Изобретение относится к области молекулярной биологии, конкретно, к белкам, регулирующим клеточную дифференцировку путем ингибирования сигнального каскада, акивируемого белком Wnt8 семейства Wnt, и может найти применение в медицине и клеточных технологиях.
Внутриклеточный сигнальный каскад, активируемый гликолипопротеинами семейства Wnt играет ключевую роль в широком спектре клеточных процессов, таких как пролиферация и поляризация клеток, а также их дифференцировка в эмбриональном развитии (Logan CY, Nusse R., 2004, The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol.; 20:781-810). Нарушения функционирования этого каскада приводят к аномалиям развития - отсутствию конечностей (Niemann et al., 2004, Homozygous WNT3 mutation causes tetra-amelia in a large consanguineous family. Am J Hum Genet. Mar; 74(3):558-63. Epub 2004 Feb 5), нарушению плотности костей, аномалиям развития зубов и глазных сосудов (Boyden et al., 2002, High bone density due to a mutation in LDL-receptor-related protein 5. N Engl J Med. May 16; 346 (20): 1513-21, Gong Y. et al., 2001, LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Osteoporosis-Pseudoglioma Syndrome Collaborative Group.Cell. Nov 16; 107(4):513-23., Toomes et al. 2004. Mutations in LRP5 or FZD4 underlie the common familial exudative vitreoretinopathy locus on chromosome 11q. Am; J. Hum. Genet. 74:721-30, Lammi et al. 2004. Mutations in AXIN2 cause familial tooth agenesis and redispose to colorectal cancer. Am. J. Hum. Genet. 74: 1043-50) и заболеваниям, в том числе онкологическим - раку толстой кишки (Lammi et al. 2004. Mutations in AXIN2 cause. familial tooth agenesis and redispose to colorectal cancer. Am. J. Hum. Genet. 74: 1043-50), меланоме (Morin PJ. 1999 beta-catenin signaling and cancer. Bioessays. Dec; 21(12):1021-30), карциноме (Satoh et al. 2000. AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus-mediated transfer of AXIN1. Nat. Genet. 24:245-50) и др. (Clevers H. 2006 Wnt/beta-catenin signaling in development and disease. Cell. Nov 3; 127(3):469-80).
Факторы роста семейства Wnt высококонсервативны у всех типов многоклеточных животных (Gordon and Nusse, 2006; McDonald et al., 2009).
Активация Wnt сигнального каскада происходит через взаимодействие молекул Wnt с цистеин-богатым доменом трансмембранного рецептора Frizzled в присутствии корецептора LRP (Gordon and Nusse, 2006). При активации Wnt каскада происходит накопление в цитоплазме белка бета-катенина, который переносится в ядро и регулирует активность генов-мишеней (Gordon and Nusse, 2006).
Белок Wnt8, член семейства белков Wnt, присутствует у всех классов позвоночных животных.
Показано, что он регулирует экспрессию Нох генов, в частности таких их кластеров, как hoxd, hoxa и hoxb, и таким образом участвует в передне-задней дифференцировке зародыша, а также установлении паттернов многих генов-мишеней (der Rieden et al., 2010).
В связи с этим важной задачей является поиск регуляторов функционирования белка Wnt8, в том числе поиск его антагонистов.
Идентифицирован ряд белков межклеточного матрикса, являющихся антагонистами представителей надсемейства Wnt. Секретируемые белки sFRPs, связанные с рецептором Frizzled, и ингибиторный белок WIF связывают молекулы Wnt, подавляя активность сигнального каскада (Bovolenta et al., 2008). Кроме этого выявлены два семейства белков-ингибиторов Wnt - семейство Dickkopf (DKK) (Ellwanger et al., 2008; Semenov et al., 2008; Wang et al., 2008) и семейство WISE/SOST (Semenov et al., 2005).
Белок noggin1, принадлежащий к семейству белков noggin (Noggin) позвоночных, известен в качестве антагониста внутриклетчного сигнального каскада, активируемого ростовыми факторами из суперсемейства TGF-beta BMP (bone morphogenetic proteins). В силу этого свойства белок noggin1 играет ключевую роль в широком спектре онтогенетических процессов, таких как закладка и формирование нервной системы позвоночных (Dale and Slack, 1987; Lamb et al., 1993; Smith and Harland, 1992; Smith et al., 1993; Slack and Tannahill, 1993), дифференцировка и установление паттерна зачатка нервной системы и мезодермы сомитов (Knecht and Harland; 1997; McMahon et al., 1998), развитие суставов и черепных швов (Brunet et al., 1998; Warren et al., 2003), дифференцировка волосяных луковиц (Botchkarev et al., 1999). Также показано, что Noggin1 может подавлять развитие кровеносных сосудов (Reese et al., 2004).
Известен гомолог белка позвоночных noggin1, названный noggin2 (Fletcher et al., Gene Expr Patterns. 2004, V.5(2), p.225-230; Eroshkin et al., Gene Expr Patterns. 2004, V.6, p.180-186). Было показано, что ген noggin2 имеет дифференциальный характер экспрессии в ходе развития Xenopus. Однако функция noggin2 не исследована.
Известны способы регуляции активности TGF-beta сигнальных каскадов белками семейства noggin, включающие регуляцию активности Smad1 зависимого каскада, активируемого белками семейства BMP (ADMP, BMP2, ВМР4, ВМР7, CDF6) с помощью белка noggin1, и регуляцию Smad2 зависимого каскада, активируемого белком Activin с помощью белка noggin2 (Патент РФ №2354662). Введение белков noggin1 и noggin2 в ткани организма может производиться либо путем введения матричных ДНК или РНК конструкций, кодирующих данный белок, либо непосредственным введением рекомбинантного белка noggin1 (Brunet et al., (1998) Science 280, 1455-1457; Lamb et al., (1993) Science 262, 713-718; Slack J.M. & Tannahill, D. (1993). Noggin the dorsalizer. Nature 361, 498-499; US Patent 5670481, Dorsal tissue affecting factor (noggin) and compositions comprising same, изобретатели: Harland; Richard M., Smith; William С., дата публикации: September 23, 1997).
Способность представителей семейства белков noggin регулировать активность Wnt сигнального каскада ранее не исследовалась.
Наиболее близкие аналоги предлагаемого способа описаны:
1) Zimmerman et al., (1996) Cell. 23:86(4):599-606. Данный аналог представляет собой способ блокирования Smad1 зависимого TGF-beta сигнального каскада, активируемого факторами семейства BMP (bone morphogenetic protein), путем прямого связывания молекул ВМР4 белком noggin. Однако наряду с остальными известными методами описанный способ не позволяет осуществлять регуляцию Wnt сигнального каскада с помощью белков семейства Noggin.
2) Piccolo et al. (1999) Nature, 25; 397(6721): 707-710. Данный наиболее близкий аналог предлагаемого способа представляет собой способ блокирования Wnt сигнального каскада, активируемого белком Wnt8, путем связывания молекул Wnt8 белком Cerberus.
Изобретение решает задачу блокирования сигнального пути, активируемого фактором Wnt8 в клетках животных.
Поставленная задача решается за счет способа блокирования сигнального пути, активируемого фактором Wnt8 в клетках животных, путем введения белка noggin1 или белка noggin2 в ткань, содержащую белок Wnt8, в количестве, эффективном для ингибирования активности Wnt8.
Изобретение впервые устанавливает, что белки noggin1 и noggin2 способны регулировать активность Wnt сигнального каскада и являются блокаторами одного из индукторов этого каскада - фактора Wnt8.
Сущность способа заключается во введении белка noggin1 или noggin2 в организм, ткань или клетку в количестве, эффективном для блокирования или снижения активности Wnt8, посредством препятствования связыванию этих факторов со специфическими белковыми рецепторами.
Указанный способ может быть использован для модификации клеточной дифференцировки, где он предусматривает приведение клетки или окружающей клетку среды в контакт с белком noggin1 или noggin2. При осуществлении способа может быть использована экспрессирующая конструкция, содержащая кодирующую noggin1 или noggin2 нуклеиновую кислоту под контролем подходящего промотора. В этом осуществлении способ изменения клеточной дифференцировки предусматривает создание экспрессирующей конструкции, содержащей кодирующую noggin1 или noggin2 нуклеиновую кислоту под контролем подходящего промотора; введение указанной экспрессирующей конструкции в клетку для экспрессии noggin1 или noggin2, где экспрессия noggin1 или noggin2 приводит к изменению клеточной дифференцировки.
Указанный способ можно применять для профилактики и лечения заболеваний, связанных с нарушениями функций Wnt сигнального каскада, например рака толстой кишки, меланомы, карциномы.
В тексте используются следующие сокращения:
кДНК - ДНК, комплиментарная матричной РНК
мРНК - матричная РНК
пг - пикограмм
ПЦР - полимеразная цепная реакция
Изобретение иллюстрируют графические материалы.
Фигура 1. Связывание белка Wnt8 белками noggin1 и noggin2 и влияние факторов noggin1 и noggin2 на активность Wnt сигнального каскада при его активации факатором Wnt8.
А. Белки noggin1 и noggin2 связывают молекулы костного морфогенетического белка (BMP) (положительный контроль) и молекулы белка Wnt8. Подробное описание приведено в тексте примера 1.
Б. Подавление экспрессии репортерной конструкции Top Flash при коинъекции мРНК генов noggin1 и noggin2 в смеси с мРНК гена Wnt8. Активация Wnt сигнального каскада осуществляется путем микроинъекций мРНК гена Wnt8. Уровень активности Wnt каскада приведен на оси ординат в относительных единицах, за 1 единицу принят уровень сигнала контрольных зародышей (без инъекций мРНК Wnt8).
В. Подавление экспрессии генетической мишени Wnt сигнального каскада - гена Xnr3 при коинъекции мРНК генов noggin1 и noggin2 в смеси с мРНК гена Wnt8. Активация Wnt сигнального каскада осуществляется путем микроинъекций мРНК гена Wnt8. Уровень экспрессии гена Xnr3 приведен на оси ординат в относительных единицах, за 1 единицу принят уровень сигнала контрольных зародышей (без инъекций мРНК Wnt8).
Г. Концентрационная зависимость способности факторов noggin1 и noggin2 подавлять экспрессию репортерной конструкции Top Flash при активации Wnt сигнального каскада фактором Wnt8. Уровень активности Wnt каскада приведен на оси ординат в относительных единицах, за 1 единицу принят уровень сигнала контрольных зародышей (без инъекций мРНК Wnt8). Количество noggin белков (ось абсцисс) приведено в пг на зародыш.
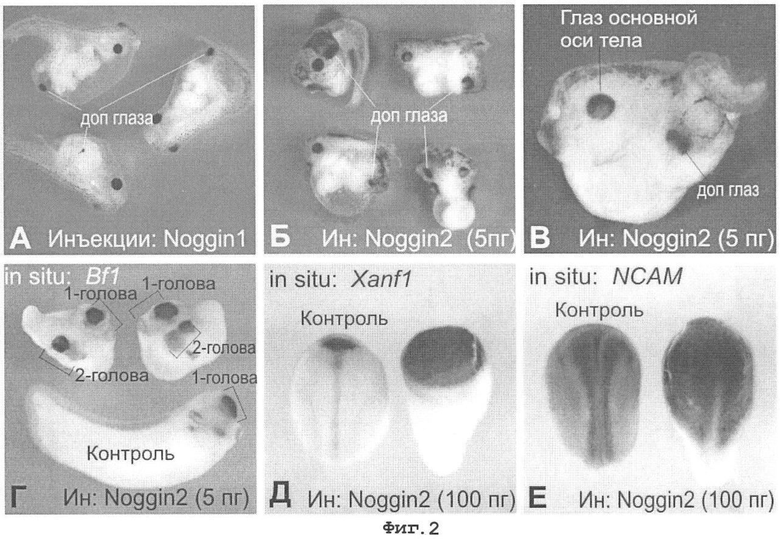
Фигура 2. Влияние noggin1 и noggin2 на формирование осей тела и дифференцировку нервной ткани в эмбрионах Xenopus laevis.
А. Вентральные микроинъекции 5 пг синтетической мРНК гена noggin1 в зародыши X. laevis на ранних стадиях развития приводят к формированию дополнительной оси тела, содержащей структуры, специфичные переднего мозга, в частности глазные структуры.
Б, В. Вентральные микроинъекции 5 пг синтетической мРНК гена noggin2 в зародыши X. laevis на ранних стадиях развития приводят к формированию дополнительной оси тела, содержащей структуры, специфичные переднего мозга, в частности глазные структуры.
Г. In situ гибридизация зародышей X. laevis, микроинъецированных 5 пг синтетической мРНК гена noggin2 в зародыши, демонстрирует активацию экспрессии молекулярного маркера переднего могза - гена Bf1 в дополнительно оси тела.
Д, Е. In situ гибридизация контрольных зародышей (слева на каждой их фотографий) и зародышей, микроинъецированных мРНК noggin2 с пробами мРНК приведенных генетических маркеров, показывает усиление нейральной дифференцировки. Сигнал показан черным.
Фигура 3. Способность гибридных молекул факторов noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP, восстанавливать деффекты развития, вызываемые экспериментальным введением в зародыши мРНК Wnt8 и экспрессионного вектора, содержащего кДНК Wnt8.
А-В'. Способность гибридных молекул факторов noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP, подавлять формирование дополнительных осей тела у зародышей, микроинъецированных на ранних стадиях развития мРНК Wnt8. Для оценки распределения инъецированного материала в инъецируемую смесь добавляют зеленый флуоресцентный краситель Fluorescein Lysine Dextran (A', Б', В').
Г-Е'. Способность гибридных молекул факторов noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP, восстанавливать развитие глазных структур, нарушаемое при микроинъециях в зародыши на ранних стадиях развития экспрессионной конструкции, содержащей кДНК Wnt8. Для оценки распределения инъецированного материала в инъецируемую смесь добавляют зеленый флуоресцентный краситель Fluorescein Lysine Dextran (Г', Д', Е').
Изобретение иллюстрируют примеры.
Пример 1.
Связывание белка Wnt8 белками noggin1 и noggin2.
Введение белков noggin1, noggin2 и Wnt8 в зародыши X. laevis осуществляют путем их экспрессии в составе плазмидных векторов pCS2-3Myc-noggin1, pCS2-3Myc-noggin2 и pCS2-3Flag-Wnt8 соответственно.
Для получения вектора pCS2-3Myc-noggin1, нуклеотидную последовательность, кодирующую три аминокислотные последовательности эпитопа белка Мус (EQKLISEEDLEQKLISEEDLEQKLISEEDL), вставляют с помощью ПЦР в кДНК noggin2 непосредственно на 3' конец последовательности, кодирующей сигнальный пептид noggin2. Для этого на первой стадии клонирования получают соответственно 5' и 3' перекрывающиеся части кДНК noggin2 в ПЦР с праймерами:
для 5' области:

На второй стадии эти фрагменты кДНК очищают от не включившихся праймеров, смешивают друг с другом, денатурируют нагреванием, отжигают и подвергают второму раунду ПЦР с концевыми праймерами:

Полученную полноразмерную кДНК noggin1, содержащую последовательность, кодирующую три аминокислотные последовательности Мус, клонируют в вектор pCS2 по рестриктным сайгам NcoI(тупой)/AgeI(тупой) и XhoI/XhoI.
Для получения вектора pCS2-3Myc-noggin2 нуклеотидную последовательность, кодирующую три аминокислотные последовательности эпитопа белка Мус (EQKLISEEDLEQKLISEEDLEQKLISEEDL), вставляют с помощью ПЦР в кДНК noggin2 непосредственно на 3' конец последовательности, кодирующей сигнальный пептид noggin2. Для этого на первой стадии клонирования получают соответственно 5' и 3' перекрывающиеся части кДНК noggin2 в ПЦР с праймерами:
для 5' области:

для 3' области:

На второй стадии эти фрагменты кДНК очищают от не включившихся праймеров, смешивают друг с другом, денатурируют нагреванием, отжигают и подвергают второму раунду ПЦР с концевыми праймерами 

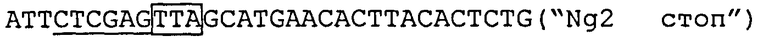 .
.
Полученную полноразмерную кДНК noggin2, содержащую последовательность, кодирующую три аминокислотные последовательности Мус, клонируют в вектор pCS2 по рестриктным сайтам NcoI(тупой)/AgeI(тупой) и XhoI/XhoI.
Вектор pCS4-3Flag-Wnt8 получают путем клонирования кДНК Wnt8, полученной методом ПЦР с праймерами:

в вектор pCS4-3Flag, содержащий три аминокислотные последовательности Flag (DYKDDDDKDYKDDDDKDYKDDDDK), по рестриктным сайтам BamHI (тупой) и NcoI/PagI.
Анализ способности экспрессированных белков 3Мyс-noggin1 и 3Myc-noggin2 связывать молекулы белка 3Flag-Wnt8 проводят методом коиммунопреципитации. Для этого 600 pg синтетической мРНК 3Мус-nogginl (или 3Myc-noggin2) и 600 pg синтетической мРНК 3Flag-Wnt8 микроинъецируют в развивающиеся зародыши X. laevis на стадии 2-х бластомеров. В дальнейшем эти зародыши инкубируют в растворе 0.1 MMR до стадии ранней гаструлы.
Выделение экспрессированных белков проводят путем лизирования зародышей на стадии ранней гаструлы по методике, описанной в статье Tanegashima et al., Int.J.Dev.Biol (2004) 48:275-283.
После выделения проводят абсорбцию зрелого белка 3Мус-noggin1 (или 3Myc-noggin2) на смоле Myc-Agarose (Sigma) согласно протоколу производителя. После промывки смолы в соответствии с протоколом производителя наличие на этой смоле связавшегося белка 3Flag-Wnt8 анализируют методом белкового блоттинга с помощью Flag-специфических моноклональных антител (Sigma).
Способность молекул белков noggin1 и noggin2 связывать молекулы белка Wnt8 показана на Фигуре 1А.
В качестве положительного контроля в эксперименте используют белок 3Flag-BMP, связываемый белками noggin1 и noggin2.
Пример 2.
Способность noggin1 и noggin2 подавлять активность Wnt сигнального каскада при его активации фактором Wnt8.
кДНК, содержащую полную кодирующую область noggin1,  ("Ng1 стоп") и клонируют в вектор рС32 (вектор pCS2-noggin2).
("Ng1 стоп") и клонируют в вектор рС32 (вектор pCS2-noggin2).
кДНК, содержащую полную кодирующую область noggin2, реамплифицируют при помощи ПЦР с клона, полученного в работе Eroshkin et al., Gene Expr Patterns. 2004, V.6, p.180-186, с  ("Ng2 стоп'') и клонируют в вектор pCS2 (вектор pCS2-noggin2).
("Ng2 стоп'') и клонируют в вектор pCS2 (вектор pCS2-noggin2).
Вектор pCS2-Wnt8 получают путем клонирования кДНК Wnt8, полученной методом ПЦР с праймерами:

в вектор pCS2 по рестриктным сайтам BamHI и XhoI.
Синтетические мРНК noggini, noggin2 и Wnt8 получают с применением набора реактивов mMESSAGE MASHINE Kit (Ambion) из векторов pCS2-noggin1, pCS2-noggin2 и вектора pCS2-Wnt8 соответственно, линеаризованных рестрикционной эндонуклеазой EcoRI (Fermentas).
В ходе данных экспериментов мРНК Wnt8, noggin1 и noggin2 инъецируют в один из вентральных вегетативных бластомеров на стадии восьми бластомеров с применением микроинъектора Eppendorf.
Для анализа влияния белков noggin1 и noggin2 на активность Wnt сигнального каскада используют метод, основанный на применении люциферазного репортера. Зародыши X. laevis на стадии 2-4 бластомеров инъецируют смесью, состоящей из люциферазной репортерной плазмиды TOP flash (50 пг/зародыш), в которой ген люциферазы находится под контролем промотора, содержащего цис-регуляторные элементы, распознаваемые транскрипционными факторами, участвующими в функционировании Wnt сигнального каскада, плазмиды pCMV-β-GAL (50 пг/зародыш), содержащей ген β-галактозидазы, для оценки общего уровня биосинтеза белка в системе, и мРНК исследуемых белков.
Измерение уровня экспрессии гена люциферазы проводят с помощью люминометра TD-20/20 Luminometer (Turner Designs) с использованием системы Lusiferase Assay system (Promega) согласно инструкции.
Уровень синтеза β-галактозидазы в лизатах зародышей измеряют с использованием субстрата ONPG (O-nitrophenil-D-galactopyranoside) на приборе iMark Microplate Reader(Bio-RAD) по стандартной методике (Martynova et al., 2008 Dev Dyn. 237(3):736-49).
Активацию Wnt сигнального каскада производят путем микроинъекции мРНК Wnt8 в один из вентральных вегетативных бластомеров на стадии восьми бластомеров.
При микроинъекции 15 пг мРНК Wnt8 на бластомер наблюдается повышение активности Wnt сигнального каскада в пять раз (Фигура 1, Б). При добавлении в микроинъецируемую смесь избыточного количества (300 пг) мРНК белков noggin1 и noggin2 уровень активности Wnt сигнального каскада понижается практически до уровня контроля. При активации Wnt сигнального каскада путем микроинъекции мРНК beta-catenin - эндогенной мишени Wnt каскада подавление активности Wnt каскада noggin белками не наблюдается. Это соответствует способности noggin белков регулировать активность Wnt каскада на экстрацеллюлярном уровне путем связывания молекул Wnt8. Инъекции мРНК Smad1 и Smad2 не влияют на способность noggin белков подавлять активность Wnt сигнального каскада, что соответствует специфичности влияния noggin белков на активность Wnt каскада.
Для анализа влияния белков noggin1 и noggin2 на активность Wnt сигнального каскада используют также метод обратной транскрипции-ПЦР (ОТ-ПЦР) в реальном времени.
ОТ-ПЦР на основе тотальной РНК из эмбрионов, подвергнутых инъекции мРНК Wnt8, а также смеси мРНК Wnt8 и мРНК noggin1 или noggin2, осуществляют с праймерами к маркеру активности и генетической мишени Wnt сигнального каскада - Xnr3: 5'-CTTCTGCACTAGATTCTG-S' и 5'-CAGCTTCTGGCCAAGACT-3'(Xanthos et al., 2002).
При микроинъекции 15 пг мРНК Wnt8 на бластомер в зародыши X. laevis наблюдается десятикратное повышение уровня экспрессии гена Xnr3 (Фигура 1В). При добавлении в микроинъецируемую смесь избыточного количества (300 пг) мРНК белков noggin1 и noggin2 уровень экспрессии гена Xnr3 понижается практически до уровня контроля. При активации Wnt сигнального каскада путем микроинъекции мРНК beta-catenin - эндогенной мишени Wnt каскада, подавление уровня экспрессии гена Xnr3 noggin белками не наблюдается. Это соответствует способности noggin белков регулировать активность Wnt каскада на экстрацеллюлярном уровне путем связывания молекул Wnt8. Инъекции мРНК Smad1 и Smad2 не влияют на способность noggin белков подавлять уровень экспрессии гена Xnr3, что соответствует специфичности влияния noggin белков на активность Wnt каскада.
Подавление активности Wnt сигнального каскада белками noggin1 и noggin2 является концентрационно зависимым и при постоянном уровне активации Wnt каскада, степень его подавления возрастает с повышением концентрации noggin белков (Фигура 1Г).
Пример 3.
Способность noggin1 и noggin2 вызывать формирование дополнительных осей тела, содержащих головные структуры, и влиять на дифференцировку нервной ткани в эмбрионах Xenopus laevis.
мРНК noggin1 и noggin2 получают, как описано в примере 2.
Микроинъекции мРНК noggin1 и noggin2 осуществляют в один из вентральных вегетативных бластомеров на стадии восьми бластомеров с применением микроинъектора Eppendorf.
Микроинъекции 5 пг синтетической мРНК гена noggin1 в зародыши X. laevis на ранних стадиях развития приводят к формированию дополнительной оси тела, содержащей глазные структуры, специфичные для переднего мозга (Фигура 2А).
Микроинъекции 5 пг синтетической мРНК гена noggin2 в зародыши X. laevis на ранних стадиях развития также приводят к формированию дополнительной оси тела, содержащей глазные структуры, специфичные для переднего мозга (Фигура 2Б, В).
Гибридизация in situ на тотальном препарате эмбрионов X. laevis с инъекцией мРНК noggin2 с зондами к мРНК маркера переднего мозга Bf1 подтверждает способность фактора noggin2 вызывать формирование дополнительных переднеголовных структур (Фигура 2Г).
Гибридизация in situ на тотальном препарате эмбрионов X. laevis с инъекцией мРНК noggin2 с зондами к мРНК нейральных (NCAM, Xanf-1) показывает гигантскую нейрализацию эктодермы (Фигура 2Д, Е).
Поскольку согласно современным представлениям (Piccolo et al. (1999) Nature, 25; 397(6721): 707-710) подавление Wnt сигнального каскада является наряду с подавлением BMP Nodal/Activin необходимым условием формирования головных структур, приведенные данные указывают на то, что факторы noggin1 и noggin2 обладают способностью подавлять активность Wnt сигнального каскада.
Анализ методом гибридизации in situ подтверждает сильную нейрализующую активность фактора noggin2 (Фигура 2Е), который при этом обнаруживает способность индуцировать экспрессию маркеров передней части нервной системы (Фигура 2Г, Д).
Пример 4.
Способность гибридных молекул белков noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP, подавлять активность Wnt сигнальнго каскада и восстанавливать деффекты развития, вызываемые экспериментальным введением в зародыши мРНК Wnt8 и экспрессионного вектора, содержащего кДНК Wnt8, соответствует способности белков noggin1 и noggin2 подавлять активность Wnt сигнального каскада.
Для экспериментов используют гибридные молекулы белков noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP - Δnoggin1 и Δnoggin2 соответственно. Нарушение функции связывания молекул BMP осуществляется путем делеции 28-ми аминокислот в N-концевой части молекул белков noggin1 и noggin2.
Для получения векторов, содержащих кДНК Δnoggin1 и Δnoggin2, методом ПЦР получают 5' и 3'- фрагменты будущих кДНК с праймерами:
для 5' фрагментов -
"Ng1 синтетический 5" и
5'-CTTCTCCTTGGGATAATGTTGGCAACCCCCTT;
"Ng2 синтетический 5" и
5'-CTGCTCCTTTGGATAAGGCTGACAGCACCCCT.
для 3' фрагментов -
5'-TTGCCAACATTATCCCAAGGAGAAGGATCTTA и "Ng1 стоп";
5'-CTGTCAGCCTTATCCAAAGGAGCAGGATCTGG и "Ng2 стоп".
На второй стадии полученные фрагменты кДНК очищают от не включившихся праймеров, смешивают друг с другом, денатурируют нагреванием, отжигают и подвергают второму раунду ПЦР с концевыми праймерами: "Ng1 синтетический 5" и "Ng1 стоп" для Δnoggin1 и "Ng2 синтетический 5" и "Ng2 стоп" для Δnoggin2. Полученные кДНК клонируют в вектор рС32 по рестриктным сайтам AgeI/BamHI (тупой) и XhoI.
Вектор pCS2-Wnt8 получают путем, описанным в Примере 2.
Микроинъекции мРНК Wnt8 осуществляют в один из вентральных вегетативных бластомеров на стадии восьми бластомеров с применением микроинъектора Eppendorf.
Для анализа влияния белков Δnoggin1 и Δnoggin2 на активность Wnt сигнального каскада используют метод, основанный на применении люциферазного репортера (подробно описан в Примере 2).
При микроинъекции 15 пг мРНК Wnt8 на бластомер наблюдается повышение активности Wnt сигнального каскада в пять раз (Фигура 1Б). При добавлении в микроинъецируемую смесь избыточного количества (300 пг) мРНК белков Δnoggin1 и Δnoggin2 уровень активности Wnt сигнального каскада понижается практически до уровня контроля.
При микроинъекции 15 пг мРНК Wnt8 на бластомер в зародыши X. laevis наблюдается десятикратное повышение уровня экспрессии гена Хnr3 (Фигура 1В). При добавлении в микроинъецируемую смесь избыточного количества (300 пг) мРНК белков Δnoggin1 и Δnoggin2 уровень экспрессии гена Xnr3 понижается практически до уровня контроля.
Эти данные соответствуют способности гибридных молекул белков noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP, подавлять активность Wnt сигнальнго каскада
Микроинъекции мРНК Wnt8 в один из вентральных вегетативных бластомеров вызывают формирование дополнителных осей тела, которое подавляется при добавлении в микроинъецируемый материал мРНК noggin1 или noggin2 (Фигура 3А-В').
Вектор pCSKA-Wnt8, содержащий кДНК Wnt8 (Christian J.L. and Moon R.Т., 1993, Genes Dev.7, 13-28), используется в экспериментах с целью предотвращения летальных последствий повышенной экспрессии Wnt8 в головной области на ранних стадиях развития, поскольку трансляция белка в случае применения этого вектора начинается на стадии ранней гаструлы. Микроинъекции линеаризованного вектора pCSKA-Wnt8 осуществляют в один из анимальных бластомеров на стадии четырех бластомеров с применением микроинъектора Eppendorf.
Микроинъекции вектора pCSKA-Wnt8 приводят к нарушениям формирования головных и, в частности, глазных структур вплоть до полного их отсутствия. Добавление в микроинъецируемый материал мРНК noggin1 или noggin2 приводит к нормализации развития глазных структур - так называемый эффект «спасения» (Фигура 3Г-Е').
Использование в эксперименте молекул белков noggin1 и noggin2 с нарушенной способностью связывать молекулы BMP соответствует специфичности наблюдаемого эффекта в отношении Wnt сигнального каскада.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ БЛОКИРОВАНИЯ СИГНАЛЬНОГО ПУТИ, АКТИВИРУЕМОГО TGF-beta ФАКТОРОМ DERRIERE В КЛЕТКАХ ЖИВОТНЫХ | 2009 |
|
RU2407799C1 |
| СПОСОБ БЛОКИРОВАНИЯ СИГНАЛЬНОГО ПУТИ, АКТИВИРУЕМОГО TGF-beta ФАКТОРОМ VgI В КЛЕТКАХ ЖИВОТНЫХ | 2008 |
|
RU2391352C1 |
| СПОСОБ БЛОКИРОВАНИЯ АКТИВНОСТИ ACTIVIN С ПОМОЩЬЮ НОГГИН2 | 2005 |
|
RU2354662C2 |
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ СТВОЛОВЫХ КЛЕТОК В МИОКАРДИАЛЬНЫЕ | 2004 |
|
RU2392315C2 |
| ОРГАНОИД ПЕЧЕНИ, ВАРИАНТЫ ЕГО ПРИМЕНЕНИЯ И СПОСОБ КУЛЬТИВИРОВАНИЯ ДЛЯ ЕГО ПОЛУЧЕНИЯ | 2011 |
|
RU2579995C2 |
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК В КАРДИОМИОЦИТЫ | 2007 |
|
RU2433174C2 |
| ДИФФЕРЕНЦИРОВКА ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2008 |
|
RU2465323C2 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ | 2017 |
|
RU2772435C2 |
| КУЛЬТУРАЛЬНАЯ СРЕДА ДЛЯ ЭПИТЕЛИАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК И ОРГАНОИДОВ, СОДЕРЖАЩИХ УКАЗАННЫЕ СТВОЛОВЫЕ КЛЕТКИ | 2010 |
|
RU2555545C2 |
| КУЛЬТУРАЛЬНАЯ СРЕДА СТВОЛОВЫХ КЛЕТОК | 2012 |
|
RU2714256C2 |

Изобретение относится к области молекулярной биологии, конкретно, к белкам, регулирующим клеточную дифференцировку путем ингибирования белка семейства Wnt, и может быть использовано в медицине для профилактики и лечения связанных с нарушениями активности Wnt сигнального пути заболеваний, например рака толстой кишки, меланомы, карциномы. Изобретение позволяет блокировать сигнальный путь, активируемый фактором семейства Wnt8 в клетках животных, путем введения белка noggin1 или noggin2 в ткань, содержащую белок Wnt8, в количестве, эффективном для ингибирования активности Wnt8. 3 ил., 4 пр.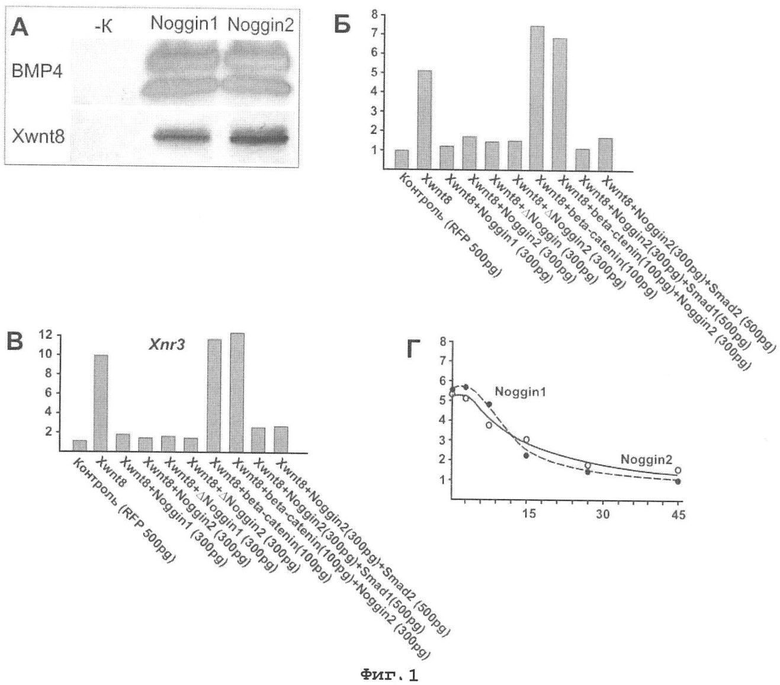
Способ блокирования сигнального пути, активируемого фактором семейства Wnt8 в клетках животных, путем введения белка noggin1 или noggin2 в ткань, содержащую белок Wnt8, в количестве, эффективном для ингибирования активности Wnt8.
| PICCOLO S | |||
| et | |||
| al., The head inducer Cerberus is a multifunctional antagonist of Nodal, BMP and Wnt signals | |||
| Nature, 1999, v.397, n.6721, p.707"710 | |||
| KAWANO Y | |||
| et | |||
| al., Secreted antagonists of the Wnt signalling pathway, J Cell Sci, 2003, v.116, p.2627-2634 | |||
| RU 2009113023 A, 20.10.2010 | |||
| PETERSEN С.P | |||
| et | |||
| al., Wnt signaling and polarity of the |

