Область изобретения
Настоящее изобретение относится к модифицированному компетентному по репликации онколитическому аденовирусу; фармацевтической композиции, содержащей указанный аденовирус; и способу лечения рака с их применением.
Предшествующий уровень техники
Восприятие роли онколитических вирусов в лечении рака резко изменилось за последнее десятилетие, поскольку приобрели популярность иммунотерапия и стимуляция собственной иммунной системы пациента для нацеливания и воздействия на рак. В начале столетия онколитические вирусы воспринимали как активные агенты в лечении рака, действующие исключительно благодаря присущей им способности лизировать опухолевые клетки посредством онколиза. В последнее время вызывает интерес их применение в качестве противораковых вакцин, и их способность высвобождать опухолевые антигены из раковых клеток при онколизе для активации иммунной системы признана важной характеристикой при разработке окончательной иммунотерапии против рака.
Аденовирусы представляют собой высокоиммуногенные вирусы, часто используемые в качестве векторов при различных подходах для создания вакцин против инфекционных заболеваний. Важно отметить, что они обладают исключительной способностью примировать и стимулировать иммунные ответы. Кроме того, присутствие онколитического аденовируса в опухоли и вызываемая им опосредуемая иммунными клетками гибель клеток, вероятно, формируют агрессивное микроокружение в опухоли в сторону более восприимчивого состояния для возникновения клинически значимого противоопухолевого иммунитета, вызывая экспрессию иммуномодуляторов TH1-типа, таких как интерферон гамма (IFN-гамма). Инфильтрация иммунных клеток в опухоль является частым следствием лечения онколитическими вирусами, и, что важно, аденовирусы вызывают инфильтрацию CD8+ Т-клетками, которые представляют собой ключевые эффекторные клетки в противораковом иммунитете. Аденовирусы вызывают иммуногенный лизис раковых клеток, после чего опухолевые антигены, включая уникальные специфичные для пациента неоантигены, ранее скрытые от иммунной системы или не презентированные в иммуногенном контексте, высвобождаются в иммуногенную среду. Это представляет собой основу для опухоле-специфичного иммунного ответа, вызываемого онколитическими аденовирусами.
Однако опухоли развили несколько иммуносупрессивных механизмов, чтобы противодействовать иммунным клеткам организма. Иммунные клетки экспрессируют молекулы клеточной поверхности, которые регулируют их активационные и эффекторные функции, в частности костимуляторные и коингибирующие молекулы. Молекулы отрицательной обратной связи, то есть молекулы контрольных точек, обеспечивают ауто-толерантность в нормальных физиологических условиях, но часто используются опухолевыми клетками, чтобы вызвать сильную иммуносупрессию. Наиболее охарактеризованные пути контрольных точек представляют собой путь белка цитотоксических Т-лимфоцитов 4 (CTLA-4) и путь белка 1 программируемой гибели клеток (PD-1/PD-L1). Из-за этих сильных иммуносупрессивных механизмов внутри опухоли вирус-индуцированный противоопухолевый иммунный ответ может быть слабым, если он не усиливается за счет использования иммуностимулирующих трансгенов. Настоящий подход решает проблему иммуносуппрессии в окружении опухоли, так как присутствие высокоиммуногенного онколитического аденовируса, кодирующего иммуностимулирующие трансгены человека, формирует микросреду опухоли в сторону «иммунновоспалительного» фенотипа, который более восприимчив к подходам иммунотерапевтическим подходам. Важно отметить, что онколитический вирус согласно данному подходу можно применять в комбинации с модуляторами контрольных точек, такими как молекулы анти-PD1, анти-PD-L1 или анти-CTLA-4, для противодействия иммуносупрессивной среде опухоли и для того, чтобы вызвать сильный антииммунный ответ.
Краткое описание изобретения
Согласно первому аспекту настоящего изобретения предложен модифицированный реплицирующийся аденовирус, обладающий литической активностью в раковых клетках-мишенях, содержащий:
a) делецию гена a E1A, при этом указанная делеция представляет собой делецию нуклеотидов, кодирующих аминокислоты 923-946;
b) химерную замену 5/3 выступа белка фибры аденовируса, при этом указанный выступ серотипа 5 Ad заменен выступом серотипа 3 Ad;
c) делецию гена 14.7k, при этом указанная делеция относится представляет собой делецию пар оснований 30448-30834 по сравнению с аденовирусом дикого типа, и при этом последовательность GGA GGA GAT GAC TGA (SEQ ID NO: 1) заменена на GGA GGA GAC GAC TGA (SEQ ID NO: 2); и
d) делецию гена gp19k и делецию гена 7.1k, при этом указанные делеции представляют собой делеции пар оснований 28541-29211 по сравнению с аденовирусом дикого типа.
В указанном выше модифицированном аденовирусе аминокислоты 923-946 делетированы из последовательности Ad5 дикого типа (wt). Эта делеция является мерой предосторожности, поскольку вирусный белок E1A не может связываться с молекулой ретинобластомы (Rb) и высвобождать фактор транскрипции E2F из Rb для транскрипции вирусного гена. Таким образом, указанный аденовирус зависит от присутствия свободного E2F в клетке-хозяине и может реплицировать свой геном либо в делящихся нормальных клетках, либо в раковых клетках, где свободный E2F постоянно доступен. Таким образом, данная модификация относительно защищает неделящиеся клетки и направлена против делящихся или раковых клеток.
Кроме того, химерная замена 5/3, то есть замена области выступа фибры аденовируса серотипа 5 на область выступа аденовируса серотипа 3; позволяет указанному вирусу обходить нативный рецептор коксаки-аденовируса Ad5 (CAR) и использовать вместо него нативный рецептор Ad3 десмоглеин 2 (DSG2) для интернализации. DSG2 в большом количестве присутствует в раковых клетках. Таким образом, указанная модификация относительно защищает неделящиеся клетки и направлена против делящихся или раковых клеток.
Аденовирусная инфекция начинается с распознавания рецепторов клетки-хозяина посредством специализированных белков на поверхности вируса, то есть белка фибры (fibre) аденовируса и, в частности, глобулярного домена на карбокси-конце белка фибры аденовируса, называемого карбокси-концевым доменом выступа (knob). Соответственно, в настоящей заявке указание на область выступа белка фибры аденовируса относится к глобулярному домену карбокси-конца белка фибры аденовируса.
Более того, делеция гена 14.7k предотвращает гибель инфицированных клеток в результате цитолиза, индуцированного TNF-альфа, и, таким образом, делеция является преимуществом. Важно отметить, что авторы настоящего изобретения наблюдали короткое перекрытие в начале гена 14.7k и вышерасположенного гена RID бета (14.5k), и, таким образом, была проведена модификация, позволяющая удалить ген 14.7k без удаления последней аминокислоты и стоп-кодона гена RID бета. В частности, нативная последовательность области соединения читается как GGAGGAGATGACTGATTAGGTA (SEQ ID NO: 3), и имеет подчеркнутую последовательность, являющуюся С-концом гена RID бета, и оставшаяся часть представляет собой N-конец гена 14.7k. Трансляция в аминокислоты читается как G G D D стоп (GGA GGA GAT GAC TGA; SEQ ID NO: 1) для гена RID бета. Таким образом, для исключения ATG в ORF гена RID бета, чтобы избежать неправильного прочтения, если трансген вставлен в сайт делеции, и чтобы убедиться, что сохраняется фукнция C-конца RID бета, авторы настоящего изобретения немного изменили последовательность RID бета: GGA GGA GAC GAC TGA (SEQ ID NO: 2). Таким образом, указанная последовательность по-прежнему читается как G G D D стоп, но не содержит ATG для любого неправильного последующего прочтения, которое может мешать экспрессии трансгена.
Наконец, gp19k представляет собой ген, который подавляет MHCI (главный комплекс гистосовместимости класса I) на инфицированной клетке, и указанный ген является избыточным для репликации аденовируса, упаковки и т.д., и поэтому может быть удален. Кроме того, поскольку этот ген представляет собой иммунорегуляторный ген, который аденовирус использует для ускользания от иммунной системы, его делеция является предпочтительной. 7.1k представляет собой ген, связанный с деградацией рецептора TRAIL 2, и ингибирует апоптоз, индуцированный высвобождением Ca2+ из эндоплазматического ретикулума (ER). Однако, поскольку стоп-кодон гена 7.1k находится в открытой рамке считывания gp19k, к этому удаленному сайту не добавляют трансгены.
В предпочтительном варианте реализации настоящего изобретения указанный аденовирус дополнительно модифицирован посредством вставки молекулы, кодирующей OX40L, в идеальном варианте OX40L человека, в область делеции гена 14.7k. OX40L представляет собой активатор Т-клеток и поэтому он является полезным для осуществления настоящего изобретения.
Ген OX40L имеет свой собственный стартовый кодон (ATG), и при вставке в область делеции гена 14.7k предпочтительно, чтобы пара оснований CC была добавлена между стоп-кодоном гена RID бета и стартовым кодоном OX40L для оптимизации трансляции (то есть для получения последовательности Козак ACCATGG).
Наиболее предпочтительно OX40L человека расположен в области E3B, заменяя делецию гена 14.7K. 3'-конец гена RID бета и 5'-конец 14.7K перекрываются в аденовирусе дикого типа (wt), и, таким образом, описанную выше модификацию T/C осуществляли на 3'-конце RID бета, чтобы обеспечить правильную транскрипцию трансгена.
В еще одном предпочтительном варианте реализации настоящего изобретения указанный аденовирус альтернативно или дополнительно модифицирован вставкой CD40L, в идеальном варианте CD40L человека. CD40L человека вставлен в область поздних генов вируса, конкретно в область позднего гена 3 (L3), в идеальном варианте после (downstream) гена 23К, или в область L5, после гена фибры. CD40L активирует антигенпрепрезентирующие клетки (APC), и поэтому он является полезным для осуществления настоящего изобретения.
Предпочтительно трансген CD40L расположен сразу после кодирующей области гена аденовируса 23К, перед его сайтом полиаденилирования, или сразу после гена фибры, перед сайтом его полиаденилирования. Обычно, но не всегда, делеции не выполняют для размещения указанного гена, и, таким образом, экспрессия указанного трансгена зависит от аппарата альтернативного сплайсинга аденовируса и акцепторного сайта сплайсинга (SAS), предшествующего трансгену (например, SAS, адаптированный из US2006/0292682 A1 и WO2006/012393).
В еще одном предпочтительном варианте реализации настоящего изобретения указанный аденовирус дополнительно модифицируют путем вставки акцепторного сайта сплайсинга (SAS) и/или последовательности Козак перед трансгеном CD40L, чтобы способствовать транскрипции CD40L.
Альтернативно и наиболее предпочтительно трансген CD40L вставляют непосредственно после OX40L, используя либо
A) сайт процессинга вируса ящура 2A (F2A) (например, см. Ad5/3-D24-OX40L-F2A-CD40L, конструкция C1 в графических материалах) или
B) сайт процессинга тешовируса свиньи-1 2A (P2A) (например, см. Ad5/3-D24-OX40L-P2A-CD40L, конструкция C3 в графических материалах).
Сайт процессинга 2A вставляют между двумя трансгенами, и в идеальном варианте обоим сайтам процессинга 2A предшествует сайт расщепления (например, сайт расщепления фурином: RKRR) и SGSG-линкер для обеспечения эффективного расщепления трансгенов.
Специалистам в данной области техники понятно, что сайты процессинга 2A представляют собой «саморасщепляющиеся» небольшие пептиды, обнаруженные в пикорнавирусах. Рибосомы клеток-хозяев пропускают синтез глицил-пролиловой пептидной связи на С-конце пептида 2A, что приводит к расщеплению между пептидом 2A и непосредственно расположенным после него пептидом. В результате расщепленный нижележащий пептид имеет пролин на своем N-конце, что означает, что белок CD40L, продуцируемый описанными в настоящей заявке вирусными конструкциями, имеет пролин на своем N-конце.
Соответственно, в предпочтительном варианте реализации настоящего изобретения указанный модифицированный аденовирус включает один и более предпочтительно два трансгена: OX40L и CD40L. Кроме того, обычно, но не исключительно, указанный трансген CD40L обеспечивают практически непосредственно после или непосредственно после указанного трансгена OX40L или наоборот.
Задействование рецептора Т-клеток комплексами антиген-MHCI/II формирует основной сигнал, сигнал 1, для активации наивных Т-клеток. Однако сигнала 1 недостаточно для инициации продуктивного образования и поддержания эффекторных Т-клеток. Полная активация CD8+ Т-клеток требует дополнительных сигналов, управляемых костимулирующими молекулами, присутствующими на активированных APC или хелперных Т-клетках, но редко на опухолях. Костимулирующие молекулы, которые авторы настоящего изобретения выбрали для использования в качестве трансгенов в настоящем изобретении, то есть CD40L и OX40L, являются членами суперсемейства лигандов фактора некроза опухоли (TNF). Большинство лигандов суперсемейства TNF преимущественно экспрессируется на клетках, связанных с иммунной системой, включая B-клетки, T-клетки, естественные киллеры (NK), моноциты и DC (дендритные клетки).
Лиганд OX40 (OX40L) и лиганд CD40 (CD40L) представляют собой трансмембранные белки типа II, которые имеют относительно длинный внеклеточный домен и короткую цитоплазматическую область. Внеклеточный домен (в частности, домен гомологии TNF) проявляет специфичность связывания рецептора, которая необходима для обеспечения функциональности лиганда. Большинство лигандов суперсемейства TNF экспрессируются в виде гомотримеров на поверхности клетки и неактивны или малоактивны как растворимые слитые тримерные белки. Таким образом, растворимая форма без трансмембранного участка не применима для подхода в рамках настоящего изобретения. Следовательно, в настоящем изобретении используют полноразмерную кДНК каждого трансгена. Таким образом, продукты трансгена содержат все домены, которые в природе присутствуют в белке, продуцируемом клеткой, то есть также трансмембранный домен, который направляет белок по секреторному пути и удерживает его в клеточной мембране, а также естественным образом возникающие сайты, которые вызывают отщепление от поверхности клетки протеиназами.
Соответственно, в предпочтительном варианте реализации настоящего изобретения молекула, кодирующая полноразмерный по меньшей мере один или каждый трансген, вставлена в указанный аденовирус, в идеальном варианте является кДНК, кодирующей полноразмерный по меньшей мере один или каждый трансген.
В большинстве подходов, в которых используют костимуляторные молекулы в качестве терапевтического агента для индукции/усиления противоопухолевого иммунитета, используют либо моноклональные агонистические антитела, специфичные к рецепторам лигандов TNF, либо растворимые формы лигандов. Этот подход требует системной доставки указанных терапевтических молекул. Напротив, подход авторов настоящего изобретения напоминает естественную ситуацию в организме, где указанные лиганды презентированы своим родственным рецепторам в виде связанных с мембраной белков, продуцируемых клеточными механизмами инфицированной раковой клетки.
CD4+ Т-клетки играют решающую роль в поддержании эффективного ответа CD8+ Т-лимфоцитов при устойчивых вирусных инфекциях. Новая вирусная конструкция согласно настоящему изобретению с костимуляторным(и) трансгеном(ами) вызывает противовирусный иммунитет Th1-типа в среде опухоли. Присутствие опухолевых антигенов на поверхности вируса и высвобождение других опухолевых антигенов в присутствии местного воспаления, а также распространение детерминант приводит к образованию противоопухолевых иммунных клонов Т-клеток. Чтобы поддерживать эффекторную фазу цитотоксических CD8+ Т-клеток, костимулирующие молекулы (а именно OX40L и CD40L) обладают уникальными свойствами, которые авторы настоящего изобретения хотят применить при лечении иммуносупрессивного рака.
Неожиданным образом, данные авторов настоящего изобретения показывают, что добавление иммуностимулирующих трансгенов, OX40L человека и CD40L человека, в локус 14.7K не снижает онколитической эффективности вирусов согласно настоящему изобретению по сравнению с вирусом-каркасом Ad5/3D24 или вирусом с иммуностимулирующим трансгеном, например GM-CSF человека, заменяющим удаленные гены gp19K/7.1K. Это является неожиданным, потому что указанные трансгены могут сильно влиять на репликацию вируса из-за размера трансгена и прямого воздействия трансгена на инфицированные клетки. Кроме того, делеция гена 14.7K не так широко изучена, как делеция генов gp19K/7.1K, и, таким образом, может иметь неожиданные последствия для репликативного аппарата вируса, особенно в контексте включения трансгенов в сайт делеции 14.7K.
Авторы настоящего изобретения также показывают, что вирус согласно настоящей заявке способен продуцировать функциональные трансгены человека из локуса гена 14.7К. Это является неожиданным, потому что авторы настоящего изобретения использовали, что несколько нетипично, кассету транскрипции с сайтом процессинга вируса 2A между двумя трансгенами.
Кроме того, авторы настоящего изобретения показывают, что вирус согласно настоящему изобретению способен вызывать MAGE-A3 и NY-ESO-1-специфичный иммунный ответ.
Соответственно, в дополнительном аспекте настоящее изобретение относится к фармацевтической композиции, содержащей по меньшей мере один компетентный по репликации аденовирус, проявляющий литическую активность в отношении клетки-мишени, согласно настоящему изобретению и подходящий носитель.
OX40/OX40L
OX40L экспрессируется на активированных APC, включая дендритные клетки, B-клетки и макрофаги. Он экспрессируется на поверхности клетки в виде тримера, позволяющего ему связываться с тремя молекулами OX40. Активация Т-клеток необходима для экспрессии OX40, рецептора OX40L, на CD8+ и CD4+ Т-клетках. Индукция OX40 происходит в течение 24 часов и достигает пика через 48-72 часов после начальной стимуляции TCR и обычно длится 3-4 дня. На относительный уровень рецептора OX40, экспрессируемого на Т-клетках, сильно влияет местная среда через контакт с профессиональными антигенпрезентирующими клетками, экспрессирующими CD80 или CD86, или через микросреду, богатую TNF-альфа или аналогичными воспалительными цитокинами. Кроме того, экспрессию OX40L в основном обычно обнаруживают в сайте воспаления. Таким образом, Т-клетки, экспрессирующие OX40, которые активируются посредством передачи сигналов TCR, получают костимуляцию посредством OX40L в сайте воспаления. Эта специфичность костимуляции посредством OX40 аналогична сигналу опасности, полученному от воспалительной среды, и, таким образом, добавляет уровень безопасности его применению. Кроме того, временная экспрессия OX40 на поверхности Т-клеток после примирования предполагает его важность для поздней пролиферации и выживания эффекторных Т-клеток. Предполагается, что активация антиапоптотических молекул BCL-2, BCL-xL и сурвивина в Т-клетках, стимулированных OX40, ответственна за усиление клональной экспансии и увеличение пула Т-клеток памяти.
CD40/CD40L
Помимо обеспечения костимулирующих сигналов непосредственно Т-клеткам, также выгодно способствовать активации APC, которые, вероятно, перекрестно презентируют опухолевый антиген, таким образом, что in situ достигается повышенная экспрессия костимулирующего лиганда. CD40 имеет решающее значение для функции B-клеток и DC, которые экспрессируют его постоянно. Лиганд CD40 (CD40L) в основном экспрессируется на активированных Т-хелперных клетках. Было показано, что лигирование CD40 «лицензирует» APC и позволяет им управлять ответами эффекторных CTL, частично за счет индукции секреции IL-12, но также за счет повышающей регуляции членов семейства B7 (CD80, CD86). Костимуляция CD40/CD40L важна для индукции эффективных противоопухолевых Т-клеточных ответов, и было показано, что включение CD40L в вакцины на основе опухолевых клеток значительно усиливает иммунные ответы на слабоиммуногенные опухоли у мышей, что подчеркивает важность косвенного влияния на адаптивный иммунитет через APC в противоопухолевом иммунитете.
В еще одном предпочтительном варианте реализации настоящего изобретения указанный аденовирус может относиться к любому типу и виду аденовирусов, например, не ограничиваясь аденовирусом человека, но чаще всего представляет собой аденовирус человека. Наиболее подходящим является то, что указанные аденовирусы способны реплицироваться и уничтожать раковые клетки, направляя противовирусный иммунный ответ против опухоли.
Из вышеизложенного следует, что указанный модифицированный аденовирус согласно настоящему изобретению был сконструирован для стимуляции иммунного ответа против рака и, в частности, в среде опухоли, где, как правило, иммунная система аномально функционирует из-за механизмов уклонения, используемых раковыми клетками.
В предпочтительном варианте реализации настоящего изобретения указанный вирус имеет по меньшей мере один из следующих полипептидов, который ковалентным или нековалентным образом присоединен к вирусному капсиду, но генетически не закодирован указанным аденовирусным вектором ,
1) VFGIELMEVDPIGHLYIFAT [SEQ ID NO: 1];
2) YLAMPFATPMEAELARRSLA [SEQ ID NO: 2];
3) RGPESRLLEFYLAMPFATPM [SEQ ID NO: 3] или
4) полипептид, идентичный указанному выше полипептиду по меньшей мере на 60%.
Соответственно, еще в одном аспекте настоящее изобретение относится к способу лечения рака у пациента, включающему введение пациенту эффективного количества композиции, содержащей по меньшей мере один компетентный по репликации модифицированный аденовирус, проявляющий литическую активность в отношении клетки-мишени, согласно настоящему изобретению.
Дополнительно или в альтернативном варианте, настоящее изобретение относится, по меньшей мере, к одному компетентному по репликации модифицированному аденовирусу, проявляющему литическую активность в отношении клетки-мишени, согласно настоящему изобретению для применения при лечении рака.
Дополнительно или в альтернативном варианте, настоящее изобретение относится к применению по меньшей мере одного компетентного по репликации модифицированного аденовируса, проявляющего литическую активность в отношении клетки-мишени, согласно настоящему изобретению для лечения рака.
Дополнительно или в альтернативном варианте, настоящее изобретение относится к применению по меньшей мере одного компетентного по репликации модифицированного аденовируса, проявляющего литическую активность в отношении клетки-мишени, согласно настоящему изобретению в изготовлении лекарственного средства для лечения рака.
Наиболее предпочтительно в настоящей заявке рак включает любой один или более из следующих видов рака: носоглоточный рак, синовиальный рак, печеночно-клеточный рак, рак почки, рак соединительных тканей, меланому, рак легкого, рак кишечника, рак толстой кишки, рак прямой кишки, колоректальный рак, рак мозга, рак горла, рак полости рта, рак печени, рак поджелудочной железы, хориокарциному, гастриному, феохромоцитому, пролактиному, Т-клеточный лейкоз/лимфому, нейрому, болезнь фон Гиппеля-Линдау, синдром Золлингера-Эллисона, рак надпочечника, рак ануса, рак желчного протока, рак мочевого пузыря, рак мочеточника, олигодендроглиому, нейробластому, менингиому, опухоль спинного мозга, рак кости, остеохондрому, хондросаркому, саркома Юинга, рак с неизвестной первичной локализацией, карциноид, карциноид желудочно-кишечного тракта, фибросаркому, рак молочной железы, болезнь Пэджета, рак шейки матки, рак пищевода, рак желчного пузыря, рак головы, рак глаза, рак шеи, рак почки, опухоль Вильмса, рак печени, саркому Капоши, рак предстательной железы, рак яичек, ходжкинскую лимфому, неходжкинскую лимфому, рак кожи, мезотелиому, множественную миелому, рак яичника, рак эндокринных клеток поджелудочной железы, глюкагонома, рак паращитовидной железы, рак пениса, рак гипофиза, саркому мягких тканей, ретинобластома, рак тонкого кишечника, рак желудка, рак вилочковой железы, рак щитовидной железы, трофобластический рак, пузырный занос (хорионаденому), рак матки, рак эндометрия, рак влагалища, рак вульвы, нейрому слухового нерва, фунгоидный микоз, инсулиному, карциноидный синдром, соматостатиному, рак десны, рак сердца, рак губы, рак оболочки головного мозга, рак ротовой полости, рак нерва, рак неба, рак околоушной железы, рак брюшины, рак зева, рак плевры, рак слюнной железы, рак языка и рак миндалины.
Из вышеизложенного следует, что настоящее изобретение относится к применению модифицированного аденовируса, оптимизированного для обеспечения безопасности и выживаемости, в качестве активного адъюванта в терапевтическом средстве для лечения рака, и в котором используется по меньшей мере один, и в идеальном варианте два иммуностимулирующих агента, то есть OX40L и CD40L.
Указанный модифицированный аденовирус действует как активный адъювант, поскольку он обеспечивает сигналы опасности, необходимые для оптимального иммунного ответа против пептида-мишени, но также сохраняет свою способность уничтожать раковые клетки, которые он инфицирует и в которых реплицирует свой геном. Онколитическок уничтожение клеток является иммуногенным по своей природе, что вызывает изменения в микросреде опухоли, которые вероятно усиливают иммунный ответ на пептиды/опухоль.
В формуле изобретения, которая приведена ниже, и в предыдущем описании изобретения, за исключением случаев, когда контекст требует иного из-за языковых выражений или необходимого смыслового значения, слово «содержит» или варианты, такие как «содержит» или «содержащий», используют в широком смысле, то есть для указания наличия заявленных признаков, без исключения или добавления дополнительных признаков в различных вариантах реализации настоящего изобретения.
Все источники, включая любые патенты или заявки на патенты, указанные в настоящей заявке, включены в настоящую заявку посредством ссылки. Не допускается, чтобы какие-либо источники рассматривались как уровень техники. Кроме того, не допускается, чтобы какой-либо из источников уровня техники рассматривался как часть общих знаний в данной области.
Предпочтительные признаки каждого аспекта настоящего изобретения могут быть такими, как описано в связи с любым из других аспектов.
Другие признаки настоящего изобретения станут понятны из следующих примеров. В целом настоящее изобретение распространяется на любой новый или любую новую комбинацию признаков, раскрытых в настоящей заявке (включая прилагаемую формулу изобретения и графические материалы). Таким образом, следует понимать, что признаки, целые числа, характеристики, соединения или химические фрагменты, описанные в связи с конкретным аспектом, вариантом реализации или примером изобретения, применимы к любому другому аспекту, варианту реализации или примеру, описанному в настоящей заявке, если они не являются несовместимыми с ними.
Более того, в настоящей заявке любой признак может быть заменен альтернативным признаком, служащим по тому же или аналогичному назначению, если не указано иное.
Во всем описании и формуле изобретения единственное число охватывает множественное число, если иное не требуется в контексте. В частности, использование единственного числа следует понимать как охватывающее как множественное число, так и единственное число, если контекст не требует иного.
Далее описан вариант реализации настоящего изобретения только в качестве примера со ссылкой на следующее, в котором:
На Фиг. 1 показан анализ путем электрофореза в агарозном геле ПЦР-амплифицированных фрагментов после реакции сборки Гибсона GA-OX40L/F2A/CD40L и GA-OX40L/P2A/CD40L. На дорожке 1 представлена ПЦР-амплификация собранных фрагментов 1, 2 и 3 с получением полноразмерного фрагмента GA-OX40L/F2A/CD40L размером 3974 п.о. На дорожке 2 представлена ПЦР-амплификация собранных фрагментов 1, 2 и 3 с получением полноразмерного фрагмента GA-OX40L/P2A/CD40L размером 3929 п.о. На дорожке 3 представлена ПЦР-амплификация плазмиды каркаса вируса pAd5/3D24 с амплификацией фрагмента размером 3564 п.о. При этом на обеих дорожках 1 и 2 можно увидеть некоторую амплификацию каркаса вируса.
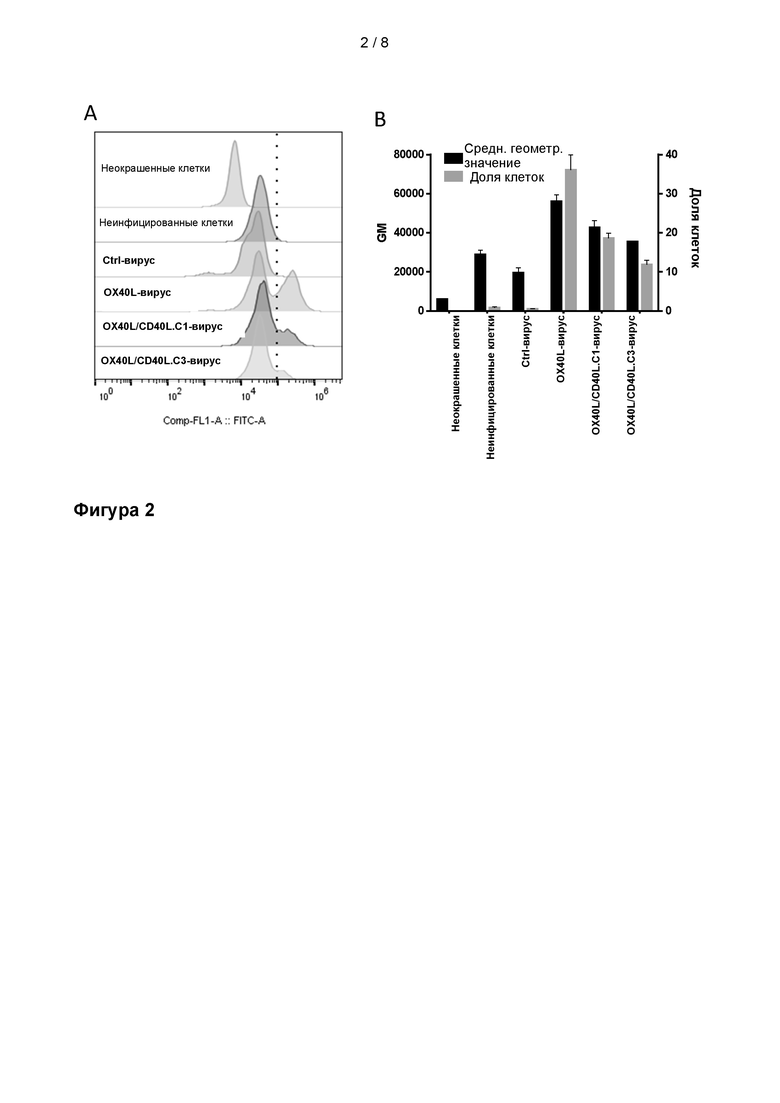
На Фиг. 2 показана способность продуцируемого вирусом белка OX40L связывать его рецептор OX40, подтвержденная проточной цитометрией. Антитело к рецептору OX40 (кролик) и антитело козы против антитела кролика, меченое Alexa fluor 488, использовали для связывания белка OX40L, экспрессированного вирусами на поверхности инфицированных клеток A549. Неокрашенные клетки, неинфицированные окрашенные клетки и окрашенные клетки, инфицированные вирусом без трансгена, использовали в качестве отрицательного контроля. Данные представлены в виде A) гистограммы или B) как среднего значения абсолютных или пропорциональных частот. GM = среднее геометрическое значение количества клеток, положительных по метке Alexa fluor 488, Freq parent = доля клеток, положительных по метке Alexa fluor 488, Ctrl-вирус = Ad5/3D24, вирус с идентичным каркасом и без трансгена, OX40L-вирус = вирус с OX40L только в качестве трансгена, OX40L/CD40L.C1 и OX40L/CD40L.C3 = вирусы с OX40L и CD40L в качестве трансгенов.
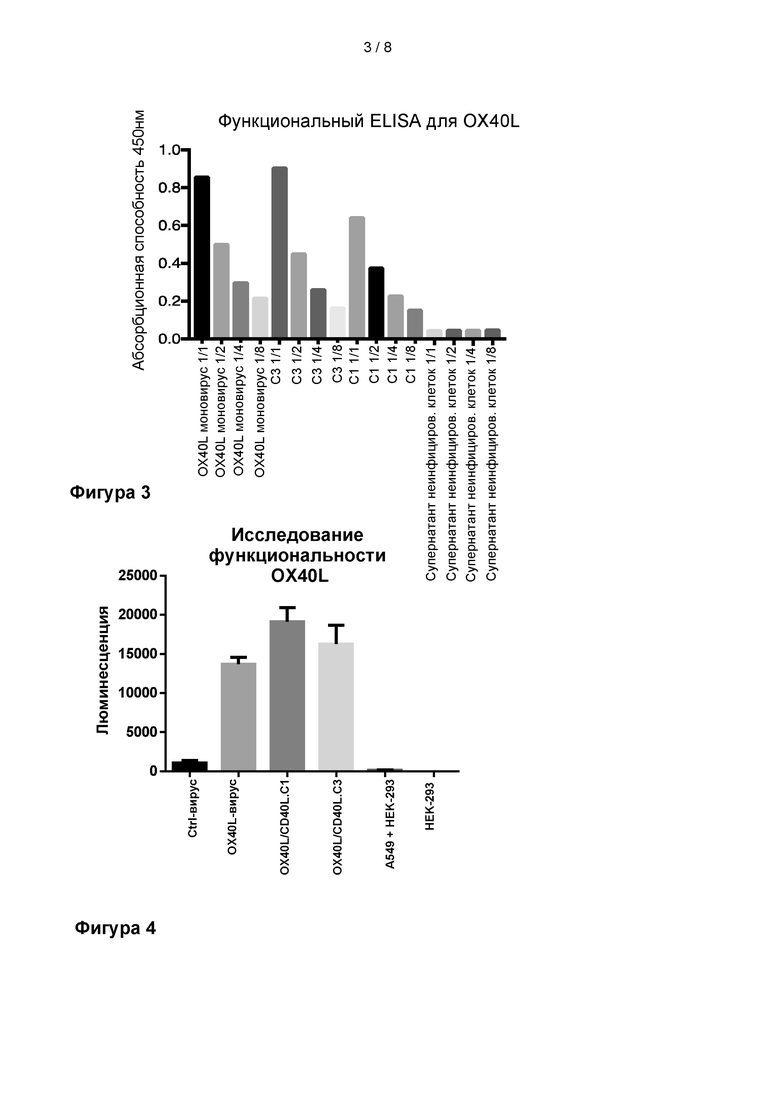
На Фиг. 3 показано, что OX40L, способный связывать рецептор OX40, экспрессируется из Ad5/3-D24-OX40L-CD40.C1 (C1 на фиг.) и Ad5/3-D24-OX40L-CD40.C3 (C3 на фиг.) вирусов с двойным трансгеном, а также из вируса, экспрессирующего только трансген OX40L (используемый в качестве контроля экспрессии). Функциональный сэндвич-ELISA использовали для обнаружения нативной формы OX40L, экспрессированной из вирусов в супернатант инфицированных клеток. Супернатант неинфицированных клеток использовали в качестве отрицательного контроля. Разведения изображены для каждого образца на фигуре.
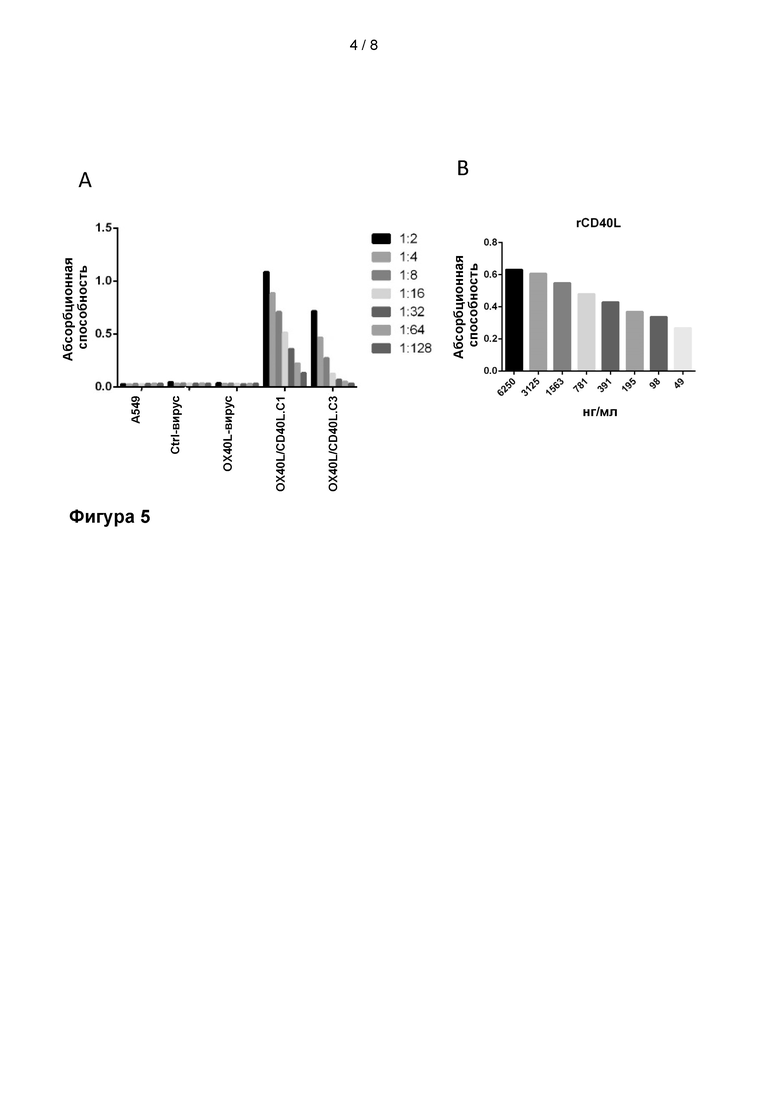
На Фиг. 4 показана функциональность экспрессируемого вирусом OX40L, определенная с использованием клеток HEK-293, экспрессирующих рецептор OX40 с репортерной люциферазной системой. Вирус без трансгена (Ctrl-вирус), вирус только с OX40L в качестве трансгена (OX40L-вирус), вирусы с OX40L и CD40L в качестве трансгенов (OX40L/CD40L.C1 и OX40L/CD40L.C3) проанализировали на предмет их способности запускать активность люциферазы, зависящую от взаимодействия OX40L/OX40. Однозначную активность люциферазы обнаруживали в случае вирусов, экспрессирующих OX40L, что указывает на то, что OX40L, экспрессируемый из вирусного генома, является функциональным.
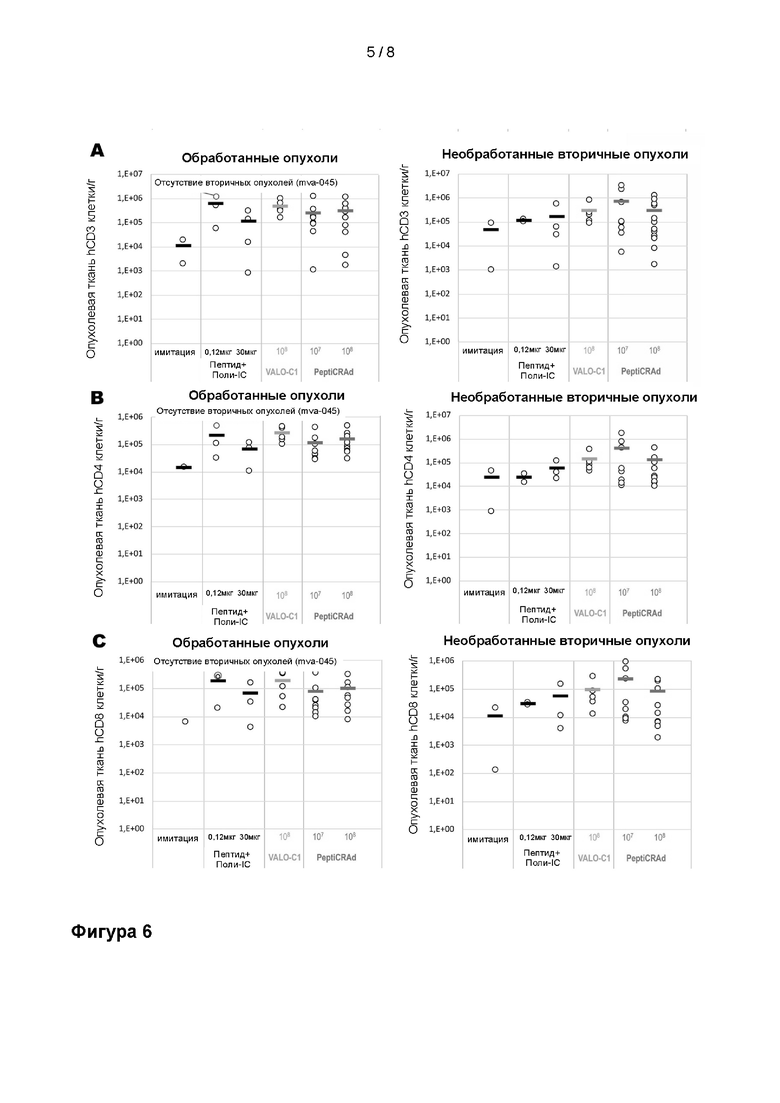
На Фиг. 5 показана абсорбционная способность, измеренная с использованием анализа клеток Ramos Blue для определения функциональных уровней экспрессии CD40L. (A) CD40L экспрессировался из вирусов с двойным трансгеном (OX40L/CD40L.C1 и OX40L/CD40L.C3), и вирус без трансгена (вирус Ctrl) или OX40L в виде одного трансгена (вирус OX40L) или неинфицированные клетки (A549) использовали в качестве отрицательного контроля. (B) Измерения абсорбционной способности клеток Ramos Blue, обработанных рекомбинантным CD40L человека.
На Фиг. 6 показана частота Т-клеток в обработанной и контрлатеральной, необработанной опухоли при обработке онколитическим вирусом без покрытия пептидами (VALO-C1), онклолитическим вирусом с покрытием пептидными антигенами NYESO-1 или MAGE-A3 (PeptiCRAd) или только пептидом. Количество CD3+ T-клеток (A), CD4+ T-клеток (B) и CD8+ T-клеток (C) показано как количество клеток на грамм опухолевой ткани в каждой группе обработки. Обработка привела к более высокой частоте Т-клеток во всех группах по сравнению с имитацией. Наибольшее количество наблюдали в опухолях, обработанных VALO-C1 или PeptiCRAd.
На Фиг. 7 показана частота всех иммунных клеток (клетки CD45+) в обработанной и контрлатеральной, необработанной опухоли при обработке онколитическим вирусом без покрытия пептидами (VALO-C1), онклолитическим вирусом с покрытием пептидными антигенами NYESO-1 или MAGE-A3 (PeptiCRAd) или только пептидом. Частоты были сходными во всех группах с несколько меньшим числом у животных, получавших имитацию.
На Фиг. 8 показано, что обработка VALO-C1 и PeptiCRAd снижает процент регуляторных Т-клеток из всех TIL в обработанных опухолях.
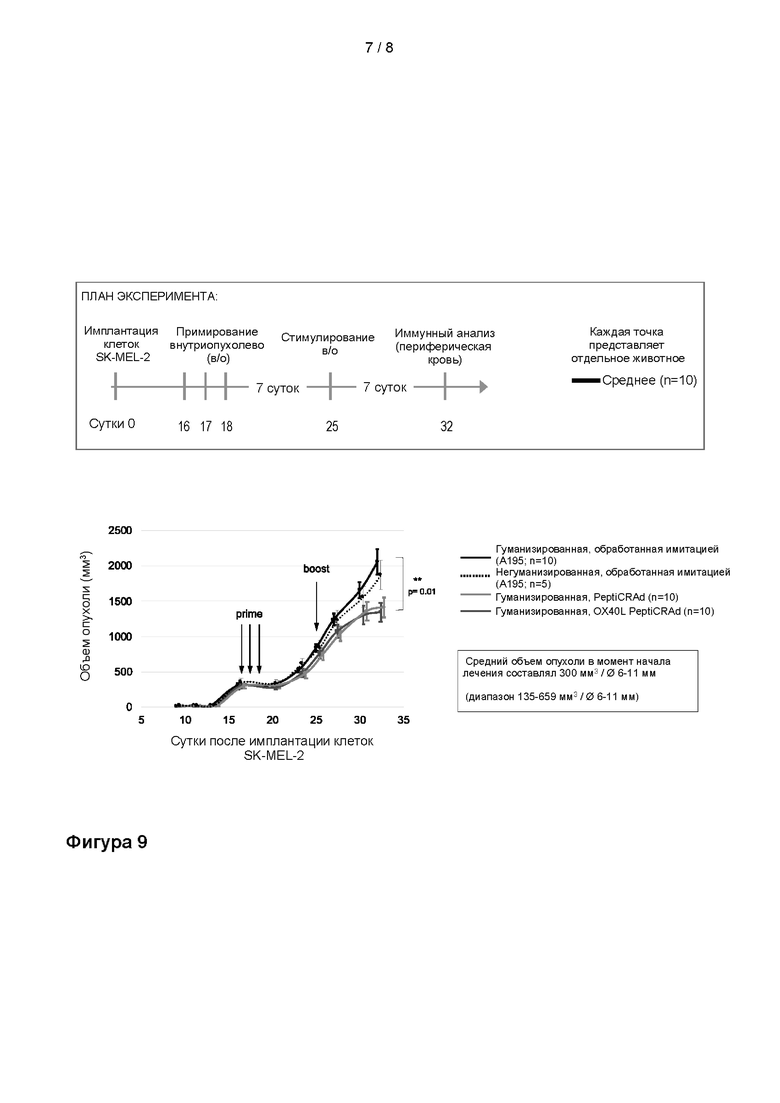
На Фиг. 9 показано, что PeptiCRAd (содержащий OX40L- и CD40L-экспрессирующие вирусы, покрытые белками NY-ESO-1 и MAGE-A3) способен останавливать рост опухоли в модели меланомы у гуманизированной мыши, даже если обработка начата в случае крупных, хорошо развившихся опухолей. Схема эксперимента: 2x106 клеток SK-MEL-2 имплантировали подкожно (одна опухоль на животное) в бок мышей с иммунодефицитом NOD/Shi-scid/IL-2Rγnull в сутки 1. В сутки 13 внутривенно вводили 5x106 PBMC (мононуклеарные клетки периферической крови). В сутки 16 внутриопухолево вводили 5x104 плазмацитоидных и миелоидных дендритных клеток. Внутриопухолевую обработку PeptiCRAd в дозе 1x109 ВЧ (вирусные частицы) проводили в сутки 16, 17, 18 (примирование) и в сутки 25 (стимулирование). Первую дозу PeptiCRad вводили сразу после инъекции DC. Наблюдали за ростом опухоли. Животных умерщвляли на 32 сутки. PeptiCRAd = Ad5/3-D24-OX40L-CD40L, онколитический аденовирус с делецией 24 п.о. в E1A, с химерным капсидом 5/3 и трансгенами CD40L и OX40L, экспрессируемыми из локуса 14.7K, покрытый пептидами NY-ESO-1 и MAGE-A3; OX40L PeptiCRAd = Ad5/3-D24-OX40L, онколитический аденовирус с делецией 24 п.о. в E1A, с химерным капсидом 5/3 и трансгеном OX40L, экспрессируемым из локуса 14.7K, покрытый пептидами NY-ESO-1 и MAGE-A3.
На Фиг. 10 показано, что OX40L(только)-PeptiCRAd увеличивал количество MAGE-A3-специфичных CD8+ Т-клеток в периферической крови по сравнению с животными, получавшими имитацию. Два животных, получавших PeptiCRAd, также показали повышенное количество MAGE-A3-специфичных CD8+ Т-клеток в крови. Т-клетки против MAGE-A3 оценивали с помощью проточной цитометрии (пентамерный анализ) в конце ранее упомянутого исследования роста опухоли на 32 сутки.
Подробное описание
Материалы и способы:
Создание онколитического аденовируса, имеющего делецию нуклеотидов, кодирующих аминокислоты 923-946, гена E1A
Делецию 24 пар оснований производили в каркасной последовательности Ad5 с использованием челночной плазмиды, нацеленной на область E1A, указаный метод клонирования описан у Kanerva et al., делеция впервые описана у Fueyo et al.
Создание онколитического аденовируса, имеющего химерную замену 5/3 белка фибры аденовируса
Выступ серотипа 5 заменяли выступом серотипа 3 с использованием челночной плазмиды с модифицированной областью фибры для введения последовательности посредством гомологичной рекомбинации в каркас вируса. Конкретные способы клонирования описаны в Kanerva et al.
Создание онколитического аденовируса, имеющего делецию пар оснований 30448-30834 гена 14.7k по сравнению с аденовирусом дикого типа и в котором последовательность GGA GGA GAT GAC TGA (SEQ ID NO: 1) заменена на GGA GGA GAC GAC TGA (SEQ ID NO: 2), и создание онколитического аденовируса, имеющего делецию пар оснований 28541-29211 гена gp19k и гена 7.1k по сравнению с аденовирусом дикого типа.
Делеция 14.7К и замена GGA GGA GAT GAC TGA (SEQ ID NO: 1) на GGA GGA GAC GAC TGA (SEQ ID NO: 2) (со вставкой трансгена OX40L на место 14.7К) и делецию генов gp19k/7.1k производили в челночную плазмиду (pShuttle-OX40L) посредством химического синтеза. На основе виртуальной последовательности, разработанной в программе Vector NTI, в GeneArt (Thermo Fisher Scientific) были созданы перекрывающиеся олигонуклеотиды, которые вместе включали всю последовательность. Синтез олигонуклеотидов осуществляли посредством твердофазного синтеза с применением стекла с контролируемыми порами в качестве твердого материала. Затем олигонуклеотиды переводили в жидкую фазу и собирали с помощью ПЦР на полностью автоматизированной станции сборки. Синтетически клонированную последовательность вводили в вектор клонирования pMX и проверяли секвенированием.
Клонирование OX40L и CD40L в pAD5/3-D24 для получения pAd5/3-D24-OX40L/F2A/CD40L и pAd5/3-D24-OX40L/P2A/CD40L
Для создания вирусов, содержащих гены OX40L и CD40L и один из сайтов процессинга вируса ящура 2A (F2A) или сайт процессинга тешовируса свиньи-1 2A (P2A), вставленных между трансгенами для ко-трансляционного процессинга, 3 фрагмента для каждой конструкции амплифицировали с помощью ПЦР.
Для конструкций, содержащих F2A, фрагмент 1, содержащий последовательность OX40L и часть сайта процессинга F2A, амплифицировали из pShuttle-OX40L с использованием праймеров Gibson OX40L и обратного праймера F2A OX40L (см. cписок всех праймеров, использованных в Таблице 1), фрагмент 2, содержащий часть сайта процессинга F2A и полную последовательность CD40L, амплифицировали из pShuttle-CD40L с использованием праймеров прямого праймера F2A CD40L и обратного праймера F2A CD40L.
Для конструкций, содержащих P2A, фрагмент 1, содержащий последовательность OX40L и часть сайта процессинга P2A, амплифицировали из pShuttle-OX40L с использованием праймеров Gibson OX40L и обратного праймера P2A OX40L (см. cписок всех праймеров, использованных в Таблице 1), фрагмент 2, содержащий часть сайта процессинга P2A и полную последовательность CD40L, амплифицировали из pShuttle-CD40L с использованием праймеров прямого праймера P2A CD40L и обратного праймера P2A CD40L.
Для конструкций с F2A и P2A фрагмент 3, содержащий геномную последовательность аденовируса, фланкирующий 3’-конец CD40L, амплифицировали из pShuttle-OX40L с использованием прямого праймера F2A к адено-концу без вставки и праймера Gibson OX40L REV. Все реакции ПЦР проводили с использованием ДНК-полимеразы Phusion High-Fidelity (Thermo Fisher, F530) в соответствии с инструкциями производителя с последующей обработкой DpnI (NEB, R0176). Реакционные смеси очищали с помощью набора NucleoSpin® Gel и PCR Clean-up (MACHEREY-NAGEL, 740609.50). Очищенные фрагменты затем собирали вместе, создавая фрагменты GA-OX40L/F2A/CD40L и GA-OX40L/P2A/CD40L с использованием мастер-микса для сборки Гибсона (NEB, E2611) с последующей амплификацией собранного фрагмента с помощью ПЦР с использованием праймеров Gibson OX40L FW и Gibson OX40L REV (см. Фиг. 1 в отношении анализа конечных фрагментов в агарозном геле).
Чтобы собрать GA-OX40L/F2A/CD40L или GA-OX40L/P2A/CD40L в каркасе вируса, pAd5/3-D24 расщепляли SrfI (NEB, R0629L) и BarI (SibEnzyme®, E548) с последующим осаждением этанолом. Расщепленный каркас вируса pAd5/3-D24 собирали с фрагментами GA-OX40L/F2A/CD40L или GA-OX40L/P2A/CD40L с использованием мастер-микса для сборки Гибсона (NEB, E2611) в соответствии с инструкциями производителя для создания pAd5/3-D24-OX40L/F2A/CD40L и pAd5/3-D24-OX40L/P2A/CD40L. Реакционные смеси сборки Гибсона трансформировали в NEB® 5-альфа компетентные клетки E.coli (NEB, C2987H) в соответствии с инструкциями производителя. Положительные колонии подвергали скринингу с помощью ПЦР, и события правильной рекомбинации дополнительно подтверждали секвенированием конструкций.
Клонирование CD40L и OX40L в pAD5/3-D24 для получения pAd5/3-D24-CD40L-OX40L
Чтобы создать фрагмент, включающий CD40L (CD40L-GA), который будет клонирован в каркас вируса pAd5/3-D24 с помощью реакции специфической гомологичной рекомбинации (торговая марка Gibson Assembly, New England Biolabs), три продукта ПЦР сначала собирали вместе с реакцией рекомбинации сборки Гибсона. Слитые вместе фрагменты содержали следующие последовательности: Фрагмент А соответствует нуклеотидам с 21376 по 22114 плазмиды pAd5/3-D24. Фрагмент B соответствует нуклеотидам от 999 до 2623 плазмиды pShuttle-CD40L. Фрагмент C соответствует нуклеотидам 22820-27107 плазмиды pAd5/3-D24 (см. список использованных праймеров и последовательностей праймеров в Таблицах 1b-3b). ПЦР-реакции проводили с использованием ДНК-полимеразы Phusion High-Fidelity (Thermo Fisher, F530) в соответствии с инструкциями производителя, и указанные реакции очищали с помощью NucleoSpin® Gel и набора для очистки ПЦР (MACHEREY-NAGEL, 740609.50). Затем для создания CD40L-GA очищенные фрагменты ПЦР собирали вместе с помощью реакции рекомбинации сборки Гибсона в соответствии с инструкциями производителя (мастер-микс сборки Гибсона, NEB E2611). После гомологичной рекомбинации вновь созданный фрагмент CD40L-GA дополнительно амплифицировали с помощью ПЦР с использованием праймеров AA и DD (последовательности праймеров см. в Таблицах 1b и 3b) с использованием ДНК-полимеразы Phusion High-Fidelity в соответствии с инструкциями производителя. CD40L-GA, амплифицированный с помощью ПЦР, очищали в геле с использованием NucleoSpin® Gel и набора для очистки ПЦР. Чтобы клонировать CD40L-GA в каркас вируса, pAd5/3-D24 расщепляли SpeI (NEB, R0133S) и AsiSI (NEB, R0630S) с последующим осаждением этанолом. Расщепленный каркас вируса pAd5/3-D24 собирали с фрагментом CD40L-GA с использованием мастер-микса для сборки Гибсона (NEB, E2611) в соответствии с инструкциями производителя для создания pAd5/3-D24-CD40L. Реакционную смесь после сборки Гибсона трансформировали в NEB® 5-альфа компетентные клетки E.coli (NEB, C2987H) в соответствии с инструкциями производителя. Положительные колонии подвергали скринингу с помощью ПЦР, и правильную рекомбинацию дополнительно подтверждали секвенированием конструкций. Для создания конечной конструкции, то есть вируса, содержащего оба гена CD40L и OX40L, фрагмент, включающий OX40L (OX40L-GA), амплифицировали с использованием следующих праймеров: Gibson OX40L FW и Gibson OX40L REV (амплификация области в pShuttle-OX40L, соответствующей нуклеотидам с 11 по 3287), см. последовательности праймеров в Таблице 4b. Реакцию ПЦР проводили с использованием ДНК-полимеразы Phusion High-Fidelity (Thermo Fisher, F530) в соответствии с инструкциями производителя, и указанные реакционные смеси очищали с помощью NucleoSpin® Gel и набора для очистки ПЦР (MACHEREY-NAGEL, 740609.50). Чтобы клонировать OX40L-GA в каркас вируса, pAd5/3-D24-CD40L расщепляли SrfI (NEB, R0629L) и BarI (SibEnzyme®, E548) с последующим осаждением этанолом. Расщепленный каркас вируса pAd5/3-D24-CD40L собирали с фрагментом OX40L-GA с использованием мастер-микса для сборки Гибсона (NEB, E2611) в соответствии с инструкциями производителя для создания pAd5/3-D24-CD40L-OX40L. Реакционную смесь после сборки Гибсона трансформировали в NEB® 5-альфа компетентные клетки E.coli (NEB, C2987H) в соответствии с инструкциями производителя. Положительные колонии подвергали скринингу с помощью ПЦР, и правильную рекомбинацию дополнительно подтверждали секвенированием конструкций.
Способы тестирования модифицированного аденовируса с одним или двумя трансгенами in vitro
Анализ методом проточной цитометрии для определения взаимодействия OX40L/OX40
Проводили анализ методом проточной цитометрии, чтобы убедиться, что OX40L, экспрессируемый вирусами, способен связывать свой нативный рецептор OX40 (Фиг. 2). Клетки человека A549 помещали на 6-луночный планшет и инфицировали вирусами с двумя трансгенами (Ad5/3-D24-OX40L-CD40L.C1 и Ad5/3-D24-OX40L-CD40L.C3, обозначенными как OX40L/CD40L.C1 и OX40L/CD40L.C3), вирусом только с OX40L (Ad5/3-D24-OX40L, обозначенный в графических материалах как OX40L-вирус) или вирусом без трансгенов (Ad5/3-D24, обозначенный как ctrl-вирус) с множественностью заражения 10 (то есть 10 вирусов на клетку).
Через 72 часа после инфицирования указанные клетки собирали и подсчитывали, и 3×105 клеток высевали на лунку 96-луночного планшета в двух экземплярах. Планшет центрифугировали при 400g в течение 5 минут и повторно суспендировали в PBS. Этот этап повторяли дважды, а затем клетки суспендировали в смеси антитела к рецептору OX40 и антитела козы против антитела кролика с Alexa fluor 488, инкубировали в течение 30 минут, промывали 3 раза и затем анализировали в проточном цитометре BD Accuri для определения среднего геометрического значения клеток с комплексом OX40L/OX40. Данные проанализировали с помощью программного обеспечения FlowJo.
Сэндвич-ELISA для проверки взаимодействия OX40L/OX40
Для дополнительной проверки того, что OX40L, экспрессируемый вирусами, способен связывать свой собственный рецептор OX40, проводили функциональный анализ сэндвич-ELISA (Фиг. 3). 96-луночный планшет покрывали 2 мкг/мл рецептора OX40 в его нативной форме в течение ночи и затем промывали 3 раза 0,05% Tween20 об./об. в PBS. В лунки добавляли супернатант от инфицированных вирусом клеток A549, и планшет инкубировали при 37°C в течение 1 часа, а затем снова промывали 3 раза. Лунки инкубировали в течение 1 часа с антителом мыши против OX40L человека (разведение 1:1000 в PBS), промывали 3 раза и инкубировали с конъюгатом антитела против антитела мыши и HRP (разведение 1:1000 в PBS) в течение 1 часа. После 3-кратного промывания планшетов добавляли 90 мкл субстрата TMB и планшет инкубировали в темноте при комнатной температуре в течение 10 минут. HRP, конъюгированная с антителом против антитела мыши, калориметрически реагирует с субстратом TMB, и интенсивность окраски измеряли при 450 нм спектрофотометрически.
Анализ функциональности с рекомбинантной клеточной линией OX40/NF-kB - HEK293
Чтобы проверить, что OX40L, продуцируемый вирусами (либо вирусами с одним трансгеном, либо вирусами с двойным трансгеном), является функциональным и способен активировать нижележащие сигналы при связывании с его рецептором OX40, проводили анализ функциональности с использованием линии клеток эмбриональной почки человека 293 (HEK- 293), постоянно экспрессирующей OX40 (Фиг. 4).
Продукт OX40L продуцируют в культуральной среде культуры клеток A549, инфицированных вирусом, экспрессирующим ген OX40L. Среду собирают и добавляют к культуре OX40/NF-κB репортерных клеток HEK293, постоянно экспрессирующих рецептор OX40. Связывание OX40L с рецептором OX40 запускает внутриклеточный сигнальный путь, который через активацию NF-κB приводит к экспрессии репортерного гена люциферазы светлячка. Активность люциферазы измеряют с использованием системы анализа люциферазы ONE-step и люминометра для определения относительной биолюминесценции известной концентрации стандарта OX40L. Затем можно проанализировать концентрацию OX40L в образце вируса на основании показаний люминесценции и стандартной кривой. Прежде чем можно будет определить концентрацию OX40L, необходимо определить стандартную кривую для OX40L с использованием известных концентраций рекомбинантного человеческого OX40L.
Вкратце, 1,5×104 клеток A549 высевали на 96-луночный планшет в 10% DMEM. На следующий день клетки A549 инфицировали с множественностью заражения 10, т.е. 10 вирусов на клетку, либо вирусом с одним трансгеном (Ad5/3-D24-OX40L, в графических материалах обозначенным как OX40L-вирус), либо вирусами с двойным трансгеном (Ad5/3-D24-OX40L-CD40L.C1 и Ad5/3-D24-OX40L-CD40L.C3, обозначенные как OX40L/CD40L.C1 и OX40L/CD40L.C3) или вирусом без трансгена (Ad5/3-D24, обозначенный как ctrl-вирус) в 2% DMEM.
Некоторые лунки оставляли неинфицированными, чтобы использовать их в качестве отрицательного контроля. Через 72 часа после инфицирования клетки центрифугировали при 500g в течение 5 минут, среду удаляли и поверх клеток A549 добавляли 2×105 клеток OX40/NF-kB-HEK293 в 100 мкл 10% MEM. После центрифугирования при 400g в течение 1 минуты клетки инкубировали при 37°C в течение 6 часов перед лизированием буфером для лизиса и добавлением 20 мкл каждого лизата в лунки прозрачного 96-луночного планшета. После добавления 100 мкл реагента люциферазы в качестве субстрата люминесценцию сразу же считывали на люминометре.
Анализ функциональности трансгенного продукта CD40L
Чтобы проверить, что белок CD40L, экспрессируемый из вирусов с двойным трансгеном, способен связывать свой нативный рецептор CD40 и активировать нижележащие сигналы на экспрессирующих CD40 клетках, проводили анализ функциональности клеток Ramos Blue (Фиг. 5).
Ramos Blue представляет собой клеточную линию B-лимфоцитов, которая стабильно экспрессирует NF-κB-индуцируемый репортерный ген SEAP (секретируемая эмбриональная щелочная фосфатаза). Продукт CD40L продуцируется в культуральной среде культуры клеток, инфицированных вирусом PeptiCRAd, экспрессирующим ген CD40L. Среду собирают и добавляют к культуре клеток Ramos Blue, постоянно экспрессирующих рецептор CD40. Связывание CD40L с CD40 запускает внутриклеточный сигнальный путь, который приводит к секреции SEAP, который окрашивает субстрат в синий цвет, что можно измерить спектрофотометрически при 620-655 нм. Относительную концентрацию функционального CD40L определяют с помощью стандартной кривой для рекомбинантного человеческого CD40L.
Вкратце, клетки A549 человека высевали на 6-луночный планшет и инфицировали вирусами с двумя трансгенами (OX40L/CD40L.C1 или OX40L/CD40L.C3), вирусом только с OX40L (OX40L-вирус) в качестве трансгена или вирусом без трансгенов (Ctrl-вирус) с множественностью заражения 10. Супернатант собирали через 72 часа, и любые клетки и клеточный дебрис удаляли центрифугированием при 500 g в течение 5 минут. Готовили серию 2-кратных разведений из супернатантов и рекомбинантного белка CD40L с начальной концентрацией 100 мкг/мл. 100 мкл каждого супернатанта добавляли к 4×105 клеток Ramos Blue в 96-луночном планшете (засевали по 100 мкл) и планшет инкубировали при 37°C в течение 18 часов. После инкубации клетки осаждали центрифугированием при 400 g в течение 5 минут, и 40 мкл супернатанта добавляли в новый 96-луночный планшет. Добавляли 160 мкл субстрата QUANTI-Blue, планшет инкубировали в течение 1 часа и уровень SEAP определяли спектрофотометрически.
Способы тестирования модифицированного аденовируса с покрытием пептидным антигеном и без него in vivo
Мышей с иммунодефицитом NOD/Shi-scid/IL-2Rγnull гуманизировали с использованием гемопоэтических стволовых клеток (CD34+, HLA-B35+), выделенных из пуповинной крови человека. Опухоли меланомы человека A375 имплантировали подкожно (2 × 106 клеток на 100 мкл), и животных произвольно распределяли в группы на основании степени гуманизации и размера опухоли. Животных обрабатывали онколитическим вирусом без покрытия пептидами (VALO-C1) или онколитическим вирусом с покрытием пептидным антигеном (PeptiCRAd) (доза вируса 1 x 108 для обеих групп; субоптимальную дозу 1 x 107 также тестировали для PeptiCRAd). Пептидные вакцины (0,12 или 30 мкг) вводили внутрикожно с Poly-IC в качестве адъюванта.
Обработку начинали через 25 дней после рандомизации (С0) болюсной дозой циклофосфамида (1 мг/мышь внутривенно (в/в)). Обработку проводили внутриопухолево (имитация, вирус и PeptiCRAd) или внутрикожно (пептидный контроль) на сутки 1, 2, 3 и 12. Вторичные опухоли имплантировали в контрлатеральный бок через двое суток после третьей обработки (сутки 5). Обработку вторичных опухолей не проводили.
Мононуклеарные клетки периферической крови (PBMC) и инфильтрирующие опухоль CD8+ лимфоциты (TIL) анализировали на CD8+ Т-клетки, специфичные к пептидным антигенам NY-ESO-1 и MAGE методом проточной цитометрии с помощью декстрамерного анализа. Оценивали различные субпопуляции иммунных клеток среди PBMC и TIL. Анализ методом проточной цитометрии выполняли на проточном цитометре Attune NxT (Life Technologies).
Иммунизация в модели мышей с PBMC
35 мышам с иммунодефицитом (NCG) NOD-Prkdcem26Cd52/IL-2Rγ em26Cd22 /NjuCrl в возрасте восьми недель прививали 2,106 опухолевых клеток SKMEL-2(HLA-B35+) в правый бок (сутки 0). На сутки 13 внутривенно вводили 5x106 HLA-B35+ мононуклеарных клеток периферической крови человека (PBMC) от двух разных доноров. Внутриопухолевоую обработку с помощью содержащего NYESO-1 и MAGE-A3 в комплексе с капсидом 5/3, OX40L-экспрессирующего вируса (“OX40L PeptiCRAd”) или содержащего NYESO-1 и MAGE-A3 в комплексе с капсидом 5/3, OX40L- и CD40L-экспрессирующего вируса (“PeptiCRAd”) - начинали в сутки 16 с дозой вируса 1 x 109 ВЧ в комплексе с каждым пептидом. Одновременно с первой обработкой PeptiCRAd внутриопухолево вводили 50000 аутологичных плазмацитоидных и миелоидных дендритных клеток. В сутки 17, 18 (примирование с первичной обработкой) и 25 (стимулирование) опухоли обрабатывали внутриопухолевыми инъекциями PeptiCRAd без добавления дендритных клеток. Схема обработки представлена на Фиг. 10. Наблюдали за ростом опухоли. Животных умерщвляли на 32 сутки. OX40L-PeptiCRAd и PeptiCRad содержат делецию 24 п.о. в E1A, удаленную область gp19k/7.1K, трансген OX40L человека, экспрессируемый из локуса 14.7K, и химерный белок фибры 5/3.
Результаты
Функциональность OX40L, экспрессируемого вирусами
Анализ методом проточной цитометрии, а также сэндвич-ELISA показывают, что продукт трансгена OX40L, экспрессируемый вирусами на клеточной поверхности инфицированной клетки, а также в некоторой степени выделяющийся с поверхности клетки, способен связывать свой рецептор OX40 (Фиг. 2 и 3 соответственно). Наиболее важно то, что при анализе функциональности OX40L, экспрессируемого из вирусов, была замечена четкая активация нижележащего гена при использовании клеток HEK-293, стабильно экспрессирующих рецептор OX40 (Фиг. 4). Связывание OX40L с OX40 запускает внутриклеточный сигнальный путь в этих клетках, который через активацию NF-κB приводит к экспрессии репортерного гена люциферазы светлячка. Уровни биолюминесценции, полученные с использованием клеток A549, инфицированных OX40L-экспрессирующими вирусами, четко показывают, что белок OX40L является функциональным и активирует нижележащую передачу сигналов при связывании с OX40 по сравнению с уровнями биолюминесценции, полученными с использованием вируса без трансгена или отрицательного клеточного контроля.
Функциональность CD40L, экспрессируемого из вирусов, экспрессирующих OX40L/CD40L
Четкую активацию нижележащего гена наблюдали при анализе функциональности CD40L с использованием клеток Ramos Blue, стабильно экспрессирующих рецептор CD40 (Фиг. 5). Связывание CD40L с CD40 запускает внутриклеточный сигнальный путь в этих клетках, который через активацию NF-κB приводит к экспрессии гена SEAP. Уровни поглощения, полученные при использовании клеток A549, инфицированных CD40L-экспрессирующими вирусами, четко показывают, что белок CD40L является функциональным и активирует нижележащую передачу сигналов при связывании с CD40, по сравнению с уровнями поглощения, полученными с использованием вирусов без CD40L в качестве трансгена или отрицательного клеточного контроля.
Модифицированный онколитический вирус с поверхностным пептидным антигеном и без него вызывает иммунный ответ, специфический к пептиду, в гуманизированной модели мыши
Все активные виды обработки (только пептид, вирус без пептида [VALO-C1] и вирус с пептидом [PeptiCRAd]) увеличивали количество иммунных клеток в первичных опухолях по сравнению с животными, получавшими имитацию. Животные, получавшие как VALO-C1, так и PeptiCRAd-1, показали больше Т-клеток (CD3, CD4, CD8) в первичных опухолях по сравнению с животными, получавшими пептидную вакцину или имитацию, после обработки, в то время как общее количество инфильтрирующих иммунных клеток (CD45) было одинаковым во всех группах (Фиг. 6 и 7 соответственно).
Кроме того, количество Т-регуляторных клеток (CD3+/CD4+/FoxP3+) было меньше в первичных опухолях, обработанных VALO-C1 и PeptiCRAd-1, по сравнению с первичными опухолями животных, получавших пептидную вакцину или имитацию (Фиг. 8). Это позволяет предположить, что внутриопухолево введенный иммуногенный аденовирус (непокрытый вирус VALO-C1 или PeptiCRAd-1) модулирует микроокружение опухоли, снижая местную иммуносупрессию.
Онколитический аденовирус с (PeptiCRAd) или без пептидного антигена (VALO-C1) превосходит стандартную пептидную вакцинацию в отношении запуска системных нацеленных на опухоль ответов CD8+ Т-клеток и инфильтрации CD8+ TIL в необработанные отдаленные опухоли. Указанные данные свидетельствуют о том, что PeptiCRAd улучшает специфичность стандартного онколитического вируса в отношении нацеливания на опухоль.
PeptiCRAd вызывает специфичный к пептиду иммунный ответ в PBMC мышиной модели
Обработки PeptiCRAd, включающим в комплексе NY-ESO-1 и MAGE-A3, приводило к остановке роста опухоли в модели гуманизированной меланомы мыши, даже когда обработка была начата для крупных, хорошо развившихся опухолей (Фиг. 9). Мыши, получавшие OX40L-PeptiCRAd, показали значительно больше MAGE-A3-специфичных CD8+ Т-клеток среди всех CD8+ Т-клеток PBMC, чем мыши, получавшие имитацию, что указывает на то, что обработка PeptiCRAd способна вызывать специфичный к пептиду ответ у гуманизированных мышей (Фиг. 10).
Ссылки
Kanerva et al 2003 Mol Ther 12:449-458.
Fueyo et al 2000. Oncogene 19:2-12.
Таблица 1 Праймеры, используемые для клонирования pAd5/3-D24-CD40L-OX40L.
(SEQ ID NO: 4)
TCGAAACATACAACCAAAC 3’ (SEQ ID NO: 5)
TCCTCTTCCTAAGGACACAGAATTCACCAGG 3’ (SEQ ID NO: 7)
CGTGGAGAGCAACCCCGGCCCCATGATCGAAACATACAACCAAAC3’ (SEQ ID NO: 8)
Таблица 1b Праймеры к ПЦР фрагменту А с pAd5/3D24 в качестве матрицы.
(SEQ ID NO: 13)
(SEQ ID NO: 14)
(SEQ ID NO: 15)
Таблица 2b Праймеры к ПЦР фрагменту B с pShuttle-CD40L в качестве матрицы.
(SEQ ID NO: 16)
(SEQ ID NO: 17)
(SEQ ID NO: 18)
(SEQ ID NO: 19)
Таблица 3b Праймеры к ПЦР фрагменту С с pAd5/3-D24 в качестве матрицы.
(SEQ ID NO: 20)
(SEQ ID NO: 21)
(SEQ ID NO: 22)
(SEQ ID NO: 23)
Таблица 4b Праймеры для ПЦР вставки OX40L с pShuttle-OX40L в качестве матрицы.
(5’->3’)
(SEQ ID NO: 24)
(SEQ ID NO: 25)
(SEQ ID NO: 26)
(SEQ ID NO: 27)



Настоящее изобретение относится к биотехнологии. Предложен модифицированный реплицирующийся аденовирус серотипа 5, обладающий литической активностью в раковых клетках-мишенях, содержащий: а) делецию гена E1A, при этом указанная делеция представляет собой делецию пар оснований 923-946; b) химерную замену 5/3 выступа белка фибры аденовируса, при этом указанный выступ серотипа 5 Ad заменен выступом серотипа 3 Ad; c) делецию гена 14.7k, при этом указанная делеция представляет собой делецию пар оснований 30448-30834 по сравнению с аденовирусом дикого типа, и при этом последовательность С-конца гена RID бета GGA GGA GAT GAC TGA заменена на GGA GGA GAC GAC TGA; и d) делецию гена gp19k и делецию гена 7.1k, при этом указанные делеции представляют собой делеции пар оснований 28541-29211 по сравнению с аденовирусом дикого типа. Предложена фармацевтическая композиция, содержащая по меньшей мере один модифицированный компетентный по репликации аденовирус, проявляющий литическую активность в отношении клетки-мишени. Предложено применение по меньшей мере одного модифицированного компетентного по репликации аденовируса, проявляющего литическую активность в отношении клетки-мишени или фармацевтической композиции в производстве лекарственного средства для лечения рака. Предложен способ лечения рака у пациента, включающий введение пациенту эффективного количества фармацевтической композиции или композиции, содержащей по меньшей мере один модифицированный компетентный по репликации аденовирус, проявляющий литическую активность в отношении клетки-мишени. Заявленное изобретение решает проблему иммуносупрессии в окружении опухоли, так как присутствие высокоиммуногенного онколитического аденовируса, кодирующего иммуностимулирующие трансгены человека, формирует микросреду опухоли в сторону «иммуновоспалительного» фенотипа, который более восприимчив к иммунотерапевтическим подходам. 4 н. и 18 з.п. ф-лы, 10 ил., 5 табл.
1. Модифицированный реплицирующийся аденовирус серотипа 5, обладающий литической активностью в раковых клетках-мишенях, содержащий:
а) делецию гена E1A, при этом указанная делеция представляет собой делецию пар оснований 923-946;
b) химерную замену 5/3 выступа белка фибры аденовируса, при этом указанный выступ серотипа 5 Ad заменен выступом серотипа 3 Ad;
c) делецию гена 14.7k, при этом указанная делеция представляет собой делецию пар оснований 30448-30834 по сравнению с аденовирусом дикого типа, и при этом последовательность С-конца гена RID бета GGA GGA GAT GAC TGA заменена на GGA GGA GAC GAC TGA; и
d) делецию гена gp19k и делецию гена 7.1k, при этом указанные делеции представляют собой делеции пар оснований 28541-29211 по сравнению с аденовирусом дикого типа.
2. Модифицированный аденовирус по п. 1, отличающийся тем, что указанный аденовирус дополнительно модифицирован вставкой молекулы, кодирующей OX40L.
3. Модифицированный аденовирус по п. 2, отличающийся тем, что указанный OX40L представляет собой OX40L человека.
4. Модифицированный аденовирус по п. 2 или 3, отличающийся тем, что указанный OX40L вставлен в область E3B, на место делеции гена 14.7К.
5. Модифицированный аденовирус по любому из предыдущих пунктов, отличающийся тем, что указанный аденовирус модифицирован посредством вставки молекулы, кодирующей CD40L.
6. Модифицированный аденовирус по п. 5, отличающийся тем, что указанный CD40L представляет собой CD40L человека.
7. Модифицированный аденовирус по любому из пп. 5, 6, отличающийся тем, что указанная молекула CD40L вставлена непосредственно после OX40L с использованием сайта процессинга 2A.
8. Модифицированный аденовирус по п. 7, отличающийся тем, что сайт процессинга 2A вставлен между двумя трансгенами, и указанному сайту процессинга 2A предшествуют сайт расщепления и SGSG-линкер для обеспечения эффективного расщепления указанных трансгенов.
9. Модифицированный аденовирус по п. 7 или 8, отличающийся тем, что указанный сайт процессинга 2A представляет собой сайт процессинга вируса ящура 2A (F2A) или сайт процессинга тешовируса-1 свиньи 2A.
10. Модифицированный аденовирус по п. 5 или 6, отличающийся тем, что указанная молекула CD40L вставлена в область поздних генов вируса, конкретно в область позднего гена 3 (L3).
11. Модифицированный аденовирус по п. 10, отличающийся тем, что указанная молекула CD40L вставлена после гена 23К, перед его сайтом полиаденилирования.
12. Модифицированный аденовирус по п. 10 или 11, отличающийся тем, что акцепторный сайт сплайсинга и/или последовательность Козак обеспечены или вставлены перед молекулой CD40L.
13. Модифицированный аденовирус по любому из пп. 2-12, отличающийся тем, что молекула, кодирующая полноразмерную кДНК по меньшей мере одного или каждого из OX40L и CD40L, вставлена в указанный аденовирус.
14. Модифицированный аденовирус по любому из пп. 1-13 для применения при лечении рака.
15. Фармацевтическая композиция, содержащая по меньшей мере один модифицированный компетентный по репликации аденовирус, проявляющий литическую активность в отношении клетки-мишени, по любому из пп. 1-14 и подходящий носитель.
16. Фармацевтическая композиция по п. 15, отличающаяся тем, что указанная композиция приготовлена для внутриопухолевой, внутримышечной, внутриартериальной, внутривенной, внутриплевральной, внутрипузырной, внутрикожной, внутриполостной или перитонеальной инъекции или для перорального введения.
17. Фармацевтическая композиция по п. 15 или 16 для применения при лечении рака.
18. Применение по меньшей мере одного модифицированного компетентного по репликации аденовируса, проявляющего литическую активность в отношении клетки-мишени, по любому из пп. 1-14 или фармацевтической композиции по п. 15 или 16 в производстве лекарственного средства для лечения рака.
19. Способ лечения рака у пациента, включающий введение пациенту эффективного количества фармацевтической композиции по п. 15 или 16 или композиции, содержащей по меньшей мере один модифицированный компетентный по репликации аденовирус, проявляющий литическую активность в отношении клетки-мишени, по любому из пп. 1-14.
20. Способ лечения рака по п. 19, отличающийся тем, что указанный по меньшей мере один модифицированный компетентный по репликации аденовирус, проявляющий литическую активность в отношении клетки-мишени, по любому из пп. 1-14 вводят с модулятором контрольной точки клетки.
21. Способ лечения рака по п. 20, отличающийся тем, что указанный модулятор контрольной точки представляет собой молекулу анти-PD1, молекулу анти-PD-L1 или молекулу анти-CTLA-4.
22. Аденовирус по п. 14 или применение по п. 18, или способ по любому из пп. 19-21, отличающиеся тем, что указанный рак выбран из перечня, содержащего или состоящего из: носоглоточного рака, синовиального рака, печеночно-клеточного рака, рака почки, рака соединительных тканей, меланомы, рака легкого, рака кишечника, рака толстой кишки, рака прямой кишки, колоректального рака, рака мозга, рака горла, рака полости рта, рака печени, рака кости, рака поджелудочной железы, хориокарциномы, гастриномы, феохромоцитомы, пролактиномы, Т-клеточного лейкоза/лимфомы, нейромы, болезни фон Гиппеля-Линдау, синдрома Золлингера-Эллисона, рака надпочечника, рака ануса, рака желчного протока, рака мочевого пузыря, рака мочеточника, олигодендроглиомы, нейробластомы, менингиомы, опухоли спинного мозга, остеохондромы, хондросаркомы, саркомы Юинга, рака с неизвестной первичной локализацией, карциноида, карциноида желудочно-кишечного тракта, фибросаркомы, рака молочной железы, болезни Пэджета, рака шейки матки, рака пищевода, рака желчного пузыря, рака головы, рака глаза, рака шеи, рака почки, опухоли Вильмса, рака печени, саркомы Капоши, рака предстательной железы, рака яичка, ходжкинской лимфомы, неходжкинской лимфомы, рака кожи, мезотелиомы, множественной миеломы, рака яичника, рака эндокринных клеток поджелудочной железы, глюкагономы, рака паращитовидной железы, рака пениса, рака гипофиза, саркомы мягких тканей, ретинобластомы, рака тонкого кишечника, рака желудка, рака вилочковой железы, рака щитовидной железы, трофобластического рака, пузырного заноса, рака матки, рака эндометрия, рака влагалища, рака вульвы, нейромы слухового нерва, фунгоидного микоза, инсулиномы, карциноидного синдрома, соматостатиномы, рака десны, рака сердца, рака губы, рака оболочки головного мозга, рака ротовой полости, рака нерва, рака неба, рака околоушной железы, рака брюшины, рака зева, рака плевры, рака слюнной железы, рака языка и рака миндалины.
| WO 2014170389 A1, 23.10.2014 | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Термосно-паровая кухня | 1921 |
|
SU72A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| RU | |||

