В данном документе представлены способы, предназначенные, среди прочего, для увеличения выхода биологически активных полипептидов, полученных рекомбинантным способом.
Уровень техники
В настоящее время большинство биофармацевтических препаратов производятся в клетках животных (млекопитающих), которые обычно секретируют рекомбинантный полипептид, представляющий интерес, с высокой эффективностью, качеством и соответствующими посттрансляционными (вторичными) модификациями (такими как, например, гликозилирование) в культивационную среду. Тем не менее, некоторые гибридные полипептиды, в особенности сложные гибридные полипептиды, полипептиды с низкой растворимостью или трудностями сворачивания, а также полипептиды, взаимодействующие с клеткой, экспрессирующей их, часто получают с очень низким выходом.
Например, антитела, как правило, экспрессируются в клетках млекопитающих с высокой эффективностью в биологически активной форме. Тем не менее, гибридные полипептиды, содержащие антитело, например, конъюгаты антител с зеленым флуоресцентным белком (GFP), совсем не экспрессируются и/или не секретируются, хотя такие белки были бы весьма интересны для экспериментальных и диагностических подходов (см., например, WO 2011/135040).
В некоторых случаях трудно экспрессируемые полипептиды могут быть получены в виде растворимых секретируемых неактивных белков-предшественников, например, так называемых зимогенов в случае протеаз, которые впоследствии могут подвергаться созреванию in vitro, например, путем протеолитической активации. В других случаях полипептиды, которые являются вредными для конкретной клетки-хозяина, могут быть экспрессированы в виде неактивных нерастворимых белковых агрегатов в клетке (телец включения (inclusion bodies, IB)), а потом подвергнуты рефолдингу in vitro. Тем не менее, процессинг прополипептидов может быть трудным или вообще невозможным. Кроме того, полученные зрелые полипептиды не содержат все посттрансляционные модификации.
Примерами слабо экспрессируемых полипептидов являются нейротрофические факторы, такие как NGF, BDNF, GDNF и NT-3 (см., например, Xia, Ch.-F., J. Gene Med. 10 (2008) 306-315; Boado, R.J., Pharm. Res. 24 (2007) 1772-1787; Negro, A., et al., J. Neurochem. 62 (1994) 471-478).
В публикации WO 2008/005847 сообщается о способе получения белков фактора VIII с помощью рекомбинантных методик. Удлиненные белком гликозилированные полипептиды описаны в WO 02/02597. В WO 2007/044323 сообщается о гибридных белках для доставки через гематоэнцефалический барьер. Модуляторы роста нервных клеток (амфитела) описаны в WO 00/64482. В WO 2012/087835 представлены композиции и способы улучшения сворачивания белков. Стабильные и растворимые антитела, ингибирующие TNF-альфа, описаны в WO 2006/131013. В WO 00/23473 сообщается о гибридных белках интерферона бета и их применении.
Сущность изобретения
В данном документе представлены способы улучшения процесса получения рекомбинантно продуцируемого полипептида.
Улучшение может представлять собой, например, повышенный выход, более надежный процесс получения, более простой процесс и/или уменьшение сложности последующей обработки.
Нужно указать, что улучшение достигается без ухудшения биофизических и/или биохимических свойств и/или биологических функций полипептида. В некоторых случаях одно или более чем одно из этих свойств даже улучшено.
Первый аспект, описанный в данном документе, заключается в том, что было обнаружено, что путем введения одного или более чем одного сайта гликозилирования может быть улучшена рекомбинантная продукция полипептида в клетке млекопитающего.
Один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида с использованием вариантного полипептида, включающий следующие этапы:
- культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую вариантный полипептид, где аминокислотная последовательность полипептида была модифицирована с помощью i) одной или более чем одной мутации поверхностно расположенных аминокислотных остатков, что приводит к более низкой изоэлектрической точке вариантного полипептида по сравнению с данным полипептидом, и/или ii) линкерного пептида, соединяющего два полипептида гибридного полипептида, и/или iii) N- или С-концевой слитой метки, содержащей аминокислоты, приводящие к более низкой изоэлектрической точке полипептида,
- извлечение рекомбинантного вариантного полипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
Таким образом, в данном документе описан способ получения рекомбинантного полипептида с использованием вариантного полипептида, включающий следующие этапы:
- культивирование эукариотической клетки (в одном из воплощений клетки млекопитающего), содержащей нуклеиновую кислоту, кодирующую вариантный полипептид, где аминокислотная последовательность полипептида была модифицирована так, чтобы включать один или более чем один искусственный сайт гликозилирования,
- извлечение вариантного рекомбинантного полипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
Один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида с использованием вариантного полипептида, включающий следующие этапы:
- получение нуклеиновой кислоты, кодирующей полипептид,
- модификация нуклеиновой кислоты, чтобы она кодировала вариантный полипептид, где аминокислотная последовательность полипептида была модифицирована так, чтобы включать один или более чем один искусственный сайт гликозилирования,
- введение нуклеиновой кислоты в эукариотическую клетку (в одном из воплощений в клетку млекопитающего),
- культивирование эукариотической клетки, и
- извлечение вариантного рекомбинантного полипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
Способы, описанные в данном документе, особенно подходят, например, для полипептидов, которые слабо или вовсе не экспрессируются/секретируются в клетках млекопитающих.
Введенный сайт (сайты) гликозилирования может быть расположен (расположены независимо от других) либо в самом полипептиде, либо в линкерном пептиде, соединяющем два полипептида гибридного полипептида, либо он может быть специфической меткой гликозилирования.
В одном воплощении полипептид содержит метку гликозилирования.
В одном воплощении полипептид содержит искусственный сайт гликозилирования. В одном воплощении искусственный сайт гликозилирования вводится путем точечной мутации поверхностно расположенной аминокислоты.
Второй аспект, описанный в данном документе, заключается в том, что было обнаружено, что если заменить в прополипептиде эндогенный сайт расщепления протеазой между просегментом и зрелым полипептидом на экзогенный (относительно происхождения полипептида) или искусственный сайт расщепления протеазой, то выход зрелого полипептида может быть повышен. Это изменение улучшает процессинг прополипептида в зрелую форму.
Таким образом, один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида с использованием вариантного прополипептида, включающий следующие этапы:
- культивирование эукариотической клетки (в одном из воплощений клетки млекопитающего), содержащей нуклеиновую кислоту, кодирующую полипептид в виде прополипептида (гибридного полипептида просегмента и полипептида), в котором эндогенный сайт ферментативного расщепления между просегментом и полипептидом заменяется экзогенным сайтом расщепления протеазой,
- извлечение рекомбинантного вариантного полипептида или вариантного прополипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
Один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида с использованием вариантного прополипептида, включающий следующие этапы:
- получение нуклеиновой кислоты, кодирующей полипептид в виде прополипептида (гибридного полипептида из просегмента и полипептида),
- модификация нуклеиновой кислоты, чтобы она кодировала вариантный прополипептид, в котором эндогенный сайт ферментативного расщепления между просегментом и полипептидом заменяется экзогенным сайтом расщепления протеазой,
- введение нуклеиновой кислоты в эукариотическую клетку (в одном из воплощений в клетку млекопитающего),
- культивирование эукариотической клетки, и
- извлечение рекомбинантного вариантного полипептида или вариантного прополипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
В одном воплощении экзогенный сайт расщепления протеазой выбирают из группы, содержащей сайт расщепления плазмином, сайт расщепления фурином, сайт расщепления IgA-протеазой, сайт расщепления протеазой TEV (от англ. tobacco etch virus - вируса табачной мозаики), сайт расщепления гранзимом В, сайт расщепления тромбином, сайт расщепления фактором 10, сайт расщепления энтерокиназой, сайт расщепления субтилизином, сайт расщепления катепсином, сайт расщепления металлопротеиназой, сайт расщепления протеазой IDES (от англ. IgG-degrading enzyme of Streptococcus pyogenes - фермент Streptococcus pyogenes, расщепляющий IgG), сайт расщепления протеазой PreScission или их функциональные варианты. В одном воплощении экзогенный сайт расщепления протеазой представляет собой сайт расщепления IgA-протеазой.
В одном воплощении расщепление прополипептида осуществляется в среде культивирования. В одном воплощении экзогенную протеазу, способную к расщеплению экзогенного сайта расщепления протеазой, добавляют в среду культивирования. В одном воплощении добавление происходит на этапе культивирования. В одном воплощении добавление происходит после этапа культивирования.
В одном воплощении экзогенную протеазу экспрессировали совместно из клетки, экспрессирующей прополипептид.
В одном воплощении культивирование является совместным культивированием клетки, экспрессирующей прополипептид, и клетки, экспрессирующей экзогенную протеазу.
В одном воплощении расщепление осуществляется после отделения клеток от среды культивирования. В одном воплощении расщепление происходит во время последующей обработки. В одном воплощении расщепление осуществляется на хроматографической колонке.
Третий аспект, описанный в данном документе, заключается в том, что было обнаружено, что с понижением изоэлектрической точки полипептида рекомбинантная продукция полипептида в клетке млекопитающего может быть повышена.
Таким образом, один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида с использованием вариантного полипептида, включающий следующие этапы:
- культивирование эукариотической клетки (в одном из воплощений клетки млекопитающего), содержащей нуклеиновую кислоту, кодирующую вариантный полипептид, в котором аминокислотная последовательность полипептида была изменена с помощью одной или более чем одной мутации поверхностно расположенных аминокислотных остатков, что давало более низкую изоэлектрическую точку вариантного полипептида по сравнению с полипептидом,
- извлечение рекомбинантного вариантного полипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
Один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида с использованием вариантного полипептида, включающий следующие этапы:
- получение нуклеиновой кислоты, кодирующей полипептид,
- модификация нуклеиновой кислоты, чтобы она кодировала вариантный полипептид, в котором аминокислотная последовательность полипептида была изменена с помощью одной или более чем одной мутации поверхностно расположенных аминокислотных остатков, что давало более низкую изоэлектрическую точку вариантного полипептида по сравнению с данным полипептидом,
- введение нуклеиновой кислоты в эукариотическую клетку (в одном из воплощений в клетку млекопитающего),
- культивирование эукариотической клетки, и
- извлечение рекомбинантного вариантного полипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
В одном воплощении полипептид имеет изоэлектрическую точку выше 9 (высокая изоэлектрическая точка, основная изоэлектрическая точка), а вариантный полипептид имеет изоэлектрическую точку, которая на 0,5 или более единиц рН ниже (более кислая) по сравнению с (родительским/дикого типа) полипептидом.
В одном воплощении полипептид представляет собой нейтрофин/нейротрофический фактор.
В одном воплощении изоэлектрическая точка понижается за счет введения отрицательно заряженной группировки.
В одном воплощении отрицательно заряженная группировка представляет собой линкерный пептид.
В одном воплощении отрицательно заряженная группировка представляет собой остаток поверхностно расположенный аминокислотный остаток. В одном воплощении один или более чем один аминокислотный остаток основного характера заменен нейтральным гидрофильным аминокислотным остатком и/или аминокислотным остатком кислотного характера или их комбинацией.
Четвертый аспект, описанный в данном документе, заключается в том, что было обнаружено, что путем корректировки длины и соединения линкерного пептида можно улучшить рекомбинантную продукцию гибридного полипептида в эукариотической клетке, такой как, например, клетка млекопитающего.
Таким образом, один из аспектов, описанных в данном документе, представляет собой способ получения (рекомбинантного) гибридного полипептида, включающий следующие этапы:
- культивирование эукариотической клетки (в одном из воплощений клетки млекопитающего), содержащей нуклеиновую кислоту, кодирующую гибридный полипептид,
- извлечение (рекомбинантного) гибридного полипептида из клетки или культивационной среды и, таким образом, получение (рекомбинантного) гибридного полипептида.
В одном воплощении одна часть гибридного полипептида представляет собой нейротрофин, а другая часть гибридного полипептида представляет собой антитело или фрагмент антитела.
Пятый аспект, описанный в данном документе, заключается в том, что было обнаружено, что
i) путем введения одного или более чем одного сайта гликозилирования, и/или
ii) путем замены в прополипептиде эндогенного сайта расщепления протеазой между просегментом и полипептидом на экзогенный сайт (относительно происхождения частей гибридного полипептида) или искусственный сайт расщепления протеазой, и/или
iii) путем понижения изоэлектрической точки полипептида, и/или
iv) путем корректировки длины, соединения и заряда линкерного пептида
можно улучшить рекомбинантную продукцию полипептида в клетке млекопитающего.
Таким образом, один из аспектов, описанных в данном документе, представляет собой способ получения (рекомбинантного) (гибридного) полипептида, включающий следующие этапы:
- культивирование эукариотической клетки (в одном из воплощений клетки млекопитающего), содержащей нуклеиновую кислоту, кодирующую вариантный (гибридный) полипептид, где аминокислотная последовательность (гибридного) полипептида была изменена
i) путем введения одного или более чем одного искусственного сайта гликозилирования, и/или
ii) путем замены в про(гибридном)полипептиде эндогенного сайта расщепления протеазой между просегментом и (гибридным) полипептидом на экзогенный сайт (относительно происхождения частей гибридного полипептида) или искусственный сайт расщепления протеазой, и/или
iii) путем понижения изоэлектрической точки (гибридного) полипептида, и/или
iv) путем корректировки длины линкерного пептида, корректировки соединения линкера, корректировки заряда линкера, введения одной или более чем одной мутации поверхностно расположенных аминокислотных остатков, дающих более низкую изоэлектрическую точку (гибридного) полипептида,
- извлечение (гибридного) полипептида или (гибридного) прополипептида из клетки или культивационной среды и, таким образом, получение (рекомбинантного) (гибридного) полипептида.
Один из аспектов, описанных в данном документе, представляет собой способ получения (рекомбинантного) гибридного полипептида, включающий следующие этапы:
- получение нуклеиновой кислоты, кодирующей гибридный полипептид,
- модификация нуклеиновой кислоты, чтобы она кодировала вариантный гибридный полипептид, в котором аминокислотная последовательность гибридного полипептида была изменена путем корректировки длины линкерного пептида, корректировки соединения линкера, корректировки заряда линкера, введения одной или более чем одной мутации поверхностно расположенных аминокислотных остатков, дающих более низкую изоэлектрическую точку гибридного полипептида,
- введение нуклеиновой кислоты в эукариотическую клетку (в одном из воплощений в клетку млекопитающего),
- культивирование эукариотической клетки, и
- извлечение (рекомбинантного) вариантного гибридного полипептида из клетки или культивационной среды и, таким образом, получение (рекомбинантного) гибридного полипептида.
В одном из воплощений предыдущих аспектов гибридный полипептид содержит биологически активную группу, линкерный пептид и одновалентную связывающую группу, которая связывается с рецептором гематоэнцефалического барьера (от англ. blood-brain-barrier, ВВВ).
В одном из воплощений рецептор гематоэнцефалического барьера выбран из группы, включающей рецептор трансферрина, рецептор инсулина, рецептор инсулиноподобного фактора роста, белок, связанный с рецептором липопротеина низкой плотности (LRP)/рецептор альфа-2-макроглобулина, белок 8, связанный с рецептором липопротеина низкой плотности (также известный как рецептор 2 аполипопротеина Е (ApoER2)), белок 1, связанный с рецептором липопротеина низкой плотности (также известный как рецептор альфа-2-макроглобулина (A2MR)), и гепарин-связывающий фактор роста, подобный эпидермальному фактору роста.
В одном из воплощений линкерный пептид содержит один или более чем один отрицательно заряженный аминокислотный остаток. В одном из воплощений линкерный пептид содержит два или более двух отрицательно заряженных аминокислотных остатков. В одном из воплощений линкерный пептид содержит два, или три, или четыре, или пять отрицательно заряженных аминокислотных остатков.
В одном из воплощений биологически активная группа представляет собой нейротрофический фактор. В одном из воплощений нейротрофический фактор представляет собой нейротрофический фактор головного мозга (brain derived neurotrophic factor, BDNF).
В одном из воплощений одновалентная связывающая группа связывается с рецептором гематоэнцефалического барьера и представляет собой одновалентный фрагмент антитела, предпочтительно, выбранный среди scFv, Fv, scFab, Fab, VHH.
В одном из воплощений гибридный полипептид представляет собой одноцепочечный гибридный полипептид, содержащий в качестве первой части человеческий нейротрофический фактор головного мозга, а в качестве второй части отдельный Fab- или scFv-фрагмент антитела против рецептора трансферрина, которые конъюгированы друг с другом либо напрямую, либо через линкерный пептид.
Один из аспектов, описанных в данном документе, представляет собой гибридный полипептид, включающий
- ровно один полипептид нейротрофического фактора дикого типа, или вариантный полипептид нейротрофического фактора, или его фрагмент с активностью нейротрофического фактора,
- связывающий домен, и
- линкерный пептид между полипептидом нейротрофического фактора и фрагментом антитела.
Один из аспектов, описанных в данном документе, представляет собой гибридный полипептид с активностью нейротрофического фактора (например, димерный комплекс), включающий
- ровно один тип полипептида нейротрофического фактора дикого типа, или вариантного полипептида нейротрофического фактора, или его фрагмента с активностью нейротрофического фактора,
- связывающий домен, и
- линкерный пептид между полипептидом нейротрофического фактора и фрагментом антитела.
В одном из воплощений связывающий домен представляет собой сайт связывания фрагмента антитела. В одном из воплощений связывающий домен представляет собой фрагмент антитела, содержащий сайт связывания антитела. В одном из воплощений фрагмент антитела специфически связывается с рецептором гематоэнцефалического барьера. В одном из воплощений фрагмент антитела представляет собой одновалентный фрагмент антитела, выбранный из группы, включающей scFv, Fv, scFab, Fab, VHH.
В одном из воплощений рецептор гематоэнцефалического барьера выбран из группы, включающей рецептор трансферрина, рецептор инсулина, рецептор инсулиноподобного фактора роста, белок, связанный с рецептором липопротеина низкой плотности (LRP)/рецептор альфа-2-макроглобулина, белок 8, связанный с рецептором липопротеина низкой плотности (также известный как рецептор 2 аполипопротеина Е (ApoER2)), белок 1, связанный с рецептором липопротеина низкой плотности (также известный как рецептор альфа-2-макроглобулина (A2MR)), и гепарин-связывающий фактор роста, подобный эпидермальному фактору роста.
В одном из воплощений гибридный полипептид содержит вторую одновалентную или двухвалентную связывающую группу, которая не связывается с рецептором ВВВ.
Один из аспектов, описанных в данном документе, представляет собой комплекс, содержащий
- в качестве первого компонента гибридный полипептид, описанный в данном документе, и
- в качестве второго компонента полипептид нейротрофического фактора дикого типа, или вариантный полипептид нейротрофического фактора, или его фрагмент с активностью нейротрофического фактора.
Один из аспектов, описанных в данном документе, представляет собой многомерный комплекс с активностью нейротрофического фактора, включающий
- в качестве первого компонента гибридный полипептид, описанный в данном документе, и
- в качестве второго компонента полипептид нейротрофического фактора дикого типа, или вариантный полипептид нейротрофического фактора, или его фрагмент с активностью нейротрофического фактора.
В одном из воплощений указанный комплекс представляет собой димерный комплекс.
В одном из воплощений всех аспектов нейротрофический фактор выбран среди фактора роста нервов (NGF), нейротрофического фактора головного мозга (BDNF), нейротрофического фактора глиальной клеточной линии (GDNF), нейротрофина 3 (NT-3) и нейротрофина 4 (NT-4). В одном предпочтительном воплощении нейротрофический фактор роста представляет собой BDNF.
Все аминокислотные последовательности, представленные в данном документе, являются конкретными аспектами данного изобретения.
Подробное описание изобретения
Причина (причины) низкого экспрессионного выхода рекомбинантно полученных полипептидов в клетках млекопитающих часто не известна и, как правило, трудно определима.
Некоторые полипептиды могут мешать функциям клетки-хозяина при экспрессии. Это может быть, например, неправильная сортировка в неправильное клеточное местоположение или экспрессия в неправильном пространственном (например, тип клеток) и/или временном (например, при зависимости от клеточного цикла) контексте. Кроме того, увеличение рекомбинантной экспрессии и секреции полипептидов может привести к неблагоприятным условиям сворачивания белка, например, приводящим к агрегации белка и/или клеточным стрессовым реакциям. В заключение, рекомбинантная экспрессия полипептидов может быть затрудненной (например, давая низкий экспрессионный выход) или даже невозможной в конкретной клетке-хозяине.
Способы, приведенные в данном документе, являются иллюстративными в отношении конкретных полипептидов, гибридных полипептидов и полипептидов, слитых с антителами. Эти молекулы используются только в качестве примера и не должны быть истолкованы как ограничивающие объем данного изобретения.
Нейротрофические белки вызывают высокий терапевтический интерес. Например, нейротрофический фактор головного мозга (BDNF) предлагается использовать для лечения или облегчения симптомов болезни Альцгеймера, Паркинсона, Хантингтона и перинатальной болезни белого вещества, болезни Дауна и аутизма/синдрома Ретта, шизофрении, депрессии, нарушения пищевого поведения, бокового амиотрофического склероза, рассеянного склероза, нервных повреждений, например, спинного мозга, а также других заболеваний (Apfel, S.C., Clin. Chem. Lab Med. 39 (2001) 351-355; Nagahara, A.H. and Tuszynski, M.H., Nat. Rev. Drug Discov. 10 (2011) 209-219; Zuccato, C. and Cattaneo, E., Nature Reviews Neurology 5 (2009) 311-322; Thoenen, H. and Sendtner, M., Nature Neuroscience Supplement 5 (2002) 1046-1050; Gharami, K., et al., J. Neurochem. 105 (2008) 369-379).
В человеческом организме нейротрофические белки находятся на очень низком уровне и часто экспрессируются специализированными типами клеток, расположенными в очень сложной микросреде (см., например, Greenberg, М.Е., et al., J. Neurosci. 29 (2009) 12764-12767). В соответствии с их природными функциями уровни экспрессии нейротрофинов в эукариотических клетках, обычно используемых для рекомбинантной промышленной продукции таких полипептидов (например, клетках СНО, клетках COS, клетках NSO и клетках НЕК), являются очень низкими (Acklin, С. et al., Int. J. Pept. Protein Res. 41 (1993) 548-552).
I. Повышение экспрессии путем введения сайтов N-гликозилирования
Одним из аспектов, описанных в данном документе, является введение одного или более чем одного (искусственного) сайта гликозилирования в полипептид для его рекомбинантной продукции. Путем ведения одного или более чем одного (искусственного) сайта гликозилирования может быть улучшена рекомбинантная продукция полипептида в эукариотической клетке, особенно в клетке млекопитающего. Этот способ особенно подходит, например, для полипептидов, которые слабо или совсем не экспрессируются/секретируются в клетках млекопитающих. Введенный сайт гликозилирования может находиться либо в самом полипептиде, либо в линкерном пептиде, соединяющем два полипептида гибридного полипептида, либо он может быть меткой специфического гликозилирования. Особенно предпочтительно введение сайта гликозилирования путем точечной мутации поверхностно локализованной аминокислоты для создания (искусственного) мотива N-гликозилирования.
Пример: Гибридные полипептиды, содержащие часть антитела и GFP
Были сконструированы иллюстративные гибридные полипептиды, содержащие антитело и группировку GFP.
GlySer-линкер (GGGGSGGGGSG; SEQ ID NO 01) был использован для слияния либо i) группировки eGFP (усиленный зеленый флуоресцентный белок, последовательность SEQ ID NO 02), либо ii) группировки emGFP (зеленый флуоресцентный белок Emerald; SEQ ID NO 03), либо iii) группировки tagGFP (SEQ ID NO 04) с С-концами тяжелых цепей (НС) анти-IGF-1R-антитела подкласса IgG1 (НС: SEQ ID NO 05; LC: SEQ ID NO 06). Различные гибриды тяжелой цепи имеют аминокислотную последовательность SEQ ID NO 07, 08 и 09.
Секреция гибридных полипептидов-антител не могла быть выявлена во всех клеточных культуральных супернатантах временно трансфицированных HEK293-клеток с использованием вестерн-блота и/или высокоэффективной жидкостной хроматографии (HPLC) на основе белка А. Также биологически активный GFP нельзя было контролировать за счет его биолюминесцентных свойств (GFP-специфическая флуоресценция). Но гибридные полипептиды могли быть обнаружены во фракции клеточного осадка путем вестерн-блота. Результаты приведены в следующей таблице.

Без привязки к этой теории GFP является цитоплазматическим мономерным белком с тенденцией к формированию (слабых) димеров. Таким образом, GFP дикого типа "от природы" (природная клетка медузы Aequorea Victoria) не предназначен для секреции, и, следовательно, молекула GFP должна быть сделана подходящей для секреции из-за несовместимости с системой секреции клеток млекопитающих.
Для того чтобы получить секретируемые GFP-содержащие гибридные полипептиды, в гибридный полипептид также был включен полипептид, полученный из человеческого светочувствительного мембраносвязанного рецептора опсина, связанного с G-белком, найденного в фоторецепторных клетках сетчатки, например, в качестве метки гликозилирования. С помощью этой метки гликозилирования в гибридный полипептид вводят дополнительные (искусственные) сайты N-гликозилирования.
Полипептид, полученный из рецептора опсина, был слит непосредственно, т.е. без промежуточного линкерного пептида, с С-концом GFP-группировки. Полипептид, полученный из опсина (NGTEGPNFYVPFSNATGVV; опсин-метка, SEQ ID NO 10), содержит два мотива сайта N-гликозилирования: мотив NGT и мотив NAT (обычный мотив сайта N-гликозилирования: N×S/T; Asn с последующей любой аминокислотой, за исключением Pro, а затем Ser или Thr). Временная экспрессия гибридных полипептидов, содержащих опсиновую метку гликозилирования (SEQ ID NO 11), в клетках HEK293 приводит к секреции гибридного полипептида. Результаты приведены в следующей таблице.

Из SDS-PAGE-анализа (размывание полос) можно видеть, что секретируемые гибридные полипептиды содержат дополнительные углеводы.
Гибридные полипептиды "антитело-GFP-опсиновая метка" очищали путем двухэтапной процедуры, в частности, аффинной хроматографии на белке А с последующей эксклюзионной хроматографией. Функциональность группировки антитела в гибридном полипептиде была продемонстрирована путем связывания с рецепторным белком IGF-1R с использованием поверхностного плазмонного резонанса (BIAcore) и интернализации в клетки посредством рецептор-опосредованного эндоцитоза в клетках, сверхэкспресирующих IGF-1R, с помощью FACS и/или конфокальной микроскопии. Функциональность GFP-группировки была показана благодаря ее характеристикам зеленой флуоресценции.
Пример: Нейротрофические белки
Иллюстративным нейротрофическим белком является нейротрофический фактор головного мозга (BDNF).
i) Метка гликозилирования
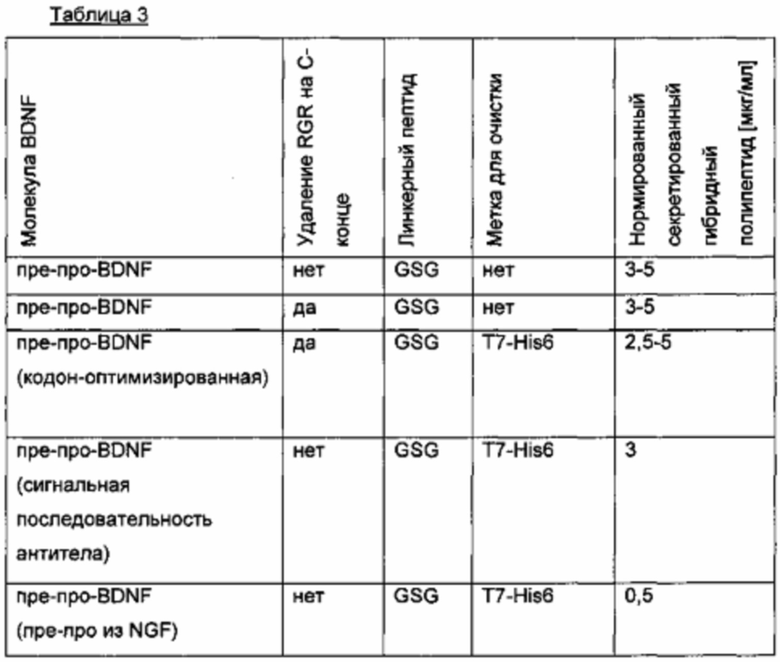
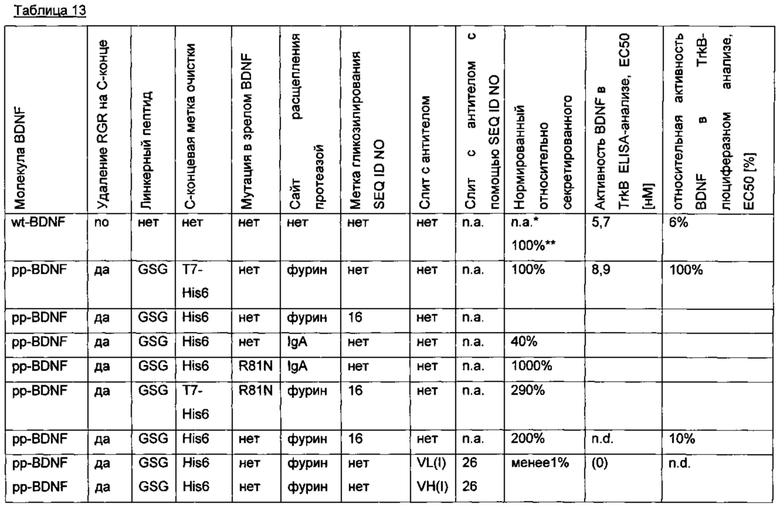
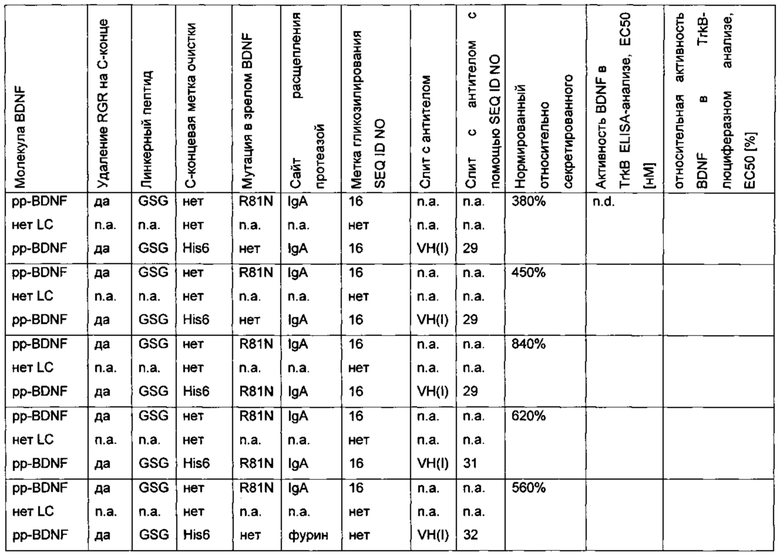
Человеческий пре-про-BDNF дикого типа, содержащий С-концевую Т7 His6-метку (MASMTGGQQMG-HHHHHH, используется для аффинной очистки; SEQ ID NO 12), слитую через GSG-линкер, экспрессируют в клетках HEK239 (пре-про-BDNF-T7-His6; SEQ ID NO 13). Аминокислотная последовательность зрелого BDNF не содержит мотив N-гликозилирования и, следовательно, не является N-гликозилированным. Зрелый BDNF получают с низким выходом, который составляет лишь несколько мкг/мл. Низкий выход экспрессии гена не может быть улучшен путем оптимизации кодонов гена, или путем удаления потенциальных сайтов расщепления протеазой (пре-про-BDNF(-RGR)-T7-His6), или путем замены нативной сигнальной последовательности BDNF сигнальной последовательностью хорошо экспрессируемого антитела (MGWSCIILFL VATATGVHS; SEQ ID NO 14), или путем замены препросегмента BDNF таковым из NGF. Результаты приведены в следующей таблице (значения нормированы по концентрации зрелого BDNF).
Для того чтобы улучшить (секреторный) выход BDNF, в молекулу BDNF были введены (искусственные) сайты гликозилирования с использованием опсиновых меток гликозилирования различной длины: 16 аминокислотных остатков (NGTEGPNFYVPFSNAT; SEQ ID NO 15), 19 аминокислотных остатков (NGTEGPNFYVPFSNATGVV; SEQ ID NO 10) и 20 аминокислотных остатков (NGTEGPNFYVPFSNATGVVR; SEQ ID NO 16). С помощью этой модификации экспрессионный выход (нормированный по концентрации зрелого BDNF) может быть улучшен. Результаты приведены в следующей таблице (нормированы по концентрации зрелого BDNF).

Неожиданно варианты BDNF, содержащие метку гликозилирования, также имели улучшенную биологическую активность по сравнению с нативным негликозилированным BDNF (CHO-TrkB-люциферазный анализ с репортерным геном). Результаты приведены в следующей таблице.
Другими возможными сайтами гликозилирования являются AAANGTGGA (один мотив сайта N-гликозилирования; SEQ ID NO 17), ANITVNITV (два мотива сайта N-гликозилирования; SEQ ID NO 18) и NATGADNGTGAS (два мотива сайта N-гликозилирования; SEQ ID NO 19).
ii) Введенный мотив(ы) сайта N-гликозилирования
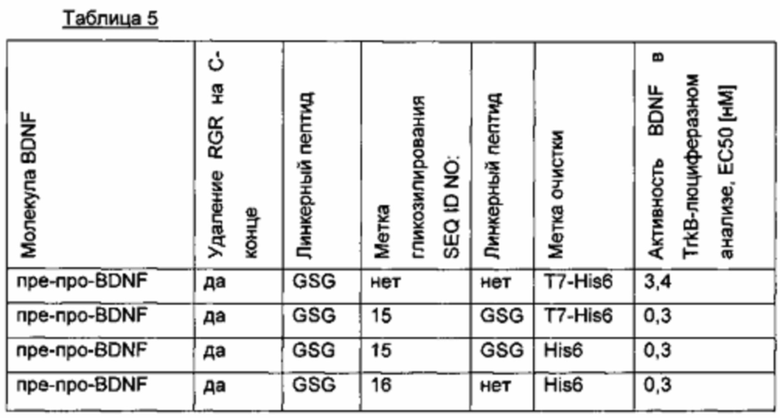
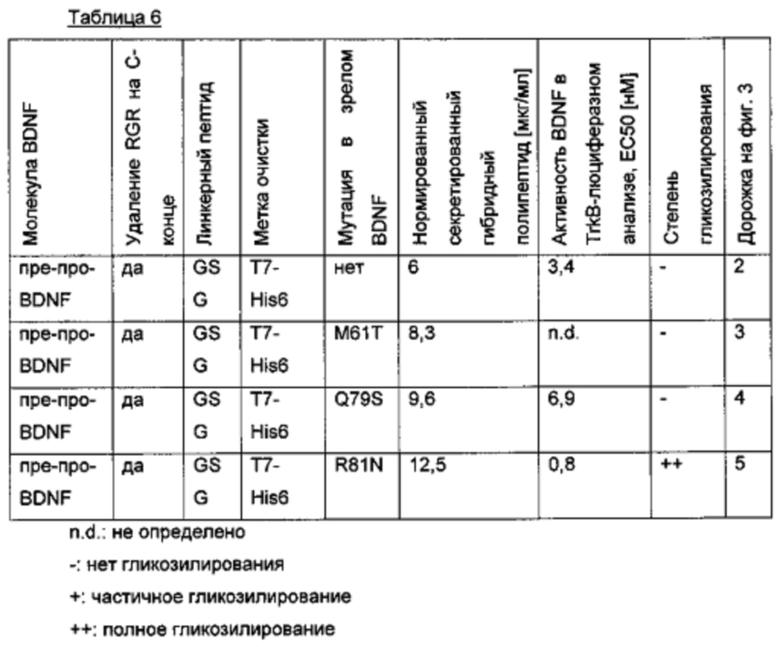
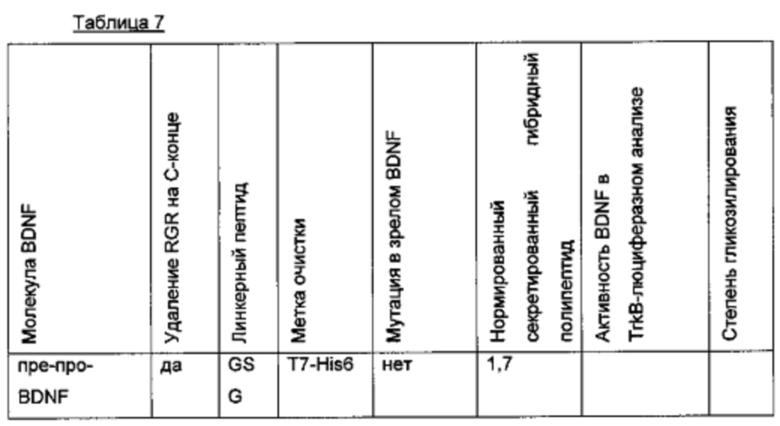
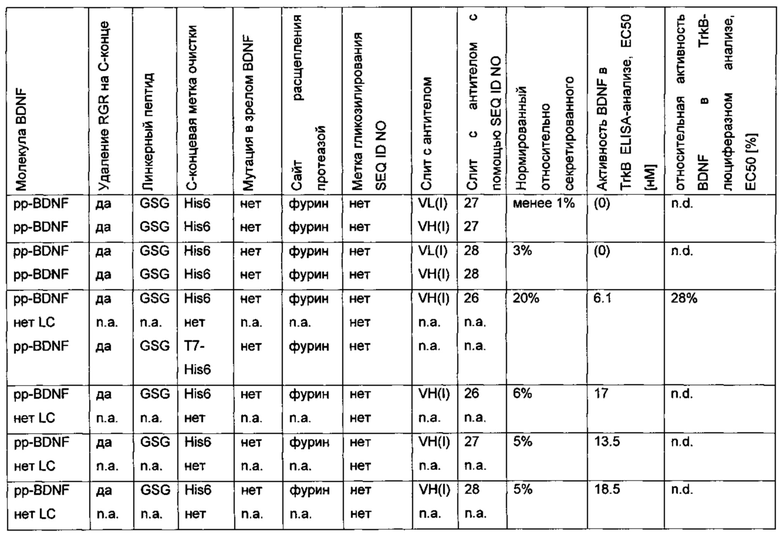
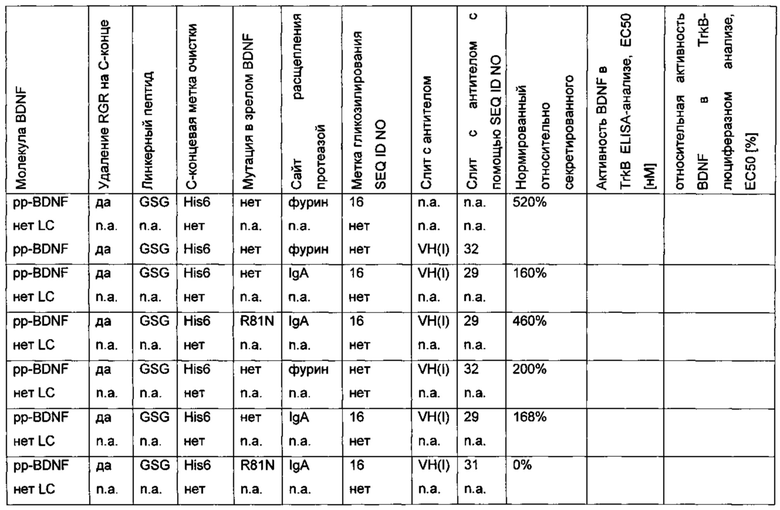
Для того чтобы улучшить (секреторный) выход BDNF, в аминокислотную последовательность BDNF были введены искусственные сайты N-гликозилирования. Искусственный сайт N-гликозилирования может быть введен, например, путем точечной мутации в аминокислотной последовательности BDNF (мотив сайта N-гликозилирования: Asn-Xxx-Ser/Thr; Ххх является любой аминокислотой кроме Pro). Соответствующие кодирующие нуклеиновокислотные последовательности были получены и временно экспрессированы в клетках HEK293. Нумерация мутаций основана на аминокислотной последовательности зрелого BDNF (SEQ ID NO 25). Для секретированных вариантов BDNF анализировали уровень экспрессии/секреции, степень N-гликозилирования (выведенную из миграции полосы из-за увеличения MW примерно на 3-5 кДа на каждый введенный и имеющийся сайт N-гликозилирования (по сравнению с негликозилированным референсом BDNF) в анализе иммуноблоттинга) и функциональность/связывание рецептора с помощью анализа с репортерным геном CHO-TrkB-люциферазы. Результаты представлены в следующих таблицах (экспрессионный выход и биологические активности, нормированные по концентрации зрелого BDNF) и на фиг. 3.
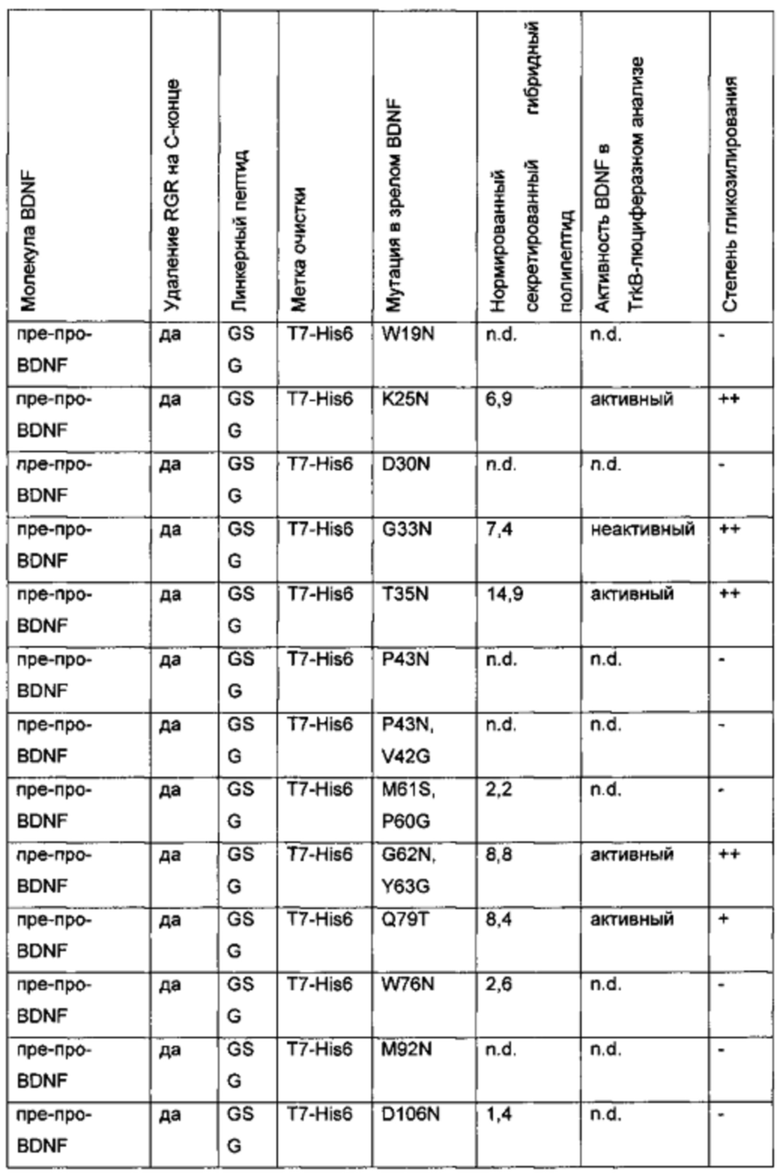

Активный: BDNF-активность в сравнении с активностью негликозилированного зрелого BDNF дикого типа, экспрессированного в Е.coli, подвергнутого рефолдингу и очищенного
Неактивный: никакой активности не выявлено
Функциональность/биологическая активность гликозилированных вариантов BDNF также была показана в анализе дорсальных корешковых ганглиев (dorsal root ganglion DRG). Активность, определенная в анализе DRG, была сравнима с негликозилированным BDNF дикого типа.
II. Повышение экспрессии путем модификации сайтов ферментативного расщепления
Часто секретируемые полипептиды синтезируются в виде препрополипептидов. Пресегмент представляет собой так называемую сигнальную последовательность. Просегмент может быть, например, необходим для сворачивания белка, клеточного нацеливания/транспортировки в конкретный клеточный компартмент (например, в лизосомы) или инактивации зрелого белка во время транспортировки и хранения в клетке или внеклеточной транспортировки (например, секреции в культуральную среду). Пре- и просегменты не являются необходимыми для биологической активности зрелого полипептида. Если просегмент не процессируется правильно, т.е. (пре-)просегмент не отщепляется от зрелого белка, например, в продуцирующей/секретирующей клетке, то он может изменять биофизические, биохимические и/или биологические активности зрелого полипептида.
Способ, представленный ниже, подходит для полипептидов, которые производятся in vivo из прополипептида путем ферментативного расщепления.
Один из аспектов, описанных в данном документе, представляет собой способ получения рекомбинантного полипептида из прополипептида с использованием вариантного прополипептида, включающий следующие этапы:
- культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую прополипептид (гибридный полипептид из просегмента и полипептида), в котором эндогенный сайт ферментативного расщепления между просегментом и полипептидом заменяется экзогенным сайтом расщепления протеазой,
- извлечение рекомбинантного вариантного прополипептида из клетки или культивационной среды и, таким образом, получение рекомбинантного полипептида.
Один из аспектов, описанных в данном документе, представляет собой способ получения гибридного полипептида из гибридного прополипептида, включающий следующие этапы:
- культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую вариантный гибридный прополипептид, в котором аминокислотная последовательность гибридного прополипептида была модифицирована путем замены в нативном гибридном прополипептиде эндогенного сайт расщепления протеазой между просегментом и гибридным полипептидом на экзогенный (относительно происхождения частей гибридного полипептида) или искусственный сайт расщепления протеазой,
- извлечение гибридного прополипептида из клетки или культивационной среды, отщепление гибридного прополипептида и, таким образом, получение (рекомбинантного) гибридного полипептида.
В одном из аспектов, описанных в данном документе, сайт расщепления протеазой между просегментом и зрелым полипептидом в прополипептиде заменяется экзогенным (относительно происхождения полипептида или частей гибридного полипептида) или искусственным сайтом расщепления протеазой.
Термин "эндогенный сайт расщепления протеазой», используемый в данном контексте, обозначает сайт расщепления протеазой, который в природе находится между просегментом и зрелым полипептидом.
Термин "экзогенный сайт расщепления протеазой», используемый в данном контексте, обозначает сайт расщепления протеазой, который в природе не обнаружен между просегментом и зрелым полипептидом.
При замене эндогенного сайта расщепления протеазой экзогенным сайтом расщепления протеазой процессинг и расщепление секретируемого прополипептида в его зрелой форме могут быть улучшены.
Экзогенный сайт расщепления протеазой может быть получен из любой протеазы, пока протеаза не присутствует/экспрессируется в клетке, из которой получен прополипептид, т.е. в которой прополипептид встречается в природе (частично или полностью).
Экзогенный сайт расщепления протеазой (по отношению к происхождению полипептида) может представлять собой любой экзогенный сайт расщепления протеазой, такой как, например, сайт расщепления IgA-протеазой, сайт расщепления протеазой TEV (от англ. tobacco etch virus - вирус табачной мозаики), сайт расщепления гранзимом В, сайт расщепления тромбином, сайт расщепления фактором 10, сайт расщепления энтерокиназой, сайт расщепления субтилизином, сайт расщепления катепсином, сайт расщепления металлопротеиназой, сайт расщепления протеазой IDES (от англ. IgG-degrading enzyme of Streptococcus pyogenes - фермент Streptococcus pyogenes, расщепляющий IgG), сайт расщепления протеазой PreScission или их функциональные варианты.
Расщепление прополипептида может происходить в различные моменты времени во время рекомбинантной продукции полипептида.
Расщепление прополипептида может происходить в культивационной среде. При этом экзогенную протеазу (обеспечивающую расщепление прополипептида) добавляют в культивационную среду, либо экзогенная протеаза (обеспечивающая расщепление прополипептида) коэкспрессируется в культивационной среде.
Кроме того, можно расщеплять прополипептид после его выделения из клеток, например, до, во время или после последующей обработки (но после культивирования).
В одном предпочтительном воплощении экзогенный сайт расщепления протеазой получен из другого организма, отличающегося от источника прополипептида и от рекомбинантной клетки, экспрессирующей этот прополипептид. Преимущество этого заключается в том, что можно определить точку процессинга прополипептида. Если клетка, экспрессирующая вариантный прополипептид, не коэкспрессирует экзогенную протеазу, и экзогенная протеаза не добавляется в культивационную среду, то расщепление уже не может быть осуществлено клеткой, экспрессирующей вариантный прополипептид, или во время культивирования, соответственно.
В одном воплощении расщепление прополипептида осуществляют во время очистки.
В одном воплощении расщепление прополипептида осуществляют на колонке во время процесса очистки. В одном конкретном воплощении колонна представляет собой аффинную колонку.
В одном предпочтительном воплощении расщепление прополипептида осуществляют путем инкубирования прополипептида с протеазой после того, как прополипептид был отделен от культивационной среды. В одном воплощении инкубацию осуществляют после первого этапа (хроматографической) очистки. В одном воплощении инкубацию осуществляют на колонке.
Эта методика также позволяет, например, включать в просегмент искусственную метку очистки, такую как, например, His6-метка, myc-метка, НА-метка или биотин/авидин-метка, для улучшения/упрощения очистки.
В одном воплощении прополипептид содержит просегмент и зрелый полипептид, где просегмент содержит метку очистки. В одном воплощении метка очистки выбрана из группы меток очистки, включающей His6-метку, myc-метку, НА-метку и биотин/авидин-метку.
В одном воплощении расщепление прополипептида, включающего метку очистки, осуществляют после этапа очистки с использованием метки очистки.
В любом случае отщепление просегмента от зрелого полипептида осуществляют перед введением зрелого полипептида пациенту. В одном воплощении расщепление осуществляют in vitro.
В одном предпочтительном воплощении эндогенный сайт расщепления протеазой представляет собой сайт расщепления IgA-протеазой, а метка очистки представляет собой гексагистидиновую (His6) метку. В одном воплощении гексагистидиновую метку конъюгируют через пептид GSG с сайтом расщепления протеазой.
Пример: пре-про-BDNF-вариант с экзогенным сайтом расщепления протеазой в прополипептиде BDNF
Примером полипептида, который экспрессируется в виде (пре)прополипептида и расщепляется в зрелую форму, является BDNF.
Человеческий BDNF дикого типа экспрессировали в клетках HEK293. Во время процессинга полипептида (т.е. во время экспрессии/рекомбинантной продукции) просегмент только частично удалялся под действием протеаз клетки-хозяина, что приводило к образованию смеси зрелого BDNF и непроцессированного про-BDNF. Человеческий про-BDNF процессировался во время секреции в клетке-хозяине, главным образом, с помощью фурина, секреторной конвертазы пробелков.
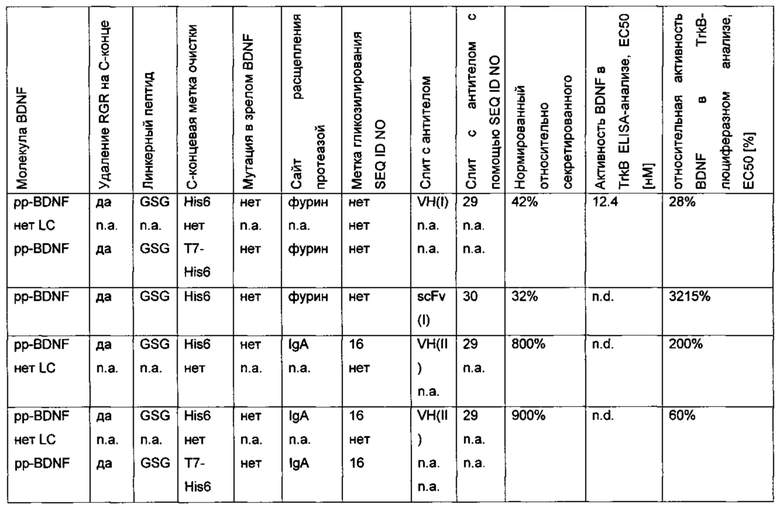
Был сконструирован про-BDNF-вариант, в котором природный сайт расщепления протеазы фурина был заменен экзогенным сайтом расщепления IgA-протеазой. IgA-протеаза распознает и расщепляет белки, содержащие аминокислотную последовательность N-X-Z-Pro-Pro/-Y-Pro-C (X - предпочтительно Pro или Ser; Y - Thr, Ser или Ala; Z - предпочтительно Arg или Thr). Аминокислотная последовательность про-BDNF-полипептида содержит сайт расщепления фурина (RVRR; SEQ ID NO 21). Сайт расщепления фурина был заменен сайтом расщепления IgA-протеазой (GSVVAPPAP; SEQ ID NO 22). Кроме того, через линкерный пептид GSG с С-концом BDNF был слит His6-сайт (НННННН, используется для аффинной очистки; SEQ ID NO 23). Также был удален потенциальный сайт расщепления протеазой (удаление С-концевой аминокислотной последовательности RGR; SEQ ID NO 24). В сравнительных вариантах мутация R54A была введена в прополипептид BDNF (пре-про-BDNF-полипептид имеет в аминокислотной позиции 54 аминокислотный остаток Ala вместо Arg), и/или вместо His6-метки использовали T7-His6-метку. Результаты приведены в следующей таблице (биологическая активность нормирована по концентрации зрелого BDNF).
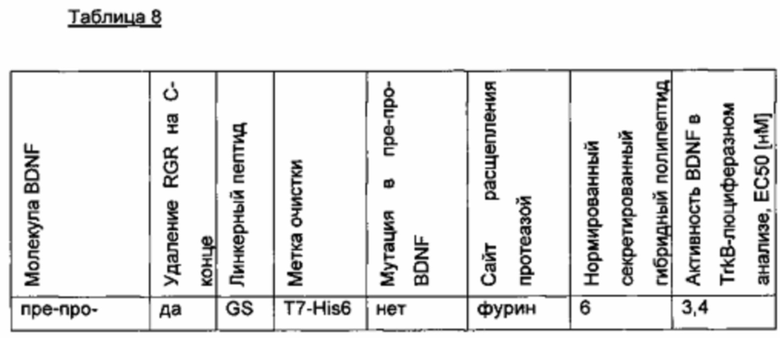
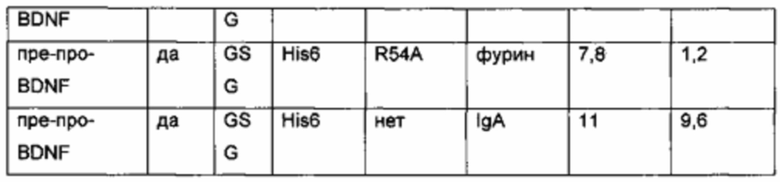
Нумерация мутации R54A основана на аминокислотной последовательности пре-про-BDNF дикого типа (SEQ ID NO 20).
Сконструированный пре-про-BDNF-полипептид экспрессировали в клетках HEK293 с высоким выходом. Секретированный сконструированный про-BDNF-полипептид эффективно превращался in vitro в зрелый нативный BDNF путем расщепления IgA-протеазой.
Таким образом, в отличие от экспрессии нативного пре-про-BDNF-полипептида, содержащего природный сайт расщепления фурина/конвертазы пропептида, сконструированный про-BDNF полипептид, включающий экзогенный сайт расщепления IgA-протеазой, был получен в виде одного продукта экспрессии. Кроме того, были получены улучшенные экспрессионные выходы. Кроме того, зрелый BDNF-полипептид, полученный из сконструированного про-BDNF-полипептида, обладает повышенной биологической активностью, как зрелый BDNF-полипептид, полученный из пре-про-BDNF дикого типа, в клеточном анализе с репортерным геном люциферазы TrkB.
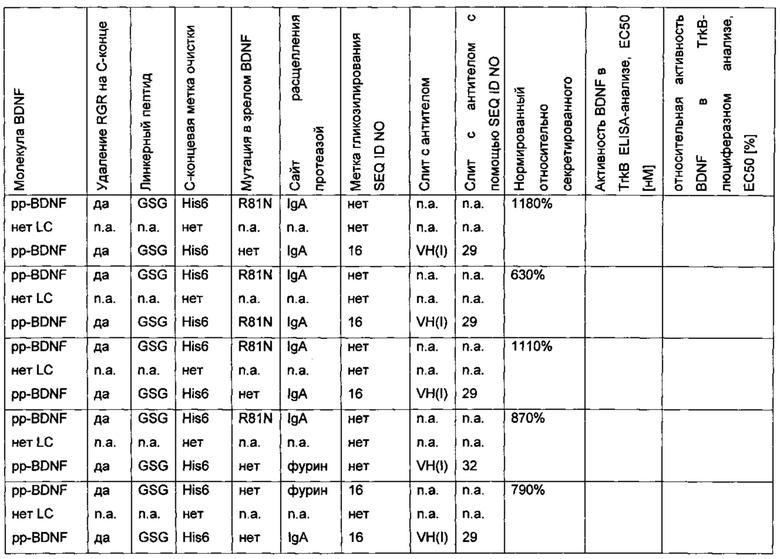
Пример: пре-про-BDNF-вариант с экзогенным сайтом расщепления протеазой и сконструированной меткой очистки в прополипептиде BDNF
Метки аффинности очень полезны для простой и эффективной очистки рекомбинантно полученных полипептидов. Тем не менее, искусственные метки очистки, которые остаются в конечном терапевтическом белке, неприемлемы для клинического применения по нескольким причинам, в том числе из-за потенциальной иммуногенности, изменений в биофизических и биохимических свойствах, а также биологической активности. Чтобы преодолеть эти ограничения, полезной является удаляемая метка очистки.
В одном воплощении (BDNF) прополипептид включает метку очистки.
Был сконструирован про-BDNF-вариант, в котором в прополипептид введена His6-метка. Кроме того, природный сайт расщепления фурином был заменен экзогенным сайтом расщепления IgA-протеазой, и/или в прополипептид была введена мутация R54A.
В сравнительном варианте His6-метка была слита через линкерный пептид GSG с С-концом BDNF. Кроме того, вместо His6-метки была использована T7-His6-метка. Результаты приведены в следующей таблице (биологическая активность нормирована по концентрации зрелого BDNF) и на фиг. 2.
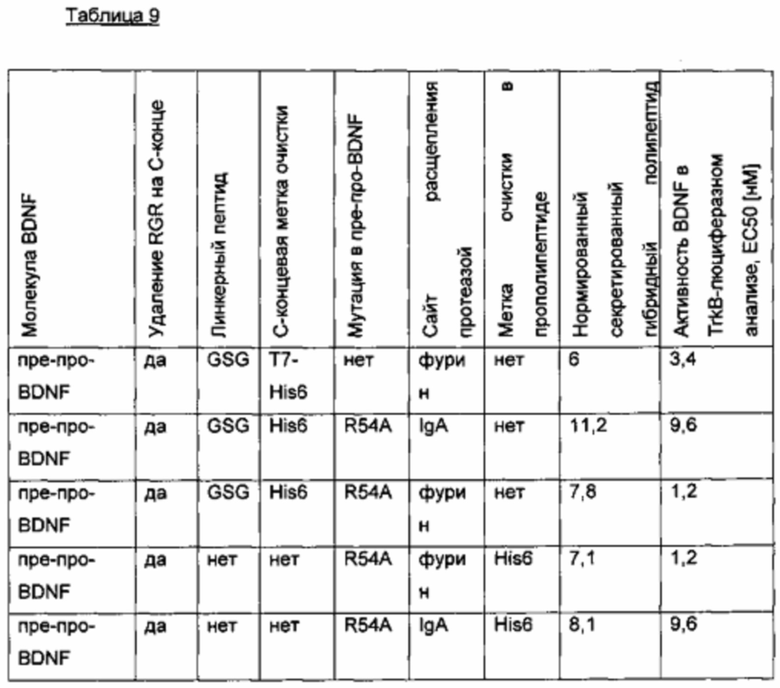
Биологическую активность/функциональность полипептидов BDNF, полученных из различных вариантных про-BDNF-полипептидов, подтверждали в анализах in vitro и in vivo. Конструкции, содержащие сайт расщепления IgA-протеазой, показали улучшенную активность.
Удаляемая метка очистки (например, такая как гекса-гистидиновая метка), вставленная в N-концевую область пропептида, может быть использована для эффективного очищения, и будет отщепляется во время созревания белка in vitro. Таким образом, никакие потенциально иммуногенные пептидные последовательности не будут сохранены в зрелом полипептиде, который будет использоваться для введения in vivo.
III. Улучшение экспрессии путем снижения изоэлектрической точки
Некоторые полипептиды, например, нейротрофины, с основной изоэлектрической точкой (IEP), т.е. изоэлектрической точкой выше 9, имеют тенденцию к образованию агрегатов.
В одном из аспектов, описанных в данном документе, снижение изоэлектрической точки полипептида используется для увеличения выхода рекомбинантно продуцируемого полипептида с использованием клеток млекопитающих. Этот способ особенно подходит для полипептидов, имеющих изоэлектрическую точку выше 9 (высокая изоэлектрическая точка, основная изоэлектрическая точка), таких как, например, нейротрофины. Снижение (уменьшение) изоэлектрической точки может быть достигнуто за счет увеличения числа отрицательных зарядов в полипептиде, например, путем введением отрицательно заряженных аминокислотных остатков и/или слияния с отрицательно заряженной группировкой, такой как линкерный пептид. В качестве альтернативы, снижение IEP может быть достигнуто путем удаления положительных зарядов с поверхности полипептида, например, путем введения отрицательно заряженных аминокислотных остатков на поверхность полипептида. Это может быть сделано, например, путем замены одного или более чем одного основного аминокислотного остатка (остатков) остатком нейтральной гидрофильной аминокислоты и/или остатком кислой аминокислоты или их комбинацией.
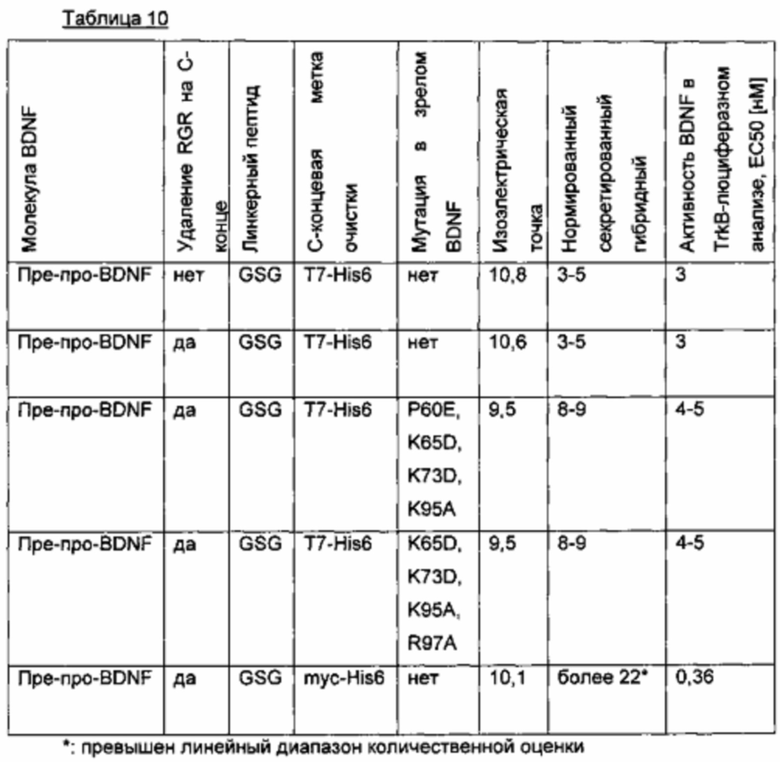
Пример: варианты BDNF со сниженной изоэлектрической точкой
BDNF имеет изоэлектрическую точку примерно 10 и представляет собой липкую основную/щелочную молекулу (см., например, Leibrock, J., et al., Nature 341 (1989), 149-152).
Для того чтобы улучшить (секреторный) выход, были сконструированы варианты BDNF, в которых аминокислотные остатки были изменены таки образом, чтобы понизить изоэлектрическую точку молекулу. Кроме того, с С-концом BDNF сливали T7-His6-метку через линкерный пептид GSG. Кроме того, в одном сравнительном варианте удаляли потенциальный сайт расщепления протеазой (удаление С-концевой аминокислотной последовательности RGR; SEQ ID NO 23). В первом варианте были введены следующие аминокислотные мутации: Р60Е, K65D, K73D и K95A. Во втором варианте были введены следующие аминокислотные мутации: K65D, K73D, K95A и R97A. Значения IEP рассчитывали с использованием способа EMBOSS IEP (Alan Bleasby (ajb © ebi.ac.uk); European Bioinformatics Institute, Wellcome Trust Genome Campus, Hinxton, Cambridge CB10 1SD, UK). Результаты приведены в следующей таблице (биологические активности нормированы по концентрации зрелого BDNF).
В отличие от BDNF дикого типа сконструированные варианты BDNF с пониженной IEP стабильно накапливались в течение 3-6 дней в клеточной культуральной среде/супернатанте во временно трансфицированных клетках HEK293. Это также было показано с помощью исследования культивирования клеток, в котором очищенный зрелый BDNF (полученный в Е.coli) и очищенный myc-меченый вариант BDNF со сниженной IEP добавляли к растущей культуре клеток-хозяев HEK293 (концентрация BDNF, 10 мкг/мл). После 4 дней культивирования клеток определяли концентрацию оставшегося BDNF в клеточном культуральном супернатанте с помощью анализов SDS PAGE в восстанавливающих условиях и вестерн-блота: зрелый BDNF был полностью выведен из клеточного культурального супернатанта через 4 дня, в то время как myc-меченный вариант BDNF был почти полностью стабильным (см. фиг. 1).
IV. Повышение экспрессии путем сочетания различных модификаций
В одном из аспектов, приведенных в данном документе, для повышения рекомбинантной продукции полипептида была сделана/введена одна или более чем одна из следующих модификаций:
i) введение одного или более чем одного сайта гликозилирования, и/или
ii) замена эндогенного сайта расщепления протеазой между просегментом и зрелым полипептидом на экзогенный сайт (относительно происхождения полипептида) или искусственный сайт расщепления протеазой, и/или
iii) снижение изоэлектрической точки полипептида, и/или
iv) корректировка длины, соединения и заряда линкерного пептида.
Для дальнейшего повышения рекомбинантной продукции полипептида в некоторых воплощениях используются в комбинации любые две, любые три или все четыре модификации, описанные выше.
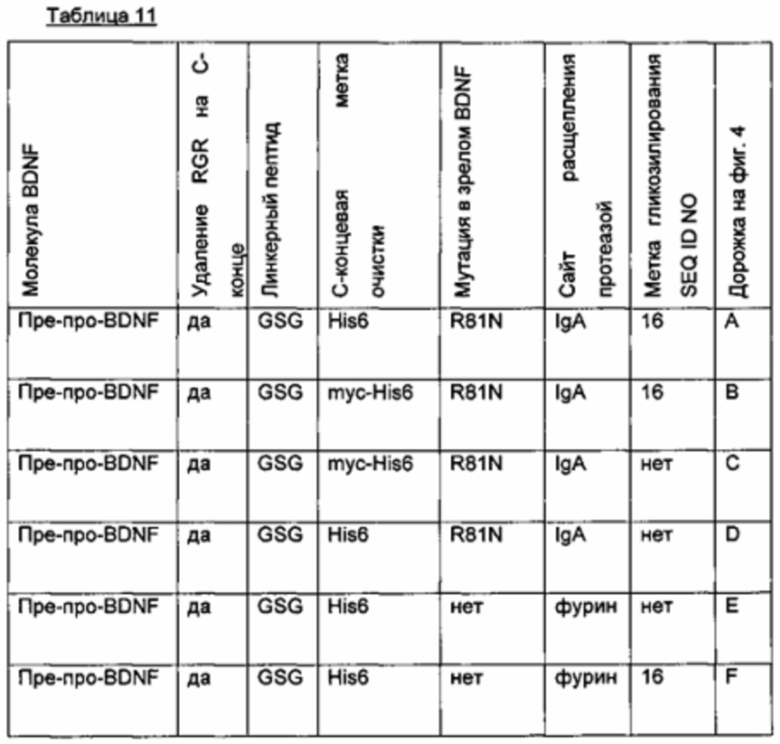
Пример: Нейротрофические белки
Иллюстративный нейротрофический белок представляет собой нейротрофический фактор головного мозга (BDNF).
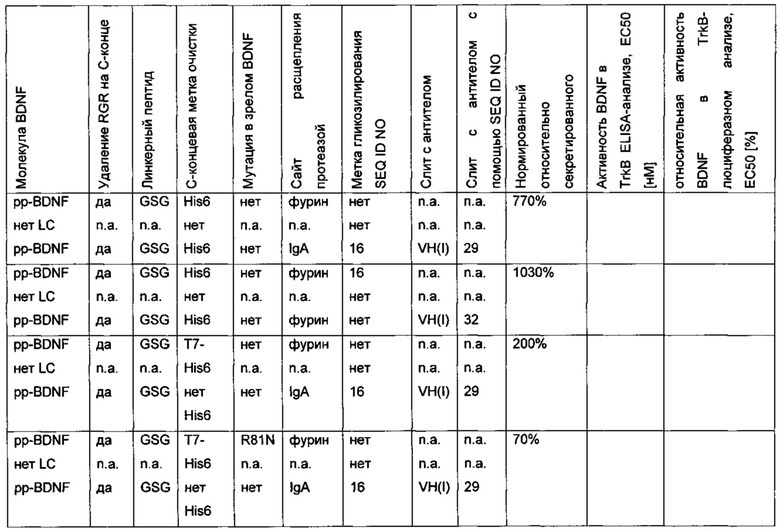
В аминокислотной последовательности BDNF были сделаны две или более двух из следующих модификаций:
- введение одного или более чем одного искусственного сайта гликозилирования,
- введение опсиновой метки в качестве метки N-гликозилирования, и
- замена эндогенного сайта расщепления фурина экзогенным сайтом расщепления IgA-протеазой.
Результаты представлены в следующих таблицах (биологические активности нормированы по концентрации зрелого BDNF).
На фиг. 4 можно видеть, что экспрессионный/секреторный выход вариантных полипептидов BDNF, содержащих один или более чем один дополнительный сайт N-гликозилирования, улучшается (дорожка А, В и F на фиг. 4) по сравнению с вариантами BDNF без дополнительного N-гликозилирования (дорожка С, D и Е на фиг. 4).

V. Повышение экспрессии гибридных полипептидов, транспортирующихся через гематоэнцефалический барьер
Проникновение крупных биотерапевтических препаратов в мозг строго ограничено обширным и непроницаемым гематоэнцефалическим барьером (ВВВ) совместно с другими клеточными компонентами в нейрососудистом комплексе (neurovascular unit, NVU). Для преодоления этого препятствия были испытаны многие стратегии, и одна из них заключается в использовании путей трансцитоза, опосредованных эндогенными рецепторами, экспрессированными в эндотелии капилляров мозга. Против этих рецепторов были сконструированы рекомбинантные белки, такие как моноклональные антитела или полипептиды, для обеспечения рецептор-опосредованной доставки биотерапевтических препаратов в мозг.
Биотерапевтические препараты имеют огромный терапевтический потенциал для лечения патологии центральной нервной системы (ЦНС, central nervous system, CNS). Тем не менее, их путь в мозг предотвращается гематоэнцефалическим барьером. Предыдущие исследования продемонстрировали, что очень небольшой процент (примерно 0,1%) IgG, введенного в кровоток, способен проникать в компартмент ЦНС (Felgenhauer, Klin. Wschr. 52 (1974) 1158-1164). Это, безусловно, ограничивает любой фармакологический эффект из-за низкой концентрации биотерапевтического препарата в ЦНС. Таким образом, носители, включающие фрагменты антител, которые опосредуют транспорт в мозг биотерапевтических препаратов, таких как нейротрофические факторы, имеют большое медицинское значение.
Таким образом, были сконструированы гибридные полипептиды, содержащие эффекторную группу и одновалентную или двухвалентную связывающую группу, которая связывается с рецептором ВВВ (см., например, ЕР 12182181.3).
В одном воплощении одновалентная связывающая группа, специфически связывающаяся с рецептором гематоэнцефалического барьера, или одновалентный фрагмент антитела, специфически связывающийся с рецептором гематоэнцефалического барьера, предпочтительно выбраны среди scFv, Fv, scFab, Fab и VHH.
В одном воплощении эффекторная группа представляет собой нейротрофический полипептид, такой как BDNF.
В одном воплощении рецептор гематоэнцефалического барьера выбран из группы, включающей рецептор трансферрина, рецептор инсулина, рецептор инсулиноподобного фактора роста, белок 8, связанный с рецептором липопротеина низкой плотности, белок 1, связанный с рецептором липопротеина низкой плотности, и гепарин-связывающий фактор роста, подобный эпидермальному фактору роста
Пример: гибридные полипептиды, содержащие часть антитела и BDNF
Boado et al. (Biotechnol. Bioeng. 97 (2007) 1376-1386) сообщают о гибридном полипептиде антитело-BDNF, в котором амино-конец человеческого BDNF слит с карбоксильным концом тяжелой цепи химерного антитела, распознающим рецептор инсулина человека. Тем не менее, экспрессия этого гибридного полипептида не представляется возможной, поскольку гибридный полипептид агрегирует в клетке.
В целом, BDNF дикого типа или вариант BDNF может быть слит с полипептидной цепью антитела либо через С-конец, либо через N-конец. Поскольку BDNF действует только в виде гомодимера, разработка биологически активных гибридных полипептидов антитело-BDNF требует, чтобы и i) фолдинг, созревание и сборка BDNF-части, и ii) фолдинг и сборка из полипептидных цепей/доменов антитела были правильными. Таким образом, возможны различные форматы (комплексы) гибридных полипептидов, такие как, например, полное антитело, состоящее из двух тяжелых и легких цепей, Fab-фрагмент антитела, состоящий из легкой цепи и фрагмента тяжелой цепи, одноцепочечный Fab (scFab) или одноцепочечный Fv (scFv) в комбинации с N- или С-концевым слиянием с полипептидом BDNF. Также комплекс гибридного полипептида может содержать две или более двух различных полипептидных цепей, например, комбинацию BDNF-scFv-гибридного полипептида и зрелого (негибридного) BDNF-полипептида, BDNF-Fab(тяжелая цепь)-гибридного полипептида и BDNF-Fab(легкая цепь)-гибридного полипептида или BDNF-Fab(тяжелая цепь)-гибридного полипептида и Fab(легкая цепь)-гибридного полипептида и зрелого (не гибридного)-BDNF-полипептида. Кроме того, полипептид BDNF может быть слит либо непосредственно, либо через линкерный пептид с соответствующей полипептидной цепью антитела. Линкерные пептиды могут различаться по i) числу аминокислот, ii) виду аминокислот (например, отрицательно заряженных и/или гидрофобных аминокислот), и iii) сконструированным предполагаемым мотивам посттрансляционной модификации (например, мотивам сайтов N-гликозилирования). Например, полипептид BDNF может быть конъюгирован с различными фрагментами антител (например, Fab, scFab и scFv), соединенными с помощью различных линкерных пептидов, образуя различные биологически активные гибридные полипептиды.
Один из аспектов, описанных в данном документе, представляет собой гибридный полипептид, включающий
- ровно один полипептид BDNF дикого типа, или вариантный полипептид BDNF, или его фрагмент с активностью BDNF,
- фрагмент антитела, и
- линкерный пептид между полипептидом BDNF и фрагментом антитела.
Один из аспектов, описанных в данном документе, представляет собой гибридный полипептид с активностью BDNF (например, димерный комплекс), включающий
- ровно один тип из полипептида BDNF дикого типа, или вариантного полипептида BDNF, или его фрагмента с активностью BDNF,
- фрагмент антитела, и
- линкерный пептид между полипептидом BDNF и фрагментом антитела.
Другой аспект, описанный в данном документе, представляет собой димерный комплекс, содержащий
- в качестве первого компонента гибридный полипептид, описанный в данном документе, и
- в качестве второго компонента полипептид BDNF дикого типа, или вариант полипептида BDNF, или его фрагмент с активностью BDNF.
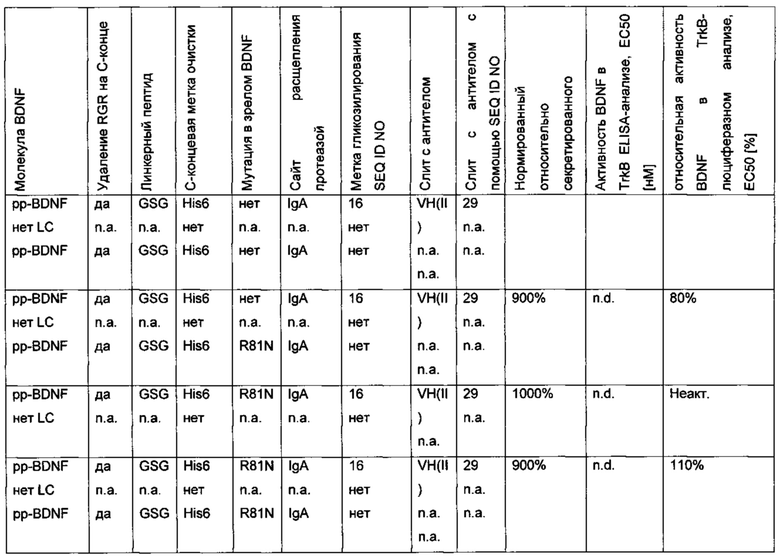
Было обнаружено, что только гибридные форматы "BDNF-антитело" экспрессируются и должным образом собираются в биологически активной форме в комплексе со вторым негибридным полипептидом BDNF, если формат содержит ровно один полипептид BDNF, ковалентно слитый через линкерный пептид с фрагментом антитела, и ровно один не гибридный полипептид BDNF.
Далее было установлено, что экспрессия таких конструкций может быть дополнительно улучшена с помощью линкерного пептида, содержащего один или более чем один отрицательно заряженный аминокислотный остаток.
Различные форматы гибридного полипептида подвергали временной экспрессии в клетках HEK293. Анализ проводили с использованием вестерн-блота и/или высокоэффективной жидкостной хроматографии (HPLC) на белке А. Результаты приведены в следующей ниже таблице (биологические активности нормированы по концентрации зрелого BDNF).

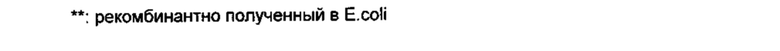
Описание графических материалов
Фиг. 1 Стабильность BDNF-вариантов во время культивирования клеток. BDNF дикого типа (А), полученный в Е.coli, и myc-меченый BDNF, полученный путем временной экспрессии в клетках HEK293 (В), добавляли к растущей культуре клеток HEK293 (10 мкг/мл) и определяли концентрацию BDNF в образцах в день 0 и день 4 с помощью анализов SDS-PAGE в восстанавливающих условиях и вестерн-блота с использованием антитела против BDNF для окрашивания/визуализации; дорожка 1: референсный материал от Peprotech (Е.coli); дорожка 2: маркер молекулярного веса; вестерн-блот был вырезан и повторно собран для простоты.
Фиг. 2 Анализ SDS-PAGE/вестерн-блот клеточных культуральных супернатантов, содержащих экспрессированные/секретированные вариантные полипептиды BDNF; дорожка 1: маркер молекулярного веса; дорожка 2: ряд 1 из таблицы 9; дорожка 3: ряд 5 из таблицы 9; дорожка 4: ряд 2 из таблицы 9; дорожка 5: ряд 4 из таблицы 9; дорожка 6: ряд 3 из таблицы 9; дорожка 7 - референс mBDNF; вестерн-блот был вырезан и повторно собран для простоты.
Фиг. 3 Анализ SDS-PAGE/вестерн-блот клеточных культуральных супернатантов, содержащих экспрессированные/секретированные вариантные BDNF-полипептиды; дорожка 1: маркер молекулярного веса; дорожка 2: ряд 1 из таблицы 6 (гликозилированный про-BDNF); дорожка 3: ряд 2 из таблицы 6 (про-BDNF); дорожка 4: ряд 3 из таблицы 6 (гликозилированный зрелый BDNF); дорожка 5: ряд 4 из таблицы 6 (зрелый BDNF).
Фиг. 4 Анализ SDS-PAGE/вестерн-блот клеточных культуральных супернатантов, содержащих экспрессированные/секретированные вариантные полипептиды BDNF, указанные в таблице 11; дорожка А: ряд 1 из таблицы 11; дорожка В: ряд 2 из таблицы 11; дорожка С: ряд 3 из таблицы 11; дорожка D: ряд 4 из таблицы 11; дорожка Е: ряд 5 из таблицы 11; дорожка F: ряд 6 из таблицы 11; звездочка означает, что были использованы различные условия экспрессии.
Следующие примеры, последовательности и графические материалы приведены для облегчения понимания данного изобретения, истинный объем которого изложен в прилагаемой формуле изобретения. Понятно, что в процедурах могут быть сделаны модификации без отклонения от сущности данного изобретения.
Примеры
Рекомбинантные методики ДНК
Для манипуляций с ДНК использовали стандартные способы, такие как описанные в Sambrook, J. et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярно-биологические реагенты использовали в соответствии с инструкциями изготовителей.
Генный синтез
Нужные гены и сегменты генов получали путем химического синтеза в Geneart GmbH (Регенсбург, Германия). Синтезированные гены и фрагменты генов клонировали в плазмиды для размножения/амплификации в Е.coli. Последовательность ДНК субклонированных генов и фрагментов генов проверяли путем секвенирования ДНК.
Определение белка
Белковую концентрацию очищенных полипептидов оценивали путем определения оптической плотности (OD) при 280 нм, используя коэффициент молярной экстинкции, рассчитанный на основании аминокислотной последовательности полипептида.
Описание основной/стандартной плазмиды для экспрессии в клетках млекопитающих
Нужные гены/полипептиды подвергали экспрессии путем временной трансфекции клеток эмбриональной почки человека (HEK293). Для экспрессии нужного гена/полипептида (например, гибридного полипептида "антитело-GFP", BDNF дикого типа, вариантных полипептидов BDNF, гибридных полипептидов BDNF-Fab и BDNF-scFv) использовали транскрипционную единицу, включающую следующие функциональные элементы:
- немедленный ранний энхансер и промотор из человеческого цитомегаловируса (HCMV), включая интрон А,
- 5'-нетранслируемую область (5'-UTR) тяжелой цепи человеческого иммуноглобулина,
- ген, который должен быть экспрессирован, и
- последовательность полиаденилирования из бычьего гормона роста (BGH рА).
Помимо экспрессионной единицы/кассеты, содержащей нужный ген, который должен быть экспрессирован, основная/стандартная плазмида для экспрессии в клетках млекопитающих содержит
- сайт начала репликации из вектора pUC18, который делает возможной репликацию этой плазмиды в Е.coli, и
- ген бета-лактамазы, который придает устойчивость к ампициллину в Е.coli.
Пример 1
Создание плазмид для экспрессии антител
a) Создание плазмид для экспрессии родительского человеческого антитела против человеческого IGF-1R
Сегменты гена, кодирующие вариабельные области человеческой легкой каппа-цепи (Vk) и тяжелой цепи (VH), присоединяли к сегментам гена, кодирующим константную область человеческой легкой каппа-цепи (Ck) или константную область человеческой тяжелой цепи гамма-1 (СН1-шарнир-СН2-СН3), соответственно. Гены обеих цепей антитела экспрессировали из двух отдельных экспрессионных плазмид, включающих геномную экзон-интронную структуру генов антител. Аминокислотные последовательности зрелых (без сигнальных последовательностей) тяжелой и легкой цепей антитела против человеческого IGF-1R показаны в SEQ ID NO 05 и SEQ ID NO 06.
Экспрессия цепей антитела контролировалась укороченным (с делецией интрона А) немедленным ранним энхансером и промотором из человеческого цитомегаловируса (HCMV), включая 5'-нетранслируемую область (5'-UTR) человеческой тяжелой цепи иммуноглобулина, сигнальную последовательность тяжелой цепи мышиного иммуноглобулина и сигнал полиаденилирования бычьего гормона роста (BGH рА). Экспрессионные плазмиды также содержали сайт начала репликации и ген бета-лактамазы из вектора pUC18 для амплификации плазмиды в Escherichia coli (см. Kopetzki, Е., et al., Virol. J. 5 (2008) 56; Ji, C, et al., J. Biol. Chem. 284(2009) 5175-5185).
b) Создание плазмиды для экспрессии легкой цепи антитела против рецептора трансферрина
Для того чтобы получить легкую цепь антитела против рецептора трансферрина, химически синтезировали ген легкой цепи, кодирующий сигнальную последовательность тяжелой цепи мышиного иммуноглобулина MGWSCIILFLVATATGVHS (SEQ ID NO 14), вариабельный домен VL крысиного антитела против мышиного рецептора трансферрина и константную область человеческой легкой цепи Vkappa. Аминокислотную последовательность VL-домена антитела крысы получали из Boado, R.J., et al., Biotechnol. Bioeng. 102 (2009) 1251-1258. Аминокислотная последовательность легкой цепи химерного крысиного/человеческого антитела против рецептора трансферрина показана в SEQ ID NO 80.
Пример 2
Создание плазмид для экспрессии гибридного полипептида "антитело - GFP"
a) Создание плазмид для экспрессии гибридных полипептидов "анти-IGF-1R-антитело - GFP"
Все гены, кодирующие гибридные полипептиды "тяжелая цепь человеческого анти-IGF-1R-антитела - GFP", собирали путем слияния химически синтезированного фрагмента ДНК, кодирующего соответствующий вариант GFP, и глицин-серинового линкера, состоящего из двух повторов Gly4Ser и дополнительного Gly (тяжелая цепь…LSPG-gggsggggsg-GFP), с 3'-концом гена тяжелой цепи анти-IGF-1R-антитела, кодирующего слегка усеченную константную область человеческой тяжелой цепи гамма-1 (удаление последней природной аминокислоты Lys). Аминокислотная последовательность гибридного белка, состоящего из тяжелой цепи анти-IGF-1R-антитела и eGFP, emGPF и tagGFP, показана в SEQ ID NO 07, SEQ ID NO 08 и SEQ ID NO 09, соответственно.
Гены тяжелой и легкой цепи антитела экспрессировали из двух отдельных экспрессионных плазмид, включающих геномную экзон-интронную структуру генов антитела.
b) Создание экспрессионных плазмид для гибридных полипептидов, состоящих из тяжелой цепи анти-IGF-1R-антитела и eGFP-опсиновой метки
Экспрессионные плазмиды для временной экспрессии в клетках HEK293 гибридных полипептидов, состоящих из тяжелой цепи анти-IGF-1R-антитела и GFP-опсиновой метки, получали из экспрессионных векторов, описанных выше. Они отличаются только в сегменте ДНК, кодирующем GFP-опсиновую метку, где пептид из 19 аминокислот (NGTEGPNFYVPFSNATGVV; опсин(М); SEQ ID NO 10) сливали непосредственно с С-концом соответствующего GFP. В качестве примера, аминокислотная последовательность гибридного полипептида, состоящего из тяжелой цепи анти-IGF-1R-антитела и eGFP-опсиновой (М) метки, показана в SEQ ID NO 33.
Пример 3
Создание экспрессионных плазмид BDNF
a) Создание экспрессионных плазмид для пре-про-BDNF дикого типа
Сегмент ДНК, кодирующий ген пре-про-BDNF человека, получали путем химического синтеза и вставляли в основной экспрессионный вектор, описанный выше. С этой целью ген пре-про-BDNF лигировали с CMV-промотором на его 5'-конце и с последовательностью полиаденилирования бычьего гормона роста на его 3'-конце. Аминокислотная последовательность пре-про-BDNF-белка дикого типа показана в SEQ ID NO 20.
b) Создание экспрессионных плазмид для вариантов BDNF
Для того чтобы получить оптимальный выход продукции, было сконструировано несколько вариантов BDNF (см. таблицу ниже):
- в некоторых вариантах сигнальная последовательность BDNF дикого типа (пресегмент) была заменена на сигнальную последовательность, полученную из высоко экспрессированной тяжелой цепи мышиного иммуноглобулина (MGWSCIILFLVATATGVHS);
- в некоторых вариантах использование кодонов в гене, кодирующем BDNF, было заменено на использование оптимизированных кодонов в просегменте и/или в зрелой части BDNF; гены BDNF с оптимизированными кодонами были получены путем обратной трансляции аминокислотной последовательности с использованием алгоритмов из Geneart (см., например, Fath, S., et al., PLOS One 6 (2011) e17596);
- в некоторых вариантах использовали T7-His6-метку (SEQ ID NO 12), так как она, как правило, увеличивает экспрессию белка (см., например, Luan, С.Н., et al., Genome Res. 14 (2004) 2102-2110);
- в большинство вариантов BDNF была включена His6-метка для упрощения получения/очистки образца;
- в некоторых вариантах последние три С-концевые аминокислоты, RGR-мотив зрелого BDNF, были удалены, так как они могут функционировать как зашифрованный сайт расщепления протеазой для протеаз, таких как фурин, или других конвертаз пробелка;
- в другом варианте сигнальная последовательность и просегмент BDNF были заменены соответствующей аминокислотной последовательностью человеческого NGF, так как было опубликовано, что этот вариант повышает экспрессию другого нейротрофина (Iwane et al., Appl. Microbiol. Biotechnol. 41 (1994) 225-232).
1: С-концевой RGR-мотив BDNF удален (deltaRGR)
2: сигнальная последовательность, полученная из высоко экспрессируемой тяжелой цепи иммуноглобулина мышиного антитела (MGWSCIILFLVATATGVHS), определенная аминокислотной последовательностью, представленной в SEQ ID NO 14
3: сигнальная последовательность и про-фрагмент BDNF, замененные соответствующей последовательностью из человеческого NGF
с) Создание экспрессионных плазмид для вариантов пре-про(IgA)-BDNF и пре-про(IgA; His6). частично включающих His6-метку с N-конца зрелого BDNF в просегменте
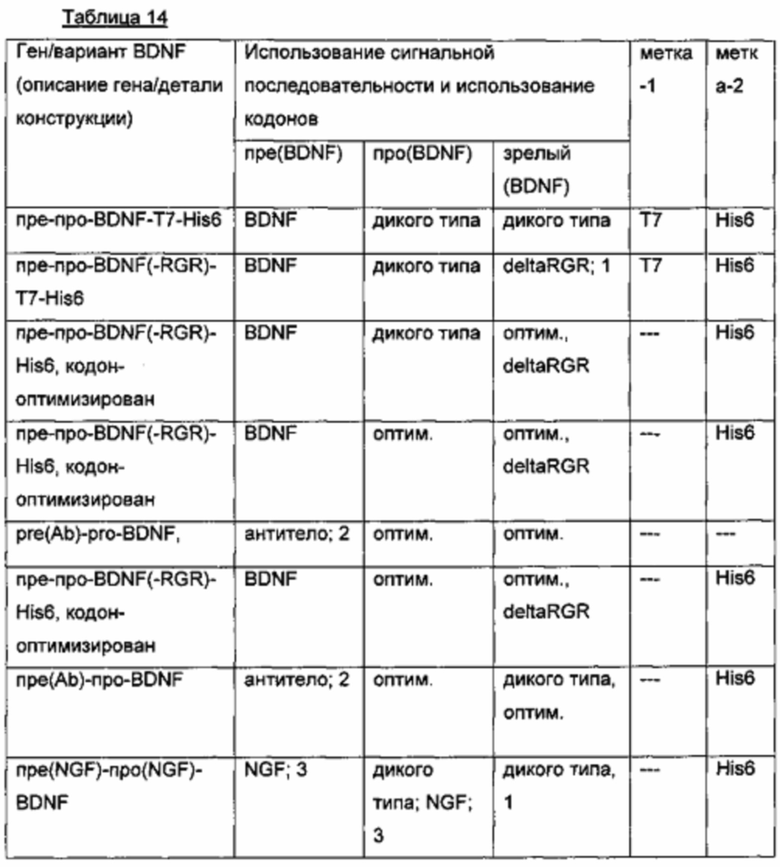
Ген пре-про-(IgA)-BDNF кодирует вариант прополипептида, в котором природный сайт расщепления фурина (RVRR) заменен сконструированным сайтом расщепления IgA-протеазой с последовательностью GSVVAPPAP (см. таблицу ниже).
Кроме того, в некоторых вариантах в прополипептид BDNF вводили точечную мутацию R54A, чтобы удалить предполагаемый сайт расщепления протеазой (см. Mowla, S.J., et al., J. Biol. Chem. 276 (2001) 12660-12666). Кроме того, в некоторых вариантах в профрагмент также включали удаляемую His6-метку с N-конца зрелого BDNF. Эта метка упрощает белковую очистку про(IgA; His6)-BDNF-вариантного белка. После окончательного созревания белка in vitro с помощью IgA-протеазы про(IgA; His6-фрагмент) удаляли и, таким образом, избегали потенциального риска иммуногенности.
Экспрессионные плазмиды для временной экспрессии пре-про(IgA)-BDNF- и пре-про(IgA; His6)-BDNF-вариантных генов/белков в клетках HEK293 получали из экспрессионного вектора, описанного выше, который кодирует пре-про-BDNF(-RGR)-Т7-His6-белок. Они дифференцируются по следующим характеристикам:

Нумерация мутации R54A основана на аминокислотной последовательности пре-про-BDNF дикого типа (SEQ ID NO 20)
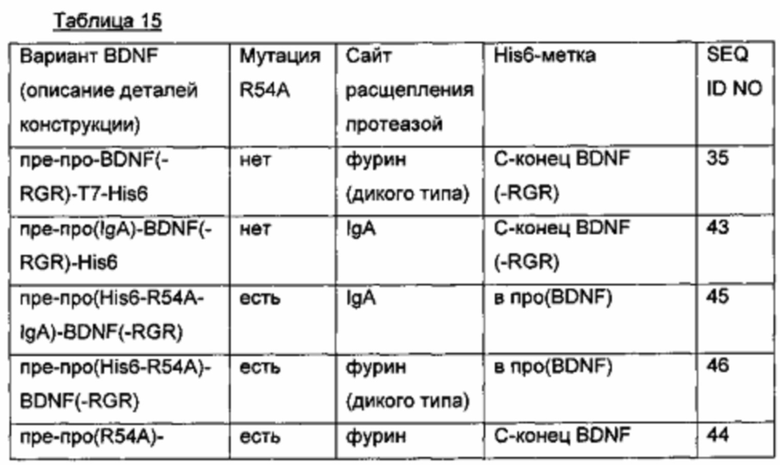
d) Создание экспрессионных плазмид для вариантов BDNF со сконструированной изоэлектрической точкой
Ранее были описаны некоторые экспрессированные в Е.coli и подвергнутые рефолдингу варианты BDNF со сконструированной пониженной IEP. Ген пре-про-BDNF(-RGR)-T7-His6 подвергали мутированию соответственно, и полученные мутантные гены BDNF временно экспрессировали в клетках HEK293 (см. таблицу ниже). Кроме того, был сконструирован myc-меченый вариант BDNF, так как (1) myc-метка (EQKLISEEDL; SEQ ID NO 90) вводит изменение заряда примерно -3, и (2) myc-метка имеет человеческое происхождение и, таким образом, должна быть менее иммуногенной.
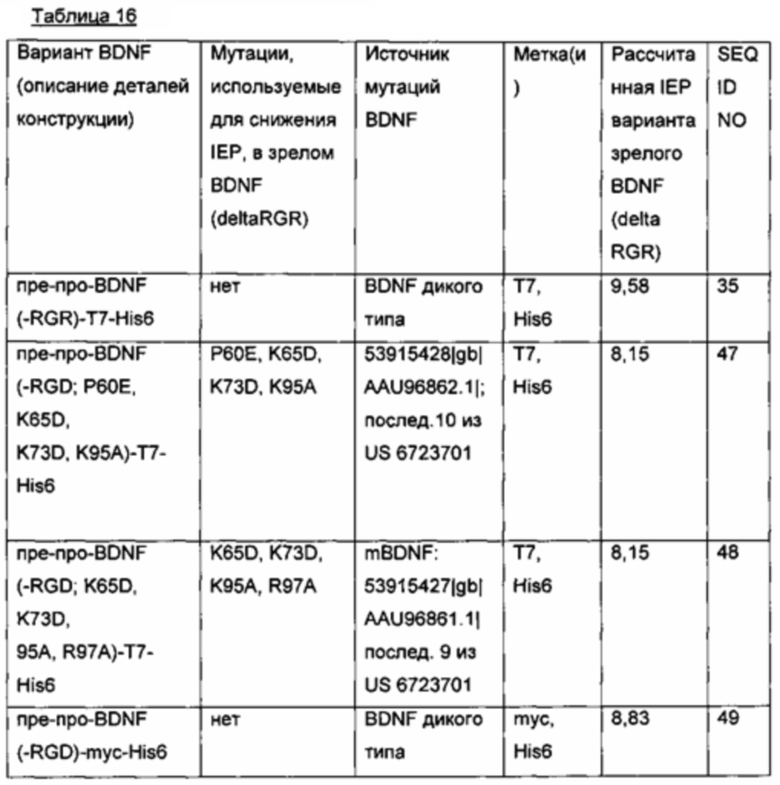
-RGD и (deltaRGR); удаление последних трех С-концевых аминокислот зрелого BDNF;
нумерация аминокислотных мутаций начинается с первой аминокислоты зрелого BDNF;
IEP зрелых BDNF(-RGR)-вариантов рассчитывали с использованием статистической программы Pepstats от European Molecular Biology Open Software Suite (EMBOSS).
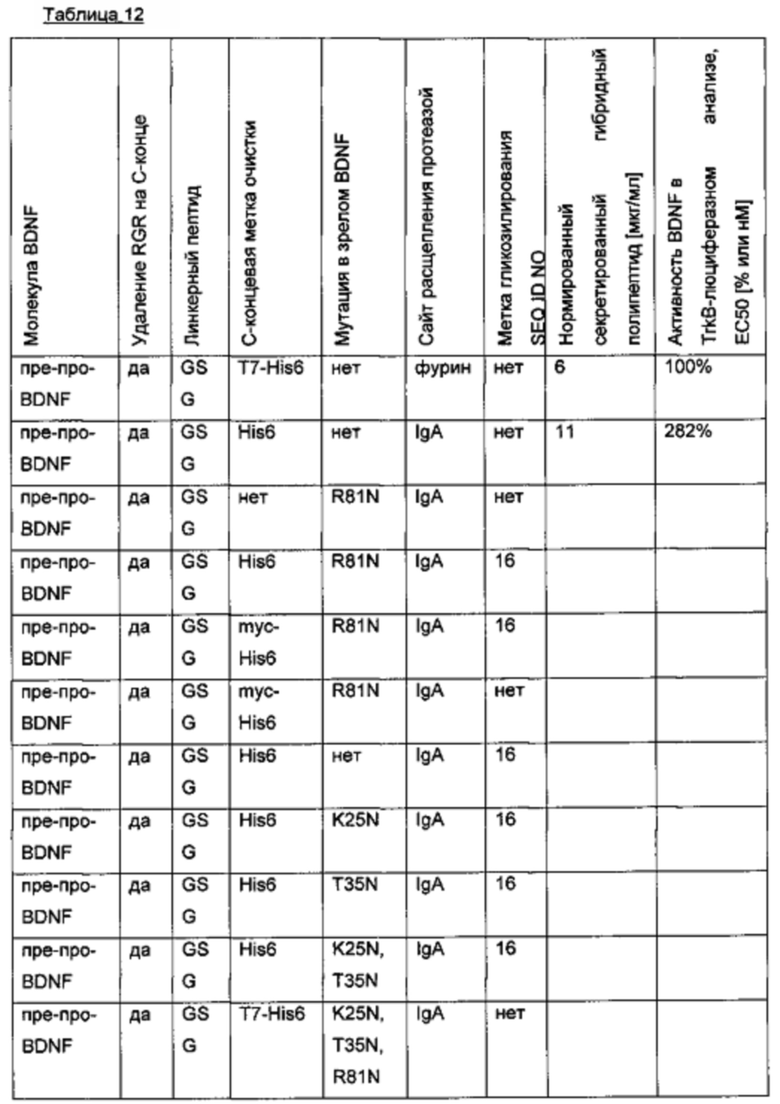
e) Создание экспрессионных плазмид для вариантов BDNF с дополнительными сайтами гликозилирования
Были получены варианты BDNF, которые несут (1) С-концевую метку, содержащую сайты гликозилирования или (2) один сконструированный сайт гликозилирования в пределах зрелой BDNF-группировки.
Метки гликозилирования были выведены из опубликованных последовательностей (например, Meder, D., et al., J. Cell Biol. 168 (2005) 303-313; Bulbarelli, A., et al., J. Cell Sci. 115 (2002) 1689-1702; Perlman, S., et al., J. Clin. Endocrinol. Metab. 88 (2003) 3227-3235; WO 2002/002597), и предположительные сайты N-гликозилирования были предсказаны с помощью искусственной нейронной сети (сервер NetNglyc; http://www.cbs.dtu.dk/services/NetNGlyc/).
Для введения сайтов N-гликозилирования в зрелую BDNF-группировку последовательность проверяли на наличие аспарагинов, серинов или треонинов в последовательности зрелого BDNF. Затем на основе трехмерной белковой структуры человеческого BDNF (1bnd; www.rcsb.org) были исключены все не поверхностно расположенные остатки Asn, Ser или Thr. Для остальных поверхностно расположенных остатков Asn, Ser и Thr были идентифицированы смежные аминокислотные остатки, чтобы сконструировать предполагаемый сайт N-гликозилирования (консенсусный мотив: N-X-(S/T), X - любая аминокислота кроме Pro) путем сайт-направленного мутагенеза. Аминокислотные позиции этих предполагаемых сконструированных сайтов N-гликозилирования были использованы для идентификации соответствующих аминокислот в структурно и функционально гомологичных нейротрофинах NGF и NT-3 путем выравнивания последовательностей и сравнения 3D-структур белка. Были исключены те аминокислотные позиции, которые, как ожидается, будут частью поверхности взаимодействия нейротрофин::p75NTR или нейротрофин::Trk(А, В), на основании гомологичных кристаллических структур рецептор:лиганд (например, 3buk, 3ij2 и 2ifg). Отдельные мутации для поверхностно экспонированных предполагаемых сайтов N-гликозилирования вне предполагаемых поверхностей взаимодействия BDNF-TrkB/p75 указаны в приведенной ниже таблице (второй столбец).
Экспрессионные плазмиды для временной экспрессии N-гликозилированных вариантных белков BDNF в клетках HEK293 получены из экспрессионного вектора, кодирующего вариант BDNF пре-про-BDNF(-RGR)-T7-His6 (-RGR: усеченный зрелый BDNF дикого типа, в котором последние три С-концевые аминокислоты RGR были удалены; Т7-метка и His6-метка присоединены к усеченному С-концу BDNF через GSG-линкер). Сегменты BDNF, метки, гликолинкеры и мутации, введенные для создания дополнительных искусственных сайтов N-гликозилирования, представлены в следующей таблице.
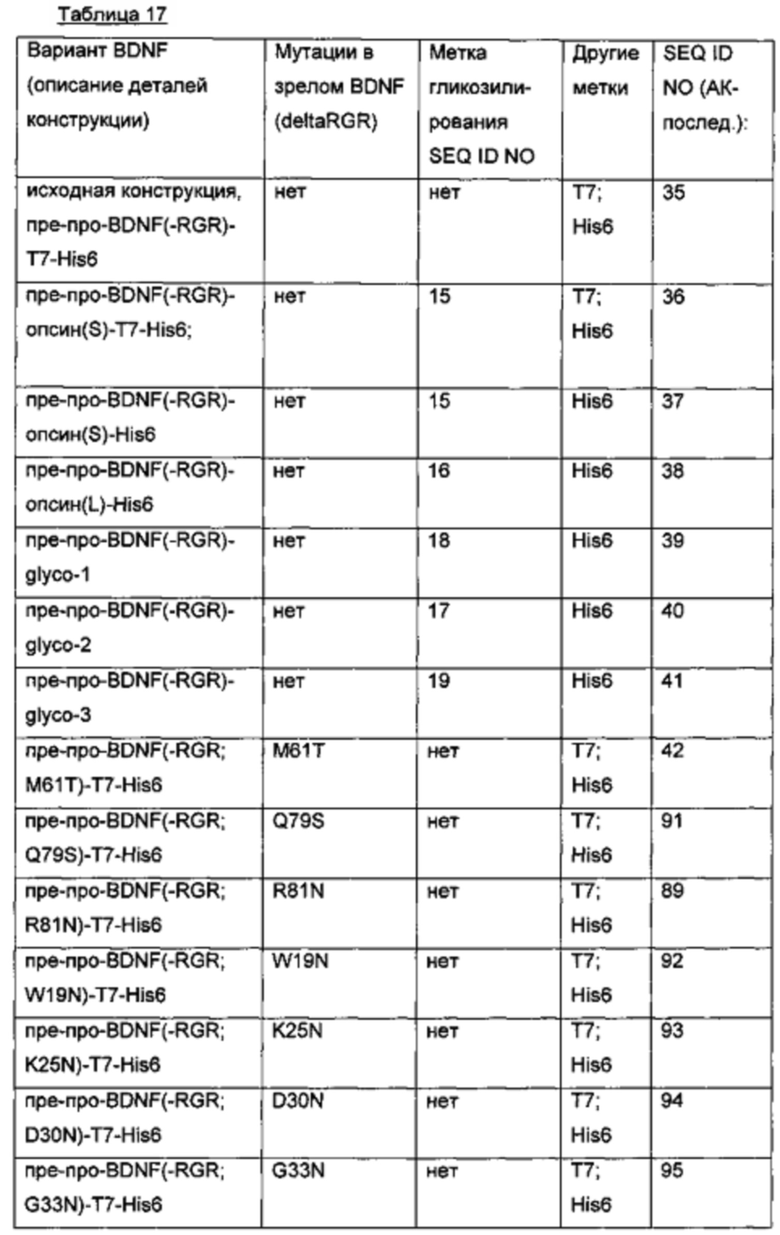
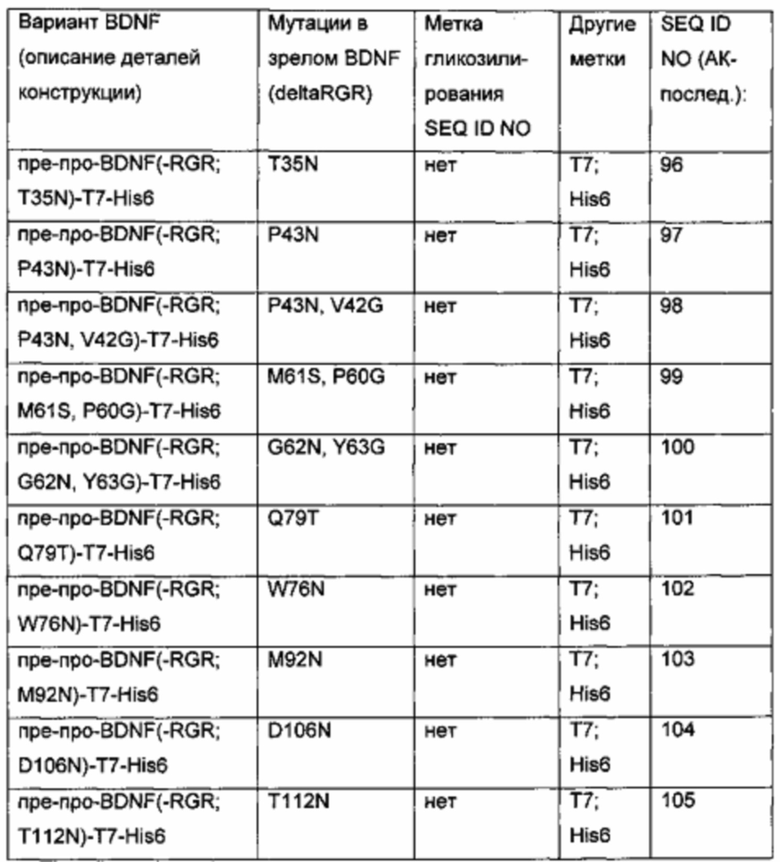
SEQ ID NO № для аминокислотных мутаций, введенных путем сайт-направленного мутагенеза в зрелый BDNF (deltaRGR), иллюстративно приведены для BDNF-вариантов пре-про-BDNF(-RGR; M61T)-T7-His6 и пре-про-BDNF(-RGR; R81N)-T7-His6.
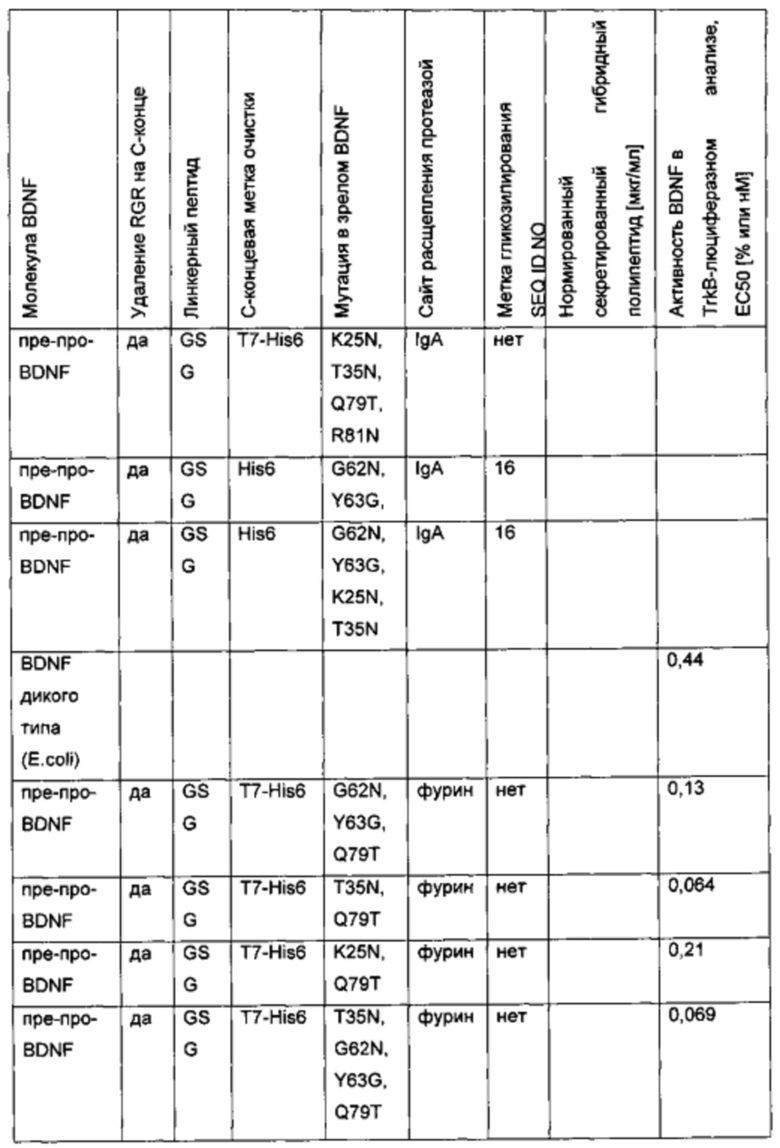
f) Создание экспрессионных плазмид для вариантов BDNF, содержащих комбинированные мутации для введения множественных (двух или более) сайтов N-гликозилирования и сайта расщепления IgA
Для того чтобы создать варианты BDNF с множественными сайтами N-гликозилирования, были объединены некоторые из дополнительных сайтов N-гликозилирования, выявленных в предыдущих экспериментах. Исходная конструкция вариантного полипептида пре-про(IgA)-BDNF(-RGR)-His6 характеризуется сайтом расщепления IgA-протеазой вместо нативного сайта фурина в просегменте, усечением с С-конца зрелого BDNF (удаление последних трех аминокислот RGR) и С-концевой His6-меткой. Желательные мутации и метки были введены/присоединены, как показано в приведенной ниже таблице.
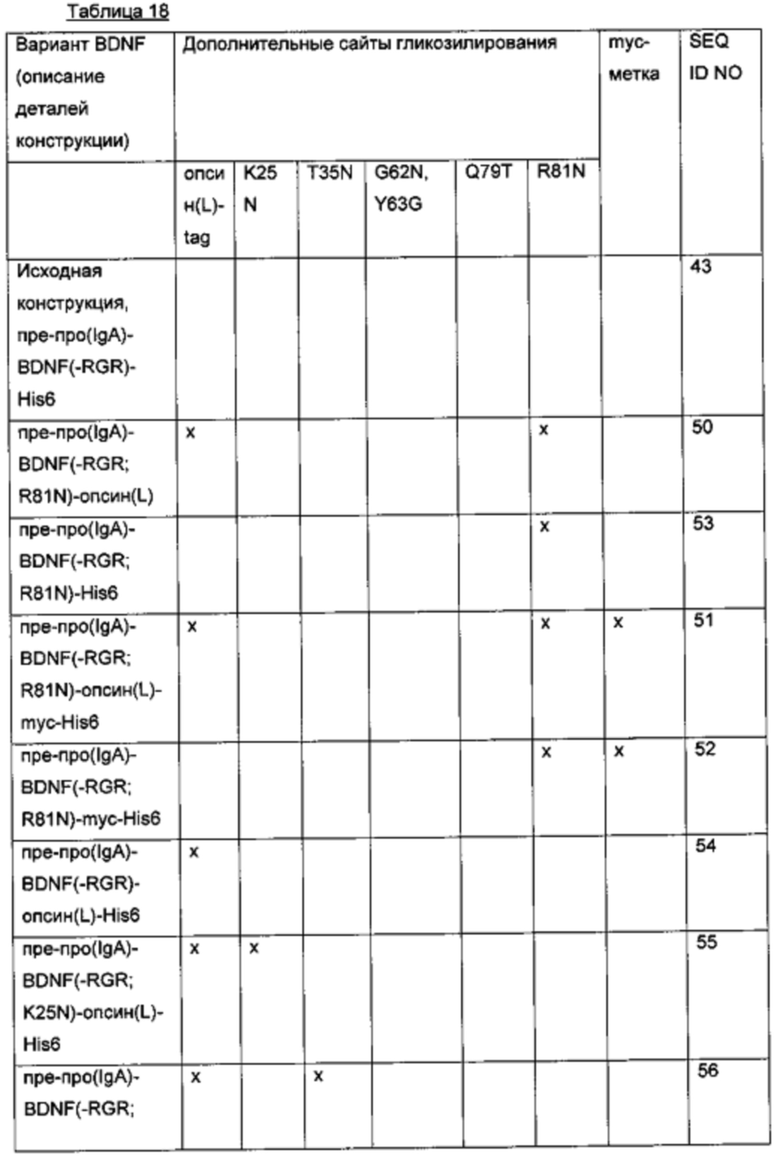
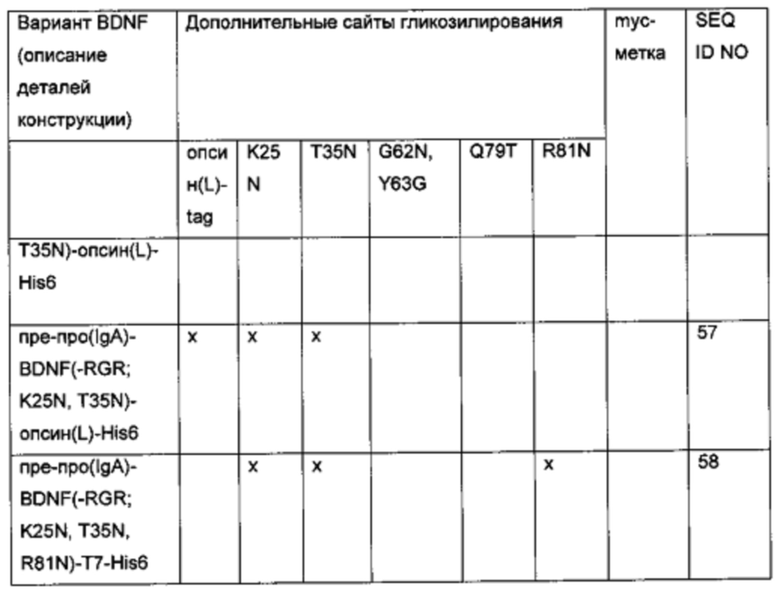
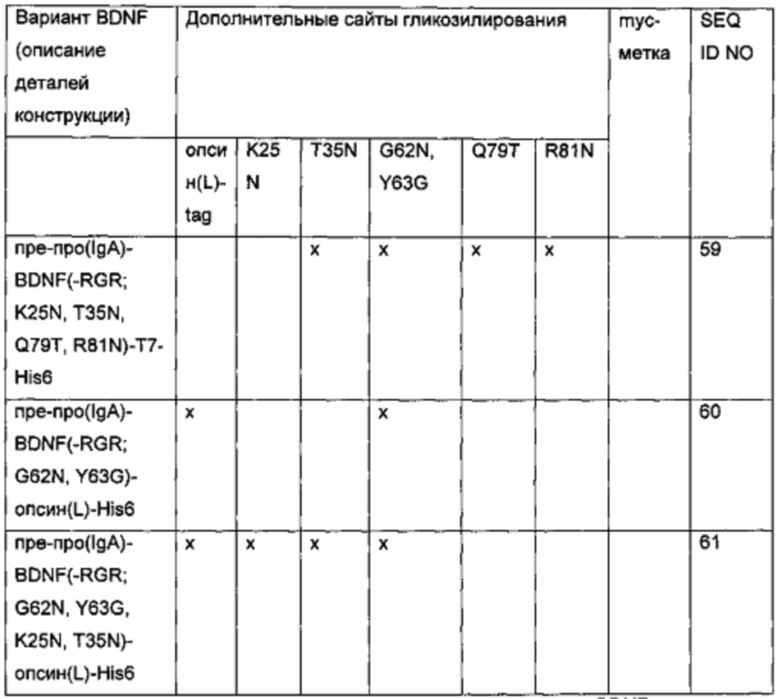
g) Создание экспрессионных плазмид для вариантов BDNF. содержащих комбинированные мутации для введения множественных (двух или более) сайтов N-гликозилирования и сайта расщепления IgA
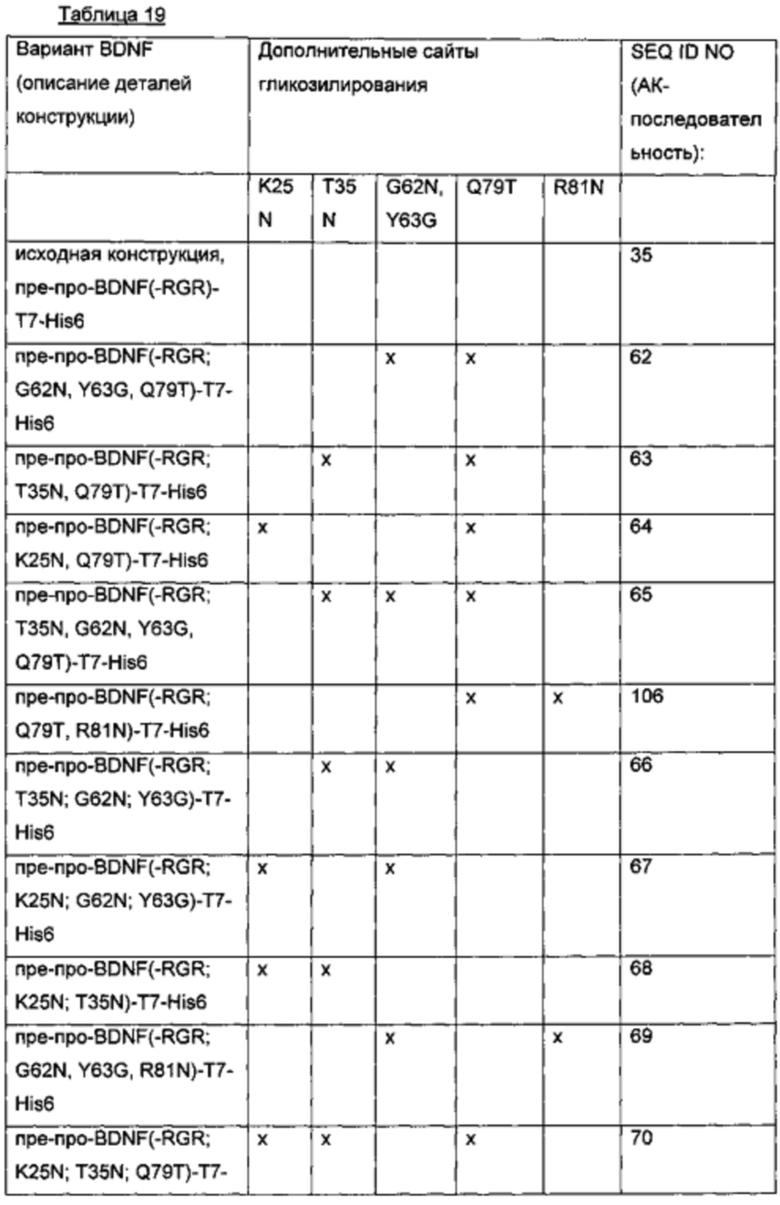
Для того чтобы создать варианты BDNF с множественными сайтами N-гликозилирования, были объединены некоторые из дополнительных сайтов N-гликозилирования, выявленных в предыдущих экспериментах. С этой целью в качестве исходного материала использовали вариантный белок пре-про-BDNF(-RGR)-T7-His6, который характеризуется усечением с С-конца зрелого BDNF (удалением последних трех аминокислот RGR) и С-концевой Т7-His6-меткой. Желательные мутации были вставлены, как показано в приведенной ниже таблице.
Пример 4
Создание экспрессионных плазмид для гибридных полипептидов, состоящих из фрагментов антитела и BDNF
a) Создание экспрессионных плазмид для гибридных полипептидов, состоящих из BDNF и Fab тяжелой цепи антитела
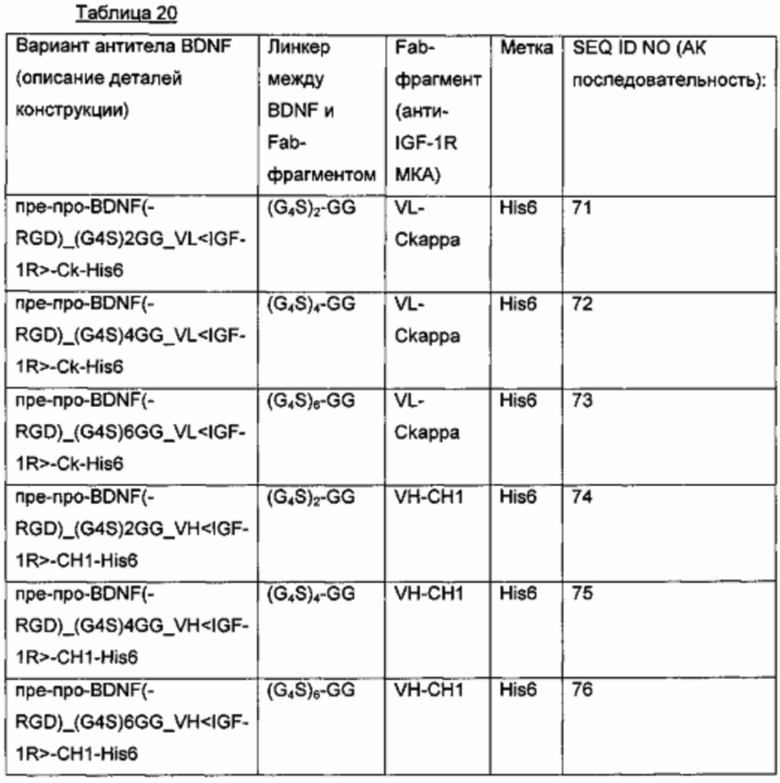
Для того чтобы получить гибридные полипептиды BDNF-(Gly4Ser)n-Fab(тяжелой цепи анти-IGF-1R-антитела), были сконструированы плазмиды для временной экспрессии в клетках HEK293, которые несут химически синтезированный фрагмент ДНК с CDS (кодирующей ДНК-последовательностью), кодирующей полипептиды со следующими характеристиками:
- пре-про-BDNF-группировка дикого типа с удаленным С-концевым RGR-мотивом слита на С-конце с глицин-богатым линкером с последующей Fab-частью тяжелой цепи (VH-CH1) человеческого анти-IGF-1R-антитела и С-концевой His6-меткой;
- глицин-богатый линкер состоит из мотива (G4S)2-GG, или (G4S)4-GG, или (G4S)6-GG (см. таблицу ниже).
b) Создание экспрессионных плазмид для гибридных полипептидов, состоящих из BDNF и Fab легкой цепи антитела
Для того чтобы получить гибридные полипептиды BDNF-(Gly4Ser)n-Fab, были сконструированы плазмиды для временной экспрессии в клетках HEK293, которые несут химически синтезированный фрагмент ДНК с CDS, кодирующей полипептиды со следующими характеристиками:
- пре-про-BDNF-группировка дикого типа с удаленным С-концевым RGR-мотивом слита на С-конце с глицин-богатым линкером с последующими Fab-доменами легкой цепи VL-Ckappa человеческого анти-IGF-1R-антитела и С-концевой His6-метки;
- глицин-богатый линкер состоит из мотива (G4S)2-GG, или (G4S)4-GG, или (G4S)6-GG (см. таблицу ниже).
c) Создание экспрессионной плазмиды для легкой цепи анти-IGF-1R-антитела
Нативную легкую цепь анти-IGF-1R-антитела использовали для создания BDNF-Fab-комплексов на основе анти-IGF-1R. Создание плазмиды для экспрессии легкой цепи анти-IGF-1R-антитела описано в примере 1.
d) Создание экспрессионной плазмиды для гибридных полипептидов BDNF-Fab(тяжелой цепи анти-IGF-1R-антитела). содержащих отрицательно заряженный Gly-Asp-линкер
Для того чтобы получить гибридный полипептид BDNF-(G3D)4-РаЬ(тяжелой цепи анти-IGF-1R-антитела), конструировали плазмиду для временной экспрессии в клетках HEK293, которая скрывала химически синтезированный фрагмент ДНК с CDS, кодирующей полипептид со следующими характеристиками:
- пре-про-BDNF-группировка дикого типа с удаленным С-концевым RGR-мотивом слита на С-конце с глицин-богатым отрицательно заряженным линкером с последующими Fab-доменами тяжелой цепи VH-CH1 человеческого анти-IGF-1R-антитела и С-концевой His6-меткой;
- глицин-богатый отрицательно заряженный линкер состоит из мотива (G3D)4-GGGS.
е) Создание экспрессионных плазмид для гибридных полипептидов BDNF-Fab(тяжелой цепи анти-IGF-1R-антитела), несущих сайт расщепления IgA в про-BDNF-сегменте, отрицательно заряженный GlyAsp-линкер и множественные сайты N-гликозилирования
Для того чтобы получить гибридные полипептиды про(IgA)-BDNF-(G3D)4-Fab (тяжелой цепи анти-IGF-1R-антитела) с множественными сайтами N-гликозилирования, конструировали плазмиды для временной экспрессии в клетках HEK293, которые несли химически синтезированный фрагмент ДНК с CDS, кодирующей полипептиды со следующими характеристиками:
- пре-про(IgA)-BDNF-группировка с удаленным С-концевым RGR-мотивом зрелого BDNF слита на С-конце с удлиненной опсин-меткой (NGTEGPNFYVPFSNATGVVR; опсин(L); SEQ ID NO 16) с последующими отрицательно заряженным линкером, богатым глицином и аспарагиновой кислотой, и Fab-частью тяжелой цепи (VH-CH1, частично удлинен пептидом EPKSC из шарнира) человеческого моноклонального антитела, направленного против инсулиноподобного фактора роста человека 1 (IGF-1R) и С-концевой His6-метки;
- отрицательно заряженный линкер, богатый глицином и аспарагиновой кислотой, состоит либо из мотива (G3D)4-GGGS, либо из мотива (G2D)5-G2SG.
Детали конструкций приведены в таблице ниже.

е) Создание экспрессионных плазмид для гибридных полипептидов BDNF-Fab(тяжелой цепи антитела против рецептора трансферрина), несущих сайт расщепления IgA в про-BDNF-сегменте, отрицательно заряженный GlyAsp-линкер и множественные сайты N-гликозилирования
Для того чтобы получить гибридные полипептиды про(IgA)-BDNF-(G3D)4-Fab(тяжелой цепи анти-TfR-антитела) с отрицательно заряженным GlyAsp-линкером и множественными сайтами N-гликозилирования, конструировали плазмиды для временной экспрессии в клетках HEK293, которые несли химически синтезированный фрагмент ДНК с CDS, кодирующей полипептиды со следующими характеристиками:
- пре-про(IgA)-BDNF-группировка с удаленным С-концевым RGR-мотивом слита на С-конце с опсин(L)-меткой с последующими отрицательно заряженным линкером, богатым глицином и аспарагиновой кислотой, и химерной крысиной/человеческой Fab-частью тяжелой цепью, в которой вариабельный домен VH получен из крысиного моноклонального антитела 8D3, которое направлено против мышиного рецептора трансферрина (mTfR), и в которой домен СН1 получен из человеческого IgG1, и С-концевой His6-меткой; аминокислотная последовательность VH-домена антитела получена из Boado, R.J. et al., Biotechnol. Bioeng. 102 (2009) 1251-1258);
- отрицательно заряженный линкер, богатый глицином и аспарагиновой кислотой, состоит из мотива (G3D)4-GGGS;
- в большинстве вариантов эндогенный сайт расщепления фурина/РС конвертазы между просегментом и зрелой частью BDNF заменен на сайт расщепления IgA-протеазой (GSVVAPPAP);
- в некоторых вариантах усеченная зрелая BDNF-группировка дикого типа (BDNF; deltaRGR) заменена на усеченный зрелый вариант BDNF, несущий искусственный сайт гликозилирования R209N (BDNF (deltaRGR; R209N)).
Детали конструкций приведены в таблице ниже.
Пример 6
е) Создание экспрессионной плазмиды для гибридных полипептидов BDNF-scFv
Для того чтобы получить гибридные полипептиды BDNF(G4S)3-scFv-тяжелая цепь анти-IGF-1R-антитела, конструировали плазмиды для временной экспрессии в клетках HEK293, которые несли химически синтезированный фрагмент ДНК с CDS, кодирующей полипептид со следующими характеристиками:
- пре-про-BDNF-группировка дикого типа с удаленным С-концевым RGR-мотивом слита на С-конце с (G4S)3-линкером с последующей scFv-группировкой человеческого антитела против человеческого IGF-1R;
- scFv-группировка антитела против человеческого IGF-1R построена из домена VL, за которым следует (G4S)4-GG-линкер, VH-область и His6-метка; аминокислотная последовательность гибридного белка пре-про-BDNF(-RGR)_(G4S)3_scFv-His6<IGF-1R> показана в SEQ ID NO 78.
Пример 7
Временная экспрессия, очистка и аналитическая характеризация полипептидов
Временная экспрессия
Полипептиды были получены путем временной трансфекции клеток HEK293 (линия, полученная из эмбриональных клеток почки человека 293), культивированных в среде F17 (Invitrogen Corp.). Для трансфекции использовали реагент "293-Free" Transfection Reagent (Novagen). Гибридные полипептиды, содержащие антитело и фрагмент антитела (Fab и scFv), экспрессировали с одной, двух или трех различных плазмид с использованием эквимолярном соотношения плазмиды после трансфекции. Трансфекцию выполняли согласно инструкциям производителя. Клеточные культуральные супернатанты, содержащие рекомбинантный полипептид, собирали в течение 4-7 дней после трансфекции. Супернатанты хранили при пониженной температуре до очистки.
Общая информация касательно рекомбинантной экспрессии человеческих иммуноглобулинов, например, в клетках HEK293, дана, например, в Meissner, P., et al., Biotechnol. Bioeng. 75 (2001) 197-203.
Очистка
a) GFP-гибридные полипептиды очищали с помощью двухэтапной процедуры, включающей хроматографию на белке А и эксклюзионную хроматографию на колонке Superdex 200™
Клеточные культуральные супернатанты, содержащие GFP-гибридные полипептиды, фильтровали. После этого GFP-гибридные полипептиды захватывали путем аффинной хроматографии с использованием колонки HiTrap MabSelectSuRe (GE Healthcare), уравновешенной PBS (1 мМ KH2PO4, 10 мМ NaHPO4, 137 мМ NaCl, 2.7 мМ KCl, рН 7,4). Несвязанные полипептиды удаляли путем промывания уравновешивающего буфера, и гибридный полипептид извлекали с помощью 0,1 М цитратного буфера, рН 2,8. Сразу после элюирования фракции нейтрализовали до рН 6,0 с помощью 1 М Трис-основания, рН 9,0.
Эксклюзионную хроматографию на Superdex 200™ (GE Healthcare, Упсала, Швеция) использовали в качестве второго этапа очистки. Эксклюзионную хроматографию проводили в 50 мМ гистидиновом буфере, 0,15 М NaCl, рН 6,8. Выделенные GFP-гибридные полипептиды хранили при -80°С.
b) Гистидин-меченные белки очищали с помощью двухэтапного протокола, начиная с аффинной хроматографии с иммобилизованным ионом металла (IMAC), и продолжая эксклюзионной хроматографией на колонке с Superdex 75™.
Клеточные культуральные супернатанты, содержащие гистидин-меченные полипептиды, подводили с помощью NaCl до конечной концентрации NaCl 500 мМ. Отфильтрованный культуральный супернатант наносили на колонку Ni-Sepharose™ 6 Fast Flow, предварительно уравновешенную NiA-буфером (50 мМ Tris, 300 мМ NaCl, 5 мМ имидазол, содержащий таблетку смеси ингибиторов протеаз без EDTA, как указано в инструкции производителя; EDTA-free Complete Mini Tablets; Roche Applied Science) со скоростью потока 1 мл/мин с использованием системы  explorer 100 (GE Healthcare, Упсала, Швеция). Колонку промывали NiA-буфером до тех пор, пока показатели УФ не возвращались к исходному уровню. Гистидин-меченный полипептид элюировали линейным градиентом имидазола от 5 мМ до 300 мМ в 50 мМ TRIS и 500 мМ NaCl, рН 8,0, в 10 объемах колонки.
explorer 100 (GE Healthcare, Упсала, Швеция). Колонку промывали NiA-буфером до тех пор, пока показатели УФ не возвращались к исходному уровню. Гистидин-меченный полипептид элюировали линейным градиентом имидазола от 5 мМ до 300 мМ в 50 мМ TRIS и 500 мМ NaCl, рН 8,0, в 10 объемах колонки.
Эксклюзионную хроматографию на Superdex 75™ (GE Healthcare, Упсала, Швеция) использовали в качестве второго этапа очистки. Эксклюзионную хроматографию проводили в 50 мМ гистидиновом буфере, 0,15 М NaCl, рН 6,8. Элюированные гистидин-меченные белки хранили при -80°С.
Аналитическая характеризация
Белковые концентрации очищенных полипептидов определяли путем измерения оптической плотности (OD) при 280 нм, используя коэффициент молярной экстинкции, рассчитанный на основании аминокислотной последовательности. Чистоту и надлежащее образование димеров полипептидов анализировали с помощью SDS-PAGE в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотреитол) и окрашивания Кумасси бриллиантовым синим. Содержание агрегатов в препаратах Fc-гибридных полипептидов определяли с помощью высокоэффективной SEC с использованием аналитической эксклюзионной колонки Superdex 200™ (GE Healthcare, Упсала, Швеция). Целостность аминокислотного остова выделенных полипептидов подтверждали с помощью масс-спектрометрии на Nano Electrospray QTOF после удаления N-гликанов путем ферментативной обработки комбинацией нейраминидазы, О-гликаназы и пептид-N-гликозидазы F (Roche Applied Science, Мангейм, Германия).
Определение концентрации BDNF в культуральных супернатантах
Концентрацию BDNF дикого типа, вариантов BDNF и гибридных полипептидов, содержащих BDNF, в культуральных супернатантах определяли путем полуколичественного анализа вестерн-блота с использованием рекомбинантного человеческого BDNF от Peprotech (каталоговый номер: 450-02) в качестве референсного стандарта. Для окрашивания использовали кроличье анти-BDNF-антитело (Santa Cruz, каталоговый номер: SC-20981) (первое антитело), конъюгированную с пероксидазой хрена овечью противокроличью антисыворотку (разведение 1:5000, Roche Diagnostics GmbH, Германия) (вторичное антитело) и усиленный хемилюминесцентный субстрат (LUMI-Light plus Western Blotting substrate, Roche Diagnostics GmbH, Германия).
Концентрацию BDNF-гибридных полипептидов также определяли путем BDNF ELISA с использованием набора BDNF Emax® ImmunoAssay Kit от Promega (каталоговый номер: G7610) в соответствии с инструкциями поставщика.
Пример 8
Функциональная характеризация in vitro
Определение биологической активности GFP дикого типа и GFP-содержащих гибридных полипептидов
Биологическую активность очищенного GFP дикого типа и GFP-содержащих гибридных полипептидов контролировали с помощью их свойств биолюминесценции (GFP-специфическая флуоресценция).
Определение аффинности связывания BDNF с помощью поверхностного плазмонного резонанса (SPR, BIAcore)
Аминное сочетание примерно 750 резонансных единиц (RU) в системе захвата (захват МКА, специфичного для человеческого IgG, Jackson Immunoresearch) выполняли на чипе СМ5 при рН 4,5 с использованием набора для аминного связывания в соответствии с инструкцией производителя (поставляется компанией GE Healthcare, Упсала, Швеция). Человеческий Fc-меченый TrkB (R&D Systems, каталоговый номер: 688-ТК-100) захватывали в концентрации 5 мкг/мл. Избыточные сайты связывания блокировали путем введения смеси человеческих Fc в концентрации 1,25 мкМ (Biodesign, каталоговый номер: 50175). Различные концентрации BDNF-содержащих гибридных полипептидов в интервале от 0,1 нМ до 50 нМ пропускали со скоростью потока 10 мкл/мин через проточные ячейки при 298 К в течение 120-240 с. Фазу диссоциации наблюдали в течение периода до 600 с и прерывали путем переключения с раствора образца на проточный буфер. Поверхность подвергали регенерации путем промывания в течение 1 мин 100 мМ раствором фосфорной кислоты со скоростью потока 30 мкл/мин. Для всех экспериментов был выбран буфер HBS-P+, поставляемый компанией GE Healthcare (10 мМ HEPES ((4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота)), рН 7,4, 150 мМ NaCl, 0,05% (об/об) поверхностно-активного вещества Р20).
Разницу в объемных рефракционных показателях корректировали путем вычитания ответа, полученного для поверхности, связанной с бланком. Пустые введения также вычитали (двойной референс).
Константу равновесной диссоциации (Kd), определенную как ka/kd, определяли с помощью анализа кривых сенсограммы, полученных с несколькими различными концентрациями, используя пакет программного обеспечения BIAevaluation 4.1. После подгонки данных следовала подходящая модель связывания.
Определение аффинности связывания BDNF с помощью ELISA
Связывающие свойства BDNF-содержащих гибридных белков определяли с помощью ELISA TrkB. Планшеты Maxisorb покрывали 1 мкг/мл гибридного TrkB-Fc (R&D Systems) в PBS в течение ночи при температуре 4°С. После блокировки планшета с помощью PBSTC (PBS с 0,05% Tween-20 и 2% куриной сыворотки (Gibco)) в течение 1 ч при комнатной температуре и трех промываний с помощью PBST BDNF-содержащие гибридные белки или в только BDNF добавляли в лунки в концентрациях от 15 до 500 нг/мл в PBSTC и инкубировали в течение 2 ч при комнатной температуре. После шести промываний лунки инкубировали с мышиным анти-BDNF-антителом (клон 4F11.1A1, 1 мкг/мл в PBSTC) и, после дополнительных промываний, с противомышиным HRP-антителом (1:10000 в PBSTC), обе инкубации в течение 1 ч при комнатной температуре. После трех промываний с помощью PBST определяли активность HRP с использованием ABTS-субстрата и фотометрической оценки при длине волны 492 нм.
Определение биологической активности BDNF-содержащих гибридных белков с помощью анализа с репортерным геном TrkB
Биологическую активность BDNF-вариантов и BDNF-содержащих гибридных белков определяли с помощью TrkB-трансфицированных клеток линии СНО, содержащий стабильно трансфицированный ген-репортер люциферазы под контролем SRE (элемент ответа сыворотки)-содержащего промотора (СНО-K1-hTrkB/pSRE-Luc). За день до экспериментов клеточную среду заменяли с ростовой среды (среда Хэма F12, содержащая 10% FCS, 2 мМ L-глутамина, 300 мкг/мл G418 и 3 мкг/мл пуромицина) на такую же среду без FCS. На следующий день, 105 клеток на лунку высевали в 96-луночный планшет в 50 мкл среды, а затем BDNF добавляли гибридные белки в концентрациях от 0,02 нМ до 115 нМ в 50 мкл среды. После инкубации в течение 4 ч при 37°С, 7,5% CO2, клетки инкубировали в течение 30 мин при комнатной температуре и в каждую лунку добавляли 100 мкл реагента BrightGlo Luciferase Assay reagent (Promega). Люминисценцию мчитывали после 5-минутной инкубации, используя планшетный ридер Tecan (время интегрирования 100 мс).
Определение биологической активности BDNF-содержащих гибридных белков в анализе роста невритов SH-SY5Y
Биологическую активность BDNF-вариантов и BDNF-содержащих гибридных белков определяли с помощью анализа роста нейритов с использованием клеток человеческой нейробластомы SH-SY5Y. Вкратце, клетки SH-SY5Y высевали в 96-луночный планшет с плотностью 4000 клеток на лунку в нормальной ростовой среде (среда Хэма F12, 1х заменимые аминокислоты (PAN), 10% FCS, 2 мМ L-глутамина, 1х пируват натрия (PAN)) с добавлением 10 мкМ ретиноевой кислоты (Sigma), чтобы индуцировать дифференцировку нейронов. Через три дня среду заменяли ростовой среды, содержащей различные концентрации BDNF-гибридных белков. После трех дополнительных дней клетки фиксировали с использованием 4% параформальдегида в PBS в течение 10 мин при комнатной температуре, промывали, быстро пермеабилизовали (0,1% Тритон-Х-100), блокировали 1% BSA в PBS и окрашивали на анти-бета-тубулиновую иммунореактивность с использованием антитела TuJ1 (Covance) в разведении 1:1000 в PBS/1% BSA, с последующими тремя промываниями и инкубацией с Alexa-488-меченным противокозлиным антителом (Invitrogen). Число невритов определяли путем флуоресцентной микроскопии, оценивая одно поле зрения на лунку.
Определение связывающей активности гибридных белков, содержащих антитело/фрагмент антитела, с помощью FACS
Связывающая активность BDNF-гибридных белков с соответствующими целевыми рецепторами (рецептор трансферрина, IGF-1R) определяли с помощью FACS. Клетки, экспрессирующие соответствующий рецептор (мышиный рецептор трансферрина: мышиные эмбриональные фибробласты MEF-1; IGF-1R: фибробласты 3Т3, стабильно трансфицированные человеческим IGF-1R) собирали из их ростовых сред, промывали PBS и ресуспендировали в буфере FACS (PBS + 5% FCS; 100 мкл, содержащие 3×105 клеток на лунку в 96-луночном круглодонном планшете). Первичное антитело (в зависимости от используемого BDNF-гибридного белка, например, противочеловеческий Fab (Jackson ImmunoResearch) или анти-His6-антитело (Roche)) добавляли в концентрации 1-10 мкг/мл и клетки инкубировали в течение 2 ч на льду. После трех промываний буфером FACS связанное антитело обнаруживали с использованием РЕ-меченого вторичного антитела (Jackson ImmunoResearch, 1:5000 - 1:10000) в течение одного часа на льду. Клетки снова промывали и измеряли среднюю флуоресценцию на цитометре FACS Canto (Becton-Dickinson).
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКОВ В ДВУХЦЕПОЧЕЧНОЙ ФОРМЕ С ДИСУЛЬФИДНЫМ МОСТИКОМ | 2006 |
|
RU2412253C2 |
| СПОСОБ ПОЛУЧЕНИЯ И ОТБОРА МОЛЕКУЛ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2646159C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2607038C2 |
| СИНТЕТИЧЕСКИЕ ВАРИАНТЫ ГИПЕРГЛИКОЗИЛИРОВАННОГО ПРОТЕАЗО-РЕЗИСТЕНТНОГО ПОЛИПЕПТИДА, ПЕРОРАЛЬНЫЕ КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ ТАКИХ ВАРИАНТОВ | 2005 |
|
RU2392963C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |
| ГЛИКОЗИЛИРОВАННЫЕ КОНЪЮГАТЫ МОЛЕКУЛ С ПОВТОРЯЮЩИМСЯ МОТИВОМ | 2010 |
|
RU2574201C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2011 |
|
RU2573588C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2015 |
|
RU2748026C2 |
| БАКТЕРИЯ Е.coli, ПРОДУЦИРУЮЩАЯ ГЕТЕРОЛОГИЧНЫЙ ПОЛИПЕПТИД, И СПОСОБ ПРОДУЦИРОВАНИЯ ПОЛИПЕПТИДА | 2001 |
|
RU2287574C2 |
| НАПРАВЛЕННЫЕ ТЕРАПЕВТИЧЕСКИЕ ХИМЕРНЫЕ БЕЛКИ ЛИЗОСОМАЛЬНЫХ ФЕРМЕНТОВ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2680581C2 |






















































































































































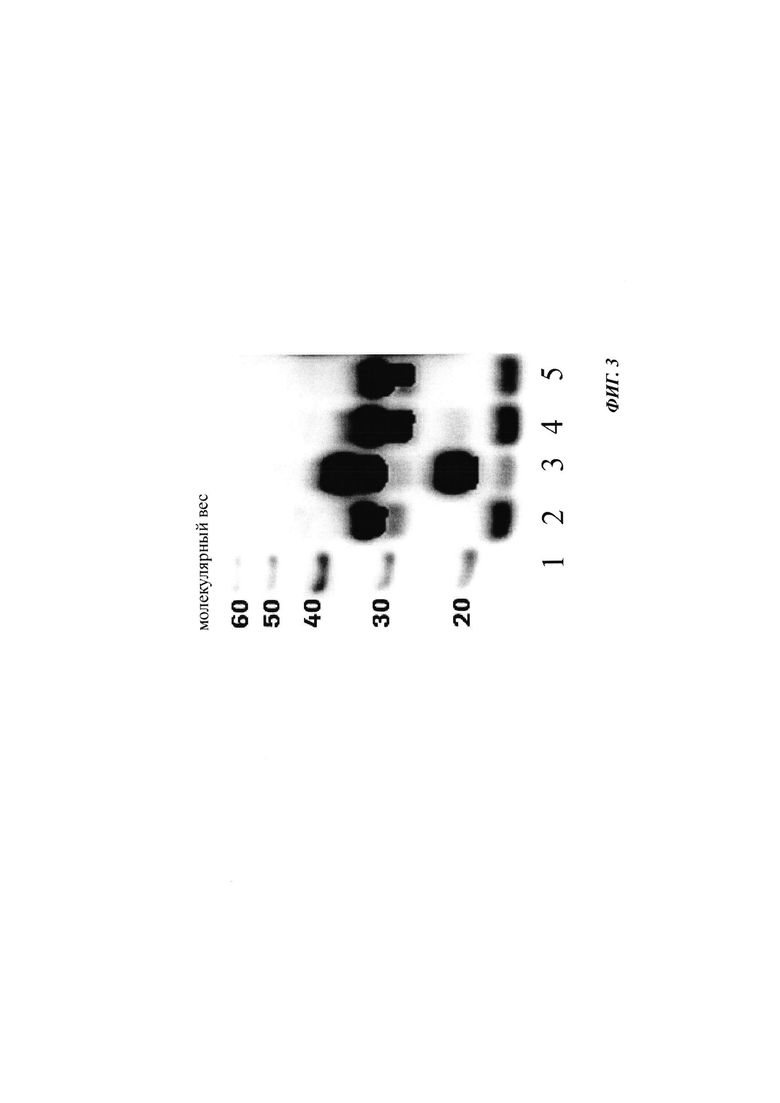
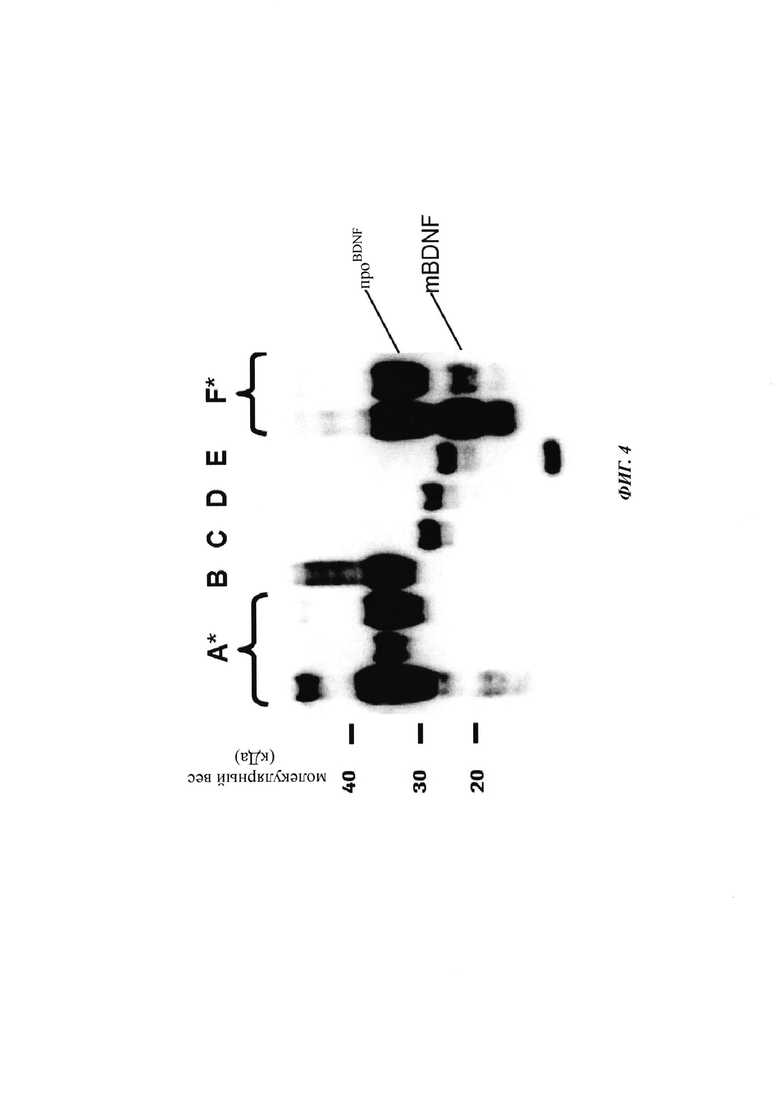
Изобретение относится к области биотехнологии, конкретно к способам получения рекомбинантного полипептида нейротрофина посредством расщепления прополипептида нейротрофина IgA-протеазой, где эндогенный сайт ферментативного расщепления между просегментом и полипептидом нейротрофина заменен сайтом расщепления IgA-протеазой. Изобретение позволяет высокоэффективно получать активные полипептиды фактора роста нервов (NGF), нейротрофического фактора головного мозга (BDNF), нейротрофина 3 (NT-3) и нейротрофина 4 (NT-4). 2 н. и 1 з.п. ф-лы, 4 ил., 22 табл., 8 пр.
1. Способ получения рекомбинантного полипептида нейротрофина, включающий следующие этапы:
- культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую полипептид нейротрофина в виде прополипептида, состоящего из просегмента и полипептида, в котором эндогенный сайт ферментативного расщепления между просегментом и полипептидом заменен сайтом расщепления IgA-протеазой, где указанная клетка млекопитающего экспрессирует указанный прополипептид;
- добавление lgA-протеазы в культивационную среду с расщеплением прополипептида на просегмент и полипептид;
- извлечение рекомбинантного полипептида из культивационной среды и, таким образом, получение рекомбинантного полипептида нейротрофина,
при этом указанная клетка млекопитающего не является эмбриональной клеткой человека.
2. Способ получения рекомбинантного полипептида нейротрофина, включающий следующие этапы:
- культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую полипептид нейротрофина в виде прополипептида, состоящего из просегмента и полипептида, в котором эндогенный сайт ферментативного расщепления между просегментом и полипептидом заменен сайтом расщепления IgA-протеазой, где указанная клетка млекопитающего экспрессирует указанный прополипептид;
- извлечение прополипептида из культивационной среды или клетки;
- обработка прополипептида IgA-протеазой с расщеплением прополипептида на просегмент и полипептид и, таким образом, получение рекомбинантного полипептида нейротрофина,
при этом указанная клетка млекопитающего не является эмбриональной клеткой человека.
3. Способ по п. 1 или 2, где нейротрофин выбран из фактора роста нервов (NGF), нейротрофического фактора головного мозга (BDNF), нейротрофина 3 (NT-3) и нейротрофина 4 (NT-4).
| LO K | |||
| M | |||
| et al., High level expression and secretion of Fc-X fusion proteins in mammalian cells, Protein engineering, 1998, V | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| WO 2008025527, 06.03.2008 | |||
| WO 2009126616, 15.10.2009 | |||
| KJELDSEN T | |||
| et al., A removable spacer peptide in an α-factor-leader/insulin precursor fusion protein improves processing and concomitant yield of | |||

