Область техники, к которой относится изобретение
Изобретение относится к области биотехнологии, молекулярно-генетической диагностики, в частности к оценке однонуклеотидного полиморфизма rs6677 (T>G) гена C11orf58 молекулярно-генетическим методом исследования.
Уровень техники
Ген C11orf58 кодирует белок, открытый в 2020 г. японскими учеными [Tsuboyama K. et al. A widespread family of heat-resistant obscure (Hero) proteins protect against protein instability and aggregation //PLoS Biology. - 2020. - Vol. 18. - №. 3. - P. e3000632]. Данный белок характеризуется высокой устойчивостью к воздействию неблагоприятных физических и химических факторов и способствует восстановлению нативной третичной или четвертичной структуры других белков в условиях стресса, что доказывает его выраженные шапероноподобные свойства. Потенциально значимая роль данного белка в широком спектре заболеваний (нейродегенеративных, атеросклероза и др.) определяет необходимость проведения молекулярно-генетических исследований гена C11orf58 и разработки методик генотипирования полиморфных вариантов данного гена. Ген C11orf58 (Gene ID: 10944) локализован на хромосоме 11p15.2. Полиморфный вариант rs6677 гена C11orf58, позиция chr11:16755585 (GRCh38.p14) [https://www.ncbi.nlm.nih.gov/snp/rs6677] локализован в 3′-нетранслируемой области и характеризуется заменой T>G. Согласно SNPinfo Web Server/ LD TAG SNP Selection [https://snpinfo.niehs.nih.gov/snpinfo/snptag.html] rs6677 гена C11orf58 является таргетным (то есть репрезентативным однонуклеотидным полиморфизмом в геномной области с высокой степенью неравновесия по сцеплению с группами других полиморфных локусов, составляющих гаплотип).
Генетический вариант rs6677 (T>G) характеризуется высоким регуляторным потенциалом. Согласно биоинформатическим ресурсам, данный генетический вариант влияет на уровень экспрессии гена C11orf58 посредством cis-eQTL-эффектов [https://gtexportal.org/home/snp/rs6677]; ассоциирован с модификациями гистонов, маркирующими энхансеры [https://pubs.broadinstitute.org/mammals/haploreg/detail_v4.2.php?query=&id=rs6677]. Также установлено влияние rs6677 на связывание с транскрипционными факторами в зависимости от носительства референсного/альтернативного аллелей [http://atsnp.biostat.wisc.edu/search]. Это доказывает высокую функциональную значимость данного генетического варианта и создает потребность в создании простого в исполнении, недорого и доступного исследователям, работающим в области генетической эпидемиологии, метода идентификации однонуклеотидного полиморфизма rs6677 (T>G) гена C11orf58.
Известен способ анализа генетических вариаций в геноме человека методом секвенирования амплифицированных участков ДНК [Mardis E. R. DNA sequencing technologies: 2006-2016 //Nature protocols. - 2017. - Vol. 12. - №. 2. - P. 213-218]. Недостатками метода являются высокая стоимость оборудования и реагентов, что исключает широкое внедрение метода в экспериментальные исследования, особенно изучение многофакторных заболеваний, которые требуют большого размера выборок для обеспечения высокой мощности исследований.
За прототип выбран способ анализа генетических вариаций в геноме человека методом матричноактивированной лазерной десорбционно-ионизационной масс-спектрометрии (MALDI). Метод заключается в том, анализируемая ДНК переносится на подложку, где она кристаллизуется с матрицей. Затем кристаллизованные аналиты переносят, облучают лазером, вызывая десорбцию и ионизацию молекул в вакуумной камере. Положительно заряженные ионы ДНК ускоряются и мигрируют через вакуумную трубку к высокочувствительному детектору с разной скоростью в зависимости от массы ионов, что приводит к различному времени пролета. Используя время пролета отдельных ионизированных ДНК-аналитов, система определяет массу и отображает масс-спектр, идентифицирующий различные генетические мишени [Li D. et al. MALDI-TOF mass spectrometry in clinical analysis and research //ACS Measurement Science Au. - 2022. - Vol. 2. - №. 5. - P. 385-404]. Однако, данный метод характеризуется высокой стоимостью, поскольку требует приобретения наборов спектральных чипов и дорогостоящих реактивов для проведения генотипирования, характеризуется высокой стоимостью оборудования, требует наличия высококвалифицированного персонала, в связи с чем он не может быть воспроизведен при наличии стандартного набора оборудования и реактивов.
Методов таргетного генотипирования rs6677 (T>G) гена C11orf58 с применением аллель-специфических флюоресцентных зондов до настоящего времени разработано не было.
Таким образом, существует реальная потребность в создании быстрого, недорогого и легко воспроизводимого способа идентификации полиморфизма rs6677 (T>G) гена C11orf58, с доступной всем исследователям структурой праймеров и аллель-специфических зондов, который мог бы использоваться в качестве «рутинного» метода генотипирования в любой ПЦР-лаборатории.
- Раскрытие сущности изобретения
Техническим результатом данного изобретения является разработка простого в исполнении и экономически целесообразного способа генотипирования однонуклеотидного полиморфизма rs6677 (T>G), локализованного в позиции chr11:16755585 (GRCh38.p14) гена C11orf58 (Gene ID: 10944) методом полимеразной цепной реакции в режиме «реального времени» с применением аллель-специфических сигнальных зондов, содержащих флуорофоры FAM и ROX.
Технический результат достигается тем, что идентификацию аллельных вариантов rs6677 (T>G) гена C11orf58 осуществляют с использованием прямого праймера rs6677 5′-TTGTTATAGTAGAGCTGTTCATTATGG-3′ (SEQ ID NO 1), обратного праймера rs6677 5′-TGTGAAAAGTACTTGCTCTCATGTT-3′ (SEQ ID NO 2), rs6677-T-аллель-специфичного флуоресцентно-меченого зонда 5′-(FAM)CTTAGTTTCCTGTTACT(RTQ1)-3′ (SEQ ID NO 3),
rs6677-G-аллель-специфичного флуоресцентно-меченого зонда 5′-(ROX)CTTAGTTGCCTGTTACT(BHQ2)-3′ (SEQ ID NO 4).
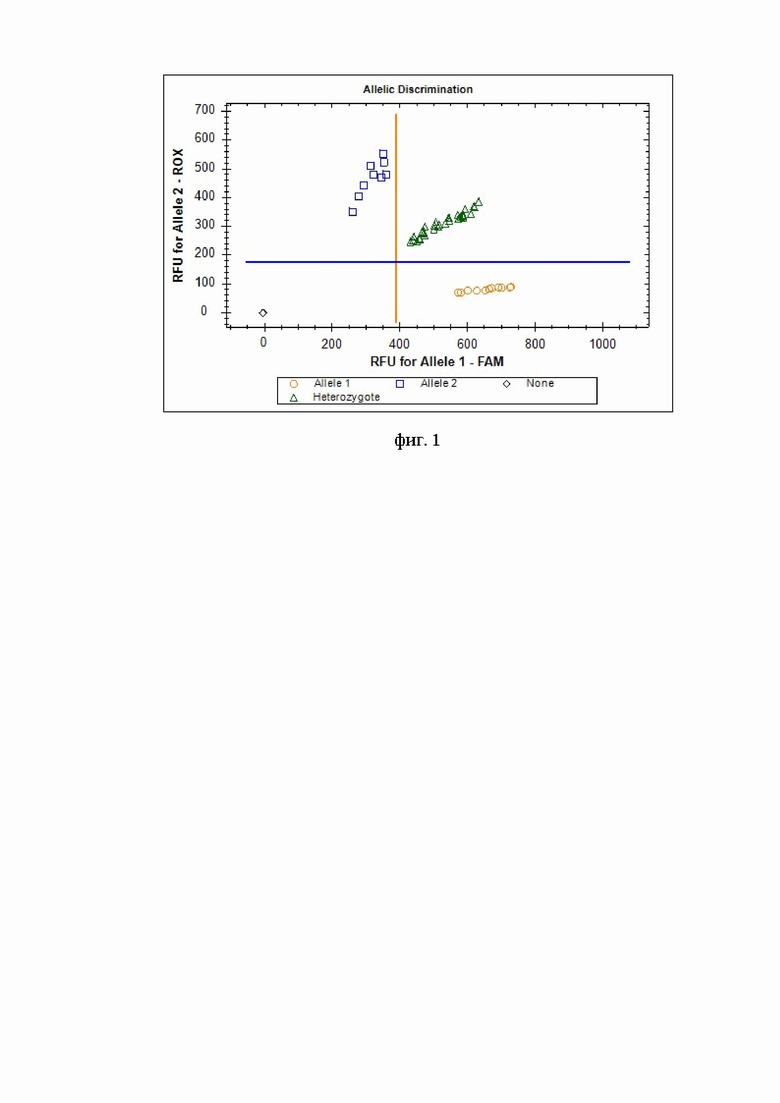
Изобретение поясняется следующей фигурой: дискриминация аллелей по локусу rs6677 (T>G) гена C11orf58 при генотипировании методом ПЦР в режиме «реального времени» с применением аллель-специфических флуоресцентных зондов по данным величин RFU (относительные единицы флуоресценции) на амплификаторе CFX96: генотипы rs6677-T/T показаны оранжевыми кругами, генотипы rs6677-T/G показаны зелеными треугольниками, генотипы rs6677-G/G показаны голубыми квадратами; черным ромбом отмечен отрицательный контроль.
Работа над дизайном олигонуклеотидов включала несколько этапов:
1) С применением открытой базы данных Ensembl genome browser 109 [https://www.ensembl.org/index.html] выбран сиквенс, фланкирующий искомую однонуклеотидную замену [T/G] rs6677 C11orf58, и затем с помощью доступного онлайн программного обеспечения Primer3web version 4.1.0 [https://primer3.ut.ee/] подобрана последовательность олигонуклеотидов, используемых для проведения ПЦР-реакции:
прямой общий праймер rs6677 5′-TTGTTATAGTAGAGCTGTTCATTATGG-3′ (SEQ ID NO 1),
обратный общий праймер rs6677 5′-TGTGAAAAGTACTTGCTCTCATGTT-3′ (SEQ ID NO 2).
Размер амплифицируемого в ходе ПЦР фрагмента гена C11orf58 составляет 150 пар нуклеотидов:
TTGTTATAGTAGAGCTGTTCATTATGGATATTTCTGCTGCCAGTCACAATCTAAATTAATTTTGGCAAAAGATTGGGTACTTAGTT[T/G]CCTGTTACTGAGTTAGCTCTACTCTTTTGGACCAAAGCAACATGAGAGCAAGTACTTTTCACA.
2). Для дизайна зондов пользовались практическими рекомендациями [Basu C. (ed.). PCR primer design. - New york : Humana Press, 2015]. В реакции использовались гидролизные зонды. Последовательность зонда подбирали таким образом, чтобы он отжигался на матрицу между прямым и обратным праймерами. Каждый зонд снабжали флуорофором и гасителем флуоресценции, спектр поглощения которого соответствует длинам волн спектра флуорофора. Для гашения флуоресценции FAM пользовались гасителем RTQ1; для гашения флуоресценции ROX - гасителем BHQ2.
На основании изложенных критериев и практических рекомендаций были подобраны зонды со следующей структурой:
rs6677-T-аллель-специфичный флуоресцентно-меченый зонд 5′-(FAM)CTTAGTTTCCTGTTACT(RTQ1)-3′ (SEQ ID NO 3),
rs6677-G-аллель-специфичный флуоресцентно-меченый зонд 5′-(ROX)CTTAGTTGCCTGTTACT(BHQ2)-3′ (SEQ ID NO 4).
3) Изготовление праймеров и зондов осуществлялось в сервисном центре НПК «Синтол», Москва.
4) С помощью практических экспериментов подобраны оптимальные условия для проведения генотипирования, которые включают следующие этапы: 50°C в течение 2 минут, 95°C в течение 10 минут, затем 39 циклов [95°C в течение 10 секунд и 48°C в течение 1 минуты].
5) Разработанный способ был апробирован в лаборатории геномных исследований на 200 образцах ДНК здоровых индивидуумов биобанка НИИ генетической и молекулярной эпидемиологии КГМУ. Генотипирование осуществляли по данным величин RFU (относительные единицы флуоресценции) зондов с флуоресцентными красителями. По результатам генотипирования rs6677 139 человек (69,5%) оказались гомозиготами по аллелю T (генотип T/T); 53 человека (26,5%) - гетерозиготами (генотип T/G), 8 человек (4%) индивидуумов были гомозиготами по аллелю G (генотип G/G).
6) Валидацию способа проводили методом масс-спектрометрического анализа на геномном времяпролетном масс-спектрометре MassArray analyzer 4 (Agena Bioscience). Результаты обоих способов генотипирования полностью (100% генотипов) совпали. Однако патентуемый способ генотипирования полиморфного локуса rs6677 (T>G) гена C11orf58 методом ПЦР в режиме «реального времени» с применением аллель-специфических зондов позволяет значительно (на 6 часов) сократить время проведения анализа, а также снижает себестоимость анализа (в 4-5 раз).
Осуществление изобретения
Способ осуществляют следующим образом:
1. Выделение ДНК из периферической венозной крови. На первом этапе к 0,5 мл крови добавляли 0,5 мл PBS и центрифугировали 10 мин при 12 тыс. об/мин. Надосадочную жидкость сливали, добавляли 1 мл PBS и вновь центрифугировали при тех же условиях. Надосадочную жидкость сливали, добавляли 200 мкл ТЕ-буфера, пипетировали до растворения осадка и затем последовательно добавляли 10 мкл 1% раствора додецилсульфата натрия SDS и 5 мкл протеиназы К. Пробирки инкубировали в термостате при t=37°C 12 ч. В ходе второго этапа проводили четыре последовательных центрифугирования с фенолом и хлороформом согласно протоколу методики (10 мин, 8 тыс. об/мин), после чего ДНК осаждали ледяным раствором 95% этилового спирта и центрифугировали 10 мин при 14,3 тыс. об/мин. По испарении спирта ДНК растворяли в 100 мкл деионизированной дистиллированной воды. Получаемый раствор ДНК в воде имел чистоту в диапазоне А260/280=1,5-2,0 и среднюю концентрацию около 180-200 нг/мкл.
2. Подготовка образцов ДНК к генотипированию. Качество выделенной ДНК оценивали по степени чистоты и концентрации раствора на спектрофотометре NanoDrop (Thermo Fisher Scientific, США). Все анализируемые образцы ДНК были разведены деионизированной водой до концентрации 15-20 нг/мкл при А260/280=1,5-2,0.
3. Анализ полиморфизма rs6677 (T>G) гена C11orf58 с помощью полимеразной цепной реакции в реальном времени с использованием аллель-специфических зондов. Для генотипирования использовали два фланкирующих праймера, прямой (SEQ ID NO 1) и обратный (SEQ ID NO 2), а также аллель-специфические зонды: T-аллель-специфичный флуоресцентно-меченый зонд (SEQ ID NO 3), G-аллель-специфичный флуоресцентно-меченый зонд (SEQ ID NO 4).
- ПЦР в «реальном времени» проводили в 25 мл реакционной смеси, содержащей 1,25 ЕД ДНК-полимеразы Hot Start Taq («Биолабмикс», Новосибирск, Россия), 20 нг ДНК, по 10 мкМ каждого праймера, по 5 мкМ каждого зонда, 0,03 мМ каждого dNTP, 3,0 мМ MgCl2; 1хПЦР-буфер [67 мМ Tris-HCl, pH 8,8, 16,6 мМ (NH4)2SO4, 0,01% Tween-20]. Реакция амплификации состояла из стадии нагревания до 50°C в течение 2 минут, 95°C в течение 10 минут, затем 39 циклов [95°C в течение 10 секунд и 48°C в течение 1 минуты].
4. Генотипирование. При проведении ПЦР в амплификаторе с флуоресцентной детекцией (Bio-Rad CFX96 или аналогичном амплификаторе) генотипирование осуществляют по данным величин RFU (относительных единиц флуоресценции). Для rs6677 (T>G) гена C11orf58 зонд с флуоресцентным красителем FAM соответствует аллелю T, зонд с красителем ROX - аллелю G (фиг. 1). На фигуре видно четкое разделение образцов на кластеры, где черный ромб соответствуют отрицательному контролю, кластер оранжевых кругов - соответствует зонду с флуоресцентным красителем FAM и позволяет идентифицировать гомозигот T/T. Кластер синих квадратов соответствует зонду с красителем ROX и позволяет идентифицировать гомозигот G/G. Кластер зеленых треугольников позволяет идентифицировать гетерозигот T/G.
Резюме
Таким образом, разработан эффективный и недорогой способ для экспресс-идентификации полиморфного варианта rs6677 (T>G) гена C11orf58 у человека методом ПЦР в режиме «реального времени» с применением аллель-специфических флуоресцентных зондов, который может быть использован в медицине при определении наследственной предрасположенности к развитию заболеваний, ассоциированных с носительством полиморфизмов гена C11orf58, а также в научных целях.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Способ
генотипирования полиморфного локуса rs6677 (T G) гена C11orf58 у
человека методом ПЦР в режиме «реального времени» с применением
аллель-специфических флуоресцентных зондов.xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2024-02-14">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023134714/10(076249)</ApplicationNumberText>
<FilingDate>2023-12-22</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>1830</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное образовательное учреждение высшего образования
"Курский государственный медицинский университет"
Министерства здравоохранения Российской Федерации,</ApplicantName>
<ApplicantNameLatin>Kursk State Medical
University</ApplicantNameLatin>
<InventionTitle languageCode="ru">Способ генотипирования
полиморфного локуса rs6677 (T>G) гена C11orf58 у человека методом
ПЦР в режиме «реального времени» с применением аллель-специфических
флуоресцентных зондов</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>27</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..27</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ttgttatagtagagctgttcattatgg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>25</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..25</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tgtgaaaagtacttgctctcatgtt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>17</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..17</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cttagtttcctgttact</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>17</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..17</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cttagttgcctgttact</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к области биотехнологии, молекулярно-генетической диагностике, в частности к способу генотипирования полиморфного локуса rs6677 (T>G) гена C11orf58 у человека методом ПЦР в режиме «реального времени» с применением аллель-специфических флуоресцентных зондов, предусматривающий проведение ПЦР с использованием специально подобранных праймеров (прямого 5'-TTGTTATAGTAGAGCTGTTCATTATGG-3' и обратного 5'-TGTGAAAAGTACTTGCTCTCATGTT-3') и зондов c флуорофорами (T-аллель-специфичного флуоресцентно-меченого зонда 5'-(FAM)CTTAGTTTCCTGTTACT(RTQ1)-3' и G-аллель-специфичного флуоресцентно-меченого зонда 5'-(ROX)CTTAGTTGCCTGTTACT(BHQ2)-3') в амплификаторе с детекцией флуоресценции. Изобретение позволяет расширить арсенал способов генотипирования полиморфных вариантов гена C11orf58, отличается простотой, точностью и низкой себестоимостью. 1 ил.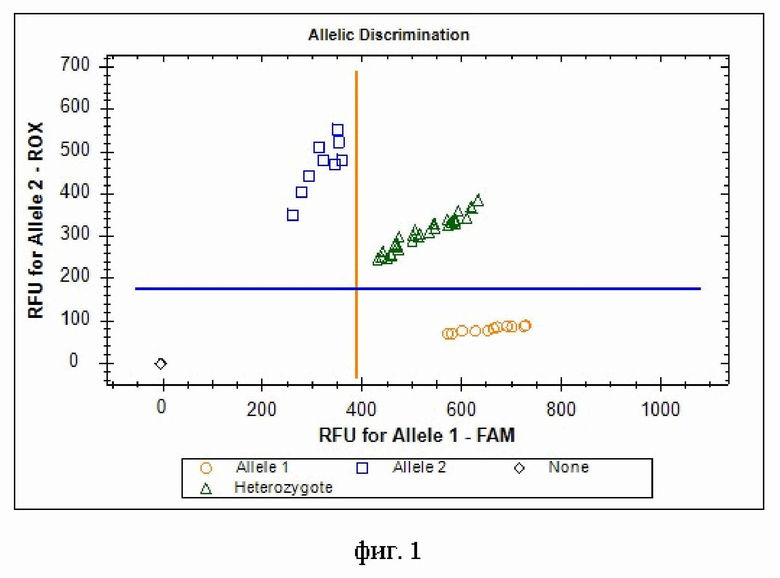
Способ генотипирования полиморфного локуса rs6677 (T>G) гена C11orf58 у человека методом ПЦР в режиме «реального времени» с применением аллель-специфических флуоресцентных зондов, отличающийся тем, что идентификацию аллельных вариантов rs6677 (T>G) гена C11orf58 осуществляют с использованием прямого праймера rs6677 5'-TTGTTATAGTAGAGCTGTTCATTATGG-3' (SEQ ID NO 1), обратного праймера rs6677 5'-TGTGAAAAGTACTTGCTCTCATGTT-3' (SEQ ID NO 2), rs6677-T-аллель-специфичного флуоресцентно-меченого зонда 5'-(FAM)CTTAGTTTCCTGTTACT(RTQ1)-3' (SEQ ID NO 3), rs6677-G-аллель-специфичного флуоресцентно-меченого зонда 5'-(ROX)CTTAGTTGCCTGTTACT(BHQ2)-3' (SEQ ID NO 4).
| Способ генотипирования полиморфного локуса rs346158 (T>C) гена C19orf53 у человека методом ПЦР в режиме "реального времени" с применением аллель-специфических флуоресцентных зондов | 2023 |
|
RU2808839C1 |
| Способ генотипирования полиморфного локуса rs10104 (A>G) гена C19orf53 у человека методом ПЦР в режиме «реального времени» с применением аллель-специфических флуоресцентных зондов | 2023 |
|
RU2808842C1 |
| EP 4032988 A1, 27.07.2022 | |||
| ZHU W | |||
| et al., Gene-based GWAS analysis for consecutive studies of GEFOS, Osteoporos Int., 2018, Volume 29(12), pp | |||
| Дирижабль | 1923 |
|
SU2645A1 |
| BUSHUEVA O | |||
| et al., Genes, encoding heat-resistant obscure (Hero) proteins: new players in ischemic stroke pathogenesis, Abstracts: | |||

