Изобретение относится к области экспериментальной медицины и биотехнологии.
Пороки развития - это стойкие морфологические изменения в органе, системе или организме, выходящие за рамки вариаций его строения и возникающие в период эмбриогенеза. Результатом развившегося нарушения являются последующие аномалии формирования органа. Врожденные ампутации или редукционные пороки развития - это группа гетерогенных аномалий развития, которые проявляются частичным или полным отсутствием как проксимальных, так и дистальных отделов верхних и нижних конечностей.
Общая распространенность редукционных пороков составляет приблизительно 1 случай на 20000 новорожденных [Tonkin М.А., Oberg K.С. The ОМТ Classification of congenital anomalies of the hand and upper limb. Hand Surg. 2015; 20(3):336-342. DOI: 10.1142/S0218810415400055].
Изучение патогенетических механизмов развития абрахии затруднено вследствие отсутствия экспериментальной модели этого заболевания. Ученые исследователи и клинические ортопеды вынуждены анализировать патофизиологические процессы уже сформированного редукционного порока, тем самым, не имея возможности прогнозирования и коррекции развивающегося патологического процесса. Отсутствие возможности анализировать течение процесса в ходе самого заболевания значительно сужает рамки изучения абрахии.
В доступной зарубежной и отечественной литературе отсутствуют данные об экспериментальных моделях исследуемого редукционного порока - абрахии.
Известен способ моделирования сколиоза на курином эмбрионе (Патент №2728927 «Способ моделирования сколиоза на курином эмбрионе», регистрация от 03 августа 2020, Зайдман A.M., Пахомова Н.Ю., Строкова Е.Л., Черноловская Е.Л., Мелешко Е.М., Шило А.Р.). В Новосибирском НИИТО в процессе исследования 50 случаев идиопатического сколиоза A.M. Зайдман впервые в мире выявлено наличие клеток нервного гребня на вогнутой стороне деформации позвоночника при идиопатическом сколиозе (A.M. Zaydman, E.L. Strokova, E.V. Kiseleva, L.A. Suldina, A.A. Strunov, A.I. Shevchenko, P.P. Laktionov, V.M. Subbotin. A New Look at Etiological Factors of Idiopathic Scoliosis: Neural Crest Cells. International Journal of Medical Sciences. 2018; 15(5):436-446. doi:10.7150/ijms.22894.) В результате была высказана гипотеза: причиной идиопатического сколиоза является депонирование клеток нервного гребня в формирующемся позвоночнике эмбриона. Методическое выполнение способа проводится на курином эмбрионе с целью стимулирования развития деформации позвоночника путем введения в нервную трубку куриного эмбриона на 11 стадии развития по классификации Гамбургер и Гамильтон siРНК к гену РАХ3;
Наиболее близким к предлагаемому является способ индуцирования врожденного вывиха бедра на курином эмбрионе (Патент №2792009 «Способ индуцирования врожденного вывиха бедра на курином эмбрионе», регистрация от 15 марта 2023, Пахомова Н.Ю., Строкова Е.Л., Корыткин А.А., Черноловская Е.Л., Гутт А.А., Мелешко Е.М., Шило О.В., Шило А.Р., Зайдман A.M.). Введение липофильной siPHK для ингибирования миграции клеток нервного гребня осуществлялось в период формирования почек нижних конечностей - период гаструляции, уровень 26-32 пар сомитов.
Задача (технический результат) предлагаемого изобретения заключается в создании модели индуцирования абрахии (отсутствие двух верхних конечностей) на экспериментальном животном (куриный эмбрион).
Поставленная задача решается тем, что в способе моделирования индуцированной абрахии на курином эмбрионе осуществляют контроль развития эмбриона и введение в нервную трубку куриного эмбриона липофильной siРНК к гену Рах3 в объеме, обеспечивающем заполнение нервной трубки. Согласно предлагаемому изобретению введение липофильной siРНК осуществляют в период формирования 16 пары сомитов.
Этиология и патогенез развития редукционных пороков верхних конечностей (абрахия) остаются неизвестными по настоящее время. Существует множество гипотез возникновения данной патологии. Многие исследователи считают, что пороки развития возникают в результате генетических, хромосомных и геномных мутаций, а также комбинированных воздействий генетических мутаций и факторов окружающей среды, внешних по отношению к эмбриону (Kantaputra P.N., Carlson В.М. Genetic regulatory pathways of splithand/foot malformation. Clin Genet. 2019; 95(1):132e139. DOI:10.1111/cge.13434; Sowinska-Seidler A., Socha M., Jamsheer A. Split-hand/foot malformation-molecular cause and implications in genetic counseling. J Appl Genet. 2014; 55(1):105el 15. DOI: 10.1007/s13353-013-0178-5). Большинство предлагаемых теорий формирования редукционных пороков, в частности абрахия, основываются на том, что патогенетические механизмы начинают определяться еще в период раннего эмбриогенеза - в период закладки эмбриональных слоев (Naruse Т., Takahara М., Takagi М., Oberg K.С., Ogino Т. Busulfaninduced central Polydactyly, syndactyly and cleft hand or foot: a common mechanism of disruption leads to divergent phenotypes. Dev Growth Differ. 2007; 49(6):533e541. DOI: 10.1111/j.l440-169X.2007.00949.x; Hall J.G., Kimber E., van Bosse H.J.P. Genetics and classifications. J Pediatr Orthop. 2017; 37(suppl 1):S4eS8. DOI: 10.1097/BPO.0000000000000997).
Почки верхних конечностей формируются на уровне 15-20 пар сомитов. По мере образования боковых складок тела конечности располагаются вдоль боковых стенок тела. Почки конечностей состоят из оболочки, которая формируется эктодермой, заключающей в себе ядро мезодермы. Эктодерма происходит из эктодермы боковой стенки тела, а мезодерма формируется из соматической латеральной пластинки, хотя впоследствии она дополняется клетками, мигрирующими из сомитов. Формирование конечностей начинается в морфогенетическом поле конечностей. Развитие конечностей происходит в результате серии взаимных тканевых взаимодействий между мезенхимой мезодермы боковой пластинки и вышележащими эпителиальными клетками эктодермы. Клетки из мезодермы боковой пластинки и миотома мигрируют в область конечности и делятся до такой степени, что они вызывают выпячивание эктодермы над ней, образуя зародыш конечности. Мезодермальные клетки боковой пластинки секретируют факторы роста фибробластов, чтобы побудить вышележащую эктодерму сформировать на конце зачатка конечности апикальный эктодермальный гребень, который направляет дальнейшее развитие конечности.
В этот период времени эмбрион представляет собой совокупность многочисленных зависимых друг от друга разносторонних процессов, которые происходят не последовательно, а параллельно, именно параллельно, но в конечном итоге все направлено только на достижение одной цели - формирование нового организма.
В последние годы интенсивно изучаются процесс миграции клеток нервного гребня и их роль. Клетки нервного гребня - это мультипотентная эмбриональная клеточная популяция, которая формируется на границе нервной пластинки и впоследствии мигрирует по всему эмбриону. Изменения в процессе миграции клеток нервного гребня могут инициировать структурные нарушения нормального эмбриогенеза, тем самым способствуя развитию морфологических инверсий в структурах развивающегося организма. Миграция клеток нервного гребня происходит волнообразно от переднего к задней части тела. Важным аспектом является то, что миграция следует непосредственно за волной сегментации сомитов, каждая пара сомитов влияет на выброс клеток нервного гребня в своей окрестности (Sela-Donenfeld, D., Kalcheim, С., 2000. Inhibition of noggin expression in the dorsal neural tube by somitogenesis: a mechanism for coordinating the timing of neural crest emigration. Development. 2000; 127(22):4845-54. doi: 10.1242/dev. 127.22.4845). Клетки нервного гребня нервной трубки эмбриона на определенных осевых уровнях мигрируют по нескольким путям, чтобы достичь целевых тканей и генерируют соответствующий репертуар производных нервного гребня. Миграционный процесс клеток нервного гребня сложный поликонтролируемый процесс, в котором принимают участие взаимопересекающиеся сигнальные системы самих клеток нервного гребня, экстрацеллюлярного матрикса, по которому осуществляется процесс миграции. Клетки нервного гребня проходят через переднюю половину каждого сомита, но никогда не проходят через заднюю половину. Контроль за миграцией клеток нервного гребня, зависит от экспрессии эмбриональных генов группы PAX (PAX1, РАХ3, РАХ9) и др. Но нарушение миграции по туловищному пути (миграция через склеротом) может быть связано с нарушением экспрессии РАХ3 гена.
Основываясь на эмбриональных закономерностях формирования почек верхних конечностей эмбриона и гипотезе A.M. Зайдман о депонировании клеток нервного гребня, ингибирование миграции было произведено при прохождении клеток нервного гребня через формирующийся апикальный эктодермальный гребень верхних конечностей. Изменения эпигенетических механизмов регуляции миграции клеток нервного гребня (ингибирование экспрессии РАХ3) в период формирования апикального эктодермального гребня почек верхних конечностей может рассматриваться как патогенетический механизм формирования абрахии. Изменения миграции клеток нервного гребня - ингибирование экспрессии РАХ3-создают условия, в результате которых клетки нервного гребня оказываются не в конечном месте и времени их эволюционного финиша. Моделируемая фенотипическая структура клеток нервного гребня и процесс формирования апикального эктодермального гребня создают предпосылки для «участия» клеток нервного гребня в морфологической структуре почек верхних конечностей, тем самым реализовывают условия для возможного нарушения формирования верхних конечностей.
Экспериментально установлено, что на этапе формирования 16 пары сомитов ингибирование гена РАХ3 приводит к нарушению процесса миграции клеток нервного гребня с последующим индуцированием абрахии.
Принципиальная новизна настоящего изобретения заключается в создании модели индуцирования абрахии (отсутствие двух верхних конечностей) и периоде введения (формирование апикального эктодермального гребня почек верхних конечностей) с целью ингибирования экспрессии РАХ3 гена липофильными интерферирующими siPHK в склеротоме куриного эмбриона.
Исследование проводилось в эмбриональном периоде развития цыпленка как экспериментального биологического объекта. Стадии приводятся в соответствии классификацией по Гамбургер и Гамильтон.
siPHK представляют собой 21 звенные дуплексы с 2 выступающими нуклеотидами на 3'-концах (19 в дуплексе, 2 выступают). Выбор последовательности siPHK, направленной на мРНК гена РАХ3 проводили следующим образом. Последовательность мРНК гена РАХЗ курицы брали из банка нуклеотидных последовательностей GenBank. Последовательности siРНК выбирали исходя из температуры плавления, наличия термодинамической ассимметрии дуплекса, комплементарности антисмысловой цепи мРНК гена-мишени с помощью программ для выбора последовательности siPHK BLOCK-iT ™ RNAi Designer (Invitrogen) или siDESIGN Center (Darmacon). Для увеличения устойчивости siPHK к действию рибонуклеаз в CpG, UpA, СрА мотивы последовательности вводили 2'-O-метильные модификации. siРНК были получены синтетически с использованием твердофазного автоматического синтезатора, их биологическая активность была определена по способности подавлять экспрессию гена-мишени в культуре фибробластов кур. Последовательности наиболее активной siPHK были использованы для конструирования холестеринсодержащих конъюгатов, конъюгат анти- РАХ3 siРНК.
siРНК проникает в цитоплазму клетки и связывается с клеточными белками, образуя комплекс RISC (RNA induced silencing complex), после активации этого комплекса, он связывается с комплементарной последовательностью в составе мРНК и находящийся в составе комплекса белок Ago2, обладающий рибонуклеазной активностью, расщепляет мРНК-мишень. После этого комплекс диссоциирует и может взаимодействовать с другой молекулой мРНК, а расщепленная мРНК деградирует. В результате этого снижается концентрация в клетке мРНК-мишени и перестает синтезироваться белок РАХ3, который кодируется мРНК-мишенью.
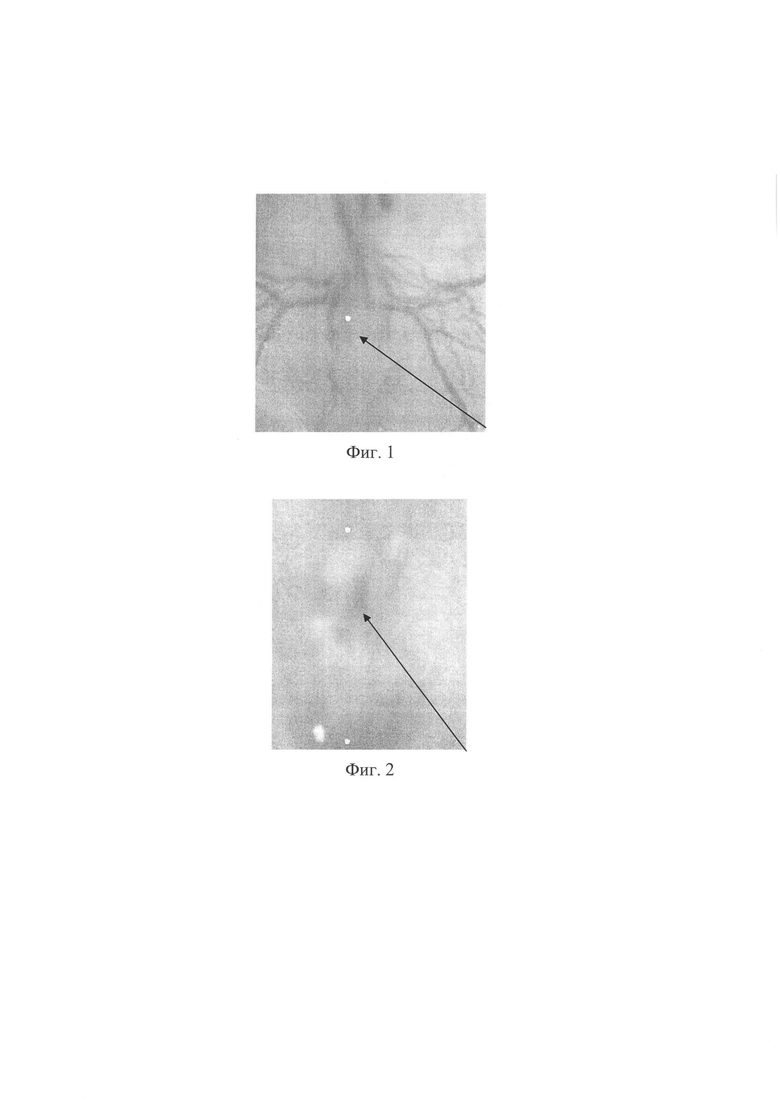
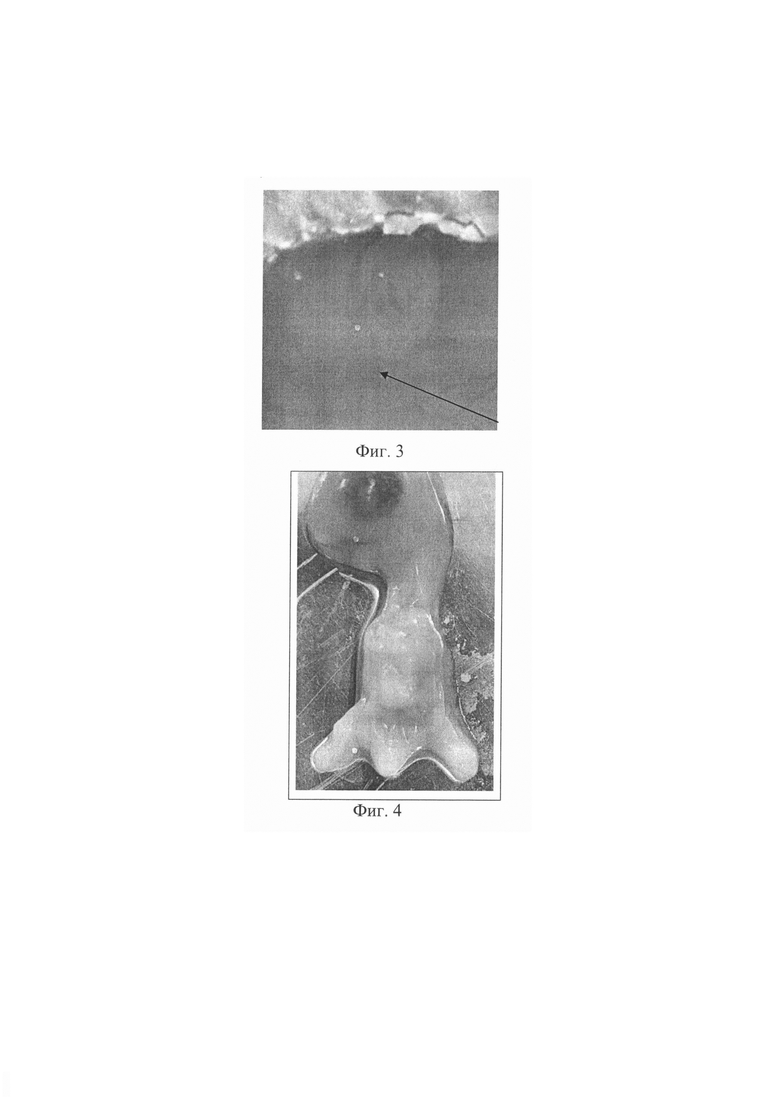
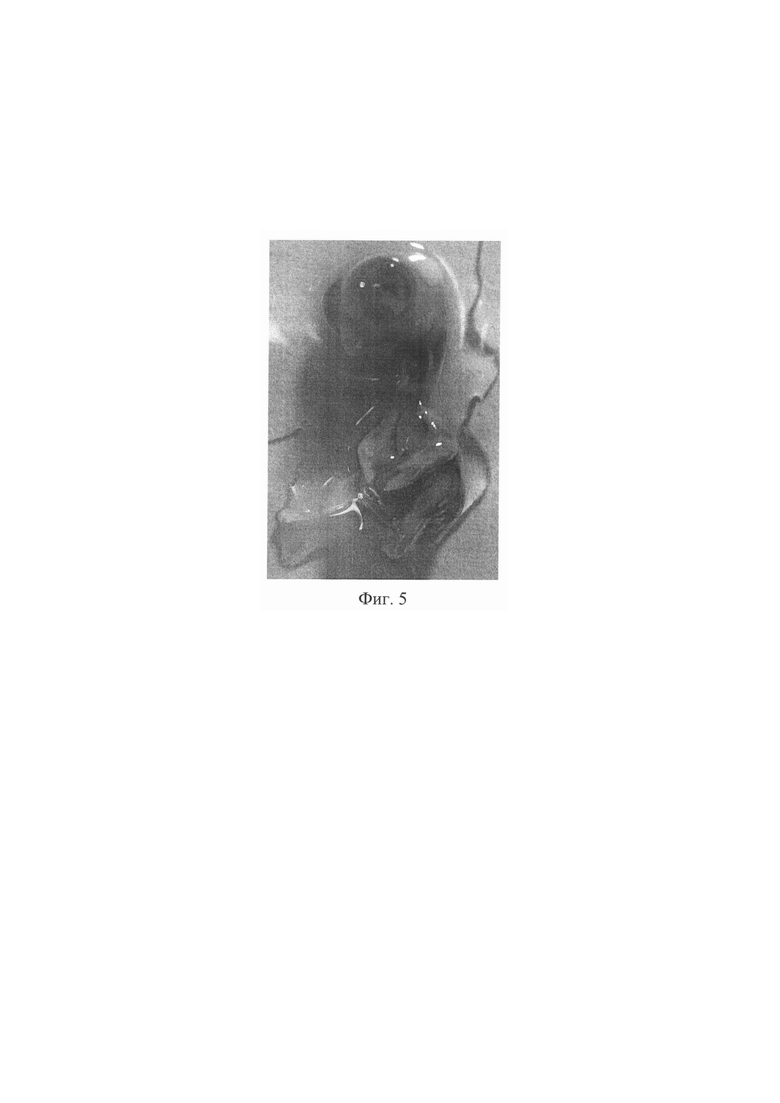
Предлагаемое изобретение поясняется фотографиями, где на фиг. 1 - 3 представлена последовательность метода: фиг. 1 - полость нервной трубки куриного эмбриона, фиг. 2 - диффузия введенной липофильной siPHK из нервной трубки в сомиты, фиг. 3 - куриный эмбрион после введения вещества. Фигура 4. - абрахия у куриного эмбриона. На фиг. 5 представлен нормальный куриный эмбрион.
Реализация предлагаемого способа поясняется примером конкретного выполнения.
Эксперимент проводился в стерильных условиях. Для реализации поставленной задачи, при выполнении эксперимента, скорлупу фертильного куриного яйца и яйца-донора обрабатывали 70% спиртом. Далее экспериментальное яйцо укладывали на подложку горизонтально, делали прокол стерильной иглой с вертикальной стороны, далее стерильными хирургическими ножницами делали отверстие в размере 0,8x1,5 см, отрезанную скорлупу убирали. Также удаляли подскорлуповую оболочку, контролировали развитие эмбриона и при визуализации формирования 16 пары сомитов, что соответствует 45 часам эмбрионального развития, производили введение липофильной siРНК.
На фиг. 1 видна полость нервной трубки 1 куриного эмбриона. При помощи инжекторного шприца фирмы NARISHAGE и стеклянного капилляра под визуальным контролем через конфокальный микроскоп фирмы (Ziess Discovery V12 SteREO) в нервную трубку 1 куриного эмбриона вводили липофильную siРНК, визуально контролируя заполнение нервной трубки до мозговых пузырей при помощи микроскопа фирмы (Ziess Discovery V12 SteREO) на увеличении 90. Для лучшей визуализации siРНК была предварительно подкрашена трипановым синим.
На фиг.2 визуализируется диффузия введенной siРНК в сегментированной мезодерме (сомиты) развивающегося эмбриона. На фиг. 3 хорошо видно, что анатомическая целостность эмбриона сохранена.
Из скорлупы яйца-донора стерильными ножницами вырезали фрагмент для закрытия отверстия на скорлупе экспериментального яйца. После выполнения манипуляций по введению липофильной siРНК накладывали крышечку из скорлупы яйца-донора вместе с подскорлуповой оболочкой. Оставляли на сутки дефектом вверх, после ротировали яйцо в соответствии с физиологией развития куриного эмбриона.
Экспериментально установлено, что ингибирование РАХ3 гена в период формирования 16 пары сомитов может рассматриваться как патогенетический механизм формирования абрахии. Ингибирование РАХ3 гена в период 45 часов приводит к нарушению миграции клеток нервного гребня через формирующийся апикальный эктодермальный гребень почек верхних конечностей, как следствие, к возникновению абрахии. Изменения миграции клеток нервного гребня - ингибирование экспрессии РАХ3 - создают условия, в результате которых клетки нервного гребня оказываются не в конечном месте и времени их эволюционного финиша.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИНДУЦИРОВАНИЯ ВРОЖДЕННОГО ВЫВИХА БЕДРА НА КУРИНОМ ЭМБРИОНЕ | 2022 |
|
RU2792009C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ БОЛЕЗНИ БЛАУНТА НА КУРИНОМ ЭМБРИОНЕ | 2022 |
|
RU2798219C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ СКОЛИОЗА НА КУРИНОМ ЭМБРИОНЕ | 2019 |
|
RU2728927C1 |
| СПОСОБ БЛОКИРОВАНИЯ СИГНАЛЬНОГО ПУТИ, АКТИВИРУЕМОГО TGF-beta ФАКТОРОМ DERRIERE В КЛЕТКАХ ЖИВОТНЫХ | 2009 |
|
RU2407799C1 |
| СПОСОБ БЛОКИРОВАНИЯ СИГНАЛЬНОГО ПУТИ, АКТИВИРУЕМОГО TGF-beta ФАКТОРОМ VgI В КЛЕТКАХ ЖИВОТНЫХ | 2008 |
|
RU2391352C1 |
| СПОСОБ БЛОКИРОВАНИЯ АКТИВНОСТИ ACTIVIN С ПОМОЩЬЮ НОГГИН2 | 2005 |
|
RU2354662C2 |
| СПОСОБ ЗАКРЫТИЯ ДЕФЕКТА ОПЛОДОТВОРЕННОГО ЯЙЦА ПРИ СОЗДАНИИ ЭКСПЕРИМЕНТАЛЬНОЙ МОДЕЛИ | 2019 |
|
RU2729370C1 |
| СПОСОБ БЛОКИРОВАНИЯ СИГНАЛЬНОГО ПУТИ, АКТИВИРУЕМОГО ФАКТОРОМ Wnt8 В КЛЕТКАХ ЖИВОТНЫХ С ПОМОЩЬЮ БЕЛКОВ СЕМЕЙСТВА NOGGIN | 2011 |
|
RU2473561C1 |
| СПОСОБ ПОЛУЧЕНИЯ ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК ИЗ СТВОЛОВЫХ КЛЕТОК | 2009 |
|
RU2576003C2 |
| СПОСОБ АНАЛИЗА И/ИЛИ ОБРАБОТКИ ОПЛОДОТВОРЕННОГО ЯЙЦА | 2017 |
|
RU2665136C1 |
Изобретение относится к биотехнологии и представляет собой cпособ моделирования индуцированной абрахии на курином эмбрионе, заключающийся во введении в нервную трубку куриного эмбриона липофильной siРНК к гену Рах3 2’-О-метильной модификации в объеме, обеспечивающем заполнение нервной трубки, причем введение липофильной siРНК осуществляют в период формирования 16 пары сомитов. Изобретение обеспечивает моделирование индуцированной абрахии на курином эмбрионе. 5 ил., 1 пр.
Способ моделирования индуцированной абрахии на курином эмбрионе, заключающийся в введении в нервную трубку куриного эмбриона липофильной siРНК к гену Рах3 2’-О-метильной модификации в объеме, обеспечивающем заполнение нервной трубки, причем введение липофильной siРНК осуществляют в период формирования 16 пары сомитов.
| HAMBURGER V | |||
| and al | |||
| A series of normal stages in the development of the chick embryo, Journal of Morphology, 1951, vol | |||
| Шланговое соединение | 0 |
|
SU88A1 |
| TAHARA Y | |||
| et al | |||
| A novel shell-less culture system for chick embryos using a plastic film as culture vessels, The Journal of Poultry Science, 2014, vol | |||
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |
| СПОСОБ МОДЕЛИРОВАНИЯ БОЛЕЗНИ БЛАУНТА НА КУРИНОМ ЭМБРИОНЕ | 2022 |
|
RU2798219C1 |
| СПОСОБ ИНДУЦИРОВАНИЯ ВРОЖДЕННОГО ВЫВИХА БЕДРА НА КУРИНОМ ЭМБРИОНЕ | 2022 |
|
RU2792009C1 |
