ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет по предварительной заявке на патент США № 62/698459, поданной 16 июля 2018 г., и предварительной заявке на патент США № 62/867477, поданной 27 июня 2019 г., полное содержание обеих из которых включено в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[0002] Настоящее изобретение относится к генетически модифицированным животным отряда грызуны и моделям заболеваний человека на грызунах. Более конкретно, настоящее изобретение относится к генетически модифицированным грызунам, геном которых содержит гуманизированный ген Il1rl2 (кодирующий субъединицу IL1rl2 IL-36R) и человеческие гены лигандов IL-36 α, β и γ. Описанные в данном документе генетически модифицированные грызуны демонстрируют усиленное воспаление кожи и кишечника в доклинической модели псориаза и воспалительного заболевания кишечника (IBD) соответственно и могут служить в качестве модели заболевания, проявляющегося дефицитом антагониста рецептора интерлейкина-36 (DITRA) человека на грызунах.
ВКЛЮЧЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ПОСРЕДСТВОМ ССЫЛКИ
[0003] Перечень последовательностей в виде текстового файла с кодировкой ASCII под названием 35950_10404US01_SequenceListing.txt, размером 65 кб, созданный 9 июля 2019 г. и поданный в Ведомство по патентам и товарным знакам США через EFS-Web, включен в данный документ посредством ссылки во всей своей полноте.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[0004] Подсемейство интерлейкина (IL)-36 состоит из трех агонистов (IL-36α, β и γ, ранее обозначенных как IL1F6, IL1F8 и IL1F9 соответственно), и одного антагониста (IL36Ra), которые передают сигнал через гетеродимерный рецептор (IL-36R), что приводит к активации ядерного фактораκB (NFκB) и митоген-активируемых протеинкиназ (MAPK). Подобно классическим представителям семейства IL-1, цитокины IL-36 участвуют в инициации и регуляции иммунных ответов. Впервые было продемонстрировано, что представители семейства IL-36 преимущественно экспрессируются в тканях плоского эпителия и, в частности, в псориатических поражениях кожи. Связь между IL-36R и эпителиально опосредованными заболеваниями была подтверждена у людей на основании мутаций с утратой функции в IL-36Ra, что привело к постановке диагноза генерализованного пустулезного псориаза (сокращенно «GPP»), который теперь считается разновидностью GPP, называемой дефицитом антагониста рецептора интерлейкина-36 (сокращенно «DITRA»). О повышенной экспрессии IL-36 сообщалось при GPP, ладонно-подошвенном пустулезном псориазе (PPPP), воспалительном заболевании кишечника (IBD), ревматоидном и псориатическом артрите, астме, хроническом обструктивном заболевании легких, хроническом заболевании почек и ихтиозе.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0005] В данном документе раскрыты некоторые варианты осуществления грызунов, генетически модифицированных таким образом, что их геном содержит гуманизированный ген Il1rl2, кодирующий белок Il1rl2, имеющий эктодомен, практически идентичный человеческому эктодомену IL1RL2, а также человеческие гены Il1f6, Il1f8 и Il1f9 , кодирующие человеческие IL1F6, IL1F8 и IL1F9 соответственно. Il1rl2 представляет собой субъединицу гетеродимерного белка IL-36R, который связывается с тремя агонистами: IL1F6, IL1F8 и IL1F9 (также известными как IL-36α, β и γ соответственно). Раскрытые в данном документе грызуны, хотя и содержат гуманизированный ген Il1rl2 и экспрессируют три человеческих гена лигандов-агонистов, сохраняют эндогенный антагонист IL-36Ra, где эндогенный IL-36Ra приблизительно в 20 раз менее активен при подавлении передачи сигнала человеческого IL-36R. Такие четырехкратно гуманизированные грызуны (например, с гуманизированным Il1rl2 и человеческими IL1F6, IL1F8 и IL1F9), как было показано в данном документе, проявляют симптомы пациентов с DITRA, у которых мутация в IL-36Ra приводит к усилению передачи сигнала IL-36R. Соответственно, в данном документе предусмотрены генетически модифицированные грызуны, композиции и способы получения таких грызунов, а также способы применения таких грызунов в качестве модели для скрининга и разработки терапевтических средств.
[0006] В одном аспекте некоторых вариантов осуществления в настоящем документе раскрыт генетически модифицированный грызун, геном которого содержит (1) гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, практически идентичный эктодомену человеческого белка IL1RL2; (2) человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна; (3) человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна; и (4) человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна.
[0007] В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна кодирует гуманизированный белок Il1rl2, который содержит трансмембранно-цитоплазматическую последовательность, практически идентичную трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна. В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна кодирует гуманизированный белок Il1rl2, который содержит сигнальный пептид, практически идентичный сигнальному пептиду эндогенного белка Il1rl2 грызуна.
[0008] В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна кодирует гуманизированный белок Il1rl2, содержащий эктодомен, практически идентичный эктодомену человеческого белка IL1RL2 , где человеческий белок IL1RL2 содержит аминокислотную последовательность, представленную под SEQ ID NO: 2.
[0009] В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна кодирует гуманизированный белок Il1rl2, содержащий эктодомен, где эктодомен содержит аминокислоты 22-337 из SEQ ID NO: 7.
[0010] В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна кодирует гуманизированный белок Il1rl2, который содержит аминокислотную последовательность, представленную под SEQ ID NO: 7.
[0011] В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна функционально связан с эндогенным промотором Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна.
[0012] В некоторых вариантах осуществления гуманизированный ген Il1rl2 у грызуна является результатом замены геномного фрагмента эндогенного гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна на нуклеотидную последовательность человеческого гена IL1RL2.
[0013] В некоторых вариантах осуществления нуклеотидная последовательность человеческого гена IL1RL2 представляет собой геномный фрагмент человеческого гена IL1RL2, который кодирует по существу эктодомен человеческого белка IL1RL2. В некоторых вариантах осуществления геномный фрагмент человеческого гена IL1RL2 содержит экзоны 3-8 человеческого гена IL1RL2.
[0014] В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены для гуманизации, содержит экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна. В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены для гуманизации, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна.
[0015] В конкретных вариантах осуществления гуманизированный ген Il1rl2 у грызуна, раскрытый в данном документе, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна, экзоны 3-8 человеческого гена IL1RL2 и оставшиеся экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
[0016] В некоторых вариантах осуществления человеческий ген IL1F6 у грызуна, раскрытый в данном документе, заменяет эндогенный ген Il1f6 грызуна в эндогенном локусе Il1f6 грызуна.
[0017] В некоторых вариантах осуществления человеческий ген IL1F8 у грызуна, раскрытый в данном документе, заменяет эндогенный ген Il1f8 грызуна в эндогенном локусе Il1f8 грызуна.
[0018] В некоторых вариантах осуществления человеческий ген IL1F9 у грызуна, раскрытый в данном документе, заменяет эндогенный ген Il1f9 грызуна в эндогенном локусе Il1f9 грызуна.
[0019] В некоторых вариантах осуществления раскрытый в данном документе грызун является гомозиготным по одному или более из четырех гуманизированных и/или человеческих генов. В некоторых вариантах осуществления раскрытый в данном документе грызун является гомозиготным по каждому из четырех гуманизированных и/или человеческих генов.
[0020] В некоторых вариантах осуществления раскрытый в данном документе грызун характеризуется укороченной толстой кишкой по сравнению с грызуном дикого типа. В некоторых вариантах осуществления раскрытый в данном документе грызун демонстрирует усиленное воспаление в модели экспериментально индуцированного воспаления по сравнению с грызуном дикого типа. В некоторых вариантах осуществления модель экспериментально индуцированного воспаления представляет собой модель воспаления кожи, индуцированного имиквимодом (IMQ). В некоторых вариантах осуществления модель экспериментально индуцированного воспаления представляет собой модель воспаления кишечника, индуцированного декстрансульфатом натрия (DSS) или оксазолоном.
[0021] В некоторых вариантах осуществления раскрытый в данном документе грызун представляет собой мышь или крысу.
[0022] Дополнительно в данном документе раскрыта выделенная клетка или ткань грызуна, описанного в данном документе.
[0023] В дополнительном аспекте некоторых вариантов осуществления в настоящем документе раскрыт способ получения генетически модифицированного грызуна, причем способ включает модификацию генома грызуна таким образом, чтобы он содержал (1) гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, практически идентичный эктодомену человеческого белка IL1RL2; (2) человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна; (3) человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна; и (4) человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна; и получение грызуна, содержащего модифицированный геном.
[0024] В некоторых вариантах осуществления геном грызуна модифицируют с помощью способа, включающего (i) получение первого грызуна, содержащего гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна; (ii) получение второго грызуна, содержащего человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна, человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна, и человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна; и (iii) скрещивание первого грызуна со вторым грызуном с получением модифицированного генома грызуна.
[0025] В некоторых вариантах осуществления грызун, содержащий гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна, получен посредством получения эмбриональной стволовой (ES) клетки грызуна, вставки нуклеотидной последовательности человеческого гена IL1RL2 в локус Il1rl2 грызуна ES-клетки грызуна с образованием гуманизированного гена Il1rl2 в локусе Il1rl2 грызуна, с получением таким образом ES-клетки грызуна, содержащей гуманизированный ген Il1rl2, и получением грызуна с применением ES-клетки грызуна, содержащей гуманизированный ген Il1rl2.
[0026] В некоторых вариантах осуществления нуклеотидная последовательность вставляемого человеческого гена IL1RL2 заменяет геномный фрагмент гена Il1rl2 грызуна в локусе Il1rl2 грызуна. В некоторых вариантах осуществления нуклеотидная последовательность человеческого гена IL1RL2 представляет собой геномный фрагмент человеческого гена IL1RL2 , который кодирует по существу эктодомен человеческого белка IL1RL2. В некоторых вариантах осуществления геномный фрагмент человеческого гена IL1RL2 содержит экзоны 3-8 человеческого гена IL1RL2. В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены, содержит экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна. В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены для гуманизации, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна. В некоторых вариантах осуществления гуманизированный ген Il1rl2, образованный в результате замены для гуманизации, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна, экзоны 3-8 человеческого гена IL1RL2 и оставшиеся экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
[0027] В некоторых вариантах осуществления грызун, содержащий человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9, получен посредством: получения эмбриональной стволовой (ES) клетки грызуна; вставки человеческого гена IL1F6 в локус Il1f6 грызуна ES-клетки грызуна, человеческого гена IL1F8 в локус Il1f8 грызуна ES-клетки грызуна и человеческого гена IL1F9 в локус Il1f9 грызуна ES-клетки грызуна, с получением таким образом ES-клетки грызуна, содержащей человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9; и получения грызуна с использованием ES-клетки грызуна, содержащей человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9.
[0028] В некоторых вариантах осуществления человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9 представлены в непрерывной молекуле нуклеиновой кислоты.
[0029] В некоторых вариантах осуществления способов человеческий ген IL1F6 заменяет эндогенный ген Il1f6 грызуна в эндогенном локусе Il1f6 грызуна.
[0030] В некоторых вариантах осуществления способов человеческий ген IL1F8 заменяет эндогенный ген Il1f8 грызуна в эндогенном локусе Il1f8 грызуна.
[0031] В некоторых вариантах осуществления способов человеческий ген IL1F9 заменяет эндогенный ген Il1f9 грызуна в эндогенном локусе Il1f9 грызуна.
[0032] В другом аспекте некоторых вариантов осуществления в данном документе раскрыта эмбриональная стволовая (ES) клетка грызуна, содержащая (1) гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, практически идентичный эктодомену человеческого белка IL1RL2; (2) человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна; (3) человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна; и (4) человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна. В некоторых вариантах осуществления такую ES-клетку можно получить с помощью способов, раскрытых в данном документе. В некоторых вариантах осуществления также раскрыто применение такой ES-клетки для получения грызуна.
[0033] В еще одном аспекте некоторых вариантов осуществления в данном документе раскрыты нацеливающиеся конструкции нуклеиновой кислоты, пригодные для модификации генома грызуна (например, в ES-клетке грызуна) для получения модифицированного грызуна.
[0034] В некоторых вариантах осуществления в данном документе раскрыта нацеливающаяся конструкция нуклеиновой кислоты, содержащая нуклеотидную последовательность человеческого гена IL1RL2, фланкированную 5'- и 3'-нуклеотидными последовательностями грызуна, способными опосредовать гомологичную рекомбинацию и интеграцию нуклеотидной последовательности человеческого гена IL1RL2 в локус Il1rl2 грызуна с образованием гуманизированного гена Il1rl2, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, практически идентичный эктодомену человеческого белка IL1RL2.
[0035] В некоторых вариантах осуществления в данном документе раскрыта нацеливающаяся конструкция нуклеиновой кислоты, содержащая непрерывную последовательность нуклеиновой кислоты, которая содержит: человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9, где непрерывная последовательность нуклеиновой кислоты фланкирована 5'- и 3'-нуклеотидными последовательностями грызуна, способными опосредовать гомологичную рекомбинацию и интеграцию непрерывной последовательности нуклеиновой кислоты в локус грызуна, охватывающий ген Il1f6 грызуна, ген Il1f8 грызуна и ген Il1f9 грызуна.
[0036] В дополнительном аспекте некоторых вариантов осуществления в данном документе раскрыто применение раскрытого в данном документе грызуна в качестве модели заболеваний человека на грызунах, ассоциированных с нарушением регуляции передачи сигнала IL-36. В качестве неограничивающих примеров вариантов осуществления, раскрытых в данном документе, грызуна можно использовать для изучения патологии и молекулярных основ заболеваний человека, ассоциированных с нарушением регуляции передачи сигнала IL-36 (таких как без ограничения DITRA), или для скрининга, тестирования и разработки терапевтических соединений, пригодных для лечения таких заболеваний.
[0037] В еще одном дополнительном аспекте некоторых вариантов осуществления в настоящем документе раскрыт способ оценки терапевтической эффективности соединения-кандидата для лечения заболевания, ассоциированного с нарушением регуляции передачи сигнала IL-36, включающий введение грызуну, раскрытому в данном документе, средства для индукции воспаления, введение грызуну соединения-кандидата и определение того, подавляет ли и/или снижает ли соединение-кандидат индуцированное воспаление.
[0038] В некоторых вариантах осуществления средство, вводимое для индукции воспаления, представляет собой DSS или оксазолон, который индуцирует воспаление кишечника у грызуна. В некоторых вариантах осуществления средство, вводимое для индукции воспаления, представляет собой IMQ, который индуцирует воспаление кожи у грызуна.
[0039] В некоторых вариантах осуществления соединение-кандидат вводят грызуну до, во время или после введения средства, которое индуцирует воспаление. В некоторых вариантах осуществления соединение-кандидат может представлять собой низкомолекулярное соединение, ингибитор на основе нуклеиновой кислоты или антигенсвязывающий белок, такой как антитело. В некоторых вариантах осуществления соединение-кандидат представляет собой антитело, которое специфически связывает IL-36R человека.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0040] Файл данного патента содержит по меньшей мере один графический материал, выполненный в цвете. Копии данного патента с цветным(-и) графическим(-и) материалом(-ами) будут предоставлены Ведомством по патентам и товарным знакам после запроса и уплаты необходимой пошлины.
[0041] На фиг. 1A-1D показаны варианты осуществления иллюстративной стратегии гуманизации мышиного Il1rl2. На фигуре 1A без соблюдения масштаба показана диаграмма, демонстрирующая геномную организацию мышиных и человеческих генов Il1rl2 и IL1RL2. Экзоны представлены тонкими полосами, расположенными поперек геномных последовательностей. Указаны мышиный геномный фрагмент длиной приблизительно 25324 пары оснований (п.о.), подлежащий удалению, и вставляемый человеческий геномный фрагмент длиной приблизительно 32389 п.о. Указаны местоположения зондов, использованных в анализе, описанном в примере 1. На фигуре 1B без соблюдения масштаба показан иллюстративный модифицированный вектор ВАС для гуманизации эндогенного мышиного гена Il1rl2 вместе с соединительными последовательностями внизу (SEQ ID NOS: 26-28) для осуществления (i) замены мышиных экзонов 3-8 и промежуточных интронов Il1rl2, вместе с 155 п.о. на 3'-конце мышиного интрона 2 и 642 п.о. на 5'-конце мышиного интрона 8, на человеческие экзоны 3-8 и промежуточные интроны IL1RL2, а также с 346 п.о. на 3'-конце человеческого интрона 2 и 1101 п.о. на 5'-конце человеческого интрона 8, и (ii) вставки кассеты loxP-hUb1-em7-Neo-pA-mPrm1-Crei-loxP (4996 п.о.) ниже вставки человеческого геномного фрагмента в интрон 8. На фигуре 1C проиллюстрирован без соблюдения масштаба гуманизированный аллель Il1rl2 после делеции кассеты устойчивости к неомицину, вместе с соединительными последовательностями внизу (SEQ ID NOS: 26 и 29). На фигуре 1D представлено выравнивание последовательностей мышиного белка Il1rl2 (SEQ ID NO: 4), человеческого белка IL1RL2 (SEQ ID NO: 2) и гуманизированного белка Il1rl2 (SEQ ID NO: 7).
[0042] На фиг. 2A-2D показаны варианты осуществления иллюстративной стратегии замены мышиных Il1f8, Il1f9 и Il1f6 на человеческие IL1F8, IL1F9 и IL1F6. На фигуре 2A без соблюдения масштаба показана диаграмма, демонстрирующая геномную организацию мышиных генов Il1f8, Il1f9 и Il1f6 и человеческих генов IL1F8, IL1F9 и IL1F6. Экзоны представлены тонкими полосами, расположенными поперек геномных последовательностей. Указаны мышиный геномный фрагмент длиной приблизительно 76548 п.о., подлежащий удалению, и вставляемый человеческий геномный фрагмент длиной приблизительно 88868 п.о. Указаны местоположения зондов, использованных в анализе, описанном в примере 1. На фигуре 2B без соблюдения масштаба показан иллюстративный модифицированный вектор BAC для замены эндогенных мышиных генов Il1f8, Il1f9 и Il1f6 вместе с соединительными последовательностями (SEQ ID NOS: 30-32) внизу, для осуществления (i) замены кодирующих последовательностей и нетранслируемых областей (UTR), а также межгенной последовательности мышиных Il1f8, Il1f9 и Il1f6 на соответствующую человеческую последовательность IL1F8, IL1F9 и IL1F6; и (ii) вставки кассеты loxP-hUb1-em7-Hygro-pA-mPrm1-Crei-loxP (5218 п.о.) ниже вставки человеческого геномного фрагмента. На фиг. 2C проиллюстрирован без соблюдения масштаба гуманизированный локус после делеции кассеты устойчивости к гигромицину, вместе с соединительными последовательностями внизу (SEQ ID NOS: 30 и 33). На фигуре 2D представлены выравнивания последовательностей мышиных белков Il1f6 (SEQ ID NO: 11) и человеческих IL1F6 (SEQ ID NO: 9); мышиных белков Il1f8 (SEQ ID NO: 17) и человеческих IL1F8 (SEQ ID NO: 15); и мышиных белков Il1f9 (SEQ ID NO: 23) и человеческих IL1F9 (SEQ ID NO: 21).
[0043] На фиг. 3A-3E показаны варианты осуществления настоящего изобретения, демонстрирующие усиление воспаления кожи, индуцированного IMQ, у гуманизированных DITRA-подобных мышей. DITRA-подобных (также сокращенно обозначенных «DITRA» на фигурах в данном документе) мышей и мышей дикого типа (WT) обрабатывали посредством местного нанесения IMQ в течение последовательных 4 дней. На 5 день собирали образцы кожи спины для последующей гистопатологической оценки и qRT-PCR. (3A) Иллюстративный внешний вид кожи DITRA-подобных мышей и мышей WT на 5 день, местно обработанных вазелином («Vas», контроль) или IMQ-содержащим кремом (Aldara) в течение 4 последовательных дней. (3B) Иллюстративное окрашивание гематоксилином и эозином («H&E») обработанной вазелином и IMQ кожи DITRA-подобных мышей и мышей WT. (3C) Экспрессия мРНК провоспалительных молекул в коже мышей DITRA и WT, которых ежедневно обрабатывали вазелином или IMQ-содержащим кремом (n=10 на каждую группу). После местного нанесения IMQ в течение 4 последовательных дней на 5 день образцы кожи спины собирали от гуманизированных одиночных мышей Il1rl2 ("1H"), DITRA-подобных мышей и мышей дикого типа для последующей гистопатологической оценки и qRT-PCR. В каждой группе столбцов слева направо показаны: WT Vas, DITRA Vas, WT IMQ и DITRA IMQ. (3D) Иллюстративный внешний вид кожи мышей 1H, DITRA-подобных мышей и мышей WT на день 5, местно обработанных вазелином (контроль) или IMQ-содержащим кремом (Aldara) в течение 4 последовательных дней. (3E) Экспрессия мРНК провоспалительных молекул в коже мышей 1H, DITRA и WT, которых ежедневно обрабатывали вазелином или IMQ-содержащим кремом (n=10 на каждую группу). В каждой группе столбцов слева направо показаны: WT Vas, 1H Vas, DITRA-ПОДОБНЫЕ Vas, WT IMQ, 1H IMQ и DITRA-ПОДОБНЫЕ IMQ.
[0044] Фиг. 4A-4B.В некоторых вариантах осуществления DITRA-подобные мыши демонстрируют укороченную толстую кишку в устойчивом состоянии. (4A) Иллюстративные изображения толстой кишки, полученной от DITRA-подобных мышей и их совместно содержащихся однопометников IL-36R KO и WT через 3 и 10 месяцев. (4B) Количественная оценка длины толстой кишки DITRA-подобных мышей в возрасте 3 и 10 месяцев по сравнению с их совместно содержащимися однопометниками IL-36R KO и WT (n=6 на каждую группу). Планки погрешностей представляют среднее±SD.
[0045] Фиг. 5A-5E.В некоторых вариантах осуществления DITRA-подобные мыши демонстрируют дефект заживления слизистой оболочки при хроническом колите, индуцированном DSS. DITRA-подобных мышей (n=16) и их контрольных совместно содержащихся однопометников WT (n=14) подвергали хроническому DSS-индуцированному колиту посредством введения 2,5% DSS в течение 7 дней с последующим введением воды в течение 11 дней и 1,5% DSS в течение 4 дней с последующим введением воды в течение 5 дней, всего в течение 27 дней. Контрольные мыши (n=5) получали обычную воду. (5A) Потеря веса тела у DITRA-подобных мышей и их контрольных совместно содержащихся однопометников WT, рассчитанная как процентная разница между исходным и фактическим весом тела в любой конкретный день. (5B) Длина толстой кишки у обработанных водой и DSS DITRA-подобных мышей и мышей WT. (5C) Окрашивание гематоксилином и эозином (H&E) и оценка патологии толстой кишки у обработанных водой и DSS DITRA-подобных мышей и мышей WT. (5D) Активность миелопероксидазы (MPO) в гомогенатах толстой кишки у обработанных водой и DSS DITRA-подобных мышей и мышей WT. (5E) Уровни провоспалительных цитокинов в гомогенатах толстой кишки у обработанных водой и DSS DITRA-подобных мышей и мышей WT. Данные являются репрезентативными для по меньшей мере трех независимых экспериментов. Планки погрешностей представляют среднее±SD. В каждой группе столбцов слева направо показаны: WT/вода, DITRA/вода, WT/DSS и DITRA/DSS.
[0046] Фиг. 6A-6B.В некоторых вариантах осуществления антагонизм человеческого IL-36R с помощью антитела REGN к человеческому IL-36R облегчает IMQ-индуцированное воспаление кожи у DITRA-подобных мышей при профилактическом введении доз. Кожу на спине DITRA-подобных мышей брили за 3 дня до начала применения IMQ, и кожу местно обрабатывали вазелином (контроль) или IMQ-содержащим кремом (Aldara) в течение 4 последовательных дней. PBS, αmAb к hIL-36R и изотипический контроль hIgG4 вводили подкожно путем инъекции при дозе 10 мг/мл в дни -3 и 1 (n=9 для каждой группы обработки). (6A) Гистологические срезы, окрашенные H&E, и оценка патологии кожи спины, полученной от DITRA-подобных мышей в день 5. (6B) Уровни провоспалительных цитокинов в гомогенатах кожи DITRA-подобных мышей в день 5, которых ежедневно обрабатывали посредством нанесений IMQ и инъекций PBS, αmAb к IL-36R и изотипического контроля в дни -3 и 1. Данные являются репрезентативными для трех независимых экспериментов. Планки погрешностей представляют среднее±SD. В каждой группе столбцов слева направо показаны: PBS, αmAb к hIL-36R и изотипический контроль hIgG4.
[0047] Фиг. 7A-7C. В некоторых вариантах осуществления терапевтическое введение антитела к человеческому IL-36R облегчает хроническое IMQ-индуцированное воспаление кожи у DITRA-подобных мышей. Кожу на спине DITRA-подобных мышей брили за 3 дня до начала введения, и кожу местно обрабатывали вазелином (контроль) или IMQ-содержащим кремом (Aldara) в течение двух циклов (5 последовательных дней с нанесением в течение первого цикла, за которыми следовали 2 дня без нанесения, затем второй цикл из 4 последовательных дней с нанесением, всего в течение 11 дней). PBS, антитело к человеческому IL-36R и изотипический контроль hIgG4 вводили подкожно путем инъекции при дозе 10 мг/мл в дни 5 и 9 (n=10 для каждой группы обработки). (7A) Толщину кожи измеряли у DITRA-подобных мышей в день 12. Толщина указана в μм. (7B) Гистологические срезы, окрашенные H&E и оценка патологии кожи DITRA-подобных мышей в день 12. (7C) Уровни провоспалительных цитокинов измеряли в гомогенатах кожи DITRA-подобных мышей в день 12. Данные являются репрезентативными для двух независимых экспериментов. Планки погрешностей представляют среднее±SD. В каждой группе столбцов слева направо показаны: забуференный фосфатом солевой раствор (PBS), αmAb к hIL-36R и изотипический контроль hIgG4.
[0048] Фиг. 8A-8E. В некоторых вариантах осуществления терапевтическое введение антитела к человеческому IL-36R, аналогично блокаде IL-12p40, облегчает DSS-индуцированное хроническое воспаление у DITRA-подобных мышей и устраняет их неспособность к заживлению DSS-индуцированного повреждения слизистой оболочки. DITRA-подобных мышей (n=45) подвергали хроническому DSS-индуцированному колиту посредством введения 3% DSS в течение 7 дней и 2% DSS в течение 13 дней с последующим введением воды всего в течение 30 дней. Контрольные мыши (n=5) получали обычную воду. PBS (n=11), αmAb к IL-36R (n=11), изотипический контроль hIgG4 (n=6), αmIL-12p40 (BioxCell) (n=11) и изотипический контроль, представляющий собой крысиный IgG2a (n=6), вводили интраперитонеально путем инъекции при дозе 10мг/мл в день 7, 10, 14, 17, 21 и 24. (8A) Потеря веса тела у DITRA-подобных мышей, рассчитанная как процентная разница между исходным и фактическим весом тела в любой конкретный день. (8B) Уровни фекального липокалина-2 (Lcn2) у DITRA-подобных мышей, измеренные в день 0, 7, 17, 23 и 30 на протяжении хронического колита. (8C) Длина толстой кишки у DITRA-подобных мышей в день 30. (8D) Активность миелопероксидазы (MPO) в гомогенатах толстой кишки DITRA-подобных мышей в день 30. (8E) Уровни провоспалительных цитокинов в гомогенатах толстой кишки у DITRA-подобных мышей в день 30. Данные являются репрезентативными для двух независимых экспериментов. Планки погрешностей представляют среднее±SD. В каждой группе столбцов слева направо показаны: PBS/вода, PBS/DSS, αhIL-36R/DSS, изотипический контроль hIgG4/DSS, αmIL-12p40/DSS и изотипический контроль, представляющий собой крысиный IgG2a/DSS.
[0049] Фиг. 9A-9C. Антагонизм человеческого IL-36R облегчает индуцированный оксазолоном колит у DITRA-подобных мышей. DITRA-подобных мышей предварительно сенсибилизировали 3% раствором оксазолона, растворенного в 100% этаноле, и интраректально вводили 1,5% оксазолон и средство-носитель (50% этанол) в дни 5, 6 и 7. Мышам интраперитонеально вводили путем инъекции PBS, mAb к человеческому IL-36R и изотипический контроль hIgG4 в дни 2, 5 и 7 после предварительной сенсибилизации. (A) Вес тела DITRA-подобных мышей, обработанных с помощью PBS, mAb к человеческому IL-36R и контроля hIgG4 во время введения оксазолона. *p<0,05 для группы, обработанной с помощью PBS. (B) Длина толстой кишки, измеренная в день 8 у DITRA-подобных мышей. (C) Уровни провоспалительных цитокинов в гомогенатах толстой кишки у DITRA-подобных мышей, обработанных оксазолоном и средством-носителем, которым путем инъекции вводили PBS, mAb к человеческому IL-36R и изотипический контроль hIgG4. * - представляет собой значимость для группы, обработанной с помощью PBS, # - значимость для группы, обработанной изотипом. Данные являются репрезентативными для двух независимых экспериментов с 5 мышами на группу. Планки погрешностей представляют среднее значение ± SEM. *p<0,05, **p<0,005, ***p<0,0005, ****p<0,00001.
ПОДРОБНОЕ ОПИСАНИЕ
[0050] В настоящем документе раскрыты грызуны (такие как без ограничения мыши), генетически модифицированные таким образом, что они содержат гуманизированный ген Il1rl2 (кодирующий гуманизированный белок IL1RL2, имеющий эктодомен, практически идентичный эктодомену человеческого IL1RL2) и человеческие гены, кодирующие лиганды IL-36α, β, γ, в то же время сохраняя эндогенный антагонист IL-36Ra, который в 20 раз менее активен при подавлении передачи сигнала человеческого IL-36R. Было показано, что генетически модифицированные грызуны, раскрытые в данном документе, проявляют симптомы пациентов с DITRA, у которых мутация в IL-36Ra приводит к усилению передачи сигнала IL-36R. Следовательно, раскрытые в данном документе генетически модифицированные грызуны могут служить в качестве новой функциональной модели DITRA, подходящей для тестирования терапевтических средств-кандидатов для лечения DITRA и подобных нарушений. Различные варианты осуществления дополнительно описаны ниже.
Генетически модифицированные грызуны: Четырехкратная гуманизация
[0051] В одном аспекте некоторых вариантов осуществления в настоящем изобретении предусмотрены животные-грызуны, которые содержат в своем геноме гуманизированный ген Il1rl2, кодирующий гуманизированный белок IL1RL2 (субъединица IL-36R), и человеческие гены, кодирующие человеческие лиганды IL-36α, β, и γ. Такие грызуны также упоминаются в данном документе как четырехкратно гуманизированные грызуны (т.е. 4H или DITRA-подобные).
[0052] Термин «гуманизированный», когда его применяют в контексте нуклеиновых кислот или белков, включает нуклеиновые кислоты или белки грызуна, структуры которых (т.е., нуклеотидные или аминокислотные последовательности) были модифицированы полностью или частично для включения структур, обнаруженных в соответствующих человеческих нуклеиновых кислотах или белках. Грызун, содержащий гуманизированный ген или экспрессирующий гуманизированный белок, является «гуманизированным» грызуном.
[0053] В некоторых вариантах осуществления грызуны по настоящему изобретению включают, в качестве неограничивающих примеров, мышь, крысу и хомяка. В некоторых вариантах осуществления грызуны по настоящему изобретению включают, в качестве неограничивающих примеров, мышь и крысу. В некоторых вариантах осуществления грызун выбран из надсемейства Muroidea. В некоторых вариантах осуществления грызун по настоящему изобретению относится к семейству, выбранному из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяк, крысы и мыши Нового света, полевки), Muridae (настоящие мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (лазающие мыши, скальные мыши, белохвостые крысы, малагасийские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В некоторых вариантах осуществления грызун по настоящему изобретению выбран из настоящей мыши или крысы (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. В некоторых вариантах осуществления мышь по настоящему изобретению является представителем семейства Muridae.
Гуманизация IL-1RL2
[0054] IL-36R является представителем семейства рецепторов интерлейкина 1 и представляет собой гетеродимер, который состоит из субъединицы рецептора, называемой IL1RL2 (также известной как IL-1Rrp2), и вспомогательного белка корецепторной субъединицы рецептора интерлейкина-1, IL-1RAcP (Garlanda C et al., Immunity 39, 1003-1018 (2013); Towne JE et al., J. Biol. Chem. 279, 13677-13688 (2004), которые включены в данный документ посредством ссылки во всей своей полноте). Рецептор (IL-36R) способен распознавать и связывать три различных агониста, IL-36α, IL-36β и IL-36γ (также известных как IL1F6, IL1F8 и IL1F9), с индукцией экспрессии воспалительных цитокинов, а также антагонист, IL-36Ra, который связывается с IL-36R и снижает экспрессию воспалительных цитокинов.
[0055] IL1RL2 содержит сигнальный пептид, внеклеточный домен («ECD» или «эктодомен»), трансмембранный домен и внутриклеточный или цитоплазматический домен. См. фиг. 1D, например. Иллюстративные последовательности IL1RL2, включая человеческие, мышиные, крысиные и гуманизированные последовательности нуклеиновой кислоты и белка Il1rl2, раскрыты в перечне последовательностей и кратко описаны в следующей таблице.
Таблица 1
CDS: 127-1854
12 экзонов: 1-114, 115-184, 185-419, 420-615, 616-775, 776-850, 851-980, 981-1117, 1118-1261,1262-1423, 1424-1804, 1805-2615.
Сигнальный пептид: 1-19
Зрелый: 20-575
Эктодомен: 20-335
CDS: 238-1962
11 экзонов: 1-231, 232-301, 302-536, 537-735, 736-898, 899-970, 971-1100, 1101-1237, 1238-1381, 1382-1543, 1544-4072.
Белок NP_573456.1
Сигнальный пептид: 1-21
Зрелый: 22-574
Эктодомен: 22-338
CDS: 89-1774
Белок NP_598259.1
Сигнальный пептид: 1-21
Зрелый: 22-561
Эктодомен: 22-338
Белок
Сигнальный пептид: 1-21
Зрелый: 22-573
Эктодомен: 22-337
[0056] В некоторых вариантах осуществления раскрытый в данном документе грызун содержит в своем геноме гуманизированный ген Il1rl2, который содержит нуклеотидную последовательность эндогенного гена Il1rl2 грызуна и нуклеотидную последовательность человеческого гена IL1RL2. Используемый в данном документе термин «нуклеотидная последовательность гена» включает геномную последовательность, последовательность мРНК или кДНК гена полностью или частично. В качестве неограничивающего примера нуклеотидная последовательность человеческого гена IL1RL2 содержит геномную последовательность, последовательность мРНК или кДНК человеческого гена IL1RL2 полностью или частично. Нуклеотидная последовательность эндогенного гена Il1rl2 грызуна и нуклеотидная последовательность человеческого гена IL1RL2 функционально связаны друг с другом таким образом, что гуманизированный ген Il1rl2 в геноме грызуна кодирует белок Il1rl2, т.е. белок, который имеет структуру белка Il1rl2 (состоит из эктодомена, трансмембранного домена и цитоплазматического домена), и который выполняет функции Il1rl2 (например, образует гетеродимер со вспомогательным белком рецептора интерлейкина-1 (IL-1RacP) и распознает три лиганда: IL-36α, IL-36β и IL-36γ).
[0057] В некоторых вариантах осуществления генетически модифицированный грызун содержит гуманизированный ген Il1rl2 в своем геноме, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, который практически идентичен эктодомену человеческого белка IL1RL2. В некоторых вариантах осуществления эктодомен, который практически идентичен эктодомену человеческого белка IL1RL2, проявляет ту же функциональность (например, свойства связывания лиганда), что и эктодомен человеческого белка IL1RL2. Под эктодоменом или полипептидом, который «практически идентичен эктодомену человеческого белка IL1RL2», в некоторых вариантах осуществления подразумевается полипептид, который на по меньшей мере 95%, 98%, 99% или 100% идентичен по последовательности эктодомену человеческого белка IL1RL2; в некоторых вариантах осуществления полипептид, который отличается от эктодомена человеческого белка IL1RL2 не более, чем на 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислоту(аминокислот); и в некоторых вариантах осуществления полипептид, который отличается от эктодомена человеческого белка IL1RL2 только в N- или C-концевой части эктодомена, например, добавлением, делецией или заменой аминокислот, но не более 5, 4, 3, 2 или 1 аминокислоты в N- или C-концевой части эктодомена. Под «N- или C-концевой частью эктодомена» подразумевается 5-10 аминокислот от N- или C-конца эктодомена. В некоторых вариантах осуществления человеческий белок IL1RL2 имеет аминокислотную последовательность, практически идентичную (например, на по меньшей мере 95%, 98%, 99% или 100% идентичную) аминокислотной последовательности, указанной под SEQ ID NO: 2. В конкретных вариантах осуществления человеческий белок IL1RL2 содержит аминокислотную последовательность, указанную под SEQ ID NO: 2, и его эктодомен состоит из аминокислот 20-335 из SEQ ID NO: 2. В некоторых вариантах осуществления гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, который практически идентичен эктодомену человеческого белка IL1RL2, указанному под SEQ ID NO: 2, т.е. аминокислотам 20-335 под SEQ ID NO: 2. В некоторых вариантах осуществления гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, содержащий аминокислоты 22-337 из SEQ ID NO: 7.
[0058] В некоторых вариантах осуществления гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит трансмембранно-цитоплазматическую последовательность (т.е. последовательность, которая содержит как трансмембранный домен, так и цитоплазматический домен), которая практически идентична трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна. В некоторых вариантах осуществления трансмембранно-цитоплазматическая последовательность, которая практически идентична трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна, проявляет ту же функциональность (например, передачу сигнала и/или взаимодействие с внутриклеточными молекулами), что и трансмембранно-цитоплазматическая последовательность эндогенного белка Il1rl2 грызуна. Под трансмембранно-цитоплазматической последовательностью или полипептидом, который «практически идентичен трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна», в некоторых вариантах осуществления подразумевается полипептид, который на по меньшей мере 95%, 98%, 99% или 100% идентичен по последовательности трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна; в некоторых вариантах осуществления полипептид, который отличается от трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна не более, чем на 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислоту(аминокислот); в некоторых вариантах осуществления полипептид, который отличается от трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна только на N- или C-конце, например, наличием добавления, делеции или замены аминокислот, но не более 5, 4, 3, 2 или 1 аминокислоты в N- или C-концевой части трансмембранно-цитоплазматической последовательности. Под «N- или C-концевой частью трансмембранно-цитоплазматической последовательности» подразумевается 5-10 аминокислот от N-конца трансмембранного домена или 5-10 аминокислот от C-конца цитоплазматического домена. В некоторых вариантах осуществления гуманизированный белок Il1rl2 содержит трансмембранно-цитоплазматическую последовательность, которая практически идентична трансмембранно-цитоплазматической последовательности мышиного белка Il1rl2, например, мышиного белка Il1rl2, практически идентичного (на по меньшей мере 95%, 98%, 99% или 100% идентичного) SEQ ID NO: 4, или трансмембранно-цитоплазматической последовательности крысиного белка Il1rl2, например, крысиного белка Il1rl2, практически идентичного (на по меньшей мере 95%, 98%, 99% или 100% идентичного) SEQ ID NO: 6.
[0059] В некоторых вариантах осуществления гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит сигнальный пептид, который практически идентичен сигнальному пептиду эндогенного белка Il1rl2 грызуна. Под сигнальным пептидом, который «практически идентичен сигнальному пептиду эндогенного белка Il1rl2 грызуна», в некоторых вариантах осуществления подразумевается полипептид, который на по меньшей мере 95%, 98%, 99% или 100% идентичен по последовательности сигнальному пептиду эндогенного белка Il1rl2 грызуна; в некоторых вариантах осуществления полипептид, который отличается от сигнального пептида эндогенного белка Il1rl2 грызуна не более, чем на 3, 2 или 1 аминокислоту(аминокислоты); в некоторых вариантах осуществления полипептид, который отличается от сигнального пептида эндогенного белка Il1rl2 грызуна только на N- или C-конце, например, наличием добавления, делеции или замены аминокислот, но не более 3, 2 или 1 аминокислоты в N- или C-концевой части сигнального пептида. Под «N- или C-концевой частью сигнального пептида» подразумевается в пределах 5 аминокислот от N- или C-конца сигнального пептида. В некоторых вариантах осуществления гуманизированный белок Il1rl2 содержит сигнальный пептид, практически идентичный сигнальному пептиду мышиного белка Il1rl2, например, мышиного белка Il1rl2, практически идентичного (на по меньшей мере 95%, 98%, 99% или 100% идентичного) SEQ ID NO: 4, или сигнальному пептиду крысиного белка Il1rl2, например, крысиного белка Il1rl2, практически идентичного (на по меньшей мере 95%, 98%, 99% или 100% идентичного) SEQ ID NO: 6.
[0060] В некоторых вариантах осуществления гуманизированный ген Il1rl2 в геноме генетически модифицированного грызуна содержит нуклеотидную последовательность человеческого гена IL1RL2 (или «нуклеотидную последовательность человеческого IL1RL2») и нуклеотидную последовательность эндогенного гена Il1rl2 грызуна (или «эндогенную нуклеотидную последовательность Il1rl2 грызуна»), где нуклеотидная последовательность человеческого IL1RL2 кодирует полипептид, практически идентичный эктодомену человеческого белка IL1RL2, кодируемого человеческим геном IL1RL2. Такая нуклеотидная последовательность человеческого IL1RL2 также упоминается как кодирующая по существу эктодомен человеческого белка IL1RL2. В некоторых вариантах осуществления нуклеотидная последовательность человеческого IL1RL2 представляет собой геномный фрагмент человеческого гена IL1RL2. В некоторых вариантах осуществления нуклеотидная последовательность человеческого IL1RL2 представляет собой геномный фрагмент, содержащий экзоны 3-8 человеческого гена IL1RL2. В некоторых вариантах осуществления нуклеотидная последовательность человеческого IL1RL2 представляет собой геномный фрагмент, содержащий 3'-концевую часть интрона 2, экзоны 3-8 и 5'-концевую часть интрона 8 человеческого гена IL1RL2. В некоторых вариантах осуществления нуклеотидная последовательность человеческого IL1RL2 представляет собой последовательность кДНК.
[0061] В некоторых вариантах осуществления гуманизированный ген Il1rl2 в геноме генетически модифицированного грызуна содержит эндогенную нуклеотидную последовательность Il1rl2 грызуна и нуклеотидную последовательность человеческого IL1RL2 , где эндогенная нуклеотидная последовательность Il1rl2 грызуна кодирует полипептид, практически идентичный трансмембранно-цитоплазматической последовательности эндогенного белка Il1rl2 грызуна. Такая нуклеотидная последовательность Il1rl2 грызуна также упоминается как кодирующая по существу трансмембранно-цитоплазматическую последовательность эндогенного белка Il1rl2 грызуна.. В некоторых вариантах осуществления эндогенная нуклеотидная последовательность Il1rl2 грызуна, присутствующая в гуманизированном гене Il1rl2, кодирует трансмембранно-цитоплазматическую последовательность эндогенного белка Il1rl2 грызуна. В некоторых вариантах осуществления эндогенная нуклеотидная последовательность Il1rl2 грызуна, присутствующая в гуманизированном гене Il1rl2, содержит оставшиеся экзоны ниже экзона 8 в эндогенном гене Il1rl2 грызуна. В некоторых вариантах осуществления эндогенная нуклеотидная последовательность Il1rl2 грызуна, присутствующая в гуманизированном гене Il1rl2, содержит 3'-концевую часть интрона 8 и оставшиеся экзоны ниже экзона 8 в эндогенном гене Il1rl2 грызуна.
[0062] В некоторых вариантах осуществления гуманизированный ген Il1rl2 в геноме генетически модифицированного грызуна содержит эндогенную нуклеотидную последовательность Il1rl2 грызуна выше (5')-конца нуклеотидной последовательности человеческого IL1RL2 , где эндогенная нуклеотидная последовательность Il1rl2 грызуна кодирует полипептид, практически идентичный сигнальному пептиду эндогенного белка Il1rl2 грызуна. Такая нуклеотидная последовательность Il1rl2 грызуна также упоминается как кодирующая по существу сигнальный пептид эндогенного белка Il1rl2 грызуна. В некоторых вариантах осуществления эндогенная нуклеотидная последовательность Il1rl2 грызуна, кодирующая полипептид, практически идентичный сигнальному пептиду эндогенного белка Il1rl2 грызуна, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна; и в некоторых вариантах осуществления эндогенная нуклеотидная последовательность Il1rl2 грызуна содержит экзоны 1-2 и 5'-концевую часть интрона 2 эндогенного гена Il1rl2 грызуна.
[0063] В некоторых вариантах осуществления гуманизированный ген Il1rl2 функционально связан с эндогенными регуляторными последовательностями Il1rl2 грызуна, например, с 5'-концевой последовательностью(последовательностями) для регуляции транскрипции, такой как промотор и/или энхансеры, например, экспрессия гуманизированного гена Il1rl2 находится под контролем 5'-концевой регуляторной(регуляторных) последовательности(последовательностей) Il1rl2 грызуна.
[0064] В некоторых вариантах осуществления гуманизированный ген Il1rl2 находится в эндогенном локусе Il1rl2 грызуна. В некоторых вариантах осуществления гуманизированный ген Il1rl2 находится в локусе, отличном от эндогенного локуса Il1rl2 грызуна; например, в результате случайной интеграции. В некоторых вариантах осуществления, где гуманизированный ген Il1rl2 находится в локусе, отличном от эндогенного локуса Il1rl2 грызуна, грызуны неспособны экспрессировать белок Il1rl2 грызуна, например, в результате инактивации (например, полной или частичной делеции) эндогенного гена Il1rl2 грызуна.
[0065] В некоторых вариантах осуществления, где гуманизированный ген Il1rl2 находится в эндогенном локусе Il1rl2 грызуна, гуманизированный ген Il1rl2 является результатом замены нуклеотидной последовательности эндогенного гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна на нуклеотидную последовательность человеческого гена IL1RL2.
[0066] В некоторых вариантах осуществления нуклеотидная последовательность эндогенного гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, которая подлежит замене, представляет собой геномный фрагмент эндогенного гена Il1rl2 грызуна, который кодирует по существу эктодомен белка Il1rl2 грызуна. В некоторых вариантах осуществления подлежащий замене геномный фрагмент грызуна содержит экзоны 3-8 эндогенного гена Il1rl2 грызуна.
[0067] В некоторых вариантах осуществления нуклеотидная последовательность человеческого гена IL1RL2, которая заменяет геномный фрагмент гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, представляет собой последовательность кДНК. В некоторых вариантах осуществления нуклеотидная последовательность человеческого IL1RL2, которая заменяет геномный фрагмент гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, представляет собой геномный фрагмент человеческого гена IL1RL2. В некоторых вариантах осуществления геномный фрагмент человеческого гена IL1RL2, который заменяет геномный фрагмент гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, содержит экзоны, полностью или частично, человеческого гена IL1RL2, которые кодируют по существу эктодомен человеческого белка IL1RL2. В некоторых вариантах осуществления человеческий геномный фрагмент содержит экзоны 3-8 человеческого гена IL1RL2.
[0068] В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, которая остается в эндогенном локусе Il1rl2 грызуна после замены для гуманизации и функционально связана со вставленной нуклеотидной последовательностью человеческого IL1RL2, кодирует по существу трансмембранно-цитоплазматическую последовательность эндогенного белка Il1rl2 грызуна. В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, которая остается в эндогенном локусе Il1rl2 грызуна после замены для гуманизации, содержит экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
[0069] В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, которая остается в эндогенном локусе Il1rl2 грызуна после замены для гуманизации и функционально связана со вставленной нуклеотидной последовательностью человеческого IL1RL2, кодирует по существу сигнальный пептид эндогенного белка Il1rl2 грызуна. В некоторых вариантах осуществления геномная последовательность эндогенного гена Il1rl2 грызуна, которая остается в эндогенном локусе Il1rl2 грызуна после замены для гуманизации, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна.
[0070] В некоторых вариантах осуществления в случаях, когда эндогенный белок Il1rl2 грызуна и человеческий белок IL1RL2 имеют общие аминокислоты вблизи места соединения трансмембранного домена и эктодомена, может не потребоваться вставка нуклеотидной последовательности человеческого IL1RL2, которая точно кодирует эктодомен человеческого белка IL1RL2. Можно вставить немного более длинную или более короткую нуклеотидную последовательность человеческого гена IL1RL2, которая кодирует по существу эктодомен человеческого белка IL1RL2, в функциональной связи с геномной последовательностью эндогенного гена Il1rl2 грызуна, которая кодирует по существу трансмембранный домен (и цитоплазматический домен) эндогенного белка Il1rl2 грызуна таким образом, что гуманизированный белок Il1rl2, кодируемый полученным в результате гуманизированным геном Il1rl2, содержит эктодомен, который идентичен эктодомену человеческого белка IL1RL2, и трансмембранный домен, который идентичен трансмембранному домену эндогенного белка Il1rl2 грызуна.
[0071] В некоторых вариантах осуществления геномный фрагмент, содержащий экзоны 3-8 эндогенного гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, был заменен на геномный фрагмент, содержащий экзоны 3-8 человеческого гена IL1RL2. В результате гуманизированный ген Il1rl2 образуется в эндогенном локусе Il1rl2 грызуна и содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна, экзоны 3-8 человеческого гена IL1RL2 и оставшиеся экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
[0072] В некоторых вариантах осуществления грызун, предусмотренный в данном документе, является гетерозиготным по гуманизированному гену Il1rl2 в своем геноме. В некоторых вариантах осуществления грызун, предусмотренный в данном документе, является гомозиготным по гуманизированному гену Il1rl2 в своем геноме.
[0073] В некоторых вариантах осуществления гуманизированный ген Il1rl2 приводит к экспрессии кодируемого гуманизированного белка Il1rl2 у грызуна. В некоторых вариантах осуществления гуманизированный белок Il1rl2 экспрессируется по паттерну, сравнимому с соответствующим белком Il1rl2 грызуна или практически такому же, у контрольного грызуна (например, грызуна без гуманизированного гена Il1rl2). В некоторых вариантах осуществления гуманизированный белок Il1rl2 экспрессируется на уровне, сравнимом с соответствующим белком Il1rl2 грызуна или практически таком же, у контрольного грызуна (например, грызуна без гуманизированного гена Il1rl2). В некоторых вариантах осуществления гуманизированный белок Il1rl2 экспрессируется и выявляется на поверхности клетки, например без ограничения на поверхности клеток, таких как кератиноциты, моноциты, макрофаги, нейтрофилы, эпителиальные клетки бронхов и кишечника среди прочих. В контексте сравнения гуманизированного гена или белка у гуманизированного грызуна с эндогенным геном или белком грызуна у контрольного грызуна, термин «сравнимый» означает, что сравниваемые молекулы или уровни могут не быть идентичными друг другу, но быть достаточно сходными для обеспечения возможности их сравнения, чтобы можно было сделать обоснованные выводы на основании наблюдаемых различий или сходств. В некоторых вариантах осуществления термин «практически одинаковый» применительно к уровням экспрессии включает сравниваемые уровни, которые не отличаются друг от друга на более чем 20%, 19%, 18%, 17%, 16%, 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1%.
[0074] В некоторых вариантах осуществления раскрытые в данном документе грызуны неспособны экспрессировать белок Il1rl2 грызуна, например, в результате инактивации (например, полной или частичной делеции) или замены (полной или частичной) эндогенного гена Il1rl2 грызуна.
Гуманизация лигандов IL-36
[0075] Раскрытые в данном документе грызуны также содержат гены зародышевого типа, кодирующие человеческие IL-36α, β, и γ лиганды.
Таблица 2
CDS: 412-888
Зрелый: 6-158
CDS: 162-644
Белок NP_062323.1
Зрелый: 8-160
CDS: 162-644
Белок, NP_001100024.1
[0076] В некоторых вариантах осуществления раскрытый в данном документе грызун имеет геном, содержащий человеческий ген IL-36α, который кодирует человеческий белок IL-36α. Упоминания «человеческого гена IL-36α» включают человеческую геномную ДНК, которая кодирует человеческий белок IL-36α и содержит человеческий промотор IL-36α. Человеческий белок IL-36α может представлять собой зрелую форму или форму предшественника человеческого белка IL-36α. В некоторых вариантах осуществления человеческий белок IL-36α содержит аминокислотную последовательность под SEQ ID NO: 9.
[0077] В некоторых вариантах осуществления человеческий ген IL-36α находится в эндогенном локусе Il-36α грызуна. В некоторых вариантах осуществления человеческий ген IL-36α находится в локусе, отличном от эндогенного локуса Il-36α грызуна; например, в результате случайной интеграции. В некоторых вариантах осуществления, где человеческий ген IL-36α находится в локусе, отличном от эндогенного локуса Il-36α грызуна, грызуны неспособны экспрессировать белок Il-36α грызуна, например, в результате инактивации (например, полной или частичной делеции) эндогенного гена Il-36α грызуна.
[0078] В некоторых вариантах осуществления человеческий ген IL-36α заменяет ген Il-36α грызуна в эндогенном локусе Il-36α грызуна.
[0079] В некоторых вариантах осуществления грызун, предусмотренный в данном документе, является гетерозиготным по человеческому гену IL-36α в своем геноме. В других вариантах осуществления грызун, предусмотренный в данном документе, является гомозиготным по человеческому гену IL-36α в своем геноме.
[0080] В некоторых вариантах осуществления человеческий ген IL-36α приводит к экспрессии кодируемого человеческого белка IL-36α у грызуна, например, в сыворотке крови, на участках слизистой оболочки, таких как кожа, эпителий кишечника, легкие, и в различных типах клеток иммунной системы (например, в моноцитах, макрофагах, Т-клетках, дендритных клетках). В некоторых вариантах осуществления человеческий белок IL-36α экспрессируется по паттерну, сравнимому с соответствующим белком Il-36 грызуна или практически такому же, α у контрольного грызуна (например, грызуна без человеческого гена IL-36α). В некоторых вариантах осуществления человеческий белок IL-36α экспрессируется на уровне, сравнимом с соответствующим белком Il-36 грызуна или практически таком же, αу контрольного грызуна (например, грызуна без человеческого гена IL-36α), например, в сыворотке крови, на участках слизистой оболочки (например, таких как кожа, эпителий кишечника, легкие) и/или в клетках иммунной системы (например, моноцитах, макрофагах, Т-клетках, дендритных клетках).
[0081] В некоторых вариантах осуществления раскрытые в данном документе грызуны неспособны экспрессировать белок Il-36α грызуна, например, в результате инактивации (например, полной или частичной делеции) или замены (полной или частичной) эндогенного гена Il-36α грызуна.
Таблица 3
CDS: 109-603
CDS: 66-617
Белок NP_081439.1
CDS: 92-415
Белок, XP_006233676
[0082] В некоторых вариантах осуществления раскрытые в данном документе грызуны имеют геном, содержащий человеческий ген IL-36β, который кодирует человеческий белок IL-36β. Упоминания «человеческого гена IL-36β» включают человеческую геномную ДНК, которая кодирует человеческий белок IL-36β и содержит человеческий промотор IL-36β. Человеческий белок IL-36β может представлять собой зрелую форму или форму предшественника человеческого белка IL-36β. В некоторых вариантах осуществления человеческий белок IL-36β содержит аминокислотную последовательность под SEQ ID NO: 15.
[0083] В некоторых вариантах осуществления человеческий ген IL-36β находится в эндогенном локусе Il-36β грызуна. В некоторых вариантах осуществления человеческий ген IL-36β находится в локусе, отличном от эндогенного локуса Il-36β грызуна; например, в результате случайной интеграции. В некоторых вариантах осуществления, где человеческий ген IL-36β находится в локусе, отличном от эндогенного локуса Il-36β грызуна, грызуны неспособны экспрессировать белок Il-36β грызуна, например, в результате инактивации (например, полной или частичной делеции) эндогенного гена Il-36β грызуна.
[0084] В некоторых вариантах осуществления человеческий ген IL-36β заменяет ген Il-36β грызуна в эндогенном локусе Il-36β грызуна.
[0085] В некоторых вариантах осуществления грызун, предусмотренный в данном документе, является гетерозиготным по человеческому гену IL-36β в своем геноме. В других вариантах осуществления грызун, предусмотренный в данном документе, является гомозиготным по человеческому гену IL-36β в своем геноме.
[0086] В некоторых вариантах осуществления человеческий ген IL-36β приводит к экспрессии кодируемого человеческого белка IL-36β у грызуна, например, в сыворотке крови, на участках слизистой оболочки (например, таких как кожа, эпителий кишечника, легкие) и/или в клетках иммунной системы (например, моноцитах, макрофагах, Т-клетках, дендритных клетках). В некоторых вариантах осуществления человеческий белок IL-36β экспрессируется по паттерну, сравнимому с соответствующим белком Il-36 грызуна или практически такому же, β у контрольного грызуна (например, грызуна без человеческого гена IL-36β). В некоторых вариантах осуществления человеческий белок IL-36β экспрессируется на уровне, сравнимом с соответствующим белком Il-36 грызуна или практически таком же, βу контрольного грызуна (например, грызуна без человеческого гена IL-36β), например, в сыворотке крови, на участках слизистой оболочки, таких как кожа, эпителий кишечника, легкие, и в различных типах клеток иммунной системы (например, моноцитах, макрофагах, Т-клетках, дендритных клетках).
[0087] В некоторых вариантах осуществления раскрытые в данном документе грызуны неспособны экспрессировать белок I Il-36β грызуна, например, в результате инактивации (например, полной или частичной делеции) или замены (полной или частичной) эндогенного гена Il-36β грызуна.
Таблица 4
CDS: 80-589
CDS: 295-876
Белок, Q8R460.1
CDS: 166-663
Белок, NP_001107262.1
[0088] В некоторых вариантах осуществления в настоящем изобретении предусмотрен грызун, геном которого содержит человеческий ген IL-36γ, который кодирует человеческий белок IL-36γ. Упоминания «человеческого гена IL-36γ» включают человеческую геномную ДНК, которая кодирует человеческий белок IL-36γ и содержит человеческий промотор IL-36γ. В некоторых вариантах осуществления человеческий белок IL-36γ может представлять собой зрелую форму или форму предшественника человеческого белка IL-36γ. В некоторых вариантах осуществления человеческий белок IL-36γ содержит аминокислотную последовательность под SEQ ID NO: 21.
[0089] В некоторых вариантах осуществления человеческий ген IL-36γ находится в эндогенном локусе Il-36γ грызуна. В некоторых вариантах осуществления человеческий ген IL-36γ находится в локусе, отличном от эндогенного локуса Il-36γ грызуна; например, в результате случайной интеграции. В некоторых вариантах осуществления, где человеческий ген IL-36γ находится в локусе, отличном от эндогенного локуса Il-36γ грызуна, грызуны неспособны экспрессировать белок Il-36γ грызуна, например, в результате инактивации (например, полной или частичной делеции) эндогенного гена Il-36γ грызуна.
[0090] В некоторых вариантах осуществления человеческий ген IL-36γ заменяет ген Il-36γ грызуна в эндогенном локусе Il-36γ грызуна.
[0091] В некоторых вариантах осуществления грызун, предусмотренный в данном документе, является гетерозиготным по человеческому гену IL-36γ в своем геноме. В других вариантах осуществления грызун, предусмотренный в данном документе, является гомозиготным по человеческому гену IL-36γ в своем геноме.
[0092] В некоторых вариантах осуществления человеческий ген IL-36γ приводит к экспрессии кодируемого человеческого белка IL-36γ (например, белка, идентичного человеческому белку IL-36γ) у грызуна, например, в сыворотке крови, на участках слизистой оболочки, таких как кожа, эпителий кишечника, легкие, и в различных типах клеток иммунной системы (например, моноцитах, макрофагах, Т-клетках, дендритных клетках). В некоторых вариантах осуществления человеческий белок IL-36γ экспрессируется по паттерну, сравнимому с соответствующим белком Il-36 грызуна или практически такому же, γ у контрольного грызуна (например, грызуна без человеческого гена IL-36γ). В некоторых вариантах осуществления человеческий белок IL-36γ экспрессируется на уровне, сравнимом с соответствующим белком Il-36 грызуна или практически таком же, γу контрольного грызуна (например, грызуна без человеческого гена IL-36γ), например, в сыворотке крови, на участках слизистой оболочки (например, таких как кожа, эпителий кишечника, легкие) и/или в клетках иммунной системы (например, моноцитах, макрофагах, Т-клетках, дендритных клетках).
[0093] В некоторых вариантах осуществления раскрытые в данном документе грызуны неспособны экспрессировать белок Il-36γ грызуна, например, в результате инактивации (например, полной или частичной делеции) или замены (полной или частичной) эндогенного гена Il-36γ грызуна.
[0094] В некоторых вариантах осуществления предусмотрены грызуны, геном которых содержит замену непрерывного геномного фрагмента, охватывающего кодирующие последовательности всех трех лигандов Il-36 в эндогенном локусе, на непрерывную нуклеиновую кислоту, содержащую кодирующие последовательности трех лигандов, которые практически идентичны человеческому IL-36α, β, и γ, соответственно. В некоторых вариантах осуществления полученный в результате локус содержит от 5'-конца к 3'-концу (i) человеческий ген IL-36β, (ii) человеческий ген IL-36γ и (iii) обратную цепь человеческого гена IL-36α.
Фенотипы четырехкратно гуманизированных грызунов
[0095] У раскрытых в данном документе генетически модифицированных грызунов не развиваются какие-либо спонтанные заболевания в устойчивом состоянии; однако эти грызуны действительно демонстрируют укороченную толстую кишку и повышенную экспрессию провоспалительных медиаторов в коже по сравнению с контрольными грызунами того же возраста без гуманизации, как в устойчивом состоянии, так и в состоянии заболевания (например, после обработки с помощью DSS или IMQ). В некоторых вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на по меньшей мере 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45% или 50% короче, чем у контрольного грызуна в устойчивом состоянии или в состоянии заболевания.
[0096] В некоторых вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 10-15% (+/- 5%) короче, чем у контрольного грызуна в устойчивом состоянии. В некоторых вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 15-20% (+/- 5%) короче, чем у контрольного грызуна в устойчивом состоянии. В некоторых вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 10-20% (+/- 5%) короче, чем у контрольного грызуна в устойчивом состоянии.
[0097] В других вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 20-40% (+/- 5%) короче, чем у контрольного грызуна в состоянии заболевания (например, после обработки с помощью DSS или оксазолона). В других вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 30-40% (+/- 5%) короче, чем у контрольного грызуна в состоянии заболевания (например, после обработки с помощью DSS или оксазолона). В других вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 20-30% (+/- 5%) короче, чем у контрольного грызуна в состоянии заболевания (например, после обработки с помощью DSS или оксазолона). В других вариантах осуществления генетически модифицированный грызун демонстрирует толстую кишку, длина которой на 25-35% (+/- 5%) короче, чем у контрольного грызуна в состоянии заболевания (например, после обработки с помощью DSS или оксазолона).
[0098] Хотя у генетически модифицированных грызунов, раскрытых в данном документе, не развиваются какие-либо спонтанные заболевания в устойчивом состоянии, было показано, что они демонстрируют усиленное воспаление кожи и кишечника в моделях экспериментально индуцированного воспаления кожи и кишечника (например, доклинических моделях псориаза и IBD, соответственно).
[0099] В некоторых вариантах осуществления DSS применяют для индукции хронического колита. В некоторых вариантах осуществления DSS вводят грызунам с питьевой водой в количестве, составляющем по меньшей мере 0,5%, по меньшей мере 1%, по меньшей мере 1,5%, по меньшей мере 2,5%. В некоторых вариантах осуществления DSS вводят грызунам с питьевой водой в количестве, составляющем не более чем 10%, 9%, 8%, 7%, 6% или 5%. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 1,5-3% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 0,5-3% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 1-3% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 2-3% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 2,5-3% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 0,5-2,5% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 0,5-2% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 0,5-1,5% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 0,5-1% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 1-2,5% DSS. В некоторых вариантах осуществления грызунам дают питьевую воду, содержащую 1,5-2% DSS.
[0100] В некоторых вариантах осуществления введение DSS можно осуществлять в течение периода времени, составляющего 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или больше дней, и его можно осуществлять непрерывно или прерывать на несколько дней без введения DSS. В некоторых вариантах осуществления грызунов обеспечивают питьевой водой, содержащей 2,5% DSS, в течение 7 дней, затем 1,5% DSS в течение 5 дней, а затем дистиллированной водой до 27-30 дней перед анализом. В некоторых вариантах осуществления грызунов обеспечивают питьевой водой, содержащей 3% DSS, в течение 7 дней, затем 2% DSS в течение 13 дней, а затем дистиллированной водой до 27-30 дней перед анализом. В некоторых вариантах осуществления DSS не вводят в течение всего периода времени. В некоторых вариантах осуществления грызунов обеспечивают питьевой водой, содержащей 2,5% DSS, в течение 7 дней, затем дистиллированной водой в течение 11 дней, затем 1,5% DSS в течение 4 дней, затем дистиллированной водой в течение 5 дней - всего в течение 27 дней перед анализом. В некоторых вариантах осуществления грызуны получают питьевую воду, содержащую 3% DSS, в течение 7 дней, затем воду в течение 13 дней, затем 2% DSS в течение 4 дней, затем дистиллированную воду в течение 6 дней до 30 дней перед анализом.
[0101] В некоторых вариантах осуществления оксазолон применяют для индукции колита. В некоторых вариантах осуществления оксазолон вводят грызунам интраректально для индукции колита. В некоторых вариантах осуществления оксазолон вводят грызунам интраректально посредством трех введений для индукции колита. В некоторых вариантах осуществления оксазолон наносят грызунам местно для предварительной сенсибилизации перед интраректальным введением. В некоторых вариантах осуществления оксазолон применяют по отношению к грызунам для предварительной сенсибилизации путем местного нанесения раствора оксазолона (например, 3% раствора оксазолона, растворенного в 100% этаноле) на выбритую кожу с последующими тремя интраректальными введениями раствора оксазолона (например, 1,0-2,0% оксазолона, растворенного в 50% этаноле). В некоторых вариантах осуществления для предварительной сенсибилизации применяют раствор 1,5%, 2,0%, 2,5%, 3,0%, 3,5% или 4,0% оксазолона, растворенного в 100% этаноле. В некоторых вариантах осуществления для интраректальных введений применяют раствор 1,0%, 1,1%, 1,2%, 1,3%, 1,5%, 1,6%, 1,7%, 1,8%, 1,9% или 2,0% оксазолона, растворенного в 50% этаноле. В некоторых вариантах осуществления интраректальное введение осуществляют через несколько дней, например, через 3, 4, 5, 6 или 7 дней после предварительной сенсибилизации. В некоторых вариантах осуществления оксазолон применяют по отношению к грызунам путем местного нанесения на кожу в виде раствора 3,0% оксазолона, растворенного в 100% этаноле, с последующими тремя интраректальными введениями раствора 1,0-2,0% оксазолона, растворенного в 50% этаноле, в дни 5, 6 и 7.
[0102] В некоторых вариантах осуществления степень колита оценивают посредством определения следующих признаков: воспаление (тяжесть и степень), эпителиальные изменения (эрозия/язва), изменения в криптах (потеря крипт, криптит/абсцесс крипт, регенерация/гиперплазия, потеря бокаловидных клеток), отек подслизистой оболочки и процент площади ткани с патологией относительно общей площади ткани на препарате. Применяют шкалу для оценивания 0-4: 0-0 - в пределах нормы, 1 - минимальная, 2 - легкая, 3 - умеренная и 4 - тяжелая степень. Общий показатель патологии рассчитывают для каждого грызуна посредством сложения отдельных показателей гистопатологических признаков. В некоторых вариантах осуществления колит оценивают посредством измерения уровня липокалина-2 (Lcn2) в образцах фекалий. В некоторых вариантах осуществления колит измеряют посредством измерения уровня активности миелопероксидазы (MPO) в гомогенатах толстой кишки. В некоторых вариантах осуществления колит оценивают посредством измерения уровней воспалительных цитокинов в гомогенатах толстой кишки.
[0103] В некоторых вариантах осуществления генетически модифицированные грызуны демонстрируют повышенный показатель патологии кишечника, например, на по меньшей мере 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400% или больше по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению DSS. В некоторых вариантах осуществления генетически модифицированные грызуны демонстрируют повышенный показатель патологии кишечника, например, на 50-400%, 50-300%, 50-200%, 50-100%, 100-400%, 100-300%, 100-200% или 200-400% по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению DSS. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенный уровень миелопероксидазы («MPO») в гомогенатах толстой кишки, например, на по меньшей мере 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400% или больше по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению DSS. В некоторых вариантах осуществления генетически модифицированные грызуны демонстрируют повышенный уровень MPO в гомогенатах толстой кишки, например, на 50-400%, 50-300%, 50-200%, 50-100%, 100-400%, 100-300%, 100-200% или 200-400% по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению DSS. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенный уровень фекального Lcn, например, на по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400% или больше по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению DSS. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов (например, KC-GRO, IL-6, IL-1β, TNFα, IL-21, IL-12p40, IL-17f, IL-17a и IL-17c) в гомогенатах толстой кишки, например, на по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500%, 600%, 700%, 800%, 900% или больше по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению DSS. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов в гомогенатах толстой кишки, например, на 20%-900%, 20%-800%, 20%-700%, 20%-600%, 20%-500%, 20%-400%, 20%-300%, 20%-200% или 20%-100%. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов в гомогенатах толстой кишки, например, на 30%-900%, 30%-800%, 30%-700%, 30%-600%, 30%-500%, 30%-400%, 30%-300%, 30%-200% или 30%-100%. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов в гомогенатах толстой кишки, например, на 40%-900%, 40%-800%, 40%-700%, 40%-600%, 40%-500%, 40%-400%, 40%-300%, 40%-200% или 40%-100%. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов в гомогенатах толстой кишки, например, на 50%-900%, 50%-800%, 50%-700%, 50%-600%, 50%-500%, 50%-400%, 50%-300%, 50%-200% или 50%-100%.
[0104] В некоторых вариантах осуществления IMQ наносят местно на кожу грызуна для индукции воспаления кожи. В некоторых вариантах осуществления IMQ применяют в носителе, подходящем для местного нанесения, например, креме, геле, включая коммерчески доступные кремы на основе IMQ (например, такие как кремы от Aldara). В некоторых вариантах осуществления для индукции воспаления кожи IMQ наносят на кожу грызуна ежедневно при суточной дозе, составляющей 1-5 мг, 2-4 мг или 3-3,5 мг в течение периода времени, составляющего 2, 3, 4, 5, 6, 7, 8, 9, 10 дней или дольше. В некоторых вариантах осуществления ежедневное местное нанесение при суточной дозе, составляющей приблизительно 3,125 мг в течение 4 дней, подходит для индукции острого воспаления кожи, а ежедневное местное нанесение при суточной дозе, составляющей приблизительно 3,125 мг в течение 9 дней, подходит для индукции хронического воспаления кожи. В некоторых вариантах осуществления IMQ наносят местно в течение множества циклов (например, 2, 3 или 4 циклов) с 4-5 последовательными днями нанесения IMQ, за которыми следуют 2 дня без нанесения IMQ в каждом цикле, перед проведением анализа или исследования. В конкретном варианте осуществления IMQ наносят местно в течение двух циклов с 5 последовательными днями нанесения IMQ, за которыми следуют 2 дня без нанесения IMQ в первом цикле, затем 4 последовательных дня нанесения IMQ во втором цикле, перед проведением анализа (см., например, фиг. 7A-7C). В некоторых вариантах осуществления тяжесть воспаления можно оценить посредством (i) использования адаптированной версии клинического индекса распространенности и тяжести псориаза на основании измерения эритемы, шелушения и утолщения кожи; (ii) осуществления гистопатологического анализа тканей кожи, например, для оценки наличия паракератоза, ортокератоза, микроабсцесса Мунро, акантоза, эпидермального изъязвления, воспаления в дерме и гиподерме, закупорки кровеносных сосудов в дерме и гиподерме, фолликулярного гиперкератоза и эпителиальной гиперплазии, и для определения общего показателя патологии; (iii) измерения экспрессии мРНК и/или уровней белков провоспалительных медиаторов в гомогенатах кожи, включая, например, экспрессию мРНК и/или уровни белков Cxcl1, IL-17f, IL-17a, IL-23a, S100A8 и Defb4, среди прочего; и (iv) комбинации (i)-(iii).
[0105] В некоторых вариантах осуществления генетически модифицированные грызуны демонстрируют повышенный показатель патологии кожи, например, на по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200% или больше по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению IMQ. В некоторых вариантах осуществления генетически модифицированные грызуны проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов (например без ограничения Cxcl1, IL-17f, IL-17a, IL-23a, S100A8 и Defb4) в гомогенатах кожи, например, на по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300% или больше по сравнению с контрольными грызунами дикого типа, которые подвергались такому же введению IMQ. В некоторых вариантах осуществления генетически модифицированные грызуны в устойчивом состоянии проявляют повышенную экспрессию мРНК и/или уровень белка одного или более провоспалительных медиаторов (например без ограничения Cxcl1, IL-17f, IL-17a, IL-23a, S100A8 и Defb4) в гомогенатах кожи, например, на по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300% или больше по сравнению с контрольными грызунами дикого типа в устойчивом состоянии.
[0106] В случае IMQ-индуцированного воспаления кожи отдельные мыши с гуманизированным Il1rl2 проявляли фенотипы, сходные с фенотипами грызуна дикого типа, что находит отражение в гистопатологии и экспрессии РНК провоспалительных молекул в коже (см. фиг. 3D и 3E). С другой стороны, у DITRA-подобных мышей (т.е. четырехкратно гуманизированных мышей, содержащих гуманизированный Il1rl2 и человеческие IL1F6, IL1F8 и IL1F9) после нанесения IMQ развивалось усиленное воспаление кожи по сравнению как с мышами WT, так и мышами 1H (см. фиг. 3D и 3E).
Ткани и клетки генетически модифицированных грызунов
[0107] В некоторых вариантах осуществления в данном документе раскрыта выделенная клетка или ткань животного-грызуна, описанного в данном документе. В некоторых вариантах осуществления клетка выбрана из дендритной клетки, лимфоцита (например, B- или T-клетки), макрофага и моноцита. В некоторых вариантах осуществления ткань выбрана из жировой ткани, ткани мочевого пузыря, головного мозга, молочной железы, костного мозга, глаза, сердца, кишечника, почки, печени, легкого, лимфатического узла, мышцы, поджелудочной железы, плазмы крови, сыворотки крови, кожи, селезенки, желудка, тимуса, яичка, яичника и их комбинации. В некоторых вариантах осуществления выделенная клетка или ткань содержит в своем геноме признаки четырехкратной гуманизации, описанные в данном документе.
Композиции и способы получения четырехкратно гуманизированных грызунов
[0108] Дополнительные аспекты настоящего изобретения направлены на способы получения генетически модифицированного грызуна, описанного выше, а также векторов на основе нуклеиновых кислот и эмбриональных стволовых клеток грызунов, подходящих для использования для получения генетически модифицированного грызуна.
[0109] Четырехкратно гуманизированного грызуна, т.е. грызуна, содержащего гуманизированный Il1rl2 и человеческие гены IL1F6, IL1F8 и IL1F9, можно получить посредством создания линии грызунов с одним гуманизированным Il1rl2 , и линии грызунов с тремя человеческими Il1f6, Il1f8 и Il1f9, с последующим скрещиванием линий с одним гуманизированным геном и с тремя человеческими генами друг с другом с получением четырехкратно гуманизированного грызуна. Термин «размножение» или «скрещивание», используемый в данном документе по отношению к грызунам, относится к спариванию грызунов для получения потомства. Специалисты в данной области техники поймут, что для достижения гомозиготности может потребоваться более одного скрещивания.
[0110] В некоторых вариантах осуществления в данном документе раскрыт нацеливающийся вектор (или конструкция нуклеиновой кислоты), который содержит человеческую нуклеотидную последовательность, которую необходимо интегрировать в локус грызуна. В некоторых вариантах осуществления человеческая нуклеотидная последовательность может представлять собой нуклеотидную последовательность человеческого гена IL1RL2, который кодирует эктодомен, практически идентичный эктодомену человеческого IL1RL2, например, нуклеотидную последовательность, содержащую экзоны 3-8 человеческого гена IL1RL2. В некоторых вариантах осуществления человеческая нуклеотидная последовательность может представлять собой нуклеотидную последовательность, охватывающую кодирующую последовательность человеческого IL-36α, кодирующую последовательность человеческого Il-36β и комплементарную цепь кодирующей последовательности человеческого IL-36γ. Нацеливающийся вектор также содержит 5'- и 3'-концевые последовательности грызуна, фланкирующие человеческую нуклеотидную последовательность, подлежащую интеграции, также известные как плечи гомологии, которые опосредуют гомологичную рекомбинацию и интеграцию человеческой нуклеотидной последовательности в целевой локус грызуна (например, локус Il1rl2 или локус, в котором расположены гены Il1f6, Il1f8 и Il1f9 грызуна). В некоторых вариантах осуществления 5'- и 3'-концевые фланкирующие последовательности грызуна представляют собой нуклеотидные последовательности, которые фланкируют соответствующую нуклеотидную последовательность грызуна в целевом локусе грызуна, которая подлежит замене на человеческую нуклеотидную последовательность. Например, в вариантах осуществления, где нуклеотидная последовательность, кодирующая эктодомен грызуна (например, экзоны 3-8 гена Il1rl2 грызуна), заменена на последовательность, кодирующую человеческий эктодомен (например, экзоны 3-8 человеческого гена IL1RL2), 5'-фланкирующая последовательность может содержать экзоны 1-2 гена Il1rl2 грызуна, а 3'-фланкирующая последовательность может содержать оставшиеся экзоны ниже экзона 8 гена Il1rl2 грызуна. В некоторых вариантах осуществления, где нуклеотидная последовательность грызуна, кодирующая все три лиганда IL-36, подлежит замене на человеческую нуклеотидную последовательность, 5'-фланкирующая последовательность может содержать нуклеотидную последовательность грызуна выше кодирующей последовательности гена Il1f6, а 3'-фланкирующая последовательность может содержать нуклеотидную последовательность грызуна выше кодирующей последовательности гена Il1f9.
[0111] В некоторых вариантах осуществления нацеливающийся вектор содержит селектируемый маркерный ген. В некоторых вариантах осуществления нацеливающийся вектор содержит один или более сайтов сайт-специфической рекомбинации. В некоторых вариантах осуществления нацеливающийся вектор содержит селектируемый маркерный ген, фланкированный сайтами сайт-специфической рекомбинации таким образом, что селектируемый маркерный ген может быть удален в результате рекомбинации между сайтами.
[0112] В иллюстративных вариантах осуществления клон бактериальной искусственной хромосомы (BAC), несущий геномный фрагмент грызуна, можно модифицировать с применением бактериальной гомологичной рекомбинации и технологии VELOCIGENE® (см., например, U.S. 6586251 и Valenzuela et al. (2003) Nature Biotech. 21(6):652-659; которые включены в данный документ посредством ссылки во всей своей полноте). В результате геномная последовательность грызуна удаляется из исходного клона ВАС и вставляется человеческая нуклеотидная последовательность, что приводит к получению модифицированного клона ВАС, несущего человеческую нуклеотидную последовательность, фланкированную 5'- и 3'-плечами гомологии грызуна. В некоторых вариантах осуществления человеческая нуклеотидная последовательность может представлять собой последовательность кДНК или человеческую геномную ДНК, кодирующую (i) человеческий IL1RL2 полностью или частично (например, эктодомен человеческого белка IL1RL2), или (ii) все три человеческих IL1F6, IL1F8 и IL1F9. После линеаризации модифицированный клон BAC можно вводить в эмбриональные стволовые (ES) клетки грызуна.
[0113] В некоторых вариантах осуществления в настоящем изобретении предусмотрено применение нацеливающегося вектора, описанного в данном документе, для получения модифицированной эмбриональной стволовой (ES) клетки грызуна. Например, нацеливающийся вектор можно вводить в ES-клетку грызуна, например, с помощью электропорации. Как мышиные ES-клетки, так и крысиные ES-клетки были описаны в данной области техники. См., например, US 7576259, US 7659442, US 7294754 и US 2008-0078000 A1 (все из которых включены в настоящий документ посредством ссылки во всей своей полноте), в которых описаны мышиные ES-клетки и способ VELOCIMOUSE® для получения генетически модифицированной мыши; US 2014/0235933 A1 (Regeneron Pharmaceuticals, Inc.), US 2014/0310828 A1 (Regeneron Pharmaceuticals, Inc.), Tong et al. (2010) Nature 467:211-215, и Tong et al. (2011) Nat Prot°C. 6(6): doi:10.1038/nprot.2011.338 (все из которых включены в настоящий документ посредством ссылки во всей своей полноте), в которых описаны крысиные ES-клетки и способы получения генетически модифицированной крысы, которые можно применять для получения модифицированного эмбриона грызуна, которого, в свою очередь, можно применять для получения животного-грызуна.
[0114] В некоторых вариантах осуществления можно отбирать ES-клетки, содержащие человеческую нуклеотидную последовательность, интегрированную в геном. В некоторых вариантах осуществления ES-клетки выбраны на основании анализов потери аллеля грызуна и/или приобретения человеческого аллеля. В некоторых вариантах осуществления отобранные ES-клетки затем применяют в качестве донорных ES-клеток для инъекции в эмбрион на стадии развития, предшествующей моруле (например, эмбрион на стадии 8 клеток), посредством применения способа VELOCIMOUSE® (см., например, US 7576259, US 7659442, US 7294754 и US 2008-0078000 A1, все из которых включены в данный документ посредством ссылки во всей своей полноте) или способов, описанных в US 2014/0235933 A1 и US 2014/0310828 A1, оба из которых включены в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления эмбрион, содержащий донорные ES-клетки, инкубируют до стадии бластоцисты и затем имплантируют суррогатной матери для получения грызуна F0, полностью полученного из донорных ES-клеток. Детенышей грызунов, несущих человеческую нуклеотидную последовательность, можно идентифицировать посредством генотипирования ДНК, выделенной из отрезанных фрагментов хвостов, с использованием анализов потери аллеля грызуна и/или приобретения человеческого аллеля.
[0115] В некоторых вариантах осуществления грызунов, гетерозиготных по гуманизированному гену, можно скрещивать с получением гомозиготных грызунов.
Способы использования четырехкратно гуманизированных грызунов
[0116] Грызуны, раскрытые в данном документе, обеспечивают полезную in vivo систему и источник биологических материалов для идентификации и тестирования соединений, пригодных для лечения заболевания или состояния, ассоциированного с нарушением регуляции передачи сигнала IL-36.
[0117] Под «заболеванием, ассоциированным с нарушением регуляции передачи сигнала IL-36», подразумевается заболевание, при котором проявляется аномальная передача сигнала IL-36, которая может непосредственно или опосредованно вызывать заболевание или обострять симптомы заболевания. Неограничивающие примеры заболеваний, ассоциированных с нарушением регуляции передачи сигнала IL-36, включают генерализованный пустулезный псориаз (GPP или DITRA) (Marrakchi S. et al., N Engl J. Med. 365:620-628 (2011) и Onoufriadis A. et al., Am J. Hum Genet 89:432-437 (2011), которые включены в данный документ посредством ссылки во всей своей полноте), ладонно-подошвенный пустулезный псориаз (PPPP) (Bissonnette R. et al., PLoS One 11:e0155215 (2016), который включен в данный документ посредством ссылки во всей своей полноте), воспалительное заболевание кишечника (IBD) (Medina-Contreras et al., J Immunol 196:34-38 (2016; Nishida A. et al., Inflamm Bowel Dis 22:303-314 (2016) и Russell SE et al., Mucosal Immunol. 9:1193-1204 (2016), которые включены в данный документ посредством ссылки во всей своей полноте), ревматоидный и псориатический артрит (Frey S. et al., Ann Rheum Dis 72:1569-1574 (2013), который включен в данный документ посредством ссылки во всей своей полноте), астму, хроническое обструктивное заболевание легких (Chen H. et al., J. Proteomics 75:2835-2843 (2012), который включен в данный документ посредством ссылки во всей своей полноте), хроническое заболевание почек (Shaik Y. et al., Int J Immunopathol Pharmacol 26:27-36 (2013), который включен в данный документ посредством ссылки во всей своей полноте) и ихтиоз (Paller AS et al., J Allergy Clin Immunol 139:152-165 (2017), который включен в данный документ посредством ссылки во всей своей полноте).
[0118] В некоторых вариантах осуществления соединения, которые можно оценивать с использованием раскрытых грызунов, включают кандидатные ингибиторы передачи сигнала IL-36, например без ограничения низкомолекулярный ингибитор, ингибитор на основе нуклеиновой кислоты (например, siRNA, рибозим, антисмысловая конструкция и т. д.), антигенсвязывающий белок (например, антитело или его антигенсвязывающий фрагмент) или блокирующий пептид/пептидный ингибитор.
[0119] В некоторых вариантах осуществления кандидатный ингибитор представляет собой антитело или его антигенсвязывающий фрагмент. Как моноклональные, так и поликлональные антитела подходят для тестирования на грызунах, раскрытых в данном документе. В некоторых вариантах осуществления антитело специфически связывается с человеческим белком IL-36R. В некоторых вариантах осуществления антитело специфически связывается с субъединицей IL1RL2 человеческого белка IL-36R.
[0120] Соединения-кандидаты можно оценивать посредством индукции воспаления, например, IMQ-индуцированного воспаления кожи или DSS-индуцированного воспаления кишечника, у грызуна, раскрытого в данном документе, и определения того, может ли соединение-кандидат лечить или подавлять индуцированное воспаление. Термин «лечение» или «подавление» включает уменьшение степени тяжести, замедление прогрессирования, устранение, отсрочку или предупреждение возникновения индуцированного воспаления и симптомов, или их комбинацию.
[0121] В некоторых вариантах осуществления грызуну вводят соединение-кандидат до, во время или после введения средства, которое индуцирует воспаление. Дозы соединений-кандидатов можно вводить любым требуемым путем введения, включая парентеральный и непарентеральный пути введения. Парентеральные пути включают, например, внутривенный, внутриартериальный, интрапортальный, внутримышечный, подкожный, интраперитонеальный, интраспинальный, интратекальный, интрацеребровентрикулярный, интракраниальный, интраплевральный или другие пути инъекции. Непарентеральные пути включают, например, пероральный, интраназальный, трансдермальный, внутрилегочный, ректальный, трансбуккальный, вагинальный, внутриглазной. Введение также можно осуществлять путем непрерывной инфузии, местного введения, замедленного высвобождения из имплантатов (гелей, мембран и т.п.) и/или внутривенной инъекции.
[0122] В некоторых вариантах осуществления подходящие контрольные грызуны могут включать, например, гуманизированных грызунов, не подвергнутых индуцированному воспалению; гуманизированных грызунов, подвергнутых индуцированному воспалению без какого-либо соединения или с контрольным соединением, не обладающим какой-либо терапевтической эффективностью (например, антителом изотипического контроля); и гуманизированных грызунов, подвергнутых индуцированному воспалению, с применением соединения, которое, как известно, является терапевтически эффективным.
[0123] Для оценки эффективности соединения-кандидата при воспалении кожи соединение можно вводить грызуну до, во время или после обработки с помощью IMQ. В конкретных вариантах осуществления соединение-кандидат вводят подкожно в область кожи или рядом с ней, на которую наносят IMQ. Соединение считается эффективным, если оно подавляет воспаление кожи по сравнению с контрольными грызунами, которым не вводили соединение. Например, соединение считается эффективным, если общий показатель патологии снижается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%); или если концентрация(концентрации) одного или нескольких провоспалительных медиаторов снижается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%).
[0124] Для оценки эффективности соединения-кандидата при воспалении толстой кишки соединение можно вводить грызуну до, во время или после обработки с помощью DSS или оксазолона. В некоторых вариантах осуществления соединение-кандидат вводят интраперитонеально через некоторое количество дней (например, 5, 6, 7, 8 или 9 дней) после начала обработки с помощью DSS. В некоторых вариантах осуществления соединение-кандидат вводят интраперитонеально множество раз во время обработки оксазолоном; например, через 2-3 дня после местного нанесения оксазолона, при одном или более интраректальных введениях оксазолона (например, при первом интраректальном введении оксазолона и при третьем интраректальном введении оксазолона). Соединение считается эффективным, если оно подавляет колит по сравнению с контрольными грызунами DITRA, которым не вводили соединение. Например, соединение считается эффективным, если общий показатель патологии снижается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%); если концентрация одного или более провоспалительных медиаторов снижается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%); если уровень фекального Lcn2 снижается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%); если активность MPO в гомогенатах толстой кишки снижается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%); если длина толстой кишки увеличивается на 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше (например, снижается на 20%-90%, 20%-80%, 20%-70%, 20%-60%, 20%-50%, 20%-40%, 30%-90%, 30%-80%, 30%-70%, 30%-60%, 30%-50% или 30%-40%); или при их комбинации.
[0125] Настоящее описание дополнительно проиллюстрировано следующими примерами, которые не следует понимать как ограничивающие каким-либо образом. Содержание всех приведенных ссылок (включая ссылки на литературу, выданные патенты и опубликованные заявки на патенты, приводимые в настоящей заявке) явным образом включено в данный документ посредством ссылки во всей своей полноте.
Пример 1. Получение линии четырехкратно гуманизированных мышей
[0126] Две генетически сконструированные линии мышей получали с использованием технологии VelociGene® (Poueymirou et al., Nat Biotechnol. 2007 Jan; 25(1): 91-9; Valenzuela et al., Nat Biotechnol 2003 Jun; 21:652-59, которые включены в данный документ посредством ссылки во всей своей полноте): линию однократно гуманизированных мышей с гуманизированным Il1rl2 и линию трехкратно гуманизированных мышей с гуманизированными Il1f6, Il1f8, Il1f9. «Четырехкратно гуманизированную» линию получали посредством скрещивания однократно гуманизированных и трехкратно гуманизированных линий друг с другом.
Получение линии однократно гуманизированных мышей с гуманизированным Il1rl2
[0127] Получали «однократно гуманизированную» линию, в которой часть мышиного гена Il1rl2, кодирующую внеклеточный домен мышиного Il1rl2 (белок 2, подобный рецептору интерлейкина-1) заменяли на фрагмент человеческого гена IL1RL2, кодирующий соответствующий внеклеточный домен человеческого IL1RL2 (экзоны 3-8 Il1rl2/IL1RL2 с промежуточными интронами и частями фланкирующих интронов соответственно) (фиг. 1A). Полученный гуманизированный ген кодирует химерный рецептор, который поддерживает внутриклеточную сигнальную специфичность мышей, при этом делая внеклеточный домен человеческим и способным связывать человеческие лиганды IL1F6, IL1F8 и IL1F9, также называемые IL36A, B и G соответственно. Гомозиготных мышей с гуманизированным Il1rl2 обозначают как Il1rl2hu/hu.
[0128] Более конкретно, использовали клон RP23-235L22 мышиной бактериальной искусственной хромосомы (ВАС), содержащий мышиный ген Il1rl2, и модифицировали его следующим образом с получением нацеливающегося вектора. Получали фрагмент ДНК, содержащий 5'-концевую мышиную гомологичную нуклеотидную последовательность, человеческую геномную ДНК IL1RL2 размером приблизительно 32389 п.о. (содержащую 346 п.о. на 3'-конце интрона 2, экзоны 3-8 со всеми промежуточными интронами и 1101 п.о. на 5'-конце интрона 8), самоудаляющуюся кассету устойчивости к неомицину размером приблизительно 4996 п.о. и 3'-концевую гомологичную последовательность мыши (фиг. 1B). Этот фрагмент ДНК применяли для модификации клона BAC RP23-235L22 посредством гомологичной рекомбинации в бактериальных клетках. В результате мышиный геномный фрагмент Il1rl2, кодирующий эктодомен (размером приблизительно 25324 п.о., включая 155 п.о. на 3'-конце мышиного интрона 2, экзоны 3-8 со всеми промежуточными интронами и 642 п.о. на 5'-конце мышиного интрона 8), в клоне BAC заменяли на человеческую геномную ДНК IL1RL2, за которой следовала самоудаляющаяся кассета устойчивости к неомицину (фиг. 1B). Полученный модифицированный клон BAC содержал в направлении от 5' к 3' (i) мышиное 5'-плечо гомологии, содержащее приблизительно 112,2 т.п.о. мышиной геномной ДНК, экзоны 1-2 мышиного Il1rl2 и 5'-концевую часть интрона 2; (ii) человеческий геномный фрагмент IL1RL2, включая 3'-концевую часть интрона 2, экзоны 3-8 и 5'-концевую часть интрона 8; (iii) самоудаляющуюся кассету устойчивости к неомицину размером приблизительно 4996 п.о., после которой следует (iv) мышиное 3'-плечо гомологии размером 31,3 т.п.о., содержащее оставшиеся мышиные экзоны Il1rl2 ниже экзона 8, все промежуточные интроны и 3'-UTR. См. фиг. 1B. Соединительные последовательности также представлены в нижней части фиг. 1B.
[0129] Модифицированный клон ВАС, содержащий гуманизированный ген Il1rl2, описанный выше, использовали в качестве нацеливающегося вектора для электропорации мышиных эмбриональных стволовых клеток линии F1H4 (50% C57BL/6NTac/50%129S6/SvEvTac) для получения модифицированных ES-клеток, содержащих гуманизированный ген Il1rl2. Целевые ES-клетки, положительные по содержанию гуманизированного гена Il1rl2, идентифицировали с помощью анализа (Valenzuela et al., выше), посредством которого выявляли наличие человеческих последовательностей IL1RL2 и подтверждали потерю и/или сохранение мышиных последовательностей Il1rl2. Праймеры и зонды, которые использовали для подтверждения гуманизации, описанной выше, представлены в таблице 5. Как только необходимый целевой клон ES-клетки выбран, кассета для отбора по устойчивости к неомицину может быть вырезана. Локус гуманизированного Il1rl2 после делеции кассеты изображен на фиг. 1C, при этом соединительные последовательности также показаны на фиг. 1C.
Таблица 5
[0130] Отобранные клоны ES-клеток вводили путем микроинъекции в 8-клеточные эмбрионы мышей-альбиносов из Charles River Laboratories Swiss Webster с получением VelociMice® F0, которые на 100% происходили из целевых клеток (Poueymirou et al. 2007, выше). В случае мышей, несущих гуманизированный локус, снова проводили подтверждение и идентификацию посредством генотипирования ДНК, выделенной из отрезанных фрагментов хвостов, с применением модификации анализа аллелей (Valenzuela et al., выше), который позволяет выявить наличие последовательностей человеческих генов. Животных, гомозиготных по гуманизированному локусу, получали посредством скрещивания гетерозиготных животных.
[0131] Выравнивание аминокислотных последовательностей полученного гуманизированного/химерного рецептора Il1rl2 (SEQ ID NO: 7), мышиного белка Il1rl2 (SEQ ID NO: 4) и человеческого белка IL1RL2 (SEQ ID NO: 2) представлено на фиг. 1D.
Получение линии трехкратно гуманизированных мышей
[0132] Получали «трехкратно гуманизированную линию», в которой полная кодирующая последовательность каждого из мышиных генов Il1f6, Il1f8 и Il1f9 была заменена на полную кодирующую последовательность каждого из человеческих генов IL1F6, IL1F8 и IL1F9 соответственно. Эта стратегия привела к получению гуманизированных генов, кодирующих человеческие лиганды, которые способны связывать человеческий внеклеточный домен химерного рецептора Il1rl2. Гомозиготных мышей с гуманизированными Il1f6, Il1f8, Il1f9 обозначают как Il1f6hu/hu, Il1f8hu/hu, Il1f9hu/hu.
[0133] Более конкретно, использовали клон RP23-90G23 мышиной бактериальной искусственной хромосомы (ВАС), содержащий мышиные гены Il1f8, Il1f9 и Il1f6 и межгенные последовательности, и модифицировали его следующим образом с получением нацеливающегося вектора. Получали фрагмент ДНК, содержащий 5'-концевую мышиную гомологичную нуклеотидную последовательность, человеческую геномную ДНК размером приблизительно 88868 п.о. (содержащую промоторы, нетранслируемые области и кодирующие последовательности человеческих IL1F8, IL1F9 и IL1F6), самоудаляющуюся кассету устойчивости к гигромицину размером приблизительно 5218 п.о. и мышиную 3'-концевую гомологичную последовательность (фиг. 2B). Этот фрагмент ДНК применяли для модификации клона BAC RP23-90G23 посредством гомологичной рекомбинации в бактериальных клетках. В результате мышиный геномный фрагмент размером приблизительно 76548 п.о. в клоне ВАС заменяли на человеческую геномную ДНК, после которой следовала самоудаляющаяся кассета (фиг. 2A-2B). Полученный модифицированный клон ВАС содержал в направлении от 5 'к 3' (i) мышиное 5'-плечо гомологии, содержащее приблизительно 5,7 т.п.о. мышиной геномной ДНК, (ii) человеческий геномный фрагмент размером приблизительно 88868 п.о. (содержащий промоторы, нетранслируемые области и кодирующие последовательности человеческих IL1F8, IL1F9 и IL1F6), (iii) самоудаляющуюся кассету устойчивости к гигромицину размером приблизительно 5218 п.о. и (iv) мышиную 3'-концевую гомологичную последовательность размером приблизительно 29,7 т.п.о. См. фиг. 2B. Соединительные последовательности также представлены в нижней части фиг. 2B.
[0134] Модифицированный клон BAC, содержащий последовательности человеческих генов IL1F8, IL1F9 и IL1F6, описанные выше, использовали в качестве нацеливающегося вектора для электропорации мышиных эмбриональных стволовых клеток линии F1H4 (50% C57BL/6NTac/50%129S6/SvEvTac) для получения модифицированных ES-клеток, содержащих последовательности человеческих генов IL1F8, IL1F9 и IL1F6. Целевые ES-клетки, положительные по содержанию последовательности человеческих генов IL1F8, IL1F9 и IL1F6, идентифицировали с помощью анализа (Valenzuela et al., выше), который позволил выявить наличие человеческих последовательностей и подтвердить потерю и/или сохранение мышиных последовательностей. Праймеры и зонды, которые использовали для подтверждения гуманизации, описанной выше, представлены в таблице 6. Как только необходимый целевой клон ES-клетки выбран, кассета для отбора по устойчивости к гигромицину может быть вырезана. Гуманизированный локус после делеции кассеты изображен на фиг. 2C, при этом соединительные последовательности также показаны на фиг. 2C.
Таблица 6
[0135] Отобранные клоны ES-клеток вводили путем микроинъекции в 8-клеточные эмбрионы мышей-альбиносов из Charles River Laboratories Swiss Webster с получением VelociMice® F0, которые на 100% происходили из целевых клеток (Poueymirou et al. 2007, выше). В случае мышей, несущих гуманизированный локус, снова проводили подтверждение и идентификацию посредством генотипирования ДНК, выделенной из отрезанных фрагментов хвостов, с применением модификации анализа аллелей (Valenzuela et al., выше), который позволяет выявить наличие последовательностей человеческих генов. Животных, гомозиготных по гуманизированному локусу, получали посредством скрещивания гетерозиготных животных.
Получение линии трехкратно гуманизированных мышей
[0136] «Четырехкратно гуманизированную» линию получали посредством скрещивания однократно гуманизированной и трехкратно гуманизированной линий друг с другом, скрещивание проводили до гомозиготности по обоим локусам на фоне 100% C57BL/6NTac. Гомозиготную четырехкратно гуманизированную линию обозначают как Il1rl2hu/hu Il1f6hu/hu Il1f8hu/hu Il1f9hu/hu или «DITRA-подобная» мышь. Кассеты устойчивости к неомицину и гигромицину удаляли в зародышевой линии F0 с помощью технологии самоудаления.
Пример 2. Характеристика мышей DITRA
Материалы и способы
[0137] Индукция острого и хронического IMQ-индуцированного воспаления кожи и обработка антителами у DITRA-подобных мышей. Для индукции воспаления кожи самкам гуманизированных DITRA-подобных мышей в возрасте 8-10 недель выбривали волосы на спине с использованием триммера для мышиных волос (Oster, MiniMax, № по каталогу 78049-100) и депилировали кожу с использованием 0,5 г геля для удаления волос Veet за три дня до нанесения крема на основе IMQ. Местную ежедневную дозу, составляющую 62,5 мг коммерчески доступного крема на основе IMQ (5%) (Aldara, GM Health Care Limited, NDC 99207-206-12, лот № QJ044A) или вазелина (CVS Pharmacy, NDC 59779-902-88), наносили на выбритую кожу спины мышей в течение четырех последовательных дней для индукции острого заболевания и девяти дней для индукции хронического заболевания. Ежедневную местную дозу, составляющую 62,5 мг Aldara, переводили в суточную дозу, составляющую 3,125 мг активного соединения. При остром IMQ-индуцированном воспалении кожи антитело к человеческому IL-36R вводили подкожно в кожу спины при 10 мг/кг за 3 дня до и через 1 день после начала применения IMQ. Контрольная группа получала инъекции PBS и 10 мг/кг изотипического контроля, представляющего собой hIgG4. При хроническом IMQ-индуцированном воспалении кожи такое же антитело к человеческому IL-36R вводили подкожно в кожу спины при 10 мг/кг терапевтически в дни 4 и 8 после начала применения IMQ. Через два или три дня после обработок кожа спины мышей начала демонстрировать признаки эритемы, шелушения и утолщения. Тяжесть воспаления измеряли ежедневно с использованием адаптированной версии клинического индекса распространенности и тяжести псориаза. Эритему, шелушение и утолщение оценивали независимо по шкале 0-4: 0 - отсутствует; 1 - слабое; 2 - умеренное; 3 - выраженное; и 4 - сильно выраженное (van der Fits et al., J Immunol 2009, 182:5836-5845, который включен в данный документ посредством ссылки во всей своей полноте). В день 4 острого и день 11 хронического IMQ-индуцированного воспаления кожи толщину кожи измеряли с использованием штангенциркуля (Kaefer).
[0138] Гистопатология - образцы ткани кожи диаметром 6 мм с мышиной спины фиксировали в 10% буферном формалине и μзалитые парафином срезы размером 4-5 м окрашивали гематоксилином и эозином. Срезы кожи оценивали вслепую на наличие паракератоза, ортокератоза, микроабсцесса Мунро, акантоза, эпидермального изъязвления, воспаления в дерме и гиподерме, закупорки кровеносных сосудов в дерме и гиподерме, фолликулярного гиперкератоза и эпителиальной гиперплазии. Применяли шкалу для оценивания 0-4: 0 - в пределах нормы, 1 - минимальная, 2 - легкая, 3 - умеренная и 4 - тяжелая степень (van der Fits et al., J Immunol 2009, 182:5836-5845, который включен в данный документ посредством ссылки во всей своей полноте). Общий показатель патологии рассчитывали для каждой мыши посредством сложения отдельных показателей гистопатологических признаков. Анализ данных проводили с использованием программного обеспечения GraphPad Prism™ .
[0139] Измерение уровня цитокинов в гомогенатах кожи - полнослойные образцы ткани кожи диаметром 6 мм с мышиной спины отбирали и помещали в пробирку объемом 15 мл, содержащую буфер T-per (Thermo Scientific, № по каталогу 378510, лот № RF236217), 1x смесь ингибиторов протеаз Halt (Thermo Scientific, № по каталогу 87786, лот № QG221763) и 5М раствор EDTA (Thermo Scientific, Cat3 78429). Образцы ткани кожи разрушали при 28000 об./мин в течение 1 минуты с использованием Polytron (PT10-35 GT-D, № по каталогу 9158158) и помещали на лед. Полученные гомогенаты кожи центрифугировали при 1500 об./мин в течение 8 минут при 4°C и супернатанты собирали в 96-луночные планшеты. Гомогенаты кожи подвергали анализу белка по Брэдфорду с использованием красителя для анализа белка (BioRad, № по каталогу 500-0006, лот № 210008149) для количественного определения общего содержания белка. Концентрации цитокинов в гомогенатах кожи измеряли с применением провоспалительной панели 1 (мышь) набора для мультиплексного иммуноанализа (MesoScale Discovery, № по каталогу K15048D) в соответствии с инструкциями производителя. Вкратце, 50μмкл/лунку калибраторов и образцов (разбавленных в разбавителе Diluent 41) добавляли в планшеты, предварительно покрытые антителами для захвата, и инкубировали при комнатной температуре при встряхивании при 700 об./мин в течение 2 часов. Затем планшеты промывали 3 раза с помощью 1xPBS, содержащего 0,05% (вес/объем) Tween-20, с последующим добавлением 25 мкл раствора антител для выявления, разбавленного в разбавителе Diluent 45. После 2-часовой инкубации при комнатной температуре при встряхивании планшеты промывали 3 раза и в каждую лунку добавляли 150 мкл 2x буфера для считывания. Электрохемилюминесценцию немедленно считывали с помощью прибора MSD Spector®. Анализ данных проводили с использованием программного обеспечения GraphPad Prism™ . Уровни цитокинов нормализовывали по отношению к общему содержанию белка.
[0140] Индукция DSS-индуцированной модели хронического колита и обработка антителами у DITRA-подобных мышей - для индукции хронического DSS-опосредованного колита самкам DITRA-подобных мышей в возрасте 12-20 недель со средним весом тела, составляющим более 23 г, давали 1,5-3% DSS (Sigma-Aldrich № по каталогу 87786, лот № PJ203966B) в питьевой воде в течение 7 дней, затем дистиллированную воду в течение 11-13 дней. Второй цикл обработки с помощью DSS (в течение 4 дней), а затем водой (5-6 дней) осуществляли до дня 27-30. Контрольная группа получала дистиллированную воду на протяжении исследования. Антитела к человеческим IL-36R и mIL-12p40 (Bioxcell, № по каталогу BE0051, клон C17.8) вводили интраперитонеально при дозе 10 мг/кг один раз в две недели, начиная с дня 7. Контрольная группа получала PBS и инъекции соответствующих изотипических контролей, представляющих собой hIgG4 и ratIgG2a (Bioxcell, № по каталогу BE0089, клон 2A3), при дозе 10мг/кг. Мышей взвешивали и наблюдали за клиническими признаками колита (например, консистенцией стула и кровью в фекалиях) на ежедневной основе. В день 27-30 мышей умерщвляли и измеряли длину толстой кишки. Для оценки колита, вызванного DSS, оценивали следующие признаки: воспаление (тяжесть и степень), эпителиальные изменения (эрозия/язва), изменения в криптах (потеря крипт, криптит/абсцесс крипт, регенерация/гиперплазия, потеря бокаловидных клеток), отек подслизистой оболочки и процент площади ткани с патологией относительно общей площади ткани на препарате. Применяли шкалу для оценивания 0-4: 0-0 - в пределах нормы, 1 - минимальная, 2 - легкая, 3 - умеренная и 4 - тяжелая степень. Общий показатель патологии рассчитывали для каждой мыши посредством сложения отдельных показателей гистопатологических признаков.
[0141] Измерение уровня Lcn-2 в образцах фекалий - для мониторинга воспаления кишечника на протяжении всего исследования фекалии отдельных DITRA-подобных мышей собирали в планшеты с глубокими лунками объемом 2 мл на еженедельной основе и хранили при -80°C. По завершении исследования фекалии, собранные в разные дни, подвергали гомогенизации. Вкратце, образцы фекалий восстанавливали с помощью 1 мл PBS, содержащего 0,1% Tween-20, 1x смесь ингибиторов протеаз Halt (Thermo Scientific, № по каталогу 87786, лот № QG221763) и 5M раствор EDTA (Thermo Scientific, № по каталогу 378429). После добавления в лунки 2 шариков из карбида вольфрама размером 3 мм (Qiagen, № по каталогу 69997) планшеты помещали на встряхиватель с максимальной скоростью в течение ночи при 4°C. Гомогенные суспензии фекалий центрифугировали при 1200 об./мин в течение 10 минут при 4°C и супернатанты собирали в 96-луночные планшеты. Уровни фекального липокалина-2 (Lcn2) измеряли с применением набора Duoset Lipocalin-2/NGAL ELISA для мышей (R&D Systems, № по каталогу DY1857, лот № P116359) в соответствии с инструкциями производителя. Анализ данных проводили с использованием программного обеспечения GraphPad Prism™ .
[0142] Измерение активности миелопероксидазы (MPO) в гомогенатах толстой кишки - кусочки дистальной части толстой кишки помещали в микроцентрифужные пробирки объемом 2 мл, содержащие 2 шарика из карбида вольфрама размером 3 мм (Qiagen, № по каталогу 69997), содержащие буфер T-per (Thermo Scientific, № по каталогу 378510, лот № RF236217), 1x смесь ингибиторов протеаз Halt (Thermo Scientific, № по каталогу 87786, лот № QG221763) и 5M раствор EDTA (Thermo Scientific, № по каталогу 78429). Ткани толстой кишки разрушали с применением Qiagen Tissue Lyser II с частотой 27,5 с-1 в течение 10 минут. Пробирки центрифугировали при 1500 об./мин в течение 8 минут при 4°C и супернатанты собирали в 96-луночные планшеты. Гомогенаты толстой кишки подвергали анализу белка по Брэдфорду с использованием красителя для анализа белка (BioRad, № по каталогу 500-0006, лот № 210008149) для количественного определения общего содержания белка. Активность миелопероксидазы (MPO) в гомогенатах толстой кишки измеряли с применением набора MPO ELISA для мышей (Hycult Biotech, № по каталогу HK210-02, лот № 21022KO617-Y) в соответствии с инструкциями производителя. Анализ данных проводили с использованием программного обеспечения GraphPad Prism™ . Уровни MPO нормализовывали по отношению к общему содержанию белка.
[0143] Измерение уровня цитокинов в гомогенатах толстой кишки - концентрации цитокинов в гомогенатах толстой кишки измеряли с применением провоспалительной панели 1 (мышь) набора для мультиплексного иммуноанализа (MesoScale Discovery, № по каталогу K15048D) в соответствии с инструкциями производителя. Вкратце, 50μл/лунку калибраторов и образцов (разбавленных в разбавителе Diluent 41) добавляли в планшеты, предварительно покрытые антителами для захвата, и инкубировали при комнатной температуре при встряхивании при 700 об./мин в течение 2 часов. Затем планшеты промывали 3 раза с помощью 1xPBS, содержащего 0,05% (вес/объем) Tween-20, с последующим добавлением 25 мкл раствора антител для выявления, разбавленного в разбавителе Diluent 45. После 2-часовой инкубации при комнатной температуре при встряхивании планшеты промывали 3 раза и в каждую лунку добавляли 150 мкл 2x буфера для считывания. Электрохемилюминесценцию немедленно считывали с помощью прибора MSD Spector®. Анализ данных проводили с использованием программного обеспечения GraphPad Prism™ . Уровни цитокинов нормализовывали по отношению к общему содержанию белка.
[0144] Статистический анализ - статистическую значимость в пределах групп определяли с помощью однофакторного анализа Anova и апостериорного критерия множественного сравнения Тьюки (*p<0,05, **p<0,005, ***p<0,0005, ****p<0,0001) (одинаково для обоих * - статистическая значимость в группе, обработанной с помощью PBS, и # - статистическая значимость в соответствующей группе, обработанной изотипическим контролем).
Результаты
[0145] Гуманизированные DITRA-подобные мыши проявляют усиленное воспаление кожи в модели IMQ-индуцированного псориаза
[0146] Для исследования роли нарушения регуляции передачи сигнала IL-36R, как и в случае с пациентами, страдающими GPP или DITRA («дефицитом антагониста рецептора интерлейкина тридцать шесть»), получали мышей с гуманизированным IL-36R (например, Il1rl2) и человеческими лигандами IL-36α, β, γ, но без антагониста IL-36Ra, как описано в примере 1. Полученных мышей hIL-36R/hIL-36α, β, γ обозначили как DITRA-подобные мыши в связи с 20-кратным снижением аффинности мышиного IL-36Ra к человеческому IL-36R, приводящим к усилению передачи сигнала IL-36R, как и в случае с пациентами, страдающими DITRA (для более подробного рассмотрения пациентов с DITRA см., например, Marrakchi et al., N Engl J Med 2011, 365:620-628, который включена в данный документ посредством ссылки во всей своей полноте).
[0147] В интактном состоянии у DITRA-подобных мышей не развивались какие-либо спонтанные заболевания. Напротив, в доклинической модели IMQ-индуцированного псориазиформного дерматита, который очень напоминает псориатические поражения у человека по фенотипическим и гистологическим характеристикам (для более подробного рассмотрения характеристик псориазиформного дерматита см. van der Fits et al., J Immunol 2009, 182:5836-5845; Swindell et al., PLoS One 2011, 6: e18266, все из которых включены в данный документ посредством ссылки во всей своей полноте), DITRA-подобные мыши демонстрировали усиленное воспаление кожи по сравнению с их однопометниками WT (фиг. 3A-3B). Вкратце, IMQ наносили ежедневно на выбритую кожу спины DITRA-подобных мышей и мышей WT в течение четырех последовательных дней. В день 5 кожу собирали для последующей гистопатологической оценки, выделения белка и РНК. У DITRA-подобных мышей, обработанных с помощью IMQ, развивались более тяжелые псориатические поражения, такие как шелушение, эритема и утолщение кожи, по сравнению с однопометниками WT (фиг. 3A). В соответствии с клиническими признаками, гистопатологическая оценка кожи позволила выявить более выраженные акантоз, паракератоз и нарушение дифференцировки кератиноцитов у DITRA-подобных мышей (фиг. 3B). DITRA-подобные мыши, обработанные с помощью IMQ, также демонстрировали значительно увеличенное количество абсцессов Мунро или пустул, как и у пациентов с GPP. Более того, нанесение IMQ привело к повышению уровней провоспалительных молекул с нарушением регуляции при псориазе (провоспалительных молекул, включая, например, IL-17a, IL-17f, IL-23a, S100A8, Defb4) в коже DITRA-подобных мышей по сравнению с однопометниками WT (фиг. 3C). Кроме того, экспрессия мРНК IL-36 α, IL-36β, и IL-36γ была одинаково повышена у DITRA-подобных мышей, обработанных с помощью IMQ, и мышей дикого типа, обработанных с помощью IMQ, по сравнению с DITRA-подобными мышами, обработанными вазелином, и мышами дикого типа, обработанными вазелином, соответственно; однако уровни белков цитокинов IL-36α и IL-36β были выявлены в 2 раза более высоких концентрациях в воспаленной коже DITRA-подобных мышей по сравнению с уровнями в воспаленной коже мышей дикого типа.
[0148] Гуманизированные DITRA-подобные мыши демонстрируют дефект заживления слизистой оболочки в DSS-индуцированной модели хронического колита - в последние годы несколько исследований на подгруппах пациентов продемонстрировали нарушение регуляции экспрессии оси IL-36 при IBD и ее возможное участие в воспалении кишечника. Было показано, что экспрессия IL-36α и IL-36γ повышена в воспаленной слизистой оболочке пациентов с язвенным колитом (см. Medina-Contreras et al., J Immunol 2016, 196:34-38; Nishida et al., Inflamm Bowel Dis 2016, 22:303-314, и Russell, Mucosal Immunol 2016, 9:1193-1204, все из которых включены в данный документ посредством ссылки во всей своей полноте). В доклинических моделях дефицит IL-36R защищал от DSS- и оксазолон-индуцированного колита (Medina-Contreras et al., J Immunol 2016, 196:34-38; Harusato et al., Mucosal Immunol 2017, 10:1455-1467, все из которых включены в данный документ посредством ссылки во всей своей полноте). В устойчивом состоянии у DITRA-подобных мышей не развивался спонтанный илеит/колит, но они демонстрировали значительное укорочение толстой кишки как в молодом, так и в старшем возрасте (в возрасте 3 и 10 месяцев соответственно) (фиг. 4A и 4B). Кроме того, в стабильном состоянии и по сравнению с мышами дикого типа DITRA-подобные мыши проявляли повышенные уровни цитокинов IL-36, а также тенденцию к повышению уровней IL-17F и IL-17A и снижению уровней IL-21 в толстой кишке.
[0149] Для оценки роли оси IL-36 в воспалении кишечника применяли химически индуцированную модель повреждения кишечника посредством применения перорального введения DSS, который повреждает эпителий толстой кишки (Okayasu et al., Gastroenterology 1990, 98:694-702, включенный в данный документ посредством ссылки во всей своей полноте) и запускает мощные воспалительные реакции (Rakoff-Nahoum et al., Cell 2004, 118: 229-241, включенный в данный документ посредством ссылки во всей своей полноте) , проявляя основные признаки IBD, в частности язвенного колита. DITRA-подобных мышей и их совместно содержащихся однопометников WT подвергали режиму с DSS-индуцированным хроническим колитом путем введения 2,5% DSS в течение 7 дней с последующим введением воды в течение 11 дней (первый цикл) и 1,5% DSS в течение 4 дней с последующим введением воды в течение 5 дней (второй цикл). В острой фазе заболевания у DITRA-подобных мышей развивалось воспаление кишечника, как и у совместно содержащихся однопометников WT (фиг. 5A). Интересно, что в восстановительной фазе заболевания DITRA-подобные мыши демонстрировали неспособность восстановиться от DSS-индуцированного повреждения слизистой оболочки, что отражалось в устойчивой потере веса тела (фиг. 5A), значительном уменьшении длины толстой кишки (фиг. 5B), показателе более тяжелой патологии (ассоциированной с более тяжелым изъязвлением, обширной эпителиальной эрозией и нейтрофильной инфильтрацией) (фиг. 5C), значительно повышенных уровнях фекального липокалина-2 (Lcn2) и активности миелопероксидазы (фиг. 5D), повышение экспрессии провоспалительных цитокинов, таких как KC-GRO, TNF-α и IL-6, и снижении уровней IL-22 в толстой кишке (фиг. 5E). Кроме того, DITRA-подобные мыши проявляли уровень смертности, составляющий приблизительно 62,5% (данные не показаны). Напротив, мыши WT действительно выздоравливали в восстановительной фазе колита и демонстрировали уровень смертности, составляющий 25%. Целостность эпителия кишки также исследовали путем введения DITRA-подобным мышам и мышам дикого типа (FITC)-декстрана, меченого флуоресцеинизотиоцианатом, с помощью желудочного зонда. Значительное повышение уровней FITC-декстрана наблюдали в сыворотке крови DITRA-подобных мышей в день 14 обработки с помощью DSS, что указывает на то, что путь IL-36 участвует в регуляции проницаемости кишечника во время повреждения кишки. Таким образом, усиление передачи сигнала IL-36 у DITRA-подобных мышей приводит к обострению воспаления кишечника и дефекту восстановления слизистой оболочки, что указывает на роль IL-36 в регуляции механизмов, участвующих в ремоделировании ткани кишечника.
[0150] Моноклональное антитело к человеческому IL-36R подавляет острое воспаление кожи у DITRA-подобных мышей при профилактическом введении доз - для изучения роли IL-36R при воспалении кожи моноклональное антитело к человеческому IL-36R тестировали в IMQ-индуцированной модели псориазиформного дерматита. IMQ наносили ежедневно на выбритую кожу спины DITRA-подобных мышей в течение четырех последовательных дней. Моноклональное антитело к человеческому IL-36R вводили при дозе 10 мг/кг за 3 дня до (-3d) и через один день после (d1) начала нанесения IMQ. Контрольные группы получали инъекции PBS и изотипического контроля hIgG4 при 10мг/кг. В день 5 кожу собирали для последующей гистопатологической оценки и выделения белка. Моноклональное антитело к человеческому IL-36R значительно снижало общий показатель IMQ-индуцированной патологии, включая паракератоз и микроабсцесс Мунро, по сравнению с группами, обработанными PBS и изотипическим контролем (фиг. 6A). Блокада человеческого IL-36R также приводила к снижению уровня KC-GRO, IL-6 и TNF на 66-93% α (фиг. 6B). Важно, что обработка антителом к человеческому IL-36R значительно снижает уровни провоспалительных цитокинов с нарушением регуляции при псориазе (например, IL-12p40, IL-17f и IL-17a) (фиг. 6B), что указывает на строго регулируемое взаимодействие между этими цитокиновыми путями. Наблюдаемая эффективность моноклонального антитела к человеческому IL-36R при купировании острого IMQ-индуцированного воспаления кожи была дозозависимой, при этом доза, составляющая 10 мг/кг, приводила к более сильному подавлению воспаления кожи по сравнению с дозой, составляющей 1 мг/кг, как определено на основании показателя патологии, толщины кожи и уровней провоспалительных цитокинов в гомогенатах кожи (данные не показаны).
[0151] Моноклональное антитело к человеческому IL-36R ингибирует хроническое воспаление кожи при введении терапевтических доз - для дополнительного изучения терапевтической эффективности антагонизма человеческого IL-36R in vivo такое же моноклональное антитело к человеческому IL-36R тестировали в модели хронического IMQ-индуцированного воспаления кожи. На протяжении двух недель IMQ наносили на выбритую кожу спины DITRA-подобных мышей в течение девяти дней в два цикла, разделенных двумя днями без обработок. Моноклональное антитело к человеческому IL-36R вводили подкожно в дни 5 и 9 после начала нанесения IMQ при дозе, составляющей 10 мг/кг. Контрольные группы получали инъекции PBS и изотипического контроля hIgG4 при 10мг/кг. В день 12 измеряли толщину кожи и собирали ткани для последующей гистопатологической оценки и выделения белка. Как и при остром IMQ-индуцированном воспалении, длительное нанесение IMQ приводило к повышению экспрессии провоспалительных медиаторов в коже DITRA-подобных мышей (данные не показаны). Моноклональное антитело к человеческому IL-36R продемонстрировало значительную эффективность при снижении IMQ-индуцированного утолщения кожи и показателей патологического поражения у DITRA-подобных мышей (фиг. 7A и 7B соответственно). Кроме того, введение моноклонального антитела к человеческому IL-36R приводило к значительному ингибированию IMQ-индуцированного продуцирования провоспалительных цитокинов в коже DITRA-подобных мышей (фиг. 7C).
[0152] В целом, данные продемонстрировали профилактическую и терапевтическую эффективность антитела к человеческому IL-36R при уменьшении степени тяжести острого и хронического IMQ-индуцированного воспаления кожи in vivo.
[0153] Моноклональное антитело к человеческому IL-36R уменьшает степень тяжести DSS-индуцированного хронического колита у DITRA-подобных мышей при введении терапевтических доз - после демонстрации дефекта заживления слизистой оболочки в результате усиления передачи сигнала IL-36 у DITRA-подобных мышей дополнительно исследовали, может ли блокада IL-36 спасти наблюдаемый фенотип. DITRA-подобных мышей подвергали хроническому DSS-индуцированному колиту путем введения 3% DSS в течение 7 дней с последующим 13-дневным введением воды в течение двух циклов. Такое же моноклональное антитело к человеческому IL-36R и моноклональное антитело к mIL-12p40 (мышиный заменитель устекинумаба, одобренного для лечения болезни Крона) вводили при дозе 10 мг/кг один раз в две недели, начиная в день 7. Контрольные группы получали инъекции PBS и соответствующих изотипических контролей, представляющих собой hIgG4 и крысиный IgG2a, при 10мг/кг. Обработка моноклональным антителом к человеческому IL-36R избавляла DITRA-подобных мышей от невозможности восстанавливаться после DSS-индуцированного повреждения слизистой оболочки и снижала тяжесть заболевания по сравнению с обработками PBS и изотипическим контролем у DITRA-подобных мышей (фиг. 8A). Для мониторинга воспаления кишечника на разных стадиях заболевания фекалии отдельных мышей собирали на недельной основе для измерения белка фекального липокалина-2 (Lcn2), неинвазивного биомаркера воспаления при повреждении кишечника (процедура описана, например, в Thorsvik et al., J Gastroenterol Hepatol 2017, 32:128-135, который включен в данный документ посредством ссылки во всей своей полноте). Как показано на фиг. 8B, группы, обработанные с помощью PBS, hIgG4 и крысиного IgG2a, демонстрировали значительное повышение уровней фекального Lcn2 в день 17, 23 и 30, по сравнению с только водой. Введения моноклонального антитела к человеческому IL-36R привели к значительному снижению уровней Lcn2 по сравнению с группами, обработанными PBS и изотипическим контролем. Устойчивое снижение уровней фекального Lcn2 наблюдали в группах, обработанных антителом к человеческому IL-36, в день 17, 23 и 30 (фиг. 8B).
[0154] Дополнительные результаты включали то, что блокада hIL-36R моноклональным антителом к человеческому IL-36R приводила к увеличению длины толстой кишки (фиг. 8C) и снижению активности миелопероксидазы (MPO) (фиг. 8D), а также снижению на 61-95% уровней провоспалительных цитокинов (фиг. 8E) в толстой кишке DITRA-подобных мышей, обработанных DSS. Моноклональное антитело к человеческому IL-36R продемонстрировало эффективность, сопоставимую с блокадой IL-12p40 при уменьшении степени тяжести DSS-индуцированного хронического колита, что отразилось в аналогичном снижении уровней фекального Lcn2, активности MPO и провоспалительных цитокинов в толстой кишке DITRA-подобных мышей (фиг. 8A-8E).
[0155] Блокада IL-36R при оксазолон-индуцированном колите - эффективность блокады IL-36R тестировали при оксазолон-индуцированном колите, другой доклинической модели IBD с гистологическим сходством с язвенным колитом человека (Heller et al., Immunity 17, 629-638 (2002), который включен в данный документ посредством ссылки во всей своей полноте). Профилактическое введение антитела к человеческому IL-36R значительно снижало тяжесть оксазолон-индуцированного заболевания у DITRA-подобных мышей по сравнению с группами, обработанными PBS и изотипическим контролем, что отражалось в средней потере веса тела и средней длине толстой кишки (фиг. 9A-9B). Кроме того, антагонизм IL-36R приводил к значительному снижению уровней IL-4 и TNFα в толстой кишке DITRA-подобных мышей, обработанных оксазолоном (фиг. 9C).
[0156] В тексте описания упоминаются различные публикации, в том числе патенты, патентные заявки, опубликованные патентные заявки, номера доступа, технические статьи и научные статьи. Каждая из данных упоминаемых публикаций включена в данный документ посредством ссылки во всей своей полноте и для всех целей.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
<120> Модели заболевания DITRA на животных, отличных от человека, и пути
их применения
<130> 35950PCT (10404WO01)
<150> 62/698,459
<151> 2018-07-16
<150> 62/867,477
<151> 2019-06-27
<160> 57
<170> Версия PatentIn 3.5
<210> 1
<211> 2628
<212> ДНК
<213> Homo sapiens
<400> 1
cccgcccacg gtggcgggga aatacctagg catggaagtg gcatgacagg gctcgtgtcc 60
ctgtcatatt ttccactctc cacgaggtcc tgcgcgcttc aatcctgcag gcagcccggt 120
ttggggatgt ggtccttgct gctctgcggg ttgtccatcg cccttccact gtctgtcaca 180
gcagatggat gcaaggacat ttttatgaaa aatgagatac tttcagcaag ccagcctttt 240
gcttttaatt gtacattccc tcccataaca tctggggaag tcagtgtaac atggtataaa 300
aattctagca aaatcccagt gtccaaaatc atacagtcta gaattcacca ggacgagact 360
tggattttgt ttctccccat ggaatggggg gactcaggag tctaccaatg tgttataaag 420
ggtagagaca gctgtcatag aatacatgta aacctaactg tttttgaaaa acattggtgt 480
gacacttcca taggtggttt accaaattta tcagatgagt acaagcaaat attacatctt 540
ggaaaagatg atagtctcac atgtcatctg cacttcccga agagttgtgt tttgggtcca 600
ataaagtggt ataaggactg taacgagatt aaaggggagc ggttcactgt tttggaaacc 660
aggcttttgg tgagcaatgt ctcggcagag gacagaggga actacgcgtg tcaagccata 720
ctgacacact cagggaagca gtacgaggtt ttaaatggca tcactgtgag cattacagaa 780
agagctggat atggaggaag tgtccctaaa atcatttatc caaaaaatca ttcaattgaa 840
gtacagcttg gtaccactct gattgtggac tgcaatgtaa cagacaccaa ggataataca 900
aatctacgat gctggagagt caataacact ttggtggatg attactatga tgaatccaaa 960
cgaatcagag aaggggtgga aacccatgtc tcttttcggg aacataattt gtacacagta 1020
aacatcacct tcttggaagt gaaaatggaa gattatggcc ttcctttcat gtgccacgct 1080
ggagtgtcca cagcatacat tatattacag ctcccagctc cggattttcg agcttacttg 1140
ataggagggc ttatcgcctt ggtggctgtg gctgtgtctg ttgtgtacat atacaacatt 1200
tttaagatcg acattgttct ttggtatcga agtgccttcc attctacaga gaccatagta 1260
gatgggaagc tgtatgacgc ctatgtctta taccccaagc cccacaagga aagccagagg 1320
catgccgtgg atgccctggt gttgaatatc ctgcccgagg tgttggagag acaatgtgga 1380
tataagttgt ttatattcgg cagagatgaa ttccctggac aagccgtggc caatgtcatc 1440
gatgaaaacg ttaagctgtg caggaggctg attgtcattg tggtccccga atcgctgggc 1500
tttggcctgt tgaagaacct gtcagaagaa caaatcgcgg tctacagtgc cctgatccag 1560
gacgggatga aggttattct cattgagctg gagaaaatcg aggactacac agtcatgcca 1620
gagtcaattc agtacatcaa acagaagcat ggtgccatcc ggtggcatgg ggacttcacg 1680
gagcagtcac agtgtatgaa gaccaagttt tggaagacag tgagatacca catgccgccc 1740
agaaggtgtc ggccgtttcc tccggtccag ctgctgcagc acacaccttg ctaccgcacc 1800
gcaggcccag aactaggctc aagaagaaag aagtgtactc tcacgactgg ctaagacttg 1860
ctggactgac acctatggct ggaagatgac ttgttttgct ccatgtctcc tcattcctac 1920
acctattttc tgctgcagga tgaggctagg gttagcattc tagacaccca gttgagctca 1980
ggcgtagaga agaggaggat gggataagaa ctggggccat ccccatgtca tggtgggtga 2040
gagctggggc catccccgtg gtcatggagg gtgagagctg ggggttatcc ccatggtcat 2100
ggagggtgag ggctggtcgg gggaggcatc cccaagtcat ggtgggtgag agctcggagc 2160
atccccatgt catggtgggt gagatctggg ggtatccctg tgtcatggtg ggtgagggcg 2220
ggtggtcatc cacatggtca tagtgggtga gagctggggg tatccctaca tcatggtggg 2280
tgagagctgg gagcatcccc atgtcatggt gggcgagatc tggggggtat ccccacgtca 2340
tggtggatga gagctggggg aatcaccatg tcatggtggg tgagatcttg ggggatcacc 2400
tgtcatggtg ggtgagagct ggggggatca cctgtcatgg tgggtgagag ttgggattca 2460
tccccatgtc atggtgggct gagcccacat ggaagcctgt gcttggacag cgtatgccct 2520
tttctctgtt tttccacaat gaacaattaa actgtaaatg ttaaaaatat cagtaatttg 2580
tgaaataaat tttattctca tttgagcaac ataaaaaaaa aaaaaaaa 2628
<210> 2
<211> 575
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Met Trp Ser Leu Leu Leu Cys Gly Leu Ser Ile Ala Leu Pro Leu Ser
1 5 10 15
Val Thr Ala Asp Gly Cys Lys Asp Ile Phe Met Lys Asn Glu Ile Leu
20 25 30
Ser Ala Ser Gln Pro Phe Ala Phe Asn Cys Thr Phe Pro Pro Ile Thr
35 40 45
Ser Gly Glu Val Ser Val Thr Trp Tyr Lys Asn Ser Ser Lys Ile Pro
50 55 60
Val Ser Lys Ile Ile Gln Ser Arg Ile His Gln Asp Glu Thr Trp Ile
65 70 75 80
Leu Phe Leu Pro Met Glu Trp Gly Asp Ser Gly Val Tyr Gln Cys Val
85 90 95
Ile Lys Gly Arg Asp Ser Cys His Arg Ile His Val Asn Leu Thr Val
100 105 110
Phe Glu Lys His Trp Cys Asp Thr Ser Ile Gly Gly Leu Pro Asn Leu
115 120 125
Ser Asp Glu Tyr Lys Gln Ile Leu His Leu Gly Lys Asp Asp Ser Leu
130 135 140
Thr Cys His Leu His Phe Pro Lys Ser Cys Val Leu Gly Pro Ile Lys
145 150 155 160
Trp Tyr Lys Asp Cys Asn Glu Ile Lys Gly Glu Arg Phe Thr Val Leu
165 170 175
Glu Thr Arg Leu Leu Val Ser Asn Val Ser Ala Glu Asp Arg Gly Asn
180 185 190
Tyr Ala Cys Gln Ala Ile Leu Thr His Ser Gly Lys Gln Tyr Glu Val
195 200 205
Leu Asn Gly Ile Thr Val Ser Ile Thr Glu Arg Ala Gly Tyr Gly Gly
210 215 220
Ser Val Pro Lys Ile Ile Tyr Pro Lys Asn His Ser Ile Glu Val Gln
225 230 235 240
Leu Gly Thr Thr Leu Ile Val Asp Cys Asn Val Thr Asp Thr Lys Asp
245 250 255
Asn Thr Asn Leu Arg Cys Trp Arg Val Asn Asn Thr Leu Val Asp Asp
260 265 270
Tyr Tyr Asp Glu Ser Lys Arg Ile Arg Glu Gly Val Glu Thr His Val
275 280 285
Ser Phe Arg Glu His Asn Leu Tyr Thr Val Asn Ile Thr Phe Leu Glu
290 295 300
Val Lys Met Glu Asp Tyr Gly Leu Pro Phe Met Cys His Ala Gly Val
305 310 315 320
Ser Thr Ala Tyr Ile Ile Leu Gln Leu Pro Ala Pro Asp Phe Arg Ala
325 330 335
Tyr Leu Ile Gly Gly Leu Ile Ala Leu Val Ala Val Ala Val Ser Val
340 345 350
Val Tyr Ile Tyr Asn Ile Phe Lys Ile Asp Ile Val Leu Trp Tyr Arg
355 360 365
Ser Ala Phe His Ser Thr Glu Thr Ile Val Asp Gly Lys Leu Tyr Asp
370 375 380
Ala Tyr Val Leu Tyr Pro Lys Pro His Lys Glu Ser Gln Arg His Ala
385 390 395 400
Val Asp Ala Leu Val Leu Asn Ile Leu Pro Glu Val Leu Glu Arg Gln
405 410 415
Cys Gly Tyr Lys Leu Phe Ile Phe Gly Arg Asp Glu Phe Pro Gly Gln
420 425 430
Ala Val Ala Asn Val Ile Asp Glu Asn Val Lys Leu Cys Arg Arg Leu
435 440 445
Ile Val Ile Val Val Pro Glu Ser Leu Gly Phe Gly Leu Leu Lys Asn
450 455 460
Leu Ser Glu Glu Gln Ile Ala Val Tyr Ser Ala Leu Ile Gln Asp Gly
465 470 475 480
Met Lys Val Ile Leu Ile Glu Leu Glu Lys Ile Glu Asp Tyr Thr Val
485 490 495
Met Pro Glu Ser Ile Gln Tyr Ile Lys Gln Lys His Gly Ala Ile Arg
500 505 510
Trp His Gly Asp Phe Thr Glu Gln Ser Gln Cys Met Lys Thr Lys Phe
515 520 525
Trp Lys Thr Val Arg Tyr His Met Pro Pro Arg Arg Cys Arg Pro Phe
530 535 540
Pro Pro Val Gln Leu Leu Gln His Thr Pro Cys Tyr Arg Thr Ala Gly
545 550 555 560
Pro Glu Leu Gly Ser Arg Arg Lys Lys Cys Thr Leu Thr Thr Gly
565 570 575
<210> 3
<211> 4072
<212> ДНК
<213> Mus musculus
<400> 3
ggcggacaga gaccggaagc tgggagatga tggcttctga tgtttgttgc ttacgcgaca 60
cagcttgttt ggaattatgt ggcccttgac aactgcgtgg cacacacttc aagatacaaa 120
agtgagaaga agaggggggt caccagagga gactgaagtc agtaagtttc taccaaagct 180
gggtttcttc tggccactcc aaacaaagct gaaattgtcc ttcagacctc tcctgagatg 240
ggggttacat ctttgctctt ctgtggggtg tttttcctgc ttctgctttt cgtggcagca 300
gatacgtgtg aggacatttt tatgcacaat gtgataattt cagagggcca gccttttcct 360
ttcaactgca catacccgcc agaaacaaac ggggcagtaa atctgacatg gtacaaaaca 420
cctagcaaaa gcccagtatc taacaacaga caccttagag ttcaccagga ccagacctgg 480
atcttgtttc ttccattgac actggaggac tccggtatct atcagtgtgt tataaggaat 540
gcccacaact gctaccaaat agctgtgaac ctaaccgttt taaaaaacca ctggtgtgac 600
tcttccatgg aggggagtcc cgtaaattca ccagatgtgt accagcaaat attacccata 660
ggaaaatcgg gcagtctgaa ttgtcatctc tacttcccag aaagttgtgc tttggattca 720
ataaaatggt ataagggttg tgaagagatt aaagcgggga aaaagtacag cccttcagga 780
gcaaagcttc ttgtgaacaa cgttgctgtg gaggacggcg ggagctatgc gtgctcagcc 840
agactgactc acttggggag acacttcacc attagaaact acattgctgt gaacaccaag 900
gaagttgagt atggaagaag gatccctaac atcacgtatc caaagaacaa ctccattgaa 960
gttccacttg gctccaccct catcgtgaac tgcaatataa cagacacgaa ggagaataca 1020
aacctgaggt gctggagagt caacaacacc ctggtggatg actactacaa agactccaaa 1080
cgcatccagg aaggaatcga aaccaatgtg tccttgaggg atcaaattcg gtacacagtg 1140
aacataacat tcttaaaagt gaaaatggag gactacggcc gtcctttcac gtgtcatgct 1200
ggagtgtccg cagcctacat cattctgata tacccagttc cagacttcag ggcttacctc 1260
ttaggagggc ttatggcgtt cctacttctg gttgtatctg ttctgttcat ctacaacagc 1320
tttaagatcg acatcatgct ttggtacaga agcgccttcc acactgccca ggctccagat 1380
gatgagaagc tgtatgatgc ctatgtctta taccccaagt acccgagagg aagccagggc 1440
catgatgtag acacactggt actgaagatc ttgcccgagg tgctggagaa acagtgtgga 1500
tataagttat ttatatttgg cagggatgaa ttccctggac aagctgtggc cagcgtcatt 1560
gatgaaaaca ttaagctgtg taggaggctg atggtctttg tggcaccaga gtcgtctagc 1620
ttcggctttc taaagaactt gtcagaagaa caaattgccg tctacaatgc cctcatccag 1680
catggcatga aggtcattct gatcgaactg gagaaagtca aagactacag caccatgccc 1740
gagtccattc agtatatccg acagaagcac ggggccatcc aatgggatgg ggacttcaca 1800
gagcagtcac agtgtgccaa gaccaagttc tggaagaaag tgagatacca tatgccaccc 1860
aggaggtatc cagcatcttc cccagtccag ctgctgggac acataccctg caattgcaag 1920
gcaggcaaat gcaatgctgc cacagggctc ataactccct gagagtggtt agtgtgtgtt 1980
ggctcacact acatcctctc tgaattgtct actcatgtag ctggctcttt tgtgcttgtg 2040
agcgaccttg tccttgccct tgtagctttt tgtttatttg tattgtttgc tggatgcttt 2100
cagtcatagc tgatccttat tactcctgtt tgcttcatgg ctcctgaaaa cccatatact 2160
ccccaagcat ggggtggctc taacgttggg cctccgtgtc agtgggatac tgaggatgac 2220
agatcaggtg tccaggatgc ttgatcctgg tttgagatgg gaggctgggc agtctgagga 2280
taccaagtag atagccttag ggaagatcag caaggacagt agtgcaatgt ctgtgtttgt 2340
ccagggtcca caagattcta gtcctcaggc agtgtagact gcatttgaat ctccttcatt 2400
ttcctattgc atttagttaa ctaaccattg tgagaacagg acagaggact gggagttggg 2460
ggtgggggga ggtgacagag gatgggagtg tgtgtgtaag aggtgacagt tacatgatca 2520
atcctgggaa atcagactca ggaaaccagc attagggtgc agatctgagt tttttgtgca 2580
gcccttaagc ttatatacag cgttcgagcc aatcccaagc ccctaaaccc tcggccaatc 2640
atatcttgcc acatcatcac tatgcctcaa tgaaagccct gcctgatgag gtaactcaaa 2700
aggaattggg gtgggaagca tcatcccctg tcagaacttc cgtgaagatg gaggacgtag 2760
ccactacccc acctcagttt ctttttgttg tctcaggttt gcaccagagc catcttaaat 2820
ctagtgacat gtgcagcaaa tcatgactct tcaaggagct gcgggagcct gtggtgcctc 2880
gctctccatc ccactttttc cttcacaggg ttgacttcca cgtcccccaa accatggtat 2940
tagcacgatg cccctagaag ccgcttagat gtcgccagga ccaatggtct tcatttccct 3000
ctttcccagc ccctgtgaat cttactattc taggaaattc aaatgagaga cacggtgcat 3060
tcacggtttc tcattttatg actggcttct ttcatacacc acggatttct caagtgctct 3120
cgagattcat ccattgatta gccttttgtt ttattattat cactactgtc tttctcttct 3180
ttttcttcct ctttaaattt ctccccctcc tcacgagcat ccggagagga gcatagactc 3240
tctgcatgcc tacagcgtga tgagctcacc gagggtccct tcaatcacaa gcttgtatgt 3300
tttgcttttc ggtttctctt tgcaccatgg gaattgtaac tccttacaga ggatttgaag 3360
gttcgggtac aaaggcgtca ctgactggtt ccttcttctg tcttcctcac cctgaatggg 3420
ctttgcttct gggtgattcc tggaaattgt cagcagtgat ggagaaacct agggacaggg 3480
ctgactaatg gggcctggcc tttcttcttt gaagcccagg acacagtgcc tgcctagtgg 3540
gtcctggaca caccaggtgg ttccactctg ctcagtggta acatttttta gctgagtcat 3600
caacatcggg tcctttagtg aggaggtgcg ggcatctctc cccagttaga tttggactct 3660
tacttccctg ctcatttccc tttgaaagat tcaggagact atgtcagaag cagggagcca 3720
gcaccctgtg tcttctccac gtggctcctt gtcacacctg tgaccaccaa ttttagaaaa 3780
tgtgtttcct tgttctgacc acacttatgc ggacattagt tgaggctgtt gtcagaaagg 3840
aaaattaact catttgaagt aaacattttc tggaaatctc tgacttctgt caggccaatg 3900
gtgctgaaaa atttaaaact gaattcactt ttttggccag aaagctttcc tcttgccaac 3960
atgtcttaat tttccttttg cttaaactta ttggagatgt attatgaatg ctaagatgtg 4020
agtgtgttat atcagtacat tgacatttta aataaagtat attttaataa aa 4072
<210> 4
<211> 574
<212> БЕЛОК
<213> Mus musculus
<400> 4
Met Gly Val Thr Ser Leu Leu Phe Cys Gly Val Phe Phe Leu Leu Leu
1 5 10 15
Leu Phe Val Ala Ala Asp Thr Cys Glu Asp Ile Phe Met His Asn Val
20 25 30
Ile Ile Ser Glu Gly Gln Pro Phe Pro Phe Asn Cys Thr Tyr Pro Pro
35 40 45
Glu Thr Asn Gly Ala Val Asn Leu Thr Trp Tyr Lys Thr Pro Ser Lys
50 55 60
Ser Pro Val Ser Asn Asn Arg His Leu Arg Val His Gln Asp Gln Thr
65 70 75 80
Trp Ile Leu Phe Leu Pro Leu Thr Leu Glu Asp Ser Gly Ile Tyr Gln
85 90 95
Cys Val Ile Arg Asn Ala His Asn Cys Tyr Gln Ile Ala Val Asn Leu
100 105 110
Thr Val Leu Lys Asn His Trp Cys Asp Ser Ser Met Glu Gly Ser Pro
115 120 125
Val Asn Ser Pro Asp Val Tyr Gln Gln Ile Leu Pro Ile Gly Lys Ser
130 135 140
Gly Ser Leu Asn Cys His Leu Tyr Phe Pro Glu Ser Cys Ala Leu Asp
145 150 155 160
Ser Ile Lys Trp Tyr Lys Gly Cys Glu Glu Ile Lys Ala Gly Lys Lys
165 170 175
Tyr Ser Pro Ser Gly Ala Lys Leu Leu Val Asn Asn Val Ala Val Glu
180 185 190
Asp Gly Gly Ser Tyr Ala Cys Ser Ala Arg Leu Thr His Leu Gly Arg
195 200 205
His Phe Thr Ile Arg Asn Tyr Ile Ala Val Asn Thr Lys Glu Val Glu
210 215 220
Tyr Gly Arg Arg Ile Pro Asn Ile Thr Tyr Pro Lys Asn Asn Ser Ile
225 230 235 240
Glu Val Pro Leu Gly Ser Thr Leu Ile Val Asn Cys Asn Ile Thr Asp
245 250 255
Thr Lys Glu Asn Thr Asn Leu Arg Cys Trp Arg Val Asn Asn Thr Leu
260 265 270
Val Asp Asp Tyr Tyr Lys Asp Ser Lys Arg Ile Gln Glu Gly Ile Glu
275 280 285
Thr Asn Val Ser Leu Arg Asp Gln Ile Arg Tyr Thr Val Asn Ile Thr
290 295 300
Phe Leu Lys Val Lys Met Glu Asp Tyr Gly Arg Pro Phe Thr Cys His
305 310 315 320
Ala Gly Val Ser Ala Ala Tyr Ile Ile Leu Ile Tyr Pro Val Pro Asp
325 330 335
Phe Arg Ala Tyr Leu Leu Gly Gly Leu Met Ala Phe Leu Leu Leu Val
340 345 350
Val Ser Val Leu Phe Ile Tyr Asn Ser Phe Lys Ile Asp Ile Met Leu
355 360 365
Trp Tyr Arg Ser Ala Phe His Thr Ala Gln Ala Pro Asp Asp Glu Lys
370 375 380
Leu Tyr Asp Ala Tyr Val Leu Tyr Pro Lys Tyr Pro Arg Gly Ser Gln
385 390 395 400
Gly His Asp Val Asp Thr Leu Val Leu Lys Ile Leu Pro Glu Val Leu
405 410 415
Glu Lys Gln Cys Gly Tyr Lys Leu Phe Ile Phe Gly Arg Asp Glu Phe
420 425 430
Pro Gly Gln Ala Val Ala Ser Val Ile Asp Glu Asn Ile Lys Leu Cys
435 440 445
Arg Arg Leu Met Val Phe Val Ala Pro Glu Ser Ser Ser Phe Gly Phe
450 455 460
Leu Lys Asn Leu Ser Glu Glu Gln Ile Ala Val Tyr Asn Ala Leu Ile
465 470 475 480
Gln His Gly Met Lys Val Ile Leu Ile Glu Leu Glu Lys Val Lys Asp
485 490 495
Tyr Ser Thr Met Pro Glu Ser Ile Gln Tyr Ile Arg Gln Lys His Gly
500 505 510
Ala Ile Gln Trp Asp Gly Asp Phe Thr Glu Gln Ser Gln Cys Ala Lys
515 520 525
Thr Lys Phe Trp Lys Lys Val Arg Tyr His Met Pro Pro Arg Arg Tyr
530 535 540
Pro Ala Ser Ser Pro Val Gln Leu Leu Gly His Ile Pro Cys Asn Cys
545 550 555 560
Lys Ala Gly Lys Cys Asn Ala Ala Thr Gly Leu Ile Thr Pro
565 570
<210> 5
<211> 2044
<212> ДНК
<213> Rattus norvegicus
<400> 5
ccggctggcc taggatcagg caagaaaagg ctgaacgcct ttctaaggac ggactctttc 60
tgtacagctc cacttgggga agcccgaaat ggggatgcca cccttgctct tctgttgggt 120
gtctttcgtg cttccacttt ttgtggcagc aggtaactgt actgatgtct atatgcacca 180
tgagatgatt tcagagggcc agcctttccc cttcaactgc acataccctc cagtaacaaa 240
cggggcagtg aatctgacat ggcatagaac acccagtaag agcccaatct ccatcaacag 300
acacgttaga attcaccagg accagtcctg gattttgttt cttccgttgg cattggagga 360
ctcaggcatc tatcaatgtg ttataaagga tgcccacagc tgttaccgaa tagctataaa 420
cctaaccgtt tttagaaaac actggtgcga ctcttccaac gaagagagtt ccataaattc 480
ctcagatgag taccagcaat ggttacccat aggaaaatcg ggcagtctga cgtgccatct 540
ctacttccca gagagctgtg ttttggattc aataaagtgg tataagggtt gtgaagagat 600
taaagtgagc aagaagtttt gccctacagg aacaaagctt cttgttaaca acatcgacgt 660
ggaggatagt gggagctatg catgctcagc cagactgaca cacttgggga gaatcttcac 720
ggttagaaac tacattgctg tgaataccaa ggaagttggg tctggaggaa ggatccctaa 780
catcacgtat ccaaaaaaca actccattga agttcaactt ggctccaccc tcattgtgga 840
ctgcaatata acagacacga aggagaatac gaacctcaga tgctggcgag ttaacaacac 900
cctggtggac gattactaca acgacttcaa acgcatccag gaaggaatcg aaaccaatct 960
gtctctgagg aatcacattc tgtacacagt gaacataaca ttcttagaag tgaaaatgga 1020
ggactacggc catcctttca catgccacgc tgcggtgtcc gcagcctaca tcattctgaa 1080
acgcccagct ccagacttcc gggcttacct cataggaggt ctcatggctt tcctacttct 1140
ggccgtgtcc attctgtaca tctacaacac ctttaaggtc gacatcgtgc tttggtatag 1200
gagtaccttc cacactgccc aggctccaga tgacgagaag ctgtatgatg cctatgtctt 1260
ataccccaag tacccaagag aaagccaggg ccatgatgtg gacacactgg tgttgaagat 1320
cttgcccgag gtgctggaga aacagtgtgg atataagtta ttcatatttg gcagggatga 1380
attccctgga caagctgtgg ccagcgtcat tgatgaaaac attaagctgt gtaggaggct 1440
gatggtcctc gtggcaccag agacatccag cttcagcttt ctgaagaact tgactgaaga 1500
acaaatcgct gtctacaatg ccctcgtcca ggacggcatg aaggtcattc tgattgaact 1560
ggagagagtc aaggactaca gcaccatgcc cgagtccatt cagtacatcc gacagaagca 1620
cggggccatc cagtgggatg gggacttcac agagcaggca cagtgcgcca agacgaaatt 1680
ctggaagaaa gtgagatatc atatgccacc caggaggtac ccggcatctc cccccgtcca 1740
gctgctagga cacacacccc gcataccagg ctagtgcagt gccaccgcca cggggctcat 1800
aactccttaa gagcggttag tgtgtggtgg ctcgcactac aacctctctg gatcatctac 1860
ccccgtagct tgctcttttg tgcttgtgag cgacctcgtc cttagccacg tcatattttg 1920
attttttgtt tgttttgttt gtttgttgta tgcttttagt catagctgat tcgtactact 1980
cctgtttgct tcatggttcc tgaatcccag agactccctg agcatgggtg gctatcatgt 2040
tggg 2044
<210> 6
<211> 561
<212> БЕЛОК
<213> Rattus norvegicus
<400> 6
Met Gly Met Pro Pro Leu Leu Phe Cys Trp Val Ser Phe Val Leu Pro
1 5 10 15
Leu Phe Val Ala Ala Gly Asn Cys Thr Asp Val Tyr Met His His Glu
20 25 30
Met Ile Ser Glu Gly Gln Pro Phe Pro Phe Asn Cys Thr Tyr Pro Pro
35 40 45
Val Thr Asn Gly Ala Val Asn Leu Thr Trp His Arg Thr Pro Ser Lys
50 55 60
Ser Pro Ile Ser Ile Asn Arg His Val Arg Ile His Gln Asp Gln Ser
65 70 75 80
Trp Ile Leu Phe Leu Pro Leu Ala Leu Glu Asp Ser Gly Ile Tyr Gln
85 90 95
Cys Val Ile Lys Asp Ala His Ser Cys Tyr Arg Ile Ala Ile Asn Leu
100 105 110
Thr Val Phe Arg Lys His Trp Cys Asp Ser Ser Asn Glu Glu Ser Ser
115 120 125
Ile Asn Ser Ser Asp Glu Tyr Gln Gln Trp Leu Pro Ile Gly Lys Ser
130 135 140
Gly Ser Leu Thr Cys His Leu Tyr Phe Pro Glu Ser Cys Val Leu Asp
145 150 155 160
Ser Ile Lys Trp Tyr Lys Gly Cys Glu Glu Ile Lys Val Ser Lys Lys
165 170 175
Phe Cys Pro Thr Gly Thr Lys Leu Leu Val Asn Asn Ile Asp Val Glu
180 185 190
Asp Ser Gly Ser Tyr Ala Cys Ser Ala Arg Leu Thr His Leu Gly Arg
195 200 205
Ile Phe Thr Val Arg Asn Tyr Ile Ala Val Asn Thr Lys Glu Val Gly
210 215 220
Ser Gly Gly Arg Ile Pro Asn Ile Thr Tyr Pro Lys Asn Asn Ser Ile
225 230 235 240
Glu Val Gln Leu Gly Ser Thr Leu Ile Val Asp Cys Asn Ile Thr Asp
245 250 255
Thr Lys Glu Asn Thr Asn Leu Arg Cys Trp Arg Val Asn Asn Thr Leu
260 265 270
Val Asp Asp Tyr Tyr Asn Asp Phe Lys Arg Ile Gln Glu Gly Ile Glu
275 280 285
Thr Asn Leu Ser Leu Arg Asn His Ile Leu Tyr Thr Val Asn Ile Thr
290 295 300
Phe Leu Glu Val Lys Met Glu Asp Tyr Gly His Pro Phe Thr Cys His
305 310 315 320
Ala Ala Val Ser Ala Ala Tyr Ile Ile Leu Lys Arg Pro Ala Pro Asp
325 330 335
Phe Arg Ala Tyr Leu Ile Gly Gly Leu Met Ala Phe Leu Leu Leu Ala
340 345 350
Val Ser Ile Leu Tyr Ile Tyr Asn Thr Phe Lys Val Asp Ile Val Leu
355 360 365
Trp Tyr Arg Ser Thr Phe His Thr Ala Gln Ala Pro Asp Asp Glu Lys
370 375 380
Leu Tyr Asp Ala Tyr Val Leu Tyr Pro Lys Tyr Pro Arg Glu Ser Gln
385 390 395 400
Gly His Asp Val Asp Thr Leu Val Leu Lys Ile Leu Pro Glu Val Leu
405 410 415
Glu Lys Gln Cys Gly Tyr Lys Leu Phe Ile Phe Gly Arg Asp Glu Phe
420 425 430
Pro Gly Gln Ala Val Ala Ser Val Ile Asp Glu Asn Ile Lys Leu Cys
435 440 445
Arg Arg Leu Met Val Leu Val Ala Pro Glu Thr Ser Ser Phe Ser Phe
450 455 460
Leu Lys Asn Leu Thr Glu Glu Gln Ile Ala Val Tyr Asn Ala Leu Val
465 470 475 480
Gln Asp Gly Met Lys Val Ile Leu Ile Glu Leu Glu Arg Val Lys Asp
485 490 495
Tyr Ser Thr Met Pro Glu Ser Ile Gln Tyr Ile Arg Gln Lys His Gly
500 505 510
Ala Ile Gln Trp Asp Gly Asp Phe Thr Glu Gln Ala Gln Cys Ala Lys
515 520 525
Thr Lys Phe Trp Lys Lys Val Arg Tyr His Met Pro Pro Arg Arg Tyr
530 535 540
Pro Ala Ser Pro Pro Val Gln Leu Leu Gly His Thr Pro Arg Ile Pro
545 550 555 560
Gly
<210> 7
<211> 573
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированный Il1r12
<400> 7
Met Gly Val Thr Ser Leu Leu Phe Cys Gly Val Phe Phe Leu Leu Leu
1 5 10 15
Leu Phe Val Ala Ala Asp Gly Cys Lys Asp Ile Phe Met Lys Asn Glu
20 25 30
Ile Leu Ser Ala Ser Gln Pro Phe Ala Phe Asn Cys Thr Phe Pro Pro
35 40 45
Ile Thr Ser Gly Glu Val Ser Val Thr Trp Tyr Lys Asn Ser Ser Lys
50 55 60
Ile Pro Val Ser Lys Ile Ile Gln Ser Arg Ile His Gln Asp Glu Thr
65 70 75 80
Trp Ile Leu Phe Leu Pro Met Glu Trp Gly Asp Ser Gly Val Tyr Gln
85 90 95
Cys Val Ile Lys Gly Arg Asp Ser Cys His Arg Ile His Val Asn Leu
100 105 110
Thr Val Phe Glu Lys His Trp Cys Asp Thr Ser Ile Gly Gly Leu Pro
115 120 125
Asn Leu Ser Asp Glu Tyr Lys Gln Ile Leu His Leu Gly Lys Asp Asp
130 135 140
Ser Leu Thr Cys His Leu His Phe Pro Lys Ser Cys Val Leu Gly Pro
145 150 155 160
Ile Lys Trp Tyr Lys Asp Cys Asn Glu Ile Lys Gly Glu Arg Phe Thr
165 170 175
Val Leu Glu Thr Arg Leu Leu Val Ser Asn Val Ser Ala Glu Asp Arg
180 185 190
Gly Asn Tyr Ala Cys Gln Ala Ile Leu Thr His Ser Gly Lys Gln Tyr
195 200 205
Glu Val Leu Asn Gly Ile Thr Val Ser Ile Thr Glu Arg Ala Gly Tyr
210 215 220
Gly Gly Ser Val Pro Lys Ile Ile Tyr Pro Lys Asn His Ser Ile Glu
225 230 235 240
Val Gln Leu Gly Thr Thr Leu Ile Val Asp Cys Asn Val Thr Asp Thr
245 250 255
Lys Asp Asn Thr Asn Leu Arg Cys Trp Arg Val Asn Asn Thr Leu Val
260 265 270
Asp Asp Tyr Tyr Asp Glu Ser Lys Arg Ile Arg Glu Gly Val Glu Thr
275 280 285
His Val Ser Phe Arg Glu His Asn Leu Tyr Thr Val Asn Ile Thr Phe
290 295 300
Leu Glu Val Lys Met Glu Asp Tyr Gly Leu Pro Phe Met Cys His Ala
305 310 315 320
Gly Val Ser Thr Ala Tyr Ile Ile Leu Gln Leu Pro Val Pro Asp Phe
325 330 335
Arg Ala Tyr Leu Leu Gly Gly Leu Met Ala Phe Leu Leu Leu Val Val
340 345 350
Ser Val Leu Phe Ile Tyr Asn Ser Phe Lys Ile Asp Ile Met Leu Trp
355 360 365
Tyr Arg Ser Ala Phe His Thr Ala Gln Ala Pro Asp Asp Glu Lys Leu
370 375 380
Tyr Asp Ala Tyr Val Leu Tyr Pro Lys Tyr Pro Arg Gly Ser Gln Gly
385 390 395 400
His Asp Val Asp Thr Leu Val Leu Lys Ile Leu Pro Glu Val Leu Glu
405 410 415
Lys Gln Cys Gly Tyr Lys Leu Phe Ile Phe Gly Arg Asp Glu Phe Pro
420 425 430
Gly Gln Ala Val Ala Ser Val Ile Asp Glu Asn Ile Lys Leu Cys Arg
435 440 445
Arg Leu Met Val Phe Val Ala Pro Glu Ser Ser Ser Phe Gly Phe Leu
450 455 460
Lys Asn Leu Ser Glu Glu Gln Ile Ala Val Tyr Asn Ala Leu Ile Gln
465 470 475 480
His Gly Met Lys Val Ile Leu Ile Glu Leu Glu Lys Val Lys Asp Tyr
485 490 495
Ser Thr Met Pro Glu Ser Ile Gln Tyr Ile Arg Gln Lys His Gly Ala
500 505 510
Ile Gln Trp Asp Gly Asp Phe Thr Glu Gln Ser Gln Cys Ala Lys Thr
515 520 525
Lys Phe Trp Lys Lys Val Arg Tyr His Met Pro Pro Arg Arg Tyr Pro
530 535 540
Ala Ser Ser Pro Val Gln Leu Leu Gly His Ile Pro Cys Asn Cys Lys
545 550 555 560
Ala Gly Lys Cys Asn Ala Ala Thr Gly Leu Ile Thr Pro
565 570
<210> 8
<211> 888
<212> ДНК
<213> Homo sapiens
<400> 8
aaaacccaag tgcagtagaa gccattgttc ataatggtag ggatacaggg tccttcgtaa 60
cagattatca gtgtggccta tgctggaaag tctggtgacc tctgattttt tttgcttcca 120
ggtctttggc cttggcactc tttgtcatat tagagttcct gggtctaggc ctgggcagga 180
ttcataggtg cagctgcttc tgctggaggt agactgcatc caacaaagta agggtgctgg 240
gtgagttctg ggagtataga ttctgactgg ggtcactgct gggctggccg ccagtctttc 300
atctgaccca gggttaaact gtggcttggg actgactcag gtcctctctt ggggtcggtc 360
tgcacataaa aggactccta tccttggcag ttctgaaaca acaccaccac aatggaaaaa 420
gcattgaaaa ttgacacacc tcagcagggg agcattcagg atatcaatca tcgggtgtgg 480
gttcttcagg accagacgct catagcagtc ccgaggaagg accgtatgtc tccagtcact 540
attgccttaa tctcatgccg acatgtggag acccttgaga aagacagagg gaaccccatc 600
tacctgggcc tgaatggact caatctctgc ctgatgtgtg ctaaagtcgg ggaccagccc 660
acactgcagc tgaaggaaaa ggatataatg gatttgtaca accaacccga gcctgtgaag 720
tcctttctct tctaccacag ccagagtggc aggaactcca ccttcgagtc tgtggctttc 780
cctggctggt tcatcgctgt cagctctgaa ggaggctgtc ctctcatcct tacccaagaa 840
ctggggaaag ccaacactac tgactttggg ttaactatgc tgttttaa 888
<210> 9
<211> 158
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Met Glu Lys Ala Leu Lys Ile Asp Thr Pro Gln Gln Gly Ser Ile Gln
1 5 10 15
Asp Ile Asn His Arg Val Trp Val Leu Gln Asp Gln Thr Leu Ile Ala
20 25 30
Val Pro Arg Lys Asp Arg Met Ser Pro Val Thr Ile Ala Leu Ile Ser
35 40 45
Cys Arg His Val Glu Thr Leu Glu Lys Asp Arg Gly Asn Pro Ile Tyr
50 55 60
Leu Gly Leu Asn Gly Leu Asn Leu Cys Leu Met Cys Ala Lys Val Gly
65 70 75 80
Asp Gln Pro Thr Leu Gln Leu Lys Glu Lys Asp Ile Met Asp Leu Tyr
85 90 95
Asn Gln Pro Glu Pro Val Lys Ser Phe Leu Phe Tyr His Ser Gln Ser
100 105 110
Gly Arg Asn Ser Thr Phe Glu Ser Val Ala Phe Pro Gly Trp Phe Ile
115 120 125
Ala Val Ser Ser Glu Gly Gly Cys Pro Leu Ile Leu Thr Gln Glu Leu
130 135 140
Gly Lys Ala Asn Thr Thr Asp Phe Gly Leu Thr Met Leu Phe
145 150 155
<210> 10
<211> 883
<212> ДНК
<213> Mus musculus
<400> 10
taacctcctc ttttcgtgtt gtggataaaa ggcttggacc ttagtagagc ttgtcagaga 60
agctgagcac agtagtgggt gtagttctgt agtgtgcaga cacattccta ttcaatcagg 120
gtcaatctgc agattggcag ctcagaaaca acatcaccat aatgaataag gagaaagaac 180
taagagcagc atcaccttcg cttagacatg ttcaggatct tagtagtcgt gtgtggatcc 240
tgcagaacaa tatcctcact gcagtcccaa ggaaagagca aacagttcca gtcactatta 300
ccttgctccc atgccaatat ctggacactc ttgagacgaa caggggggat cccacgtaca 360
tgggagtgca aaggccgatg agctgcctgt tctgcacaaa ggatggggag cagcctgtgc 420
tacagcttgg ggaagggaac ataatggaaa tgtacaacaa aaaggaacct gtaaaagcct 480
ctctcttcta tcacaagaag agtggtacaa cctctacatt tgagtctgca gccttccctg 540
gttggttcat cgctgtctgc tctaaaggga gctgcccact cattctgacc caagaactgg 600
gggaaatctt catcactgac ttcgagatga ttgtggtaca ttaaggtttt tagacacatt 660
gctctgtggc actctctcaa gatttcttgg attctaacaa gaagcaatca aagacacccc 720
taacaaaatg gaagactgaa aagaaagctg agccctccct gggctgtttt tccttggtgg 780
tgaatcagat gcagaacatc ttaccatgtt ttcatccaaa gcatttactg ttggttttta 840
caaggagtga attttttaaa ataaaatcat ttatctcatt tgc 883
<210> 11
<211> 160
<212> БЕЛОК
<213> Mus musculus
<400> 11
Met Asn Lys Glu Lys Glu Leu Arg Ala Ala Ser Pro Ser Leu Arg His
1 5 10 15
Val Gln Asp Leu Ser Ser Arg Val Trp Ile Leu Gln Asn Asn Ile Leu
20 25 30
Thr Ala Val Pro Arg Lys Glu Gln Thr Val Pro Val Thr Ile Thr Leu
35 40 45
Leu Pro Cys Gln Tyr Leu Asp Thr Leu Glu Thr Asn Arg Gly Asp Pro
50 55 60
Thr Tyr Met Gly Val Gln Arg Pro Met Ser Cys Leu Phe Cys Thr Lys
65 70 75 80
Asp Gly Glu Gln Pro Val Leu Gln Leu Gly Glu Gly Asn Ile Met Glu
85 90 95
Met Tyr Asn Lys Lys Glu Pro Val Lys Ala Ser Leu Phe Tyr His Lys
100 105 110
Lys Ser Gly Thr Thr Ser Thr Phe Glu Ser Ala Ala Phe Pro Gly Trp
115 120 125
Phe Ile Ala Val Cys Ser Lys Gly Ser Cys Pro Leu Ile Leu Thr Gln
130 135 140
Glu Leu Gly Glu Ile Phe Ile Thr Asp Phe Glu Met Ile Val Val His
145 150 155 160
<210> 12
<211> 869
<212> ДНК
<213> Rattus norvegicus
<400> 12
taatctcctc ccttcatgtt ttggataaaa ggcctggacc ctagcagagc ttgtcagaga 60
agctgagcac agtagtggct gtagttctgt agtgtgcaga tacattcctc tgcaatcagg 120
atcagtctgc agattggcag ttcagaaaca acatcaccat aatgaataag gataaagaac 180
taagtgcagc aacaccttgg tttagacata ttcaggatct tagtagtcgt gtgtgggtcc 240
ttcaagacaa tattcttact gcagtcccaa ggaaagagca aacggttcca gtcactatca 300
ccttactccc atgccaatat ctggacactc ttgagaagaa caaaggggat cccatgtacc 360
tgggagtgaa gaaacctcaa agttgtctgt cctgcacaaa gaatggggag cagcctgtac 420
tacaacttcg ggaagggaac atactggata tgtaccacca gaaggaacgt gtaaaagcct 480
ctctcttcta tcacaagaag agtggtacaa cctctacatt cgagtctgca gccttccctg 540
gttggttcat tgctgtctgc tccaaaggga gctgcccact cgttttgacc caagaacttg 600
ggaaaacctt catcactgat tttgagatga ctgtagtaca ttaagattgc tctgcagcac 660
tctctgaaga tctctcggat tctaacaagc aaccaaacac acccccaaca aaatacaaga 720
ctgaaaagaa agctgagccc tccctgggct tttttccctt ggcaggtgga tcagatgcag 780
aacttcttac catgttttca tccaaagcat ttactgatgg tttttacaag aagaaaacgt 840
ttttaattaa aatcatttat ctcacttgc 869
<210> 13
<211> 160
<212> БЕЛОК
<213> Rattus norvegicus
<400> 13
Met Asn Lys Asp Lys Glu Leu Ser Ala Ala Thr Pro Trp Phe Arg His
1 5 10 15
Ile Gln Asp Leu Ser Ser Arg Val Trp Val Leu Gln Asp Asn Ile Leu
20 25 30
Thr Ala Val Pro Arg Lys Glu Gln Thr Val Pro Val Thr Ile Thr Leu
35 40 45
Leu Pro Cys Gln Tyr Leu Asp Thr Leu Glu Lys Asn Lys Gly Asp Pro
50 55 60
Met Tyr Leu Gly Val Lys Lys Pro Gln Ser Cys Leu Ser Cys Thr Lys
65 70 75 80
Asn Gly Glu Gln Pro Val Leu Gln Leu Arg Glu Gly Asn Ile Leu Asp
85 90 95
Met Tyr His Gln Lys Glu Arg Val Lys Ala Ser Leu Phe Tyr His Lys
100 105 110
Lys Ser Gly Thr Thr Ser Thr Phe Glu Ser Ala Ala Phe Pro Gly Trp
115 120 125
Phe Ile Ala Val Cys Ser Lys Gly Ser Cys Pro Leu Val Leu Thr Gln
130 135 140
Glu Leu Gly Lys Thr Phe Ile Thr Asp Phe Glu Met Thr Val Val His
145 150 155 160
<210> 14
<211> 1186
<212> ДНК
<213> Homo sapiens
<400> 14
cacgggttcc tccccactct gtctttctca cctctccttc acttttccta gcctcctcac 60
caccatctga tctatcttgt tctcttcaca aaaggctctg aagacatcat gaacccacaa 120
cgggaggcag cacccaaatc ctatgctatt cgtgattctc gacagatggt gtgggtcctg 180
agtggaaatt ctttaatagc agctcctctt agccgcagca ttaagcctgt cactcttcat 240
ttaatagcct gtagagacac agaattcagt gacaaggaaa agggtaatat ggtttacctg 300
ggaatcaagg gaaaagatct ctgtctcttc tgtgcagaaa ttcagggcaa gcctactttg 360
cagcttaagc ttcagggctc ccaagataac atagggaagg acacttgctg gaaactagtt 420
ggaattcaca catgcataaa cctggatgtg agagagagct gcttcatggg aacccttgac 480
caatggggaa taggagtggg tagaaagaag tggaagagtt cctttcaaca tcaccatctc 540
aggaagaagg acaaagattt ctcatccatg cggaccaaca taggaatgcc aggaaggatg 600
tagaaataag gggaggaaga ttcccatctc tacaatcttt gagtgggttt gctatcaatg 660
aaatgctaca aatggaataa gttgcagaaa tttttctctt ttcttgggtt ctggagagtt 720
tgtaaaacaa ggacactatg tatttttaaa gagttggtaa atcttacctg taaagctaga 780
gaaggtcgga gtctttttag gagtagattt ggactacata acctgtaaat gtgttttgtc 840
cagtccttag agtgtttttt aaaaaattgt aaagtcaagg ttttcatgaa aaatgggaag 900
atcagacaac attgctcctg aattcccaca gagcagcaag ctactagagc tcaatctgtt 960
atttcttttc ctgatgtaca ggggttaagt cctatggaag aaacagcaga attattcaaa 1020
attatttaca taatgtgcaa ttattcacta gagcatgagg agtgaaacgc tctgtttagt 1080
atgtataact taaaaggaac acatacaatt aaaagtaatt gaaagacatt tcttcttaaa 1140
aattctataa tcttacactg gtaaaataaa ctagtttttc ccatgt 1186
<210> 15
<211> 164
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Met Asn Pro Gln Arg Glu Ala Ala Pro Lys Ser Tyr Ala Ile Arg Asp
1 5 10 15
Ser Arg Gln Met Val Trp Val Leu Ser Gly Asn Ser Leu Ile Ala Ala
20 25 30
Pro Leu Ser Arg Ser Ile Lys Pro Val Thr Leu His Leu Ile Ala Cys
35 40 45
Arg Asp Thr Glu Phe Ser Asp Lys Glu Lys Gly Asn Met Val Tyr Leu
50 55 60
Gly Ile Lys Gly Lys Asp Leu Cys Leu Phe Cys Ala Glu Ile Gln Gly
65 70 75 80
Lys Pro Thr Leu Gln Leu Lys Leu Gln Gly Ser Gln Asp Asn Ile Gly
85 90 95
Lys Asp Thr Cys Trp Lys Leu Val Gly Ile His Thr Cys Ile Asn Leu
100 105 110
Asp Val Arg Glu Ser Cys Phe Met Gly Thr Leu Asp Gln Trp Gly Ile
115 120 125
Gly Val Gly Arg Lys Lys Trp Lys Ser Ser Phe Gln His His His Leu
130 135 140
Arg Lys Lys Asp Lys Asp Phe Ser Ser Met Arg Thr Asn Ile Gly Met
145 150 155 160
Pro Gly Arg Met
<210> 16
<211> 790
<212> ДНК
<213> Mus musculus
<400> 16
gactggcctc attgcctgct gtcataactt cgatcccaga gacaagactt ctttggacca 60
aggcaatgat ggctttccct ccacaatctt gtgtacatgt tcttcctcca aagagtattc 120
aaatgtggga accgaatcat aacactatgc atggatcctc acaatctccc agaaactaca 180
gggttcatga ctcacaacag atggtatggg tcctgactgg aaatacttta acagcagttc 240
ctgctagcaa caatgtcaag cctgtcattc ttagcttgat agcatgtaga gacacggaat 300
tccaagatgt aaagaaaggt aatctagttt tcctgggaat caagaacaga aatctctgct 360
tctgctgtgt tgagatggag ggcaaaccaa ctttgcagct taaggaagta gacatcatga 420
atttgtacaa agagagaaaa gcacaaaaag cctttctgtt ctatcatggc atagagggct 480
ccacttctgt ctttcagtca gtcctctatc ctggctggtt tatagccacc tcttccatag 540
aaagacagac aatcatcctc acacatcagc ggggtaaatt ggttaacact aacttctaca 600
tagagtctga gaagtaaatc caacatgggt catatgtggc cagctccagg ccacagaatc 660
aaactgttga agaatcttct acttgaaaac atagcaagcc tcttagaaga ccaccatgcc 720
atgtcttcac cagacatcca tgtctaaaat gccactcatt tgaatatcat tccagcatga 780
agcccatgaa 790
<210> 17
<211> 183
<212> БЕЛОК
<213> Mus musculus
<400> 17
Met Met Ala Phe Pro Pro Gln Ser Cys Val His Val Leu Pro Pro Lys
1 5 10 15
Ser Ile Gln Met Trp Glu Pro Asn His Asn Thr Met His Gly Ser Ser
20 25 30
Gln Ser Pro Arg Asn Tyr Arg Val His Asp Ser Gln Gln Met Val Trp
35 40 45
Val Leu Thr Gly Asn Thr Leu Thr Ala Val Pro Ala Ser Asn Asn Val
50 55 60
Lys Pro Val Ile Leu Ser Leu Ile Ala Cys Arg Asp Thr Glu Phe Gln
65 70 75 80
Asp Val Lys Lys Gly Asn Leu Val Phe Leu Gly Ile Lys Asn Arg Asn
85 90 95
Leu Cys Phe Cys Cys Val Glu Met Glu Gly Lys Pro Thr Leu Gln Leu
100 105 110
Lys Glu Val Asp Ile Met Asn Leu Tyr Lys Glu Arg Lys Ala Gln Lys
115 120 125
Ala Phe Leu Phe Tyr His Gly Ile Glu Gly Ser Thr Ser Val Phe Gln
130 135 140
Ser Val Leu Tyr Pro Gly Trp Phe Ile Ala Thr Ser Ser Ile Glu Arg
145 150 155 160
Gln Thr Ile Ile Leu Thr His Gln Arg Gly Lys Leu Val Asn Thr Asn
165 170 175
Phe Tyr Ile Glu Ser Glu Lys
180
<210> 18
<211> 587
<212> ДНК
<213> Rattus norvegicus
<400> 18
ggcgtgggtc ctggctggaa attctttgac agcggttcct gctagcaaca gcgtcaaatc 60
tgtcattctt agcttgatag catgtagaga catggaattc caagatgaaa agaagggtaa 120
tctagttttc ctgggaatca aaggcagaag tctctgcctc ttctgtgctg agatcgaggg 180
caaaccgact ttgcagctta aggatgtaga catcatggat ttgtacaatg agaaaaaagc 240
acagaaagcc tttctcttct accatggcat agagggatct acttctgtct ttcagtcagt 300
cttgtatcct ggctggttta tagccacctc ttccacagca agacaaacaa tcattcttac 360
acaggagagg ggtgaagcta ataacactaa cttctactta gagtctgaga attagatcta 420
ccatgggcca tatgtggcca gttgcaggcc aaagaatcaa actgttggag aatcttttac 480
ttgaaaacat agcaagacat cttagaagac caccatgttt cctcttctcc tgacatccat 540
gtctgaaatg ccattcattg gtataagaga ataaaaaata gggctgg 587
<210> 19
<211> 179
<212> БЕЛОК
<213> Rattus norvegicus
<400> 19
Met Met Ala Phe Pro Pro Gln Ser His Val Cys Phe Thr Pro Pro Lys
1 5 10 15
Ser Ser Asp Phe Asp Asn Ile Asn Lys Gly Val Gln Ala Phe Pro Ser
20 25 30
Thr Tyr Arg Val His Asp Ser Gln His Lys Ala Trp Val Leu Ala Gly
35 40 45
Asn Ser Leu Thr Ala Val Pro Ala Ser Asn Ser Val Lys Ser Val Ile
50 55 60
Leu Ser Leu Ile Ala Cys Arg Asp Met Glu Phe Gln Asp Glu Lys Lys
65 70 75 80
Gly Asn Leu Val Phe Leu Gly Ile Lys Gly Arg Ser Leu Cys Leu Phe
85 90 95
Cys Ala Glu Ile Glu Gly Lys Pro Thr Leu Gln Leu Lys Asp Val Asp
100 105 110
Ile Met Asp Leu Tyr Asn Glu Lys Lys Ala Gln Lys Ala Phe Leu Phe
115 120 125
Tyr His Gly Ile Glu Gly Ser Thr Ser Val Phe Gln Ser Val Leu Tyr
130 135 140
Pro Gly Trp Phe Ile Ala Thr Ser Ser Thr Ala Arg Gln Thr Ile Ile
145 150 155 160
Leu Thr Gln Glu Arg Gly Glu Ala Asn Asn Thr Asn Phe Tyr Leu Glu
165 170 175
Ser Glu Asn
<210> 20
<211> 1212
<212> ДНК
<213> Homo sapiens
<400> 20
gaagctgctg gagccacgat tcagtcccct ggactgtaga taaagaccct ttcttgccag 60
gtgctgagac aaccacacta tgagaggcac tccaggagac gctgatggtg gaggaagggc 120
cgtctatcaa tcaatgtgta aacctattac tgggactatt aatgatttga atcagcaagt 180
gtggaccctt cagggtcaga accttgtggc agttccacga agtgacagtg tgaccccagt 240
cactgttgct gttatcacat gcaagtatcc agaggctctt gagcaaggca gaggggatcc 300
catttatttg ggaatccaga atccagaaat gtgtttgtat tgtgagaagg ttggagaaca 360
gcccacattg cagctaaaag agcagaagat catggatctg tatggccaac ccgagcccgt 420
gaaacccttc cttttctacc gtgccaagac tggtaggacc tccacccttg agtctgtggc 480
cttcccggac tggttcattg cctcctccaa gagagaccag cccatcattc tgacttcaga 540
acttgggaag tcatacaaca ctgcctttga attaaatata aatgactgaa ctcagcctag 600
aggtggcagc ttggtctttg tcttaaagtt tctggttccc aatgtgtttt cgtctacatt 660
ttcttagtgt cattttcacg ctggtgctga gacaggggca aggctgctgt tatcatctca 720
ttttataatg aagaagaagc aattacttca tagcaactga agaacaggat gtggcctcag 780
aagcaggaga gctgggtggt ataaggctgt cctctcaagc tggtgctgtg taggccacaa 840
ggcatctgca tgagtgactt taagactcaa agaccaaaca ctgagctttc ttctaggggt 900
gggtatgaag atgcttcaga gctcatgcgc gttacccacg atggcatgac tagcacagag 960
ctgatctctg tttctgtttt gctttattcc ctcttgggat gatatcatcc agtctttata 1020
tgttgccaat atacctcatt gtgtgtaata gaaccttctt agcattaaga ccttgtaaac 1080
aaaaataatt cttgtgttaa gttaaatcat ttttgtccta attgtaatgt gtaatcttaa 1140
agttaaataa actttgtgta tttatataat aataaagcta aaactgatat aaaataaaga 1200
aagagtaaac tg 1212
<210> 21
<211> 169
<212> БЕЛОК
<213> Homo sapiens
<400> 21
Met Arg Gly Thr Pro Gly Asp Ala Asp Gly Gly Gly Arg Ala Val Tyr
1 5 10 15
Gln Ser Met Cys Lys Pro Ile Thr Gly Thr Ile Asn Asp Leu Asn Gln
20 25 30
Gln Val Trp Thr Leu Gln Gly Gln Asn Leu Val Ala Val Pro Arg Ser
35 40 45
Asp Ser Val Thr Pro Val Thr Val Ala Val Ile Thr Cys Lys Tyr Pro
50 55 60
Glu Ala Leu Glu Gln Gly Arg Gly Asp Pro Ile Tyr Leu Gly Ile Gln
65 70 75 80
Asn Pro Glu Met Cys Leu Tyr Cys Glu Lys Val Gly Glu Gln Pro Thr
85 90 95
Leu Gln Leu Lys Glu Gln Lys Ile Met Asp Leu Tyr Gly Gln Pro Glu
100 105 110
Pro Val Lys Pro Phe Leu Phe Tyr Arg Ala Lys Thr Gly Arg Thr Ser
115 120 125
Thr Leu Glu Ser Val Ala Phe Pro Asp Trp Phe Ile Ala Ser Ser Lys
130 135 140
Arg Asp Gln Pro Ile Ile Leu Thr Ser Glu Leu Gly Lys Ser Tyr Asn
145 150 155 160
Thr Ala Phe Glu Leu Asn Ile Asn Asp
165
<210> 22
<211> 1647
<212> ДНК
<213> Mus musculus
<400> 22
atcattacac ctgggagctg cccaagctgg gatttggtct tctagacttt ccataaggac 60
tctttcttgc catgtaccta caaacctaca aaaccctttg agtgcgcctc tgagtgtgga 120
gctttctgca gttcagggtt cagtgaatgg acagatctgt cttgctgtaa gtgatcctgt 180
gagcgatcct gtgagcgatc ctgtgagcga tcctgtgagc gatcctgtga gcgatcctgt 240
gagtgatcct gtgagcaatc ctgtgagagc tctgtgcacc agaacaagat cacgatggaa 300
aacaatgaaa aaaaaaacat tgtgtatgga agtgatgttg agatggaaca cgagagagct 360
gggctatttg tatcttcagc tatgttttct aaacacccat tttctacaca catctcagga 420
agagaaactc ctgactttgg ggaggttttt gacttggacc agcaggtgtg gatctttcgt 480
aatcaggccc ttgtgacagt tccacgaagc cacagagtaa ccccagtcag cgtgactatc 540
ctcccatgca agtacccaga gtctcttgaa caggacaaag ggattgccat ttatttggga 600
attcagaatc cagataaatg cctgttttgt aaggaagtta atggacaccc tactttgctg 660
ctaaaggaag agaagatttt ggatttgtac caccaccctg agccaatgaa gccattcctg 720
ttttaccaca cccggacagg tggaacatcc acctttgaat cagtggcttt ccctggccac 780
tatattgcct cctccaagac tggcaacccc atcttcctca catcaaaaaa gggagaatat 840
tacaacatta acttcaattt agatataaag tcttaaactc agcatggaag tggagggttg 900
gttagaactc ttcctcttaa aacatcaaac ctctaatatg ttatcatttg tttgatctat 960
ggtattttca tactgaaagg taagcaggac caacattatc tcctttcata gatgaagaag 1020
caacaaaata atgcatactc caaagtcggt gggattggag tggtgtaagg atgttctcta 1080
agactgaagt ggtccaatct ataagacatc atgtctgaga aatagggtgc tgtgccacag 1140
tgtataaaac ctagaatgcc ctgtgtgtga gtggtgacat cctcatttcc agccattggc 1200
ctgattcata ccatgttttc acacatttgt aatatcaact ccttgcattt ggatagcaga 1260
ggcctcacaa tgacatttgc ctcaacagct aatagctttg atctttcaat gctgtgtaga 1320
tctgaataca ctagaaaagg aattagaatc ctgagtgaat gaactgtgag gagccgccct 1380
cacatttgcc attataagat ggcactgaca gctgtgttct aagtggtaaa catagtctgc 1440
acacatgcag gggcagtttt cccaccatgt gttctgcctt tcccgtgatg acaactgggc 1500
cgatgggctg cagccaatca gggagtaata cgtcctaggt ggaggataat tctccttaaa 1560
agggacgggg ttctggcact ctctcccttc cttgcttgct ctcttgcact ctggctcctg 1620
aagatgtaag caataaagct ttgccgc 1647
<210> 23
<211> 164
<212> БЕЛОК
<213> Mus musculus
<400> 23
Met Phe Ser Lys His Pro Phe Ser Thr His Ile Ser Gly Arg Glu Thr
1 5 10 15
Pro Asp Phe Gly Glu Val Phe Asp Leu Asp Gln Gln Val Trp Ile Phe
20 25 30
Arg Asn Gln Ala Leu Val Thr Val Pro Arg Ser His Arg Val Thr Pro
35 40 45
Val Ser Val Thr Ile Leu Pro Cys Lys Tyr Pro Glu Ser Leu Glu Gln
50 55 60
Asp Lys Gly Ile Ala Ile Tyr Leu Gly Ile Gln Asn Pro Asp Lys Cys
65 70 75 80
Leu Phe Cys Lys Glu Val Asn Gly His Pro Thr Leu Leu Leu Lys Glu
85 90 95
Glu Lys Ile Leu Asp Leu Tyr His His Pro Glu Pro Met Lys Pro Phe
100 105 110
Leu Phe Tyr His Thr Arg Thr Gly Gly Thr Ser Thr Phe Glu Ser Val
115 120 125
Ala Phe Pro Gly His Tyr Ile Ala Ser Ser Lys Thr Gly Asn Pro Ile
130 135 140
Phe Leu Thr Ser Lys Lys Gly Glu Tyr Tyr Asn Ile Asn Phe Asn Leu
145 150 155 160
Asp Ile Lys Ser
<210> 24
<211> 1395
<212> ДНК
<213> Rattus norvegicus
<400> 24
ggaagctgcc caagctggga tttgatcttc tgaactttcc ataaggactc tttcttgcca 60
ggcaccagaa caagctcatg atggaaaaca atgaaaaaaa tcgttacata tgggtgtgat 120
gttgagatgt gacactaacg aactgggcta tttgtatctt cagctatgtc ctctaaatac 180
ccacattctc catgtactgc ctcagcagga aaagaaactc ctgaccttgg gcaggtttct 240
gatgtggatc agcaggtgtg gatctttcgt gatcaggccc ttgtgacagt tccacgaagc 300
cacactgtaa ctccagtcac tgtgactgtc ctcccatgca agtacccaga gtctcttgag 360
cagggcaaag ggactcccat ttatttggga attcaaaatc cagataaatg cctgttttgt 420
aaggaagtta atggacaccc cactttgctc ctgaaggaag agaagatttt gaatttgtac 480
caccatcctg agccaatgaa gccattcctg ttttaccaca ccctgacagg tgcaacgtcc 540
acctttgaat cagtggtttt ccctggcagc tttattgcct cctccaagat tggcaaaccc 600
atcttcctca catcaaaaaa gggagaacat tacaacattc acttcagttt agatataatt 660
tagatataaa gtcttgaact cagaatggag gtggagggtt ggttagaact cttataactt 720
caaacctcta atatgctatc atttgtttga tgtgtggttg tttcatactg agaagtgagc 780
aaaaccaaca ttatctcatt tcatagatga agaagcaaca aaacagaatg tgtactccaa 840
agtaggttgg atgggagtgg tgtaaggctc tcctctaaga ctgaagtggt ccaacccata 900
aggcatcatg cctttctcag gtgctattat agggtgctgt gccacagtat ataaaaacta 960
gaatgcccca tgtgtgagta gcaacatcct cacttccagt cattggcctg attcatacca 1020
ggttttcaca cacttgtaat accaattcct tgcatttgga tagcaggagc ctcacaatga 1080
catttgctgc caacagctga tagctttgat ccttcagcaa ctgactgcta cgtggatctg 1140
aagacgccag aaaaggaatt agaatcctaa gtgaacgaac agtttttaaa atgtctagaa 1200
ctatttaaac tatgttagaa tacagtggtt tatccttgaa aggtaaggat actgcctggt 1260
aaatcaaaaa cagttaggtc aataaactca gcttgaacaa ttctttcctg gaaacaatgt 1320
gtacaagtaa tgattaaaac attacctttt attattctag acttccataa aaaaaaaaaa 1380
aaaaaaaaaa aaaaa 1395
<210> 25
<211> 165
<212> БЕЛОК
<213> Rattus norvegicus
<400> 25
Met Ser Ser Lys Tyr Pro His Ser Pro Cys Thr Ala Ser Ala Gly Lys
1 5 10 15
Glu Thr Pro Asp Leu Gly Gln Val Ser Asp Val Asp Gln Gln Val Trp
20 25 30
Ile Phe Arg Asp Gln Ala Leu Val Thr Val Pro Arg Ser His Thr Val
35 40 45
Thr Pro Val Thr Val Thr Val Leu Pro Cys Lys Tyr Pro Glu Ser Leu
50 55 60
Glu Gln Gly Lys Gly Thr Pro Ile Tyr Leu Gly Ile Gln Asn Pro Asp
65 70 75 80
Lys Cys Leu Phe Cys Lys Glu Val Asn Gly His Pro Thr Leu Leu Leu
85 90 95
Lys Glu Glu Lys Ile Leu Asn Leu Tyr His His Pro Glu Pro Met Lys
100 105 110
Pro Phe Leu Phe Tyr His Thr Leu Thr Gly Ala Thr Ser Thr Phe Glu
115 120 125
Ser Val Val Phe Pro Gly Ser Phe Ile Ala Ser Ser Lys Ile Gly Lys
130 135 140
Pro Ile Phe Leu Thr Ser Lys Lys Gly Glu His Tyr Asn Ile His Phe
145 150 155 160
Ser Leu Asp Ile Ile
165
<210> 26
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 26
tacatttgaa ggtcttagca ttatgcttct aaatgactgg gctagtttgg gcgatcgcga 60
caagaagggt tgatttactc tgtaggtgag tacagagtaa agaactct 108
<210> 27
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 27
caccctgagc acatggggaa tccttcttgg gttgcaacat gtggaaggac ctcgagataa 60
cttcgtataa tgtatgctat acgaagttat atgcatggcc tccgcgccgg gttttggcgc 120
ctcccgcggg cgcccccctc 140
<210> 28
<211> 181
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 28
tttcactgca ttctagttgt ggtttgtcca aactcatcaa tgtatcttat catgtctgga 60
ataacttcgt ataatgtatg ctatacgaag ttatgctagt aactataacg gtcctaaggt 120
agcgagctag ccagaatcaa tgacaaagat gaagaatata tttaacctca taaacttggt 180
t 181
<210> 29
<211> 177
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 29
caccctgagc acatggggaa tccttcttgg gttgcaacat gtggaaggac ctcgagataa 60
cttcgtataa tgtatgctat acgaagttat gctagtaact ataacggtcc taaggtagcg 120
agctagccag aatcaatgac aaagatgaag aatatattta acctcataaa cttggtt 177
<210> 30
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 30
ctggctctgt caactagatt ggaaggcatg cggagctgcc agtctcaact cgtcgacgtc 60
actctccttt cccactgcat gtgaagcatt tgagtgtctg 100
<210> 31
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 31
tgcactcaga aaagacaagt attcacattt tttcttgtgg ctgatctgga ctcgagataa 60
cttcgtataa tgtatgctat acgaagttat atgcatggcc tccgcgccgg gttttggcgc 120
ctcccgcggg cgcccccctc 140
<210> 32
<211> 181
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 32
tttcactgca ttctagttgt ggtttgtcca aactcatcaa tgtatcttat catgtctgga 60
ataacttcgt ataatgtatg ctatacgaag ttatgctagt aactataacg gtcctaaggt 120
agcgagctag cctatgcctg cacagccctt ccataggtac taaggattta aactctcgtt 180
t 181
<210> 33
<211> 177
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 33
tgcactcaga aaagacaagt attcacattt tttcttgtgg ctgatctgga gtcgagataa 60
cttcgtataa tgtatgctat acgaagttat gctagtaact ataacggtcc taaggtagcg 120
agctagccta tgcctgcaca gcccttccat aggtactaag gatttaaact ctcgttt 177
<210> 34
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 34
gggtccatta tttgagactt tcca 24
<210> 35
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 35
agctttctgc tggccccatt acttg 25
<210> 36
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 36
gcatgtgcct gtcttcaca 19
<210> 37
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 37
gagtggtaca acctctacat ttgag 25
<210> 38
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 38
ctgcagcctt ccctggttgg ttc 23
<210> 39
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 39
agtgggcagc tccctttaga 20
<210> 40
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 40
cccactgcat gtgaagcat 19
<210> 41
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 41
ttgagtgtct gcacctgagc ca 22
<210> 42
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 42
ggagacctct tattagcctg tga 23
<210> 43
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 43
cacctgccct aagtcatctc 20
<210> 44
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 44
tacagcaaga gcagaggcca ca 22
<210> 45
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 45
gcctgcagag aagcaatgtt c 21
<210> 46
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 46
cagccgctac acaccacaa 19
<210> 47
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 47
ccagctgcta cacaaatgca gggc 24
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 48
cgtcatctcc tgccagttca 20
<210> 49
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 49
ctgctgctca gagcattgaa a 21
<210> 50
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 50
ccatggccag ggaaggctta cta 23
<210> 51
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 51
cgcctccgac tgaacatatg ac 22
<210> 52
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 52
cctgatatgc atctttccct atgga 25
<210> 53
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 53
atggcacctc agaccagacc cac 23
<210> 54
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 54
ggcagcaata atactgggac aaac 24
<210> 55
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 55
ggccaggaaa cacatctgaa g 21
<210> 56
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 56
agcagtgact ctaaatgctc agtgtga 27
<210> 57
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 57
gcctcactcc aacaacatta tga 23
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН КЛАСТЕРА ДИФФЕРЕНЦИРОВКИ 47 | 2015 |
|
RU2728412C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН КЛАСТЕРА ДИФФЕРЕНЦИРОВКИ 47 | 2015 |
|
RU2827833C1 |
| АНТИТЕЛО ПРОТИВ IL-17A И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2816204C2 |
| НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ ГУМАНИЗИРОВАННЫЙ КОМПЛЕКС CD3 | 2015 |
|
RU2726446C2 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ДЛЯ ВЫРАБОТКИ ТЕРАПЕВТИЧЕСКИХ АНТИТЕЛ ПРОТИВ КОМПЛЕКСОВ ПЕПТИД-MHC, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ВАРИАНТЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2819525C2 |
| АНТИТЕЛО, СВЯЗЫВАЮЩЕЕ ЧЕЛОВЕЧЕСКИЙ IL-4R, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2779649C1 |
| ГУМАНИЗИРОВАННЫЕ МЫШИ С НОКИНОМ SIRPA-IL15 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2822370C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ РЕЦЕПТОРА ИНТЕРЛЕЙКИНА 36 (IL-36R) | 2016 |
|
RU2745898C2 |
| ГРЫЗУНЫ, ХАРАКТЕРИЗУЮЩИЕСЯ ГУМАНИЗИРОВАННЫМ ГЕНОМ TMPRSS | 2017 |
|
RU2749715C2 |
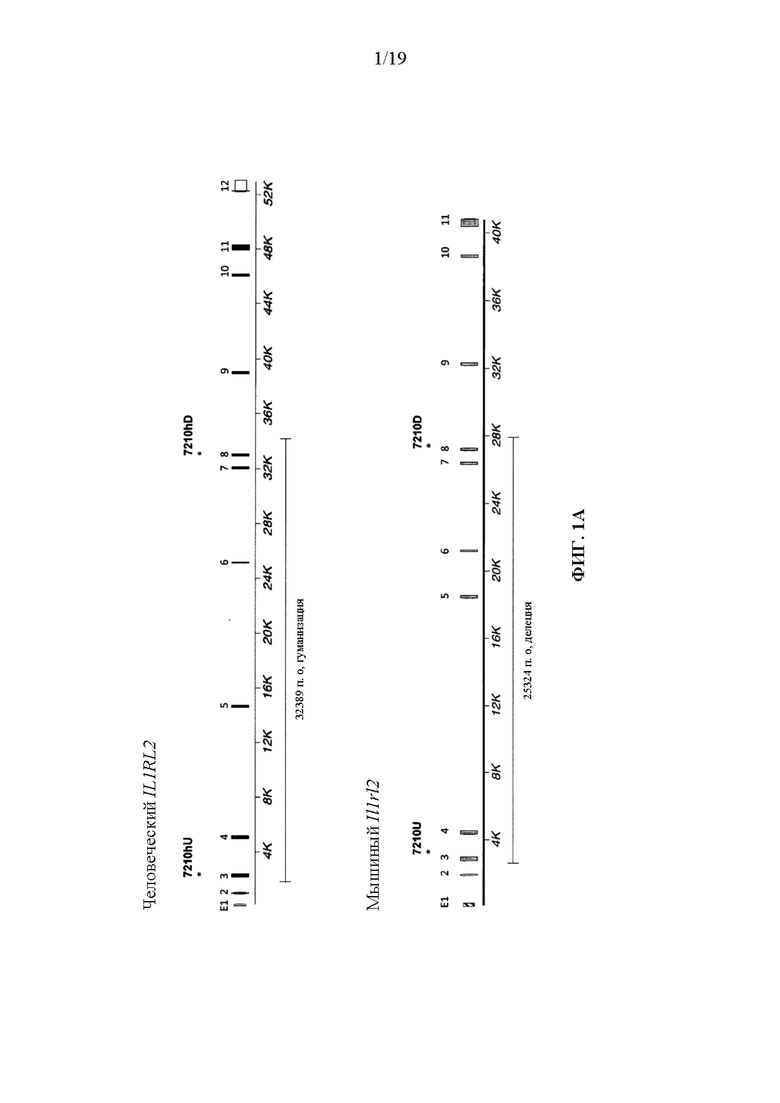
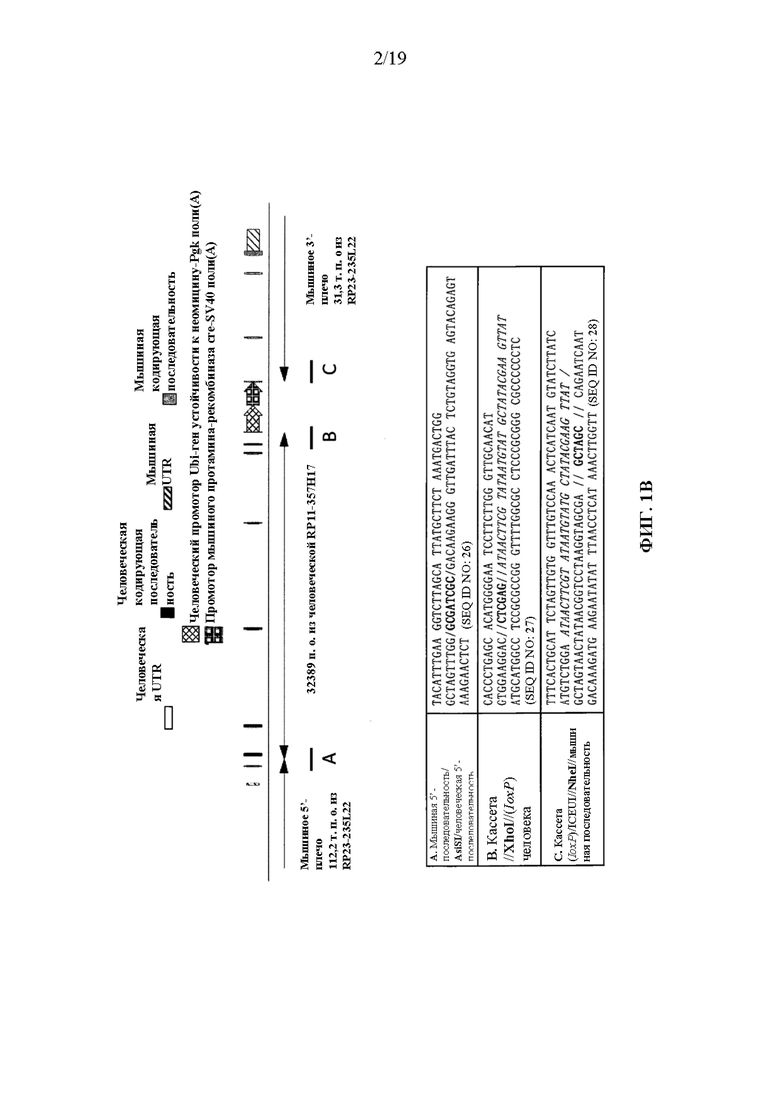
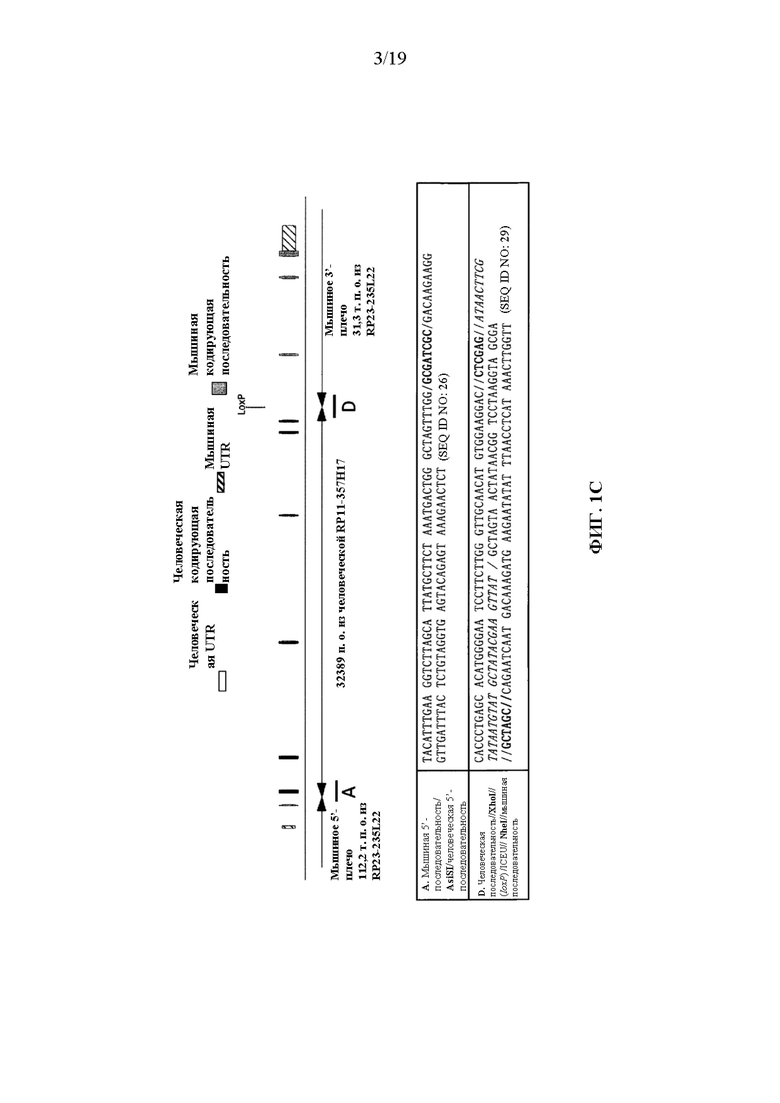
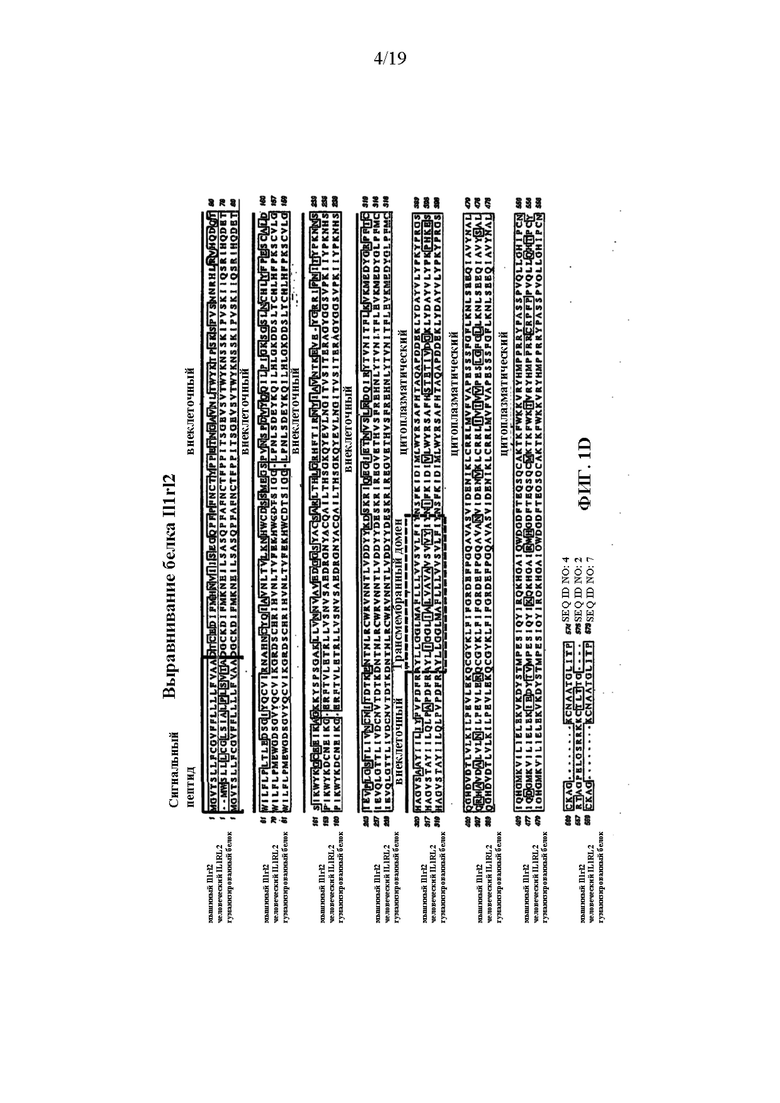
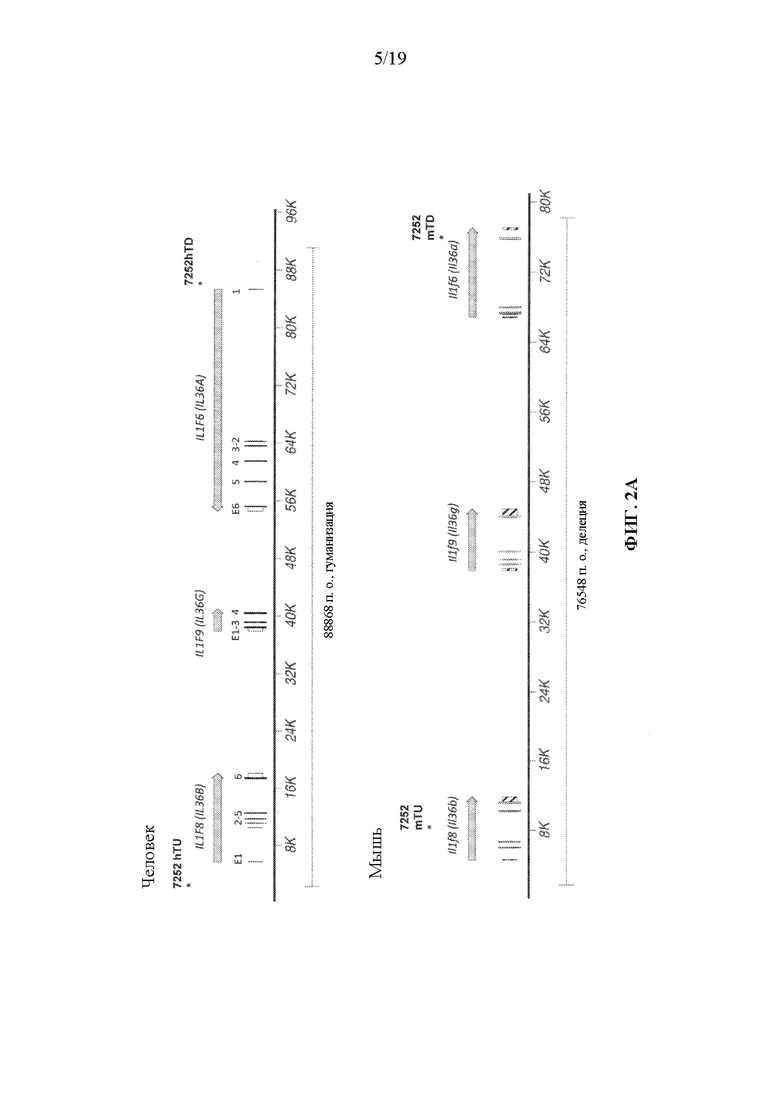

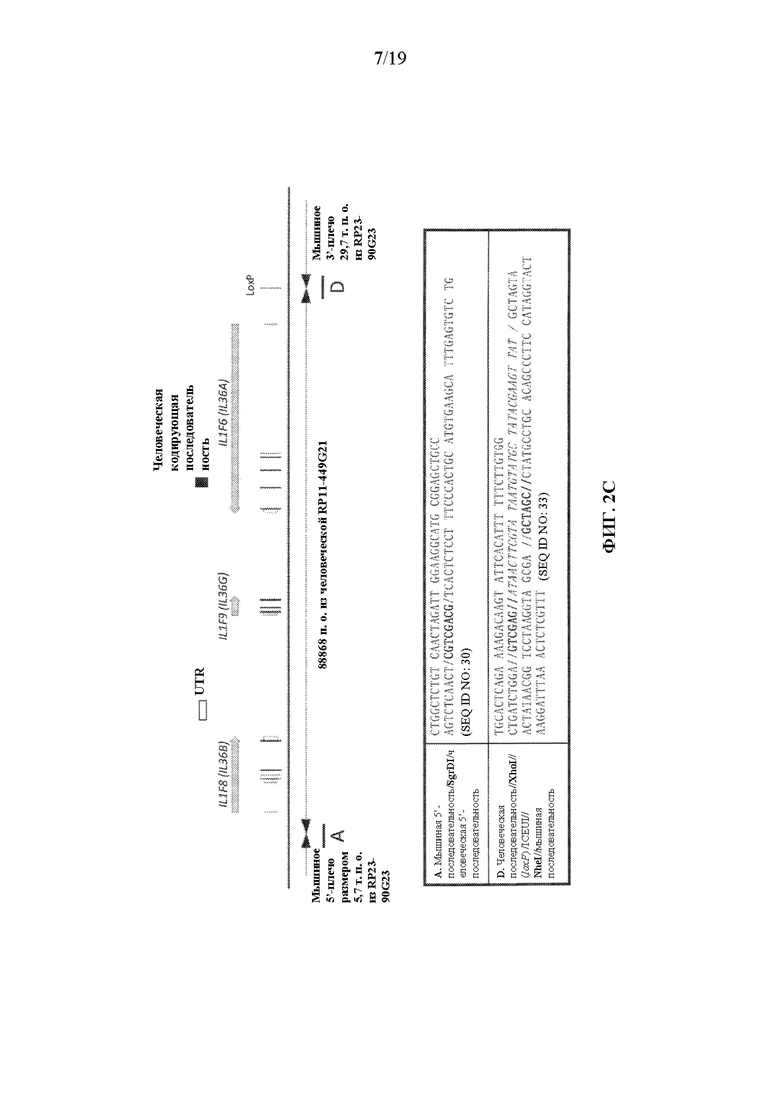

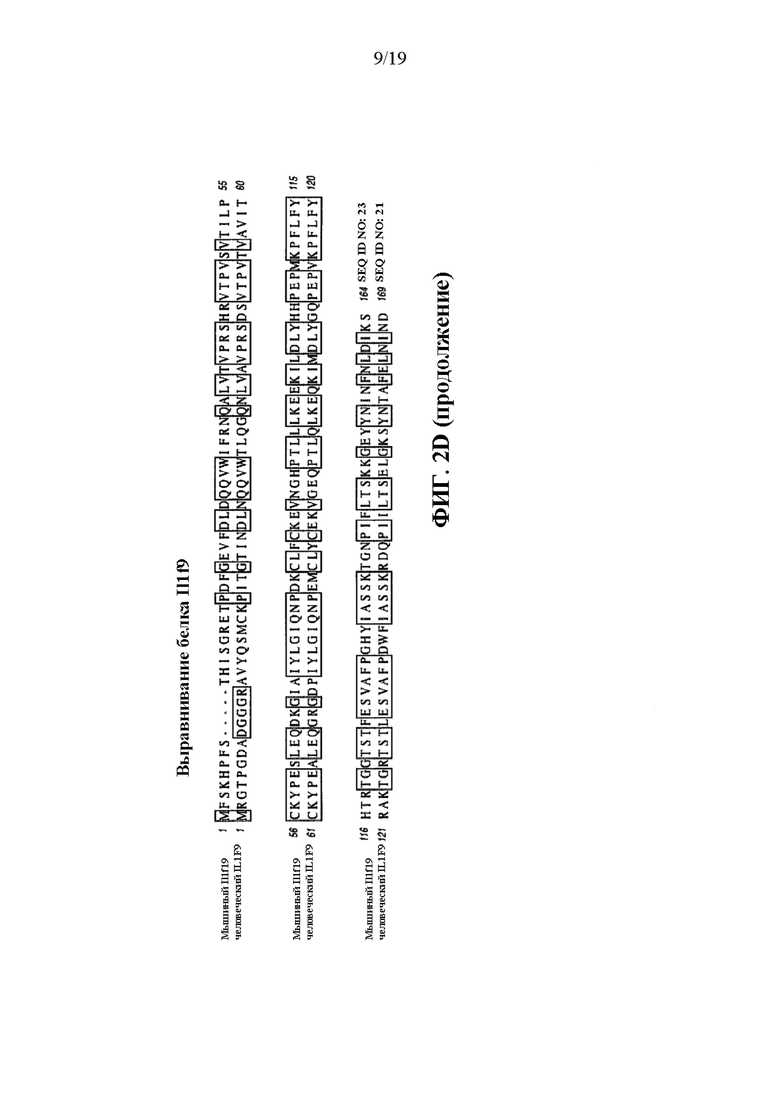
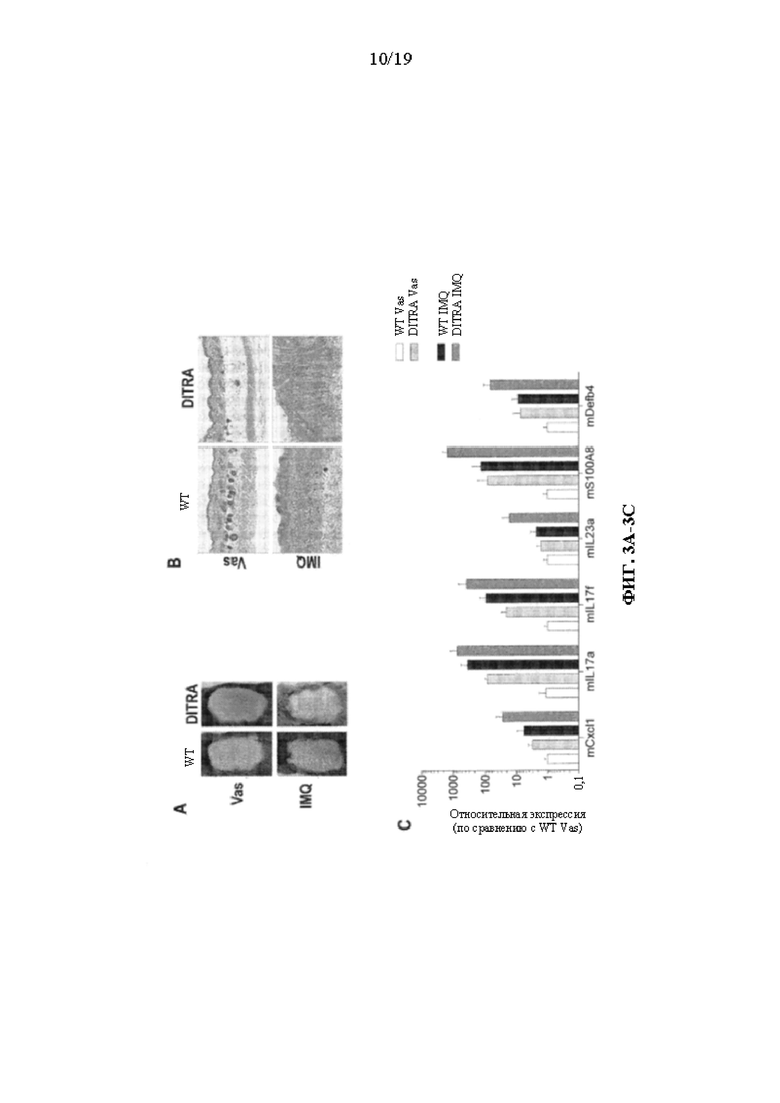
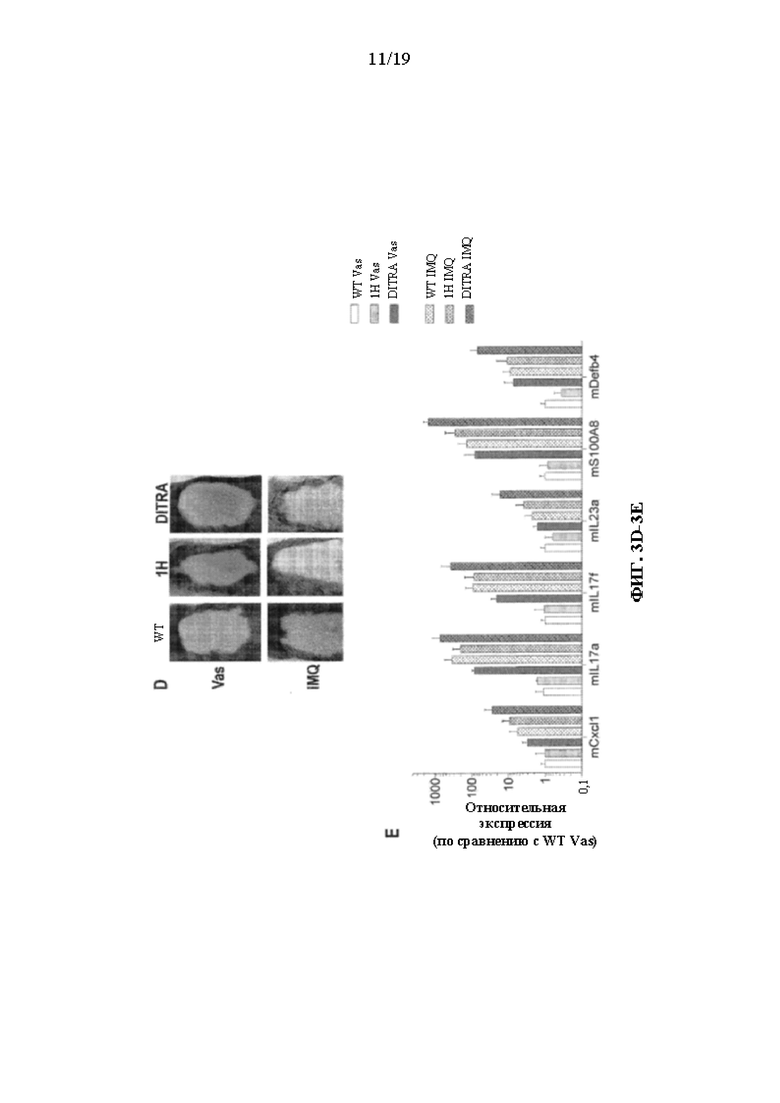
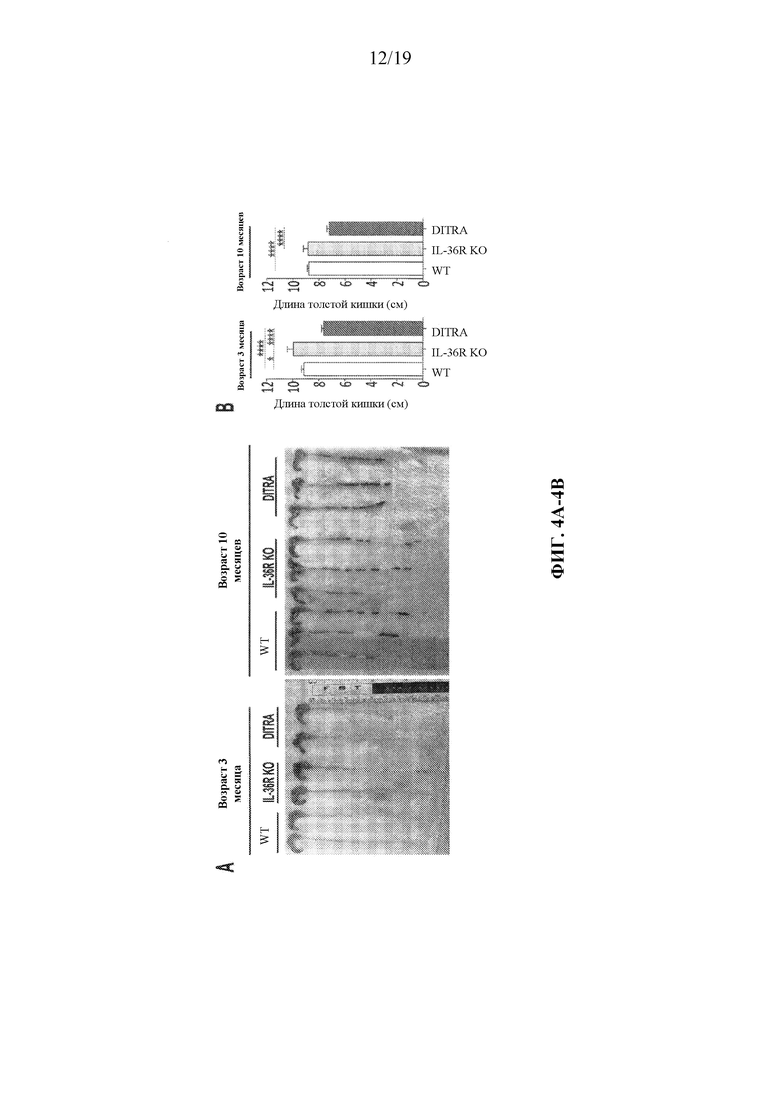
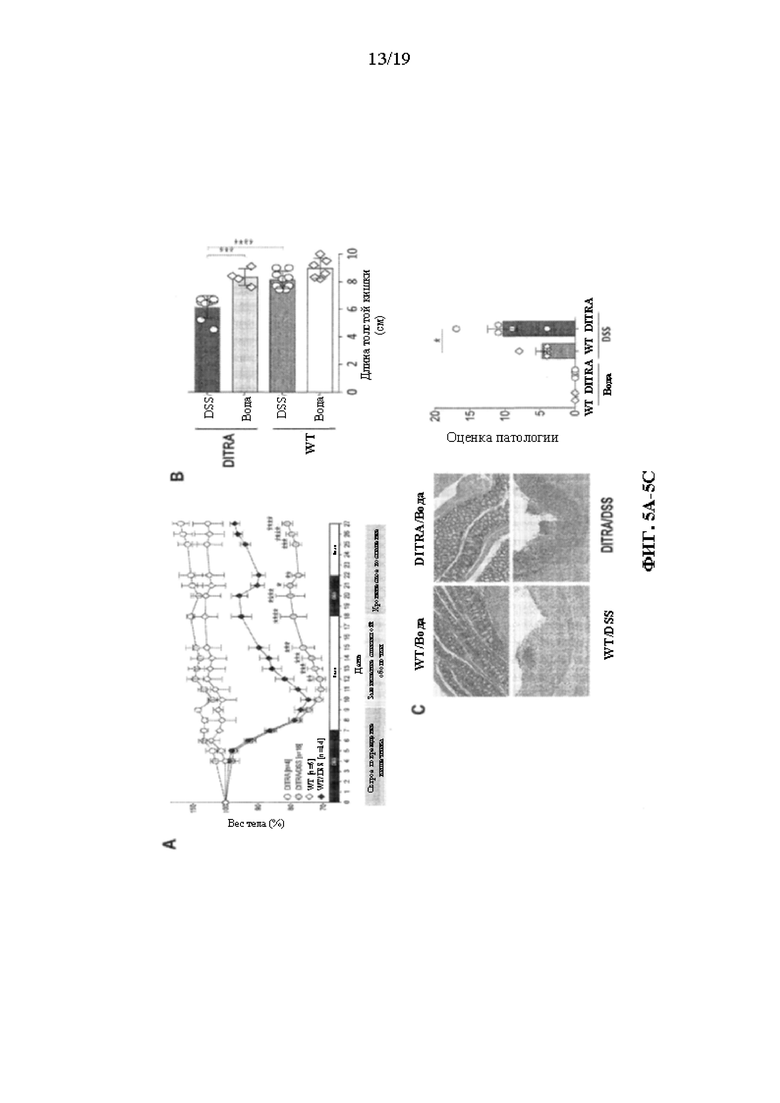
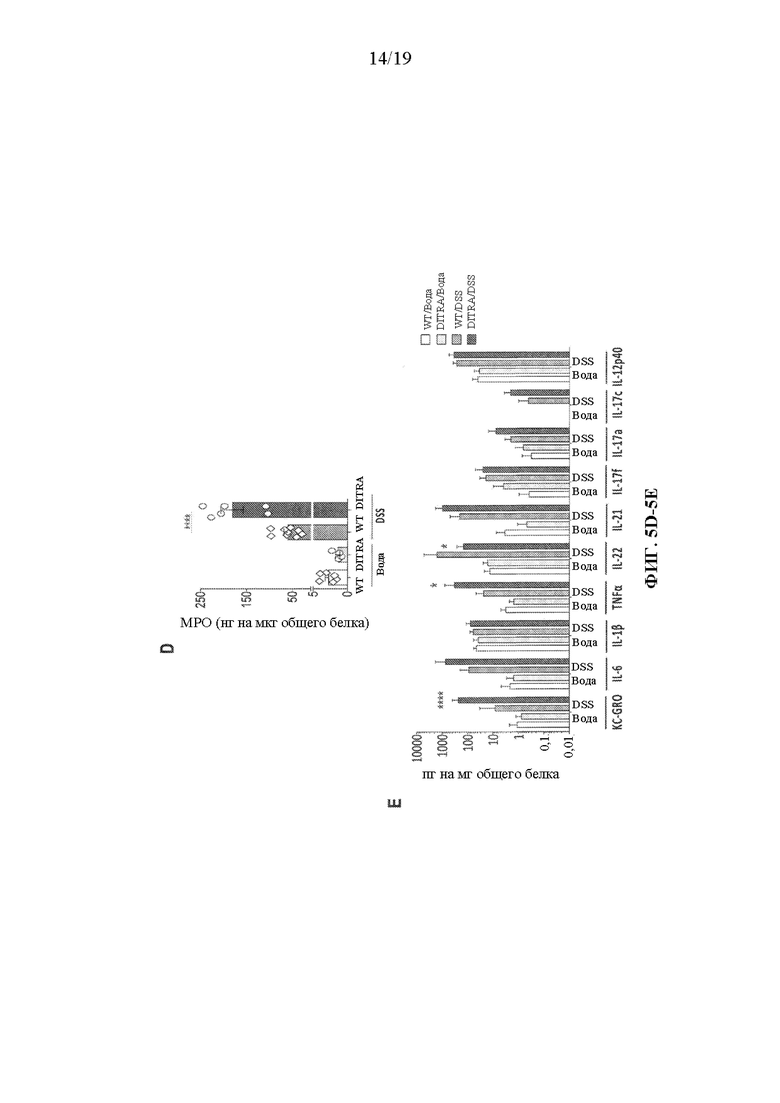

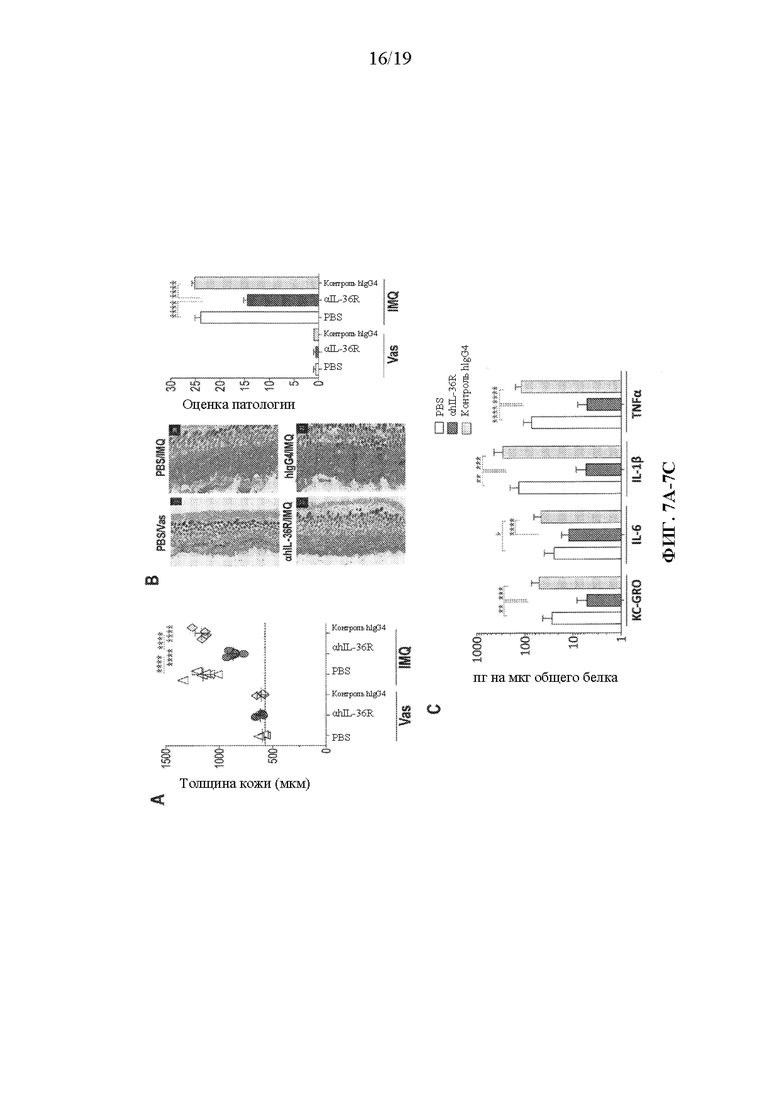
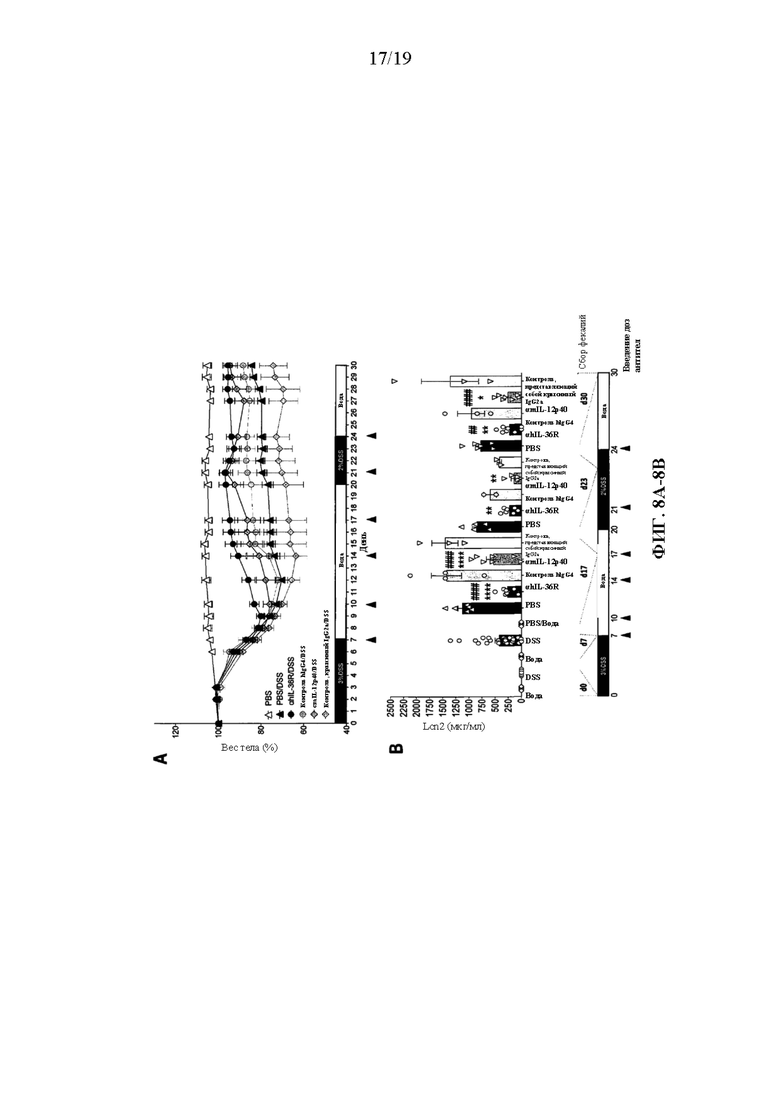

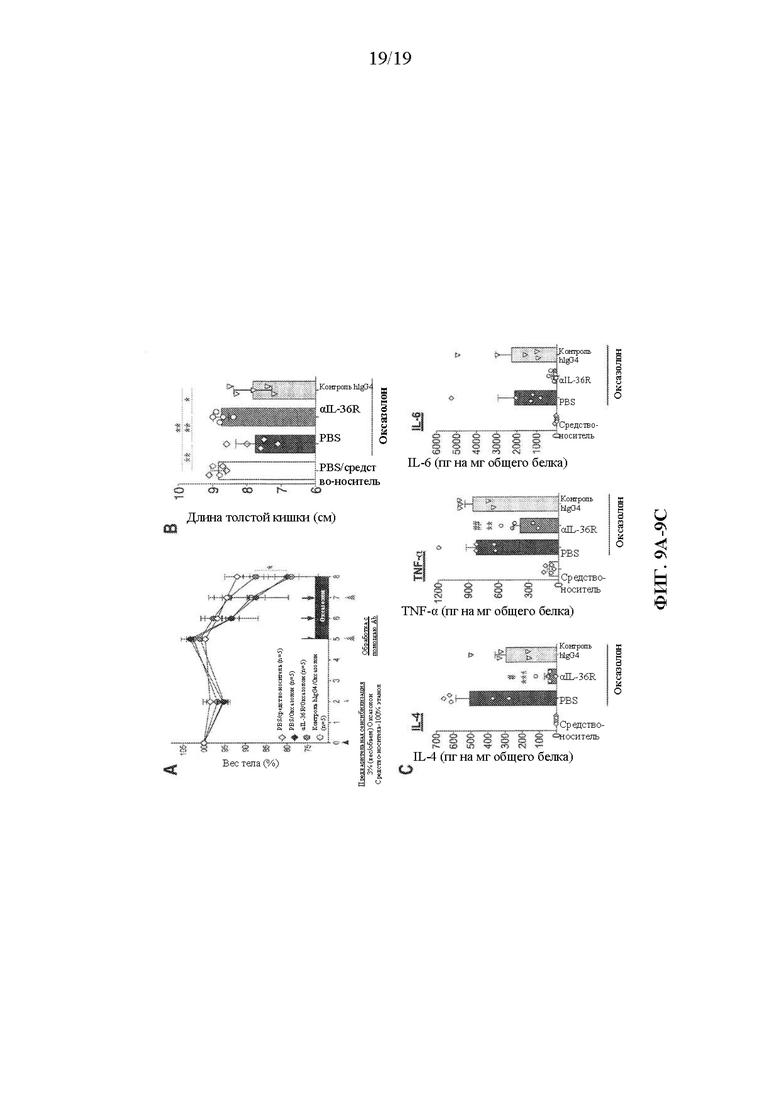
Изобретение относится к области биотехнологии, в частности к генетически модифицированному грызуну, геном которого содержит гуманизированный ген Il1rl2 функционально связанный с эндогенным промотором Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна, человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна и человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна, к его клетке и ткани, а также к способу его получения. Также раскрыты нацеливающаяся конструкция нуклеиновой кислоты для образования гуманизированного гена Il1rl2 в геноме грызуна и нацеливающаяся конструкция нуклеиновой кислоты для интеграции человеческого гена IL1F6, человеческого гена IL1F8 и человеческого гена IL1F9 в геном грызуна. Изобретение эффективно для оценки терапевтической эффективности соединения-кандидата для лечения заболевания, ассоциированного с нарушением регуляции передачи сигнала IL-36. 10 н. и 35 з.п. ф-лы, 9 ил., 6 табл., 2 пр.
1. Генетически модифицированный грызун для экспрессии гуманизированного гена Il1rl2, человеческого гена Il1f6, человеческого гена Il1f8 и человеческого гена Il1f9, геном которого содержит:
(1) гуманизированный ген Il1rl2 функционально связанный с эндогенным промотором Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит сигнальный пептид, эктодомен, характеризующийся по меньшей мере 95% идентичностью последовательности эктодомена человеческого белка IL1RL2 и трансмембранно-цитоплазматическую последовательность;
(2) человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна;
(3) человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна; и
(4) человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна,
где грызун представляет собой мышь или крысу и не способен экспрессировать ген Il1rl2 грызуна, ген Il1f6 грызуна, ген Il1f8 грызуна и ген Il1f9 грызуна.
2. Генетически модифицированный грызун по п. 1, где транс мембранно-цитоплазматическая последовательность гуманизированного белка Il1rl2 характеризуется по меньшей мере 95% идентичностью с трансмембранно-цитоплазматической последовательностью эндогенного белка Il1rl2 грызуна.
3. Генетически модифицированный грызун по п. 1 или 2, где сигнальный пептид гуманизированного белка Il1rl2 характеризуется по меньшей мере 95% идентичностью с сигнальным пептидом эндогенного белка Il1rl2 грызуна.
4. Генетически модифицированный грызун по любому из пп. 1-3, где человеческий белок IL1RL2 содержит аминокислотную последовательность, представленную под SEQ ID NO: 2.
5. Генетически модифицированный грызун по любому из пп. 1-3, где эктодомен гуманизированного белка Il1rl2 содержит аминокислоты 22-337 из SEQ ID NO: 7.
6. Генетически модифицированный грызун по любому из пп. 1-3, где гуманизированный белок Il1rl2 содержит аминокислотную последовательность, представленную под SEQ ID NO: 7.
7. Генетически модифицированный грызун по любому из пп. 1-6, где гуманизированный ген Il1rl2 является результатом замены геномного фрагмента эндогенного гена Il1rl2 грызуна в эндогенном локусе Il1rl2 грызуна на нуклеотидную последовательность человеческого гена IL1RL2.
8. Генетически модифицированный грызун по п. 7, где нуклеотидная последовательность человеческого гена IL1RL2 представляет собой геномный фрагмент человеческого гена IL1RL2, который кодирует полипептид, характеризующийся по меньшей мере 95% идентичностью последовательности эктодомена человеческого белка IL1RL2.
9. Генетически модифицированный грызун по п. 8, где геномный фрагмент человеческого гена IL1RL2 содержит экзоны 3-8 человеческого гена IL1RL2.
10. Генетически модифицированный грызун по любому из пп. 7-9, где геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены, содержит экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
11. Генетически модифицированный грызун по любому из пп. 7-10, где геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна.
12. Генетически модифицированный грызун по п. 7, где гуманизированный ген Il1rl2 содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна, экзоны 3-8 человеческого гена IL1RL2 и оставшиеся экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
13. Генетически модифицированный грызун по любому из предыдущих пунктов, где человеческий ген IL1F6 заменяет эндогенный ген Il1f6 грызуна в эндогенном локусе Il1f6 грызуна.
14. Генетически модифицированный грызун по любому из предыдущих пунктов, где человеческий ген IL1F8 заменяет эндогенный ген Il1f8 грызуна в эндогенном локусе Il1f8 грызуна.
15. Генетически модифицированный грызун по любому из предыдущих пунктов, где человеческий ген IL1F9 заменяет эндогенный ген Il1f9 грызуна в эндогенном локусе Il1f9 грызуна.
16. Генетически модифицированный грызун по любому из предыдущих пунктов, где грызун является гомозиготным по каждому из гуманизированного гена Il1rl2, человеческого гена IL1F6, человеческого гена IL1F8 и человеческого гена IL1F9.
17. Генетически модифицированный грызун по любому из пунктов 1-16, где грызун представляет собой мышь.
18. Генетически модифицированный грызун по любому из пунктов 1-16, где грызун представляет собой крысу.
19. Способ получения генетически модифицированного грызуна, включающий
модификацию генома грызуна для содержания
(1) гуманизированного гена Il1rl2, функционально связанного с эндогенным промотором Il1rl2 грызуна в эндогенном локусе Illrl2 грызуна, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит сигнальный пептид, эктодомен, характеризующийся по меньшей мере 95% идентичностью последовательности эктодомена человеческого белка IL1RL2 и трансмембранно-цитоплазматическую последовательность;
(2) человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна;
(3) человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна; и
(4) человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна; и
получение грызуна, содержащего модифицированный геном, где грызун представляет собой мышь или крысу и не способен экспрессировать ген Il1rl2 грызуна, ген Il1f6 грызуна, ген Il1f8 грызуна и ген Il1f9 грызуна.
20. Способ по п. 19, где геном грызуна модифицируют с помощью способа, включающего
(i) получение первого грызуна, содержащего гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна;
(ii) получение второго грызуна, содержащего человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна, человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна и человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна; и
(iii) скрещивание первого грызуна со вторым грызуном с получением грызуна, содержащего модифицированный геном грызуна.
21. Способ по п. 20, где грызуна, содержащего гуманизированный ген Il1rl2 в эндогенном локусе Il1rl2 грызуна, получают посредством
получения эмбриональной стволовой (ES) клетки грызуна,
вставки нуклеотидной последовательности человеческого гена IL1RL2 в локус Il1rl2 грызуна ES-клетки грызуна с образованием гуманизированного гена Il1rl2 в локусе Il1rl2 грызуна, с получением таким образом ES-клетки грызуна, содержащей гуманизированный ген Il1rl2, и
получения грызуна с использованием ES-клетки грызуна, содержащей гуманизированный ген Il1rl2.
22. Способ по п. 21, где нуклеотидная последовательность человеческого гена IL1RL2 заменяет геномный фрагмент гена Il1rl2 грызуна в локусе Il1rl2 грызуна.
23. Способ по п. 22, где нуклеотидная последовательность человеческого гена IL1RL2 представляет собой геномный фрагмент человеческого гена IL1RL2, который кодирует полипептид, характеризующийся по меньшей мере 95% идентичностью последовательности эктодомена человеческого белка IL1RL2.
24. Способ по п. 23, где геномный фрагмент человеческого гена IL1RL2 содержит экзоны 3-8 человеческого гена IL1RL2.
25. Способ по любому из пп. 22-24, где геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены, содержит экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
26. Способ по любому из пп. 22-25, где геномная последовательность эндогенного гена Il1rl2 грызуна, оставшаяся после замены, содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна.
27. Способ по п. 21, где гуманизированный ген Il1rl2 содержит экзоны 1-2 эндогенного гена Il1rl2 грызуна, экзоны 3-8 человеческого гена IL1RL2 и оставшиеся экзоны, расположенные ниже экзона 8 эндогенного гена Il1rl2 грызуна.
28. Способ по п. 20, где второго грызуна, содержащего человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9, получают посредством:
получения эмбриональной стволовой (ES) клетки грызуна,
вставки:
(i) человеческого гена IL1F6 в локус Il1f6 грызуна ES-клетки грызуна,
(ii) человеческого гена IL1F8 в локус Il1f8 грызуна ES-клетки грызуна, и
(iii) человеческого гена IL1F9 в локус Il1f9 грызуна ES-клетки грызуна,
с получением таким образом ES-клетки грызуна, содержащей человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9, и
получения второго грызуна с использованием ES-клетки грызуна, содержащей человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9.
29. Способ по п. 28, где человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9 представлены в непрерывной молекуле нуклеиновой кислоты.
30. Способ по любому из пп. 19-29, где человеческий ген IL1F6 заменяет эндогенный ген Il1f6 грызуна в эндогенном локусе Il1f6 грызуна.
31. Способ по любому из пп. 19-30, где человеческий ген IL1F8 заменяет эндогенный ген Il1f8 грызуна в эндогенном локусе Il1f8 грызуна.
32. Способ по любому из пп. 19-31, где человеческий ген IL1F9 заменяет эндогенный ген Il1f9 грызуна в эндогенном локусе Il1f9 грызуна.
33. Выделенная клетка грызуна по любому из пп. 1-18 для экспрессии человеческого или гуманизированного генов, где выделенная клетка содержит гуманизированный ген Il1rl2, человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9.
34. Выделенная ткань грызуна по любому из пп. 1-18 для экспрессии человеческого или гуманизированного генов, где выделенная ткань содержит гуманизированный ген Il1rl2, человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9.
35. Эмбриональная стволовая (ES) клетка грызуна для получения грызуна, содержащая
(1) гуманизированный ген Il1rl2 функционально связанный с эндогенным промотором Il1rl2 грызуна в эндогенном локусе Illrl2 грызуна, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит сигнальный пептид, эктодомен, характеризующийся по меньшей мере 95% идентичностью последовательности эктодомена человеческого белка IL1RL2 и трансмембранно-цитоплазматическую последовательность;
(2) человеческий ген IL1F6 в эндогенном локусе Il1f6 грызуна;
(3) человеческий ген IL1F8 в эндогенном локусе Il1f8 грызуна; и
(4) человеческий ген IL1F9 в эндогенном локусе Il1f9 грызуна;
где ES клетка грызуна представляет собой ES клетку мыши для получения мыши или ES клетку крысы для получения крысы, и не способна экспрессировать ген Il1rl2 грызуна, ген Il1f6 грызуна, ген Il1f8 грызуна и ген Il1f9 грызуна.
36. Применение ES-клетки грызуна по п. 35 для получения грызуна.
37. Применение грызуна по любому из пп. 1-18 в качестве модели заболеваний, ассоциированных с нарушением регуляции передачи сигнала IL-36, на грызунах.
38. Способ оценки терапевтической эффективности соединения-кандидата для лечения заболевания, ассоциированного с нарушением регуляции передачи сигнала IL-36, включающий:
получение грызуна по любому из пп. 1-18,
введение грызуну средства для индукции воспаления,
введение грызуну соединения-кандидата и
определение того, подавляет ли соединение-кандидат индуцированное воспаление.
39. Способ по п. 38, где средство представляет собой декстрансульфат натрия (DSS) или оксазолон, и у грызуна индуцируют воспаление кишечника.
40. Способ по п. 38, где средство представляет собой имиквимод (IMQ), и у грызуна индуцируют воспаление кожи.
41. Способ по любому из пп. 38-40, где соединение-кандидат вводят грызуну до, во время или после введения средства.
42. Способ по любому из пп. 38-41, где соединение-кандидат представляет собой низкомолекулярное соединение, ингибитор на основе нуклеиновой кислоты или антитело.
43. Способ по п. 42, где соединение-кандидат представляет собой антитело.
44. Нацеливающаяся конструкция нуклеиновой кислоты для образования гуманизированного гена Il1rl2 в геноме грызуна, содержащая
нуклеотидную последовательность человеческого гена IL1RL2, фланкированную 5'- и 3'-нуклеотидными последовательностями грызуна, способными опосредовать гомологичную рекомбинацию и интеграцию нуклеотидной последовательности человеческого гена ILRL2 в локус Il1rl2 грызуна с образованием гуманизированного гена Il1rl2, где гуманизированный ген Il1rl2 кодирует гуманизированный белок Il1rl2, который содержит эктодомен, характеризующийся по меньшей мере 95% идентичностью последовательности эктодомена человеческого белка IL1RL2.
45. Нацеливающаяся конструкция нуклеиновой кислоты для интеграции человеческого гена IL1F6, человеческого гена IL1F8 и человеческого гена IL1F9 в геном грызуна, содержащая непрерывную последовательность нуклеиновой кислоты, содержащую человеческий ген IL1F6, человеческий ген IL1F8 и человеческий ген IL1F9,
где непрерывная последовательность нуклеиновой кислоты фланкирована 5'- и 3'-нуклеотидными последовательностями грызуна, способными опосредовать гомологичную рекомбинацию и интеграцию непрерывной последовательности нуклеиновой кислоты в локус грызуна, охватывающей ген Il1f6 грызуна, ген Il1f8 грызуна и ген Il1f9 грызуна.
| WO2013063556 A1, 02.05.2013 | |||
| WO2015171861 A1, 12.11.2015 | |||
| AKITAKA SHIBATA M.D., PH.D | |||
| et al., Toll-like receptor 4 antagonist TAK-242 inhibits autoinflammatory symptoms in DITRA, Journal of Autoimmunity, 2017, Vol | |||
| Капельная масленка с постоянным уровнем масла | 0 |
|
SU80A1 |
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| COWEN E.W | |||
| et al., DIRA, DITRA, and New Insights Into Pathways of Skin Inflammation, Archives of Dermatology, 2012, | |||
