Изобретение относится к области медицины, и может быть использовано создания средства, обладающего противовирусными свойствами в отношении респираторно-синцитиального вируса (РСВ) и вируса гриппа.
Эпидемиологические исследования позволяют утверждать, что среди всех вирусных патогенов именно респираторные вирусы (грипп, респираторно-синцитиальный вирус, риновирусы и т.д.) являются самыми распространенными (более 90% от всех случаев инфекционных заболеваний). Один из самых распространенных вирусных патогенов -респираторно-синцитиальный вирус (РСВ). Он поражает нижние дыхательные пути, что примерно в 40% случаев, сопровождается тяжелым бронхиолитом и пневмонией. Особенно РСВ инфекция опасна для новорожденных, у которых часто приводит к летальному исходу, для лиц с иммунодефицитами и для астматиков [Pretorius и др., 2016]. Несмотря на десятилетия исследований респираторных вирусных инфекций к настоящему моменту не существует эффективных, безопасных и при этом доступных средств лечения и профилактики. Попытки создать профилактическую вакцину оказались неудачными, поэтому в настоящее время для лечения РСВ-инфекции применяется только виразол (рибавирин), однако он вызывает многочисленные побочные эффекты [Janai НK, 1990]. Многие анти-РСВ терапевтические средства еще находятся в разработке [Osminkina и др., 2014; Хаитов и др., 2010]. Препараты на основе моноклональных антител малодоступны ввиду их высокой стоимости.
Один из перспективных подходов к созданию антивирусных средств - это использование природных пептидов. Природные пептиды являются неотъемлемой частью врожденного иммунного ответа с бактерицидными, иммуномодулирующими и противовирусными свойствами. У млекопитающих два основных семейства таких пептидов: дефензины и кателицидины [Gwyer Findlay, Currie, Davidson, 2013].
Основные механизмы противовирусного действия природных пептидов это:
1. блокирование входа вируса в клетку (путем конкурентного ингибирования либо клеточного рецептора, либо лиганда на поверхности вируса),
2. дестабилизация вириона,
3. блокирование репликации вируса,
4. активация противовирусных интерферонов.
О перспективности данного подхода свидетельствует тот факт, что около 15-ти пептид-содержащих антимикробных препаратов проходят клинические исследования [Fjell и др., 2012; Greber, Dawgul, 2017].
Применение природных пептидов в медицинской практике сопряжено с рядом проблем; а именно: процесс выделения пептидов из природных источников затратен, химический синтез крупных (40-50 а.о.) природных пептидов обходится ненамного дешевле, не высокая стабильность - линейные пептиды легко разрушаются протеазами.
Несмотря на вышеуказанные сложности, существуют способы их преодоления. В первую очередь это картирование крупных пептидов и выявление в их структуре биологически-значимой части. Благодаря такому подходу можно добиться снижения размера целевого пептида (с 40-50 до 22-35 а.о.). Во-вторых, это стабилизация структуры пептидов за счет введения точечных аминокислотных замен или дополнительных химических связей. В-третьих, путем дендримеризации пептидов (синтез разветвленных пептидов) можно добиться повышения их стабильности за счет наличия неприродных ε-связей и биологической активности за счет повышения валентности молекулы пептида по отношению к биологическому рецептору, при сокращенном количестве циклов присоединения аминокислот к наращиваемой пептидной цепи.
В данном изобретении мы описываем катионный разветвленный пептид КК-45, который проявляет выраженные антивирусные свойства по отношению к респираторным вирусам (в частности, к РСВ и вирусу гриппа).
Применение пептида в качестве антивирусных средств является перспективным направлением. В международной базе данных представлено множество изобретений в данной области.
В изобретении US9102712 B2 описаны фармацевтические композиции, содержащие активные дендримерные пептиды, которые демонстрируют противомикробные, противогрибковые или противовирусное свойства. Отличие, представленного нами изобретения по сравнению с изобретением, описанным в US9102712 B2, состоит в том, что использована другая химическая формула дендримерного пептида, а также пептиды, описанные в US9102712 B2, предлагается использовать совместно с антибиотиками для синергетического эффекта, а не в качестве индивидуальных соединений.
Изобретение, описанное в ES2365685 A1, представляет собой разветвленные макромолекулы, используемые в качестве противовирусного, антибактериального и противогрибкового средства. Основное отличие от нашего изобретения заключается в том, что данные пептиды, синтезированны из полифенольного ядра, карбосилановой структуры и функционализированны на периферии анионными группами, которые придают макромолекуле отрицательный суммарный заряд и способны связываться с ВИЧ и ВПГ. То же время пептид КК-45 имеет иную химическую структуру, обеспечивающую положительный, а не отрицательный заряд, в водных растворах.
Дендримерные пептиды, описанные в изобретении ES2444490 A1 близки с описанными в патенте ES2365685 A1. Указанные в данном патенте ES2444490 A1 молекулы были синтезированы из поливалентного ядра, в частности полифенольного ядра или ядра атома кремния, были получены в виде сферических дендримеров, так же, тест вирусного ингибирования проводился с анион-связывающим вирусом - ВИЧ-1, что и является главным отличием от нашего изобретения. Еще одним важным отличием пептида КК-45 вот соединение описанных в ES2444490 A1 является отсутствие элемента кремния в структуре.
Новые дендримерные пептидные соединения в патенте US9079937B2, которые проявляют антимикробную активность, могут быть приготовлены в виде фармацевтических соединений и использованы для профилактики и лечения различных заболеваний у млекопитающих, включая человека. Настоящие пептиды особенно ценны, так как их действие быстрое, широкое по спектру и практически не влияет на резистентность, вызываемую стандартными антибиотиками. Основным отличием является спектр действия. В патенте US9079937B2 был описан антибактериальный спектр действия днендримерных соединений, в то время как пептид КК-45 проявляет антивирусный эффект в отношении респираторных вирусов.
Настоящее изобретение ES2255060 T3 относится к противовирусным пептидным агентам и, в частности, оно относится к дендримерам, которые, как было обнаружено, обладают значительной противовирусной активностью против вируса иммунодефицита человека (ВИЧ) и других оболочечных вирусов. Отличие от нашего изобретения заключается в том, что дендримерный пептид КК-45 направлен на респираторные вирусы.
Наиболее близким к нашей разработке является изобретение RU2451026C2, где описаны химерные пептиды, способные ингибировать или аттенуировать вирусную инфекцию. Пептиды содержат первичную структуру, которая содержит, по меньшей мере, один сегмент, который замедляет активацию протеазы NS3 вируса семейства Flaviviridae, и сегмент, проникающий в клетку. В заявленном изобретении мишенью является респираторно - синцитиальный вирус, вирус гриппа, которые относятся к роду вирусов из семейства ортомиксовирусов (Orthopneumovirus).
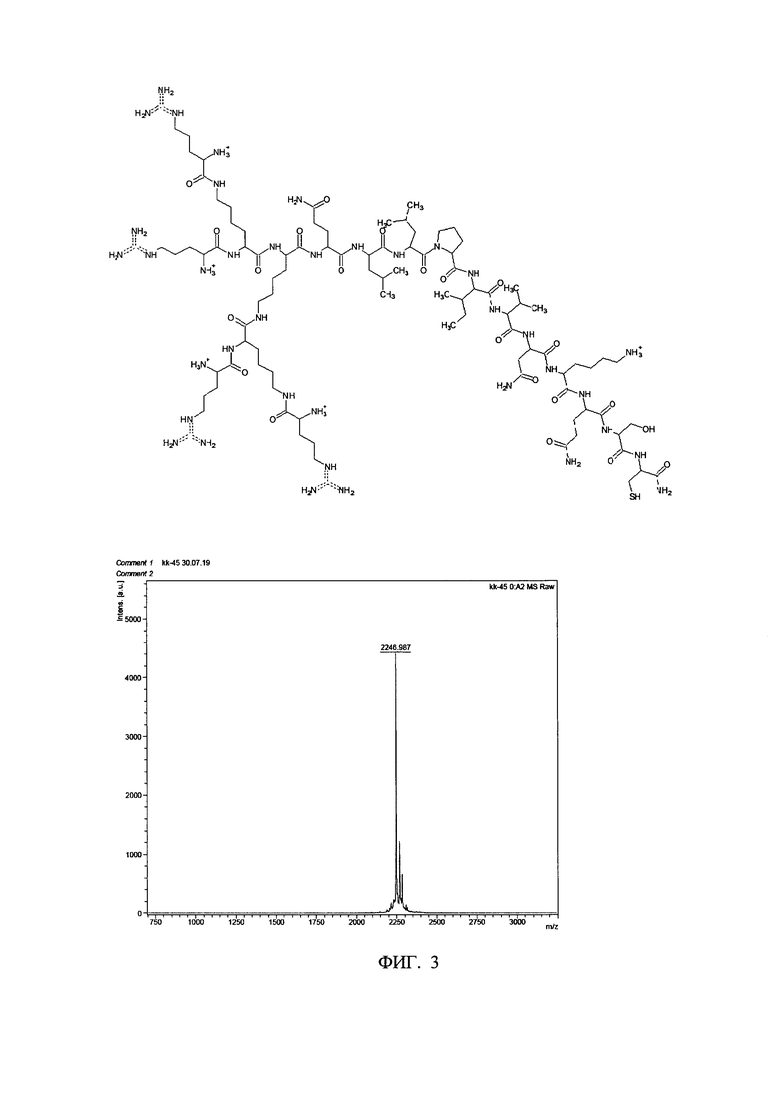
В настоящем изобретении описана разработка катионного разветвленного пептида, который способны ингибировать вирусную инфекцию, вызванную вирусами семейства Orthopneumovirus. Разработанные пептиды характеризуются структурной формулой R4K2KQLLPIVNKQSC, где R - аргинин; K - лизин; Q - глутамин, L - лейцин, Р - пролин, I - изолейцин, V - валин, N - аспарагин, S - серии, С - цистеин.
Средство для профилактики или лечения респираторной вирусной инфекции, вызванной респираторно-синцитиальным вирусом или вирусом гриппа содержит эффективное количество указанного катионного разветвленного пептида.
Средство может быть произведено в лекарственной форме, предназначенной для ингаляционного или интраназального введения введения. В одном из вариантов изобретения твердая лекарственная форма (лиофилизат) растворяется или диспергируется в жидкой среде перед применением и полученная суспензия, эмульсия, или раствор вводится ингаляционно или интраназально. Средство может быть введено на ранней стадии инфекции во время инкубационной фазы, или во время активной инфекции после возникновения симптомов.
Эффективное количество означает количество агента (пептида), которое является достаточным при введении субъекту, страдающему от инфекции с целью лечения, улучшения симптомов, предупреждения и/или отсрочки возникновения инфекции, заболевания. Растворители включают, но не ограничиваются ими, фосфатные буферные растворы, не содержащую пирогенов воду, изотонический солевой раствор.
Технический результат настоящего изобретения состоит в том, что результирующая структура катионного разветвленного пептида специфически связывается с вирионами посредством линейной С-концевой последовательности, а за счет наличия разветвленной структуры на N-конце, стерически препятствует нормальной функции белка F вируса, а именно, нарушает его структурную перестройку из префузионной в постфузионную форму. Полученный синтетический пептид с дендримерной (разветвленной) структурой ингибирует прикрепление вируса семейства Orthopneumovirus к клеткам-мишеням, при этом высокая антивирусная активность пептида, объясняется его способностью конкурентно связываться с рецептором -нуклеолином. Таким образом, полученное средство способно ингибировать вирусную инфекцию, вызванную вирусами семейства Orthopneumovirus, путем комбинированного действия полученного синтетического дендримерного пептида -ингибирования проникновения вируса в клетку и прямого вирулицидного действия -повреждающего действия пептида на оболочку вируса за счет его высокой плотности положительного заряда.
Сущность заявленного изобретения поясняется чертежами.
Краткое описание графических материалов.
Фигура 1. Структура белка F вируса РСВ и структуры изучаемых пептидов. А. Структура белка F вируса РСВ. Белок F разделен на две субъединицы, F1 и F2. В его структуре выделяют следующие участки: FP - пептид слияния для взаимодействия с клеткой; ТМ - трансмембранный домен для закрепления белка F в оболочке РСВ; HR-1 и HR-2 - области, образующие α-спиральную структуру. Домены D1, D2 и D3 образуют основной глобулярный домен. Антигенный ∅-сайт имеет важное значение для процесса слияния вируса с клеткой, его локализация отмечено на схеме.
Б. Структуры изучаемых пептидов. Синим цветом выделена разветвленная (дендримерная) часть пептида, красным цветом выделена линейная часть пептида, имитирующая антигенной ∅-сайт.
Фигура 2. Схема получение дендримерного пептида КК-45.
Фигура 3. Строение и масс-спектр синтезированного пептида КК-45 (+ позитивные заряды на остатках аргинина и лизина, всего 9).
Фигура 4. Цитотоксичность пептидов in vitro в культуре клеток МА-104.
× - статистически значимо отличается от «КК-44» по U-критерию Манна-Уитни. N=10.
* - статистически значимо отличается от «D-12» по U-критерию Манна-Уитни. N=10.
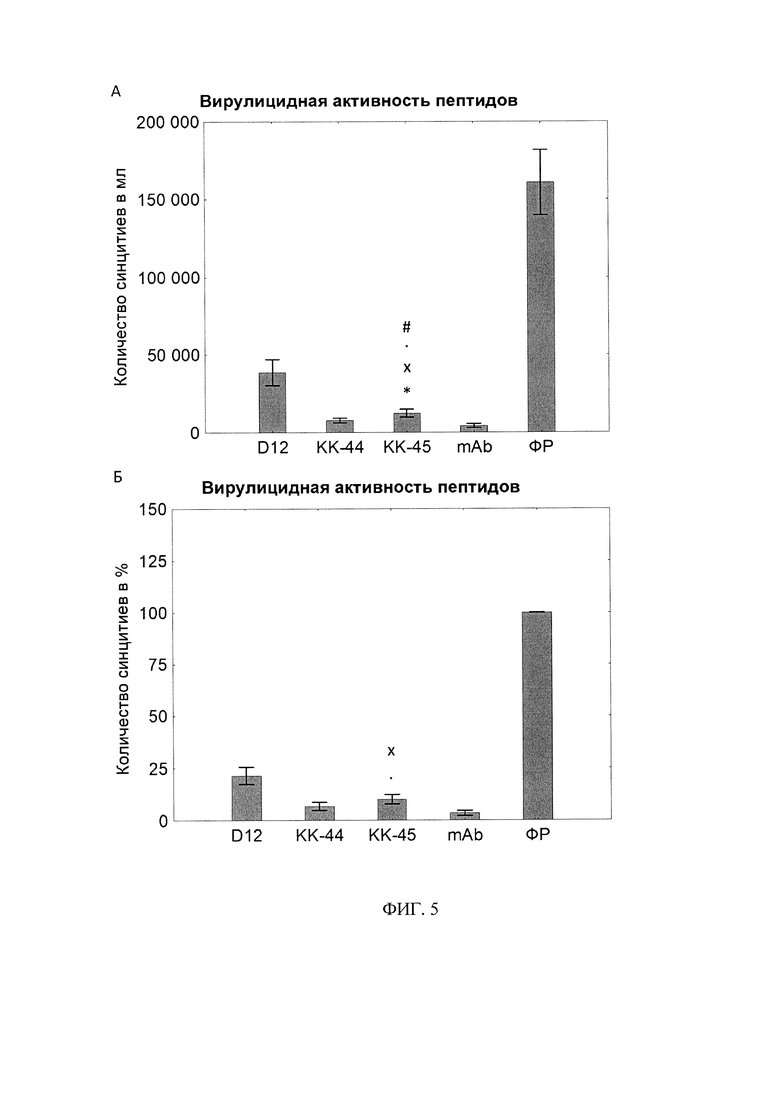
Фигура 5. Вирулицидная активность пептидов.
А - количество синцитиев в 1 мл.
Б - количество синцитиев в %.
* - статистически значимо отличается от «КК-44» по U-критерию Манна-Уитни. N=10.
# - статистически значимо отличается от «D-12» по U-критерию Манна-Уитни. N=10.
х - статистически значимо отличается от «mAb» по U-критерию Манна-Уитни. N=10.
. - статистически значимо отличается от «ФР» по U-критерию Манна-Уитни. N=10.
Фигура 6. Блокирование лигандов на поверхности РСВ.
А - Блокирование лигандов на поверхности РСВ в процентах относительно 0 мг/мл.
Б - Связывание пептидов с РСВ.
* - статистически значимо отличается от «КК-44» по U-критерию Манна-Уитни. N=10.
# - статистически значимо отличается от «D-12» по U-критерию Манна-Уитни. N=10.
х - статистически значимо отличается от «mAb» по U-критерию Манна-Уитни. N=10.
. - статистически значимо отличается от «Heparin» по U-критерию Манна-Уитни. N=10
∧ - статистически значимо отличается от «РСВ» по U-критерию Манна-Уитни. N=10
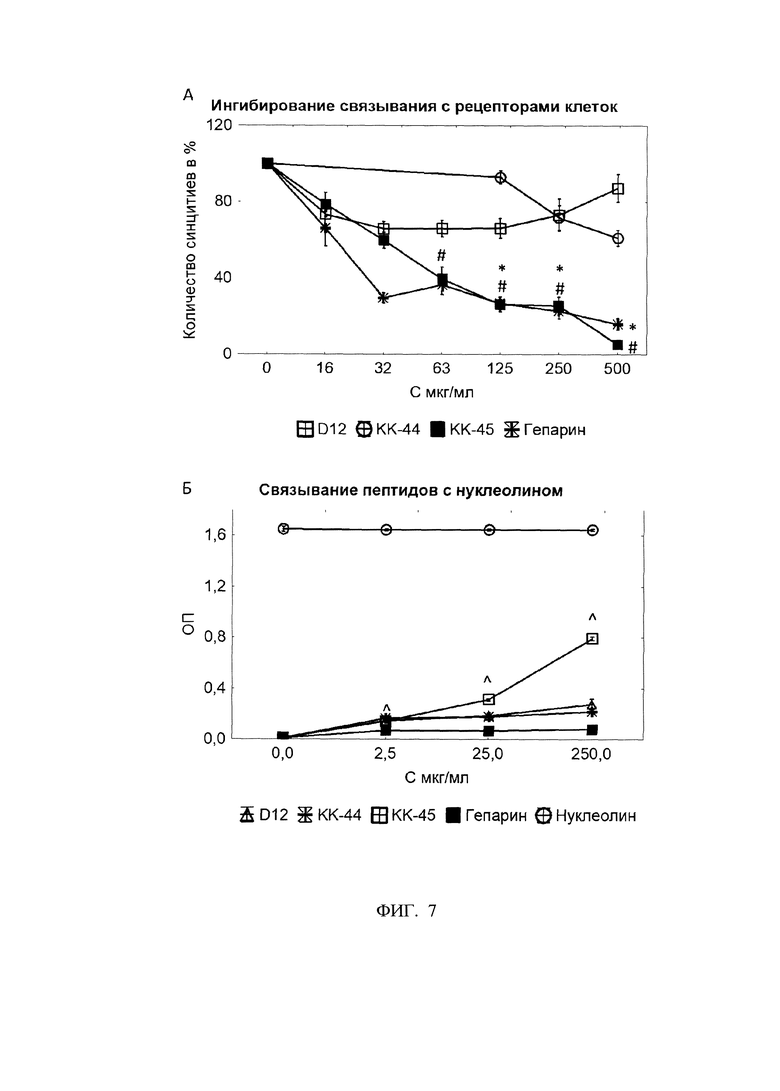
Фигура 7. Связывание пептидов с нуклеолином.
А - Ингибирование связывания с рецепторами клеток. Количество синцитиев в % относительно 0 мкг/мл.
Б - Связывание пептидов с нуклеолином.
* - статистически значимо отличается от «КК-44» по U-критерию Манна-Уитни. N=10.
# - статистически значимо отличается от «D-12» по U-критерию Манна-Уитни. N=10.
. - статистически значимо отличается от «Гепарин» по U-критерию Манна-Уитни. N=10
∧ - статистически значимо отличается от «Нуклеолин» по U-критерию Манна-Уитни. N=10
Фигура 8. Вирусная нагрузка в легких.
А - Вирусную нагрузку оценивали методом RT-PCR с использованием специфических праймеров и зонда. vRNA - РНК вируса, tRNA - общая РНК.
Б - Вирусная нагрузка в ткани легких методом титрования БАЛ на монослое чувствительных клеток с последующим подсчетом количество синцитиев.
х - статистически значимо отливается от «РСВ/PBS» по U-критерию Манна-Уитни. N=10.
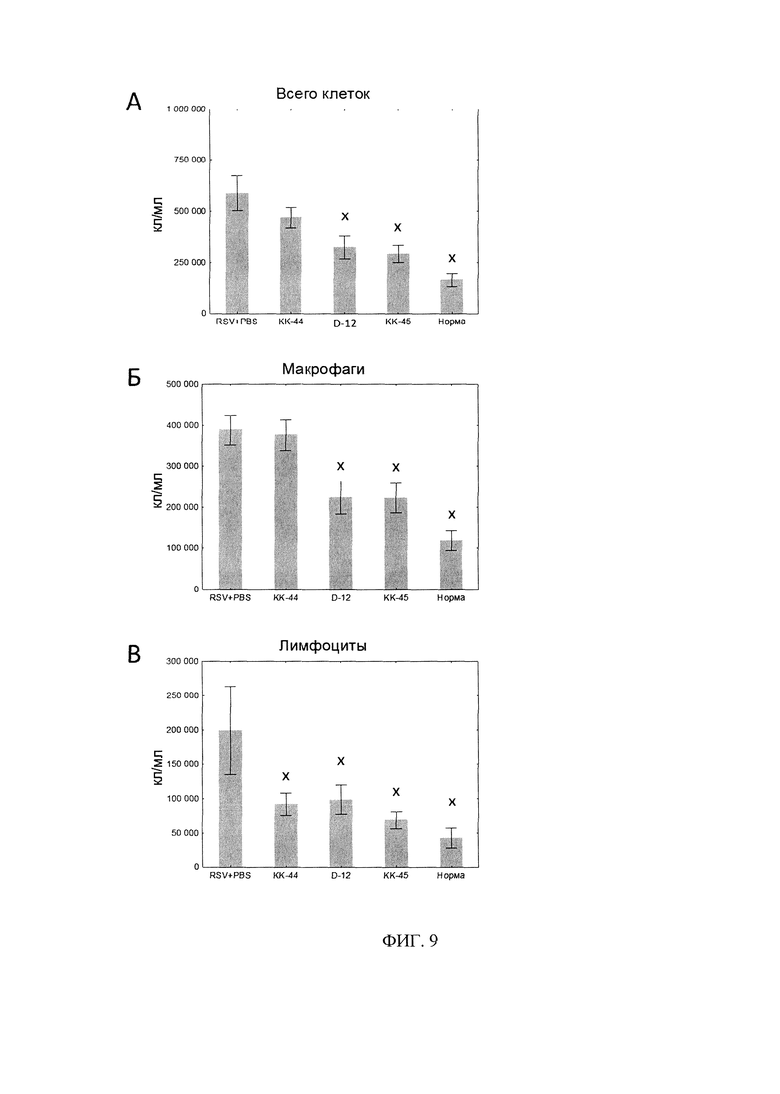
Фигура 9. Клеточный состава БАЛ.
А - Общее количество клеток в 1 мл БАЛ.
Б - Количество макрофагов в 1 мл БАЛ.
В - Количество лимфоцитов в 1 мл БАЛ.
× - статистически значимо отливается от «РСВ/PBS» по U-критерию Манна-Уитни. N=10.
Фигура 10. Гистологическое изменение ткани легких.
А - Площадь перибронхиального инфильтрата измеряемая в пике.
Б - Общее количество клеток в инфильтрате.
В - Толщина стенки бронхов измеряемая в пике.
× - статистически значимо отливается от «РСВ/PBS» по U-критерию Манна-Уитни. N=10.
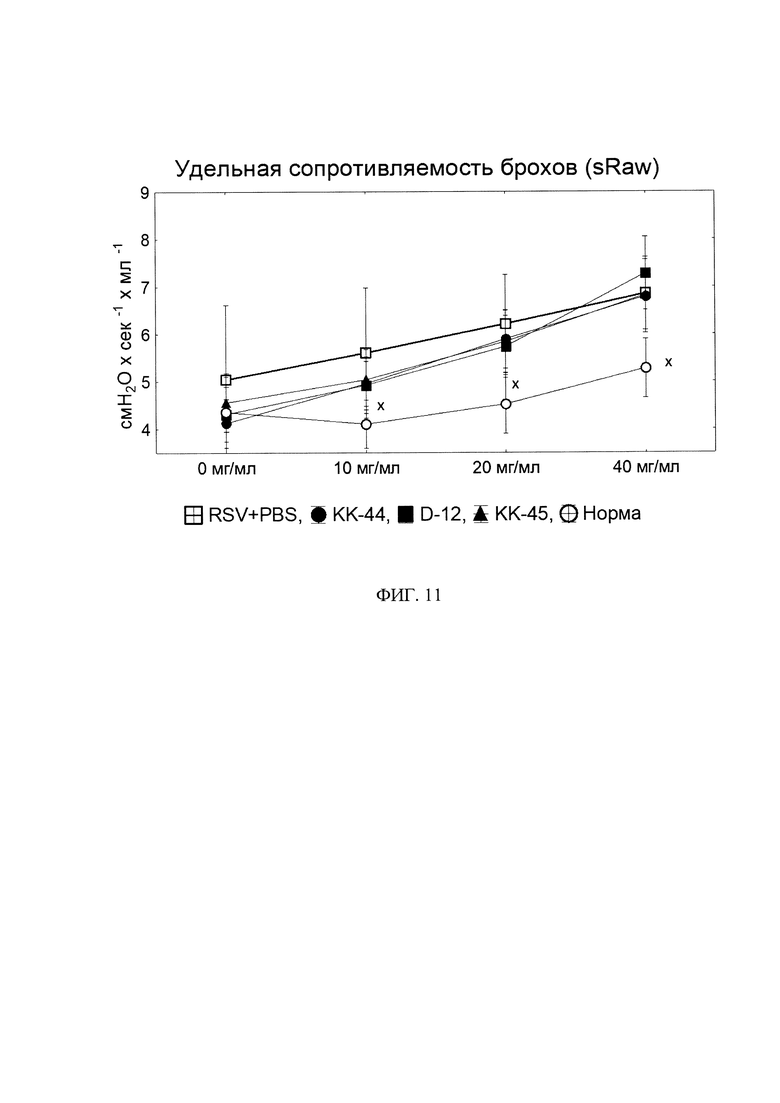
Фигура 11. Удельная сопротивляемость бронхов.
х - статистически значимо отливается от «РСВ/PBS» по U-критерию Манна-Уитни. N=10.
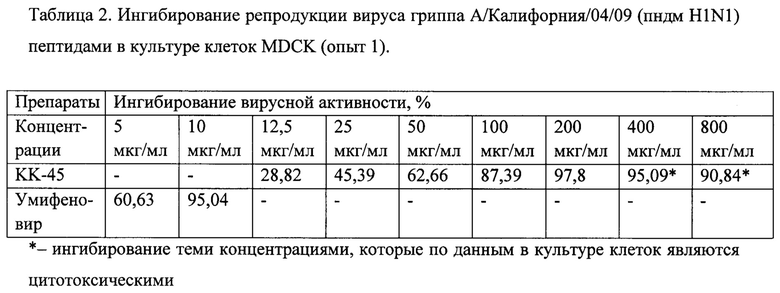
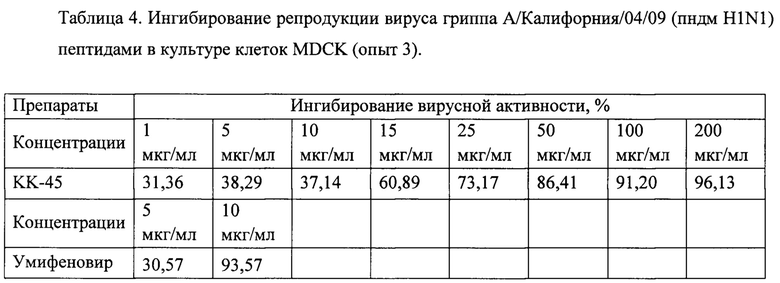
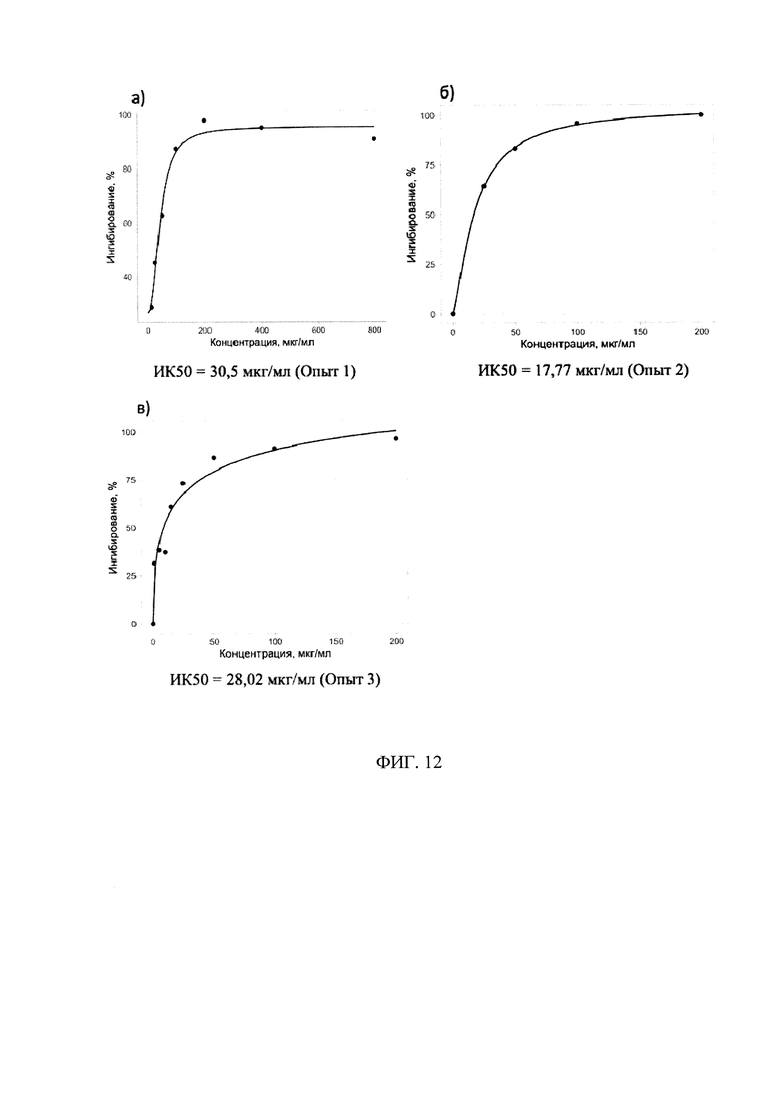
Фигура 12. Противовирусная активность препарата КК-45 в культуре клеток MDCK (1, 2 и 3 опыты)
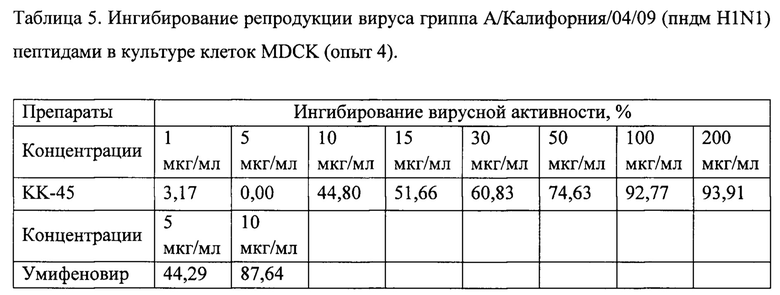
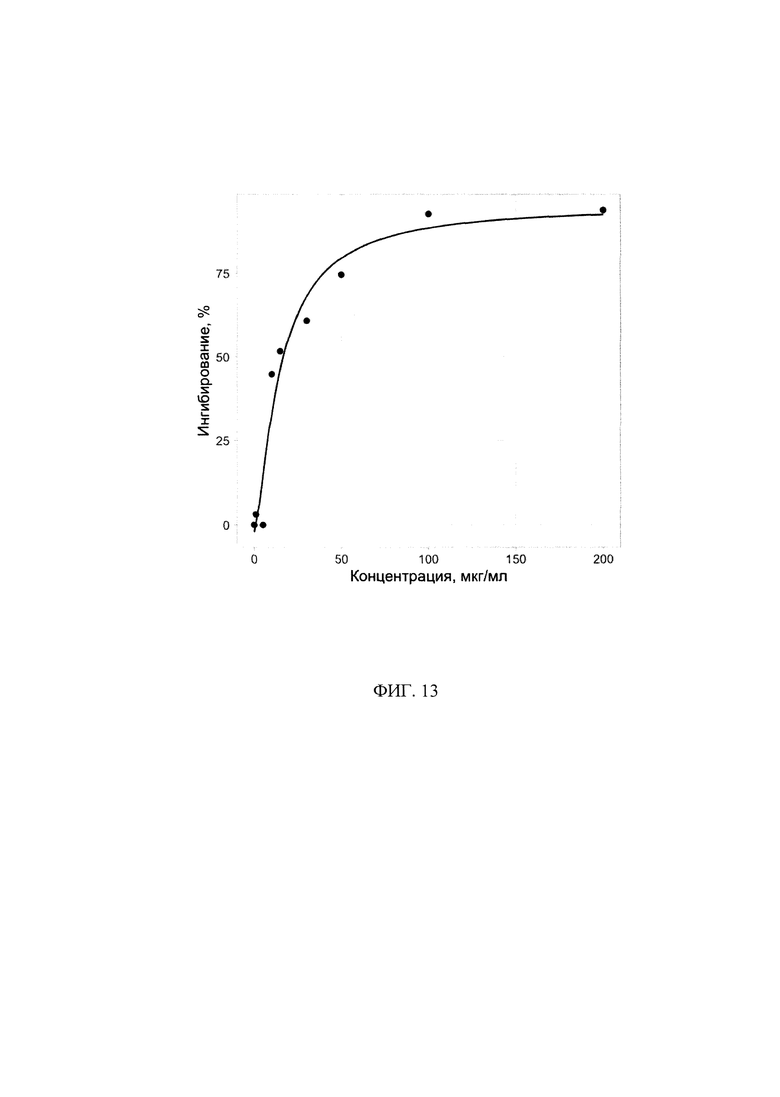
Фигура 13. Противовирусная активность препарата КК-45 в культуре клеток MDCK (4 опыт).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Пример 1. Получение пептида, имитирующего фрагмент ∅-сайта (HRA-домена) F-белка вируса РСВ.
Один из подходов к разработке антивирусных препаратов - создание пептидных ингибиторов, предотвращающих процессы прикрепления и слияния вируса с клетками-мишенями. В одной из работ [Lambert и др., 1996]. Место для ввода текста. была продемонстрирована антивирусная активность пептидов из HR2-домена F-белка. Домены HR2 и HR1 участвуют в структурных перестройках белка F, что необходимо для слияния вируса с клеткой и инициации инфекционного процесса. Спроектированные Lambert и коллегами [Lambert и др., 1996] пептиды связывались с доменом HR2 и тем самым препятствовали инфекции.
В то же время домен HR1 белка F вируса РСВ также имеет большое значение для инфекционного процесса. Недавние исследования показали важность нулевого (∅) антигенного сайта белка слияния F в инфекции. Этот антигенный сайт располагается на границе HR1-домена и домена D1 (Фиг. 1А). Антитела, направленные против этого сайта, в значительной степени предотвращали перестройку белка F из префузионной в пост-фузионную конформацию, что приводило к уменьшению инфекции.
Учитывая это, мы спроектировали линейный пептид из нулевого сайта (АА201-212) белка F - КК-44. Также к линейной части QLLPIVNKQSC мы добавили дендримерный фрагмент с N-конца (пептид КК-45). Также был синтезирован малый дендримерный пептид D-12, использованный в качестве контроля влияния дендримерной части пептида (Фиг. 1Б, таблица 1).
Для синтеза пептида использовали автоматический синтезатор пептидов PS3 (Protein Technologies Inc., США). Схема синтеза представлена на фигуре 2.
Для очистки пептида использовалась высокоэффективная жидкостная хроматография (ВЭЖХ) в виде системы LC-20 Prominence (Shimadzu, Япония). При элюировании использовали линейный градиент 5-70% элюента В (25 минут) при 30 мл/мин с детектированием при 226/280 нм. Элюент А содержал 0,1% TFA/H20; элюент В содержал чистый ацетонитрил (Sigma-Aldrich). Стационарна фаза - Kromasil Eternity С18.
Функциональные группы боковых цепей Fmoc-аминокислот (AnaSpec, США/ Sigma Aldrich, Германия / Iris Biotech, Германия) были защищены трет-бутилом (t-Bu) для гидроксильных и карбоксильных групп, трет-бутилоксикарбонилом (Boc) или флуоренилметоксикарбонилом (Fmoc) для е-аминогруппы лизина, тритильной группой (Trt) для SH-группы цистеина и амидной группы аспарагина и глутамина, и 2,2,4,6,7-пентаметил-дигидробензофуран-5-сульфонил (Pbf) для гуанидиновой группы аргинина. Синтез осуществляли автоматически на пептидном синтезаторе PS3 (Protein Technologies Inc., США) с использованием сополимера 1% дивинилбензола с полистиролом (смола Fmoc-Rink Amide-MBHA) или смол на ПЭГ-основе (Rink-amide chemmatrix, TGR-, TGA-).
Для конденсации использовали смесь HBTU/NMM. Растворителем, использованным для всех стадий синтеза, был N,N-диметилформамид (ДМФА) или смесь ДМФА с N-метилпирролидоном (МП); снятие Fmoc-защиты после каждой стадии конденсации проводили в растворе 4-метилпиперидина (для Fmoc), а полное снятие защит осуществляли путем использования трифторуксусной кислоты (TFA) (все защитные группы боковых цепей, кроме Fmoc, были лабильными для TFA).
Схема твердофазного синтеза основана на повторяющемся цикле стандартных операций, который включает:
1) На старте навеску исходной смолы, например Fmoc-Rink Amide-MBHA, вносят в реактор, добавляют ДМФА, энергично перемешивают суспензию до полного набухания смолы, операцию повторяют 3 раза по 10-30 мин, удаляя растворитель фильтрованием под давлением.
2) Удаление Fmoc-защиты 20% раствором 4-метилпиперидина (МПП) в течение 10-20 мин 2 раза.
3) Промывку смолы ДМФА при тщательном перемешивании, операцию повторяют 3 раза, удаляя отработанный растворитель фильтрованием под давлением.
4) Присоединение аминокислоты путем добавления в реактор активированной Fmoc-аминокислоты в среде ДМФА или N-метилпирролидона (МП) и инкубацией в течение 30 мин -2 ч.
5) Промывку смолы с пептидом ДМФА при тщательном перемешивании, операцию повторяют 3 раза, удаляя отработанный растворитель фильтрованием под давлением.
6) Операции по пунктам 2-5 повторяются.
Отщепление пептида от смолы проводят смесью трифторуксусной кислоты (95%) с добавлением тиоанизола, 1,2-этандитиола, триизопропилсилана и деионизированной воды. Сырой пептидный продукт из этой смеси осаждают сухим диэтиловым эфиром, осадок дополнительно промывают диэтиловым эфиром и экстрагируют 5% водной уксусной кислотой и экстракт высушивали лиофильно, получая «сырой пептид». Очистку сырого пептида проводят путем высокоэффективной жидкостной хроматографии на обращенной фазе (С18), используя градиентное элюирование.
Заявляемый ДКП - стабильное вещество при хранении в сухом виде. В результате был получен дендримерный пептид общей формулы:
[((Arg)2Lys)2LysGlnLeuLeuProIleValAsnLysGlnSerCys-C(O)NH2]+CF3COO-n, где карбоксильная группа цистеина находится в форме амида с (C(O)NH2), и где CF3COOHn обозначает трифторуксусную кислоту, которая в форме аниона формирует соль с пептидом-противоином, имеющим щелочные/основные свойства (n=9). Содержание в молях кислоты-аниона соответствует числу молей позитивного заряда пептидного катиона. Число зарядов при нейтральном рН равно 9 (фиг. 3).
Пример 2. Цитотоксическая активность дендримерного пептид КК-45.
Цитотоксичность пептидов изучали с помощью МТТ-теста на культуре клеток МА-104 (эмбриональная почка обезьяны) в широком диапазоне концентраций от 0,04 до 2,5 мг/мл. Данная культура была выбрана в связи с тем, что последующие исследования антивирусной активности проводились на этой же культуре клеток. Для этого в 96-лучночный планшет засевали 20000 клеток/лунку в объеме 100 мкл полной среды Игла-Мем (10% ЭТС, 300 мг/л L-глутамина, 50 мкг/мл гентамицина, 25 мМ HEPES). На следующий день в клетки вносили пептиды в различных концентрациях. Через сутки в клетки вносили 25 мкл раствора МТТ (4 мг/мл) и инкубировали 4 ч. После чего клетки лизировали раствором (20% SDS на 0,02н водном растворе H2SO4). Далее измеряли оптическую плотность при длинах волн 570 и 650 нм и на основе этих данных рассчитывали значения СС50 (50%-ая цитотоксическая концентрация - концентрация тестируемого вещества, необходимая для гибели 50% клеток).
В результате тестирования 3-х пептидов установлено, что линейный пептид КК-44 и контрольный дендримерный пептид D12 являются не токсичными в данном диапазоне концентраций. В то же время комплексный дендримерный пептид КК-45 проявлял более выраженный цитотоксический эффект; значение СС50 составляло 0,50±0,09 мг/мл (Фиг. 4). По всей видимости, за счет высокой плотности положительного заряда и относительно крупного размера, данный пептид оказывают более выраженное повреждающее действие на мембрану клетки, приводя к потере ее жизнеспособности.
Пример 3. Вирулицидная активность к РСВ пептидов.
Была изучена вирулицидная активность этих пептидов по методике, описанной в статье [Donalisio и др., 2012] с некоторыми модификациями. Кратко, пептиды в диапазоне нетоксических концентрации 250 мкг/мл были инкубированы с вирусом (Титр=1000 синцитиеобразующих единиц/мл) 2 ч при комнатной температуре после чего вирус титровали на заранее подготовленном монослое клеток МА-104, а через 5 суток подсчитывали количество синцитиев методом световой микроскопии. В качестве отрицательного контроля вирус обрабатывали физиологическим раствором (ФР), а в качестве положительного контроля вирус обрабатывали нейтрализующими моноклональными антителами (mAb) в концентрации 10 мкг/мл.
В итоге наибольшим вирулицидным эффектом обладали нейтрализующие моноклональные антитела; общее количество синцитиев составило 4350±3165, а количество синцитиев в процентах от отрицательного контроля (вирус обрабатывался физиологическим раствором - ФЗ) составило 4±3%. Дендрмерный пептид КК-45 оказывал выраженной прямое вирулицидное действие на вирионы РСВ, количество синцитиев составило 12400±6396, процентная доля от отрицательного контроля составила 10±6%. Линейный пептид КК-44 также проявлял высокую вирулицидную активность, количество синцитиев 7767±3801, а процент синцитиев от группы отрицательного контроля составил 7±5%. В то же время контрольный дендрмерный пептид D-12 не оказывали такого эффекта (Фиг. 5А, 5Б). По всей видимости крупные дендримерные пептиды с высокой плотностью позитивного заряда (+8 для КК-45) оказывают прямое дестабилизирующее действие на оболочку РСВ, что в итоге приводит к снижению жизнеспособности вируса.
Пример 4. Блокирование лигандов на поверхности РСВ.
Пептиды были изучены в тесте на ингибирование лигандов на поверхности вирионов РСВ. Для этого засевали клетки МА-104 в 96-луночный планшет в количестве 15 тыс/лун в полной среде ДМЕМ, инкубировали ночь до образования монослоя и отмывали от сывороточной среды физиологическим раствором, вносили 100 мкл/лун холодной бессывороточной среды ДМЕМ и охлаждали планшет до +4С. Далее пептиды титровали до получения следующих концентраций 500, 250, 125, 62,5, 31,25 мкг/мл, охлаждали на льду и вносили охлажденный до +4С вирус в количестве 50 бляшкообразующих единиц (БОЕ) на лунку объемом 100 мкл бессывороточной среды ДМЕМ. Инкубировали смесь пептид/РСВ при +4С в течение 1 ч после чего вносили ее к клеткам МА-104 в лунки предварительно отмытого и охлажденного до +4С планшета. Инкубировали смесь пептид/РСВ с клетками 2 ч при +4С, удаляли содержимое из лунок вместе с не связавшимся вирусом, меняли среду на бессывороточную ДМЕМ и инкубировали при +37С в СО2-инкубаторе в течение 3-5 суток до образования синцитиев. Далее количество синцитиев в лунке подсчитывали методом световой микроскопии (учитывая число блящек). При таком дизайне эксперимента пептиды, обладающие способностью взаимодействовать с вирионами, будут связываться с лигандами (скорее всего с белками F и G вируса) на их поверхности и при внесении к клеткам препятствовать процессу прикрепления/слияния.
Проведенные исследования показали, что инкубация линейного КК-44 с вирусом не приводит к снижению инфекционного процесса; ЕС50 составляет более 500 мкг/мл. Это может свидетельствовать о его неспособности нарушать структурные перестройки в белке F, необходимые для слияния вируса с клетками, или о низкой стабильности пептида. В то же время дендримерная форма пептида - КК-45 значительно подавляет образование синцитиев (ЕС50=33±3 мкг/мл). Стоит отметить, что контрольный дендримерный пептид D-12 также обладал анти-РСВ эффектом в данном тесте, но значительно (в 6 раз) более слабым (ЕС50=197±48 мкг/мл) (фиг. 6А).
В дополнительных экспериментах мы исследовали способность пептидов взаимодействовать с РСВ методом ИФА по методике сходной с опубликованной в работе [Donalisio и др., 2012]. Для этого пептиды в концентрациях 0,25, 2,5, 25 и 250 мкг/мл сорбировали на планшет при +37С в течение 2 ч, после чего двукратно отмывали (0,25%Tween-20/PBS) и осуществляли забивку 2% обезжиренным сухим молоком (SIGMA) при +4С в течение ночи. В качестве отрицательного контроля осуществляли сорбцию 1% BSA (ПанЭко) (соответствует значениям 0 мг/мл на фигуре 6А) и гепарина в аналогичном диапазоне концентраций (от 0,25 до 250 мкг/мл). Далее после трехкратной отмывки вносили РСВ в бессывороточной среде ДМЕМ (Т=10000 БОЕ/мл) и инкубировали 2 ч при +37С.После двукратной отмывки вносили антитела против F-белка вируса (РСВ Fusion Protein Monoclonal Antibody (C106D) (ThermoFisher)) в разведении 1/200 и инкубировали 1 ч при комнатной температуре. После двукратной отмывки вносили вторичные биотинилированные антитела (rat anti-mouse IgG1-biotin (BD)). Отмывали 2 раза и вносили конъюгат SaV-HRP (BD) в разведении 1/1000 с последующей инкубацией 1 час при комнатной температуре. После трехкратной отмывки вносили ТМБ (BD) и инкубировали 10-20 минут. Реакцию останавливали 2н H2SO4. Оптическую плотность (ОП) измеряли на планшетном фотометре Multiskan GO (ThermoFisher) при двух длинах волн: 450 и 620 нм. Программным обеспечением прибора высчитывалась разница между ОП450 и ОП620 (ΔОП).
Проведенное исследование показало, что линейный пептид КК-44 значительно связывался с РСВ; ΔОП составляла 0,97±0,10 (фиг. 6Б). Дендримерная форма этого пептида - КК-45 и контрольный дендримерный пептид D-12 также связывались РСВ, но с меньшей интенсивностью; ΔОП составили 0,67±0,14 и 0,73±0,13 (фиг. 6Б). Эти данные свидетельствуют о том, что линейный пептид КК-44, несмотря на связывание его с вирусом не оказывает ингибирующего действия на инфекционный процесс (фиг. 6Б). По всей видимости, данный пептид не приводит к нарушению функционирования белка F и/или не стабилен в условиях длительного культивирования. В то же время пептид КК-45 значительно ингибирует дальнейший инфекционный процесс (фиг. 6Б). Скорее всего, это обусловлено тем, что пептид КК-45, специфически связывается с вирионами посредством линейной С-концевой последовательности, за счет наличия разветвленной структуры на N-конце, стерически препятствует нормальной функции белка F вируса, а именно, нарушает его структурную перестройку из префузионной в постфузионную форму.
Контрольный пептид D-12 также значительно связывался с РСВ, в данном случае может иметь место его неспецифическое связывание за счет электростатического взаимодействия; D-12 с высокой плотностью положительного заряда взаимодействует с отрицательно заряженными участками вириона (например, липидными рафтами в бислойной мембране). Слабо выраженная антивирусная активность данного пептида (D-12) в тесте на ингибирование лигандов РСВ (фиг. 6Б) вероятнее всего объясняется его способностью связываться с поверхностью вириона, что дестабилизирует его оболочку.
Пример 5. Ингибирование связывания РСВ с рецепторами клеток.
Для оценки способности пептидов ингибировать вход вируса в клетки-мишени клеточную линию МА-104 засевали в 96-луночный планшет в полной питательной среде ДМЕМ (10% ЭТС, 300 мг/л L-глутамин, 25 мМ HEPES, 50 мкг/мл Гентамицина) и культивировали в 5% CO2 при 37°С так, чтобы через сутки конфлюентность монослоя составила 100%. Отмывали планшет от полной среды ДМЕМ физиологическим раствором (который был охлажден до +4С). Далее манипуляции проводили на льду при +4С, т.к. при этой температуре РСВ способен прикрепляться к клеткам-мишеням, но не способен осуществлять слияние с ними. После отмывки в лунки планшета вносили по 50 мкл холодной бессывороточной среды ДМЕМ (600 мг/л Л-глутамина, 25 мМ HEPES, 50 мкг/мл Гентамицина) и тестируемые пептиды, которые титровали в диапазоне нетоксичных концентраций, и инкубировали 30 минут на льду. В качестве положительного контроля применяли гепарин, который связывается с РСВ и конкурентно препятствует связыванию вируса с клеточными рецепторами - гепаран-сульфатами [Donalisio и др., 2012]. Далее в лунки вносили РСВ А2 в количестве 50 БОЕ на лунку в объеме 50 мкл/лунку бессывороточной ДМЕМ, инкубировали вирус 2 часа на льду. Затем отмывали не связавшийся вирус и пептиды, вносили в лунки 100 мкл бессывороточной ДМЕМ и инкубировали при +37С в CO2-инкубаторе 2-3 суток. Подсчитывали количество синцитиев методом световой микроскопии [Donalisio и др., 2012]. На основе данных о влиянии различных концентраций пептидов на количество синцитиев были рассчитаны значения ЕС50 и ЕС75, т.е. концентрации пептидов, необходимых для ингибирования прикрепления вируса к клетке на 50% (в 2 раза) и 75% (в 4 раза), соответственно.
Проведенные исследования показали, что КК-44 в незначительной степени ингибирует прикрепления вируса к клетке; значения ЕС50 составили 979±244 мкг/мл. Пептида D-12 не демонстрировал ингибирующего эффекта. Самую высокую способность ингибировать прикрепление РСВ к клеткам-мишеням продемонстрировал пептид КК-45 значение ЕС50 составило 47±8 мкг/мл, что сопоставимо с положительным контролем гепарином (ЕС50 - 48±11 мкг/мл) (фиг 7А).
В отдельных экспериментах была изучена способность пептидов связываться с клеточным рецептором для РСВ - белком нуклеолином. Для этого пептиды в концентрациях 2,5, 25 и 250 мкг/мл сорбировали на планшет при +37С в течение 2 ч после чего двукратно отмывали (0,25%Tween-20/PBS) и осуществляли забивку 2% обезжиренным сухим молоком (SIGMA) при +4С в течение ночи. В качестве отрицательного контроля осуществляли сорбцию 1% BSA (ПанЭко) (соответствует значениям 0 мг/мл на фигуре 6) и гепарина в аналогичном диапазоне концентраций (от 2,5 до 250 мкг/мл). Далее после трехкратной отмывки вносили 100 мкл 1 мкг/мл раствора нуклеолина NCL (Human NCL / Nucleolin Protein (Recombinant 6His, N-terminus) (aa2-482) (LSBio)) и инкубировали 2 ч при +4С. После двукратной отмывки вносили кроличьи поликлональные антитела против нуклеолина (Nucleolin Polyclonal Antibody (ThermoFisher)) в разведении 1/500 и инкубировали 1 ч при комнатной температуре. После двукратной отмывки вносили детектирующие анти-кроличьи IgG-антитела меченые HRP (Goat anti-Rabbit IgG (H+L) Secondary Antibody, HRP (ThermoFisher)). Отмывали 3 раза и вносили ТМБ (BD) инкубировали 15-30 минут.Реакцию останавливали 2н H2SO4. Оптическую плотность (ОП) измеряли на планшетном фотометре Multiskan GO (ThermoFisher) при двух длинах волн: 450 и 620 нм. Программным обеспечением прибора высчитывалась разница между ОП450 и ОП620.
Как было сказано выше пептиды КК-44 и КК-45 моделируют участок белка F вируса, необходимый для связывания с клеткой-мишенью; при этом КК-45 -дендримерный пептид, содержащий линейный участок F-белка вируса АА201-212 и дендримерную часть (R4K2K), пептид КК-44 имеет линейную структуру АА201-212. В качестве контроля был дополнительно синтезирован дендримерный пептид D-12 (R4K2K).
Проведенные исследования показали, что дендримерная форма КК-45 значительно связывается с нуклеолином (ΔОП=0,79±0,01), в то время как линейная форма пептида КК-44, а также контрольный дендримерный пептид D-12 связываются заметно слабее; ΔОП - 0,22±0,01 и 0,28±0,04 (фиг. 7Б). Высокая антивирусная активность пептида КК-45, скорее всего, объясняется его способностью конкурентно связываться с рецептором -нуклеолином. Линейный пептид КК-44 не проявлял выраженной анти-РСВ-активности, что также может объясняться значительно более слабой способностью связываться с этим рецептором (фиг. 6).
Пример 6. Антивирусная активность пептида КК-45 в легких лабораторных животных.
После того как была установлена антивирусная активность пептида КК-45 в экспериментах in vitro мы провели исследования антивирусных свойств пептида на лабораторных животных.
Мыши самки, линии BALB/c весом 20-22 г возрастом 10-12 недель был разделены на пять групп (по 10 мышей в каждой). Животных с 1 по 4 группы инфицировали (интраназально) РСВ А2 однократно в объеме 50 мкл PBS («ПанЭко», Россия) в дозе 5×106 ТЦД50/мышь. Сухой пептид разводили в деионизованной воде, после чего терапия полученной композицией осуществлялась интраназально в объеме 50 мкл/мышь. Отдельные группы животных получали пептиды КК-45, КК-44 и D12 в дозе 0,0045 мкМ/мышь, что соответствует 10 мкг/мышь, 6 мкг/мышь и 4,7 мкг/мышь соответственно. Терапия длилась 5 дней по одному интраназальному введению в сутки. Животные 5-ой группы (Норма) никаким манипуляциям не подвергались. На 6-й день эксперимента мышей забивали методом цервикальной дислокации и отбирали образцы бронхоальвеолярного лаважа (БАЛ) для последующего определения вирусной нагрузки. Правое легкое отбирали и замораживали для выделения РНК и определения вирусной нагрузки методом количественной ПЦР.
Для получения БАЛ вскрывали грудную клетку мышей и сепарировали трахею, далее с использованием стерильного шприца (2 мл) промывали легкие через трахею 1 мл культуральной среды PRMI1640 (300 мг/л L-глутамина, 50 мкг/мл гентамицина, 25 мМ HEPES). Образцы БАЛ центрифугировали 350g 7 минут, после чего отбирали надосадок для последующего титрования вируса на монослое клеток. В итоге вирусная нагрузка в легких оценивалась двумя методами: количественной ПЦР гомогенатов легких и титрованием образцов БАЛ на монослое клеток.
Для оценки вирусной нагрузки методом количественного ПЦР из гомогенатов легких выделяли общую РНК набором RNeasy Mini Kit (Qiagen, Германия) согласно протоколу производителя. Концентрацию РНК определяли спектрофотометрически с использованием NanoDrop2000. Синтез кДНК проводили набором «Реверта-L» №К3-4-100 в соответствии с методикой производителя. Полимеразная цепная реакция в реальном времени проводилась набором «Синтол» ПЦР-Микс №М-428 по методике производителя. Количество копий рассчитывали по калибровочной кривой и нормализовали по концентрации общей РНК. Вирусную нагрузку выражали как количество копий РНК вируса на 1 мкг общей РНК.
Для количественного определения вирусной нагрузки методом титрования БАЛ готовили монослой чувствительных клеток МА-104. За день до эксперимента клетки засевали в 96-лучночный планшет (SPL Lifesciences, Южная Корея) в количестве 15000 клеток/лунку в 100 мкл полной среды Игла-MEM. После ночной инкубации, непосредственно перед титрованием, клетки промывали физиологическим раствором, и вносили в лунки 100 мл бессывороточной среды Игла-MEM. Далее осуществляли десятичное титрование БАЛ и в качестве позитивного контроля использовали РСВ А2 с известным титром. После чего клетки инкубировали при +37С в 5% атмосфере СО2 в течение 4 часов и меняли среду в лунках на полную питательную среду Игла-MEM. После чего клетки инкубировали при +37С в 5% атмосфере СО2 в течение 1 суток и меняли полную питательную среду Игла -MEM на среду OverLay (0,6% агароза в бессывороточной среде Игла-MEM). Инкубировали при +37С в 5% атмосфере СО2 в течение 5-6 суток. Далее осуществляли фиксацию клеток формалином (10% нейтральный забуференный) по 100 мкл/лун в течение ночи. На следующий день для удаления агарозной среды из лунок аккуратно промывали планшеты под проточной водой и вытряхивали среду из лунок до полного удаления среды. После фиксации монослоя осуществляли 3 последовательных отмывки буфером PBS-T (на 1 л PBS/ 0,5 мл Tween 20). Далее в планшеты вносили по 200 мкл блокирующего буфера (1 л PBS/ 0,5 мл Tween 20+25 г сухого обезжиренного молока) на лунку, после чего инкубировали их в течение 30 минут на шейкере при комнатной температуре. После инкубации плейты трехкратно отмывали и вносили 100 мкл раствора антител РСВ Fusion Protein Monoclonal Antibody (Invitrogen) в буфере для разведения образцов в разведении 1/50. После инкубации планшеты трехкратно отмывали и вносили 100 мкл раствора антител, меченных биотином против rat biotin-ani-mouse-IgG1 (BD) в разведении 1/1000. Затем планшеты инкубировали в течение 1 часа при комнатной температуре, после чего последовательно отмывали их три раза и вносили 100 мкл раствора конъюгата пероксидазы хрена со стрептовидином SR-HRP (BD) в разведении 1/1000. Планшеты инкубировали при комнатной температуре в течение часа. После инкубации планшеты отмывали 4 раза и вносили в них по 50 мкл на лунку субстрата (ТМБ мембранный, Sigma, США). Далее планшеты инкубировали в темноте до проявки (в среднем 25-30 минут). Для остановки реакции использовали 50 мкл дистиллированной воды. Окрашенный синцитий подсчитывали методом световой микроскопии. Вирусную нагрузку выражали как количество синцитиев в 1 мл БАЛ.
Методом количественной ПРЦ показано снижение числа копий вирусной РНК у мышей, получавших все три пептида КК-45, D-12 и КК-44 (фиг. 8А). Количество копий vRNA на 1 мкг РНК в группе, получавший пептид КК-44, уменьшилось в 1,5 раза по сравнению с группой РСВ. Самый высокий элиминирующий эффект в отношении vRNA продемонстрировал пептид КК-45, почти в 3 раза меньше копий по сравнению с группой РСВ/PBS. Применение короткого дендримерного пептида D12 также приводит к снижению вирусной нагрузки, но не так значительно, приблизительно в 1,8 раза. Полученные данные в целом подтверждают результаты исследований in vitro, проведенных на предыдущем этапе, в которых дендримерный пептид КК-45 проявлял самую высокую антивирусную активность.
Следовательно, эти дендримерная структура пептида обладает более эффективным профилем биологической активности, в сравнении с линейными пептидами.
Методом титрования БАЛ показано, что интраназальное применение дендримерного пептида КК-45 в наибольшей степени уменьшало количество синцитиев в БАЛ в 13 раз. Применение короткого дендримерного пептида D12 также приводит к снижению вирусной нагрузки, но не так значительно в 2,2 раза, в то время как линейный пептид КК-44 не оказывает антивирусного эффекта (фиг. 8Б). Различия в полученных данных объясняются особенностью методов детекции; методом титрования выявляются жизнеспособные вирионы, в то время как ПЦР дополнительно выявляет аберрантные вирионы и внутриклеточную РНК вируса.
Пример 7. Оценка влияния пептида КК-45 на воспалительную реакцию легких и гиперреактивность дыхательных путей.
Помимо оценки вирусной нагрузки в легких также оценивалась выраженность воспалительных реакций в этом органе: для этого оценивали клеточный состав БАЛ, гистологические изменения в ткани легких, а также гиперреактивность бронхов.
При оценке клеточного состава БАЛ дыхательные пути через трахею с помощью шприца промывались дважды 1 мл бессывороточной среды Игла-MEM. После чего измеряли клеточность БАЛ на автоматическом гематологическом анализаторе Abacus Junior Vet (Diatron, Австрия). Результат выражали в количестве клеток в 1 мл БАЛ. После измерения клеточности образцы центрифугировали 10 мин при 150g. Осажденные клетки наносились на предметное стекло, высушивались и фиксировались в метаноле в течение 10 минут, после чего окрашивались азуром и эозином. Подсчет клеточного состава БАЛ проводили методом световой микроскопии.
В проведенном эксперименте мы оценивали воспаление по изменению количества клеток в образцах БАЛ. Мы показали, что происходит статистически значимое снижение общего количества клеток в БАЛ у животных, получавших пептиды D-12 и КК-45 на 45% и 50%, соответственно (фиг. 9А). Это снижение достигается за счет двукратного уменьшения количества инфильтрирующих макрофагов (фиг. 9Б) и лимфоцитов (фиг. 9В). Нейтрофилы и эозинофилы были представлены единичными клетками (данные не представлены). В то же время интраназальное введение линейного пептида КК-44 не приводит к уменьшению общего количества клеток и макрофагов в БАЛ, но при этом количество лимфоцитов уменьшалось в 2 раза по сравнению с мышами контрольной группы (фиг. 8). Эти данные показывают, что дендримерный пептид КК-45, снижая вирусную нагрузку в легких, тем самым способствуют уменьшению воспаления респираторного тракта.
Для гистологического анализа отбирали левое легкое для приготовления микропрепаратов и помещали его в 10% формалин, обезвоживали путем проводки по спиртам и заливали образцы в парафин. Микротомированием парафиновых блоков получали срезы легких толщиной 2-4 мкм. Полученные препараты окрашивали гематоксилином и эозином (Bio-Optica, Италия) для идентификации эозинофилов, нейтрофилов и лимфоцитов. Гистологическое исследование микропрепаратов легких осуществляли на световом микроскопе (Olympus, Япония). Толщину бронхиального эпителия измеряли в пикселях с использованием микрофотографий ткани легкого при увеличении в 200 раз и программного обеспечения Altami Studio (Альтами, Россия). Анализировали 5 бронхов у каждого экспериментального животного.
Проведенный гистологический анализ ткани легких подтвердил уменьшение количества инфильтрирующих клеток; площадь перибронхиальных инфильтратов и количество в них клеток было существенно снижено у мышей, получавших пептиды КК-44, D-12 и КК-45; при этом максимальное уменьшение этих показателей было в группе животных, которым интраназально вводили дендримерный пептид КК-45 (фиг. 10А, 10Б). Значения толщины стенки бронхов статистически значимо отличалось только в группе норма (фиг. 10В). Эти данные подтверждают снижение РСВ-индуцированного воспаления легких.
Оценивали также гиперактивность бронхов (ГРБ) с использованием прибора для неинвазивной плетизмографии - FinePointe NAM (Buxco, США). ГРБ определяли при появлении бронхоспазма у экспериментальных мышей, вызванного аэрозольным введением метахолина (Sigma-Aldrich, США) в возрастающих концентрациях (6,25; 12,5; 25 мг/мл), после чего в автоматическом режиме оценивали удельную сопротивляемость дыхательных путей (ее выражали в см вод. ст. × с).
Воспаление, вызванное РСВ - инфекцией, приводит к нарушению дыхательной функции легких, что выражается в увеличении сопротивляемости бронхов. Мы измерили этот показатель методом неинвазивной плетизмографии и установили, что инфицирование мышей РСВ в дозе 107 TCID50 значительно увеличивает удельную сопротивляемость бронхов (sRaw) на 25%-35% в сравнении с неинфицированными животными. В то же время интраназальное введение пептидов КК-45, D12 и КК-44 не приводит к восстановлению этого показателя несмотря на снижение уровня воспаления (фиг. 11). Скорее всего сопротивляемость бронхов в большей степени вызвана иными факторами, связанными с протеканием инфекции, а не с воспалением.
Пример 8. Определение антивирусной активности пептида КК-45 в отношении вируса гриппа.
Учитывая, что оболочечные вирусы имеют сходство в строении вириона и в механизмах проникновения в клетку, было важно изучить обладает ли разработанный пептид КК-45 специфичным эффектом в отношении РСВ или его спектр антивирусной активности более широкий. Для этого были изучены его антивирусные свойства в отношении вируса гриппа в экспериментах in vitro.
Для исследования противовирусной активности пептидов в отношении вируса гриппа традиционной моделью является инфекция в культуре клеток MDCK. В связи с поставленной целью в задачи исследования входило:
1) Оценить цитотоксичность предоставленных образцов in vitro, а именно определение ЦТД50% (цитотоксической дозы 50%) и МПД (максимально переносимой дозы) катионных пептидов в культуре клеток MDCK;
2) Оценить противовирусную эффективность предоставленных образцов in vitro, а именно определение ИК50 (ингибирующей концентрации 50%) пептидов в отношении вируса гриппа при лечебно-профилактической схеме добавления.
В качестве препарата сравнения использовали Умифеновир. Клетки MDCK (Madin Darby canine kidney) - перевиваемая культура клеток почки собаки). Штамм вируса гриппа А/Калифорния/04/2009 (пндм H1N1 2009), полученный из ВОЗ). Вирус выращивали в аллантоисной полости 9-дневных куриных эмбрионов, аликвоты вируса были заморожены и хранились при температуре -80°С. Титр стока составлял не менее 106-107 ЭИД50/мл или 105-106 БОЕ/мл. Во всех трех экспериментах использовали одну и ту же аликвоту из одного стока.
Использовали стандартный МТТ-текст для определения жизнеспособности клеток MDCK после их культивирования с пептидами в течение 3 суток. Каждая точка эксперимента была поставлена в 4 повторностях. Максимальная концентрация препарата, не изменяющая жизнеспособность клеток по сравнению с контролем принималась за максимально переносимую концентрацию (МПК). Концентрация субстанций, уменьшающая значение оптической плотности на 50% по сравнению с контролем, принималась за 50% цитотоксическую дозу (ЦТД50), вычисление которой проводили при построении кривой зависимости (выживания) с использованием пакета "drc" в Rstudio.
Изучение противовирусной активности образцов проводили методом ИФА, модифицированном для определения активности препаратов в культуре клеток как рекомендовано разрешительными органами РФ [Методические указания по изучению ИЛП: Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ, 2012].
Клетки MDCK рассаживали в 96-луночные планшеты для культивирования клеток с плоским дном (35000 клеток/лунку) и выращивали до полного монослоя в КС. Перед заражением вирусом клетки 2 раза промывали средой MEM без сыворотки. Далее исследуемые соединения добавляли в концентрациях, указанных в таблицах 2 и 3 в объеме 100 мкл/лунку PC (среда MEM с двойным набором аминокислот и витаминов без сыворотки с 10 мМ глутамина и антибиотиков). Часть лунок использовали для контроля вируса и клеток. Каждая точка эксперимента была поставлена в 4 повторностях. Далее были проведены две модификации опытов. В первой модификации (количество экспериментов n=3) после инкубации клеток с исследуемыми препаратами в течение 2 часов при 37°С в лунки, исключая клеточный контроль, добавляли вирус на PC (100 мкл), при этом множественность заражения составляла приблизительно 0,01 БОЕ/ на клетку.
Во второй модификации экспериментов (количество экспериментов n=1) после инкубации препаратов с клетками, препараты удаляли, в лунки добавляли вирус на PC (100 мкл), множественность заражения составляла приблизительно 0,1 БОЕ на клетку. После инкубации в течение 1 часа, вирус удаляли и к клеткам опять добавляли препараты на PC (100 мкл), а затем к каждой лунке с препаратами добавляли по 100 мкл PC. В лунки клеточного и вирусного контроля добавляли по 200 мкл PC. Далее в обоих модификациях планшеты инкубировали в течение 24 часов в атмосфере 5% СО2 при 37°С. После чего клетки планшета просматривали под инвертированным микроскопом, чтобы убедиться в отсутствии в них цитотоксических и цитопатических изменений.
Далее среду удаляли, клетки фиксировали 80% ацетоном на буфере 0,01М PBS (20 минут), высушивали и отмывали 3 раза PBS с 0,05% Твин-20. Все дальнейшие процедуры проводили в соответствии с техникой постановки ИФА для выявления антигена. К клеткам добавляли по 100 мкл раствора PBS с 1% фетальной сывороткой и 0,05% Твин-20 и инкубировали при 37°С в течение 30 минут. После удаления раствора к клеткам добавляли по 100 мкл моноклональных антител (МКА) к внутренним белкам вируса гриппа A (NP+M1), разведенным 1:1000 на ИФА-растворе. После инкубации с антителами в течение 1 часа при 37°С и последующей 3-х-кратной промывки в лунки вносили по 100 мкл IgG козы против IgG мыши, меченных пероксидазой хрена (Sigma) в разведении 1:5000 и инкубировали еще 1 час при 37°С. После 4-кратной промывки связанную пероксидазу выявляли добавлением в лунки 100 мкл 0,05% раствора 3,3',5,5'-тетраметилбензидина в 0,003% цитратном буфере рН=5,0, содержащем 0,003% Н2О2. Планшеты выдерживали 15-30 минут в темноте до появления окраски, реакцию останавливали добавлением 50 мкл 4Н H2SO4, далее оптическую плотность (ОП) измеряли на спектрофотометре для измерения оптического поглощения в 96-луночных планшетах при длине волны 450 нм. В качестве клеточного контроля использовали лунки, не зараженные вирусом. Процент ингибирования вирусной репродукции изучаемым соединением определяли по формуле:
Ингибирование, %=100 - ((ОПопыт - ОПкл.контр)/(ОПвир.контр - ОПкл.контр)) Для одной точки опыта использовали 4 лунки планшета, из которых определяли среднее значение, далее строили кривую доза- ответ, используя программу RStudio. Концентрация препарата, уменьшающая значение величины ОП на 50%, принималась за ингибирующую концентрацию 50 (ИК50) и также определялась автоматически с использованием пакета "drc" в RStudio. Индекс селективности (ИС) определяли по следующей формуле ЦТД50/ИК50.
Цитотоксичность в культуре MDCK
На основании полученных данных с использованием количественного метода МТТ были построены кривые зависимости выживаемости клеток от концентрации препаратов, которые обобщены в Таблице 2. По результатам трех экспериментов для КК-45 была выявлена ЦТД50=318,45±36,74 мкг/мл, МПК, (максимально переносимая концентрация), при которой не было изменения клеток по сравнению с клеточным контролем для КК-45 составляла 200 мкг/мл.
Антивирусная активность в отношении вируса гриппа
Изучение противовирусного действия препаратов в культуре клеток MDCK было проведено методом ИФА, модифицированном для определения активности препаратов в культуре клеток. Для начального исследования (опыт 1) был выбран диапазон концентраций, которые были изучены при определении цитотоксичности, для второго опыта основываясь на данных по цитотоксичности были выбраны концентрации ниже ЦТД50, а также концентрация 1 мкг/мл. В опытах для заражения клеточной культуры была использована множественность заражения клеток вирусами гриппа А/Калифорния/04/09 (пндм H1N1) - 0.01 БОЕ/на клетку. В качестве препарата сравнения был использован умифеновир в концентрациях 5 и 10 мкг/мл.
Полученные результаты представлены в Таблицах 2 и 3. Специфическим (селективным) считали подавление вирусной репродукции выше 30%, и при том, что эффективность подавления увеличивалась с увеличением концентрации. Из полученных данных в обоих опытах видно, что в нетоксических концентрациях КК-45 специфически ингибировал репродукцию вируса гриппа, ингибирование увеличивалось пропорционально с увеличением концентрации. Активность умифеновира, взятого в качестве препарата сравнения, также увеличивалась с увеличением его концентрации, соответствуя его активности в ранее проведенных исследованиях [Leneva и др., 2009; Y S Boriskin, I A Leneva, E-I Pecheur, 2008].
Поскольку изученные нами в 1 и 2 опытах некоторые концентрации являются токсическими, в следующей серии опытов мы уменьшили область концентраций, увеличив их количество внутри этого диапазона. Из полученных данных (таблица 4) видно, что пептид КК-45 специфически ингибировал репродукцию вируса гриппа, поскольку ингибирование увеличивалось пропорционально с увеличением концентрации. Построение кривых доза-ответ по результатам, полученным в 1-3 серии опытов, представлены на Фиг. 12. ИК50, определенные из этих кривых, для КК-45 составляли 30,5 мкг/мл (Опыт 1), 17,77 мкг/мл (опыт 2) и 28,02 мкг/мл (Опыт 3).
Опыты 1-3, проведенные нами показали, что некоторые из пептидов в нетоксичных для клеток MDCK концентрациях оказывали невысокое ингибирующее действие на вирусную репликацию, при этом для них не была выявлена зависимость эффекта от их дозы (Таблица 5). Предполагается, что предоставленные для изучения пептиды блокируют вход вируса в клетки. В связи с вышесказанным для более детальной проверки наличия активности препаратов нами была проведена серия опытов, в которой используемый нами метод определения активности был модифицирован таким образом, чтобы выявить ингибиторы ранних стадий вирусной репликации. Исследуемые препараты инкубировали с клетками в течение 2 часов при 37°С, далее препараты удаляли, в лунки добавляли вирус и инкубировали в течение 1 часа. Далее вирус удаляли и к клеткам опять добавляли препараты и инкубировали в течение 24 часов. Данные, полученные при такой модификации, совпали с данными, полученными ранее. Из изученных препаратов только препарат КК-45 оказывал вирусоспецифическое действие, при его низких концентрациях (1-5 мкг/мл) ингибирования практически не наблюдалось, с увеличением концентрации (от 10 до 50 мкг/мл) процент ингибирования увеличивался (45%-75%), достигая практически полного ингибирования вирусной репликации при концентрациях 100-200 мкг/мл.
Построение кривой доза-ответ по результатам этого опыта представлено на Фиг. 13. ИК50, определенная из этой кривой, для препарата КК-45 составила 16,7 мкг/мл (Таблица 6). Используемый в качестве препарата сравнения умифеновир также как и в 1-3 опытах показал специфическую противовирусную активность, при этом процент ингибирования при изученных концентрациях не отличались от таковых, полученных в предыдущих исследования и входили в диапазон концентраций, оцениваемых как активных, для лабораторных и эталонных штаммов вирусов гриппа для умифеновира [Boriskin и др., 2008],[Leneva и др., 2009]. Выведенные из кривых доза-ответ для КК-45 значения ИК50 во всех опытах, обобщены в Таблице 6. Принимая во внимание полученные в опытах по цитотоксичности данные, можно вывести индекс селективности СИ, который для КК-45 составлял 13,8 (Таблица 6). Из представленных данных следует, что противовирусные эффективные концентрации пептида КК-45 для вируса гриппа сопоставимы с данными эффективной концентрации пептида в отношении РСВ.
Таким образом, полученное средство, содержащее эффективное количество синтетического пептида с дендримерной (разветвленной) структурой КК-45 обладает противовирусными свойствами в отношении респираторно-синцитиального вируса (РСВ) и вируса гриппа. Проведенные исследования показали, что пептид КК-45 демонстрирует противовирусную активность как в экспериментах in vitro, так и in vivo. Механизм антивирусного действия пептида КК-45 состоит в блокировании входа вируса в клетку как за счет связывания с клеточным рецептором нуклеолином, так и за счет связывания с лигандами (белком F) на поверхности РСВ. Ингибирование проникновения в вируса клетку позволяет использовать полученный пептид КК-45 для профилактики респираторной вирусной инфекции, вызванной Orthopneumovirus. Дендримерный пептид КК-45, в виду высокой протеолитической стабильности, являются перспективным соединением для создания новых противовирусных препаратов.



ЛИТЕРАТУРА
1. Boriskin Y. S. и др. Arbidol: A Broad-Spectrum Antiviral Compound that Blocks Viral Fusion // Current Medicinal Chemistry. 2008. T. 15. C. 997-1005.
2. Donalisio M. и др. Inhibition of human respiratory syncytial virus infectivity by a dendrimeric heparan sulfate-binding peptide // Antimicrobial Agents and Chemotherapy. 2012. T. 56. №10. C. 5278-5288.
3. Fjell C. D. и др. Designing antimicrobial peptides: Form follows function // Nature Reviews Drug Discovery. 2012. T. 11. №1. C. 37-51.
4. Greber K. E., Dawgul M. Antimicrobial Peptides Under Clinical Trials // Current Topics in Medicinal Chemistry. 2017. T. 17. C. 620-628.
5. Gwyer Findlay E., Currie S. M., Davidson D. J. Cationic host defence peptides: Potential as antiviral therapeutics // BioDrugs. 2013. T. 27. №5. C. 479-493.
6. Janai HK M. M. Z. M. S. HR. Ribavirin_adverse_drug_reactions,_1986_to_1988.13 // The Pediatric Infectious Disease Journal. 1990. T. 9. №3. C. 209-211.
7. Lambert D. M. и др. Peptides from conserved regions of paramyxovirus fusion (F) proteins are potent inhibitors of viral fusion.: Merck & Co., Inc, 1996. 2186-2191 c.
8. Leneva I. А. и др. Characteristics of arbidol-resistant mutants of influenza virus: Implications for the mechanism of anti-influenza action of arbidol // Antiviral Research. 2009. T. 81. №2. C. 132-140.
9. Osminkina L. А. и др. Porous silicon nanoparticles as scavengers of hazardous viruses // Journal of Nanoparticle Research. 2014. T. 16. №6.
10. Pretorius M. А. и др. The role of influenza, RSV and other common respiratory viruses in severe acute respiratory infections and influenza-like illness in a population with a high HIV sero-prevalence, South Africa 2012-2015 // Journal of Clinical Virology. 2016. T. 75. C. 21-26.
11. Y S Boriskin, I A Leneva, E-I Pecheur S. J. P. Arbidol a broad-spectrum antiviral compound that blocks viral fusion // Curr Med Chem. 2008. T. 15. №10. C. 997-1005.
12. Хаитов M.P. и др. Интерференция РНК. Новые подходы к разработке противовирусных препаратов // Иммунология. 2010. Т. 31. №2. С. 69-76.
13. Методические указания по изучению ИЛП: Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ: Elsevier Inc, 2012.
| название | год | авторы | номер документа |
|---|---|---|---|
| Комбинированное лекарственное средство, обладающее противовирусным эффектом в отношении нового коронавируса SARS-CoV-2 | 2021 |
|
RU2746362C1 |
| СПОСОБ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ БРОНХИАЛЬНОЙ АСТМЫ, ОСЛОЖНЯЮЩИХ ЕЕ РЕСПИРАТОРНЫХ ВИРУСНЫХ ИНФЕКЦИЙ И ДРУГИХ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ ДЫХАТЕЛЬНЫХ ПУТЕЙ | 2012 |
|
RU2526146C2 |
| Композиция на основе пептида, подавляющего репликацию вируса гриппа А | 2018 |
|
RU2695336C1 |
| Штамм респираторно-синцитиального вируса RSV/Novosibirsk/66Hl/2018 для использования в диагностике респираторно-синцитиальной вирусной инфекции и исследования эффективности противовирусных препаратов in vitro | 2020 |
|
RU2746280C1 |
| Использование композиции, состоящей из катионного пептида LTP и молекул РНК против респираторных вирусов | 2015 |
|
RU2609857C1 |
| Способ получения профилактической противовирусной композиции на основе эпигаллокатехин-3-галлата (EGCG) | 2021 |
|
RU2771898C1 |
| ТЕСТ-СИСТЕМА ДЛЯ ДИАГНОСТИКИ ВОЗБУДИТЕЛЕЙ ОСТРЫХ РЕСПИРАТОРНЫХ ВИРУСНЫХ ИНФЕКЦИЙ | 2019 |
|
RU2733379C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К РАЗЛИЧНЫМ ШТАММАМ РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА | 2018 |
|
RU2713340C1 |
| F-БЕЛОК РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2464316C2 |
| ПРОТИВОВИРУСНОЕ СРЕДСТВО НА ОСНОВЕ МЕЛАНИНА | 2011 |
|
RU2480227C2 |






Изобретение относится к области медицины. Предложено применение способа интраназального введения пептида R4K2KQLLPIVNKQSC для лечения респираторной вирусной инфекции, вызванной респираторно-синцитиальным вирусом или вирусом гриппа, где R - аргинин; K - лизин; Q - глутамин, L - лейцин, Р - пролин, I - изолейцин, V - валин, N - аспарагин, S - серин, С - цистеин. Катионный разветвленный пептид R4K2KQLLPIVNKQSC блокирует вход вируса в клетку как за счет связывания с клеточным рецептором нуклеолином, так и за счет связывания с лигандами (белком F) на поверхности РСВ. 13 ил., 6 табл., 8 пр.
Применение способа интраназального введения пептида R4K2KQLLPIVNKQSC для лечения респираторной вирусной инфекции, вызванной респираторно-синцитиальным вирусом или вирусом гриппа, где R - аргинин; K - лизин; Q - глутамин, L - лейцин, Р - пролин, I - изолейцин, V - валин, N - аспарагин, S - серин, С - цистеин.
| Shilovskiy I | |||
| P | |||
| at al "Dendrimeric form of the peptide from antigenic zero-site of F protein exhibit antiviral effect against respiratory syncytial virus", Allergy, 2020, Volume 75: Abstracts from the European Academy of Allergy and Clinical Immunology Digital Congress, 06-08 June 2020, Abstract 0471, p.140, https://doi.org/10.1111/all.14506 | |||
| RU |

