ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к антителу и его антигенсвязывающему фрагменту, которые специфически связываются с IL-17A. Настоящее изобретение, в частности, относится к антителам и к их антигенсвязывающим фрагментам, которые ингибируют IL-17А-опосредованную биологическую активность, и к композициям, содержащим антитела или их антигенсвязывающие фрагменты, а также к способам лечения соответствующих заболеваний. Настоящее изобретение, в частности, относится к применению антител против IL-17А и их антигенсвязывающих фрагментов в лечении иммунных патологических заболеваний, включающих аутоиммунные и воспалительные заболевания, такие как ревматоидный артрит, псориаз, анкилозирующий спондилит, рассеянный склероз, системная красная волчанка (SLE), волчаночный нефрит или хроническая обструктивная болезнь легких, астма, инфекционная гранулема, муковисцидоз или рак.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Интерлейкин 17 (IL-17), также известный как CTLA-8 или IL-17A, играет ключевую роль в иммунной системе. Семейство IL-17 включает шесть представителей, т.е. IL-17A, IL-17B, IL-17C, IL-17D, IL-17E и IL-17F, все из которых содержат 4 высококонсервативных остатка цистеина, которые являются очень важными. Однако биологические эффекты этих представителей значительно различаются. Из них, IL-17A и IL-17F имеют наиболее близкую гомологию и биологические функции и в настоящее время наиболее глубоко изучаются. IL-17A, экспрессируемый in vivo, имеет N-концевой сигнальный пептид, состоящий из 23 аминокислот, который отщепляется с образованием зрелого IL-17A. Зрелый IL-17A связан дисульфидной связью и обычно секретируется и присутствует в виде гомодимера. Иногда он также связывается с IL-17F, образуя гетеродимер IL-17AF. В общем случае, IL-17A или IL-17 относится к белку гомодимеру IL-17A, который продуцируется преимущественно хелперными Т-клетками 17 (Т-хелперами 17 или Th17), а также может синтезироваться и секретироваться другими иммунными клетками, такими как γδТ-клетки, клетки-индикаторы лимфоидной ткани (LTi), врожденные лимфоидные клетки (ILC) и естественные киллерные Т-клетки (NKT) (Cua DJ, Tato СМ., Nature reviews. 2010, 10:479-489). Регуляция экспрессии IL-17А является очень сложной. Было обнаружено, что, когда цитокины, такие как IL-6, IL-1β и TGFβ, индуцируют дифференцировку исходных CD4+ Т-клеток в Th17, Th17-клетки секретируют небольшое количество IL-17A из-за их слабой стабильности, и оказывают слабый эффект повреждения тканей. IL-23, когда он присутствует, вызывает вспышку воспаления и повреждение тканей, способствуя стабильности клеток Th17 для их непрерывной секреции IL-17A, увеличение экспрессии провоспалительных факторов (IL-22, CSF-2 и IFN-γ) и снижение экспрессии противовоспалительных факторов (IL-2, IL-27 и IL-12) и другим образом (McGeachy MJ, et al., Nature immunology. 2009, 10:314-324). Таким образом, путь IL-17A играет критическую роль в повреждении тканей, при аномальной экспрессии IL-23 в тканях.
IL-17 обычно секретируется в определенном месте и действует в местных тканях путем связывания с рецептором IL-17 (IL-17R) на поверхности клеток-мишеней. Семейство IL-17R включает пять представителей, то есть IL-17RA, IL-17RB, IL-17RC, IL-17RD и IL-17RE, которые широко экспрессируются на различных клеточных мембранах (Iwakura Y, et al., Immunity. 2011; 34:149-162). IL-17 в основном оказывает эффекты, связываясь с комплексом IL-17RA/IL-17RC на поверхности клеток негемопоэтического происхождения (таких как эпителиальные клетки и мезенхимальные клетки) (Ishigame Н, et al., Immunity. 2009; 30:108-119), и повреждают ткани, стимулируя секрецию клетками цитокинов (таких как IL-6, G-CSF, GM-CSF, IL-10, TGF-β и TNF-α), хемокинов (включая IL-8, CXCL1 и МСР-1) и простагландина (например PGE2), индуцируя агрегацию нейтрофилов и макрофагов и высвобождая активные формы кислорода (ROS) (Stark MA, et al., Immunity. 2005; 22:285-294).
Аутоиммунные заболевания, такие как псориаз, ревматоидный артрит, анкилозирующий спондилит, болезнь Крона и рассеянный склероз, представляют серьезную угрозу для здоровья человека. Установлено, что диссекретоз IL-17 тесно связан с возникновением и развитием таких заболеваний. Антитела, нацеленные на IL-17, эффективны в облегчении симптомов аутоиммунных заболеваний путем ингибирования сигнального пути IL-17-IL-17R (Sarah L, et al., Nat Rev Immunol. 2014, 14(9): 585-600). Cosentyx (секукинумаб), разработанный компанией Novartis, является первым в мире моноклональным антителом к IL-17. Его одобренные показания включают бляшечный псориаз в степени от средней до тяжелой, обеспечивая важный вариант биологического лечения первой линии для обширной группы больных псориазом. Однако крайне актуально и важно разработать антитела против IL-17 с разными свойствами, такими как разные структуры, лучшая эффективность и более широкие диапазоны показаний для лечения аутоиммунных заболеваний, таких как псориаз, ревматоидный артрит и анкилозирующий спондилит, а также других заболеваний, связанных с IL-17.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложено антитело или его антигенсвязывающий фрагмент, которые специфически связываются с человеческим IL-17A, содержащий по меньшей мере одну последовательность области, определяющей комплементарность (CDR), выбранную из SEQ ID NO: 1-24 и 60-65.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытый в настоящем документе, содержит по меньшей мере один домен CDR тяжелой цепи, выбранный из SEQ ID NO: 1-3, 4-6, 7-9, 10-12 и 60-62, и/или по меньшей мере один домен CDR легкой цепи, выбранный из SEQ ID NO: 13-15, 16-18, 19-21, 22-24 и 63-65.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые здесь, содержат вариабельную область тяжелой цепи (VH), где VH содержит HCDR1, выбранный из SEQ ID NO: 1, 4, 7, 10 и 60, HCDR2, выбранный из SEQ ID NO: 2, 5, 8, 11 и 61, и HCDR3, выбранный из SEQ ID NO: 3, 6, 9, 12 и 62.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые здесь, содержат вариабельную область тяжелой цепи (VH), где VH содержит совокупность аминокислотных последовательностей HCDR1, HCDR2 и HCDR3, выбранных из любой совокупности А-Е:


В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытый здесь, содержат вариабельную область легкой цепи (VL), где VL содержит LCDR1, выбранный из SEQ ID NO: 13, 16, 19, 22 и 63, LCDR2, выбранный из SEQ ID NO: 14, 17, 20, 23 и 64, и LCDR3, выбранный из SEQ ID NO: 15, 18, 21, 24 и 65.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, содержат вариабельную область легкой цепи (VL), где VL содержит совокупность аминокислотных последовательностей LCDR1, LCDR2 и LCDR3, выбранных из любой совокупности F - J:

В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые здесь, содержат вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), причем эти вариабельные области содержат совокупность из 6 аминокислотных последовательностей CDR, выбранных из любой совокупности I - VI:


В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, содержат вариабельную область тяжелой цепи (VH) и/или вариабельную область легкой цепи (VL), где VH имеет аминокислотную последовательность, выбранную из SEQ ID NO: 25, 26, 27, 28, 33, 35, 38 и 40, и/или VL имеет аминокислотную последовательность, выбранную из SEQ ID NO: 29, 30, 31, 32, 34, 36, 38, 39 и 41.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые здесь, содержат вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), причем эти вариабельные области имеют совокупность аминокислотных последовательностей, выбранных из любой совокупности 1-7:

В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, содержат легкую цепь (LC) и/или тяжелую цепь (НС), причем НС имеет аминокислотную последовательность, выбранную из SEQ ID NO: 42, 44, 46 и 49, и/или LC имеет аминокислотную последовательность, выбранную из SEQ ID NO: 43, 45, 47, 48 и 50.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, содержат легкую цепь (LC) и тяжелую цепь (НС), причем LC имеет аминокислотную последовательность, указанную в SEQ ID NO: 43, а НС имеет аминокислотную последовательность, указанную в SEQ ID NO: 42 или 44; LC имеет аминокислотную последовательность, указанную в SEQ ID NO: 45, и НС имеет аминокислотную последовательность, указанную в SEQ ID NO: 44; LC имеет аминокислотную последовательность, указанную в SEQ ID NO. 47 или 48, и НС имеет аминокислотную последовательность, указанную в SEQ ID NO: 46; или LC имеет аминокислотную последовательность, указанную в SEQ ID NO: 50, и НС имеет аминокислотную последовательность, указанную в SEQ ID NO: 49.
В одном воплощении антитело, раскрытое здесь, является интактным антителом, предпочтительно IgG и более предпочтительно IgG4.
В одном конкретном воплощении антитело, раскрытое в настоящем документе, представляет собой 1F8, 2В2, 2F5, ch1, ch2, ch16, hu31, hu43, hu44, hu59, hu60 или hu250.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, характеризуются: а) специфическим связыванием с гомодимером IL-17AH гетеродимером IL-17AF; б) блокированием связывания IL-17A c его рецептором; и/или в) ингибированием опосредованной IL-17A биологической активности.
В одном воплощении IL-17A, IL-17AF или IL-17F, описанные здесь, выбраны из одного или более из яванского макака, мыши и человека.
В одном воплощении, антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, специфически не связывается с а) любым одним или более гомодимером IL-17F человека, гомодимером IL-17B человека, гомодимером IL-17C человека, гомодимером IL-17D человека и гомодимером IL-17E человека и/или б) любым одним или более гомодимером IL-17F яванского макака и гомодимером IL-17F мыши.
В другом аспекте антитело или его антигенсвязывающий фрагмент, раскрытый в настоящем документе, выполняет функцию ингибирования связывания IL-17A с его рецептором и/или снижения трансдукции клеточного сигнала и/или опосредованной IL-17A биологической активности.
В одном воплощении антитело или его антигенсвязывающий белок, раскрытые в настоящем документе, способны ингибировать индукцию посредством IL-17A секреции CXCL1 эпителиальными клетками при оценке активности in vitro.
В одном воплощении, антитело или его антигенсвязывающий белок, раскрытые в настоящем документе, способны ингибировать индукцию посредством IL-17A секреции CXCL1 у мышей при оценке активности in vivo.
В одном воплощении антитело или его антигенсвязывающий белок, раскрытые в настоящем документе, способны препятствовать возникновению псориаза, индуцированного имиквимодом в мышиной модели, и снижать клинический показатель начала псориаза у мышей и степень проявления ушного аллерготеста у мышей, при оценке активности in vivo.
В одном воплощении выделенное антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, способны ингибировать отек коленного сустава в модели антиген-индуцированного артрита, такой как AIA-модель у яванского макака, при оценке in vivo.
В настоящем изобретении также предложено применение антитела или его антигенсвязывающего фрагмента (предпочтительно hu31, hu43, hu44, hu59, hu60 или hu250) в качестве лекарственного средства, предпочтительно в качестве лекарственного средства для лечения патологических заболеваний, опосредованных IL-17A, и/или тех, которые лечат посредством ингибирования IL-17A сигнальной трансдукции.
В одном конкретном воплощении патологическое заболевание, опосредованное IL-17A, представляет собой воспалительное заболевание или состояние, такое как артрит, ревматоидный артрит, псориаз, анкилозирующий спондилит, хроническая обструктивная болезнь легких, системная красная волчанка (SLE), волчаночный нефрит, астма, рассеянный склероз или муковисцидоз.
В настоящем изобретении предложен способ лечения патологических заболеваний, опосредованных IL-17A, включающий введение эффективного количества выделенного антитела или его антигенсвязывающего фрагмента, раскрытого в настоящем изобретении, предпочтительно антитела hu31, hu43, hu44, hu59, hu60 или hu250, для облегчения состояния.
Настоящее изобретение также относится к способу получения антитела или его антигенсвязывающего фрагмента, раскрытым в настоящем изобретении. Такие способы включают выделенные молекулы нуклеиновых кислот, кодирующие по меньшей мере вариабельную область тяжелой и/или легкой цепи антитела, или белок, раскрытый в настоящем документе, или вектор клонирования/экспрессии, содержащий такие нуклеиновые кислоты, в частности для рекомбинации в клетке-хозяине с получением антитела или белка, раскрытых в настоящем документе, например hu31, hu43, hu44, hu59, hu60 или hu250.
Настоящее изобретение также относится к одному или более векторам клонирования и к клетке-хозяину, содержащей экспрессионные векторы, и к способу получения антитела или белка, содержащего его антигенсвязывающий фрагмент, раскрытому в настоящем документе, в частности такому, как антитело hu31, hu43, hu44, hu59, hu60 или hu250, где способ включает культивирование клетки-хозяина, очистку и выделение антитела или белка.
В одном воплощении выделенное антитело или белок, содержащий его антигенсвязывающий фрагмент, раскрытые в настоящем документе, конъюгированы с другой активной группировкой.
В одном воплощении антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, может быть моноклональным антителом или его антигенсвязывающий фрагментом, предпочтительно химерным антителом, гуманизированным антителом или человеческим антителом, или его частью.
В одном аспекте настоящего изобретения предлагается фармацевтическая композиция, содержащая антитело или белок, содержащий его антигенсвязывающий фрагмент в соответствии с любым из воплощений настоящего изобретения, в сочетании с одним или более фармацевтически приемлемыми эксципиентами, разбавителями или носителями.
В одном воплощении фармацевтическая композиция содержит один или более дополнительных активных ингредиентов.
В одном конкретном воплощении фармацевтическая композиция представляет собой лиофилизированный порошок. В другом конкретном воплощении фармацевтическая композиция представляет собой стабильную жидкую композицию, содержащую терапевтически приемлемое количество антитела или молекулы, раскрытой в настоящем документе.
В одном воплощении настоящее изобретение относится к применению антитела или его антигенсвязывающего фрагмента для получения лекарственного средства для лечения любого из патологических расстройств, опосредованных IL-17A.
В настоящем изобретении предлагается выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент, экспрессионный вектор или рекомбинантный вектор, содержащий молекулу нуклеиновой кислоты, и клетка-хозяин, трансформированная вектором.
В настоящем изобретении предлагается фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, молекулу нуклеиновой кислоты, вектор или клетку-хозяина и фармацевтически приемлемый носитель или эксципиент.
В настоящем изобретении предлагается применение антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, вектора или клетки-хозяина, или фармацевтической композиции для получения лекарственного средства для лечения и/или предупреждения заболеваний или расстройств, опосредованный IL-17A.
В некоторых воплощениях лекарственное средство является лекарственным средством для лечения артрита, ревматоидного артрита, псориаза, анкилозирующего спондилита, хронической обструктивной болезни легких, системной красной волчанки (SLE), волчаночного нефрита, астмы, рассеянного склероза или муковисцидоза.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 показана электрофореграмма SDS-PAGE рекомбинантного человеческого белка IL-17A-mFc.
На Фиг. 2 показано связывание человеческого IL-17A-mFc с IL-17RA на клетках 293F, исследованное с помощью FACS (MFI представляет среднюю интенсивность флуоресценции; группа, подвергнутая лечению, против контрольной группы: 7695 против 308).
На Фиг. 3 показано влияние гибридомных антител на блокирование опосредованной IL-17A биологической активности in vivo, исследованное методом ELIS А (твердофазный иммуноферментный анализ). Гибридомное антитело значительно ингибирует индукцию экспрессии CXCL1 посредством IL-17Ay мышей.
На Фиг. 4 показано связывание химерных антител с человеческим IL-17A, исследованное посредством ELISA. Химерные антитела ch1, ch2, ch4, ch7 и ch16 связываются с человеческим IL-17Ac более высокой специфичностью, со значениями ЕС50, составляющими 6,62 нг/мл, 5,17 нг/мл, 88,48 нг/мл, 39,96 нг/мл и 15,42 нг/мл, соответственно.
На Фиг. 5 показано влияние химерных антител на блокирование связывания человеческого IL-17A с IL-17RA на клетках 293F, анализируемых посредством FACS (сортировка флуоресцентно-активированных клеток). Химерные антитела ch1, ch2, ch7 и ch16 эффективно блокируют связывание, со значениями IC50, составляющими 4,46 мкг/мл, 3,145 мкг/мл, 1,220 мкг/мл и 1,445 мкг/мл соответственно.
На Фиг. 6 показано связывание гуманизированных антител с человеческим IL-17A, исследованное посредством ELISA. Антитела hu31, hu43, hu44, hu59, hu60 и hu250 связываются с человеческим IL-17A с более высокой специфичностью, со значениями ЕС50, составляющими 8,13 нг/мл, 8,64 нг/мл, 6,764 нг/мл, 6,102 нг/мл, 5,776 нг/мл и 6,351 нг/мл соответственно.
На Фиг. 7 показан эффект блокирования гуманизированными антителами связывания человеческого IL-17A с IL-17RA на клетках 293F, исследованный посредством FACS. Все антитела hu31, hu43, hu44, hu59, hu60 и hu250 эффективно блокируют связывание со значениями IC50, составляющими 867,6 нг/мл, 780,8 нг/мл, 828,5 нг/мл, 467,4 нг/мл, 482,8 нг/мл и 577,8 нг/мл соответственно.
На Фиг. 8 показано влияние гуманизированных антител на блокирование IL-17А-опосредованной секреции CXCL1 эпителиальными клетками, исследованное посредством ELISA. Все антитела hu31, hu43, hu44, hu59, hu60 и hu250 эффективно ингибируют индукцию экспрессии CXCL1 посредством IL-17A в эпителиальных клетках и обладают более сильным блокирующим эффектом, чем контрольное антитело.
На Фиг. 9 показано влияние гуманизированных антител на блокирование IL-17А-опосредованной биологической активности in vivo, исследованное посредством ELISA. Антитела hu31, hu43, hu44, hu60 и hu250 эффективно ингибируют IL-17A-индуцируемую экспрессии CXCL1 у мышей и обладают более сильным блокирующим эффектом, чем контрольное антитело.
На Фиг. 10-1 показано влияние введения гуманизированного антитела на клинический показатель индуцированного имиквимодом псориаза у мышей. Введение hu31 и hu44 может значительно ингибировать шелушение кожи, уплотнение, отечность и другие состояния индуцированного имиквимодом псориаза в мышиной модели, то есть уменьшать клинический показатель (*Р<0,05 в сравнении с KLH (гемоцианин лимфы улитки)).
На Фиг. 10-2 показано влияние гуманизированных антител на степень отечности ушей у мышей с имиквимод-индуцированным псориазом. Введение hu31 и hu44 может значительно улучшить степень отечности ушей. (*Р<0,05 в сравнении с KLH, **Р<0,01 в сравнении с KLH).
На Фиг. 11-1 показано влияние гуманизированных антител на массу тела самок яванских макак с артритом, индуцированным коллагеном II типа. hu31 и hu59 в некоторой степени улучшают потерю массы, вызванную артритом (**Р<0,01, ****Р<0,0001 по сравнению с "G2: контрольной группой"; однофакторный дисперсионный анализ ANOVA/Даннетт).
На Фиг. 11-2 показано влияние гуманизированных антител на показатель артрита, индуцированного коллагеном II типа, у самок яванских макак. hu31 значительно подавляет тенденцию к увеличению клинического показателя артрита у яванских макак (***Р<0,001, #Р<0,05 по сравнению с G2: контрольная группа; однофакторный дисперсионный анализ ANOVA/Даннетт).
На Фиг. 12 показано влияние гуманизированных антител на индуцированную NIH3T3-IL-17 клетками отечность суставов у мышей.
На Фиг. 13 показано влияние гуманизированных антител на индуцированное NIH3T3-IL-17 клетками воспаление в мышиной модели "воздушный мешок".
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителу, которое специфически связывается с человеческим IL-17A и блокирует опосредованную IL-17A биологическую активность. Настоящее изобретение относится к полноразмерному антителу IgG и его антигенсвязывающему фрагменту, описанным ниже.
Определения
Если не указано иное, в воплощениях настоящего изобретения будут использованы традиционные методики молекулярной биологии (включая рекомбинантные методики), микробиологии, цитобиологии, биохимии и иммунологии, все из которых находятся в пределах знаний специалиста.
Для облегчения понимания настоящего изобретения некоторые технические и научные термины конкретно определены следующим образом. Если иное конкретно не определено в настоящем документе, все технические и научные термины, используемые в настоящем документе, имеют такое же значение, которое обычно подразумевают специалисты в области техники, к которой относится настоящее изобретение. За определениями и терминологией в данной области специалист может обратиться непосредственно к современным протоколам молекулярной биологии (Ausubel). Сокращенные обозначения аминокислотных остатков представляют собой стандартные 3-буквенные и/или 1-буквенные коды, используемые в данной области техники для обозначения одной из 20 обычно используемых L-аминокислот.
Формы единственного числа, используемые в настоящем документе (включая пункты формулы изобретения), включают их формы множественного числа, если иное явным образом не указано в контексте.
IL-17 или IL-17A представляют собой интерлейкин-17. Если не указано иное, IL-17 обычно относится к человеческому IL-17A.
IL-17A экспрессируемый in vivo, его номер в NCBI NP-443104.1, имеет N-концевой сигнальный пептид, состоящий из 23 аминокислот, который отщепляется с получением зрелого IL-17A. В одном конкретном воплощении IL-17A, раскрытый здесь, относится к зрелому IL-17A, который не содержит N-концевого сигнального пептида, с аминокислотной последовательностью, указанной в SEQ ID NO: 66, и нуклеотидной последовательностью, указанной в SEQ ID NO: 67.
IL-17F, экспрессируемый in vivo, его номер в NCBI NP-443104.1, имеет N-концевой сигнальный пептид, состоящий из 30 аминокислот, который отщепляется с получением зрелого IL-17F. В одном конкретном воплощении раскрытый здесь IL-17F относится к зрелому IL-17F, который не содержит N-концевого сигнального пептида, с аминокислотной последовательностью, указанной в SEQ ID NO: 68.
Раскрытый здесь IL-17AF представляет собой гетеродимер субъединицы IL-17A и субъединицы IL-17F в понимании специалистов в данной области техники.
Термин "иммунный ответ" относится к действию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитов, гранулоцитов и растворимых макромолекул, продуцируемых вышеуказанными клетками или печенью (включая антитела, цитокины и комплементы), которое приводит к избирательному повреждению, разрушению или элиминации из организма человека инвазивных патогенов, клеток или тканей, инфицированных патогенами, раковых клеток или, в случаях аутоиммунитета или патологического воспаления, нормальных клеток или тканей человека.
Термин "путь передачи сигнала" или "активность передачи сигнала" относится к биохимической причинно-следственной связи, обычно инициируемой белок-белковым взаимодействием, такой как связывание фактора роста с рецептором, приводящей к передаче сигнала от одной части клетки к другой части клетки. В общем случае, передача включает специфическое фосфорилирование одного или более остатков тирозина, серина или треонина на одном или более белках в ряде реакций, вызывающих передачу сигнала. Предпоследний процесс обычно включает в себя ядерное событие, приводящее к изменению экспрессии генов.
В отношении антитела или его антигенсвязывающего фрагмента, раскрытого в настоящем документе, термин "активность" или "биологическая активность", или термин "биологическое свойство" или "биологическая характеристика" могут использоваться здесь взаимозаменяемо и включают, без ограничения ими, сродство к эпитопу/антигену и специфичность, способность нейтрализовать или противодействовать активности IL-17А in vivo или in vitro, IC50, стабильность антитела in vivo и иммуногенные свойства антитела. Другие идентифицируемые биологические свойства или характеристики антитела, известные в данной области техники, включают, например, перекрестную реактивность (т.е. перекрестную реактивность с нечеловеческими гомологами целевого пептида или с другими белками или тканями в общем смысле) и способность поддерживать высокий уровень экспрессии белка в клетках млекопитающих. Вышеупомянутые свойства или характеристики наблюдаются, определяются или оцениваются с использованием методик, хорошо известных в данной области техники, включая, но без ограничения ими, ELISA, FACS или анализ плазменного резонанса BIACORE, неограниченные анализы нейтрализации in vitro или in vivo, связывание рецепторов, продуцирование и/или секрецию цитокинов или факторов роста, трансдукцию сигналов и иммуногистохимию участков тканей разного происхождения (включая человека, приматов или имеющих любое другое происхождение).
"Антитело" относится к любой форме антител, имеющей желательную биологическую активность. Таким образом, этот термин используется в самом широком смысле и конкретно включает, без ограничения ими, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, мультиспецифические антитела (например биспецифические антитела), гуманизированные антитела, полностью человеческие антитела, химерные антитела и верблюдизированные однодоменные антитела.
"Изолированное антитело" относится к очищенному состоянию конъюгата и в этом случае означает, что молекула по существу свободна от других биомолекул, таких как нуклеиновые кислоты, белки, липиды, сахара или другие вещества, такие как клеточный дебрис и среда для роста. Термин "изолировать(нный)" не означает полного отсутствия таких веществ или отсутствие воды, буферов или солей, если только они не присутствуют в количествах, которые будут существенно мешать экспериментальному или терапевтическому применению конъюгатов, описанных в настоящем документе.
"Моноклональное антитело" относится к антителу, полученному из по существу однородной популяции антител, то есть антитела, составляющие популяцию, идентичны, за исключением возможных естественных мутаций, которые могут присутствовать в незначительном количестве. Моноклональное антитело является высокоспецифичными и нацелено на один эпитоп антигена. Напротив, обычные (поликлональные) препараты антител обычно включают большое количество антител, нацеленных на различные эпитопы (или специфичных к ним). Модификатор "моноклональный" указывает характеристику антитела, полученного из по существу однородной популяции антител, и его не следует толковать как получение антитела каким-либо конкретным способом.
"Полноразмерное антитело" относится к молекуле иммуноглобулина, содержащей четыре пептидные цепи, когда они присутствуют в природе, включая две тяжелые (Н) цепи (примерно 50-70 кДа при полной длине) и две легкие (L) цепи (примерно 25 кДа при полной длине), связанные друг с другом дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначаемой здесь как VH) и константной области тяжелой цепи (сокращенно обозначаемой здесь как СН). Константная область тяжелой цепи состоит из 3 доменов CH1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой здесь как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. VH и VL области могут быть дополнительно разделены на области, определяющие комплементарность (CDR), с высокой изменчивостью и более консервативными областями, называемыми каркасными участками (FR), которые расположены друг от друга на расстоянии CDR. Каждая область VH или VL состоит из 3 CDR и 4 FR, в следующем порядке от аминоконца к карбоксильному концу: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающие домены, которые взаимодействуют с антигенами. Константные области антитела могут опосредовать связывание иммуноглобулинов с тканями или факторами хозяина, включая связывание различных клеток иммунной системы (например эффекторных клеток) с первым компонентом (C1q) классической системы комплемента.
"Антигенсвязывающий фрагмент" антитела ("родительского антитела") включает фрагмент или производное антитела, обычно включающий(ее) по меньшей мере один фрагмент антигенсвязывающей области или вариабельной области (например одну или более CDR) родительского антитела, который сохраняет по меньшей мере часть специфичности связывания родительского антитела. Примеры связывающих фрагментов антитела включают, без ограничения ими, фрагменты Fab, Fab', F(ab')2 и Fv; диатело; линейное антитело; молекулу одноцепочечного антитела, такую как sc-Fv; и нанотело и мультиспецифическое антитело, образованное фрагментами антитела. Связывающий фрагмент или производное обычно сохраняет по меньшей мере 10% своей антигенсвязывающей активности, когда антигенсвязывающая активность присутствует в молярной концентрации. Предпочтительно, связывающий фрагмент или производное сохраняет по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% или более антигенсвязывающего аффинности родительского антитела. Также предполагается, что антигенсвязывающий фрагмент антитела может включать консервативные или неконсервативные аминокислотные замены, которые существенно не изменяют их биологическую активность (называемые "консервативными вариантами" или "функционально-консервативными вариантами" антитела). Термин "конъюгат" относится как к антителу, так и к его связывающему фрагменту.
"Одноцепочечное антитело Fv" или "scFv" относится к фрагменту антитела, содержащему домены VH и VL антитела, где эти домены присутствуют в одной полипептидной цепи. В общем случае, полипептид Fv также содержит полипептидный линкер между доменами VH и VL, который позволяет scFv формировать нужную структуру для связывания антигена.
"Доменное антитело" представляет собой иммунофункциональный фрагмент иммуноглобулина, который содержит только вариабельную область тяжелой или легкой цепи. В некоторых случаях две или более областей VH ковалентно связаны с пептидным линкером с образованием двухвалентного доменного антитела. Две VH-области двухвалентного доменного антитела могут быть нацелены на одни и те же или разные антигены.
"Двухвалентное антитело" содержит 2 антигенсвязывающих сайта. В некоторых случаях эти 2 сайта связывания имеют одинаковую антигенную специфичность. Однако двухвалентное антитело может быть биспецифичным.
"Диатело" относится к небольшому фрагменту антитела, имеющему два антигенсвязывающих сайта, который содержит вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в той же полипептидной цепи (VH-VL или VL-VH). При использовании линкера, который слишком короток для спаривания между двумя доменами в одной цепи, домены вынуждены спариваться с комплементарными доменами другой цепи, образуя два антигенсвязывающих сайта.
"Химерное антитело" представляет собой антитело, имеющее вариабельные домены первого антитела и константные домены второго антитела, причем первое и второе антитела принадлежат к разным видам. Обычно вариабельный домен получают из антитела экспериментального животного, такого как грызун ("родительское антитело"), а последовательность константного домена получают из человеческого антитела, так что полученное в результате химерное антитело с меньшей вероятностью индуцирует неблагоприятный иммунный ответ у человека по сравнению с родительским антителом грызуна.
"Гуманизированное антитело" относится к форме антител, содержащей последовательности как человеческих, так и нечеловеческих (таких как мыши и крысы) антител. В общем случае, гуманизированное антитело содержит по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют петлям иммуноглобулина, не являющегося человеческим, и все или по существу все каркасные участки (FR) являются участками последовательности иммуноглобулина человека. Гуманизированное антитело возможно может содержать по меньшей мере часть константной области иммуноглобулина человека (Fc).
"Полностью человеческое антитело" относится к антителу, которое содержит только последовательности иммуноглобулинов человека. Полностью человеческое антитело может содержать мышиные гликоцепи, если оно получено в мышах, мышиных клетках или гибридомах, полученных из мышиных клеток. Аналогично, "мышиное антитело" относится к антителу, которое содержит только последовательности мышиного иммуноглобулина. Альтернативно, полностью человеческое антитело может содержать гликоцепи крысы, если оно продуцируется в крысах, клетках крыс или гибридомах, полученных из клеток крыс.Аналогично, "крысиное антитело" относится к антителу, которое содержит только последовательности крысиных иммуноглобулинов.
"Изотип" относится к типу антител (например IgM, IgE, IgG, такому как IgG1 или IgG4), обеспечиваемому генами константной области тяжелой цепи. Изотип также включает модифицированные формы одного из этих типов, в которые были внесены модификации для изменения функции Fc, например для усиления или ослабления эффекторной функции или связывания с Fc-рецепторами.
Термин "нуклеиновая кислота" или "полинуклеотид" относится к дезоксирибонуклеиновой кислоте (ДНК) или рибонуклеиновой кислоте (РНК) и их полимерам в одно- или двухцепочечной форме. Если он явно не ограничен, этот термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают аналогичными связывающими свойствами, как и эталонная нуклеиновая кислота, и метаболизируются образом, аналогичным природным нуклеотидам (см патент US 8278036 - Kariko et al., который раскрывает молекулу мРНК, в которой уридин замещен псевдоуридином, а также способ синтеза молекулы мРНК и способ доставки терапевтических белков in vivo). Могут быть использованы способы с модифицированной мРНК, например, те, которые раскрыты в патенте US 8278036 - Kariko et al. и в патентной заявке WO 2013/090186 A1 - Moderna. Если не указано иное, конкретная нуклеиновокислотная последовательность также неявно включает ее консервативно модифицированные варианты (например замены вырожденных кодонов), аллели, ортологи, SNP (однонуклеотидные полиморфизмы) и комплементарные последовательности, а также явно указанную последовательность. В частности, замены вырожденных кодонов могут быть достигнуты путем образования последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов заменено смешанными основаниями и/или остатками дезоксиинозина (Batzer et al., Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); и Rossolini et al., MoA Cell. Probes 8:91-98 (1994)).
"Конструкция" относится к любой рекомбинантной полинуклеотид ной молекуле (такой как плазмида, космида, вирус, автономно реплицирующаяся полинуклеотидная молекула, фаг, или линейная или кольцевая одно- или двухцепочечная полинуклеотидная молекула ДНК или РНК), полученная из любого источника, способная к геномной интеграции или автономной репликации, содержащая полинуклеотидную молекулу, в которой одна или более полинуклеотидных молекул связаны функциональным образом (т.е. функционально связаны). Рекомбинантная конструкция, как правило, содержит полинуклеотид по настоящему изобретению, функционально связанный с регуляторными последовательностями инициации транскрипции, которые направляют транскрипцию полинуклеотида в клетке-хозяине. Как гетерологичные, так и негетерологичные (т.е. эндогенные) промоторы могут быть использованы для прямой экспрессии нуклеиновых кислот по настоящему изобретению.
"Вектор" относится к любой рекомбинантной полинуклеотидной конструкции, которая может быть использована для трансформации (т.е. введения гетерологичной ДНК в клетку-хозяина). Одним из типов вектора является "плазмида", которая относится к двухцепочечной петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например бактериальные векторы, имеющие бактериальное начало репликации, и эписомные векторы млекопитающих). После введения в клетку-хозяина другие векторы (например неэписомные векторы млекопитающих) интегрируются в геном клетки-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Кроме того, определенные векторы способны направлять экспрессию функционально связанных генов. Такие векторы упоминаются здесь как "экспрессионные векторы".
Термин "экспрессионный вектор", используемый здесь, относится к молекуле нуклеиновой кислоты, способной к репликации и экспрессии целевого гена при трансформации, трансфекции или трансдукции в клетку-хозяина. Экспрессионный вектор содержит один или несколько фенотипических селектируемых маркеров и точку начала репликации для обеспечения поддержания вектора и для обеспечения амплификации в хозяине, если это необходимо.
Используемый здесь термин "антагонист IL-17A" или "блокатор IL-17A" относится к антителу или его антигенсвязывающему белку, который ингибирует активность передачи сигнала IL-17A, индуцированного IL-17R, тем самым снижая или нейтрализуя активность IL-17A. Это может быть показано в анализах для клеток человека, таких как анализы зависимого от IL-17A продуцирования CXCL1 для клеток человека. Такие анализы более подробно описаны в приведенных ниже примерах.
Если иное явно не указано в контексте, то "активация", "стимуляция" и "лечение" для клетки или рецептора могут иметь одно и то же значение. Например, клетка или рецептор активируется, стимулируется или обрабатывается лигандом. "Лиганды" включают природные и синтетические лиганды, такие как цитокины, варианты цитокинов, аналоги, мутантные белки и связывающие соединения, полученные из антител. "Лиганды" также включают небольшие молекулы, такие как пептидомиметики цитокинов и пептидомиметики антител. "Активация" может относиться к активации клетки, регулируемой внутренними механизмами, а также внешними факторами или факторами окружающей среды. "Ответ/реакция", например реакция клетки, ткани, органа или организма, включает изменения в биохимическом или физиологическом поведении (например концентрации, плотности, адгезии или миграции, скорости экспрессии генов или состоянии дифференцировки в биологическом компартменте), где изменения связаны с активацией, стимуляцией или воздействием, или связаны с внутренним механизмом, таким как генетическое программирование.
Используемый здесь термин "лечить", "лечение" или "обработка" любого заболевания или расстройства относится в одном воплощении к уменьшению интенсивности заболевания или расстройства (т.е. к замедлению или остановке, или к уменьшению развития заболевания или по меньшей мере одного из его клинических симптомов). В другом воплощении термины "лечить", "лечение" или "обработка" относится к улучшению состояния или улучшению по меньшей мере одного физического параметра, включая те физические параметры, которые могут быть не заметны пациенту. В другом воплощении термины "лечить", "лечение" или "обработка" относятся к изменению заболевания или расстройства, физически (то есть стабилизация явных симптомов), физиологически (например стабилизация физических параметров), или к обоими изменениям. Если явно не оговорено в описании изобретения, то методы оценки лечения и/или предупреждения заболевания, как правило, известны в данной области.
"Субъект" включает любого человека или любое животное, не являющееся человеком. Термин "животное, не являющееся человеком" включает всех позвоночных, например млекопитающих и не млекопитающих, таких как приматы, не являющиеся человеком, овца, собака, кошка, лошадь, крупный рогатый скот, куры, амфибии и рептилии. Используемый здесь термин "супо" относится к яванским макакам.
"Терапевтически эффективное количество", "терапевтически эффективная доза" и "эффективное количество", при использовании в настоящем документе, относятся к количеству антитела IL-17A или его антигенсвязывающего фрагмента, раскрытого в настоящем документе, которое является эффективным для предупреждения или улучшения одного или более симптомов заболевания или состояния, или развития заболевания или состояния при введении отдельно или в комбинации с другими терапевтическими агентами в клетку, ткань или субъекта. Терапевтически эффективная доза также относится к количеству антитела или его антигенсвязывающего фрагмента, достаточному для того, чтобы вызвать улучшение симптомов, например к количеству для лечения, излечения, предупреждения или улучшения соответствующего состояния или для содействия лечению, излечению, предупреждению или улучшению такого состояния. Когда активный ингредиент вводят субъекту сам по себе, терапевтически эффективная доза относится к количеству этого ингредиента. При введении комбинации, терапевтически эффективная доза относится к объединенному количеству активных ингредиентов, которое оказывает терапевтический эффект, независимо от того, вводят эти активные ингредиенты в комбинации, последовательно или одновременно. Эффективное количество терапевтического агента приводит к увеличению диагностического показателя или параметра по меньшей мере на 10%, обычно по меньшей мере на 20%; предпочтительно по меньшей мере примерно на 30%, более предпочтительно по меньшей мере на 40%, наиболее предпочтительно по меньшей мере на 50%.
Получение антител
Любой подходящий способ получения антител может быть использован для получения антитела, раскрытого в настоящем документе. Любая подходящая форма человеческого IL-17A может быть использована в качестве иммуногена (антигена) для продуцирования антител. В качестве примера, а не ограничения ими, любой изотип IL-17A человека или его фрагмент могут быть использованы в качестве иммуногена. Примеры включают, без ограничения ими, естественный зрелый человеческий IL-17A (имеющий аминокислотную последовательность, указанную в SEQ ID NO: 66), как описано здесь.
В предпочтительном воплощении гибридомные клетки, продуцирующие мышиные моноклональные антитела против IL-17A человека, могут быть получены методами, хорошо известными в данной области. Эти методы включают, без ограничения ими, гибридомные технологии, первоначально разработанные Kohler et al., (1975) (Nature 256:495-497). Предпочтительно, мышиные спленоциты выделяют и сливают с мышиной миеломной клеточной линией с использованием ПЭГ или электрослияния в соответствии со стандартными схемами. Полученные гибридомы, продуцирующие антиген-специфические антитела, затем могут быть подвергнуты скринингу. Например, в случае 50% ПЭГ, одноклеточная суспензия лимфоцитов селезенки, полученных от иммунизированной мыши, может быть слита с 1/6 количеством мышиных миеломных клеток SP20 (АТСС). Клетки могут быть посеяны в микротитровальный планшет с плоским дном с плотностью примерно 2×105 клеток/мл, с последующей 2 недельной инкубацией в полной среде, содержащей 20% фетальной бычьей сыворотки, и селекционной среде, содержащей 1 × HAT (среда с гипоксантином, аминоптерином и тимидином) (Sigma; HAT добавляли через 24 ч после слияния). Через 2 недели клетки можно культивировать в среде, где HAT заменили на НТ (HAT без аминоптерина). Затем лунки могут быть подвергнуты скринингу посредством ELISA на наличие моноклональных антител IgG к IL-17A человека. В общем случае, через 10-14 суток крупномасштабного роста гибридом можно оценивать среду. Гибридомы, секретирующие антитела, могут быть посеяны и снова подвергнуты скринингу. Если полученные гибридомы остаются положительными в отношении IgG человека, гибридомы с моноклональными антителами к IL-17A человека могут быть субклонированы по меньшей мере дважды посредством серийных разведений. Стабильные субклоны затем можно культивировать in vitro с получением небольших количеств антител в тканевой среде для их характеристики.
В предпочтительном воплощении моноклональными гибридомными клетками, полученными в настоящем изобретении, являются 1F8, 2F5 и 2В2, которые секретируют антитела, связывающиеся с IL-17A с высокой специфичностью, блокирующие связывание IL-17A с IL-17RA и ингибирующие IL-17А-опосредованную биологическую активность, такую как ингибирование секреции CXCL1.
В предпочтительном воплощении последовательности ДНК вариабельных областей иммуноглобулина гибридомных клеток-кандидатов 1F8, 2F5 и 2В2 определяют в настоящем изобретении с использованием ПЦР на основе дегенеративных праймеров. Гибридомная клетка 1F8 продуцирует два гена легкой цепи антител и один ген тяжелой цепи антитела, a 2F5 продуцирует два гена тяжелой цепи антитела и один ген легкой цепи антитела. Следовательно, каждое из антител, секретируемых 1F8 и 2F5, может включать два смешанных интактных антитела. В данном описании антитела, секретируемые 1F8, обозначены как 1F8-1 и 1F8-2, а антитела, секретируемые 2F5, обозначены как 2F5-1 и 2F5-2.

Антитела, полученные от грызунов (например мыши), могут индуцировать нежелательную иммуногенность антител при использовании в качестве терапевтических агентов in vivo. Повторное применение этих антител вызывает иммунный ответ в организме человека на терапевтические антитела. Такие иммунные реакции приводят, по меньшей мере, к потере терапевтической эффективности и, в худшем случае, к потенциально смертельной аллергической реакции. Один из способов снижения иммуногенности антител грызунов включает получение химерных антител, в которых мышиная вариабельная область слита с константной областью человека (Liu et al., (1987) Proc. Natl. Acad. Sci. USA 84:3439-43). Однако сохранение интактной вариабельной области грызунов в химерных антителах все еще может индуцировать опасную иммуногенность у пациентов. Пересадку петель области, определяющей комплементарность (CDR), вариабельного домена грызунов на человеческий каркас (т.е. гуманизация) использовали для дополнительной минимизации последовательностей грызунов (Jones et al., (1986) Nature 321:522; Verhoeyen et al., (1988) Science 239:1534).
В некоторых воплощениях, химерные или гуманизированные антитела, раскрытые здесь, могут быть получены на основе последовательностей моноклональных антител, полученных с помощью гибридомы семейства мышиных. ДНК, кодирующая тяжелые и легкие цепи иммуноглобулина, может быть получена из представляющей интерес гибридомы мышиных и сконструирована так, чтобы включать последовательности немышиных (например человеческих) иммуноглобулинов, с использованием стандартных методов молекулярной биологии.
В некоторых воплощениях для химерных антител, раскрытых здесь, химерная тяжелая цепь и химерная легкая цепь могут быть получены путем функционального связывания вариабельных областей тяжелой цепи и легкой цепи иммуноглобулина гибридомного происхождения с константными областями IgG человека соответственно, с использованием методов, известных в данной области (см, например, US 4816567 - Cabilly et al.). Человеческий IgG может быть выбран из любого подтипа, такого как IgG1, IgG2, IgG3, IgG4, предпочтительно IgG4.
В конкретном воплощении химерные антитела, раскрытые здесь, могут быть получены посредством "смешивания и подбора" экспрессионной плазмиды для химерной легкой цепи с экспрессионной плазмидой для химерной тяжелой цепи для трансфекции экспрессирующих клеток. Связывание таких "смешанных и подобранных" антител с IL-17A может быть проанализировано с использованием вышеупомянутого анализа связывания и других обычных анализов связывания (например ELISA). Предпочтительные ch1, ch2, ch4, ch7 и ch16 обладают оптимальной связывающей и блокирующей активностью, и их аминокислотные последовательности вариабельной области показаны в Таблице 2.
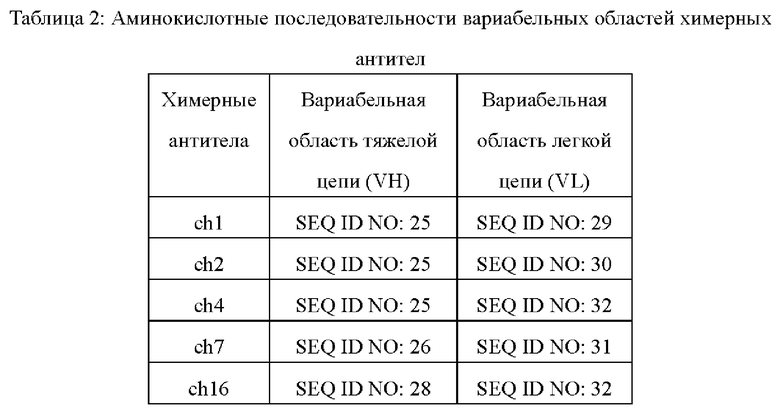
Точные границы аминокислотных последовательностей CDR вариабельной области в антителах, раскрытых в настоящем документе, могут быть определены с использованием любой из хорошо известных схем, включая схему Кабат, описанную в Kabat et al., (1991), Sequences of Proteins of Immunological Interest, 5th edition, Public Health Service, National Institutes of Health, Bethesda, MD. (схема нумерации "Kabat") и схему IMGT, описанную в Lefranc М.-Р. et al., (1999 Nucleic Acids Research, 27:209-212). В некоторых воплощениях конкретные схемы определения CDR и аминокислотные последовательности вариабельных областей предпочтительных мышиных антител, раскрытых в настоящем документе, показаны в таблице 3.

В некоторых воплощениях, для гуманизированных антител, раскрытых здесь, области CDR семейства мышиных могут быть вставлены в каркасные участки зародышевой линии человека с использованием методов, известных в данной области. См. патент US 5225539 - Winter et al. и патенты US 5530101; 5585089; 5693762 и 6180370 - Queen et al. В кратком изложении, гены IgG зародышевой линии человека, гомологичные последовательности кДНК вариабельных областей мышиных антител, были получены в базе данных генов иммуноглобулинов человека NCBI (http://www.ncbi.nlm.nih.gov/igblast/) автором изобретения и, в принципе, гуманизация достигается путем прививки выбранных CDR. Однако обмен петлями CDR, по-прежнему не позволяет однотипно произвести антитело с теми же свойствами связывания, что и исходное антитело. В гуманизированных антителах часто требуются изменения в каркасных остатках (FR) (остатках, участвующих в поддержке петли CDR) для поддержания антигенсвязывающей аффинности. Kabat et al., (1991) J. Immunol. 147:1709. В кратком изложении, гуманизация включает следующие стадии: А. последовательности генов антител-кандидатов сравнивают с последовательностью генов антител зародышевой линии человека для отбора последовательностей с высокой гомологией; В, с помощью анализа аффинности HLA-DR выбирают каркасную последовательность зародышевой линии человека с низкой аффинностью; и С, каркасные аминокислотные последовательности вариабельных областей и их периферию анализируют с помощью компьютерного моделирования и молекулярного докинга, и устанавливают пространственное и стерео-расположение. Ключевые аминокислотные единицы, которые взаимодействуют с IL-17A и поддерживают пространственный каркас в последовательности генов антител-кандидатов анализируют путем расчета электростатической силы, силы Ван-дер-Ваальса, гидрофильности и гидрофобности, и значения энтропии и прививают к выбранному каркасу зародышевой линии человека. Аминокислотные положения каркасных участков, которые должны быть сохранены, отображают на карте. После этого гуманизированные антитела синтезируют.
В некоторых воплощениях предпочтительные гуманизированные антитела, полученные в настоящем изобретении, представляют собой hu31, hu43, hu44, hu59, hu60 и hu250.
Вариабельные области гуманизированных антител hu31, hu43, hu44, hu59, hu60 и hu250 и их соответствующие аминокислотные последовательности CDR показаны в таблице 4.


Аминокислотные и нуклеотидные последовательности легкой/тяжелой цепи гуманизированных антител hu31, hu43, hu44, hu59, hu60 и hu250 показаны в Таблице 5.


Другие антитела, раскрытые здесь, включают антитела, имеющие аминокислотную последовательность, которая мутирована посредством удаления, вставки или замены аминокислоты, но все еще имеют по меньшей мере 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 98%, 99% или 100% идентичность с вышеуказанными антителами (особенно в областях CDR, представленных в вышеуказанных последовательностях). В некоторых воплощениях антитело, раскрытое здесь, представляет собой мутант любого из hu31, hu43, hu44, hu59, hu60 и hu250, где мутант содержит мутантную аминокислотную последовательность, в которой не более 1, 2, 3, 4 или 5 аминокислот мутированы путем делеции, вставки или замены аминокислот в области CDR по сравнению с областями CDR, представленными в вышеуказанных последовательностях.
Другие нуклеиновые кислоты, кодирующие антитело, раскрытое здесь, включают нуклеиновые кислоты, которые мутированы путем делеции, вставки или замены нуклеотидов, но все еще имеют по меньшей мере 60%, 70%, 80%, 90%, 95% или 100% идентичность с соответствующими областями кодирования CDR, представленными в указанных выше последовательностях.
Экспрессия антител
В некоторых воплощениях, настоящее изобретение относится к одному или более экспрессионный векторам или к клетке-хозяину, содержащей экспрессионные векторы, и к способу получения антитела или его антигенсвязывающего фрагмента, раскрытому в настоящем документе, где способ включает культивирование клетки-хозяина, очистку и выделение антитела или его антигенсвязывающего фрагмента.
Раскрытые здесь антитела могут быть получены в клетках-хозяев ах с использованием, например, комбинацию метода рекомбинантной ДНК и метода трансфекции генов, хорошо известных в данной области, (e.g., Morrison, S. 1985, Science 229:1202). Например, для экспрессии антитела или фрагмента антитела, ДНК, кодирующая части легких и тяжелых цепей или полноразмерные легкие и тяжелые цепи, может быть получена с использованием стандартных методов молекулярной биологии или биохимии (например химический синтез ДНК, ПЦР-амплификация или клонирование кДНК с использованием гибридом, экспрессирующих интересующее антитело), и ДНК может быть вставлена в экспрессионный вектор таким образом, чтобы гены были функционально связаны с транскрипционными и трансляционными контрольными последовательностями. В данном описании термин "функционально связанные" означает, что гены антител связаны в векторе таким образом, что последовательности контроля за транскрипцией и трансляцией в этом векторе выполняют свои запланированные функции регулирования транскрипции и трансляции генов антител. Выбирают вектор экспрессии и последовательности управления экспрессией, которые могут быть совместимы с используемой экспрессирующей клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть встроены в разные векторы или, более типично, оба гена встраивают в один и тот же экспрессионный вектор. Ген антитела встраивают в экспрессионный вектор стандартными методами (например лигированием фрагмента гена антитела и комплементарных сайтов рестрикции на векторе или лигированием тупого конца, если сайт рестрикции отсутствует). Полноразмерные гены антител любого изотипа антител могут быть получены путем вставки вариабельных областей легкой и тяжелой цепи антитела, описанного здесь, в экспрессионный вектор, который уже кодирует константные области тяжелой и легкой цепи необходимого изотипа, чтобы обеспечить функциональное связывание сегментов VH с сегментами СН в векторе и функциональное связывание сегментов VL с сегментами CL в векторе. Дополнительно или альтернативно, рекомбинантный экспрессионный вектор может кодировать сигнальный пептид (также называемый лидерной последовательностью), который облегчает секрецию цепи антител из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор таким образом, чтобы сигнальный пептид был связан с аминоконцом гена цепи антител в той же рамке считывания. Сигнальный пептид может быть сигнальным пептидом иммуноглобулина или гетерологичным сигнальным пептидом (т.е. сигнальным пептидом из неиммуноглобулинового белка).
Клетки-хозяева млекопитающих для экспрессии рекомбинантных антител, раскрытых в настоящем документе, включают ряд иммортализованных клеточных линий, доступных в Американской коллекции клеточных культур (АТСС). Они включают, в частности, клетки яичников китайского хомяка (СНО), NS0, клетки SP2/0, клетки HeLa, клетки почек детеныша хомяка (BHK), клетки почек обезьяны (COS), клетки гепатоцеллюлярной карциномы человека, клетки А549, клетки 293Т и многие другие клеточные линии. Клетки-хозяева млекопитающих включают клетки человека, мыши, крысы, собаки, обезьяны, свиньи, козы, коровы, лошади и хомяка. Особенно предпочтительные клеточные линии выбирают путем определения того, какая клеточная линия имеет высокий уровень экспрессии.
Когда рекомбинантный экспрессионный вектор, кодирующий тяжелую цепь или ее антигенсвязывающий фрагмент, или его фрагмент, легкую цепь и/или ее антигенсвязывающий фрагмент вводят в клетку-хозяина млекопитающего, антитело получают посредством культивирования клетки-хозяина в течение периода времени, достаточного для экспрессии антитела в этой клетке-хозяине, или более предпочтительно, путем секреции антитела в среду, в которой выращивают клетку-хозяина. Антитело может быть выделено из среды с использованием стандартных методов очистки белка.
Вполне вероятно, что антитела, экспрессируемые разными клеточными линиями или трансгенными животными, отличаются друг от друга гликозилированием. Однако все антитела, кодируемые представленными здесь молекулами нуклеиновых кислот или содержащие представленные здесь аминокислотные последовательности, являются неотъемлемыми частями настоящего изобретения, независимо от гликозилирования антитела. Аналогично, в некоторых воплощениях нефукозилированные антитела являются более полезными, поскольку они, как правило, обладают более высокой эффективностью in vitro и in vivo, чем их фукозилированные аналоги, и вряд ли будут иммуногенными, поскольку их гликановые структуры являются обычными компонентами природных человеческих сывороточных IgG.
Медицинское применение
Антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, имеет диагностическое и терапевтическое применение in vitro и in vivo. Предпочтительно, гуманизированные антитела hu31, hu43, hu44, hu59, hu60 или hu250 полезны для лечения заболеваний или расстройств, связанных с IL-17А.
В одном аспекте, выделенное антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, способны противодействовать возникновению индуцированного имиквимодом псориаза в мышиной модели и снижать клинический показатель возникновения псориаза у мышей и степень отечности ушей у мышей при оценке активности in vivo.
В конкретном воплощении, в модели индуцированного имиквимодом псориаза, гуманизированные антитела hu31 и hu44 могут значительно противодействовать возникновению у мышей и снижать клинический показатель возникновения псориаза у мышей и степень отечности ушей у мышей.
В одном аспекте, выделенное антитело или его антигенсвязывающий фрагмент, раскрытые здесь, способны ингибировать отечность коленного сустава в модели артрита, индуцированного антигеном, таких как AIA-модель яванского макака, при оценке in vivo.
В конкретном воплощении, в AIA-модели яванского макака, гуманизированное антитело hu31 значительно подавляет тенденцию к увеличению клинического показателя артрита у яванского макака
В одном аспекте настоящего изобретения предложен способ лечения патологических заболеваний, опосредованных IL-17A, включающий введение эффективного количества выделенного антитела или его антигенсвязывающего фрагмента в соответствии с настоящим изобретением, в частности антитела hu31, hu43, hu44, hu59, hu60 или hu250 для облегчения расстройства.
В одном воплощении выделенное антитело или белок, содержащий его антигенсвязывающий фрагмент, раскрытые в настоящем документе, конъюгирован с другой активной группировкой.
В одном воплощении выделенное антитело или белок, содержащий его антигенсвязывающий фрагмент, раскрытые в настоящем документе, может быть моноклональным антителом или его антигенсвязывающим фрагментом, предпочтительно химерным антителом, гуманизированным антителом или человеческим антителом, или его частью.
В аспектах настоящего изобретения предлагается фармацевтическая композиция, содержащая антитело или белок, содержащий его антигенсвязывающий фрагмент, согласно воплощениям, описанным в настоящем документе, в сочетании с одним или более фармацевтически приемлемыми эксципиентами, разбавителями или носителями.
В воплощениях фармацевтическая композиция содержит один или более дополнительных активных ингредиентов.
В одном конкретном воплощении фармацевтическая композиция представляет собой лиофилизированный порошок. В другом конкретном воплощении фармацевтическая композиция представляет собой стабильную жидкую композицию, содержащую терапевтически приемлемое количество антитела или молекулы, раскрытых в настоящем документе.
В частности, в настоящем изобретении предложен способ лечения расстройств, связанных с IL-17A, и/или аутоиммунных и воспалительных расстройств. В некоторых воплощениях способ включает введение субъекту, нуждающемуся в этом, выделенного антитела или его антигенсвязывающего фрагмента по настоящему изобретению.
В настоящем изобретении также предложен способ ослабления или ингибирования реакции передачи сигнала, индуцированного IL-17A или IL-17AF в клетке-мишени или ткани, путем контакта клетки с композицией, содержащей терапевтически эффективную дозу антитела, раскрытого в настоящем документе.
В настоящем изобретении термин "заболевание, опосредованное IL-17A" или "расстройство, связанное с IL-17A", включает все заболевания и состояния, в которых IL-17A или IL-17AF играют роль (прямо или косвенно), включая причину, развитие, прогрессирование, персистенцию или патологию заболевания или состояния. Таким образом, эти термины включают состояния, ассоциированные с или характеризующиеся аномальным уровнем IL-17A или IL-17AF, и/или заболевания или состояния, которые можно лечить путем ослабления или ингибирования IL-17А/АР-индуцированной активности (например CXCL1) в клетке-мишени или ткани. Такие заболевания или состояния включают воспалительные состояния и аутоиммунные заболевания, такие как артрит, ревматоидный артрит, анкилозирующий спондилит, рассеянный склероз или псориаз. Такие заболевания также включают аллергии и аллергические состояния, реакции гиперчувствительности, хроническую обструктивную болезнь легких, муковисцидоз и отторжение трансплантата органов или тканей.
При использовании здесь, "подавлять" или "лечить" или "лечение" включает задержку в развитии симптомов, ассоциированных с расстройством, и/или уменьшение тяжести симптомов таких расстройств. Этот термин также включает уменьшение интенсивности существующих неконтролируемых или вредных симптомов, предупреждение других симптомов и устранение или предупреждение основных причин таких симптомов. Таким образом, данный термин указывает на то, что полезный результат был обеспечивается позвоночному субъекту, страдающему или вероятно страдающему расстройством, болезнью или состоянием.
Термины "терапевтически эффективное количество", "терапевтически эффективная доза" и "эффективное количество", используемые в настоящем документе, относятся к раскрытому здесь количеству конъюгата IL-17, которое эффективно для предупреждения или улучшения одного или более симптомов заболевания или состояния, или развития заболевания или состояния при введении отдельно или в комбинации с другими терапевтическими агентами в клетку, ткань или субъекта. Терапевтически эффективная доза также относится к количеству конъюгата, достаточному для того, чтобы вызвать улучшение симптомов, например к количеству для лечения, излечения, предупреждения или улучшения соответствующего состояния или содействия лечению, излечению, предупреждению или улучшению такого состояния. Когда активный ингредиент вводят субъекту сам по себе, терапевтически эффективная доза относится к количеству этого ингредиента. При введении комбинации, терапевтически эффективная доза относится к объединенному количеству активных ингредиентов, которое оказывает терапевтический эффект, независимо от того, вводят эти активные ингредиенты в комбинации, последовательно или одновременно. Эффективное количество терапевтического агента приводит к увеличению диагностического индекса или параметра по меньшей мере на 10%, обычно по меньшей мере на 20%; предпочтительно по меньшей мере примерно на 30%, более предпочтительно по меньшей мере на 40%, наиболее предпочтительно по меньшей мере на 50%.
Ревматоидный артрит (RA)
RA прогрессирующее системное заболевание, характеризующееся воспалением синовиальных суставов, поражающее примерно 0,5% населения земного шара. См. Emery, (2006) BMJ 332:152-155. Воспаление суставов может привести к деформации, боли, скованности и разбуханию, и в конечном итоге к необратимой дегенерации суставов. Пораженные суставы включают коленные, локтевые, шейные суставы и суставы конечностей. Традиционные методы лечения включают симптоматическое лечение с помощью NSAID (нестероидные противовоспалительные средства), с последующим лечением противоревматическими лекарственными средствами, модифицирующими заболевание (DMRD), такими как золото, пеницилламин, сульфасалазин и метотрексат.Последние достижения включают лечение ингибиторами TNF-α, в том числе моноклональными антителами, такими как инфликсимаб, адалимумаб и голимумаб, и рецепторными слитыми белками, такими как этанерцепт.Лечение такими ингибиторами TNF-a значительно уменьшает структурные повреждения, вызванные заболеванием.
Антитела против IL-17A, раскрытые в настоящем документе, могут быть использованы для лечения RA у субъекта, нуждающегося в таком лечении. Антитела против IL-17A, раскрытые в настоящем документе, также могут быть объединены с другими методами лечения RA, такими как метотрексат, азатиоприн, циклофосфамид, этилмикофенолат, NSAID или ингибиторы TNF-α.
Псориаз
Кожа является важным барьером между внутренней и внешней средой, предотвращающим контакт с потенциально вредными антигенами. В случае инвазии антигена/патогена Т-клетки, полиморфноядерные клетки, макрофаги и т.п.в месте контакта с кожей локально инфильтрируются и инициируют воспалительную реакцию для устранения антигена (см., например, Williams and Kupper, (1996) Life Sci., 58:1485-1507). Как правило, эта воспалительная реакция, вызванная патогеном, находится под строгим контролем и прекращается, когда патоген устранен. В некоторых случаях такая воспалительная реакция возникает без внешнего раздражения и без надлежащего контроля, что приводит к воспалению кожи. В настоящем изобретении предложен способ лечения и диагностики такого воспаления кожи. Воспаление кожи (следствие вышеупомянутой клеточной инфильтрации и цитокинов, секретируемых клетками) включает несколько воспалительных состояний, таких как рубцевой пемфигоид, склеродермия, гнойный гидраденит, токсический эпидермальный некролиз, акне, остеит, болезнь "трансплантат против хозяина (GVHD)", гангренозная пиродермия и синдром Бехчета (см., например, Williams and Griffiths, (2002) Clin. Exp.Dermatol., 27:585-590). Наиболее распространенным воспалением кожи является псориаз.
Псориаз характеризуется гиперпролиферацией кератиноцитов, опосредованной Т-клетками, с воспалительной инфильтрацией. Заболевание имеет некоторые общие проявления, включая бляшечные поражения, сыпь и гнойничковые поражения (см., например, Gudjonsson et al., (2004) Clin. Exp.Immunol. 135:1-8). Примерно у 10% пациентов с псориазом может развиться артрит.Заболевание имеет сильную и сложную генетическую предрасположенность с 60% идентичностью у однояйцевых близнецов.
Типичные псориатические поражения представляют собой красные бляшки с четким краем, покрытые толстыми серебристыми чешуйками. Воспаление и гиперпролиферация псориатической ткани связаны с разными гистологическими, антигенными и цитокиновыми профилями по сравнению с нормальной кожей.
Цитокинами, ассоциированными с псориазом, являются: TNF-α, IL-19, IL-18, IL-15, IL-12, IL-7, IFN-γ, IL-17A и IL-23 (см. Gudjonsson et al., выше).
Антитела против IL-17A, раскрытые здесь, отдельно или в комбинации с другими агентами, также могут быть использованы для предупреждения, лечения, диагностики и прогнозирования возникновения псориаза.
Для получения фармацевтической или стерильной композиции IL-17A антитела, раскрытого в настоящем документе, антитело смешивают с фармацевтически приемлемым носителем или эксципиентом. См., например, Remington's Pharmaceutical Sciences and U.S. Pharmacopeia: National Formulary, Mack Publishing Company, Easton, PA (1984).
Следующие лекарственные формы терапевтических и диагностических средств могут быть получены путем смешивания антитела с приемлемым носителем, эксципиентом или стабилизатором: лиофилизированный порошок, паста, водный раствор или суспензия. В одном воплощении, антитело IL-17A, раскрытое здесь, разбавляют до подходящей концентрации раствором ацетата натрия (рН 5-6) и добавляют NaCl или сахарозу для регулировки осмоляльности. Для повышения стабильности могут быть добавлены другие вещества (например полисорбат 20 или полисорбат 80).
Режим дозирования зависит от нескольких факторов, включая оборот терапевтического антитела в сыворотке или тканях, тяжесть симптомов, иммуногенность терапевтического антитела и доступность клеток-мишеней в биологической матрице. Предпочтительно, режим дозирования обеспечивает достаточное количество терапевтического антитела для достижения улучшения состояния целевого заболевания при минимизации неблагоприятных побочных эффектов. Таким образом, количество доставляемых биологических агентов частично зависит от конкретного терапевтического антитела и тяжести состояния, подлежащего лечению. Имеются рекомендации по выбору соответствующих доз для терапевтических антител. Подходящие дозы могут быть определены врачом-клиницистом, например, с использованием параметров или факторов, известных или предполагаемых в этой области, как влияющие на лечение. Как правило, дозировка начинается с количества, немного меньшего оптимальной дозы, и впоследствии будет увеличиваться небольшими приращениями до тех пор, пока не будет достигнут желательный или оптимальный эффект по сравнению с любыми негативными побочными эффектами. Важные методы диагностики включают, например, методы диагностики, основанные на воспалительных симптомах или уровнях продуцируемых воспалительных цитокинов. Предпочтительно используют биологические агенты, полученные из того же вида, что и животное, для целевой терапии, тем самым сводя к минимуму воспалительные, аутоиммунные или пролиферативные реакции на агента. Например, в случае людей предпочтительными являются химерные, гуманизированные и полностью человеческие антитела.
Настоящее изобретение включает любые комбинации описанных конкретных воплощений. Дополнительные воплощения настоящего изобретения и полный объем применимости станут очевидными из подробного описания, представленного ниже. Однако следует понимать, что подробное описание и конкретные примеры, указывающие на предпочтительные воплощения настоящего изобретения, приведены только в качестве иллюстрации, поскольку различные изменения и модификации в пределах сущности и объема настоящего изобретения станут очевидными специалистам в данной области из подробного описания. Все публикации, патенты и патентные заявки, приведенные в настоящем документе, включая ссылки, включены посредством ссылки в полном объеме для всех целей.
ПРИМЕРЫ
Пример 1. Рекомбинантный человеческий белок IL-17A-mFc
Плазмида HG12047-G, содержащая последовательность кДНК, кодирующую полноразмерный человеческий IL-17A (регистрационный номер NCBI NP 002181.1), была приобретена у Sino Biological, Inc., и зрелый фрагмент человеческого IL-17A (аминокислоты 24-155 регистрационного номера NP 002181.1, аминокислотная последовательность SEQ ID NO: 66, нуклеотидная последовательность SEQ ID NO: 67) амплифицировали обычным методом ПЦР. Амплифицированный фрагмент переваривали с помощью BSPQI и клонировали в эукариотическую экспрессионную плазмидную систему (MXT1-Fc, содержащую домен Fc тяжелой цепи мышиного IgG), сконструированную собственными силами, тем самым получая экспрессионную плазмиду для рекомбинантного белка слияния IL-17A-mFc. Верифицированную плазмиду трансфицировали в экспрессирующую клетку 293F обычными методами, и после экспрессии и очистки был получен рекомбинантный человеческий белок IL-17A-mFc. На Фиг. 1 показана SDS-PAGE электрофореграмма рекомбинантного человеческого белка IL-17A-mFc.
Пример 2. Создание стабильных клеточных линий 293F, экспрессирующих человеческий IL-17RA
Плазмида HG10895-G, содержащая последовательность кДНК, кодирующую полноразмерный человеческий IL-17RA, была приобретена у Sino Biological, Inc. и последовательность ДНК, кодирующая полноразмерный человеческий IL-17RA, была амплифицирована с помощью обычной ПЦР (SEQ ID NO: 69). Амплифицированные фрагменты были клонированы в эукариотическую экспрессионную плазмидную систему (НХР), сконструированную собственными силами с помощью обычных методов клонирования и содержащую пуромициновую систему скрининга. Верифицированная рекомбинантная экспрессионная плазмида для IL-17RA была трансфицирована в клетки 293F (АТСС). Через 24 ч после трансфекции клетки отбирали с помощью пуромицина (2 мкг/мл) с получением стабильных клеток IL-17RA 293F. Отдельные клоны выделяли обычными методами, например клонированием с методом серийных разведении, и переносили в 96-луночные планшеты по 0,8 клетки на лунку. Через 15 суток моноклоны IL-17RA-293F отбирали и пересевали с получением стабильных клеточных линий IL-17RA 293F. Все клоны были подвергнуты скринингу с помощью FACS (флуоресцентного сортинга) или тому подобного, и клоны с наивысшим уровнем экспрессии были отобраны с помощью анализа связывания FACS для гибридомных моноклональных антител или для использования в функциональных анализах.
Пример 3. Связывание рекомбинантного белка IL-17A-mFc со стабильными клеточными линиями IL-17RA 293F
Специфичность связывания рекомбинантного белка IL-17A-mFc с IL-17RA на клетках 293F определяли с помощью FACS. В кратком изложении, клетки (стабильные клеточные линии IL-17RA 293F) готовили в виде суспензий клеток в концентрации 1×10б/мл и добавляли в 96-луночные планшеты в количестве 20 мкл/лунку с фактическим числом клеток 2×104/лунка. Рекомбинантный белок IL-17A-mFc (3 мкг/мл, 20 мкл/лунка; группа лечения) или 1% BSA (20 мкл/лунка, группа отрицательного контроля) смешивали с клеточной суспензией, и после инкубации при 37°С в течение 30 минут клетки промывали буфером FACS 3 раза. Был добавлен анти-мышиный IgG (1:200), и клетки инкубировали при комнатной температуре в течение 30 минут.Клетки промывали 3 раза буфером FACS и определяли с помощью проточного цитометра среднюю интенсивность флуоресценции (MFI) групп. Как показано на Фиг. 2, рекомбинантный белок IL-17A-mFc может специфически связываться с IL-17RA на клетках 293F.
Пример 4. Получение и скрининговый анализ гибридомных антител
Гибридомные антитела получали с использованием стандартных молекулярно-биологических методов. В кратком изложении, нативный человеческий белок IL-17A, купленный у HumanZyme, в качестве антигена смешивали с равным количеством иммуноадъюванта. 5 самок мышей FVB в возрасте 6 недель были иммунизированы. Одну бустер-иммунизация проводили еженедельно после первичной иммунизации, в общей сложности было проведено семь иммунизаций. После последней бустер-иммунизации для слияния клеток были отобраны мыши с высокими титрами антител против IL-17A в сыворотке. Клетки селезенки были выделены и слиты с клетками мышиной миеломы SP2/0 (АТСС) посредством стандартных гибридомных методов. Слитые клетки ресуспендировали в полной среде RPMI-1640, содержащей HAT, и помещали в лунки с питающим подслоем перитонеальных клеток.
Секреторные супернатанты моноклональной гибридомы были идентифицированы на основе первоначально желательных характеристик связывания антител/антигенов (таких как сродство к IL-17A, способность блокировать связывание IL-17A с его рецептором, перекрестная реактивность и способность блокировать опосредованные IL-17A биологические эффекты in vitro). Антитела в супернатанте от гибридом 1F8, 2В2, 2F5, 2F2, 2Н1 и 2Н5 использовали для дальнейшей характеристики.
Пример 5. Эффект гибридомных антител на блокировании биологической активности IL-17A in vivo
Большое количество исследований показывает, что IL-17A способствует экспрессии и высвобождению цитокина CXCL1 in vivo. Таким образом, изменение экспрессии CXCL1 в сыворотке мыши может быть количественно обнаружено с помощью ELISA, чтобы определить влияние гибридомных антител на IL-17А-опосредованную биологическую активность у мышей. В кратком изложении, 40 самок мышей Balb/c в возрасте 10 недель были отобраны и разделены на 8 групп по 5 мышей в каждой. За 4 суток до введения собирали сыворотку и измеряли экспрессию CXCL1 в качестве исходного уровня. В сутки введения гибридомные антитела-кандидаты, физиологический раствор (контроль) или эталонное антитело mAb317 (имеющееся в продаже антитело против IL-17A, от R&D) вводили внутрисердечно в дозе 1 мг/кг.Через 1 час после введения нативный человеческий IL-17A (HumanZyme) вводили подкожно в дозе 150 мкг/кг; через 2 часа после введения человеческого IL-17A собирали сыворотку, измеряли концентрацию CXCL1 в крови и сравнивали с исходным уровнем. Для каждой группы рассчитывали кратное изменение (среднее значение ± стандартная ошибка (среднее значение ±SEM)) концентрации CXCL1 до и после введения. Сравнительный анализ между группами лечения и контрольной группой считается значимо отличающимся, когда Р составляет <0,05 в t-тесте студента, *Р<0,05, **Р<0,01, ***Р<0,001.
Как показано на Фиг. 3, и гибридомные антитела, полученные в Примере 4, и имеющееся в продаже антитело mAb317 могут значительно ингибировать IL-17A, индуцирующий экспрессию CXCL1 у мышей.
Пример 6. Определение последовательностей вариабельных областей антител-кандидатов
Последовательности ДНК вариабельных областей антител, экспрессируемых гибридомами 1F8, 2В2 и 2F5, определяли с помощью ПЦР с использованием дегенеративных праймеров. В кратком изложении, гибридомные клеточные линии 1F8, 2В2 и 2F5 размножали и собирали центрифугированием при 1000 об/мин. Общую РНК экстрагировали с использованием тризола. Используя общую РНК в качестве матрицы, синтезировали кДНК первой нити. Последовательности ДНК соответствующих вариабельных областей амплифицировали методом ПЦР с использованием кДНК первой нити в качестве последующей матрицы. Используемые ПЦР-праймеры были основаны на совокупности Ig-праймеров. Продукты ПЦР собирали, очищали, секвенировали и анализировали с получением последовательностей вариабельных областей тяжелых и легких цепей гибридомных антител-кандидатов. Гибридомная клеточная линия 1F8 продуцировала две последовательности генов вариабельной области легкой цепи антител и одну последовательность генов вариабельной области тяжелой цепи антител, a 2F5 продуцировала две последовательности генов вариабельной области тяжелой цепи антител и одну последовательность генов вариабельной области легкой цепи антител. Следовательно, каждое из антител, секретируемых 1F8 и 2F5, может включать два интактных антитела, где антитела, секретируемые 1F8, были обозначены как 1F8-1 и 1F8-2, а антитела, секретируемые 2F5, были обозначены как 2F5-1 и 2F5-2.
Аминокислотные последовательности вариабельных областей тяжелой и легкой цепей антител, экспрессируемых 1F8, 2F5, 2В2, показаны в Таблице 1 (см. ПОДРОБНОЕ ОПИСАНИЕ).
Пример 7. Конструирование экспрессионных векторов рекомбинантных химерных антител
Fc-фрагмент константной области тяжелой цепи человеческого IgG4 и константную область легкой цепи каппа клонировали из человеческих клеток крови (Beijing Blood Institute) и лигировали с помощью плазмиды pCDNA3.1 для конструирования. Фрагменты последовательностей вариабельной области тяжелой цепи и легкой цепи были синтезированы с помощью GenScript. Тяжелые и легкие цепи были расщеплены Bspq I, а затем лигированы с соответственно модифицированной плазмидой pcDNA3.1. Экспрессионные плазмиды химерной тяжелой цепи IgG4 (ch-HC) или легкой цепи (ch-LC) верифицировали путем секвенирования. Экспрессионные плазмиды для разных химерных тяжелых и легких цепей спаривали для трансфекции экспрессирующих клеток с получением 16 химерных антител с номерами от ch1 до ch16 (см. Таблицу 6). Все последующие экспериментальные материалы были получены с помощью экспрессирующих клеток, трансфицированных этими плазмидами.
Пример 8. Специфичность связывания химерных антител с человеческим IL-17A Специфичность связывания химерных антител с человеческим IL-17A определяли посредством обычного ELISA. А именно, 0,5 мкг/мл человеческого IL-17A-mFc иммобилизовали на 96-луночном планшете и инкубировали в течение 60-90 минут при 37°С. Раствор в лунках затем отбрасывали, и лунки 3 раза промывали промывочным буфером и блокировали в течение 60 минут раствором PBS, содержащим 2% BSA. Планшет 3 раза промывали промывочным буфером и добавляли разбавленные химерные антитела в разных концентрациях. Антитела инкубировали при 37°С в течение 60 минут.Затем планшет 3 раза промывали промывочным буфером и добавляли 10000-кратно разбавленное биотинилированное антитело против IgG4. Систему инкубировали при 37°С в течение 1 часа, трижды промывали промывочным буфером, добавляли HRP-Strep, 10000-кратно разбавленный промывочным буфером, и инкубировали при комнатной температуре в течение 1 часа. После 3 промывок промывочным буфером для проявления цвета добавляли 100 мкл субстрата ТМВ (тетраметилбензидин). Систему инкубировали при комнатной температуре в течение 30 минут, и реакцию завершали добавлением 100 мкл 2 М раствора соляной кислоты. Поглощение при 450 нм измеряли с помощью планшет-ридера.
Как показано на Фиг. 4, химерные антитела ch1, ch2, ch4, ch7 и ch16 связываются с человеческим IL-17A с более высокой специфичностью, с ЕС50 химерных антител, составляющей 6,62 нг/мл, 5,17 нг/мл, 88,48 нг/мл, 39,96 нг/мл и 15,42 нг/мл соответственно.
Пример 9. Блокирование связывания IL-17A человека с IL-17RA с помощью химерных антител
Блокирование связывания IL-17A с IL-17RA на клетках определяли при помощи анализа на основе конкурентной клеточной проточной цитометрии (FACS). В кратком изложении, разведения химерных антител с разными концентрациями (3-кратно разведенные 10 мкг/мл) смешивали с предварительно биотинилированным человеческим IL-17A-mFC (3 мкг/мл), полученным в примере 1, и инкубировали при комнатной температуре в течение 30 минут. Затем смесь инкубировали совместно с суспензией клеток (стабильная клеточная линия IL-17RA 293F, полученная в Примере 2, 1,5×105 клетки/лунка) при 37°С в течение 15 минут. После 3 промывок PBS добавляли 5 мкг/мл анти-мышиного IgG и систему инкубировали при комнатной температуре в течение 30 минут.После 3 промывок PBS химерные антитела анализировали методом проточной цитометрии в отношении ингибирования связывания IL-17A с IL-17RA на поверхности клеток 293F.
Как показано на Фиг. 5, химерные антитела ch1, ch2, ch7 и ch16 могут значительно ингибировать связывание человеческого IL-17 с IL-17RA на поверхности клеток 293F. Совместно с другими экспериментальными результатами ch1 и ch16 были отобраны для дальнейшей гуманизации.
Пример 10. Гуманизация антител
Для гуманизации антител гены IgG человека, гомологичные последовательности кДНК вариабельных областей антител мышей, были получены в базе данных генов иммуноглобулинов человека NCBI (http://www.ncbi.nlm.nih.gov/igblast/). Аминокислотные последовательности CDR вариабельных областей и их границы затем определяли в соответствии с системой нумерации Kabat или системой нумерации IMGT. Человеческие IGVH и IGVk с высокой гомологией с вариабельными областями мышиного антитела были выбраны в качестве матриц для гуманизации и были гуманизированы путем прививки CDR. В кратком изложении, гуманизация включает следующие этапы: А. последовательности генов антител-кандидатов сравнивали с последовательностью генов антител зародышевой линии человека для отбора последовательностей с высокой гомологией; В. с помощью анализа аффинности HLA-DR выбирали последовательность каркаса зародышевой линии человека с низкой аффинностью; и С.каркасные аминокислотные последовательности вариабельных областей и их периферию проанализировали с помощью компьютерного моделирования и молекулярного докинга, и верифицировали пространственное и стерео-расположение. Ключевые аминокислотные единицы, которые взаимодействуют с IL-17A и поддерживают пространственный каркас в последовательности генов антител-кандидатов, были проанализированы путем расчета электростатической силы, силы Ван-дер-Ваальса, гидрофильности и гидрофобности, а также значения энтропии, и привиты на выбранный каркас зародышевой линии человека. На карту должны быть нанесены аминокислотные положения каркасных участков, которые необходимо сохранить. После этого гуманизированные антитела были синтезированы.
Схема определения CDR вариабельных областей мышиных антител и их аминокислотные последовательности приведены в Таблице 3 Подробного описания изобретения.
Основываясь на вышеупомянутых результатах, химерные антитела ch1 и ch16 были отобраны для гуманизации. После первичного скрининга ряда антитело/антигенсвязывающих свойств (таких как аффинность связывания с IL-17A, способность блокировать связывание IL-17A с его рецептором), гуманизированные антитела hu31, hu43, hu44, hu59, hu60 и hu250 были отобраны для последующей проверки.
Вариабельные области гуманизированных антител hu31, hu43, hu44, hu59, hu60 и hu250 и аминокислотные последовательности их CDR показаны в Таблице 4 Подробного описания изобретения.
Пример 11. Специфичность связывания гуманизированных антител с человеческим IL-17A
Специфичность связывания гуманизированных антител с человеческим IL-17A определяли с помощью обычного ELISA. Метод и методики описаны в Примере 8. Как показано на Фиг. 6, гуманизированные антитела hu31, hu43, hu44, hu59, hu60 и hu250 специфически связываются с IL-17A. ЕС50 составлял 8,13 нг/мл, 8,64 нг/мл, 6,76 нг/мл, 6,10 нг/мл, 5,78 нг/мл и 6.35 нг/мл соответственно.
Пример 12. Блокирование связывания человеческого IL-17A с IL-17RA при помощи гуманизированных антител
Блокирование связывания IL-17A с IL-17RA на клетках обнаруживали посредством анализа методом конкурентной клеточной проточной цитометрии (FACS). Метод и методики описаны в Примере 9. Как показано на Фиг. 7, гуманизированные антитела hu31, hu43, hu44, hu59, hu60 и hu250 значительно ингибируют специфическое связывание IL-17A с IL-17RA на клетках. Величина IC50 составляла 867,6 нг/мл, 780,8 нг/мл, 828,5 нг/мл, 467,4 нг/мл, 482,8 нг/мл и 577,8 нг/мл соответственно.
Пример 13. Антагонизм в отношении индуцированной IL-17A экспрессии CXCL1 в эпителиальных клетках с помощью гуманизированных антител
IL-17A может стимулировать экспрессию и высвобождение цитокина CXCL1 в различных эпителиальных клетках и других клетках. Изменение уровня экспрессии CXCL1 в супернатанте клеток может быть количественно определено с помощью ELISA для определения влияния гуманизированных антител на биологические активности, опосредованные IL-17A, в клетках.
Клетки НТ-29 (эпителиальные клетки аденокарциномы толстой кишки человека, АТСС) инкубировали в среде для культивирования/анализа в колбах, обработанных тканевыми культурами в соответствии со стандартными методами. НТ-29 выращивали в колбах, обработанных тканевыми культурами, до тех пор, пока они не достигли слияния 50-80% в день анализа. В день анализа клетки промывали PBS и отделяли от колбы трипсином + ЭДТА с получением клеточной суспензии. Разведения (3-кратное последовательное разведение исходных 55 мкг/мл) гуманизированных антител hu31, hu59, hu60, hu250 или эталонного антитела (секукинумаб, Novartis) смешивали с человеческим IL-17A (1 мкг/мл), переносили на 96-луночные планшеты и инкубировали в течение 1 часа. 100 мкл суспензии (2×104 клеток) клеток НТ-29 (АТСС, эпителиальных клеток колоректальной аденокарциномы человека) добавляли в каждую лунку и инкубированы при 37°С/7% СО2 в течение 48 ч. Клетки центрифугировали при 500xg в течение 5 мин, и супернатант переносили на новые 96-луночные планшеты. Экспрессию CXCL1 определяли с помощью набора ELISA.
Как показано на Фиг. 8, гуманизированные антитела hu31, hu59, hu60 и hu250 демонстрировали более сильные антагонизм IL-17 А-стимуляции высвобождения CXCL1 в эпителиальных клетках, чем эталонное антитело секукинумаб.
Пример 14. Антагонизм экспрессии CXCL1, индуцированной IL-17A, у мышей, с помощью гуманизированных антител
Влияние гуманизированных антител на IL-17А-опосредованную биологическую активность in vivo определяли путем измерения изменений уровней CXCL1 в сыворотке крови мышей, как описано в Примере 5. В кратком изложении, 40 самок мышей Balb/c в возрасте 10 недель были отобраны и разделены на 8 групп по 5 мышей в каждой. За 4 суток до введения собирали сыворотку и измеряли экспрессию CXCL1 в качестве исходного уровня. В сутки введения внутрикардиально вводили антитела-кандидаты (гуманизированные антитела hu31, hu43, hu44, hu60 и hu250) или контроль изотипа IgG4 (hlgG) в дозе 1 мг/кг.Через 1 час после введения человеческий IL-17A вводили подкожно в дозе 150 мкг/кг; через 2 часа после введения человеческого IL-17A собирали сыворотку, измеряли концентрацию CXCL1 в крови и сравнивали с исходным уровнем. Кратное изменение (среднее значение ± стандартная ошибка (среднее значение ± SEM)) концентрации CXCL1 до и после введения рассчитывали для каждой группы. Сравнительный анализ между антителами-кандидатами и контролем изотипа IgG4 считается значимо различным, когда Р составляет <0,05 в t-тесте Стьюдента, *Р<0,05, **Р<0,01, ***Р<0,001.
Как показано на Фиг. 9, гуманизированные антитела-кандидаты hu31, hu43, hu44, hu60 и hu250 демонстрировали более мощный антагонизм IL-17А-опосредованной стимуляции высвобождения CXCL1 в эпителиальных клетках, чем контроль изотипа IgG4.
Пример 15. Исследованием эффективности гуманизированных антител в улучшении индуцированного имиквимодом псориаза у мышей
Имиквимод, введенный на кожу уха и спины мыши, может индуцировать псориазо-подобные патологические характеристики, включая гиперпролиферацию кератиноцитов, агрегацию воспалительных клеток, гиперплазию сосочковых сосудов кожи и тому подобное, таким образом, создается мышиная модель с псориазом. Эффективность антител на мышах с псориазом определяли по конечным результатам, таким как клинические показатели, степень отечности уха и тому подобное.
15.1. Методология
Были отобраны 48 самок мышей C57BL/6 в возрасте 6-8 недель (приобретенные в Центре исследований модельных животных Нанкинского университета, сертификат №201605578). Волосы мышей на спине были выбриты, и мышей, не входящие в группу плацебо-хирургии, сенсибилизировали через двое суток. За двое суток до сенсибилизации мышей рандомизировали на 5 групп (по 8 в каждой): группа I была группой плацебо-хирургии; группа II была группой PBS, группа III была контрольной группой KLH (изотип IgG), получавшей KLH (гемоцианин лимфы улитки); группа IV была группой лечения hu31; группа V была группой лечения hu43; группа VI была группой лечения hu44. Доза для всех групп составляла 50 мг/кг, и вышеуказанным группам вводили соответствующие лекарственные средства внутрибрюшинно один раз на 0-е и 3-е сутки. В сутки сенсибилизации (сутки 1), мышам в группах II-VI вводили примерно 62,5 мг крема имиквимода (Aldara, 5%, 3М Health Саге Limited) на кожу правого уха и спины в течение 4 суток подряд.
15.2. Оценка
Толщину правого уха мыши измеряли ежедневно микрометром, начиная со дня сенсибилизации. Вместе с толщиной уха в первые сутки 1, в качестве исходной отметки, записывали толщину отечности уха. Мышей ежедневно взвешивали, наблюдали за шелушением кожи, уплотнением и покраснением, и классифицировали по 4-балльной шкале: 0 - нет; 1 - легкая; 2 - умеренная; 3 - тяжелая; 4 - серьезная. Результаты записывали в виде среднего ± стандартная ошибка (среднее ± SEM). Различия между группами выявляли с помощью однофакторного дисперсионного анализа (ANOVA) и подтверждали t-критерием Стьюдента, где Р<0,05 указывает на значимую разницу.
Как показано на Фиг. 10-1, введение антител-кандидатов, раскрытых здесь, может значительно ингибировать шелушение кожи, уплотнение, отечность и другие состояния индуцированного имиквимодом псориаза в мышиной модели, демонстрируя снижение баллов.
Как показано на Фиг. 10-2, крем имиквимода, вводимый в правое ухо мыши, начиная со дня сенсибилизации, вызывал сильную отечность и увеличил толщину правого уха, в то время как гуманизированное антитело по настоящему изобретению значительно улучшало отечность уха.
В мышиной модели с имиквимод-индуцированным псориазом, гуманизированные антитела, раскрытые в данном документе, очевидным образом могут противостоять заболеваемости мышей, демонстрируя снижение клинического показателя мышей и степень отечности уха.
Пример 16. Исследование эффективности гуманизированного антитела в улучшения артрита, индуцированного коллагеном II типа у самок яванского макака
Животные с артритом, индуцированным коллагеном II типа, являются моделями, обычно используемыми в исследованиях ревматоидного артрита (RA). Такие модели имеют те же гистопатологические особенности, что и RA человека, и характеризуются воспалением в фасеточных суставах и прогрессирующей эрозией хрящей и костей. Человеческие/гуманизированные биологические макромолекулы, включая антитела, обычно обладают лучшей перекрестной реактивностью с антигенами яванского макака. Таким образом, модель яванского макака с артритом является эффективной системой для анализа противоревматической эффективности гуманизированного антитела IL-17A, раскрытого здесь. В эксперименте оценивали эффективность антител-кандидатов у яванского макака с артритом.
16.1. Методология
Коллаген бычьего типа II (CII, Сычуаньский университет) растворяли в уксусной кислоте (САТ# 10000218; Sinopharm, Шанхай, Китай), перемешивали в течение ночи в холодильнике при температуре 4°С и эмульгировали равным объемом полного адъюванта Фрейнда (САТ# F5881, Sigma-Aldrich, США) с получением коллагеновой эмульсии с конечной концентрацией 2 мг/мл. В 0-е сутки животных анестезировали золетилом (1,5-5 мг/кг, внутримышечно) и вводили коллагеновую эмульсию в несколько мест на спине и хвосте с 1,5% -5% изофлурана для поддержания анестезирующего эффекта по мере необходимости. Животным повторно вводили коллаген через 3 недели (21-е сутки) тем же методом, что и раньше. Затем животных делили на 4 группы. G1 представляла собой нормальную группу без индукции артрита; G2 представляла собой контрольную группу носителя; G3 представляла собой группу лечения антителом hu31; и G4 представляла собой группу лечения антителом hu59. Животных с баллом в 5% от максимального клинического балла (192х5%≈10) последовательно разделяли на группы, пока все животные, отвечающие этому требованию, не были распределены. После распределения препарат вводили при помощи инфузионного насоса в дозе 7,5 мг/кг в течение 30 минут один раз в неделю в течение 5 недель.
16.2. Оценка
Масса тела: Массу тела животных измеряли за сутки до индукции артрита и один раз в неделю после этого до конца исследования.
Оценка артрита: обезьян классифицировали по воспалению в 0-е и 21-е сутки и один раз в неделю после 21-ых суток до конца исследования (если наблюдалось раннее начало, еженедельная оценка должна начинаться соответственно). Критерии оценки приведены в Таблице 2. Было оценено 15 суставов в каждой лапе: 5 пястно-фаланговых суставов (МСР), 4 проксимальных межфаланговых сустава (PIP), 5 дистальных межфаланговых суставов (DIP), 1 лучезапястный сустав и 1 голеностопный сустав. Также желательно оценить тяжесть поражения коленных/локтевых суставов конечностей. Сумма индивидуальных баллов по суставам была оценкой артрита у животного, с максимальным баллом 192 (16×3×4). Критерии оценки артрита: 0 - нормальный; 1 - легкий артрит с небольшим, но заметным поражением; 2 - умеренная отечность; 3 - тяжелый артрит с тяжелой отечностью или очевидной деформацией сустава.
Экспериментальные данные представлены в виде среднего ± стандартная ошибка (среднее значение ± S. Е. М). Суммировали различия в параметрах между контрольной группой носителя, группой эталонного лекарственного средства и группой испытуемого образца, где р<0,05 указывает на статистическое различие (однофакторный дисперсионный анализ ANOVA/Даннетт).
Как показано на Фиг. 11-1, масса тела яванского макака в нормальной группе (G1) была стабильной; у яванского макака с индуцированным артритом средний вес в контрольной группе носителя (G2) постоянно снижался, и снижения массы тела у обезьян, получавших антитела hu31 и hu59, контролировались. Таким образом, в условиях данного исследования hu31 и hu59 в некоторой степени улучшают потерю массы тела, вызванную артритом (**Р<0,01, ****р<0,0001, по сравнению с "G2: группой плацебо"; Однофакторный дисперсионный анализ ANOVA/Даннетт).
Как показано на Фиг. 11-2, клинические баллы артрита у животных в нормальной группе остаются 0 после распределения; балл артрита у животных в контрольной группе с носителем (G2) постепенно повышался, в то время как исследуемые антитела hu31 и hu59 значительно подавляли тенденцию к увеличению клинического балла артрита. Таким образом, антитела hu31 и hu59 демонстрировали ингибирующее действие на прогрессирование артрита, и hu31 значительно подавлял тенденцию к увеличению клинического балла артрита у яванского макака (***р<0,001, #Р<0,05, по сравнению с G2: группа с носителем; Однофакторный дисперсионный анализ NAOVA/Даннетт).
Пример 17. Эффективность гуманизированных антител при опухании суставов у мышей, индуцированном клетками NIH3T3-IL-17
1. Животные: C57BL/6, самки, 6-8 недель, Beijing Vital River Laboratory Animal Technology Co., Ltd.
2. Клетки: клетки NIH3T3, клетки NIH3T3, экспрессирующие человеческий IL-17.
3. Группы и режим дозирования:
Группа NIH3T3;
Группа NIH3T3-IL-17 + эталонное антитело IgG (30 мг/кг);
Группа NIH3T3-IL-17 + высокая доза лечебного антитела (hu31, 3 мг/кг);
Группа NIH3T3-IL-17 + средняя доза лечебного антитела (hu31, 10 мг/кг);
Группа NIH3T3-IL-17 + низкая доза лечебного антитела (hu31, 30 мг/кг);
Группа NIH3T3-IL-17 + положительное лекарственное средство (Cosentyx, 10 мг/кг).
4. Моделирование и введение:
Клетки NIH3T3-IL-17 и контрольные клетки NIH3T3 (2,5×105 клеток/мышь, с объемом инъекции 25 мкл) были введены в полость правого голеностопного сустава мышей.
Внутрибрюшинные инъекции антител hu31 (3, 10, 30 мг/кг) и Cosentyx (10 мг/кг) вводили один раз в 3 суток, начиная с 1 суток до выбора модели. Была изучена эффективность инъекции антитела hu31 в модели мыши с артритом.
5. Детектирование:
Толщину лодыжки измеряли штангенциркулем с нониусом и рассчитывали степень отечности.
6. Результаты:
Как показано на Фиг. 12, после того, как клетки NIH3T3, экспрессирующие hIL-17, были введены в полость сустава мышей, на следующий день наблюдалась тяжелая отечность сустава. Скорость ингибирования отечности рассчитывали в соответствии с толщиной лодыжки мыши, при этом формула расчета выглядит следующим образом: степень ингибирования (%) = (толщина лодыжки группы NIH3T3-IL-17 IgG толщина лодыжки группы лечения) / (толщина лодыжки группы NIH3T3-IL-17 IgG толщина лодыжки группы NIH3T3) × 100. Результаты показали, что на 2-е сутки после введения наблюдалось ингибирование отечности лодыжки у мышей во всех группах лечения до конца исследования, а максимальный ингибирующий эффект наблюдался на 6-е сутки после введения. На 10-е сутки уровни ингибирования для антител hu31 при 3, 10 и 30 мг/кг составляли 49,4%, 65,9% и 74,1% соответственно. Уровень ингибирования отечности с помощью Cosentyx (10 мг/кг) составлял 67,1%. Средние показатели ингибирования были рассчитаны в разные сутки в группах лечения и результаты показали, что средние показатели ингибирования составили 45,2%, 57,0% и 73,9% для групп лечения 3, 10 и 30 мг/кг соответственно. Средний уровень ингибирования отечности с помощью Cosentyx (10 мг/кг) составлял 60,4%. Результаты показали, что антитело hu31 может ингибировать IL-17-индуцированную отечность лодыжки у мышей дозо-зависимым образом и эффективность при 10 мг/кг эквивалентна положительному лекарственному средству Cosentyx (10 мг/кг). Эффективность при 30 мг/кг превосходит эффективность эталонного средства Cosentyx (10 мг/кг).
Пример 18. Эффективность гуманизированных антител в модели воздушного мешка у мышей при воспалении, индуцированном клетками NIH3T3-IL-17
1. Животные: C57BL/6, самцы, 6-8 недель, приобретены у компании Beijing Vital River Laboratory Animal Technology Co., Ltd.
2. Клетки: клетки NIH3T3, клетки NIH3T3, экспрессирующие человеческий IL-17.
3. Реагент: антитело Grl-FITC, BioLegend
4. Группы и режим дозирования:
Группа NIH3T3
Группа NIH3T3-IL-17 + эталонное антитело IgG (30 мг/кг);
Группа NIH3T3-IL-17 + высокая доза лечебного антитела (hu31, 30 мг/кг);
Группа NIH3T3-IL-17 + средняя доза лечебного антитела (hu31, 10 мг/кг);
Группа NIH3T3-IL-17 + низкая доза лечебного антитела (hu31, 3 мг/кг);
Способ введения: внутрибрюшинно.
5. Моделирование:
Воздушный мешок: Мышам вводили 2,5 мл воздуха на спину в 0 и 3-е сутки. Начиная с 5-ых суток в воздушный мешок вводили клетки. Каждая мышь получала 2×105 клеток/500 мл PBS. Мышей распределяли по группам, каждая из которых содержала 8 мышей.
6. Детектирование:
Лейкоциты мигрировали в воздушный мешок. Клетки в промывочной жидкости подсчитывали и долю клеток Grl+ определяли посредством проточной цитометрии для расчета количества нейтрофилов.
Количество нейтрофилов = общее количество клеток х доля клеток Grl+ 7.
7. Результаты
Как показано на Фиг. 13, после того, как мышам с воздушным мешком вводили клетки NIH3T3, экспрессирующие hIL-17A, по сравнению с группой NIH3T3, количество инфильтрирующих лейкоцитов в воздушном мешке группы NIH3T3-IL-17 значительно увеличивалось, а доля и количество клеток Grl+ также были значительно повышенными, что указывает на успешный выбор модели. В день моделирования антитело hu31 (3, 10 и 30 мг/кг) вводили внутрибрюшинно. Уровень ингибирования рассчитывали в соответствии с общим количеством инфильтрирующих клеток и количеством клеток Grl+, при этом формула расчета выглядит следующим образом: уровень ингибирования (%) = (количество клеток группы NIH3T3-IL-17-IgG -количество клеток в группе лечения)/(количество клеток группы NIH3T3-IL-17-IgG - количество клеток группы NIH3T3) × 100. Результаты показали, что общее количество инфильтрирующих клеток в группах лечения антителом hu31 (3, 10 и 30 мг/кг) было ингибировано на 50,0%, 56,7% и 78,3% соответственно. Был рассчитан уровень ингибирования количества клеток Grl+, демонстрирующий, что: средние уровни ингибирования клеток Grl+ во всех группах лечения антителом hu31 (3, 10 и 30 мг/кг) составляли 59,1%, 54,5% и 81,8% соответственно. Антитело hu31 может ингибировать общую клеточную инфильтрацию, индуцированную IL-17, и воспалительную клеточную инфильтрацию у мышей дозозависимым образом.
--->
Список последовательностей
<110> SHANGHAI JUNSHI BIOSCIENCES CO., LTD.
SUZHOU JUNMENG BIOSCIENCES CO., LTD.
<120> ANTI-IL-17A ANTIBODY AND USE THEREOF
<130> 199971PCWO
<150> CN 201811515045.7
<151> 2018-12-12
<160> 69
<170> SIPOSequenceListing 1.0
<210> 1
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR1
<400> 1
Gly Tyr Ile Phe Thr Asn Tyr Gly
1 5
<210> 2
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR2
<400> 2
Ile Asn Asp His Thr Gly Glu Pro
1 5
<210> 3
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR3
<400> 3
Ala Asn Tyr Gly Phe Gly Tyr Phe Asp Tyr
1 5 10
<210> 4
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR1
<400> 4
Asn Tyr Asp Ile His
1 5
<210> 5
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR2
<400> 5
Val Ile Trp Ala Gly Gly Arg Thr His Ser Asp Ser Ala Leu Met Ser
1 5 10 15
<210> 6
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR3
<400> 6
Glu Gly Gly Asp Tyr Tyr Lys Tyr Leu Asp Phe
1 5 10
<210> 7
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR1
<400> 7
Asp Thr Tyr Ile His
1 5
<210> 8
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR2
<400> 8
Arg Ile Asp Pro Ala Asn Gly Asn Thr Lys Tyr Asp Pro Lys Phe Gln
1 5 10 15
Gly
<210> 9
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR3
<400> 9
Asp Tyr Tyr Gly Ser Ser Leu Leu Asp Tyr
1 5 10
<210> 10
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR1
<400> 10
Ser Asp Tyr Ala Trp
1 5
<210> 11
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR2
<400> 11
Phe Ile Ser Tyr Ser Gly Pro Thr Ser Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 12
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR3
<400> 12
Gly Gly Asp Gly Asp Ser Phe Asp Tyr
1 5
<210> 13
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR1
<400> 13
Gln Ser Leu Val His Ser Asn Gly Asn Ser Tyr
1 5 10
<210> 14
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR2
<400> 14
Lys Val Ser
1
<210> 15
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR3
<400> 15
Ser Gln Ser Thr His Val Pro Tyr Thr
1 5
<210> 16
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR1
<400> 16
Arg Ser Gly Gln Ser Leu Val His Ser Asn Gly His Thr Tyr Phe His
1 5 10 15
<210> 17
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR2
<400> 17
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 18
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR3
<400> 18
Ser Gln Ser Thr His Val Pro Tyr Thr
1 5
<210> 19
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR1
<400> 19
Lys Ala Ser His Asp Ile Asn Ser Tyr Leu Ser
1 5 10
<210> 20
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR2
<400> 20
Arg Ala Asn Gly Leu Val Asp
1 5
<210> 21
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR3
<400> 21
Leu Gln Tyr Glu Met Leu Pro Leu Thr
1 5
<210> 22
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR1
<400> 22
Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr Leu Asp
1 5 10 15
<210> 23
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR2
<400> 23
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 24
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR3
<400> 24
Phe Gln Gly Ser His Phe Pro Thr
1 5
<210> 25
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 25
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Met Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Asp His Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Lys Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Asn Tyr Gly Phe Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 26
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 26
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ile Asn Tyr
20 25 30
Asp Ile His Trp Leu Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ala Gly Gly Arg Thr His Ser Asp Ser Ala Leu Met
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Asn Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Asn Val His Ser Leu Arg Pro Glu Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Glu Gly Gly Asp Tyr Tyr Lys Tyr Leu Asp Phe Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 27
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 27
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Thr Lys Tyr Asp Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Thr Gly Asp Lys Ser Ser Asn Ala Ala Tyr
65 70 75 80
Leu Gln Leu Ser Arg Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Thr Asp Tyr Tyr Gly Ser Ser Leu Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 28
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 28
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Asp
20 25 30
Tyr Ala Trp Ser Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Ala Phe Ile Ser Tyr Ser Gly Pro Thr Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Lys Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Asp Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 29
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 29
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Ser Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 30
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 30
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Gly Gln Ser Leu Val His Ser
20 25 30
Asn Gly His Thr Tyr Phe His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 31
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 31
Asp Val Lys Met Thr Gln Ser Pro Ser Ser Met Tyr Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Ile Thr Cys Lys Ala Ser His Asp Ile Asn Ser Tyr
20 25 30
Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Thr Leu Ile
35 40 45
Tyr Arg Ala Asn Gly Leu Val Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Gln Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Tyr
65 70 75 80
Glu Asp Met Gly Val Tyr Tyr Cys Leu Gln Tyr Glu Met Leu Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 32
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 32
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 33
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 33
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Met Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Asp His Thr Gly Glu Pro Thr Tyr Ala Asp Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Leu Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Tyr Gly Phe Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 34
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 34
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Ser Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 35
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 35
Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Met Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Asp His Thr Gly Glu Pro Thr Tyr Ala Asp Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Leu Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Tyr Gly Phe Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 36
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 36
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Ser Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 37
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 37
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Ser Ser Asp
20 25 30
Tyr Ala Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Pro Thr Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Asp Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 38
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 38
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Lys Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 39
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 39
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 40
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 40
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
Tyr Ala Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Pro Thr Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Lys Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Asp Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 41
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 41
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 42
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> HC
<400> 42
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Met Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Asp His Thr Gly Glu Pro Thr Tyr Ala Asp Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Leu Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Tyr Gly Phe Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 43
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> LC
<400> 43
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Ser Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 44
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> HC
<400> 44
Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Met Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Asp His Thr Gly Glu Pro Thr Tyr Ala Asp Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Leu Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Tyr Gly Phe Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 45
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> LC
<400> 45
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Ser Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 46
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> HC
<400> 46
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Ser Ser Asp
20 25 30
Tyr Ala Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Pro Thr Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Asp Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 47
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> LC
<400> 47
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Lys Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 48
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> LC
<400> 48
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 49
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> HC
<400> 49
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
Tyr Ala Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Pro Thr Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Lys Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Asp Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 50
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> LC
<400> 50
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 51
<211> 1335
<212> DNA
<213> Artificial Sequence
<220>
<223> HC
<400> 51
caggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg 60
agctgcaagg ccagcggcta catcttcacc aactacggca tgaactggat gagacaggcc 120
cccggccagg gcctggagtg gatgggctgg atcaacgacc acaccggcga gcccacctac 180
gccgacaagt tccagggcag agtgaccttc accctggaca ccagcatcag caccgcctac 240
atggagctga gcagactgag aagcgacgac accgccgtgt actactgcgc caactacggc 300
ttcggctact tcgactactg gggccagggc accctggtga ccgtgagcag cgctagcacc 360
aagggcccca gcgtgttccc cctggcccct tgcagcagaa gcaccagcga gagcacagcc 420
gccctgggct gcctggtgaa ggactacttc cccgagcccg tgaccgtgtc ctggaacagc 480
ggcgctctga ccagcggcgt gcataccttc cccgccgtgc tccagagcag cggactgtac 540
tccctgagca gcgtggtgac cgtgccttcc agcagcctgg gcaccaagac ctacacctgc 600
aacgtggacc acaagcccag caacaccaag gtggacaaga gagtggagag caagtacggc 660
cctccctgcc ccccttgccc tgcccccgag ttcctgggcg gacctagcgt gttcctgttc 720
ccccccaagc ccaaggacac cctgatgatc agcagaaccc ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccaggagga ccccgaggtc cagtttaatt ggtacgtgga cggcgtggaa 840
gtgcataacg ccaagaccaa gcccagagag gagcagttca acagcaccta cagagtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aggaatacaa gtgcaaggtc 960
tccaacaagg gcctgcctag cagcatcgag aagaccatca gcaaggccaa gggccagcca 1020
cgggagcccc aggtctacac cctgccacct agccaagagg agatgaccaa gaaccaggtg 1080
tccctgacct gtctggtgaa aggcttctat cccagcgata tcgccgtgga gtgggagagc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggcagc 1200
ttcttcctgt actccagact gaccgtggac aagtccagat ggcaggaggg caacgtcttc 1260
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgagcctg 1320
agcctgggca agtga 1335
<210> 52
<211> 660
<212> DNA
<213> Artificial Sequence
<220>
<223> LC
<400> 52
gacgtggtga tgacccagag ccccctgagc ctgcccgtga ccctgggcca gcccgccagc 60
atcagctgca gaagcagcca gagcctggtg cacagcaacg gcaacagcta cctgcactgg 120
tacctgcaga agcccggcca gagcccccag ctgctgatct acaaggtgag caacagattc 180
agcggcgtgc ccgacagatt cagcggcagc ggcagcggca ccgacttcac cctgaagatc 240
agcagagtgg aggccgagga cgtgggcgtg tactactgca gccagagcac ccacgtgccc 300
tacaccttcg gccagggcac caagctggag atcaagcgaa ctgtggctgc accatctgtc 360
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 420
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 480
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 540
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 600
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttag 660
<210> 53
<211> 1335
<212> DNA
<213> Artificial Sequence
<220>
<223> HC
<400> 53
cagatccagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaagatc 60
agctgcaagg ccagcggcta catcttcacc aactacggca tgaactggat gagacaggcc 120
cccggccagg gcctggagtg gatgggctgg atcaacgacc acaccggcga gcccacctac 180
gccgacaagt tccagggcag agtgaccttc accctggaca ccagcgccag caccgcctac 240
atggagctga gcagactgag aagcgacgac accgccgtgt actactgcgc caactacggc 300
ttcggctact tcgactactg gggccagggc accctggtga ccgtgagcag cgctagcacc 360
aagggcccca gcgtgttccc cctggcccct tgcagcagaa gcaccagcga gagcacagcc 420
gccctgggct gcctggtgaa ggactacttc cccgagcccg tgaccgtgtc ctggaacagc 480
ggcgctctga ccagcggcgt gcataccttc cccgccgtgc tccagagcag cggactgtac 540
tccctgagca gcgtggtgac cgtgccttcc agcagcctgg gcaccaagac ctacacctgc 600
aacgtggacc acaagcccag caacaccaag gtggacaaga gagtggagag caagtacggc 660
cctccctgcc ccccttgccc tgcccccgag ttcctgggcg gacctagcgt gttcctgttc 720
ccccccaagc ccaaggacac cctgatgatc agcagaaccc ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccaggagga ccccgaggtc cagtttaatt ggtacgtgga cggcgtggaa 840
gtgcataacg ccaagaccaa gcccagagag gagcagttca acagcaccta cagagtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aggaatacaa gtgcaaggtc 960
tccaacaagg gcctgcctag cagcatcgag aagaccatca gcaaggccaa gggccagcca 1020
cgggagcccc aggtctacac cctgccacct agccaagagg agatgaccaa gaaccaggtg 1080
tccctgacct gtctggtgaa aggcttctat cccagcgata tcgccgtgga gtgggagagc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggcagc 1200
ttcttcctgt actccagact gaccgtggac aagtccagat ggcaggaggg caacgtcttc 1260
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgagcctg 1320
agcctgggca agtga 1335
<210> 54
<211> 660
<212> DNA
<213> Artificial Sequence
<220>
<223> LC
<400> 54
gacgtggtga tgacccagag ccccctgagc ctgcccgtga ccctgggcca gcccgccagc 60
atcagctgca gaagcagcca gagcctggtg cacagcaacg gcaacagcta cctgcactgg 120
tacctgcaga agcccggcca gagcccccag ctgctgatct acaaggtgag caacagattc 180
agcggcgtgc ccgacagatt cagcggcagc ggcagcggca ccgacttcac cctgaagatc 240
agcagagtgg aggccgagga cgtgggcgtg tacttctgca gccagagcac ccacgtgccc 300
tacaccttcg gccagggcac caagctggag atcaagcgaa ctgtggctgc accatctgtc 360
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 420
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 480
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 540
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 600
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttag 660
<210> 55
<211> 1338
<212> DNA
<213> Artificial Sequence
<220>
<223> HC
<400> 55
caggtgcagc tgcaggagag cggccccggc ctggtgaagc ccagcgagac cctgagcctg 60
acctgcaccg tgaccggcta cagcatcagc agcgactacg cctggagctg gatcagacag 120
ccccccggca agggcctgga gtggatcggc ttcatcagct acagcggccc caccagctac 180
aaccccagcc tgaagtccag agtgaccatc agcagagaca ccagcaagaa ccagttcagc 240
ctgaagctga gcagcgtgac cgccgccgac accgccgtgt actactgcgc cagaggcggc 300
gacggcgaca gcttcgacta ctggggccag ggcaccctgg tgaccgtgag cagcgctagc 360
accaagggcc ccagcgtgtt ccccctggcc ccttgcagca gaagcaccag cgagagcaca 420
gccgccctgg gctgcctggt gaaggactac ttccccgagc ccgtgaccgt gtcctggaac 480
agcggcgctc tgaccagcgg cgtgcatacc ttccccgccg tgctccagag cagcggactg 540
tactccctga gcagcgtggt gaccgtgcct tccagcagcc tgggcaccaa gacctacacc 600
tgcaacgtgg accacaagcc cagcaacacc aaggtggaca agagagtgga gagcaagtac 660
ggccctccct gccccccttg ccctgccccc gagttcctgg gcggacctag cgtgttcctg 720
ttccccccca agcccaagga caccctgatg atcagcagaa cccccgaggt gacctgcgtg 780
gtggtggacg tgtcccagga ggaccccgag gtccagttta attggtacgt ggacggcgtg 840
gaagtgcata acgccaagac caagcccaga gaggagcagt tcaacagcac ctacagagtg 900
gtgtccgtgc tgaccgtgct gcaccaggac tggctgaacg gcaaggaata caagtgcaag 960
gtctccaaca agggcctgcc tagcagcatc gagaagacca tcagcaaggc caagggccag 1020
ccacgggagc cccaggtcta caccctgcca cctagccaag aggagatgac caagaaccag 1080
gtgtccctga cctgtctggt gaaaggcttc tatcccagcg atatcgccgt ggagtgggag 1140
agcaacggcc agcccgagaa caactacaag accacccccc ctgtgctgga cagcgacggc 1200
agcttcttcc tgtactccag actgaccgtg gacaagtcca gatggcagga gggcaacgtc 1260
ttcagctgct ccgtgatgca cgaggccctg cacaaccact acacccagaa gtccctgagc 1320
ctgagcctgg gcaagtga 1338
<210> 56
<211> 657
<212> DNA
<213> Artificial Sequence
<220>
<223> LC
<400> 56
gacgtggtga tgacccagag ccccctgagc ctgcccgtga ccctgggcca gcccgccagc 60
atcagctgca gaagcagcca gagcatcgtg cacagcaacg gcaacaccta cctggactgg 120
tacctgcaga agcccggcaa gagccccaag ctgctgatct acaaggtgag caacagattc 180
agcggcgtgc ccgacagatt cagcggcagc ggcagcggca ccgacttcac cctgaagatc 240
agcagagtgg aggccgagga cctgggcgtg tactactgct tccagggcag ccacttcccc 300
accttcggcc agggcaccaa gctggagatc aagcgaactg tggctgcacc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttag 657
<210> 57
<211> 657
<212> DNA
<213> Artificial Sequence
<220>
<223> LC
<400> 57
gacgtggtga tgacccagag ccccctgagc ctgcccgtga ccctgggcca gcccgccagc 60
atcagctgca gaagcagcca gagcatcgtg cacagcaacg gcaacaccta cctggactgg 120
tacctgcaga agcccggcca gagccccaag ctgctgatct acaaggtgag caacagattc 180
agcggcgtgc ccgacagatt cagcggcagc ggcagcggca ccagcttcac cctgaagatc 240
agcagagtgg aggccgagga cctgggcgtg tactactgct tccagggcag ccacttcccc 300
accttcggcc agggcaccaa gctggagatc aagcgaactg tggctgcacc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttag 657
<210> 58
<211> 1338
<212> DNA
<213> Artificial Sequence
<220>
<223> HC
<400> 58
caggtgcagc tgcaggagag cggccccggc ctggtgaagc ccagcgagac cctgagcctg 60
acctgcaccg tgagcggcta cagcatcacc agcgactacg cctggagctg gatcagacag 120
ccccccggca agggcctgga gtggatcggc ttcatcagct acagcggccc caccagctac 180
aaccccagcc tgaagtccag agtgaccatc agcagagaca ccagcaagaa ccagttcttc 240
ctgcagctga agtccgtgac cgccgccgac accgccgtgt actactgcgc cagaggcggc 300
gacggcgaca gcttcgacta ctggggccag ggcaccaccg tgaccgtgag cagcgctagc 360
accaagggcc ccagcgtgtt ccccctggcc ccttgcagca gaagcaccag cgagagcaca 420
gccgccctgg gctgcctggt gaaggactac ttccccgagc ccgtgaccgt gtcctggaac 480
agcggcgctc tgaccagcgg cgtgcatacc ttccccgccg tgctccagag cagcggactg 540
tactccctga gcagcgtggt gaccgtgcct tccagcagcc tgggcaccaa gacctacacc 600
tgcaacgtgg accacaagcc cagcaacacc aaggtggaca agagagtgga gagcaagtac 660
ggccctccct gccccccttg ccctgccccc gagttcctgg gcggacctag cgtgttcctg 720
ttccccccca agcccaagga caccctgatg atcagcagaa cccccgaggt gacctgcgtg 780
gtggtggacg tgtcccagga ggaccccgag gtccagttta attggtacgt ggacggcgtg 840
gaagtgcata acgccaagac caagcccaga gaggagcagt tcaacagcac ctacagagtg 900
gtgtccgtgc tgaccgtgct gcaccaggac tggctgaacg gcaaggaata caagtgcaag 960
gtctccaaca agggcctgcc tagcagcatc gagaagacca tcagcaaggc caagggccag 1020
ccacgggagc cccaggtcta caccctgcca cctagccaag aggagatgac caagaaccag 1080
gtgtccctga cctgtctggt gaaaggcttc tatcccagcg atatcgccgt ggagtgggag 1140
agcaacggcc agcccgagaa caactacaag accacccccc ctgtgctgga cagcgacggc 1200
agcttcttcc tgtactccag actgaccgtg gacaagtcca gatggcagga gggcaacgtc 1260
ttcagctgct ccgtgatgca cgaggccctg cacaaccact acacccagaa gtccctgagc 1320
ctgagcctgg gcaagtga 1338
<210> 59
<211> 657
<212> DNA
<213> Artificial Sequence
<220>
<223> LC
<400> 59
gacgtggtga tgacccagac ccccctgagc ctgagcgtga cccccggcca gcccgccagc 60
atcagctgca agagcagcca gagcatcgtg cacagcaacg gcaacaccta cctggactgg 120
tacctgcaga agcccggcca gagccccaag ctgctgatct acaaggtgag caacagattc 180
agcggcgtgc ccgacagatt cagcggcagc ggcagcggca ccgacttcac cctgaagatc 240
agcagagtgg aggccgagga cgtgggcgtg tactactgct tccagggcag ccacttcccc 300
accttcggcg gcggcaccaa ggtggagatc aagcgaactg tggctgcacc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttag 657
<210> 60
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR1
<400> 60
Gly Tyr Ser Ile Thr Ser Asp Tyr Ala
1 5
<210> 61
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR2
<400> 61
Ile Ser Tyr Ser Gly Pro Thr
1 5
<210> 62
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> HCDR3
<400> 62
Ala Arg Gly Gly Asp Gly Asp Ser Phe Asp Tyr
1 5 10
<210> 63
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR1
<400> 63
Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
1 5 10
<210> 64
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR2
<400> 64
Lys Val Ser
1
<210> 65
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> LCDR3
<400> 65
Phe Gln Gly Ser His Phe Pro Thr
1 5
<210> 66
<211> 132
<212> PRT
<213> Homo sapiens
<400> 66
Gly Ile Thr Ile Pro Arg Asn Pro Gly Cys Pro Asn Ser Glu Asp Lys
1 5 10 15
Asn Phe Pro Arg Thr Val Met Val Asn Leu Asn Ile His Asn Arg Asn
20 25 30
Thr Asn Thr Asn Pro Lys Arg Ser Ser Asp Tyr Tyr Asn Arg Ser Thr
35 40 45
Ser Pro Trp Asn Leu His Arg Asn Glu Asp Pro Glu Arg Tyr Pro Ser
50 55 60
Val Ile Trp Glu Ala Lys Cys Arg His Leu Gly Cys Ile Asn Ala Asp
65 70 75 80
Gly Asn Val Asp Tyr His Met Asn Ser Val Pro Ile Gln Gln Glu Ile
85 90 95
Leu Val Leu Arg Arg Glu Pro Pro His Cys Pro Asn Ser Phe Arg Leu
100 105 110
Glu Lys Ile Leu Val Ser Val Gly Cys Thr Cys Val Thr Pro Ile Val
115 120 125
His His Val Ala
130
<210> 67
<211> 396
<212> DNA
<213> Homo sapiens
<400> 67
ggaatcacaa tcccacgaaa tccaggatgc ccaaattctg aggacaagaa cttcccccgg 60
actgtgatgg tcaacctgaa catccataac cggaatacca ataccaatcc caaaaggtcc 120
tcagattact acaaccgatc cacctcacct tggaatctcc accgcaatga ggaccctgag 180
agatatccct ctgtgatctg ggaggcaaag tgccgccact tgggctgcat caacgctgat 240
gggaacgtgg actaccacat gaactctgtc cccatccagc aagagatcct ggtcctgcgc 300
agggagcctc cacactgccc caactccttc cggctggaga agatactggt gtccgtgggc 360
tgcacctgtg tcaccccgat tgtccaccat gtggcc 396
<210> 68
<211> 133
<212> PRT
<213> Homo sapiens
<400> 68
Arg Lys Ile Pro Lys Val Gly His Thr Phe Phe Gln Lys Pro Glu Ser
1 5 10 15
Cys Pro Pro Val Pro Gly Gly Ser Met Lys Leu Asp Ile Gly Ile Ile
20 25 30
Asn Glu Asn Gln Arg Val Ser Met Ser Arg Asn Ile Glu Ser Arg Ser
35 40 45
Thr Ser Pro Trp Asn Tyr Thr Val Thr Trp Asp Pro Asn Arg Tyr Pro
50 55 60
Ser Glu Val Val Gln Ala Gln Cys Arg Asn Leu Gly Cys Ile Asn Ala
65 70 75 80
Gln Gly Lys Glu Asp Ile Ser Met Asn Ser Val Pro Ile Gln Gln Glu
85 90 95
Thr Leu Val Val Arg Arg Lys His Gln Gly Cys Ser Val Ser Phe Gln
100 105 110
Leu Glu Lys Val Leu Val Thr Val Gly Cys Thr Cys Val Thr Pro Val
115 120 125
Ile His His Val Gln
130
<210> 69
<211> 2601
<212> DNA
<213> Homo sapiens
<400> 69
atgggggccg cacgcagccc gccgtccgct gtcccggggc ccctgctggg gctgctcctg 60
ctgctcctgg gcgtgctggc cccgggtggc gcctccctgc gactcctgga ccaccgggcg 120
ctggtctgct cccagccggg gctaaactgc acggtcaaga atagtacctg cctggatgac 180
agctggattc accctcgaaa cctgaccccc tcctccccaa aggacctgca gatccagctg 240
cactttgccc acacccaaca aggagacctg ttccccgtgg ctcacatcga atggacactg 300
cagacagacg ccagcatcct gtacctcgag ggtgcagagt tatctgtcct gcagctgaac 360
accaatgaac gtttgtgcgt caggtttgag tttctgtcca aactgaggca tcaccacagg 420
cggtggcgtt ttaccttcag ccactttgtg gttgaccctg accaggaata tgaggtgacc 480
gttcaccacc tgcccaagcc catccctgat ggggacccaa accaccagtc caagaatttc 540
cttgtgcctg actgtgagca cgccaggatg aaggtaacca cgccatgcat gagctcaggc 600
agcctgtggg accccaacat caccgtggag accctggagg cccaccagct gcgtgtgagc 660
ttcaccctgt ggaacgaatc tacccattac cagatcctgc tgaccagttt tccgcacatg 720
gagaaccaca gttgctttga gcacatgcac cacatacctg cgcccagacc agaagagttc 780
caccagcgat ccaacgtcac actcactcta cgcaacctta aagggtgctg tcgccaccaa 840
gtgcagatcc agcccttctt cagcagctgc ctcaatgact gcctcagaca ctccgcgact 900
gtttcctgcc cagaaatgcc agacactcca gaaccaattc cggactacat gcccctgtgg 960
gtgtactggt tcatcacggg catctccatc ctgctggtgg gctccgtcat cctgctcatc 1020
gtctgcatga cctggaggct agctgggcct ggaagtgaaa aatacagtga tgacaccaaa 1080
tacaccgatg gcctgcctgc ggctgacctg atccccccac cgctgaagcc caggaaggtc 1140
tggatcatct actcagccga ccaccccctc tacgtggacg tggtcctgaa attcgcccag 1200
ttcctgctca ccgcctgcgg cacggaagtg gccctggacc tgctggaaga gcaggccatc 1260
tcggaggcag gagtcatgac ctgggtgggc cgtcagaagc aggagatggt ggagagcaac 1320
tctaagatca tcgtcctgtg ctcccgcggc acgcgcgcca agtggcaggc gctcctgggc 1380
cggggggcgc ctgtgcggct gcgctgcgac cacggaaagc ccgtggggga cctgttcact 1440
gcagccatga acatgatcct cccggacttc aagaggccag cctgcttcgg cacctacgta 1500
gtctgctact tcagcgaggt cagctgtgac ggcgacgtcc ccgacctgtt cggcgcggcg 1560
ccgcggtacc cgctcatgga caggttcgag gaggtgtact tccgcatcca ggacctggag 1620
atgttccagc cgggccgcat gcaccgcgta ggggagctgt cgggggacaa ctacctgcgg 1680
agcccgggcg gcaggcagct ccgcgccgcc ctggacaggt tccgggactg gcaggtccgc 1740
tgtcccgact ggttcgaatg tgagaacctc tactcagcag atgaccagga tgccccgtcc 1800
ctggacgaag aggtgtttga ggagccactg ctgcctccgg gaaccggcat cgtgaagcgg 1860
gcgcccctgg tgcgcgagcc tggctcccag gcctgcctgg ccatagaccc gctggtcggg 1920
gaggaaggag gagcagcagt ggcaaagctg gaacctcacc tgcagccccg gggtcagcca 1980
gcgccgcagc ccctccacac cctggtgctc gccgcagagg agggggccct ggtggccgcg 2040
gtggagcctg ggcccctggc tgacggtgcc gcagtccggc tggcactggc gggggagggc 2100
gaggcctgcc cgctgctggg cagcccgggc gctgggcgaa atagcgtcct cttcctcccc 2160
gtggaccccg aggactcgcc ccttggcagc agcaccccca tggcgtctcc tgacctcctt 2220
ccagaggacg tgagggagca cctcgaaggc ttgatgctct cgctcttcga gcagagtctg 2280
agctgccagg cccagggggg ctgcagtaga cccgccatgg tcctcacaga cccacacacg 2340
ccctacgagg aggagcagcg gcagtcagtg cagtctgacc agggctacat ctccaggagc 2400
tccccgcagc cccccgaggg actcacggaa atggaggaag aggaggaaga ggagcaggac 2460
ccagggaagc cggccctgcc actctctccc gaggacctgg agagcctgag gagcctccag 2520
cggcagctgc ttttccgcca gctgcagaag aactcgggct gggacacgat ggggtcagag 2580
tcagaggggc ccagtgcatg a 2601
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К TIGIT И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2827445C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ PD-1 В ИЗГОТОВЛЕНИИ ЛЕКАРСТВЕННОГО СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ СОЛИДНЫХ ОПУХОЛЕЙ | 2020 |
|
RU2825845C2 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| ГЕТЕРОДИМЕРНОЕ БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ TNFα/ПРОТИВ IL-17A, ПО СТРУКТУРЕ НАПОМИНАЮЩЕЕ ПРИРОДНОЕ АНТИТЕЛО, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2019 |
|
RU2781816C1 |
| IL-5-СВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ | 2021 |
|
RU2811518C1 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| АНТИТЕЛО, СВЯЗЫВАЮЩЕЕ ЧЕЛОВЕЧЕСКИЙ IL-4R, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2779649C1 |






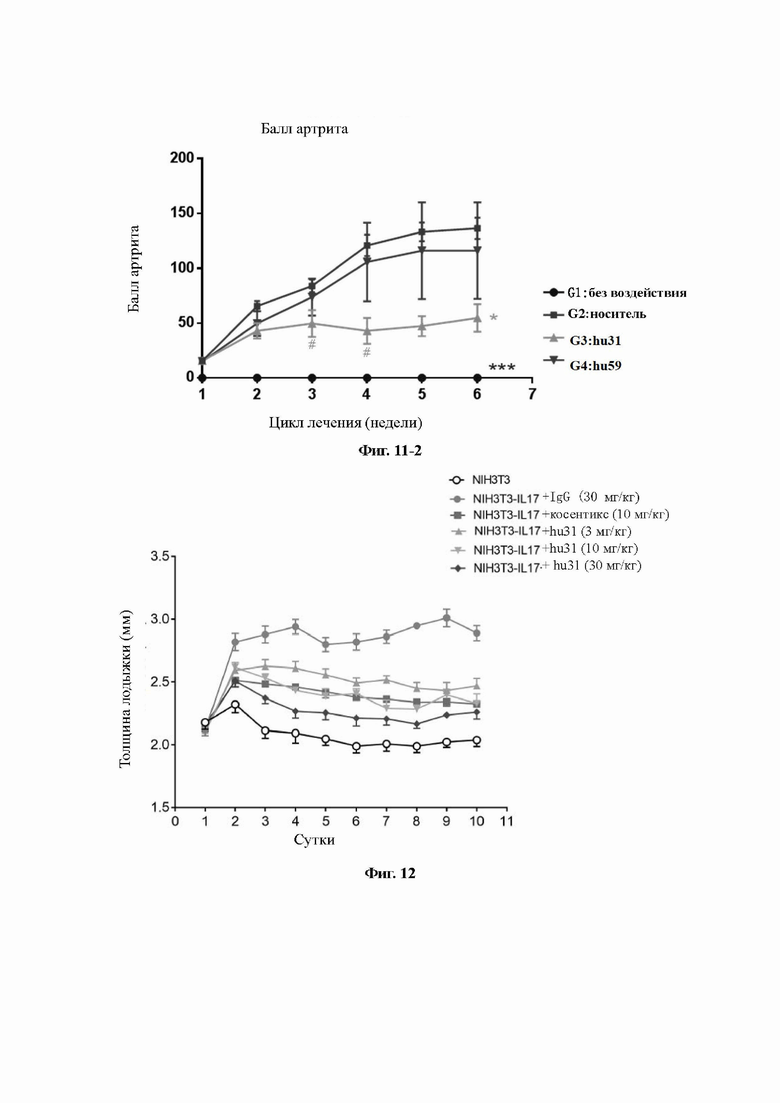

Изобретение относится к области биотехнологии, в частности к антителу или его антигенсвязывающему фрагменту, которые специфически связываются с интерлейкином 17А (IL-17A), а также к композиции, его содержащей. Также раскрыты молекула нуклеиновой кислоты, кодирующая вышеуказанное антитело или его фрагмент, а также клетка и вектор, ее содержащие. Изобретение эффективно для лечения или предупреждения заболеваний или расстройств, опосредованных IL-17A. 6 н. и 9 з.п. ф-лы, 13 ил., 6 табл., 18 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые специфически связываются с интерлейкином 17А (IL-17A), содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где вариабельные области содержат совокупность из 6 аминокислотных последовательностей CDR, выбранных из любой совокупности I-VI:

2. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащие вариабельную область тяжелой цепи (VH), имеющую аминокислотную последовательность, выбранную из SEQ ID NO: 25, 26, 27, 28, 33, 35, 37 и 40, и вариабельную область легкой цепи (VL), имеющую аминокислотную последовательность, выбранную из SEQ ID NO: 29, 30, 31, 32, 34, 36, 38, 39 и 41.
3. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где вариабельные области содержат аминокислотные последовательности, выбранные из любой совокупности 1-7:

4. Антитело или его антигенсвязывающий фрагмент по п. 1, где антитело или его антигенсвязывающий фрагмент представляет собой мышиное антитело, химерное антитело, гуманизированное антитело или полностью человеческое антитело.
5. Антитело или его антигенсвязывающий фрагмент по п. 1, где антитело представляет собой интактное антитело, и антигенсвязывающий фрагмент выбран из одноцепочечного антитела, Fab-антитела, Fab'-антитела и (Fab')2-антитела.
6. Антитело или его антигенсвязывающий фрагмент по п. 1, где антитело или его антигенсвязывающий фрагмент имеет любой из подтипов IgG.
7. Антитело или его антигенсвязывающий фрагмент по п. 6, где антитело или его антигенсвязывающий фрагмент представляет собой IgG1, IgG2, IgG3 или IgG4.
8. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащие тяжелую цепь (НС), имеющую аминокислотную последовательность, выбранную из SEQ ID NO: 42, 44, 46 и 49, и легкую цепь (LC), имеющую аминокислотную последовательность, выбранную из SEQ ID NO: 43, 45, 47, 48 и 50.
9. Антитело или его антигенсвязывающий фрагмент по п. 8, где легкая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 43, и тяжелая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 42 или 44; легкая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 45, и тяжелая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 44; легкая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 47 или 48, и тяжелая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 46; или легкая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 50, и тяжелая цепь имеет аминокислотную последовательность, представленную в SEQ ID NO: 49.
10. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-9.
11. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п. 10.
12. Клетка-хозяин для продуцирования антитела по п. 1, трансформированная экспрессионным вектором по п. 11.
13. Фармацевтическая композиция для лечения или предупреждения заболеваний или расстройств, опосредованных IL-17A, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-9, молекулу нуклеиновой кислоты по п. 10, экспрессионный вектор по п. 11 или клетку-хозяина по п. 12 и фармацевтически приемлемый носитель или эксципиент.
14. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-9, молекулы нуклеиновой кислоты по п. 10, экспрессионного вектора по п. 11 или клетки-хозяина по п. 12 или фармацевтической композиции по п. 13 для изготовления лекарственного средства для лечения или предупреждения заболеваний или расстройств, опосредованных IL-17A.
15. Применение по п. 14, где лекарственное средство представляет собой лекарственное средство для лечения артрита, ревматоидного артрита, псориаза, анкилозирующего спондилита, хронической обструктивной болезни легких, системной красной волчанки (SLE), волчаночного нефрита, астмы, рассеянного склероза или муковисцидоза.
| WO2018050028 A1, 22.03.2018 | |||
| ERIC TOUSSIROT, Ixekizumab: an anti- IL-17A monoclonal antibody for the treatment of psoriatic arthritis, Expert Opinion on Biological Therapy, 2017, 18(1), 101-107 | |||
| SARAH L | |||
| GAFFEN et al | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| АНТИ-IL-17-АНТИТЕЛА, СПОСОБ ИХ ПОЛУЧЕНИЯ И СПОСОБ ПРИМЕНЕНИЯ | 2014 |
|
RU2577228C2 |

