Изобретения относятся к области биохимии, клеточной биологии, фармакологии, биотехнологии и медицины, в частности, к способам выделения и применения углевод-распознающих белков, способных связывать специфические молекулярные паттерны в структуре гликанов различного строения, локализованных на клеточных поверхностях и во внеклеточном матриксе, которые выполняют в природе роль патоген-распознающих рецепторов и применяются в биотехнологии и медицине в качестве молекул, специфический аффинитет которых позволяет маркировать, иммобилизовать и инактивировать биологические системы с характерным профилем гликозилирования; изобретение может быть использовано как в научных исследованиях, так и в медицине.
Специфическое взаимодействие белков с углеводами лежит в основе разнообразных межклеточных коммуникаций и взаимодействия клеток с внеклеточным матриксом. Наиболее известной группой углевод-связывающих белков являются лектины, для которых показано сродство к самым разнообразным углеводным детерминантам. Первоначально все углевод-связывающие белки относили к лектинам, однако обширные геномные исследования выявили и другие группы белков, распознающих гликаны, в частности в структурах патоген-ассоциированных молекулярных паттернов (Wang W., Song X., Wang L., Song L. Pathogen-Derived Carbohydrate Recognition in Molluscs Immune Defense // Int. J. Mol. Sci. 2018. Vol. 19. P. 721). Среди этих биополимеров C1q домен-содержащие (C1q domain-containing, C1qDC) белки образуют очень интересное семейство, широко распространенное в различных группах беспозвоночных и особенно среди двустворчатых моллюсков. При этом C1qDC белки нередко классифицируют и как лектины (Yang J., Wei X., Liu X., Xu J., Yang D., Yang J., Fang J., Hu X. Cloning and transcriptional analysis of two sialic acid-binding lectins (SABLs) from razor clam Solen grandis // Fish Shellfish Immunol. 2012. Vol. 32, P. 578-585), и как лектин-подобные белки (Gerdol M., Greco S., Pallavicini A. Extensive Tandem Duplication Events Drive the Expansion of the C1q-Domain-Containing Gene Family in Bivalves // Mar. Drugs. 2019, Vol. 17, P. 583).
В последние годы большое внимание уделяется связывающим углеводы белкам, полученным из морских организмов. Двустворчатые моллюски обладают самым обширным репертуаром лектинов и C1qDC белков, что позволяет им процветать в среде, насыщенной патогенами (Gerdol M., Greco S., Pallavicini A. Extensive Tandem Duplication Events Drive the Expansion of the C1q-Domain-Containing Gene Familyin Bivalves // Mar. Drugs 2019. Vol. 17, P. 583; Gerdol M., Gomez-Chiarri M., Castillo M.G., Figueras A., Fiorito G., Moreira R., Novoa B., Pallavicini A., Ponte G., Roumbedakis K., et al. Immunity in Molluscs: Recognition and Effector Mechanisms, with a Focus on Bivalvia // In Advances in Comparative Immunology. Springer International Publishing: Cham, Switzerland. 2018. P. 225-341). Помимо антимикробных свойств, лектины двустворчатых моллюсков обладают многообещающей противоопухолевой и противовирусной активностью, и, кроме того, могут быть использованы в качестве инструментов в различных областях биологии и медицины.
Широчайшая представленность и характерная функциональность лектинов привела к их активному использованию в биотехнологии, которое началось еще с рицина и арбина, открытых и выделенных более 100 лет назад. Лектины сыграли немаловажную роль на ранних этапах становления иммунологии и гематологии, в первом случае как модельные антигены, а во втором - как вещества способные распознавать группы крови. При этом они все так же остаются важным инструментом иммуногематологии, продолжая классически использоваться для обнаружения специфических антигенов эритроцитов и активации различных типов лимфоцитов, а кроме того стала обсуждаться перспектива их использования как инструментов выявления стволовых клеток через углеводные маркеры (Gorakshakar A.C., Ghosh K. Use of lectins in immunohematology // Asian J Transfus Sci. 2016. Vol. 10, № 1. P. 12–21; Carvalho E.V.M.M., Oliveira W.F., Coelho L.C.B.B., Correia M.T.S. Lectins as mitosis stimulating factors: Briefly reviewed // Life Sci. 2018. Vol. 207. P. 152-157). Также не теряет актуальности их использование в картировании гликанов в гистохимии и клеточной биологии(Manning J.C., Romero A., Habermann F.A., García Caballero G., Kaltner H., Gabius H.-J. Lectins: a primer for histochemists and cell biologists // Histochem. Cell Biol. 2017. Vol. 147, № 2. P. 199–222). Внедрение лектинов в ставшую классической технологию иммуноферментного анализа привело к появлению его нового аналога – ELLSA (enzyme-linked lectin sorbent assay), отличающегося направленностью на выявление углеводных компонент (Wu A.M., Liu J.H. Lectins and ELLSA as powerful tools for glycoconjugate recognition analyses // Glycoconj. J. 2019. Vol. 36, № 2. P. 175–183). Кроме того, современные аналитические методы с применением лектинов включают лектиноаффинную хроматографию, лектиновый блоттинг, анализ на микропланшетах, микрочипах и биосенсорные технологии (Hendrickson O.D., Zherdev A.V. Analytical Application of Lectins // Crit Rev Anal Chem. 2018. Vol. 48, № 4. P. 279-292). Огромный потенциал данной группы белков и современная тенденция к миниатюризации технологий анализа, как в науке, так и в клиницистике, привела к активному обсуждению использования лектинов в системах типа "лаборатория на чипе" (lab-on-a-chip), идеология которых заключается в проведении сложных комплексных процедур с использованием минимального количества аналитов на микрочипах в один шаг (Hassan S.-U., Donia A., Sial U., Zhang X., Bokhari H. Glycoprotein- and Lectin-Based Approaches for Detection of Pathogens // Pathogens. 2020. Vol. 9, № 9).
Другим интенсивно развивающимся направлением является применение лектинов в составе биокомпозитных материалов и структур самого различного назначения и типа, от гликозилированных органических макроструктур до гликонаночастиц на основе неорганических матриц (Bojarová P., Křen V. Sugared biomaterial binding lectins: achievements and perspectives // Biomater Sci. 2016. Vol. 4, № 8. P. 1142-1160). При этом особенность использования в составе наночастиц заключается в их нацеливании по углеводным детерминантам, что может быть использовано в адресной доставке лекарств, таргетной терапии и пероральной иммунизации (Devi R.V., Basil-Rose M.R. Lectins as Ligands for Directing Nanostructured Systems // Curr Drug Deliv. 2018. Vol. 15, № 4. P. 448-452).
Поскольку злокачественная трансформация тесно связана с изменением углеводного репертуара клеточной поверхности и внеклеточного матрикса, то лектины могут представлять собой ценный инструмент для диагностики опухолей и потенциально быть использованными в составе терапевтических средств. Для ряда лектинов уже показаны цитотоксические эффекты, проявляющиеся главным образом за счет индукции апоптотических и аутофагических путей в злокачественных клетках (Mazalovska M., Kouokam J.C. Plant-Derived Lectins as Potential Cancer Therapeutics and Diagnostic Tools // Biomed Res Int. 2020. Vol. 2020. P. 1631394; Catanzaro E., Calcabrini C., Bishayee A., Fimognari C. Antitumor Potential of Marine and Freshwater Lectins // Mar Drugs. 2019. Vol. 18, № 1). При этом рассматриваются варианты, включающие их использование в нанотераностике, представляющей собой объединение диагностических и терапевтических функций в единой системе на основе нанотехнологий (Višnjar T., Romih R., Zupančič D. Lectins as possible tools for improved urinary bladder cancer management // Glycobiology. 2019. Vol. 29, № 5. P. 355-365). Кроме того, известно изменение структуры и функционирования галектинов на поверхности ряда трансформированных клеток, что привело к разработке возможных стратегий лечения на основе влияния гликомиметиков и неогликоконъюгатов на опухолевые галектины (Laaf D., Bojarová P., Elling L., Křen V. Galectin-Carbohydrate Interactions in Biomedicine and Biotechnology // Trends Biotechnol. 2019. Vol. 37, № 4. P. 402-415). Что касается конкретных примеров в изменении углеводного репертуара поверхности опухолевых клеток, то ранее уже была показана роль маннозы и полимеров на ее основе в данных процессах (Stavenhagen K., Laan L.C., Gao C., Mehta A.Y., Heimburg-Molinaro J., Glickman J.N., van Die I., Cummings R.D. Tumor cells express pauci- and oligomannosidic N-glycans in glycoproteins recognized by the mannose receptor (CD206) // Cell. Mol. Life Sci. 2021. Vol. 78. P. 5569-5585), что позволило использовать манноз-специфичные лектины при разработке систем селективной доставки лекарств (Sugahara T., Ohama Y., Fukuda A., Hayashi M., Kawakubo A., Kato K. The cytotoxic effect of Eucheuma serra agglutinin (ESA) on cancer cells and its application to molecular probe for drug delivery system using lipid vesicles // Cytotechnology. 2001. Vol. 36. P. 93-99; Omokawa Y., Miyazaki T., Walde P., Akiyama K., Sugahara T., Masuda S., Inada A., Ohnishi Y., Saeki T., Kato K. In vitro and in vivo anti-tumor effects of novel Span 80 vesicles containing immobilized Eucheuma serra agglutinin // Int. J. Pharm. 2010. Vol. 389, P. 157-167). Кроме того, было показано онкодиагностическое применение различных лектинов, обладающих сродством к β-разветвленным галактанам и маннанам (Chen S., Zheng T., Shortreed M.R., Alexander C., Smith L.M. Analysis of Cell Surface Carbohydrate Expression Patterns in Normal and Tumorigenic Human Breast Cell Lines Using Lectin Arrays // Anal. Chem. 2007. Vol. 79. P. 5698-5702; Qiu Y., Patwa T.H., Xu L., Shedden K., Misek D.E., Tuck M., Jin G., Ruffin M.T., Turgeon D.K., Synal S. et al. Plasma Glycoprotein Profiling for Colorectal Cancer Biomarker Identification by Lectin Glycoarray and Lectin Blot // J. Proteome Res. 2008. Vol. 7. P. 1693-1703).
Многие из лектинов показывают противовирусную активность, в результате чего, некоторые из них были выдвинуты в качестве кандидатов на разработку методов профилактики или лечения вирусных инфекций (Mitchell C.A., Ramessar K., O’Keefe B.R. Antiviral lectins: Selective inhibitors of viral entry // Antiviral Res. 2017. Vol. 142. P. 37-54). Среди потенциальных мишеней при этом отмечают ВИЧ, гепатит, грипп, энцефалит, коронавирус, вирус простого герпеса и другие (Singh R.S., Walia A.K. Lectins from red algae and their biomedical potential // J. Appl. Phycol. 2018. Vol. 30, № 3. P. 1833-1858). Особое внимание при этом уделяют исследованиям возможности применения лектинов в борьбе со СПИДом и сопутствующими инфекциями у ВИЧ-инфицированных (Mazalovska M., Kouokam J.C. Lectins as Promising Therapeutics for the Prevention and Treatment of HIV and Other Potential Coinfections // Biomed Res Int. 2018. Vol. 2018. P. 3750646).
Проблемы развития у инфекционных бактерий резистентности к антибиотикам и сложностей в поисках их новых эффективных представителей толкает на поиски новых антимикробных агентов. Лектины считаются одной из перспективных в этом направлении групп, поскольку множество из них имеют иммуномодулирующую активность, в том числе путем запуска цитокиновых каскадов и ускорения фагоцитоза, а также способны напрямую уничтожать данные патогены (Breitenbach Barroso Coelho L.C., Marcelino Dos Santos Silva P., Felix de Oliveira W., de Moura M.C., Viana Pontual E., Soares Gomes F. et al. Lectins as antimicrobial agents // J. Appl. Microbiol. 2018. Vol. 125, № 5. P. 1238-1252; Mishra A., Behura A., Mawatwal S., Kumar A., Naik L., Mohanty S.S. et al. Structure-function and application of plant lectins in disease biology and immunity // Food Chem. Toxicol. 2019. Vol. 134. P. 110827). Кроме того, рассматривается возможность применения вакцин на основе комплексов инактивированных нагреванием бактерий с лектинами, которые должны вызывать активную выработку антител с более разнообразным репертуаром распознаваемых антигенных детерминант (Mishra A., Behura A., Mawatwal S., Kumar A., Naik L., Mohanty S.S. et al. Structure-function and application of plant lectins in disease biology and immunity // Food Chem. Toxicol. 2019. Vol. 134. P. 110827), а также применение лектинов и какпротивогрибковых агентов (Breitenbach Barroso Coelho L.C., Marcelino Dos Santos Silva P., Felix de Oliveira W., de Moura M.C., Viana Pontual E., Soares Gomes F. et al. Lectins as antimicrobial agents // J. Appl. Microbiol. 2018. Vol. 125, № 5. P. 1238-1252). В основе структуры клеточной стенки бактерий лежат две основные группы сложных биополимеров: пептидогликаны (peptidoglycans, PDG) и липополисахарадиды (lipopolysaccharides, LPS), которые могут быть расположены в различной последовательности, что лежит в основе дифференциальной окраски по Граму. При этом для многих C1qDC белков Bivalvia показан высоких аффинитет в отношении данных биополимеров (Yang J., Wei X., Liu X., Xu J., Yang D., Yang J., Fang J., Hu X. Cloning and transcriptional analysis of two sialic acid-binding lectins (SABLs) from razor clam Solen grandis // Fish Shellfish Immunol. 2012. Vol. 32, р. 578-585; Gestal C., Pallavicini A., Venier P., Novoa B., Figueras A. MgC1q, a novel C1q-domain-containing protein involved in the immune response of Mytilus galloprovincialis // Dev. Comp. Immunol. 2010. Vol. 34. P. 926-934). Кроме того, особое место в составе бактерий занимает кислое производное галактозы - D-галактуроновая кислота, а также производное маннозы - L-глицеро-D-манно-гептоза (Herget S., Toukach P.V., Ranzinger R., Hull W.E., Knirel Y.A., Von der Lieth C.W. Statistical analysis of the Bacterial Carbohydrate Structure Data Base (BCSDB): characteristics and diversity of bacterial carbohydrates in comparison with mammalian glycans // BMC structural biology. 2008. Vol. 8. P. 1-20).
Среди лектинов двустворчатых моллюсков можно отметить лектины мидий MytiLec из Mytilus galloprovincialis и CGL из Crenomytilus grayanus, которые проявляют иммуномодулирующую активность в отношении макрофагов млекопитающих, а также проапоптотическую и противоопухолевую активность за счет связывания с глоботриозой на гликолипидах клеточных поверхностей (Chernikov O., Kuzmich A., Chikalovets I., Molchanova V., Hua K.-F. Lectin CGL from the sea mussel Crenomytilus grayanus induces Burkitt’s lymphoma cells death via interaction withsurface glycan // Int. J. Biol. Macromol. 2017. Vol. 104, № Pt A.P. 508-514; Chernikov O.V., Wong W.-T., Li L.-H., Chikalovets I.V., Molchanova V.I., Wu S.-H. et al. A GalNAc/Gal-specific lectin from the sea mussel Crenomytilus grayanusmodulates immune response in macrophages and in mice // Sci Rep. 2017b. Vol. 7, № 1. P. 6315). При этом такая комбинация свойств показала свою перспективность в качестве эффективных диагностических и терапевтических средств и уже привела к созданию искусственного лектина под названием Мицуба-1 (Mitsuba-1), со смоделированным по образу MytiLec и CGL дизайном (Terada D., Voet A.R.D., NoguchiH., Kamata K., Ohki M., Addy C. et al. Computational design of a symmetrical β-trefoil lectin with cancer cell binding activity // Sci Rep. 2017. Vol. 7, № 1. P. 5943). Кроме того, выраженная антимикробная активность была показана дляразличных растворимых лектинов, выделенных из Modiolus modiolus (Tunkijjanukij S., Mikkelsen H.V., Olafsen J.A. A heterogeneous sialic acid-binding lectin with affinity for bacterial LPS from horse mussel (Modiolus modiolus) hemolymph // Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 1997. Vol. 117, № 2. P. 273-286; Tunkijjanukij S., Olafsen J.A. Sialic acid-binding lectin with antibacterial activity from the horse mussel: further characterization and immunolocalization // Dev. Comp. Immunol. 1998. Vol. 22, № 2. P. 139-150), Argopecten irradians (Zhu L., Song L., Xu W., Qian P.-Y. Identification of a C-type lectin from the bay scallop Argopecten irradians // Mol. Biol. Rep. 2009. Vol. 36, № 5. P. 1167-1173; Huang M., Zhang H., Jiang S., Wang L., Liu R., Yi Q. et al. An EPD/WSD motifs containing C-type lectin from Argopectens irradians recognizes and binds microbes with broad spectrum // Fish Shellfish Immunol. 2015b. Vol. 43, № 1. P. 287-293), Crassostrea hongkongensis (He X., Zhang Y., Yu F., Yu Z. A novel sialic acid binding lectin with anti-bacterial activity from the Hong Kong oyster (Crassostrea hongkongensis) // Fish Shellfish Immunol. 2011. Vol. 31, № 6. P. 1247-1250), Mytilus californianus (García-Maldonado E., Cano-Sánchez P., Hernández-Santoyo A. Molecular and functional characterization of a glycosylated Galactose-Binding lectin from Mytilus californianus // Fish Shellfish Immunol. 2017. Vol. 66. P. 564-574) и многих других видов Bivalvia.
На существующем уровне техники известны лектины и C1qDC белки с антибактериальными свойствами и преобладающей специфичностью к маннозе, галактозе или сиаловой кислоте и, соответственно, ряду поли- и олигосахаридов, включающих их в свою структуру (Cheung R.C.F., Wong J.H., Pan W., Chan Y.S., Yin C., Dan X., Ng T.B. Marine lectins and their medicinal applications // Applied microbiology and biotechnology. 2015. Vol. 99. P. 3755-3773; Wang W., Song X., Wang L., & Song L. Pathogen-derived carbohydrate recognition in molluscs immune defense // International journal of molecular sciences. 2018. Vol. 19. P. 721). Однако в доступной литературе нет ни одного упоминания об углевод-распознающих белках с высоким аффинитетом к кислым галактанам и маннанам, которые являются уникальными паттернами распознавания, в том числе бактериального происхождения (Herget S., Toukach P.V., Ranzinger R., Hull W.E., Knirel Y.A., Von der Lieth C.W. Statistical analysis of the Bacterial Carbohydrate Structure Data Base (BCSDB): characteristics and diversity of bacterial carbohydrates in comparison with mammalian glycans // BMC structural biology. 2008. Vol. 8. P. 1-20; Nazarenko E. L., Crawford R.J. & Ivanova E.P. The structural diversity of carbohydrate antigens of selected gram-negative marine bacteria // Marine Drugs. 2011. Vol. 9. P. 1914-1954).
В охранном документе RU2644332C1 описано выделение лектина вискумин (mestletoe lectin I, ML1) из омелы белой (Viscum album). Способ предусматривает экстракцию белка из сухих листьев омелы белой в 0,25М растворе NaCl, проведение трех этапов очистки, включая аффинную хроматографию с лактозил-содержащим сорбентом и ионно-обменную хроматографию с катионообменной смолой типа monoS, и концентрирование до 2 мг/мл на мембране с размерами пор от 10 до 30 кДА. Композиция содержит 0,04-0,15 мг/мл белка ML1 и водный изотонический солевой раствор или 0,1-10 мкг/мл белка ML1, 0,1-1 мг/мл ферромагнитных наночастиц и водный изотонический солевой раствор. Изобретения обеспечивают получение лектина ML1 из омелы белой с чистотой не менее 95% и фармацевтической композиции на его основе, обладающей иммуностимулирующим, противоопухолевым действием и стабильностью в течение 2,5 года.
Однако ML1 имеет специфичность к углеводам иной структуры, содержащим в терминальном положении галактозу, и композиция на его основе не отмечена противомикробной активностью. Кроме того, способ включает множество этапов, включая длительную экстракцию, два типа хроматографии и дополнительное концентрирование.
Известно выделение специфичного к маннозе лектина EIL из гемолимфы морского краба Erimacrus isenbeckii, которое описано в охранном документе KR20070117156. EIL потенциально может быть использован для снижения риска сердечных приступов и их последствий, а также противораковой терапии. Специфичность EIL к маннозе и гликопротеинам на ее основе была выявлена с помощью метода ингибирования гемагглютинации, а также была определена N-концевая последовательность аминокислот: Gly-Ala-X3-Glu-Ala-Ser-Ile-Leu-Ala-Ser-Phe-Asn-X13-Val-Lys-Phe-X17-Leu-Lys, где X3 и X13 представляют собой либо остатки Cys, либо модифицированные аминокислоты, которые не определяются при аминокислотном секвенировании использованным методом. Способ получения данного белка включает стадии: разбавления гемолимфы морского краба хлоридом натрия (NaCl), фильтрации раствора для удаления осадка, центрифугирования отфильтрованного раствора для отделения супернатанта и нанесения его на колонку с сефакрилом S-200 или аффинным носителем, включающем маннозу.
Недостатком данного технического решения является узкая специфичность только к маннозе и гликопротеинам на ее основе, без способности распознавать разнообразные, в том числе кислые производные маннозы. К другим недостаткам аналога относится отсутствие противомикробной активности и отсутствие полной аминокислотной последовательности белка, для которого установлена ограниченной лишь структура N-концевого пептида.
Известен способ получения специфичного к кислому производному галактозы (галактозо-6-сульфату) лектина, включающий сложную многоэтапную биоинженерную схему. Содержащие галактозо-6-сульфат углеводы играют особую роль в злокачественных новообразованиях или дифференцировке клеток. В качестве зонда для выявления таких углеводов предложен модифицированный белок на основе EW29Ch-α - α-домена C-концевого функционального домена лектина, полученного из дождевого червя. Недостатком аналога в данном случае является чрезвычайно узкая специфичность и, соответственно, ограниченное применение полученного лектина(JP 2013039043).
Из наиболее близких изобретений, выбранных в качестве прототипа, следует назвать получение противовирусного манноз-специфичного лектина GLC, выделенного из морской красной водоросли Grateloupia chiangii (KR102099154B1, US20200157156A1). GLC имеет селективный индекс (selective index, SI) не менее 28 и 140 против вирусов простого герпеса HSV1 и HSV2 соответственно, молекулярную массу по данным электрофореза белков в денатурирующих условиях (SDS-PAGE) около 25 кДа, агглютинирующую активность в отношении эритроцитов лошади и специфичность к маннозе, фруктозе и фетуину по данным метода ингибирования реакции гемагглютинации. Также белок устойчив к широкому диапазону температур, сохраняя 50% от исходной активности после 30 минут инкубации при 40°С и 15% после 90°С, а также не меняет активности в присутствии Ca2+, Mg2+ или без бивалентных катионов. Методами молекулярной биологии мыла получена кодирующая структура GLC в виде кДНК длиной 900 п.н. с открытой рамкой считывания (open reading frame, ORF) в 696 п.н., на основании чего были определены полная аминокислотная последовательность (231 аминокислотный остаток), молекулярная масса (24,9 кДа) и теоретическая изоэлектрическая точка (pI 6,97). Очистку лектина производили из сырого экстракта с помощью аффинной хроматографии на носителе с D-маннозой. Из 30 г сухой массы водоросли Grateloupia chiangii было получено 0,65 мг очищенного GLC.
Согласно формуле изобретения антивирусный лектин из Grateloupia chiangii (GLC) получают следующим способом (1):
приготавливают грубый экстракт Grateloupia chiangii, являющейся красной водорослью;
проводят очистку лектина из Grateloupia chiangii (GLC)из грубого экстракта посредством афинной хроматографии на D-маннозе.
2. Другим существенным признаком изобретения-прототипа - лектина, веделенного из Grateloupia chiangii по способу (1), являются антивирусная активность, проявляемая против HSV-1 и HSV-2.
3. Для антивирусного лектина, выделенного и очищенного из Grateloupia chiangii установлена его аминокислотная последовательность, соответствующая молекулярной массе белка 25 кДа.
4. Для лектина из Grateloupia chiangii с аминокислотной последовательностью, заявленной в пункте (3), установлена последовательность азотистых оснований (нуклеотидов) кодирующего его гена.
К существенным недостаткам прототипа следует отнести:
1. Неспособность белка распознавать кислые производные галактозы и маннозы, распространенные в составе патоген-ассоциированных молекулярных паттернов, представленные уроновыми кислотами, их олигомерными и полимерными формами.
2. Отсутствие простого механизма регуляции активности белка, основанного на ассоциации/диссоциации двухвалентных катионов или иных кофакторов, изменение концентрации которых позволяет активировать\инактивировать углевод-связывающие свойства белка.
3. Отсутствие выраженной антибактериальной активности белка.
Указанные недостатки прототипа решаются следующим способом и характеризуются следующими существенными признаками заявляемого изобретения:
(1) в качестве источника белка с углевод-связывающей активностью используют другой биологический объект - морского двустворчатого моллюска Modiolus kurilensis, используя не всю его биомассу, а гемолимфу - ткань, обогащенную углевод-связывающим белком, проявляющем агглютинирующие свойства в отношении эритроцитов млекопитающих;
(2) для последующей очистки белка используют не грубый (тотальный) экстракт, а предварительно очищенную (обогащенную) фракцию, представляющую собой результат высаливания белка с целевой активностью с помощью двуступенчатого фракционирования сульфатом аммония, что обеспечивает более эффективную очистку белка, с увеличением скорости и степени чистоты препарата (не менее 99%);
(3) конечную очистку препарата осуществляют аффинной хроматографией на другом носителей - иммобилизованном гликане, включающем в качестве основных структур молекулярных паттернов разнообразные кислые галактаны и/или маннаны;
(4) способность белка связывать разнообразные производные галактозы и маннозы с преимущественным распознаванием кислых форм доказывается реакциями гемагглютинации и ее ингибирования соответствующими углеводами;
(5) способность в отличие от прототипа к антибактериальной активности, выражена в агглютинировании и подавлении роста разнообразных грамположительных и грамотрицательных микроорганизмов;
(6) в отличие от прототипа углевод-связывающий белок относится не к лектинам, а лектин-подобным белкам, содержащим C1q домен, характерный для компонентов системы комплемента, имеет уникальную, отличную от прототипа и других известных белков, аминокислотную последовательность.
Технической проблемой, поставленной перед данным изобретением является получение нового белка, способного преимущественно распознавать кислые производные галактозы и маннозы в составе разнообразных углеводных структур, разработка эффективного способа его выделения, установление его первичной структуры, физико-химических параметров регуляции его биологической активности, доказательство его антибактериальных свойств.
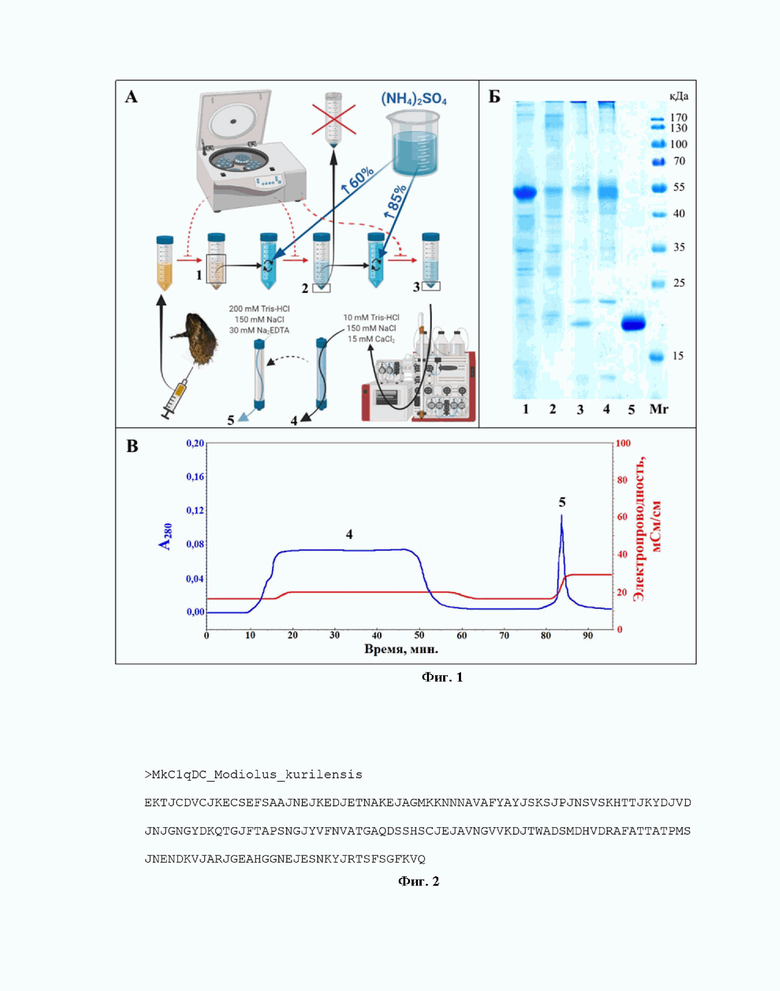
Техническим решением поставленной проблемы является выделение белка MkC1qDC из бесклеточной гемолимфы (плазмы) двустворчатого моллюска Мodiolus kurilensis (Bivalvia, Mytilidae; Bernard, 1983). Представители данного вида являются широко распространенными в северо-западной Пацифике и имеют небольшую глубину массовых поселений до 6 м, что делает его удобным для сбора объектом. Схема выделения включает три этапа и представлена на фиг. 1.
На первом этапе производят сбор гемолимфы из синуса заднего мускула-замыкателя одноразовыми стерильными шприцами, что позволяет получить в чистом виде значительные ее объемы нелетальным способом и, таким образом, производить повторные взятия после периода восстановления животных. Образцы гемолимфы до момента отделения клеток хранят в пробирках на ледяной бане (0 °C), что снижает активность гемоцитов, их дегрануляцию, агрегацию и разрушение, сохраняя состав плазмы в максимально нативном состоянии. Клетки отделяют путем центрифугирования при минимально травмирующих их условиях (12 мин при 800 g, 4 °C), что также предохраняет плазму от дополнительных загрязнений компонентами гемоцитов, включая опасные для белков протеолитические ферменты лизосом. Супернатант (плазму) отбирают в криопробирки, которые сразу же помещают для экспресс-заморозки в жидкий азот (-196 °С), что позволяет сохранить максимальную активность белков, с последующим переносом в низкотемпературную морозильную камеру (-80 °С) для длительного хранения, что позволяет собирать плазму в течение длительного времени без потери активности белков.
На втором этапе белки плазмы фракционируют высаливанием сульфатом аммония (СА). Сначала к плазме медленно приливали раствор насыщенного СА, забуференного Трис (рН 7,5) для предотвращения закисления раствора, опасного для белков, до 60 % насыщения СА при постоянном помешивании, после чего оставляют раствор без перемешивания на 30 мин для формирования белковых коагулятов. Все процедуры проводят на ледяной бане (0 °C) для замедления возможных ферментативных реакций, в том числе наиболее опасных - протеолитических. Затем раствор центрифугируют 30 мин при 3200 g, 0 °С. Полученный на данном этапе осадок, содержащий множество примесей (главным образом нецелевых белков). удаляют. К полученному супернатанту, содержащему целевые белки, снова приливают СА до 85 % насыщения раствора по описанной выше схеме для осаждения оставшихся белков, включая MkC1qDC. Полученный раствор вновь центрифугируют при тех же условиях и полученный осадок растворяют в TBS (Tris-buffered saline: 10 мМ Tris-HCl, 150 мМ NaCl, pH 7,5) с 15 мМ CaCl2.
На третьем этапе проводят аффинную хроматографию на пектин-сефарозе CL-4B, поскольку пектин имеет полигалактуронид в качестве центральной части молекулы, т.е. полимер на основе кислой галактозы. Уравновешивание колонки, нанесение образца и промывку колонки от не связавшихся белков проводят в присутствии ионов Ca2+ (15 мМ CaCl2), поскольку MkC1qDC имеет выраженную кальций-зависимость. По этой же причине элюцию проводят раствором, содержащим хелатирующий (связывающий бивалентные катионы) агент, а также с большой буферной емкостью для предотвращения изменения рН при изменении конформации белка в процессе потери им кофактора и отсоединения от лиганда: 200 мМ Tris-HCl, 150 мМ NaCl, 30 мМ Na2ЭДТА, рН 7,5. Нанесение образца проводят при небольшой скорости элюции 0,2 мл/мин для эффективного связывания MkC1qDC с носителем. Элюат, содержащий очищенный MkC1qDC, собирают и диализуют против пятидесятикратного объема TBS без добавок в три смены при 4 °С для удаления блокирующего активность хеланта.
Техническим результатом изобретения является получение нового углевод-связывающего белка с преимущественным аффинитетом к структурам гликанов, содержащим кислые производные галактозы и маннозы. Для подтверждения данного результата проводится оценка спектра углеводной специфичности белка методом реакции гемагглютинации и ее ингибирования растворами углеводов различного строения. В этом случае в качестве модельных частиц используют эритроциты, которые в случае наличия агглютинирующей способности белка, агрегируют и формируют характерную картину реакции. При добавлении в реакцию углеводов интересующей структуры при наличии аффинитета в ним наблюдается эффект отмены за счет конкурентного связывания активных центров белков углеводом. Важными техническими особенностями при проведении реакции является фиксация эритроцитов для избегания их спонтанного гемолиза, точная стандартизация концентрации их суспензии с помощью гемоцитометра и наличие в растворе ионов Ca2+ по причине кальций-зависимости MkC1qDC. Результат оценки специфичности нового белка данным подходом представлен в таблице (см. в графической части).
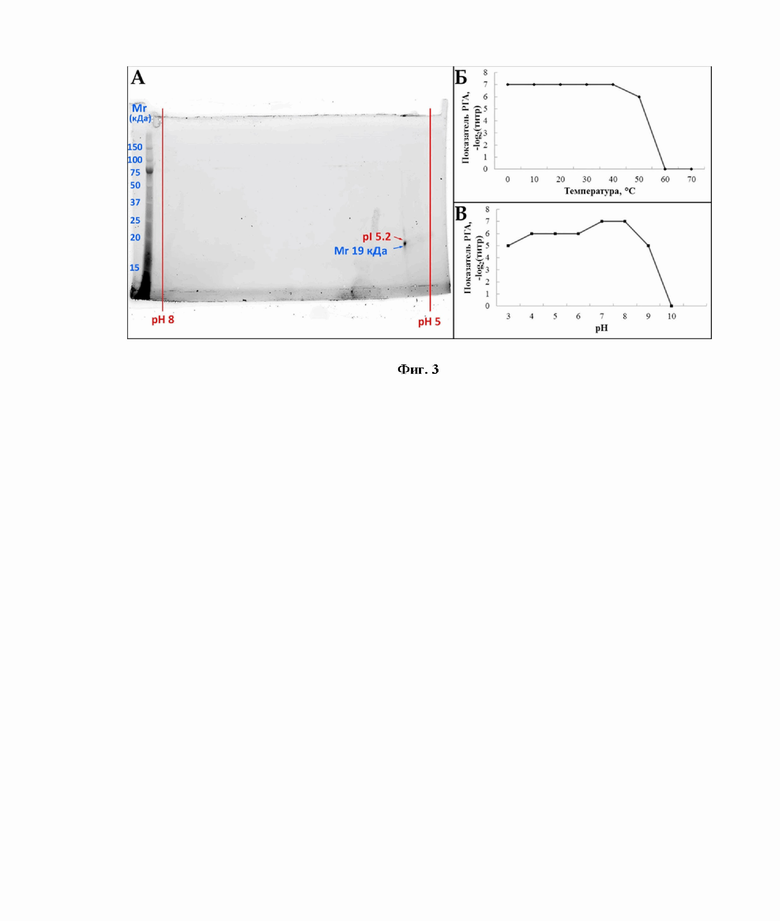
Другим техническим результатом является установление первичной структуры нового белка методами секвенирования его аминокислотной последовательности двумя независимыми подходами - N-концевым секвенированием фрагмента белка по Эдману и масс-спектрометрическим MS/MS анализом со сборкой полноразмерной первичной последовательности de novo. Совпадение результатов двух различных методических подходов в совокупности с повторяемостью результатов позволяют с высокой достоверностью установить структуру нового белка. Пять обособленных экспериментов N-концевого секвенирования методом Эдмана, проведенных на независимо полученных образцах очищенного MkC1qDC, и его масс-спектрометрический de novo анализ, в результате которого были получены пептидные фрагменты, совпадающие с результатами другого метода, позволили получить полную аминокислотную последовательность MkC1qDC (фиг. 2, SEQ ID NO.1), анализ которой с помощью помощью веб-инструмента «Basic Local Alignment Search Tool» (BLAST, https://blast.ncbi.nlm.nih.gov/Blast.cgi, доступ от 26 мая 2021 г.) показывает его уникальность, содержание консервативного C1q домена и высокую гомологию с C1qDC белками других представителей Bivalvia. Данные теоретического расчета молекулярной массы и изоэлектрической точки совпадают с фактическими значениями, полученными методом 2D-электрофореза очищенного белка (фиг. 3), что дополнительно подтверждает достоверность результатов.
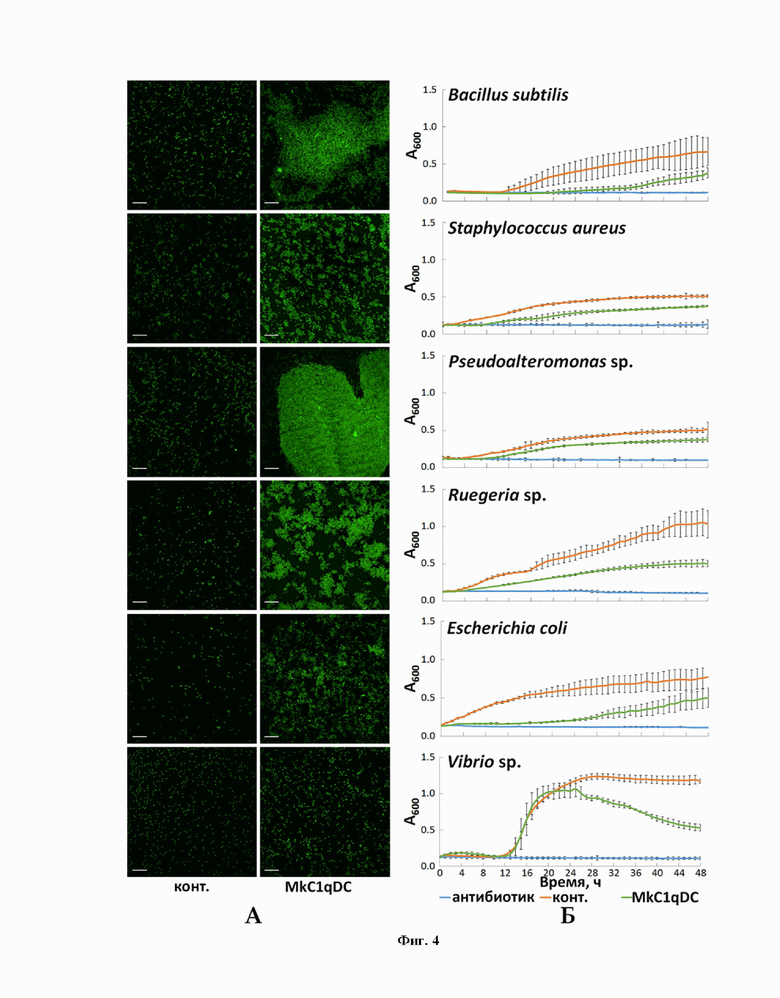
Еще одним техническим результатом изобретения является доказательство антимикробной активности MkC1qDC двумя методами. Первый основан на оценке способности агглютинировать белком термически инактивированные, фиксированные и флуоресцентно-окрашенные бактерии с визуализацией полученного результата с помощью флуоресцентной микроскопии. Второй метод заключается в оценке подавления роста живых бактериальных культур при их длительной инкубации с раствором белка. Динамику роста культур оценивали по их оптической плотности при длине волны 600 нм с использованием планшетного спектрофотометра в 6 независимых повторах. Результаты показали агглютинирующие и бактериостатические свойства MkC1qDC в отношении как грамотрицательных, так и грамположительных бактерий, что продемонстрировано на фиг.4.
Изобретение иллюстрируется следующими фигурами:
фиг. 1. Этапы выделения MkC1qDC. А - общая схема выделения; Б - электрофореграмма с образцами различных этапов выделения лектина; В - профиль элюции белков при выделении целевого лектина аффинной хроматографией. 1 - бесклеточная гемолимфа (плазма) Modiolus kurilensis; 2 - образец после высаливания плазмы раствором сульфата аммония со степенью насыщения 60 %; 3 - образец после второй ступени высаливания с доведением насыщения раствора сульфата аммония до 85 %; 4 - образец белков, не связавшихся с носителем в аффинной хроматографии; 5 - образец после аффинной хроматографии, снятый с носителя раствором, содержащим ЭДТА; Mr - стандарты молекулярных масс PageRuler™ Prestained Protein Ladder;
фиг. 2 показана полная аминокислотная последовательность MkC1qDC в формате «fasta»;
фиг. 3. Основные физико-химические свойства MkC1qDC. А - 2D-электрофорез очищенного MkC1qDC; Б - активность MkC1qDC после 1 ч инкубации при разных температурах; В - активность MkC1qDC после 1 ч инкубации при различных pH. РГА - реакция гемагглютинация; Mr - молекулярная масса; pI - изоэлектрическая точка;
фиг. 4. Антибактериальные свойства MkC1qDC (0,1 мг/мл). А - агглютинация фиксированных параформальдегидом FITC-меченых бактерий (масштабный отрезок - 10 мкм); Б - оптическая плотность живых бактериальных культур (среднее ± 95% доверительный интервал). А600 - оптическая плотность суспензий живых культур при длине волны 600 нм.
Таблица (см. в графической части). Углеводы, используемые в реакции ингибирования гемагглютинации с MkC1qDC и их концентрации полумаксимального ингибирования (IC50).
Изобретение иллюстрируется следующими примерами.
Пример 1
В результате выполнения всех описанных ниже этапов в среднем нам удавалось очистить 2-4 мг MkC1qDC из 1000 мл плазмы гемолимфы M. kurilensis.
На первом этапе у двустворчатых моллюсков вида Мodiolus kurilensis (Bivalvia, Mytilidae; Bernard, 1983) производили сбор гемолимфы в пробирки на ледяной бане (0 °C), которые затем центрифугировали в течение 12 мин при 800 g, 4 °C. Для последующей работы супернатант (плазму), полученную от разных особей, объединяли.
На втором этапе белки плазмы фракционировали высаливанием сульфатом аммония (СА). Сначала к плазме медленно приливали раствор насыщенного СА, забуференного Трис (рН 7,5), с перемешиванием на ледяной бане (0 °C) до 60 % насыщения СА, после чего оставляли без перемешивания при 0 °C на 30 мин для формирования белковых коагулятов и затем центрифугировали 30 мин при 3200 g, 0 °С. Осадок удаляли, к полученному супернатанту снова приливали СА до 85 % насыщения раствора, вновь центрифугировали при тех же условиях и полученный осадок растворяли в TBS (Tris-buffered saline: 10 мМ Tris-HCl, 150 мМ NaCl, pH 7,5) с 15 мМ CaCl2.
На третьем этапе проводили аффинную хроматографию. В качестве носителя использовали сефарозу CL-4B с ковалентно иммобилизованными дивинилсульфоном молекулами пектина. Колонку уравновешивали раствором TBS с 15 мМ CaCl2, образец наносили в том же растворе, и им же промывали тремя полными объемами колонки носитель от не связавшихся компонентов. Элюцию проводили раствором TBS большой буферной емкости, содержащим хелатирующий агент: 200 мМ Tris-HCl, 150 мМ NaCl, 30 мМ Na2ЭДТА, рН 7,5. Все этапы проводили при скорости элюции 0,2 мл/мин. Элюат, содержащий очищенный MkC1qDC, собирали и диализировали против пятидесятикратного объема TBS без добавок в три смены при 4 °С. Полученный раствор разделяли на аликвоты, содержащие по 100 мкг белка и хранили при -80 °С.
Описанная схема выделения представлена на фиг. 1А. На электрофореграмме с образцами различных этапов выделения MkC1qDC (фиг. 1Б) видно, что полоса с массой 19 кДа, соответствующей MkC1qDC, является единственной после последнего этапа очистки. При этом на хроматограмме выход белка виден в виде узкого пика повышения оптической плотности раствора при 280 нм при смене раствора элюата, заметного по увеличению его электропроводимости (фиг. 1В).
Пример 2
Проведение описанных ниже процедур позволяет определить структуру MkC1qDC в виде первичной последовательности белка, его предсказанной и фактической массы и изоэлектрической точки, необходимых для описания и подтверждения новизны выделенной молекулы.
N-концевое секвенирование очищенного MkC1qDC выполняли согласно методу деградации Эдмана с помощью прибора «Procise 492 Protein Sequencer» (Applied Biosystems), используя протокол производителя.
Для масс-спектрометрии MkC1qDC растворяли в 6 М гуанидин-HCl, 100 мМ Трис 10 мМ хлорацетамиде, 5 мМ Трис (2-карбоксиэтил) фосфин гидрохлорид, pH 8, нагревали при 95 °C в течение 5 мин. Образец разбавляли водой в 10 раз и добавляли трипсин/субстрат в соотношении 1/100. Ферментацию проводили при 37 °C в течение 4 ч. Далее раствор подкисляли до конечной концентрации 1% трифторуксусной кислоты, центрифугировали при 15000g, 10 мин, 23 °C и обессоливали. Пептиды анализировали на масс-спектрометре «Fusion Lumos» (Thermo Fisher), соединенном с насосом «RSLnano uHPLC» (Thermo Fisher). Пептиды разделяли на колонке «C18 Aurora» 15 см (IonOptiks), используя 40-минутный градиент от 5-35% ацетонитрила с 0,05% уксусной кислотой. Прибор работал в режиме «high/high» с разрешением 120k/35k (MS и MS/MS). Секвенирование de novo выполняли с использованием программного пакета «PEAKS» (Bioinformatics Solutions) с точностью масс 5ppm/0,01Да (МС и МС/МС). Были отобраны пептиды со средней локальной достоверностью (Average Local Confidence) >80%, которые с высокой степенью достоверности были собраны в более длинные участки с использованием ассемблера де Брюйна «ALPS», используя k-value равное 7, что привело к сборке 11 белковых последовательностей. Первоначальный поиск с помощью веб-инструмента «Basic Local Alignment Search Tool» (BLAST, https://blast.ncbi.nlm.nih.gov/Blast.cgi, доступ от 26 мая 2021 г.) в базе данных неизбыточных последовательностей Bivalvia выявил значительную гомологию с CBX41653.1. Поэтому данную последовательность использовали в качестве референса для сборки полной последовательности MkC1qDC с помощью веб-инструмента «Clustal Omega» (https://www.ebi.ac.uk/Tools/msa/clustalo/, доступ от 26 мая 2021 г.). Теоретическая молекулярная масса и pI собранного MkC1qDC были определены с помощью веб-инструмента «ExPASy» (http://web.expasy.org/compute_pi/, доступ от 26 мая 2021 г.).
2D-электрофорез проводили с изофокусированием в первом направлении с использованием коммерческих полимерных полосок (стрипов) с градиентом pH 4–7 "ReadyStrip™ IPG Strips" (Bio-Rad) и электрофорезом в градиентном 4–20 % полиакриламидном геле во втором направлении.
Полная последовательность MkC1qDC представлена на фиг. 2 и имеет длину в 176 аминокислот, предсказанную молекулярную массу 19181 Да и pI 5,2. Данные значения полностью соответствуют фактическим показателям, полученным в результате 2D-электрофореза и представленным на фиг. 3А.
Пример 3
Результатом выполнения всех описанных ниже процедур являлась максимальная биологическая активность MkC1qDC, проявляющаяся в виде агглютинации эритроцитов в модели реакции гемагглютинации (РГА), а также в виде связывания сахаров в модели ингибирования реакции гемагглютинации (ИРГА) с максимальным аффинитетом к кислым галактанам и маннанам.
РГА проводили в круглодонных 96-луночных иммунологических планшетах с эритроцитами человека группы крови 0. Предварительно эритроциты отмывали раствором PBS (Phosphate buffered saline: 10 мM Na2HPO4-КH2PO4, 137 мМ NaCl, 2,7 мМ KCl, pH 7,4) центрифугированием в течение 15 мин при 4 °С, 300 g до полного осветления супернатанта. Далее проводили фиксацию 0,25 % раствором глютарового альдегида на PBS в течение 30 мин, перемешивая аккуратным покачиванием, с последующей трехкратной отмывкой раствором PBS с центрифугированием при тех же условиях, нейтрализацией оставшихся альдегидных групп 1 % раствором глицина на PBS в течение 4 ч и трехкратными отмывками раствором TBS (Tris-buffered saline: 10 мМ Tris-HCl, 150 мМ NaCl, pH 7,5) с центрифугированием при тех же условиях. Непосредственно в реакции использовали суспензии эритроцитов с концентрацией 6×107 клеток/мл (подсчет проводили в камере Горяева) на растворе TBS с 15 мМ CaCl2.
В каждую лунку планшета помещали по 100 мкл TBS с 15 мМ CaCl2, в первый ряд дополнительно вносили по 100 мкл MkC1qDC с концентрацией 0,1 мг/мл на TBS с 15 мМ CaCl2 и получали в последующих рядах серию двукратных разведений (от 1/2 до 1/2048), оставляя последний ряд без образца для отрицательного контроля, и далее добавляли 50 мкл суспензии эритроцитов. Результаты РГА визуально фиксировали через 2 ч инкубации при 23 °С. Полученный титр использовали для получения активных разведений на следующем этапе постановки ИРГА с целью оценки концентрации полумаксимального ингибирования (half maximal inhibitory concentration, IC50).
ИРГА также проводили в круглодонных 96-луночных иммунологических планшетах с суспензиями эритроцитов, полученных описанным выше способом. В каждую лунку планшета помещали по 50 мкл TBS с 15 мМ CaCl2, в первый ряд дополнительно вносили по 50 мкл раствора одного из сахаров на TBS с 15 мМ CaCl2 и получали в последующих рядах серию их двукратных разведений, оставляя последний ряд без углеводов для отрицательного контроля. Далее во лунки вносили по 50 мкл раствора MkC1qDC на TBS с 15 мМ CaCl2 в концентрации, соответствующей (титр)×4 на основании результатов РГА. Планшеты инкубировали 30 мин при 23 °С, после чего добавляли суспензии эритроцитов. Результаты ИРГА визуально фиксировали через 2 ч инкубации при 23 °С. За IC50 считали последнее разведение углевода, которое вызывало эффект отмены агглютинации.
Полученные результаты представлены в таблице. Из 23 моносахаридов, протестированных методом ИРГА, только шесть были ингибирующими. L-гулоза, сиаловая (N-ацетилнейраминовая), D-галактуроновая и D-глюкуроновая кислоты показали самый высокий ингибирующий эффект среди моносахаридов IC50 в диапазоне 1,16-1,46 мг/мл. D-галактоза и 2-дезокси-D-галактоза были эффективны в диапазоне 2,46-5,4 мг/мл. Кроме того, дисахариды, такие как 2α-маннобиоза и D-лактоза, были отмечены как ингибирующие молекулы, показывающие IC50, начиная с 5,13 мг/мл и выше. Маннан из Saccharomyces cerevisiae, LPS из Escherichia coli, PDG из Staphylococcus aureus и муцин типа II из желудка свиньи были умеренно ингибирующими при концентрациях на три порядка выше, чем для наиболее эффективных сахаридов (0,10-0,49 мг/мл). Альгинат, κ-каррагинаны, фукоидан и пектин были высокоэффективными ингибиторами с самым низким значением IC50 (менее 0,002 мг/мл).
Пример 4
Результаты описанных ниже процедур описывают активность MkC1qDC после воздействия на него различных pH и температур и, таким образом, являются обоснованием возможности выполнения приведенных в Примерах 1 и 3 процедур при различных значениях данных параметров.
Определение рН-зависимости MkC1qDC проводили методом РГА по описанной в Примере 1 методике на TBS с повышенной буферной емкостью (50 мМ Tris-HCl, 150 мМ NaCl, 15 мМ CaCl2, рН 7,5). В качестве образцов использовали аликвоты очищенного, диализированного против деионизированной воды и лиофильно высушенного лектина, которые предварительно растворяли в концентрации 0,1 мг/мл и инкубировали 1 ч в следующих 10 мМ буферных растворах, содержащих 150 мМ NaCl: pH 3 - глицин-HCl, pH 4 и pH 5 - CH3COOH-CH3COONa, pH 6 и pH 7 - (CH3)2AsO2Na-HCl, pH 8 - трис-HCl, pH 9 - глицин-NaOH, pH 10 - NaHCO3-Na2CO3.
Определение термолабильности MkC1qDC также проводили методом РГА по описанной в Примере 1 методике. Для этого отдельные аликвоты MkC1qDC с концентрацией 0,1 мг/мл в TBS (10 мМ Tris-HCl, 150 мМ NaCl, 15 мМ CaCl2, рН 7,5) предварительно инкубировали по 1 ч при температурах 0 °C, 10 °C, 20 °C, 30 °C, 40 °C, 50 °C, 60 °C, 70 °C с последующим доведением до 23 °С.
Очищенный MkC1qDC обладал высокой агглютинирующей активностью после инкубации в течение 1 ч при температурах 0-40 °С, которая немного снижалась при 50 °С и полностью исчезала при 60 °С и выше (фиг. 3Б). В то же время максимальная активность была обнаружена через 1 ч инкубации в диапазоне pH 7-8 (фиг. 3В). Повышение pH до 9 приводило к небольшому снижению активности и полной ее потере при pH 10. Кислые растворы до pH 3 оказывали лишь слабое влияние на титры РГА, что указывает на широкий диапазон pH-стабильности MkC1qDC.
Таким образом оптимальными условиями для выделения и работы MkC1qDC являются рН растворов от 7 до 8 и их температура от 0 °C до 40 °C. Однако возможны процедуры выделения и оценки активности при рН 3-9 и температурах 0-50 °C.
Пример 5
Приведенные ниже процедуры и их результаты являются обоснованием возможности применения MkC1qDC в качестве антибактериального агента широкого спектра воздействия.
Антимикробные свойства MkC1qDC проверяли в отношении условно патогенных грамотрицательных бактерий (Vibrio sp., Ruegeria sp., Escherichia coli, Pseudoalteromonas sp.) и грам(+) бактерий (S. aureus, Bacillus subtilis), выделенных из морских организмов.
Для оценки агглютинирующей активности MkC1qDC бактерий термически инактивировали нагреванием до 72 °С в течение 1 ч. Далее суспензии отмывали тремя циклами центрифугирования (3000 g, 20 мин, 4 °С) и ресуспендирования в растворе HBS (HEPES buffered saline: 10 мМ HEPES-NaOH, 150 мМ NaCl, pH 7,4). Осадки бактерий тщательно ресуспендировали, чтобы разбить крупные конгломераты на отдельные клетки, и фиксировали 4 % раствором параформальдегида (ПФА) на HBS при постоянном покачивании, после чего снова трижды отмывали раствором HBS. Далее проводили окраску в течение 2 ч флуоресцентным красителем флуоресцеин-5-изотиоцианатом (0,5 мг/мл fluorescein isothiocyanate (FITC), 0,1 М NaHCO3-Na2CO3, рН 9,3) при постоянном помешивании, после чего трижды отмывали раствором HBS от не связавшегося красителя. Реакцию агглютинации осуществляли в микропробирках на 1,5 мл в объеме реакционной смеси 200 мкл при постоянном перемешивании и конечной концентрации компонентов: суспензии бактерий с оптической плотностью A600 = 1,5, 65 мкг/мл MkC1qDC, 10 мМ HEPES-NaOH, 150 мМ NaCl, 50 мM CaCl2, pH 7,5. В качестве негативного контроля использовали смесь того же состава, но без MkC1qDC. После инкубирования суспензии наносили на предметные стекла и после высушивания заключали в среду «Sub-X» (Leica Biosystems) под покровные стекла. Визуализацию проводили, используя объективы с увеличением 100х, на системе лазерного имиджинга на базе микроскопа «FluoView FV1200MPE» (Olympus), получая серию изображений по оси Z с последующим объединением для создания Z-проекции.
Для оценки влияния MkC1qDC на бактериальный рост культуры выращивали до log-фазы (фазы роста) в жидкой среде Йошимицу-Кимура на основе морской воды, после чего разводили в жидкой среде PBM (Poor broth medium: 1,5 % пептон, 1,5 % NaCl, pH 7,2) до A600 = 0,05 (оптическая плотность при длине волны 600 нм) в кварцевой кювете с длиной оптического пути 10 мм и вносили по 50 мкл в лунки 96-луночного плоскодонного планшета. В экспериментальные лунки дополнительно вносили по 50 мкл раствора MkC1qDC с концентрацией 0,4 мг/мл на PBM, в положительный контроль по 50 мкл PBM без добавок, в отрицательный - 50 мкл PBM с пенициллином (2000 ед/мл) и стрептомицином (2 мг/мл). Для каждого типа контрольных и экспериментальных образцов использовали по 6 лунок для каждого вида бактерий. Результаты ингибирования бактериального роста измеряли при помощи планшетного ридера «Cytation 5 imaging reader» (BioTek Instruments) с использованием программы анализа данных «Gen 5». Оптическую плотность растворов в лунках измеряли каждый час в течение 48 ч при длине волны 600 нм в режиме «Endpoint» (измерение по конечной точке) при температуре инкубации 37 °C с перемешиванием аппаратным встряхиванием перед каждым измерением.
MkC1qDC показал антибактериальную активность в отношении всех использованных бактерий (фиг. 4). В то же время наиболее эффективно агглютинация протекала у Pseudoalteromonas sp. и B. subtilis, которые в присутствии MkC1qDC образовывали крупные конгломераты, в отличие от Vibrio sp., которые были объединены в небольшие разреженные группы по 4-15 клеток. Кроме того, MkC1qDC проявлял бактериостатические свойства по отношению к большинству вышеперечисленных штаммов, со значимым подавлением роста культур через 4-12 ч (p <0,05). Единственным исключением являлись культуры Vibrio sp., уменьшение плотности которых в присутствии MkC1qDC происходило только через 26 ч культивирования (p <0,05), когда контрольные культуры уже находились в стационарной фазе (фиг. 4).
--->
<?xml version=”1.0” encoding=“UTF-8”?>
<!DOCTYPE ST26SequenceListing PUBLIC “-//WIPO//DTD Sequence Listing
1.3//EN” “ST26SequenceListing_V1_3.dtd”>
<ST26SequenceListing originalFreeTextLanguageCode="ru"
dtdVersion="V1_3" fileName="MkC1qDC aa" softwareName="WIPO Sequence"
softwareVersion="2.3.0" productionDate="2024-06-20">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2021139868</ApplicationNumberText>
<FilingDate>2021-12-30</FilingDate>
</ApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
автономное образовательное учреждение высшего образования
"Дальневосточный федеральный университет" (ДВФУ)</ApplicantName>
<ApplicantNameLatin>Federalnoe gosudarstvennoe avtonomnoe
obrazovatelnoe uchrezhdenie vysshego obrazovaniia "Dalnevostochnyi
federalnyi universitet" (DVFU)</ApplicantNameLatin>
<InventorName languageCode="ru">Кумейко Вадим
Владимирович</InventorName>
<InventorNameLatin>Kumeiko Vadim Vladimirovich</InventorNameLatin>
<InventionTitle languageCode="ru">Углевод-связывающий белок с
антибактериальными свойствами, распознающий кислые галактаны и
маннаны, и способ его выделения</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>176</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..176</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Modiolus kurilensis</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>EKTJCDVCJKECSEFSAAJNEJKEDJETNAKEJAGMKKNNNAVAFYAYJSKS
JPJNSVSKHTTJKYDJVDJNJGNGYDKQTGJFTAPSNGJYVFNVATGAQDSSHSCJEJAVNGVVKDJTWA
DSMDHVDRAFATTATPMSJNENDKVJARJGEAHGGNEJESNKYJRTSFSGFKVQ</INSDSeq_sequen
ce>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЦЕНКИ ЗДОРОВЬЯ МОРСКИХ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ И СОСТОЯНИЯ СРЕДЫ ИХ ОБИТАНИЯ | 2014 |
|
RU2571817C1 |
| РЕКОМБИНАНТНЫЙ МАННАН-СВЯЗЫВАЮЩИЙ ЛЕКТИН ЧЕЛОВЕКА | 2000 |
|
RU2292217C2 |
| ПОЛИСАХАРИД И ЕГО ПРОИЗВОДНЫЕ С АКТИВНОСТЬЮ К ФИКОЛИНУ-3, СПОСОБ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2011 |
|
RU2575752C2 |
| Способ определения активности маннан-связывающих лектин-ассоциированных сериновых протеаз в тесте коагуляции фибриногена | 2019 |
|
RU2739113C1 |
| ЛЕКТИН ROBINIA PSEUDOACACIA И ЕГО ПРИМЕНЕНИЕ | 1998 |
|
RU2214259C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТИТЕЛЬНОГО ЛЕКТИНА ML1 ИЗ ОМЕЛЫ БЕЛОЙ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ (ВАРИАНТЫ). | 2016 |
|
RU2644332C1 |
| РЕКОМБИНАНТНЫЙ ЛЕКТИН ОМЕЛЫ БЕЛОЙ (RML) | 1996 |
|
RU2241750C2 |
| ЭКСПРЕССИРУЮЩАЯ КОНСТРУКЦИЯ, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР PEDII CS150 И СПОСОБ ПРОДУЦИРОВАНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА | 1999 |
|
RU2241035C2 |
| Способ получения и определения специфической активности фактора активации макрофагов GcMAF | 2023 |
|
RU2823530C1 |
| Определение активности классического пути системы комплемента в тесте коагуляции фибриногена | 2019 |
|
RU2707568C1 |

Изобретения относятся к области биотехнологии. Предложены углевод-связывающий белок C1q-домен-содержащий белок из морского двустворчатого моллюска Modiolus kurilensis (MkC1qDC) с аминокислотной последовательностью SEQ ID NO.1 и способ его выделения. Получают гемолимфу из морского двустворчатого моллюска Modiolus kurilensis. Отделяют плазму от клеточной фракции. Получают обогащенную фракцию из бесклеточной гемолимфы методом высаливания сульфатом аммония. Полученная фракция обладает наибольшей гемагглютинирующей активностью, эффективно ингибируемой кислыми производными галактозы и маннозы, включая их олигомерные и полимерные формы. Затем выделяют очищенный углевод-распознающий белок MkC1qDC методом аффинной хроматографии с иммобилизованными гликанами, содержащими кислые производные галактозы или маннозы. Углевод-связывающий белок MkC1qDC из двустворчатого моллюска Modiolus kurilensis проявляет антибактериальные свойства в отношении грамположительных и грамотрицательных микроорганизмов. 2 н.п. ф-лы, 4 ил., 1 табл., 5 пр.
1. Способ выделения углевод-связывающего C1q-домен-содержащего белка с аминокислотной последовательностью SEQ ID NO.1 из морского двустворчатого моллюска Modiolus kurilensis (MkC1qDC), заключающийся в получении гемолимфы из морского двустворчатого моллюска Modiolus kurilensis с последующим отделением плазмы от клеточной фракции; получение обогащенной фракции из бесклеточной гемолимфы методом высаливания сульфатом аммония, которая обладает наибольшей гемагглютинирующей активностью, эффективно ингибируемой кислыми производными галактозы и маннозы, включая их олигомерные и полимерные формы; с последующим выделением очищенного углевод-распознающего белка методом аффинной хроматографии с иммобилизованными гликанами, содержащими кислые производные галактозы или маннозы.
2. Углевод-связывающий белок MkC1qDC с аминокислотной последовательностью SEQ ID NO.1 из двустворчатого моллюска Modiolus kurilensis, полученный способом по п.1, проявляющий антибактериальные свойства в отношении грамположительных и грамотрицательных микроорганизмов.
| СПОСОБ ПОЛУЧЕНИЯ УГЛЕВОД-БЕЛКОВОГО КОМПЛЕКСА ИЗ ПРИМОРСКОГО ГРЕБЕШКА | 1996 |
|
RU2121844C1 |
| Приспособление для заливки выводных конусов в аккумуляторах | 1934 |
|
SU41653A1 |
| GERDOL M | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Developmental & Comparative | |||

