Изобретение относится к области генной инженерии, индустриальной биотехнологии и представляет собой способ получения рекомбинантного белка ORF2 цирковируса свиней второго типа в форме вирусоподобных частиц.
Настоящее изобретение включает метод получения и очистки вирусоподобных частиц белка ORF2 цирковируса свиней второго типа при использовании штамма метилотрофных дрожжей Komagataella phaffii (Pichia pastoris) BG11/pPic-CV2-7, несущего плазмиду pPic-CV2, содержащую оптимизированную нуклеотидную последовательностью гена белка ORF2 под контролем сильного индуцибельного AOX1 промотора, обеспечивающую синтез и секрецию белка ORF2 цирковируса свиней второго типа.
Цирковирус свиней 2-го типа относится к ДНК-вирусам семейства Circoviridae, содержит одноцепочную ковалентную ДНК и является основным возбудителем мультисистемного истощения свиней [1]. Вирус преимущественно поражает лимфоидные ткани, что приводит к истощению лимфоидов и подавлению иммунитета у свиней. С точки зрения биотехнологии наибольший интерес вызывает капсидный белок, отвечающий за способность вируса прикрепляться к рецепторам поверхности клеток. Этот белок, кодируемый последовательностью нуклеотидов открытой рамки считывания 2 (ORF2), благодаря своим иммуногенным свойствам, является перспективной мишенью при производстве вакцин от цирковируса второго типа [2].
Несмотря на то, что большинство вакцинных препаратов для животных разработаны по традиционным технологиям и представляют собой либо аттенуированный, либо убитый вирус, в настоящее время активно исследуются и создаются новые субъединичные вакцины. Одним из лидеров в этой области стали вакцины на основе вирусоподобных частиц (ВПЧ), поскольку присущие им свойства - стабильность, легкость в использовании и полная безопасность - обуславливают их неоспоримые преимущества и высокую эффективность [3].
В настоящее время на международном рынке доступны коммерческие вакцины, основанные на экспрессии антигена белка ORF2 цирковируса свиней второго типа [4]: препараты Circumvent PCV (Intervet, Нидерланды) и Ingelvac CircoFLEX (Boehringer Ingelheim, Германия), полученные в бакуловирусах; препараты Fostera PCV2 (Pfizer, США), Porcilis PCV (MSD Animal Health, Нидерланды) Circovac (Merial, Франция), Suvaxyn PCV (Zoeitis, Нидерланды), содержащие инактивированный антиген ORF2 цирковируса второго типа или инактивированный вирус PCV2 в качестве антигена.
Несмотря на наличие эффективных импортных вакцин против цирковирусной инфекции второго типа, высокая стоимость препаратов значительно ограничивает их использование в животноводстве, нанося существенный экономический эффект. Создание эффективной рекомбинантной вакцины от цирковируса свиней второго типа является легитимной задачей отечественной биотехнологии.
Известны способы получения рекомбинантного антигена вирусного капсида ORF2 цирковируса свиней второго типа и его модификаций путем экспрессии в прокариотической системе бактерий E. Сoli [5], [6], Bacillus sp. [7]. Авторы сообщают о достаточном уровне экспрессии и возможности использования белка ORF2 в качестве антигена для создания вакцин. Существенным недостатком метода является сложная процедура очистки, рефолдинга и сборки вирусоподобных частиц целевого белка, поскольку он экспрессируется в тельцах включения в мономерной форме.
Известен способ применение белка ORF-2 цирковируса свиней 2 типа для производства вакцины для защиты положительных по антителам материнского происхождения (MDA-положительных) поросят от инфекции PCV-2. Вакцина по изобретению может содержать рекомбинантный белок ORF-2, который получают экспрессией в бакуловирусной системе в клетках насекомых, указанный бакуловирусный экспрессионный вектор содержит последовательность гена ORF-2 PCV-2 под контролем промотора р10. Преимущество изобретения заключается в защите от инфекции MDA-положительных поросят [8].
Известно применение иммуногенной композиции, которая содержит по меньшей мере 2 мкг рекомбинантного белка PCV2 ORF2 полученного в бакуловирусной системе экспрессии для получения лекарственного средства для индукции иммунного ответа против PCV2, для уменьшения тяжести клинических симптомов, связанных с инфекцией PCV2, или для профилактики инфекции PCV2, где лекарственное средство вводят по одной дозе на свинью [9]. Недостатками бакуловирусной системы экспрессии является сложная процедура выделения целевого белка и его высокая себестоимость.
Известно о получении рекомбинантного белка ORF2 цирковируса свиней второго типа в дрожжах Saccharomyces cerevisiae, авторы сообщаю о внутриклеточной локализации целевого белка [10], [11]. Вирусоподобные частицы белка ORF2 выделяли при использовании метода ультрацентрифугирования в градиенте сахарозы (10-40%), так же использовали полигистидиновую вставку. Авторы сообщают о получении иммунохимически активных, гомогенных по форме и размерам вирусоподобных частиц белка ORF2, способных индуцировать моноклональные тела к цирковирусной инфекции свиней второго типа.
Известно о создании штамма-продуцента белка ORF2 цирковируса свиней второго типа в дрожжах K. phaffii GS115 под контролем AOX1 промотора, способных к внутриклеточной продукции целевого белка на уровне 174 мг/л [12], [13], [14]. Полученным после разрушения клеток экстрактом (содержание белка ORF2 составило 80 мкг) проводили иммунизацию поросят. В работе показано появление специфических антител к белку ORF2 через 7 дней после повторной иммунизации, высокий уровень антител держался в течение двух месяцев. Однако, культуральная жидкость не подвергалась дополнительной очистке и не может использоваться в качестве конечной формы вакцинного препарата.
Метилотрофные дрожжи K. phaffii считаются универсальными продуцентами секреторных белков в различных отраслях биотехнологии [15]. Такое широкое применение K. phaffii в биотехнологии связано с высокой продуктивностью, низкой аллергенностью, возможностью посттрансляционных модификаций белков, низкой себестоимостью и относительной простотой культивирования. В последнее время появились публикации, касающихся активного использования дрожжей K. phaffii для гетерологичной экспрессии и получения вирусоподобных частиц [16].
В качестве штамма-реципиента для продукции рекомбинантного белка ORF2 цирковируса свиней второго типа нами были выбраны метилотрофные дрожжи K. phaffii BG11.
Для экспрессии гетерологичных белков в K. phaffii активно применяется сильный промотор метанол-индуцированной алкогольоксидазы (AOX1) [17]. Тот факт, что при росте на метаноле индукция AOX1 промотора может достигать 1000-кратного уровня, а доля общего растворимого белка может увеличиваться до 30% [17], обусловило выбор AOX1 промотора для создания системы экспрессии белка ORF2 цирковируса свиней второго типа в метилотрофных дрожжах K. phaffiii BG11.
Целью изобретения являлось создание способа получения вирусоподобных частиц рекомбинантного белка ORF2 цирковируса свиней второго типа при использовании дрожжей K. phaffii BG11/pPic-CV2-7, способных секретировать белок ORF2 в культуральную жидкость.
Поставленная задача достигается получением штамма дрожжей K. phaffii BG11/pPic-CV2-7, который содержит интеграционный вектор pPic-CV2, несущий ген белка ORF2 цирковируса свиней второго типа под контролем высокоэффективного индуцибельного AOX1 промотора, и способен секретировать целевой белок в культуральную жидкость с образованием вирусоподобных частиц, разработкой метода выделения и очистки капсидной формы белка ORF2.
Техническим результатом заявленного изобретения является способ получения вирусоподобных частиц рекомбинантного белка ORF2 цирковируса свиней второго типа, которые, сохраняя правильную структуру белка оболочки ORF2, способного формировать иммунохимически активный эпитоп, необходимый для образования эффективного иммунного ответа, являются перспективным агентом для создания вакцины против цирковирусной инфекции второго типа у свиней.
Штамм K. phaffii BG11/pPic-CV2-7 - продуцент рекомбинантного белка ORF2 цирковируса свиней второго типа, был получен в результате генно-инженерной модификации дрожжей K. phaffii BG11 (aox1Δ (MutS)), в которые методом электропорации вводился интегративный вектор pPic-CV2-7. Изобретение обеспечивает получение стабильного, имуннологически активного рекомбинантного белка ORF2 цирковируса свиней второго типа в форме вирусоподобных частиц.
Способ продукции рекомбинантных вирусоподобных частиц цирковируса свиней второго типа, состоящих из рекомбинантного белка ORF2, заключается в культивировании штамма-продуцента Komagataella phaffii BG11/pPic-CV2-7, созданного путем трансформации плазмидой pPic-CV2, и являющегося продуцентом рекомбинантного белка ORF2 под контролем сильного индуцибельного AOX1 промотора, с последующим выделением и очисткой рекомбинантного ORF2.
Плазмида pPic-CV2 по п. 1 длиной 9975 п. о., определяющая синтез рекомбинантного ORF2, состоит из элементов (Рисунок 1):
- селективный маркер устойчивости к ампицилину для отбора трансформантов в клетках Е.coli (8805-9665, 861 п. о.);
- селективный маркер канамицина KanR для отбора трансформантов дрожжей Komagataella phaffii (5627-6442, 816 п. о.);
- маркер PpHis4 для селекции на минимальной среде - ген гистидинол дегидрогеназы (2679-5213, 2535 п. о.);
- сайт начала репликации плазмидной конструкции ori (8046-8634, 589 п. о.);
- сигнальный пептид α-фактора, (949-1215, 267 п. о.), обеспечивающий эффективную секрецию гетерологичных белков;
- кодон-оптимизированная для Komagataella phaffii нуклеотидная последовательность гена белка ORF2 цирковируса свиней второго типа (1222-1920, 699 п. о.);
- промотор AOX1 (2-937, 936 п. о.);
- терминатор транскрипции AOX1 (2020-2266, 247 п. о.)
- локус интеграции AOX1 (6821-7577, 757 п. о. и 209-937, 729 п. о.), позволяющий интеграцию целевого белка без нарушения интегративной области нативного гена. В качестве потенциального локуса интеграции могут выступать другие локусы, в том числе HIS4, 18S, ENO1.
Функциональная карта интегративного вектора pPic-CV2 представлена на Рисунке 1. Нуклеотидная последовательность синтетического гена, кодирующего белок ORF2 цирковируса свиней второго типа (SEQ ID NO 1), а также аминокислотная последовательность белка ORF2 (SEQ ID NO 2).
Для создания интеграционной кассеты использовался вектор pPIC9K (Invitrogen). Синтезированный в оптимальных кодонах для дрожжей Komagataella phaffii ген белка ORF2 цирковируса свиней второго типа, фланкированный сайтами эндонуклеаз рестрикции BstSNI - NotI, переносили на плазмиду pPIC9K, предварительно обработанную рестриктазами BstSNI - NotI. Клоны E.coli выращивали и отбирали методом рестрикции эндонуклеазами BstSNI - NotI по наличию фрагмента 716 п. о. В результате был получен интеграционный вектор pPic-CV2.
Трансформацию дрожжей K. phaffii BG11 вектором pPic-CV2 осуществляли методом электропорации согласно протоколу [18]. Перед процедурой электропорации плазмида pPic-CV2 была линеаризована рестриктазой SacI. Трансформантов отбирали на селективной среде YPD, содержащей генетицин (4,0 мг/мл).
Полученные клоны анализировали на наличие целевой вставки методом полимеразно-цепной реакции при использовании праймеров SEQ ID NO 3 и SEQ ID NO 4 по наличию фрагмента 745 п. о.
Отобранные клоны дрожжей K. phaffii BG11/pPic-CV2 культивировались в 100 мл среды BMMY в колбах при температуре 30°С, 250 об/мин в течение 72 часов. Индукция поддерживалась добавлением метанола до 0,5% по объёму каждые 24 часа. Анализ целевого белка в культуральной жидкости проводился методом дот-блоттинга согласно протоколу [19]. Полученный штамм дрожжей K. phaffii BG11/ pPic-CV2-7 - продуцент рекомбинантного белка ORF2 цирковируса свиней второго типа, использовали для наработки целевого белка.
Высокоплотное культивирование дрожжей K. phaffii BG11/ pPic-CV2-7 - проводили в 2 л ферментерах BIOSTAT B+ (Sartorius, Германия) на среде BSM согласно протоколам Invitrogen. В качестве источника углерода на первой стадии ферментации использовали глицерин (40 г/л), вторую стадию ферментации начинали только после полной утилизации углерода в среде. Перед началом индукции в ферментер добавляли сорбитол (2%), индукцию метанолом проводили дробно (концентрация в среде не превышала 1%). Утилизацию метанола отслеживали по изменению скорости потребления кислорода, следующую порцию подпитки добавляли только после полной утилизации предыдущей. Ферментацию проводили в течение 135 ч., конечная оптическая плотность культуральной жидкости OD600 после окончания ферментации составила 260.
Определение содержания белка ORF2 в культуральной жидкости проводили методом денситометрического сканирования полиакриламидных гелей после электрофоретического разделения белков в денатурирующих условиях. Выход секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа в результате культивирования штамма дрожжей K. phaffii BG11/ pPic-CV2-7 составил 80 мг/л.
Выделение белка ORF2 из культуральной жидкости дрожжей проводили методом тангенциальной ультрафильтрации на лабораторной системе фильтрации (BioTechno, Россия). Культуральная жидкость последовательно пропускалась через фильтры с пористостью 1,4мкм - собирали пермеат; 300 кДа и 100 кДа, - собирали ретентант. Анализ ретентанта, полученного после фильтрации через мембрану с пористостью 300 кДа, методами электрофореза в денатурирующих условиях и MALDI-TOF масс-спектрометрии (Bruker, Германия) показал, что рекомбинантный белок ORF 2 цирковируса свиней второго типа представлен на электрофореграмме в виде полос с молекулярной массой 30, 45 и 66 кДа (Рисунок 2). Такой широкий диапазон молекулярных масс обусловлен способностью вирусного белка капсида формировать олигомерные структуры даже в денатурирующих условиях[20]. Тот факт, что белки с молекулярной массой в диапазоне 30-66 кДа не проходят сквозь фильтр с пористостью 300 кДа, указывает на капсидную форму белка. Данная гипотеза была подтверждена при помощи аналитического ультрацентрифугирования в градиенте сахарозы (10-40%) фракций до и после прохождения фильтра 300 кДа.
Полученный после 100кДа фильтра ретентант наносили на колонку с сорбентом DEAE-Sepharose Fast Flow (GE-Healthcare, США). Анализ фракций методом дот-блоттинга показал, что сформировавшиеся вирусоподобные частицы не связывались с хроматографическим сорбентом, тогда как остатки нуклеиновых кислот связывались с сорбентом и элюировались подачей 1M NaCl. Содержащие ORF2 белок фракции концентрировали на фильтре 15 кДа.
Представленная схема очистки, сочетающая тангенциальную ультрафильтрацию (на фильтрах 1,4 мкм -целевой пермеат, 300 кДа, 100 кДа, 15 кДа - целевой ретентант) и анионобменную хроматографию на DEAE Sepharose Fast Flow позволила получить целевой белок с чистотой более 90%, выход белка составил 20-25%. Анализ полученного препарата на трансмиссионном электронном микроскопе JEM-1400 (JEOL, Япония) подтвердил формирование вирусоподобных частиц (Рисунок 3) размером 26-30 нм, что сопоставимо с размером нативных частиц цирковируса свиней второго типа.
Пример №1. Создание интегративного вектора pPic-CV2
Нуклеотидная последовательность гена ORF2 цирковируса свиней второго типа, синтезированная в оптимальных для K. phaffii BG11 кодонах (SEQ ID NO 1), была фланкирована сайтами эндонуклеаз рестрикции BstSNI - NotI. Ген ORF2 цирковируса свиней второго типа по сайтам рестрикции BstSNI - NotI переносили на интегративный вектор pPIC9K, предварительно обработанный этими рестриктазами. Лигазной смесью трасформировали клетки E. coli, целевые клоны отбирали методом рестрикции эндонуклеазами по наличию фрагмента 716 п. о. после обработки рестриктазами BstSNI - NotI. В результате был получен интегративный вектор pPic-CV2 (Рисунок 1).
Пример №2. Трансформация дрожжей K. phaffii штамма BG11 интегративным вектором pPic-CV2
Для трансформации дрожжей K. phaffii BG11 использовали 4 мкг предварительно линеаризованного эндонуклеазой рестрикции SaсI интегративного вектора pPic-CV2. Трансформацию осуществляли методом электропорации согласно протоколу [18]. Целевые клоны отбирали на чашках Петри с селективной средой YPD, содержащей генетицин (4,0 мг/мл). После высева на чашки Петри добивались проращивания отдельных колоний при 30°С 48 ч.
Пример №3. Культивирование и проверка отдельных клонов штамма-продуцента
Полученные клоны анализировали на наличие целевой вставки методом полимеразно-цепной реакции при использовании праймеров SEQ ID NO 3 и SEQ ID NO 4 по наличию фрагмента 745 п. о.
Отобранные в результате полимеразно-цепной реакции клоны выращивались в пробирках с 5 мл среды BMGY при температуре 30°С до OD600=20. Затем клетки собирались центрифугированием (3000×g, 5 минут, 4°С), ресуспендировались в 100 мл среды BMMY и культивировались в колбах при температуре 30°С, 250 об/мин в течение 72 часов. Индукция поддерживалась добавлением метанола до 0,5% по объёму каждые 24 часа. После центрифугирования (3000×g, 5 минут, 4°С) культуральная жидкость отбиралась и хранилась при 4°С. Анализ целевого белка в культуральной жидкости проводился методом дот-блоттинга. Для этого образцы культуральной жидкости (1-5 мкл) наносились на нитроцеллюлозную мембрану (Boster Biological Technology, США). В качестве стандартного образца использовали вакцинный препарат Ингельвак ЦиркоФЛЕКС (Boehriger Ingelheim, Германия). Процедуру дот-блоттинга проводили согласно протоколу [19]. В качестве первичных антител использовали поликлональные кроличьи антитела к цирковирусу свиней второго типа (GeneTex, США, Cat. No. GTX128121). В качестве вторичных антител использовали антикроличьи антитела козы, конъюгированные с пероксидазой хрена HRP (GeneTex, США, GTX213110-01). Для визуализации активности пероксидазы хрена использовали раствор, содержащий 0,5 мг/мл 3,3’-диаминобензидина (Sigma, Германия) в 50 мМ буфере Трис-HCl и 0,01% перекиси водорода (добавляется непосредственно перед проявлением). Развитие цветной окраски дожидались в течение 3 мин, затем мембрану двукратно промывали 100 мМ натрий-фосфатным буфером (рН 7,2), содержащим 150 мМ хлорида натрия и 0,05% Tween-20, для остановки развития окраски.
Полученный штамм дрожжей K. phaffii BG11/ pPic-CV2-7 - продуцент рекомбинантного белка цирковируса свиней второго типа с максимальной продуктивностью согласно данным дот-блоттинга, использовали для наработки и очистки целевого белка.
Пример №4. Культивирование штамма дрожжей K. phaffii BG11/ pPic-CV2-7-продуцента секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа
Высокоплотное культивирование штамма K. phaffii BG11/ pPic-CV2-7 -продуцента секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа проводили в 2 л ферментерах BIOSTAT B+ (Sartorius, Германия) на среде BSM, содержащей согласно протоколам Invitrogen. Поскольку белок ORF2 цирковируса свиней второго типа содержит участки с высоким содержанием аминокислотных остатков аргинина, в среду для культивирования дополнительно добавляли 1,5 г/л аргинина. В качестве пеногасителя при культивировании использовался Софэксил-1520 («Софекс-Силикон», Россия) в концентрации 0,5 г/л. В качестве титрующих агентов использовались 10% р-р аммиака и 5% р-р H2SO4.
Посевной материал выращивался в 150 мл среды в колбе в течение 24 ч. при температуре 28оС и 250 об/мин на среде, содержащей 10 г/л дрожжевого экстракта, 20 г/л пептона, 3,4 г/л YNB и 20 г/л глицерина.
Первую стадию культивирования штамма-продуцента проводили в условиях 30оС, рН 4,0, 700 об/мин, в качестве источника углерода использовали глицерин (40 г/л). Вторую стадию культивирования штамма-продуцента начинали после полной утилизации глицерина и проводили при 26оС, рН 5,5, 700 об/мин, перед началом индукции метанолом в ферментер добавляли сорбитол (2%). Метанол добавляли дробно - так, чтобы его концентрация в среде не превышала 1%. Утилизацию метанола отслеживали по изменению скорости потребления кислорода, следующую порцию подпитки добавляли только после полной утилизации предыдущей.
Ферментацию проводили в течение 135 ч, конечная оптическая плотность культуральной жидкости OD600 после окончания ферментации составила 260.
Пример №5. Оценка выхода секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа в результате культивирования штамма дрожжей K. phaffii BG11/ pPic-CV2-7
Определение содержания целевого белка в культуральной жидкости проводили методом денситометрического сканирования полиакриламидных гелей после электрофоретического разделения белков в денатурирующих условиях. Изображение геля (Рисунок 2) документировано с помощью прибора ChemiDoc Imaging System (Bio-Rad, США) и обработано с помощью программы обработки Image Lab Software (Bio-Rad). Анализу подвергали отдельные дорожки геля таким образом, что программа обсчитывала пропорциональное содержание (в %) индивидуальных полипептидных полос. Полученное значение содержания целевого белка представляет процент его содержания суммарно от всех остальных белков, т.е. процент содержания от суммарного клеточного белка. Количество целевого белка оценивали, проводя сравнительный анализ с белками-стандартами (бычий сывороточный альбумин) в известном количестве после электрофоретического анализа.
Выход секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа в результате культивирования штамма дрожжей K. phaffii BG11/pPic-CV2-7 составил 80 мг/л.
Пример №6. Выделение и очистка секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа, получение вирусоподобных частиц
В качестве первой ступени выделения секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа была выбрана очистка методом тангенциальной ультрафильтрации на лабораторной системе фильтрации (BioTechno, Россия). Культуральную жидкость дрожжей штамма K. phaffii BG11/ pPic-CV2-7 (1,8 л), полученную после ферментации, центрифугировали (1500 g, 30 мин, 4оС). Полученный супернатант подавали на керамический фильтр с пористостью 1,4 мкм, промывали двукратным объемом 50 мМ натрий-фосфатного буфера (pH 7,2), содержащего 150 мМ NaCl. Полученный пермеат подавали на фильтр 300 кДа, промывали двукратным объемом 50 мМ натрий-фосфатного буфера (pH 7,2), содержащего 150 мМ NaCl. Анализ полученного ретентанта методом электрофореза в денатурирующих условиях показал, что белки с молекулярной массой 30-66 кДа не проходят сквозь фильтр в 300 кДа, что указывает на формирование в ретентанте вирусоподобных частиц с молекулярной массой превышающей 300 кДа.
Это было также подтверждено при помощи аналитического ультрацентрифугирования XL-1 (Beckman Coulter, США) 50000 RPM An-60 Ti ротор при 10°C в градиенте 10-40% сахарозы фракции до и после прохождения фильтра 300 кДа.
Для удаления примесных белков с молекулярной массой менее 100 кДа полученный ретентант пропускали через фильтр 100 кДа, промывали двукратным объемом 50 мМ натрий-фосфатного буфера (pH 7,2), содержащего 150 мМ NaCl. Полученный после 100кДа фильтра ретентант наносили на колонку с сорбентом DEAE-Sepharose Fast Flow (GE-Healthcare, США). Очистка производилась при скорости потока 0,5 мл/мин. Анализ фракций методом дот-блоттинга показал, что сформировавшиеся вирусоподобные частицы не связывались с хроматографическим сорбентом. Тогда как остатки нуклеиновых кислот связывались с сорбентом и элюировались подачей 1M NaCl.
Содержащие ORF2 белок фракции концентрировали на фильтре 15 кДа, промывали 2-кратным объемом 50 мМ натрий-фосфатного буфера (pH 7,0), содержащего 150 мМ NaCl. Представленная схема очистки, сочетающая тангенциальную ультрафильтрацию (на фильтрах 1,4 мкм -целевой пермеат, 300 кДа, 100 кДа, 15 кДа - целевой ретентант) и анионобменную хроматографию на DEAE-Sepharose Fast Flow позволила получить целевой белок с чистотой более 90%, выход белка составил 20-25%. Анализ полученного белка методом MALDI-TOF масс-спектрометрии (Bruker, Германия), обнаружил 90% покрытие последовательности белка, тогда как детекция целевого белка методом дот-блоттинга указывала на правильную структуру целевого белка, способного формировать иммунохимически активный эпитоп, необходимый для образования эффективного иммунного ответа.
Анализ полученного препарата на трансмиссионном электронном микроскопе JEM-1400 (JEOL, Япония) подтвердил формирование вирусоподобных частиц (Рисунок 3) размером 26-30 нм, что сопоставимо с размером нативных частиц цирковируса свиней второго типа.
Список литературы
[1] G. M. Allan и J. A. Ellis, «Porcine Circoviruses: A Review», J Vet Diagn Invest., т. 12, вып. 1, сс. 3-14, 2000, doi: 10.1177/104063870001200102.
[2] X. J. Meng, «Porcine circovirus type 2 (PCV2): pathogenesis and interaction with the immune system», Annu Rev Anim Biosci., вып. 1, сс. 43-64, 2013, doi: 10.1146/annurev-animal-031412-103720.
[3] G. T. Jennings и M. F. Bachmann, «The coming of age of virus-like particle vaccines», Biol Chem., т. 389, вып. 5, сс. 521-36, 2008, doi: 10.1515/bc.2008.064.
[4] B. R. Trible и R. R. R. Rowland, «Genetic variation of porcine circovirus type 2 (PCV2) and its relevance to vaccination, pathogenesis and diagnosis», Virus Res., т. 164, вып. 1-2, сс. 68-77, 2012, doi: 10.1016/j.virusres.2011.11.018.
[5] P. C. Wu, T. Y. Chen, J. N. Chi, M. S. Chien, и C. Huang, «Efficient expression and purification of porcine circovirus type 2 virus-like particles in Escherichia coli», J Biotechnol., вып. 220, сс. 78-85, 2016, doi: 10.1016/j.jbiotec.2016.01.017.
[6] М. Р. Ди-Алмиэда и др., «Рекомбинантные антигены цирковируса свиней типа 2 (pcv-2) для композиций вакцин, диагностический набор и их применение», RU 2 687 001 C2.
[7] и др., «ORF2 recombinant gene of porcine circovirus type 2 and vaccine composition using the same», KR20190014263A [Онлайн]. Доступно на: https://patents.google.com/patent/KR20190014263A/en?q=(ORF2+recombinant)&oq=ORF2+recombinant
[8] Ф. Руринк и П. В. Вунсел, «Вакцина против рсv-2», RU2389506C2 [Онлайн]. Доступно на: https://patents.google.com/patent/RU2389506C2/ru
[9] М. Эйчмейер, Г. Нитцель, и М. Шеффер, «Иммуногенные композиции PCV2 и способы получения этих композиций», RU2007128943A [Онлайн]. Доступно на: https://patents.google.com/patent/RU2575615C2/en
[10] S. A. Bucarey и др., «The optimized capsid gene of porcine circovirus type 2 expressed in yeast forms virus-like particles and elicits antibody responses in mice fed with recombinant yeast extracts», Vaccine, т. 27, вып. 42, сс. 5781-90, 2009, doi: 10.1016/j.vaccine.2009.07.061.
[11] J. Nainys и др., «Generation in yeast of recombinant virus-like particles of porcine circovirus type 2 capsid protein and their use for a serologic assay and development of monoclonal antibodies», BMC Biotechnol, вып. 14, с. 100, 2014.
[12] «Pichia expression PCV2 (porcine circovirus 2) ORF2 (open reading frame 2) protein and subunit vaccine», CN102115755A [Онлайн]. Доступно на: https://patents.google.com/patent/CN102115755A/en
[13] Y. Tu и др., «High-level expression and immunogenicity of a porcine circovirus type 2 capsid protein through codon optimization in Pichia pastoris», Appl Microbiol Biotechnol., т. 97, вып. 7, сс. 2867-75, 2013, doi: 10.1007/s00253-012-4540-z.
[14] J. G. Silva и др., «Secretory expression of Porcine Circovirus Type 2 capsid protein in Pichia pastoris», J Virol Methods., т. 207, сс. 226-31, 2014, doi: https://doi.org/10.1016/j.jviromet.2014.07.021.
[15] Z. Yang и Z. Zhang, «Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review», Biotechnol Adv., т. 36, вып. 1, сс. 182-195, 2018, doi: 10.1016/j.biotechadv.2017.11.002.
[16] S. de Sá Magalhães и E. Keshavarz-Moore, «P. pastoris (Komagataella phaffii) as a Cost-Effective Tool for Vaccine Production for Low- and Middle-Income Countries (LMICs)», Bioengineering (Basel)., т. 31, вып. 8(9), с. 119, 2021, doi: 10.3390/bioengineering8090119.
[17] A. T. Özçelik, S. Yılmaz, и M. Inan, «Pichia pastoris Promoters», Methods Mol Biol., т. 1923, сс. 97-112, 2019, doi: 10.1007/978-1-4939-9024-5_3.
[18] Lin-Cereghino J., Naranjo C.A., и Lin-Cereghino G.P., «Competent Cell Preparation and Transformation of Pichia pastoris. In: Mapelli, V., Bettiga, M. (eds) Yeast Metabolic Engineering. Methods in Molecular Biology, vol 2513. Humana, New York, NY. https://doi.org/10.1007/978-1-0716-2399-2_7», 2022 г.
[19] M. Taesuji, K. Rattanamas, U. Kulthonggate, T. Mamom, и S. Ruenphet, «Sensitivity and specificity for African horse sickness antibodies detection using monovalent and polyvalent vaccine antigen-based dot blotting», Vet World., т. 15, вып. 12, сс. 2760-2763, 2022, doi: 10.14202/vetworld.2022.2760-2763.
[20] R. L. C. Leung и др., «Monitoring the Disassembly of Virus-like Particles by 19F-NMR», J Am Chem Soc., т. 139, вып. 15, сс. 5277-5280, апр. 2017, doi: 10.1021/jacs.6b11040.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="ORF2.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-12-19">
<ApplicantFileReference>До присвоения номера
заявки</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
учреждение «Федеральный исследовательский центр «Фундаментальные
основы биотехнологии» Российской академии наук»</ApplicantName>
<ApplicantNameLatin>Federal State Institution Federal Research
Centre Fundamentals of Biotechnology of the Russian Academy of
Sciences</ApplicantNameLatin>
<InventorName languageCode="ru">Макарова Анна </InventorName>
<InventorNameLatin>Makarova Anna</InventorNameLatin>
<InventionTitle languageCode="ru">Способ получения секреторного
рекомбинантного белка ORF2 цирковируса свиней второго типа в дрожжах
Komagataella phaffii BG11/ pPic-CV2-7 в форме вирусоподобных
частиц.</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>699</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..699</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q7">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acttacccacgtcgaagataccgtcgaaggcgtcatagaccaagaagtc
atttgggtcaaattttgcgaaggcgtccatggttggttcatccaagacaccgttacaggtggagacgtaa
gaacggaattttcaacaccagattgtccagaaccattggatacactgttaagaagaccaccgttagaacc
ccatcctggaacgttgatatgatgagattcaacattaacgatttcttgccaccaggaggaggatccaacc
cattgaccgttccattcgagtactacagaattagaaaggtcaaggttgaattctggccatgttctccaat
tactcagggagatagaggtgttggatccactgctgttattttggatgataacttcgttaccaaggctaac
gctttgacttacgatccatacgttaactactcttccagacataccattacccagccattctcctaccatt
ccagatacttcactccaaagccagttttggatagaaccattgattacttccagccaaacaacaagagaaa
ccagttgtggttgagattgcagactaccggaaacgttgatcatgttggattgggaactgctttcgaaaac
tctatttacgatcagaactacaacattagaattactatgtacgttcaattcagagagttcaacttgaagg
atccaccattgaacccataa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>232</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..232</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Sus scrofa</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TYPRRRYRRRRHRPRSHLGQILRRRPWLVHPRHRYRWRRKNGIFNTRLS
RTIGYTVKKTTVRTPSWNVDMMRFNINDFLPPGGGSNPLTVPFEYYRIRKVKVEFWPCSPITQGDRGVGS
TAVILDDNFVTKANALTYDPYVNYSSRHTITQPFSYHSRYFTPKPVLDRTIDYFQPNNKRNQLWLRLQTT
GNVDHVGLGTAFENSIYDQNYNIRITMYVQFREFNLKDPPLNP</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>25</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..25</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acttacccacgtcgaagataccgtc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>27</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..27</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q9">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cagtcatgtctaaggcgaattaattcg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
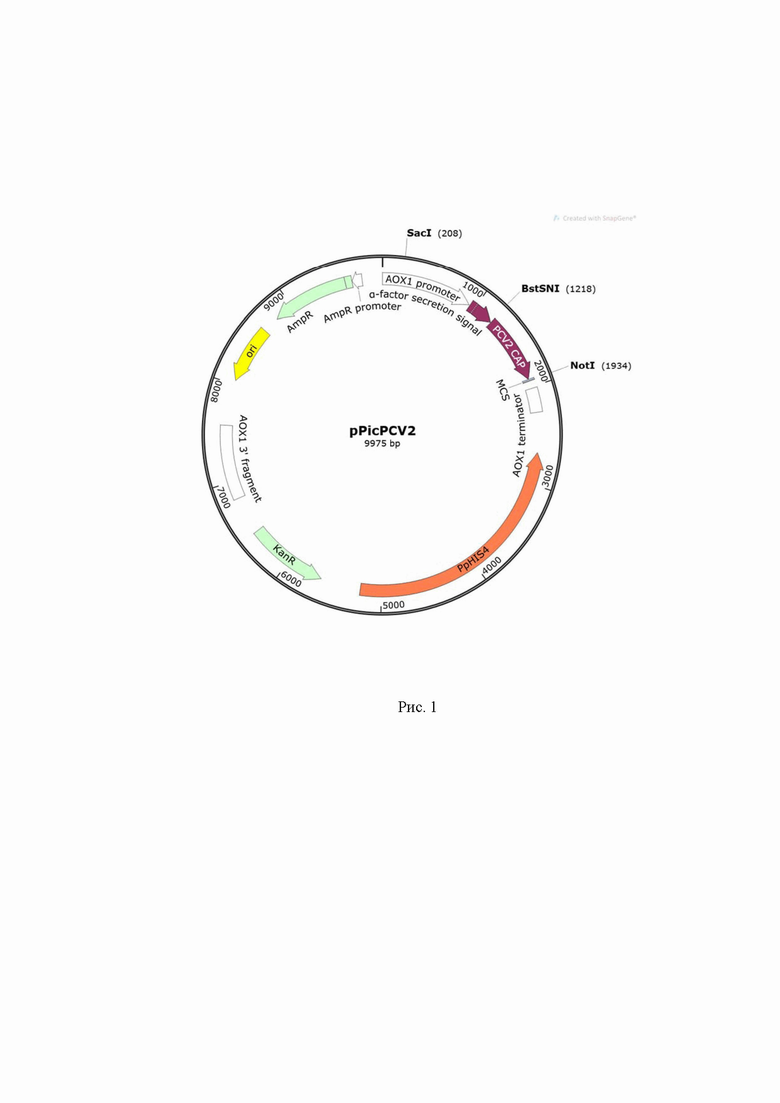

Изобретение относится к области биотехнологии, а именно к плазмиде pPic-CV2, определяющей синтез рекомбинантного ORF2. Плазмида включает маркер устойчивости к ампицилину, селективный маркер канамицина KanR, маркер PpHis4 - ген гистидинол дегидрогеназы, сайт начала репликации плазмидной конструкции ori, сигнальный пептид α-фактора, кодон-оптимизированная для Komagataella phaffii нуклеотидная последовательность гена белка ORF2 цирковируса свиней второго типа, промотор AOX1, терминатор транскрипции AOX1, локус интеграции AOX1. Изобретение эффективно для продукции рекомбинантных вирусоподобных частиц цирковируса свиней второго типа, состоящих из рекомбинантного белка ORF2. 2 н.п. ф-лы, 3 ил., 6 пр.
1. Плазмида pPic-CV2, длиной 9975 п.о., определяющая синтез рекомбинантного ORF2, состоящая из элементов:
- селективный маркер устойчивости к ампицилину для отбора трансформантов в клетках Е.coli 8805-9665, 861 п.о.;
- селективный маркер канамицина KanR для отбора трансформантов дрожжей Komagataella phaffii 5627-6442, 816 п.о.;
- маркер PpHis4 для селекции на минимальной среде - ген гистидинол дегидрогеназы 2679-5213, 2535 п.о.;
- сайт начала репликации плазмидной конструкции ori 8046-8634, 589 п.о.;
- сигнальный пептид α-фактора, 949-1215, 267 п.о., обеспечивающий эффективную секрецию гетерологичных белков;
- кодон-оптимизированная для Komagataella phaffii нуклеотидная последовательность гена белка ORF2 цирковируса свиней второго типа 1222-1920, 699 п.о.;
- промотор AOX1 2-937, 936 п.о.;
- терминатор транскрипции AOX1 2020-2266, 247 п.о.;
- локус интеграции AOX1 6821-7577, 757 п.о. и 209-937, 729 п.о., позволяющий интеграцию целевого белка без нарушения интегративной области нативного гена.
2. Способ продукции рекомбинантных вирусоподобных частиц цирковируса свиней второго типа, состоящих из рекомбинантного белка ORF2, заключающийся в культивировании штамм-продуцента Komagataella phaffii BG11/pPic-CV2-7, созданного путем трансформации плазмидой pPic-CV2 по п.1, и являющегося продуцентом рекомбинантного белка ORF2 под контролем сильного индуцибельного AOX1 промотора, с последующим выделением и очисткой рекомбинантного ORF2.
| CN 104651316 A, 27.05.2015 | |||
| TOME-AMAT, JAIME et al., Secreted production of assembled Norovirus virus-like particles from Pichia pastoris, Microbial cell factories, 2014, v | |||
| Насос | 1917 |
|
SU13A1 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКА ORF2 PCV2b В КЛЕТКАХ НАСЕКОМЫХ | 2018 |
|
RU2779423C2 |
| MAZALOVSKA, M | |||
| et al., (2020) | |||
| Progress in the Production of Virus-Like Particles for Vaccination against Hepatitis E Virus, 2020, | |||

