Область техники, к которой относится изобретение
Первым объектом настоящей заявки на патент является капсидный очищенный рекомбинантный белок PCV-2, полученный в E. coli, в качестве антигенов вакцин и диагностического антигена. Вторым объектом изобретения являются вирусоподобные частицы (ВПЧ) PCV2, полученные с помощью E. coli, в качестве антигена вакцин или диагностического антигена. Третьим объектом настоящей заявки является рекомбинантная вакцина против PCV-2, содержащая очищенные антигены, или рекомбинантные ВПЧ PCV2, полученные в E. coli, к которым добавлен гидроксид алюминия в качестве адъюванта. Четвертым объектом настоящей заявки на патент является набор для ELISA с захватом (антигенов), предназначенный для количественной оценки антигенов PCV-2 в вакцине. Антигены вакцин (объекты изобретения 1 и 2) и вакцина (объект изобретения 3), указанные в настоящем описании, можно применять для иммунизации животных в программах контроля ассоциированных с PCV-2 болезней в общепринятых системах свиноводства, и они являются альтернативой поступающим в продажу вакцинам. Набор ELISA (объект изобретения 4) можно применять для оценки качества коммерческих и/или экспериментальных вакцин против PCV-2. Упомянутые выше продукты можно применять в области фармации или защиты здоровья животных.
Известный уровень техники
Цирковирус свиней (PCV) идентифицирован впервые в 1974 г.в качестве персистентной контаминации, присутствующей в культуре клеток почек свиней. Однако связанная с этим вирусом патогенность для свиней была обнаружена только в конце 90-х годов. Она представляла собой синдром мультисистемного истощения после отъема (PMWS), который впервые был описан в Канаде. Из-за генетических и иммунологических различий между указанным новым PCV, выделенным из животных, вызывающим симптомы PMWS, и указанными контаминантами, полученными из лабораторных клеток, эти два вируса были обозначены как цирковирус свиней-2 (PCV-2) и цирковирус свиней-1 (PCV-1) соответственно (Silva Junior A., Carvalho O.V., Bulos L.H.S., Fietto J.L.R., Moraes M.P., Almeida M.R., Porcine circovirus 2: immunopathogenesis and recent developments in vaccines; World journal vaccine, т. 2, 2012, cc. 96-104).
Поскольку с PCV-2 ассоциирован различный набор синдромов у свиней помимо PMWS, то в 2006 г. Американская ассоциация по ветеринарии свиней (AASV) ввела понятие «ассоциированная с цирковирусом свиней болезнь» (PCVAD) для группы, включающей все эти синдромы. Понятие включает энтериты, недостаточность репродуктивной функции, дерматит и нефропатию свиней, главным образом на фазах роста и заключительного периода откорма. Позднее в 2011 г. был описан новый синдром, характеризующийся острым легочным отеком, который поражает молодых и вакцинированных против PCV-2 животных, при котором уровень смертности составляет примерно 20% (Cino-Ozuna A.G., Henry S., Hesse R., Nietfeld J.C., Bai J., Scott H.M., Rowland R.R.R., Characterization of a new disease syndrome associated with porcine circovirus type 2 in previously vaccinated herds. Journal of Clinical Microbiology, т. 49, 2011, cc. 2012-2016). В настоящее время PCV-2 широко распространен во всем мире и болезни группы PCVAD являются эндемическими заболеваниями в наиболее развитых странах.
Не у всех животных, инфицированных PCV-2, проявляются признаки этих болезней. Таким образом, для точного установления диагноза необходимо обнаружить присутствие соответствующих клинических признаков и присутствие характерных микроскопических повреждений помимо выявления вирусов у животного с помощью лабораторных методик, таких как полимеразная цепная реакция (ПЦР), иммуногистохимия (ИГХ) и некоторых других (Silva Junior A., Carvalho O.V., Bulos L.H.S., Fietto J.L.R., Moraes M.P., Almeida M.R., Porcine circovirus 2: immunopathogenesis and recent developments in vaccines World journal vaccine, т. 2, 2012, cc. 96-104).
Поскольку у больных животных обычно имеет место лимфопения, то часто встречаются вторичные инфекции, вызываемые условно-патогенными организмами, что повышает важность контроля PCVAD. В проведенных в связи с этим исследованиях высказано предположение о том, что развитие PCVAD непосредственно связано с взаимодействием вирусов, а также других условно-патогенных инфекционных агентов с иммунной системой животного, что свидетельствует о многофакторной природе этих болезней (Silva Junior А., Carvalho O.V., Bulos L.H.S., Fietto J.L.R., Moraes M.P., Almeida M.R., Porcine circovirus 2: immunopathogenesis and recent developments in vaccines World journal vaccine, т. 2, 2012, cc. 96-104).
PCV-2 может распространяться в окружающей среде посредством выделений из рта и носа, с экскрементами и кровью. Передача вируса происходит назальным, оральным путем, через экскременты и мочевые пути. Инфекционная вирусная частица является устойчивой к физическим и химическим факторам, и обладает устойчивостью в кислой среде (pH 3), к инактивации нагреванием (120°С в течение 30 мин), замораживанию и воздействию ультрафиолетового света. Дезинфектанты на основе моноперсульфата натрия, перуксусная кислота с пероксидом водорода, соединения, содержащие четвертичные аммониевые соли, гипохлорит натрия или гидроксид натрия обладают эффективностью в отношении снижения количества инфекционных частиц. С другой стороны, продукты, содержащие йод или фенольные соединения, не обладают способностью снижать инфекционность вируса in vitro. Благодаря многофакторной природе PCVAD, ее контроль следует осуществлять с использованием комбинации факторов, эффективность которых доказана, таких как надлежащая практика управления (численностью), рекомендованная хорошо известным руководством «Madec's 20 points», уменьшение стресса у животных, уменьшение смешения партий, адаптация к глобальному проекту по обеспечению доступа к системе охраны здоровья (Health gaps), а также вакцинация и серологический надзор стада (Madec F., Rose N., Grasland В., Cariolet R., Jestin A., Post-weaning multisystemic wasting syndrome and other PCV2-related problems in pigs: a 12-year experience. Transboundary and Emerging Diseases, т. 55, 2008, cc. 273-283).
После обнаружения PMWS и подтверждения того, что PCV-2 является и его этиологическим агентом, в мире начались повторные исследования, посвященные развитию путей контроля указанной болезни, прежде всего с помощью вакцин. Применяли несколько стратегий для создания иммуногенов. К ним относятся вакцины на основе инактивированного вируса, выращенного в клеточной культуре, ДНК-вакцины и субъединичные вакцины, содержащие белок вирусного капсида, полученный в созданных модифицированных клетках насекомых (Silva Junior A., Carvalho O.V., Bulos L.H.S., Fietto J.L.R., Moraes M.P., Almeida M.R., Porcine circovirus 2: immunopathogenesis and recent developments in vaccines World journal vaccine, т. 2, 2012, cc. 96-104).
Структурный белок капсида цирковируса, SEQ ID NO: 04, кодируется открытой рамкой считывания ОРС2, SEQ ID NO: 03, и имеет молекулярную массу примерно 28 кДа и теоретическую изоэлектрическую точку (ИЭТ) примерно 10,7. Поскольку он специфически взаимодействует с рецепторами на клеточной поверхности, то он является наиболее вероятным белком, участвующим в иммунном ответе хозяина. Он имеет сигнал ядерной локализации в аминоконцевой области (первый 41 аминокислотный остаток) с большим количеством положительно заряженных аминокислотных остатков, из которых примерно 40% представляют собой остатки аргинина. Исследования продемонстрировали, что присутствие трех иммуногенных эпитопов на капсидном белке PCV-2 обусловливает создание нейтрализующих антител. Они содержат аминокислотные последовательности, включающие остатки с 47 по 57, с 165 по 200 и последние четыре аминокислотных остатка с С-конца (Lekcharoensuk P., Morozov I., Marsh P.S., Thangthumniyom N., Wajjawalku W., Meng X.J., Epitope mapping of the major capsid protein of type 2 porcine circovirus (PCV2) by using chimeric PCV1 and PCV2. Journal of Virology, т. 78, 2004, cc. 8135-8145).
Вирусный капсидный белок PCV-2 впервые был получен в Escherichia coli Liu с соавторами в 2001 г. в виде полипептидной цепи, слитой со связывающим мальтозу белком (МВР) и гистидиновым хвостом - (MBP)-His8 (Liu Q., Tikoo S.K., Babiuk L.A., Nuclear localization of the ORF2 protein encoded by porcine circovirus type 2. Virology, т. 285, 2001, cc. 91-99). Этот рекомбинантный белок, очищенный с помощью аффинной хроматографии, специфически взаимодействовал как в кроличьей поликлональной сывороткой, предварительно иммунизированной PCV-2, так и с сывороткой свиней, инфицированных вирусом.
Экспрессия больших количеств капсидного белка PCV-2 в клетках Escherichia coli является затруднительной из-за большого количества остатков аргинина в сигнале ядерной локализации и присутствия других аминокислотных остатков, синтезируемых с редких для этого организма кодонов. Другим путем решения проблемы, связанной с низким уровнем экспрессии капсидного белка PCV-2, является применение бактериальных штаммов, сверхэкспрессирующих тРНК редких кодонов, таких как BL21 кодон -плюс-DE3-RIL. Преимущество этого пути состоит в сохранении всех аминокислотных остатков белка (первичная структура является консервативной), которые гарантируют возможность создания антител к эпитопам, присутствующим в области сигнала ядерной локализации. Однако для экспрессии можно использовать и другие бактериальные системы, поступающие в продажу или разработанные компаниями.
В настоящее время экспрессию рекомбинантного Cap-PCV-2 осуществляют главным образом в двух системах: в первой используют клетки насекомых, зараженные рекомбинантным бакуловирусом, а в другой осуществляют индукцию рекомбинантного белка в трансформированных бактериальных системах. В последнем случае система экспрессии, как правило, находится под контролем оператора lac, поэтому для ее индукции обычно в среду для роста добавляют изопропил-β-D-1-тиогалактопиранозид (ИПТГ). Однако для экспрессии в бактериальных системах в продажу поступают несколько векторов различных компаний. Таким образом, другие промоторы, гомологичные или гетерологичные, можно применять также в качестве промоторов, модулируемых арабинозой. Вирусоподобные частицы (ВПЧ) представляют собой структурные белки вирусного капсида, которые имеют структуру, сходную со структурой вирусной частицы дикого типа, формирующей капсид, лишенной генетического материала. Указанные неинфекционные частицы обладают высокой иммуногенностью из-за присутствия структурной формы вирионов (вирусная инфекционная частица). Таким образом, применение ВПЧ в рекомбинантных вакцинах повышает эффективность таких вакцин в результате стимуляции иммунной системы посредством пути, сходного с путем, характерным для вирусной частицы дикого типа. Была продемонстрирована способность капсидных белков PCV-2 образовывать ВПЧ как в эукариотических экспрессионных системах (Bucarey S.A., Noriega J., Reyes P., Tapia C., Saenz L. Zuniga A. Tobar J.A., The optimized capsid gene of porcine circovirus type 2 expressed in yeast forms virus-like particles and elicits antibody responses in mice fed with recombinant yeast extracts. Vaccine, т. 27, 2009, cc. 5781-5790), так и в прокариотических системах (Lou Z.Z., Li Z.Y., Wang G., Li J.Q., Lan X., Li X.R., Yin X.P., Liu J.X., Liu S.D., Prokaryotic expression and potential application of the truncated PCV-2 capsid protein. Virological Sinica, т. 25, 2010, cc. 86-97).
В настоящем изобретении применяли методологию, основанную на бактериальной системе. С ее помощью можно получать антиген в количествах от десятков до сотен миллиграммов на литр среды при индукции в течение нескольких часов. Однако при использовании системы экспрессии, основанной на клетках насекомых, для получения таких количеств требуется несколько дней. Другим преимуществом являются затраты, ассоциированные с таким получением. В бактериальной системе применят более дешевые средства индукции по сравнению с клетками насекомых. При этом, однако, в системах, в которых используются клетки насекомых, после экспрессии не требуется стадии концентрирования и очистки антигена. Однако в настоящем изобретении рекомбинантные антигены для последующего использования выделяют с помощью менее дорогостоящих технологий.
Другим возможным недостатком бактериальной системы является отсутствие пост-трансляционных модификаций капсидного белка, поскольку эта система является прокариотической. Однако при создании настоящего изобретения продемонстрировано, что вирусный капсидный белок при экспрессии в бактериальной системе обладает способностью к самосборке и к усиленному созданию неинфекционных ВПЧ, которые по форме и размеру сходны с вирусными частицами дикого типа, аналогично тому, что имеет место при экспрессии в эукариотических системах.
Другой трудностью, связанной с производством применяемого в вакцинах соединения, содержащего вирусный капсидный белок PCV-2, является его количественная оценка в композиции вакцины. Для поступающих в продажу рекомбинантных вакцин, как правило, в качестве метода количественной оценки используют денситометрию в полиакриламидном геле, применяя в качестве эталона стандартный белок, такой как бычий сывороточный альбумин (БСА). В настоящей заявке описано применение антител к PCV-2 для количественной оценки иммуногенного белка. Указанные антитела специфически взаимодействуют с капсидным белком PCV-2, присутствующим в конкретном образце, благодаря специфическому маркеру на антителе происходит образование сигнала (цвет, радиоактивность и т.д.), пропорционального количеству антигена, присутствующего в образце. Таким образом, количественная оценка происходит при считывании указанного количественного сигнала и путем сравнения с образцом, содержащим очищенный рекомбинантный белок.
Из литературы известно несколько усилителей иммунных ответов и согласно настоящему изобретению их можно применять в сочетании с композициями, указанными в настоящем описании. Предпочтительными являются водные адъюванты, такие как гидроксид алюминия или углеродные полимеры, такие как карбоксиметилцеллюлоза, поскольку они вызывают меньше воспаления и боли, чем масляные адъюванты.
Применяемые в вакцинах соединения, разработанные при создании настоящего изобретения, можно вводить любыми путями, известными из литературы, предпочтительно внутримышечно.
При осуществлении поиска в патентных базах данных были обнаружены патентные документы, касающиеся процессов получения иммуногенов для свиней, однако ни в одном из этих патентов не применяли продукты или процессы, родственные объекту настоящего патента. При указанном поиске был обнаружен патентный документ PI 0412855-9 от 07/26/2004, озаглавленный «КОМПОЗИЦИИ ВАКЦИН (VACCINE FORMULATIONS)», который относится к «новой эмульсии масло-вода с повышенной стабильностью в присутствии бактериальных или вирусных суспензий, прежде всего концентрированных и неочищенный или мало очищенных», в котором описан раствор, содержащий инактивированный PCV-2. Объект настоящего изобретения отличается от описанного в указанном патенте, поскольку он сфокусирован на применение рекомбинантного белка, продуцируемого в бактериальной системе, в дополнение к предложенным конкретным новым процедурам выделения и количественной оценки указанного антигена.
Другая обнаруженная заявка на патент представляет собой заявку под номером BRPI0615862-5 от 09/08/2006, озаглавленную «ВАКЦИНА ПРОТИВ PCV-2 И СПОСОБ ПОЛУЧЕНИЯ ВАКЦИНЫ (VACCINE PCV-2, AND, METHOD FOR MANUFACTURING A VACCINE)», в которой описана «вакцина против цирковируса свиней (PCV-2) и метод производства указанной вакцины для защиты свиней от заражения PCV», в которой заявители разработали процесс получения рекомбинантных антигенов, предпочтительно в эукариотической системе (клетки насекомых) для решения проблемы низкого выхода вирусных частиц в культивируемых клетках. Аналогично вышеуказанному патенту, в патенте США 770028, зарегистрированному 29 декабря 2005 г. в патентной базе данных США, который озаглавлен: «Содержащие PCV2 иммуногенные композиции и способы получения таких композиций (PCV2 immunogenic compositions and methods of producing such compositions)», описан метод повышенного выхода капсидного рекомбинантного белка PCV-2 в супернатант индуцированных клеток, начиная примерно с 5 дня после заражения. В противоположность этому, в настоящем изобретении предложено получение от десятков для сотен миллиграммов белков в течение нескольких часов индукции, после лизиса и выделения рекомбинантного белка.
Другим депозитом, обнаруженным в патентной базе данных США, являлся US 8008001, зарегистрированный 11 декабря 2009 г., озаглавленный «Вакцина против PCV-2 (PCV-2 VACCINE)», в котором описана вакцина и метод защиты поросят от заражения PCV-2 путем введения вакцины, содержащей капсидный белок PCV-2 в количестве по меньшей мере 20 мкг/дозу. В указанном патенте описано предпочтительное применение эукариотической экспрессионной системы (клетки насекомых, инокулированные рекомбинантным бакуловирусом). Эта система отличается от предложенной в настоящем изобретении, поскольку в нем используют бактериальные системы, а также новые способы выделения и количественной оцени антигена.
Кроме того, в патентной базе США обнаружен патент US 6287856 от 03/12/1999, озаглавленный «Вакцины против цирковирусных инфекций (Vaccines against circovirus infections)», в котором описаны композиции вакцин, обеспечивающие защитное действие от заражения PCV. Их основой являются векторы на основе нуклеиновой кислоты, содержащей кодирующую последовательность полипептида PCV, при этом в нуклеиновой кислоте отсутствует вирусный сайт инициации репликации, таким образом, патент относится к ДНК-вакцинам, отличным от предлагаемых в настоящей заявке, согласно которой антиген получают в E. coli.
Первым объектом настоящего изобретения является очищенный рекомбинантный капсидный белок PCV-2, полученный в генетически модифицированной E. coli, в качестве антигена вакцины и диагностического антигена. Вторым объектом изобретения являются вирусоподобные частицы (ВПЧ) PCV2, полученные с помощью генетически модифицированной E. coli, в качестве антигена вакцины. Оба объекта можно применять в программах контроля и предупреждения инфекции, вызываемой PCV-2. Третьим объектом настоящего изобретения является рекомбинантная вакцина против PCV-2, содержащая очищенный антиген PCV2 или рекомбинантные ВПЧ, или их варианты, полученные в генетически модифицированной E. coli, в сочетании с гидроксидом алюминия в качестве адъюванта. Ее также можно применять в программах контроля и предупреждения инфекций, вызываемых PCV-2.
Кроме того, объектом настоящего изобретения является набор для ELISA с захватом антигенов, предназначенный для дополнительной количественной оценки полученного рекомбинантного белка в композициях вакцин. Основными преимуществами этой методики является простота и автоматическое выполнение, точность, технологичность и низкая стоимость. Кроме того, основным преимуществом предложенного ELISA с захватом антигенов является то, что для анализа не требуется ни очистка образцов, ни высокие концентрации, поскольку антиген фиксируется с помощью специфических антител, и это взаимодействие пропорционально концентрации указанных белков, что приводит к большей воспроизводимости результатов, когда требуется количественная оценка. Можно обнаруживать очень низкие количества белка, такие как 50 мкг, разведенные в 1 л раствора. Таким образом, благодаря его полезным свойствам и простоте, этот анализ сам является перспективным кандидатом для применения в качестве эталона безопасного тестирования и анализа для оценки зависимости от дозы с целью подтверждения силы вакцины, что требуется при выпуске различных коммерческих вакцин, при необходимости после адаптации.
Создание иммуногенных соединений, обладающих способностью обеспечивать защиту от заражения PCV-2, является целевым исследованием в общей рамке исследований для контроля PCVAD. Капсидный рекомбинантный белок PCV-2, полученный в бактериальной системе и выделенный указанным выше путем, рассматривается в качестве потенциального иммуногена. Кроме того, замена трудоемких методик детекции и количественной оценки PCV-2 в качестве диагностического метода, представляет определенный интерес в применяемой в ветеринарии диагностической индустрии. Таким образом, вклад в производство и выделение рекомбинантных антигенов, представленных в настоящем описании, для получения требуемых соединений для целей диагностики и создания вакцин, а также другие пути их применения являются очень многообещающими.
Описание чертежей
На чертежах показано:
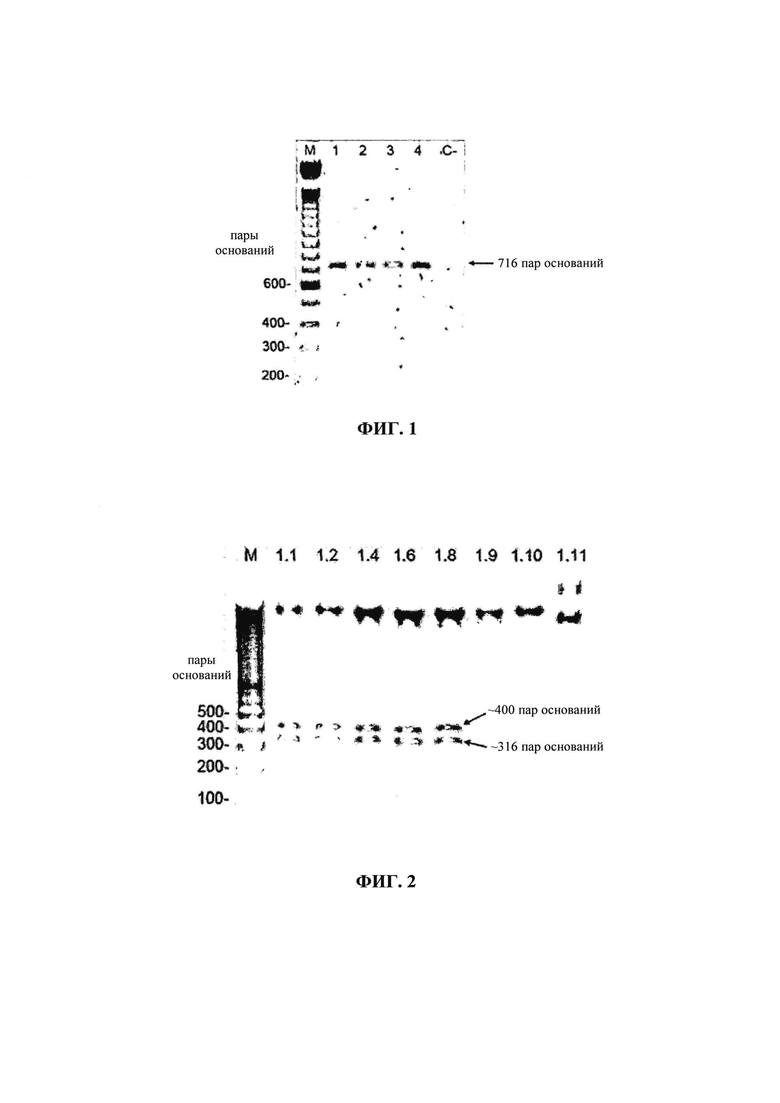
на фиг. 01 - результаты анализа в 1%-ном агарозном геле ПЦР-продуктов. М: маркер длин ДНК (ДНК-лестница) 100 пар оснований; 1, 2, 3 и 4: ДНК, соответствующие ОРС2 амплифицированного PCV-2; С-: отрицательный контроль (ДНК, экстрагированная из SK6-клеток, свободных от PCV-2);
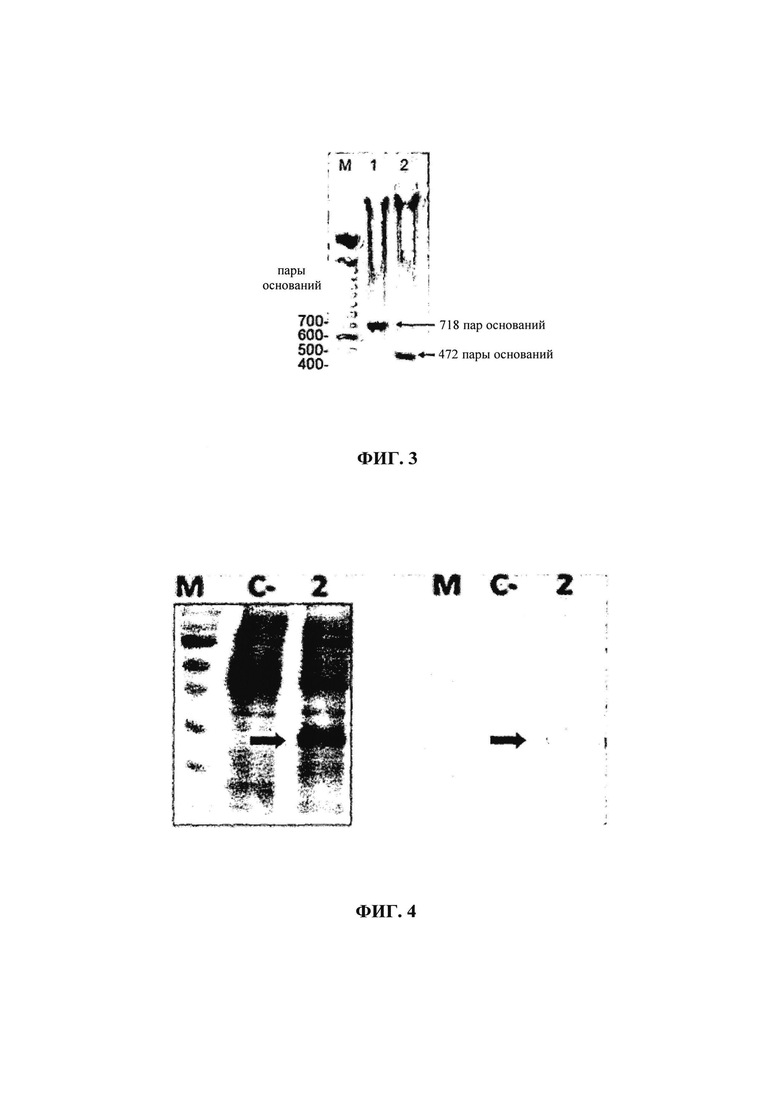
на фиг. 02 - результаты анализа в 1%-ном агарозном геле продуктов реакций расщепления плазмид с помощью EcoRI, демонстрирующие полосы, соответствующие 400 и 316 парам оснований, полученные в результате расщепления ОРС2. М: маркер длин ДНК (ДНК-лестница) 100 пар оснований; 1.1, 1.2, 1.4, 1.6 и 1.8: колонии белого цвета, произвольно выбранные 1.9, 1.10 и 1.11: отрицательный контроль (колонии голубого цвета);
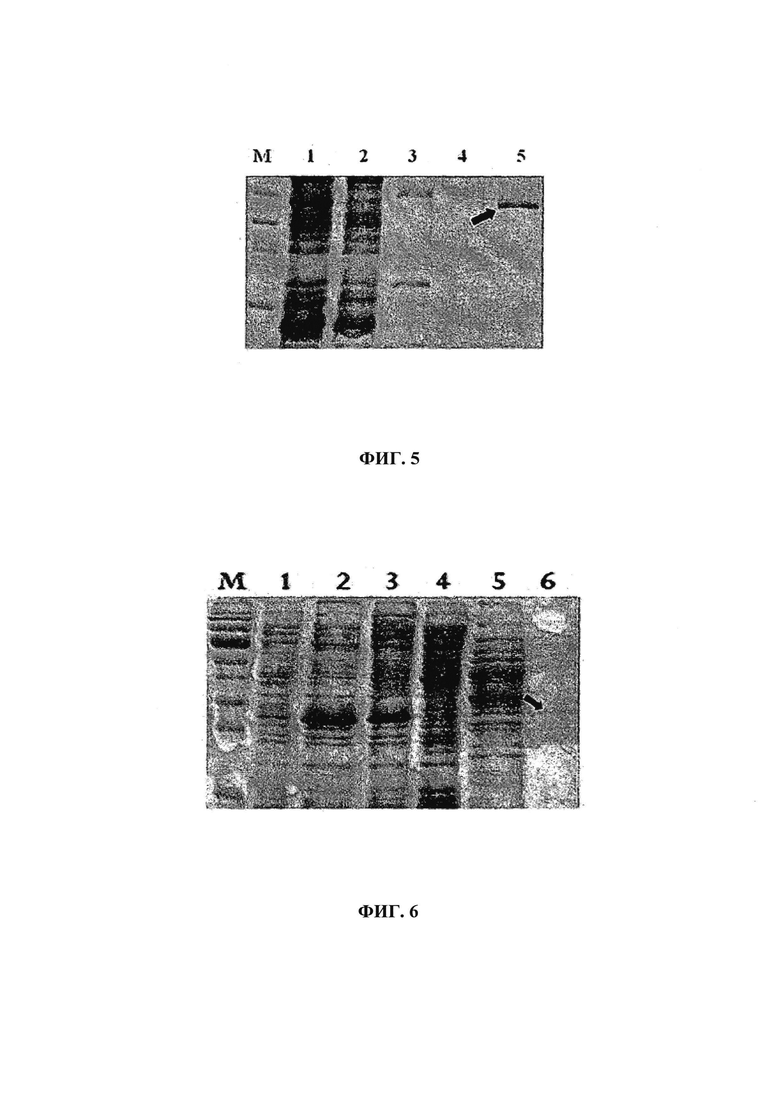
на фиг. 03 - результаты анализа в 1%-ном агарозном геле продуктов реакций расщепления рекомбинантной плазмиды pCap-PCV2 с помощью EcoRI (канал 1) и XbaI (канал 2). М: маркер длин ДНК (ДНК-лестница) 100 пар оснований;
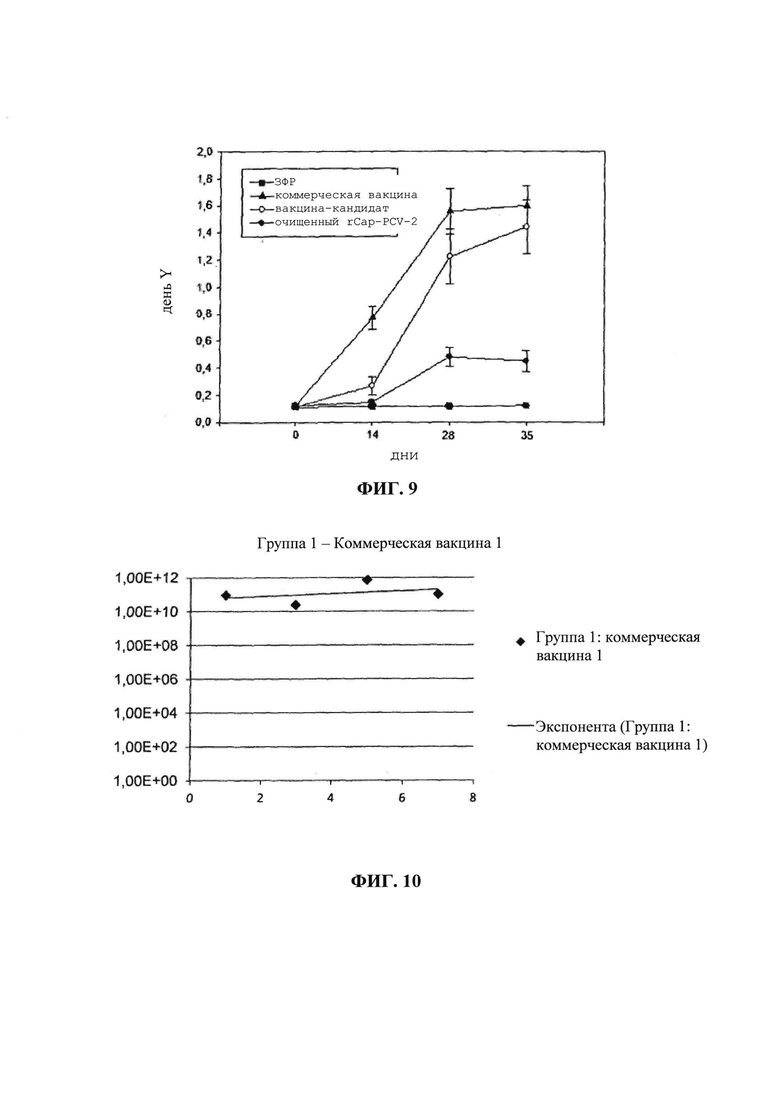
на фиг. 04 - результаты, подтверждающие экспрессию rCap-PCV-2. Представлены результаты, полученные на геле после анализа с помощью 15% ДСН-ПААГ (слева), и на нитроцеллюлозной мембране, полученные с помощью Вестерн-блоттинга (справа). М: маркер молекулярной массы; С-: отрицательный контроль (бактериальная экспрессия в растворимой фракции экстракта E. coli, трансформированной пустой плазмидой, и индуцированной ИПТГ); и 2: образец (растворимая фракция экстракта E. coli, трансформированной pCap-rPCV2 и индуцированной ИПТГ). Стрелкой обозначена полоса массой примерно 30 кДа, соответствующая rCap-PCV-2;
на фиг. 05 - белковый профиль, полученный в процессе очистки из растворимой фракции. М: маркер молекулярной массы; 1: растворимая фракция индуцированного экстракта E. coli; 2: невзаимодействующий объем; 3: промывка с помощью 10%-ного элюирующего буфера; 4: промывка с помощью 20%-ного элюирующего буфера; 5: элюция с помощью 100%-ного элюирующего буфера. Стрелкой обозначен очищенный rCap-PCV-2;
на фиг. 06 - белковый профиль полученных фракций после процесса осаждения растворимой фракции с различными уровнями насыщения сульфатом аммония. М: маркер молекулярной массы; 1: растворимая фракция, 2: фракция 0-20%, 3: фракция 20-40%, 4: фракция 40-60%, 5: фракция 60-80%, 6: очищенный rCap. Стрелкой обозначен rCap-PCV-2;
на фиг. 07А, Б и В - изображения, полученные с помощью электронной микрографии: образование вирусоподобных частиц (ВПЧ) rCap-PCV-2.
Увеличение 140000× (А и Б). Увеличение 250000× (В). Образцы анализировали с помощью трансмиссионного электронного микроскопа, применяя микроскоп с решеткой 200 меш, покрытой формваром/углеродом. ВПЧ обозначены стрелками;
на фиг. 08 - данные о разведениях образцов: rCap-PCV-2: разведения от 10 мг/л до 0,004883 мг/л. Вакцина-кандидат: разведения от 1:50 до 1:103200. Лизат У. coli, применяемый в качестве отрицательного контроля: разведения от 1:50 до 1:103200. Полученное уравнение представляет собой y=0,875x+0,196 (R2=0,996), в котором у обозначает ОП при 492 нм, а x обозначает концентрацию белка, подлежащего обнаружению;
на фиг. 09 - данные о специфическом гуморальном ответе, индуцированном rCap-PCV-2 у мышей, до и после вакцинации, полученные с помощью непрямого ELISA. Животных инокулировали в дни 0 и 14 и осуществляли контрольное заражение в день 35. У животных, инокулированных вакциной-кандидатом, обнаружены существенные количества антител, особенно после второй инокуляции;
на фиг. 10 - результаты оценки виремии у животных, вакцинированных поступающей в продажу вакциной 1, полученные в момент отбора образцов. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=5E+10е0,1847x. Не обнаружено существенного снижения вирусной нагрузки в течение периода после вакцинации (R2=0,1195);
на фиг. 11 - результаты оценки виремии у животных, вакцинированных поступающей в продажу вакциной, полученные при использовании 2 доз образцов. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=2E+13e-1,323x. Не обнаружено существенного снижения вирусной нагрузки в течение периода после вакцинации (R2=0.5703);
на фиг. 12 - результаты оценки виремии у животных, вакцинированных вакциной-кандидатом, применяемой в дозе 50 пг, полученные в соответствующие моменты сбора. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=2E+12е-0,766x. Не обнаружено существенного снижения вирусной нагрузки в течение периода после вакцинации (R2=0,9696);
на фиг. 13 - результаты оценки виремии у животных, вакцинированных вакциной-кандидатом, применяемой в виде двух доз по 50 мкг, полученные в соответствующие моменты сбора. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=1E+12е-0,79x. Обнаружено существенное снижение вирусной нагрузки в течение периода после вакцинации (R2=0,9418);
на фиг. 14 - результаты оценки виремии у животных, вакцинированных вакциной-кандидатом, применяемой в дозе 150 мкг, полученные в соответствующие моменты сбора. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=2E+17е-4,106x. Не обнаружено существенного снижения вирусной нагрузки в течение периода после вакцинации (R2=0,7912).
на фиг. 15 - результаты оценки виремии у животных, вакцинированных вакциной-кандидатом, применяемой в виде двух доз по 150 мкг, полученные в соответствующие моменты сбора. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=4E+13е-1,594x. Не обнаружено существенного снижения вирусной нагрузки в течение периода после вакцинации (R2=0,6673);
на фиг. 16 - результаты оценки виремии у невакцинированных животных (отрицательный контроль), полученные в соответствующие моменты сбора. На вертикальной оси указана вирусная нагрузка, на горизонтальной оси указано время 4 стадий сбора: свиньи возрастом 21, 63, 105 и 154 дней. Линейная регрессия представлена формулой y=2Е+13е-1,304x. Не обнаружено существенного снижения вирусной нагрузки в течение периода после обработки (R2=0,7425).
Подробное описание изобретения
Выделение и клонирование кодирующей области капсидного белка цирковируса свиней
ДНК цирковируса свиней 2 (PCV-2), применяемую для амплификации кодирующей области капсидного белка (ОРС2), SEQ ID NO: 03, выделяли из образцов ткани пораженных PCV-2 свиней из области Понти Нова, штат Минас-Жирайс. ОРС2 амплифицировали с помощью полимеразной цепной реакции (ПЦР), используя «прямой» олигонуклеотид 5'-CGCCATATGACGTATCCAAGGAGG-3' («прямой праймер»), который встраивали в сайт рестрикции, распознаваемый ферментом NdeI, на 5'-конце, и «обратный» олигонуклеотид 5'-CCCTCGAGTTAGGGTTTAAGTGGG-3' («обратный праймер»), который создавали для сайта, распознаваемого XhoI, на конце кодирующей области. Эти олигонуклеотиды конструировали на основе геномной последовательности бразильского изолята PCV-2 (DQ364650), депонированной в GenBank. Амплификацию ДНК-фрагментов осуществляли в термоячейке, используя ДНК-полимеразу Taq (5 ед./мкл). Амплифицированные образцы хранили при -4°С и анализировали с помощью горизонтального электрофореза на 1%-ном агарозном геле. На геле при использовании бромида этидия в концентрации 0,4 г/л в ультрафиолетовом свете обнаружен ДНК-фрагмент размером 716 пар оснований, что продемонстрировано на фиг. 01.
ПЦР-продукт очищали из агарозного геля. Затем очищенную ОРС2 PCV-2 клонировали в векторе для амплификации (векторная система pGEM®-T Easy, фирма PROMEGA). Полученную с помощью набора согласно инструкциям производителя связывающуюся смесь добавляли примерно к 100 мкл раствора, содержащего бактерии штамма E. coli DH5α, которым перед этим придана компетентность (Sambrook J., Russell D.W., Molecular Cloning: A laboratory manual. 3-е изд., изд-во Cold Spring Harbor Laboratory Press, New York, 2001), и инкубировали в течение 30 мин на льду. Затем смесь клеток и плазмидной ДНК подвергали тепловому шоку в водяной бане при 42°С в течение 1 мин и вновь помещали на лед на 2 мин. Затем добавляли 900 мкл среды LB-Net (1% триптона, 0,5% дрожжевого экстракта, 1% NaCl; pH 7,5) без антибиотиков и клетки инкубировали при 37°С в течение 2 ч при 250 об/мин на орбитальном шейкере. Указанную смесь высевали на твердую содержащую LB-агар среду (1% триптона, 0,5% дрожжевого экстракта, 1% NaCl, 1,5% агара Select; pH 7,5), включающую ампициллин (от 5 до 10 мг/мл), и инкубировали в течение 12-14 ч при 37°С.
Выделяли устойчивые к ампициллину клоны и применяли их для подтверждения присутствия плазмиды, содержащей представляющий интерес ген, с помощью ПЦР и ферментативного анализа экстрагированных плазмид с использованием препаратов мини-плазмид (Sambrook J., Russell D.W., Molecular Cloning: A laboratory manual. 3-е изд., изд-во Cold Spring Harbor Laboratory Press, New York, 2001). Ферментативный анализ осуществляли с помощью рестриктазы EcoRI и продукт реакции расщепления анализировали электрофоретически в 1%-ном агарозном геле (фиг. 02). ПЦР колонии осуществляли таким же методом, что и описанная выше ПЦР, однако вместо геномной ДНК из зараженных тканей добавляли с помощью стерильных палочек аликвоту трансформированных колоний. ПЦР-продукты анализировали также с помощью электрофореза на 1%-ном агарозном геле.
Полученные образцы плазмид, содержащие вирусную ДНК, правильность которых подтверждена с помощью ферментативного анализа и ПЦР-анализа колонии, очищали, используя набор для очистки ПЦР-продуктов, и затем отправляли в Genomics Laboratory, BIOAGRO/UFV для последующего секвенирования с целью подтверждения правильной сборки вектора. Реакции секвенирования основывались на технологии терминации цепей с помощью дидезоксинуклеотидов (ддНТФ) (Sanger F., Nicklen S., Coulson A.R., DNA sequencing with chain-terminating inhibitors. Proceedings of the National Academy of Sciences, т. 74, 1977, cc. 5463-54677). Созданные последовательности редактировали и собирали в виде «контига» (непрерывная нуклеотидная последовательность, составленная из отдельных перекрывающихся коротких нуклеотидных последовательностей), используя программы для биоинформатики, а затем сравнивали друг с друг с другом и доступными последовательностями, депонированными в GenBank, с помощью программы BLAST. Транслированная последовательность ОРС2 представлена в SEQ ID NO: 04. Содержащие конструкцию клоны хранили в среде LB-Net в микроцентрифужных пробирках с 20% глицерина и хранили при -80°С.
Перенос кодирующей области капсидного белка цирковируса свиней 2 в экспрессионный вектор в бактериальной системе
После проверки правильности последовательности вставки в векторе образец плазмидной ДНК pCapPCV-2e, SEQ ID NO: 03 (вектор для амплификации) подвергали ферментативному анализу, для которого использовали специфические сайты рестрикции для вставки гена ОРС2 в бактериальный экспрессионный вектор (рЕТ-16b, фирма Novagen), которая находилась под контролем промотора lac Т7 и имела последовательность, кодирующую состоящий из 10 остатков гистидина хвост на N-конце (применяемый для очистки посредством аффинности с содержащей никель смолой). Последовательность ОРС2 в pCapPCV-2 расщепляли с помощью NdeI и XhoI, аналогично описанному для бактериального экспрессионного вектора. После расщепления продукты (расщепленный бактериальный экспрессионный вектор и ОРС2-вставка, удаленная из вектора для амплификации) связывали с помощью ДНК-лигазы Т4. Затем продукты реакции связывания использовали для трансформации штамма E. coli DH5α согласно описанному выше методу. Трансформированные клоны произвольно отбирали из колоний для идентификации плазмид, несущих вставку. Для подтверждения клонирования осуществляли ПНР-анализы колонии, которые продемонстрировали, что механизм сайтнаправленного клонирования действовал ожидаемым образом для двух групп, произвольно выбранных из колоний, приводя к образованию фрагмента размером 716 пар оснований. Отбирали позитивные колонии и каждую колонию индивидуально подвергали ПЦР. Для колонии, содержащей рекомбинантную плазмиду с фрагментом размером примерно 716 пар оснований, осуществляли реакции расщепления плазмиды PCAP-RPCV-2, SEQ ID NO: 01, для подтверждения их правильной ориентации. Так, полосы ожидаемого размера 718 пар оснований и 472 пары оснований обнаружены после расщепления с помощью EcoRI и XbaI соответственно, при этом высокая молекулярная масса соответствовала оставшемуся фрагменту плазмиды (фиг. 03). Полученные рекомбинантные плазмиды обозначали как PCAP-rPCV2 и хранили в микроцентрифужных пробирках, содержащих 15-30% глицерина, и выдерживали при -70°С.
Экспрессия рекомбинантного белка капсида цирковируса свиней 2 и анализ с помощью ДСН-ПААГ-геля
Полную экспрессию рекомбинантных белков осуществляли в 1000 мл ТВ-среды (триптон, 12 г/л, дрожжевой экстракт, 24 г/л, глицерин, 4 мл, первичный кислый фосфат калия, 2,31 г/л и вторичный кислый фосфат калия, 12,54 г/л). Для этого компетентных бактерий штамма E. coli BL21-DE3-RIL codon plus, трансформировали конструкциями PCAP-rPCV2 с использованием методов, аналогичных применяемым для вектора для амплификации. При этом добавляли примерно 20 нг рекомбинантной плазмиды PCAP-rPCV2 к 100 мкл компетентных клеток, и смесь инкубировали на льду в течение 30 мин. Затем смесь клеток и плазмидной ДНК подвергали тепловому шоку в водяной бане при 42°С в течение 1 мин и вновь выдерживали на льду в течение 2 мин. Затем добавляли 900 мкл LB-среды (бакто-триптон, 10 г/л, дрожжевой экстракт, 5 г/л и хлорид натрия, 10 г/л) без антибиотиков и клетки инкубировали при 37°С в течение 2 ч при 250 об/мин. Клетки разводили стократно (1:100) в LB-среде, содержащей 50 мкг/мл ампициллина, и инкубировали при 37°С и 200 об/мин в течение 12 ч (преинокуляция). Культуру отрицательного контроля (нетрансформированные бактерии BL21) также выращивали в среде LB-Net, pH 7,0, дополненной хлорамфениколом (17 мкг/мл). Затем клетки разводили в соотношении 1:100 в жидкой ТВ-среде, pH 7,0, дополненной ампициллином (100 мкг/мл), и культуру выращивали при 30°С/250 об/мин в течение примерно 3 ч вплоть до достижения оптической плотности (ОП600) 0,6-0,8. Такую же процедуру осуществляли для отрицательного контроля, используя хлорамфеникол (17 мкг/мл). После достижения указанной ОП аликвоты культуры переносили в новые пробирки, в которых осуществляли индукцию в течение 1, 2, 3, 4 и 5 ч для определения наиболее оптимальной продолжительности индукции. Тестируемые концентрации ИПТГ составляли 0,1; 0,5 и 1 мМ, а применяемые температуры составляли 30°С и 37°С, всегда осуществляли интенсивное перемешивание и достаточную аэрацию. Отрицательной контроль подвергали такой же процедуре. Собирали по 1 мл образцов, полученных в каждом из вариантов культивирования, центрифугировали при 10000×g в течение 1 мин, супернатант отбрасывали и клеточный дебрис хранили при -20°С для анализа с помощью ДСН-ПААГ.
Для очистки индуцировали 1 л растущих колоний, содержащих рекомбинантную плазмиду pCap-rPCV2, используя установленные наиболее оптимальные параметры, такие как продолжительность стадии получения (4 ч), температура (30°С) и концентрация ИПТГ (0,25 мМ). После индукции в оптимальных условиях образец центрифугировали при 10000×g в течение 20 мин при 4°С. Супернатант отбрасывали и клеточный дебрис хранили при -20°С.
Осадок, полученный из объема 100 мл индуцированной среды, подвергали оттаиванию и ресуспендировали в лизирующем буфере (NaH2PO4, 20 мМ; NaCl, 300 мМ; имидазол, 5 мМ, pH 8,0) до конечного объема, составляющего примерно 2,5 мл. К этому буферу добавляли лизоцим в концентрации 1 мг/мл, и затем образец инкубировали при 0°С в течение 30 мин. Процесс клеточного лизиса осуществляли с использованием 6 циклов облучения ультразвуком каждый раз по 10 с при 200-300 Вт с 10-секундными интервалами, помещая пробирки на лед для предупреждения нагревания образца. Клеточный дебрис и тельца включения осаждали путем центрифугирования при 15000×g в течение 30 мин при 4°С. Супернатант (растворимая фракция) собирали в новую пробирку и применяли для очистки капсидного рекомбинантного белка PCV-2, обозначенного как rCap-PCV-2 (SEQ ID NO: 02).
Образцы (включая отрицательные контроли) анализировали в 15%-ном полиакриламидном геле (Sambrook J., Russell D.W., Molecular Cloning: A laboratory manual. 3-е изд., изд-во Cold Spring Harbor Laboratory Press, New York, 2001). После разделения гель проявляли с помощью раствора красителя (кумасси бриллиантовый голубой R-250, 0,1%, уксусная кислота, 9%, этанол 45%). Электрофоретическое разделение подтвердило наличие полосы 30 кДа, присутствующей в индуцированном образце. Указанная масса соответствует белку, кодируемому ОРС2 PCV-2, с добавленным гистидиновым хвостом. Подтверждение экспрессии осуществляли с помощью анализа методом Вестерн-блоттинга (фиг. 04).
Очистка капсидного рекомбинантного белка цирковируса свиней 2 и анализ с помощью ДСН-ПААГ-геля
Вирусный капсидный рекомбинантный белок PCV-2 очищали из растворимой фракции с помощью аффинной хроматографии на агарозной смоле с иммобилизованным никелем. Применяемый метод представлял собой «жидкостную хроматографию быстрого разрешения» (FPLC), в которой использовали хроматографическую колонку, загруженную никелем, для очистки рекомбинантных белков, слитых с гистидиновыми хвостами. Очистку стандартизовали с помощью нескольких анализируемых параметров, таких как: линейный поток, объемы, применяемые для промывки и элюции и концентрации имидазола и мочевины. После очистки раствор, содержащий белок, подвергали диализу трижды в противотоке 50 объемов карбонатного буфера (50 мМ, pH 7,2), содержащего 300 мМ NaCl, в целом в течение 9 ч (трехчасовой диализ). Образцы, полученные на всех стадиях очистки, анализировали на 15%-ном полиакриламидном геле согласно описанной выше процедуре. Белок успешно был очищен, что продемонстрировано на фиг. 05.
Определение концентрации Cap-PCV-2
Количество рекомбинантного белка вирусного капсида PCV-2 после очистки и диализа определяли с использованием метода, описанного Брэндфордом (Bradford М.М., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the Principle of protein-dyebinding. Analytical Biochemistry, т. 72, 1976, cc. 248-254). Расчет осуществляли с помощью линейной регрессии, при которой получали уравнение y=0,3267+0,0108x путем наилучшего подбора относительно величин оптической плотности для тестируемых разведений БСА.
Выделение рекомбинантного капсида белка цирковируса свиней 2 путем осаждения сульфатом аммония и получение применяемого в вакцине соединения и подтверждение образования вирусоподобных частиц (ВПЧ)
Для этого в растворимую фракцию, содержащую rCap-PCV-2, добавляли Na2SO4 в количестве, достаточном для 20% насыщения (до 0°), и затем образец инкубировали на льду при 100 об/мин в течение 30 мин. Затем белковый экстракт центрифугировали при 15000×g в течение 20 мин при 0°С. После завершения центрифугирования супернатант собирали и осадок ресуспендировали в таком же объеме карбонатного буфера, pH 7,0, содержащего 300 мМ NaCl (белковая фракция в сульфате аммония 0-20%). К супернатанту добавляли Na2SO4 в количестве, достаточном для 40% насыщения, начиная с первоначального раствора этой соли с насыщением 20%. Затем повторяли стадии инкубации и перемешивания на льду, и центрифугирования с получением фракций сульфата аммония с насыщением 20-40%, 40-60% и 60-80%. Фракции подвергали диализу в противотоке карбонатного буфера, pH 7,0, содержащего 300 мМ NaCl, и затем хранили при 4°С в течение максимум 2 дней для анализа методом ДСН-ПААГ (фиг. 06), Вестерн-блоттинга и ELISA с захватом антигена. Доза вакцины объемом 1 мл содержала в качестве адъюванта гель гидроксида алюминия1 (фирма OMEGA - Chemicals Inc.).
Для верификации ВПЧ фракции, полученные с помощью градиента CsCl, дающие положительный результат по данным Вестерн-блоттинга, по отдельности подвергали диализу в противотоке 500 мл карбонатного буфера (NaCl, 300 мМ, карбонат, 50 мМ, pH 7,0), 2 раза, каждый раз по 4 ч. Примерно 10 мкл каждой фракции добавляли на решетки 200 меш, покрытые формваром/углеродом, и давали выстояться в течение 1 мин при комнатной температуре. Затем избыток образца удаляли с помощью фильтровальной бумаги и на каждую решетку добавляли каплю 2%-ного уранилацетата и давали действовать в течение 1 мин. Избыток этого контрастного вещества удаляли фильтровальной бумагой и решетки выдерживали в эксикаторе в течение 2 дней. Анализ осуществляли с помощью трансмиссионного микроскопа и изображения фотографировали (фиг. 07А, Б и В).
Количественная оценка антигенов вакцин против PCV2 с помощью (антиген)захватывающего ELISA
При осуществлении указанного ELISA-анализа осаждали антитела к rcap-PCV-2, образовавшиеся в организме кролика, и применяли в качестве захватывающего и идентифицирующего антител, последнее конъюгировали с пероксидазой. При стандартизации установлено, что оптимальная концентрация антигена составляла 0,625 г/мл, оптимальное разведение захватывающего антитела составляло 15 мкг/мл и разведение конъюгированного с пероксидазой идентифицирующего антитела составляло 1:800. Все процедуры осуществляли согласно методу «шахматной доски» (Crowther J.R., ELISA. Theory and Practice. Methods in Molecular Biology, т. 42, 1995, с.с. 1-223). После оптимизации рабочих условий (антиген)захватывающего ELISA концентрацию rCap-PCV-2 применяли для создания стандартной кривой (y=0,196+0,875x) с помощью линейной регрессии (фиг. 08). Дл создания стандартной кривой рассчитывали средние величины абсорбции, полученные с использованием 2-кратного серийного разведения белка, начиная с 5 мг/л до 0,0048 мг/л, в 96-луночном микропланшете, сенсибилизированном захватывающим антителом в концентрации 0,625 г/мл. Рабочий диапазон анализа определяли как линейную область кривой с коэффициентом корреляции R2=0,996. Таким образом, калибровка по семи точкам с концентрацией rCap-PCV-2 от 0,625 до 0,0391 мг/л соответствовала результату анализа методом линейной регрессии.
Для количественного определения белка в соединении, применяемом в вакцине, использовали 0,6 г осадка, что эквивалентно 100 мл индуцирующей клетки среды, содержащей трансформированные E. coli с рекомбинантной плазмидой pCap-2-RPCV, содержащей ген, соответствующий ОРС2 PCV-2, и индуцированной ИПТГ для получения рекомбинантного белка. В качестве образца, применяемого для отрицательного контроля, использовали такое же количество экстракта клеток, трансформированных пустым (незагруженным) бактериальным экспрессионным вектором. Осадки этих клеток подвергали оттаиванию и ресуспендировали в 2 мл лизирующего буфера (NaH2PO4, 20 мМ, NaCl, 300 мМ, имидазол, 5 мМ, pH 8,0). К лизирующему буферу добавляли лизоцим в концентрации 1 мг/мл, 1 мМ PMSF и затем образец инкубировали на льду в течение 30 мин. Процесс клеточного лизиса осуществляли с использованием 6 циклов облучения ультразвуком каждый раз по 10 с при 200-300 Вт с 10-секундными интервалами, помещая пробирки на лед для предупреждения нагревания образца. Неизвестную концентрацию rCap-PCV-2 в вакцине-кандидате определяли методом Holden с соавторами (Holden L., Faeste C.K., Egaas E., Quantitative Sandwich ELISA for the determination of lupine-lupinus spp. in foods. Journal of Agriculture Food Chemistry, т. 53, 2005, cc. 5866-5871), при осуществлении которого получали серийные разведения вакцины, и разведение, при котором величина ОП при 492 нм была наиболее близкой к средней точке линейной области стандартной кривой, применяли для расчета концентрации белка в вакцине. Так, концентрация rCap-PCV-2 (SEQ ID NO: 02) в вакцине-кандидате после расчета с учетом фактора разведения образца, составляла 0,73 мг/мл.
Представленные в качестве иллюстрации эксперименты
Эксперименты на мышах
Самок мышей возрастом 5 недель разделяли на 4 следующие группы по 5 животных в каждой: одну группу обрабатывали разработанной вакциной-кандидатом (белковый осадок после диализа в противотоке сульфата аммония 0-40%); одну группу обрабатывали очищенным rCap-PCV-2; одной группе инокулировали фосфатный буфер ЗФР (отрицательный контроль) и одной группе инокулировали поступающие в продажу вакцины против цирковируса свиней (положительный контроль). При обработке групп, которые вакцинировали вакциной-кандидатом, очищенным белком, и группы, которой инокулировали ЗФР, добавляли в качестве адъюванта гидроксид алюминия, исключением были животные, которых инокулировали поступающей в продажу вакциной. Животных вакцинировали дважды (две дозы) i.p., с интервалом между дозами 14 дней. Количество rCap-PCV-2, инокулированного животным из группы, обработанной вакциной-кандидатом, и из группы, инокулированной очищенным белком, составляло 30 мкг для первой дозы и 15 мкг для второй дозы. Собирали образцы крови посредством пункции глазного синуса в дни 0, 14, 28 и 35 после вакцинации для серологического анализа. Животных умерщвляли через 14 дней после контрольного заражения. Все этические принципы проведения экспериментов с животными соответствовали Бразильской коллегии по проведению экспериментов с участием животных (Brazilian College of Animal Experimentation (COBEA)) (Сертификат по этике использования животных (Certificate of Ethics for Animal Use) (CEUA)AJFV, Case №054/2011).
Оценку гуморального иммунного ответа осуществляли методом непрямого ELISA. Оптимальные рабочие концентрации антигена rCap-PCV-2 и наилучшее разведение сыворотки (первичное антитело) определяли титрованием методом «шахматной доски» (Crowther J.R., ELISA. Theory and Practice. Methods in Molecular Biology, т. 42, 1995, cc. 1-223). При использовании для этого PCV-2-позитивной мышиной сыворотки было установлено, что оптимальная концентрация антигена (rcap-PCV-2) составляет 0,156 мкг/лунку и разведение составляет 1:4000 соответственно. У мышей, инокулированных вакциной-кандидатом, обнаружены уровни, сходные с уровнями, обнаруженными у мышей, инокулированных поступающими в продажу вакцинами, прежде всего после введения второй дозы вакцины (фиг. 09), возможно из-за присутствия ВПЧ (фиг. 07) и адъюванта в применяемой концентрации.
Эксперименты на свиньях
Свиней из стандартной фермы по коммерческому разведению возрастом 21 день вакцинировали вакциной-кандидатом и для сравнения поступающими в продажу вакцинами. Оценивали у свиней антигенный потенциал применяемого в вакцине соединения, содержащего рекомбинантный белок вирусного капсида PCV-2 (rCap-PCV-2). Для этого использовали 140 свиней возрастом примерно 21 день из стандартной фермы по коммерческому разведению из области Понти Нова, штат Минас-Жирайс. Эксперимент осуществляли согласно принятому на ферме руководству. Указанный полевой опыт проводили согласно стандартам Руководства по применению животных для учебных, исследовательских и пропагандистских целей (Standards of Conduct for the use of Animals in Teaching, Research, and Extension) Департамента ветеринарной медицины DVT/UFV, регистрационный номер 37/2012.
Животных разделяли на 7 групп по 20 животных. Группы 1 и 2 вакцинировали поступающими в продажу вакцинами согласно рекомендациям производителя. Группы 3 и 4 вакцинировали вакциной- кандидатом для поступления на рынок, содержащей 50 мкг рекомбинантного белка, при этом животные из группы 3 получали одну дозу, когда они имели возраст 21 день, а животные из группы 4 получали две дозы в возрасте 21 и 35 дней. В противоположность этому, животных из групп 5 и 6 вакцинировали вакциной-кандидатом, содержащей 150 мкг рекомбинантного белка, используя одну и две дозы соответственно, согласно такой же схеме вакцинации, что и применяемая в группах 3 и 4. Контрольной группе 7 вводили фосфатный буфер (ЗФР). Все иммунизации осуществляли внутримышечным путем. Для количественной оценки вирусной нагрузки образцы крови собирали, начиная с адаптационной фазы вплоть до дня убоя. Образцы крови собирали в дня жизни животных 21, 63, 105 и 154. Экстракцию общей ДНК осуществляли с помощью поступающего в продажу набора для экстракции. Количественную оценку нагрузки вирусом PCV-2 в собранной сыворотке осуществляли с помощью ПЦР в реальном времени согласно протоколу, описанному у Silva с соавторами. (Silva F.M.F., Silva Jr.A., Vidigal P.M., Oliveira C.R., Viana V.W., Silva C.H.O., Vargas I., Fietto J.L.R., Almeida M.R., Porcine Circovirus-2 Viral Load versus Lesions in Pigs: Perspectives for Post-weaning multisystemic Wasting Syndrome. Journal of Comparative Pathology, т. 144, 2011, cc. 296-302). Реакции осуществляли в трех повторностях, и все проанализированные значения Ct (пороговый цикл) представляли собой среднее значение Ct для каждого образца, таким образом, из среднего значения Ct каждого образца рассчитывали усредненное значение и стандартное отклонение для группы и изучаемой популяции. Установленные средние значения применяли для оценки тенденции к снижению вирусной нагрузки с помощью экспоненциального регрессионного анализа. При применении поступающих в продажу вакцин 1 и 2 не обнаружено статистически значимой тенденции к снижению вирусной нагрузки (фиг. 10 и 11). Статистически значимое снижение вирусной нагрузки продемонстрировано в группах 3 и 4 (rCap-PCV2 50 мкг, одна и две дозы) R2=0,96 и R2=0,94 соответственно (фиг. 12 и 13), при этом тенденция к снижению обнаружена, начиная с третьей недели анализа. В группах 5 и 6 (rCap-PCV2 150 мкг - одна и две дозы) не обнаружено статистически значимой тенденции к снижению вирусной нагрузки (R2=0,7184 и R2=0,638, см. фиг. 14 и 15 соответственно). В группе отрицательного контроля не обнаружено никакой статистически значимой тенденции к снижению вирусной нагрузки (фиг. 16).
Заключение
В настоящее время на рынке ветеринарных средств присутствуют коммерческие вакцины против PCV-2 различных иностранных компаний, и они производятся вне Бразилии, что добавляет связанную с импортом стоимость к стоимости партий вакцин. Антигены вакцин можно применять для вакцинации животных в программах контроля ассоциированных с PCV-2 болезней в общепринятых системах свиноводства, и они представляют собой альтернативу поступающим на рынок вакцинами. Набор для ELISA можно применять для количественной оценки вакцинных антигенов PCV-2, который можно использовать в качестве тестов для гарантии качества поступающей в продажу и/или экспериментальной вакцины.







Изобретение относится к области биотехнологии. Описано получение рекомбинантного антигена вирусного капсида цирковируса свиней 2 (PCV-2) и его модификации путем экспрессии в прокариотической системе, очистки в мономерной форме, выделения вирусоподобных частиц (ВПЧ) и его применение в композициях вакцин, диагностических наборах и системе для количественной оценки антигена PCV-2 в партиях вакцин путем анализа с помощью (антиген)захватывающего ELISA. Антигены и композиции вакцин можно применять в программах иммунизации животных для борьбы с ассоциированными с PCV-2 болезнями в общепринятых системах разведения свиней, и они представляют собой альтернативы поступающим в продажу вакцинам. Набор для ELISA можно использовать для оценки качества поступающих в продажу и/или экспериментальных вакцин против PCV-2. 11 н.п. ф-лы, 16 ил., 2 пр.
1. Рекомбинантный антиген цирковируса свиней 2 (PCV-2) для получения вакцин против PCV-2, отличающийся тем, что он имеет SEQ ID NO: 02.
2. Рекомбинантный антиген цирковируса свиней 2 (PCV-2) для получения наборов для диагностики PCV-2, отличающийся тем, что он имеет SEQ ID NO: 02.
3. Вирусоподобная частица (ВПЧ) PCV-2 для получения вакцин против PCV-2, отличающаяся тем, что она содержит продукт олигомеризации SEQ ID NO: 02.
4. Вирусоподобная частица (ВПЧ) PCV-2 для получения наборов для диагностики PCV-2, отличающаяся тем, что она содержит продукт олигомеризации SEQ ID NO: 02.
5. Вакцина против цирковируса свиней 2, включающая один или несколько выделенных антигенов по п. 1 или вирусоподобных частиц (ВПЧ) по п. 3 в эффективном количестве и фармацевтически приемлемый адъювант.
6. Диагностический набор для обнаружения цирковируса свиней 2, отличающийся тем, что содержит
(1) антиген по п. 2; и
(2) реагенты для проведения ELISA с захватом антигенов.
7. Применение антигена по п. 1 или вирусоподобной частицы (ВПЧ) по п. 3, для получения соединений, применяемых в вакцинах против PCV-2.
8. Применение антигена по п. 2 или вирусоподобной частицы (ВПЧ) по п. 4 для создания наборов для диагностики PCV-2.
9. Применение вакцины по п. 5, отличающееся тем, что ее используют для контроля болезней, связанных с PCV-2.
10. Применение диагностического набора по п. 6, отличающееся тем, что его используют для количественной оценки капсидного белка PCV-2, более конкретно для контроля качества композиций вакцин против PCV-2.
11. Способ выделения антигенов по п. 1 или 2, отличающийся тем, что осуществляют следующие стадии, на которых:
- добавляют сульфат аммония (NH4)2SO4 к растворимой фракции экстракта Е. coli, трансформированной pCap-rPCV2, которая содержит капсидный рекомбинантный белок PCV-2 с аминокислотной последовательностью SEQ ID NO: 02 (PCV-2-rcap) в количестве, достаточном для уровня насыщения 0-20%, после чего инкубируют образец при 0°С, 100 об/мин в течение 30 мин;
- центрифугируют белковый экстракт при 15000 × g в течение 20 мин при 0°С, собирают супернатант и ресуспендируют осадок в начальном объеме, равном начальному объему карбонатного буфера рН 7,0, содержащего 300 мМ хлорид натрия (NaCl);
- добавляют сульфат аммония в количестве, достаточном для 40%-ного насыщения супернатанта, начиная с применения начального раствора указанной соли с уровнем насыщения 20%;
- повторяют стадии инкубации и перемешивания на льду, и центрифугирования с получением фракций с уровне м насыщения сульфатом аммония 20-40%, 40-60% и 60-80%;
- осуществляют диализ фракций в карбонатном буфере, рН 7,0, содержащем 300 мМ NaCl, с получением выделенного антигена;
- хранят выделенный антиген при 4°С.
| WO 2010061000 A1, 03.06.2010 | |||
| WO 2009085912 A1 | |||
| WO 2011048215 A1 | |||
| ЦИРКОВИРУС СВИНЕЙ ТИПА II И ЕГО ПРИМЕНЕНИЕ | 1998 |
|
RU2283862C2 |
| WU P.-CH | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Appl | |||
| microbial | |||
| biotechnol | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| YIN SH | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Virology journal | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |

