ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее раскрытие относится к антителам против IL-5 (интерлейкин-5) и их антигенсвязывающим фрагментам. Кроме того, настоящее раскрытие также относится к химерным антителам, гуманизированным антителам, содержащим области CDR (гипервариабельная область) антител против IL-5, и настоящее раскрытие также относится к фармацевтической композиции, содержащей антитело против IL-5 и его антигенсвязывающий фрагмент, и к его применению в качестве диагностического и терапевтического средства для заболеваний, связанных с IL-5.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Интерлейкин-5 (IL-5) представляет собой одного из важных членов семейства интерлейкинов, также известного как фактор замены Т-клеток (TRF), фактор-II роста В-клеток (BCGF-II), фактор усиления IgA (IgA-EF) или фактор дифференциации эозинофилов (EDF). Он представляет собой гомодимерный гликопротеин, секретируемый, главным образом, хэлперной Т-клеткой 2 (Th2). Человеческий IL-5 состоит из 134 аминокислотных остатков, включающих сигнальный пептид, состоящий из 22 аминокислот и двух сайтов гликозилирования. Человеческий IL-5 имеет 70%-ную идентичность с мышиным IL-5 на уровне аминокислот. Активный IL-5 находится в форме олигодимера с двумя пептидными цепями, связанными друг с другом посредством дисульфидной(ных) связи(зей) и в антипараллельной конфигурации, тогда как мономеры IL-5 не являются биологически активными (Adv Immunol. 1994; 57:145-90).
Эозинофилы (EOS) ассоциированы с целым рядом воспалительных заболеваний в легком, включая аллергические заболевания, ассоциированные с анафилактической реакцией. Среди данных заболеваний астма представляет собой хроническое респираторное воспалительное заболевание, поражающее приблизительно 300 миллионов пациентов во всем мире, с заболеваемостью 10%. Ее патогенез ассоциирован с целым рядом цитокинов, и IL-5, и его рецептор IL-5R играют важную роль в патогенезе астмы. Имеется большое количество воспалительных клеток, инфильтрующих бронхо-легочную ткань пациентов с астмой, среди которых эозинофилы наиболее значительно увеличиваются в числе. Многие исследования продемонстрировали то, что эозинофилы являются одними из главных клеток, приводящих к воспалению дыхательных путей при астме (Curr Opin Pulm Med. 2005 Jan; 11(1): 1-6). IL-5 играет крайне важную роль в дифференциации, созревании, адгезии, инфильтрации и апоптозе EOS. В большом числе исследований на животных и клинических исследований продемонстрировали то, что IL-5 может активировать клетки-предшественники EOS в костном мозге и инициировать агрегацию EOS в периферической крови и дыхательных путях, приводя к хроническому воспалению и гиперчувствительности дыхательных путей (J Immunol. 2014 Oct 15; 193(8):4043-52). Кроме того, IL-5 может продлевать продолжительность выживания EOS, усиливать их дегранулирующий ответ на специфические стимулирующие факторы (такие как IgA или IgG) и опосредовать хемотактическую активность эозинофилов (J Asthma Allergy. 2015 Nov 3; 8: 125-34). Повышенную экспрессию IL-5 выявляли как у пациентов с астмой, так и в человеческих моделях, индуцированных бронхиальными антигенами (Greenfeder et al., Respiratory Research, 2: 71-79, 2001). Рекомбинантный человеческий белок IL-5, принимаемый пациентами с астмой, будет приводить к увеличенному числу эозинофилов, бронхиальной гиперчувствительности и высвобождению токсичных частиц эозинофилами, указывая на то, что IL-5 представляет собой ключевой фактор в патогенезе астмы.
В настоящее время самым эффективным способом лечения астмы является ингибирование экспрессии некоторых ключевых медиаторов (включая IL-5) при астме посредством назального или перорального введения стеринов для облегчения воспаления в легком. Однако долговременное применение стеринов имеет много побочных эффектов. Следовательно, необходимо находить новые фармацевтические мишени для лечения астмы. Исследования продемонстрировали то, что посредством ингибирования связывания IL-5 с его рецептором антитела против IL-5 могут значительно снижать накопление эозинофилов в легком, уменьшать уровень эозинофилов в крови, ткани и мокроте, уменьшать опосредованный эозинофилами воспалительный ответ, улучшать функцию легкого и демонстрировать хорошую эффективность для лечения тяжелой эозинофильной астмы и рецидивирующей астмы (Drugs. 2017 May; 77(7): 777-784). В настоящее время только антитела против IL-5 меполизумаб от GSK и реслизумаб от Teva Pharma имеются в продаже. Другие антитела против мишени - IL-5 - находятся в фазе доклинического исследования. Родственными патентами являются, например, WO 2017033121, WO 2016040007, WO 2015095539, WO 2012083370, WO 2012158954, WO 2006046689, WO 9621000, WO 9535375 и т.д. Однако все же имеется потребность в улучшении индуцированного IL-5 устранения эозинофилов и улучшении функции легкого. Следовательно, необходимо продолжать разрабатывать антитела с высокой селективностью, высокой аффинностью и хорошей эффективностью для обеспечения более предпочтительных схем лечения против IL-5 при астме.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно настоящему раскрытию предложены моноклональные антитела или антигенсвязывающие фрагменты (также именуемые молекулы, связывающиеся с IL-5), которые специфично связываются с аминокислотной последовательностью IL-5 или трехмерной структурой.
В одном аспекте согласно данному раскрытию предложено моноклональное антитело или его антигенсвязывающийся фрагмент, связывающийся с человеческим IL-5, где данное моноклональное антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(i) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 16-18, или варианты HCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 16-18, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 19-21, или варианты LCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 19-21 соответственно; или
(ii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 22-24, или варианты HCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 22-24, соответственно, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 25-27, или варианты LCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 25-27 соответственно; или
(iii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 28-30, или варианты HCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 28-30, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 31-33, или варианты LCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 31-33 соответственно; или
(iv) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 34-36, или варианты HCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 34-36, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 37-39, или варианты LCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 37-39 соответственно; или
(v) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 40-42, или варианты HCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 40-42, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 43-45, или варианты LCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 43-45 соответственно; или
(vi) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 34-36, или варианты HCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 34, 82 и 36, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 37-39, или варианты LCDR, имеющие 3, 2 или 1 отличие(чия) аминокислот от области LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 37-39 соответственно.
В некоторых воплощениях варианты моноклонального антитела или CDR антигенсвязывающего фрагмента (включающие 3 CDR тяжелой цепи и 3 CDR легкой цепи), имеющие 3, 2 или 1 отличие(чия) аминокислот, представляют собой варианты моноклонального антитела или CDR антигенсвязывающего фрагмента, которые получают способами созревания аффинности.
В некоторых воплощениях моноклональные антитела или антигенсвязывающие фрагменты связываются с IL-5 с аффинностью (KD) меньше, чем 10-8 М, меньше, чем 10-9 М, меньше, чем 10-10 М или меньше, чем 10-11 М.
В некоторых воплощениях моноклональное антитело или антигенсвязывающий фрагмент специфично связывается с человеческим IL-5, причем данное моноклональное антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(vii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 16-18, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 19-21; или
(viii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 22-24, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 25-27; или
(ix) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 28-30, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 31-33; или
(x) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 34-36, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 37-39; или
(xi) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 40-42, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 43-45; или
(xii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в SEQ ID NO: 34, 82 и 36, и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в SEQ ID NO: 37-39.
В некоторых воплощениях моноклональное антитело представляет собой рекомбинантное антитело.
В некоторых воплощениях моноклональное антитело выбрано из группы, состоящей из мышиного антитела, химерного антитела, рекомбинантного антитела гуманизированного антитела или их антигенсвязывающего фрагмента.
В некоторых воплощениях последовательности области FR (каркас) легкой и тяжелой цепи на вариабельной области легкой и тяжелой цепи гуманизированного антитела происходят, соответственно, из легкой и тяжелой цепи человеческой зародышевой линии или их мутировавших последовательностей.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, гуманизированное антитело содержит вариабельную область тяжелой цепи SEQ ID NO: 49, 57, 63, 69 или 75, или ее вариант; причем данный вариант имеет 1-10 аминокислотную(ные) мутацию(ции) на вариабельной области тяжелой цепи, как определено в SEQ ID NO: 49, 57, 63, 69 или 75.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, вариант имеет 1-10 аминокислотных обратных мутаций на FR области вариабельной области тяжелой цепи, как определено в SEQ ID NO: 49, 57, 63, 69 или 75; предпочтительно данная обратная мутация выбрана из группы, состоящей из S49T, V93T и K98S или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 49, или обратная мутация выбрана из группы, состоящей из S49T, V93T и K98T или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 57, или обратная мутация выбрана из группы, состоящей из R38K, M48I, R67K, V68A, M70L, R72V, T74K и L83F или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 63, или обратная мутация выбрана из группы, состоящей из F29I, R38K, V48I, R72A, T97F и N55V или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 69, или обратная мутация выбрана из группы, состоящей из R38K, M48I, R67K, V68A, R72A, T74K, M81L, L83F и D89E или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 75.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, гуманизированное антитело содержит вариабельную область тяжелой цепи SEQ ID NO: 50 или 51, или содержит вариабельную область тяжелой цепи SEQ ID NO: 58 или 59, или содержит вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 64, 65 и 66, или содержит вариабельную область тяжелой цепи SEQ ID NO: 70 или 71, или содержит вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 76-79.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, гуманизированное антитело содержит вариабельную область легкой цепи SEQ ID NO: 46, 54, 60, 67 или 72, или ее вариант; данный вариант имеет 1-10 замен аминокислот на вариабельной области легкой цепи, как определено в SEQ ID NO: 46, 54, 60, 67 или 72.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, вариант имеет 1-10 обратных мутаций аминокислот на области FR вариабельной области легкой цепи, как определено в SEQ ID NO: 46, 54, 60, 67 или 72; предпочтительно указанная обратная мутация выбрана из группы, состоящей из A43S, L47V, G66R, T69S, F71Y и Y87F или их комбинации на вариабельной области легкой цепи SEQ ID NO: 46; или обратная мутация выбрана из группы, состоящей из A43S, L47M, F71Y и Y87F или их комбинации на вариабельной области легкой цепи SEQ ID NO: 54; или обратная мутация выбрана из группы, состоящей из E1D, I2T, I57V, V84T и Y86F или их комбинации на вариабельной области легкой цепи SEQ ID NO: 60; или обратная мутация выбрана из группы, состоящей из M4L, A42S, L45P и L46W или их комбинации на вариабельной области легкой цепи SEQ ID NO: 67; или обратная мутация выбрана из группы, состоящей из A43S, I48V и F71Y или их комбинации на вариабельной области легкой цепи SEQ ID NO: 72.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, гуманизированное антитело содержит вариабельную область легкой цепи SEQ ID NO: 47 или 48; или содержит вариабельную область легкой цепи SEQ ID NO: 55 или 56; или содержит вариабельную область легкой цепи SEQ ID NO: 61 или 62; или содержит вариабельную область легкой цепи SEQ ID NO: 68; или содержит вариабельную область легкой цепи SEQ ID NO: 73 или 74.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, гуманизированное антитело содержит:
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 49-51, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 46-48; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 57-59, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 54-56; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 63-66, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 60-62; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 69-71, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 67-68; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 75-79, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 72-74.
В некоторых воплощениях моноклональное антитело или его антигенсвязывающий фрагмент, антитело представляет собой полноразмерное антитело, дополнительно содержит константную область человеческого антитела, где константная область тяжелой цепи предпочтительно представляет собой константную область тяжелой цепи человеческого антитела IgG1, IgG2, IgG3 и IgG4. Более предпочтительно данное полноразмерное антитело содержит константную область тяжелой цепи человеческого антитела, как определено в SEQ ID NO: 52, и константную область легкой цепи человеческого антитела, как определено в SEQ ID NO: 53.
В некоторых воплощениях антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, Fab', F(ab')2, одноцепочечного антитела (scFv), димеризованной области V (диатело), области V, стабилизированной дисульфидом (dsFv), и пептида, содержащего CDR.
Согласно настоящему раскрытию также предложено выделенное моноклональное антитело или его антигенсвязывающий фрагмент, которое конкурирует за связывание с человеческим IL-5 с моноклональным антителом или его антигенсвязывающим фрагментом, описанным выше.
Согласно настоящему раскрытию также предложена фармацевтическая композиция, содержащая терапевтически эффективное количество моноклонального антитела или его антигенсвязывающего фрагмента согласно настоящему раскрытию и один или более чем один фармацевтически приемлемый носитель, разбавитель, буфер или эксципиент. Количество моноклонального антитела или его антигенсвязывающего фрагмента, содержащееся в единичной дозе фармацевтической композиции, предпочтительно составляет от 0,1 до 2000 мг, более предпочтительно составляет от 1 до 1000 мг.
Согласно настоящему раскрытию также предложена выделенная молекула нуклеиновой кислоты, кодирующая моноклональное антитело или его антигенсвязывающий фрагмент согласно настоящему раскрытию.
Согласно настоящему раскрытию также предложен рекомбинантный вектор, содержащий молекулу нуклеиновой кислоты, описанную выше.
Согласно настоящему раскрытию также предложена клетка-хозяин, трансформированная рекомбинантным вектором согласно настоящему раскрытию, причем данная клетка-хозяин выбрана из группы, состоящей из прокариотических клеток и эукариотических клеток, предпочтительно эукариотических клеток, более предпочтительно клеток млекопитающих.
Согласно настоящему раскрытию также предложен способ получения моноклонального антитела или его антигенсвязывающего фрагмента согласно настоящему раскрытию, включающий культивирование описанной выше клетки-хозяина в культуре с образованием и накоплением вышеописанного моноклонального антитела или его антигенсвязывающего фрагмента, и выделение данного моноклонального антитела или его антигенсвязывающего фрагмента из культуры.
Согласно настоящему раскрытию также предложен способ выявления или определения человеческого IL-5, причем данный способ включает применение вышеописаного моноклонального антитела или его антигенсвязывающих фрагментов.
Согласно настоящему раскрытию также предложено средство для выявления или определения человеческого IL-5, которое содержит моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из описанных выше.
Согласно настоящему раскрытию также предложено диагностическое средство для заболевания, ассоциированного с человеческим IL-5, причем данное диагностическое средство содержит вышеописанное моноклональное антитело или его антигенсвязывающий фрагмент.
Согласно настоящему раскрытию также предложен способ осуществления диагностики заболевания, ассоциированного с человеческим IL-5, включающий выявление или определение человеческого IL-5 или клеток, позитивных в отношении IL-5, с использованием вышеописанного моноклонального антитела или его антигенсвязывающего фрагмента.
Согласно настоящему раскрытию также предложено применение вышеописанного моноклонального антитела или его антигенсвязывающего фрагмента для получения диагностического средства для заболевания, ассоциированного с человеческим IL-5.
Согласно настоящему раскрытию также предложено лекарственное средство для лечения заболевания, ассоциированного с человеческим IL-5, содержащее вышеописанное моноклональное антитело или его антигенсвязывающий фрагмент, или содержащее вышеописанную фармацевтическую композицию, или содержащее вышеописанную молекулу нуклеиновой кислоты.
Согласно настоящему раскрытию также предложен способ лечения заболевания, ассоциированного с человеческим IL-5, включающий введение субъекту фармацевтически эффективного количества вышеописанного моноклонального антитела или его антигенсвязывающего фрагмента, или содержащей их фармацевтической композиции, или вышеописанной молекулы нуклеиновой кислоты для предупреждения или лечения заболевания, ассоциированного с человеческим IL-5.
Согласно настоящему раскрытию также предложено применение вышеописанного моноклонального антитела или его антигенсвязывающего фрагмента, или содержащей их фармацевтической композиции, или вышеописанной молекулы нуклеиновой кислоты для получения терапевтического средства для заболевания, ассоциированного с человеческим IL-5.
Вышеописанное заболевание или состояние предпочтительно выбрано из группы, состоящей из астмы, злокачественного приступа астмы, хронической пневмонии, аллергического ринита, аллергического бронхолегочного аспергиллеза, эозинофилии, синдрома Чарга-Стросса, атопического дерматита, онхоциркозного дерматита, эпизодического ангиоотека, синдрома эозинофильной миалгии, эозинофильного гастроэнтерита, инфекции гельминтами, болезни Ходжкина, полипов носа, синдрома Леффлера, уртикарии, эозинофильного гиперпластического бронхита, узелкового артериита, синусита, эозинофильного эзофагита, аллергического эозинофильного эзофагита, аллергического конъюнктивита, онхоциркозного дерматита, эндометриоза и зависимого от стероидов эозинофильного бронхита.
Моноклональные антитела против IL-5 или антигенсвязывающие фрагменты по настоящему раскрытию имеют высокую специфичность и высокую аффинность с IL-5. Гуманизированные антитела имеют значительно пониженную иммуногенность и полностью сохраняют специфичность от мышиного антитела, и демонстрируют высокую аффинность и превосходные активности in vitro и in vivo.
Моноклональные антитела или антигенсвязывающие фрагменты против IL-5 по настоящему раскрытию имеют хорошую селективность в отношении просто специфичного распознавания IL-5.
Моноклональные антитела или антигенсвязывающие фрагменты против IL-5 по настоящему раскрытию имеют хорошие характеристики метаболической динамики у крыс, демонстрируют длительный период полувыведения и высокую биодоступность.
Молекулы гуманизированного антитела против IL-5 по настоящему раскрытию имеют хорошую долговременную стабильность, не имеют очевидной ненормальной химической модификации, не имеют очевидной агрегации при высокой концентрации и имеют высокую чистоту и термостабильность.
Помимо уменьшения пролиферации эозинофилов, моноклональные антитела или антигенсвязывающие фрагменты против IL-5 по настоящему раскрытию имеют хорошие свойства в улучшении функции легкого.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: антитела против IL-5 блокируют связывание IL-5 с рецептором IL-5 в эксперименте FACS (флуоресцентная сортировка клеток);
Фиг. 2: выявление специфичности связывания антител против IL-5 с цитокином Th2;
Фиг. 3: антитела против IL-5 увеличивают респираторное периодическое значение (Penh). G1: нормальная контрольная группа (PBS (фосфатно-солевой буферный раствор)); G2: модельная группа (IgG); G3: группа с 10 mpk (миллиграммов на килограмм) антитела h1705-008; G4: группа с 2 mpk антитела h1705-008; G5: группа с 10 mpk антитела h1706-009; G6: группа с 2 mpk антитела h1706-009; G7: группа с 10 mpk Hu39D10; *р меньше 0,05, ** меньше 0,01 (при сравнении с группой G2 посредством ANOVA (дисперсионный анализ)/поправки Бонферрони);
Фиг. 4А: уровень эозинофилов BALF (бронхоальвеолярный лаваж) в легком астматических мышей; Фиг. 4В: Баллы толщины слизистой трахеи астматических мышей. G1: нормальная контрольная группа; G2: модельная группа; G3: группа с 10 mpk антитела h1705-008; G4: группа с 2 mpk антитела h1705-008; G5: группа с 10 mpk антитела h1706-009; G6: группа с 2 mpk антитела h1706-009; G7: группа с 10 mpk Hu39D10; Фиг. 4С: процентная доля эозинофилов BALF в легком астматических мышей;
На Фиг. 5А и Фиг. 5Б показана способность mAb (моноклональное антитело) против IL5 уменьшать уровень эозинофилов в BALF.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
1. Терминология
Для того чтобы легче понять настоящее раскрытие, некоторые технические и научные термины конкретно определяются ниже. Если в данном документе прямо не определено иначе, все другие технические и научные термины, использованные в данном документе, имеют значение, обычно понятное обычному специалисту в области, к которой принадлежит данное раскрытие.
Трехбуквенные коды и однобуквенные коды аминокислот, используемые в настоящем раскрытии, являются такими, как описано в J. biol. chem, 243, р3558 (1968).
Термин «антитело» в том виде, в котором он используется в данном документе, относится к иммуноглобулину - структуре из четырех пептидных цепей, соединенных друг с другом дисульфид ной связью между двумя идентичными тяжелыми цепями и двумя идентичными легкими цепями. Разные константные области тяжелой цепи иммуноглобулина демонстрируют разные аминокислотные составы и порядки по рангу, следовательно, представляют разную антигенность. Соответственно, иммуноглобулины могут быть разделены на пять типов, или они именуются изотипами иммуноглобулинов, а именно: IgM, IgD, IgG, IgA и IgE, с тяжелой цепью μ, δ, γ, α и ε соответственно. Согласно аминокислотному составу его шарнирной области и числу, и положению дисульфидных связей тяжелой цепи, тот же самый тип Ig может дополнительно подразделяться на разные подтипы, например, IgG может подразделяться на IgG1, IgG2, IgG3 и IgG4. Легкая цепь может быть подразделена на цепь κ и λ на основе разной константной области. Каждый из пяти типов Ig может иметь цепь κ или λ.
В настоящем раскрытии легкая цепь антитела, упомянутая в настоящем раскрытии, дополнительно содержит константную область легкой цепи, которая содержит человеческую или мышиную цепь κ, λ или ее вариант.
В настоящем раскрытии тяжелая цепь антитела, упомянутая в настоящем раскрытии, дополнительно содержит константную область тяжелой цепи, которую содержит человеческий или мышиный IgG1, IgG2, IgG3, IgG4 или его вариант.
Примерно 110-аминокислотные последовательности, смежные с N-концом тяжелой и легкой цепей антитела, являются высоковариабельными, известными как вариабельная область (область Fv); остальные аминокислотные последовательности, близкие к С-концу, являются относительно стабильными, известными как константная область. Вариабельная область включает три гипервариабельные области (HVR) и четыре относительно консервативные каркасные области (FR). Три гипервариабельные области, которые определяют специфичность антитела, также известны как области, определяющие комплементарность (CDR). Каждая вариабельная область легкой цепи (LCVR) и каждая вариабельная область тяжелой цепи (HCVR) состоит из трех областей CDR и четырех областей FR с последовательным порядком от аминоконца к карбоксильному концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Три области CDR легкой цепи относятся к LCDR1, LCDR2 и LCDR3, и три области CDR тяжелой цепи относятся к HCDR1, HCDR2 и HCDR3. Число и положение аминокислотных остатков CDR в областях LCVR и HCVR антитела или его антигенсвязывающих фрагментов в данном документе согласуются с известными критериями нумерации Kabat (LCDR1-3, HCDR1-3).
Антитело по настоящему раскрытию включает мышиное антитело, химерное антитело и гуманизированное антитело, предпочтительно оно представляет собой гуманизированное антитело.
Термин «мышиное антитело» в настоящем раскрытии относится к моноклональному антителу против человеческого IL-5, полученному согласно знаниям и квалификации в данной области. Во время получения анализируемому субъекту может быть инъецирован антиген IL-5, и затем выделяется гибридома, экспрессирующая антитело, которое обладает желательной последовательностью или функциональными характерстиками. В предпочтительном воплощении настоящего раскрытия мышиное антитело против IL-5 или его антигенсвязывающий фрагмент дополнительно содержит константную область легкой цепи мышиной цепи κ, λ или ее вариант, или дополнительно содержит константную область тяжелой цепи мышиного IgG1, IgG2, IgG3 или IgG4, или ее вариант.
«Химерное антитело» представляет собой антитело, полученное посредством слияния вариабельной области мышиного антитела с константной областью человеческого антитела, и данное химерное антитело может облегчать иммунный ответ, индуцированный мышиным антителом. Для получения химерного антитела может быть создана гибридома, секретирующая специфичное мышиное моноклональное антитело, и ген вариабельной области клонируется из мышиной гибридомы. Затем можно клонировать желательный ген константной области человеческого антитела и соединить с мышиным геном вариабельной области с образованием химерного гена, который может быть затем вставлен в экспрессионный вектор. Наконец, молекула химерного антитела будет экспрессироваться в эукариотической или прокариотической системе. В предпочтительном воплощении настоящего раскрытия легкая цепь химерного антитела против IL-5 дополнительно содержит константную область легкой цепи, происходящую из человеческой цепи κ, λ или ее варианта. Тяжелая цепь химерного антитела против IL-5 дополнительно содержит константную область тяжелой цепи, происходящую из человеческого IgG1, IgG2, IgG3, IgG4 или его варианта, предпочтительно содержит константную область тяжелой цепи, происходящую из человеческого IgG1, IgG2 или IgG4, или содержит вариант константной области тяжелой цепи человеческого IgG1, IgG2 или IgG4 с аминокислотной(ными) мутацией(ями), такой(кими) как мутация(ции) YTE или обратная(ные) мутация(ции).
Термин «гуманизированное антитело» относится к антителу, полученному прививкой мышиных последовательностей CDR в каркас вариабельной области человеческого антитела, т.е. антитела, продуцированного в разных типах последовательностей каркаса антитела человеческой зародышевой линии. Гуманизированное антитело может преодолевать гетерологичные ответы, индуцированные большим числом компонентов мышиного белка, которые несет химерное антитело. Такие каркасные последовательности могут быть получены из общедоступной базы данных ДНК, охватывающей последовательности генов антител зародышевой линии, и опубликованных ссылок. Например, последовательности ДНК зародышевой линии генов вариабельной области человеческой тяжелой и легкой цепи могут быть найдены в базе данных последовательностей человеческой зародышевой линии «VBase» (доступна в сети: www.mrccpe.com.ac.uk/vbase), а также в Kabat, ЕА, et al. 1991 Sequences of Proteins of Immunological Interest, 5th Ed. Для того чтобы избежать уменьшения активности, вызванного пониженной иммуногенностью, каркасные последовательности в вариабельной области человеческого антитела могут быть подвергнуты минимальным реверсивным мутациям или обратным мутациям для подержания активности. Гуманизированное антитело по настоящему раскрытию также включает гуманизированное антитело, на котором созревание аффинности CDR осуществляется посредством фагового дисплея. В предпочтительном воплощении настоящего раскрытия последовательность CDR гуманизированного антитела против IL-5 выбрана из группы, состоящей из SEQ ID NO: 16-21, 22-27, 28-33, 34-39 и 40-45. Сконструирован и выбран каркас вариабельной области человеческого антитела, в котором последовательность области FR на вариабельной области тяжелой цепи антитела происходит из последовательности тяжелой цепи человеческой зародышевой линии и последовательности легкой цепи человеческой зародышевой линии. Для того чтобы избежать уменьшения активности, вызванного пониженной иммуногенностью, вариабельная область человеческого антитела может быть подвергнута минимальным реверсивным мутациям (обратным мутациям, то есть, аминокислотные остатки области FR, происходящей из человеческого антитела, заменяются аминокислотными остатками, соответствующими исходному антителу) для поддержания активности.
Прививка CDR может приводить к снижению аффинности образующегося антитела или его антигенсвязывающего фрагмента против IL-5 к антигену из-за изменения остатков каркаса, контактирующих с антигеном. Такие взаимодействия могут быть результатом высокосоматических мутаций. Следовательно, все еще может быть необходимым перенос донорных аминокислот каркаса в каркас гуманизированного антитела. Аминокислотные остатки, происходящие из антитела против IL-5, не являющегося человеческим, или его антигенсвязывающего фрагмента, которые участвуют в связывании антигена, могут быть идентифицированы посредтвом проверки последовательности и структуры вариабельной области мышиного моноклонального антитела. Аминокислотные остатки в каркасе донорного CDR, которые отличаются от аминокислотных остатков в зародышевых линиях, могут считаться родственными. Если не возможно определение самой близкородственной зародышевой линии, данную последовательность можно сравнивать с общей последовательностью, которую имеют подтипы, или с общей последовательностью мышиных последовательностей, имеющих высокую процентную долю сходства. Полагают то, что редкие остатки каркаса являются результатом высокой мутации в соматических клетках, которая играет важную роль в связывании.
Термин «антигенсвязывающий фрагмент» или «функциональный фрагмент» в том виде, в котором он используется в данном документе, относится к одному или более чем одному фрагменту антитела, сохраняющему способность к связыванию с антигеном (например, с IL-5). Было показано то, что фрагменты полноразмерного антитела можно использовать для достижения функции связывания с антигеном. Примеры связывающих фрагментов в термине «антигенсвязывающий фрагмент» включают: (i) фрагмент Fab - одновалентный фрагмент, состоящий из домена VL, VH, CL и СН1; (ii) фрагмент F(ab')2 - двухвалентный фрагмент, содержащий два фрагмента Fab, связанных дисульфидной связью в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и СН1; (iv) фрагмент Fv, состоящий из доменов VH и VL неполного антитела; (v) один домен или фрагмент dAb (Ward et al. (1989) Nature 341:544-546), состоящий из домена VH; и (vi) отдельную область, определяющую комплементарность (CDR), и (vii) комбинацию двух или более чем двух отдельных CDR, возможно связанных синтетическим линкером. Кроме того, хотя домен VL и домен VH фрагмента Fv и кодируются двумя отдельными генами, они могут быть связаны синтетическим линкером посредством применения способов генной инженерии, генерируя, посредством этого, одну цепь белка одноваленной молекулы, образованной спариванием домена VL и VH (называется одноцепочечный Fv (scFv); см., например, Bird et al. (1988); Science 242: 423-426 и Huston et al (1988) Proc. Natl. Acad. Sci USA 85:5879-5883). Также подразумевается то, что данное одноцепочечное антитело также включается в термин «антигенсвязывающий фрагмент» антитела. Такие фрагменты антитела получают с использованием традиционных методик, известных в данной области, и подвергают скринингу на функциональные фрагменты с использованием того же самого способа, что и для интактного антитела. Антигенсвязывающие части могут быть получены методикой генной инженерии или ферментативным, или химическим расщеплением интактного иммуноглобулина. Антитела могут находиться в виде разных изотипов, например, антитело IgG (например, подтип IgG1, IgG2, IgG3 или IgG4), IgA1, IgA2, IgD, IgE или IgM.
Антигенсвязывающий фрагмент в настоящем раскрытии включает Fab, F(ab')2, Fab', одноцепочечное антитело (scFv), димеризованную область V (диатело), область V, стабилизированную дисульфидом (dsFv), и пептид, содержащий CDR.
Fab представляет собой фрагмент антитела, полученный обработкой молекулы антитела IgG папаином (который расщепляет аминокисотный остаток в положении 224 цепи Н). Фрагмент Fab, в котором примерно половина N-концевой стороны цепи Н и вся цепь L связываются друг с другом дисульфидной связью, имеет молекулярную массу примерно 50000 и имеет антигенсвязывающую активность.
Fab по настоящему раскрытию может быть получен обработкой папаином моноклонального антитела по настоящему изобретению, которое специфично распознает человеческий IL-5 и связывается с его аминокислотной последоваельностью внеклеточной области или трехмерной структурой. Также Fab может быть получен вставкой ДНК, кодирующей Fab антитела, в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и введением данного вектора в прокариота или эукариота для экспрессии Fab.
F(ab')2 представляет собой фрагмент антитела, имеющий молекулярную массу примерно 100000, имеющий антигенсвязывающую активность и содержащий две области Fab, которые связываются в положении шарнира, F(ab')2 получают расщеплением пепсином расположенной ниже части с двумя дисульфидными связями в шарнирной области IgG.
F(ab')2 по настоящему раскрытию может быть получен обработкой пепсином моноклонального антитела по настоящему изобретению, которое специфично распознает человеческий IL-5 и связывается с его аминокислотной последоваельностью внеклеточной области или трехмерной структурой. Также F(ab')2 может быть получен связыванием Fab', описанного ниже, посредством тиоэфирной связи или дисульфидной связи.
Fab' представляет собой фрагмент антитела, имеющий молекулярную массу примерно 50000 и имеющий антигенсвязывающую активность. Fab' получают расщеплением дисульфидной связи в шарнирной области вышеописанного F(ab')2. Fab' по настоящему раскрытию может быть получен обработкой восстановителем, таким как дитиотрейтол, F(ab')2 по настоящему изобретению, который специфично распознает IL-5 и связывается с его аминокислотной последовательностью внеклеточной области или трехмерной структурой.
Также Fab' может быть получен вставкой ДНК, кодирующей фрагмент Fab' антитела, в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и введением данного вектора в прокариота или эукариота для экспрессии Fab'.
Термин «одноцепочечное антитело», «одноцепочечный Fv» или «scFv» относится к молекуле, содержащей вариабельный домен тяжелой цепи антитела (или область; VH) и вариабельный домен легкой цепи антитела (или область; VL), соединенные линкером. Такие молекулы scFv имеют общую структуру NH2-VL-линкер-VH-COOH или NH2-VH-линкер-VL-СООН. Подходящий линкер в предшествующем уровне техники состоит из повторяющейся аминокислотной последовательности GGGGS или ее варианта, например, с использованием варианта с 1-4 повторами (Holliger et al. (1993), Proc. Natl. Acad. Sci. USA 90:6444-6448). Другие линкеры, которые можно использовать для настоящего раскрытия, описываются Alfthan et al. (1995), Protein Eng. 8:725-731, Choi et al. (2001), Eur. J. Immunol. 31:94-106, Hu et al. (1996), Cancer Res. 56:3055-3061, Kipriyanov et al. (1999), J. Mol. Biol. 293:41-56 и Roovers et al. (2001), Cancer Immunol.
scFv по настоящему раскрытию может быть получен посредством следующих стадий: получение кДНК, кодирующей VH и VL моноклонального антитела по настоящему раскрытию, которое специфично распознает человеческий IL-5 и связывается с его аминокислотной последоваельностью внеклеточной области или трехмерной структурой, построение ДНК, кодирующей scFv, вставка данной ДНК в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и затем вставка данного экспрессионного вектора в прокариота или эукариота для экспрессии scFv.
Диатело представляет собой фрагмент антитела, в котором scFv димеризуется, и представляет собой фрагмент антитела, имеющий двухвалентную антигенсвязывающую активность. При двухвалентной активности связывания два антигена могут быть одинаковыми или разными.
Диатело по настоящему раскрытию может быть получено посредством следующих стадий: получение кДНК, кодирующих VH и VL моноклонального антитела по настоящему раскрытию, которое специфично распознает человеческий IL-5 и связывается с его аминокислотной последоваельностью внеклеточной области или трехмерной структурой, построение ДНК, кодирующей scFv, таким образом, что длина линкерного пептида составляет 8 или меньше аминокислотных остатков, вставка данной ДНК в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и затем вставка данного экспрессионного вектора в прокариота или эукариота для экспрессии диатела.
dsFv получают заменой одного аминокислотного остатка в каждой VH и VL остатком цистеина и затем соединением данных полипептидов с заменой посредством дисульфидной связи между двумя остатками цистеина. Аминокислотный остаток, подлежащий замене остатком цистеина, может быть выбран на основе прогнозирования трехмерной структуры антитела согласно известным способам (Protein Engineering, 7, 697 (1994)).
dsFv по настоящему раскрытию может быть получен посредством следующих стадий: получение кДНК, кодирующих VH и VL моноклонального антитела по настоящему раскрытию, которое специфично распознает человеческий IL-5 и связывается с его аминокислотной последовательностью внеклеточной области или трехмерной структурой, построение ДНК, кодирующей dsFv, вставка данной ДНК в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и затем вставка данного экспрессионного вектора в прокариота или эукариота для экспрессии dsFv.
Пептид, содержащий CDR, построен одной или более чем одной областью CDR VH и VL. Пептиды, содержащие несколько CDR, могут соединяться непосредственно или через подходящий пептидный линкер.
Пептид, содержащий CDR по настоящему раскрытию, может быть получен посредством следующих стадий: построение ДНК, кодирующей CDR VH и VL моноклонального антитела по настоящему раскрытию, которые специфично распознают человеческий IL-5 и связываются с его аминокислотной последовательностью внеклеточной области или трехмерной структурой, вставка данной ДНК в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и затем введение данного экспрессионного вектора в прокариота или эукариота для экспрессии данного пептида. Пептид, содержащий CDR, также может быть получен способом химического синтеза, такого как способ с Fmoc или способ с tBoc.
Термин «каркас антитела» в том виде, в котором он используется в данном документе, относится к части вариабельного домена: либо VL, либо VH, которая служит в качестве каркаса для антигенсвязывающих петель (CDR) данного вариабельного домена. По существу, он представляет собой вариабельный домен без CDR.
Термин «отличие аминокислот» относится к отличиям в одном или более чем одном положении аминокислоты по длине полипептидного фрагмента между полипептидом и его вариантом, где данный вариант может быть получен заменой, вставкой или делецией одной или более чем одной аминокислоты на полипептиде.
Термин «эпитоп» или «антигенная детерминанта» относится к сайту на антигене, с которым специфично связывается иммуноглобулин или антитело (например, к специфическому сайту на молекуле IL-5). Эпитопы типично включают по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 последовательных или непоследовательных аминокислот в уникальной третичной конформации. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, ed. G.E. Morris (1996).
Термин «специфично связываются с», «селективно связываются с», «селективно связывается с» или «специфично связывается с» относится к связыванию антитела с заданным эпитопом на антигене. Типично антитело связывается с аффинностью (KD) меньше, чем примерно 10-8 М, например, меньше, чем примерно 10-9 М, 10-10 М или 10-11 М, или даже меньше.
Термин «KD» относится к равновесной константе диссоциации в отношении конкретного взаимодействия антитело-антиген. Типично антитело по настоящему раскрытию связывается с IL-5 с равновесной константой диссоциации (KD) меньше, чем примерно 10-7 М, как, например, меньше, чем примерно 10-8 М, 10-9 М или 10-10 М, или даже меньше, например, при определении с использованием методик поверхностного плазмонного резонанса (SPR) на приборе BIACORE.
При использовании термина «конкуренция» в контексте антигенсвязывающих белков (например, нейтрализующие антигенсвязывающие белки или нейтрализующие антитела), которые конкурируют за тот же самый эпитоп, он означает то, что конкуренция происходит среди антигенсвязывающих белков, что определяется посредством следующих анализов: антигенсвязывающий белок, подлежащий анализу (например, антитело или его иммунологически функциональный фрагмент), предотвращает или ингибирует (например, уменьшает) специфичное связывание между эталонным антигенсвязывающим белком (например, лигандом или эталонным антителом) и общим антигеном (например, антиген IL-5 или его фрагмент). Доступны многие типы конкурентных анализов связывания для определения того, конкурирует ли антигенсвязывающий белок с другим антигенсвязывающим белком. Данными анализами являются, например, твердофазный прямой или непрямой радиоиммуноанализ (RIA), твердофазный прямой или непрямой ферментативный иммуноанализ (EIA), конкурентный сэндвич-анализ (см., например, Stahli et al, 1983, Methods in Enzymology 9: 242-253); твердофазный прямой EIA биотин-авидин (см., например, Kirkland et al, 1986, J. Immunol. 137: 3614-3619), твердофазный прямой анализ мечения, твердофазный прямой сэндвич-анализ мечения (см., например, Harlow and Lane, 1988, Antibodies, A Laboratory Manual, Cold Spring Harbor Press); твердофазный прямой RIA мечения с меткой I-125 (см., например, Morel et al, 1988, Molec. Immunol. 25: 7-15); твердофазный прямой EIA биотин-авидин (см., например, Cheung, et al, 1990, Virology 176: 546-552); и прямой RIA мечения (Moldenhauer et al, 1990, Scand. J. Immunol. 32: 77-82). Типично данный анализ включает применение очищенного антигена (либо на твердой поверхности, либо на поверхности клетки), способного связываться как с немеченым антигенсвязывающим белком, подлежащим анализу, так и с меченым эталонным антигенсвязывающим белком. Конкурентное ингибирование определяется измерением количества метки, связавшейся с твердой поверхностью или с клеткой, в присутствии антигенсвязывающего белка, подлежащего анализу. Обычно антигенсвязывающий белок, подлежащий анализу, присутствует в избытке. Антигенсвязывающие белки, идентифицированные конкурентным анализом (конкурирующие с антигенсвязывающим белком) включают: антигенсвязывающие белки, которые связываются с таким же эпитопом, что и эталонный антигенсвязывающий белок; и антигенсвязывающие белки, которые связываются с эпитопом, который является достаточно близким к эпитопу, с которым связывается эталонный антигенсвязывающий белок, где два эпитопа пространственно мешают друг другу с затруднением связывания. Дополнительные подробности относительно способов опеделения конкурентного связывания предоставляются в данном документе в Примерах. Типично, когда конкурирующий антигенсвязывающий белок присутствует в избытке, он будет ингибировать (например, уменьшать) специфичное связывание между эталонным антигенсвязывающим белком и общим антигеном по меньшей мере на 40-45%, 45-50%, 50-55%, 55-60%, 60-65%, 65-70%, 70-75% или 75%, или даже больше. В некоторых случаях связывание ингибируется по меньшей мере на 80-85%, 85-90%, 90-95%, 95-97% или 97%, или даже больше.
Термин «молекула нуклеиновой кислоты» в том виде, в котором он используется в данном документе, относится к молекулам ДНК и молекулам РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно представляет собой двухцепочечную ДНК. Нуклеиновая кислота «связывается функциональным образом» при ее помещении в функциональную связь с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер связывается функциональным образом с кодирующей последовательностью, если он воздействует на транскрипцию данной последовательности.
Термин «вектор» относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. В одном воплощении вектор представляет собой «плазмиду», что относится к кольцевой двухцепочечной петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. В другом воплощении вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Векторы, раскрытые в данном документе, способны к саморепликации в клетке-хозяине, в которую они вводятся (например, бактериальные векторы, имеющие бактериальный репликатор, и эписомные векторы млекопитающих), или могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и, посредством этого, реплицируются наряду с геномом хозяина (например, неэписомные векторы млекопитающих).
Способы получения и очистки антител и антигенсвязыващих фрагментов хорошо известны в данной области и могут быть найдены, например, в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, главы 5-8 и 15. Например, мыши могут быть иммунизированы человеческим IL-5 или его фрагментами, и образующиеся антитела могут быть затем ренатурированы, очищены и секвенированы на аминокислотные последовательности с использованием традиционных способов, хорошо известных в данной области. Антигенсвязывающие фрагменты также могут быть получены традиционными способами. Антитела или антигенсвязывающие фрагменты по настоящему раскрытию конструируются так, чтобы содержать одну или более чем одну человеческую область FR на CDR, происходящих из антитела, не являющегося человеческим. Человеческие последовательности FR зародышевой линии могут быть получены посредством выравнивания с использованием базы данных генов вариабельных областей человеческих антител зародышевой линии и программы МОЕ от ImMunoGeneTics (IMGT) посредством их веб-сайта http://imgt.cines.fr, или из The Immunoglobulin Facts Book, 2001, ISBN 012441351.
Термин «клетка-хозяин» относится к клетке, в которую был введен экспрессионный вектор. Клетки-хозяева могут включать микробные (например, бактериальные), растительные или животные клетки. Бактерии, которые являются чувствительными к трансформации, включают членов энтеробактерий, таких как штаммы Escherichia coli или Salmonella; бациллы, такие как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Подходящие микроорганизмы включают Saccharomyces cerevisiae и Pichia pastoris. Подходящие линии клеток-хозяев животных включают СНО (линия клеток яичника китайского хомяка), кетки HEK (в качестве неограничивающих примеров, клетки HEK293E) и клетки NS0, но не ограничиваются ими.
Сконструированные антитела или антигенсвязывающие фрагменты по настоящему раскрытию могут быть получены и очищены с использованием известных способов. Например, последовательность кДНК, кодирующая тяжелую цепь и легкую цепь, может быть клонирована и сконструирована в экспрессионном векторе GS. Экспрессионный вектор для рекомбинантного иммуноглобулина может быть затем стабильно трансфицирован в клетки СНО. В качестве более рекомендованного способа, хорошо известного в данной области, системы экспрессии на основе клеток млекопитающих будут приводить к гликозилированию антитела, типично на высококонсервативных N-концевых сайтах области Fc. Стабильные клоны могут быть подтверждены на экспрессию антитела, специфично связывающегся с человеческим IL-5. Позитивные клоны могут быть размножены в бессывороточной культуральной среде в биореакторах для продукции антител. Культуральная среда, в которую секретировалось антитело, может быть очищена традиционными методиками. Например, очистка может осуществляться на колонке сефарозы FF с белком А или G, которая была уравновешена подведенным буфером. Колонку промывают для удаления неспецифично связывающихся компонентов, и затем связавшееся антитело элюируется градиентом рН, и фракции антитела выявляются SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), и затем собираются. Данные антитела можно фильтровать и концентрировать с использованием обычных методик. Подходящие смеси и полимеры можно удалять обычными методиками, такими как гель-фильтрация или обмен ионов. Образующийся продукт затем немедленно замораживается, например, при -70°С, или он может быть лиофилизирован.
Термин «введение», «осуществление введения» или «обработка» в том виде, в котором он применяется к животному, человеку, субъекту, клетке, ткани, органу или биологической жидкости, относится к приведению в контакт экзогенного фармацевтического, терапевтического, диагностического средства или композиции с животным, человеком, субъектом, клеткой, тканью, органом или биологической жидкостью. Термин «введение», «осуществление введения» или «обработка» может относиться, например, к терапевтическим, фармакокинетическим, диагностическим, исследовательским и экспериментальным способам. Обработка клетки охватывает приведение в контакт реактива с клеткой, а также приведение в контакт реактива с жидкостью, где данная жидкость находится в контакте с клеткой. Термин «введение», «осуществление введения» или «обработка» также означает обработки клетки in vitro и ex vivo реактивом, диагностической, связывающей композицией или другой клеткой. Термин «обработка» в том виде, в котором он применяется к человеку, ветеринарному или исследовательскому субъекту, относится к терапевтическому лечению, профилактическим или предупредительным мерам, к исследовательским и диагностическим приложениям.
«Лечить» означает введение терапевтического средства, такого как композиция, содержащая любые антитела или их фрагменты по настоящему раскрытию, внутренне или наружно пациенту, имеющему один или более чем один симптом заболевания, для которого данное средство имеет известную терапевтическую активность. Типично данное средство вводится в эффективном количестве для облегчения одного или более чем одного симптома заболевания у пациента или популяции, подлежащей лечению, посредством индуцирования регрессии или ингибирования прогрессирования такого(ких) симптома(мов) на любой клинически измеримый уровень. Количество терапевтического средства, которое является эффективным для облегчения любого конкретного симптома заболевания (также именуемое «терапевтически эффективное количество»), может варьировать согласно таким факторам, как состояние заболевания, возраст, масса пациента и способность данного лекарственного средства вызывать желательный ответ у пациента. Был ли облегчен симптом заболеания может оцениваться посредством любого клинического измерения, типично используемого лечащими врачами или другими квалифицированными поставщиками медицинских услуг, для оценки тяжести или статуса прогрессирования данного симптома. В то время как воплощение настоящего раскрытия (например, способ лечения или производственное изделие) может не быть эффективным в облегчении целевого(вых) симптома(мов) заболевания у каждого пациента, оно должно облегчать целевой(вые) симптом(мы) заболевания у статистически значимого числа пациентов при определении любым статистическим критерием, известным в данной области, таким как t-критерий Стьюдента, критерий хи-квадрат, U-критерий согласно Манну и Уитни, критерий Крускала-Уоллиса (Н-критерий), критерий Джонкхиера-Терпстра и критерий Вилкоксона.
Термин «консервативная модификация» или «консервативная замена» относится к заменам аминокслот в белке другими аминокислотами, имеющими аналогичные характеристики (например, заряд, размер боковой цепи, гидрофобность/гидрофильность, конформация остова и жесткость, и т.д.), таким образом, что можно часто делать изменения без изменения биологической активности белка. Специалистам в данной области известно то, что, в общем, замена одной аминокислоты в несущественных областях полипептида по существу не изменяет биологическую активность (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.)). Кроме того, замены структурно или функционально аналогичными аминокислотами с меньшей вероятностью нарушат биологическую активность.
«Эффективное количество» охватывает достаточное количество для уменьшения интенсивности или предупреждения симптома или признака медицинского состояния. Эффективное количество также означает достаточное количество для обеспечения или облегчения постановки диагноза. Эффективное количество для конкретного пациента или ветеринарного субъекта может варьировать, в зависимости от таких факторов, как состояние, которое лечат, общее состояние здоровья пациента, путь и доза введения и тяжесть побочных эффектов. Эффективное количество может представлять собой максимальную дозу или протокол дозирования, в котором избегаются значительные побочные эффекты или токсические эффекты.
Термин «экзогенный» относится к веществам, которые продуцируются вне организма, клетки или человеческого тела, в зависимости от контекста. Термин «эндогенный» относится к веществам, которые продуцируются в пределах клетки, организма, или человеческого тела, в зависимости от контекста.
Термин «гомология» относится к сходству последовательностей между двумя полинуклеотидными последовательностями или между двумя полипептидными последовательностями. Когда положение в обеих из двух сравниваемых последовательностях занято одинаковой мономерной субъединицей основания или аминокислоты, например, если положение в каждой из двух молекул ДНК занято аденином, тогда данные молекулы являются гомологичными в данном положении. Процент гомологии между двумя последовательностями является функцией числа совпадающих или гомологичных положений, которые делят две последовательности, поделенного на число положений, подлежащих сравнению, и затем умноженного на 100. Например, если 6 из 10 положений в двух последовательностях являются совпадающими или гомологичными при оптимальном выравнивании данных последовательностей, тогда две данные последовательности имеют 60%-ную гомологию; если 95 из 100 положений в двух последовательностях являются совпадающими или гомологичными, тогда две данные последовательности имеют 95%-ную гомологию. Обычно сравнение осуществляется при выравнивании двух последовательностей с получением максимального процента гомологии.
Выражения «клетка», «линия клеток» и «культура клеток» в том виде, в котором они используются в данном документе, используются взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают клетки первичного субъекта и полученные из них культуры, независимо от числа пассажей. Также следует понимать то, что все потомство может не быть точно идентичным по содержанию ДНК из-за преднамеренных или случайных мутаций. Включается мутантное потомство, которое имеет такую же функцию или биологическую активность, на которую проводился скрининг в исходно трансформированных клетках. Когда подразумеваются отличные обозначения, это будет отчетливо понятно из контекста.
Термин «полимеразная цепная реакция» или «ПЦР» в том виде, в котором он используется в данном документе, относится к процедуре или методике, в которой крошечные количества специфической части нуклеиновой кислоты, РНК и/или ДНК амплифицируются, как описано, например, в патенте США №4683195. В общем, должна быть доступна информация по последовательности около концов интересующей области или за пределами интересующей области, таким образом, что могут быть сконструированы олигонуклеотидные праймеры; данные праймеры будут идентичными или аналогичными по последовательности противоположным нитям матрицы, подлежащей амплификации. 5'-концевые нуклеотиды двух праймеров согласуются с концами вещества, подлежащего амплификации. ПЦР может использоваться для амплификации специфических последовательностей РНК, специфических последовательностей ДНК из общей геномной ДНК и кДНК, транскрибированной из общей клеточной РНК, последовательностей бактериофага или плазмиды и т.д. См., в общем, Mullis et al. (1987) Cold Spring Harbor Symp. Ouant. Biol. 51:263; Erlich, ed., (1989) PCR TECHNOLOGY (Stockton Press, N.Y.). ПЦР-анализ, использованный в настоящем раскрытии, считается одним, но не единственным примером способа полимеразной реакции для амплификации опытного образца нуклеиновой кислоты. Данный способ включает применение известных последовательностей нуклеиновой кислоты в качестве праймеров и полимеразы нуклеиновой кислоты для амплификации или получения специфической части нуклеиновой кислоты.
Термин «возможный» или «возможно» означает то, что событие или ситуация, которые следуют, могут, но не обязательно случаются, и данное описание включает примеры, в которых событие или обстоятельство случается или не случается. Например, фраза «возможно содержит 1-3 вариабельные области тяжелой цепи антитела» означает то, что вариабельная область тяжелой цепи антитела со специфической последовательностью может присутствовать, но не обязательно присутствует.
Термин «фармацевтическая композиция» относится к смеси, содержащей одно или более чем одно соединение согласно настоящему раскрытию или его физиологически/фармацевтически приемлемую соль или пролекарство и другие химические компоненты, такие как физиологически/фармацевтически приемлемые носители и эксципиенты. Данная фармацевтическая композиция нацелена на стимуляцию введения в организм, облегчение поглощения активного ингредиента и, посредством этого, оказание биологического эффекта.
Кроме того, настоящее раскрытие включает средство для лечения заболеваний, ассоциированных с IL-5, и данное средство содержит моноклональное антитело по настоящему раскрытию или его фрагмент антитела в качестве активного ингредиента.
Нет ограничения заболеваний, ассоциированных с IL-5, при условии, что они ассоциированны с IL-5. Например, терапевтические ответы, индуцированные молекулами по настоящему раскрытию, могут быть получены посредством связывания с человеческим IL-5 и, вследствие этого, репрессии или ингибирования стимуляции, индуцированной эозинофилами. Таким образом, при нахождении в препаратах и композициях, подходящих для терапевтических применений, молекулы по настоящему раскрытию являются очень полезными для индивидов, которые страдают от аллергических и/или атопических ответов, или ответов, ассоциированных с эозинофилами, например, астмы, обострения астмы, опасного приступа астмы, хронической пневмонии, аллергического ринита, круглогодичного аллергического ринита, аллергического бронхолегочного аспергиллеза, эозинофилии, синдрома Чарга-Стросса, атопического дерматита, онхоциркозного дерматита, эпизодического ангиоотека, синдрома эозинофильной миалгии, эозинофильного гастроэнтерита, инфекции гельминтами, болезни Ходжкина, полипов носа, синдрома Леффлера, уртикарии, эозинофильного гиперпластического бронхита, узелкового артериита, синусита, эозинофильного эзофагита, аллергического эозинофильного эзофагита, аллергического конъюнктивита, онхоциркозного дерматита, эндометриоза, зависимого от стероидов эозинофильного бронхита и тому подобных, но не ограничиваясь ими. В предпочтительном воплощении такое лечение ингибирует или уменьшает инфильтрующие эозинофилы в легочной ткани. Антитела или их фрагменты могут вводиться от трех раз в сутки до одного раза каждые шесть месяцев и могут вводиться внутривенно, подкожно, внутримышечно, парентерально или местно.
Кроме того, настоящее раскрытие относится к способу иммуновыявления или иммуноанализа IL-5, реактивам для иммуновыявления или иммуноанализа IL-5, иммуновыявлению или иммуноанализу клеток, экспрессирующих IL-5, и диагностическому средству для осуществления диагностики заболевания, ассоциированного с IL-5, которое содержит моноклональное антитело или фрагмент антитела по настоящему раскрытию, специфично распознающий человеческий IL-5 и связывающийся с его аминокислотной последовательностью внеклеточной области или трехмерной структурой, в качестве активного ингредиента.
В настоящем раскрытии способом выявления или определения количества IL-5 может быть любой способ, известный в данной области. Например, он включает иммуновыявление или иммуноанализ.
Иммуновыявление или иммуноанализ представляет собой способ выявления или определения количества антитела или антигена посредством применения меченого антигена или антитела. Примеры иммуновыявления или иммуноанализа включают способ мечения радиоактивным веществом иммунологического антитела (RIA), ферментативный иммуноанализ (EIA (иммуноферментный анализ) или ELISA (твердофазный иммуноферментный анализ)), флуоресцентный иммуноанализ (FIA), люминисцентный иммуноанализ, вестерн-блоттинг, физико-химические анализы и тому подобное.
Вышеупомянутые заболевания, ассоциированные с IL-5, могут быть диагностированы посредством выявления или определения клеток, экспрессирующих IL-5, с использованем моноклонального антитела или фрагмента антитела по настоящему раскрытию.
Для того чтобы выявлять клетки, экспрессирующие полипептид, можно использовать известный иммуноанализ, предпочтительно иммуноосаждение, флуоресцентное окрашивание клеток, иммуногистохимическое окрашивание и тому подобное. Кроме того, можно использовать способ окрашивания флуоресцентным антителом с использованием системы FMAT8100HTS (Applied Biosystem) и тому подобного.
В настоящем раскрытии живой образец, используемый для выявления или определения IL-5, не является особенно ограниченным, при условии, что он вероятно содержит клетки, экспрессирующие IL-5, например, можно использовать клетки тканей, кровь, плазму, сыворотку, панкреатический сок, мочу, кал, тканевую жидкость или культуральную среду.
Диагностическое средство, содержащее моноклональное антитело или фрагмент антитела по настоящему раскрытию, может дополнительно содержать средство для осуществления реакции антиген-антитело или средство для выявления реакции, в зависимости от желательного диагностического способа. Средство для осуществления реакции антиген-антитело включает такие компоненты, как буфер и соли. Средство для выявления реакции включает реактивы, обычно используемые в способе иммуновыявления или иммуноанализа, например, такие как меченое вторичное антитело, распознающее моноклональное антитело, его фрагмент антитела или содержащий его конъюгат и субстрат, соответсвующий меткам.
2. Примеры и опытные примеры
Следующие примеры предоставляются для дополнительного описания настоящего раскрытия, но не предназначены для ограничения объема данного раскрытия. Экспериментальные способы, для которых специфические условия конкретно не указываются, обычно проводятся согласно традиционным условиям, см. Molecular Cloning, Laboratory Manual of antibody technology, Cold Spring Harbor Laboratory; или согласно условиям, рекомендованным изготовителем веществ или продуктов. Реактивы, для которых источники конкретно не указываются, представляют собой имеющиеся в продаже реактивы.
Пример 1. Получение антигенов IL-5 и белков для выявления
Конструирование и экспрессия антигена IL-5
Последовательности, кодирующие меченный His человеческий IL-5, IL-5 макака-резуса, мышиный IL-5, крысиный IL-5 или внеклеточный домен человеческого рецептора IL-5Rα, слитый с фрагментом человеческого IgG1-Fc, вставляли в вектор phr для конструирования экспрессионных плазмид, которые затем трансфицировали в HEK293. На сутки 6 после трансфекции образцы центрифугировали при 4500 об./мин в течение 10 мин, и собирали супернатанты клеток. Супернатант, содержащий рекомбинантный белок IL-5 или рецептора IL-5α, очищали с использованием никелевой колонки, и рекомбинантный слитый белок человеческий IL-5-Fc очищали с использованием колонки для аффинной хроматографии с белком А. Очищенный белок можно использовать в следующих примерах. Показаны следующие последовательности белка антигена:
1. Аминокислотная последовательность человеческого IL-5 с His меткой (rhIL-5-his)

Примечание: текст курсивом показывает His6-метку.
SEQ ID NO: 1
2. Аминокислотная последовательность IL-5 яванского макака с His меткой

Примечание: текст курсивом показывает His6-метку.
3. Аминокислотная последовательность мышиного IL-5 с His меткой

Примечание: текст курсивом показывает His6-метку.
4. Аминокислотная последовательность крысиного IL-5 с His меткой

Примечание: текст курсивом показывает His6-метку.
5. Аминокислотная последовательность человеческого рецептора IL-5α, слитого с человеческим фрагментом Fc

Примечание: текст курсивом показывает человеческую метку Fc.
Пример 2: конструирование и идентификация линий клеток с рекомбинантным рецептором IL-5α и рецептором IL-5α/β
Для скрининга функциональных антител в настоящем раскрытии сконструирована линия клеток CHO-S/IL-5α, экспрессирующая IL-5α, и линия клеток CHO-S/IL-5α/IL-5β, экспрессирующая и IL-5α, и IL-5β.
В частности, полноразмерный человеческий ген IL-5α (Q01344) клонировали в экспрессионный вектор для клетки млекопитающего - pTargeT, а линеаризованную плазмиду трансфицировали в клетки CHO-S посредством электропорации. Скрининг проводили под G418 в течение 2 недель, с последующими двумя циклами предельных разведений. Ген IL-5α, экспрессируемый на поверхности клетки, выявляли посредством FACS (флуоресцентная сортировка клеток), отбирали линии клеток CHO-S/IL-5α с высоким уровнем экспресии IL-5α и трансфицировали в линеаризованную pcDNA3.1-IL-5β посредством электропорации. Осуществляли скрининг с G418 и зеоцином в течение 2 недель, с последующими двумя циклами предельных разведений. Ген IL-5α и IL-5β, экспрессируемых на поверхности клетки, выявляли посредством FACS, и отбирали линии клеток CHO-S/IL-5α/IL-5β с высокой экспрессией IL-5α и IL-5β.
Пример 3: получение мышиного моноклонального антитела против человеческого IL-5
Две группы мышей Balb/c (5 мышей/группу) и четыре группы мышей SJL (5 мышей/группу) иммунизировали рекомбинантным белком rhIL-5-his и адъювантом Фрейнда CFA (Sigma, № партии SLBQ1109V); или IFA (Sigma, № партии SLBJ2845V) в двух дозировках 100 г/50 г/50 г (группа с высокой дозой) и 25 г/12,5 г/12,5 г (группа с низкой дозой) соответсвенно. Специфичный иммунный ответ на IL-5 определяли посредством выявления сывороточного титра ELISA, анализом блокирования лиганда-рецептора и анализом ингибирования пролиферации TF-1. Отбирали мышей с лучшим специфичным иммунным ответом, умерщвляли, отбирали клетки селезенки и сливали с миеломными клетками.
Первичный скрининг проводили с использованием анализа связывания ELISA против человеческого IL-5. Гибридомные клетки переносили в 24-луночные планшеты, и супернатанты подвергали повторному скринингу посредством анализа связывания ELISA против IL-5 человека, яванского макака или мыши посредством анализа блокирования на основе ELISA против рецептора IL-5 и посредством анализа ингибирования пролиферации TF-1. После такого скрининга полученные позитивные клоны подвергали двум циклам субклонирования с получением гибридомных клонов для продукции антител. Полученные антитела очищали аффинной хроматографией.
Очищенные антитела подвергали следующим анализам: SEC-HPLC (гель-фильтрация на основе высокоэффективной жидкостной хроматографии), выявление содержания эндотоксинов, анализ Biacore на аффинность к разным IL-5, анализ блокирования против рецептора IL-5 на основе FACS, анализ ингибирования пролиферации TF-1, анализ адгезии эозинофилов и оценка эффективности мышиной модели астмы и модели нейтрализации in vivo морских свинок. Отбирали линии клеток моноклональной гибридомы mAb1705, mAb1706, mAb1780, mAb1773 и mAb1779 из-за их превосходных активностей in vitro и in vivo.
Клонировали последовательности из позитивной гибридомы следующим образом. Отбирали гибридомные клетки в логарифмической фазе роста, выделяли РНК с использованием Trizol (Invitrogen, кат. №15596-018) согласно инструкции или набору изготовителя, и обратную транскрипцию проводили с использованием набора обратной транскриптазы PrimeScript™ (Takara, кат. №2680А). кДНК, полученные обратной транскрипцией, подвергали ПЦР-амплификации с использованием набора праймеров для мышиного Ig (Novagen, ТВ326 Rev. В 0503), и образующиеся продукты секвенировали. Получали аминокислотные последовательности, соответствующие последовательностям ДНК вариабельных областей тяжелой и легкой цепи mAb1705, mAb1706, mAb1780, mAb1773 и mAb1779 (аминокислотные остатки CDR VH/VL определяли и аннотировали посредством системы нумерации Kabat):
последовательность вариабельной области тяжелой цепи мышиного mAb1705
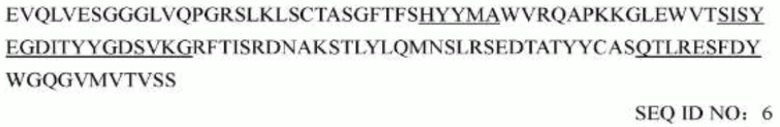
последовательность вариабельной области легкой цепи мышиного mAb1705

последовательность вариабельной области тяжелой цепи мышиного mAb1706
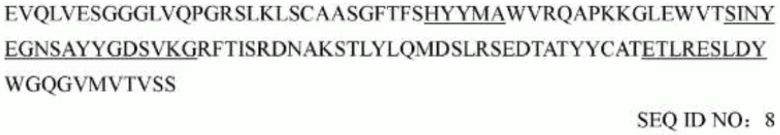
последовательность вариабельной области легкой цепи мышиного mAb1706

последовательность вариабельной области тяжелой цепи мышиного mAb1780

последовательность вариабельной области легкой цепи мышиного mAb1780
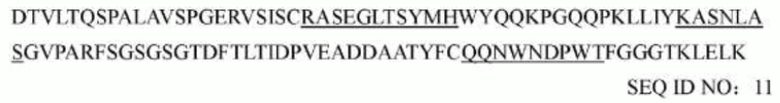
последовательность вариабельной области тяжелой цепи мышиного mAb1773
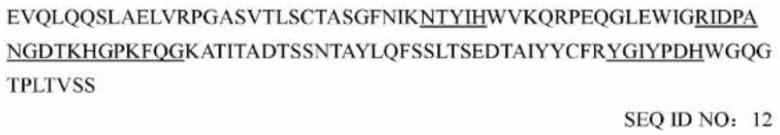
последовательность вариабельной области легкой цепи мышиного mAb1773

последовательность вариабельной области тяжелой цепи мышиного mAb1779

последовательность вариабельной области легкой цепи мышиного mAb1779

Последовательности CDR легкой и тяжелой цепи каждого антитела показаны в Таблице 1.
Результаты активности анализа Biacore показаны в Таблице 2.
Данные результаты показывают то, что мышиные антитела по настоящему раскрытию имеют высокую аффинность к данному антигену.
Пример 4: очистка рекомбинантных белков, родственных IL-5, и очистка гибридомных антител и рекомбинантных антител
4.1 Стадии для очистки рекомбинантных белков IL-5-Flag-His:
Образцы центрифугировали при высокой скорости для удаления примесей и концентрировали до подходящего объема. Аффинную колонку NI-NTA (QIAGEN, кат. №30721) уравновешивали PBS (фосфатно-солевой буферный раствор) и промывали 2-5 объемами колонки. Экспрессированные клетками супернатанты без примесей загружали на колонку, которую затем промывали PBS, пока показание А280 (поглощение при 280 нм) не падало до исходного уровня. Затем колонку промывали PBS для удаления неочищенного белка. Интересующий белок элюировали промывочным буфером (20 мМ имидазол) и затем элюирующим буфером (300 мМ имидазол), и собирали элюированный пик.
Отобранный элюат дополнительно очищали посредством обмена ионов (колонка Hiload 16/600 superdex 200). Данную колонку уравновешивали примерно 2 объемами колонки PBS для обеспечения рН 7,4. Элюирующий буфер, содержащий интересующий белок, концентрировали и загружали на колонку для последующей очистки. Данные образцы собирали, идентифицировали SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и LC-MS (жидкостная хроматография-масс-спектрометрия), и затем аликвотировали для применения.
4.2. Очистка экспрессированного гибридомой антитела и слитых с Fc белков
Образцы экспрессированного клетками супернатанта центрифугировали при высокой скорости для удаления примесей, и затем экспрессированные гибридомой супернатанты очищали с использованием колонки с белком G, супернатанты, экспрессирующие слитый белок Fc, очищали с использованием колонки с белком А. Колонку промывали PBS, пока показание А280 не падало до исходного уровня. Интересующие белки элюировали 100 мМ уксусной кислотой, рН 3,0, и нейтрализовали 1 М Tris-HCl, рН 8,0. Элюированные образцы подходящим образом концентрировали и дополнительно очищали посредством гель-хроматографии на Superdex 200 (GE), предварительно уравновешенном PBS. Пики, лишенные агрегатов, собирали и аликвотировали для применения.
Пример 5: конструирование гуманизации моноклональных антител против человеческого IL-5
Гуманизацию мышиных моноклональных антител против человеческого IL-5 проводили, как раскрыто в литературе в данной области. Вкратце, константные области мышиных антител заменяли человеческими константными областями, CDR мышиных антител прививали на человеческую матрицу, имеющую наивысшую гомологию FR, и некоторые аминокислотные остатки в области FR, которые имеют ключевое влияние на поддержание конформации антитела и влияют на связывание антитела с антигеном, подвергали обратной мутации.
Посредством выравнивания с базой данных IMGT генов зародышевой линии вариабельной области тяжелой и легкой цепи человеческого антитела в качестве матриц, соответственно, выбирали гены вариабельной области тяжелой и легкой цепи человеческой зародышевой линии, которые имеют высокую идентичность аминокислотной последовательности с антителом mAb-1705, mAb-1706, mAb1780, mAb1773 и mAb1779. CDR данных мышиных антител отдельно прививали на соответствующие матрицы человеческого происхождения с образованием последовательности вариабельной области в порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Аминокислотные остатки определяли и аннотировали посредством системы нумерации Kabat.
Выбор человеческой области FR и обратная мутация ключевых аминокислот
На основе типичной структуры CDR VH/VL полученного мышиного антитела из базы данных человеческой зародышевой линии выбирали гомологичные последовательности вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH). Полученные в результате последовательности VL и VH человеческой зародышевой линии ранжировали от высокого балла до низкого на основе гомологии FR, и последовательности зародышевой линии с наивысшей гомологией FR выбирали в качестве главных матриц. CDR мышиных антител прививали на человеческие матрицы. И затем, используя программное обеспечение и на основе трехмерной структуры мышиных антител, вставленные остатки, остатки, которые непосредственно взаимодействуют с областями CDR, и остатки, которые имеют значительное влияние на конформацию VL и VH, подвергали обратным мутациям. Кроме того, химически нестабильные аминокислотные остатки оптимизировали для получения конечных гуманизированных молекул.
5.1 Выбор гуманизированного каркаса для клона гибридомы mAb1705
IGHV3-23*04 был выбран в качестве матрицы для VH h1705, и IGKV1-12*01 был выбран в качестве матрицы для VL. CDR мышиного mAb1705 прививали на человеческую матрицу. Вставленные остатки и остатки, которые непосредственно взаимодействуют с областями CDR, были обнаружены программой и были подвергнуты обратной мутации. Вариабельные области легкой и тяжелой цепи гуманизированных антител были сконструированы, как показано в Таблице 3.


Конкретные последовательности вариабельных областей
гуманизированного антитела h1705 являются следующими:
>h1705_VL.1 (SEQ ID NO: 46)

>h1705_VL.1A (SEQ ID NO: 47)

>h1705_VL.1B (SEQ ID NO: 48)
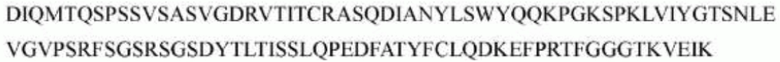
>h1705_VH.1 (SEQ ID NO: 49)
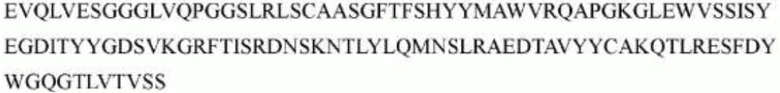
>h1705_VH.1A (SEQ ID NO: 50)
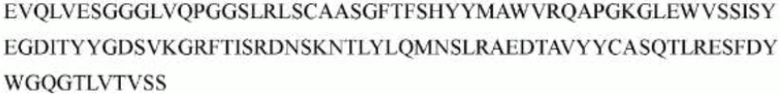
>h1705_VH.1B (SEQ ID NO: 51)

Каждую из приведенных выше вариабельных областей легкой цепи объединяли с константной областью легкой цепи, как определено в SEQ ID NO: 53, с образованием конечных интактных последовательностей легкой цепи. Каждую вариабельную область тяжелой цепи объединяли с константной областью тяжелой цепи, как определено в SEQ ID NO: 52, с образованием конечных последовательностей тяжелой цепи.
Последовательность константной области гуманизированного антитела
>Константная область тяжелой цепи IgG1:

Примечание: подчеркнутый текст представляет сконструированную мутацию M252Y, S254T или Т256Е
>Константная область каппа легкой цепи:
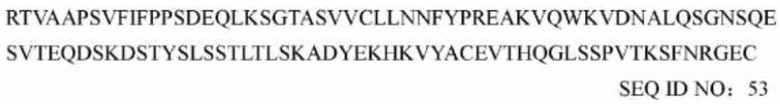
5.2 Выбор гуманизированного каркаса для клона гибридомы mAb1706
IGHV3-23*04 был выбран в качестве матрицы для VH h1706, и IGKV1-12*01 был выбран в качестве матрицы для VL. CDR мышиного mAb1706 прививали на человеческую матрицу. Вставленные остатки и остатки, которые непосредственно взаимодействуют с областями CDR, были обнаружены программой и были подвергнуты обратной мутации. Вариабельные области легкой и тяжелой цепи гуманизированных антител были сконструированы, как показано в Таблице 5.

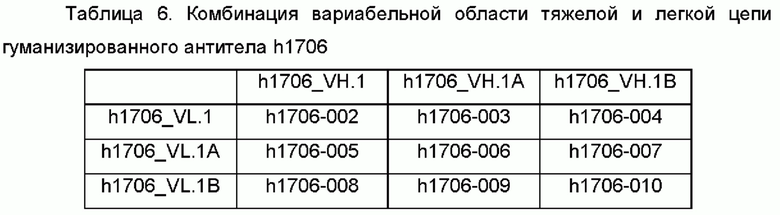
Показаны следующие конкретные последовательности вариабельных областей гуманизированного антитела h1706:
>h1706_VL.1 (SEQ ID NO: 54)
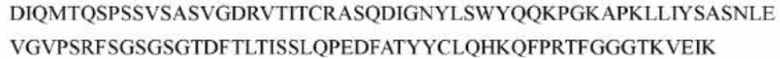
>h1706_VL.1A (SEQ ID NO: 55)
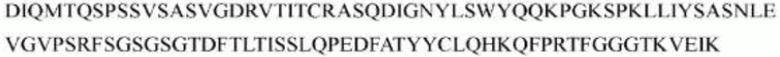
>h1706_VL.1B (SEQ ID NO: 56)

>h1706_VH.1 (SEQ ID NO: 57)
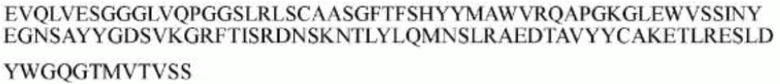
>h1706_VH.1A (SEQ ID NO: 58)

>h1706_VH.1B (SEQ ID NO: 59)
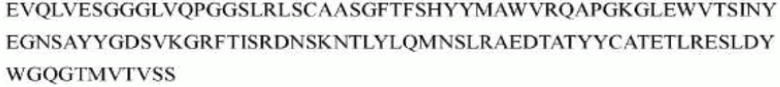
Каждую из приведенных выше вариабельных областей легкой цепи объединяли с константной областью легкой цепи, как определено в SEQ ID NO: 53, с образованием конечных последовательностей интактной легкой цепи. Каждую вариабельную область тяжелой цепи объединяли с константной областью тяжелой цепи, как определено в SEQ ID NO: 52, с образованием конечных последовательностей тяжелой цепи.
5.3 Выбор гуманизированного каркаса для клона гибридомы mAb1780
IGHV1-2*02 был выбран в качестве матрицы для VH h1780, и IGKV3-11*01 был выбран в качестве матрицы для VL. CDR мышиного mAb1780 прививали на человеческую матрицу. Вставленные остатки и остатки, которые непосредственно взаимодействуют с областями CDR, были обнаружены программой и были подвергнуты обратной мутации. Вариабельные области легкой и тяжелой цепи гуманизированных антител были сконструированы, как показано в Таблице 7.

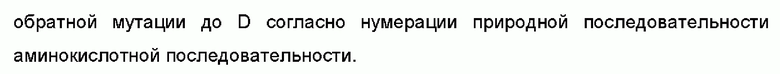
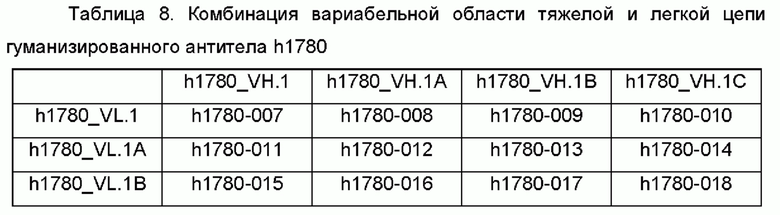
Показаны следующие конкретные последовательности вариабельных областей гуманизированного антитела h1780:
>h1780_VL.1 (SEQ ID NO: 60)
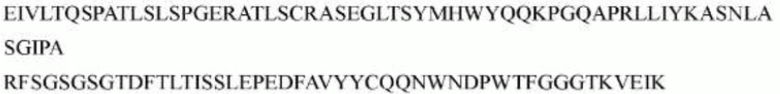
>h1780_VL.1A (SEQ ID NO: 61)
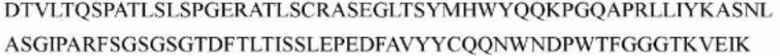
>h1780_VL.1B (SEQ ID NO: 62)

>h1780_VH.1 (SEQ ID NO: 63)
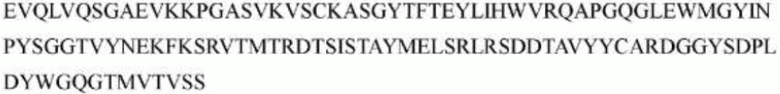
>h1780_VH.1A (SEQ ID NO: 64)
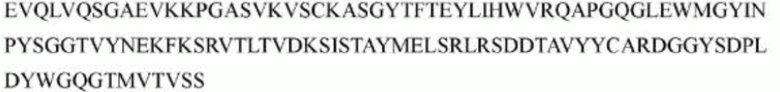
>h1780_VH.1B (SEQ ID NO: 65)

>h1780_VH.1C (SEQ ID NO: 66)

Каждую из приведенных выше вариабельных областей легкой цепи объединяли с константной областью легкой цепи, как определено в SEQ ID NO: 53, с образованием конечных последовательностей интактной легкой цепи. Каждую вариабельную область тяжелой цепи объединяли с константной областью тяжелой цепи, как определено в SEQ ID NO: 52, с образованием конечных последовательностей тяжелой цепи.
5.4 Выбор гуманизированного каркаса для клона гибридомы mAb1773
IGHV3-73*01 был выбран в качестве матрицы для VH h1773, и IGKV1-39*01 был выбран в качестве матрицы для VL. CDR мышиного mAb1773 прививали на человеческую матрицу. Вставленные остатки и остатки, которые непосредственно взаимодействуют с областями CDR, были обнаружены программой и были подвергнуты обратной мутации. Вариабельные области легкой и тяжелой цепи гуманизированных антител были сконструированы, как показано в Таблице 9. Тем временем, для того чтобы удалить изомеризованные сайты в областях CDR, N, расположенную в HCDR2 (RIDPANGDTK HGPKFQG) h1773 заменяли на V (т.е., N55V) с образованием вариабельной области тяжелой цепи и антитела, содержащего вариант HCDR2 (последовательность мутировавшей HCDR2 показана в виде SEQ ID NO: 82: RIDPAVGDTKHGPKFQG).
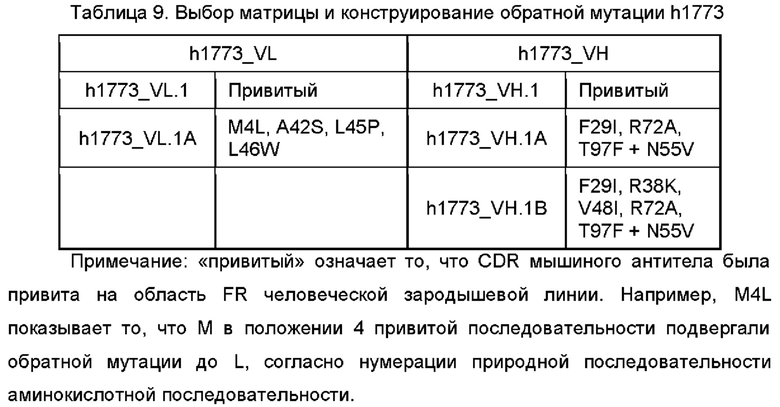
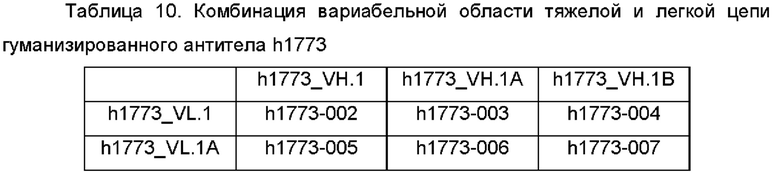
Показаны следующие конкретные последовательности вариабельных областей гуманизированного антитела h1773:
>h1773_VL.1 (SEQ ID NO: 67)
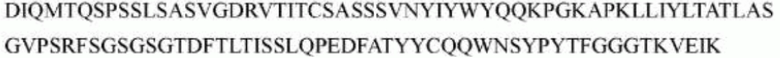
>h1773_VL.1A (SEQ ID NO: 68)
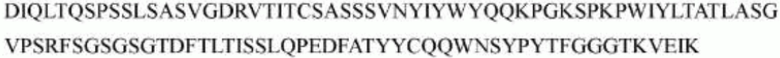
>h1773_VH.1 (SEQ ID NO: 69)
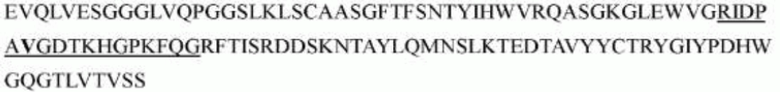
>h1773_VH.1A (SEQ ID NO: 70)

>h1773_VH.1B (SEQ ID NO: 71)

Каждую из приведенных выше вариабельных областей легкой цепи объединяли с константной областью легкой цепи, как определено в SEQ ID NO: 53, с образованием конечных последовательностей интактной легкой цепи. Каждую вариабельную область тяжелой цепи объединяли с константной областью тяжелой цепи, как определено в SEQ ID NO: 52, с образованием конечных последовательностей тяжелой цепи.
5.5 Выбор гуманизированного каркаса для клона гибридомы mAb1779
IGHV1-2*02 был выбран в качестве матрицы для VH h1779, и IGKV1-33*01 был выбран в качестве матрицы для VL. CDR мышиного h1773 прививали на человеческую матрицу. Вставленные остатки и остатки, которые непосредственно взаимодействуют с областями CDR, были обнаружены программой и были подвергнуты обратной мутации. Вариабельные области легкой и тяжелой цепи гуманизированных антител были сконструированы, как показано в Таблице 11.
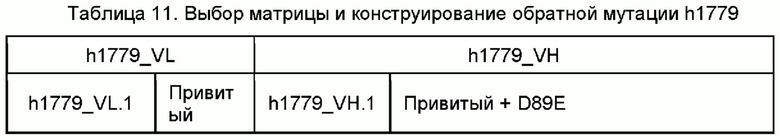
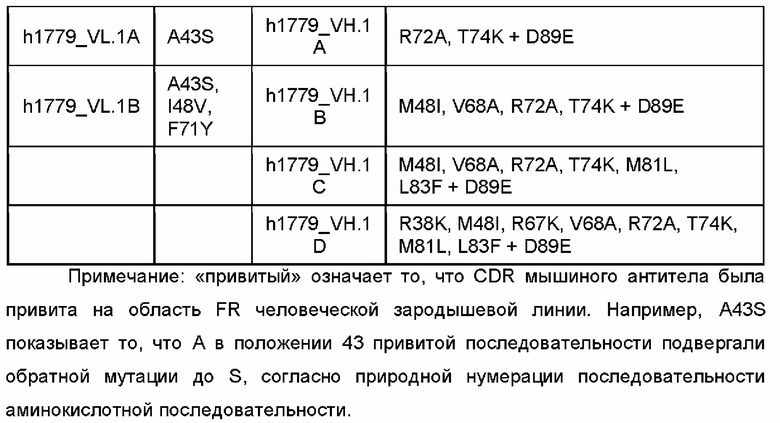
Сконструированные гуманизированные молекулы объединяли с разными молекулами, как показано в Таблице 12.
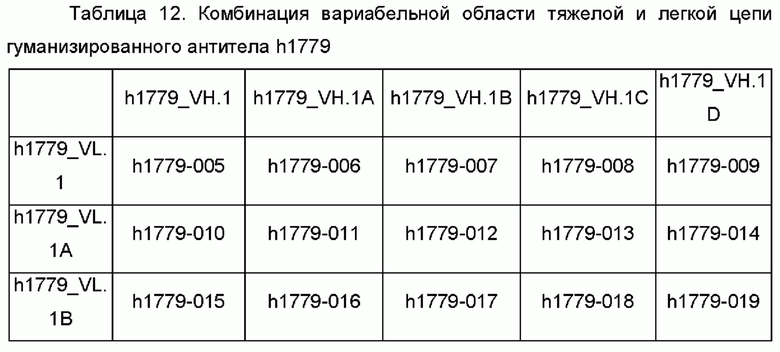
Показаны следующие конкретные последовательности вариабельных областей гуманизированного антитела h1779:
>h1779_VL.1 (SEQ ID NO: 72)
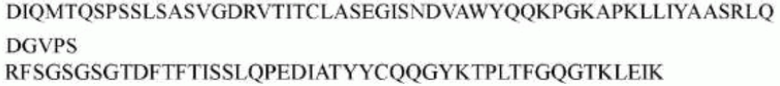
>h1779_VL.1A (SEQ ID NO: 73)
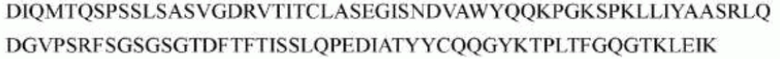
>h1779_VL.1B (SEQ ID NO: 74)
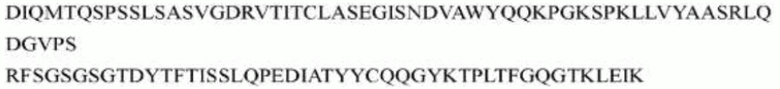
>h1779_VH.1 (SEQ ID NO: 75)
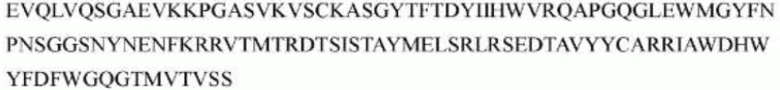
>h1779_VH.1A (SEQ ID NO: 76)
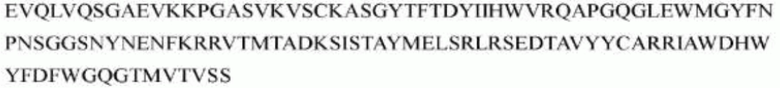
>h1779_VH.1B (SEQ ID NO: 77)

>h1779_VH.1C (SEQ ID NO: 78)
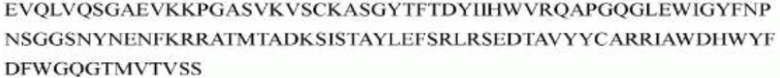
>h1779_VH.1D (SEQ ID NO: 79)

Каждую из приведенных выше вариабельных областей легкой цепи объединяли с константной областью легкой цепи, как определено в SEQ ID NO: 53, с образованием конечных последовательностей интактной легкой цепи. Каждую вариабельную область тяжелой цепи объединяли с константной областью тяжелой цепи, как определено в SEQ ID NO: 52, с образованием конечных последовательностей тяжелой цепи.
Тем временем, антитело Hu39D10 против IL-5, раскрытое в WO 2012083370 A1, использовали в настоящем раскрытии в качестве позитивного контроля, и его последовательность тяжелой и легкой цепи показана в SEQ ID NO: 80 и SEQ ID NO: 81 соотвественно.
Последовательность тяжелой цепи Hu39D10


Последовательность легкой цепи Hu39D10

Пример 6. Получение рекомбинантных химерных антител и гуманизированных антител
1. Молекулярное клонирование рекомбинантных химерных антител
Позитивные молекулы антител, полученные скринингом гибридомы, секвенировали с получением последовательностей генов, кодирующих вариабельные области. На основе полученных последовательностей конструировали прямые и обратные праймеры, и фрагмент гена VH/VK каждого антитела конструировали посредством ПЦР с использованием последовательности гена в качестве матрицы. Фрагмент гена VH/VK затем подвергали гомологичной рекомбинации с экспрессионным вектором pHr (содержащим сигнальный пептид и фрагмент гена константной области hIgG1/hkappa (CH1-Fc/CL) для конструирования полноразмерной экспрессионной плазмиды для рекомбинантного химерного антитела VH-CH1-Fc-pHr/VL-CL-pHr с образованием пяти химерных антител Ch1705, Ch1706, Ch1780, Ch1773 и Ch1779.
2. Молекулярное клонирование гуманизированных антител
На последовательностях сконструированного гуманизированного антитела проводили оптимизацию кодонов с получением кодирующих последовательностей генов с предпочтением к человеческим кодонам. Конструировали ПЦР-праймеры для построения фрагмента гена VH/VK каждого антитела, и затем фрагмент гена VH/VK подвергали гомологичной рекомбинации с экспрессионным вектором pHr (содержащим сигнальный пептид и фрагмент гена константной области hIgG1/hkappa (CH1-Fc/CL)) с построением полноразмерной экспрессионной плазмиды гуманизированного антитела VH-CH1-Fc-pHr/VL-CL-pHr.
3. Экспрессия и очистка рекомбинантных химерных антител и гуманизированных антител
Плазмиду, экспрессирующую легкую или тяжелую цепь антитела, трансфицировали в клетки HEK293E в соотношении 1:1,2. Через 6 суток экспрессионные супернатанты отбирали, центрифугировали при высокой скорости для удаления примесей и очищали посредством колонки с белком А. Колонку промывали PBS, пока показание А280 не падало до исходного уровня. Интересующий белок элюировали кислотным элюционным буфером - рН 3,0-рН 3,5 - и нейтрализовали 1 М Tris-HCl, рН 8,0-9,0. Элюированные образцы подходящим образом концентрировали и дополнительно очищали посредством гель-хроматографии на Superdex 200 (GE), предварительно уравновешенном PBS, для удаления агрегатов. Пики мономера собирали и аликвотировали для применения.
Эффективность и полезные эффекты антител в настоящем раскрытии подтверждали посредством следующих способов анализа.
Оценка биологической активности in vitro
Опытный пример 1: связывание мышиных антител против IL-5 с IL-5 разных видов, определенное посредством анализа Biacore
Аффинность между опытными мышиными антителами против IL-5 и человеческим IL-5 определяли с использованием прибора Biacore Т200 (GE).
Молекулы, подлежащие анализу, аффинно захватывали посредством биосенсорного чипа на основе белка А, и затем антиген (рекомбинантный человеческий, обезьяний и мышиный IL-5, полученный в Примере 1) протекал через поверхность чипа, и сигналы реакции выявляли в реальном времени посредством применения прибора Biacore Т200 с получением кривых связывания и диссоциации. После диссоциации каждый цикл эксперимента завершали, биосенсорный чип промывали и регенерировали регенерирующим раствором на основе глицина-соляной кислоты (рН 1,5). Данные аппроксимировали к модели Лангмюра (1:1) посредством применения программы BIAevaluation версии 4.1 GE, и значения аффинности получали, как показано в Таблице 13.

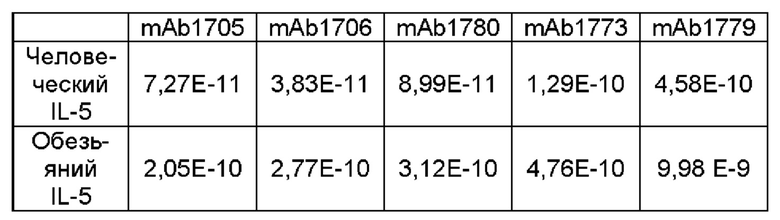
Данный пример демонстрирует то, что все антитела mAb 1705, mAb 1706, mAb 1780, mAb 1773 и mAb 1779 по настоящему раскрытию имеют высокую аффинность к IL-5 разных видов (человек, обезьяна).
Опытный пример 2: аффинность гуманизированных антител против IL-5 к IL-5 разных видов, определенная анализом Biacore
Аффинность между опытными гуманизированными антителами против IL-5 и человеческим IL-5 определяли с использованием прибора Biacore Т200 (GE).
Молекулы, подлежащие анализу, аффинно захватывали посредством биосенсорного чипа на основе белка А, и затем антиген (полученный в Примере 1) протекал через поверхность чипа, и сигналы реакции выявляли в реальном времени посредством применения прибора Biacore Т200 с получением кривых связывания и диссоциации. После диссоциации каждый цикл эксперимента завершали, биосенсорный чип промывали и регенерировали регенерирующим раствором на основе глицина-соляной кислоты (рН 1,5). Данные аппроксимировали к модели Лангмюра (1:1) посредством применения программы BIAevaluation версии 4.1 GE, и значения аффинности получали, как показано в Таблице 14.

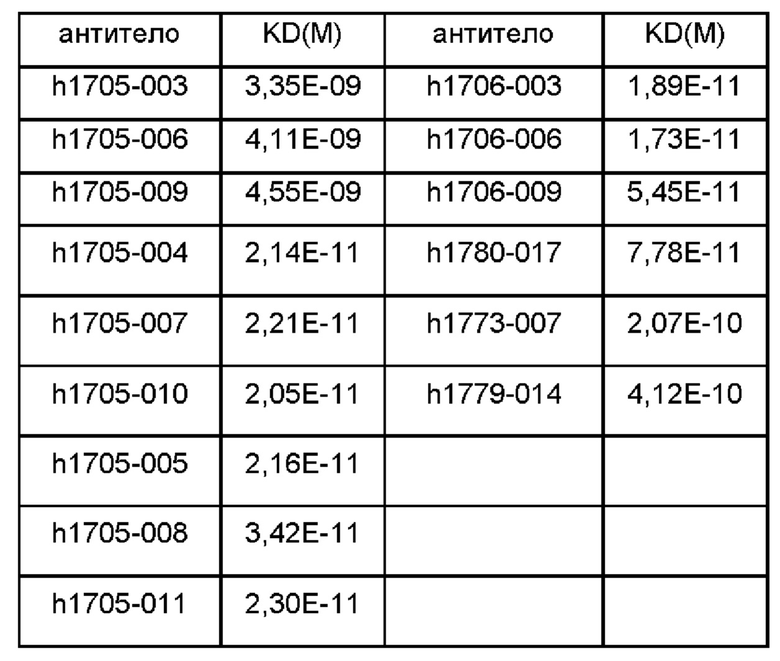
Данные результаты показывают то, что гуманизированные антитела против IL-5 все еще имеют высокую аффинность к человеческому IL-5 (в том, что касается гуманизированных вариантов других мышиных антител, были приведены только типичные данные, за исключением каждого гуманизированного варианта h1705).
Опытный пример 3: мышиные антитела против IL-5 блокируют связывание между IL-5 и рецептором IL-5α при определении посредством анализа ELISA
Для демонстрации способности антител против IL-5 предотвращать связывание IL-5 с внеклеточной областью рекомбинантно экспрессированного белка рецептора IL-5α, планшет ELISA покрывали IL-5 (5 мкг/мл в PBS), инкубировали при 37°С в течение 1 часа, жидкость отбрасывали, затем добавляли блокирующий раствор с 5% обезжиренного молока, разведенного PBS, в объеме 200 мкл/лунку и блокировали при 37°С в течение 2,5 часов в инкубаторе. После завершения блокирования блокирующий раствор отбрасывали, данный планшет 5 раз промывали буфером PBST (PBS, содержащий 0,05% Tween-20, рН 7,4), добавляли 25 мкл 10 мкг/мл IL-5Rα (в 1% BSA (бычий сывороточный альбумин)), который был мечен набором для мечения биотином (Dojindo Chemical, LK03), и затем добавляли 25 мкл градиентно разведенного антитела, которое конкурировало с IL-5Rα за связывание с IL-5, и инкубировали при 37°С в течение 1 часа. После инкубации реакционный раствор в планшете отбрасывали, планшет 5 раз промывали PBST, добавляли стрептавидин-пероксидазный полимер (Sigma, S2438-250UG), разведенный 1:600 раствором для разведения образца, в объеме 50 мкп/лунку и инкубировали при 37°С в течение 1 часа. После промывки планшета 5 раз PBST добавляли 50 мкл/лунку хромогенного субстрата ТМВ (тетраметилбензидин) (KPL, 52-00-03), инкубировали при комнатной температуре в течение 3-10 мин, и добавляли 50 мкл/лунку 1 М H2SO4 для остановки реакции. Значение поглощения считывали при 450 нм с использованием микропланшет-ридера NOVOStar. Рассчитывали значения IC50 (полумаксимальное ингибирование) для блокирования антителами против IL-5 связывания между IL-5 и IL-5Rα. Результаты показаны в Таблице 15, оба антитела по настоящему раскрытию могут эффективно ингибировать связывание IL-5 с его рецептором.

Опытный пример 4: антитела против IL-5 блокируют связывание между IL-5 и рецептором IL-5 при определении посредством FACS
Для идентификации образующихся антител против IL-5, которые могут блокирвать рецептор IL-5 на поверхности клетки, авторы данного изобретения сконструировали рекомбинантную линию клеток CHOS, которая сильно экспрессирует оба рецептора IL-5Rα/β. Данный эксперимент идентифицировал то, что антитела против IL-5 могут предотвращать связывание IL-5 с рекомбинантным рецептором IL-5α/β на поверхности линии клеток CHOS соответственно.
Конкретный способ: CHO-S-IL-5Rα и β культивировали с CD-CHO, содержащей 100 нг/мл G418 и 25 нг/мл зеоцина, и концентрация во время культивирования клеток составляла не больше, чем 3×106 клеток/мл. Клетки IL-5Rα/β-CHOS в хорошем состоянии центрифугировали (1000 об./мин, 5 мин), один раз промывали 10% FBS (фетальная телячья сыворотка) в PBS, подсчитывали, концентрацию клеток доводили до 4×106 клеток/мл, и 25 мкл которых добавляли в 96-луночный планшет с круглым дном. Антитела, подлежащие анализу, разводили раствором PBS, содержащим 10% FBS, с исходной концентрацией 200 мкг/мл, и посредством разведения 1:10 были получены 8 градиентов. 25 мкл 100 нг/мл IL-5, меченого набором для мечения биотином (Dojindo Chemical, LK03), однородно смешивали с 50 мкл каждого разведенного антитела, добавляли в 96-луночный планшет, в который были добавлены клетки, и инкубировали при 4°С в течение 1 часа. После инкубации, центрифугирования при 4°С (400 g, 5 мин), супернатант отбрасывали, центрифугировали и промывали 200 мкл предварительно охлажденного PBS, повторяли дважды; добавляли вторичное антитело, конъюгированное с РЕ(фикоэритрин)-авидином, разведенное 1:1333, и инкубировали в темноте при 40°С в течение 40 мин, супернатант отбрасывали после центрифугирования при 4°С (400 g, 5 мин), добавляли 200 мкл предварительно охлажденного PBS, клетки диссоциировали, центрифугировали и промывали при 4°С, повторяли три раза и добавляли 100 мкл PBS. Планшет считывали на приборе, и рассчитывали значения IC50 блокирования антителами против IL-5 связывания между IL-5 и IL-5Rα/β на основе значений флуоресцентных сигналов. Результаты показаны в Таблице 16 и на Фиг. 1.

Результаты показывают то, что антитела h1705-008, h1706-009, h1780-017, h1773-007 и h1779-014 демонстрировали более сильную способность блокировать связывание IL-5 с рецептором IL-5 на поверхности клетки.
Опытный пример 5: антитела против IL-5 ингибируют индуцированную IL5 пролиферацию клеток TF1
IL-5 может индуцировать пролиферацию клеток TF-1, и антитела против IL-5 могут предотвращать стимуляцию IL-5 пролиферации клеток TF-1.
В частности, клетки TF-1 (АТСС, CRL-2003) культивировали в RPMI1640, содержащей 10% FBS и 2 нг/мл rhGM-CSF (рекомбинантный человеческий гранулоцитарно-макрофагальный колониестимулирующий фактор) (Lianke Bio, каталожный №96-AF-300-03-20), инкубировали в инкубаторе при 37°С, 5% CO2, с плотностью клеток не больше, чем 1×106 клеток/мл. Для выявления антител клетки, находящиеся в логарифмической фазе роста, три раза промывали PBS посредством центрифугирования при 800 об./мин в течение 5 мин, и плотность клеток доводили до 6000 клеток/лунку/90 мкл с использованием RPMI1640 (FBS: 2%, рекомбинантный человеческий IL-5: 10 нг/мл). Добавляли в 96-луночный планшет 10 мкл градиентных разведений антител, подлежащих анализу, и культивировали в течение 3 суток. Добавляли 30 мкл клеточного титра и перемешивали. Значения IC50 рассчитывали на основе показаний. Результаты показаны в Таблице 17 ниже.
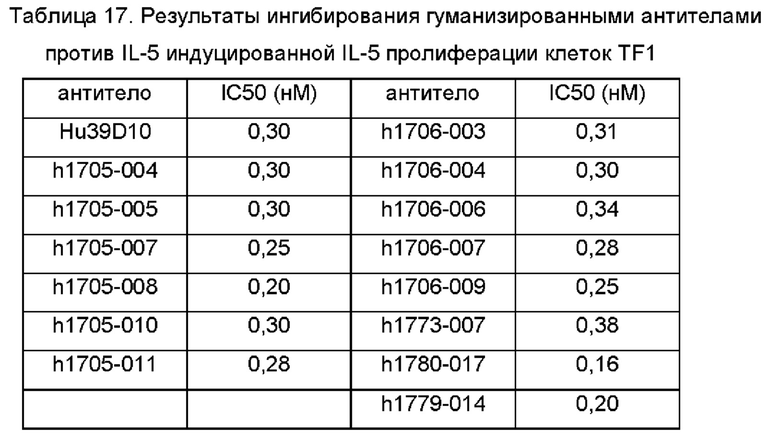
Опытный пример 6: антитела против IL-5 ингибируют индуцированную IL-5 адгезию эозинофилов
IL-5 может индуцировать дифференциацию, созревание, миграцию и активацию эозинофилов, вызывать воспаление в дыхательной системе и приводить к астме. Согласно тому принципу, что цитокин IL-5 может стимулировать адгезию эозинофилов и активировать эозинофилы, в данном опытном примере определяли эффекты IL-5-специфичных антител на блокирование пути, опосредованного IL-5, посредством сбора и очистки эозинофилов из человеческой периферической крови и выявления эффектов in vitro антител против IL-5 на блокирование опосредованной IL-5 адгезии эозинофилов.
В частности, человеческая периферическая кровь была в 5 раз разведена PBS, содержащим 2 мМ EDTA (этилендиаминтетрауксусная кислота), моноциты и гранулоциты разделяли посредством Percoll™ (с градиентом плотности 1,088), слой эритроцитов, содержащий гранулоциты, аккуратно отсасывали, и эритроциты удаляли с использованием лизисного буфера для эритроцитов. Остающиеся клетки подсчитывали, добавляли отделяющие магнитные шарики, покрытые человеческим антителом против CD16 (Miltenyi, каталожный №130-045-701), инкубировали в течение 30 мин и пропускали через колонку с магнитными шариками. Элюент клеточной фракции, который, главным образом, состоял из эозинофилов, прямо собирали. Образующиеся эозинофилы подсчитывали и добавляли в 96-луночный планшет для культуры клеток, предварительно покрытый антителом против IgG, примерно 1×104 клеток на лунку, добавляли человеческий IL-5 (20 нг/мл) и разные концентрации молекул антитела против IL-5 (начиная с 10 мкг/мл, с 3-кратным разведением, 10 уровней концентраций). Планшет для культуры клеток инкубировали в инкубаторе при 37°С, 5% CO2, в течение 1 ч, и затем добавляли 0,3% СТАВ (цетилтриметиламмоний бромид) для лизирования клеток. Наконец, добавляли реакционный субстрат пероксидазы ТМВ для развития окрашивания. На микропланшет-ридере считывали значения поглощения ОП450. Максимальное значение поглощения считывали в лунке с добавлением только IL-5, и лунки, не содержащие ни IL-5, ни препарата в виде антитела, принимали за контроль фона. Значение ингибирования каждой концентрации препаратов антитела относительно максмального значения поглощения рассчитывали как (максимальное значение поглощения - [препарат антитела] / (максимальное значение поглощения - значение контроля фона) × 100%, и рассчитывали IC50. Результаты показаны в Таблице 18.

Данные результаты показывают то, что гуманизированные антитела по настоящему раскрытию демонстрируют сильную способность ингибировать опосредованную IL-5 адгезию эозинофилов.
Опытный пример 7: оценка специфичности гуманизированных антител против IL-5 к цитокинам Th2
IL-5 представляет собой один из цитокинов Th2. Для подтверждения того, что антитела против IL-5 специфично нацелены только на IL-5 без перекрестной реактивности с другими цитокинами, в анализе Fortebio использовали 12 типов цитокинов Th2 и родственных цитокинов, включая IL2(R&D, 202-IL-010/CF), IL4(R&D, 204-IL-050/CF), IL-5(R&D, 205-IL-025/CF), IFNгамма, IL6(R&D, 7270-IL-025/CF), IL9(R&D, 209-IL-010/CF), IL10(R&D, 217-IL-025/CF), IL13(R&D, 213-ILB-025/CF), IL25(R&D, 8134-IL-025/CF), IL31(R&D, 2824-IL-010/CF), IL3, делящий α рецептор с IL-5 (203-IL-050/CF), и GMCSF (R&D, 215-GM-010/CF).
В частности, антитела захватывали с использованием биосенсора с белком A (PALL Fortebio, 18-5010), записывали сигнал захвата, затем добавляли 40 нМ каждого цитокина и записывали новые сигналы связывания. Наконец, сигнал связывания для IL-5 определяли как 100%, и наблюдали сигналы связывания между антителами и другими цитокинами. Результаты показаны на Фиг. 2.
Данные результаты показывают то, что среди 12 типов родственных цитокинов гуманизированные антитела против IL-5 h1705-008 и h1706-009 просто специфично связываются с IL-5 и не демонстрируют перекрестной реактивности с другими цитокинами Th2.
Фармакокинетическая оценка
Опытный пример 8: фармакокинетическая оценка гуманизированных антител против IL-5 у крыс
Экспериментальных крыс SD (предоставленных Sipple-BK Lab Animal Co., Ltd.), 18 самцов, делили на 6 групп с 3 крысами в каждой группе. Hu39D10, h1705-008 и h1706-009 вводили внутривенно и подкожно. Другим 9 крысам SD только внутривенно вводили h1773-007, h1779-014 или h1780-017. Для группы внутривенного введения отбирали 0,2 мл цельной крови без введения противосвертывающего средства до введения и через 5 мин, 8 ч, 1 сут., 2 сут., 4 сут., 7 сут., 10 сут., 14 сут., 21 сут. и 28 сут. после введения. Образцы крови помещали при 4°С на 30 мин, центрифугировали при 1000 g в течение 15 мин, супернатанты (сыворотку) помещали в пробирки ЕР и хранили при -80°С. Для группы подкожного введения цельную кровь отбирали до введения и через 1 ч, 2 ч, 4 ч, 8 ч, 1 сут., 2 сут., 4 сут., 7 сут., 10 сут., 14 сут., 21 сут. и 28 сут. после введения. Концентрацию антитела в сыворотке определяли посредством ELISA.
Результаты показывают то, что молекулы гуманизированного антитела по настоящему раскрытию имеют длительный период полувыведения и высокую биодоступность у крыс.
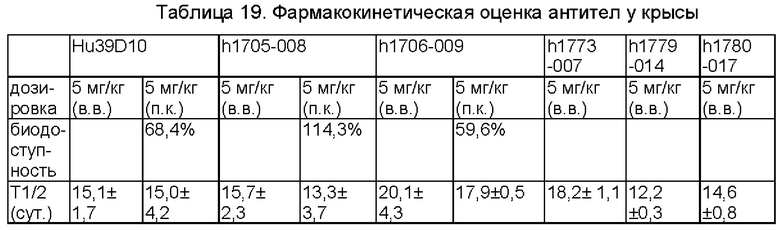
Биологическая оценка in vivo
Опытный пример 9: оценка эффективностей антител против IL-5 в мышиной модели астмы, индуцированной OVA
Данный опытный пример предназначен для того, чтобы оценить эффективности антител против IL-5 в модели индуцированной аэрозолем овальбумина (OVA) астмы у мышей BALB/c на основе воспаления дыхательных путей и ремоделирования дыхательных путей.
Мышей случайным образом делили на 7 групп согласно массе тела, 10 мышей в каждой группе: нормальная контрольная группа (G1); модельная группа (G2); группы обработки для антитела h1705-008 (G3 и G4 для двух дозировок 10 mpk и 2 mpk соответственно) и h1706-009 (G5 и G6 для двух дозировок 10 mpk и 2 mpk соответственно); и контрольная группа для позитивного антитела Hu39D10 (G7, 10 mpk). В сутки 1 и сутки 14 всех мышей сенсибилизировали внутрибрюшинной инъекцией сенсибилизирующего раствора. В сутки 28, 29 и 30 шесть групп мышей, исключая группу 1, стимулировали стимулирующим раствором на основе аэрозоля OVA в течение 30 минут. За два часа до стимуляции группе 2 (G2) внутрибрюшинно инъецировали фосфатный буфер, и мышам группы 3-7 (G3-G7) внутрибрюшинно инъецировали разные дозы разных антител, один раз в сутки в течение трех последующих суток. Свежие растворы антител, подлежащих анализу, готовили непосредственно перед каждой инъекцией, и все инъекции завершали в пределах получаса. В качестве нормального контроля, через 2 часа после внутрибрюшинной инъекции фосфатного буфера мышей в группе 1 стимулировали аэрозолем PBS в течение 30 минут, один раз в сутки в течение трех последовательных суток.
В сутки 31 животных анализировали на гиперчувствительность дыхательных путей с использованием системы WBP. Всем животным ингалировали спрей метахолина в дважды возрастающей концентрации: 1,5625; 3,125; 6,25; 12,5; 25, 50 мг/мл для определения повторяющегося значения усиленного выдоха при соответствующей концентрации.
В сутки 31, через 1 час после анализа ответа дыхательных путей с использованием системы WBP вставляли в трахею и фиксировали трахейную канюлю с диаметром 1,2 мм, дважды осуществляли отбор легочного лаважа, каждый раз использовали 0,8 мл фосфатного буфера, содержащего 1% BSA и 0,6 мМ EDTA. Записывали извлеченный объем жидкости лаважа.
BALF центрифугировали при 300 g при 4°С в течение 5 минут, и супернатант сохраняли для анализа цитокинов. После центрифугирования клетки ресуспендировали в 1,5 мл PBS (содержащего 1% BSA и 0,6 мМ EDTA) для подсчета клеток. Общее число клеток в BALF подсчитывали с использованием гемоцитометра и анализа окрашивания трипановым синим. Клетки размазывали, окрашивали красящим раствором Райта в течение одной минуты и затем окрашивали Giemsa в течение 7 минут для различения эозинофилов, нейтрофилов, макрофагов и лимфоцитов. Клетки подсчитывали под оптическим микроскопом.
После лаважа отбирали легочные ткани, погружали в 10%-ный нейтральный раствор формальдегида и затем фиксировали в 10%-ном нейтральном растворе формальдегида. Фиксированные ткани затем подвергали заливке в парафин, подрезке, окрашиванию Н & Е (гематоксилин и эозин) и бальной оценке. Результаты анализа показаны на Фиг. 3 и Фиг. 4А и 4В.
Данные результаты показывают то, что молекулы антитела h1705-008 и h1706-009 по настоящему раскрытию могут значительно улучшать функцию легкого дозозависимым образом, в то время как высокая доза (10 mpk) позитивного соединения не может улучшать функцию легкого (см. Фиг. 3). В то же самое время два данных антитела значимо снижали уровень эозинофилов и толщину слизистой, и демонстрировали более сильную способность уменьшать количество эозинофилов по сравнению с позитивным антителом в такой же дозировке (10 mpk) (см. Фиг. 4А и 4В). Посредством повторения эксперимента в таком же типе мышиной модели астмы авторы данного изобретения также подтвердили то, что 1 мг/мл всех из h1705-008, h1706-009 и h1780-017 значительно снижали уровень эозинофилов в BalF по сравнению с позитивным антителом (см. Фиг. 4С).
Опытный пример 10: оценка эффективности in vivo антител против IL-5 при индуцированной экзогенным человеческим IL-5 острой астме в модели морских свинок
В данном опытном примере самцов морских свинок использовали для создания модели острой астмы, индуцированной человеческим IL-5, и hu39D10 использовали в качестве позитивного антитела для оценки того, имеют ли пять гуманизированных моноклональных антител против IL-5 по настоящему раскрытию ингибирующие эффекты на индуцированное человеческим IL-5 увеличением уровня эозинофилов в жидкости бронхоальвеолярного лаважа (BALF) у морских свинок. Морских свинок делили на 9 групп, по 8-10 в каждой группе: нормальная контрольная группа, модельная группа, группа hu39D10 (1 мг/кг), группа h1705-008 (1 мг/кг), группа h1706-009 (1 мг/кг), группа h1780-017 (1 мг/кг), группа h1773-007 (1 мг/кг) и группа h1779-014 (1 мг/кг). В сутки 1 морским свинкам в модельной группе и группах с введением лекарственного средства внутритрахеально инъецировали 100 мкл человеческого IL-5 (содержащего 5 мкг антигена IL-5) для примирования, и группе нормального контроля внутритрахеально инъецировали PBS. Через 2 часа после примирования группам с введением лекарственного средства внутрибрюшинно вводили 1 мг/кг описанных выше моноклональных антител против IL5 (при объеме введения 5 мл/кг), соответствующее антитело IgG вводили модельной группе, и PBS внутрибрюшинно вводили группе нормального контроля. Через 24 часа после внутритрахеальной инъекции морских свинок анестезировали и извлекали жидкость бронхоальвеолярного лаважа. Концентрацию клеток доводили до 5×106/мл, и 15 мкл их суспензии капали на стеклянное предметное стекло, сушили и фиксировали для окрашивания НЕ, и общее число клеток, и число эозинофилов подсчитывали под микроскопом с 400× увеличением. Рассчитывали процентную долю эозинофилов. Результаты показаны на Фиг. 5А и 5В, указывая то, что все пять гуманизированных антител по настоящему раскрытию могут значимо уменьшать уровень эозинофилов в BALF.
Оценка стабильности
Опытный пример 11: стабильность гуманизированных антител против IL-5
1. Химическая стабильность антител
Модификация в виде деамидирования является обычной химической модификацией антител, которая может влиять на долговременную стабильность. В частности, высокой степени модификации в виде деамидирования, окисления или изомеризации части аминокислот в областях CDR обычно следует избегать, или следует ее снижать посредством мутации. 100 мкг образцы, отобранные в разные моменты времени, растворяли в 100 мкл раствора 0,2 М His-HCl, 8 М Gua-HCl, рН 6,0, добавляли 3 мкл 0,1 г/мл DTT (дитиотрейтол), инкубировали в водной бане при 50°С в течение 1 часа и затем дважды подвергали ультрафильтрации с 0,02 М His-HCl, рН 6,0. Добавляли 3 мкл 0,25 мг/мл трипсина, инкубировали в водной бане при 37°С в течение ночи для ферментативного расщепления. Анализ ЖХ-МС (жидкостная хроматография-масс-спектрометрия) проводили с использованием Agilent 6530 Q-TOF. Результаты показывают то, что химическая стабильность гуманизированных антител по настоящему раскрытию была хорошей, и данные антитела не демонстрировали ненормальной модификации после ускоряющей реакции в буферной системе 529 при 40°С в течение одного месяца.
2. Исследование по степени агрегации антител при условиях высокой концентрации
Стабильность опытных антител оценивали при высоких концентрациях в условиях разных буферных систем и разных температур в течение одного месяца. Стабильность исследовали при концентрации 50 мг/мл в трех буферных системах: РВ9, His и 529 при 40°С, 25°С, 4°С и -80°С с повторным замораживанием и оттаиванием. Степень агрегации отслеживали посредством SEC-HPLC. Использовали хроматограф Waters е2695 с колонкой Waters Xbridge ВЕН 200А SEC, и подвижной фазой был PBS (рН доводили до 6,8 разбавленной соляной кислотой). Загружали 50 мкг белка, и изократическую элюцию проводили при скорости тока 0,5 мл/мин. Наблюдали то, что ни одно из гуманизированных антител против IL-5 значительно не агрегировало в условиях высокой концентрации. После 4 недель ускоряющей реакции при 40°С чистота мономера антитела была больше, чем 95% во всех трех системах.
3. Исследование чистоты антител при условиях высокой концентрации
Стабильность анализируемых антител оценивали при высоких концентрациях и в условиях разных буферных систем и разной температуры в течение одного месяца. Стабильность исследовали при концентрации 50 мг/мл в системе His и при 40°С. Чистоту отслеживали посредством CE-SDS (капиллярный электрофорез с додецилсульфатом натрия). Отбирали 100 мкг белка, и добавляли буфер для образца с достижением 95 мкл. Для анализа в восстанавливающем режиме добавляли 5 мкл димеркаптоэтанола; для анализа в невосстанавливающем режиме добавляли 5 мкл IAA. Инкубировали в водной бане при 70°С в течение 10 мин, центрифугировали и загружали супернатант. Данные получали и анализировали с использованием прибора для электрофореза Beckman PA800plus. Наблюдали то, что белки антител имеют хорошую стабильность в отношении чистоты при условиях высокой кнцентрации, и после 28 суток ускоряющей реакции при 40°С анализ CE-SDS показал то, что главный пик антител уменьшался только на 2%.
4. Выявление термостабильности разных антител посредством UNIT
Образцы растворяли в соответствующем буфере (буфер PBS), и концентрацию данных образцов контролировали при примерно 1 мг/мл. Загружали 9 мкл. Установка параметров: исходная температура - 20°С; инкубация - 0 с; скорость нагревания - 0,3°С/мин; удерживание на плитке - 5 с; завершающая температура - 95°С. Выявляли значения Tm антител. Данные антитела имеют высокое значение Tm и демонстрируют хорошую термостабильность.
Вышеописанное изобретение было подробно описано с помощью сопровождающих графических материалов и примеров. Однако данное описание и примеры не следует истолковывать как ограничивающее объем данного раскрытия. Раскрытия всех патентов и научной литературы, процитированные в данном документе, прямо включаются посредством ссылки во всей их полноте.
--->
Перечень последовательностей
<110> JIANGSU HENGRUI MEDICINE CO., LTD.; SHANGHAI HENGRUI PHARMACEUTICAL CO., LTD.
<120> АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО
МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ
<130> 370337CG
<160> 82
<170> PatentIn version 3.3
<210> 1
<211> 140
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность человеческого IL-5 с his
меткой (rhIL-5-his)
<400> 1
Met Arg Met Leu Leu His Leu Ser Leu Leu Ala Leu Gly Ala Ala Tyr
1 5 10 15
Val Tyr Ala Ile Pro Thr Glu Ile Pro Thr Ser Ala Leu Val Lys Glu
20 25 30
Thr Leu Ala Leu Leu Ser Thr His Arg Thr Leu Leu Ile Ala Asn Glu
35 40 45
Thr Leu Arg Ile Pro Val Pro Val His Lys Asn His Gln Leu Cys Thr
50 55 60
Glu Glu Ile Phe Gln Gly Ile Gly Thr Leu Glu Ser Gln Thr Val Gln
65 70 75 80
Gly Gly Thr Val Glu Arg Leu Phe Lys Asn Leu Ser Leu Ile Lys Lys
85 90 95
Tyr Ile Asp Gly Gln Lys Lys Lys Cys Gly Glu Glu Arg Arg Arg Val
100 105 110
Asn Gln Phe Leu Asp Tyr Leu Gln Glu Phe Leu Gly Val Met Asn Thr
115 120 125
Glu Trp Ile Ile Glu Ser His His His His His His
130 135 140
<210> 2
<211> 140
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность IL-5 яванского макака с his
меткой
<400> 2
Met Arg Met Leu Leu His Leu Ser Leu Leu Ala Leu Gly Ala Ala Tyr
1 5 10 15
Val Tyr Ala Ile Pro Thr Glu Ile Pro Ala Ser Ala Leu Val Lys Glu
20 25 30
Thr Leu Ala Leu Leu Ser Thr His Arg Thr Leu Leu Ile Ala Asn Glu
35 40 45
Thr Leu Arg Ile Pro Val Pro Val His Lys Asn His Gln Leu Cys Thr
50 55 60
Glu Glu Ile Phe Gln Gly Ile Gly Thr Leu Glu Ser Gln Thr Val Gln
65 70 75 80
Gly Gly Thr Val Glu Arg Leu Phe Lys Asn Leu Ser Leu Ile Lys Lys
85 90 95
Tyr Ile Gly Gly Gln Lys Lys Lys Cys Gly Glu Glu Arg Arg Arg Val
100 105 110
Asn Gln Phe Leu Asp Tyr Leu Gln Glu Phe Leu Gly Val Met Asn Thr
115 120 125
Glu Trp Ile Ile Glu Ser His His His His His His
130 135 140
<210> 3
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность IL-5 мыши с his меткой
<400> 3
Met Glu Ile Pro Met Ser Thr Val Val Lys Glu Thr Leu Thr Gln Leu
1 5 10 15
Ser Ala His Arg Ala Leu Leu Thr Ser Asn Glu Thr Met Arg Leu Pro
20 25 30
Val Pro Thr His Lys Asn His Gln Leu Cys Ile Gly Glu Ile Phe Gln
35 40 45
Gly Leu Asp Ile Leu Lys Asn Gln Thr Val Arg Gly Gly Thr Val Glu
50 55 60
Met Leu Phe Gln Asn Leu Ser Leu Ile Lys Lys Tyr Ile Asp Arg Gln
65 70 75 80
Lys Glu Lys Cys Gly Glu Glu Arg Arg Arg Thr Arg Gln Phe Leu Asp
85 90 95
Tyr Leu Gln Glu Phe Leu Gly Val Met Ser Thr Glu Trp Ala Met Glu
100 105 110
Gly His His His His His His
115
<210> 4
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность IL-5 крысы с his меткой
<400> 4
Met Glu Ile Pro Met Ser Thr Val Val Lys Glu Thr Leu Ile Gln Leu
1 5 10 15
Ser Thr His Arg Ala Leu Leu Thr Ser Asn Glu Thr Met Arg Leu Pro
20 25 30
Val Pro Thr His Lys Asn His Gln Leu Cys Ile Gly Glu Ile Phe Gln
35 40 45
Gly Leu Asp Ile Leu Lys Asn Gln Thr Val Arg Gly Gly Thr Val Glu
50 55 60
Ile Leu Phe Gln Asn Leu Ser Leu Ile Lys Lys Tyr Ile Asp Gly Gln
65 70 75 80
Lys Glu Lys Cys Gly Glu Glu Arg Arg Lys Thr Arg His Phe Leu Asp
85 90 95
Tyr Leu Gln Glu Phe Leu Gly Val Met Ser Thr Glu Trp Ala Met Glu
100 105 110
Val His His His His His His
115
<210> 5
<211> 560
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> человеческий IL-5 альфа, слитый в человеческим фрагментом Fc
<400> 5
Asp Leu Leu Pro Asp Glu Lys Ile Ser Leu Leu Pro Pro Val Asn Phe
1 5 10 15
Thr Ile Lys Val Thr Gly Leu Ala Gln Val Leu Leu Gln Trp Lys Pro
20 25 30
Asn Pro Asp Gln Glu Gln Arg Asn Val Asn Leu Glu Tyr Gln Val Lys
35 40 45
Ile Asn Ala Pro Lys Glu Asp Asp Tyr Glu Thr Arg Ile Thr Glu Ser
50 55 60
Lys Cys Val Thr Ile Leu His Lys Gly Phe Ser Ala Ser Val Arg Thr
65 70 75 80
Ile Leu Gln Asn Asp His Ser Leu Leu Ala Ser Ser Trp Ala Ser Ala
85 90 95
Glu Leu His Ala Pro Pro Gly Ser Pro Gly Thr Ser Ile Val Asn Leu
100 105 110
Thr Cys Thr Thr Asn Thr Thr Glu Asp Asn Tyr Ser Arg Leu Arg Ser
115 120 125
Tyr Gln Val Ser Leu His Cys Thr Trp Leu Val Gly Thr Asp Ala Pro
130 135 140
Glu Asp Thr Gln Tyr Phe Leu Tyr Tyr Arg Tyr Gly Ser Trp Thr Glu
145 150 155 160
Glu Cys Gln Glu Tyr Ser Lys Asp Thr Leu Gly Arg Asn Ile Ala Cys
165 170 175
Trp Phe Pro Arg Thr Phe Ile Leu Ser Lys Gly Arg Asp Trp Leu Ala
180 185 190
Val Leu Val Asn Gly Ser Ser Lys His Ser Ala Ile Arg Pro Phe Asp
195 200 205
Gln Leu Phe Ala Leu His Ala Ile Asp Gln Ile Asn Pro Pro Leu Asn
210 215 220
Val Thr Ala Glu Ile Glu Gly Thr Arg Leu Ser Ile Gln Trp Glu Lys
225 230 235 240
Pro Val Ser Ala Phe Pro Ile His Cys Phe Asp Tyr Glu Val Lys Ile
245 250 255
His Asn Thr Arg Asn Gly Tyr Leu Gln Ile Glu Lys Leu Met Thr Asn
260 265 270
Ala Phe Ile Ser Ile Ile Asp Asp Leu Ser Lys Tyr Asp Val Gln Val
275 280 285
Arg Ala Ala Val Ser Ser Met Cys Arg Glu Ala Gly Leu Trp Ser Glu
290 295 300
Trp Ser Gln Pro Ile Tyr Val Gly Asn Asp Glu His Lys Pro Leu Arg
305 310 315 320
Glu Trp Ile Glu Gly Arg Met Asp Glu Pro Lys Ser Cys Asp Lys Thr
325 330 335
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
340 345 350
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
355 360 365
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
370 375 380
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
385 390 395 400
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
405 410 415
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
420 425 430
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
435 440 445
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
450 455 460
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
465 470 475 480
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
485 490 495
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
500 505 510
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
515 520 525
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
530 535 540
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
545 550 555 560
<210> 6
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области тяжелой цепи мышиного
mAb1705
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu Glu Trp Val
35 40 45
Thr Ser Ile Ser Tyr Glu Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Ser Gln Thr Leu Arg Glu Ser Phe Asp Tyr Trp Gly Gln Gly Val
100 105 110
Met Val Thr Val Ser Ser
115
<210> 7
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области легкой цепи мышиного
mAb1705
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Met Ser Val Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Ile Ala Arg Ser Pro Lys Leu Val Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Asn Thr Leu Glu Ser
65 70 75 80
Glu Asp Thr Gly Ile Tyr Phe Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Arg Leu Glu Leu Lys
100 105
<210> 8
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области тяжелой цепи мышиного
mAb1706
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu Glu Trp Val
35 40 45
Thr Ser Ile Asn Tyr Glu Gly Asn Ser Ala Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Thr Glu Thr Leu Arg Glu Ser Leu Asp Tyr Trp Gly Gln Gly Val
100 105 110
Met Val Thr Val Ser Ser
115
<210> 9
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области легкой цепи мышиного
mAb1706
<400> 9
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Met Ser Val Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Leu Gly Lys Ser Pro Lys Leu Met Ile
35 40 45
His Ser Ala Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Asn Thr Leu Glu Ser
65 70 75 80
Glu Asp Pro Gly Ile Tyr Phe Cys Leu Gln His Lys Gln Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 10
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области тяжелой цепи мышиного
mAb1780
<400> 10
Gln Val Lys Leu Leu Gln Ser Gly Ala Ala Leu Val Lys Pro Gly Asp
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Asp Tyr Thr Phe Thr Glu Tyr
20 25 30
Leu Ile His Trp Val Lys Gln Ser Gln Gly Arg Ser Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Gly Thr Val Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Lys Ala Thr Leu Thr Val Asp Lys Phe Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Arg Arg Leu Thr Phe Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Ser Asp Pro Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Val Met Val Thr Val Ser Ser
115
<210> 11
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области легкой цепи мышиного
mAb1780
<400> 11
Asp Thr Val Leu Thr Gln Ser Pro Ala Leu Ala Val Ser Pro Gly Glu
1 5 10 15
Arg Val Ser Ile Ser Cys Arg Ala Ser Glu Gly Leu Thr Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Gln Pro Lys Leu Leu Ile Tyr
35 40 45
Lys Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asp Pro Val Glu Ala Asp
65 70 75 80
Asp Ala Ala Thr Tyr Phe Cys Gln Gln Asn Trp Asn Asp Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 12
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области тяжелой цепи мышиного
mAb1773
<400> 12
Glu Val Gln Leu Gln Gln Ser Leu Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asp Thr Lys His Gly Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Leu Gln Phe Ser Ser Leu Thr Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Phe Arg Tyr Gly Ile Tyr Pro Asp His Trp Gly Gln Gly Thr Pro Leu
100 105 110
Thr Val Ser Ser
115
<210> 13
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области легкой цепи мышиного
mAb1773
<400> 13
Gln Ile Val Leu Thr Gln Ser Pro Ala Leu Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Arg Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Leu Thr Ala Thr Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Phe Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Asn Ser Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Glu
100 105
<210> 14
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области тяжелой цепи мышиного
mAb1779
<400> 14
Gln Val Lys Leu Leu Gln Ser Gly Ala Ala Leu Val Lys Pro Gly Asp
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ile Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Arg Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Leu Glu Phe Ser Arg Val Thr Ser Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Gly Arg Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe Trp Gly Pro
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 15
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области легкой цепи мышиного
mAb1779
<400> 15
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Thr Val Ser Ile Glu Cys Leu Ala Ser Glu Gly Ile Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Ser Gly Arg Ser Pro Gln Leu Leu Val
35 40 45
Tyr Ala Ala Ser Arg Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Arg Tyr Phe Phe Lys Ile Ser Gly Met Gln Pro
65 70 75 80
Glu Asp Glu Ala Asp Tyr Phe Cys Gln Gln Gly Tyr Lys Thr Pro Leu
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 16
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR1 mAb1705
<400> 16
His Tyr Tyr Met Ala
1 5
<210> 17
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR2 mAb1705
<400> 17
Ser Ile Ser Tyr Glu Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val Lys
1 5 10 15
Gly
<210> 18
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR3 mAb1705
<400> 18
Gln Thr Leu Arg Glu Ser Phe Asp Tyr
1 5
<210> 19
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR1 mAb1705
<400> 19
Arg Ala Ser Gln Asp Ile Ala Asn Tyr Leu Ser
1 5 10
<210> 20
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR2 mAb1705
<400> 20
Gly Thr Ser Asn Leu Glu Val
1 5
<210> 21
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR3 mAb1705
<400> 21
Leu Gln Asp Lys Glu Phe Pro Arg Thr
1 5
<210> 22
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR1 mAb1706
<400> 22
His Tyr Tyr Met Ala
1 5
<210> 23
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR2 mAb1706
<400> 23
Ser Ile Asn Tyr Glu Gly Asn Ser Ala Tyr Tyr Gly Asp Ser Val Lys
1 5 10 15
Gly
<210> 24
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR3 mAb1706
<400> 24
Glu Thr Leu Arg Glu Ser Leu Asp Tyr
1 5
<210> 25
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR1 mAb1706
<400> 25
Arg Ala Ser Gln Asp Ile Gly Asn Tyr Leu Ser
1 5 10
<210> 26
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR2 mAb1706
<400> 26
Ser Ala Ser Asn Leu Glu Val
1 5
<210> 27
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR3 mAb1706
<400> 27
Leu Gln His Lys Gln Phe Pro Arg Thr
1 5
<210> 28
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR1 mAb1780
<400> 28
Glu Tyr Leu Ile His
1 5
<210> 29
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR2 mAb1780
<400> 29
Tyr Ile Asn Pro Tyr Ser Gly Gly Thr Val Tyr Asn Glu Lys Phe Lys
1 5 10 15
Ser
<210> 30
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR3 mAb1780
<400> 30
Asp Gly Gly Tyr Ser Asp Pro Leu Asp Tyr
1 5 10
<210> 31
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR1 mAb1780
<400> 31
Arg Ala Ser Glu Gly Leu Thr Ser Tyr Met His
1 5 10
<210> 32
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR2 mAb1780
<400> 32
Lys Ala Ser Asn Leu Ala Ser
1 5
<210> 33
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR3 mAb1780
<400> 33
Gln Gln Asn Trp Asn Asp Pro Trp Thr
1 5
<210> 34
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR1 mAb1773
<400> 34
Asn Thr Tyr Ile His
1 5
<210> 35
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR2 mAb1773
<400> 35
Arg Ile Asp Pro Ala Asn Gly Asp Thr Lys His Gly Pro Lys Phe Gln
1 5 10 15
Gly
<210> 36
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR3 mAb1773
<400> 36
Tyr Gly Ile Tyr Pro Asp His
1 5
<210> 37
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR1 mAb1773
<400> 37
Ser Ala Ser Ser Ser Val Asn Tyr Ile Tyr
1 5 10
<210> 38
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR2 mAb1773
<400> 38
Leu Thr Ala Thr Leu Ala Ser
1 5
<210> 39
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR3 mAb1773
<400> 39
Gln Gln Trp Asn Ser Tyr Pro Tyr Thr
1 5
<210> 40
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR1 mAb1779
<400> 40
Asp Tyr Ile Ile His
1 5
<210> 41
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR2 mAb1779
<400> 41
Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe Lys
1 5 10 15
Arg
<210> 42
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> HCDR3 mAb1779
<400> 42
Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe
1 5 10
<210> 43
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR1 mAb1779
<400> 43
Leu Ala Ser Glu Gly Ile Ser Asn Asp Val Ala
1 5 10
<210> 44
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR2 mAb1779
<400> 44
Ala Ala Ser Arg Leu Gln Asp
1 5
<210> 45
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> LCDR3 mAb1779
<400> 45
Gln Gln Gly Tyr Lys Thr Pro Leu Thr
1 5
<210> 46
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1705_VL.1
<400> 46
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 47
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1705_VL.1A
<400> 47
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 48
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1705_VL.1B
<400> 48
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Val Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 49
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1705_VH.1
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Tyr Glu Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gln Thr Leu Arg Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 50
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1705_VH.1A
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Tyr Glu Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Gln Thr Leu Arg Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 51
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1705_VH.1B
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Thr Ser Ile Ser Tyr Glu Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Ser Gln Thr Leu Arg Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 52
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Константная область тяжелой цепи IgG1
<400> 52
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 53
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Константная область легкой цепи каппа
<400> 53
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 54
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1706_VL.1
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Lys Gln Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 55
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1706_VL.1A
<400> 55
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Lys Gln Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 56
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1706_VL.1B
<400> 56
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Met Ile
35 40 45
Tyr Ser Ala Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Leu Gln His Lys Gln Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 57
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1706_VH.1
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asn Tyr Glu Gly Asn Ser Ala Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Thr Leu Arg Glu Ser Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 58
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1706_VH.1A
<400> 58
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asn Tyr Glu Gly Asn Ser Ala Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Glu Thr Leu Arg Glu Ser Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 59
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1706_VH.1B
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Thr Ser Ile Asn Tyr Glu Gly Asn Ser Ala Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Thr Glu Thr Leu Arg Glu Ser Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 60
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VL.1
<400> 60
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Gly Leu Thr Ser Tyr
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asn Trp Asn Asp Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 61
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VL.1A
<400> 61
Asp Thr Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Gly Leu Thr Ser Tyr
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asn Trp Asn Asp Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 62
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VL.1B
<400> 62
Asp Thr Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Gly Leu Thr Ser Tyr
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Asn Trp Asn Asp Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 63
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VH.1
<400> 63
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Leu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Gly Thr Val Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Ser Asp Pro Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 64
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VH.1A
<400> 64
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Leu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Gly Thr Val Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Ser Asp Pro Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 65
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VH.1B
<400> 65
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Leu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Gly Thr Val Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Ala Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Ser Asp Pro Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 66
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1780_VH.1C
<400> 66
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Leu Ile His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Gly Thr Val Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Lys Ala Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Ser Asp Pro Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 67
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1773_VL.1
<400> 67
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Leu Thr Ala Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Asn Ser Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 68
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1773_VL.1A
<400> 68
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Leu Thr Ala Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Asn Ser Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 69
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1773_VH.1
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Ser Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Asp Pro Ala Val Gly Asp Thr Lys His Gly Pro Lys Phe
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Tyr Gly Ile Tyr Pro Asp His Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 70
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1773_VH.1A
<400> 70
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Ile Ser Asn Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Ser Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Asp Pro Ala Val Gly Asp Thr Lys His Gly Pro Lys Phe
50 55 60
Gln Gly Arg Phe Thr Ile Ser Ala Asp Asp Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Phe Arg Tyr Gly Ile Tyr Pro Asp His Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 71
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1773_VH.1B
<400> 71
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Ile Ser Asn Thr
20 25 30
Tyr Ile His Trp Val Lys Gln Ala Ser Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Val Gly Asp Thr Lys His Gly Pro Lys Phe
50 55 60
Gln Gly Arg Phe Thr Ile Ser Ala Asp Asp Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Phe Arg Tyr Gly Ile Tyr Pro Asp His Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 72
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VL.1
<400> 72
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Gly Ile Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Arg Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Lys Thr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 73
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VL.1A
<400> 73
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Gly Ile Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Arg Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Lys Thr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 74
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VL.1B
<400> 74
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Gly Ile Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Val
35 40 45
Tyr Ala Ala Ser Arg Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Lys Thr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 75
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VH.1
<400> 75
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ile Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Arg Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 76
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VH.1A
<400> 76
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ile Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Arg Arg Val Thr Met Thr Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 77
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VH.1B
<400> 77
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ile Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Arg Arg Ala Thr Met Thr Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 78
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VH.1C
<400> 78
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ile Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Arg Arg Ala Thr Met Thr Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Glu Phe Ser Arg Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 79
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> h1779_VH.1D
<400> 79
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ile Ile His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Gly Ser Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Arg Lys Ala Thr Met Thr Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Glu Phe Ser Arg Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ile Ala Trp Asp His Trp Tyr Phe Asp Phe Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 80
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Последовательность тяжелой цепи Hu39D10
<400> 80
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Ser Leu Thr Ser Asn
20 25 30
Ser Val Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Trp Ser Asn Gly Asp Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Ser Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Tyr Tyr Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 81
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Последовательность легкой цепи Hu39D10
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Asn Ser Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Ala Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Lys Phe Pro Asn
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Val Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 82
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариант HCDR2 тяжелой цепи антитела h1773
<400> 82
Arg Ile Asp Pro Ala Val Gly Asp Thr Lys His Gly Pro Lys Phe Gln
1 5 10 15
Gly
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ КЛАУДИНА 18.2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2822550C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| АНТИТЕЛО К PD-1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2807484C2 |
| АНТИТЕЛО К CD3 И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2802272C2 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО ПРОТИВ IL-5, И ЕЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2824390C2 |
| АНТИТЕЛО ПРОТИВ БЕТА-АМИЛОИДА, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2777844C1 |
| АНТИТЕЛО К PD-L1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2778085C2 |
| АНТИТЕЛО К CD40, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2779128C2 |
| АНТИТЕЛО, СВЯЗЫВАЮЩЕЕ ЧЕЛОВЕЧЕСКИЙ IL-4R, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2779649C1 |
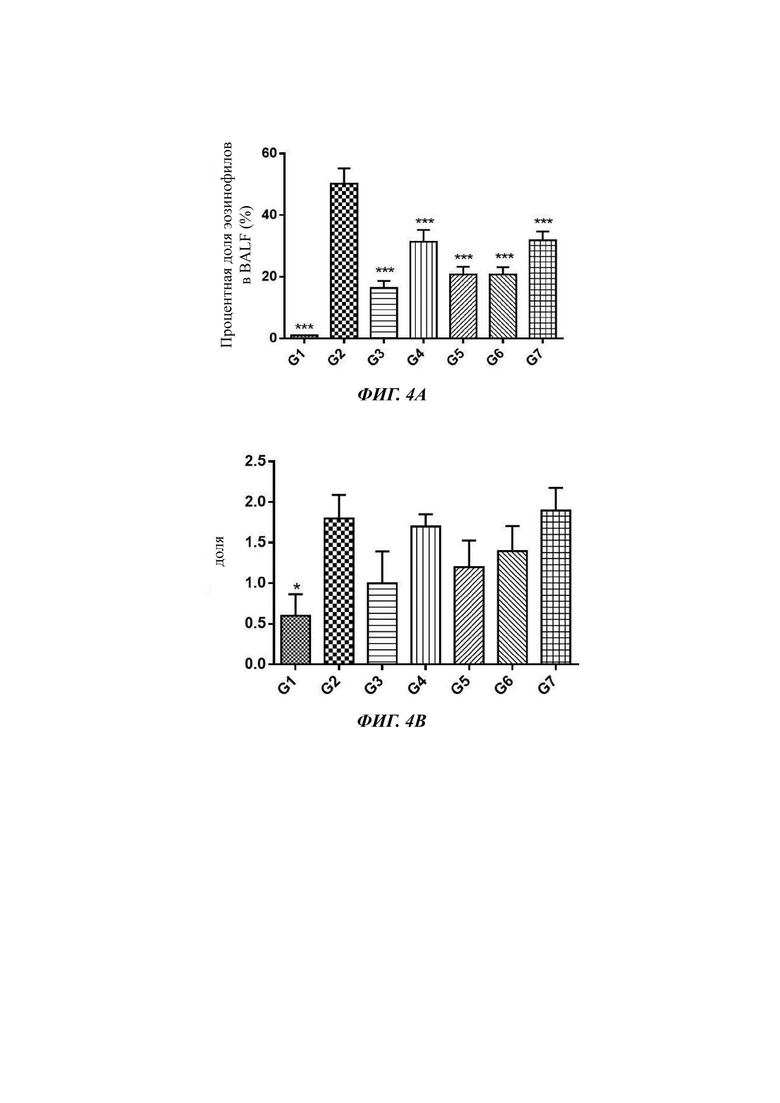
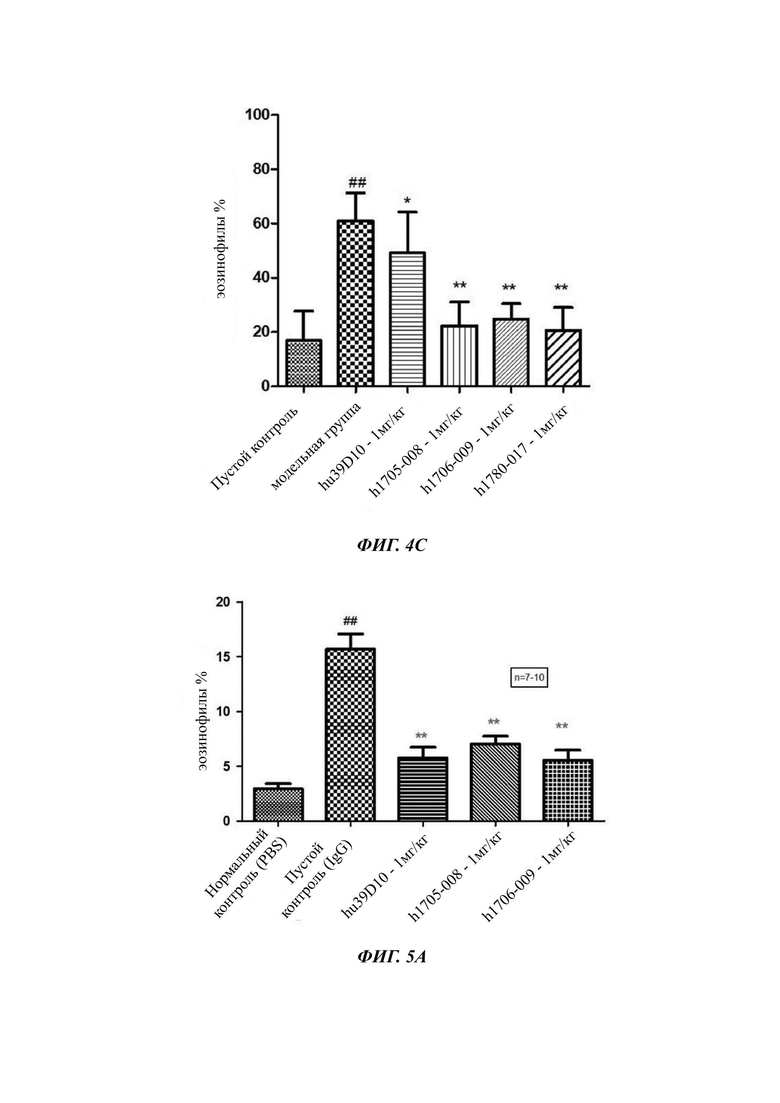
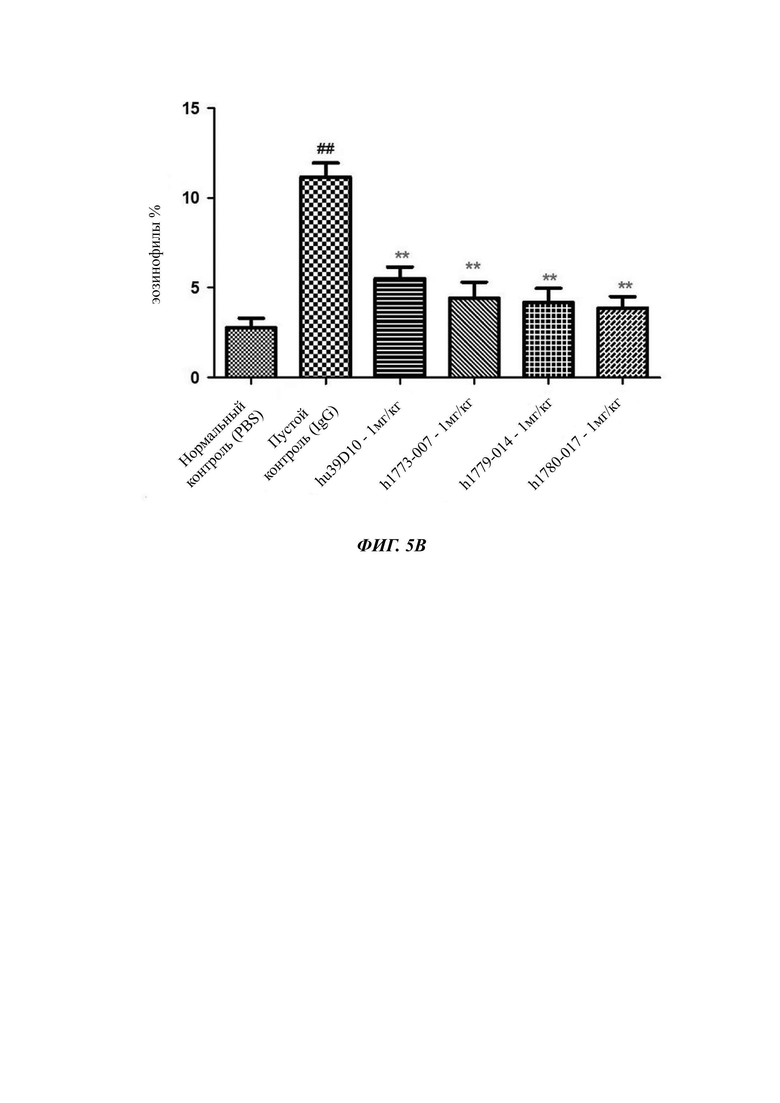
Изобретение относится к области биотехнологии и медицины, в частности к моноклональному антителу или его фрагменту против IL-5; к фармацевтической композиции для лечения заболевания, ассоциированного с человеческим IL-5; к выделенной молекуле нуклеиновой кислоты, кодирующей моноклональное антитело или его антигенсвязывающий фрагмент против IL-5; к рекомбинантному экспрессионному вектору, содержащему вышеупомянутую молекулу нуклеиновой кислоты; к экспрессирующей клетке-хозяину, трансформированной вышеупомянутым вектором; к способу получения моноклонального антитела или его антигенсвязывающего фрагмента против IL-5; к способу и средству выявления или определения человеческого IL-5; к способу лечения заболевания, ассоциированного с человеческим IL-5; а также к применению вышеупомянутого антитела или его антигенсвязывающего фрагмента, фармацевтической композиции, выделенной молекулы нуклеиновой кислоты, или их комбинации в получении лекарственного средства для предупреждения или лечения заболевания, ассоциированного с человеческим IL-5. 10 н. и 10 з.п. ф-лы, 5 ил., 19 табл., 6 пр.
1. Моноклональное антитело или его антигенсвязывающий фрагмент, специфично связывающееся с человеческим интерлейкином-5 (IL-5), где указанное моноклональное антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(i) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 16-18, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 19-21, соответственно; или
(ii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 22-24, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 25-27, соответственно; или
(iii) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 28-30, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 31-33, соответственно; или
(iv) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 34-36, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 37-39, соответственно; или
(v) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 40-42, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 43-45, соответственно; или
(vi) вариабельная область тяжелой цепи содержит область HCDR1, HCDR2 и HCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 34, 82 и 36, соответственно; и вариабельная область легкой цепи содержит область LCDR1, LCDR2 и LCDR3, как определено в аминокислотных последовательностях SEQ ID NO: 37-39, соответственно.
2. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где указанное моноклональное антитело представляет собой рекомбинантное антитело, предпочтительно выбранное из группы, состоящей из мышиного антитела, химерного антитела и гуманизированного антитела.
3. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 2, где указанное гуманизированное антитело содержит вариабельную область тяжелой цепи SEQ ID NO: 49, 57, 63, 69 или 75, или ее вариант, при этом указанный вариант имеет 1-10 мутаций аминокислот на вариабельной области тяжелой цепи, как определено в SEQ ID NO: 49, 57, 63, 69 или 75.
4. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 3, где указанный вариант имеет 1-10 обратных мутаций аминокислот на области FR (каркас) вариабельной области тяжелой цепи, как определено в SEQ ID NO: 49, 57, 63, 69 или 75, при этом, предпочтительно, указанная обратная мутация выбрана из группы, состоящей из S49T, V93T и K98S или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 49, или указанная обратная мутация выбрана из группы, состоящей из S49T, V93T и K98T или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 57, или указанная обратная мутация выбрана из группы, состоящей из R38K, M48I, R67K, V68A, M70L, R72V, T74K и L83F или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 63, или указанная обратная мутация выбрана из группы, состоящей из F29I, R38K, V48I, R72A, T97F на вариабельной области тяжелой цепи SEQ ID NO: 69 и N55V на CDR или их комбинации, или указанная обратная мутация выбрана из группы, состоящей из R38K, M48I, R67K, V68A, R72А, T74K, М81L, L83F и D89E или их комбинации на вариабельной области тяжелой цепи SEQ ID NO: 75.
5. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 4, где указанное гуманизированное антитело содержит вариабельную область тяжелой цепи SEQ ID NO: 50 или 51, или содержит вариабельную область тяжелой цепи SEQ ID NO: 58 или 59, или содержит вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 64, 65 и 66, или содержит вариабельную область тяжелой цепи SEQ ID NO: 70 или 71, или содержит вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 76-79.
6. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 2, где гуманизированное антитело содержит вариабельную область легкой цепи SEQ ID NO: 46, 54, 60, 67 или 72, или ее вариант, при этом указанный вариант имеет 1-10 замен аминокислот на вариабельной области легкой цепи, как определено в SEQ ID NO: 46, 54, 60, 67 или 72.
7. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 6, где указанный вариант имеет 1-10 обратных мутаций аминокислот на области FR вариабельной области легкой цепи, как определено в SEQ ID NO: 46, 54, 60, 67 или 72, при этом, предпочтительно, указанная обратная мутация выбрана из группы, состоящей из A43S, L47V, G66R, T69S, F71Y и Y87F или их комбинации на вариабельной области легкой цепи SEQ ID NO: 46, или указанная обратная мутация выбрана из группы, состоящей из A43S, L47M, F71Y и Y87F или их комбинации на вариабельной области легкой цепи SEQ ID NO: 54, или указанная обратная мутация выбрана из группы, состоящей из E1D, I2T, I57V, V84T и Y86F или их комбинации на вариабельной области легкой цепи SEQ ID NO: 60, или указанная обратная мутация выбрана из группы, состоящей из M4L, A42S, L45P и L46W или их комбинации на вариабельной области легкой цепи SEQ ID NO: 67, или указанная обратная мутация выбрана из группы, состоящей из A43S, I48V и F71Y или их комбинации на вариабельной области легкой цепи SEQ ID NO: 72.
8. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 7, где указанное гуманизированное антитело содержит вариабельную область легкой цепи SEQ ID NO: 47 или 48 или содержит вариабельную область легкой цепи SEQ ID NO: 55 или 56, или содержит вариабельную область легкой цепи SEQ ID NO: 61 или 62, или содержит вариабельную область легкой цепи SEQ ID NO: 68; или содержит вариабельную область легкой цепи SEQ ID NO: 73 или 74.
9. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-8, где указанное гуманизированное антитело содержит:
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 49-51, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 46-48; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 57-59, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 54-56; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 63-66, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 60-62; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 69-71, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 67-68; или
вариабельную область тяжелой цепи, выбранную из любой из SEQ ID NO: 75-79, и вариабельную область легкой цепи, выбранную из любой из SEQ ID NO: 72-74.
10. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-9, где указанное антитело представляет собой полноразмерное антитело и дополнительно содержит константную область человеческого антитела, при этом, предпочтительно, указанное полноразмерное антитело содержит константную область тяжелой цепи человеческого антитела, как определено в SEQ ID NO: 52, и константную область человеческой легкой цепи, как определено в SEQ ID NO: 53.
11. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-9, где указанный антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, Fab', F(ab')2, одноцепочечного антитела (scFv), димеризованной области V (диатело), области V, стабилизированной дисульфидом (dsFv), и пептида, содержащего CDR.
12. Фармацевтическая композиция для лечения заболевания, ассоциированного с человеческим IL-5, содержащая терапевтически эффективное количество моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-11 и один или более фармацевтически приемлемых носителей, разбавителей, буферов или эксципиентов.
13. Выделенная молекула нуклеиновой кислоты, кодирующая моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-11.
14. Рекомбинантный экспрессионный вектор, содержащий выделенную молекулу нуклеиновой кислоты по п. 13.
15. Экспрессирующая клетка-хозяин, трансформированная рекомбинантным экспрессионным вектором по п. 14, при этом указанная клетка-хозяин выбрана из группы, состоящей из прокариотических клеток и эукариотических клеток, предпочтительно эукариотических клеток, более предпочтительно клеток млекопитающих.
16. Способ получения моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-11, включающий культивирование клетки-хозяина по п. 15 в культуральной среде с образованием и накоплением моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-11, и выделение моноклонального антитела или его антигенсвязывающего фрагмента из культуры.
17. Способ выявления или определения человеческого IL-5, включающий применение моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-11.
18. Средство для выявления или определения человеческого IL-5, которое содержит моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-11.
19. Способ лечения заболевания, ассоциированного с человеческим IL-5, причем указанный способ включает введение субъекту терапевтически эффективного количества моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-11, или фармацевтической композиции по п. 12, или выделенной молекулы нуклеиновой кислоты по п. 13 для лечения заболевания, ассоциированного с человеческим IL-5, причем указанное заболевание предпочтительно выбрано из группы, состоящей из астмы, злокачественного приступа астмы, хронической пневмонии, аллергического ринита, аллергического бронхолегочного аспергиллеза, эозинофилии, синдрома Чарга-Стросса, атопического дерматита, онхоциркозного дерматита, эпизодического ангиоотека, синдрома эозинофильной миалгии, эозинофильного гастроэнтерита, инфекции гельминтами, болезни Ходжкина, полипов носа, синдрома Леффлера, уртикарии, эозинофильного гиперпластического бронхита, узелкового артериита, синусита, эозинофильного эзофагита, аллергического эозинофильного эзофагита, аллергического конъюнктивита, онхоциркозного дерматита, эндометриоза и зависимого от стероидов эозинофильного бронхита.
20. Применение моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-11, или фармацевтической композиции по п. 12, или выделенной молекулы нуклеиновой кислоты по п. 13, или их комбинации в получении лекарственного средства для предупреждения или лечения заболевания или состояния, где указанное заболевание или состояние представляет собой заболевание, ассоциированное с человеческим IL-5, причем указанное заболевание или состояние предпочтительно выбрано из группы, состоящей из астмы, злокачественного приступа астмы, хронической пневмонии, аллергического ринита, аллергического бронхолегочного аспергиллеза, эозинофилии, синдрома Чарга-Стросса, атопического дерматита, онхоциркозного дерматита, эпизодического ангиоотека, синдрома эозинофильной миалгии, эозинофильного гастроэнтерита, инфекции гельминтами, болезни Ходжкина, полипов носа, синдрома Леффлера, уртикарии, эозинофильного гиперпластического бронхита, узелкового артериита, синусита, эозинофильного эзофагита, аллергического эозинофильного эзофагита, аллергического конъюнктивита, онхоциркозного дерматита, эндометриоза и зависимого от стероидов эозинофильного бронхита.
| WO 017033121 A1, 02.03.2017 | |||
| RU2019123112 A, 26.01.2021 | |||
| CN0001175263 A, 04.03.1998 | |||
| Fala L | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| American health & drug benefits, 2016 | |||
| Vol | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Светоэлектрический измеритель длин и площадей | 1919 |
|
SU106A1 |

