Изобретение относится к синтезу белков и может быть использовано для получения и определения специфической активности фактора активации макрофагов GcMAF, который относится к Gc-белкам и является носителем витамина D, который также нейтрализует G-актин, выделяющийся при клеточном лизисе.
Известен способ получения Gc-белка (DBP), который является исходным материалом фактора активации макрофагов GcMAF (DBP, предшественник), согласно которому предшественник витамина D-связывающего белка (DBF) выделяют из плазмы крови человека на хроматографическом субстрате с аффинным сорбентом 25-hydroxyvitamin D3-Sepharose[1].
Недостатком способа является его относительно высокая сложность.
Известен также способ получения фактора активации макрофагов (GcMAF), так называемого GcMAF второго поколения, согласно которому осуществляют обработку плазмы крови здоровых доноров ферментами β-галактозидазой и сиалидазой, сорбированными на твердом носителе, причем, считается, что поскольку в плазме крови находится до 500 мг/л DBP предшественника, то этого количества достаточно для клинического применения обогащенной активированным GcMAF плазмы [2].
Однако такая субстанция не является высокоспециализированным фармацевтическим препаратом и не поддается фармакологической стандартизации.
Известен и способ получения фактора активации макрофагов GcMAF, так называемого GcMAF третьего поколения [3], однако, полученное вещество не является истинно GcMAF, а представляет собой молозиво крупного рогатого скота, в котором содержится определенное количество белков, не имеющих онтологического отношения к активатору макрофагов, которые несут сайт гликозилирования GalNac/сиаловая кислота/галактоза (по-видимому, в различных комбинациях).
Поэтому этот препарат по своим молекулярным характеристикам является малоэффективным и не может выполнять функцию специфического активатора макрофагов, направляющего синтез про- или противовоспалительных цитокинов.
Наиболее близким к предложенному является способ получения и фармацевтическая композиция для применения при заживлении ран или лечении аллергических заболеваний, аутоаллергических заболеваний, побочных реакций, вызванных терапевтическими агентами, или ангиогенных заболеваний [4], содержащая один или множество дегалактозилированных Gc глобулинов человека, которые выбраны из группы, состоящей из (i)-(iii):
(i) дегалактозилированный Gc-глобулин, который получают обработкой Gc-глобулина подтипа If и/или подтипа Is, полученного из плазмы или сыворотки крови человека, только β-галактозидазой;
(ii) дегалактозилированный Gc-глобулин человека, содержащий аминокислотную последовательность, как показано в следующем идентификаторе №1, и имеющий структуру сахарной цепи, в которой N-ацетилгалактозамин связан с треонином либо в положении 418, либо в положении 420 аминокислотной последовательности, как показано в следующем идентификаторе №1, и сиаловая кислота дополнительно связывается с N-ацетилгалактозамином;
(iii) дегалактозилированный Gc-глобулин человека, содержащий аминокислотную последовательность, как показано в SEQ ID NO: 2, и имеющий структуру сахарной цепи, в которой N-ацетилгалактозамин связан с треонином либо в положении 418, либо в положении 420 аминокислотной последовательности, как показано в SEQ ID NO: 2, а а-манноза является далее связывается с N-ацетилгалактозамином.
Таким образом, в наиболее близком техническом решении получение фактора активации макрофагов GcMAF включает выделение предшественника DBP из плазмы крови человека на хроматографическом субстрате с последующей элюцией белка в раствор и ферментативной обработкой β-галактозидазой, фиксированной на твердом носителе или активированными лейкоцитами или кондиционной средой от активированных лейкоцитов.
Недостатком наиболее близкого технического решения является относительно низкая эффективность применения полученного продукта, вызванная относительно узкой специфической активностью применения. Это вызвано тем, что для создания высокоспецифичного фармацевтического препарата полученный указанными способом фактор активации макрофагов GcMAF требует дополнительной хроматографической очистки. Кроме того, для конвертации DBP в GcMAF используется только фермент β-галактозидаза, что создает условия для активации только одного вектора воспаления (провоспалительного) макрофагального звена иммунитета. И, наконец, согласно известному способу, не предусмотрена оценка направления воспалительного ответа макрофагов, что является абсолютно необходимой частью создания высокоспецифичного лекарственного средства.
Задачей изобретения является разработка способа получения фактора активации макрофагов GcMAF, обладающего более широкой областью применения и повышенной эффективностью применения.
Требуемый технический результат заключается в расширении области применения и повышении эффективности применения.
Поставленная задача решается, а требуемый технический результат достигается тем, что в способе получения и определения специфической активности фактора активации макрофагов GcMAF, основанного на выделении, конвертации и определении специфической активности, согласно изобретению, выделение и конвертация фактора активации макрофагов GcMAF осуществляют на оригинальном твердом носителе Sephacril/25-OH D3 в условиях аффинной сорбции фактора активации макрофагов на частицах смолы, для его валидации используют специфические антитела и матричную рибонуклеиновую кислоту (мРНК), синтез которой активируется фактором активации макрофагов GcMAF в перитонеальных макрофагах (ПМФ) мышей линии C57BL/6, набор специфических праймеров и процедуру полимеразной цепной реакции (ПЦР) в реальном времени, при этом, в качестве исходного сырья для получения активатора макрофагов GcMAF используют смесь плазм доноров, разбавленную в половину физиологическим раствором или фосфатно-солевым буфером, а стандартизацию фактора активации макрофагов GcMAF проводят путем выделения фактора на оригинальном Sephacril/25-OH D3 субстрате, конвертации DBP в фактор активации макрофагов GcMAF в связанном с субстратом Sephacril/25-OH D3 состоянии, использовании для определения активности каждого выделения препарата клеточной системы ПМФ мышей линии C57BL/6 и определении способности DBP и GcMAF индуцировать синтез мРНК ПМФ четырех цитокинов -медиаторов воспаления TNF-α и IL-1β и противовоспалительных цитокинов TGF-β и IL-10, при этом, при конвертации используют очищенные ферменты β-галактозидазу и сиалидазу, присутствующие в конвертационной среде, а способность ПМФ синтезировать цитокины определяют с использованием процедуры ПЦР в реальном времени, где в качестве специфических праймеров используют праймеры на провоспалительные цитокины TNF-α и IL-β и противовоспалительные цитокины TGF-β и IL-10.
Ниже приводятся теоретические и экспериментальные данные, а также методики, подтверждающие, что изобретение отвечает критериям новизны и промышленной применимости изобретения.
Изобретение иллюстрируется чертежами.
На фиг. 1 - представлена структурно-функциональная характеристика DBP, где А - полиакриламидный гель с образцами DBP, полученными аффинной хроматографией (слева) с использованием хитин-актиновой колонки (1) и колонки с 25-ОН-Д3/Сефароза (2) и вестерн-блоттинга этих образцов с антителами против Gc-группы (справа); В - Полиакриламидные гели с образцами DBP, полученными с помощью двух последовательных хроматографий: 25-ОН-Д3/Сефароза//хитин-актин (1-3) и хитин-актин//25-ОН-Д3/Сефароза (4-6); проскоки после второй хроматографии (1 и 5); образцы DBP, очищенные с помощью двойной последовательной хроматографии (2 и 4); исходные образцы DBP, полученные первой хроматографией (3 и 6); М -молекулярный белковый маркер "The Thermo Scientific™ Page Ruler™ Prestained protein Ladder" (Thermo Fisher Scientific Inc., USA).
На фиг. 2 - представлена количественная оценка экспрессии мРНК генов цитокинов в перитонеальных макрофагах мышей, обработанных лейкоцитами и DBP, полученными от разных доноров, где А - экспрессия мРНК генов цитокинов в перитонеальных макрофагах мышей, обработанных лейкоцитами в дозе 1х, 10х и 100х по сравнению с контролем (ПМФ, необработанные лейкоцитами), чей уровень экспрессии был принят за 1 (красная сплошная линия); В-Е - экспрессия мРНК генов цитокинов в ПМФ, обработанных DBP, полученных от разных доноров (В - DBP LEV; С - DBP 1; D - DBP2 и Е - DBP3), в дозах 0,02 мкг, 0,2 мкг и 2 мкг по отношению к контролю (ПМФ, необработанные DBP), уровень экспрессии которого был принят за 1 (красная сплошная линия). Приведены медианные значения, межквартильный диапазон, а также минимум-максимум диапазон.
На фиг. 3 - представлено влияние различных образцов GcMAF полученных от одного донора (LEV), на экспрессию цитокинов клетками культуры макрофагов J774 и ПМФ в условиях ex vivo и in vivo, где А-С -экспрессия мРНК генов цитокинов в клетках J774 и в перитонеальных макрофагах мышей, обработанных С GcMAF LEV L pure 37°C (A - J774 cells, ex vivo; В - перитонеальные макрофаги, ex vivo; С - перитонеальные макрофаги, in vivo), дозами 0,02 мкг, 0,2 мкг, 2 мкг и 10-20 мкг по отношению к контролю (ПМФ, необработанные GcMAF), уровень экспрессии которого был принят за 1 (красная сплошная линия); D - экспрессия мРНК генов цитокинов в перитонеальных макрофагах мышей, обработанных ex vivo Ex GcMAF LEV L pure LysoPC 37°C дозами 0,02 мкг, 0,2 мкг и 2 мкг по отношению к контролю (ПМФ необработанные), уровень экспрессии которого был принят за 1 (красная сплошная линия); Е, F - Оценка фагоцитарной активности перитонеальных макрофагов, обработанных С GcMAF LEV L pure 37°C, по их способности интернализовать металлические частицы. Е -изображения частиц, фагоцитированных нативными макрофагами (контроль) и макрофагов, обработанных С GcMAF LEV L pure 37°C; F - количественная оценка фагоцитарной активности перитонеальных макрофагов (IBN), обработанных С GcMAF LEV L pure 37°C в дозах 0,2 мкг, 0,5 мкг и 1 мкг.Фагоцитарную активность нативных макрофагов принимали за 1 (красная сплошная линия). G - Сопоставление фагоцитарной активности (пунктирная линия) и уровня синтеза мРНК противовоспалительных цитокинов (куполообразная кривая) мышиными перитонеальными макрофагами, активированными разными дозами С GcMAF LEV L pure 37°C: 0,02 мкг, 0,2 мкг и 1-2 мкг.
На фиг. 4 - представлено влияние различных образцов GcMAF, полученных от разных доноров и разными способами, на синтез мРНК цитокинов ПМФ мышей, а также на фагоцитарную активность ПМФ мыши, где А-С экспрессия мРНК генов цитокинов в перитонеальных макрофагах мышей, обработанных GcMAF, полученных от разных доноров и разными способами (А - Ex GcMAF 1 L pure LysoPC 37°С; В - Ex GcMAF 1 L plasma 39°C; С - С GcMAF2 L plasma 39°C) дозами 0,02 мкг, 0,2 мкг и 2 мкг по отношению к контролю (ПМФ необработанные), уровень экспрессии которого был принят за 1 (красная сплошная линия). D, Е - оценка фагоцитарной активности перитонеальных макрофагов, обработанных С GcMAF2, по их способности интернализовать металлические частицы. D - изображения частиц, фагоцитированных нативными макрофагами (контроль) и макрофагами, обработанными С GcMAF2; Е - количественная оценка фагоцитарной активности перитонеальных макрофагов (IBN), обработанных DBP, С GcMAF2, Ex GcMAF 1 в дозах 0,2 мкг и 2 мкг, а также LPS. В качестве контроля использовали нативные макрофаги, фагоцитарную активность которых принимали за 1 (красная сплошная линия). F - Сопоставление фагоцитарной активности и уровня синтеза мРНК провоспалительных цитокинов мышиных ПМФ, активированных С GcMAF2 дозами 0,02 мкг, 0,2 мкг и 2 мкг; G-I -экспрессия мРНК генов цитокинов в перитонеальных макрофагах мышей, обработанных GcMAF3, полученного от одного и того же DBP3, но разными способами (G - Ex GcMAF3 L plasma 39°С; Н - Ex GcMAF3 L plasma 37°C; I - Ex GcMAF3 L plasma LysoPC 37°C) дозами 0,02 мкг, 0,2 мкг и 2 мкг по отношению к контролю (ПМФ необработанные), уровень экспрессии которого был принят за 1 (красная сплошная линия).
На фиг. 5 - представлены результаты флуоресцентно-цитологического анализа взаимодействия ПМФ и Су 5-GcMAF, присутствующего в инкубационной среде в различных дозах, где А - общий вид ПМФ, инкубируемых в присутствии трех доз Cy5-GcMAF: 0,02 мкг, 0,2 мкг, 20 мкг.В - ПМФ, инкубированные в присутствии 0,02 мкг лиганда. Незначительное количество Cy5-GcMAF распределено в цитоплазме ПМФ. С - ПМФ, инкубированные в присутствии 0,2 мкг лиганда. Меченый материал Су5-GcMAF четко детектируется в цитоплазме ПМФ. D - ПМФ, инкубированные в присутствии 2-20 мкг лиганда, где детектируется два варианта распределения меченного материала Су5-GcMAF: Су5-GcMAF занимает большую часть внутреннего пространства клетки (1); Cy5-GcMAF расположен на внешней поверхности клетки в форме четко оформленного пятна (2-4).
На фиг. 6 - представлены результаты анализа взаимодействия GcMAF и CLEC10A, для каждого блока приведены окраска Кумасси и вестерн-блот, где А - взаимодействие белка CLEC10A и антител к CLEC10A в вестерн-блот анализе. Четко визуализируются высокомолекулярные дериваты белка CLEC10A. В - вестерн-блот анализ взаимодействия белков лизата ПМФ и белка CLEC10A с антителами к CLEC10A. С - вестерн-блот анализ перекрестного взаимодействия антител к GcMAF и белка CLEC10A. D -анализ прямого взаимодействия GcMAF и CLEC10A в cross-link экспериментах с использованием парафармальдегида в качестве кросслинкирующего агента и антител к CLEC10A. На имидже вестерн блота отчетливо видно отсутствие перекрестной гомологии между GcMAF и антителами к CLEC10A (линия 3). Стрелкой указан продукт кросслинка, по молекулярной массе соответствующий комплексу GcMAF/CLEC10A.
На фиг. 7 - результаты анализа прямого взаимодействия GcMAF и CLEC10A, где А - сэндвич анализ прямого взаимодействия фракций белков, выделяемых с 25-ОН Sepharose - DBP и аполипопротеина, а также GcMAF этого же образца с белком CLEC10A. Окраска Кумасси и вестерн-блот.Для анализа взят образец GcMAF3. Результат свидетельствует, что и DBP, и GcMAF3 прямо связываются с CLEC10A. Аполипопротеин прямо не взаимодействует с CLEC10A; В - сэндвич анализ, демонстрирующий необходимость ионов Са2+ для прямого взаимодействия высокомолекулярных дериватов CLEC10A и GcMAF. Результат свидетельствует, что присутствие 20 мМ Са2+ в инкубационной системе вестерн-блота высокомолекулярный дериват CLEC10A (-65 кДа) прямо связывается с белком GcMAF. Белок CLEC10A не связывается с предшественником DBP при выбранных концентрациях Са2+. Стрелкой отмечен специфический сигнал. Для анализа взят образец GcMAF3; С - анализ влияния температуры на прямое взаимодействие белков CLEC10A и GcMAF. Сэндвич анализ. Окраска Кумасси и вестерн-блот.Стрелками отмечен специфический сигнал. Для анализа взят образец GcMAF3; D - аватар графика синтеза мРНК анализируемых цитокинов ПМФ, обработанных GcMAF 1 (фиг.4, А) свидетельствующий, что провоспалительный ответ ПМФ зависит от связывания GcMAF с дериватом CLEC10A 65 к Да; Е - сэндвич-блот анализ взаимодействия CLEC10A и GcMAF в условиях конкурентного присутствия свободного GalNac. Результаты свидетельствуют, что при количестве 10 мкг свободного GalNac в инкубационной системе вестерн-блота происходит разрушение связи 65 кДа CLEC10A деривата и GcMAF. Стрелкой указано разрушение специфического сигнала. Для анализа взят образец GcMAF3; F - анализ перекрестного взаимодействия свободного GalNac и Gc антител. Стрелкой указан специфический сигнал перекрестного взаимодействия; G - in vivo анализ синтеза ПМФ мРНК анализируемых цитокинов в условиях конкурентного взаимодействия GcMAF и свободного GalNac. Для анализа взят образец GcMAF3; Н - сэндвич анализ прямого взаимодействия CLEC10A и двух вариантов GcMAF, характеризующихся противоположной способностью активировать ПМФ к синтезу мРНК про- и противовоспалительных цитокинов. Стрелками отмечены изменения во взаимодействии высокомолекулярных дериватов CLEC10A 63 кДа и 65 кДа с GcMAF. В варианте с использованием GcMAF с более выраженными противовоспалительными свойствами (GcMAF3 L plasma LysoPC 37°С) специфически взаимодействует с GcMAF преимущественно высокомолекулярный CLEC10A дериват 63 кДа; I - аватары графиков синтеза мРНК анализируемых цитокинов ПМФ, обработанных С GcMAF LEV L pure 37°C и GcMAF3 L plasma LysoPC 37°C (фиг. 3, В и фиг. 4, А, соответственно).
На фиг. 8 - результаты анализа прямого взаимодействия GcMAF и белков лизата ПМФ, где А - сэндвич анализ прямого взаимодействия лизата ПМФ с белком GcMAF в присутствии и отсутствии ионов Са2+. Окраска Кумасси и сэндвич вестерн-блот. На блоте выявляются специфически реагирующие фракции белков, соответствующие классическому CLEC10A с молекулярным весом 29 кДа, и две высокомолекулярные фракции, по подвижность соответствующих высокомолекулярным CLEC10A дериватам. Отсутствие в инкубационной среде и инкубационной системе сэндвич вестерн-блота ионов Са2+ значительно снижает силу специфического сигнала, характеризующего прямое взаимодействие двух белков. В - аватары графиков синтеза мРНК анализируемых цитокинов ПМФ, обработанных GcMAF3 L plasma LysoPC 37°С (фиг. 4, А). При отсутствии ионов Са2+ банд 63 кДа реагирует с GcMAF в меньшей степени, чем с белком 65 кДа. В присутствии ионов Са2+ обе фракции с большей эффективностью взаимодействуют с GcMAF, причем фракция 63 кДа связывает лиганд в условиях сэндвич вестерн блота с аналогичной эффективностью, что и банд 65 кДа. Это свидетельствует о появлении специфического аффиннитета GcMAF и CLEC10A 63 кДа. С - график синтеза мРНК анализируемых цитокинов ПМФ, обработанных препаратом GcMAF, полученным конвертацией очищенными ферментами; D - сэндвич анализ прямого взаимодействия лизата ПМФ с Ex GcMAF LEV L pure LysoPC 37°C, демонстрирующего синтез мРНК цитокинов разнонаправленного действия, и Ex GcMAF sum enzyme 37°С, демонстрирующего выраженный противовоспалительный вектор синтеза мРНК анализируемых цитокинов. В первом случае оба высокомолекулярных деривата CLEC10A взаимодействуют с GcMAF. Во втором случае с GcMAF преимущественно связывается дериват 63 кДа.
Способ получения и определения специфической активности фактора активации макрофагов GcMAF реализуется следующим образом.
Способ получения и определение специфической активности фактора активации макрофагов GcMAF основан на выделении, конвертации и определении специфической активности, причем, выделение и конвертацию фактора активации макрофагов GcMAF осуществляют на оригинальном твердом носителе Sephacril/25-OH D3 в условиях аффинной сорбции фактора активации макрофагов на частицах смолы, для его валидации используют матричную рибонуклеиновую кислоту (мРНК), синтез которой активируется фактором активации макрофагов GcMAF в перитонеальных макрофагах (ПМФ) мышей линии C57BL/6, набор специфических праймеров и процедуру полимеразной цепной реакции (ПЦР) в реальном времени, при этом, в качестве исходного сырья для получения активатора макрофагов GcMAF используют смесь плазм доноров, разбавленную в половину физиологическим раствором или фосфатно-солевым буфером, а стандартизацию фактора активации макрофагов GcMAF проводят путем выделения фактора на оригинальном Sephacril/25-OH D3 субстрате, конвертации DBP в фактор активации макрофагов GcMAF в связанном с субстратом Sephacril/25-OH D3 состоянии, использовании для определения активности каждого выделения препарата клеточной системы ПМФ мышей линии C57BL/6 и определении способности DBP и GcMAF индуцировать синтез мРНК ПМФ четырех цитокинов - медиаторов воспаления TNF-α и IL-1β и противовоспалительных цитокинов TGF-β и IL-10, при этом, при конвертации используют очищенные ферменты β-галактозидазу и сиалидазу, присутствующие в конвертационной среде, а способность ПМФ синтезировать цитокины определяют с использованием процедуры ПЦР в реальном времени, где в качестве специфических праймеров используют праймеры на провоспалительные цитокины TNF-α и IL-1β и противовоспалительные цитокины TGF-β и IL-10.
Сущность способа заключается в том, что создан оригинальный субстрат для выделения предшественника 25-OH/Sepharose, найдены оптимальные технологические параметры выделения DBP и его конвертации в GcMAF на твердом носителе, разработана уникальная система анализа вектора воспалительной реакции, индуцируемого препаратом GcMAF с использованием ПМФ мышей линии C57BL/6 и их способностью синтезировать мРНК медиаторов воспаления TNF-α, IL-β и основных противовоспалительных цитокинов TGF-β IL-10.
Для выделения фактора используется оригинальный аффинный субстрат 25-OH/Sepharose.
Кроме того, требуемый результат достигается тем, что предшественник DBP конвертируется в активатор макрофагов GcMAF на твердом носителе ферментами, находящимися в растворе, что создает условия получения высокоочищенного субстрата для получения фармацевтического препарата. При этом, требуемый результат достигается тем, что каждое выделение препарата GcMAF стандартизируется оценкой его способности активировать направление воспалительной реакции макрофагов, используя в качестве модели мышиные ПМФ, а требуемый результат достигается тем, что оценивается синтез ПМФ мРНК специфических про- или противовоспалительных цитокинов TNF-α, IL-1β, TGF-β, IL-10.
Актуальность данного изобретения подтверждается описанными в современной научной литературе векторами терапевтического воздействия препарата GcMAF.
Используемые методики.
Экспериментальные животные.
В работе использовали самцов и самок мышей линии C57BL/6 в возрасте от 2 до 6 мес (масса 18-24 г), выращенных в Центре коллективного пользования Виварии конвенционных животных Института цитологии и генетики СО РАН (Новосибирск, Россия). Животных содержали группами по 6-10 мышей в клетке при свободном доступе к пище и воде. Животных умертвляли методом шейной дислокации. Все эксперименты с участием животных проводились в строгом соответствии с принципами гуманности в соответствии с Директивами Совета Европейского сообщества (86/609/ЕЕС) и были одобрены Комиссией по уходу и использованию животных Института цитологии и генетики СО РАН.
Получение GcMAF.
Витамин Д3-связывающий белок (DBP) выделяли из человеческой плазмы индивидуальных доноров с использованием либо аффинной хроматографии на колонке с 25-ОН-D3/сефарозой (сефароза с ковалентно пришитым к ней витамином D3, гидроксилированным в 25-ом положении), либо в отдельных оговоренных случаях с помощью актин-сефарозной или актин-хитиновой аффинной хроматографии [5, 6]. DBP, полученный от индивидуального донора, обозначали буквами или цифрой, например, DBP LEV, означает, что DBP был получен от донора LEV. Плазма от других доноров помечалась цифрами 1-3. В работе полученный DBP конвертировали в GcMAF несколькими способами.
Первый способ. Convention conditions (С). Для получения GcMAF посаженный на 25-ОН-D3/сефарозу DBP элюировали 3 М гуанидинхлоридом, диализовали против фосфатно-солевого буфера и инкубировали с предварительно очищенными от эритроцитов лейкоцитами (L pure). Лизис эритроцитов осуществляли буфером (0.15 М NH4Cl, 0.01М KHCO3, 0.1 тМ EDTA). Инкубацию DBP с очищенными лейкоцитами проводили с добавлением 10% эмбриональной бычьей сыворотки (FBS) и CaCl2 в конечной концентрации 1,5 тМ, в атмосфере СО2 при 37°С в течение 24 ч. После конвертации в convention conditions (С) препарат GcMAF стерилизовали фильтрованием. Обозначение препарата GcMAF (С GcMAF L pure 37°С) указывает на то, что препарат получен в convention conditions (С) с использованием очищенных неактивированных лейкоцитов при инкубации при температуре 37°С. Обозначение С GcMAF L plasma 39°С указывает, что препарат получен в convention conditions (С) с использованием лейкоцитов, предварительно активированных в родной плазме температурой 39°С (L plasma 39°С) в течение 2х часов.
Второй способ получения GcMAF обозначен нами как экспресс-способ (Ех). Конвертация проводилась во время нахождения предшественника в связанном состоянии с субстратом на смоле. В конвертационную среду с находящейся в ней смолой, ассоциированной с DBP, добавляли лейкоциты, обработанные различными способами: очищенные не активированные лейкоциты (L pure); предварительно активированные с помощью лизофосфатидилхолина (LysoPC, Sigma) (L LysoPC); предварительно активированные в родной плазме повышенной температурой 39°С (L plasma 39°С); предварительно активированные в плазме при 37°С (L plasma 37°С). Активация очищенных лейкоцитов с помощью LysoPC или температурой 39°С проводилась в буфере А (2 mM Tris-HCl рН=8, 0,5 тМ β-меркаптоэтанол, 0,2 тМ CaCl2). Препараты GcMAF, полученные таким экспресс-способом обозначены нами следующим образом: обозначение Ex GcMAF L plasma 37°С указывает на то, что этот препарат получен экспресс-способом, а именно, добавлением к посаженному на колонку DBP лейкоцитов, активированных в плазме при 37°С в течение 3 ч; обозначение Ex GcMAF L plasma 39°С указывает, что препарат получен экспресс-способом, добавлением к посаженному на колонку DBP предварительно активированных в родной плазме лейкоцитов при температуре 39°С в течение Зх часов и совместной инкубации при 37°С в течение 20 ч в присутствии 1.5 мМ CaCl2; Ex GcMAF L plasma LysoPC 37°С обозначает, что препарат получен экспресс-способом, добавлением к посаженному на колонку DBP предварительно активированных LysoPC лейкоцитов в родной плазме при 37°С в течение 3х часов и дальнейшей совместной инкубации при 37°С в течение 20 ч в присутствии 1.5 мМ CaCl2; Ex GcMAF L pure LysoPC 37°С обозначает, что препарат получен экспресс-методом, добавлением к посаженному на колонку DBP предварительно очищенных лейкоцитов, активированных LysoPC при 37°С в течение 3х часов и совместной инкубации при 37°С в течение 20 ч в присутствии 1.5 мМ CaCl2. Полученные в результате проведенных обработок препараты GcMAF отмывались от конвертационной среды через 20 ч инкубации, элюировались 3 М гуанидинхлоридом, диализовались против фосфатно-солевого буфера и использовались для активации ПМФ. Цифры и буквы, уточняющие происхождение GcMAF, указывают на донора плазмы крови, используемой для получения предшественника GcMAF - DBP. Данные препараты являются собственностью компании ООО «АКТИВАТОР МАФ».
Активация ПМФ с помощью DBP и GcMAF ex vivo. ПМФ выделяли из интактных мышей линии C57BL/6 и осаждали центрифугированием при 400 g в течение 7 мин. Клетки ресуспендировали в RPMI; концентрацию подсчитывали в камере Горяева. ПМФ (1 ×106 клеток/лунку) культивировали в среде RPMI-1640 («BioloT», Санкт-Петербург, Россия), содержащей 10% FBS (HyClone) и 40 мкг/мл гентамицина, в 24-луночных планшетах в течение 12 ч. Затем среду меняли на RPMI-1640 в присутствии DBP или GcMAF или в их отсутствие (контроль). Активаторы добавляли в каждую лунку (четыре раза в каждую дозу): 0,02 мкг, 0,2 мкг, 2 мкг и 20 мкг. Клетки инкубировали в атмосфере СО2 в течение 3 часов. Через 3 часа клетки отмывали от активатора.
Активированные и контрольные ПМФ лизировали TRlzol Reagent (Thermo Fisher Scientific, Waltham, США) для получения тотальной РНК.
Активация клеток J774 с помощью GcMAF ex vivo. Для активации культуральных макрофагов J774 препаратом GcMAF клетки культивировали в среде DMEM (БиолоТ, Санкт-Петербург, Россия), содержащей 10% FBS (HyClone) и 40 мкг/мл гентамицина, в 24-луночных планшетах (1×106 клеток/лунку) в течение 12 ч. Затем процедура активации проводилась по схеме, аналогичной активации ПМФ препаратом GcMAF ex vivo.
Активация макрофагов препаратом GcMAF in vivo. Для активации ПМФ препаратом GcMAF in vivo было взято по 3 мыши C57BL/6 в каждой из 4-х опытных групп и одной контрольной группе. GcMAF вводили внутривенно однократно в дозах 0,02 мкг, 0,2 мкг, 2 мкг и 10 мкг в 200 мкл физиологического раствора. Контрольной группе мышей вводили только физ. раствор. Через 5 часов после введения препарата ПМФ были выделены в RPMI индивидуально от каждой мыши, переведены в осадок и лизированы в TRlzol Reagent (Thermo Fisher Scientific, Waltham, USA) для получения total RNA.
Анализ активации фагоцитарной активности перитонеальных макрофагов у мышей. Фагоцитарную функцию макрофагов оценивали по методике, представленной в работе [7]. ПМФ мышей линии C57BL/6 (5x10^ клеток/лунку) культивировали в среде RPMI-1640 («БиолоТ», Санкт-Петербург, Россия), содержащей 10% FBS (HyClone) и 40 мкг/мл гентамицина в 24 луночных планшетах в течение 2-5 ч. Затем среду заменили на RPMI-1640 с добавлением 10% FBS либо в отсутствие (контроль), либо в присутствии GcMAF: 10 мкг/мл липополисахарида E.coli 0114:В4 (Sigma, St. Louis, МО, USA) в качестве положительного контроля и GcMAF в дозах 0,2 мкг, 0,5 мкг и 1 мкг на лунку. Клетки инкубировали в атмосфере СО2 в течение 3 ч. Затем в каждую лунку добавляли магнитные гранулы (шарики) (Dynabeads М-280, Invitrogen, Carlsbad, СА, USA) в дозе 60 мкг/лунку. После 30-минутной инкубации макрофаги трижды промывали фосфатно-солевым буфером для удаления неинтернализированных гранул (шариков). Макрофаги фотографировали в проходящем свете с помощью инвертирующего микроскопа Axio Observer Zl (Zeiss, Jena, Germany) и подсчитывали количество интернализованных шариков (IBN). Фагоцитарную активность макрофагов оценивали по формуле: IBN=количество интернализованных шариков/количество макрофагов. Для статистического анализа IBN использовали данные четырех независимых экспериментов; всего в каждом опыте оценивали 300-500 клеток. Клетки подсчитывали в нескольких полях, расположенных в разных частях лунки планшета.
Масс-спектрометрический анализ витамин-D3 связывающего белка. Информация о структуре пептидов была получена при помощи тандемного масс-спектрометрического анализа, в ходе которого проводилось разделение ионов-предшественников, фрагментация этих ионов с образованием вторичных ионов и анализ фрагментных ионов. Для масс-спектрометрического анализа образцов, предварительно разделенных методом 1D электрофореза, проводили ферментативное расщепление компонентов образцов трипсином с очисткой от примесей и последующим раздельным анализом получившихся смесей пептидов. Для этого выбранные бэнды были вырезаны из геля, помещены в 1,5 мл полипропиленовые пробирки и двукратно промыты водой, затем 50% ацетонитрилом (Merk) в 0,2 М бикарбонате аммония (Sigma Aldrich), высушены 100%) ацетонитрилом (Merk). Для редукции добавляли по 2 мкл 20 мМ дитиотрейтола (BioRad) в буфере, содержащем 0,2 М бикарбонат аммония (Sigma Aldrich) и инкубировали 30 мин в термошейкере (Biosan TS-100) при 600 об/мин и 57°С. Для алкилирования добавляли 2 мкл 50 мМ йодацетамида (BioRad) в буфере, содержащем 0,2 М бикарбонат аммония (Sigma Aldrich) и инкубировали в темноте 30 мин в термошейкере (Biosan TS-100) при 600 об/мин. Промывали и сушили 100% ацетонитрилом (Merk). Для проведения триптического гидролиза белка добавляли 0,02 мМ трипсина (Trypsin Gold, Mass Spectrometry Grade, Promega, USA) в 50 мМ бикарбоната аммония (Sigma Aldrich) и инкубировали 14 часов в термошейкере (Biosan TS-100) при 600 об/мин и 37°С.
Экстракцию пептидов проводили 1% трифторуксусной кислотой (Sigma Aldrich). Очистку пептидов проводили на Millipore ZIPTIP С18 ((Merk) согласно протоколу производителя. Пептидные смеси высушивали и перерастворяли в 20 мкл 0,1% раствора трифторуксусной кислоты (Sigma Aldrich) в смеси ацетонитрил (Мегк)/вода (W6-1, Fisher Chemical) (2%/98% по объему). Для хроматографического разделения использовалась ВЭЖХ система Thermo Fisher Scientific Ultimate 3000 Series (Nano/Cap System NCS-3500RS), автосамплер WPS-3000 TPL RS (с петлей 20 мкм), система охлаждения образцов. Разделение веществ проводили на колонке Acclaim РерМар RSLC С18 (75 мкм × 150 мм, 2 мкм, 100 Å) в градиентном режиме. Состав подвижной фазы (элюента): Компонент А: 0.1% раствор муравьиной кислоты (Thermo Scientific) в воде (W6-1, Fisher Chemical). Компонент В: 0.1%) раствор муравьиной кислоты (Thermo Scientific) в смеси ацетонитрил (Мегк)/вода (W6-1, Fisher Chemical) (80%/20% по объему). Подвижная фаза нагрузочного насоса: 0.1% раствор трифторуксусной кислоты (Sigma Aldrich) в смеси ацетонитрил (Мегк)/вода (W6-1, Fisher Chemical) (2%/98% по объему).
Определение соединений осуществляли на масс-спектрометре Orbitrap Fusion Lumos в следующих условиях: ионизация проводилась электростатическим распылением при атмосферном давлении, в качестве детектора использовался орбитрап, режимы сканирования - MS ОТ, ddMS2 от HCD, разрешающая способность 60 000, использовалась ячейка высокоэнергетической соударительной диссоциации, энергия ионизации 30%, детектируемое зарядовое состояние от 2 до 7, для изоляции прекурсоров использовался квадрупольный фильтр масс, ширина изоляции масс - 1,6 m/z, диапазон детектируемых масс 300-1500 m/z, проводилось детектирование положительных ионов, порог интенсивности 5×104, время накопления ионов 60 мс, число сканирований 1, длительность динамического исключения 60 с, напряжение на распылителе 2,0 кВ, температура на капилляре 305°С, температура распылителя 270°С. Для сбора и обработки масс-спектрометрических данных использовалась программа Xcalibur v.2.4.
В программу Proteome Discovere (2.4.0) загружали масс-спектрометрические данные в формате первичных файлов с расширением raw. Обработку проводили базовым рабочим процессом (Prosessing step), включающим следующие основные узлы: Spectrum files, Spectrum selector, SequestHT, Percolator и IMP-ptmRS, согласованный рабочий процесс (Consensus step) включал узлы: MSF Files, PSM Grouper, Peptide Validator, Peptide and Protein Filter со ссылкой на аннотацию белка, Protein Score со ссылкой на Protein FDR Validator и Protein Grouping. Полученные данные об аминокислотной последовательности при обработке MS1 и MS2 масс-спектрограмм сравнивали с аминокислотной последовательностью человеческих генов из протеома UP000005640.
Получение кДНК.
Суммарную РНК из ПМФ выделяли с использованием реагента TRlzol (Thermo Fisher Scientific, Waltham, США) в соответствии с инструкциями производителя. Количество РНК измеряли на флуорометре Qubit 4 (Thermo Fisher Scientific, Waltham, США). ПЦР с обратной транскрипцией проводили на матрице поли-А мРНК с использованием усилителя Т100 Thermal Cycler (Bio-Rad Laboratories, Inc., Геркулес, США) и набора MMLV RT (Евроген, Москва, Россия) согласно протоколу производителя.
ПЦР-анализ.
Праймеры ПЦР для кодирующих областей каждого гена провоспалительных и противовоспалительных цитокинов были сконструированы с использованием программного обеспечения Vector NTI v.9 (Life Technologies, Wilmington, DE, USA) и синтезированы компанией BIOSSET Ltd. (BIOSSET, Новосибирск, Россия).
Последовательности праймеров, использованных в данном исследовании (for - прямой праймер; rev - обратный праймер).
Праймеры Олигонуклеотидные последовательности
TNF-α-for 5' - AAGCCTGTAGCCCACGTCGTA - 3'
TNF-α-rev 5' - GGCACCACTAGTTGGTTGTCTTTG - 3'
IL-1β-for 5' - TCCAGGATGAGGACATGAGCAC - 3'
IL-1β-rev 5' - GAACGTCACACACCAGCAGGTTA - 3'
TGF-β-for 5' - GTGTGGAGCAACATGTGGAACTCTA - 3'
TGF-β-rev 5' - TTGGTTCAGCCACTGCCGTA - 3'
IL-10-for 5' - GACCAGCTGGACAACATACTGCTAA - 3'
IL-10-rev 5' - GATAAGGCTTGGCAACCCAAGTAA - 3'
GAPDH-for5' - AAATGGTGAAGGTCGGTGTG - 3'
GAPDH-rev5' - TGAAGGGGTCGTTGATGG- 3'
Полимеразную цепную реакцию проводили в общем объеме 50 мкл; реакционная смесь содержала Taq-буфер, 2 мМ MgCl2, 0,2 мМ dNTP, 0,2 пмоль прямого и обратного праймеров, 1 нг матрицы и 5 ед. Taq-полимеразы (реагенты для ПЦР были предоставлены компанией «Медиген», Новосибирск, Россия). Схема ПЦР была следующей: цикл 1 (×1): 95°С - 3 мин; цикл 2 (×33): 95°С - 30 с; 59°С - 30 с; 72°С - 40 с; цикл 3 (×1): 72°С - 5 мин; хранение при 10°С.
ПЦР в реальном времени.
ПЦР в реальном времени проводили в 96-луночных планшетах с использованием BioMaster HS-qPCR SYBR (2х) (ООО «БИОЛАБМИКС», Новосибирск, Россия) по протоколу производителя на ПЦР-системе QuantStudio5 (Thermo Fisher Scientific, Waltham, CILJA). Анализ qlTLIP в реальном времени каждого образца проводили в трех повторностях. Относительный уровень экспрессии определяли методом 2-AACt. В качестве контрольной группы использовали интактные ПМФ без обработки; уровень экспрессии целевого гена в них принимался равным 1. В качестве эталона использовали ген GAPDH. Параметры циклирования были следующие: 95°С -10 мин, 40 циклов 95°С - 30 с, 59°С - 30 с, 72°С - 30 с. с конечной стадией плавления при медленном нагреве от 6 до 95°С.
Взаимодействие GcMAF и чистого GalNac с антителами на Gc-группу. Для сравнения взаимодействия GcMAF и чистого галактозамина (GalNac) с антителами на Gc-группу, образцы веществ наносили на нитроцеллюлозную мембрану: GalNac в эквимолярном количестве, 10-кратном и 100-кртном избытке по отношению к количеству GcMAF. GalNac наносили в 110 мкл 0.01 М фосфатно-солевого буфера с 0.02% Tween 20 в указанных количествах в лунку вакуумного приспособления для дот-блотинга. В четвертую лунку прибора наносили 20 мкл (4.5 мкг) GcMAF. Прибор подключали к вакууму, и после того, как содержимое лунок прошло через мембрану, было проведено последовательно три фиксирующих обработки (вакуумное просасывание) мембраны метанолом по 100 мкл и три промывки по 50 мкл трансферного буфера. После завершения прокачивания мембрану разрезали на две части: в первой части были три нанесения GalNac, а в другой - GcMAF. Мембраны были обработаны блокирующим буфером 0.01 М фосфатно-солевым буфером с 0.02%) Tween 20 в течение 1.5 часов. Затем были добавлены антитела к Gc группе с последующей инкубацией в течение ночи на качалке при температуре +9°С.
Western-blot анализ лизата макрофагов с антителами против CLEC 10А.
Для доказательства наличия CLEC 10А на поверхности макрофагов был проведен Western-blot анализ. Свежевыделенные перитонеальные макрофаги (ПМФ, 10 млн клеток) лизировали в 100 мкл физ. раствора, включающего 15 мкл 0.1 М PMFS, 40 мкл Sample Bufer (0.066 М Трис-HCl рН=6.8; 26.3% глицерола; 2.1% SDS, 0.011%) Bromophenol Blue) в течение 10 мин, затем кипятили 10 минут и центрифугировали. На электрофорез наносили лизат из 2 млн клеток ПМФ на дорожку. В качестве контроля использовали коммерчески доступный CLEC10A - 2 мкг на дорожку. После электрофореза проводили Western-blot с антителами к CLEC10A.
Анализ белок-белкового взаимодействия GcMAF//CLEC10A (сэндвич-анализ).
Прямое взаимодействие GcMAF и CLEC10A изучали методом сэндвича, который заключался в следующем. CLEC 10А или GcMAF после электрофореза переносили из полиакриламидного геля на мембрану описанным выше методом и обрабатывали контрагентом. Количество наносимого на гель фактора оставляло 2 мкг, а для обработки мембраны использовали 15 мкг контрагента в 10 мл 0.01 М фосфатно-солевого буфера с 0.02% Tween 20 и 20 мМ CaCl2. После инкубации в течение ночи, мембраны отмывали 3 раза по 5 минут буфером (0.01 М фосфатно-солевой буфер с 0.1% Tween 20) и инкубировали с 5 мкг анти-Gc или анти-CLEC10A в 10 мл 0.01 М фосфатно-солевого буфера с 0.02% Tween 20 в течение 10 часов. После отмывки выявляли реакцию антител.
Флуоресцентно-цитологический анализ взаимодействия ПМФ и Су5-GcMAF. Свежевыделенные ПМФ (1×106 клеток/лунку) культивировали в 500 мкл среды RPMI-1640 («BioloT», Санкт-Петербург, Россия), содержащей 10% FBS (HyClone) и 40 мкг/мл гентамицина, в 24-луночных планшетах (Eppendorf SE, ФРГ) в течение 12 ч. Затем в каждую лунку было добавлено по 2,5 мкг Су5-GcMAF. Планшет анализировали на конфокальном микроскопе LSM 780 NLO (Carl Zeiss AG, ФРГ). Интенсивность свечения зонда внутри клетки анализировали в программе ZEN (Carl Zeiss AG, ФРГ).
Статистический анализ.
Статистический анализ проводили с использованием программного обеспечения Statistica 8 (StatSoft, Tulsa, USA).
Рассмотрим дополнительные теоретические и экспериментальные предпосылки настоящего изобретения.
Активатор макрофагов GcMAF является одним из наиболее противоречивых и не до конца понятных факторов, вызывающих постоянные споры и недоверие научного сообщества. Фактору приписывают разнообразные свойства, а проведенные эксперименты свидетельствуют о значительном терапевтическом потенциале GcMAF [8]. Разные научные группы продемонстрировали противораковую активность активатора макрофагов [9-13]. Показана способность фактора купировать нейродегенеративные заболевания, включая аутический синдром [14-17].
Проведенные эксперименты свидетельствуют, что разнообразные терапевтические свойства препарата напрямую связаны с его воздействием на макрофагальное звено иммунитета [18-23]. Считается, что предшественник GcMAF, витамин Дз связывающий белок, конвертируется в активатор макрофагов частичным дегликозилированием трисахарида, связанного О-гликозидной связью с треонином в 420 положении, приводящим к появлению концевого GalNac остатка. Имеющиеся экспериментальные данные свидетельствуют, что именно с этим карбогидратом связана способность GcMAF активировать макрофаги [24-29]. Известны препараты, имеющие название GcMAF, но по сути таковыми не являющиеся. Это GcMAF второго поколения, который является смесью обработанных ферментами плазматических белков и, так называемый, GcMAF третьего поколения, представляющий собой молозиво, характеризующееся присутствием свободного GalNac [2, 3, 23, 30].
В процессе работ по настоящему изобретению авторы столкнулись с неопределенностью процедуры выделения белка DBP из плазмы крови и его конвертации в GcMAF [6, 16, 24, 31]. В этой связи разработан оригинальный способ получения, конвертации и анализа активирующей способности GcMAF. Одновременно были выяснены молекулярные причины, определяющие направление воспалительной реакции макрофагов, активированных GcMAF, и разработали принципы и элементы стандартизации препарата GcMAF.
Рассмотрим преимущества использования для выделения DBP (в последующем GcMAF) аффинный субстрат Sephacril/25-OH D3.
В наших предыдущих работах[6] активатор макрофагов GcMAF-RF получали из витамин Дз связывающего белка (DBP), после аффинной хроматографии плазмы донорской крови на колонке Актин/хитин с последующей конвертацией DBP в специфический активатор макрофагов GcMAF-RF обработкой лейкоцитарной взвесью в суспензии в течение 12-24 ч в атмосфере СО2 (фиг. 1, А). Специфичность полученного фактора определяли либо вестерн-блот анализом на антитела против Gc группы, либо в специальных экспериментах на взаимодействие с лектином Н. pomatia [5]. Во всех выделениях вместе с целевым белком всегда выделялись примесные белки, также специфично взаимодействующие с актином. Несколько таких примесных мажорных фракций охарактеризованы в других работах [27, 32-36]. Такая процедура выделения не позволяла стандартизовать целевой белок, что было необходимо для детальных молекулярных исследований и создания медицинского препарата на основе специфического активатора макрофагов.
В этой связи была разработана система аффинного выделения предшественника, где в качестве аффинной матрицы использовался 25-OH/Sepharose. Получение матрицы полностью отличалось от способа, описанного в работе [31], и является предметом промышленной собственности компании ООО «АКТИВАТОР МАФ». Продукт, полученный с использованием указанной системы аффинной хроматографии, представлял собой два белка (фиг. 1, А2). Один мигрировал на уровне GcMAF, описанного в литературе, и специфически взаимодействовал с антителами к Gc группе. Другой мигрировал на уровне ~28-30 kDa и не взаимодействовал с антителами к Gc группе (фиг. 1, В).
И таким образом в настоящем изобретении использовался аффинный сорбент 25-OH/Sepharose® со следующим обоснованием: 1) стабильная химически модифицированная матрица, позволяющая проводить большое количество хроматографий (до 9); 2) гомогенный продукт хроматографии, состоящий всегда из двух белков, описанных ниже, и как следствие, возможность дальнейшей стандартизации препарата.
Рассмотрим результаты тандемного масс-спектрометрического анализа двух белков 58 кДа и 29 кДа, стабильно выделяющихся хроматографией на Sephacril/25-OH D3.
На следующем этапе было проведено хроматографическое разделение компонентов бэндов методом ВЭЖХ на системе Thermo Fisher Scientific Ultimate 3000 Series с последующим определением соединений на тандемном масс-спектрометре Orbitrap Fusion Lumos. Применение электрораспылительной ионизации позволило получить многозарядные ионы, достоинством которых является возможность получения масс-спектров вторичных ионов при фрагментации ионов-предшественников с различным зарядовым состоянием, что увеличивает степень идентификации аминокислотной последовательности. С помощью программы Proteome Discoverer 2.4. была установлена идентичность верхнего банда (58 кДа) на 63.66% с витамин Д3 связывающим белком, что достаточно для идентификации продукта, получаемого с 25-ОН колонки, как DBP. Полученный результат позволял нам использовать в наших дальнейших исследованиях общепринятый термин для обозначения фактора как GcMAF без добавления related factor (RF). Короткий белок (29 кДа) имел идентичность 74,54% с аполипопротеином А1, что достаточно для идентификации бека как ApoAl.
Рассмотрим принципы стандартизации препарата GcMAF.
Основой настоящего изобретения является разработанный протокол стандартизации каждого выделения препарата GcMAF, включающий: выделение фактора на оригинальном Sephacril/25-OH D3® субстрате; конвертация DBP в активатор макрофагов в связанном с субстратом Sephacril/25-OH D3 состоянии; использование для определения активности каждого выделения препарата клеточной системы ПМФ, получаемых от мышей линии C57BL/6; способность DBP и GcMAF индуцировать синтез мРНК ПМФ четырех цитокинов: медиаторов воспаления TNF-α и IL-1β и основных противовоспалительных цитокинов TGF-β и IL-10; использование в качестве конвертационной системы очищенные ферменты β-галактозидазу и сиалидазу, присутствующие в конвертационной среде в точных количествах.
Рассмотрим процедуру конвертации DBP в GcMAF на твердом носителе и охарактеризуем режимы конвертации.
Был разработан быстрый и эффективный способ конвертации DBP в GcMAF непосредственно на смоле, названный нами экспресс-выделение. В этом случае все обработки DBP проводят тогда, когда белок ассоциирован с матрицей. Именно этот способ конвертации является одним из основных элементов настоящего изобретения.
Проведена оценка эффективности конвертации лейкоцитами, обработанными различными техниками и чистыми ферментами. Полученный в результате различных вариантов конвертации GcMAF анализировался на эффективность активации синтеза мРНК выбранных цитокинов ПМФ мышей.
Обозначения выбранных режимов конвертации.
Были проанализированы различные режимы конвертации предшественника с использованием лейкоцитов. Также в сравнение были взяты результаты конвертации DBP обработкой ферментами сиалидазой и β-галактозидазой. Мы систематизировали используемые режимы и внесли соответствующие обозначения. Выделение в обычных условиях - с (conventional); выделенный на смоле - Ex (express); лейкоциты - L; в плазме -plasma; с использованием LysoPC - LysoPC; очищенные лейкоциты - pure; лейкоциты, обработанные при разных температурах, соответственно: 37°С и 39°С.
Варианты конвертации. DBP, фиксированный на смоле, обрабатывается лейковзвесью в родной плазме при 37°С или при 39°С.DBP, фиксированный на смоле, обрабатывается лейковзесью в родной плазме в присутствии LysoPC [37]. DBP, фиксированный на смоле, обрабатывается отмытыми лейкоцитами в буфере А и активированными к воспалению LysoPC в течение трех часов при 37°С в присутствии 2 мМ Са2+. Суммарный DBP, фиксированный на смоле, обрабатывается очищенными ферментами сиалидазой и β-галактозидазой.
Полученные в результате проведенных обработок препараты GcMAF отмывались от конвертационной среды, элюировались 3 М гуанидинхлоридом, диализовались против фосфатно-солевого буфера и использовались для активации ПМФ.
Обработка лейковзвесью предполагала, что в плазме в обязательном порядке будут продукты жизнедеятельности лейкоцитов. Также в конвертационной среде (фосфатно-солевой буфер) в некоторых вариантах присутствовало 10% FBS. Ранее было показано, что непосредственно конвертационная среда (фосфатно-солевой буфер + 10% FBS) не влияет на фагоцитарную активность ПМФ [6]. Это означало, что именно конвертированный в GcMAF DBP активирует ПМФ. Необходимо было дополнительно оценить влияние конвертационной среды на анализируемые в настоящем исследовании параметры, а именно синтез ПМФ мРНК медиаторов воспаления TNF-α и 1L-1β и основных противовоспалительных цитокинов TGF-β и IL-10. Установлено, что обработка ПМФ конвертационной средой, не содержащей DBP, но содержащей неиндуцированные к воспалению лейкоциты, не влияет на синтез указанных цитокинов ПМФ (фиг. 2, А). Это говорит о том, что лейкоциты сами по себе не способны активировать ПМФ.
Таким образом, для анализа активирующей способности DBP и GcMAF была выбрана модель оценки синтеза медиаторов воспаления TNF-α и IL-1β и основных противовоспалительных цитокинов TGF-β и IL-10 ПМФ, выделенными из мышей линии C57BL/6. Был использован препарат GcMAF, полученный аффинной хроматографией на 25-ОН, конвертированный либо в растворе, либо непосредственно на смоле различными вариантами конвертационной среды, содержащей лейкоциты, полученные от донора или чистые ферменты сиалидазу и β-галактозидазу.
Рассмотрим активацию синтеза мРНК ПМФ GcMAF, полученным в результате конвертации лейкоцитами, обработанными различными техниками.
На начальном этапе анализа было обнаружено, что DBP нескольких (4-х) доноров сами по себе активируют ПМФ к синтезу мРНК одного или нескольких цитокинов, при этом в некоторых случаях, относящихся к разнонаправленным векторам воспаления (фиг. 2, В, С, D, Е).
DBP LEV незначительно активировал ПМФ к синтезу IL-1β (фиг. 2, В). Нужно еще раз отметить, что во всех наших предыдущих работах [5, 6, 38, 39] именно этот предшественник использовался во всех проведенных экспериментах. При этом он активировал ПМФ к синтезу противовоспалительных цитокинов, взятых в анализ.
DBP1 активировал синтез мРНК TNF-α, IL-1β и TGF-β, относящихся к противоположным векторам воспаления.
DBP2 активировал синтез мРНК IL-1β и незначительно влиял на синтез мРНК противовоспалительных цитокинов.
DBP3 дозозависимо активировал синтез мРНК TNF-α и IL-1β. Синтез мРНК TGF-β при использовании всех трех доз активатора находился на уровне в ~6 раз выше контрольных значений.
Был проведен разносторонний сравнительный анализ способности препаратов предшественников и полученных из них препаратов GcMAF активировать синтез мРНК выбранных цитокинов. В сравнение попали различные дозы, температуры, способы конвертации и различным образом подготовленная лейкоцитарная взвесь. Во всех экспериментах использовались три дозы 0,02 мкг, 0,2 мкг, 2,0 мкг и в некоторых экспериментах 10 мкг целевого белка на точку (106 ПМФ в 0,5 мл полной культуральной среды DMEM).
Результаты представлены в следующей логической схеме. На фиг. 3 и фиг. 4 приведены результаты всех экспериментов. Для препаратов С GcMAF LEV L pure 37°C и GcMAF2, приведены результаты как анализа синтеза мРНК анализируемых цитокинов, так и анализ фагоцитарной активности этих же макрофагов. Мы сгруппировали полученные результаты согласно используемым в работе DBP.
С GcMAF LEV L pure 37°C. Был проведен анализ с использованием трех модельных систем: культуры макрофагов J774, ex vivo обработанных ПМФ и in vivo обработки экспериментальных животных с последующим анализом ПМФ. Установлено, что в случае культуры клеток активируется синтез мРНК IL-1β. Синтез мРНК других цитокинов не активируется. При обработке ПМФ ex vivo активируется синтез всех анализируемых цитокинов. Для in vivo системы синтез мРНК провоспалительных цитокинов незначительный. Синтез противовоспалительных TGF-β и IL-10 для in vivo варианта достигает 10-кратного превышения по сравнению с контролем. In vivo анализ свидетельствует о противовоспалительном ответе ПМФ при их активации С GcMAF LEV L pure 37°C. Анализ графика фагоцитарной активности при активации различными дозами С GcMAF LEV L pure 37°C продемонстрировал последовательный рост по ходу увеличения дозы препарата. При этом синтез мРНК TGF-β и IL-10 противовоспалительных цитокинов при двух минимальных дозах достигает максимальных значений, а при следующей максимально выбранной дозе <2,0 мкг падает до нуля (фиг. 3, А, В, С, Е, F, G). Таким образом, противовоспалительные TGF-β и IL-10 демонстрируют куполообразный график синтеза мРНК с максимумом в малых дозах. При этом доза <2,0 мкг для всех проанализированных цитокинов имеет провальные, до нулевой точки, значения. Результат активации ПМФ Ex GcMAF LEV L pure LysoPC 37°C конвертированным чистыми лейкоцитами, полученными в результате экспресс обработки LysoPC, демонстрирует максимально выраженный провоспалительный ответ фагоцитов (фиг. 3, D).
GcMAF 1 демонстрирует максимальный провоспалительный ответ ПМФ (фиг. 4, А, В).
GcMAF2 демонстрирует провоспалительный ответ ПМФ (фиг. 3, С). Анализ графика фагоцитарной активности при активации различными дозами GcMAF2 продемонстрировал следующую направленность. Повышение синтеза провоспалительных цитокинов (фиг. 4, D, Е, F) коррелирует с повышением фагоцитарной активности ПМФ, что диаметрально противоположно действию С GcMAF LEV L pure 37°C, где аналогичный эффект достигается при увеличении синтеза противовоспалительных цитокинов (фиг. 3, Е, F, G). Этот факт может говорить о том, что фагоцитарная активность ПМФ не связана с состоянием их воспалительной реакции.
GcMAF3 индуцирует колебательный ответ ПМФ в зависимости от обработок лейкоцитами. Один режим в выбранных дозах индуцирую анергию фагоцитов, что проявляется в около нулевых значениях синтеза мРНК всех анализируемых цитокинов (фиг. 4, G). Два других демонстрируют разнонаправленный эффект активации ПМФ (фиг. 4, Н, I).
В экспериментах с использованием С GcMAF LEV L pure 37°C наблюдалась куполообразная кривая эффективности синтеза мРНК при ярко выраженном эффекте ингибирования синтеза любых цитокинов в дозе GcMAF <2,0 мкг. В литературе подобное явление описано и, как считается, связано с агрегацией рецепторов при высоких дозах лиганда. Это предполагало, что при цитологическом анализе в образце с большим количеством лиганда могут наблюдаться зоны агрегации молекул рецепторов на ПМФ [40-43]. Для проверки этого предположения С GcMAF LEV L pure 37°C был помечен Су5 и проведена обработка ПМФ. Были выбраны три рабочих дозы 0,02 мкг, 0,2 мкг и 2,0 мкг. Конфокальный анализ живых клеток показал, что в случае двух меньших доз меченный материал локализуется в цитоплазме и ядре клеток. При использовании дозы 2,0 мкг детектируется 1) очень сильная метка, захватывающая всю клетку; 2) метка внутри клеток и 3) специфические пятна флуоресценции на поверхности макрофагов. Эти пятна могут являться агрегированными рецепторами, структурированными избытком лиганда (фиг. 5), что находится в согласии с имеющимся в литературе мнением.
Проведенный анализ свидетельствовал, что GcMAF, обработанный лейкоцитами, активированным различными техниками, действует разнонаправленно и непредсказуемо на синтез мРНК выбранных цитокинов ПМФ.
Рассмотрим причины разнонаправленного и непредсказуемого действия на синтез мРНК выбранных цитокинов ПМФ, активированных GcMAF, полученным в результате конвертации лейкоцитами, обработанными различными техниками.
Охарактеризуем возможный механизм активации ПМФ GcMAF.
GcMAF, как это следует из экспериментальной научной литературы, несет свободный GalNac в 420 положении (треонин) аминокислотной последовательности [24-27, 29, 44-48]. Хорошо изучены рецепторы с высокой аффинностью, связывающие карбогидраты [49]. Для GalNac описано два специфических рецептора, это ASGR1 (CLEC4H1) и CLEC10A (MGL или CD301). В отсутствии патологических проявлений CLEC10A экспрессируется на толерогенных тканевых дендритных клетках, макрофагах кожи, легких, перитонеальных макрофагах. При различных индуцирующих событиях экспрессия С-лектинового рецептора значительно возрастает и при аджаджировании с лигандом толерогенные антиген презентирующие клетки индуцируют или развитие Т-регуляторных лейкоцитов, или анергию иммунных клеток, в частности, Т-лимфоцитов MGL (CLEC10A) зависимым механизмом [50-53]. Выше сказанное означает, что основным кандидатом на взаимодействие с GcMAF является CLEC10A. Факт того, что DBP, у которого GalNac должен быть закрыт остатками двух других сахаров, в определенных случаях дозозависимо или не зависимо от дозы индуцирует синтез мРНК различных анализируемых цитокинов, может говорить о двух возможностях:
1) часть DBP уже дегликозилирована и имеет концевой GalNac. Хотя в этом случае кривые активации должны совпадать с кривыми GcMAF, а они в значительной степени отличаются (фиг. 2, 3, 4);
2) возможно частичное дегликозилирование;
3) DBP активирует синтез мРНК ПМФ за счет иного механизма активации.
Сравнивая кривые синтеза мРНК под влиянием DBP и GcMAF, можно предположить, что обработка DBP лейкоцитами меняет структуру DBP так, что мода активации полностью меняется. DBP связывается с определенным фактором и активирует ПМФ. Конвертация модифицирует молекулу DBP и, либо отменяет предыдущую связь DBP с ПМФ и образует новую, либо добавляет новую, которая меняет моду активации.
Для разъяснения этих моментов были проведены исследования, характеризующие взаимодействие DBP, GcMAF и CLEC10A.
Проанализируем взаимодействия GcMAF и CLEC10A.
GcMAF содержит открытый аддукт N-Ацетилгалактозамин (GalNAc), связанный с треонином в 420-ой позиции. Дегликозилирование трисахарида и, как например, удаление сиаловой кислоты и галактозы в очаге воспаления является принципиальным шагом в конвертации переносчика витамина Дз (DBP) в активатор макрофагов (GcMAF) [28, 54, 55].
Давно и активно изучаются свойства специфических рецепторов, связывающих карбогидраты (лектины), одним из которых является представитель большого семейства Са2+ зависимых лектиновых рецепторов С-типа, а именно CLEC10A (лектин макрофагального галактозного типа, MGL, CD301). CLEC10A содержит домен распознавания лектинов, связывающий концевой или свободный GalNAc [53, 56-59]. Он присутствует в большом количестве на цитоплазматической мембране дендритных клеток и активированных М2 макрофагов в форме тримера [43, 60-64], и согласно всем имеющимся представлениям должен связываться с GalNac треонина 420 у конвертированного в очаге воспаления GcMAF. Это означает, что CLEC10A является важнейшим кандидатом на место фактора, опосредующего активирующее действие GcMAF [5].
Был разработан метод сэндвич анализа и была проведена серия экспериментов, характеризующая взаимодействие GcMAF и CLEC10A. На фиг. 6 представлены результаты анализа взаимодействия этих факторов.
Показано, что в ПМФ C57BL/6 содержится CLEC10A, и что рецептор специфически взаимодействует с активатором макрофагов. Несколько фракций, реагирующих с антителами CLEC10A, выявляются на блоте (фиг. 6, А, В).
Анализ перекрестного взаимодействия антител к GC и CLEC10A не выявил значимого перекрестного аффиннитета, что позволяло провести эксперименты по анализу прямого взаимодействия белков GcMAF и CLEC10A (фиг. 6, С, D).
В первых экспериментах по анализу прямого взаимодействия CLEC10A и GcMAF с более выраженной провоспалительной активностью было проанализировано возможное взаимодействие анализируемых факторов с использованием техники cross-link формальдегидом. Технически анализ проводился следующим образом. Белки кросслинкировались в растворе. Проводился электорофорез и перенос на мембрану. Мембрана обрабатывалась антителами к CLEC10A. Было обнаружено, что появляется высокомолекулярная фракция по подвижности, предположительно, соответствующая комплексу GcMAFVCLEC 10 А, указано стрелкой (фиг. 6, D).
Далее были проведены эксперименты, характеризующие прямое взаимодействие двух факторов методом сэндвича. Либо CLEC10A, либо GcMAF после электрофореза переносились на мембрану и обрабатывались контрагентом. После этого образовавшийся комплекс выявлялся соответствующим конъюгированными с HRP антителами. На фиг. 7, А-С, Е, Н приведены результаты экспериментов. В обоих направлениях детектируется специфическое взаимодействие белков. В экспериментах использовались препараты GcMAF, полученные конвертацией различными режимами. В качестве основного анализируемого препарата был выбран GcMAF3, полученный с использованием режима Ex GcMAF3 L plasma 39°С, демонстрирующий околонулевые значения синтеза мРНК всех анализируемых цитокинов (фиг. 3, G). В части других экспериментов использовали другие препараты GcMAF, что оговорено непосредственно при описании результатов экспериментов.
Одним из основных вопросов был вопрос о влиянии ионов Са2+ на взаимодействие концевого карбогидрата DBP и GcMAF с CLEC10A. Считается, что именно Са2+ определяет направление ответа фагоцитов [56, 58, 65]. Для анализа был взяты предшественник DBP3 и GcMAF3, конвертированный экспресс-способом в родной плазме лейкоцитами, активированными нагреванием (39°С), с нейтральными или около нейтральными характеристиками (режим Ex GcMAF3 L plasma 39°С, фиг. 4, G). Стрипы с разогнанным CLEC10A обрабатывались активатором в присутствии разных концентраций Са2+ 2 мМ, 20 мМ и 100 мМ. После инкубации проявляли комплексы антителами к Gc. Оказалось, что фракция CLEC10A, соответствующая общеизвестной форме белка 29 кДа [66, 67] реагирует и с DBP, и с GcMAF без видимой разницы в условиях вестерн-блот анализа. Специфическое взаимодействие обнаруживалось только с высокомолекулярными фракциями препарата. В образце CLEC10A, обработанном GcMAF3 (режим Ex GcMAF3 L plasma 39°C) в присутствии 20 мМ Ca2+ появляются два белка (высокомолекулярные дериваты CLEC10A ~63, ~65 кДа), специфически связывающие GcMAF. Для этого эксперимента наиболее сильное взаимодействие демонстрирует белок ~65 кДа. При этом следующее повышение концентрации Са2+ практически отменяет связывание. Полученный результат предполагал, что 20 мМ Са2+ в условиях вестерн-блот анализа открывают сайт рецептора, связывающий концевой карбогидрат GalNac GcMAF3 (режим Ex GcMAF3 L plasma 39°C), что характерно для CLEC10A (фиг. 7, В, указано стрелкой). Дополнительно была проанализирована эффективность связывания CLEC10A и GcMAF (режим Ex GcMAF 1 L pure LysoPC 37°C), демонстрирующая выраженную провоспалительную активность при двух температурах 37°С и 39°С в присутствии оптимальной концентрации Са2+. Не было обнаружено влияния температуры на специфичность взаимодействия (фиг. 7, С). Основным реагирующим белком был белок 65 кДа. Белок 63 кДа реагировал кратно слабее.
Чтобы проверить гипотезу связывания GcMAF через концевой GalNac, были проведены эксперименты по конкурентному ингибированию связывания GalNac-GcMAF свободным GalNac. Результаты проведенного анализа представлены на фиг. 7, Е, F, G. Показано, что свободный GalNac конкурирует с концевым GalNac GcMAF за связывание специфического высокомолекулярного деривата CLEC10A, не давая ему сформировать четко оформленный комплекс с белком, что выражается в появлении размытого облака, детектируемого антителами (фиг. 7, Е, четвертая дорожка).
Для прояснения ситуации о принадлежности размытого облака к комплексу GalNac/рецептор, но не GcMAF/рецептор, была проанализирована способность антител против Gc группы взаимодействовать с чистым GalNac. Показано, что Gc антитела специфически связывают GalNac, что свидетельствует о том, что сайт белка GcMAF, несущий карбогидрат, является антигеном, индуцирует образование антител к GcMAF и, по-видимому, именно это взаимодействие выявляется как шмер (облако) при конкурентном вытеснении GcMAF из комплекса с высокомолекулярной фракцией CLEC10A 65 кДа в присутствии 20 мМ Са2+ свободным GalNac в количестве 10 мкг (фиг. 7, Е, F).
Были проведены ex vivo эксперименты, характеризующие конкурентные взаимоотношения свободного карбогидрата и концевого карбогидрата в составе GcMAF. ПМФ были обработаны GalNac, GcMAF, GalNac + GcMAF. Была проведена ПЦР в реальном времени. Показано, что оба вещества конкурируют за связывание с рецептором, от которого зависит активация синтеза мРНК анализируемых цитокинов (фиг.7, G). Выбранный GcMAF (режим Ex GcMAF3 L plasma 39°С) в этом эксперименте показал незначительную активацию ПМФ одновременно к синтезу как про-, так и противовоспалительных цитокинов. Кроме того, полученный ex vivo результат свидетельствует, что свободный GalNac не активирует ПМФ к синтезу мРНК анализируемых цитокинов.
Дополнительно проведен сравнительный анализ взаимодействия с CLEC10A двух препаратов GcMAF, полученных различными способами конвертации С GcMAF LEV L pure 37°C и Ex GcMAF3 plasma LysoPC 37°C. Первый препарат GcMAF характеризуется активацией разнонаправленных реакций ПМФ (фиг. 3, В), второй препарат демонстрирует активацию ПМФ в основном к противовоспалительной реакции, хотя в некоторых дозах наблюдается незначительный синтез мРНК TNF-α и IL-1β (фиг. 4, I). Оказалось, что для двух типов ответов (двух типов GcMAF) характерен различный паттерн связывания с двумя высокомолекулярными дериватами CLEC10A. Для С GcMAF LEV L pure 37°, активирующего оба направления воспалительной реакции ПМФ, это обе фракции с большей интенсивностью банда 65 кДа, а для Ex GcMAF3 plasma LysoPC 37°С, активирующего в большей степени противовоспалительный ответ ПМФ, - это преимущественно банд 63 кДа (фиг. 7, Н). Этот факт может свидетельствовать, о том, что за активацию только противовоспалительного ответа отвечает взаимодействие GcMAF преимущественно (или только) с дериватом CLEC10A 63 кДа. А связывание только с бандом 65 кДа отвечает за активацию провоспалительной реакции, как это следует из активации ПМФ Ex GcMAF 1 L pure LysoPC 37°С (фиг. 4, А, фиг. 7, D). Взаимодействие сразу с двумя дериватами активирует оба направления воспалительного ответа ПМФ. Также данный результат позволяет предположить, что два высокомолекулярных деривата CLEC10A 63 кДа и 65 кДа являются близкими «аллельными» формами одного и того же белкового фактора, специфически связывающими GcMAF.
В последней экспериментальной части настоящего изобретения было проанализировано несколько вариантов взаимодействия GcMAF и белков лизата ПМФ в системе сэндвич-вестерн-блот в присутствии и отсутствии ионов Са2+(как в культивационной среде, так и в растворах системы анализа). Показано, что Ex GcMAF3 L plasma LysoPC 37°С (фиг. 3,1) взаимодействует с тремя мажорными фракциями лизата ПМФ, по подвижности соответствующих CLEC10A 28 кДа, ~63 кДа и ~65 кДа. В присутствии ионов Са2+ специфическое взаимодействие с высокомолекулярными фракциями CLEC10A ~63 кДа и ~65 кДа усиливается (фиг. 8, А). В отсутствии ионов Са2+ наблюдается значительно более слабое взаимодействие в сэндвич реакции с Ex GcMAF3 L plasma LysoPC 37°С (фиг. 8, А, В). Можно отметить, что при появлении в растворе ионов Са2+ GcMAF более активно связывается с белком 63 кДа, что коррелирует с более выраженным противовоспалительным ответом ПМФ, показанным для этого препарата GcMAF. Разница в более сильном взаимодействии Ex GcMAF3 L plasma LysoPC 37°С с белком 63 кДа в случае чистого препарата CLEC10A (фиг. 7, Н, J), по сравнению с лизатом белков ПМФ, может быть связана с различиями в структуре аутентического и коммерческого CLEC10A. Также можно предположить, что с двумя белковыми фракциями взаимодействуют два разных варианта GcMAF, отличающиеся глубиной дегликозилирования. И существует стохастическая конкуренция в интернализации между аллельными формами CLEC10A, связавшимися с той или другой формой GcMAF в данном конкретном случае. Такая мода взаимодействия предполагает определенную вероятность в реализации про- или противовоспалительного ответа ПМФ с преимуществом противовоспалительного направления ответа для анализируемого препарата GcMAF.
В дополнении к описанным выше результатам установлено, что суммарный GcMAF, конвертированный экспресс способом из смеси DBP, полученных от разных доноров с использованием очищенных ферментов сиалидазы и β-галактозидазы, взаимодействует преимущественно с фракцией ~63 кДа (фиг. 8, С, Е) и активирует ПМФ к синтезу только противовоспалительного IL-10. При этом в этом же эксперименте другой препарат Ex GcMAF LEV L pure LysoPC 37°C (фиг. 3, С, D) с выраженным провоспалительным и одновременно противовоспалительным активирующим действием на ПМФ, взаимодействует с обеими фракциями ~63 и ~65 кДа в сэндвич-блот анализе (указано стрелками).
Настоящее изобретение свидетельствует, что ПМФ на своей поверхности несут несколько дериватов CLEC10A рецептора. Связь с низкомолекулярным дериватом 29 кДа не влияет на синтез про- или противовоспалительных цитокинов. Взаимодействие с двумя высокомолекулярными дериватами 63 и 65 кДа определяет направление воспалительной реакции. Только провоспалительная реакция проявляется в случае связывания лиганда с дериватом 65 кДа, только противовоспалительная - в случае связывания лиганда с дериватом 63 кДа.
Одновременно активируются оба направления воспалительной реакции ПМФ в случае связывания с обоими дериватами.
Таким образом, благодаря усовершенствованию известного технического решения обеспечивается достижение требуемого технического результата, который заключается в расширении области применения и повышении эффективностью применения.
Источники информации
1. Link R.P. et al. Purification of human serum vitamin D-binding protein by 25-hydroxyvitamin D3-Sepharose chromatography // Anal Biochem. 1986. Vol. 157, №2. P. 262-269.
2. Inui T. et al. Clinical experience of integrative cancer immunotherapy with GcMAF // Anticancer Res. 2013. Vol.33, №7. P. 2917-2920.
3. Amitani H. et al. Development of colostrum MAF and its clinical application // Neuropsychiatry. 2017. Vol.7, №3. P. 246-253.
4. Hori H. et al. Method for production of novel degalactosylated Gc globulin // Patent EP 2612921 B1.C07K 14/47, 10.07.2013.
5. Dolgova E.V. et al. Analysis of the Biological Properties of Blood Plasma Protein with GcMAF Functional Activity // Int J Mol Sci. 2022. Vol. 23, №15.
6. Левитес E.B. и др. Оценка in vitro биологической активности отечественного препарата макрофаг-активирующего фактора (GcMAF-RF) // Вавиловский журнал генетики и селекции. 2020. Vol.24, №3. Р. 284-291.
7. Ishikawa М. et al. A novel assay system for macrophage-activating factor activity using a human U937 cell line // Anticancer Res. 2014. Vol.34, №8. P. 4577-4582.
8. Saburi E., Saburi A., Ghanei M. Promising role for Gc-MAF in cancer immunotherapy: From bench to bedside // Caspian Journal of Internal Medicine. 2017. Vol.8, №4. p.228-238.
9. Kisker O. et al. Vitamin D binding protein-macrophage activating factor (DBP-maf) inhibits angiogenesis and tumor growth in mice // Neoplasia. 2003. Vol.5, №LP. 32-40.
10. Korbelik M., Naraparaju V.R., Yamamoto N. Macrophage-directed immunotherapy as adjuvant to photodynamic therapy of cancer // Br J Cancer. 1997. Vol.75, №2. P. 202-207.
11. Mohamad S. Bin et al. Preparation of Gc protein-derived macrophage activating factor (GcMAF) and its structural characterization and biological activities // Anticancer Research. 2002. Vol.22, №6 C. P. 4297-4300.
12. Pacini S. et al. Gc protein-derived macrophage-activating factor (GcMAF) stimulates cAMP formation in human mononuclear cells and inhibits angiogenesis in chick embryo chorionallantoic membrane assay // Cancer Immunology, Immunotherapy. 2011. Vol.60, №4. P. 479-185.
13. Toyohara Y. et al. Inhibitory effect of vitamin D-binding protein-derived macrophage activating factor on DMBA-induced hamster cheek pouch carcinogenesis and its derived carcinoma cell line // Oncol Lett. 2011. Vol.2, №4. P. 685-691.
14. Siniscalco D. et al. The in vitro GcMAF effects on endocannabinoid system transcriptionomics, receptor formation, and cell activity of autism-derived macrophages // J Neuroinflammation. 2014. Vol.11. P. 1-11.
15. Bradstreet et al. Initial Observations of Elevated Alpha-N-Acetylgalactosaminidase Activity Associated with Autism and Observed Reductions from GC Protein-Macrophage Activating Factor Injections // Autism Insights. 2012. P. 31.
16. Smith R. et al. Effects of Gc-Macrophage Activating Factor in Human Neurons; Implications for Treatment of Chronic Fatigue Syndrome // Am J Immunol. 2013. Vol.9, №4. P. 120-129.
17. Inui T. et al. Case Report: A Non-small Cell Lung Cancer Patient Treated with GcMAF, Sonodynamic Therapy and Tumor Treating Fields // Anticancer Res. 2016. Vol.36, №7.
18. Albracht S.P. Immunotherapy with GcMAF revisited - A critical overview of the research of Nobuto Yamamoto // Cancer Treat Res Commun. 2022. Vol.31.
19. Saburi E., Saburi A., Ghanei M. Promising role for Gc-MAF in cancer immunotherapy: From bench to bedside // Caspian J Intern Med. 2017. Vol.8, №4. P. 228-238.
20. Yamamoto N., Naraparaju V.R. Role of vitamin D3-binding protein in activation of mouse macrophages. // The Journal of Immunology. 1996. Vol.157, №4. P. 1744-1749.
21. Thyer L. et al. A novel role for a major component of the vitamin D axis: Vitamin D binding protein-derived macrophage activating factor induces human breast cancer cell apoptosis through stimulation of macrophages // Nutrients. 2013. Vol.5, №7. P. 2577-2589.
22. Thyer L. et al. GC protein-derived macrophage-activating factor decreases a-N-acetylgalactosaminidase levels in advanced cancer patients // Oncoimmunology. 2013. Vol.2, №8.
23. Sumiya Y. et al. Macrophages Exhibit a Large Repertoire of Activation States via Multiple Mechanisms of Macrophage-activating Factors // Anticancer Res. 2016. Vol.36, №7. P. 3619-3623.
24. Mohamad S.B. et al. Tumor cell alpha-N-acetylgalactosaminidase activity and its involvement in GcMAF-related macrophage activation // Comp Biochem Physiol A Mol Integr Physiol. 2002. Vol.132, №1. P. 1-8.
25. Delrue C, Speeckaert M.M. Vitamin D and Vitamin D Binding Protein in Health and Disease 2.0 // Int J Mol Sci. 2023. Vol.24, №12. P. 10316.
26. Bouillon R. et al. Vitamin D Binding Protein: A Historic Overview // Front Endocrinol. 2020. Vol.10.
27. Uto Y. et al. Degalactosylated/Desialylated Bovine Colostrum Induces Macrophage Phagocytic Activity Independently of Inflammatory Cytokine Production // Anticancer Res. 2015. Vol.35, №8. P. 4487-4492.
28. Yamamoto N., Kumashiro R. Conversion of vitamin D3 binding protein (group-specific component) to a macrophage activating factor by the stepwise action of beta-galactosidase of В cells and sialidase of T cells. // The Journal of Immunology. 1993. Vol.151, №5. P. 2794-2802.
29. Osawa M. et al. Epitope analysis of monoclonal antibodies to human Gc globulin (vitamin D-binding protein) // Biochem Mol Biol Int. 1998. Vol.44, №2. P. 293-303.
30. Kawakatsu K. et al. Characteristic Morphological Changes and Rapid Actin Accumulation in Serum-MAF-treated Macrophages // Anticancer Res. 2019. Vol.39, №8. P. 4533-4537.
31. Link R.P. et al. Purification of human serum vitamin D-binding protein by 25-hydroxyvitamin D3-Sepharose chromatography // Anal Biochem. 1986. Vol.157, №2. P. 262-269.
32. Mohamad S.B. et al. Erratum: Tumor cell alpha-N-acetylgalactosaminidase activity and its involvement in GcMAF-related macrophage activation (Comparative Biochemistry and Physiology Part A (2002) 132 (1-8) PII: SI095643315220) // Comparative Biochemistry and Physiology - A Molecular and Integrative Physiology. 2003. Vol.134, №2. P. 481.
33. De Souza M.G. et al. Actin immobilization on chitin for purifying myosin II: A laboratory exercise that integrates concepts of molecular cell biology and protein chemistry // Biochem Mol Biol Educ. 2008. Vol.36, №1. P. 55-60.
34. Miller K.G., Alberts B.M. F-actin affinity chromatography: technique for isolating previously unidentified actin-binding proteins // Proc Natl Acad Sci USA. 1989. Vol.86, №13. P. 4808-4812.
35. Trujillo G. et al. Cofactor regulation of C5a chemotactic activity in physiological fluids. Requirement for the vitamin D binding protein, thrombospondin-1 and its receptors // Mol Immunol. 2011. Vol.49, №3. P. 495-503.
36. Haddad J.G., Kowalski M.A., Sanger J.W. Actin affinity chromatography in the purification of human, avian and other mammalian plasma proteins binding vitamin D and its metabolites (Gc globulins) // Biochem J. 1984. Vol.218, №3. P. 805-810.
37. Yamamoto N., Homma S. Vitamin D3 binding protein (group-specific component) is a precursor for the macrophage-activating signal factor from lysophosphatidylcholine-treated lymphocytes // Proc Natl Acad Sci USA. 1991. Vol.88, №19. P. 8539-8543.
38. Ruzanova V.S. et al. Experimental Comparison of the In Vivo Efficacy of Two Novel Anticancer Therapies // Anticancer Res. 2021. Vol.41, №7. P. 3371-3387.
39. Кирикович C.C. и др. Влияние GcMAF-RF на ex vivo поляризацию макрофагов, активацию дендритных клеток и продукцию цитокинов клетками цельной крови человека // Медицинская иммунология. 2021. Vol.23, №2. Р. 257-274.
40. Mayes Р.А., Hance K.W., Hoos A. The promise and challenges of immune agonist antibody development in cancer // Nat Rev Drug Discov. 2018. Vol.17, №7. P. 509-527.
41. Westerfield J.M., Barrera F.N. Membrane receptor activation mechanisms and transmembrane peptide tools to elucidate them // J Biol Chem. 2020. Vol.295, №7. P. 1792-1814.
42. Martyanov A.A. et al. Heterogeneity of Integrin allb|33 Function in Pediatric Immune Thrombocytopenia Revealed by Continuous Flow Cytometry Analysis // International Journal of Molecular Sciences. 2020. Vol.21, №9. P. 3035.
43. Jegouzo S.A. et al. Organization of the extracellular portion of the macrophage galactose receptor: a trimeric cluster of simple binding sites for N-acetylgalactosamine // Glycobiology. 2013. Vol.23, №7. P. 853-864.
44. Morales E. GcMAF: a polemic or a highly promising molecule ? // World Sci News. 2017. Vol.65. P. 20-36.
45. Nagasawa H. et al. Gc protein (vitamin D-binding protein): Gc genotyping and GcMAF precursor activity // Anticancer Res. 2005. Vol.25, №6 A. P. 3689-3695.
46. Cooke N.E., David E. V. Serum vitamin D-binding protein is a third member of the albumin and alpha fetoprotein gene family // J Clin Invest. 1985. Vol.76, №6. P. 2420-2424.
47. Braun A., Bichlmaier R., Cleve H. Molecular analysis of the gene for the human vitamin-D-binding protein (group-specific component): allelic differences of the common genetic GC types // Hum Genet. 1992. Vol.89, №4. P. 401-406.
48. Останин А.А. и др. Тернистый путь макрофаг-активирующего фактора (GcMAF): от открытия к клинической практике // Вавиловский журнал генетики и селекции. 2019. Vol.25, №5. Р. 624-631.
49. Kenneth Hoober J. ASGR1 and Its Enigmatic Relative, CLEC10A // Int J Mol Sci. 2020. Vol.21, №14. P. 1-20.
50. Raes G. et al. Macrophage galactose-type C-type lectins as novel markers for alternatively activated macrophages elicited by parasitic infections and allergic airway inflammation // J Leukoc Biol. 2005. Vol.77, №3. P. 321-327.
51. van Vliet S.J., Saeland E., van Kooyk Y. Sweet preferences of MGL: carbohydrate specificity and function // Trends Immunol. 2008. Vol.29, №2. P. 83-90.
52. Kenneth Hoober J. Asgrl and its enigmatic relative, CLEC10A // International Journal of Molecular Sciences. 2020. Vol.21, №14. P. 1-20.
53. van Kooyk Y., Ilarregui J.M., van Vliet S.J. Novel insights into the immunomodulatory role of the dendritic cell and macrophage-expressed C-type lectin MGL // Immunobiology. 2015. Vol.220, №2. P. 185-192.
54. Yamamoto N. et al. Immunotherapy of metastatic colorectal cancer with vitamin D-binding protein-derived macrophage-activating factor, GcMAF // Cancer Immunology, Immunotherapy. 2008. Vol.57, №7. P. 1007-1016.
55. Yamamoto N., Suyama H., Yamamoto N. Immunotherapy for prostate cancer with Gc protein-derived macrophage-activating factor, GcMAF // Transl Oncol. 2008. Vol.1, №2. P. 65-72.
56. Drickamer K., Taylor M.E. Recent insights into structures and functions of C-type lectins in the immune system // Curr Opin Struct Biol. 2015. Vol.34. P. 26-34.
57. Iida S.I., Yamamoto K., Irimura T. Interaction of human macrophage C-type lectin with O-linked N-acetylgalactosamine residues on mucin glycopeptides // J Biol Chem. 1999. Vol.274, №16. P. 10697-10705.
58. Taylor M.E., Drickamer К. Mammalian sugar-binding receptors: known functions and unexplored roles // FEBS J. 2019. Vol.286, №10. P. 1800-1814.
59. Zelensky A.N., Gready J.E. The C-type lectin-like domain superfamily // FEBS J. 2005. Vol.272, №24. P. 6179-6217.
60. Klimmeck D. et al. Proteomic Cornerstones of Hematopoietic Stem Cell Differentiation: Distinct Signatures of Multipotent Progenitors and Myeloid Committed Cells // Mol Cell Proteomics. 2012. Vol.11, №8. P. 286.
61. Oo-puthinan S. et al. The amino acids involved in the distinct carbohydrate specificities between macrophage galactose-type C-type lectins 1 and 2 (CD301a and b) of mice // Biochim Biophys Acta. 2008. Vol.1780, №2. P. 89-100.
62. Roszer T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms // Mediators Inflamm. 2015. Vol.2015.
63. Wang Y. et al. Ml and M2 macrophage polarization and potentially therapeutic naturally occurring compounds // Int Immunopharmacol. 2019. Vol.70. P. 459-466.
64. Zhang B. et al. M2-Polarized tumor-associated macrophages are associated with poor prognoses resulting from accelerated lymphangiogenesis in lung adenocarcinoma // Clinics. 2011. Vol.66, №11. P. 1879.
65. Sancho D., Reis e Sousa C. Signaling by myeloid C-type lectin receptors in immunity and homeostasis // Annu Rev Immunol. 2012. Vol.30. P. 491-529.
66. Higashi N. et al. The macrophage C-type lectin specific for galactose/N-acetylgalactosamine is an endocytic receptor expressed on monocyte-derived immature dendritic cells // J Biol Chem. 2002. Vol.277, №23. P. 20686-20693.
67. Valladeau J. et al. Immature human dendritic cells express asialoglycoprotein receptor isoforms for efficient receptor-mediated endocytosis // J Immunol. 2001. Vol.167, №10. P. 5767-5774.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ТОРМОЖЕНИЯ РОСТА ПОДКОЖНОГО ТРАНСПЛАНТАТА ЭКСПЕРИМЕНТАЛЬНОЙ ГЛИОБЛАСТОМЫ ЧЕЛОВЕКА U-87, ПЕРЕВИТОГО ИММУНОДЕФИЦИТНЫМ МЫШАМ NU/J | 2019 |
|
RU2717218C1 |
| СПОСОБ КЛОНИРОВАНИЯ СВЯЗЫВАЮЩЕГО ВИТАМИН D БЕЛКА (БЕЛКА Gc), СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОГО ФАКТОРА АКТИВАЦИИ МАКРОФАГОВ, ФАКТОР АКТИВАЦИИ МАКРОФАГОВ (ВАРИАНТЫ), АДЪЮВАНТ (ВАРИАНТЫ), КЛОНИРОВАННЫЙ СВЯЗЫВАЮЩИЙ ВИТАМИН D БЕЛОК (Gc-БЕЛОК), КЛОНИРОВАННЫЙ ДОМЕН III СВЯЗЫВАЮЩЕГО ВИТАМИН D БЕЛКА (ВАРИАНТЫ) | 1996 |
|
RU2198218C9 |
| Средство, обладающее иммуномодулирующей активностью | 2018 |
|
RU2697526C1 |
| Средство, обладающее иммуномодулирующей активностью | 2017 |
|
RU2657819C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ИММУНОМОДУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2010 |
|
RU2423141C1 |
| ЛИНИЯ КЛЕТОК МЕЛАНОМЫ ЧЕЛОВЕКА ILG, СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2008 |
|
RU2395574C1 |
| ЛИНИЯ КЛЕТОК МЕЛАНОМЫ ЧЕЛОВЕКА 26G, СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2008 |
|
RU2395571C1 |
| ЛИНИЯ КЛЕТОК МЕЛАНОМЫ ЧЕЛОВЕКА 31G, СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2008 |
|
RU2395573C1 |
| ЛИНИЯ КЛЕТОК МЕЛАНОМЫ ЧЕЛОВЕКА IG, СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2008 |
|
RU2395572C1 |
| ЛИНИЯ КЛЕТОК МЕЛАНОМЫ ЧЕЛОВЕКА PG, СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2008 |
|
RU2395570C1 |
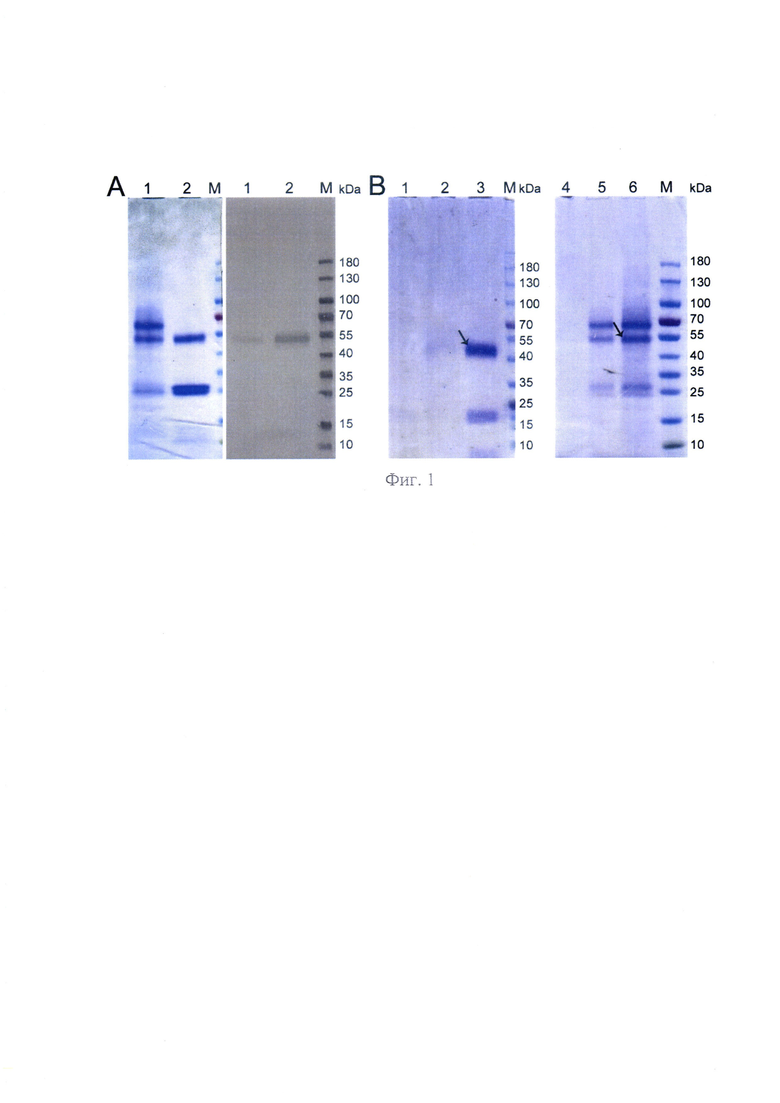
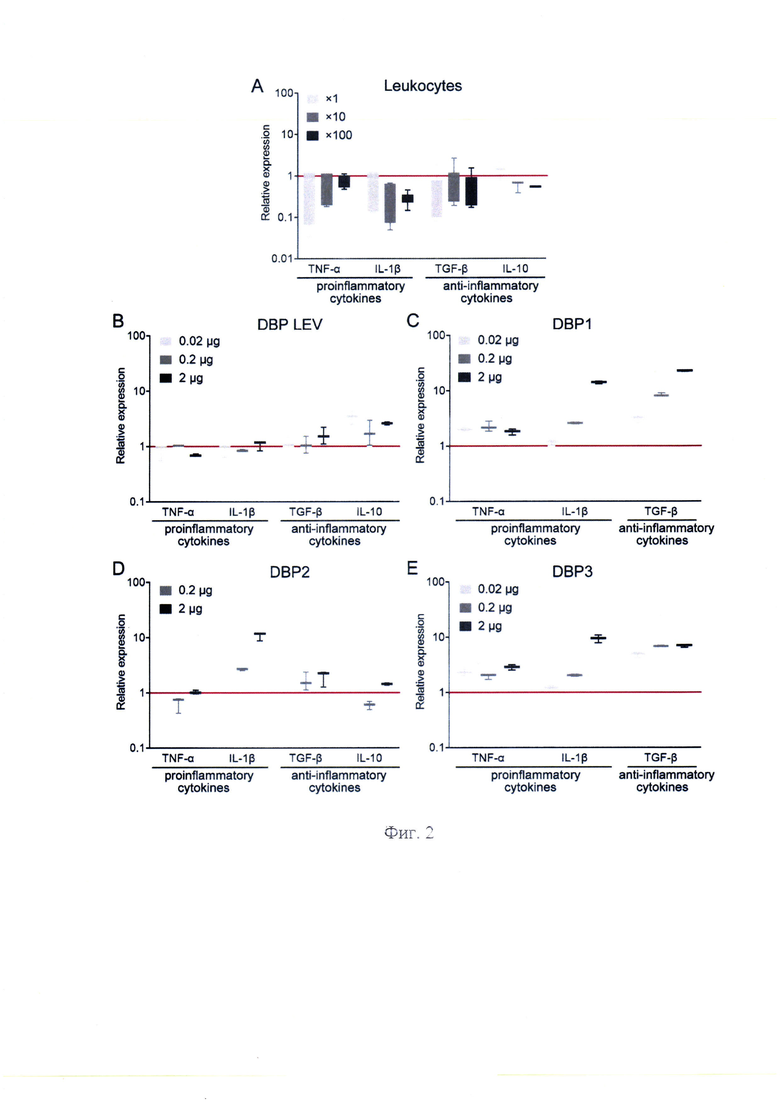
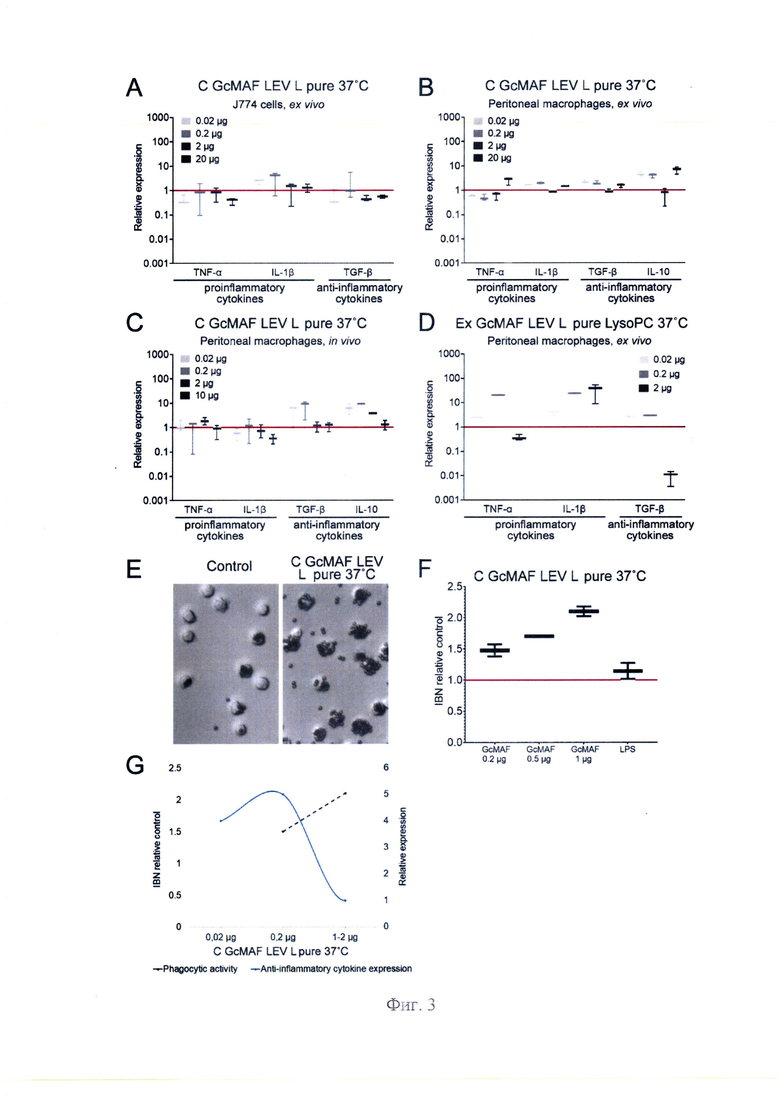


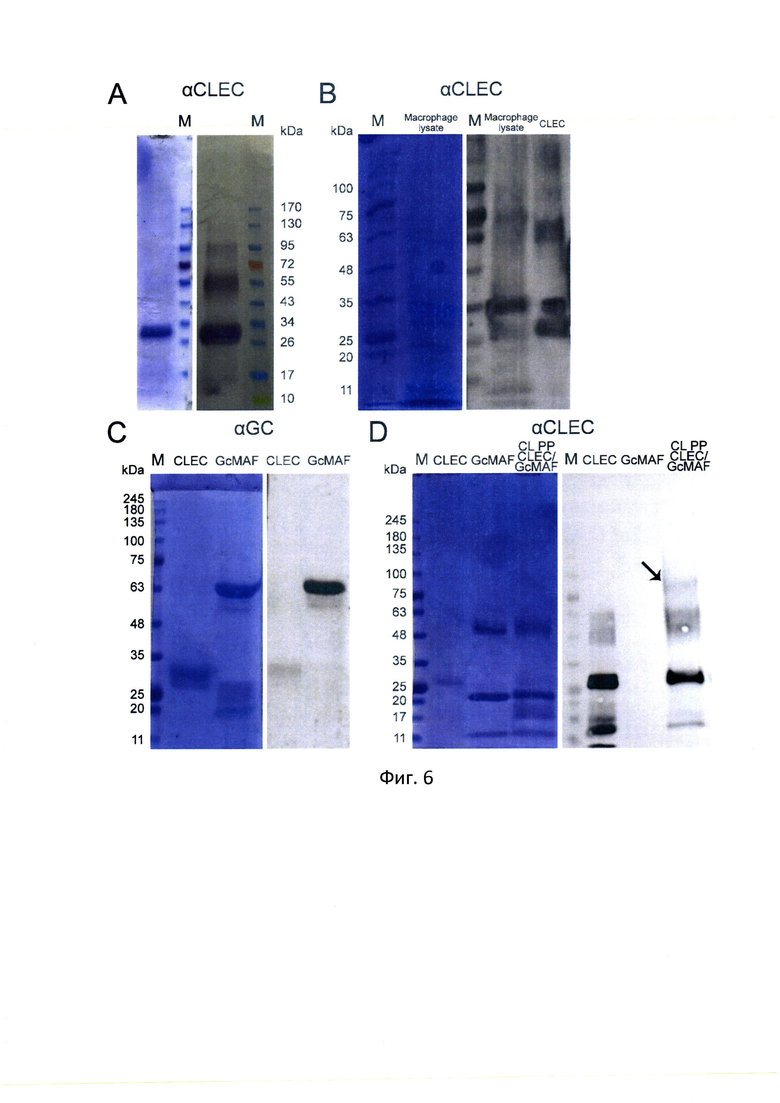


Изобретение относится к биотехнологии. Описан способ получения и определения специфической активности фактора активации макрофагов GcMAF. Способ основан на выделении, конвертации и определении специфической активности. При этом осуществляют выделение и конвертацию фактора активации макрофагов GcMAF на твердом носителе Sephacril/25-OH D3 - сефароза с ковалентно пришитым к ней витамином D3, гидроксилированным в 25-м положении, в условиях аффинной сорбции фактора активации макрофагов на частицах смолы. Для его валидации используют: специфические антитела и матричную рибонуклеиновую кислоту (мРНК), синтез которой активируется фактором активации макрофагов GcMAF в перитонеальных макрофагах (ПМФ) мышей линии C57BL/6; набор специфических праймеров и процедуру полимеразной цепной реакции (ПЦР) в реальном времени. В качестве исходного сырья используют смесь плазм доноров, разбавленную в половину физиологическим раствором или фосфатно-солевым буфером, а стандартизацию фактора активации макрофагов GcMAF проводят путем выделения фактора на Sephacril/25-OH D3 субстрате - сефароза с ковалентно пришитым к ней витамином D3, гидроксилированным в 25-м положении. Далее осуществляют конвертацию DBP в фактор активации макрофагов GcMAF в связанном с субстратом Sephacril/25-OH D3 состоянии. Определяют активность каждого выделения препарата клеточной системы ПМФ мышей линии C57BL/6 и определяют способности DBP и GcMAF индуцировать синтез мРНК ПМФ четырех цитокинов - медиаторов воспаления TNF-α и IL-1β и противовоспалительных цитокинов TGF-β и IL-10, при этом при конвертации используют очищенные ферменты β-галактозидазу и сиалидазу, присутствующие в конвертационной среде, а способность ПМФ синтезировать цитокины определяют с использованием процедуры ПЦР в реальном времени, где в качестве специфических праймеров используют праймеры на провоспалительные цитокины TNF-α и IL-1β и противовоспалительные цитокины TGF-β и IL-10. Изобретение может быть использовано для получения и определения специфической активности фактора активации макрофагов GcMAF, который является Gc-белком и носителем витамина D, который нейтрализует G-актин, выделяющийся при клеточном лизисе. Изобретение расширяет область применения и повышает эффективность. 8 ил.
Способ получения и определения специфической активности фактора активации макрофагов GcMAF, основанный на выделении, конвертации и определении специфической активности, отличающийся тем, что выделение и конвертацию фактора активации макрофагов GcMAF осуществляют на твердом носителе Sephacril/25-OH D3 - сефароза с ковалентно пришитым к ней витамином D3, гидроксилированным в 25-м положении, в условиях аффинной сорбции фактора активации макрофагов на частицах смолы, для его валидации используют специфические антитела и матричную рибонуклеиновую кислоту (мРНК), синтез которой активируется фактором активации макрофагов GcMAF в перитонеальных макрофагах (ПМФ) мышей линии C57BL/6, набор специфических праймеров и процедуру полимеразной цепной реакции (ПЦР) в реальном времени, при этом в качестве исходного сырья используют смесь плазм доноров, разбавленную в половину физиологическим раствором или фосфатно-солевым буфером, а стандартизацию фактора активации макрофагов GcMAF проводят путем выделения фактора на Sephacril/25-OH D3 субстрате - сефароза с ковалентно пришитым к ней витамином D3, гидроксилированным в 25-м положении, конвертации DBP в фактор активации макрофагов GcMAF в связанном с субстратом Sephacril/25-OH D3 состоянии, использовании для определения активности каждого выделения препарата клеточной системы ПМФ мышей линии C57BL/6 и определении способности DBP и GcMAF индуцировать синтез мРНК ПМФ четырех цитокинов - медиаторов воспаления TNF-α и IL-1β и противовоспалительных цитокинов TGF-β и IL-10, при этом при конвертации используют очищенные ферменты β-галактозидазу и сиалидазу, присутствующие в конвертационной среде, а способность ПМФ синтезировать цитокины определяют с использованием процедуры ПЦР в реальном времени, где в качестве специфических праймеров используют праймеры на провоспалительные цитокины TNF-α и IL-1β и противовоспалительные цитокины TGF-β и IL-10.
| СБОРКА РАСПРЕДЕЛИТЕЛЬНОГО УСТРОЙСТВА | 2012 |
|
RU2612921C2 |
| AU 2017254728 A1, 06.12.2018 | |||
| ПРЕПАРАТ ЦИСТЕИНОВОЙ ПРОТЕИНАЗЫ ПШЕНИЦЫ ТРИТИКАИНА-АЛЬФА, ПОЛУЧЕННОЙ В РАСТВОРИМОЙ ФОРМЕ, И СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА | 2017 |
|
RU2676322C1 |
| EP 3725883 A1, 21.10.2020. | |||

