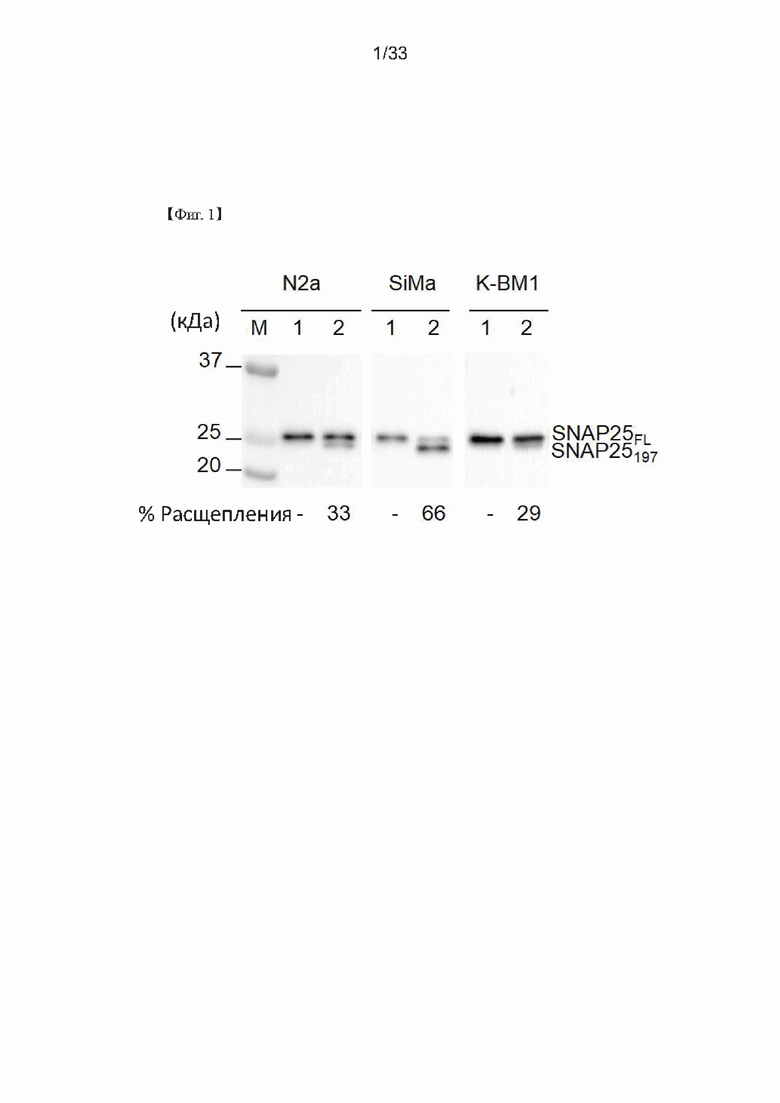

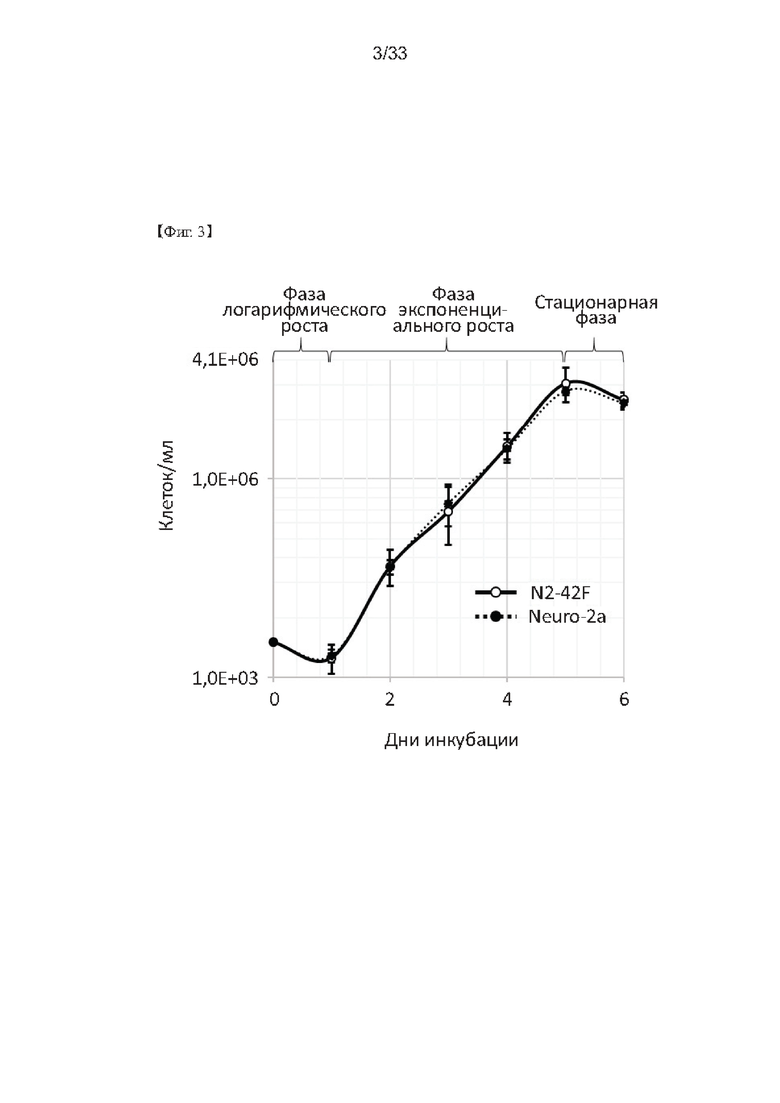
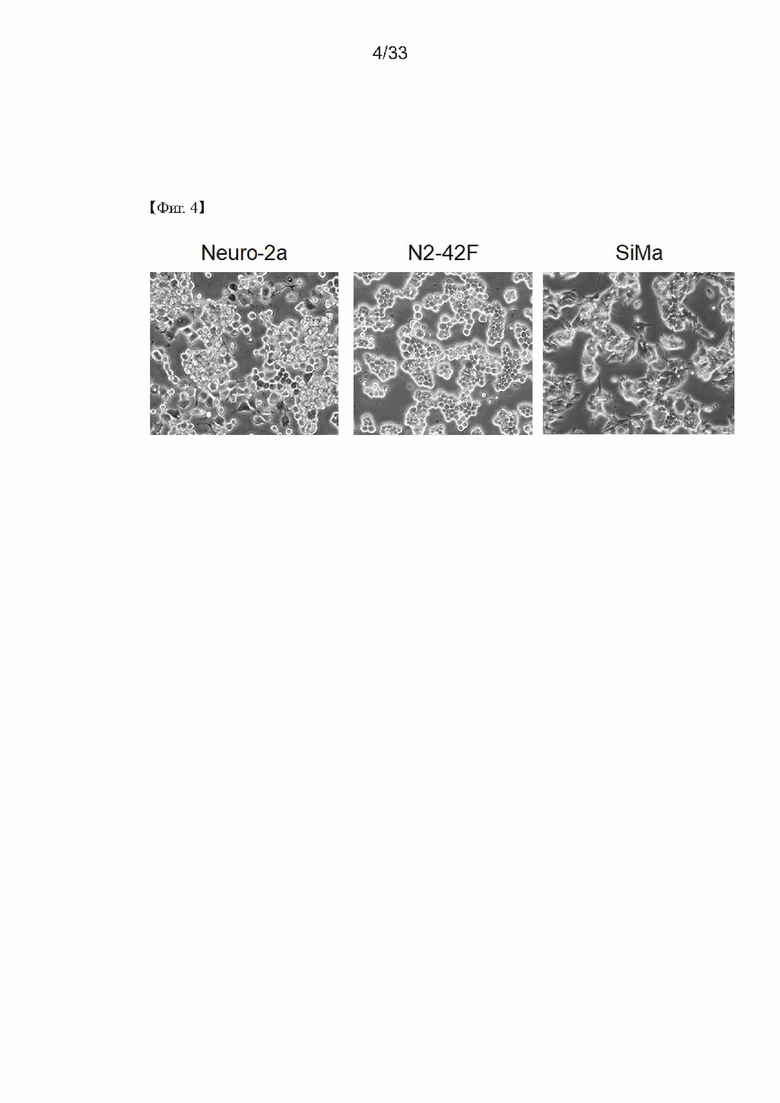
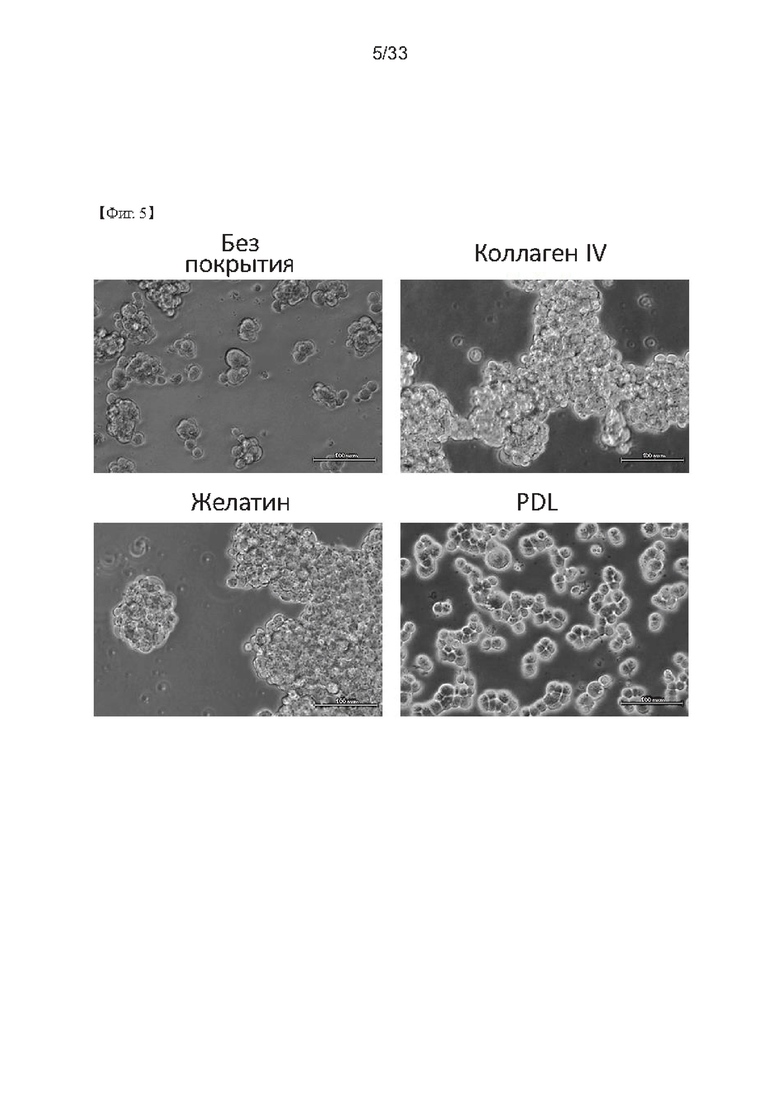
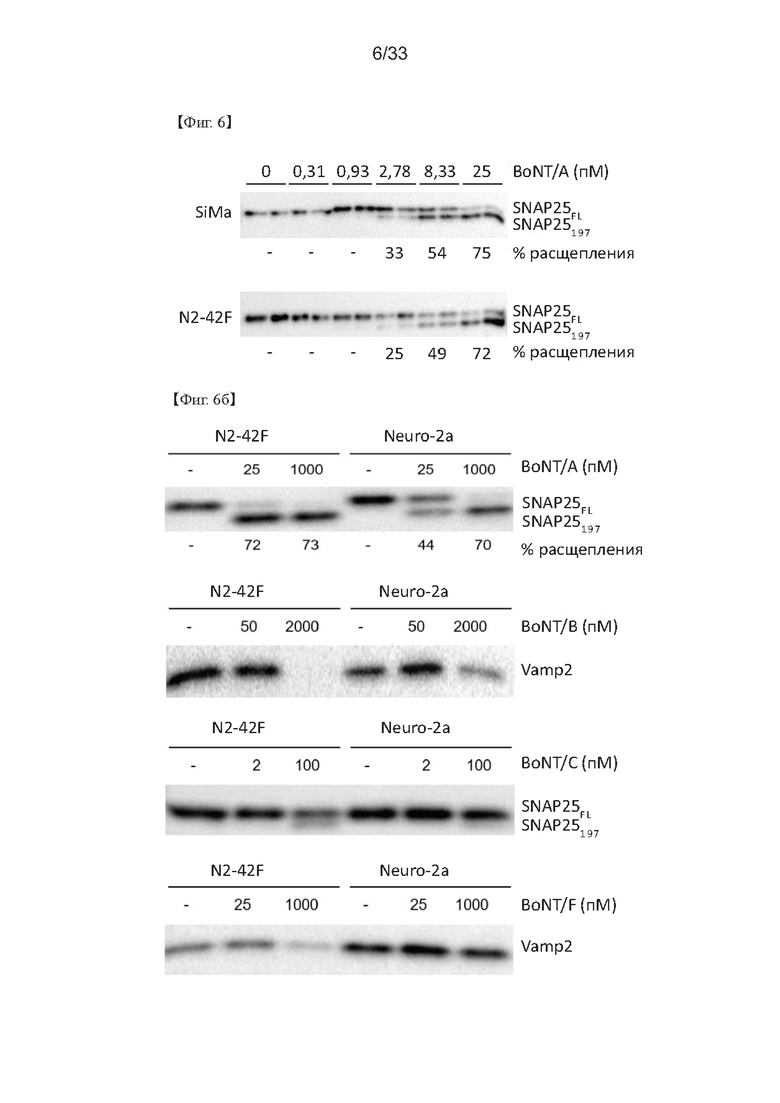
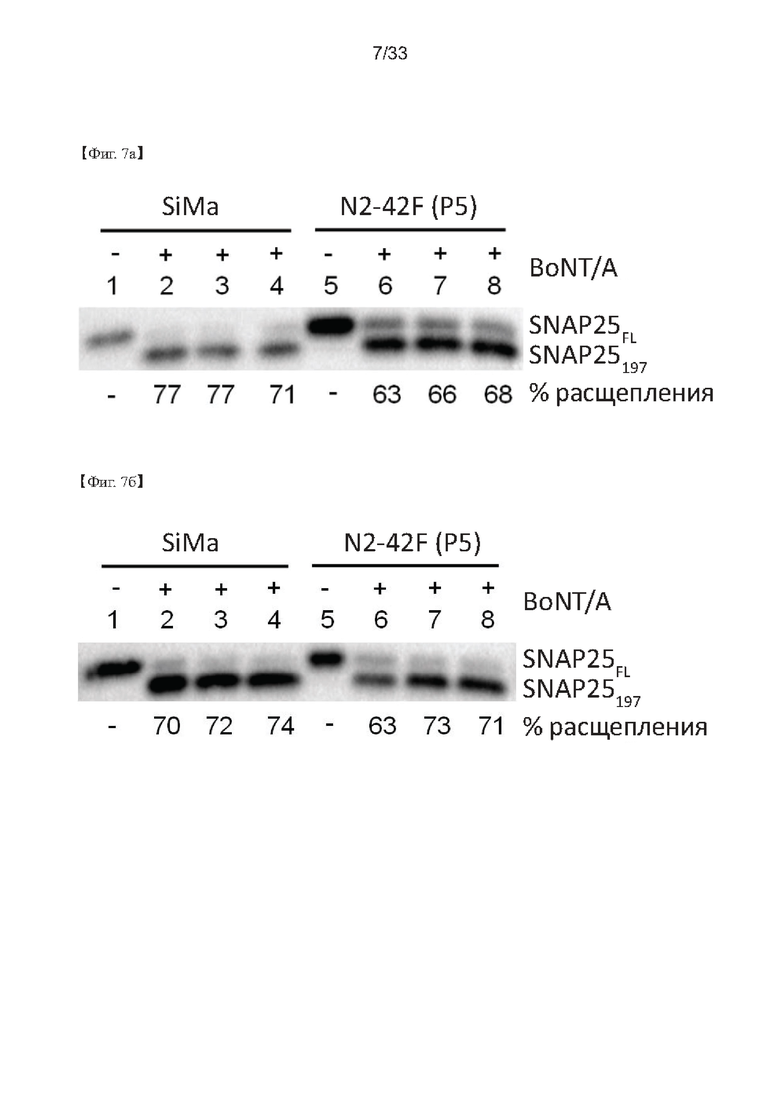
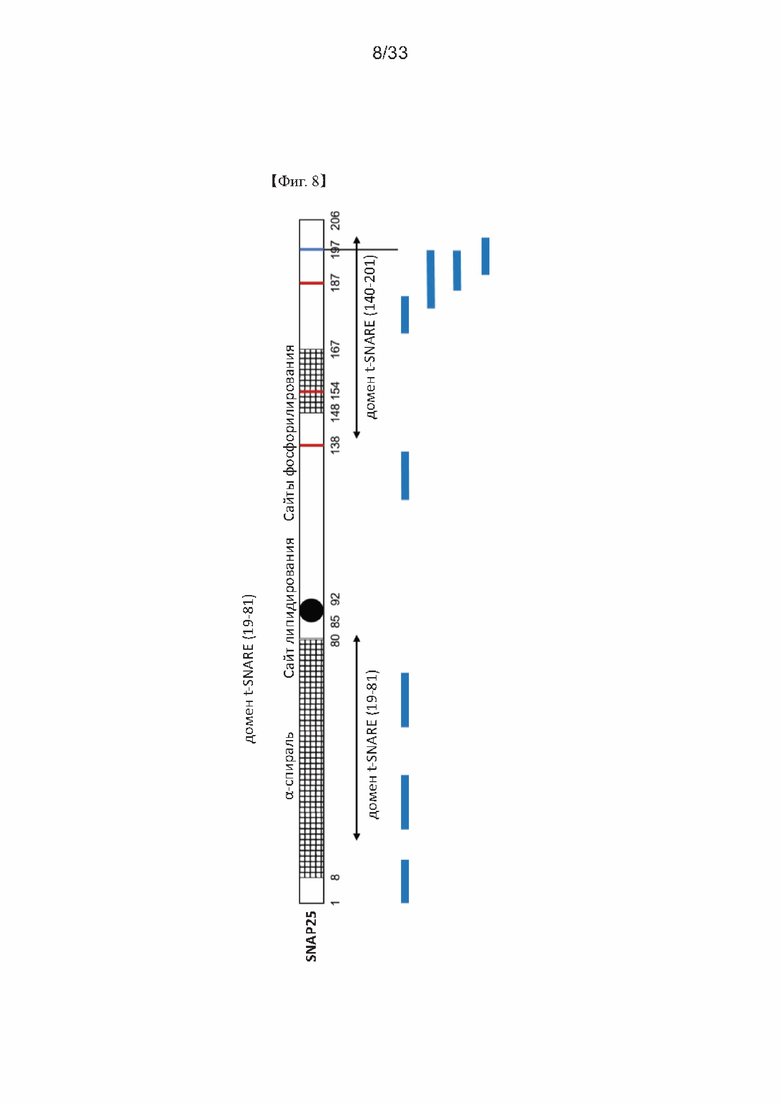

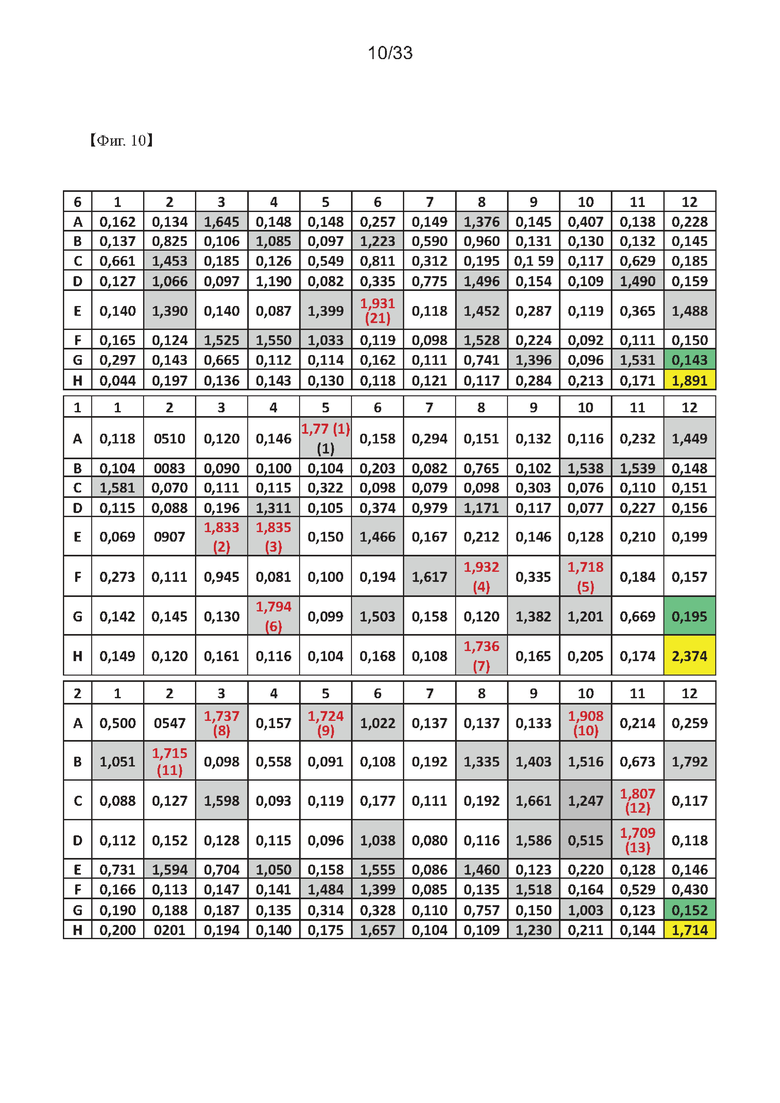
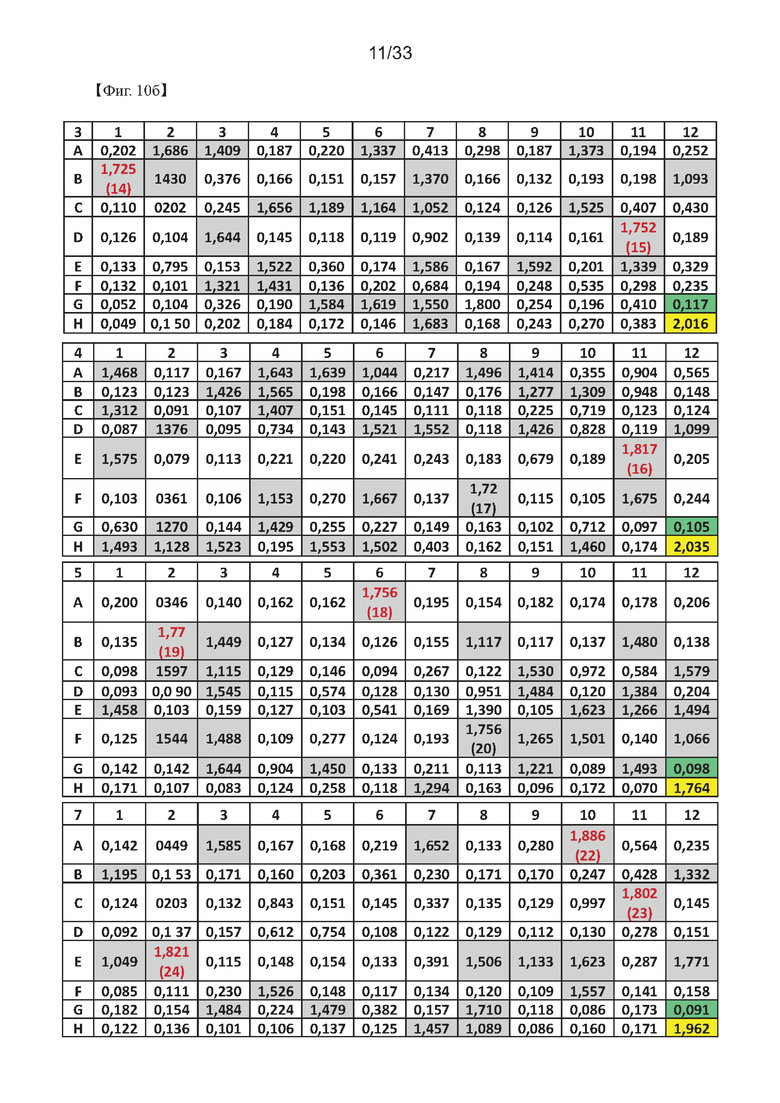
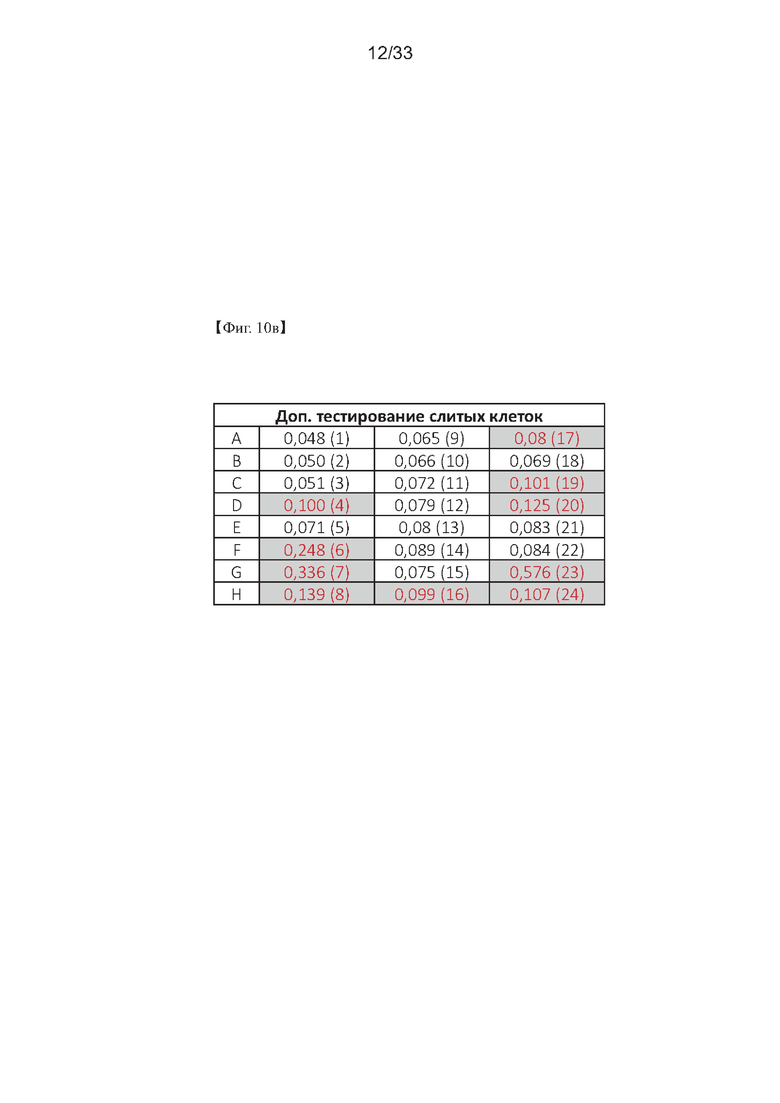
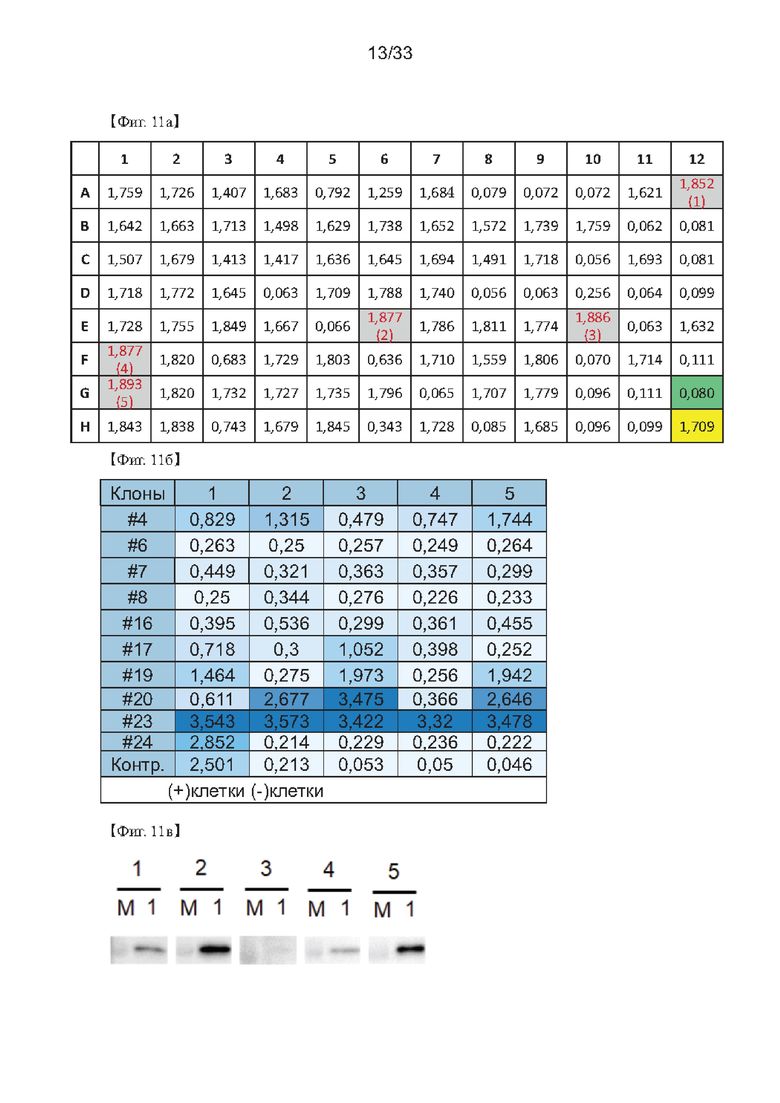
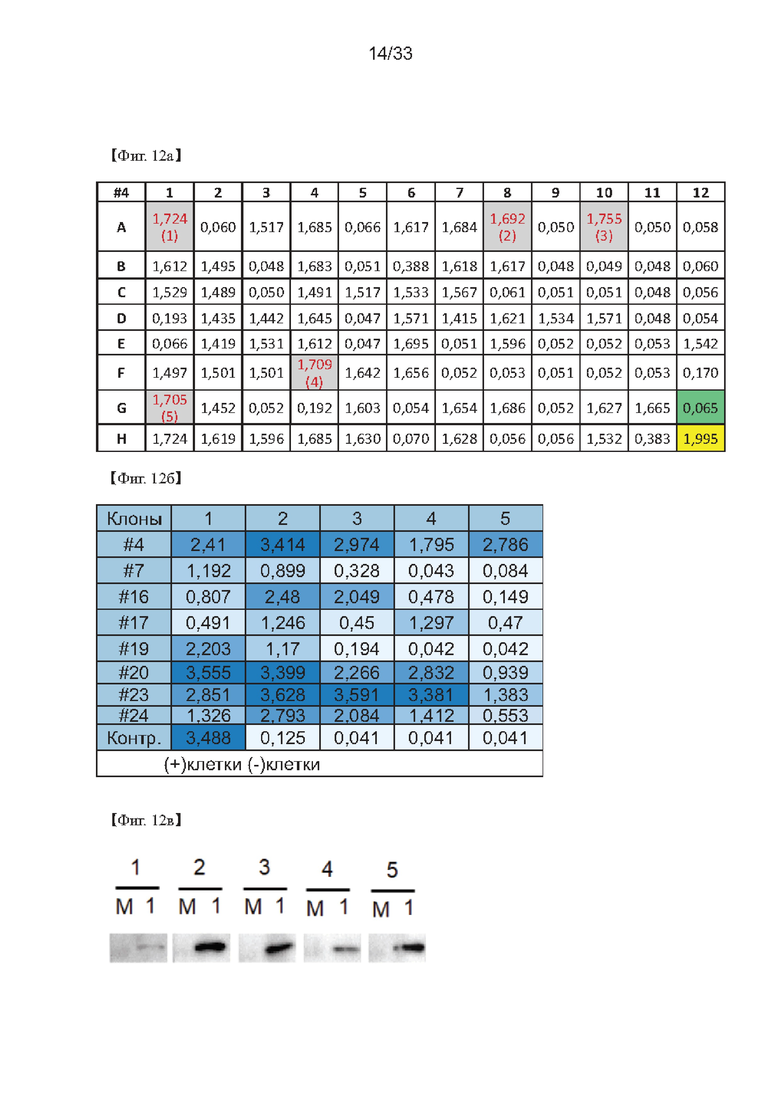
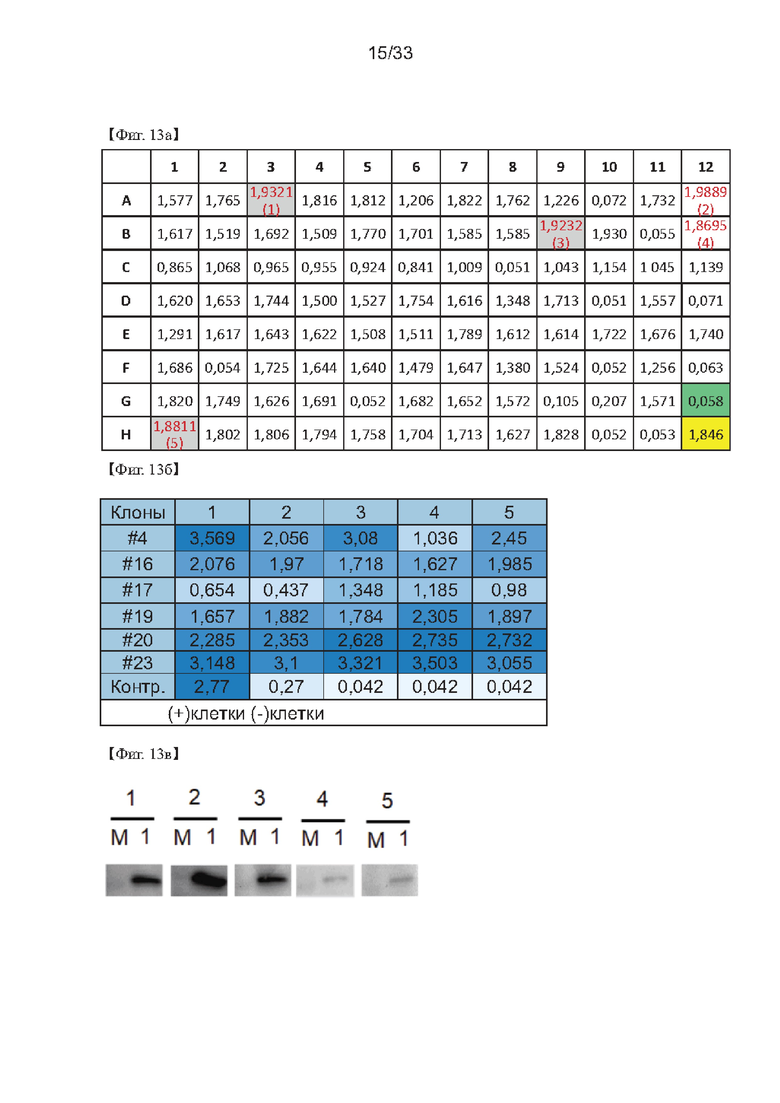
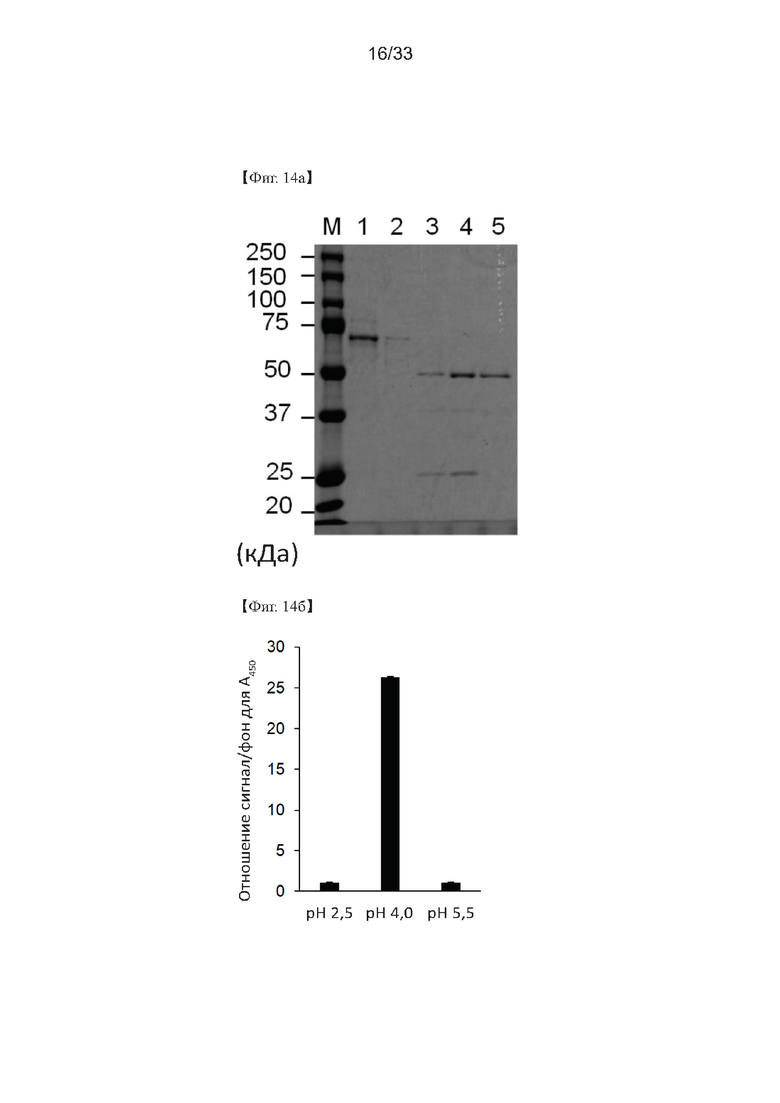
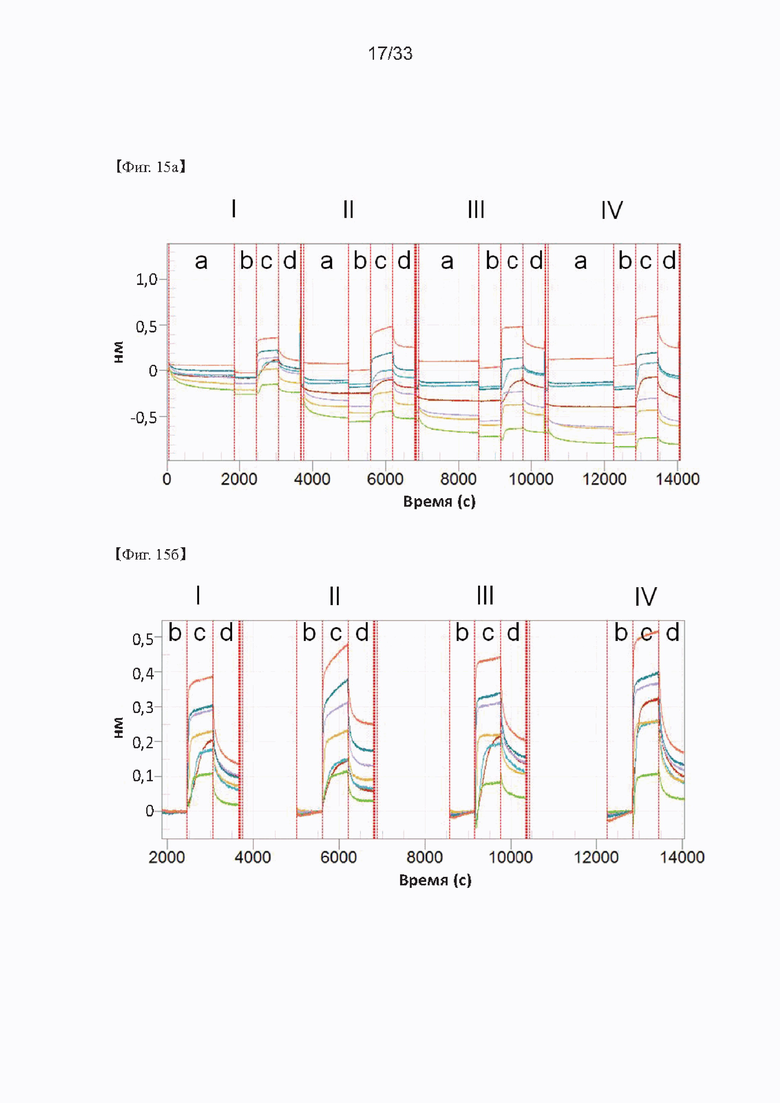
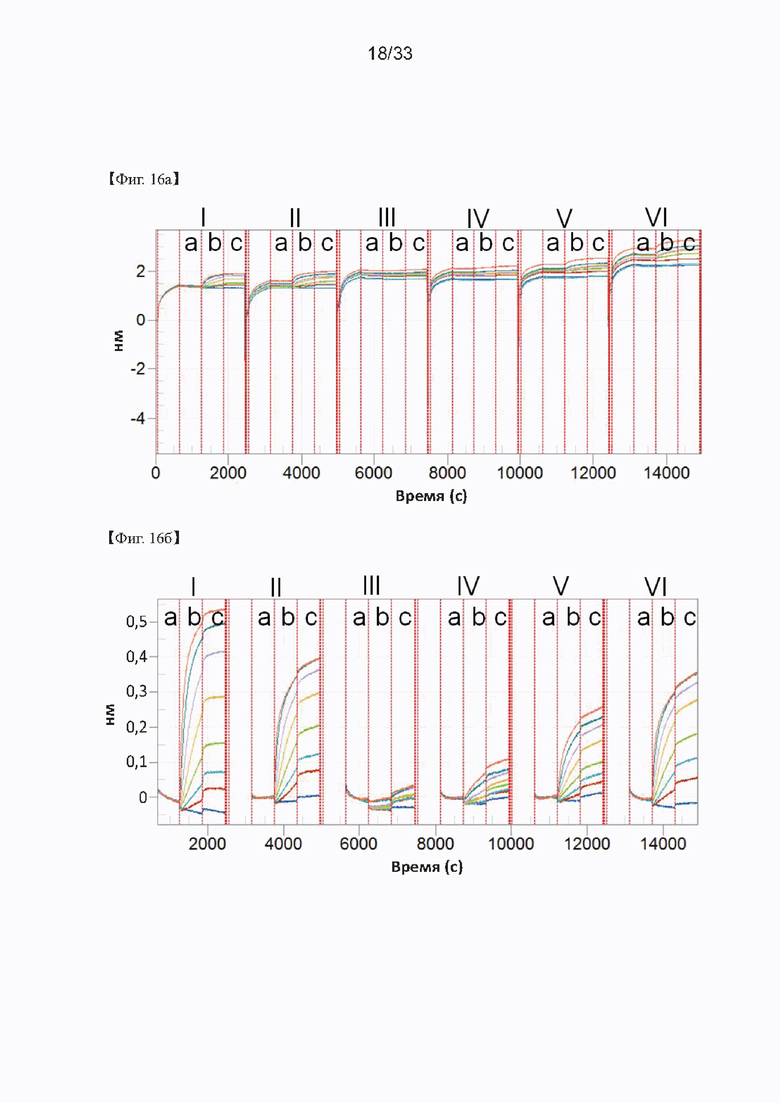
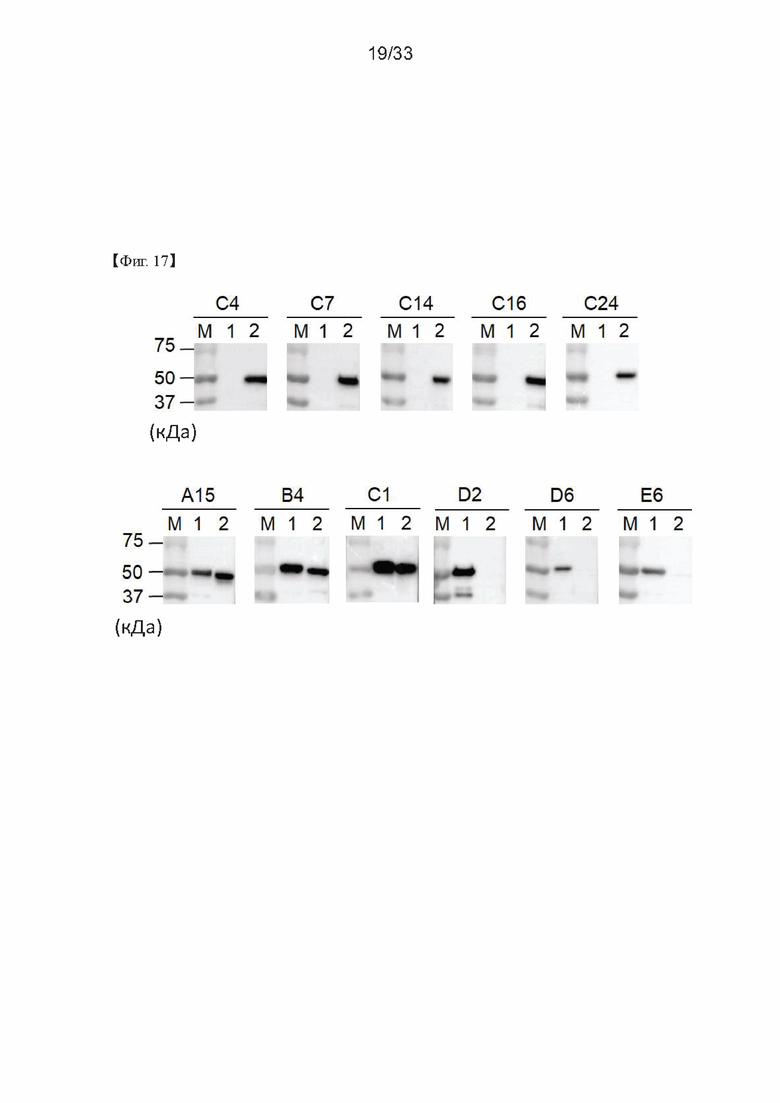
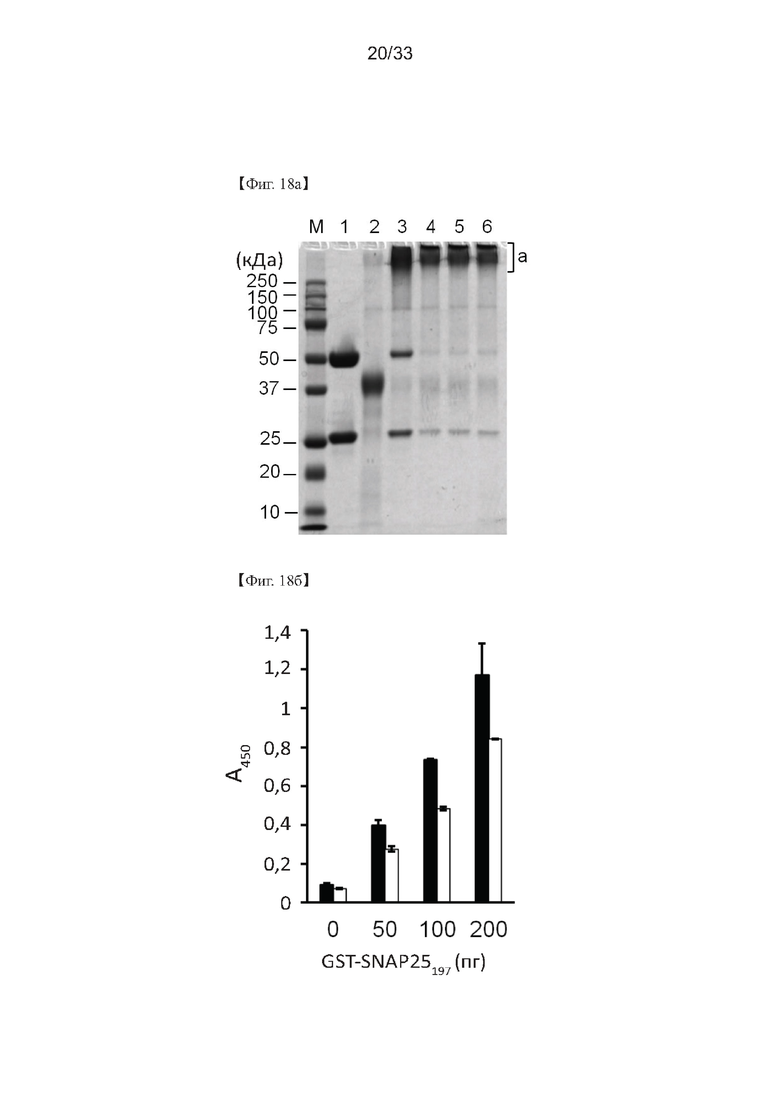
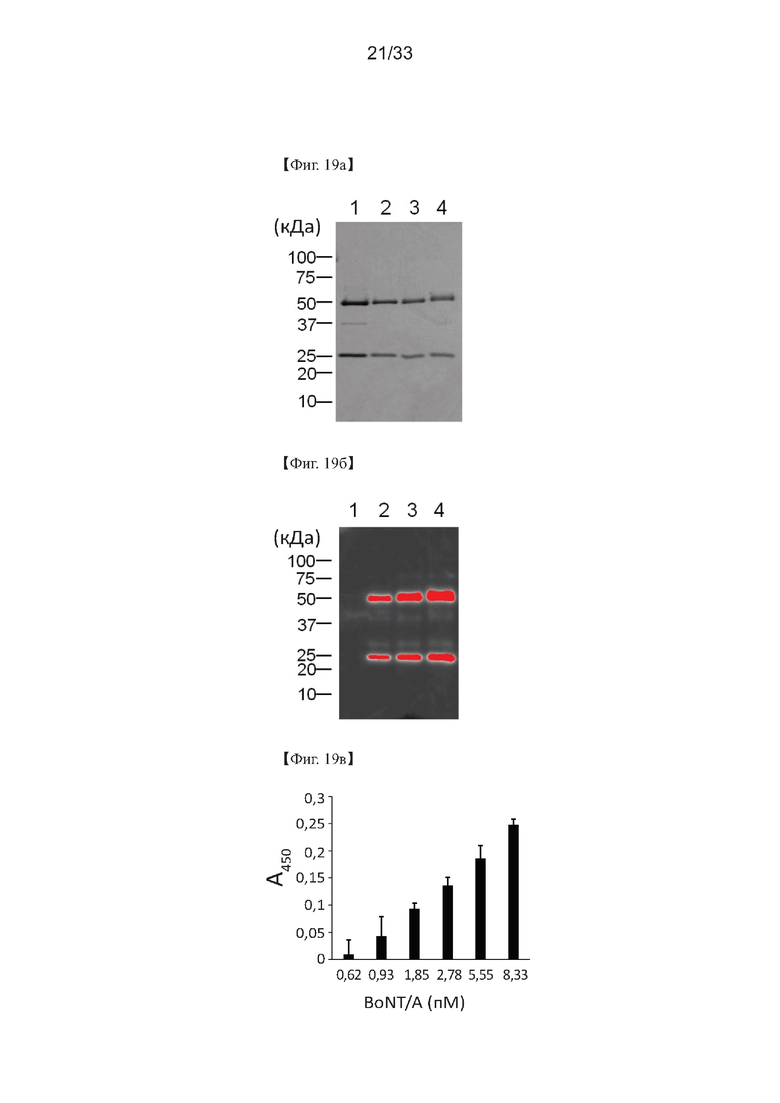
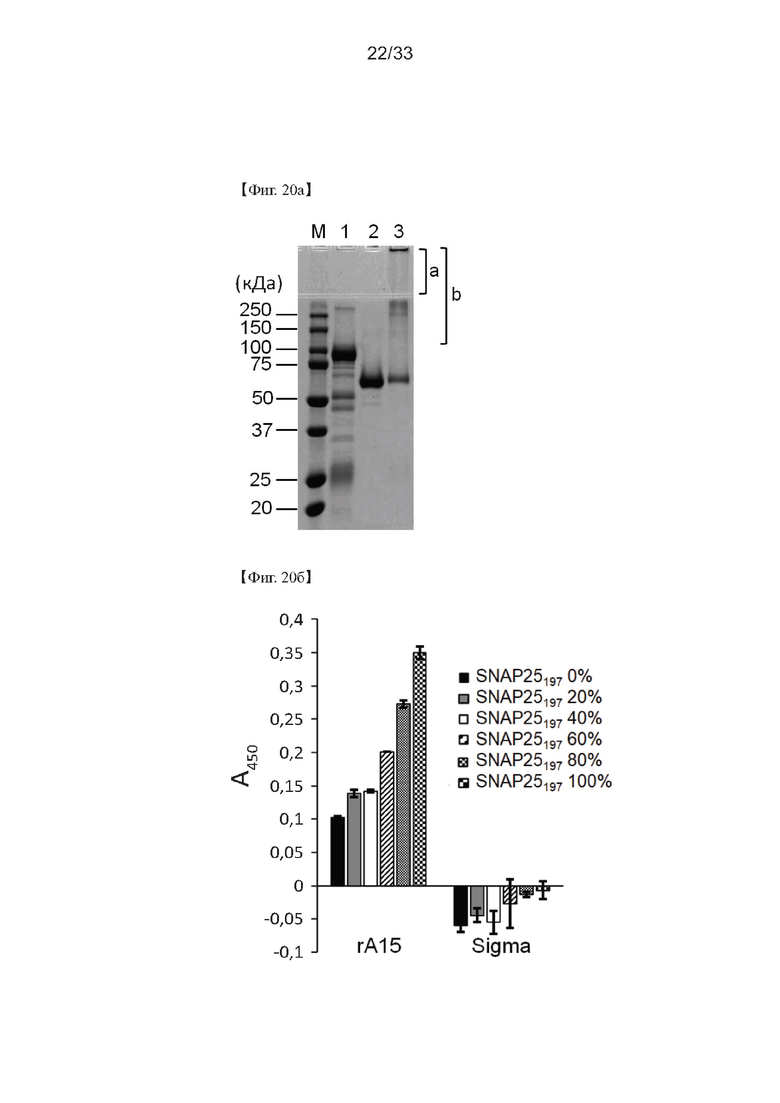
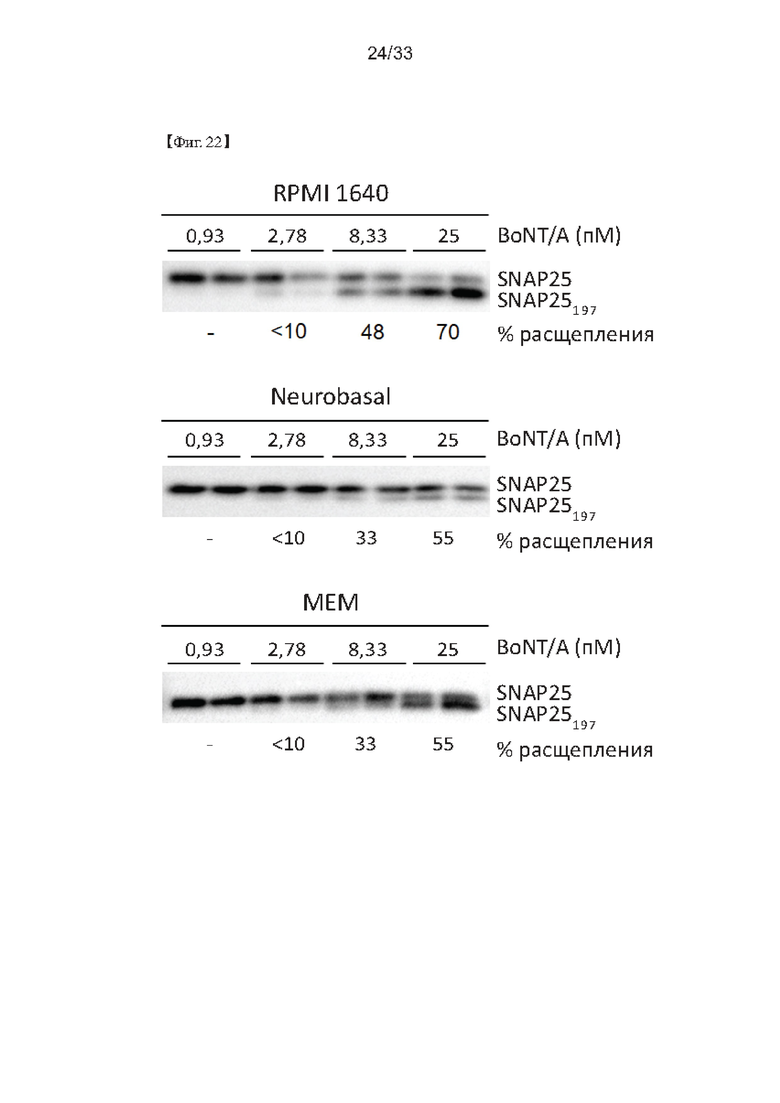
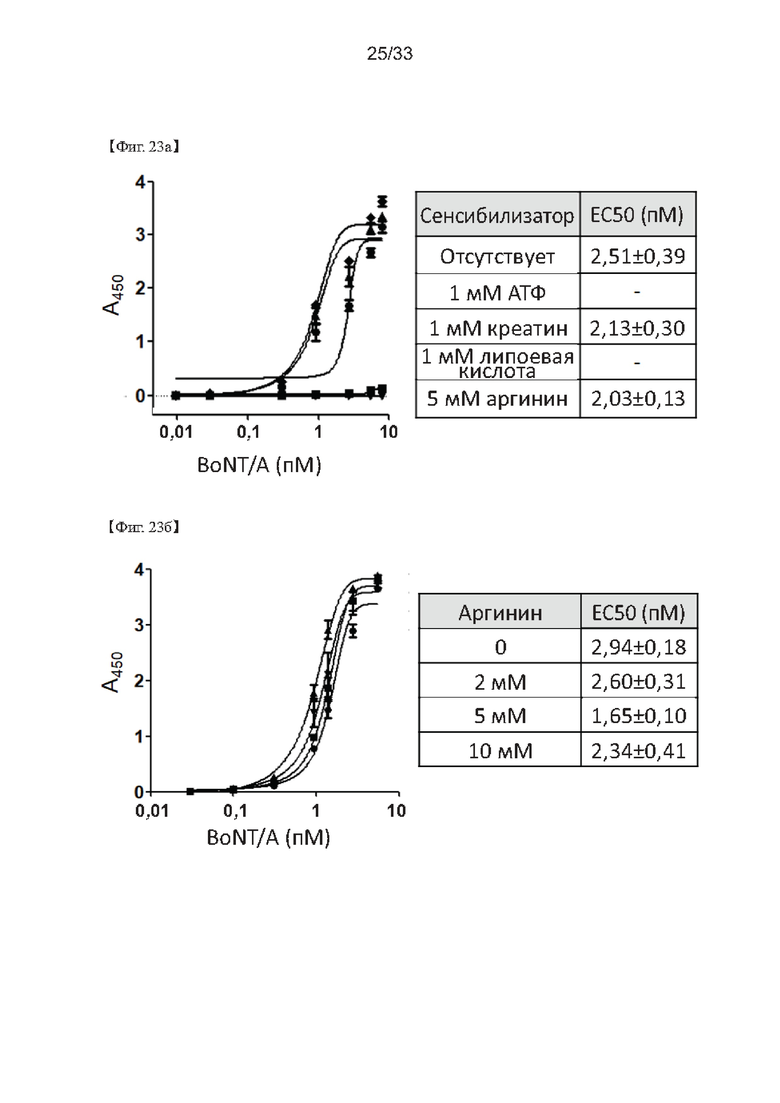
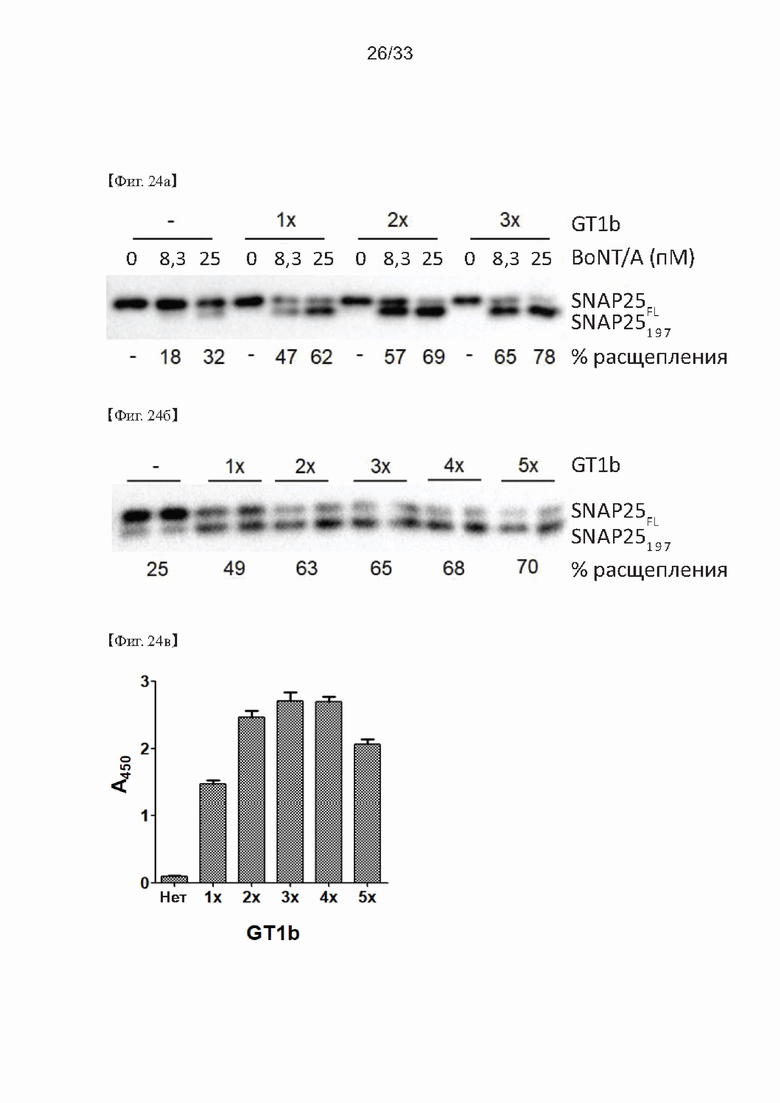
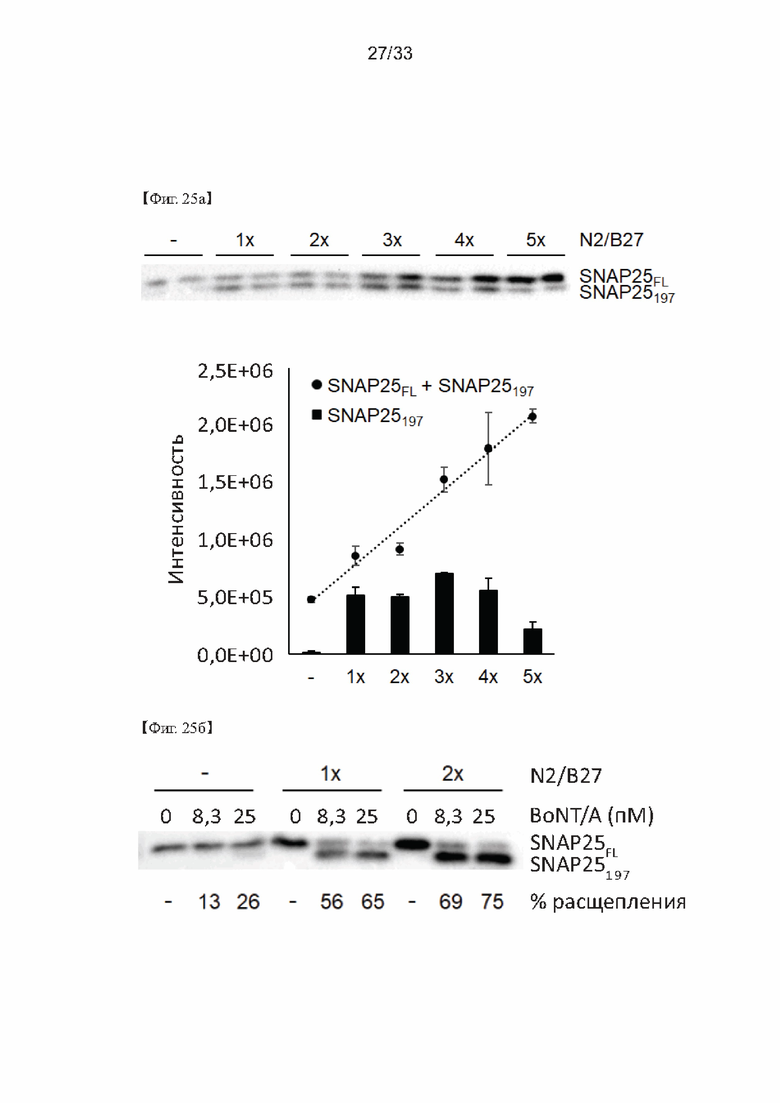
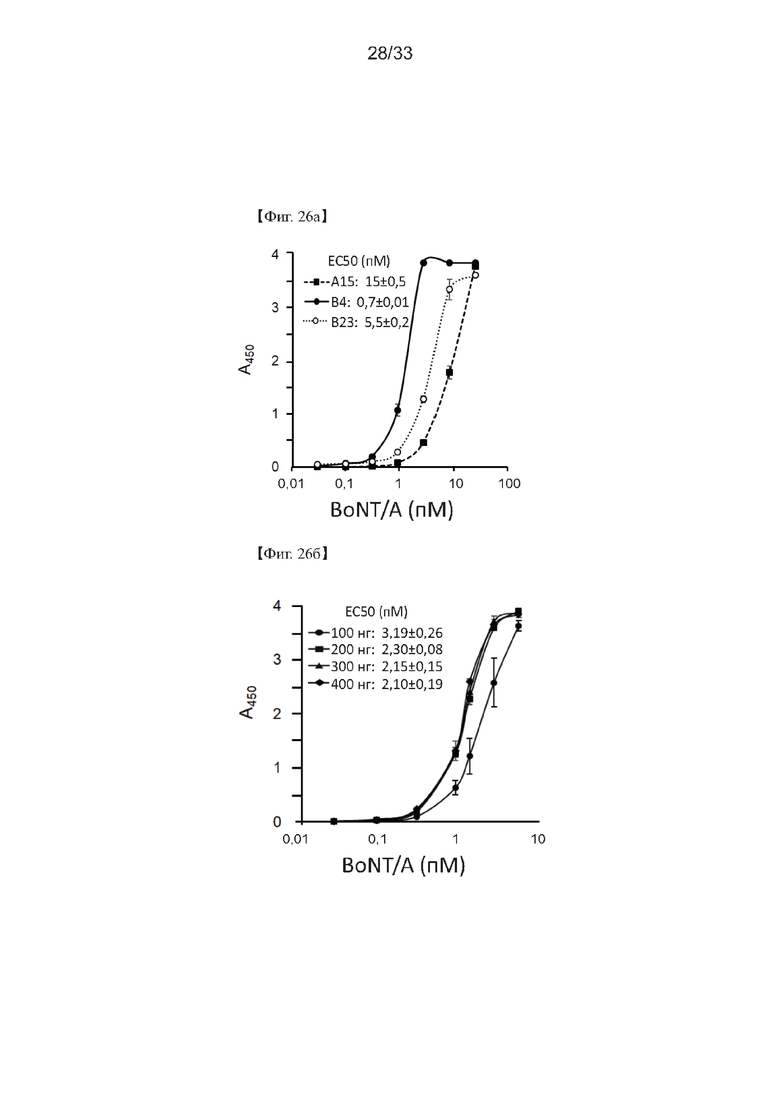
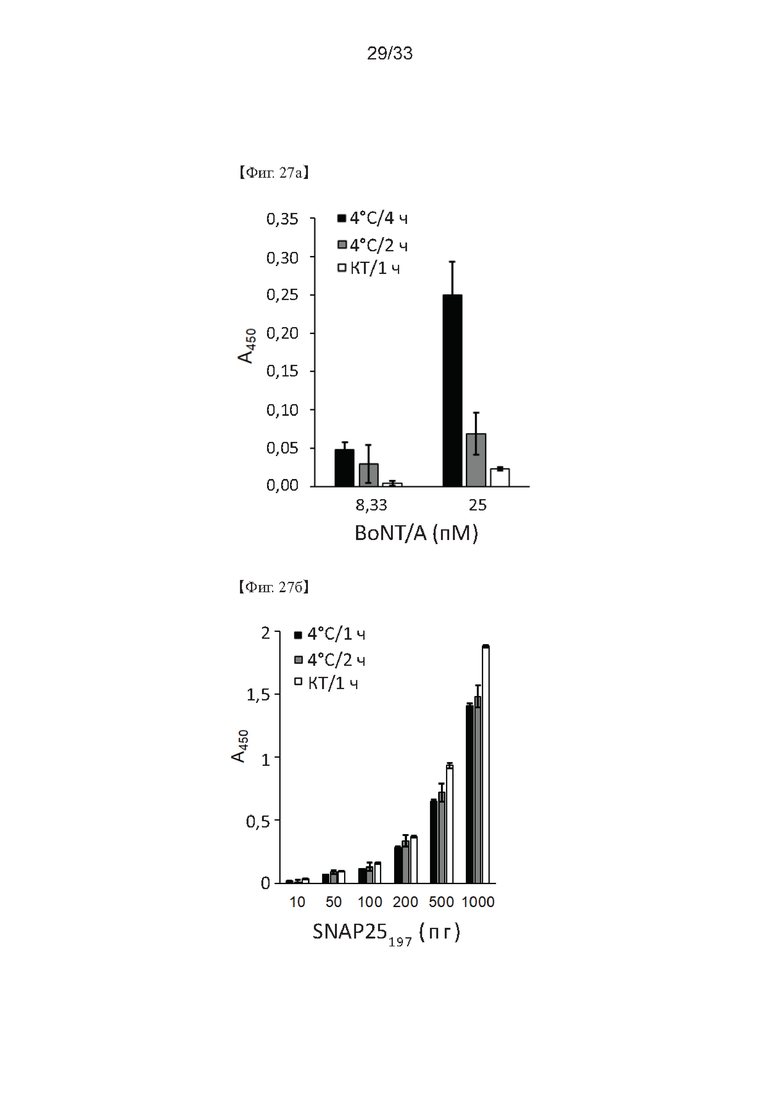
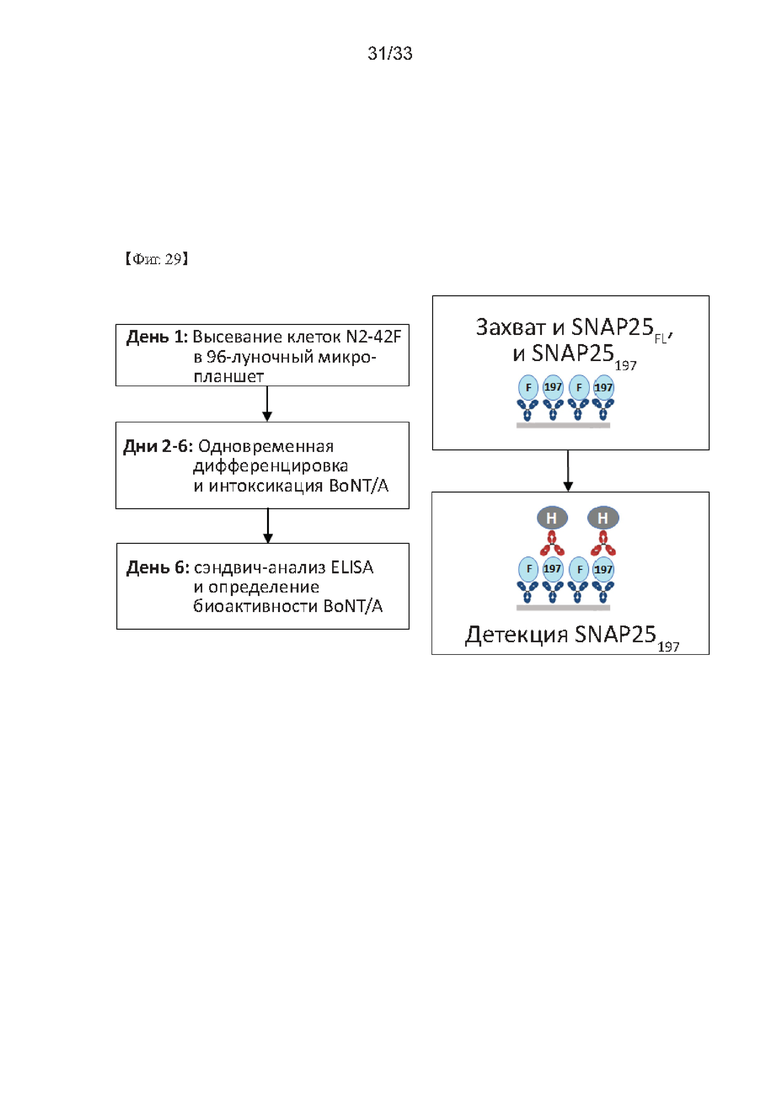

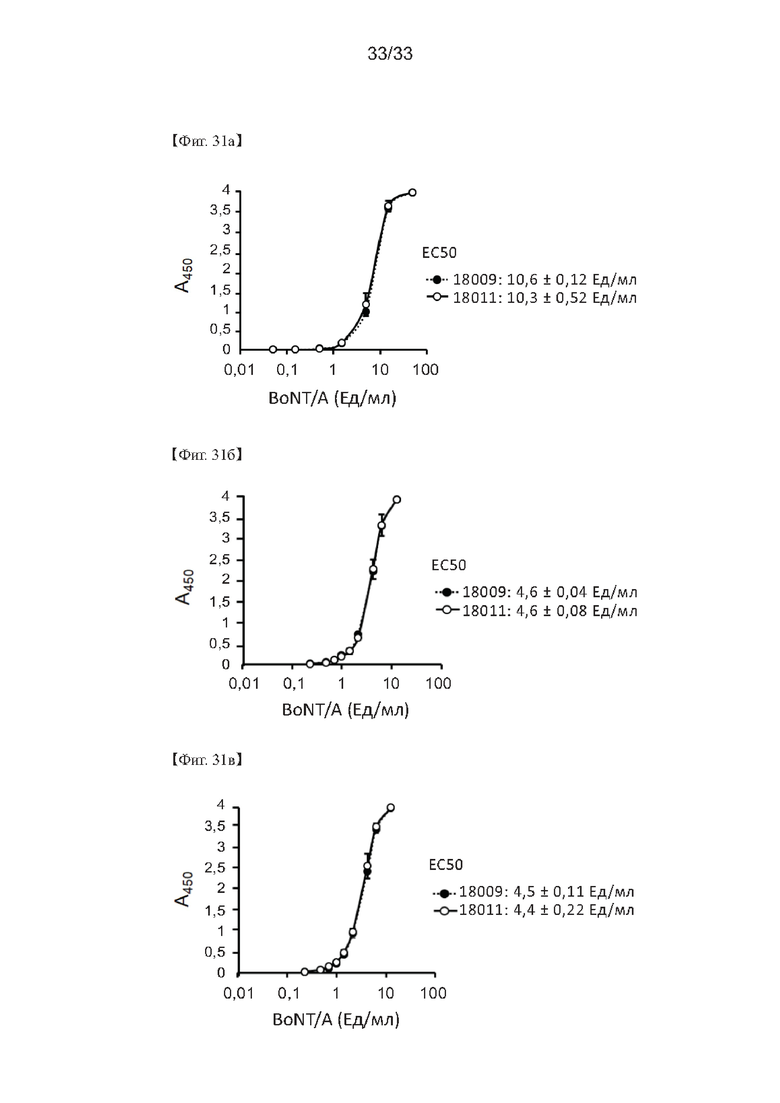
Группа изобретений относится к биотехнологии. Представлены: антитела, которые специфически связываются с SNAP25FL, композиция антител и набор для определения активности ботулинического токсина, а также клетка гибридомы для производства антитела по любому из пунктов 1-3, представляющая собой гибрид клетки селезенки и миеломной клетки мыши. Кроме того, представлены способ определения активности ботулотоксина и способ обнаружения ботулотоксина. Настоящее изобретение позволяет получить новое антитело, имеющее значительно более высокую аффинность связывания и специфичность в отношении SNAP25FL. Таким образом, ожидается, что указанное антитело будет активно использоваться в фармацевтической и косметической сферах. 8 н.п. ф-лы, 31 ил., 19 табл., 18 пр.
1. Антитело, которое специфически связывается с SNAP25FL и которое выбирается из группы, содержащей следующие (а) - (с):
(a) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 11,
участок CDR2 тяжелой цепи, состоящий SEQ ID NO: 14,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 17,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 20,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 23,
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 25;
(b) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 12,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 15,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 18,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 21,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 24, и
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 26;
(c) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 13,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 16,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 19,
участок CDR1 легкой цепи состоящий из SEQ ID NO: 22,
участок CDR2 легкой цепи состоящий из SEQ ID NO: 24; и
участок CDR3 легкой цепи состоящий из SEQ ID NO: 27.
2. Антитело, которое специфически связывается с SNAP25197 и которое выбирается из группы, содержащей следующие (а) - (g):
(а) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 28,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 34,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 40,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 47,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50,
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52;
(b) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 29,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 35,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 41,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 48,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50,
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52;
(c) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 29,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 36,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 42,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 47,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50,
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52;
(d) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 30,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 37,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 44,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 48,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50, и
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52;
(e) Антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из region consisting of SEQ ID NO: 31,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 38,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 45,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 47,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50, и
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 53;
(f) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 32,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 39,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 46,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 49,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 51, и
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 54;
(g) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 33,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 35,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 43,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 48,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 50, и
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 52.
3. Антитело, которое специфически связывается с SNAP25FL и SNAP25197 и которое выбирается из группы, содержащей следующие (а) и (b):
(a) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO:55,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 57,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 59,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 61,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 63, и
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 65;
(b) антитело, содержащее:
участок CDR1 тяжелой цепи, состоящий из SEQ ID NO: 56,
участок CDR2 тяжелой цепи, состоящий из SEQ ID NO: 58,
участок CDR3 тяжелой цепи, состоящий из SEQ ID NO: 60,
участок CDR1 легкой цепи, состоящий из SEQ ID NO: 62,
участок CDR2 легкой цепи, состоящий из SEQ ID NO: 64,
участок CDR3 легкой цепи, состоящий из SEQ ID NO: 66.
4. Композиция антител для определения активности ботулинического токсина, содержащая антитело по любому из пп. 1-3.
5. Набор для определения активности ботулинического токсина, содержащий антитело по любому из пп. 1-3 или композицию антител по п. 4.
6. Клетка гибридомы для производства антитела по любому из пп. 1-3, представляющая собой гибрид клетки селезенки и миеломной клетки мыши, содержащая нуклеиновую кислоту, кодирующую антитело по любому из пп. 1-3.
7. Способ определения активности ботулотоксина, включающий стадии:
(а) обработка нейрональной клетки указанным ботулотоксином;
b) измерение содержания SNAP25FL или SNAP25197 в нейрональной клетке с помощью антитела по любому из пп. 1-3.
8. Способ обнаружения ботулотоксина, включающий следующие стадии:
a) обработка нейрональной клетки исследуемым образцом;
b) измерение содержания SNAP25197 в нейрональной клетке с помощью антитела по любому из пп. 2, 3;
c) определение того, что если содержание комплекса антитела SNAP25197 обнаруживается, ботулотоксин присутствует в указанном образце.
| FERNANDEZ-SALAS E | |||
| et al., Botulinum neurotoxin serotype A specific cell-based potency assay to replace the mouse bioassay, PLoS One, 2012, 7(11) | |||
| US 2014248644 A1, 04.09.2014 | |||
| НИКИТИНА А.В | |||
| и др., Иммуночипы для одновременного обнаружения пяти ботулотоксинов методом фосфоресцентного анализа (ФОСФАН), Проблемы особо опасных инфекций, 2016, No | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
