Область изобретения
Настоящее изобретение относится к модифицированному онколитическому аденовирусу; фармацевтической композиции, содержащей указанный аденовирус; и способу лечения рака с его применением.
Предшествующий уровень техники
Восприятие роли онколитических вирусов в лечении рака резко изменилось за последнее десятилетие, поскольку приобрели популярность иммунотерапия и стимуляция собственной иммунной системы пациента для нацеливания и воздействия на рак. В начале столетия онколитические вирусы воспринимали как активные агенты в лечении рака, действующие исключительно благодаря присущей им способности лизировать опухолевые клетки посредством онколиза. В последнее время вызывает интерес их применение в качестве противораковых вакцин, и их способность высвобождать опухолевые антигены из раковых клеток при онколизе для активации иммунной системы признана важной характеристикой при разработке окончательной иммунотерапии против рака.
Аденовирусы представляют собой высоко иммуногенные вирусы, часто используемые в качестве векторов при различных подходах для создания вакцин против инфекционных заболеваний. Важно отметить, что они обладают исключительной способностью примировать и стимулировать иммунные ответы. Кроме того, присутствие онколитического аденовируса в опухоли и вызываемая им опосредуемая иммунными клетками гибель клеток, вероятно, формируют агрессивное микроокружение в опухоли в сторону более восприимчивого состояния для возникновения клинически значимого противоопухолевого иммунитета, вызывая экспрессию иммуномодуляторов TH1-типа, таких как IFN-гамма. Однако иммуногенность онколитических аденовирусов является палкой о двух концах; антивирусный иммунитет часто бывает настолько более сильным, что он подавляет гораздо более слабый иммунный ответ, вызванный против аутоантигенов, экспрессируемых опухолью.
Пептидные вакцины были перспективной концепцией в иммунотерапии рака, но клиническая эффективность традиционных подходов к пептидным вакцинам общепризнана низкой. Вакцины против рака доставляют раковые антигены в комбинации с адъювантом, который должен обеспечивать необходимые воспалительные сигналы для усиления противоопухолевого иммунного ответа. Однако центральная и периферическая толерантность, а также процесс иммунного редактирования при прогрессировании опухоли обычно приводят к потере реактивности Т-клеток, которая наблюдается in vitro при выборе пептидов-кандидатов для пептидных вакцин. Современные адъюванты вакцин просто недостаточно эффективны, чтобы нарушить толерантность иммуносупрессивной опухоли, даже если указанная опухоль экспрессирует выбранные антигены. Кроме того, многие подходы создания пептидных вакцин основаны на коротких пептидах, которые соответствуют точным, минимальным последовательностям эпитопов CD8+ Т-клеток, связывающихся с MHC класса I (главный комплекс гистосовместимости класса I). Несмотря на обнадеживающие результаты на доклинических моделях опухолей, было показано, что короткие пептиды вызывают системную периферическую толерантность у пациентов, которая предположительно является результатом презентации пептидов непрофессиональными APC (антиген-презентирующие клетки) или незрелыми DC (дендритные клетки), лишенными важных костимулирующих сигналов.
Настоящее изобретение объединяет два вышеуказанных подхода создания вакцин таким образом, чтобы использовать лучшее из обоих. Физическая единица, которая сочетает в себе онколитический аденовирус и раковые антигены, является решением проблемы а) чрезвычайно сильной вирус-специфичной мишени и отсутствия достаточной опухолеспецифичной мишени для иммунных клеток, б) проблемы слабого адъюванта и в) проблемы центральной толерантности, вызванной вакцинами с короткими пептидами. Таким образом, настоящее изобретение относится к аденовирусу, покрытому пептидами, с управляемой репликацией (Peptide-coated Conditionally Replicating Adenovirus - PeptiCRAd), который представляет собой инновационный и уникальный способ объединения двух подходов к иммунотерапии рака c доказанной клинической эффективностью: онколитического аденовируса и пептидной вакцины. В PeptiCRAd используют иммуногенные вирусы в качестве активных носителей опухолеспецифичных пептидов, чтобы направлять иммунную систему на специфичную мишень и уничтожать раковые клетки. Более того, в отличие от онколитических вирусных вакцин, генетически кодирующих опухолевые антигены, покрытие аденовируса иммуногенным пептидом делает технологию PeptiCRAd высокоадаптивной для всех видов рака; один и тот же вирус-носитель может быть использован для лечения всех
видов рака, и адаптация происходит просто путем покрытия вируса различными пептидами, удлиненными полилизином. Эта уникальная технология использует значительную иммуногенность аденовируса и одновременно направляет ответ CD8+ Т-клеток на опухолевую ткань.
Краткое описание изобретения
Согласно первому аспекту настоящего изобретения предложен модифицированный аденовирус или вектор, содержащий:
по меньшей мере один из следующих полипептидов, который ковалентным или нековалентным образом присоединен к вирусному капсиду, но генетически не закодирован указанным аденовирусным вектором
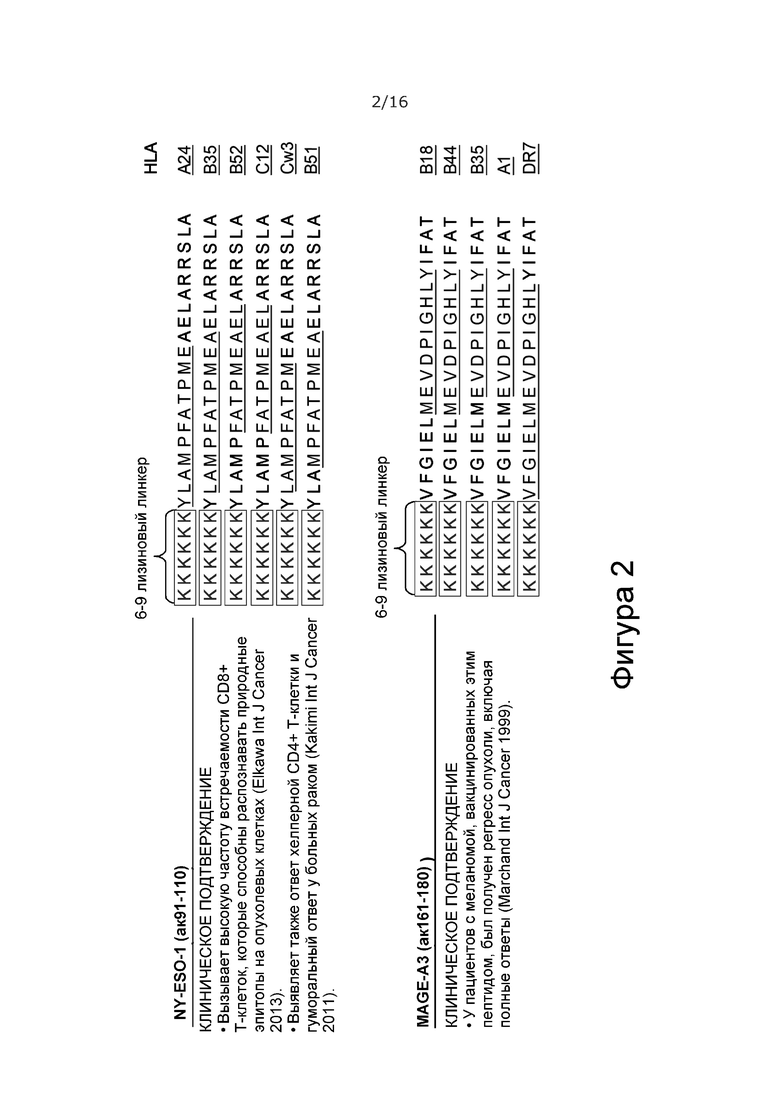
1) VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 1] MAGE-A3 161-180;
2) YLAMPFATPMEAELARRSLA [SEQ ID NO: 2] NY-ESO-1 91-110;
3) RGPESRLLEFYLAMPFATPM [SEQ ID NO: 3] NY-ESO-1 81-100; или
4) полипептид, идентичный указанному выше пептиду по меньшей мере на 60%.
В предпочтительном варианте реализации настоящего изобретения указанный аденовирус способен реплицироваться и обладает литической активностью по отношению к клетках-мишеням.
Еще более предпочтительно указанный полипептид согласно п. 4) имеет 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичности с полипептидом п. 1) или 2), или 3).
Преимущественно указанные полипептиды могут стимулировать специфичный к пептидуиммунный ответ у субъекта и, что еще более предпочтительно, поскольку указанные полипептиды не были генетически кодированы указанным аденовирусным вектором, но были присоединены к капсиду ковалентно или нековалентно, это присоединение может быть выполнено быстро и эффективно, то есть без необходимости ожидания репликации вируса в клетке-хозяине. Обычно для облегчения присоединения по меньшей мере одного, двух или трех полипептидов указанный полипептид(ы) представляет собой удлиненный полилизином полипептид с по меньшей мере 4, в идеальном варианте 5, 6, 7, 8 или 9 остатками лизина. Чаще всего используют 6 или 9 лизинов, которые наиболее предпочтительно присоединены к амино-концу полипептида.
Соответственно, указанные пептиды для прикрепления к указанному аденовирусу представляют собой:
KKKK(KKKKK)-VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 7]; и/или
KKKK(KKKKK)-YLAMPFATPMEAELARRSLA [SEQ ID NO: 8]; и/или,
KKKK(KKKKK)- RGPESRLLEFYLAMPFATPM [SEQ ID NO: 9]; в альтернативном варианте:
KKKKKK-VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 4]; и/или
KKKKKK-YLAMPFATPMEAELARRSLA [SEQ ID NO: 5]; и/или
KKKKKK- RGPESRLLEFYLAMPFATPM [SEQ ID NO: 6].
В еще одном предпочтительном варианте реализации настоящего изобретения указанный аденовирус может относиться к любому типу и виду аденовирусов, например, не ограничиваясь аденовирусом человека, но чаще всего представляет собой аденовирус человека. Наиболее благоприятно то, что указанные аденовирусы способны реплицироваться и уничтожать раковые клетки, направляя противовирусный иммунный ответ против опухоли, таким образом, указанный модифицированный аденовирус или вектор является компетентным по репликации и онколитическим.
Указанный аденовирус или вектор, используемый в настоящем изобретении, может быть создан опухолеспецифичным в отношении репликации, например, указанный аденовирусный вектор может содержать модификации генов E1, E3, E4 и/или L3, такие как вставка опухолеспецифичных промоторов, делеции генов и вставка трансгенов.
В идеальном варианте указанный аденовирус или вектор являются онколитическими, то есть способны инфицировать и уничтожать раковые клетки путем избирательной (селективной) репликации в опухоли, но не в нормальных клетках.
Каркас указанного аденовируса или вектора может быть любого серотипа. В одном варианте реализации настоящего изобретения серотип каркаса аденовируса или вектора выбран из серотипа 3 или 5. В настоящей заявке термин «каркас нуклеиновой кислоты аденовируса серотипа 5 (Ad5)» относится к геному Ad5, а «каркас нуклеиновой кислоты аденовируса серотипа 3 (Ad3)» относится к геному Ad3.
Кроме того, вирус или вектор могут быть химерными, например, векторы Ad5/3, Ad3/5 или Ad5/35. В качестве примера, «вектор Ad5/3» относится к химерному вирусу или вектору, имеющему части обоих векторов Ad5 и Ad3.
Термин «капсид» аденовируса или вектора относится к белковой оболочке вируса. Капсид состоит из нескольких олигомерных структурных субъединиц, составленных из белков, называемых протомерами.
Наиболее предпочтительно указанный аденовирус или вектор относится к серотипу 5, который выбран, поскольку он оптимален для примирования и стимулирования ответа Т-клеток; он индуцирует CD8+ Т-клетки, необходимые для уничтожения опухолей; любой ранее существовавший иммунитет против онколитических вирусов может повысить эффективность внутриопухолевой обработки (лечения) и имеет надлежащие показатели клинической безопасности.
В еще одном предпочтительном варианте реализации настоящего изобретения указанный аденовирус или вектор дополнительно модифицированы с тем, чтобы включить любой один или более из следующих признаков, включая любые и все их комбинации.
Химерная замена Ad5/3, то есть замена области выступа белка фибры аденовируса серотипа 5 на область выступа аденовируса серотипа 3; что позволяет указанному вирусу или вектору обходить нативный рецептор коксаки-аденовируса Ad5 (CAR) и использовать вместо него нативный рецептор Ad3 десмоглеин 2 (DSG2) для интернализации. DSG2 в большом количестве присутствует в раковых клетках.
Аденовирусная инфекция начинается с распознавания рецепторов клетки-хозяина посредством специализированных белков на поверхности вируса, то есть белка фибры (fibre) аденовируса и, в частности, глобулярного домена карбокси-конца белка фибры аденовируса, называемого карбокси-концевым доменом выступа. Соответственно, в настоящей заявке указание на выступ (knob) белка фибры аденовируса относится к глобулярному домену карбокси-конца белка фибры аденовируса.
Делеция гена E1A, где указанная делеция представляет собой делецию нуклеотидов, кодирующих аминокислоты 122-129 (LTCHEACF); указанная делеция является мерой предосторожности, поскольку вирусный или векторный белок E1A не может связываться с молекулой ретинобластомы (Rb) и высвобождать фактор транскрипции E2F из Rb для транскрипции вирусного гена. Таким образом, указанный аденовирус или вектор зависят от присутствия свободного E2F в клетке-хозяине и могут реплицировать свой геном либо в делящихся нормальных клетках, либо в раковых клетках, где свободный E2F постоянно доступен. Таким образом, данная модификация относительно защищает неделящиеся клетки и направлена против делящихся или раковых клеток.
Частичная делеция гена E3, которая, учитывая иммуносупрессивную роль этого генного продукта, усиливает иммуногенность. В идеальном варианте ген 14.7К должен быть частично или полностью делетирован.
В предпочтительном варианте реализации указанный модифицированный аденовирус или вектор содержит вставку по меньшей мере одного трансгена, который кодирует костимуляторную молекулу, и, в идеальном варианте, двух трансгенов, где один из указанных генов приводит к активации врожденной иммунной системы, а другой приводит к активации адаптивной иммунной системы. Предпочтительные трансгены включают CD40L для активации врожденной иммунной системы за счет использования APC для управления ответами CD8+ Т-клеток и OX40L для активации адаптивной иммунной системы за счет увеличения клональной экспансии, выживаемости CD8+ Т-клеток и формирования большого пула Т-клеток памяти.
Ряд трансгенов может быть помещен в разные положения аденовируса или вектора. Один трансген, например, может быть помещен в подвергаемую частичной или полной делеции область E3, либо под контролем промотора E3, либо под контролем экзогенного промотора, или в подвергаемую частичной или полной делеции область E1, либо под контролем промотора E1, либо под контролем экзогенного промотора, или подвергаемую частичной или полной делеции область L3 под контролем промотора L3 или под контролем экзогенного промотора.
Наиболее предпочтительно OX40L, в идеальном варианте OX40L человека, расположен в области E3B, заменяя делецию гена 14.7K.
Также наиболее предпочтительно, CD40L, в идеальном варианте CD40L человека, вставлен в область поздних генов вируса, конкретно в область позднего гена 3 (L3), в идеальном варианте ниже гена 23К.
В предпочтительном варианте реализации оба трансгена вставлены в указанный вирус, то есть OX40L и CD40L.
В альтернативном варианте ДНК, кодирующая OX40L и CD40L, может быть присоединена и вставлена в виде слитой молекулы с использованием известных методов генной инженерии, таких как использование внутреннего сайта связывания рибосомы (IRES) или, что более предпочтительно, с использованием самоотщепляющегося пептида 2A, преимуществом которого является небольшой размер и высокая эффективность расщепления между генами выше и ниже пептида 2А. Обычно CD40L вставляют сразу после OX40L, но можно осуществить настоящее изобретение и с обратной конфигурацией.
Вирус или вектор, используемые в настоящем изобретении, также могут содержать другие модификации, чем описано выше. Возможно могут быть использованы любые дополнительные компоненты или модификации, но они не являются обязательными для настоящего изобретения.
Из вышеизложенного следует, что аденовирус или вектор согласно настоящему изобретению сконструирован для стимуляции иммунного ответа против рака и, в частности, в среде опухоли, где, как правило, иммунная система аномально функционирует из-за механизмов уклонения, используемых раковыми клетками.
Неожиданным образом наши данные показывают, что добавление иммуностимулирующих трансгенов, OX40L человека и CD40L человека, в локус 14.7K не снижает онколитической эффективности вирусов согласно настоящему изобретению по сравнению с вирусом-каркасом Ad5/3D24 или вирусом с иммуностимулирующим трансгеном, например GM-CSF человека, заменяющим удаленные гены gp19K/7.1K. Это является неожиданным, потому что указанные трансгены могут сильно влиять на репликацию вируса из-за размера трансгена и прямого воздействия трансгена на инфицированные клетки. Кроме того, делеция гена 14.7K не так широко изучена, как делеция генов gp19K/7.1K, и, таким образом, может иметь неожиданные последствия для репликативного аппарата вируса, особенно в контексте включения трансгенов в сайт делеции 14.7K.
Авторы настоящего изобретения также демонстрируют, что вирус согласно настоящей заявке способен продуцировать функциональные трансгены человека из локуса гена 14.7К. Это было неожиданно, потому что авторы настоящего изобретения использовали, что несколько необычно, кассету транскрипции с сайтом процессинга вируса 2A между двумя трансгенами.
Кроме того, авторы настоящего изобретения демонстрируют, что вирус согласно настоящему изобретению способен вызывать MAGE-A3 и NY-ESO-1-специфичный иммунный ответ.
Соответственно, настоящее изобретение включает фармацевтическую композицию, содержащую по меньшей мере один модифицированный аденовирус или вектор согласно настоящему изобретению и подходящий носитель. В предпочтительном варианте реализации настоящего изобретения указанная фармацевтическая композиция приготовлена для внутриопухолевой, внутримышечной, внутриартериальной, внутривенной, внутриплевральной, внутрипузырной, внутриполостной или перитонеальной инъекции или для перорального введения.
Соответственно, еще в одном аспекте настоящее изобретение относится к способу лечения рака у индивидуума, включающему введение индивиду эффективного количества фармацевтической композиции, содержащей по крайней мере один аденовирус или вектор согласно настоящему изобретению.
Дополнительно или в альтернативном варианте, настоящее изобретение относится, по меньшей мере, к одному аденовирусу или вектору, или композиции согласно настоящему изобретению для применения при лечении рака.
Дополнительно или в альтернативном варианте, настоящее изобретение относится к применению по меньшей мере одного аденовируса, или вектора или композиции согласно настоящему изобретению для лечения рака.
Дополнительно или в альтернативном варианте, настоящее изобретение относится применению по меньшей мере одного аденовируса или вектора согласно настоящему изобретению в изготовлении лекарственного средства для лечения рака.
Учитывая, что опухоли развили несколько иммуносупрессивных механизмов для противодействия иммунным клеткам организма, терапию согласно настоящему изобретению также осуществляют в комбинации с применением молекулы контрольной точки. Лучше всего охарактеризованные пути контрольных точек представляют собой путь белка цитотоксических Т-лимфоцитов 4 (CTLA-4) и путь белка 1 программируемой гибели клеток (PD-1/PD-L1). Таким образом, аденовирус или вектор согласно настоящему изобретению можно использовать в комбинации с модуляторами контрольных точек, такими как молекулы анти-PD1, анти-PD-L1 или анти-CTLA-4, для противодействия иммуносупрессивной среде опухоли и для того, чтобы вызвать сильный антииммунный ответ.
Указанный модифицированный аденовирус или вектор действует как активный адъювант, поскольку он обеспечивает сигналы опасности, необходимые для оптимального иммунного ответа против целевого пептида, но также, в случае реплицирующегося аденовируса или вектора, он сохраняет свою способность лизировать раковые клетки, заражает и реплицирует в них свой геном. Онколитическое уничтожение клеток является иммуногенным по своей природе, что вызывает изменения в микросреде опухоли, которые, вероятно, усиливают иммунный ответ на пептиды/опухоль. Таким образом, применение заявленного модифицированного аденовируса, физически включенного в комплекс с длинными пептидами (20 аминокислот без полилизинового хвоста), приводит к лучшему противоопухолевому иммунному ответу по сравнению с пептидными вакцинами или только онколитическими вакцинами.
Наиболее предпочтительно в настоящей заявке рак включает любой один или более из следующих видов рака: носоглоточный рак, синовиальный рак, печеночно-клеточный рак, рак почки, рак соединительных тканей, меланому, рак легкого, рак кишечника, рак толстой кишки, рак прямой кишки, колоректальный рак, рак мозга, рак горла, рак полости рта, рак печени, рак кости, рак поджелудочной железы, хориокарциному, гастриному, феохромоцитому, пролактиному, Т-клеточный лейкоз/лимфому, нейрому, болезнь фон Гиппеля-Линдау, синдром Золлингера-Эллисона, рак надпочечника, рак ануса, рак желчного протока, рак мочевого пузыря, рак мочеточника, олигодендроглиому, нейробластому, менингиому, опухоль спинного мозга, рак кости, остеохондрому, хондросаркому, саркому Юинга, рак с неизвестной первичной локализацией, карциноид, карциноид желудочно-кишечного тракта, фибросаркому, рак молочной железы, болезнь Пэджета, рак шейки матки, рак, рак пищевода, рак желчного пузыря, рак головы, рак глаза, рак шеи, рак почки, опухоль Вильмса, рак печени, саркома Капоши, рак предстательной железы, рак яичек, ходжкинскую лимфому, неходжкинскую лимфому, рак кожи, мезотелиому, множественную миелому, рак яичника, рак эндокринных клеток поджелудочной железы, глюкагоному, рак паращитовидной железы, рак пениса, рак гипофиза, саркому мягких тканей, ретинобластому, рак тонкого кишечника, рак желудка, рак вилочковой железы, рак щитовидной железы, трофобластический рак, пузырный занос (хорионаденому), рак матки, рак эндометрия, рак влагалища, рак вульвы, нейрому слухового нерва, фунгоидный микоз, инсулинома, карциноидный синдром, соматостатиному, рак десны, рак сердца, рак губы, рак оболочки головного мозга, рак ротовой полости, рак нерва, рак неба, рак околоушной железы, рак брюшины, рак зева, рак плевры, рак слюнной железы, рак языка и рак миндалины.
В формуле изобретения, которая приведена ниже, и в предыдущем описании изобретения, за исключением случаев, когда контекст требует иного из-за языковых выражений или необходимого смыслового значения, слово «содержит» или варианты, такие как «содержит» или «содержащий», используют в широком смысле, то есть для указания наличия заявленных признаков, без исключения или добавления дополнительных признаков в различных вариантах реализации настоящего изобретения.
Все источники, включая любые патенты или заявки на патенты, указанные в настоящей заявке, включены в настоящую заявку посредством ссылки. Не допускается, чтобы какие-либо источники рассматривались как предшествующий уровень техники. Кроме того, не допускается, чтобы какой-либо из предшествующих уровней техники рассматривался как часть общих знаний в данной области.
Предпочтительные признаки каждого аспекта настоящего изобретения могут быть такими, как описано в связи с любым из других аспектов.
Другие признаки настоящего изобретения станут понятны из следующих примеров. В целом настоящее изобретение распространяется на любой новый или любую новую комбинацию признаков, раскрытых в настоящей заявке (включая прилагаемую формулу изобретения и графические материалы). Таким образом, следует понимать, что признаки, целые числа, характеристики, соединения или химические фрагменты, описанные в связи с конкретным аспектом, вариантом реализации или примером изобретения, применимы к любому другому аспекту, варианту реализации или примеру, описанному в настоящей заявке, если они не являются несовместимыми с ними.
Более того, в настоящей заявке любой признак может быть заменен альтернативным признаком, соответствующим тому же или аналогичному назначению, если не указано иное.
Во всем описании и формуле изобретения единственное число охватывает множественное число, если иное не требуется в контексте. В частности, использование единственного числа следует понимать как предполагающее как множественное число, так и единственное число, если контекст не требует иного.
Далее описан вариант реализации настоящего изобретения только в качестве примера со ссылкой на следующее, в котором:
на Фиг. 1 представлено схематическое изображение модифицированного аденовируса в соответствии с настоящим изобретением. В частности, аденовирус Ad5 модифицируют для включения в него белка области выступа серотипа 3, то есть области выступа Ad3, и посредством использования полилизиновых хвостов по меньшей мере один, а в идеальном варианте два или три полипептида согласно настоящему изобретению присоединяют к вирусному капсиду. Эти полипептиды не являются частью вирусного генома, но ковалентно или нековалентно присоединены к капсиду. Это нововведение, именуемое в настоящей заявке PeptiCRAd-1, таким образом задействует все основные звенья иммунной системы для борьбы с раком.
На Фиг. 2 показаны два выбранных пептида. Они не связываются напрямую с молекулами MHC класса I, но требуют, чтобы процессинг антигена происходил с помощью APC. Следовательно, указанные пептиды не могут связываться с молекулами MHC класса I на непрофессиональных APC, и поэтому не вызывают временной активации и последующей анергии CD8+ Т-клеток. Только активированные профессиональные APC (которые могут получать дополнительную костимуляцию от кодируемого вирусом CD40L, если он присутствует в дальнейшем модифицированном аденовирусе) могут представлять эти пептиды Т-клеткам, что приводит к устойчивой активации и пролиферации пептид-специфичных CD8+ Т-клеток (далее костимуляция и усиленный ответ Т-клеток памяти обеспечивается кодируемым вирусом OX40L, если он присутствует в дополнительном модифицированном аденовирусе).
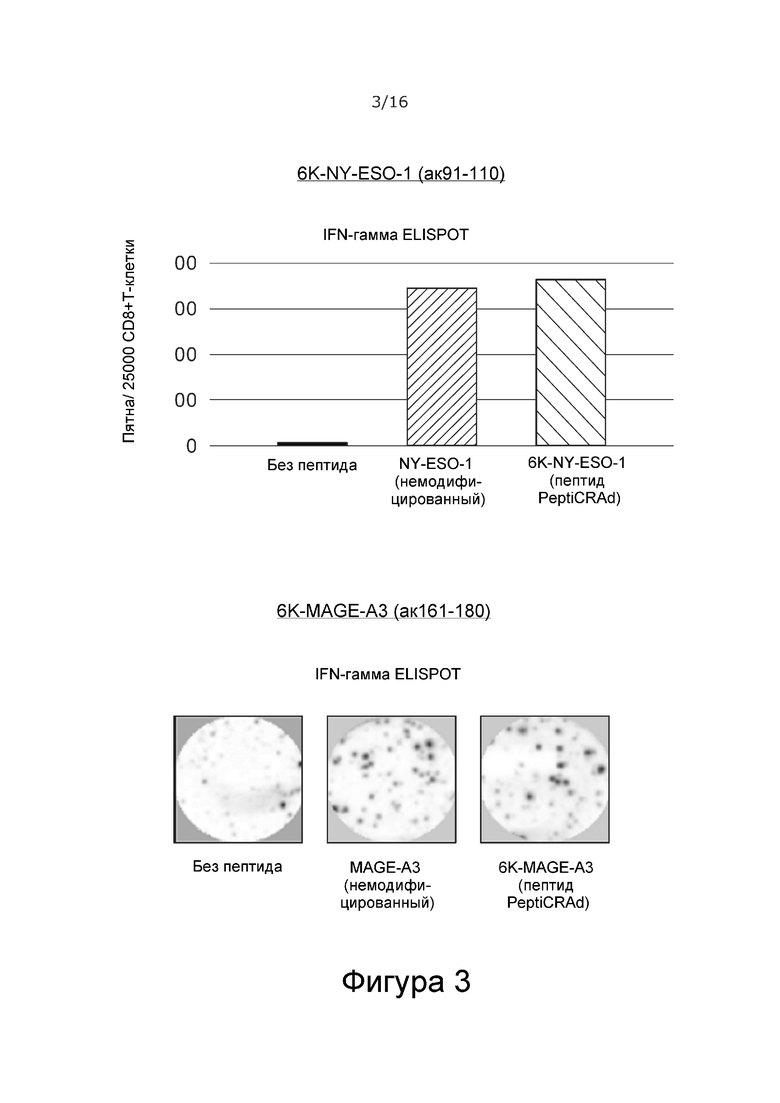
На Фиг. 3 показано, что полилизиновый хвост на N-конце пептидов из Фиг. 2 не изменяет иммунологические свойства указанных пептидов по сравнению с клинически используемыми немодифицированными пептидами с известной способностью к активации иммунной системы у больных раком.
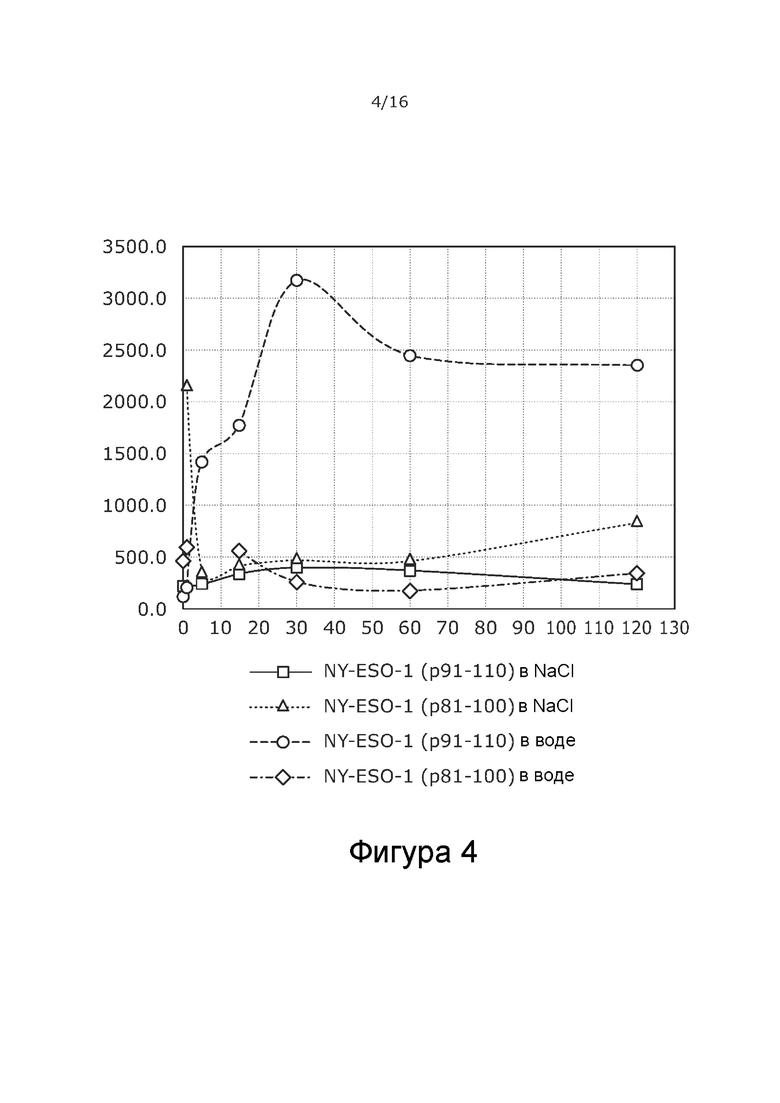
На Фиг. 4 показан средний гидродинамический диаметр обоих комплексов вирус-пептид NY-ESO-1 в воде и физиологической соли с различными концентрациями пептида.
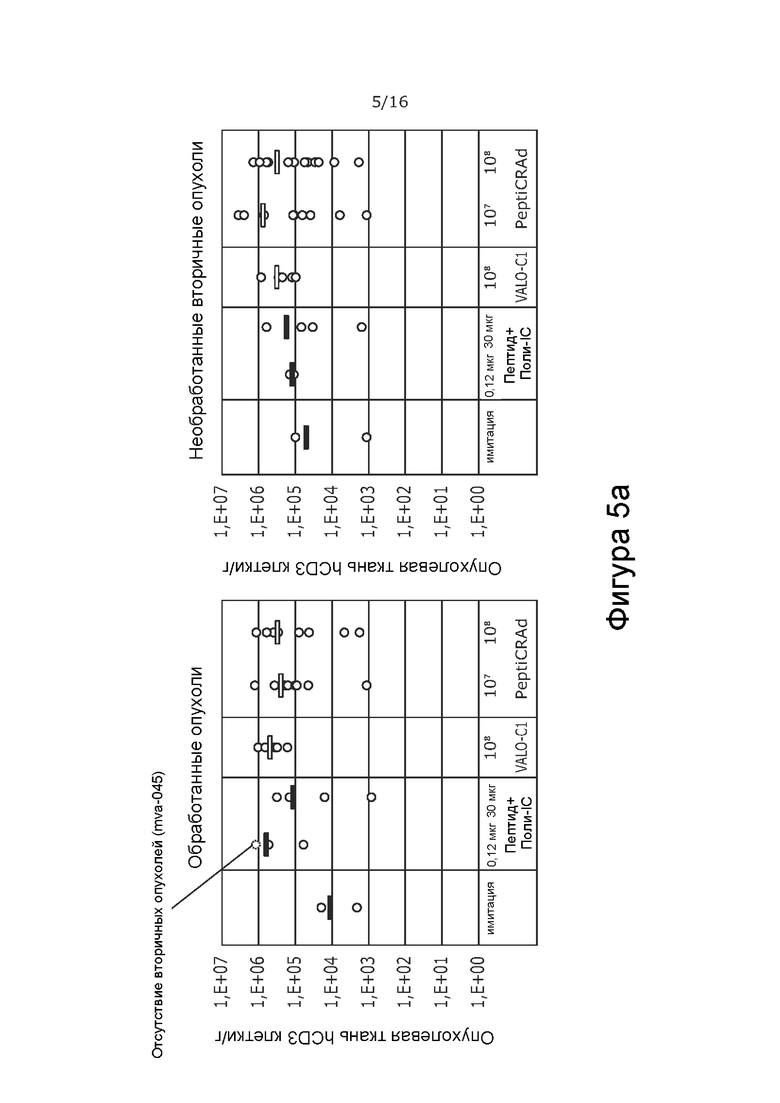
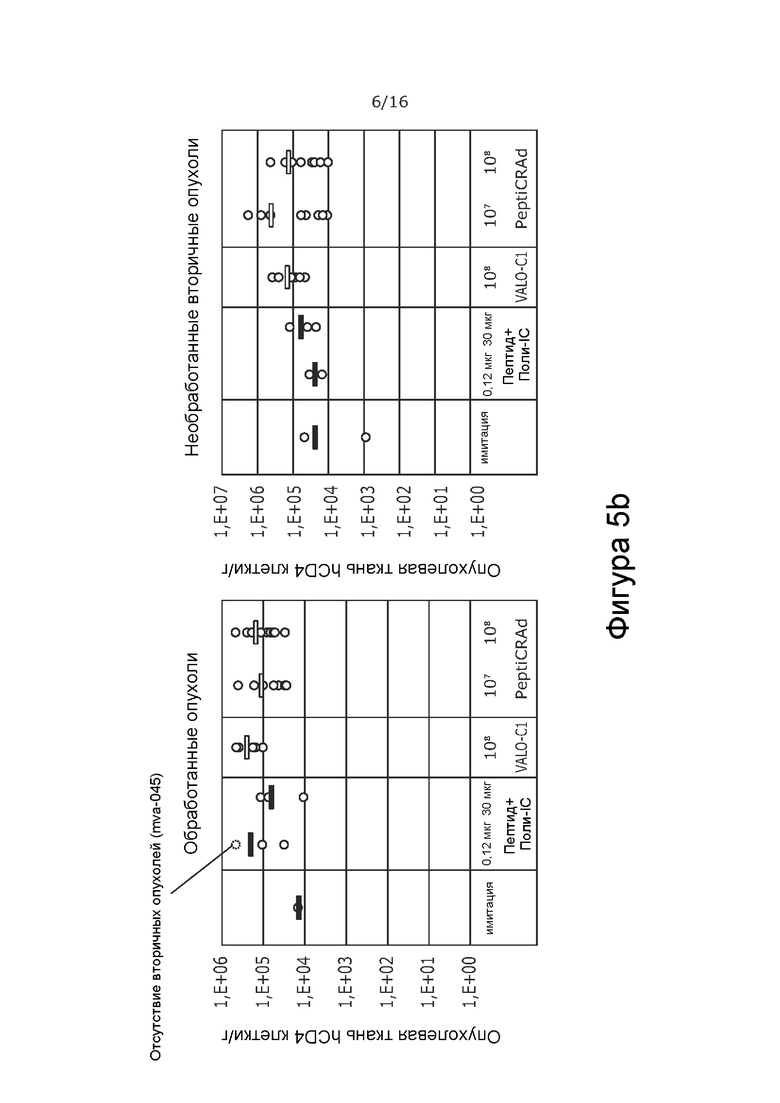
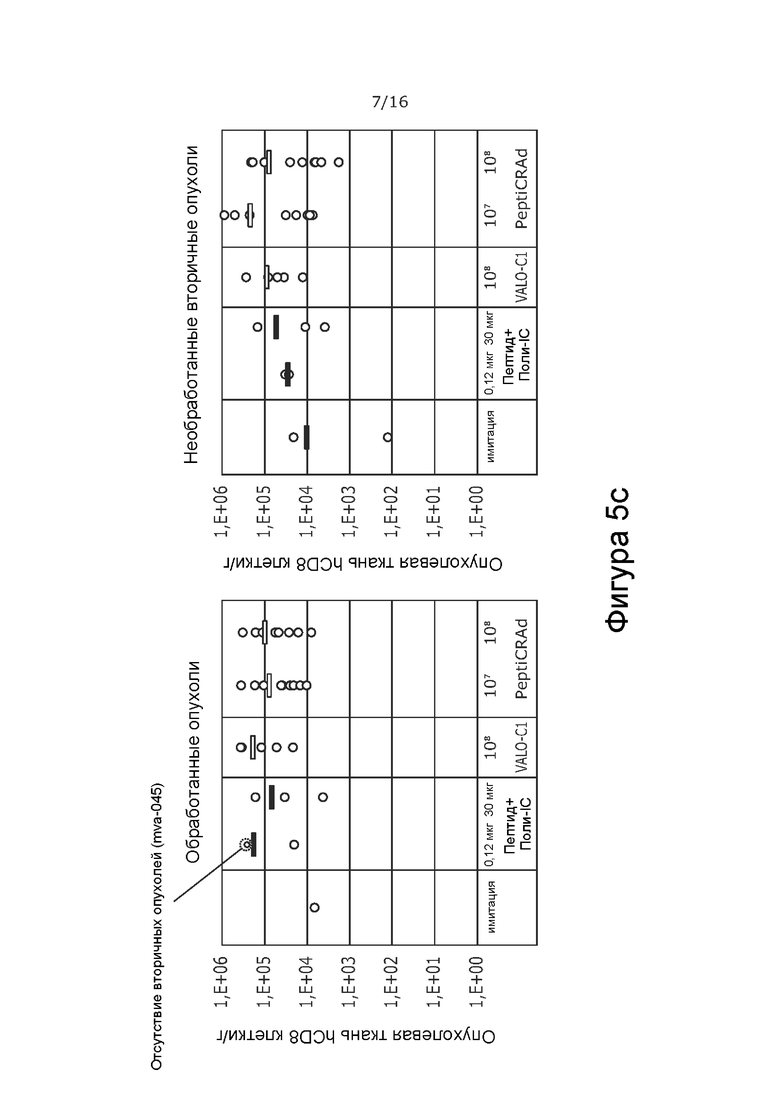
На Фиг. 5 показана частота встречаемости Т-клеток в обработанной и контралатеральной, необработанной опухоли при обработке модифицированным вирусом в соответствии с настоящим изобретением, содержащим капсид 5/3, содержащий OX40L- и CD40L-экспрессирующие вирусы (PeptiCRAd -1), покрытым NYESO-1-пептидом и MAGE-A3-пептидом, или тем же самым вирусом без покрытия NYESO-1 и MAGE-A3-пептидом (VALO-C1), или только пептидом. Количество CD3+ T-клеток (A), CD4+ T-клеток (B) и CD8+ T-клеток (C) показано как количество клеток на грамм опухолевой ткани в каждой группе обработки. Обработка привела к более высокой частоте встречаемости Т-клеток во всех группах по сравнению с имитацией. Наибольшее количество наблюдали в опухолях, обработанных VALO-C1 или PeptiCRAd.
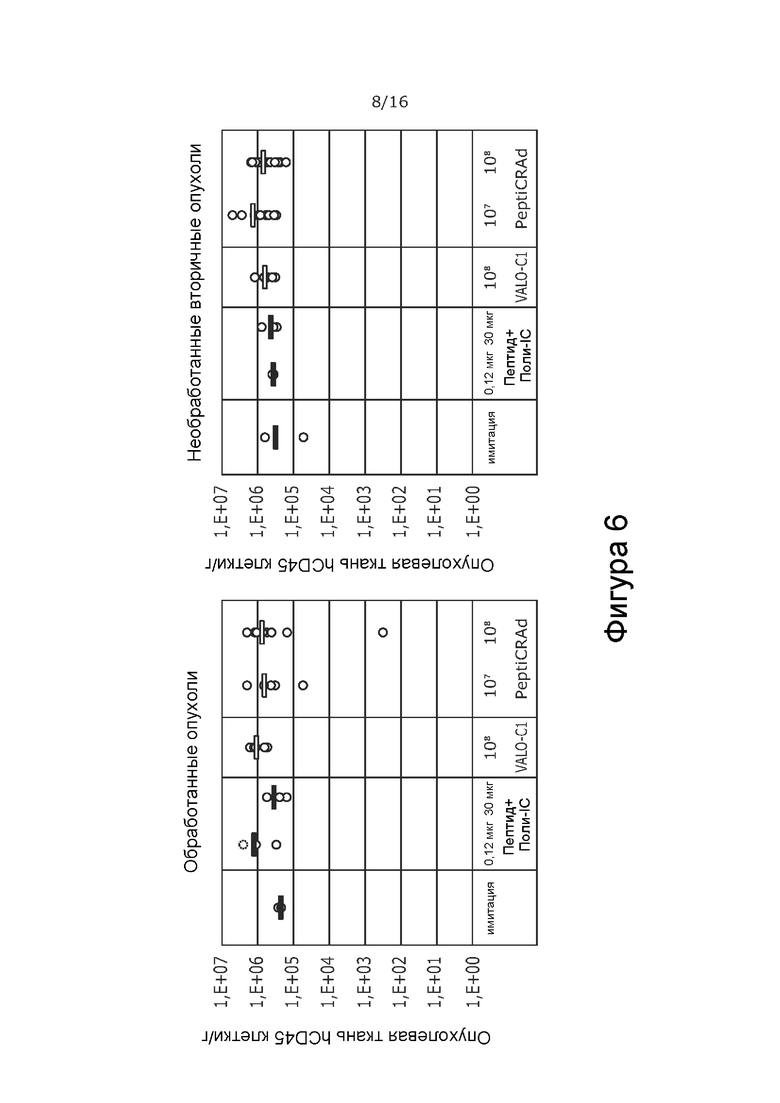
На Фиг. 6 показана частота встречаемости всех иммунных клеток (клетки CD45+) в обработанной и контрлатеральной, необработанной опухоли при обработке PeptiCRAd-1 или тем же вирусом без покрытия пептидами (VALO-C1) или только пептидом. Частота встречаемости была сходной во всех группах с несколько меньшим числом у животных, получавших имитацию.
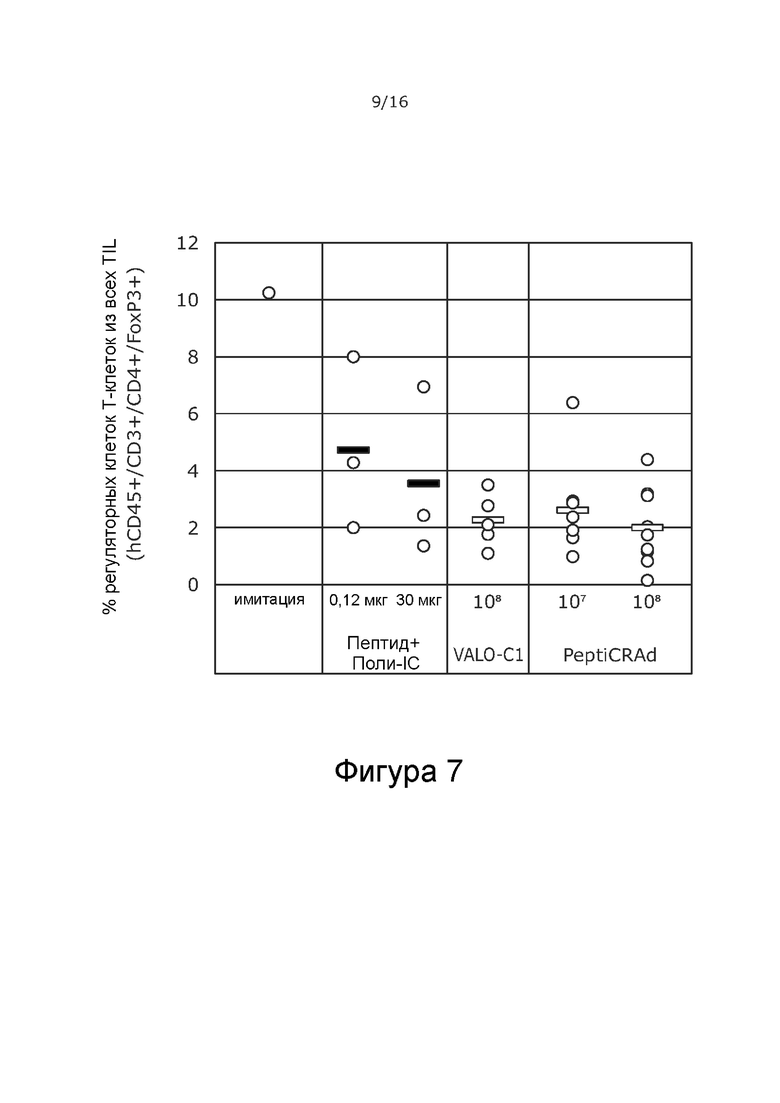
На Фиг. 7 показано, что обработка PeptiCRAd и VALO-C1 снижает процентное содержание регуляторных Т-клеток из всех TIL в обработанных опухолях.
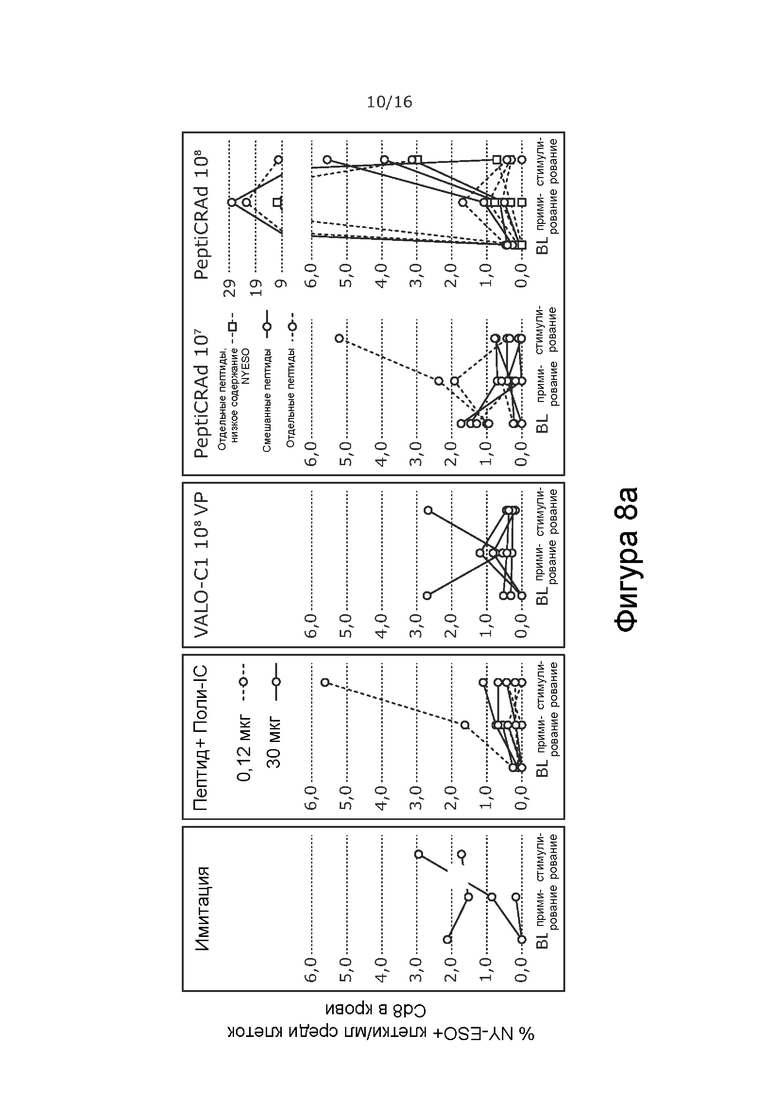
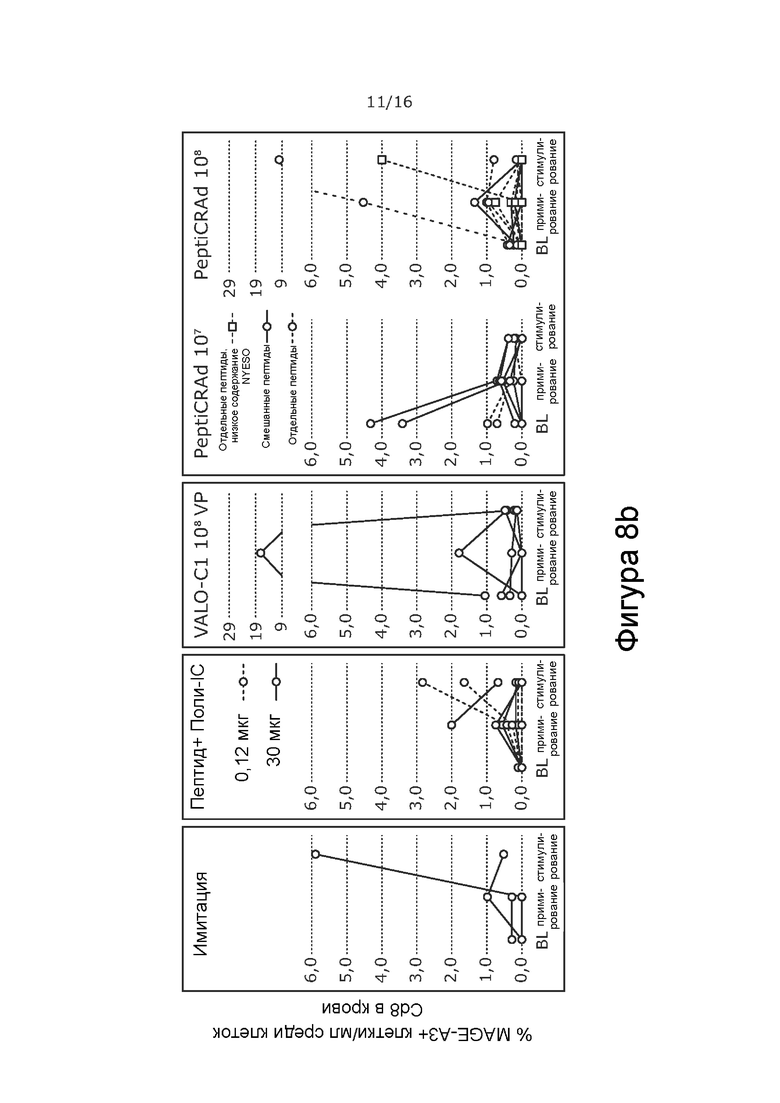
На Фиг. 8 показано процентное содержание NY-ESO-1- (A) и MAGE-A3 (B) -специфичных CD8+ Т-клеток в опухоли крови при обработке PeptiCRAd-1 или тем же вирусом без покрытия пептидами (VALO-C1) или только пептидом. Процент рассчитывали от всех CD8+ Т-клеток на 1 мл крови. Обработка PeptiCRAd привела к наивысшему проценту NY-ESO-I-специфичных CD8+ Т-клеток, а обработка как VALO-C1, так и PeptiCRAd привела к более высокому проценту MAGE-A3-специфичных клеток по сравнению с животными, получавших имитацию или пептид.
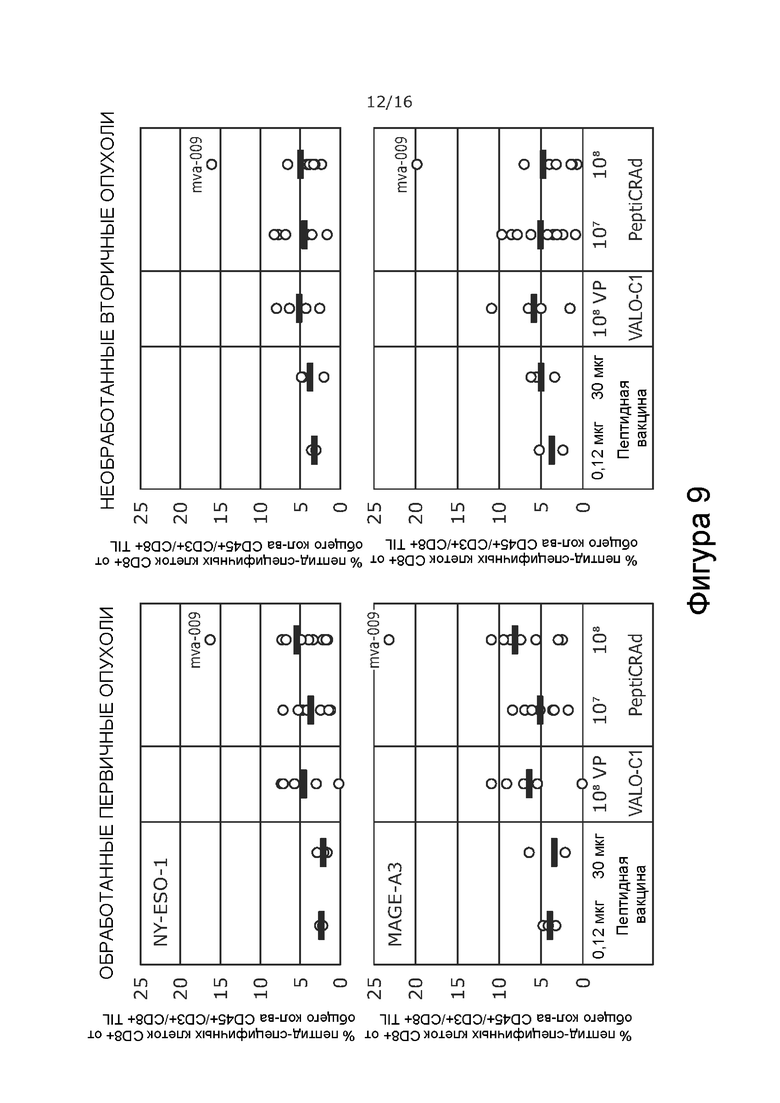
На Фиг. 9 показано процентное содержание NY-ESO-1 или MAGE-A3-специфичных CD8+ Т-клеток в обработанной или контрлатеральной, необработанной опухоли при обработке PeptiCRAd-1 или тем же вирусом без покрытия пептидами (VALO-C1) или только пептидом. Процент рассчитывали от всех CD8+ Т-клеток на грамм опухолевой ткани. VALO-C1 и PeptiCRAd одинаково эффективны в привлечении пептид-специфичных CD8+ TIL в опухолях.
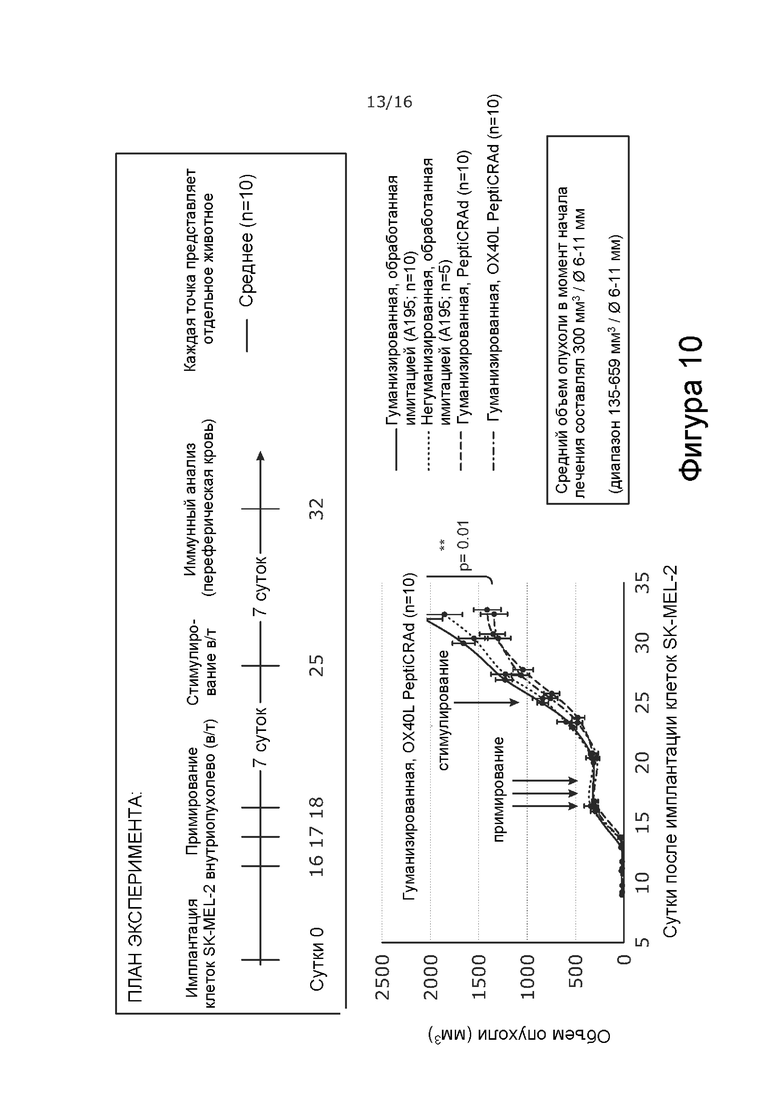
На Фиг. 10 показано, что PeptiCRAd, покрытый белками NY-ESO-1 и MAGE-A3, способен останавливать рост опухоли в модели меланомы гуманизированной мыши, даже если обработка начата в случае крупных, хорошо сформировавшихся опухолей. План эксперимента: 2x106 клеток SK-MEL-2 имплантировали подкожно (одна опухоль на животное) в бок мышей с иммунодефицитом NOD/Shi-scid/IL-2Rγnull в сутки 1. В сутки 13 внутривенно вводили 5x106 PBMC (мононуклеарные клетки периферической крови). В сутки 16 внутриопухолево вводили 5x104 плазмоцитоидных и миелоидных дендритных клеток. Внутриопухолевую обработку PeptiCRAd в дозе 1x109 ВЧ (вирусные частицы) проводили в сутки 16, 17, 18 (примирование) и в сутки 25 (стимулирование). Первую дозу PeptiCRad вводили сразу после инъекции DC. Наблюдали за ростом опухоли. Животных умерщвляли на 32 сутки. PeptiCRAd = Ad5/3-D24-OX40L-CD40L, онколитический аденовирус с делецией 24 п.н. в E1A, с химерным капсидом 5/3 и трансгенами CD40L и OX40L, экспрессируемыми из локуса 14.7K, покрытый пептидами NY-ESO-1 и MAGE-A3; OX40L PeptiCRAd = Ad5/3-D24-OX40L, онколитический аденовирус с делецией 24 п.н. в E1A, с химерным капсидом 5/3 и трансгеном OX40L, экспрессируемым из локуса 14.7K, покрытый пептидами NY-ESO-1 и MAGE-A3.
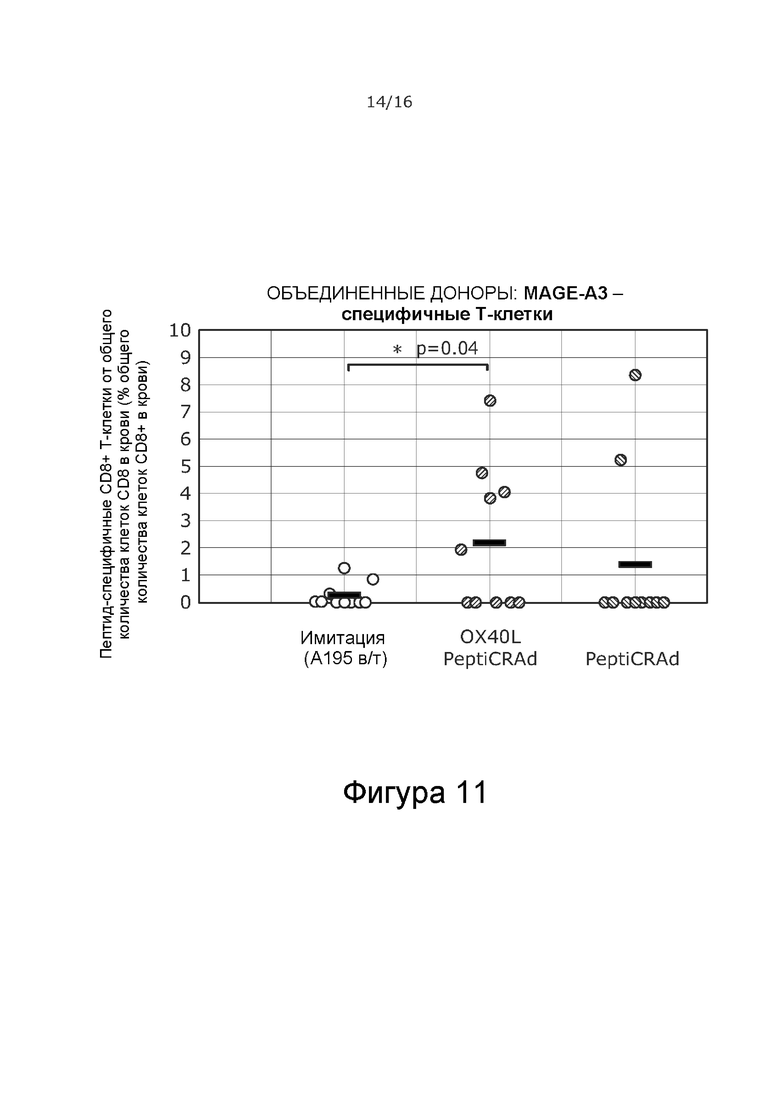
На Фиг. 11 показано, что OX40L-PeptiCRAd увеличивал количество MAGE-A3-специфичных CD8+ Т-клеток в периферической крови по сравнению с животными, получавшими имитацию. Два животных, получавших PeptiCRAd, также показали повышенное количество MAGE-A3-специфичных CD8+ Т-клеток в крови. Т-клетки против MAGE-A3 оценивали с помощью проточной цитометрии (пентамерный анализ) в конце ранее упомянутого исследования роста опухоли на 32 сутки.
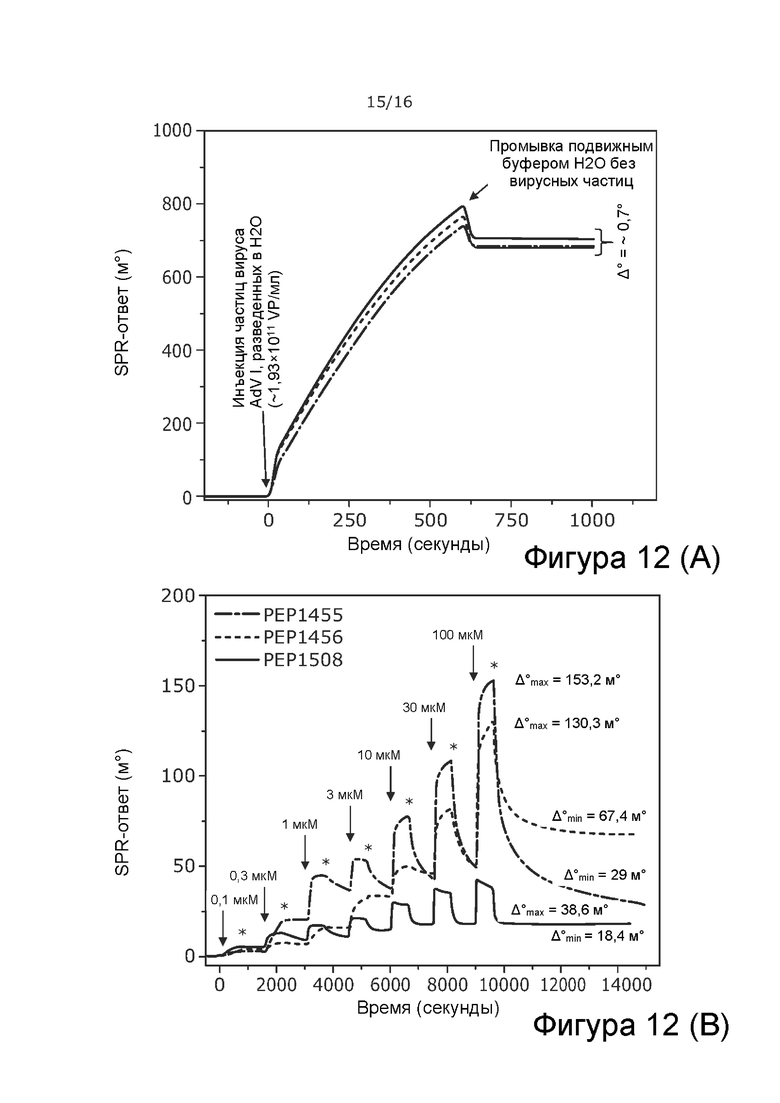
Фиг. 12 A) Три отдельных ответа SPR во время приготовления AdV, разведенного в стерильной воде и иммобилизации на датчике APTES с использованием стерильной воды в качестве подвижного буфера. С = ~1,93×1011 ВЧ/мл. B) SPR-ответы при 3 различных покрытиях пептидами: PEP1455 = KKKKKK-VFGIELMEVDPIGHLYIFAT (SEQ ID NO: 4); PEP1456 = KKKKKKKKK-VFGIELMEVDPIGHLYIFAT (SEQ ID NO: 7); PEP1508 = KKKKKK-YLAMPFATPMEAELARRSLA (SEQ ID NO: 5). Исходным растворам (5 мг/мл в стерильной воде) с возрастающими концентрациями давали возможность взаимодействовать с иммобилизованными частицами AdV. В качестве подвижного буфера использовали стерильную воду. Стрелки указывают моменты времени введения образца пептида с соответствующей концентрацией, а звездочки указывают момент времени начала промывки подвижным буфером. Значения Δmax и Δmin, отмеченные на Фиг., соответствуют SPR-ответам, используемым для расчета количества пептидов, взаимодействующих на вирусную частицу в каждой ситуации (пептид, присутствующий в 100 мкМ в растворе, Δmax, и отсутствие пептида в растворе,Δmin).
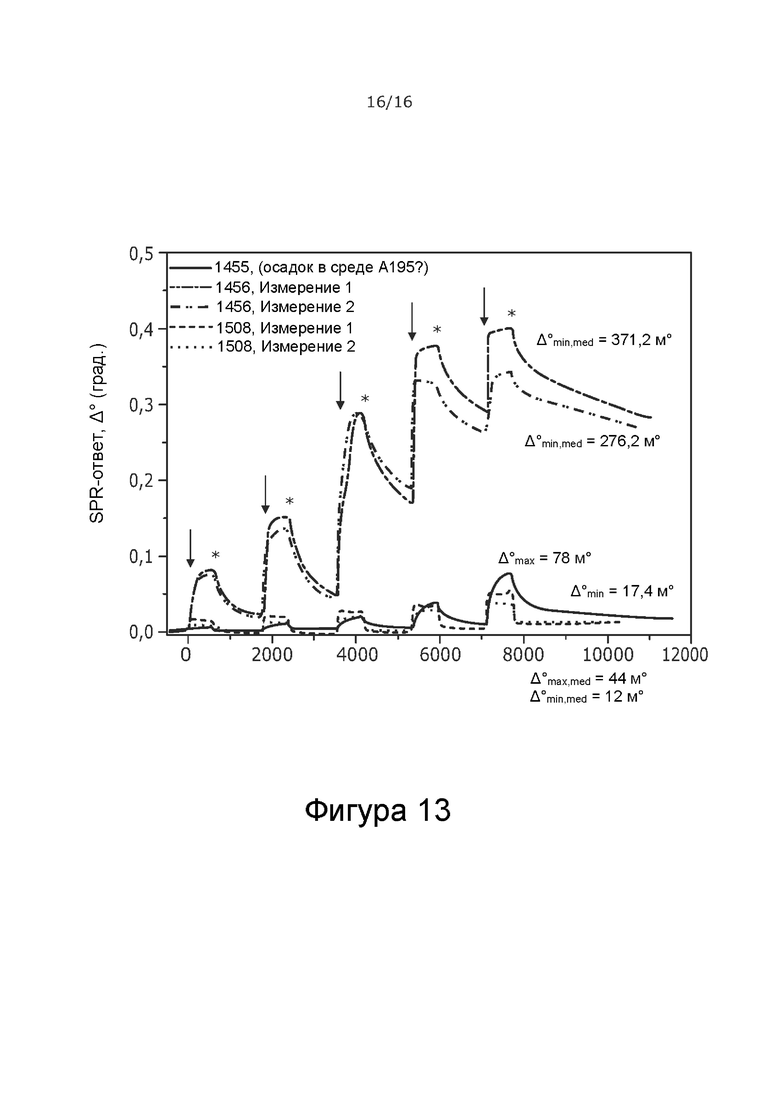
Фиг.13 Ответы SPR при увеличении концентраций образцов покрытий пептидами PEP1455, PEP1456 и PEP1508, разбавленных в буфере A195, взаимодействующих с иммобилизованными частицами AdV. Стрелки указывают момент времени введения образца пептида с соответствующей концентрацией, а звездочки указывают момент времени начала промывки подвижным буфером (среда A195). Значения Δmax и Δmin,, отмеченные на Фиг., соответствуют SPR-ответам, используемым для расчета количества взаимодействующих пептидов, на вирусную частицу в каждой ситуации (пептид, присутствующий в 100 мкМ в растворе, Δmax, и отсутствие пептида в растворе, Δmin). Индексы «min, med» и «max, med» относятся к среднему значению двух отдельных измерений, рассчитанному на основе соответствующих измерений.
Способы и материалы
Пептиды:
Пептиды выбирали с помощью противоиона ацетата, который представляет собой подходящую форму соли для пациентов, представляющих собой человека. Пептиды были произведены PepScan.
К специфичным пептидам, предназначенным для клинического использования в PeptiCRAd, относятся:
- белок MAGE-A3 (аминокислоты 161-180, последовательность KKKKKK(KKK)-VFGIELMEVDPIGHLYIFAT) пептид 20-мер с хвостом из 6 (SEQ ID NO: 4) или 9 (SEQ ID NO: 7) лизинов, примыкающим к аминоконцу;
- белок NY-ESO-1 (аминокислоты 91-110, последовательность KKKKKK(KKK)- YLAMPFATPMEAELARRSLA) пептид 20-мер с хвостом из 6 (SEQ ID NO: 5) или 9 (SEQ ID NO: 8) лизинов, примыкающим к аминоконцу;
- белок NY-ESO-1 (аминокислоты 81-100, последовательность KKKKKK(KKK)-RGPESRLLEFYLAMPFATPM) пептид 20-мер с хвостом из 6 (SEQ ID NO: 6) или 9 (SEQ ID NO: 9) лизинов, примыкающим к аминоконцу.
Аденовирус, используемый в получении PeptiCRAd:
Аденовирус delta 24, вирус серотипа 5 с модифицированной областью выступа из аденовируса серотипа 3 (AdV 5/3). Данный модельный вирус не является в точности тем вирусом, который предназначен для клинического использования, но он имеет идентичный вирусный капсид. Разница между модельным вирусом и клиническим вирусом заключается в том, что клинический вирус в идеальном варианте будет включать генетические вставки, кодирующие CD40L и OX40L, которые не влияют на капсид вируса и характеристики, изучаемые в настоящей заявке.
Образование комплекса PeptiCRAd:
Отдельные пептиды разбавляли водой или 0,5% физиологическим раствором для достижения исходной концентрации пептида 5 мкг/мкл. Исходный вирус в буфере A195 (1,45E+ 12 ВЧ/мл) разбавляли водой до достижения концентрации 1E+9 ВЧ/мкл. Комплекс вирус-пептид получали путем смешивания 1 или 3 мкл разведенного вируса (соответствующих 1E+9 или 3E+9 ВЧ) с варьируемым объемом пептида(ов) для достижения целевого соотношения пептида к вирусу.
(PeptiCRAd, отдельные пептиды); две отдельные аликвоты вирусов по отдельности покрывали отдельными пептидами. Эти два отдельных PeptiCRAd, покрытые одним пептидом (комплекс NY-ESO-1 и комплекс MAGE-A3), затем объединяют непосредственно перед инъекцией в опухоль. Независимо покрытые вирусы также могут быть введены в виде отдельных инъекций. В альтернативном варианте пептиды NY-ESO-1 и MAGE-A3 сначала смешивают вместе, а затем эту смесь пептидов используют для образования комплекса PeptiCRAd.
Проведение измерений с помощью Zetasizer для сложных размеров и заряда:
Измерения с помощью Zetasizer проводили сразу после образования комплекса, через 15 минут после смешивания компонентов или после того, как комплекс оставляли при комнатной температуре (КТ) приблизительно в течение 1,5 часов. Комплексы вирус-пептид сначала разбавляли, добавляя к образцам 700 мкл воды, а затем переносили в измерительную кювету. Измеряли гидродинамический диаметр (нм) с последующим измерением зета-потенциала (мВ). Эти параметры измеряли трижды и регистрировали средние значения размера и зета-потенциала.
Иммунологическая активность пептидов PeptiCRAd-1:
CD8+ Т-клетки, полученные у пациентов с меланомой, с известной активностью NY-ESO-1 и MAGE-A3 Т-клеток, предварительно стимулировали немодифицированными пептидами (SEQ ID NO: 2 NY-ESO-1 91-110: YLAMPFATPMEAELARRSLA или SEQ ID NO: 1 MAGE-A3 161-180: VFGIELMEVDPIGHLYIFAT). Распознавание пептидов, удлиненных полилизином (SEQ ID NO: 5 NY-ESO-1 91-110: KKKKKKYLAMPFATPMEAELARRSLA или SEQ ID NO: 4 MAGE-A3 161-180: KKKKKKVFGIELMEVDPIGHLYIFAT), исследовали стандартным способом ELISPOT (метод иммуноферментных пятен) IFN-гамма. В экспериментах использовали следующий протокол ELISPOT: CD4+/CD8+ Т-клетки, очищенные на колонке для разделения клеток MACS® (Miltenyi Biotech, Lund, Sweden), предварительно сенсибилизировали активированными пептидами (10 мкг/мл) облученными аутологичными PBMC, лишенными CD4+ и CD8+ Т-клеток (25000 клеток/лунка). Предварительно сенсибилизированные CD4+/CD8+ Т-клетки проверяли на сутки 10-12 в анализе IFNγ ELISPOT на распознавание активированных пептидами (1 мкг/мл) аутологичных антиген-презентирующих клеток (В-клетки или DC, трансформированные EBV). После 16 часов инкубации (37 C°) количество цитокин-продуцирующих антиген-специфичных Т-клеток оценивали с помощью AID EliSpot Reader Classic ELR 07 (Autoimmun Diagnostika GmbH, Strassberg, Germany).
Иммунизация in vivo
Мышей с иммунодефицитом NOD/Shi-scid/IL-2Rγnull гуманизировали с использованием гемопоэтических стволовых клеток (CD34+, HLA-B35+), выделенных из пуповинной крови человека. Опухоли меланомы человека A375 имплантировали подкожно (2 × 106 клеток на 100 мкл), и животных произвольно распределяли в группы на основании степени гуманизации и размера опухоли. Животных обрабатывали либо PeptiCRAd-1, либо тем же вирусом без покрытия пептидами (VALO-C1) (доза вируса 1 × 108 для обеих групп; субоптимальную дозу 1 × 107 также проверяли для PeptiCRAd). Пептидные вакцины (0,12 или 30 мкг) вводили внутрикожно с Poly-IC в качестве адъюванта.
Обработку начинали через 25 дней после рандомизации (D0) болюсной дозой циклофосфамида (1 мг/мышь внутривенно (в/в)). Обработку проводили внутриопухолево (имитация, вирус и PeptiCRAd) или внутрикожно (пептидный контроль) на сутки 1, 2, 3 и 12. Вторичные опухоли имплантировали в контрлатеральный бок через двое суток после третьей обработки (сутки 5). Обработку вторичных опухолей не проводили.
Мононуклеарные клетки периферической крови (PBMC) и инфильтрирующие опухоль лимфоциты CD8+ (TIL) анализировали на NNY-ESO-1 YLAMPFATPMEAELARRSLA SEQ ID NO: 2 и MAGE-A3 VFGIELMEVDPIGHLYIFAT SEQ ID NO: 1 специфичные CD8+ T-клетки методом проточной цитометрии с помощью декстамерного анализа. Оценивали различные субпопуляции иммунных клеток среди PBMC и TIL. Анализ методом проточной цитометрии выполняли на проточном цитометре Attune NxT (Life Technologies).
Иммунизация модели мышей с PBMC
35 мышам с иммунодефицитом (NCG) NOD-Prkdcem26Cd52/IL-2Rγ em26Cd22 /NjuCrl в возрасте восьми недель прививали 2.106 опухолевых клеток SKMEL-2(HLA-B35+) в правый бок (сутки 0). На сутки 13 внутривенно вводили 5x106 HLA-B35+ мононуклеарных клеток периферической крови человека (PBMC) от двух разных доноров. Внутриопухолевую обработку NYESO-1 (SEQ ID NO: 2) и MAGE-A3 (SEQ ID NO: 1) в комплексе с капсидом 5/3 и содержащим OX40L-экспрессирующий вирус (“OX40L PeptiCRAd”) или NYESO-1 (SEQ ID NO: 2) и MAGE-A3 (SEQ ID NO: 1) - в комплексе с капсидом 5/3, содержащим OX40L- и CD40L-экспрессирующий вирус (“PeptiCRAd”) - начинали в сутки 16 с дозой вируса 1 x 109 ВЧ в комплексе с каждым пептидом. Одновременно с первой обработкой PeptiCRAd внутриопухолево вводили 50000 аутологичных плазмоцитоидных и миелоидных дендритных клеток. В сутки 17, 18 (примирование с первичной обработкой) и 25 (стимулирование) опухоли обрабатывали внутриопухолевыми инъекциями PeptiCRAd без добавления дендритных клеток. Схема обработки представлена на Фиг. 10. Наблюдали за ростом опухоли. Животных умерщвляли на 32 сутки. OX40L-PeptiCRAd и PeptiCRad содержат делецию 24 п.н. в E1A, удаленную область gp19k/7.1K, трансген OX40L человека, экспрессируемый из локуса 14.7K, и химерную фибру 5/3.
Комплексообразование вирус-пептид
Иммобилизация вирусных частиц и взаимодействия между пептидами в покрытии и вирусными частицами, иммобилизованными на сенсорных поверхностях силикагеля, функционализированных APTES, измеряли с помощью прибора многопараметрического поверхностного плазмонного резонанса (SPR), оборудованного 2-канальным проточным каналом полидиметилсилоксана (PDMS) и устройством автосэмплера для 96-луночного планшета (MP-SPR 220A). Пептиды в покрытии были следующими: PEP1455 = KKKKKK-VFGIELMEVDPIGHLYIFAT (SEQ ID NO: 4); PEP1456 = KKKKKKKKK-VFGIELMEVDPIGHLYIFAT (SEQ ID NO: 7); PEP1508 = KKKKKK-YLAMPFATPMEAELARRSLA (SEQ ID NO: 5). Температура, скорость потока и длина волны лазера, использованные для всех измерений SPR, составляли +°20 C, 20 мкл/мин и 680 нм соответственно.
Для измерений в воде аденовирус с химерной фибриллой 5/3 (~ 1,93×1011 ВЧ/мл), разведенный в стерильной воде, иммобилизовали в течение 12 минут на датчике APTES, и для каждого образца проводили измерения в трех экземплярах. В качестве подвижного буфера также использовали стерильную воду. Для измерений в буфере A195 аденовирус сначала разводили в PBS и иммобилизовали в течение 12 минут на датчике APTES. После иммобилизации PBS заменяли на A195 в качестве подвижного буфера, и измерения выполняли в трех повторениях.
Для измерений в стерильной воде исходные растворы пептида готовили растворением 5 мг пептида в 1 мл воды (5 мг/мл), что приводило к соответствующим концентрациям 1,65 мМ для 6K-MAGE-A3 (PEP1455), 1,46 мМ для 9K- MAGE-A3 (PEP1456) и 1,66 мМ для 6K-NY-ESO (PEP1508). Образцы пептидов вводили последовательно в возрастающих концентрациях в течение 6 минут с последующим 8-минутным периодом промывания подвижным буфером между каждой концентрацией образца. Используемые концентрации пептидов составляли 0,1, 0,3, 1, 3, 10, 30 и 100 мкМ.
Для измерений в буфере A195 исходные растворы пептидов получали растворением 340 мкг соответствующих пептидов в среде A195 с образованием 100 мкМ растворов образцов, которые затем использовали в качестве исходных растворов для приготовления растворов образцов пептидов с более низкими концентрациями. Образцы пептидов вводили последовательно в возрастающих концентрациях в течение 10 минут с последующим 15-минутным периодом промывания подвижным буфером между каждой концентрацией образца. Используемые концентрации пептидов составляли 3,125, 6,25, 12,5, 25, 50 и 100 мкМ.
Расчеты для оценки количества пептидов, связанных на одну вирусную частицу, были основаны на использовании геометрических расчетов предполагаемого количества вирусных частиц, иммобилизованных на поверхности сенсора SPR, и максимального количества адсорбированных пептидов, определенного с помощью измерений SPR.
Результаты и обсуждение
Влияние суммарного заряда пептида на комплексообразование
Пептид MAGE-A3 имеет суммарный заряд -3, 9 без лизинового хвоста. Три кислые аминокислоты, обеспечивающие отрицательные заряды, расположены близко к аминоконцу (аминокислоты 5, 8 и 10). Исследовали, нужно ли компенсировать отрицательный суммарный заряд пептида более длинным положительным лизиновым хвостом (9 вместо 6 лизинов) для достижения надлежащего связывания пептидов на поверхности вируса. Влияние длины лизинового хвоста на образование комплекса и стабильность комплекса оценивали путем определения зета-потенциала и среднего размера комплекса покрытых пептидом вирусных образцов, содержащих 1E+9 вирусных частиц в комплексе с 20 или 40 мкг любого пептида. Измерения с помощью Zetasizer проводили сразу после образования комплекса, через 15 минут после смешивания компонентов и после того, как комплекс оставляли при комнатной температуре (КТ) приблизительно в течение 1,5 часов. Результаты представлены в Таблице 1.
Результаты размеров и зета-потенциалов показывают, что комплексы с пептидом MAGE-A3 образуются независимо от длины лизинового хвоста, что позволяет предположить, что отрицательно заряженные аминокислоты по меньшей мере в конкретном положении пептидной цепи, не препятствуют образованию комплекса. Более длинный лизиновый хвост давал комплексы приблизительно на 10% большего размера и немного более низкий зета-потенциал, чем соответствующий пептид с более коротким лизиновым хвостом. Исходя из размера комплексов свежеприготовленных образцов и образцов после выдерживания в течение 1,5 часов при КТ, комплексы с любым лизиновым хвостом были стабильными. В свежеприготовленных комплексах количество пептида, используемого при образовании комплекса, не влияло на размер, но в образцах, хранящихся в течение 1,5 ч, результаты несколько отличаются. В заключение, было решено использовать пептиды с хвостом из шести лизинов в последующих исследованиях с помощью Zetasizer.
Влияние соотношения пептида 6K-MAGE-A3 к вирусу на размер и зета-потенциал комплексов
Готовили пептиды к вирусным частицам, в соотношении, составляющем 15, 30, 45, и 60 мкг пептида на вирусные частицы 3E + 9, для оценки влияния соотношения на образование и стабильность комплекса (свежеприготовленные комплексы против комплексов, выдержанных при КТ в течение 2 часов) на основе определения размеров и зета-потенциала комплексов.
Результаты в Таблице 2 показывают тенденцию к тому, что чем ниже соотношение пептида к вирусу в пределах тестируемого диапазона, тем меньше размер комплекса, предполагая, что можно определить оптимальное соотношение, которое приводит к получению комплексов пептид-вирус, состоящих из одного вируса, или состоящих из небольших агрегатов, образованных парой вирусов. Комплексы также были достаточно стабильными по меньшей мере в течение двух часов с зета-потенциалом около +30 мВ.
Была предпринята попытка определить оптимальное соотношение пептида к вирусу с пептидом 6K-MAGE-A3, как указано в Таблице 3. По результатам размера комплекса предполагают, что всего 1-5 мкг пептида 6K-MAGE-A3 на 3E+9 вирусных частиц, что соответствует диапазону молярного соотношения от 6,6E+4 до 3,3E+5, приводят к получению стабильного комплекса. Но при более низком соотношении комплексы имеют тенденцию к агрегированию.
Влияние покрытия вируса пептидом 6K-MAGE-A3 на инфекционность
Способность вируса в комплексе инфицировать опухолевые клетки представляет собой один из важных аспектов механизма действия PeptiCRAd. Вот почему влияние покрытия пептидами на инфекционность вируса оценивали путем приготовления комплексов с различным соотношением пептидов к вирусу и определения инфекционности комплекса на клетках A549 на основе иммуноцитохимического анализа (ICC). Результаты в Таблице 4 показывают, что при использовании соотношения пептида и вирусу, равного приблизительно 1 мкг, сохранялось 62% инфекционности по сравнению с образцом только вируса, что позволяет предположить, что покрытие вируса не оказывает значительного влияния на инфекционность.
Стабильность 6K-MAGE-A3-PeptiCRAd при различных температурах
Для оценки стабильности 6K-MAGE-A3-PeptiCRAd при различных температурах, комплексы получали смешиванием 15 или 30 мкг пептида 6K-MAGE-A3 на 3E+9 ВЧ и посредством хранения образцов в различных условиях. Стабильность тестировали с помощью Zetasizer из образцов, хранившихся при КТ, +5, -20, -80 °C в течение 18-20 часов. Результаты представлены в Таблице 5. Средний гидродинамический диаметр оставался довольно стабильным во время хранения при различных температурах в диапазоне от 165 нм до 276 нм, что позволяет предположить, что никакой значительной агрегации не происходило ни при какой испытанной температуре. Зета-потенциал был выше уровня +30 мВ в каждом образце, что свидетельствует о хорошей стабильности. Наименьший размер частиц был достигнут после хранения при -20 °C, но также хранение при +4 °C казалось благоприятным для предотвращения агрегации.
Влияние соотношения пептида 6K-NY-ESO-1 (p81-100) к вирусу на размер и зета-потенциал комплексов, полученных в воде
Другой клинически интересный пептид, NY-ESO-1-пептид, изучали с точки зрения характеристик комплексообразования, размера и зета-потенциала, определенных с помощью измерений Zetasizer. Результаты представлены в Таблице 6а. Что касается размера комплекса, этот конкретный пептид вероятно следует синусоидальной кривой, начиная с меньшего размера комплекса при небольшом соотношении пептида к вирусу после увеличения размера комплекса по мере того, как соотношение становится больше, достигая локального максимума при 1 мкг пептида, после чего размер комплекса начинает уменьшаться до 60 мкг, где размер комплекса достигает минимум 180 нм. Что касается стабильности комплекса, результаты зета-потенциала позволяют предположить, что более высокое соотношение пептида к вирусу является благоприятным по сравнению с 6K-MAGE-A3. Пептид NY-ESO-1 имеет нативный заряд -1, имея две отрицательные и две положительные аминокислоты, близкие к аминоконцу (кислые в положениях 4 и 9 и основная в положениях 1 и 6 пептидной последовательности). Остальная часть последовательности NY-ESO-1 является достаточно гидрофобной, как и в случае с пептидом MAGE-A3.
Влияние соотношения пептида 6K-NY-ESO-1 (p91-110) к вирусу на размер и зета-потенциал комплексов, полученных в воде
Пептид 6K-NY-ESO-1 (p91-110) также изучали с точки зрения характеристик образования комплекса, размера и зета-потенциала, определенных с помощью измерений Zetasizer. Результаты представлены в Таблице 6b. Что касается размера комплекса, диапазон пептидов от 0,5 до 1 мкг приводит только к умеренно агрегированному комплексу, что позволяет предположить, что оптимальное соотношение с этим пептидом находится где-то между 1 и 5 мкг на 3E+9 ВЧ. Соответствующие значения зета-потенциала остаются близкими к нулю, что позволяет предположить, что стабильность комплекса может быть неоптимальной из-за тенденции к агрегации.
Пептид NY-ESO-1 (p91-110) имеет нативный заряд -1 и две отрицательные и две положительные аминокислоты, расположенные близко к карбоксиконцу (кислые в положениях 11 и 13 и основные в положениях 16 и 17 указанной пептидной последовательности). Остальная часть последовательности NY-ESO-1 довольно гидрофобна. Расположение положительных зарядов рядом с С-концом может также обеспечивать связывание С-конца с поверхностью вируса.
PeptiCRAd, покрытый обоими пептидами 6K-MAGE-A3- и 6K-NY-ESO-1
Оба клинически интересных пептида, NY-ESO-1- и MAGE-A3-пептиды, изучали в виде смесей с точки зрения признаков комплексообразования на основании измерений с помощью Zetasizer. Комплексы готовили как в воде, так и в физиологическом растворе NaCl (0,9%). Результаты представлены в Таблицах 7 а и b. Средний размер комплекса, полученного смешиванием равных количеств каждого пептида с вирусом, был больше, чем средний размер комплекса, приготовленного только из пептида 6K-MAGE-A3, и меньше, чем размер комплекса, приготовленного только из пептида 6K-NY-ESO-1 (p81-100). Средний размер менее 300 нм позволяет предположить, что во время образования комплекса происходила только умеренная агрегация. Указанный комплекс может быть получен как в воде, так и в 0,9% NaCl с сопоставимыми размерами комплексов.
Средний размер комплекса, полученного смешиванием равных количеств пептидов 6K-MAGE-A3 и 6K-NY-ESO-1 (p91-110) с вирусом, был больше, чем средний размер комплекса, приготовленного из пептидов 6K-MAGE-A3 и 6K-NY-ESO-1 (p81-100). В воде разница в средних размерах комплексов была примерно 1,8 раза, но уменьшилась до 1,3 раза в физиологическом NaCl. Средний размер 400-500 нм предполагает, что во время образования комплекса происходила только умеренная агрегация.
Влияние соотношения пептида NY-ESO-1 к вирусу на размер и зета-потенциал комплексов, полученных в физиологическом растворе NaCl.
Поскольку результаты определения размера комплексов, образованных с пептидом NY-ESO-1 (p81-100) в воде, с помощью Zetasizer позволяют предположить синусоидальное поведение в зависимости от концентрации пептида, мы изучили, может ли стадия образования комплекса в физиологическом солевом растворе предотвращать возможные неспецифичные нарушения. Результаты представлены в Таблице 8a. Результаты показывают, что размер комплекса как функция увеличения концентрации пептида следовал совершенно аналогичной тенденции, как и комплекс, полученный в воде. Размер комплекса был больше, но в любом случае вполне соответствовал соотношению пептида к вирусу в диапазоне от 5 мкг до 60 мкг. Результаты показывают, что оптимальное соотношение пептида к вирусу может быть обнаружено в диапазоне от 1 до 5 мкг на 3E+9 ВЧ.
Результаты определения среднего размера комплексов, образованных с пептидом NY-ESO-1 (p91-110) в воде, с помощью Zetasizer позволяют предположить об увеличении агрегации в случаях, когда соотношения пептида к вирусу выше 1 мкг, в результате чего размер комплекса превышает 3000 нм. Изучали, может ли стадия образования комплекса в физиологическом солевом растворе предотвратить агрегацию. Результаты представлены в Таблице 8 b и показывают, что размер комплекса как функция увеличения концентрации пептида был довольно постоянным в диапазоне 200-400 нм. Таким образом, физиологический NaCl в жидкой фазе во время образования комплекса может довольно резко предотвратить агрегацию, как можно видеть на Фиг. 1, где представлены данные как комплексов пептид-вирус NY-ESO-1 в воде, так и в физиологической соли. При соотношениях соответствующего пептида и вируса 1-5 мкг на 3E+9 ВЧ размер комплекса оставался в пределах 217-245 нм.
Иммунологическая активность пептидов PeptiCRAd-1
Удлиненные полилизином пептиды NY-ESO-1 и MAGE-A3 запускают продукцию IFN-гамма в антиген-специфичных CD8+ Т-клетках, полученных от больного раком, при стимуляции in vitro так же эффективно, как и немодифицированные пептиды (Фиг. 3), что указывает на то, что модификация полилизином не влияет на функцию.
PeptiCRAd вызывает специфичный к пептиду иммунный ответ в модели гуманизированной мыши
Все активные виды обработки (только пептид, вирус без покрытия пептидами [VALO-C1] и вирус с покрытием пептидами NY-ESO-1 и MAGE-A3 [PeptiCRAd]) увеличивали количество иммунных клеток в первичных опухолях по сравнению с животными, получавшими имитацию. Животные, обработанные как VALO-C1, так и PeptiCRAd-1, показали больше Т-клеток (CD3, CD4, CD8) в первичных опухолях по сравнению с животными, получавшими пептидную вакцину или имитацию, после обработки, в то время как общее количество инфильтрирующих иммунных клеток (CD45) было одинаковым во всех группах (Фиг. 5 и 6 соответственно).
Кроме того, количество Т-регуляторных клеток (CD3+/CD4+/FoxP3+) было меньше в первичных опухолях, обработанных VALO-C1 и PeptiCRAd-1, по сравнению с первичными опухолями животных, получавших пептидную вакцину или имитацию (Фиг. 7). Это позволяет предположить, что внутриопухолево введенный иммуногенный аденовирус (непокрытый вирус VALO-C1 или PeptiCRAd-1) модулирует микроокружение опухоли, снижая местную иммуносупрессию.
В отличие от животных, получавших VALO-C1, животные, получавшие PeptiCRAd-1, показали больше CD4+ и CD8+ Т-клеток в необработанных вторичных опухолях, чем в обработанных первичных опухолях, что позволяет предположить, что нацеливание на опухоль посредством покрытия пептидами вируса было критически важным для индукции эффекта в отдаленных необработанных опухолях (Таблица 9). Кроме того, у животных, получавших PeptiCRAd-1, было больше NYESO-специфичных CD8+ Т-клеток в крови после примирования (среднее значение равно 4,3% от общего количества CD8+ клеток) по сравнению с животными, получавшими OV (среднее значение равно 0,6%) или пептидную вакцину (среднее значение равно 0,6%) (Фиг. 8). Частота MAGE-специфичных CD8+ TIL у животных, получавших PeptiCRAd и VALOC1 - в обработанных и необработанных опухолях - была выше, чем частота MAGE-специфичных T-клеток CD8+ у животных, обработанных пептидом (Фиг.9). Интересно, что более высокую частоту MAGE-специфичных CD8+ T-клеток наблюдали в опухолях, обработанных VALOC1 и PeptiCRAd, по сравнению с NY-ESO-1-специфичными CD8+ T-клетками, в то время как в крови наблюдали обратное.
PeptiCRAd вызывает пептид-специфичный иммунный ответ в мышиной модели с PBMC
Обработка (лечение) PeptiCRAd, с входящими в комплекс NY-ESO-1 и MAGE-A3, приводила к остановке роста опухоли в модели гуманизированной меланомы мыши, даже когда обработка была начата для крупных, хорошо установленных опухолей (Фиг. 10). Мыши, получавшие OX40L-PeptiCRAd, показали значительно больше MAGE-A3-специфичных CD8+ Т-клеток среди всех CD8+ Т-клеток PBMC, чем мыши, получавшие имитацию, что указывает на то, что обработка с PeptiCRAd способно вызывать пептид-специфичный ответ у гуманизированных мышей (Фиг. 11).
Аденовирус с химерным капсидом 5/3 может образовывать комплекс с пептидами NY-ESO-I и MAGE-A3 в оптимальных условиях комплексообразования.
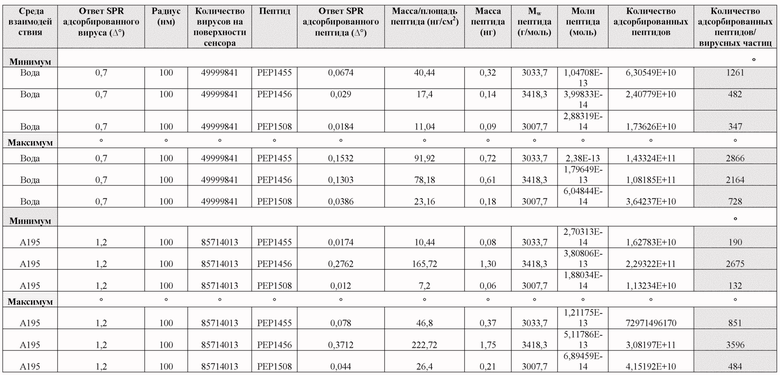
Взаимодействие между пептидами в покрытии и аденовирусными частицами (AdV) изучали с использованием метода поверхностного плазмонного резонанса (SPR). Основные цели: 1) определить динамику связывания пептидов NY-ESO-1 и MAGE-A3 с вирусами в различных средах (стерильная вода и среда A195, обычно используемая среда для вирусных препаратов), 2) оценить, какое количество молекул пептида может связываться с одной вирусной частицей в каждой среде и 3) оценить стабильность комплексов пептид-вирус. Пептиды в покрытии были следующими: PEP1455 = KKKKKK-VFGIELMEVDPIGHLYIFAT (SEQ ID NO: 4); PEP1456 = KKKKKKKKK-VFGIELMEVDPIGHLYIFAT (SEQ ID NO: 7); PEP1508 = KKKKKK-YLAMPFATPMEAELARRSLA (SEQ ID NO: 5).
SPR-ответ для частиц AdV в стерильной воде составляет приблизительно 0,7°, что соответствует 50% вирусов, покрывающих зону обнаружения (1,4° соответствует 100%-ному охвату) (Таблица 10), что позволяет дать оценку, что 5×107 вирусных частиц адсорбируется в зоне обнаружения. Когда частицы AdV обрабатывали в буфере A195, ответ SPR составлял приблизительно 1,2°, что соответствует охвату 86% поверхности обнаружения (Фиг. 12). Эти результаты позволяют приблизительно подсчитать, что 8,6 × 107 вирусных частиц адсорбированы в зоне обнаружения. Эти результаты могут указывать на то, что структура вирусов лучше сохраняется в буфере A195, чем в стерильной воде, или что буфер A195 способствует адсорбции вирусов на поверхности APTES.
Пептиды 1455 и 1508, по-видимому, лучше адсорбируются на иммобилизованных вирусных частицах в стерильной воде, чем в среде A195 (Фиг. 12B и 13). Обратное верно для пептида 1456, который адсорбируется на иммобилизованных вирусных частицах значительно лучше в буфере A195, чем в воде. Количество адсорбированных пептидов на вирус рассчитано в Таблице 10.
На основании результатов PEP1455 не был полностью растворим в буфере A195, и вероятно PEP1456 является более перспективной модификацией с точки зрения растворимости и эффективности покрытия, о чем судят по соотношению пептидов и вирусов в буфере A195. Результаты показывают, что комплексы пептида и вируса могут быть получены с использованием конструкции аденовируса с химерным капсидом 5/3 и 6/9K-пептидами в подходящей буферной среде. Комплексы достаточно стабильны без свободного пептидного окружения, как показывают результаты SPR. Кроме того, соотношение пептидов к вирусным частицам является фактором, специфичным для пептидной последовательности, и также зависит от наличия оптимальных условий для образования комплекса.
Результаты
С тремя протестированными пептидами, одним 6K-MAGE-A3 и двумя 6K-NY-ESO-1, размер комплекса вероятно зависит от соотношения пептида к вирусу и вероятно является специфичным свойством пептида, которое необходимо определять для каждого пептида и каждой комбинации пептидов и вируса. Агрегацию и возможные неспецифичные взаимодействия во время образования комплекса можно до некоторой степени предотвратить, приготовив комплекс в физиологическом солевом растворе вместо воды в случае 6K-NY-ESO-1 (p91-110), но в случае с 6K-NY-ESO-1 (p81-100) этого не наблюдали, что свидетельствует о специфичном поведении пептида.
Образование комплекса с пептидом 6K-MAGE-A3 не привело к значительному ухудшению инфекционности вируса, при этом оставалось 62% инфекционности при использовании соответствующего соотношения пептида к вирусу.
На основании результатов стабильности комплекса MAGE-A3, оптимальными условиями хранения комплекса вероятно являются +4 oC или -20 oC.
PeptiCRAd с покрытием(ями) пептидом(ами) превосходит непокрытый онколитический аденовирус (VALO-C1) или стандартную пептидную вакцинацию в отношении запуска системных нацеленных на опухоль ответов CD8+ Т-клеток и инфильтрации CD8+ TIL в необработанные отдаленные опухоли. Данные свидетельствуют о том, что PeptiCRAd улучшает специфичность стандартного онколитического вируса в отношении опухоли.
Таблица 1 Средний диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием вируса и пептида в пропорциях 1E+9 вирусных частиц с 20 или 40 мкг пептидов MAGE-A3 с хвостами из 6 или 9 лизинов.
Таблица 2 Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, измеренные сразу или через 2 часа после получения путем смешивания 3E+9 вирусных частиц с различными количествами пептида 6K-MAGE-A3, как указано в Таблице.
Таблица 3 Результаты среднего гидродинамического диаметра и зета-потенциала комплексов пептид-вирус, полученных путем смешивания 3E+9 вирусных частиц с различными количествами пептида 6K-MAGE-A3.
Таблица 4 Инфекционность комплексов PeptiCRAd, полученных с использованием различных соотношений пептида MAGE-A3 к вирусу (3E+9 ВЧ).
Таблица 5 Результаты среднего гидродинамического диаметра и зета-потенциала комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с 15 мкг или 30 мкг пептида 6K-MAGE-A3 после выдерживания при различных температурах в течение 18-20 часов.
Таблица 6 а Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с различными количествами пептида 6K-NY-ESO-1 (p81-100).
Таблица 6 b Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с различными количествами пептида 6K-NY-ESO-1 (p91-110).
Таблица 7 а Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с 1 мкг как пептида 6K-NY-ESO-1 (p81-100), так и пептида MAGE-A3 в воде или в физиологическом растворе NaCl (0,9%).
Таблица 7 b Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с 1 мкг как пептида 6K-NY-ESO-1 (p91-110), так и пептида 6K-MAGE-A3 в воде или физиологическом растворе NaCl (0,9%).
Таблица 8 а Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с различными количествами пептида 6K-NY-ESO-1 (p81-100) в физиологическом растворе NaCl (0,9%).
Таблица 8 b Средний гидродинамический диаметр и зета-потенциал комплексов пептид-вирус, полученных смешиванием 3E+9 вирусных частиц с различными количествами пептида 6K-NY-ESO-1 (p91 - 110) в физиологическом растворе NaCl (0,9%).
Таблица 9 Животные, получавшие PeptiCRAd, показали больше CD8+ и CD4+ Т-клеток в необработанных отдаленных опухолях, чем в обработанных первичных опухолях, что позволяет предположить, что обработка PeptiCRAd обеспечивает более эффективную презентацию антигена и последующее возвращение Т-клеток к удаленным опухолям по сравнению с OV. Результаты представлены как отношение количества лимфоцитов, инфильтрирующих опухоль в необработанных опухолях к обработанным опухолям ± SEM.
n = 5-6
n = 5
n = 17-20
Таблица 10 Результаты основаны на измерениях SPR для вируса и комплекса вирус-пептид.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Вало Терапьютикс Ой
<120> Терапия рака
<130> 4595P/WO
<150> GB1804468.5
<151> 2018-03-21
<150> GB1814866.8
<151> 2018-09-13
<160> 9
<170> PatentIn версии 3.5
<210> 1
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Val Phe Gly Ile Glu Leu Met Glu Val Asp Pro Ile Gly His Leu Tyr
1 5 10 15
Ile Phe Ala Thr
20
<210> 2
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Tyr Leu Ala Met Pro Phe Ala Thr Pro Met Glu Ala Glu Leu Ala Arg
1 5 10 15
Arg Ser Leu Ala
20
<210> 3
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Arg Gly Pro Glu Ser Arg Leu Leu Glu Phe Tyr Leu Ala Met Pro Phe
1 5 10 15
Ala Thr Pro Met
20
<210> 4
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Lys Lys Lys Lys Lys Lys Val Phe Gly Ile Glu Leu Met Glu Val Asp
1 5 10 15
Pro Ile Gly His Leu Tyr Ile Phe Ala Thr
20 25
<210> 5
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Lys Lys Lys Lys Lys Lys Tyr Leu Ala Met Pro Phe Ala Thr Pro Met
1 5 10 15
Glu Ala Glu Leu Ala Arg Arg Ser Leu Ala
20 25
<210> 6
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Lys Lys Lys Lys Lys Lys Arg Gly Pro Glu Ser Arg Leu Leu Glu Phe
1 5 10 15
Tyr Leu Ala Met Pro Phe Ala Thr Pro Met
20 25
<210> 7
<211> 29
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Lys Lys Lys Lys Lys Lys Lys Lys Lys Val Phe Gly Ile Glu Leu Met
1 5 10 15
Glu Val Asp Pro Ile Gly His Leu Tyr Ile Phe Ala Thr
20 25
<210> 8
<211> 29
<212> БЕЛОК
<213> Homo sapiens
<400> 8
Lys Lys Lys Lys Lys Lys Lys Lys Lys Tyr Leu Ala Met Pro Phe Ala
1 5 10 15
Thr Pro Met Glu Ala Glu Leu Ala Arg Arg Ser Leu Ala
20 25
<210> 9
<211> 29
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Lys Lys Lys Lys Lys Lys Lys Lys Lys Arg Gly Pro Glu Ser Arg Leu
1 5 10 15
Leu Glu Phe Tyr Leu Ala Met Pro Phe Ala Thr Pro Met
20 25
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ ОНКОЛИТИЧЕСКИЕ АДЕНОВИРУСЫ | 2019 |
|
RU2788638C2 |
| Вирусный вектор | 2020 |
|
RU2799418C1 |
| Покрытые онколитические аденовирусы для противораковых вакцин | 2015 |
|
RU2695375C2 |
| БИОИНФОРМАТИКА | 2021 |
|
RU2836005C1 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ЛЕГКИХ, ПРЕЖДЕ ВСЕГО, НЕМЕЛКОКЛЕТОЧНОГО РАКА ЛЕГКИХ (НМКРЛ) | 2008 |
|
RU2526510C2 |
| АНТИГЕН-СВЯЗЫВАЮЩИЙ БЕЛОК, РАСПОЗНАЮЩИЙ ПРОИСХОДЯЩИЙ ИЗ MAGE-A4 ПЕПТИД | 2018 |
|
RU2777074C2 |
| Полиэпитопная противоопухолевая вакцинная конструкция, содержащая эпитопы опухоль-ассоциированных антигенов, фармацевтическая композиция и ее применение для стимуляции специфического противоопухолевого иммунного ответа | 2016 |
|
RU2684235C2 |
| ВЕКТОР, КОЭКСПРЕССИРУЮЩИЙ МОЛЕКУЛЫ ДЛЯ ВАКЦИНАЦИИ И КОСТИМУЛИРУЮЩИЕ МОЛЕКУЛЫ | 2016 |
|
RU2714157C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2799573C2 |
| КОМБИНАЦИЯ МОДУЛЯТОРА ИММУННЫХ КОНТРОЛЬНЫХ ТОЧЕК И КОМПЛЕКСА, СОДЕРЖАЩЕГО ПРОНИКАЮЩИЙ В КЛЕТКУ ПЕПТИД, КАРГО-МОЛЕКУЛУ И ПЕПТИДНЫЙ АГОНИСТ TLR, ДЛЯ ПРИМЕНЕНИЯ В МЕДИЦИНЕ | 2017 |
|
RU2769314C1 |

Изобретение относится к биотехнологии. Описан модифицированный аденовирус для лечения рака, содержащий по меньшей мере один из следующих полипептидов, ковалентно или нековалентно присоединенный к вирусному капсиду, но не кодируемый генетически указанным аденовирусным вектором: VFGIELMEVDPIGHLYIFAT или YLAMPFATPMEAELARRSLA, и причем указанный аденовирус содержит трансген, кодирующий CD40L и/или OX40L, и дополнительно где соотношение пептида к вирусу находится в диапазоне от 1 до 5 мкг на 3E+9 вирусных частиц. Раскрыта фармацевтическая композиция для лечения рака, содержащая в эффективном количестве модифицированный указанный аденовирус в комбинации с подходящим носителем. Раскрыто применение указанного аденовируса в изготовлении лекарственного средства для лечения рака. Описан способ лечения рака у пациента, включающий введение пациенту эффективного количества указанного аденовируса или композиции. Изобретение расширяет арсенал средств для лечения рака. 4 н. и 17 з.п. ф-лы, 13 ил., 10 табл.
1. Модифицированный аденовирус для лечения рака, содержащий
по меньшей мере один из следующих полипептидов, ковалентно или нековалентно присоединенный к вирусному капсиду, но не кодируемый генетически указанным аденовирусным вектором:
1) VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 1] или
2) YLAMPFATPMEAELARRSLA [SEQ ID NO: 2], и причем указанный аденовирус содержит трансген, кодирующий CD40L и/или OX40L, и дополнительно где соотношение пептида к вирусу находится в диапазоне от 1 до 5 мкг на 3E+9 вирусных частиц.
2. Модифицированный аденовирус по п. 1, отличающийся тем, что указанный полипептид удлинен за счет полилизина с использованием по меньшей мере 3, 4, 5, 6, 7, 8 или 9 лизинов.
3. Модифицированный аденовирус по п. 2, отличающийся тем, что указанные лизины присоединены к аминоконцу указанного полипептида.
4. Модифицированный аденовирус по любому из предшествующих пунктов, отличающийся тем, что указанный полипептид представляет собой:
KKKKKK(KKK)-VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 7]; и/или
KKKKKK(KKK)-YLAMPFATPMEAELARRSLA [SEQ ID NO: 8];
в качестве альтернативы:
KKKKKK-VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 4]; и/или
KKKKKK-YLAMPFATPMEAELARRSLA [SEQ ID NO: 5].
5. Модифицированный аденовирус по любому из предшествующих пунктов, отличающийся тем, что указанный аденовирус представляет собой аденовирус человека.
6. Модифицированный аденовирус по любому из предшествующих пунктов, отличающийся тем, что указанный аденовирус содержит модификации в генах E1, и/или E3, и/или E4, и/или L3, включая вставку опухолеспецифичных промоторов, по меньшей мере одну делецию гена и вставку по меньшей мере одного трансгена.
7. Модифицированный аденовирус по любому из предшествующих пунктов, отличающийся тем, что указанный аденовирус относится к серотипу 5.
8. Модифицированный аденовирус по п. 7, отличающийся тем, что указанный аденовирус дополнительно модифицирован для включения химерной замены Ad5/3, то есть замены области выступа фибриллы аденовируса серотипа 5 на область выступа аденовируса серотипа 3.
9. Модифицированный аденовирус по любому из предшествующих пунктов, отличающийся тем, что указанный аденовирус дополнительно модифицирован для включения делеции гена E1A, при этом указанная делеция представляет собой делецию по меньшей мере тех нуклеотидов, которые кодируют аминокислоты 122-129.
10. Модифицированный аденовирус по любому из предшествующих пунктов, отличающийся тем, что указанный аденовирус дополнительно модифицирован для включения частичной или полной делеции гена 14.7К.
11. Модифицированный аденовирус по любому из пп. 1-10, отличающийся тем, что указанный трансген представляет собой трансген человека.
12. Модифицированный аденовирус по любому из пп. 1-11, отличающийся тем, что указанный OX40L расположен в области E3B, замещая делецию гена 14.7К.
13. Модифицированный аденовирус по любому из пп. 1-12, отличающийся тем, что указанный аденовирус содержит трансгены, кодирующие CD40L и OX40L, и указанная молекула CD40L вставлена непосредственно ниже OX40L с использованием сайта процессинга 2A.
14. Фармацевтическая композиция для лечения рака, содержащая в эффективном количестве модифицированный аденовирус по любому из пп. 1-13 в комбинации с подходящим носителем.
15. Фармацевтическая композиция по п. 14, отличающаяся тем, что указанная композиция приготовлена для внутриопухолевой, внутримышечной, внутриартериальной, внутривенной, внутриплевральной, внутрипузырной, внутрикожной, внутриполостной или перитонеальной инъекции или для перорального введения.
16. Аденовирус по любому из пп. 1-13 или фармацевтическая композиция по любому из пп. 14, 15 для применения при лечении рака.
17. Применение аденовируса по любому из пп. 1-13 в изготовлении лекарственного средства для лечения рака.
18. Способ лечения рака у пациента, включающий введение пациенту эффективного количества аденовируса по любому из пп. 1-13 или композиции по п. 14 или 15.
19. Способ лечения рака по п. 18, отличающийся тем, что аденовирус по любому из пп. 1-13 или композицию по п. 14 или 15 вводят с модулятором контрольной точки клетки.
20. Способ лечения рака по п. 19, отличающийся тем, что указанный модулятор контрольной точки представляет собой молекулу анти-PD1, молекулу анти-PD-L1 или молекулу анти-CTLA-4.
21. Аденовирус или применение по п. 16 или 17 или способ по пп. 18-20, отличающиеся тем, что указанный рак включает любой один или более из следующих видов рака: носоглоточный рак, синовиальный рак, печеночно-клеточный рак, рак почки, рак соединительных тканей, меланома, рак легкого, рак кишечника, рак толстой кишки, рак прямой кишки, колоректальный рак, рак мозга, рак горла, рак полости рта, рак печени, рак кости, рак поджелудочной железы, хориокарцинома, гастринома, феохромоцитома, пролактинома, Т-клеточный лейкоз/лимфома, нейрома, болезнь фон Гиппеля-Линдау, синдром Золлингера-Эллисона, рак надпочечника, рак ануса, рак желчного протока, рак мочевого пузыря, рак мочеточника, олигодендроглиома, нейробластома, менингиома, опухоль спинного мозга, рак кости, остеохондрома, хондросаркома, саркома Юинга, рак с неизвестной первичной локализацией, карциноид, карциноид желудочно-кишечного тракта, фибросаркома, рак молочной железы, болезнь Пэджета, рак шейки матки, рак пищевода, рак желчного пузыря, рак головы, рак глаза, рак шеи, рак почки, опухоль Вильмса, рак печени, саркома Капоши, рак предстательной железы, рак яичек, ходжкинская лимфома, неходжкинская лимфома, рак кожи, мезотелиома, множественная миелома, рак яичника, рак эндокринных клеток поджелудочной железы, глюкагонома, рак паращитовидной железы, рак пениса, рак гипофиза, саркома мягких тканей, ретинобластома, рак тонкого кишечника, рак желудка, рак вилочковой железы, рак щитовидной железы, трофобластический рак, пузырный занос, рак матки, рак эндометрия, рак влагалища, рак вульвы, нейрома слухового нерва, фунгоидный микоз, инсулинома, карциноидный синдром, соматостатинома, рак десны, рак сердца, рак губы, рак оболочки головного мозга, рак ротовой полости, рак нерва, рак неба, рак околоушной железы, рак брюшины, рак зева, рак плевры, рак слюнной железы, рак языка и рак миндалины.
| WO 2015177098 A2, 26.11.2015 | |||
| EVA SCHLINGLOFF AUS BRAUNSCHWEIG, Evaluierung der humoralen Immunantwort gegen Cancer-Testis-Antigene nach autologer und allogener Stammzelltransplantation bei Patienten mit Multiplem Myelom, Hamburg 2016, таблица 5, найдено в интернет 14.10.2022 https://d-nb.info/1150401583/34 | |||
| MANAMI MIYAI, SHINGO EIKAWA et al., |

