Изобретение относится к медицинской вирусологии и может быть использовано при исследовании эффективности лечебных и профилактических препаратов против респираторно-синцитиального вируса (РСВ), а также при изучении биологических характеристик респираторно-синцитиального вируса.
Штамм может быть применен в качестве контрольного референс-штамма РСВ при проведении диагностических работ методом ПЦР и при изучении биологических свойств вновь выделяемых вариантов РСВ. Штамм также может быть использован для оценки эффективности лечебных и профилактических препаратов против РСВ in vitro.
РСВ является одним из основных этиологических агентов, вызывающих острые инфекции нижних дыхательных путей. Особую опасность он представляет для младенцев и детей раннего возраста, и является наиболее частой причиной смерти, вызванной ОРВИ, у детей до года [1]. Считается, что более 95% детей переносят инфицирование респираторно-синцитиальным вирусом к возрасту 2-х лет [2]. На его долю приходится более 120000 случаев госпитализации в год в США, чей ежегодный экономический урон оценивается более чем в 500 миллионов долларов. РСВ способен вызывать тяжелые поражения нижних отделов респираторного тракта с бронхообструкцией и бронхиолитом [3, 4], что способствует в дальнейшем повышению риска развития бронхиальной астмы.
РСВ человека относится к виду Human orthopneumovirus рода Orthopneumovirus, входящего в состав семейства Pneumoviridae. Вирион представляет собой вирусную частицу плеоморфной формы со спиральной симметрией размером порядка 200 нм, состоящую из нуклеокапсида в билипидной оболочке. Геном РСВ состоит из однонитевой, несегментированной «-» РНК. Размер генома составляет 15 т.н., составляющих 10 генов, которые кодируют 11 белков [5], основными из которых являются белок нуклеокапсида N, фосфопротеин Р, большая субъединица полимеразы L, малый гидрофобный белок SH, негликозилированный матричный белок М, неструктурные белки NS1 и NS2, белок слияния F, трансмембранный гликопротеин G. Белок F обеспечивает слияние вирусных частиц с поверхностью клетки-хозяина, а также слияние инфицированных клеток друг с другом, что вызывает появление синцития. Образование синцития является показателем цитопатического эффекта РСВ. [б, 7]. На основании строения гликопротеина G выделяют две генетические группы РСВ - А и В, которые могут циркулировать одновременно во время вспышек заболевания [8, 9]. Считается, что РСВ группы А доминируют по сравнению с РСВ группы В и способны вызвать более тяжелое заболевание [10, 11]. Группа А делится на 16 генотипов, группа В - на 22 генотипа на основании антигенных свойств поверхностного гликопротеина G [12], тогда как гликопротеин F проявляет свойства консервативности среди разных генотипов РСВ [13]. В целом, РСВ группы А, как правило, более распространен по сравнению с РСВ группы В [14, 15], однако периодически сообщается о более высокой распространенности РСВ В относительно РСВ А [16]. Причины превалирования той или иной группы окончательно не выяснены, хотя были предложены модели чередования эпидемий РСВ типа А и типа В. Так, например, формирование иммунитета к РСВ одного типа в течение сезона может сделать популяцию более восприимчивой к другому типу вируса в последующие сезоны [17].
РСВ впервые был выделен и идентифицирован как новый вид вируса в 1956 году от шимпанзе, а в 1957 году РСВ был выделен от детей подросткового возраста с пневмонией и бронхиолитом. С того времени РСВ периодически регистрируется в виде эпидемических подъемов заболеваемости и является наиболее частой причиной инфекции нижних дыхательных путей у детей младшего возраста по всему миру [18], а также повышает уровни заболеваемости и смертности у пациентов, перенесших заболевания верхних дыхательных путей, бронхиолит и пневмонию. Наиболее уязвимой группой населения являются дети до двух лет [19]. Передача РСВ происходит воздушно-капельным путем с последующим распространением инфекции в верхних дыхательных путях и паренхиме легких. Особые опасения вызывает потенциальная способность РСВ из носовой полости по нейронам обонятельных рецепторов проникать в периферическую НС и ткани головного мозга, минуя гематоэнцефалический барьер, и вызывать патологию ЦНС [20].
Несмотря на масштабы заболевания РСВ, лечение остается в основном симптоматическим, хотя РСВ исследуется уже более 40 лет. Испытания бронходилататоров, кортикостероидов не показали убедительного клинического преимущества. Противовирусный препарат рибавирин продемонстрировал лишь незначительную клиническую пользу и обычно не показан при лечении заболевания РСВ [21]. Поливизумаб, представляющий собой препарат гуманизированных моноклональных антител (IgG1к), полученных путем рекомбинантной ДНК-технологии, эффективен в профилактике для детей с высоким риском тяжелой инфекции РСВ, хотя оптимальные показания, основанные на оценке финансовых затрат, не были определены ввиду высокой стоимости препарата [22]. Будущие направления в лечении и профилактике РСВ-инфекций, вероятно, включают в себя моноклональные антитела к РСВ, более мощные противовирусные соединения и более уникальные противовоспалительные агенты. Вакцинация против РСВ находится в стадии разработки, но на данный момент не является широко применяемой [23] и поэтому требует дальнейших исследований биологических свойств штаммов РСВ и их патогенного потенциала.
Одним из наиболее эффективных направлений является создание вакцин на основе аттенуированных штаммов РСВ дикого типа. Однако одной из главных проблем выделения РСВ является сложность пассирования на различных культурах клеток и нестабильность РСВ при выделении и хранении [24, 25, 26, 27].
Анализ известных аналогов
На данный момент наиболее близкими изобретениями, относящимися к РСВ человека, являются запатентованные работы в области биотехнологий по созданию субъединичных вакцинных препаратов против РСВ инфекции, основанные на выделении и технологическом модифицировании отдельных вирусных белков [28, 29].
Однако данные работы представляют лишь технологическую часть по выделению отдельных белков вируса, не характеризуя конкретные штаммы РСВ и способы наработки. Кроме того, необходимы данные о полных последовательностях вирусного генома. На данный момент существует очень мало данных о циркуляции РСВ в России. Нет информации о полногеномных сиквенсах штаммов РСВ, активных на территории России. В базе данных GenBank зарегистрировано только 30 последовательностей поверхностных белков G и F штаммов, циркулирующих в европейской части России и относящихся преимущественно к группе А. На данный момент только 5 штаммов РСВ группы В (Human respiratory syncytial virus B1 NCBI:txid79692, Human respiratory syncytial virus (subgroup В / strain 18537) NCBI:txid11251, Human respiratory syncytial virus (subgroup B/strain 8/60) NCBI:txid11258, Respiratory syncytial virus type В NCBI:txidl615611)) зарегистрированы в базе данных GenBank. Только для одного из них представлена полная последовательность генома {Human Respiratory syncytial virus 9320 AUTHORS Cheng X., Park H, Jin H. GenBank: AY353550.1), но этот штамм не актуален для России, так как не циркулирует на ее территории.
Таким образом, очевидна необходимость создания генетически актуального штамма РСВ, пригодного для использования в заявляемой области.
Задачей настоящего изобретения является получение оригинального штамма-продуцента РСВ группы В, обладающего следующими выгодными свойствами:
- антигенной актуальностью.
- пригоден для изучения и использования в диагностике РСВ группы В методом ПЦР.
- пригоден для исследования лечебной и профилактической эффективности противовирусных препаратов и вакцин in vitro.
- Выделен от человека и способен размножаться на моделях in vitro (культуры клеток НЕр2, HeLa, А549) и достигать титров не менее 3,0 lg TCID50/ml.
Указанная задача решена выделением, детальной характеристикой штамма RSVB/Novosibirsk/66Hl/2018.
Технический результат достигается получением штамма
RSVB/Novosibirsk/66Hl/2018, используемого для приготовления антигенсодержащего препарата для диагностических целей, для применения его в качестве контрольного референс-образца при оценке специфичности тест-систем на основе ПЦР, а также для исследования эффективности противовирусных препаратов in vitro.
Предлагаемый штамм является единственным известным штаммом РСВ группы В, выделенным от человека за последние 20 лет на территории России, для которого определена нуклеотидная последовательность полного генома. Этот факт подчеркивает актуальность его использования для тестирования лекарственных препаратов на модели in vitro. Очевидна пригодность его использования при разработке современной диагностической системы РСВ различных субтипов методом ПЦР.
Раскрытие сущности изобретения Характеристика штамма RSVB/Novosibirsk/66Hl/2018
Заявленный штамм респираторно-синцитиального вируса группы В RSV/Novosibirsk/66Hl/2018 является представителем респираторно-синцитиальных вирусов рода Orthopneumovirus семейства Pneumoviridae. Штамм принят на депонирование под номером V-902 в Государственную коллекцию возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора (630559, р.п. Кольцово, Новосибирская область, Россия).
Происхождение
Штамм RSV/Novosibirsk/66Hl/2018 был выделен из носоглоточного мазка, полученного от ребенка, госпитализированного с симптомами ОРВИ, в марте 2018 г. в культуре клеток HeLa (карцинома шейки матки человека). Индикация вируса в культуральной жидкости оценивалась под микроскопом по наличию специфического цитопатического эффекта, заключающегося в образовании клеточного синцития, а также с помощью полимеразной цепной реакции в режиме реального времени с применением тест-системы «AmpliSens ОРВИ-скрин» («Интерлабсервис», Россия).
Идентификация заявленного штамма РСВ
Идентификация штамма осуществлена методом полимеразной цепной реакции в режиме реального времени с помощью тест-системы «AmpliSens ОРВИ-скрин» («Интерлабсервис», Россия), с дальнейшим подтверждением секвенированием полного генома и BLAST-анализом.
Получение последовательностей
Определена последовательность полного генома исследуемого штамма методом NGS-секвенирования и зарегистрирована в базе данных GenBank под номером МК534512. Нуклеотидные последовательности заявленного штамма приведены в Перечне нуклеотидных и аминокислотных последовательностей штамма респираторно-синцитиального вируса RSV/Novosibirsk/66Hl/2018. По результатам филогенетического исследования штамм RSV/Novosibirsk/66Hl/2018 был отнесен к РСВ генетической линии В.
Культуральные свойства
Штамм RSVB/Novosibirsk/66Hl/2018 при культивировании на монослое перевиваемых культур клеток HeLa, Нер2, А549, Vero вызывает цитопатиеское действие в виде образования клеточного синцития. Условия культивирования: температура 37°С в атмосфере 5% CO2, питательная среда для культур клеток ДМЕМ с 2% фетальной бычьей сыворотки и 100 мкг/мл гентамицина сульфата.
Гемагглютинирующие свойства
Штамм RSVB/Novosibirsk/66Hl/2018 не агглютинирует эритроциты петуха, гуся, морской свинки и человека.
Патогенность штамма
Штамм RSVB/Novosibirsk/66Hl/2018 отностится к 4-й группе патогенности для человека.
При интраназальном инфицировании мышей линии Balb/c штамм не оказывал патогенного действия: отсутсвие репликации вируса в респираторном тракте экспериментальных животных.
Филогенетический анализ
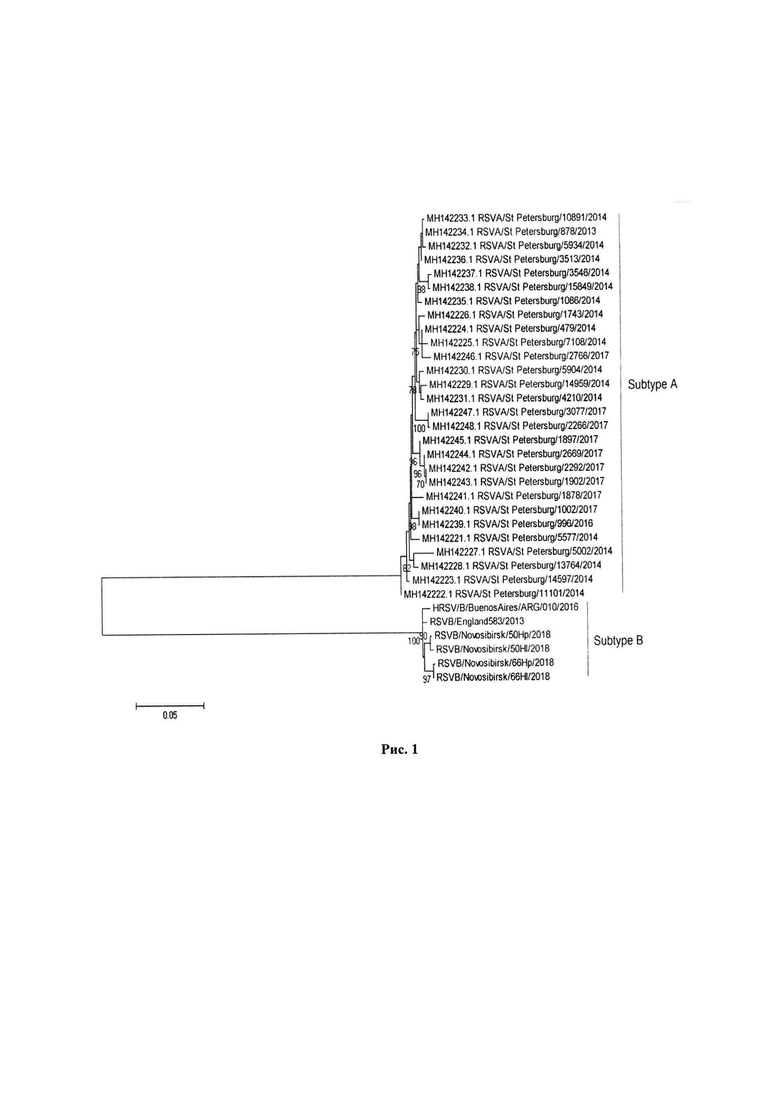
Для штамма RSV/Novosibirsk/66Hl/2018 был выполнен филогенетический анализ на основе полногеномных нуклеотидных последовательностей, а также на основе последовательностей отдельных генов. Множественные выравнивания нуклеотидных последовательностей (рассматриваемого штамма и представленных в базе данных GenBank) были выполнены посредством программного обеспечения MUSCLE. Построение филогенетических денрограмм было выполнено в программе MEGA 5, с использованием метода максимального правдоподобия (Maximum Likelihood) и модели нуклеотидных замен GTR (general time reversible). Для оценки достоверности топологии филогенетических дендрограмм использовалась бутстреп-поддержка (500 репликаций).
В результате анализа последовательностей РСВ, представленных в базе данных GenBank, было обнаружено, что ранее полногеномные последовательности штаммов РСВ, выделенных на территории России, в эту базу данных не депонировались и все российские штаммы (выделены в Санкт-Петербурге в 2014-2016 гг.) представлены сиквенсами двух генов: F и G. Ген, кодирующий G-белок респираторно-синцитиального вируса, является основным маркером принадлежности к той или иной генетической группе и именно на генетических различиях по G-гену базируется классификация респираторно-синцитиальных вирусов. F-белок является поверхностным гликопротеином. Генетические отличия между F-генами вариантов респираторно-синцитиального вируса могут обуславливать различия в их биологических свойствах. Исходя из этого, для филогенетического анализа РСВ, циркулирующих на территории России, из полногеномного сиквенса штамма RSVB/Novosibirsk/66Hp/2018, были выделены последовательности генов F и G и, в совокупности с прочими последовательностями российских штаммов, а также близкородственных (по результатам BLAST-анализа) штаммов, использованы для построения филогенетических дендрограмм (рис. 1-2).
Согласно филогенетическому анализу, доступные в базе данных последовательности G-гена штаммов из Санкт-Петербурга значительно отличаются от последовательностей G-гена штамма RSVB/Novosibirsk/66Hl/2018. Санкт-Петербургские и новосибирские штаммы формируют два отдельных филогенетических кластера, соответствующих субтипам А и В РСВ (рис. 1). При этом штамм RSV/Novosibirsk/66Hl/2018 по нуклеотидной последовательности G-гена филогенетически родственен европейским и южноамериканским вариантам вируса.
Согласно филогенетическому анализу по F-гену (рис. 2), штамм RSV/Novosibirsk/66Hl/2018 и штаммы из Санкт-Петербурга формируют две отдельные филогенетические группы, соответствующие субтипам А и В. Так же, как и в случае с G-геном, F-ген RSV/Novosibirsk/66Hl/2018 штамма РСВ генетически схож с F-генами штаммов из Европы и Южной Америки.
Перечень фигур, чертежей и иных материалов
Рисунок 1. Филогенетическая дендрограмма на основе нуклеотидных последовательностей G-гена штамма RSV/Novosibirsk/66Hl/2018.
Рисунок 2. Филогенетическая дендрограмма на основе нуклеотидных последовательностей F-гена штамма RSV/Novosibirsk/66Hl/2018
Рисунок 3. Образование синцитиоподобных структур при инфицировании культуры клеток HeLa штаммом RSV/Novosibirsk/66Hl/2018. Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), режим DIC-contrast, ув. x4000. A. - Культура клеток HeLa, инфицированная штаммом RSV/Novosibirsk/66Hl/2018, Б. - Интактная культура клеток HeLa.
Рисунок 4. Локализация РСВ в клетках культуры клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, при имуноцитохимическом окрашивании на ИФ-РС (набор «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). Фотография на конфокальном микроскопе LSM 710 (Carl Zeiss), ув. х6300, режим флуоресценции. Ядра клеток окрашены DAPI (синее свечение). В культуре клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, локализация РСВ (интенсивное зеленое свечение в центральной части синцитиоподобных образований). А - Культура клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, Б - Интактная культура клеток HeLa.
Рисунок 5. Образование синцитиоподобных структур при инфицировании культуры клеток Нер2 штаммом RSVB/Novosibirsk/66Hl/2018. Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), режим DIC-contrast, ув. x4000. A. -Культура клеток Hep2, инфицированная штаммом RSVB/Novosibirsk/66Hl/2018. Б. - Интактная культура клеток Нер2.
Рисунок 6 (был 4.2.) Локализация РСВ в клетках культуры клеток Нер2, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, при имуноцитохимическом окрашивании на ИФ-РС (набор «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). А - Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), режим DIC-contrast, ув. x4000. Б - Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), режим флуоресценции, ув. x4000. В культуре клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, локализация РСВ (интенсивное зеленое свечение в центральной части синцитиоподобного образования). В (был 4.2.Д.) - Интактная культура клеток Нер2 при имуноцитохимическом окрашивании на ИФ-РС (набор «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), режим флуоресценции, ув. х4000. В интактной культуре клеток Нер2 отсутствуют синцитиоподобные образования и интенсивное зеленое свечение клеток.
Рисунок 7. Образование синцитиоподобных структур при инфицировании культуры клеток А549 штаммом RSVB/Novosibirsk/66Hl/2018. Окраска азур-эозином. Фотография на микроскопе EVOS V5000 (ThermoFisher Scientific), режим Bright Field, ув. х2000. А - Культура клеток А549, инфицированная RSVB/Novosibirsk/66Hl/2018. Синиотиоподобная структура. Б - Интактная культура клеток А549.
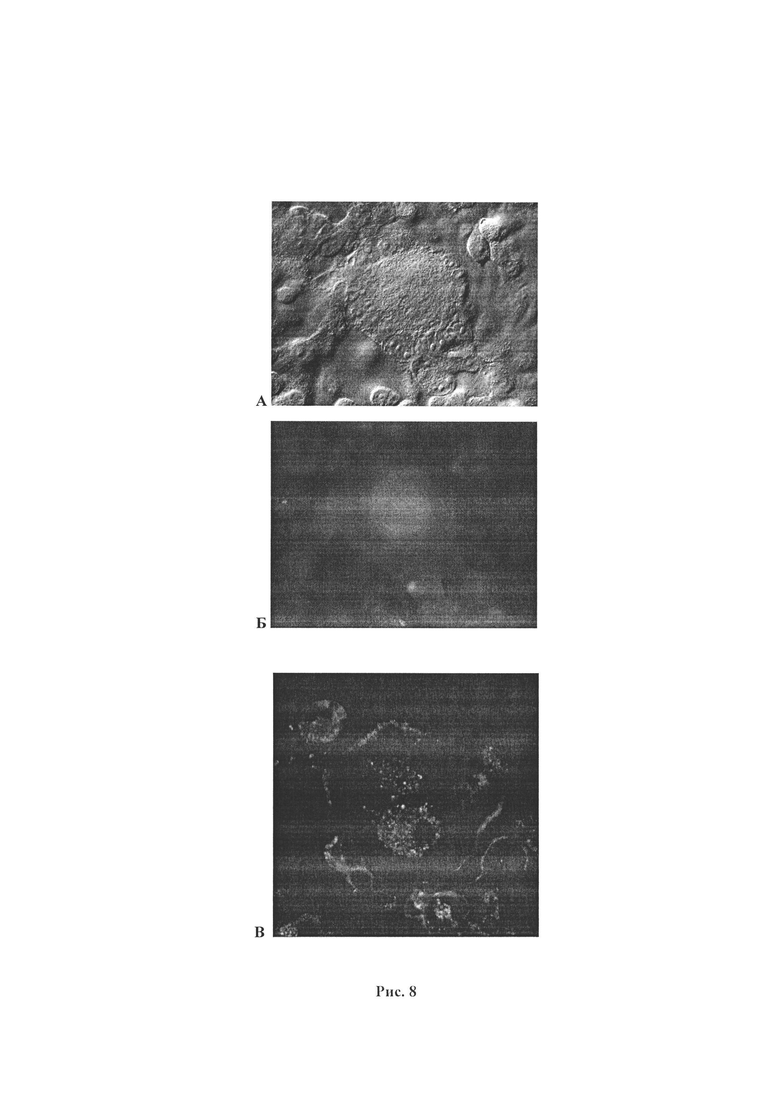
Рисунок 8. Локализация РСВ в клетках культуры клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, при имуноцитохимическом окрашивании на ИФ-РС (набор «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), режим флуоресценции, ув. х4000. А - Культура клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018. Синцитиоподобное образование. Режим DIC-contrast, ув. х4000. Б - Культура клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018. Синцитиоподобное образование. Режим флуоресценции, ув. х4000. В - Культура клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018. Синцитиоподобное образование. Фотография на конфокальном микроскопе LSM 710 (Carl Zeiss), ув. х6300, режим флуоресценции. В культуре клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, локализация РСВ (интенсивное зеленое свечение в центральной части синцитиоподобного образования). Г - Интактная культура клеток А549. Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss), DIC-contrast, ув. x2000. Д - Интактная культура клеток A549. Фотография на микроскопе Axiolmager A1 (Zeiss), камера AxioCam HRc (Zeiss). Режим флуоресценции, ув. х2000.
Рисунок 9. Образование синцитиоподобных структур в лунках контроля вируса на 6-е сутки после инфицирования культуры клеток HeLa. Фотография на микроскопе AxioVert 40 CFL (Zeiss), камера AxioCam HRc (Zeiss), режим проходящего света, ув. х2000.
Рисунок 10. Отсутствие синцитиоподобных структур в культуре клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018 100TCID50 с одновременным внесением раствора коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в концентрации 1/20 на 6-е сутки после инфицирования. Фотография на микроскопе AxioVert 40 CFL (Zeiss), камера AxioCam HRc (Zeiss), режим проходящего света, ув. х2000.
Рисунок 11. Образование синцитиоподобных структур в культуре клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018 100TCID50 с одновременным внесением раствора коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в концентрации 1/80 на 6-е сутки после инфицирования. Фотография на микроскопе AxioVert 40 CFL (Zeiss), камера AxioCam HRc (Zeiss), режим проходящего света, ув. х2000.
Рисунок 12. Интактная культура клеток HeLa (6-е сутки). Фотография на микроскопе AxioVert 40 CFL (Zeiss), камера AxioCam HRc (Zeiss), режим проходящего света, ув. х2000.
Рисунок 13. Культура клеток HeLa, инкубированная с препаратом «Интерферон человеческий лейкоцитарный» в концентрации 1/20 (6-е сутки). Фотография на микроскопе AxioVert 40 CFL (Zeiss), камера AxioCam HRc (Zeiss), режим проходящего света, ув. х2000.
Осуществление изобретения
Пример 1. Получение штамма вируса RSV/Novosibirsk/66Hl/2018 в культуре клеток HeLa
Штамм RSV/Novosibirsk/66Hl/2018 выделен из клинического материала, представляющего собой назофарингеальный мазок в культуре клеток HeLa (карцинома шейки матки человека). Для этого оптимизировали условия культивирования вируса. С этой целью аликвоты образца клинического материала вносили параллельно в планшеты с суточным монослоем и в суспензию клеток линии HeLa. Планшет с монослоем клеток предварительно отмывали от ростовой среды раствором Хенкса; после адсорбции вируса на клетках в течение часа в лунки планшета вносили поддерживающую среду (питательная среда для культур клеток DMEM, содержащая 2% фетальной бычьей сыворотки и антибиотики). Через 24 ч после инфицирования часть лунок в планшетах обрабатывали препаратом RDE (Receptor destroying enzyme), представляющим собой фермент нейраминидазу холерного вибриона, с целью активации образования синцития клеток. Для этого из планшетов удаляли культуральную среду, вносили раствор RDE и оставляли в термостате на 1 ч, после чего препарат удаляли и в лунки планшета вносили поддерживающую среду, как описано выше. В качестве контроля использовали интактные клетки линии HeLa. Ежедневно под микроскопом оценивали цитопатическое действие (ЦПД), проявляющееся в образовании синцития клеток. При наличии ЦПД проводили ПЦР в режиме реального времени вируссодержащей культуральной жидкости на наличие генетического материала РСВ с помощью тест-системы «AmpliSens ОРВИ-скрин» («Интерлабсервис», Россия). Таким образом, было обнаружено, что ЦПД наиболее выражено в лунках планшета при заражении суспензии клеток HeLa с последующей их обработкой RDE.
Морфологический анализ культуры клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, проводили под мокроскопом Axiolmager Zl (Zeiss) в режиме DIC-contrast. В качестве контроля была использована интактная культура клеток HeLa. При морфологическом анализе культуры клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, выло выявлено образование синцитиоподобных структур с расположением ядер по периферии образования (Рис 3.А). В интактной культуре клеток подобных образований не было отмечено (Рис 3.Б).
Для выявления локализации РСВ в клетках проводили имуноцитохимическое окрашивание фиксированной культуры клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, с помощью набора «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). Наличие и локализацию вирусных частиц штамма RSV/Novosibirsk/66Hl/2018 идентифицировали по зеленому свечению в клетках HeLa при анализе инфицированной клеточной культуры на конфокальном микроскопе LSM 710 (Carl Zeiss) в режиме флуоресценции. Ядра клеток окрашивали DAPI и идентифицировали по синему свечению. В культуре клеток HeLa, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, наличие и локализация РСВ были выявлены по интенсивному зеленому свечению в центральной части синцитиоподобных образований. Клеточные ядра в синцитиоподобном образовании, окрашенные DAPI, были расположены по периферии и имели синее свечение (Рис. 4.А). В интактной культуре клеток HeLa специфического зеленого свечения не было (Рис. 4.Б).
Пример 2. Получение штамма вируса RSV7Novosibirsk/66Hl/2018 в культуре клеток Нер2
Штамм RSVB/Novosibirsk/66Hl/2018, выделенный в культуре клеток HeLa, культивировали в культуре клеток Нер2 (эпидермоидная карцинома гортани человека). Для этого оптимизировали условия культивирования вируса. С этой целью аликвоты вируссодержащей культуральной жидкости вносили параллельно в планшеты с суточным монослоем и в суспензию клеток линии Нер2. Планшет с монослоем клеток предварительно отмывали от ростовой среды раствором Хенкса; после адсорбции вируса на клетках в течение часа в лунки планшета вносили поддерживающую среду (питательная среда для культур клеток DMEM, содержащая 2% фетальной бычьей сыворотки и антибиотики). Через 24 ч после инфицирования часть лунок в планшетах обрабатывали препаратом RDE (Receptor destroying enzyme), представляющим собой фермент нейраминидазу холерного вибриона, с целью активации образования синцития клеток. Для этого из планшетов удаляли культуральную среду, вносили раствор RDE и оставляли в термостате на 1 ч, после чего препарат удаляли и в лунки планшета вносили поддерживающую среду, как описано выше. В качестве контроля использовали интактные клетки линии Нер2. Ежедневно под микроскопом оценивали цитопатическое действие (ЦПД), проявляющееся в образовании синцития клеток. При наличии ЦПД проводили ПЦР в режиме реального времени вируссодержащей культуральной жидкости на наличие генетического материала РСВ с помощью тест-системы «AmpliSens ОРВИ-скрин» («Интерлабсервис», Россия). Таким образом, было обнаружено, что ЦПД наиболее выражено в лунках планшета при заражении суспензии клеток Нер2 с последующей их обработкой RDE.
Морфологический анализ культуры клеток Нер2, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, проводили под мокроскопом Axiolmager Z1 (Zeiss) в режиме DIC-contrast. В качестве контроля была использована интактная культура клеток Нер2. При морфологическом анализе культуры клеток Нер2, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, выло выявлено образование синцитиоподобных структур с расположением ядер по периферии образования (Рис 5.А). В интактной культуре клеток подобных образований не было отмечено (Рис 5.Б).
Для выявления локализации РСВ в клетках проводили имуноцитохимическое окрашивание фиксированной 4% раствором формалина культуры клеток Нер2, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, с помощью набора «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). Наличие и локализацию вирусных частиц штамма RSV/Novosibirsk/66Hl/2018 идентифицировали по зеленому свечению в клетках Нер2 при анализе инфицированной клеточной культуры на микроскопе Axiolmager Z1 (Zeiss) в режиме флуоресценции. В культуре клеток Нер2, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, наличие и локализация РСВ были выявлены по интенсивному зеленому свечению в центральной части синцитиоподобных образований (Рис. 6.А). Клеточные ядра в синцитиоподобном образовании, были расположены по периферии (Рис. 6.Б). В интактной культуре клеток Нер2 специфического зеленого свечения не было (Рис. 6.В).
Пример 3. Получение штамма вируса RSV/Novosibirsk/66Hl/2018 в культуре клеток А549
Штамм RSVB/Novosibirsk/66Hl/2018, выделенный в культуре клеток HeLa, культивировали в культуре клеток А549 (карцинома легкого человека). Для этого оптимизировали условия культивирования вируса. С этой целью аликвоты вируссодержащей культуральной жидкости вносили параллельно в планшеты с суточным монослоем и в суспензию клеток линии А549. Планшет с монослоем клеток предварительно отмывали от ростовой среды раствором Хенкса; после адсорбции вируса на клетках в течение часа в лунки планшета вносили поддерживающую среду (питательная среда для культур клеток DMEM, содержащая 2% фетальной бычьей сыворотки и антибиотики). Через 24 ч после инфицирования часть лунок в планшетах обрабатывали препаратом RDE (Receptor destroying enzyme), представляющим собой фермент нейраминидазу холерного вибриона, с целью активации образования синцития клеток. Для этого из планшетов удаляли культуральную среду, вносили раствор RDE и оставляли в термостате на 1 ч, после чего препарат удаляли и в лунки планшета вносили поддерживающую среду, как описано выше. В качестве контроля использовали интактные клетки линии А549. Ежедневно под микроскопом оценивали цитопатическое действие (ЦПД), проявляющееся в образовании синцития клеток. При наличии ЦПД проводили ПЦР в режиме реального времени вируссодержащей культуральной жидкости на наличие генетического материала РСВ с помощью тест-системы «AmpliSens ОРВИ-скрин» («Интерлабсервис», Россия). Таким образом, было обнаружено, что ЦПД наиболее выражено в лунках планшета при заражении суспензии клеток А549 с последующей их обработкой RDE.
Для морфологического анализа инфицированной культуры клеток А549 клеточные препараты фиксировали 4% раствором формалина и окрашивали азур-эозином. Морфологический анализ культуры клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, проводили под микроскопом EVOS V5000 (ThermoFisher Scientific), режим Bright Field, ув. х2000. В качестве контроля была использована интактная культура клеток А549. При морфологическом анализе культуры клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, было выявлено образование синцитиоподобных структур с расположением ядер по периферии образования (Рис 7.А). В интактной культуре клеток подобных образований не было отмечено (Рис 7.Б).
Для выявления локализации РСВ в клетках проводили имуноцитохимическое окрашивание фиксированной 4% раствором формалина культуры клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, с помощью набора «Иммуноглобулины диагностические флуоресцирующие респираторно-синцитиальные вирусные сухие» (С.-Петербург, ООО «Предприятие по производству диагностических препаратов»). Наличие и локализацию вирусных частиц штамма RSV/Novosibirsk/66Hl/2018 идентифицировали по зеленому свечению в клетках А549 при анализе инфицированной клеточной культуры на микроскопе Axiolmager Zl (Zeiss) в режиме флуоресценции. В культуре клеток А549, инфицированной штаммом RSVB/Novosibirsk/66Hl/2018, наличие и локализация РСВ были выявлены по интенсивному зеленому свечению в центральной части синцитиоподобных образований (Рис. 8.Б, 8.В.). Клеточные ядра в синцитиоподобном образовании, были расположены по периферии (Рис. 8.А). В интактной культуре клеток А549 специфического зеленого свечения не было (Рис. 8.Г, 8.Д).
Пример 4. Подтверждение специфичности праймеров, предназначенных для диагностики респираторно-синтициального вируса (РСВ) типа В посредством выявления наличия гена белка N этого патогена методом ПЦР с детекцией результатов в реальном времени.
Для типирования изолятов методом ПЦР в реальном времени амплификацию проводили с использованием набора реагентов БиоМастер HS-qPCR (ООО «Биолабмикс», РФ). Набор БиоМастер HS-qPCR (2×) содержит 2× реакционную смесь БиоМастер HS-qPCR (2×) и стерильную воду. Реакционная смесь БиоМастер HS-qPCR (2×) предназначена для проведения ПЦР в режиме реального времени с использованием флуоресцентно-меченых зондов. В состав БиоМастер HS-qPCR (2×) входят все необходимые компоненты ПЦР (исключая ДНК-матрицу, праймеры и зонд): HS-Taq ДНК-полимераза, смесь dNTP, 2× ПЦР-буфер, Mg2+. При проведении ПЦР были использованы праймеры RSVB-F 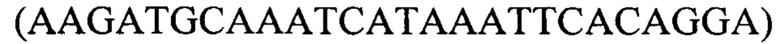 и RSVB-R
и RSVB-R  а также флуоресцентный TaqMan-зонд RSVB-Probe
а также флуоресцентный TaqMan-зонд RSVB-Probe 
 Такая комбинация праймеров и зонда позволяет специфично выявлять РСВ генотипа В, что и было доказано в ходе дальнейшей оптимизации условий реакции и апробации метода с использованием различных контрольных образцов.
Такая комбинация праймеров и зонда позволяет специфично выявлять РСВ генотипа В, что и было доказано в ходе дальнейшей оптимизации условий реакции и апробации метода с использованием различных контрольных образцов.
Состав реакционной смеси:
1) БиоМастер HS-qPCR - 12,5 мкл
2) кДНК - 2 мкл
3) Прямой праймер (RSVB-F) - 1 мкл (150 нМ)
4) Обратный праймер (RSVB-R) - 1 мкл (150 нМ)
5) Флуоресцентный зонд (RSVB-Probe) - 1 мкл (40 нМ)
6) Вода Nuclease-free - 3,5 мкл
Температурный профиль реакции:
1) Т=95С, 5 мин
45 циклов:
1) Т=95С, 15 сек
2) Т=56С, 15 сек
3) Т=68С, 30 сек (детекция флуоресценции)
Результат контрольной реакции ПЦР с использованием специфических праймеров и зонда. В качестве положительного контроля на испытаниях праймеров используется предлагаемый штамм RSVB/Novosibirsk/66Hl/2018. В качестве отрицательных контрольных образцов были использованы:
- транспортная среда, используемая для сбора и хранения мазков из зева и носа
- стерильный физиологический раствор
- генетический материал вируса гриппа типа А
- генетический материал вируса гриппа типа В
- генетический материал метапневмовируса
- генетический материал вирусов парагриппа 1, 2, 3 и 4 типов
- генетический материал коронавирусов
- генетический материал риновирусов
- генетический материал аденовирусов групп В, С и Е
- генетический материал бокавируса
В результате, посредством полимеразной цепной реакции (ПЦР) с гибридизационно-флуоресцентной детекцией было показано, что используемые праймеры и зонд позволяют специфично выявлять именно респираторно-синцитиальный вирус генотипа В.
Таким образом, штамм RSV/Novosibirsk/66Hl/2018 был использован как контрольный при постановке реакции ПЦР с использованием праймеров и зонда для идентификации РСВ генотипа В в режиме реального времени посредством гибридизационно-флуоресцентной детекции. Положительный результат получен исключительно со штаммом RSVB/Novosibirsk/66Hl/2018. Праймеры и TaqMan-зонд не проявили специфичности к отрицательным контрольным образцам.
Пример 5. Исследование противовирусной активности лекарственного препарата «Интерферон человеческий лейкоцитарный» в отношении РСВ штамма RSVB/Novosibirsk/66Hl/2018in vitro.
Коммерческий лекарственный препарат «Интерферон человеческий лейкоцитарный» представляет собой группу белков (интерфероны альфа), синтезированных лейкоцитами здоровых доноров, в ответ на воздействие вируса-индуктора интерферона (вирус болезни Ньюкасла, вирус Сендай) и оказывает непрямой противовирусный эффект, индуцируя в клетках состояние резистентности к вирусным инфекциям [30, 31].
Исследования противовирусного эффекта коммерческий лекарственный препарат «Интерферон человеческий лейкоцитарный» проводили на культуре клеток HeLa (карцинома шейки матки человека). В планшет в суспензию клеток HeLa вносили 100TCID50 штамма RSV/Novosibirsk/66Hl/2018 с одновременным внесением раствора коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» (ФГУП «НПО «Микроген» Минздрава России) в концентрации 1/20, 1/80. В качестве контроля использовали интактную культуру клеток HeLa. В качестве контроля цитотоксичности препарата использовали культуру клеток HeLa с добавлением коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в концентрации 1/20, 1/80. В качестве контроля вируса использовали культуру клеток HeLa, инфицированную в суспензию штаммом RSV/Novosibirsk/66Hl/2018 в дозе 100TCID50. Ежедневно под микроскопом оценивали цитопатическое действие (ЦПД), проявляющееся в образовании синцития клеток.
В результате ежедневного визуального анализа было выявлено ЦПД, проявляющееся в образовании синцития, в лунках контроля вируса на 6-е сутки после инфицирования культуры клеток (Рис 9). В культуре клеток, инфицированной штаммом RSV/Novosibirsk/66Hl/2018 100TCID50 с одновременным внесением раствора коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в концентрации 1/20, ЦПД не было выявлено (Рис 10.). ЦПД было выявлено в культуре клеток, инфицированной штаммом RSV/Novosibirsk/66Hl/2018 100TCID50 с одновременным внесением раствора коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в концентрации 1/80 (Рис 11.). Визуальное сравнение жизнеспособности культуры клеток в интактной культуре HeLa (Рис 12.) и в культуре клеток HeLa, инкубированной с препаратом «Интерферон человеческий лейкоцитарный» в концентрации 1/20 (Рис 13.), показало отсутствие цитотоксического действия коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в данной концентрации.
Таким образом, в результате проведенного исследования был показан противовирусный эффект коммерческого лекарственного препарата «Интерферон человеческий лейкоцитарный» в концентрации 1/20, тогда как при уменьшении концетрации препарата до 1/80 противовирусного действия не было.
Данное исследование показывает пригодность штамма RSV/Novosibirsk/66Hl/2018 в исследовании противовирусной активности коммерческих лекарственных препаратов.
Список использованных источников
1. Griffiths С, Drews S.J., Marchant D.J. Respiratory Syncytial Virus: Infection, Detection, and New Options for Prevention and Treatment // Clinical Microbiology Reviews January 2017 V. 30 Issue 1 P. 277-319.
2. Hall C.B., Weinberg G.A., Iwane M.K., Blumkin A.K., Edwards K.M., Staat M.A., Auinger P., Griffin M.R., Poehling K.A., Erdman D., Grijalva C.G., Zhu Y., Szilagyi P. The burden of respiratory syncytial virus infection in young children. // N. Engl. J. Med. 2009. 360 P. 588-598.
3. Monto A.S. Epidemiology of respiratory viruses in persons with and without asthma and COPD. // Am J Respir Crit Care Med. 1995. V. 151. P. 1653-1657.
4. Hegele R.G., Hayashi S., Hogg J.C., Pare P.D. Mechanisms of airway narrowing and hyperresponsiveness in viral respiratory tract infections. 1995. // Am J Respir Crit Care Med. V. 151.P. 1659-1664.
5. Collins P.L., Mcintosh K., Channock R.M. (2001) Respiratory syncytial virus. In: Fields, B.N., Knipe, D.M., Hawley, P. M. (eds) Virology. 4th edn, Lippincott-Raven, Philadelphia, PA, pp 1313
6. Hall, С.B. (2003) Respiratory syncytial virus. In: Feigin, R.D, Cherry, J.D. (eds) Textbook of paediatric infectious diseases. W.B. Saunders, Philadelphia, PA, P. 2084-2111.
7. Chidgey S.M., Broadley K.J. Respiratory syncytial virus infections: characteristics and treatment, J. Pharmacy and Pharmacology. 2005. V. 57. P. 1371-1381.
8. Hall C.B., Walsh E.E., Long C.E. Immunity to and frequency of infection and reinfection with RSV. J. Infect. Dis. 1991. V. 163. P. 693-698
9. Anderson L.J., Hierholzer J.C., Tsou C., Hendry R.M., Fernie B.F., Stone Y., Mcintosh K. Antigenic characterization of respiratory syncytial virus strains with monoclonal antibodies. J. Infect. Dis. 1985. V. 151(4). P. 626-633.
10. McConnochie K.M., Hall C., Walsh E.E., Roghmann K.J. Variation in severity of respiratory syncytial virus infections with subtype. J. Paediatr. 1990. V. 117. P. 52-62.
11. Walsh E.E., McConnochie K.M., Long C.E. et al. Severity of respiratory syncytial virus infection is related to virus strain. J Infect Dis. 1997. V. 175. P. 814-820.
12. Levine S., Klaiber-Franco R., Paradiso P. R. Demonstration that glycoprotein G is the attachment protein of respiratory syncytial virus. J. Gen. Virol. 1987. V. 68. P. 2521-2526.
13. Johnson P.R., Collins P.L. The fusion of glycoproteins of human respiratory syncytial virus of subgroup A and B: sequence conservation provides a structural basis for antigenic relatedness. J. Gen. Virol. 1988. V. 69. P. 2623-2628.
14. Jafri H.S., Wu X., Makari D., Henrickson K.J. Distribution of respiratory syncytial virus subtypes A and В among infants presenting to the emergency department with lower respiratory tract infection or apnea. Pediatr. Infect. Dis. J. 2013. V. 32. P. 335-340.
15. Esposito S., Piralla A., Zampiero A., Bianchini S., Di Pietro G., Scala A., Pinzani R., Fossali E., Baldanti F., Principi N. Characteristics and their clinical relevance of respiratory syncytial virus types and genotypes circulating in northern Italy in five consecutive winter seasons. PLoS One. 2015. V. 10 (6):e0129369.
16. Peret T.C., Hall C.B., Schnabel K.C., Golub J.A., Anderson L.J. Circulation patterns of genetically distinct group A and В strains of human respiratory syncytial virus in a community. J Gen Virol. 1998. V.79. P. 2221-2229.
17. White L.J., Waris M., Cane P.A., Nokes D.J., Medley G.F. The transmission dynamics of groups A and В human respiratory syncytial virus (hRSV) in England & Wales and Finland: seasonality and crossprotection. Epidemiol Infect. 2005.1 V. 33. P. 279 -289.
18. Mcintosh K. Respiratory syncytial virus. In: Evans, A., Kasiaw, R. (eds) Viral infections in humans-epidemiology and control. 4th edn, Plenum, New York. 1997. P. 691-711.
19. Chidgey S.M., Broadley K.J. Respiratory syncytial virus infections: characteristics and treatment J. Pharmacy and Pharmacology. 2005. V. 57. P. 1371-1381.
20. Bryche В., M., Albin A., D.M., L., Langevin C, D., et al. Respiratory syncytial virus tropism for olfactory sensory neurons in mice J. Neurochem. 2019. V. 7:el4936.
21. Hall C.B., Walsh E.E., Hruska J.F. et al. Ribavirin treatment of experimental respiratory syncytial viral infection. A controlled double-blind study in young adults. JAMA. 1983. V. 249. P. 2666-2670.
22. Hashmi N.A., Cosgrove J.F., MacMahon P. Prophylaxis in RSV infection (Palivizumab)-is it worthwhile?. Ir Med J. 2000. V.93(9). P. 284.
23. Nokes J.D., Cane P.A. New strategies for control of respiratory syncytial virus infection. Current Opinion in Infectious Diseases. 2008. V. 21. P. 639-643.
24. Karron R.A., Buchholz U.J., Collins P.L. Live-Attenuated Respiratory Syncytial Virus Vaccines. Curr. Top.Microbiol. Immunol. 2013. V. 372. P. 259-284.
25. Wright P.F., Ikizler M.R., Gonzales R.A., Carroll K.N., Johnson J.E., Werkhaven J.A. Growth of respiratory syncytial virus in primary epithelial cells from the human respiratory tract. 2005. J Virol. V. 79. P. 8651- 8654.
26. Smith M.C., Creutz C., Huang Y.T. Detection of respiratory syncytial virus in nasopharyngeal secretions by shell vial technique. J Clin Microbiol. 1991. V. 29. P. 463- 465.
27. Griffiths C., Drews S.J., Marchant D.J. Respiratory syncytial virus: infection, detection, and new options for prevention and treatment. Clin Microbiol Rev. 2017. V.30. P. 277-319.
28. Субъединичный вакцинный препарат респираторно-синцитиального вируса КЕЙТС Джордж А. (СА), САНУЭЗА Соня Е. (СА), ООМЕН Раймонд П. (СА), КЛАЙН Мишель X. (СА) публикация патента: 27.12.2004.
29. F-белок респираторно-синцитиального вируса и его применение ГРЮНВАЛЬД Томас (DE), ЮБЕРЛА Клаус (DE) Певион Биотек АГ (СН), Рур-Университет Бохум (DE) публикация патента: 20.10.2012.
30. Интернет-ресурс: https://www.rlsnet.ru/tn_index_id_17916.htm.
31. Крамарев С.А., Евтушенко В.В. Опыт применения назальных форм интерферона в лечении и профилактике острых респираторных инфекций Актуальная инфектология. 2019. Т. 7(4). С.217-223.
Перечень нуклеотидных и аминокислотных последовательностей штамма
респираторно-синцитиального вируса RSV/Novosibirsk/66Hl/2018
(GenBank: МK534512.1)
110 - Федеральное государственное бюджетное научное учреждение
«Федеральный исследовательский центр фундаментальной и
трансляционной медицины»
120 - Штамм респираторно-синцитиального вируса
RSV/Novosibirsk/66Hl/2018 для использования в диагностике
респираторно-синцитиальной вирусной инфекции и исследования
эффективности противовирусных препаратов in vitro
160-1
210-SEO ID NO: 1
211 - 15154
212 - кРНК;
213 - Orthopneumovirus RSV/Novosibirsk/66Hl/2018 400-1









| название | год | авторы | номер документа |
|---|---|---|---|
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К РАЗЛИЧНЫМ ШТАММАМ РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА | 2018 |
|
RU2713340C1 |
| СПОСОБ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ БРОНХИАЛЬНОЙ АСТМЫ, ОСЛОЖНЯЮЩИХ ЕЕ РЕСПИРАТОРНЫХ ВИРУСНЫХ ИНФЕКЦИЙ И ДРУГИХ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ ДЫХАТЕЛЬНЫХ ПУТЕЙ | 2012 |
|
RU2526146C2 |
| F-БЕЛОК РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2464316C2 |
| Набор синтетических олигонуклеотидных праймеров и зондов для выявления вируса респираторно-синцитиальной инфекции крупного рогатого скота и гена GAPDH крупного рогатого скота и способ выявления РНК вируса респираторно-синцитиальной инфекции крупного рогатого скота | 2020 |
|
RU2768753C2 |
| Набор олигонуклеотидов для полногеномной амплификации респираторно-синцитиальных вирусов типов А и В | 2023 |
|
RU2811576C1 |
| СИНТЕТИЧЕСКИЕ ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ И СПОСОБ ВЫЯВЛЕНИЯ РНК ВИРУСА РЕСПИРАТОРНО-СИНЦИТИАЛЬНОЙ ИНФЕКЦИИ КРУПНОГО РОГАТОГО СКОТА С ПОМОЩЬЮ СИНТЕТИЧЕСКИХ ОЛИГОНУКЛЕОТИДНЫХ ПРАЙМЕРОВ В ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ (ПЦР) | 2009 |
|
RU2405039C1 |
| Катионный разветвленный пептид для ингибирования респираторных вирусов | 2022 |
|
RU2810986C2 |
| СУБЪЕДИНИЧНЫЙ ВАКЦИННЫЙ ПРЕПАРАТ РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА | 1997 |
|
RU2243234C2 |
| МЕТОД ПЕРВИЧНОЙ ИЗОЛЯЦИИ ШТАММОВ ВИРУСА РЕСПИРАТОРНО-СИНЦИТИАЛЬНОЙ ИНФЕКЦИИ КРУПНОГО РОГАТОГО СКОТА, ШТАММ ВИРУСА BOVINE RESPIRATORY SYNCITIAL VIRUS ДЛЯ ИЗГОТОВЛЕНИЯ ДИАГНОСТИЧЕСКИХ ПРЕПАРАТОВ | 2010 |
|
RU2451073C2 |
| Производные 7-гидроксикумарина, содержащие остатки монотерпеноидов, как ингибиторы репродукции респираторно-синцитиального вируса (РСВ) | 2022 |
|
RU2787352C1 |










Изобретение относится к медицинской вирусологии. Представлен штамм респираторно-синцитиального вируса RSV/Novosibirsk/66Hl/2018, депонированный в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» под номером V-902, для использования в диагностике респираторно-синцитиальной вирусной инфекции и исследования эффективности противовирусных препаратов in vitro. Изобретение может быть использовано при исследовании эффективности лечебных и профилактических препаратов против респираторно-синцитиального вируса (РСВ), а также при изучении биологических характеристик респираторно-синцитиального вируса. 13 ил., 5 пр.
Штамм респираторно-синцитиального вируса RSV/Novosibirsk/66Hl/2018, депонированный в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» под номером V-902, для использования в диагностике респираторно-синцитиальной вирусной инфекции и исследования эффективности противовирусных препаратов in vitro.
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СПОСОБ СТИМУЛИРОВАНИЯ ИММУННОЙ СИСТЕМЫ | 1993 |
|
RU2139729C1 |
| WO 2002009749 A2, 07.02.2002 | |||
| WO 1998002457 A1, 22.01.1998. | |||

