ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к биотехнологии. Заявлен ген дарбэпоэтина альфа, клонированный в экспрессионный вектор, полученный на его основе эукариотический штамм-продуцент и способ получения дарбэпоэтина альфа.
УРОВЕНЬ ТЕХНИКИ
Эритропоэтин является цитокином, который стимулирует эритропоэз. Его рекомбинантные варианты широко используются для коррекции анемии при различных заболеваниях, включая хроническую почечную недостаточность (диализные и предиализные пациенты), онкологические заболевания, хронические воспалительные заболевания кишечника (болезнь Крона, язвенный колит), а также при трансплантации органов и тканей, в пред- и послеоперационный период, у ослабленных пациентов и так далее (см. Эритропоэтин и его биологическая роль, О.Н. Варламова и др., Medicine: theory and practice том 4 №3 2019).
Эритропоэтин (ЭПО) является первым цитокином, который был клонирован и получен в виде рекомбинантного белка. Американская компания «Amgen» разработала биотехнологию получения эпоэтина альфа в системе клеток СНО (US 677813). Эпоэтин бета, синтезируемый клетками почки хомячка (СНО), был разработан фирмами «F. Hoffmann-LaRoche» (NeoRecormon®) и «Chugai» (Эпогин®). Кроме того, был разработан препарат эпоэтин омега, фиромй Epomax. Европейские фирмы «Transkaryotic» и «Aventis» разработали технологию получения препарата Dynepo (эпоэтин дельта), который синтезируется опухолевыми клетками фибросаркомы человека (НТ-1080). В Российской Федерации, разработаны и выпускаются препараты эпоэтина альфа (ГосНИИ ОЧБ) и эпоэтина бета («Фармапарк», «Микроген», «Биннофарм», «Лэнс-Фарм» и ЗАО МТХ) (Препараты рекомбинантных эритропоэтинов и их характеристика, Меркулов В.А. и др., Федеральное государственное бюджетное учреждение «Научный центр экспертизы средств медицинского применения» Министерства здравоохранения Российской Федерации, Москва, 22.08.2013 г.
ЭПО человека представляет собой гликозилированный белок массой около 30,4 кДа, полипептидная цепь которого состоит из 165 аминокислотных остатков. В результате посттрансляционных модификаций к полипептидной цепи белка происходит присоединение N- и O-связанных углеводных цепей, на долю углеводного компонента приходится 40% молекулярной массы. Каждая из N- связанных цепей может содержать максимум 4 остатка сиаловых кислот, тогда как O-связанная цепь может содержать не более двух остатков. Таким образом, ЭПО может содержать от 4 до 14 остатков сиаловых кислот, что соответствует минимальному и максимальному уровню сиалирования (Egrie JC, Grant JR, Gillies DK, Aoki KH, Strickland TW. S7.7 The role of carbohydrate on the biological activity of erythropoietin. Glycoconjugate Journal 1993, 10:263- 263).
Различные изоформы ЭПО, отличающиеся степенью сиалирования и, как следствие, зарядом, могут быть разделены методами изоэлектрического фокусирования в геле капиллярного электрофореза.
В рамках экспериментов по повышению стабильности ЭПО в крови больных было создано множество новых молекул ЭПО, одна из них, названная NM279, содержащая 4 N-присоединенные углеводные цепи, проявила увеличенную биологическую активность (исследования проводились на полицитемических мышах). Дальнейшие опыты привели к созданию молекулы NM321 (дарбэпоэтин альфа), которая содержала 5 N-связанных углеводных цепей, дополнительные углеводные цепочки присоединялись к остаткам Asn 30 и Asn 88 (Egrie JC, Browne JK. Development and characterization of novel erythropoiesis stimulating protein (NESP). Br J Cancer 2001, 84 Suppl 1:3-10). Для создания данной молекулы было проведено пять следующих аминокислотных замен в природной молекуле ЭПО: Ala30Asn, His32Thr, Pro87Val, Trp88Asn и Pro90Thr. Введение двух дополнительных N-связанных углеводных компонентов позволило увеличить максимальное количество остатков сиаловых кислот с 14 до 22. Дополнительный углеводный фрагмент дарбэпоэтина привел к увеличению молекулярной массы белка с 30,400 (для нативного ЭПО) до 38,000. Содержание углеводной части в дарбэпоэтине составляет около 52%, тогда как в ЭПО оно составляет 40%. Сродство молекулы дарбэпоэтина к рецептору эритропоэтина оказалось меньше, чем у природного лиганда, однако данный факт компенсируется его гораздо большей стабильностью.
Таким образом дарбэпоэтин альфа представляет собой сверхсиалированный аналог человеческого эритропоэтина альфа (ЭПО-альфа), имеющий увеличенный период полувыведения из организма по сравнению с нативной и рекомбинантной формой этого гормона. Он имеет такой же механизм действия, как и ЭПО, связывается с тем же поверхностным мембранным рецептором и запускает ту же цепь молекулярных взаимодействий внутри клетки. Дополнительные N-связанные углеводные цепи дарбэпоэтина альфа приводят к большей метаболической устойчивости молекулы in vivo, таким образом время его полувыведения при внутривенном введении в три раза больше, по сравнению со временем полувыведения ЭПО- альфа. Данное свойство дарбэпоэтина позволяет вводить его пациентам менее часто. Исследования показали, что введение дарбэпоэтина раз в неделю позволяет поддерживать уровень гемоглобина у пациентов с почечной недостаточностью на постоянном уровне. Дарбэпоэтин с 2007 г. разрешен к применению в России, и так как по сравнению с природным прототипом генно- и гликомодифицированный дарбэпоэтин обладает втрое большим временем жизни в крови больных, он является более предпочтительным при лечении анемий различного происхождения (Cases A. Darbepoetin alfa: a novel erythropoiesis-stimulating protein. Drugs Today (Bare) 2003, 39:477-495, Smith R. Applications of darbepoietin-alpha, a novel erythropoiesis- stimulating protein, in oncology. Curr Opin Hematol 2002, 9:228- 233, Macdougall 1C. Darbepoetin alfa: a new therapeutic agent for renal anemia. Kidney Int Suppi 2002:55-61).
В настоящее время для биосинтеза рекомбинантных белков человека, требующих корректного гликозилирования, обычно применяют клеточные линии СНО, несущие интегрированные в геном гены целевых белков под контролем сильных промоторов клеточного или вирусного происхождения. (Hansen I.A. et al, Systematic Comparison of Constitutive Promoters and the Doxycycline- Inducible Promoter. PLoS One 2010, 5:e10611, Magnusson Т, et al Sustained, high transgene expression in liver with plasmid vectors using optimized promoter-enhancer combinations. J Gene Med 2011, 13:382-391; Tokushige K, et al., Comparison between cytomegalovirus promoter and elongation factor-1 alpha promoter- driven constructs in the establishment of cell lines expressing hepatitis С virus core protein. J Virol Methods 1997, 64:73-80).
В число наиболее эффективных входят промоторы CMV (промоторно-энхансерный элемент предранних генов цитомегаловируса человека), EF1α (промотор альфа-субъединицы фактора 1 элонгации трансляции).
Авторы настоящего изобретения обнаружили, что объединение трех элементов, а именно, промотора фактора элонгации трансляции 1α (альфа EF1), связанного с 3’-концом энхансера цитомеголовируса, и промотора убеквитина С, связанного с 5’- концом энхансера цитомегаловируса, где указанные промоторы расположены относительно друг друга в транс-положении, позволяет значительно увеличить связывание РНК полимеразы 1 с промоторными областями и, тем самым, значительно повысить экспрессию целевого белка, в частности, дарбэпоэтина. При использовании указанных промоторов отдельно, такой уровень экспрессии был значительно ниже.
Кроме того, заявителями был разработан способ очистки, позволяющий более эффективно и дешево получить необходимый вариант белка дарбэпоэтина альфа.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали систему промоторы/энхансер, которая позволяет значительно повысить уровень экспрессии белка, находящегося под контролем такой системы. В ее состав входит промотор альфа EF1, связанный с 3’- концом энхансера цитомеголовируса, и промотор убеквитина С, связанный с 5’-концом энхансера цитомегаловируса. Указанные промоторы расположены относительно друг друга в транс-положении.
Кроме того, был создан вектор экспрессии, содержащий указанную систему промотор/энхансер, и две копии гена дарбэпоэтина альфа.
Также была получена линия клеток-продуцентов, трансформированных вектором экспрессии по настоящему изобретению, а также система лабораторных клеточных банков, на основании лучших клонов этих клеток.
Наконец, был разработан способ культивирования линии клеток продуцента, включающий получение культуральной жидкости, содержащей белок дарбэпоэтина альфа, и разделение полученного белка с помощью хроматографических методов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1. Сравнение кДНК гена эритропоэтина человека и гена дарбэпоэтина альфа.
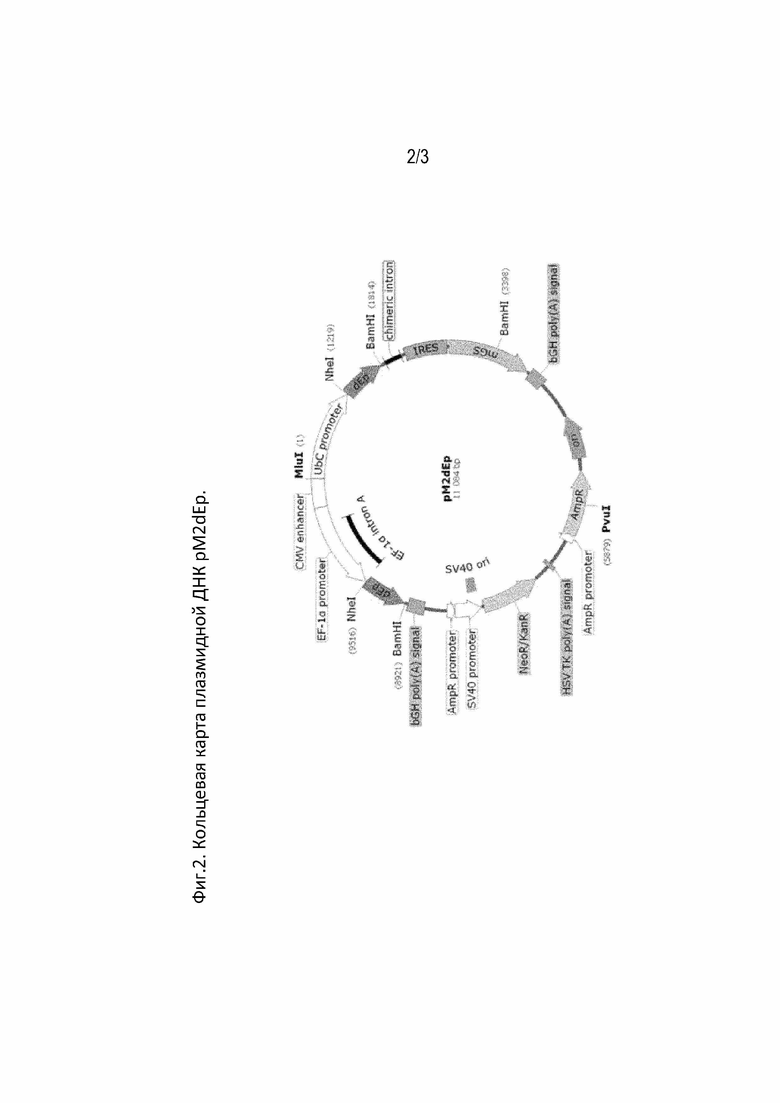
Фиг. 2. Кольцевая карта плазмидной ДНК pM2dEp.
На кольцевой карте плазмидной ДНК рМ2dEp указано следующее: CMV enhancer - энхансер CMV промотора, UbiC -промотор убиквитина
С, dEPO - ген дарбэпоэтина, chimeric intron - химерный вариант интронов аденовируса и генов тяжелой цепи иммуноглобулинов, IRES
- участок внутренней посадки рибосомы вируса энцефаломиокардита, mGS - глутаминсинтетаза Mus musculus, bGH poly (A) - сигнал полиаденилирования бычьего гормона роста, ori - точка начала репликации pUC, AmpR - β-лактамаза, промотор AmpR - промотор AmpR, сигнал HSV TK poly (A) - сигнал полиаденилирования тимидинкиназы вируса простого герпеса, NeoR/KanR - аминогликозидфосфотрансфераза Tn5, SV40 - энхансер и ранний промотор SV40, AmpR - промотор AmpR и ген устойчивости к ампициллину, dEPO - ген дарбэпоэтина, EF-1α intron A - интрон, расположенный перед старт-кодоном EF-1α человека, EF-1α - промотор фактора элонгации EF-1α человека, MluI, NheI, BamHI, XbaI, PvuI - места разрезания соответствующими эндонуклеазами.
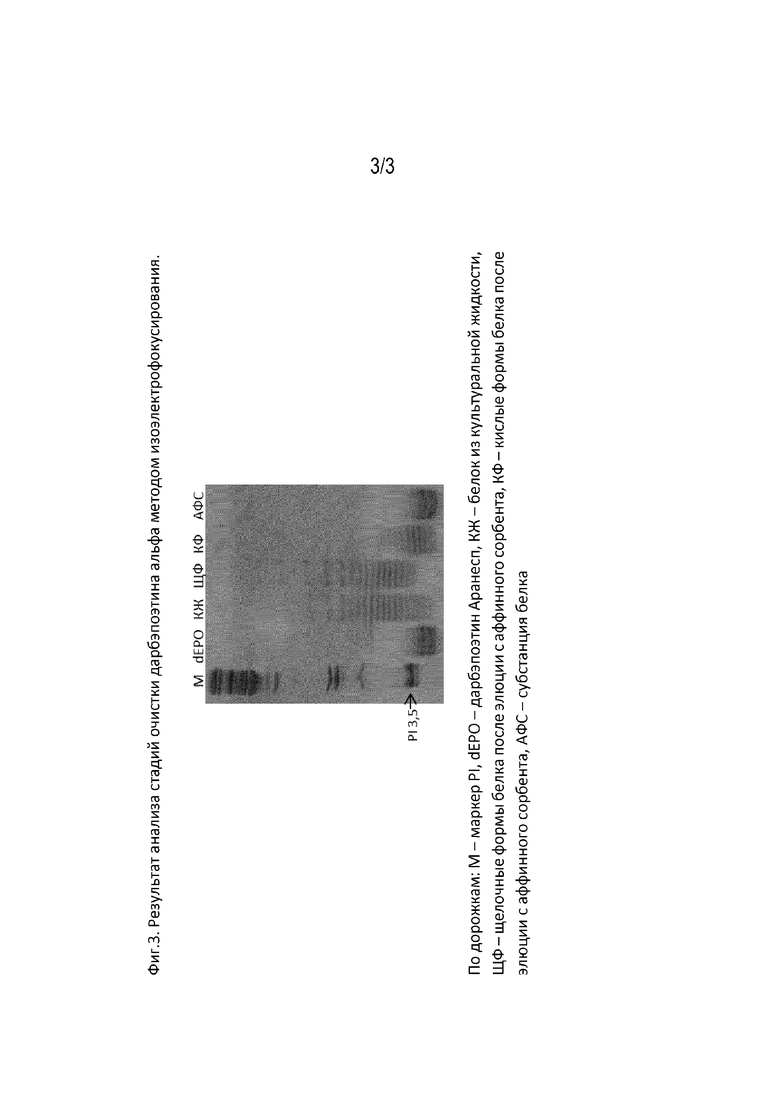
Фиг. 3. Результат анализа стадий очистки дарбэпоэтина альфа методом изоэлектрофокусирования.
Дорожки: М - маркер PI, dEPO - дарбэпоэтин «Аранесп», КЖ - белок из культуральной жидкости, ЩФ - щелочные формы белка после элюции с аффинного сорбента, КФ - кислые формы белка после элюции с аффинного сорбента, АФС - субстанция белка, полученная способом по изобретению.
ПРИМЕРЫ
Пример 1. Конструирование экспрессионных плазмид для обеспечения продукции дарбэпоэтина в клетках СНО
кДНК гена эритропоэтина человека (hEPO) бала получена методом обратной транскрипции с последующей ПЦР на матрице тотальной РНК, выделенной из клеток линии CHO-ЭПО SPM (ООО Протеиновый контур), с использованием праймеров 5’TGCTAGCCGGAGATGGGGGTGCAC3’ и 5’CGGATCCAGTCATCTGTCCCCTGTCCTGC3’
(Seq1). Внесение замен в ген hEPO: G(169)A, C(170)A, C(175)G, A(176)C, C(340)G, C(341)T, G(342)C, T(343)A, G(344)A, G(345)C и
C(349)A, проводили методом праймер специфического мутагенеза с последующим синтезом гена дарбэпоэтина (dEP) перекрывающимся ПЦР. Для этих целей использовали праймеры:
5’TGCTAGCCGGAGATGGGGGTGCAC3’ (SEQ ID NO:1)
5’CGGATCCAGTCATCTGTCCCCTGTCCTGC3’ (SEQ ID NO:2)
5’GCTGCAGGTTTCATTACAGCCCGTCGTG3’ (SEQ ID NO:3)
5’GGCTGTAATGAAACCTGCAGCTTGAATGAG3’ (SEQ ID NO:4)
5’GCAGGGTCTCGTTGACCTGGGAAGAGTTGAC3’ (SEQ ID NO:5)
5’CTCTTCCCAGGTCAACGAGACCCTGCAGCTG3’ (SEQ ID NO:6)
В результате была получена кДНК гена дарбэпоэтина альфа (SEQ ID NO:9).
Ген дарбэпоэтина был клонирован по сайтам рестрикции NheI - BamHI в вектора pSeq-EF1 и pZ-UbiC-GS (ООО Протеиновый контур). В результате были получены экспрессионные вектора pSeqdEp и pZdEp, каждый из которых нес вставку кДНК гена дарбэпоэтина. На матрице плазмиды pSeqdEp ставили ПЦР c использование праймеров
5’GTTCTTTCACGCGTTGACATTGATTATTG3’ (SEQ ID NO:7)
5’CCGACGTTGGTCGCGAGCCCTGGGCCTTCAC3’ (SEQ ID NO:8).
Амплифицированный фрагмент ДНК, несущий элементы CMVenx/hEF1/dEp/pABGH/SV40/Neo/pATK клонировали в вектор pZdEp, по сайтам рестрикции MluI и NruI. В результате был получен экспрессионный вектор pM2dEP, содержащий две последовательности гена дарбэпоэтина в двух экспрессионных кассетах: hEF1/dEp/pA BGH и UbiC/dEp/IVS/IRES/mGS/pA BGH, объединенных энхансером CMVenx (Фиг.1.).
Пример 2. Получение стабильного клона-продуцента дЭПО-альфа
Трансфекцию клеток линий CHO K1 (ИНЦ РАН) линеаризованной плазмидной ДНК pM2dEP по сайту рестрикции PvuI проводили с использованием реагента TurboFect Transfection Reagent (Thermo Scientific) в соответствии со стандартным протоколом производителя. Для этого за сутки до проведения трансфекции в лунки 24-луночного планшета (Corning) высевали по 5×104 клеток. Для трансфекции использовали 1 мкг плазмидной ДНК pM2dEP, рестрицированной по сайту PvuI, и 3 мкл реагента TurboFect на одну лунку. В контрольные лунки ДНК не добавляли. Через 24 часа после трансфекции экспериментальные и контрольные лунки были переведены в селективные условия: полная ростовая среда без глютамина, содержащая 500 мкг/мл G418. Во всех лунках культуральную среду меняли каждые 3 дня. На 9 день после начала селекции клетки отрицательных контролей умерли. Заполнение лунок клетками основного эксперимента составляло около 70% поверхности. Клетки снимали раствором Версена и использовали для клонирования в 96-луночных планшетах, методом придельных разведений, в селективных условиях. В результате микроскопического исследования было выявлено 250 индивидуальных клонов. Через 15 дней заполнение лунок индивидуальных клонов составило 80%. Анализ экспрессии дарбэпоэтина в культуральной жидкости проводили методом ИФА. 83 лучших клона были перенесены в 24-луночные планшеты. Через 7 дней культивирования лунки имели заполнение более 80%. На основании критериев роста и продуктивности были отобраны 13 лучших клонов, на клеточном материале которых был поставлен анализ по двух месячному культивированию клеток в флаконах Т-25, в накопительной среде IMDM, 1%FcII со сменой среды и забором проб каждые 2 дня. По результатам анализа были отобраны клоны 13B7-6,6 мг/л и 11D4-8,3 мг/л (средняя концентрация по эксперименту). Для каждого из клонов была создана система лабораторных клеточных банков.
Пример 3. Масштабирование культивирования
Культивирование в роллерных флаконах (Corning) с площадью поверхности 850 см2 проводили в 250 мл накопительной среды, содержащей IMDM, 1%FcII, 10мМ Галактоза, 1 мМ Уридин, 1 мкМ MnCl2, 0,25 мМ N-ацетил-D-глюкозамин. Культивирование проводили при температуре 36°С и скорости вращения 1,5 об/мин. Слив культуральной жидкости осуществляли каждые два дня. Концентрацию дарбэпоэтина в культуральной жидкости определяли методом ИФА. Всего проводили до 15 сливов. В условиях цикла масштабирования перспективный клон 13B7 показал в среднем продуктивность на уровне 8,3 мкг/мл, при содержании изоформ Аранесп в среднем более 32%
Пример 4. Полный цикл хроматографической очистки дарбэпоэтина альфа из культуральной жидкости
Перед проведением хроматографических стадий в КЖ добавляли 10% раствор азида натрия до конечной концентрации 0,05% и фильтровали через каскад фильтров: глубинный фильтр с размером пор 10 мкм и стерилизующий фильтр с размером пор 0,22 мкм. Значение pH КЖ фиксировали в диапазоне 6,8-7,2.
Для осуществления стадии аффинной хроматографии использовали иммуносорбент PC/ED7 Sepharose FF (ООО Протеиновый контур), упакованный в хроматографическую колонну высотой слоя
10 см. На колонку, уравновешенную раствором PBS+0,05% NaN3 наносили культуральную жидкость. Промывали сорбент растворами: PBS+0,05% NaN3; PBS+0,05% Tween 20; PBS+0,05% Tween 20+1M NaCl и
20 мМ ТРИС гидрохлорид pH 7,50. Элюировали щелочные формы белка раствором глицина гидрохлорида pH 3,20. Элюировали кислые формы белка раствором глицина гидрохлорида pH 2,19. Полученный элюат нейтрализовали добавлением 1М раствора ТРИС до значения pH 7,50. Измеряют концентрацию дарбэпоэтина альфа в нейтрализованном элюате спектрофотометрическим методом. Выход на стадии составлял 37%.
Для осуществления стадии анионообменной хроматографии использовали сорбент YMC BioPro Q75, упакованный в хроматографическую колонну высотой слоя 13см. Колонку уравновешивали раствором 50 мМ уксусной кислоты, 60 мМ ТРИС, 20 мМ хлорида натрия. Полупродукт после стадии аффинной хроматографии наносили на сорбент и промывали растворами, содержащими уксусную кислоту, ТРИС гидрохлорид, хлорид натрия и глицин гидрохлорид pH 2,87 Элюировали кислые формы целевого белка раствором 50 мМ уксусной кислоты, 60 мМ ТРИС гидрохлорид,
400 мМ хлорида натрия. Измеряли концентрацию дарбэпоэтина альфа в полученном элюате спектрофотометрическим методом. Выход на стадии составлял 25%.
Для осуществления стадии гель фильтрации использовали молекулярные сита Sephacryl S-200 HR, упакованные в хроматографическую колонну высотой слоя 60-90см. Колонку уравновешивали раствором для субстанции. Полупродукт после стадии анионообменной хроматографии наносили в объеме, равном 2,5-3,5% от объема колонны. Элюировали 1,2 колоночными объемами раствора для субстанций. Измеряли концентрацию дарбэпоэтина альфа в полученном элюате спектрофотометрическим методом. Полупродукт после хроматографической очистки фильтруют через стерилизующий фильтр с размером пор 0,22 мкм и хранят при минус 70°C. Анализ по стадиям очистки методом изоэлектрофокусирования представлен на фиг. 2.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Общество с ограниченной ответственностью «Протеиновый контур»
<12 0> ВЕКТОР ЭКСПРЕССИИ, ЭУКАРИОТИЧЕСКИЙ ШТАММ ПРОДУЦЕНТ И СПОСОБ
ПОЛУЧЕНИЯ ДАРБЭПОЭТИНА АЛЬФА <160> 2
<170> PatentIn версия 3.5
<210> 1 <211> 24
<212> Праймер
<400> 1
5' TGCTAGCCGGAGATGGGGGTGCAC 3'
<210> 2
<211> 29 <212>
Праймер
<400> 2
5' CGGATCCAGTCATCTGTCCCCTGTCCTGC 3'
<210> 3
<211> 28 <212>
Праймер
<400> 3
5' GCTGCAGGTTTCATTACAGCCCGTCGTG 3'
<210> 4
<211> 30
<212> Праймер
<400> 4
5' GGCTGTAATGAAACCTGCAGCTTGAATGAG 3'
<210> 5
<211> 31
<212> Праймер
<400> 5
5' GCAGGGTCTCGTTGACCTGGGAAGAGTTGAC 3'
<210> 6 <211> 31
<212> Праймер <400> 6
5' CTCTTCCCAGGTCAACGAGACCCTGCAGCTG 3'
<210> 7 <211> 29
<212> Праймер
<400> 7
5' GTTCTTTCACGCGTTGACATTGATTATTG 3'
<210> 8
<211> 31
<212> Праймер
<400> 8
5' CCGACGTTGGTCGCGAGCCCTGGGCCTTCAC 3'
<210> 9
<211>
<212> кДНК DarbEPO
<400> 9
5'atgggggtgcacgaatgtcctgcctggctgtggcttctcctgtccctgctgtcgctccctct
gggcctcccagtcctgggcgccccaccacgcctcatctgtgacagccgagtcctggagaggtac
ctcttggaggccaaggaggccgagaatatcacgacgggctgtAAtgaaACctgcagcttgaatg
agaatatcactgtcccagacaccaaagttaatttctatgcctggaagaggatggaggtcgggca
gcaggccgtagaagtctggcagggcctggccctgctgtcggaagctgtcctgcggggccaggcc
ctgttggtcaactcttcccagGTCAACgagAccctgcagctgcatgtggataaagccgtcagtg
gccttcgcagcctcaccactctgcttcgggctctgggagcccagaaggaagccatctcccctcc
agatgcggcctcagctgctccactccgaacaatcactgctgacactttccgcaaactcttccga
gtctactccaatttcctccggggaaagctgaagctgtacacaggggaggcctgcaggacagggg
acagatga3'
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм линии клеток яичника китайского хомячка | 2023 |
|
RU2805879C1 |
| ЭУКАРИОТИЧЕСКАЯ КЛЕТКА-ХОЗЯИН ДЛЯ ЭКСПРЕССИИ ДАРБЭПОЭТИНА, ВЕКТОР ЭКСПРЕССИИ И СПОСОБ ПОЛУЧЕНИЯ ДАРБЭПОЭТИНА АЛЬФА | 2013 |
|
RU2523948C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 | 2016 |
|
RU2671477C2 |
| Способ получения димерной формы мутантного иммуноглобулина IgA2m1-изотипа в клетках млекопитающих | 2023 |
|
RU2822889C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA1 | 2016 |
|
RU2656142C1 |
| Способ получения димерной формы иммуноглобулина IgA1-изотипа в клетках млекопитающих | 2023 |
|
RU2822890C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-Intht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IgA1 | 2016 |
|
RU2664184C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| ГИБРИДНЫЙ БЕЛОК НА ОСНОВЕ РЕКОМБИНАНТНОГО ЭРИТРОПОЭТИНА ЧЕЛОВЕКА, ОБЛАДАЮЩИЙ ПРОЛОНГИРОВАННЫМ ДЕЙСТВИЕМ (ВАРИАНТЫ), И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2515914C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ХИМЕРНОЕ АНТИТЕЛО ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ-АЛЬФА ЧЕЛОВЕКА, ЛИНИЯ ЭУКАРИОТИЧЕСКИХ КЛЕТОК-ПРОДУЦЕНТ ХИМЕРНОГО АНТИТЕЛА И СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО АНТИТЕЛА | 2013 |
|
RU2555533C9 |

Изобретение относится к биотехнологии и генной инженерии и представляет собой систему промоторы/энхансер для увеличения связывания РНК полимеразы 1 с промоторными областями, состоящую из промотора фактора элонгации трансляции 1α (альфа EF1), связанного с 3’-концом энхансера цитомеголовируса, и промотора убеквитина С, связанного с 5’-концом энхансера цитомегаловируса. Изобретение касается вектора экспрессии, содержащего ген целевого белка под контролем указанной системы, где ген целевого белка представляет собой две копии гена дарбэпоэтина альфа. Изобретение позволяет эффективно осуществить способ получения стабильного клона дарбэпоэтина альфа, включающий трансфекцию клеточной линии заявляемым вектором, отбор стабильного продуцента дарбэпоэтина селекцией в среде с последующим клонированием, культивирование стабильного клона в питательной среде. 4 н. и 8 з.п. ф-лы, 3 ил., 4 пр.
1. Система промоторы/энхансер для увеличения связывания РНК полимеразы 1 с промоторными областями, состоящая из промотора фактора элонгации трансляции 1α (альфа EF1), связанного с 3’-концом энхансера цитомеголовируса, и промотора убеквитина С, связанного с 5’-концом энхансера цитомегаловируса.
2. Система по п.1, где промоторы расположены относительно друг друга в транс-положении.
3. Вектор экспрессии, содержащий ген целевого белка под контролем системы по п.1, где ген целевого белка представляет собой две копии гена дарбэпоэтина альфа.
4. Вектор по п.3, где каждая копия гена расположена под контролем одного из двух промоторов, промотора фактора элонгации трансляции 1α (альфа EF1), связанного с 3’-концом энхансера цитомеголовируса, или промотора убеквитина С, связанного с 5’-концом энхансера цитомегаловируса.
5. Способ получения стабильного клона-продуцента дарбэпоэтина альфа, включающий:
а) трансфекцию клеточной линии вектором по п.3,
b) отбор стабильного продуцента дарбэпоэтина селекцией в среде с последующим клонированием,
с) культивирование стабильного клона-продуцента в питательной среде.
6. Способ по п.5, где клеточная линия представляет собой клетки китайского хомячка (СНО К1).
7. Способ по п.5-6, где селективная среда содержит G418 и не содержит глютамин.
8. Способ по п.5-7, где способ культивирования является роллерным.
9. Способ по п.5-8, где способ культивирования представляет собой 3D культивирование.
10. Способ хроматографической очистки дарбэпоэтина альфа, содержащий получение стабильного клона-продуцента дарбэпоэтина способом по любому из п.5-9, осуществление аффинной хроматографии для элюирования щелочных и кислых форм белка, последующее осуществление анионообменной хроматографии для элюирования кислых форм целевого белка.
11. Способ по п.10, включающий разделения вариантов белка с разным уровнем гликозилирования проведением дифференциальной элюции различными растворами.
12. Способ по п.10 или 11, включающий использование раствора глицина гидрохлорид с разными значениями рН.
| Egrie JC, Browne JK | |||
| Development and characterization of novel erythropoiesis stimulating protein (NESP) | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| ЭУКАРИОТИЧЕСКАЯ КЛЕТКА-ХОЗЯИН ДЛЯ ЭКСПРЕССИИ ДАРБЭПОЭТИНА, ВЕКТОР ЭКСПРЕССИИ И СПОСОБ ПОЛУЧЕНИЯ ДАРБЭПОЭТИНА АЛЬФА | 2013 |
|
RU2523948C1 |
| WO 2011070405 А1, 16.06.2011 | |||
| КАРБАМИЛИРОВАННЫЙ ДАРБЭПОЭТИН 9С-DEPO, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ ЕГО В КАЧЕСТВЕ ЛЕКАРСТВЕННОГО СРЕДСТВА С ЦИТОПРОТЕКТОРНЫМ ДЕЙСТВИЕМ | 2013 |
|
RU2575773C2 |
| НУРБАКОВ А.А, и др., Оптимизация технологии культивирования с подпиткой клеток CHO, экспрессирующих рекомбинантный | |||

