Область изобретения
Настоящее изобретение относится к устройству для идентификации опухолевого антигена и способу идентификации опухолевого антигена; опухолевому антигену, идентифицированному в соответствии с применением указанного устройства и/или способа; фармацевтической композиции, содержащей указанный опухолевый антиген; способу лечения рака с использованием указанного устройства и/или указанного способа; способу стратификации пациентов в отношении лечения рака с использованием указанного устройства и/или указанного способа; схемы лечения, включающей стратификацию пациентов в отношении лечения рака с использованием указанного устройства и/или способа и затем введение противоракового терапевтического средства; и опухолевому антигену, идентифицированному с использованием указанного устройства и/или указанного способа, для применения в качестве противораковой вакцины или иммуногенного агента или противораковой терапии.
Предшествующий уровень техники
CD8+ T-клетки играют ключевую роль в обнаружении и устранении клеток, которые презентируют аномальные пептиды на своей поверхности в результате патогенной инфекции, такой как вирусная инфекция или злокачественная трансформация. Перекрестная реактивность Т-клеточного рецептора (TCR) означает, что Т-клетки могут распознавать невероятно большое разнообразие различных пептидов-мишеней. Это явление позволяет относительно небольшому числу Т-клеток распознавать множество молекул pMHC (пептид:главный комплекс гистосовместимости), которые являются типичными для аномальных клеток и поэтому потенциально угрожают здоровью или жизни.
Тем не менее, нежелательным побочным действием этого механизма является, что иммунный ответ, направленный против патогена, может преодолеть порог толерантности для в высокой степени гомологичного аутоантигена, вызывая вредный эффект вне мишени, который опосредован перекрестной реактивностью Т-клеток; процесс, известный как молекулярная мимикрия. В области аутоиммунитета потенциально вредные последствия такой гомологии между собственными и патогенными пептидами хорошо известны, тем не менее, они не были исследованы при раке.
Действительно, считалось, что лучшими прогностическими маркерами для успешного исхода иммунотерапевтического лечения рака является высокая частота мутаций опухолевого антигена и обильная инфильтрация Т-клеток. Это суждение основано на том факте, что опухоль, которая имеет большое количество мутаций, будет иметь более высокие шансы быть распознанной и устраненной путем инфильтрации Т-клеток.
Тем не менее, некоторые исследования продемонстрировали, что качественные свойства опухолевых антигенов могут быть более важны, чем их количество. Кроме того, было обнаружено, что противовирусные Т-клетки заселяют микроокружение опухоли [28], однако до сих пор неясно, является ли их роль активной или просто пассивной.
Онкоиммунология и иммунотерапия полностью изменили путь, при помощи которого рак лечили в течение прошедшего десятилетия, в частности, когда используют ингибиторы контрольных точек (ICI), для которых были обнаружены выдающиеся клинические результаты, но, к сожалению, только для небольшого количества пациентов. Становится очевидно, что иммунотерапия при помощи ICI будет полностью работать только при нацеливании на специфические опухолевые антигены. Тем не менее, на данный момент не существует легкий и быстрый способ идентификации этих опухолевых антигенов.
Здесь авторы изобретения выдвинули гипотезу, что опухоли могут представлять пептиды, которые обладают высокой степенью гомологии или оценкой аффинности с пептидами, имеющими происхождение от патогена, в частности вирусными пептидами, и это могло бы позволить возникшим вследствие патогенеза обладающим перекрестной реактивностью T-клеткам распознавать и уничтожать опухолевые клетки.
Для этого авторы изобретения разработали биоинформатическое устройство под названием HEX (Оценка гомологии ксенопептидов) для автоматической и очень простой идентификации опухолевых антигенов, которые очень похожи на вирусные пептиды. С использованием этого устройства авторы изобретения обнаружили, что противовирусный Т-клеточный иммунитет, опосредованный пептидом с высокой гомологией или оценкой аффинности с раковым антигеном, может также активно контролировать рост опухоли как в профилактических, так и в терапевтических условиях. Это наблюдение демонстрирует перекрестную реактивность активированных Т-клеток как против гомологичных вирусных пептидов, так и против опухолевых пептидов.
Затем авторы изобретения также обнаружили, что гуморальный ответ на цитомегаловирус (CMV) способен стратифицировать ответ у пациентов, страдающих от меланомы, на терапию ингибитором контрольных точек (антитело против PD1). Действительно, было обнаружено, что пептиды, гомологичные CMV и меланоме, идентифицированные посредством использования устройства HEX, способствуют высвобождению Inf-g из мононуклеарных клеток периферической крови (PBMC) пациентов, страдающих от меланомы, сероположительных в отношении CMV.
Изложение изобретения
В соответствии с первым аспектом изобретения предложено устройство для идентификации опухолевого антигена, включающее:
по меньшей мере один проточный канал, содержащий множество подложек, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против MHC (главный комплекс гистосовместимости) или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего pMHC (пептид:главный комплекс гистосовместимости) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела.
Ссылка здесь на антитело против пан-HLA представляет собой ссылку на антитело, которое распознает или обладает специфичностью в отношении любого из различных HLA, присутствующих у млекопитающих, в частности людей.
Более идеально указанное антитело распознает или обладает специфичностью в отношении конкретного типа HLA, такого как MHC-I, и выбранного из по меньшей мере одного из следующей группы: MHC класса I A, B и C.
Дополнительно или альтернативно указанное антитело распознает или обладает специфичностью в отношении конкретного типа HLA, такого как MHC-II, выбранного из по меньшей мере одного из следующей группы: MHC класса II DP, DM, DO, DQ и DR.
Еще более предпочтительно указанное антитело является противочеловеческим антителом.
В предпочтительном воплощении изобретения указанная молекула или комплекс представляет собой комплекс тиольной и алкильной (“еновой”) функциональных групп, в идеале охватывающий стехиометрическое соотношение 1,5 к 1,0 в диапазоне 0,15:0,1-1500:1000 (тетратиол: триаллил).
Более идеально, конструкция устройства основана на инициируемой UV (ультрафиолетом) фотореакции между тиольной и алкильной (“еновой”) функциональными группами. В дополнительном предпочтительном воплощении изобретения устройство изготовлено путем UV фотополимеризации с биотин-PEG4-алкином (биотин-полиэтиленгликоль 4-алкином) (Sigma, 764213). За ней следует реакция с авидин:стрептавидином. Тем не менее, в объеме изобретения также находится присоединение авидина к биологическому объекту до присоединения биологического объекта к основному элементу и наоборот.
В еще одном предпочтительном воплощении указанное устройство функционализировано путем подвергания указанного антитела взаимодействию с указанным стрептавидином. В идеале, более чем одно антитело взаимодействует с указанным стрептавидином, причем каждый комплекс, функционализированный стрептавидином, несет множество, например 2 или 3, указанных антител, например множество антител против пан-HLA.
В еще одном предпочтительном воплощении изобретения микростолбиковую матрицу кондиционировали с белком, таким как бычий сывороточный альбумин (BSA) (в идеале 100 мкг/мл в 15 мМ PBS (забуференный фосфатом физиологический раствор), инкубация в течение 10 мин), после функционализации стрептавидином, но перед связыванием с антителом.
В более предпочтительном аспекте изобретения указанные подложки включают множество микростолбиков, таких как представленные в микрофлюидном устройстве, и, предпочтительно, указанные столбики упорядочены в матрицу в указанном устройстве. В идеале указанные микростолбики изготовлены из композиции, которая является такой же или похожа на композицию, которую используют в обычных микрофлюидных устройствах, например, для осуществления реакций с иммобилизованным ферментом.
В соответствии с дополнительным аспектом изобретения предложен способ идентификации опухолевого антигена, включающий:
1) растворение или суспендирование образца опухоли в жидкости;
2) пропускание указанной жидкости через устройство для идентификации опухолевого антигена, включающее: по меньшей мере один проточный канал, содержащий множество подложек, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против MHC (главный комплекс гистосовместимости) или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего pMHC (пептид:главный комплекс гистосовместимости) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
3) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC (пептид:главный комплекс гистосовместимости) в указанном образце;
4) возможно, извлечение указанного по меньшей мере одного связанного pMHC (пептид:главный комплекс гистосовместимости) в соответствии с частью (3) из указанного устройства;
5) сравнение указанного пептида указанного связанного pMHC (пептид:главный комплекс гистосовместимости) с библиотекой антигенов патогенов для определения того, демонстрирует ли указанный пептид гомологию или аффинность (молекулярную мимикрию) с по меньшей мере одним антигеном патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
6) идентификация указанного пептида в качестве опухолевого антигена для применения в противораковой терапии.
В предпочтительном воплощении любого аспекта изобретения указанная гомология/аффинность может представлять собой любую из следующих процентных долей: 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90 91, 92, 93, 94, 95, 96, 97, 98, 99 и 100%.
Ссылка здесь на гомологию/аффинность включает ссылку на сравнение структур последовательности в части (5) выше с использованием одного из: определенной здесь гомологии/аффинности; или идентичности, которая определяется по количеству идентичных остатков на протяжении определенной длины в данном выравнивании; или сходства, которое определяется по количеству идентичных остатков или консервативных замен, имеющих похожие физико-химические свойства на протяжении определенной длины в данном выравнивании.
В предпочтительном способе в соответствии с изобретением указанная библиотека представляет собой антигены человеческих патогенов и она содержит проверенную библиотеку известных антигенов патогенов, в идеале полученных из патогенных вирусов, или более подходящим образом их протеомов, где вирусы инфицируют млекопитающих, в идеале людей, таких как любой один или более чем один, или действительно любая комбинация следующих вирусов:
Abyssoviridae; Ackermannviridae; Actantavirinae; Adenoviridae; Agantavirinae; Aglimvirinae; Alloherpesviridae; Alphaflexiviridae; Alphaherpesvirinae; Alphairidovirinae; Alphasatellitidae; Alphatetraviridae; Alvernaviridae; Amalgaviridae; Amnoonviridae; Ampullaviridae; Anelloviridae; Arenaviridae; Arquatrovirinae; Arteriviridae; Artoviridae; Ascoviridae; Asfarviridae; Aspiviridae; Astroviridae; Autographivirinae; Avsunviroidae; Avulavirinae; Bacilladnaviridae; Baculoviridae; Barnaviridae; Bastillevirinae; Bclasvirinae; Belpaoviridae; Benyviridae; Betaflexiviridae; Betaherpesvirinae; Betairidovirinae; Bicaudaviridae; Bidnaviridae; Birnaviridae; Bornaviridae; Botourmiaviridae; Brockvirinae; Bromoviridae; Bullavirinae; Caliciviridae; Calvusvirinae; Carmotetraviridae; Caulimoviridae; Ceronivirinae; Chebruvirinae; Chordopoxvirinae; Chrysoviridae; Chuviridae; Circoviridae; Clavaviridae; Closteroviridae; Comovirinae; Coronaviridae; Corticoviridae; Crocarterivirinae; Cruliviridae; Crustonivirinae; Cvivirinae; Cystoviridae; Dclasvirinae; Deltaflexiviridae; Densovirinae; Dicistroviridae; Endornaviridae; Entomopoxvirinae; Equarterivirinae; Eucampyvirinae; Euroniviridae;
Filoviridae; Fimoviridae; Firstpapillomavirinae; Flaviviridae; Fuselloviridae; Gammaflexiviridae; Gammaherpesvirinae; Geminialphasatellitinae; Geminiviridae; Genomoviridae; Globuloviridae; Gokushovirinae; Guernseyvirinae; Guttaviridae; Hantaviridae; Hepadnaviridae; Hepeviridae; Herelleviridae; Heroarterivirinae; Herpesviridae; Hexponivirinae; Hypoviridae; Hytrosaviridae; Iflaviridae; Inoviridae; Iridoviridae; Jasinkavirinae; Kitaviridae; Lavidaviridae; Leishbuviridae; Letovirinae; Leviviridae; Lipothrixviridae; Lispiviridae; Luteoviridae; Malacoherpesviridae; Mammantavirinae; Marnaviridae; Marseilleviridae; Matonaviridae; Mccleskeyvirinae;
Mclasvirinae; Medioniviridae; Medionivirinae; Megabirnaviridae; Mesoniviridae; Metaparamyxovirinae; Metaviridae; Microviridae; Mimiviridae; Mononiviridae; Mononivirinae; Mymonaviridae; Myoviridae; Mypoviridae; Nairoviridae; Nanoalphasatellitinae; Nanoviridae; Narnaviridae; Nclasvirinae; Nimaviridae; Nodaviridae; Nudiviridae; Nyamiviridae; Nymbaxtervirinae; Okanivirinae; Orthocoronavirinae; Orthomyxoviridae; Orthoparamyxovirinae; Orthoretrovirinae;
Ounavirinae; Ovaliviridae; Papillomaviridae; Paramyxoviridae; Partitiviridae; Parvoviridae; Parvovirinae; Pclasvirinae; Peduovirinae; Peribunyaviridae; Permutotetraviridae; Phasmaviridae; Phenuiviridae; Phycodnaviridae; Picobirnaviridae; Picornaviridae; Picovirinae; Piscanivirinae; Plasmaviridae; Pleolipoviridae; Pneumoviridae; Podoviridae; Polycipiviridae; Polydnaviridae; Polyomaviridae; Portogloboviridae; Pospiviroidae; Potyviridae; Poxviridae; Procedovirinae; Pseudoviridae; Qinviridae; Quadriviridae; Quinvirinae; Regressovirinae; Remotovirinae; Reoviridae; Repantavirinae; Retroviridae; Rhabdoviridae; Roniviridae; Rubulavirinae; Rudiviridae; Sarthroviridae; Secondpapillomavirinae; Secoviridae; Sedoreovirinae; Sepvirinae; Serpentovirinae; Simarterivirinae; Siphoviridae; Smacoviridae; Solemoviridae; Solinviviridae; Sphaerolipoviridae; Spinareovirinae; Spiraviridae; Spounavirinae; Spumaretrovirinae; Sunviridae; Tectiviridae; Tevenvirinae; Tiamatvirinae; Tobaniviridae; Togaviridae; Tolecusatellitidae; Tombusviridae; Torovirinae; Tospoviridae; Totiviridae; Tristromaviridae; Trivirinae; Tunavirinae; Tunicanivirinae; Turriviridae; Twortvirinae; Tymoviridae; Variarterivirinae; Vequintavirinae; Virgaviridae; Wupedeviridae; Xinmoviridae; Yueviridae; и Zealarterivirinae.
Более предпочтительно патогенный вирус представляет собой цитомегаловирус (CMV) или вирус Эпштейна-Барр (EBV) или, предпочтительно, вирус герпеса, поксвирус, гепаднавирус, вирус гриппа, коронавирус, вирус гепатита, HIV (вирус иммунодефицита человека) или буньявирус.
Наиболее предпочтительно указанный вирус является не онколитическим, то есть его репликация конкретно не ограничена раковыми клетками.
В альтернативном воплощении указанный вирус является онколитическим, то есть способен инфицировать и уничтожать раковые клетки путем избирательной репликации в опухолевых, а не в нормальных клетках.
В предпочтительном способе указанное сравнение указанного пептида указанного связанного pMHC (пептид:главный комплекс гистосовместимости) с библиотекой антигенов патогенов включает множество оценок (оценка пептидной аффинности и оценка выравнивания, и оценка сходства, и оценка аффинности связывания с MHC) для определения указанных гомологии/аффинности или идентичности, или сходства. Наиболее предпочтительно указанная оценка представляет собой оценку гомологии/аффинности, как описано ниже.
В одном из воплощений получают матрицу из строк (или столбцов), демонстрирующую аминокислотное положение в опухолевом пептиде, и столбцов (или строк), демонстрирующих каждую из 20 стандартных аминокислот, и аминокислотным положениям в опухолевом пептиде присваивают высокую оценку, а другим положениям присваивают низкую оценку сходства с антигеном/пептидом патогена. Опухолевые пептиды с самой высокой степенью гомологии/аффинности с антигеном/пептидом патогена с точки зрения (в порядке убывания предпочтения):
а) структура полноразмерной последовательности (явным образом опухолевый пептид со 100% идентичностью с пептидом патогена получает самую высокую оценку);
б) идентичность ключевых аминокислот в “горячих точках” или ключевых сайтах связывания; и
в) наибольшее количество ключевых аминокислот в “горячих точках” или ключевых сайтах связывания получает самые высокие оценки.
Выравнивания рассчитывают попарно между пептидами, в запросе набор (имеющий происхождение из патогена) против опухолевого набора, или наоборот. Для данной пары пептидов их выравнивание рассчитывают путем суммирования оценок дистанций между парами аминокислот в одном и том же положении. Оценку взвешивают, чтобы приоритизировать сходство между более центральными аминокислотами в пептиде.
Дополнительно осуществляют прогнозы связывающей аффинности MHC класса I. Способы для достижения этого известны в области техники, такие как использование NetMHC (NetMHC4,0 или NetMHCpan4,1.) при помощи интерфейса прикладных программ IEDB (http://tools.iedb.org/main/tools-api/) и затем анализируют и сопоставляют в инструменте.
В альтернативном воплощении изобретения указанное сравнение включает введение в программное обеспечение перечней опухолевых пептидов длиной 8-12 аминокислот. Во-первых, инструмент BLAST используют для обнаружения «хитов» (похожих последовательностей) в библиотеке антигенов патогенов. Для этой задачи, как правило, используют инструмент PAM30, тем не менее, поддерживаются матрицы замещения BLOSUM и PAM через несколько эволюционных дистанций. Затем попарное оптимизирующее выравнивание с использованием по меньшей мере BLOSUM62 и, в идеале, также матрицы замещения, разработанное Kim et al (описано в BMC Bioinformatics. 2009;10:394. Published 2009 Nov 30. doi:10.1186/1471-2105-10-394 Derivation of an amino acid similarity matrix for peptide: MHC binding and its application as a Bayesian prior. Kim Y, Sidney J, Pinilla C, Sette A, Peters B.), осуществляют для того, чтобы оптимизировать выравнивание взвешенным по положению образом. Пара пептидов с высоким сходством в области взаимодействия с TCR (T-клеточный рецептор) (центральная область пептидов) получают более высокую оценку сходства. Наконец, осуществляют прогноз аффинности связывания MHC как для опухолевого, так и для вирусного когнатного пептида с использованием NetMHC4.0 или NetMHCpan4.1 в качестве отдельного инструмента командной строки. Результаты в конечном итоге автоматически анализируются и сопоставляются в рамках инструмента, и создается финальная оценка, которая обобщает вышеупомянутый анализ.
В соответствии с еще одним дополнительным аспектом изобретения предложены противораковые терапевтическое средство или иммуногенный агент, или противораковая вакцина, содержащая опухолевый антиген, идентифицированный с использованием вышеприведенного устройства и/или способа, или обоих из них.
В соответствии с еще одним дополнительным аспектом изобретения предложен опухолевый антиген для применения в противораковой терапии, идентифицированный с использованием вышеприведенного устройства и/или способа, или обоих из них.
В соответствии с еще одним дополнительным аспектом изобретения предложен опухолевый антиген для применения в изготовлении лекарственного средства для лечения рака, где указанный антиген идентифицирован с использованием вышеприведенного устройства и/или способа, или обоих из них.
Наиболее предпочтительно, указанные противораковые терапевтическое средство или иммуногенный агент, или противораковая вакцина или опухолевый антиген представляют собой одно или более чем одно из перечисленных в таблицах 1-6.
В соответствии с еще одним дополнительным аспектом изобретения предложен способ лечения пациентов, страдающих от рака, включающий:
идентификацию опухолевого антигена с использованием указанного устройства и/или способа в соответствии с изобретением и затем введение индивиду указанного опухолевого антигена или использование указанного опухолевого антигена для увеличения популяции T-клеток, активных против указанного опухолевого антигена, и затем введение пациенту указанных T-клеток.
В предпочтительном воплощении указанные T-клетки представляют собой T-клетки ex vivo и/или культивируемые T-клетки, либо аллогенные, либо аутологичные T-клетки.
В соответствии с еще одним дополнительным аспектом изобретения предложены фармацевтическая композиция или иммуногенный агент, или вакцина, содержащие опухолевый антиген в соответствии с изобретением и фармацевтически приемлемый носитель, адъювант, разбавитель или эксципиент.
Подходящие фармацевтические эксципиенты хорошо известны специалистам в данной области техники. Фармацевтические композиции могут быть приготовлены для введения при любым подходящим путем, например путем внутриопухолевой, внутримышечной, внутриартериальной, внутривенной, внутриплевральной, внутривезикулярной, внутриполостной или внутрибрюшинной инъекции, трансбуккального, назального или бронхиального (ингаляция), трансдермального или парентарального пути и могут быть приготовлены любыми способами, хорошо известными в области фармации.
Эта композиция может быть приготовлена путем объединения вышеописанного опухолевого антигена с носителем. Как правило, композиции готовят путем однородного и тщательного объединения опухолевого антигена с жидкими носителями, или тонкоизмельченными твердыми носителями, или обоими, и затем при необходимости формования продукта. Настоящее изобретение распространяется на способы получения фармацевтической композиции, включающие приведение определенного здесь опухолевого антигена в сочетание или ассоциацию с фармацевтически или ветеринарно приемлемым носителем или разбавителем.
В соответствии с еще одним аспектом изобретения предложено комбинированное терапевтическое средство для лечения рака, содержащее: опухолевый антиген, идентифицированный с использованием указанного устройства и/или способа в соответствии с изобретением, и по меньшей мере одно дополнительное противораковое терапевтическое средство.
Предпочтительно указанное дополнительное противораковое терапевтическое средство наиболее подходящим образом содержит циклофосфамид, поскольку он осуществляет понижающую регуляцию T-регуляторных клеток. Тем не менее, как понятно специалистам в данной области техники, указанное дополнительное терапевтическое средство может представлять собой любой противораковый агент, известный в области техники.
Предпочтительно, дополнительное противораковое терапевтическое средство содержит ингибитор контрольных точек (ICI).
Наиболее хорошо охарактеризованные пути для ингибирования контрольных точек представляют собой путь через цитотоксический T-лимфоцитарный белок 4 (CTLA-4) и путь через белок программируемой клеточной гибели 1 (PD-1/PD-L1(лиганд PD-1)). Таким образом, настоящее изобретение может быть использовано в комбинации с по меньшей мере одним модулятором контрольных точек, таким как молекулы антитела против CTLA-4, антитела против PD1 или антитела против PD-L1, для того, чтобы нейтрализовать иммуносупрессивное опухолевое окружение и вызывать сильный антииммунный ответ.
В соответствии с еще одним дополнительным аспектом изобретения предложен способ стратификации пациентов в отношении лечения рака ингибитором контрольных точек, включающий:
1) отбор у пациента образца опухоли;
2) растворение или суспендирование указанного образца в жидкости;
3) пропускание указанной жидкости через устройство для идентификации опухолевого антигена, включающее: по меньшей мере один проточный канал, содержащий множество подложек, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против MHC (главный комплекс гистосовместимости) или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего pMHC (пептид:главный комплекс гистосовместимости) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
4) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC (пептид:главный комплекс гистосовместимости) в указанном образце;
5) возможно, извлечение указанного по меньшей мере одного связанного pMHC (пептид:главный комплекс гистосовместимости) в соответствии с частью (4) из указанного устройства;
6) сравнение указанного пептида указанного связанного pMHC (пептид:главный комплекс гистосовместимости) с библиотекой антигенов человеческих патогенов для определения того, демонстрирует ли указанный пептид гомологию последовательности/аффинность с по меньшей мере одним антигеном человеческого патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
7) идентификацию указанного пептида в качестве опухолевого антигена; и
8) в случае обнаружения указанного пептида, введение указанному пациенту эффективного количества по меньшей мере ингибитора контрольных точек (ICI).
В соответствии с еще одним дополнительным аспектом или воплощением изобретения предложен способ стратификации пациентов в отношении лечения рака ингибитором контрольных точек, включающий:
определение того, является ли пациент CMV-сероположительным, и в случае, если это имеет место, отбор пациента для лечения с использованием эффективного количества по меньшей мере ингибитора контрольных точек (ICI).
В соответствии с еще одним дополнительным аспектом изобретения предложен способ лечения рака, включающий:
1) отбор у пациента образца опухоли;
2) растворение или суспендирование указанного образца в жидкости;
3) пропускание указанной жидкости через устройство для идентификации опухолевого антигена, включающее по меньшей мере один проточный канал, содержащий множество подложек, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против MHC (главный комплекс гистосовместимости) или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего pMHC (пептид:главный комплекс гистосовместимости) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
4) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC (пептид:главный комплекс гистосовместимости) в указанном образце;
5) возможно, извлечение указанного по меньшей мере одного связанного pMHC (пептид:главный комплекс гистосовместимости) в соответствии с частью (4) из указанного устройства;
6) сравнение указанного пептида указанного связанного pMHC (пептид:главный комплекс гистосовместимости) с библиотекой антигенов человеческих патогенов для определения того, демонстрирует ли указанный пептид гомологию последовательности/аффинность с по меньшей мере одним антигеном человеческого патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
7) идентификацию указанного пептида в качестве опухолевого антигена; и введение эффективного количества указанного пептида указанному пациенту для стимулирования или активирования T-клеток против опухолевого антигена и, таким образом, рака, из которого был отобран образец, или использование указанного опухолевого антигена для увеличения популяции T-клеток, активных против указанного опухолевого антигена, и затем введение указанных T-клеток пациенту.
В соответствии с еще одним дополнительным аспектом или воплощением изобретения предложен способ лечения рака, включающий:
определение того, является ли пациент CMV-сероположительным, и в случае, если это имеет место, введение указанному пациенту эффективного количества по меньшей мере ингибитора контрольных точек (ICI).
В соответствии с еще одним дополнительным аспектом изобретения предложен способ стратификации пациентов в отношении лечения аденовирусного рака, включающий:
1) отбор у пациента образца опухоли;
2) растворение или суспендирование указанного образца в жидкости;
3) пропускание указанной жидкости через устройство для идентификации опухолевого антигена, включающее: по меньшей мере один проточный канал, содержащий множество подложек, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против MHC (главный комплекс гистосовместимости) или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего pMHC (пептид:главный комплекс гистосовместимости) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
4) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC (пептид:главный комплекс гистосовместимости) в указанном образце;
5) возможно, извлечение указанного по меньшей мере одного связанного pMHC (пептид:главный комплекс гистосовместимости) в соответствии с частью (4) из указанного устройства;
6) сравнение указанного пептида указанного связанного pMHC (пептид:главный комплекс гистосовместимости) с библиотекой антигенов человеческих патогенов для определения того, демонстрирует ли указанный пептид гомологию/аффинность с по меньшей мере одним антигеном человеческого патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
7) идентификацию указанного пептида в качестве опухолевого антигена; и
8) в случае обнаружения указанного пептида, присоединение указанного пептида к капсиду аденовирусного вектора и введение эффективного количества указанного аденовирусного вектора указанному пациенту.
В предпочтительном воплощении указанный пептид присоединен к указанному аденовирусному вектору с использованием известной технологии, такой как технология, описанная в WO 2015/177098 и/или которая содержится здесь, см. препарат PeptiCRAd.
Ссылка здесь на “эффективное количество” относится к количеству, которое является достаточным для того, чтобы достичь желаемого биологического эффекта, такого как гибель раковой клетки.
Понятно, что эффективная доза будет зависеть от возраста, пола, состояния здоровья и массы реципиента, типа сопутствующего лечения, если такое проводится, частоты лечения и природы желаемого эффекта. Как правило, эффективное количество определяется лицом, осуществляющим лечение.
Наиболее предпочтительно рак, упомянутый здесь, включает любой один или более чем один из следующих видов рака: рак носоглотки, синовиальный рак, гепатоклеточный рак, рак почки, рак соединительных тканей, меланома, рак легкого, рак кишечника, рак толстой кишки, рак прямой кишки, колоректальный рак, рак мозга, рак горла, рак полости рта, рак печени, рак кости, рак поджелудочной железы, хориокарцинома, гастринома, феохромоцитома, пролактинома, Т-клеточный лейкоз/лимфома, неврома, болезнь фон Гиппеля-Линдау, синдром Золлингера-Эллисона, рак надпочечников, анальный рак, рак желчных протоков, рак мочевого пузыря, рак мочеточника, олигодендроглиома, нейробластома, менингиома, опухоль спинного мозга, остеохондрома, хондросаркома, саркома Юинга, рак без выявленного первичного очага, карциноид, карциноид желудочно-кишечного тракта, фибросаркома, рак молочной железы, болезнь Педжета, рак шейки матки, рак пищевода, рак желчного пузыря, рак головы, рак глаза, рак шеи, рак почки, опухоль Вильмса, рак печени, саркома Капоши, рак предстательной железы, рак яичка, болезнь Ходжкина, неходжкинская лимфома, рак кожи, мезотелиома, множественная миелома, рак яичников, эндокринный рак поджелудочной железы, глюкагонома, рак паращитовидной железы, рак полового члена, рак гипофиза, саркома мягких тканей, ретинобластома, рак тонкой кишки, рак желудка, рак тимуса, рак щитовидной железы, трофобластический рак, хорионаденома, рак матки, рак эндометрия, рак влагалища, рак вульвы, акустическая неврома, грибовидный микоз, инсулинома, карциноидный синдром, соматостатинома, рак десен, рак сердца, рак губы, рак оболочек головного мозга, рак рта, рак нервов, рак неба, рак околоушной железы, рак брюшины, рак глотки, рак плевры, рак слюнных желез, рак языка и рак миндалевидной железы.
В конкретном предпочтительном воплощении изобретения указанный образец отобран из меланомы и указанный опухолевый антиген гомологичен антигену или пептиду CMV.
В следующей формуле изобретения и в предшествующем описании изобретения за исключением того, когда в контексте требуется иное вследствие специального языка или необходимого подразумевания, фраза “содержать” или вариации, такие как “содержит” или “содержащий” используют в включающем смысле, то есть для обозначения присутствия заявленных свойств, но не мешая присутствию или добавлению дополнительных свойств в различных воплощениях изобретения.
Все ссылки, включающие любые патент или заявку на патент, цитированные в этом описании, включены сюда путем ссылки. Никакое допущение не принималось в отношении того, что любая ссылка составляет предшествующий уровень техники. Дополнительно, никакое допущение не принималось в отношении того, что любой предшествующий уровень техники составляет часть общего знания в области техники.
Предпочтительные свойства каждого аспекта изобретения могут быть такими, как описано в связи с любыми другими аспектами.
Другие признаки настоящего изобретения будут понятны из следующих примеров. В общем, настоящее изобретение распространяется на любой новый признак или любую новую комбинацию признаков, раскрытых в настоящем описании изобретения (включая прилагаемые формулу изобретения и графические материалы). Таким образом, понятно, что признаки, целочисленные значения, характеристики, соединения или химические группировки, описанные в связи с конкретным аспектом, воплощением или примером изобретения применимы к любому другому описанному здесь аспекту, воплощению или примеру, если он не противоречит им.
Кроме того, если не указано иное, то любое раскрытое здесь свойство может быть заменено на альтернативное свойство, служащее той же самой или похожей задаче.
В описании и формуле изобретения форма единственного числа охватывает множественное число, если в контексте не требуется иное. В частности, когда используют неопределенный артикль, описание следует понимать как охватывающее форму множественного числа, а также форму единственного числа, если в контексте не требуется иное.
Воплощение настоящего изобретения далее будет описано исключительно в качестве примера со ссылкой на следующее, где:
Фиг. 1. [A] Внутри PeptiCHIP. PeptiCHIP внутри состоит из тысяч столбиков, покрытых линкером, к которому присоединен биотин. Биотин затем подвергают взаимодействию со стрептавидином, который затем может взаимодействовать с тремя молекулами HLA-специфического захватывающего антитела. [B]. Микрочиповая технология в качестве новой платформы иммуноочистки для быстрого выявления антигена.
Схематический обзор, описывающий разработанную новую микрочиповую технологию. Тиоленовые микрочипы, включающие свободные поверхностные тиолы, дериватизированные биотин-PEG4-алкинтиоленом (стадия 1) и функционализированные слоем стрептавидина (стадия 2), после чего биотинилированное антитело против пан-HLA иммобилизовали на поверхности микростолбиков (стадия 3) и на микрочип наносили клеточный лизат (стадия 4). После соответствующего времени инкубации и стадий отмывания молекулы HLA элюировали путем добавления 7% уксусной кислоты (стадия 5).
Фиг. 2. Характеристика избирательности функционализации микрочипа биотинилированным антителом против пан-HLA.
А) Связывающая эффективность AlexaFluor 488-стрептавидин на тиоленовых микростолбиках, предварительно покрытых биотин-PEG4-алкином при двух отличающихся временах инкубации со стрептавидином (15 мин и 1 ч). B) Эффект концентрации стрептавидина (не флуоресцентного) в отношении количества иммобилизованного биотинилированного антитела против пан-HLA, количественно оцениваемого при помощи меченного AlexaFluor 488 вторичного антитела. C) Эффект инкубации с BSA в отношении количества иммобилизованного биотинилированного антитела против пан-HLA, определяемого при помощи меченного AlexaFluor 488 вторичного антитела. Эффективность BSA в отношении блокирования сайтов неспецифического связывания оценивали путем предварительного кондиционирования микростолбиковой матрицы с BSA до (BSA-panHLA) или после (panHLA-BSA) иммобилизации биотинилированного антитела против пан-HLA. D) Общее количество биотинилированного антитела против пан-HLA, связанного на единичном чипе в зависимости от циклов загрузки. Для каждого цикла использовали свежую партию той же самой (постоянной) концентрации антитела против пан-HLA. Значимость оценивали путем двустороннего непарного t-теста Стьюдента, * p меньше 0,05.
Фиг. 3. Свойства набора данных HLA-I пептидомов, полученного для линии клеток JY. A) Количество уникальных пептидов, элюированных из 50x106, 10x106 и 1x106 клеток JY. B) Общее распределение длин пептидов для пептидов HLA в трех наборах данных, полученных для линии клеток JY. C-E) Распределение длин пептидов HLA изображено по количеству уникальных пептидов (левая ось y) и процентной доле частоты (правая ось y) для 50x106 (C), 10x106 (D) и 1x106 (E).
Фиг. 4. Точный анализ лигандов HLA, выделенных из линии клеток JY. A) Элюированные 9-меры анализировали в отношении их аффинности связывания с HLA-A*02:01 и HLA-B*07:02. Связывающиеся (зеленые точки) и не связывающиеся (черные точки) определяли с использованием сервера NetMHCpan 4,0 (примененный ранг 2%). B) консенсусные мотивы связывания HLA-I. Кластерный анализ Гиббса осуществили для определения консенсусных мотивов связывания среди элюированных 9-мерных пептидов. Референсный мотив представлен в верхнем правом углу. Представлены кластеры с оптимальной подгонкой (более высокие величины KLD, оранжевая звезда) и последовательность logo представлена с номером HLA-I для каждого кластера.
Фиг. 5A. Блок-схема алгоритма HEX. Матрицу строили на основе аминокислотного состава опухолевого пептида (референсного пептида). Эту матрицу затем использовали для сканирования вирусной базы данных и полученные в результате вирусные пептиды ранжировали в порядке log-вероятности распознавания. Каждому вирусному пептиду присваивали оценку выравнивания и оценку прогноза связывания с MHC-I. Вирусные пептиды-кандидаты ранжировали на основе следующих критериев: оценка прогноза связывания с MHC-I > оценка выравнивания > B-оценка, и пептиды с самыми высокими оценками анализировали экспериментально.
Фиг. 5B. Блок-схема применения существующего программного обеспечения. Перечни опухолевых пептидов длиной 8-12 аминокислот могут быть использованы в качестве входных данных для программного обеспечения. Сначала используют инструмент BLAST для обнаружения «хитов» (похожих последовательностей) в библиотеке антигенов патогенов. Для этой задачи, как правило, используют PAM30, тем не менее, поддерживаются матрицы замещения BLOSUM и PAM через несколько эволюционных дистанций. Затем попарное оптимизирующее выравнивание с использованием по меньшей мере BLOSUM62 и, в идеале, также матрицы замещения, разработанной Kim et al (описано в BMC Bioinformatics. 2009;10:394. Published 2009 Nov 30. doi:10.1186/1471-2105-10-394 Derivation of an amino acid similarity matrix for peptide: MHC binding and its application as a Bayesian prior. Kim Y, Sidney J, Pinilla C, Sette A, Peters B.), осуществляют для того, чтобы оптимизировать выравнивание позиционно взвешенным образом. Пара пептидов с высоким сходством в области взаимодействия с TCR (центральная область пептидов) получают более высокую оценку сходства. Наконец, осуществляют прогноз аффинности связывания MHC как для опухолевого, так и для вирусного когнатного пептида с использованием NetMHC4,0 или NetMHCpan4.1 в качестве отдельного инструмента командной строки. Результаты в конечном итоге автоматически анализируются и сопоставляются в рамках инструмента, и создается финальная оценка, которая обобщает вышеупомянутый анализ.
Фиг. 6: Иммунизация вирусными пептидами, гомологичными опухолевым антигенам, замедляет опухолевый рост.
A: Схема эксперимента на животных: Для оценки того, могут ли вирусные пептиды, похожие на опухолевые пептиды, влиять на опухолевый рост, формировали 4 группы мышей C57BL6. Группу мышей, не подвергавшихся воздействию, использовали в качестве модельной, каждую из других трех групп иммунизировали различными пулами вирусных пептидов. Мышей иммунизировали в 2 момента времени, а именно, в 14 и 7 сутки перед трансплантацией опухоли.
B: Через две недели после первой иммунизации мышам подкожно инъецировали 3x105 клеток мышиной меланомы B16-OVA. После трансплантации за опухолевым ростом следили путем измерения при помощи цифрового штангенциркуля через сутки в течение девятнадцати суток. P-значение рассчитывали с использованием двустороннего дисперсионного анализа ANOVA с множественным сравнением с коррекцией Тьюки.
C: Мышей умерщвляли после достижения конечной точки. Спленоциты мышей в каждой группе собирали и объединяли в пул для анализа ELISpot. На каждый пул затем воздействовали соответствующими вирусными пептидами (вирусные пептиды, гомологичные TYR1, вирусные пептиды, гомологичные TRP2, вирусные пептиды, гомологичные GP100) для оценки ответа на лечение. Пунктирная линия демонстрирует фоновый уровень, образуемый отрицательным контролем.
Фиг. 7: Вирусные пептиды, обладающие более высокой аффинностью в отношении MHC, являются более иммуногенными и могут вызывать более сильный перекрестнореагирующий ответ.
A: Для оценки ответа на соответствующий исходный опухолевый эпитоп спленоциты из каждой иммунизируемой группы объединяли в пул вместе и подвергали воздействию соответствующего опухолевого пептида.
B: Сравнение между ответами, вызванными воздействием на спленоциты объединенными в группы вирусными пептидами и соответствующим исходным опухолевым пептидом.
C: Прогнозируемая аффинность исходной опухоли и соответствующего похожего объединенного вирусного пула пептидов к мышиному MHC класса I. 50 нМ рассматривали как пороговое значение для определения пептидов с “высокой-” и “промежуточной/низкой аффинностью” в соответствии с указаниями IEDB (Базы данных иммунных эпитопов).
D: Корреляция между данными в результате ответа IFN-γ (интерферон гамма) и прогнозируемой аффинностью.
E: Стратификация пептидов на основании их аффинности и способности стимулировать продукцию IFN-γ. Высокоаффинные пептиды (IC50 (средняя ингибирующая концентрация) меньше 50 нМ) вызывали значимо более высокую продукцию INF-γ по сравнению с пептидами, обладающими промежуточной/низкой аффинностью (IC50 больше 50 нМ). P-значение рассчитывали с использованием t-теста с коррекцией Манна-Уитни. Диапазон p-значений обозначали звездочками в соответствии со следующими критериями: больше 0,05 (не значимо), не более 0,05 (*), не более 0,01 (**), не более 0,001 (***), не более 0,0001 (****).
Фиг. 8. Вирусные пептиды, гомологичные опухолевым антигенам, могут уменьшать опухолевый рост при уже сформировавшихся опухолях.
A: Схема эксперимента на животных: 4 группы мышей C57BL формировали для каждой тестируемой линии опухолевых клеток. Мышам подкожно инъецировали клетки B16-OVA или B16F10 в 0 сутки. После того, как опухоль становилась пальпируемой, мышей обрабатывали физиологическим раствором (модельная группа), непокрытым аденовирусом (группа непокрытого вируса), аденовирусом, покрытым пулом вирусных пептидов, гомологичных TRP2 (Viral PeptiCRAd, VPC), или аденовирусом, покрытым пептидом TRP2 (TRP2 PeptiCRAd, TPC).
B: Индивидуальный рост опухоли B16 OVA. Пороговый уровень, который определяет успех терапии, идентифицируют по медиане для всех объемов опухоли за последние сутки.
C: B16 Объем опухоли OVA в момент конечной точки. Медиана объема опухоли, представленная в виде пунктирной линии, определяет пороговый уровень успеха терапии.
D: График вероятности B16 OVA демонстрирует количество тех, у кого проявляется реакция, в каждой обрабатываемой группе.
E: Индивидуальный рост опухоли B16 F10. Пороговый уровень, который определяет успех терапии, идентифицируют по медиане для всех объемов опухоли за последние сутки
F: Объем опухоли B16 F10 в момент конечной точки. Медиана объема опухоли, представленная в виде пунктирной линии, определяет пороговый уровень успеха терапии.
G: График вероятности B16 OVA демонстрирует количество тех, у кого проявляется реакция, в каждой обрабатываемой группе.
(C-F) P-значение рассчитывали с использованием одностороннего ANOVA с коррекцией Тьюки. Диапазон p-значений обозначали звездочками в соответствии со следующими критериями: больше 0,05 (не значимо), не более 0,05 (*), не более 0,01 (**), не более 0,001 (***), не более 0,0001 (****).
(D-G) P-значение рассчитывали с использованием теста хи-квадрат (и точного критерия Фишера) отношения шансов. Диапазон p-значений обозначали звездочками в соответствии со следующими критериями: больше 0,05 (не значимо), не более 0,05 (*), не более 0,01 (**), не более 0,001 (***), не более 0,0001 (****).
Фиг. 9. T-клеточная перекрестная реактивность между вирусными и опухолевыми антигенами, несущими степень гомологии/аффинности. На PBMC, полученные у пациента с HLA-A*02:01 и с высоким уровнем в сыворотке крови Ab (антител) против CMV, воздействовали пептидами, представленными в таблице 2. Уровень IFN-γ, секретируемого активированными CD8+ T-клетками, обнаруживали при помощи анализа ELISpot. Пунктирная линия указывает на уровень шума, возникающего в результате неспецифической активации CTL в отрицательном контроле (DMSO (диметилсульфоксид)) (A). PBMC, полученные у пациента с HLA-A*02:01 и с высоким уровнем в сыворотке крови Ab против CMV и у здорового донора (HS), который был положительным в отношении ответа CMV, тестировали в отношении ответа против CMV при помощи анализа ELISpot с использованием специфического в отношении CMV HLA-A*02:01 усеченного пептида NLVPMVATV. P-значение рассчитывали с использованием t-теста с коррекцией Манна-Уитни (B).
Фиг. 10. Платформа на основе микрочипа выявляет иммунопептидомный профиль в скудных опухолевых биопсиях. A) Здесь обобщены масса образцов до обработки, общее количество, и уникальные пептиды, и обогащение 7-13-мерными образцами. B) Распределение длин пептидов в отношении абсолютного количества и процентной доли представлены в виде столбчатой диаграммы.
Фиг. 11. Иммунопептидомный анализ органоидов, полученных у пациента (PDO), страдающего от ccRCC (светлоклеточная почечно-клеточная карцинома) и от рака мочевого пузыря. A) Множество уникальных пептидов, обнаруженных в PDO для ccRCC и рака мочевого пузыря. B) Распределение длин пептидов представлено в виде общего количества уникальных пептидов (левая ось y) и процентной доли обнаружения (правая ось y) для каждого PDO (ccRCC верхний блок, для мочевого пузыря нижний блок).
Фиг. 12. Тестирование пептидов. Мышам Balb/c подкожно инъецировали сингенную опухолевую модель CT26 в левый и правый бок (0 сутки, фиг. 12A). После формирования опухолей (7 сутки, фиг. 12A) Valo-mD901 покрывали парой каждого модифицированного полилизином пептида в составленном авторами изобретения списке (PeptiCRAd1, PeptiCRAd2, PeptiCRAd3, таблица) и инъецировали внутрь опухоли только в правую опухоль. PeptiCRAd4, состоящий из Valo-mD901, покрывали gp70423-431 (AH1-5), представляющим собой известный иммунодоминантный антиген CT26, получен из аутоантигена, кодируемого в геноме. Модельную и Valo-mD901 группы также использовали в качестве контроля. PeptiCRAd 1 и PeptiCRAd2 улучшали контроль опухолевого роста, а также Valo-mD901 в повреждениях, подвергнутых инъекции (фиг. 12B, правая часть графической панели), как представлено также в единичном опухолевом росте для каждой мыши для каждой обрабатываемой группы. Строго говоря, только PeptiCRAd1 значительно улучшал противоопухолевую активность в необработанной опухоли, тогда как Valo-mD901 не вызывал эффекта (фиг. 12B, левая часть графической панели).
Пептиды в PeptiCRAd1 получены в результате анализа HEX. PeptiCRAd4 состоит из Valo-mD9, покрытых gp70 423-431 (AH1-5).
Фиг. 13. Изображено схематическое представление изобретения. Устройство в соответствии с изобретением представлено в форме диаграммы, оно ранее было функционализировано множеством антител против MHC (главный комплекс гистосовместимости) или антител против человеческого пан-лейкоцитарного антигена (HLA), которые способны захватывать pMHC (пептид:главный комплекс гистосовместимости). Опухолевый лизат пропускают через устройство, и антитела захватывают pMHC. pMHC элюируют и пептиды из комплекса pMHC анализируют, например, посредством жидкостной хроматографии с масс-спектрометрией (LC-MS/MS), в результате, получают перечень опухолевых пептидов. Каждый опухолевый пептид сравнивают с библиотекой белков патогенов (включая антигены) для определения уровня гомологии/аффинности между каждым опухолевым пептидом, идентифицированным с использованием устройства, и белками патогенов (включая антигены патогена/пептиды патогена) в библиотеке. Последнее обеспечивает формирование перечня кандидатов, идеальным образом приоритизированных с точки зрения гомологии/аффинности для применения в противораковой терапии.
Таблица 1. Пептиды, использованные для эксперимента на животных. Известные опухолевые пептиды меланомы (TRP2180-188, hGP10025-33 и TYR208-216) анализировали при помощи HEX. Отбирали самые лучшие вирусные пептиды-кандидаты, предложенные программным обеспечением, и пул, составленный самыми лучшими 4 ксенопептидами на каждый исходный опухолевый эпитоп тестировали in иммунодоминантный vivo.
Таблица 2. Пептиды, использованные для ELISpot в отношении PBMC пациентов. Известные ассоциированные с меланомой антигены анализировали при помощи HEX. Самый лучший человеческий полученный для CMV пептид-кандидат для каждого антигена выбирали для тестирования in vitro.
Таблица 3. Сравнительный анализ между основанной на микрочипе технологии иммунопреципитации и стандартным способом. В таблице приведено общее количество антитела, нанесенного в рамках микрочиповой IP технологии, и при помощи стандартной процедуры.
Таблица 4. Демонстрирует выход после HEX для идентифицированных 13 опухолевых ограниченных MHC пептидов с их соответствующими пептидами патогена.
Таблица 5. Таблица пептидов, протестированных в анализе ELISPOT.
Таблица 6. Таблица выбранных белков для анализов PeptiCRAd in vivo, представленных на фиг. 12.
Материалы и методы
Изготовление устройства
Устройство в соответствии с изобретением (известное как PeptiCHIP) представляет собой проточную конструкцию, содержащую тысячи столбиков, которые покрыты линкером, к которому присоединен биотин. Биотин в свою очередь связан со стрептавидином, который связан с HLA-специфическим захватывающим антителом, конкретно тремя молекулами HLA-специфического захватывающего антитела.
С использованием обычных способов микрофлюидного производства изготовление PeptiCHIP основано на UV-инициированной фотореакции между тиольной и аллильной (“еновой”) функциональными группами в соответствии со следующим нестехиометрическим соотношением 150:100 (тетратиол: триаллил).
Непосредственно после изготовления PeptiCHIP дериватизируют биотин-PEG4-алкином (Sigma, 764213) путем UV фотополимеризации с последующей реакцией с авидином. Это осуществляют в соответствии со следующим.
Стадия 1: Готовили 1 мМ биотин-PEG4-алкиновый раствор (концентрированный раствор биотин-PEG4-алкин в 10 мМ этиленгликоле). Отбирали небольшую аликвоту и добавляли 1% (масс./об.) фотоинициатора (Igracure® TPO-L, BASF) с использованием 10% фотоинициаторного концентрированного раствора в метаноле, таким образом, окончательный раствор представляет собой 1 мМ биотин + 1% Lucirin в EG-MeOH 9:1*
Заполняли чип одним объемом раствора биотин-PEG4-алкина и 1% Lucirin в EG:MeOH 9:1 (об./об.), подвергали воздействию UV в течение 1 мин (с использованием LED UV лампы λ=365 нм, l=15 мВт/см2).
Тщательно промывали сначала метанолом, затем водой milli-Q, промывали 1-2 мл каждого растворителя через канал и хранили его сухим (если необходимо).
Стадия 2: После дериватизации биотином чип функционализировали стрептавидином. Готовили концентрированный раствор стрептавидина 0,01 мг/мл в PBS и его добавляли к чипу в течение 15' в темноте при комнатной температуре. Промывали 3 раза 200 мкл PBS.
Добавляли BSA 100 мкг/мл в 15 мМ в PBS в течение 10' при комнатной температуре.
Стадия 3: Реакция с антителом против пан-HLA. Добавляли 25 мкл антитела против человеческого HLA-A,B,C с биотином 1,6 мкг/мкл (Biolegend CAT. Номер по каталогу 311434) в течение 15' при комнатной температуре. Затем три раза промывали 200 мкл PBS и затем получали чип, готовый для иммунопреципитации комплекса MHC.
Приготовление образца опухоли: открепляли клетки при помощи EDTA (этилендиаминтетраукусная кислота) 4 мМ и однократно промывали PBS. Добавляли 25 мкл Igepal 1% в PBS + ингибиторы протеазы. Центрифугируют 500xg 10' +4°C. Центрифугируют 20000xg 10' +4°C.
Оптимизированные микрофлюидные столбиковые матрицы
Стадии иммунопреципитации осуществляли в одном микрофлюидном чипе путем добавления биотинилированного антитела пан-HLA к предварительно функционализированной стрептавидином структуре твердой подложки (то есть микростолбиковая матрица) и затем иммобилизовали HLA на покрытой антителом пан-HLA твердой поверхности.
В заключение, нестехиометрические тиоленовые (OSTE) полимерные микрсотолбиковые матрицы изготавливали при помощи методики UV- формования реплик и биотинилировали. Затем биотинилированные микростолбики функционализировали стрептавидином и добавляли биотинилированное антитело пан-HLA, после чего клеточный лизат загружали непосредственно в микрофлюидный чип для избирательного захвата комплексов HLA-I. После надлежащего промывания захваченные комплексы HLA-I элюировали при комнатной температуре путем применения 7% уксусной кислоты (фиг. 1B). Затем протокол осуществляли в соответствии со стандартным процессом иммунопептидомики, включающим очистку элюированных пептидов HLA при помощи SepPak®-C18 в ацетонитриле и их упаривание досуха с использованием вакуумного центрифугирования.
Эффективность функцонализации стрептавидином на микростолбиковых матрицах исследовали в отношении двух различных времен инкубации (15 мин и 1 ч) с помощью флуоресцентного AlexaFluor488-стрептавидина. Обнаружили, что более короткое время инкубации было достаточно длительным для построения первого стрептавидинового слоя (фиг. 2A). Кроме того, для определения эффекта концентрации стрептавидина на окончательное количество иммобилизованного биотинилированного антитела пан-HLA тестировали несколько концентраций нефлуоресцентного стрептавидина в присутствии фиксированного количества биотинилированного антитела пан-HLA. В этом случае биотинилированное антитело пан-HLA инкубировали в течение 15 мин с последующими тремя стадиями промывания PBS (200 мкл для каждой стадии). Для определения количества иммобилизованного биотинилированного антитела пан-HLA при каждой концентрации стрептавидина использовали флуоресцентно меченное вторичное антитело AlexaFluor488 для титрования иммобилизованного биотинилированного антитела пан-HLA. Интересно, что даже 10-кратное увеличение концентрации стрептавидина не оказывало влияния на количество иммобилизованного биотинилированного антитела пан-HLA (фиг. 2B), что, вероятно, являлось результатом стерических затруднений, ограничивающих количество доступных сайтов связывания стрептавидина. На основании этого открытия не исследовали никакие дополнительные концентрации стрептавидина, но самую высокую протестированную концентрацию стрептавидина (0,1 мг/мл) использовали во всех последующих экспериментах для обеспечения максимального связывания биотинилированного антитела пан-HLA. Тем не менее, для дополнительного изучения избирательности связывания антитела с функционализированной стрептавидином поверхности микростолбиков влияние дополнительной стадии покрытия бычьим сывороточным альбумином (BSA) на количество иммобилизованного биотинилированного антитела пан-HLA исследовали для устранения неспецифических взаимодействий. Для этого микростолбиковую матрицу предварительно кондиционировали BSA (100 мкг/мл в 15 мМ PBS, инкубация в течение 10 мин) после функционализации стрептавидином и эффективность последующего связывания биотинилированного антитела пан-HLA снова определяли с помощью флуоресцентно меченного вторичного антитела. Эта процедура существенно уменьшала количество иммобилизованного антитела пан-HLA по сравнению с поверхностями без предварительного кондиционирования (фиг.2С), позволяя предположить, что неспецифические сайты связывания могут быть блокированы с помощью простой стадии предварительной инкубации с BSA. Таким образом, стадию инкубации BSA адаптировали для всех дальнейших экспериментов.
Наконец, авторы изобретения стремились охарактеризовать максимальное количество иммобилизованного биотинилированного антитела пан-HLA, которое может быть связано с одним чипом с использованием оптимизированного протокола. Последнее оценивали путем использования нескольких циклов загрузки партии нового антитела с одинаковой концентрацией (0,5 мг/мл) на один микрофлюидный чип. В этом случае количество иммобилизованного антитела пан-HLA определяли путем сравнения количества антитела пан-HLA в подаваемом растворе с таковым в выходном растворе посредством анализа ELISA. Было обнаружено, что количество иммобилизованного антитела увеличивается почти линейно вместе с количеством циклов загрузки (фиг. 2D), что позволяет точно корректировать общее количество иммобилизованного биотинилированного антитела пан-HLA на основе количества циклов загрузки. После семи циклов количество иммобилизованного антитела достигло приблизительного количества 45 мкг, которое является достаточным, по меньшей мере теоретически, для иммунопептидомного исследования скудного количества биологического материала, поскольку 10 мг пан-HLA (3,88x1016 молекул антитела) требуется в соответствии со способами из уровня техники для исследования количества клеток 109 [22 А] (таблица 3).
С помощью микрочиповой установки можно иммобилизовать 1,74х1014 молекул антитела и технически можно исследовать 4,5х106 клеток.
Обогащение антигеном на основе микрочипа, реализуемое в технологическом процессе иммунопептидомики, позволяет идентифицировать представленные естественным образом пептиды HLA-I
Чтобы оценить, можно ли использовать разработанный тиол-еновый микрочип в качестве платформы для обнаружения антигена, осуществляли иммуноочищение пептидов HLA из лимфобластоидной линии B-клеток человека JY. Линия JY имеет высокую экспрессию HLA класса I и гомозиготна в отношении трех аллелей, распространенных в человеческой популяции (HLA-A*02:01, HLA-B*07:02 и HLA-C*07:02), и она была широко принята для лигандомного анализа. Следовательно, линию клеток JY рассматривали в качестве подходящей модели для тестирования технологии обогащения антигена путем иммунопреципитации на основе микрочипа.
Следовательно, комплексы HLA-I иммуноаффинно очищали с использованием тиол-еновых микрочипов, функционализировали количеством антитела пан-HLA, как описано выше. Кроме того, для определения чувствительности предложенного авторами изобретения подхода, протокол проверяли с использованием низкого общего количества клеток, такого как 50x106, 10x106 и 1x106. Лизаты загружали в микрочипы, и после адекватной промывки PBS пептиды элюировали 7% уксусной кислотой и анализировали посредством тандемной масс-спектрометрии. Весь технологический процесс от функционализации стрептавидином до элюирования опухолевых пептидов занимал в среднем менее 24 часов. Строгий порог частоты ложного обнаружения 1% для идентификации пептида и белка применяли для получения данных с высокой достоверностью. Авторам изобретения удалось идентифицировать 5589, 2100 и 1804 уникальных пептидов из 50х106, 10х106 и 1х106 клеток, соответственно (фиг.3А).
Поскольку авторы изобретения стремились тщательно проанализировать способность микрочиповой технологии обогащать природные связывающие HLA-I вещества и избегать возможных совместного элюируемых загрязняющих веществ, авторы изобретения широко охарактеризовали элюируемые пептиды. Во-первых, элюируемые пептиды из клеточной линии JY представляли типичное распределение по длине набора данных лигандома, причем 9-меры представляли собой наиболее обогащенный вид пептидов (фиг. 3B-E). Затем определяли спрогнозированную аффинность связывания для двух аллелей HLA-I (HLA-A*0201 и HLA-B*0702), экспрессированных в клетках JY. Клетки JY также имеют низкий уровень аллели HLA-C*0702, но мотив связывания перекрывается с мотивами HLA-A*0201 и HLA-B*0702; следовательно, только эти аллели были рассмотрены в последующем анализе.
Из уникальных 9-меров 78%, 83% и 67% были спрогнозированы как связывающиеся (описанные как связывающиеся в NetMHCpan4.0, с применяемым критерием 2% [24A-26A]) с аллелями HLA-A*0201 или HLA-B*0702 для 50x106, 10x106 и 1x106 клеток, соответственно (фиг. 4A). Кроме того, проводили анализ Гиббса для деконволюции консенсусных мотивов, связывающихся с соответствующими аллелями HLA-I из элюированных 9-мерных пептидов; они сгруппированы в две отличающиеся группы с предпочтением уменьшенной аминокислотной сложности для остатков в положениях P2 и Ω, что значительно совпадает с известными для HLA-A*0201 и HLA-B*0702 (фиг. 4В).
Затем, для того, чтобы определить роль идентифицированных пептидов, проводили анализ обогащения в терминах генной онтологии (GO) в полученном авторами изобретения списке 9-мерных связывающихся исходных белков. Авторы изобретения обнаружили обогащение ядерных и внутриклеточных белков, в основном тех, которые взаимодействуют с ДНК, РНК или вовлечены в катаболическую активность. Наконец, авторы изобретения разработали анализ уничтожения in vitro для того, чтобы дополнительно продемонстрировать возможность микрочиповой технологии для выделения пептидов в комплексе с HLA-I. Для этого набор из трех пептидов был выбран из полученного авторами изобретения набора данных JY для стимулирования соответствующих HLA РВМС; CD8+ T-клетки очищали из РВМС и принимали в качестве эффекторных клеток в совместной культуре с клетками JY. Для учета неспецифической цитотоксичности, обусловленной самими эффекторными клетками, в качестве контроля использовали нестимулированные РВМС. Затем контролировали цитолиз в реальном времени. Интересно, что CD8 + T-клетки, на которые воздействовали пептидами QLVDIIEKV (SEQ ID NO: 75; название гена PSME3) и KVLEYVIKV (SEQ ID NO: 76; название гена MAGEA1), продемонстрировали приблизительно 10% специфический цитолиз, тогда как CD8 + T-клетки, на которые воздействовали пептидом ILDKKVEKV (SEQ ID NO: 77; название гена HSP90AB3P), продемонстрировали 15% специфический цитолиз, указывая на специфический лизис в присутствии определенных пептидов.
Чтобы оценить достоверность полученных авторами изобретения списков пептидов HLA-I, идентифицированных при помощи микрочиповой технологии, авторы изобретения изучили SysteMHC, представляющее собой хранилище иммунопетидомных данных, генерируемых при помощи масс-спектрометрии. Среди уникальных 9-мерных связывающихся белков, идентифицированных в полученных авторами изобретения данных, 69%, 77% и 81% также были обнаружены в ранее опубликованном наборе лигандомных данных, полученном для линии клеток JY (ID в базе данных PRIDE PXD000394) [3А] для 50x106, 10x106 и 1x106 клеток, соответственно (фиг. 6A). Кроме того, была подтверждена положительная корреляция между количеством исходного белка и представлением HLA, причем наиболее распространенные белки являются основным источником пептидов HLA.
Следовательно, эти результаты продемонстрировали, что протокол на основе чипов может быть использован в качестве надежной платформы иммунопреципитации в рамках иммунопептидомного технологического процесса.
Использование устройства
Нанесение образца на чип и элюирование фракций для последующего анализа
Наносили образец в течение нескольких циклов и инкубировали в течение 5 минут (при 4°С). 3 раза промывали 200 мкл PBS и аспирировали последнюю промывку для опустошения чипа. Готовили раствор 50% MeOH и 50% MilliQ. С использованием предшествующего раствора готовили 7% раствор уксусной кислоты. Наносили этот раствор на чип и фракцию собрали. Время элюирования составляло 5 минут. В тот же день очищали собранные фракции. Готовили картридж SepPak для каждого образца ткани для образца пептидов HLA-I и метили их. С использованием шприца и специализированного адаптера сначала промывали картридж 1 мл 80% ACN (ацетонитрил) в 0,1% TFA (трифторуксусная кислота), затем дважды 1 мл 0,1% TFA. Наносили каждый из биологических образцов на картридж SepPak. Медленно пропускали их (скорость приблизительно 1 мл в течение 20 с). Дважды промывали картриджи 1 мл 0,1% TFA. Элюировали связывающиеся с HLA пептиды в собирающую пробирку с 300 мкл 30% ACN в 0,1% TFA.
HEX (Оценка гомологии ксенопептидов)
Оценка гомологии (/аффинности) ксенопептидов (HEX) представляет собой новую платформу in silico, которая сравнивает сходство между опухолевыми пептидами (референсные пептиды) и полученными у патогена, такими как вирусные, пептидами (пептиды, представляющие интерес). В ней используется несколько параметров для ускорения отбора пептидов-кандидатов. Это осуществляют путем включения обоих новых способов (алгоритм оценки пептида и оценки выравнивания) и интегрирования предварительно существующих способов (прогноз связывания MHC-I). HEX сопровождается множеством предварительно компилированных баз данных известных белков, таких как белки, имеющие происхождение из вирусных патогенов и человеческого протеома (33).
Ассоциированную матрицу оценки генерировали ad hoc на основе аминокислотного состава референсного пептида в противоположность экспериментальному. В частности, в матричных строках, представляющих положение аминокислоты в пептиде, и столбцах, представляющих каждую из 20 стандартных аминокислот, положениям аминокислот референсного пептида присваивали такую же высокую оценку, а другим положениям присваивали такую же низкую оценку.
Выравнивания вычисляют попарно между пептидами в наборе запросов по сравнению с референсным набором. Для данной пары пептидов их выравнивание рассчитывают путем суммирования оценок расстояния между парами аминокислот в одном и том же положении. Оценка взвешена для того, чтобы приоритизировать сходство между более центральными аминокислотами в пептиде. HEX поддерживает матрицы замещения BLOSUM и PAM на нескольких эволюционных расстояниях. В частности, BLOSUM 62 представлял собой матрицу, выбранную для этого исследования.
Прогнозы аффинности связывания MHC класса I осуществляют с использованием NetMHC [27] при помощи интерфейса прикладного программирования IEDB (http://tools.iedb.org/main/tools-api/) и затем анализировали и сопоставляли в рамках инструмента. Пользователь может указать требуемый способ оценки или вернуть ряд рекомендуемых результатов. Поддерживаются прогнозы для множества аллелей MHC-I человека и мыши.
Пользователи способны выбирать пептиды по своим собственным критериям или выбирать пептиды в соответствии с моделью случайного леса. Случайный лес был обучен на экспериментальных результатах для пептидов, выбранных авторами изобретения. Важность признака определяли путем увеличения среднеквадратичной ошибки (MSE) вне набора (OOB) и перекрестно валидировали на невидимом образце пептидов. HEX был разработан как веб-приложение с использованием пакета R Shiny и доступен по ссылке https://picpl.arcca.cf.ac.uk/hex/app/ без регистрации пользователя. Исходный код доступен по адресу https://github.com/whalleyt/hex.
В альтернативном воплощении изобретения HEX поддерживает матрицы замен как BLOSUM, так и PAM через несколько эволюционных дистанций. В частности, BLOSUM 62 была матрицей, использованной в этом исследовании. HEX сначала осуществляет поиск BLAST поиск с использованием PAM30, чтобы найти «хиты» (похожие последовательности) в референсной библиотеке. Затем HEX выполняет попарное уточняющее выравнивание с использованием как BLOSUM62, так и матрицы замен, разработанной Kim et al.
Ссылка: Kim Y, Sidney J, Pinilla C, Sette A, Peters B. Derivation of an amino acid similarity matrix for peptide: MHC binding and its application as a Bayesian prior. BMC Bioinformatics. 2009;10:394. Published 2009 Nov 30. doi:10,1186/1471-2105-10-394.
Прогнозы аффинности связывания MHC класса I получали с использованием NetMHC4.0 (или NetMHCpan4.1) в качестве автономного инструмента командной строки и затем анализировали и сопоставляли в рамках инструмента. Пользователь может указать желаемый способ оценки или вернуть множество рекомендуемых результатов. Поддерживаются прогнозы для множества аллелей MHC-I человека и мыши.
Пациенты и образцы
В общей сложности 16 пациентов, страдающих от метастатической меланомы 4 стадии, лечили моноклональным антителом против PD1 в Комплексном Онкологическом Центре Центральной больницы Хельсинского Университета (HUCH). Пациентов случайным образом отбирали для осуществления инфузий ниволумаба (n равно 7) каждую вторую неделю или инфузий пембролизумаба (n равно 9) каждую третью неделю. Исследование было одобрено этическим комитетом Центральной больницы Хельсинкского университета (HUCH) (Dnro 115/13/03/02/15). Письменное информированное согласие было получено у всех пациентов, и исследование проводили в соответствии с Хельсинкской декларацией.
Образцы периферической крови (3 мл крови с EDTA, 50 мл крови с гепарином) собирали для трех моментов времени; до начала лечения, после одного и трех месяцев лечения. Из них плазму отделяли путем центрифугирования и затем хранили при -70°C. Уровни CMV и EBV IgG измеряли в размороженных образцах плазмы крови с EDTA с использованием VIDAS CMV IgG (BioMérieux, Marcy-l'Etoile, France) и наборов Siemens Enzygnost Anti-EBV/IgG (Siemens Healthcare Diagnostics, Marburg, Germany). Иммуноглобулины (IgA, IgM, IgG) в размороженной плазме крови с гепарином измеряли в центральной лаборатории Центральной больницы Хельсинкского Университета (HUSLAB).
Клеточные линии и человеческие образцы
Линию клеток мышиной меланомы B16-F10 приобретали в Американской Коллекции Типовых Культур (ATCC; Manassas, VA, USA). Клетки выращивали в RPMI (Gibco, Thermo Fisher Scientific, US) с 10% фетальной бычьей сывороткой (FBS) (Life Technologies), 1% глутамаксом (Gibco, Thermo Fisher Scientific, US) и 1% пенициллин и стрептомицин (Gibco, Thermo Fisher Scientific, US) при 37°C/ 5% CO2.
Клеточная линия B16-OVA, представляющая собой линию клеток мышиной меланомы, модифицированная таким образом, чтобы конститутивно экспрессировать овальбумин цыпленка (OVA), любезно предоставлена профессором Richard Vile (Mayo Clinic, Rochester, MN, USA). Эти клетки выращивали в RPMI с низким уровнем глюкозы (Gibco, Thermo Fisher Scientific, US) с 10% FBS (Gibco, Thermo Fisher Scientific, US), 1% глутамаксом, 1% пенициллином и стрептомицином (Gibco, Thermo Fisher Scientific, US) и 1% генетицином (Gibco, Thermo Fisher Scientific, US) при 37°C/ 5% CO2.
Все клетки были протестированы в отношении загрязнения микоплазмой с помощью имеющегося в продаже набора для обнаружения (Lonza - Basel, Switzerland). Выделенные человеческие PBMC замораживали в FBS, дополненной 10% DMSO, и затем поддерживали в жидком азоте до использования. Криоконсервированные РВМС размораживали и выдерживали в течение ночи при 37°С/5% CO2 в полной среде RPMI, дополненной 10% FBS, 1% глутамаксом, 1% пенициллином-стрептомицином перед нанесением их на планшет для ELISPOT.
Пептиды
Все пептиды, использованные в этом исследовании, были приобретены в Zhejiang Ontores Biotechnologies Co. (Zhejiang, China) 5 мг, чистота более 90%. Последовательности всех пептидов, использованных в этом исследовании, можно найти в таблицах 1 и 2.
Приготовление PeptiCRAd
Все комплексы PeptiCRAd, описанные в данной работе, готовили путем смешивания аденовирусов и пептидов с поли-K хвостовыми группами в соответствии со следующим протоколом: 1x109 в.ч. (вирусных частиц) смешивали с 20 мкг пептидов с поли-K хвостовыми группами (ресуспендированными в воде); после перемешивания на вихревой мешалке смесь инкубировали при комнатной температуре в течение 15 мин; PBS добавляли после инкубации до объема инъекции (50 мкг/мышь), затем раствор вновь перемешивали на вихревой мешалке и использовали для анализов или для инъекций животным. Для TRP2-PeptiCRAd, 1x109 в.ч. смешивали с 20 мкг пептида 6K-TRP2180-188, тогда как Viral-PeptiCRAd готовили с использованием 1x109 в.ч., смешанных с 5 мкг каждого вирусного 6K-пептида, гомологичного TRP2180-188.
Новые PeptiCRAd готовили перед каждым экспериментом с использованием свежих реактивов. Все разбавления вируса и пептидов, требующиеся перед инкубацией для приготовления PeptiCRAd, осуществляли в стерильном PBS или воде. PeptiCRAd затем разбавляли в буфере, требующемся для анализа.
Вирусы получали, размножали и охарактеризовывали, как описано в [20].
Эксперименты на животных и этические разрешения
Все эксперименты на животных были рассмотрены и одобрены Комитетом по экспериментам на животных Университета Хельсинки и местными органами власти Южной Финляндии.
Все эксперименты осуществляли с использованием мышей C57BL/6JOlaHsd, полученных из Scanbur (Karlslunde, Denmark).
Для эксперимента с иммунизацией иммунокомпетентных самок мышей C57BL/6J в возрасте 8-9 недель распределяли на 4 группы. N = 3 мыши использовали в качестве модельной группы, n = 7 мышей использовали для формирования каждой из трех различных групп, подвергаемых обработке. Каждую из групп, подвергаемых обработке, вакцинировали отличающейся группой ксенопептидов. Мышей вакцинировали дважды и инъекции осуществляли с интервалом в одну неделю (0 и 7 сутки) в основание хвоста 40 мкг пептидов и 40 мкг адъюванта (VacciGrade poly(I:C) - Invivogen) в окончательном инъецируемом объеме 100 мкл. Мышей, не подвергавшихся воздействию (которым инъецировали PBS), использовали в качестве модельной группы. В четырнадцатые сутки мышам инъецировали 3*105 клеток B16-OVA в правый бок и за ростом опухоли следили до достижения конечной точки.
Для лечения сформировавшихся опухолей авторы изобретения тестировали 2 различные линии опухолевых клеток: клетки B16-OVA и более агрессивные клетки B16F10. 3*105 клеток B16-OVA и 1*105 клеток B16-F10 инъецировали подкожно в правый бок иммунокомпетентных самок мышей C57BL/6J в возрасте 8-9 недель. Затем этих мышей случайным образом распределяли на 4 группы по 7-8 мышей для каждой линии опухолевых клеток. Модельную группу обрабатывали PBS; вторую группу обрабатывали непокрытым аденовирусом; третью группу обрабатывали аденовирусом, покрытым TRP2180-188 (TRP2-PeptiCRAd); последнюю группу обрабатывали аденовирусом, покрытым вирусными пептидами, гомологичными TRP2-(Viral-PeptiCRAd).
Мышей дважды подвергали внутриопухолевой обработке и инъекции осуществляли с интервалом, составлявшим двое суток (10 и 12 сутки от трансплантации опухоли), и за ростом опухоли следили до достижения конечной точки. Медиана измерения объема опухоли в последние сутки идентифицирует пороговый уровень терапевтического успеха, представленный пунктирной линией. Мышей, которые в момент конечной точки демонстрировали объем опухоли ниже порогового уровня, считали отвечающими на обработку, тогда как мышей, демонстрирующих объем опухоли выше порогового уровня, считали не отвечающими на обработку.
За опухолевым ростом следили при помощи цифрового штангенциркуля, измеряющего два измерения опухоли. Затем объем математически рассчитывали в соответствии со следующей формулой:
((длинная сторона) × (короткая сторона)2)/2
Во всех экспериментах опухоли измеряли в каждые вторые или третьи сутки до тех пор, пока размер опухоли не достигал допустимого максимума, и мышей затем умерщвляли и отбирали селезенки.
Анализ ELISpot
Для оценки количества активных антигенспецифических T-клеток секрецию интерферона-γ (IFN-γ) измеряли при помощи анализа ELISPOT от IMMUNOSPOT (CTL, Ohio USA) для мышиного IFN-γ и MABTECH (Mabtech AB, Nacka Strand, Sweden) для человеческого IFN-γ.
Использовали свежие спленоциты мышей, собранные в момент конечной точки эксперимента. Этот способ осуществляли в соответствии с указаниями производителя. Кратко, для мышиного IFN-γ 3*105 спленоцитов/лунку высевали в планшет в 0 сутки. Клетки стимулировали 2 мкг пептидов/лунку. После 3 суток инкубации при 37°C/5% CO2 планшеты проявляли в соответствии с протоколом к набору.
Для ELISPOT человеческого IFN-γ человеческие PBMC размораживали и оставляли на ночь при 37°C/5% CO2 в полной среде. В следующие сутки высевали 3x105 PBMC/лунку и стимулировали 2 мкг/лунку пептидов. После 48 ч инкубации при 37°C/5% CO2 планшеты проявляли в соответствии с протоколом производителя. Для анализа планшеты отправляли в CTL-Europe GmbH.
Клеточная линия и реактивы
Трансформированную EBV человеческую лимфобластоидную линию B-клеток JY (коллекция ECACC HLA-типа, Sigma Aldrich) выращивали в RPMI 1640 (GIBCO, Invitrogen, Carlsbad, CA, USA), дополненной 1% GlutaMAX (GIBCO, Invitrogen, Carlsbad, CA, USA) и 10% инактивированной нагреванием фетальной бычьей сывороткой (HI-FBS, GIBCO, Invitrogen, Carlsbad, CA, USA).
Стрептавидин (Streptomyces avidinii, аффинно-очищенный, лиофилизированный из 10 мМ фосфата калия, не менее 13 Е/мг белка) был приобретен в Sigma-Aldrich (Saint Louis, Missouri, USA).
Конъюгированный с биотином клон против HLA-A, B, C w6/32 был приобретен в Biolegend (San Diego, CA, USA) для анализа.
Следующие пептиды, приобретенные в Ontores Biotechnologies Co., Ltd., использовали в исследовании: KVLEYVIKV (SEQ ID NO: 75; название гена MAGE A1), ILDKKVEKV (SEQ ID NO: 76; название гена HSP90), и QLVDIIEKV (SEQ ID NO: 77; название гена PSME3).
Кроме того, следующие пептиды приобретали в Chempeptide (Shangai, China): VIMDALKSSY (SEQ ID NO: 78; название гена NNMT), FLAEGGGVR (SEQ ID NO: 79; название гена FGA) и EVAQPGPSNR (SEQ ID NO: 80; название гена HSPG2).
Биопсия опухоли яичников и этические соображения
Биопсию опухоли яичников отбирали у пациентки, страдающей от метастатической опухоли яичников (высокой степени серозности), которая заполнила информированное согласие, в процессе исследований, одобренных Этическим комитетом по исследованиям Госпитального района Северного Саво под номером подтверждения 350/2020. Образцы нарезали на небольшие кусочки и обрабатывали буфером для разрушения, содержащим 1 мг/мл коллагеназы типа D (Roche), 100 мкг/мл гиалуронидазы (Sigma Aldrich) и 1 мг/мл ДНКазы I (Roche) в течение 1 ч при 37°C. Клеточную суспензию затем пропускали через 500 мкм и 300 мкм клеточное сито (pluriSelect) для получения единичных клеток.
Образцы почечно-клеточной карциномы и опухоли мочевого пузыря и этические соображения
Образцы ткани пациента для органоидных культур получали в исследовании DEDUCER (Разработка диагностики и лечения урологических видов рака) Центрального госпиталя Хельсинкского Университета под номером подтверждения HUS/71/2017, 26.04.2017, номером подтверждения этического комитета 15.03.2017 Dnro 154/13/03/02/2016, и с согласия пациента. Образец почки получали в результате нефрэктомии взрослого мужчины, страдающего от светлоклеточной почечно-клеточной карциномы (ccRCC, pTNM стадия pT3a G2). Для экспериментов использовали образец доброкачественной опухолевой ткани. Уротелиальную карциному (рак мочевого пузыря, высокая степень, стадия III, 1x1 см) получали у взрослой женщины и образец раковой ткани использовали для органоидной культуры.
Органоидная культура светлоклеточной почечно-клеточной карциномы и опухоли мочевого пузыря
Клетки выделяли из исходной ткани непосредственно после хирургической операции путем диссоциации ткани на небольшие кусочки и их обработки коллагеназой (40 единиц/мл) в течение 2-4 ч. Доброкачественные и раковые клетки почки пациентов, страдающих от светлоклеточной почечно-клеточной карциномы, выращивали в виде органоидов в среде F [3:1 (об./об.) питательной смеси F‐12 (Ham) - DMEM (среда Игла, модифицированная Дульбекко) (Invitrogen), 5% FBS, 8,4 нг/мл холерного токсина (Sigma), 0,4 мкг/мл гидрокортизона (Sigma), 10 нг/мл эпидермального фактора роста (Corning), 24 мкг/мл аденина (Sigma), 5 мкг/мл инсулина (Sigma), 10 мкМ ингибитора ROCK (Y‐27632, Enzo Life Sciences, Lausen, Switzerland) и 1% пенициллина-стрептомицина с 10% матригелем (Corning). Органоиды, полученные из опухоли мочевого пузыря, выращивали в кальциевой среде для гепатоцитов (Corning)[15A], дополненной 5% CSFBS (очищенная древесным углем фетальная бычья сыворотка) (Thermo Fisher Scientific), 10 мкМ Y-27632 ингибитор RHO (Sigma), 10 нг/мл эпидермальным фактором роста (Corning), 1% GlutaMAX (Gibco), 1% пенициллином-стрептомицином и 10% матригелем. 6x106 Клеток собирали путем центрифугирования, промывали в PBS для удаления матригеля и быстро замораживали перед анализами.
Типирование HLA
Клиническое типирование HLA опухолевых образцов (ccRCC и мочевого пузыря) осуществляли в аккредитованной Европейской Федерацией Иммуногенетики (EFI) лаборатории HLA в Службе крови Красного Креста Финляндии. Аллельное определение трех классических генов HLA-I HLA-A, -B и -C осуществляли посредством методики основанного на таргетной PCR (полимеразная цепная реакция) секвенирования следующего поколения (NGS) в соответствии с протоколом, предложенным производителем (NGSgo® Workflow, GenDx, Utrecht, The Netherlands).
Указание аллелей на 4-польном уровне разрешения осуществляли при помощи NGSengine Version: 2,11,0,11444 (GenDx, Utrecht, The Netherlands) с использованием базы данных IPD IMGT/HLA версии 3.33.0, https://www.ebi.ac.uk/ipd/imgt/hla/.
Анализ посредством проточной цитометрии
Следующие антитела использовали для анализа экспрессии HLA-A2 и HLA-A, B и C на клеточной поверхности: конъюгированные с PE антитела против HLA-A2 (клон BB7.2, BioLegend 343306 San Diego, CA, USA), конъюгированные с PE антитела против человеческого HLA-A, B и C (клон W6/32, BioLegend 311406, San Diego, CA, USA), и Human TruStain FcX block (BioLegend B247182, San Diego, CA, USA).
Данные получали с использованием проточного цитометра BDLSR Fortessa. Анализ почечно-клеточной карциномы и органоидов опухоли мочевого пузыря посредством проточной цитометрии осуществляли с использованием BD Accuri 6 plus (BD Biosciences) и анализировали при помощи программного обеспечения FlowJo (Tree Star, Ashland, OR, USA).
Результаты
Разработка инструмента оценки гомологии ксенопептидов (HEX) для идентификации вирусных и опухолевых пептидов с высокой молекулярной мимикрией
Чтобы исследовать то, может ли молекулярная мимикрия между вирусами и опухолью повлиять на рост опухоли, авторы изобретения должны были идентифицировать пептиды, обладающие высокой степенью гомологии/аффинности. Тем не менее, отсутствует инструмент, облегчающий идентификацию соответствующей мишени в этой области. Таким образом, авторы изобретения разработали HEX (оценка гомологии ксенопептидов), представляющее собой устройство, которое сравнивает входные последовательности с последовательностями из базы данных антигенов/пептидов патогенов или белковыми последовательностями, и выбирает в высокой степени гомологичные пары-кандидаты пептидов на основе по меньшей мере двух из трех следующих критериев: 1) B-оценка, которая соответствует вероятности пептидов, распознаваемых данным TCR; 2) позиционно взвешенная оценка выравнивания для приоритизации в отношении сходства в области взаимодействия с TCR; 3) предсказание аффинности связывания МНС класса I (фиг. 5А). Альтернативно, может быть использовано обычное программное обеспечение, как представлено на фиг. 5B и как описано выше.
Авторы изобретения начали с трех ассоциированных с меланомой антигенов, которые были успешно применены в ряде исследований вакцинации: TRP2180-188 (белок 2, родственный тирозиназе), GP10025-33 (он же PMEL; белок премеланосомы) и TYR1208-216 (тирозиназа 1). Не прогнозируется, что TYR1208-216 связывается с мышиным МНС, и, таким образом, она рассматривается как “ненадлежащая мишень”. При помощи HEX авторы изобретения идентифицировали вирусные пептиды, которые обладают высокой степенью гомологии/аффинности с входными опухолевыми эпитопами. Пулы из 4 вирусных пептидов на каждый исходный опухолевый эпитоп (таблица 1) были выбраны для дополнительной оценки in vivo.
Противовирусный иммунитет контролирует опухолевый рост посредством молекулярной мимикрии с опухолевыми антигенами
Чтобы оценить, будут ли вирусные пептиды влиять на опухолевый рост, авторы изобретения решили имитировать вирусную инфекцию путем иммунизации мышей C57BL6 выбранными пулами вирусных пептидов с последующей трансплантацией опухоли (фиг. 6А). У всех иммунизированных мышей наблюдали уменьшение опухолевого роста по сравнению с модельной группой. Кроме того, наблюдали значительные различия между обрабатываемыми группами. Опухолевый рост был наиболее уменьшен у мышей, иммунизированных пулом вирусных пептидов, гомологичных TRP2 или gp100 (фиг. 6В).
Чтобы дополнительно исследовать вклад выбранных вирусных пептидов в уменьшение опухолевого роста, авторы изобретения собрали спленоциты мышей в конечной точке для анализа ELISpot (фиг. 6C). Спленоциты мышей, иммунизированных пулами вирусных пептидов, гомологичных TRP2 или gp100, способствовали более высокому ответу IFN-γ по сравнению со спленоцитами мышей, иммунизированных контрольными эпитопами, гомологичными TYR1, в корреляции с исходом опухолевого роста.
Авторы изобретения дополнительно исследовали реакционноспособность этих спленоцитов предварительно иммунизированных вирусом мышей в отношении их соответствующего когнатного опухолевого антигена (фиг. 7А). При попарном сравнении только вирусные пептиды, гомологичные TRP2, продемонстрировали более высокую секрецию IFN-γ по сравнению с когнатным опухолевым пептидом (фиг. 7В). Кроме того, авторы изобретения ретроспективно оценили аффинности каждого опухолевого антигена и их гомологичные вирусные пулы к молекулам MHC класса I C57BL/6J (фиг. 7C), и обнаружили значительную корреляцию с их соответствующей способностью стимулировать секрецию IFN-γ (фиг. 7D). В соответствии с рекомендациями Базы данных иммунных эпитопов (IEDB) авторы изобретения разделили пептиды на пептиды с высокой и низкой аффинностью с использованием порогового уровня 50 нМ (http://tools.iedb.org/mhci/help/), и обнаружили, что пептиды с высокой аффинностью MHC-I способствовали продукции большего количества INF-γ по сравнению с пептидами с низкой аффинностью MHC-I (фиг. 7E). Взятые вместе эти результаты подтверждают прогнозирующую эффективность инструмента HEX, чтобы идентифицировать гомологичные пептиды и подчеркнуть, что молекулярная мимикрия между вирусами и опухолью играет роль в противоопухолевом иммунном ответе посредством перекрестно-реактивных T-клеток.
Вирусные эпитопы, обладающие высоким сходством с опухолевыми эпитопами, уменьшают рост сформировавшейся меланомы in vivo
Авторы изобретения продемонстрировали, что молекулярная мимикрия между вирусными и опухолевыми антигенами может влиять на опухолевый рост у предварительно иммунизированных мышей. Затем авторы изобретения хотели оценить, мог ли направленный молекулярной мимикрией ответ оказать влияние на уже сформировавшиеся опухоли у мышей, ранее не подвергавшихся воздействию. Для этого мышам имплантировали опухоли B16OVA или более агрессивные и иммуносупрессивные опухоли B16F10 и затем обрабатывали ранее разработанной вакцинной платформой PeptiCRAd (20) для имитации вирусной инфекции (фиг. 8А). Кратко, PeptiCRAd представляет собой вакцинную технологию, состоящую из аденовирусов, покрытых ограниченными MHC-I пептидами, несущими положительно заряженные полиаминокислотные хвостовые группировки. При этом авторы изобретения продолжили использовать наиболее эффективный пул пептидов из первого эксперимента in vivo. Вакцины готовили путем покрывания аденовирусов исходным эпитопом TRP2180-188 (TRP2-PeptiCRAd) или соответствующим пулом вирусных пептидов, похожих на TRP2 (вирусный PeptiCRAd). Внутриопухолевые инъекции PeptiCRAd значительно уменьшали развитие опухоли по сравнению с обработкой физиологическим буферным раствором или непокрытым вирусом в отношении как опухоли B16OVA (фиг. 8B-C), так и опухоли B16F10 (фиг. 8E-F). Мыши, обработанные исходным опухолевым антигеном или вирусными гомологами, демонстрировали значительно большее количество демонстрирующих ответ в обеих опухолевых моделях (фиг. 8D, 8G). В обеих опухолевых моделях авторы изобретения обнаружили значительное уменьшение опухолевого роста, когда мышей обрабатывали вирусными пептидами, похожими на опухолевые пептиды, предполагая еще раз, что молекулярная мимикрия может играть фундаментальную роль в противоопухолевом иммунитете, затрагивающем перекрестно-реактивные T-клетки.
Исследование того, может ли молекулярная мимикрия между CMV и опухолевыми антигенами объяснить более хороший прогноз
Авторы изобретения выбрали пул ассоциированных с меланомой белков [21], которые сравнивали с протеомом CMV с использованием HEX, генерируя перечень опухолевых пептидов с высоким сходством с CMV (таблица 2). Опухолевые пептиды и их аналог CMV использовали в анализе ELISPOT, показывая, что у пациента, демонстрирующего ответ, PBMC всегда реагировали как на вирусные, так и на аналогичные им опухолевые антигены, свидетельствуя о том, что CMV-инфекция увеличивала вирусные T-клеточные клоны, которые могут атаковать и уничтожать опухолевые клетки, делая сероположительных пациентов более склонными к реакции на специфические для меланомы эпитопы, похожие на CMV (фиг. 9A, B). Хотя это наблюдение было заманчивым, оно имело ограничение, которое непосредственно не показывало, увеличивали ли вирусные и опухолевые пептиды те же самые T-клеточные клоны.
Новая микрофлюидная платформа на основе чипа идентифицирует иммунопептидомный профиль в скудной ткани опухолевой биопсии
Авторы изобретения критически оценивали платформу для исследования скудной опухолевой биопсии. Таким образом, у пациента была отобрана метастатическая опухоль яичника (высокая степень серозности) и четыре кусочка были получены из границы опухоли (S1, S2, S3 и S4); также отбирали центральную часть опухоли (S5). Затем образцы взвешивали, и, как обобщено на фиг. 10А, средний размер составлял от 0,01 г до 0,06 г. После разрушения образца полученные суспензии одиночных клеток лизировали и пропускали через микрочип. При применении строгой границы уровня ложноположительных результатов 1% для идентификации белка и пептида 916, 695, 172, 1128 и 256 уникальных пептидов были идентифицированы, соответственно, в S1, S2, S3, S4 и S5 (фиг. 10A). В соответствии с типичным лигандомным профилем, общее обогащение (выше 70%) наблюдали в образцах 7-13-меров (фиг. 10А). В отношении абсолютного количества и процентной доли распределение аминокислотной длины продемонстрировало, что образцы 9-меров были наиболее репрезентативными (фиг. 10B), что подтверждает анализ авторов изобретения и другой предшествующий иммунопептидомный анализ. Затем авторы изобретения дополнительно исследовали исходные белки, обнаруженные в данных авторов изобретения, с применением анализа обогащения генной онтологии (GO). В соответствии с типичным лигандомным профилем, метаболические процессы были обогащены во всех исследованных образцах. Кроме того, анализ выявил увеличение белков пути развития кожи в соответствии с эпителиальной природой проанализированной здесь серозной опухоли яичников. В целом, эти результаты подчеркнули возможность использования разработанной микрофлюидной чиповой платформы для анализа скудной биопсии опухоли и получения персонализированного антигенного пептида для лечения опухоли.
Основанный на микрочипе протокол выявляет иммунопептидомный ландшафт в полученных у пациента органоидах.
Микрочиповую технологию критически оценивали с использованием всего 6х106 клеток из органоидов, полученных от пациентов (PDO). Авторы изобретения выбрали двух пациентов из продолжающегося исследования прецизионной медицины для урологических форма рака, образца нефрэктомии, содержащего как доброкачественную, так раковую ткань пациента, страдающего от светлоклеточной почечно-клеточной карциномы (ccRCC), и образца 1x1 см пациента, страдающего от рака мочевого пузыря, дополнительно обрабатывали как 3D первичные органоидные культуры.
Применяя разработанную микрочиповую технологию и строгий уровень ложноположительных результатов 1% для идентификации пептида и белка авторы изобретения смогли идентифицировать в общей сложности 576 и 2089 уникальных пептидов в PDO ccRCC и мочевого пузыря, соответственно (фиг. 11A). Количество извлеченных пептидов в двух образцах различалось, при этом образцы мочевого пузыря приводили к большему количеству пептидов, чем образцы ccRCC. Хорошо известно, что экспрессия HLA влияет на количество выделенных пептидов HLA-I [22A], и в соответствии с этим анализ посредством проточной цитометрии выявил более высокие поверхностные уровни HLA-A, HLA-B и HLA-C в мочевом пузыре, чем в образцах ccRCC, объясняя различные выходы полученных пептидов из образцов. Анализ пептидов продемонстрировал предпочтение 9-12-меров (56,4% в ccRCC и 47,9% в опухолях мочевого пузыря) с обогащением 9-мерной популяции, в соответствии с типичным распределением длины лигандомного анализа [33] (фиг. 11B). Чтобы различать вещества, связывающие HLA-I, и загрязнители, осуществляли кластеризацию Гиббса и анализ NetMHC4.0. Во-первых, деконволюция 9-меров продемонстрировала, что 55% и 67% в PDO мочевого пузыря и ccRCC, соответственно, соответствовали по меньшей мере одной из аллелей HLA пациентов. Затем NetMHC4.0 применяли в отношении всех 9-меров, идентифицированных в наборе данных авторов изобретения. Среди них 46% и 69% были предсказаны как связывающиеся со специфическими HLA пациентов. Затем были исследованы исходные белки, присутствующие в обоих наборах данных, полученных авторами изобретения. Для этого был проведен анализ обогащения генной онтологии (GO). В соответствии с предыдущими наблюдениями и опубликованными данными авторов изобретения, оба образца продемонстрировали обогащение внутриклеточными и ядерными белками, взаимодействующими с РНК и вовлеченными в катаболические/метаболические процессы.
Чтобы продемонстрировать, что технология может быть использована для быстрой разработки терапевтических противораковых вакцин, авторы изобретения разработали анализ уничтожения. Авторы изобретения сосредоточились на анализе образцов ccRCC. Авторы изобретения использовали транскриптомные уровни для отбора предполагаемых опухолевых антигенов с использованием PBMC и здоровой ткани почек в качестве референсных наборов. Затем на PBMC здоровых добровольцев воздействовали выбранными пептидами, и в анализе использовали CD8+ T-клетки, выделенные из этих клеток. T-клетки, на которые воздействовали пептидом EVAQPGPSNR (название гена HSPG2), продемонстрировали приблизительно 10% специфический цитолиз.
Наконец, авторы изобретения стремились исследовать T-клеточный ответ у пациента, страдающего от ccRCC. Для этого нефракционированные РВМС, полученные у пациента, стимулировали in vitro пептидом EVAQPGPSNR, тогда как нестимулированные РВМС были приняты в качестве контроля. Производные CD8 + T- клетки затем добавляли к ccRCC PDO, демонстрируя приблизительно 7% увеличение уничтожающей активности по сравнению с контрольной группой.
Продемонстрировано, что пептиды из органоидов пациентов обладают терапевтической эффективностью в исследованиях in vivo
Перечень пептидов анализировали посредством HEX. Во-первых, программное обеспечение приоритизировало пептиды, которые одновременно обладали сильными связывающими свойствами (диапазон отсечения IC50 50 нМ-500 нМ в соответствии с NetMHC4.0) и продемонстрировали более высокую взвешенную оценку выравнивания (отсечение 0,8-1 нормализованной взвешенной оценки выравнивания). Последнее фокусирует сходство пептида в области взаимодействия, который наиболее вероятно будет взаимодействовать с TCR в CD8 + T-клетках для того, чтобы вызвать опосредованный иммунный ответ; получающиеся в результате пептиды затем дополнительно распределяли по категориям на основе их полного процента идентичности с антигенами/пептидами патогенов и оценкой IC50 аффинности связывания. Конечный выход состоял из тринадцати пептидов с аналогичными им патогенными пептидами (таблица 4).
Для определения иммуногенности пептида мышей предварительно иммунизировали подкожными инъекциями каждого одиночного пептида в присутствии адъюванта поли(I:C); группе мышей инъецировали только поли(I:C) или также физиологический раствор в качестве контроля. Спленоциты этих мышей собирали и тестировали в отношении продукции IFNγ при специфической стимуляции (таблица 5) в анализе ELISpot.
Авторы изобретения затем попытались проверить, можно ли использовать пептиды-кандидаты в качестве противораковой вакцины для лечения сформировавшихся опухолей. Для этого авторы изобретения использовали PeptiCRAd, представляющую собой ранее разработанную авторами изобретения платформу противораковых вакцин, которая комбинирует онколитические аденовирусы с присоединенными к капсиду (через полилизиновый линкер) модифицированными полилизином пептидами. Использованный здесь аденовирус представляет собой VALO-mD901, то есть генетически модифицированный для экспрессии мышиных OX40L и CD40L, и ранее показано, что он вызывает контроль опухолевого роста и системный противоопухолевый ответ в мышиной модели меланомы. Таким образом, мышам Balb/c подкожно инъецировали сингенную модель опухоли CT26 в левый и правый бок (0 сутки, фиг. 12А). Когда опухоли формировались (7 сутки, Фиг. 12А), Valo-mD901 покрывали парой каждого модифицированного полилизином пептида из перечня авторов изобретения (PeptiCRAd1, PeptiCRAd2, PeptiCRAd3, Таблица 6), и осуществляли внутриопухолевую инъекцию только в правую опухоль. PeptiCRAd4 состоял из Valo-mD901, покрытых gp70423-431 (AH1-5). В качестве контроля также использовали модельную группу и группу Valo-mD901. PeptiCRAd1 и PeptiCRAd2 улучшали контроль опухолевого роста, а также Valo-mD901 в поражениях, подвергнутых инъекции (фиг. 12B, правый блок). Благоприятно, PeptiCRAd1 улучшал противоопухолевый контроль роста в необработанной опухоли в отличие от Valo-mD901, который не вызывал эффекта.
Заключение
HEX представляет собой биоинформатический инструмент, который анализирует пептидные последовательности и сравнивает их с проверенной базой данных вирусных патогенных протеомов в поисках последовательностей с высоким сходством.
Поскольку представленные пептиды в основном участвуют во взаимодействии с якорными остатками в МНС [25], разработанное авторами изобретения программное обеспечение возвращает оценку выравнивания, которая является позиционно взвешенной для того, приоритизировать сходство, возникающее в центральной части пептида, то есть положение, которое больше всего вовлечено во взаимодействие с TCR [26]. Кроме того, чтобы увеличить шансы на то, что предложенная авторами изобретения мишень является представленным эпитопом, авторы изобретения ранжировали полученные пептиды в соответствии с аффинностью связывания для выбранных MHC с использованием в одном из примеров инструмента прогнозирования IEDB NetMHC API [27]; или NetMHC4.0 или NetMHCpan4.1b в качестве самостоятельных инструментов.
Авторы изобретения использовали HEX для идентификации вирусных пептидов, гомологичных трем широко охарактеризованным ассоциированным с опухолями антигенам.
Авторы изобретения обнаружили, что предварительная иммунизация отобранными авторами изобретения пулами вирусных пептидов имитировала противовирусный иммунный статус до формирования опухоли и эффективно замедляла рост подкожно инъецированных опухолевых клеток меланомы у мышей, свидетельствуя, что предварительное воздействие вирусными пептидами может влиять на опухолевый рост.
Затем авторы изобретения обнаружили, что гомологичные опухоли вирусные пептиды оказывают аналогичный эффект на уже сформировавшиеся опухоли при введении во время прогрессирования опухоли, что указывает на то, что вирусная инфекция, происходящая во время прогрессирования опухоли, все еще может влиять на ее рост.
Вместе полученные авторами изобретения результаты предполагают, что молекулярная мимикрия между вирусными и опухолевыми антигенами способна осуществлять контроль в отношении опухолевого роста, даже не полагаясь на предварительное воздействие или предварительно приобретенный иммунитет.
Новые ICPI значительно улучшали выживание пациентов при нескольких солидных опухолях, особенно при метастатической меланоме, по сравнению с другими обычно используемыми способами терапии, такими как лучевая терапия и химиотерапия. Тем не менее, несмотря на улучшение выживаемости и эффективные уровни ответа, все еще не известно, почему некоторые пациенты не получали пользы от способов терапии ICPI. Полученные авторами изобретения результаты показали, что пациенты с высоким титром CMV-специфических уровней IgG имели значительно более длительную выживаемость без прогрессирования заболевания.
Авторы изобретения обнаружили, что РВМС из пациентов с высоким титром IgG против CMV взаимодействовали с антигенами меланомы, похожими на пептиды CMV. Эти данные указывают на то, что CMV-серопозитивность способствует специфическому для меланомы Т-клеточному иммунитету и, таким образом, обеспечивает клиническое преимущество для этих пациентов, проходящих терапию ICPI.
Здесь авторы изобретения продемонстрировали, что вирусные инфекции могут оказывать влияние на опухолевый рост и клиренс. Кроме того, авторы изобретения продемонстрировали перекрестную реактивность цитотоксических Т-клеток против вирусных и гомологичных опухолевых антигенов, отобранных с использованием HEX. На основании полученных авторами изобретения результатов авторы изобретения пришли к выводу о том, что явление молекулярной мимикрии может быть использовано в будущем для разработки новых способов лечения или при сочетании со способами иммунотерапии потенциально усиливает противоопухолевый иммунный эффект, достигаемый этими способами лечения.
Кроме того, разработка эффективного устранения опухоли и стратегий защиты требует надежной идентификации опухолевых пептидов, связывающихся с HLA-I. Прямая идентификация пептидов из комплекса HLA-I все еще представляет наилучший способ. Тем не менее, процесс иммунопептидомики является относительно сложным и, таким образом, представляет собой основное узкое место в процессе обнаружения антигена. В настоящее время неспособность анализировать иммунопептидомы из небольшого количества биологических материалов (например, тканевой биопсии с помощью иглы), обработка образца, стоимость и матрица аффинности (которая является трудоемкой и дорогостоящей в производстве) принятая в обычной платформе, были изображены в качестве основных технических проблем, которые требуется решать.
В этой работе авторы изобретения рассмотрели несколько технических вопросов, затрудняющих исследование лигандома, уделяя основное внимание ограниченной доступности материала для анализа, стоимости расходных материалов и длительных протоколов.
Путем использования хорошо охарактеризованного взаимодействия биотин-стрептавидин для того, чтобы иммобилизовать биотинилированное антитело пан-HLA на функционализированных стрептавидином поверхностях, авторы изобретения смогли заменить обычную технологию, основанную на матрицах аффинностей, полученных при помощи реакций перекрестного связывания, на микрочиповую платформу. Ограничения, обусловленные скудностью материала (например, биопсия с помощью иглы), стимулировали работу по осуществлению микрофлюидного протокола. В этой работе авторы изобретения использовали пользовательский микрочиповый протокол, охватывающий микростолбиковую матрицу на основе тиоленового полимера в качестве твердого носителя для дополнительной биофункционализации, которая позволила осуществить всю процедуру IP на одном микрофлюидном чипе.
Проверка идентифицированных пептидов в анализе уничтожения in vitro подтвердила, что пептиды, идентифицированные при осуществлении изобретения, были на самом деле представлены на поверхности клеток JY, поскольку они были уничтожены специфическим для CD8 + T-клеток образом. Обнаруженный в полученном авторами изобретения наборе данных пептид ILDKKVEKV (SEQ ID NO: 3) вызвал более высокую процентную долю специфического цитолиза.
Таким образом, подход авторов изобретения предлагает неисследованный инструмент для иммуно-аффинной очистки.
ССЫЛКИ:
20. C. Capasso, M. Hirvinen, M. Garofalo, D. Romaniuk, L. Kuryk, T. Sarvela, A. Vitale, M. Antopolsky, A. Magarkar, T. Viitala, T. Suutari, A. Bunker, M. Yliperttula, A. Urtti, V. Cerullo, Oncolytic adenoviruses coated with MHC-I tumor epitopes increase the antitumor immunity and efficacy against melanoma, Oncoimmunology 5, e1105429 (2016).
21. D. Weinstein, J. Leininger, C. Hamby, B. Safai, Weinstein 2014 Diagnostic and Prognostic biomarkers in melanoma, J. Clin. Aesthet. Dermatol. 7, 13-24 (2014).
25. J. Sidney, E. Assarsson, C. Moore, S. Ngo, C. Pinilla, A. Sette, B. Peters, Quantitative peptide binding motifs for 19 human and mouse MHC class i molecules derived using positional scanning combinatorial peptide libraries, Immunome Res. 4, 2 (2008).
26. A. K. Sharma, J. J. Kuhns, S. Yan, R. H. Friedline, B. Long, R. Tisch, E. J. Collins, Class I Major Histocompatibility Complex Anchor Substitutions Alter the Conformation of T Cell Receptor Contacts, J. Biol. Chem. 276, 21443-21449 (2001).
27. M. Andreatta, M. Nielsen, Gapped sequence alignment using artificial neural networks: Application to the MHC class i system, Bioinformatics 32, 511-517 (2016).
28. P. C. Rosato, S. Wijeyesinghe, J. M. Stolley, C. E. Nelson, R. L. Davis, L. S. Manlove, C. A. Pennell, B. R. Blazar, C. C. Chen, M. A. Geller, V. Vezys, D. Masopust, Virus-specific memory T cells populate tumors and can be repurposed for tumor immunotherapy, Nat. Commun. 10, 567 (2019).
33. S. Giguère, A. Drouin, A. Lacoste, M. Marchand, J. Corbeil, F. Laviolette, MHC-NP: Predicting peptides naturally processed by the MHC, J. Immunol. Methods 400-401, 30-36 (2013).
3A. Bassani-Sternberg M, Pletscher-Frankild S, Jensen LJ, Mann M. Mass spectrometry of human leukocyte antigen class I peptidomes reveals strong effects of protein abundance and turnover on antigen presentation. Mol Cell Proteomics. 2015;14(3):658-73.
15A. Lee SH, Hu W, Matulay JT, Silva MV, Owczarek TB, Kim K, et al. Tumor Evolution and Drug Response in Patient-Derived Organoid Models of Bladder Cancer. Cell. 2018;173(2):515-28 e17.
22A. Purcell AW, Ramarathinam SH, Ternette N. Mass spectrometry-based identification of MHC-bound peptides for immunopeptidomics. Nat Protoc. 2019;14(6):1687-707.
24A. Jurtz V, Paul S, Andreatta M, Marcatili P, Peters B, Nielsen M. NetMHCpan-4.0: Improved Peptide-MHC Class I Interaction Predictions Integrating Eluted Ligand and Peptide Binding Affinity Data. J Immunol. 2017;199(9):3360-8.
25A. Nielsen M, Andreatta M. NetMHCpan-3.0; improved prediction of binding to MHC class I molecules integrating information from multiple receptor and peptide length datasets. Genome Med. 2016;8(1):33.
26A. Hoof I, Peters B, Sidney J, Pedersen LE, Sette A, Lund O, et al. NetMHCpan, a method for MHC class I binding prediction beyond humans. Immunogenetics. 2009;61(1):1-13.


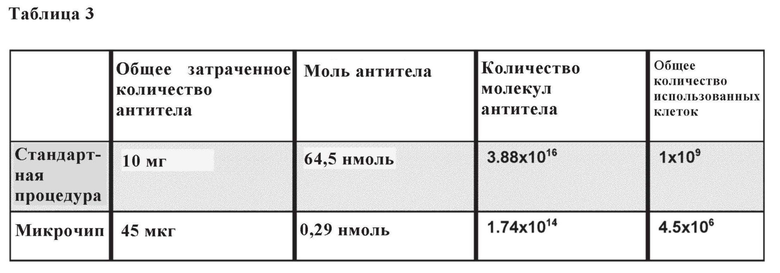

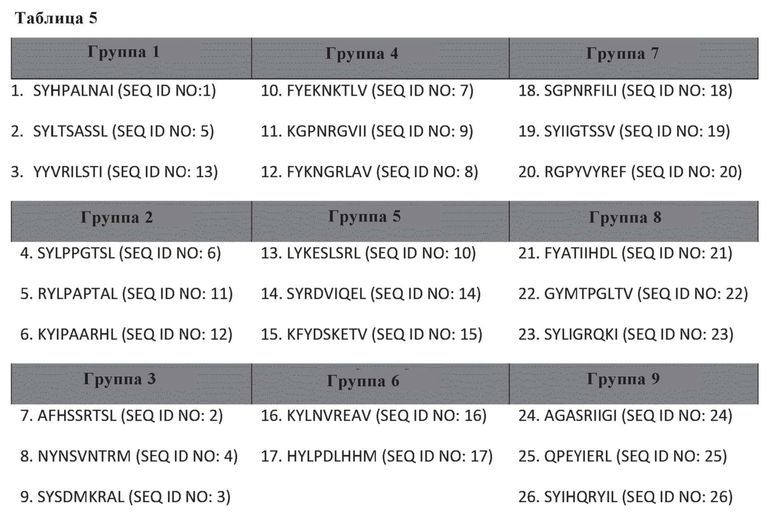
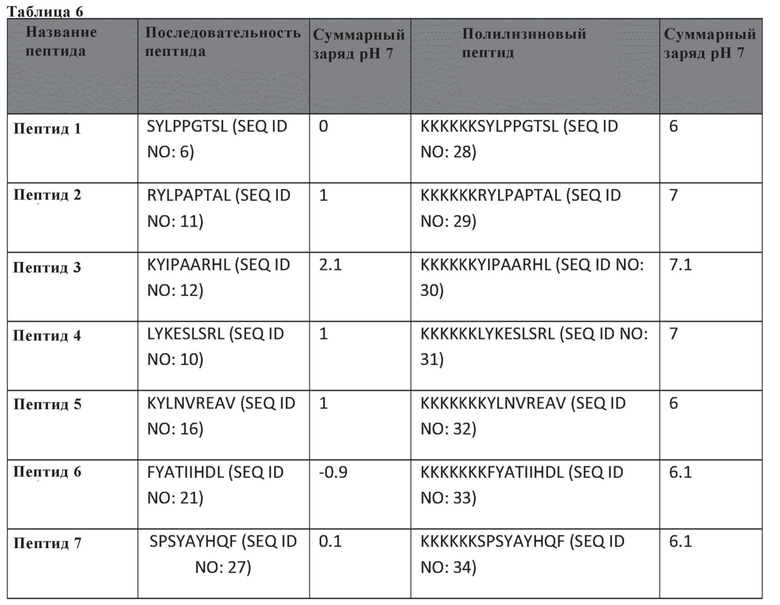
| название | год | авторы | номер документа |
|---|---|---|---|
| Покрытые онколитические аденовирусы для противораковых вакцин | 2015 |
|
RU2695375C2 |
| БЕЛКИ, ОТНОСЯЩИЕСЯ К СЕМЕЙСТВУ Всl-2, И ИХ ФРАГМЕНТЫ И ИХ ПРИМЕНЕНИЕ У ПАЦИЕНТОВ СО ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛЬЮ | 2004 |
|
RU2367468C2 |
| АНТИГЕН-СВЯЗЫВАЮЩИЙ БЕЛОК, РАСПОЗНАЮЩИЙ ПРОИСХОДЯЩИЙ ИЗ MAGE-A4 ПЕПТИД | 2018 |
|
RU2777074C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ПРОИЗВОДСТВА Т-КЛЕТОК | 2018 |
|
RU2793344C2 |
| МНС-ДИСПЛЕЙ ПЕПТИДОВ МЛЕКОПИТАЮЩЕГО КАК ИНСТРУМЕНТ ДЛЯ ВЫБОРА АНТИГЕННОЙ ДЕТЕРМИНАНТЫ ПРИ СОЗДАНИИ ВАКЦИН | 2019 |
|
RU2795983C2 |
| СПОСОБ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2770447C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ГАММА ДЕЛЬТА Т-КЛЕТКИ | 2015 |
|
RU2756247C2 |
| ДОСТАВКА БИОМОЛЕКУЛ В КЛЕТКИ ИММУННОЙ СИСТЕМЫ | 2015 |
|
RU2739794C2 |
| Т-КЛЕТКИ С ТРАНСДУЦИРОВАННЫМ В НИХ АНТИГЕНОМ, ПРИМЕНЯЕМЫЕ В КАЧЕСТВЕ СИСТЕМЫ ДОСТАВКИ АНТИГЕНОВ | 2003 |
|
RU2330884C2 |
| МОДИФИЦИРОВАННЫЕ ОНКОЛИТИЧЕСКИЕ АДЕНОВИРУСЫ | 2019 |
|
RU2788638C2 |

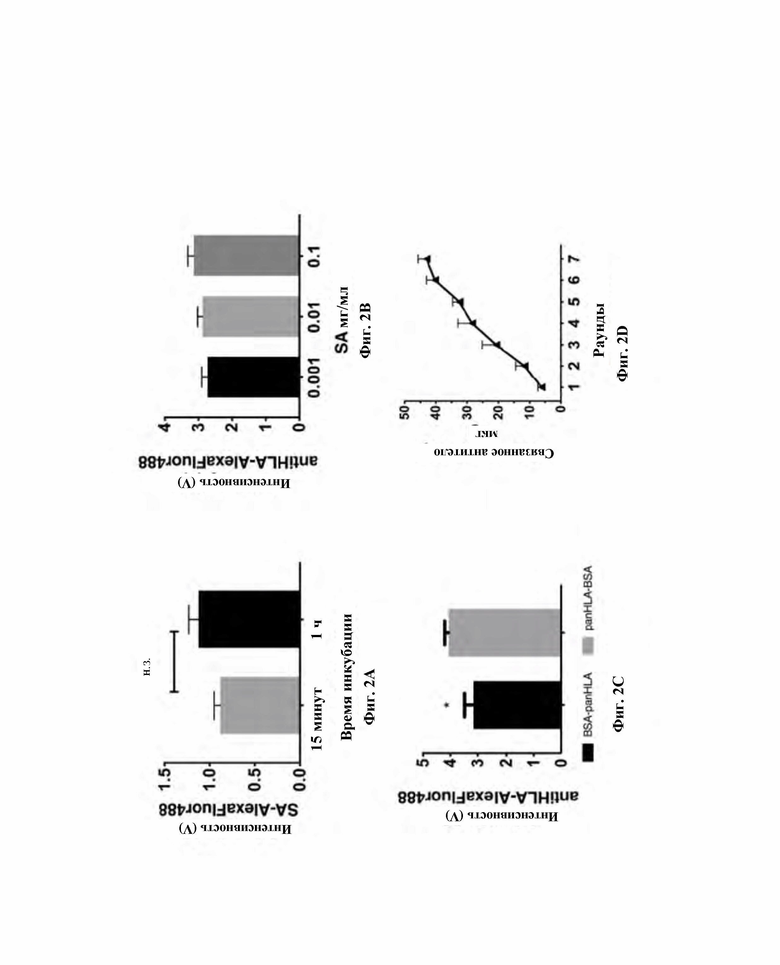
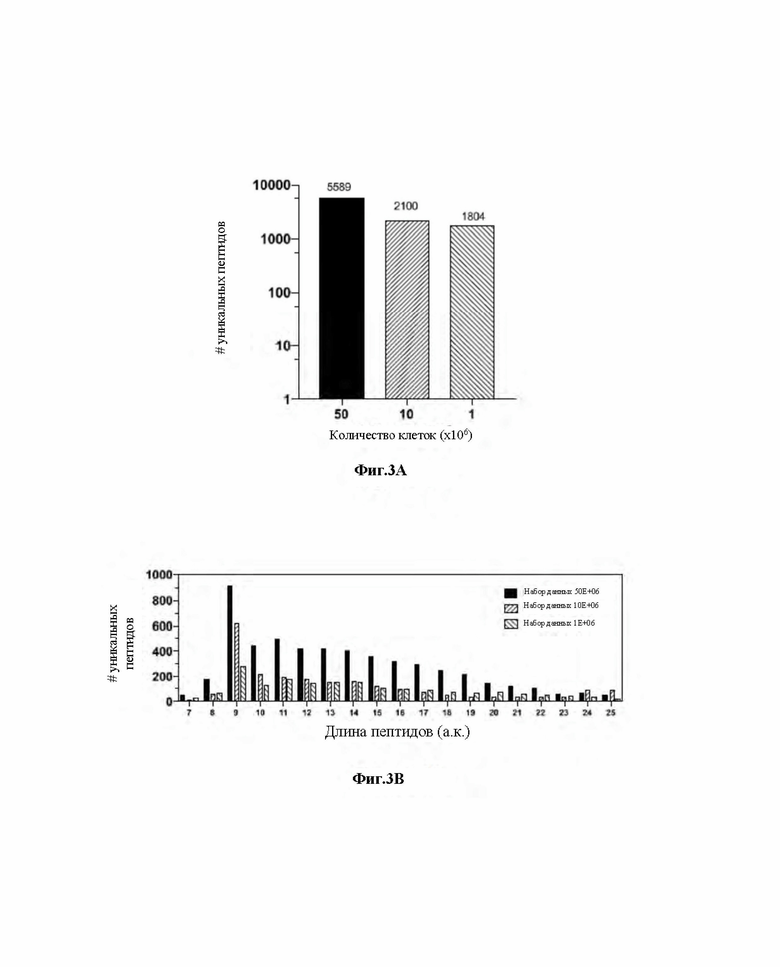
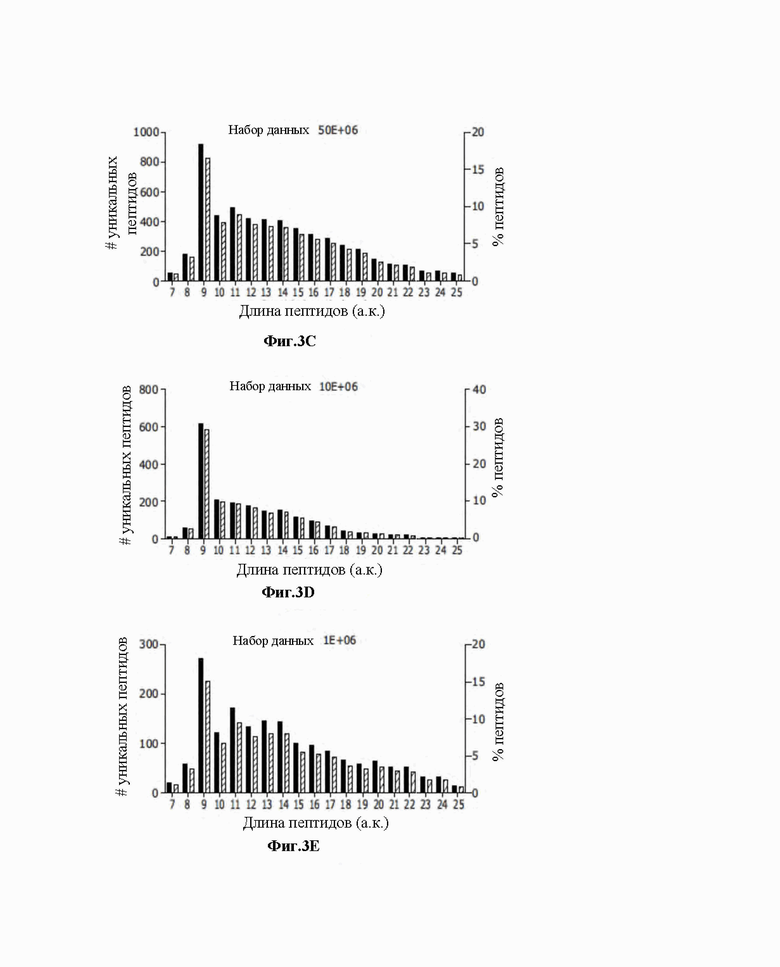
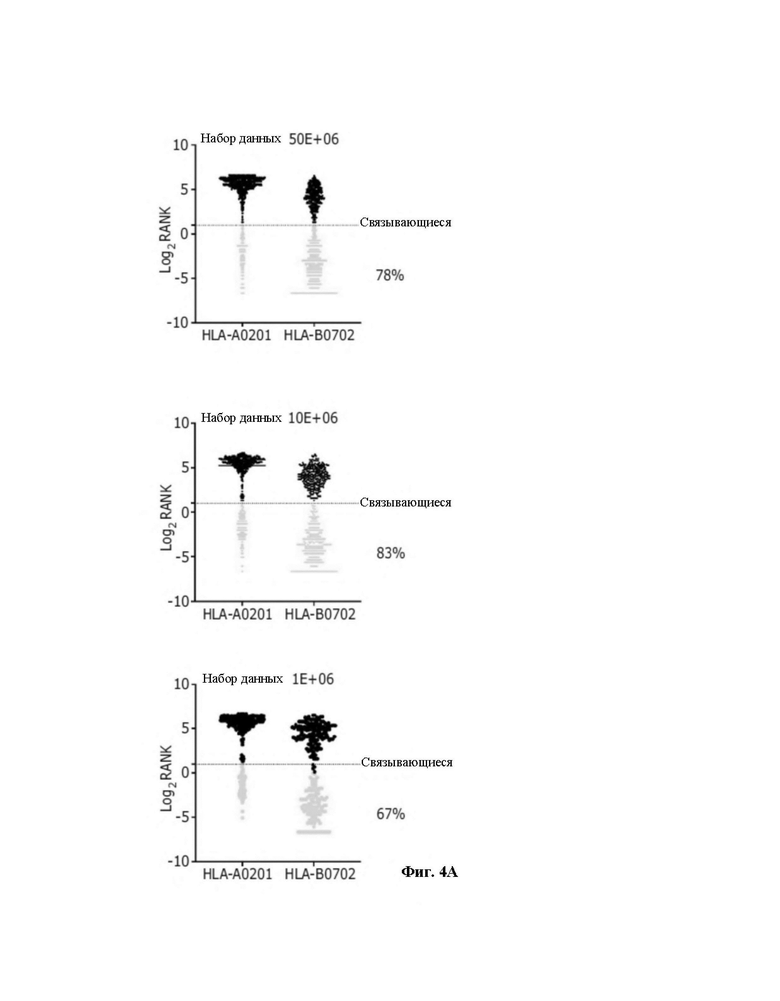
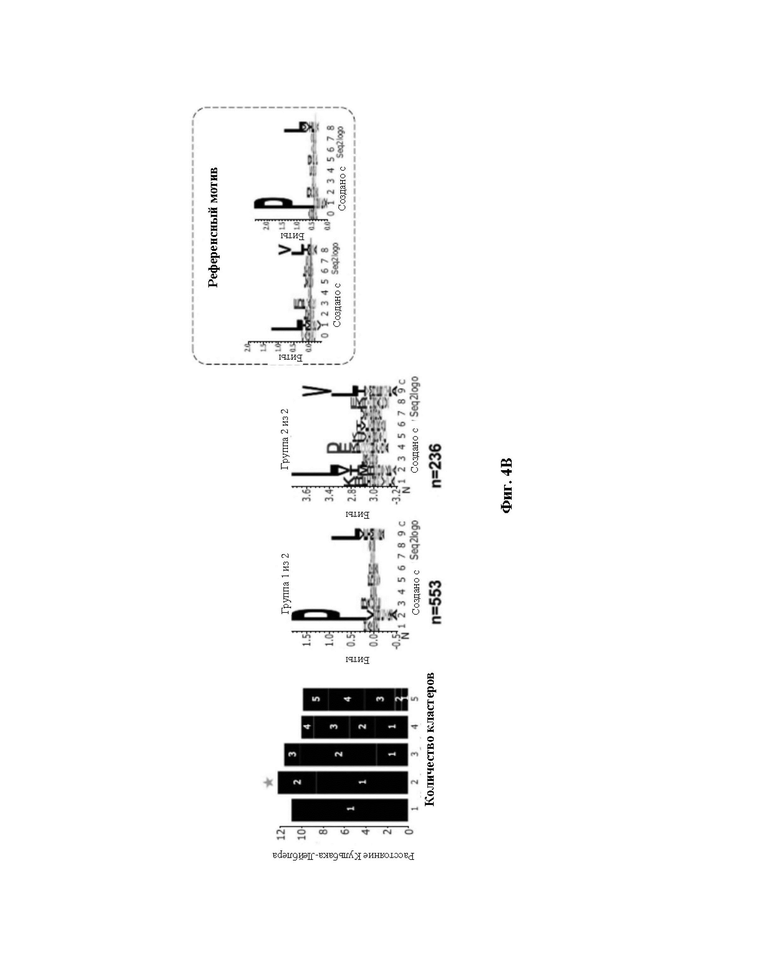
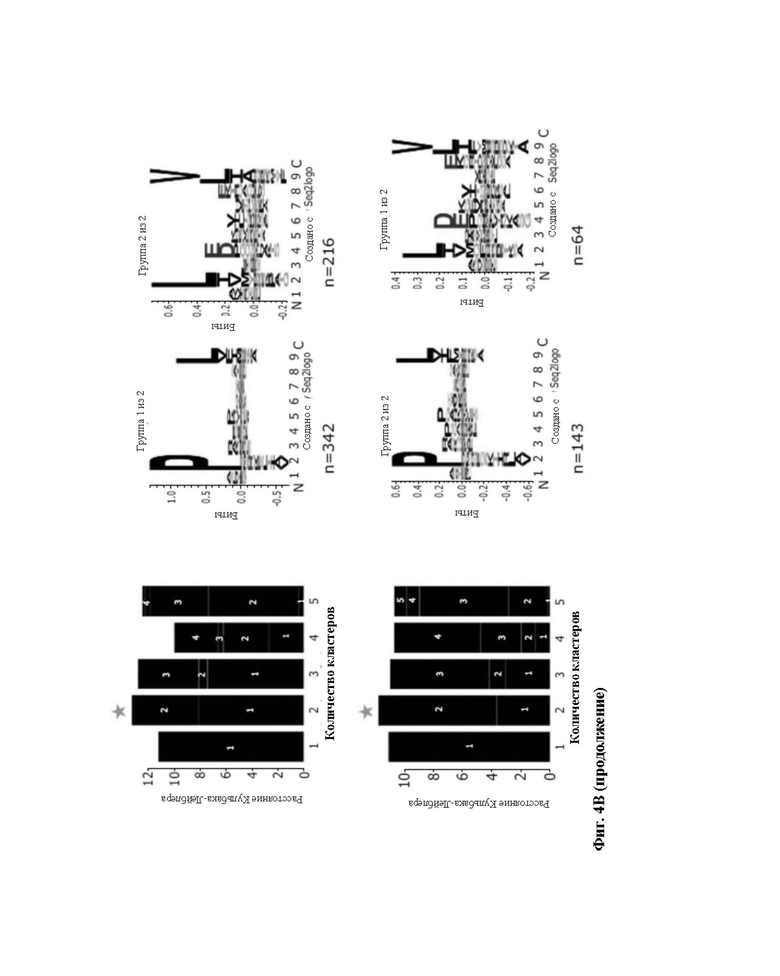
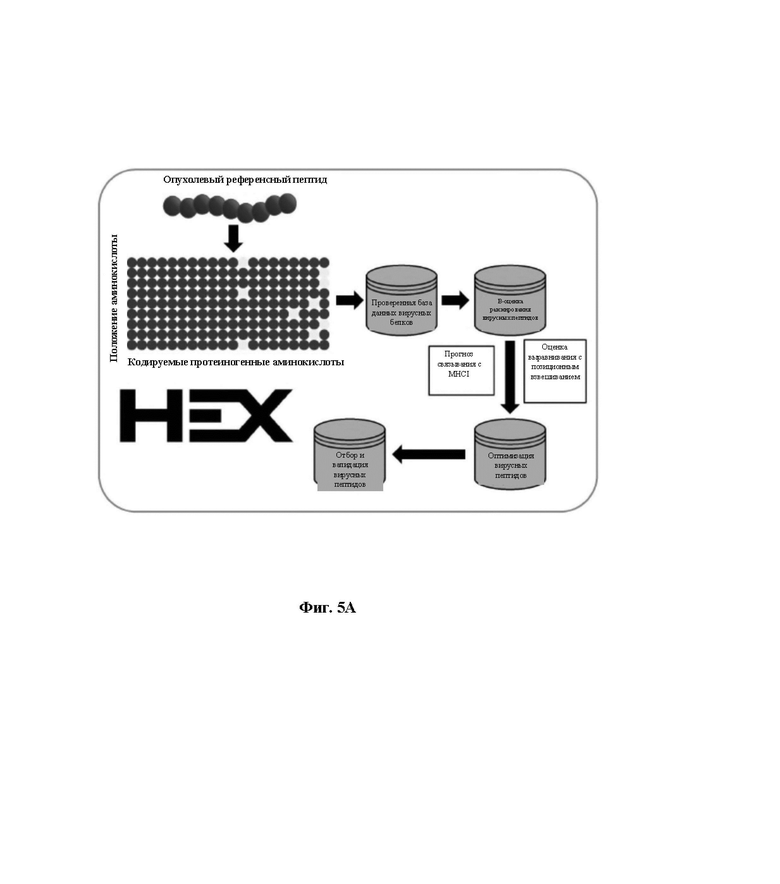
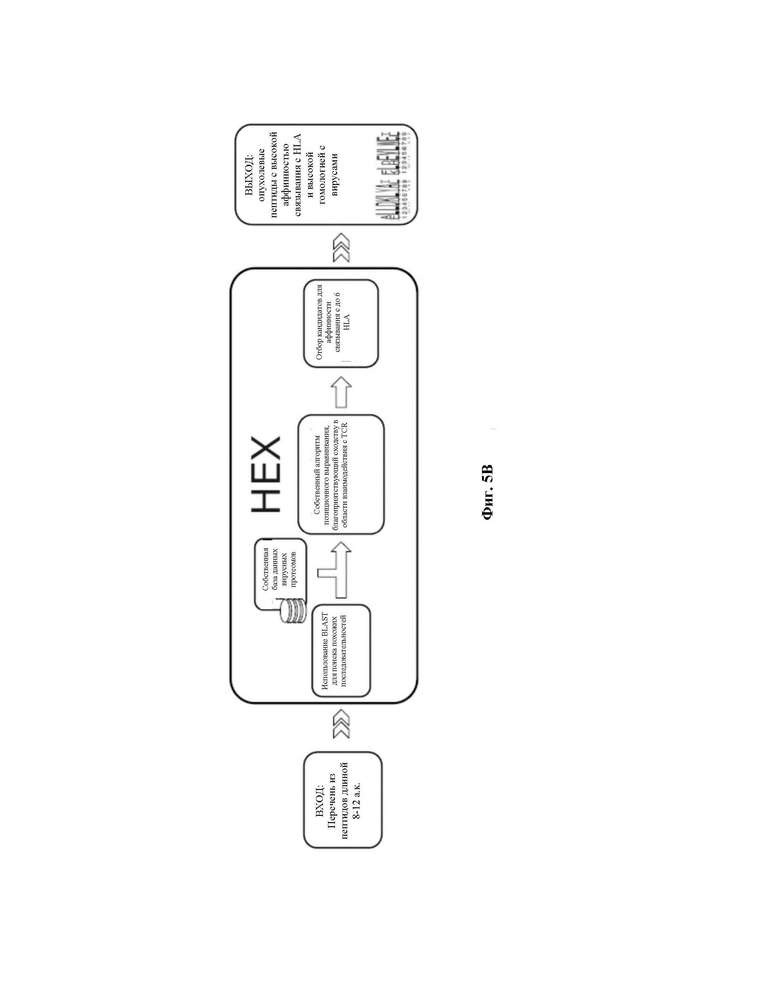
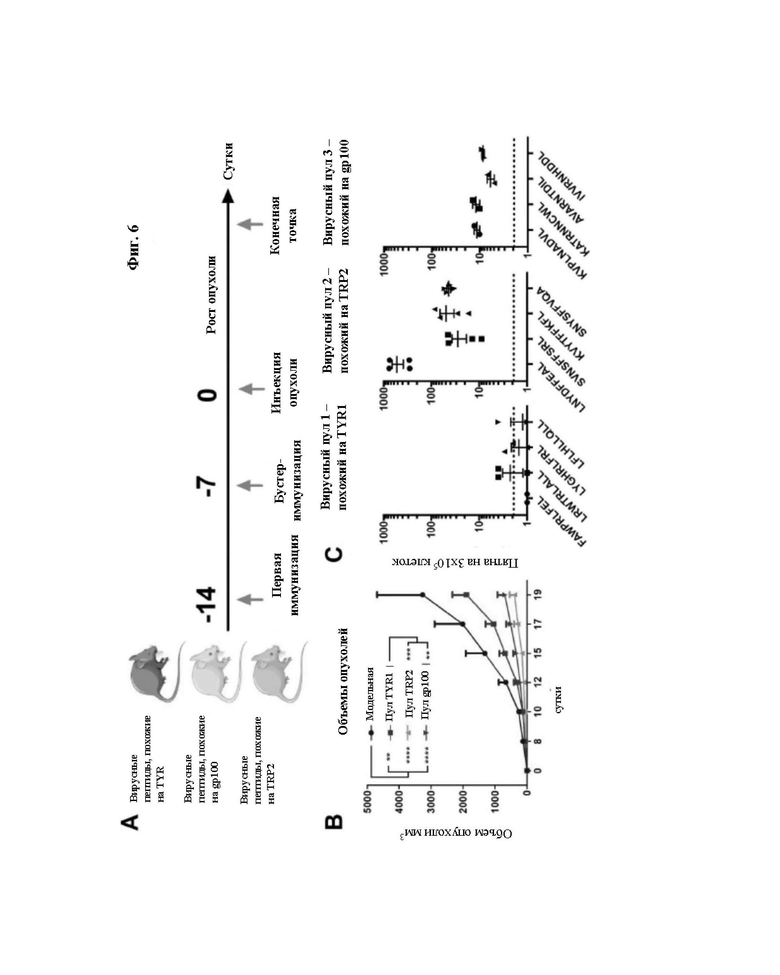
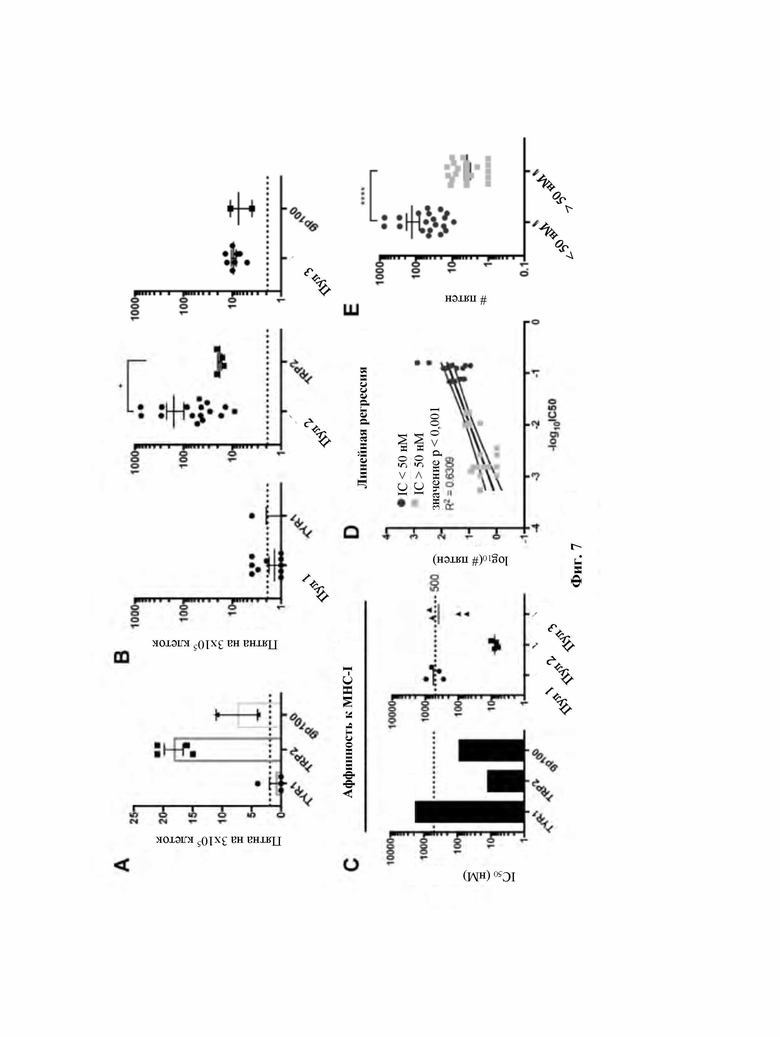

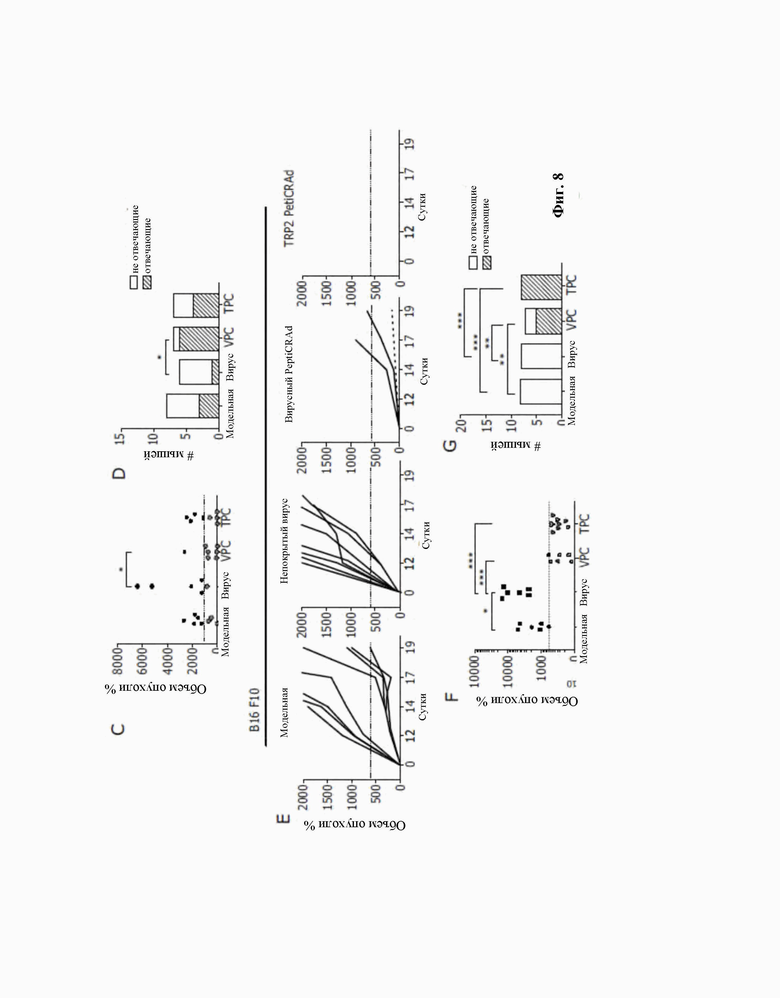
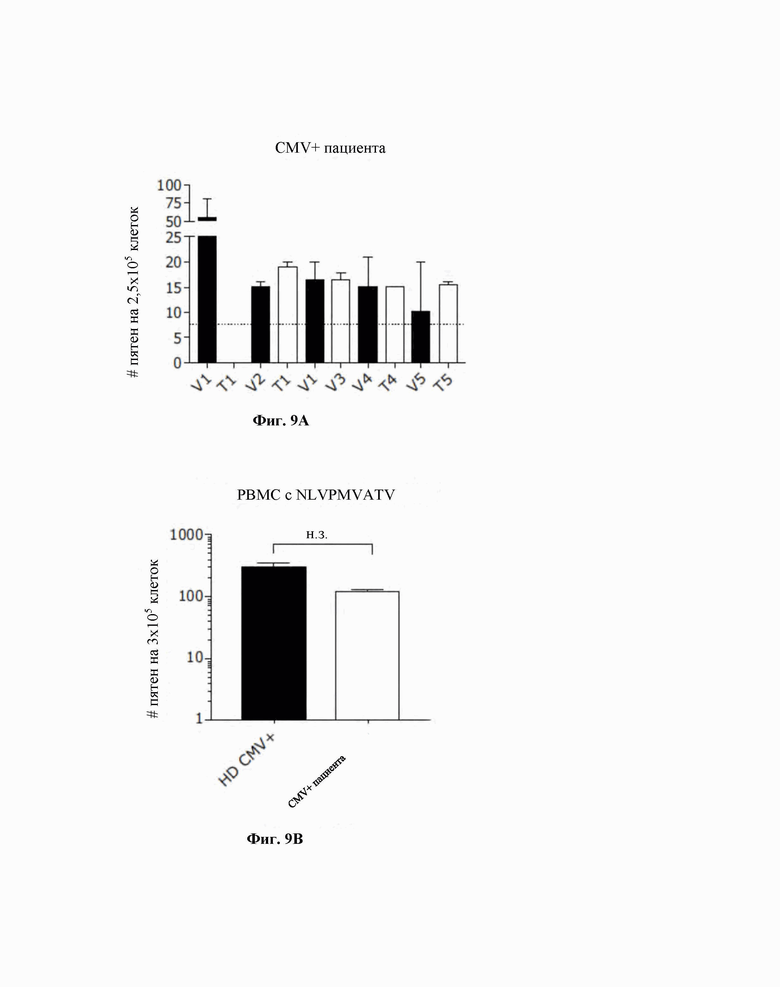
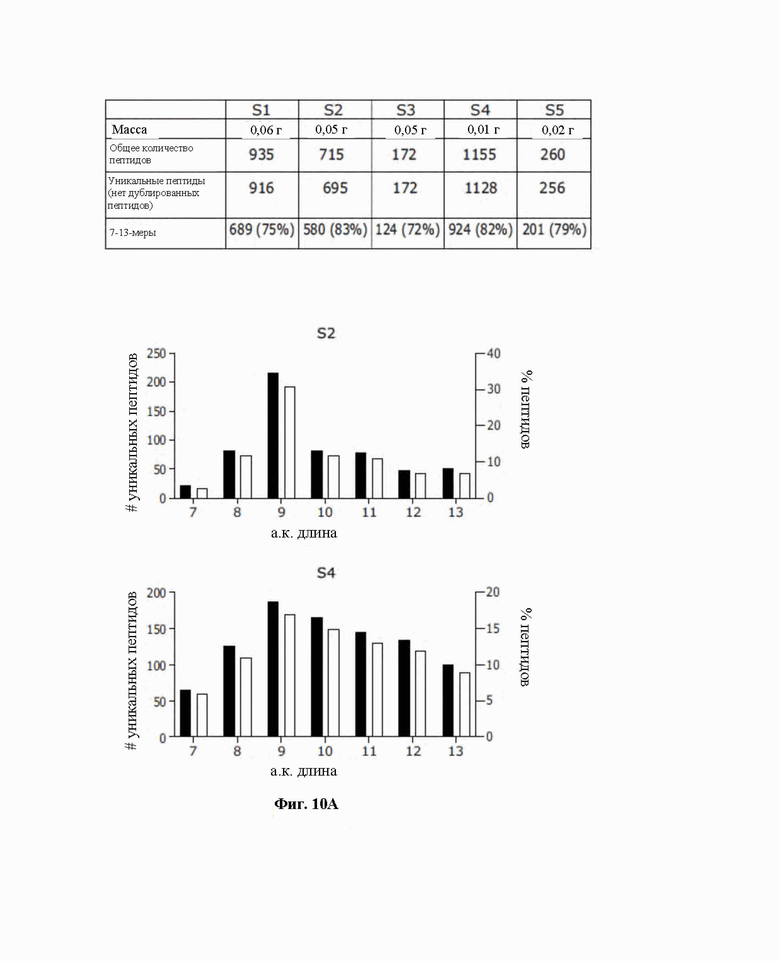
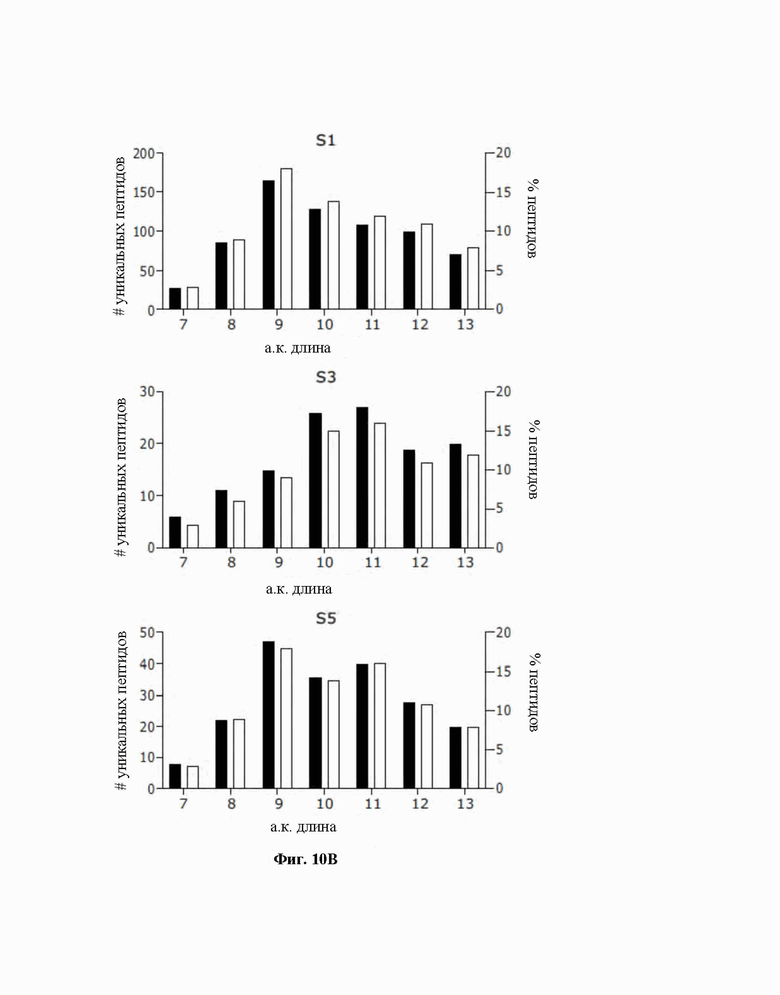
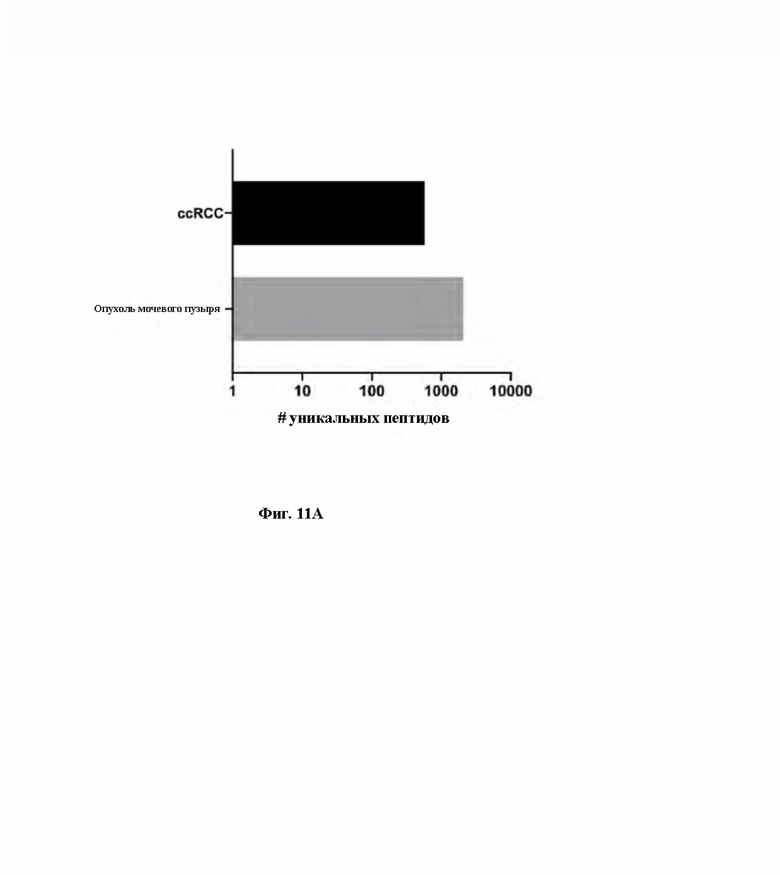
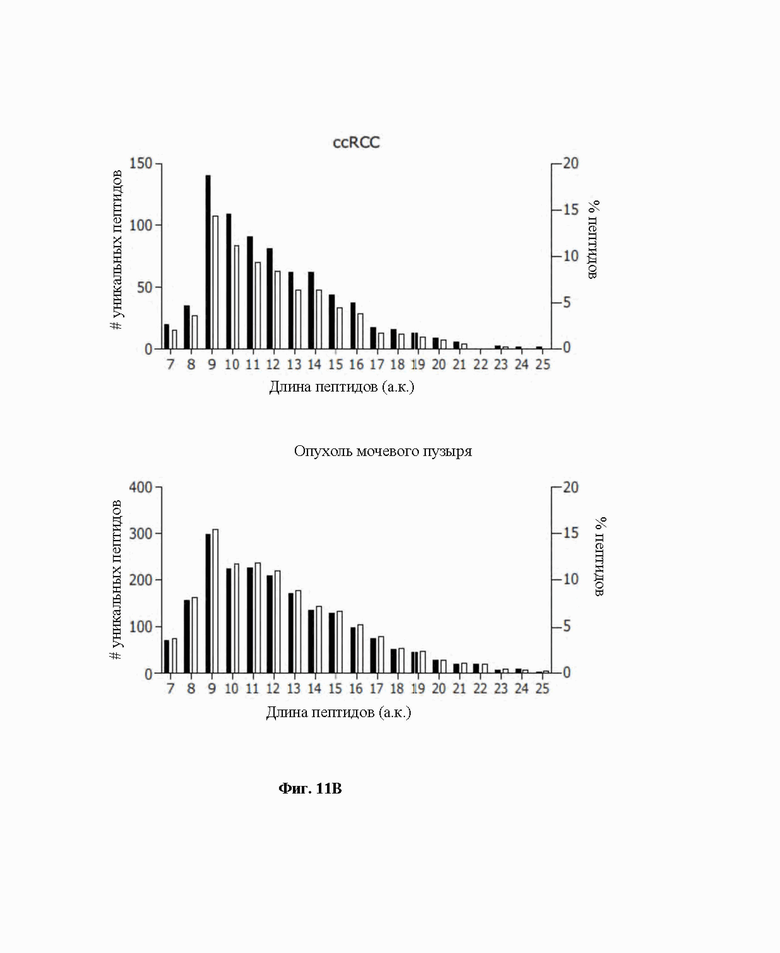
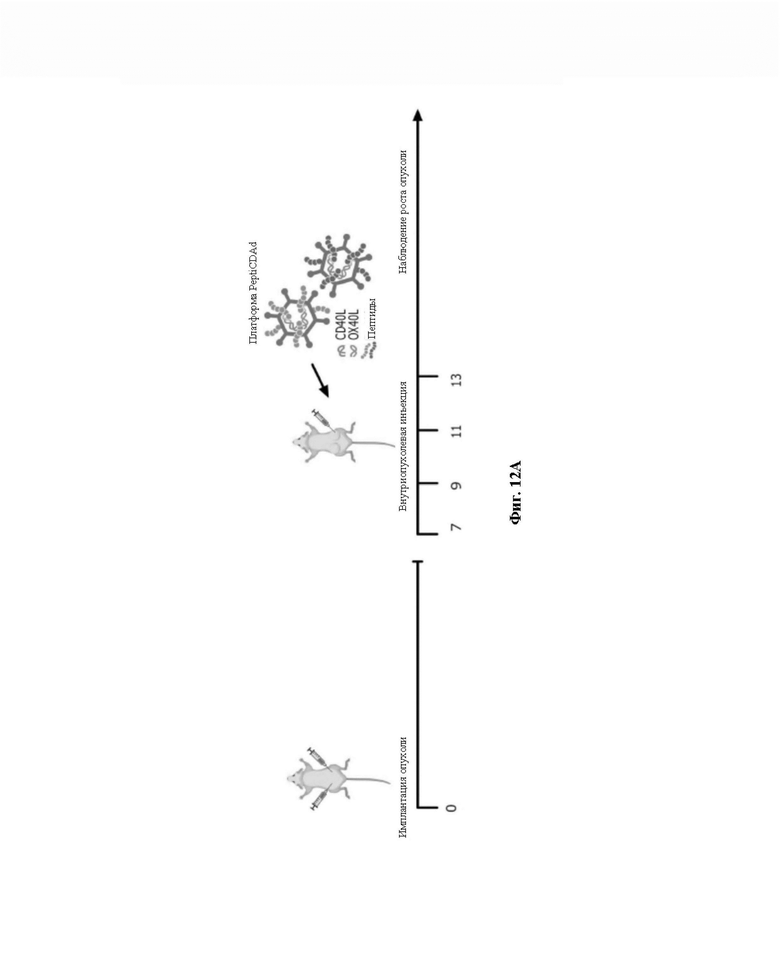
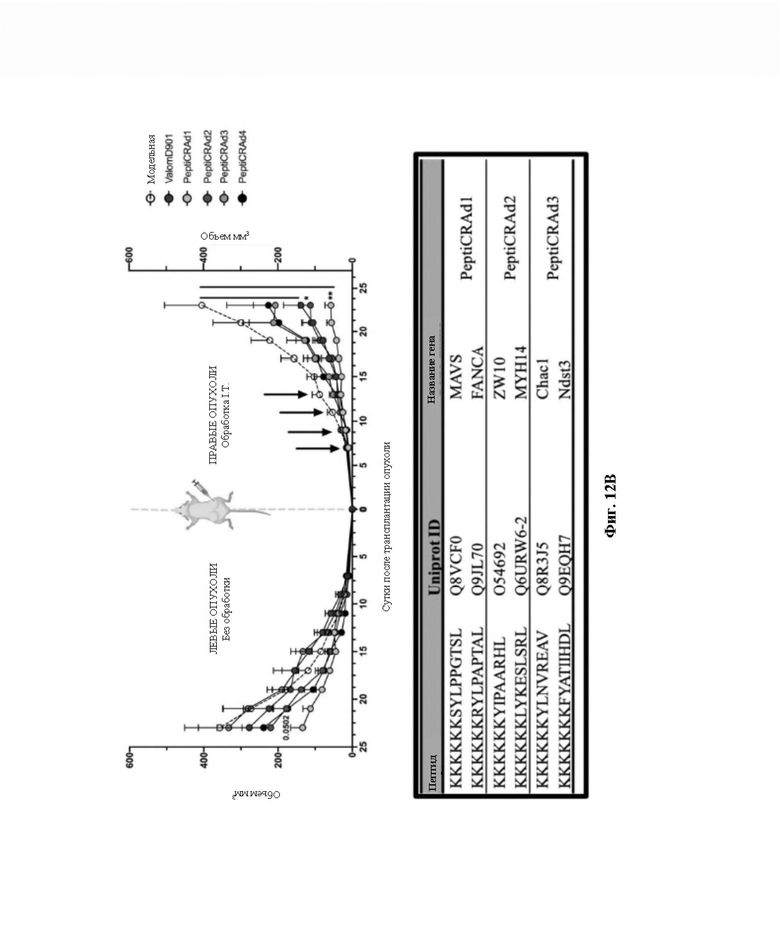
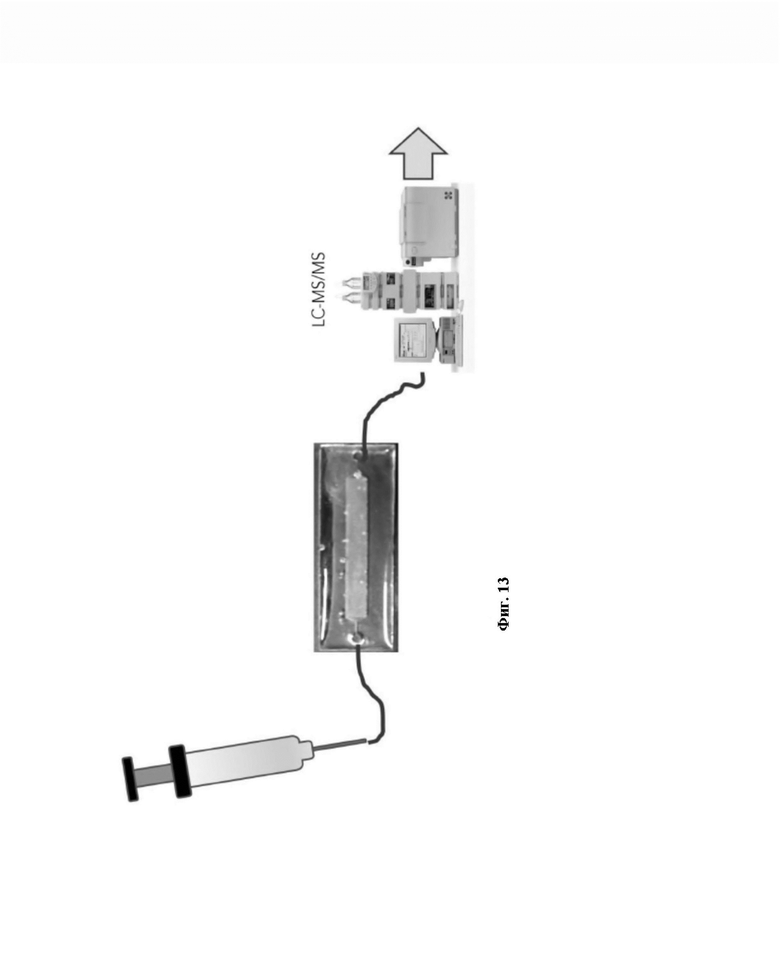
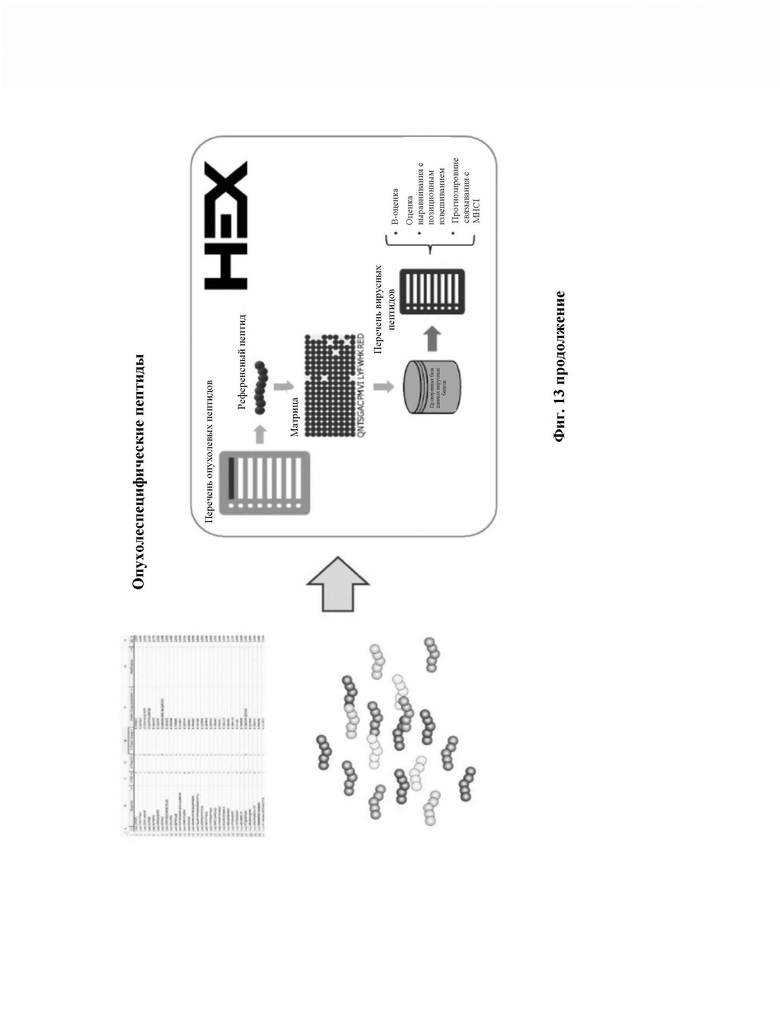
Изобретение относится к биотехнологии. Описан способ идентификации опухолеспецифического антигена с помощью следующего устройства, включающего: микрофлюидное устройство, содержащее по меньшей мере один проточный канал, содержащий множество микростолбиков, упорядоченных в матрицу, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против главного комплекса гистосовместимости или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего пептид:главный комплекс гистосовместимости (pMHC) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела; и процессор, адаптированный для идентификации указанного пептида, связанного с указанным главным комплексом гистосовместимости, и сравнения указанного пептида указанного связанного pMHC с библиотекой пептидных антигенов патогенов для определения того, демонстрирует ли указанный пептид, который связан или был связан с указанным главным комплексом гистосовместимости, гомологию/аффинность с по меньшей мере одним антигеном патогена или его частью, и где существует более чем 60%-ная гомология/аффинность; определения указанного пептида, который связан или был связан с указанным главным комплексом гистосовместимости, в качестве опухолеспецифического антигена для применения в противораковой терапии. Описан способ стратификации пациентов в отношении лечения рака ингибитором контрольных точек. Кроме того, представлен способ стратификации пациентов в отношении лечения аденовирусного рака. Представлен способ лечения рака. Изобретение расширяет арсенал средств для борьбы с раком. 5 н. и 16 з.п. ф-лы, 13 ил., 6 табл.
1. Способ идентификации опухолеспецифического антигена, включающий:
1) растворение или суспендирование образца опухоли в жидкости;
2) пропускание указанной жидкости через микрофлюидное устройство для идентификации опухолеспецифического антигена, включающее: по меньшей мере один проточный канал, содержащий множество микростолбиков, упорядоченных в матрицу, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против главного комплекса гистосовместимости или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена, посредством чего пептид:главный комплекс гистосовместимости (pMHC) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
3) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC в указанном образце;
4) извлечение указанного по меньшей мере одного связанного pMHC в соответствии с частью (3) из указанного устройства;
5) сравнение указанного пептида указанного связанного pMHC с библиотекой антигенов патогенов для определения того, демонстрирует ли указанный пептид гомологию/аффинность с по меньшей мере одним антигеном патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
6) идентификация указанного пептида в качестве опухолеспецифического антигена для применения в противораковой терапии.
2. Способ по п. 1, где указанная библиотека антигенов патогенов содержит проверенную библиотеку известных антигенов патогенов.
3. Способ по п. 1 или 2, где указанные антигены патогенов представляют собой антигены человеческих патогенов.
4. Способ по любому из пп. 1-3 где указанные антигены патогенов представляют собой вирусы.
5. Способ по любому из пп. 1-4, где указанные антигены патогенов получены из по меньшей мере одного вируса, выбранного из группы, содержащей:
Abyssoviridae; Ackermannviridae; Actantavirinae; Adenoviridae; Agantavirinae; Aglimvirinae; Alloherpesviridae; Alphaflexiviridae; Alphaherpesvirinae; Alphairidovirinae; Alphasatellitidae; Alphatetraviridae; Alvernaviridae; Amalgaviridae; Amnoonviridae; Ampullaviridae; Anelloviridae; Arenaviridae; Arquatrovirinae; Arteriviridae; Artoviridae; Ascoviridae; Asfarviridae; Aspiviridae; Astroviridae; Autographivirinae; Avsunviroidae; Avulavirinae; Bacilladnaviridae; Baculoviridae; Barnaviridae; Bastillevirinae; Bclasvirinae; Belpaoviridae; Benyviridae; Betaflexiviridae; Betaherpesvirinae; Betairidovirinae; Bicaudaviridae; Bidnaviridae; Birnaviridae; Bornaviridae; Botourmiaviridae; Brockvirinae; Bromoviridae; Bullavirinae; Caliciviridae; Calvusvirinae; Carmotetraviridae; Caulimoviridae; Ceronivirinae; Chebruvirinae; Chordopoxvirinae; Chrysoviridae; Chuviridae; Circoviridae; Clavaviridae; Closteroviridae; Comovirinae; Coronaviridae; Corticoviridae; Crocarterivirinae; Cruliviridae; Crustonivirinae; Cvivirinae; Cystoviridae; Dclasvirinae; Deltaflexiviridae; Densovirinae; Dicistroviridae; Endornaviridae; Entomopoxvirinae; Equarterivirinae; Eucampyvirinae; Euroniviridae; Filoviridae; Fimoviridae; Firstpapillomavirinae; Flaviviridae; Fuselloviridae; Gammaflexiviridae; Gammaherpesvirinae; Geminialphasatellitinae; Geminiviridae; Genomoviridae; Globuloviridae; Gokushovirinae; Guernseyvirinae; Guttaviridae; Hantaviridae; Hepadnaviridae; Hepeviridae; Herelleviridae; Heroarterivirinae; Herpesviridae; Hexponivirinae; Hypoviridae; Hytrosaviridae; Iflaviridae; Inoviridae; Iridoviridae; Jasinkavirinae; Kitaviridae; Lavidaviridae; Leishbuviridae; Letovirinae; Leviviridae; Lipothrixviridae; Lispiviridae; Luteoviridae; Malacoherpesviridae; Mammantavirinae; Marnaviridae; Marseilleviridae; Matonaviridae; Mccleskeyvirinae; Mclasvirinae; Medioniviridae; Medionivirinae; Megabirnaviridae; Mesoniviridae; Metaparamyxovirinae; Metaviridae; Microviridae; Mimiviridae; Mononiviridae; Mononivirinae; Mymonaviridae; Myoviridae; Mypoviridae; Nairoviridae; Nanoalphasatellitinae; Nanoviridae; Narnaviridae; Nclasvirinae; Nimaviridae; Nodaviridae; Nudiviridae; Nyamiviridae; Nymbaxtervirinae; Okanivirinae; Orthocoronavirinae; Orthomyxoviridae; Orthoparamyxovirinae; Orthoretrovirinae; Ounavirinae; Ovaliviridae; Papillomaviridae; Paramyxoviridae; Partitiviridae; Parvoviridae; Parvovirinae; Pclasvirinae; Peduovirinae; Peribunyaviridae; Permutotetraviridae; Phasmaviridae; Phenuiviridae; Phycodnaviridae; Picobirnaviridae; Picornaviridae; Picovirinae; Piscanivirinae; Plasmaviridae; Pleolipoviridae; Pneumoviridae; Podoviridae; Polycipiviridae; Polydnaviridae; Polyomaviridae; Portogloboviridae; Pospiviroidae; Potyviridae; Poxviridae; Procedovirinae; Pseudoviridae; Qinviridae; Quadriviridae; Quinvirinae; Regressovirinae; Remotovirinae; Reoviridae; Repantavirinae; Retroviridae; Rhabdoviridae; Roniviridae; Rubulavirinae; Rudiviridae; Sarthroviridae; Secondpapillomavirinae; Secoviridae; Sedoreovirinae; Sepvirinae; Serpentovirinae; Simarterivirinae; Siphoviridae; Smacoviridae; Solemoviridae; Solinviviridae; Sphaerolipoviridae; Spinareovirinae; Spiraviridae; Spounavirinae; Spumaretrovirinae; Sunviridae; Tectiviridae; Tevenvirinae; Tiamatvirinae; Tobaniviridae; Togaviridae; Tolecusatellitidae; Tombusviridae; Torovirinae; Tospoviridae; Totiviridae; Tristromaviridae; Trivirinae; Tunavirinae; Tunicanivirinae; Turriviridae; Twortvirinae; Tymoviridae; Variarterivirinae; Vequintavirinae; Virgaviridae; Wupedeviridae; Xinmoviridae; Yueviridae; и Zealarterivirinae, включая любую их комбинацию.
6. Способ по любому из пп. 1-5, где указанный антиген патогена представляет собой антиген цитомегаловируса, или антиген вируса Эпштейна-Барр, или антиген вируса герпеса, или антиген поксвируса, или антиген гепаднавируса, или антиген вируса гриппа, или антиген коронавируса, или антиген вируса гепатита, или антиген HIV (вируса иммунодефицита человека), или антиген буньявируса.
7. Способ по любому из пп. 1-6, где сравнение указанного пептида указанного связанного pMHC с библиотекой антигенов патогенов включает оценку пептида и/или оценку выравнивания для определения указанной гомологии/аффинности.
8. Способ по любому из пп. 1-7, где сравнение указанного пептида указанного связанного pMHC с библиотекой антигенов патогенов включает определение:
а) сходства или идентичности структур полноразмерной последовательности указанного пептида и указанных антигенов патогенов; и/или
б) сходства или идентичности ключевых аминокислот в ключевых сайтах связывания указанного пептида и указанных антигенов патогенов; и/или
в) наибольшего количества похожих или идентичных ключевых аминокислот в ключевых сайтах связывания указанного пептида и указанных антигенов патогенов.
9. Устройство для идентификации опухолеспецифического антигена, включающее:
микрофлюидное устройство, содержащее по меньшей мере один проточный канал, содержащий множество микростолбиков, упорядоченных в матрицу, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против главного комплекса гистосовместимости или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена (HLA), посредством чего пептид:главный комплекс гистосовместимости (pMHC) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела; и
процессор, адаптированный для идентификации указанного пептида, связанного с указанным главным комплексом гистосовместимости, и сравнения указанного пептида указанного связанного pMHC с библиотекой пептидных антигенов патогенов для определения того, демонстрирует ли указанный пептид, который связан или был связан с указанным главным комплексом гистосовместимости, гомологию/аффинность с по меньшей мере одним антигеном патогена или его частью, и где существует более чем 60%-ная гомология/аффинность; определения указанного пептида, который связан или был связан с указанным главным комплексом гистосовместимости, в качестве опухолеспецифического антигена для применения в противораковой терапии.
10. Устройство по п. 9, где указанное антитело против пан-HLA выбрано из группы, содержащей: MHC класса I A, B и C.
11. Устройство по п. 9, где указанное антитело против пан-HLA выбрано из группы, содержащей: MHC класса II DP, DM, DO, DQ и DR.
12. Устройство по любому из пп. 9-11, где указанное антитело является противочеловеческим.
13. Устройство по любому из пп. 9-12, где указанные молекула или комплекс представляют собой комплекс тиольной и алкеновой функциональных групп.
14. Устройство по любому из пп. 9-13, где указанные молекула или комплекс содержат биотин и стрептавидин.
15. Устройство по любому из пп. 9-14, где указанные молекула или комплекс содержат более чем одно антитело, связанное со стрептавидином.
16. Устройство по любому из пп. 9-15, где указанные молекула или комплекс кондиционированы путем покрывания белком, таким как бычий сывороточный альбумин, перед связыванием с указанным антителом.
17. Устройство по любому из пп. 9-16, где указанное устройство адаптировано для осуществления способа по любому из пп. 1-8.
18. Способ стратификации пациентов в отношении лечения рака ингибитором контрольных точек, включающий:
1) отбор у пациента образца опухоли;
2) растворение или суспендирование указанного образца в жидкости;
3) пропускание указанной жидкости через микрофлюидное устройство для идентификации опухолеспецифического антигена, включающее: по меньшей мере один проточный канал, содержащий множество микростолбиков, упорядоченных в матрицу, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против главного комплекса гистосовместимости или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена, посредством чего пептид:главный комплекс гистосовместимости (pMHC) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
4) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC в указанном образце;
5) извлечение указанного по меньшей мере одного связанного pMHC в соответствии с частью (4) из указанного устройства;
6) сравнение указанного пептида указанного связанного pMHC с библиотекой антигенов человеческих патогенов для определения того, демонстрирует ли указанный пептид гомологию/аффинность с по меньшей мере одним антигеном человеческого патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
7) идентификацию указанного пептида в качестве опухолеспецифического антигена; и
8) в случае обнаружения указанного пептида, введение указанному пациенту эффективного количества по меньшей мере одного ингибитора контрольных точек.
19. Способ стратификации пациентов в отношении лечения аденовирусного рака, включающий:
1) отбор у пациента образца опухоли;
2) растворение или суспендирование указанного образца в жидкости;
3) пропускание указанной жидкости через микрофлюидное устройство для идентификации опухолеспецифического антигена, включающее: по меньшей мере один проточный канал, содержащий множество микростолбиков, упорядоченных в матрицу, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против главного комплекса гистосовместимости или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена, посредством чего пептид:главный комплекс гистосовместимости (pMHC) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
4) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC в указанном образце;
5) извлечение указанного по меньшей мере одного связанного pMHC в соответствии с частью (4) из указанного устройства;
6) сравнение указанного пептида указанного связанного pMHC с библиотекой антигенов человеческих патогенов для определения того, демонстрирует ли указанный пептид гомологию/аффинность с по меньшей мере одним антигеном человеческого патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
7) идентификацию указанного пептида в качестве опухолеспецифического антигена; и
8) в случае обнаружения указанного пептида, присоединение указанного пептида к капсиду аденовирусного вектора и введение эффективного количества указанного аденовирусного вектора указанному пациенту.
20. Способ лечения рака, включающий:
1) отбор у пациента образца опухоли;
2) растворение или суспендирование указанного образца в жидкости;
3) пропускание указанной жидкости через микрофлюидное устройство для идентификации опухолеспецифического антигена, включающее: по меньшей мере один проточный канал, содержащий множество микростолбиков, упорядоченных в матрицу, к которым присоединены по меньшей мере одна молекула или по меньшей мере один комплекс, связанные с по меньшей мере одним антителом против главного комплекса гистосовместимости или по меньшей мере одним антителом против человеческого пан-лейкоцитарного антигена, посредством чего пептид:главный комплекс гистосовместимости (pMHC) в образце, проходящем через указанный канал, может экстрагироваться из указанного образца с использованием указанного по меньшей мере одного антитела;
4) связывание с указанным по меньшей мере одним антителом по меньшей мере одного pMHC в указанном образце;
5) извлечение указанного по меньшей мере одного связанного pMHC в соответствии с частью (4) из указанного устройства;
6) сравнение указанного пептида указанного связанного pMHC с библиотекой антигенов человеческих патогенов для определения того, демонстрирует ли указанный пептид гомологию/аффинность с по меньшей мере одним антигеном человеческого патогена или его частью, и где существует более чем 60%-ная гомология/аффинность;
7) идентификацию указанного пептида в качестве опухолеспецифического антигена; и
8) введение эффективного количества указанного пептида указанному пациенту для стимулирования или активирования T-клеток против опухолеспецифического антигена и, таким образом, рака, из которого был отобран образец; или использование указанного опухолеспецифического антигена для увеличения популяции T-клеток, активных против указанного опухолеспецифического антигена, и затем введение указанных T-клеток указанному пациенту.
21. Способ по любому из пп. 19, 20, где указанный рак выбран из группы, содержащей: рак носоглотки, синовиальный рак, гепатоклеточный рак, рак почки, рак соединительных тканей, меланому, рак легкого, рак кишечника, рак толстой кишки, рак прямой кишки, колоректальный рак, рак мозга, рак горла, рак полости рта, рак печени, рак кости, рак поджелудочной железы, хориокарциному, гастриному, феохромоцитому, пролактиному, Т-клеточный лейкоз/лимфому, неврому, болезнь фон Гиппеля-Линдау, синдром Золлингера-Эллисона, рак надпочечников, анальный рак, рак желчных протоков, рак мочевого пузыря, рак мочеточника, олигодендроглиому, нейробластому, менингиому, опухоль спинного мозга, остеохондрому, хондросаркому, саркому Юинга, рак без выявленного первичного очага, карциноид, карциноид желудочно-кишечного тракта, фибросаркому, рак молочной железы, болезнь Педжета, рак шейки матки, рак пищевода, рак желчного пузыря, рак головы, рак глаза, рак шеи, рак почки, опухоль Вильмса, рак печени, саркому Капоши, рак предстательной железы, рак яичка, болезнь Ходжкина, неходжкинскую лимфому, рак кожи, мезотелиому, множественную миелому, рак яичников, эндокринный рак поджелудочной железы, глюкагоному, рак паращитовидной железы, рак полового члена, рак гипофиза, саркому мягких тканей, ретинобластому, рак тонкой кишки, рак желудка, рак тимуса, рак щитовидной железы, трофобластический рак, хорионаденому, рак матки, рак эндометрия, рак влагалища, рак вульвы, акустическую неврому, грибовидный микоз, инсулиному, карциноидный синдром, соматостатиному, рак десен, рак сердца, рак губы, рак оболочек головного мозга, рак рта, рак нервов, рак неба, рак околоушной железы, рак брюшины, рак глотки, рак плевры, рак слюнных желез, рак языка и рак миндалевидной железы.
| WO 2019204229 A1, 24.10.2019 | |||
| RU 2017104900 A, 08.10.2018 | |||
| Т.Н | |||
| ЗАМАЙ, А.Г | |||
| БОРИСОВ и др., Микрофлюидные устройства в диагностике онкологических заболеваний, Сибирское медицинское обозрение, 2013, номер 5, стр.10-14. |

