Область техники, к которой относится изобретение
Настоящее изобретение относится к области биотехнологии, в частности к генетической инженерии, и может быть использовано в различных областях биотехнологии и научных исследованиях (тонкий органический синтез, селективное определение формиат-иона и НАД+ в сложных биологических системах, включая жидкости и ткани человека, фиксации СО2 из атмосферы и др.). Для достижения указанных целей сконструирована рекомбинантная плазмидная ДНК pHpFDH_HisA, содержащая модифицированный ген hpfdh_hisa, кодирующий мутантную `НАД+-зависимую формиатдегидрогеназу (КФ 1.2.1.2, HpFDH_HisA) c дополнительной на N-конце фермента последовательностью из шести остатков гистидина и одного остатка аланина. Полученная плазмида обеспечивает суперэкспрессию, включенного в нее гена hpfdh_hisa в клетках Escherichia coli в активной форме. Для обеспечения высокой эффективности экспрессии в целевом штамме-продуценте в модифицированном гене hpfdh_hisa в качестве стоп кодона используется тандем триплетов TAATAA вместо триплета TGA в исходном гене. Введение на N-конец фермента дополнительной последовательности His6Ala приводит к повышению термостабильности и позволяет в одну стадию получать препараты высокоочищенного фермента. В результате трансформации штамма E.coli BL21(DE3) предложенной плазмидой получен новый рекомбинантный штамм-продуцент E.coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559, обеспечивающий суперэкспрессию HpFDH_HisA в активной форме.
Уровень техники
НАД+-зависимая формиатдегидрогеназа катализирует окисление формиата до углекислого газа при сопряженном восстановлении NAД+ до НАДН.
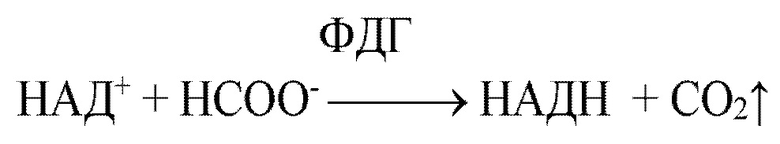
Равновесие реакция окисления формиат-иона сдвинуто в сторону образования СО2 и НАДН, продуктом является углекислый газ, который можно легко вывести из реакционной среды. Также ФДГ обладает широким pH-оптимумом активности. Поэтому формиатдегидрогеназа может быть эффективно использована для регенерации НАДН в процессах синтеза оптически активных соединений с использованием дегидрогеназ. В качестве примера можно привести крупномасштабный процесс получения трет-L-лейцина из триметилпирувата с использованием лейциндегидрогеназы с системой регенерации НАДН на основе формиатдегидрогеназы из дрожжей Candida boidinii (Bommarius A.S., Schwarm M., Stingl K., Kottenhahn M., Huthmacher K. and Drauz K., Synthesis and Use of Enantiomerically Pure tert-Leucine, Tetrahedron Asymmetry, 1995, v. 6, p. 2851-2888). Этот процесс до сих пор остается самым крупнотоннажным процессом получения хиральных соединений с помощью индивидуальных ферментов. Также применяют способ, в котором процесс синтеза и регенерацию осуществляют непосредственно в бактериальной клетке. В одной из первых работ ген ФДГ из бактерий Mycobacterium vaccae N10 был экспрессирован в клетках E.coli совместно с одним из ферментов - или лейциндегидрогеназой, или аланиндегидрогеназой, или фенилаланиндегидрогеназой. Такие клетки E.coli были использованы для синтеза L-лейцина, L-валина, L-норлейцина, L-метионина, L-фенилаланина и L-тирозина с выходом более 80% и оптической чистотой до 100% (Galkin, A., Kulakova, L., Yoshimura, T., Soda, K., & Esaki, N. Synthesis of optically active amino acids from alpha-keto acids with Escherichia coli cells expressing heterologous genes. Appl. Environ. Microbiol., 1997, v. 63, p. 4651-4656). Дальнейшим развитием работ в этом направлении стало получение химрных слитых белков. Ген ФДГ из дрожжей Candida boidini был объединен в одну полипептидную цепочку с кеторедуктазой и затем экспрессирован в E.coli . Такой биокатализатор был использован в процессах синтеза ксилитола из ксилозы и S-1-(2-хлорфенил)этанола из o-хлорацетофенона (Mädje K., Schmölzer K., Nidetzky B., Kratzer R. Host cell and expression engineering for development of an E.coli ketoreductase catalyst: Enhancement of formate dehydrogenase activity for regeneration of НАДH. Microb. Cell Fact. 2012 v. 11, p. 11-17). Отметим, что положение ферментов в полипептидной цепи имело решающее значение. Активным был только вариант слитого белка формиатдегидрогеназа-редуктаза. Одним из рекордсменов среди «слитых белков» является гибридный белок на основе мутантной НАДФ+-специфичной формиатдегидрогеназы из бактерий Pseudomonas sp.101 (NP-PseФДГН) и цитохрома Р450 монооксигеназы из Bacillus megaterium BM3 (P450BM3). P450BM3 используется в фармацевтической промышленности для получения синтонов и требует НАДФН в качестве кофермента (Kokorin A., Parshin P.D., Bakkes P.J., Pometun A.A., Tishkov V.I., Urlacher V.B. Genetic fusion of P450 BM3 and formate dehydrogenase towards self-sufficient biocatalysts with enhanced activity. Scientific Reports 2021, v. 11, paper 21706. DOI: 10.1038/s41598-021-00957-5). Было показано, что полученный гибридный биокатализатор с разными субстратами более эффективен, чем смесь отдельных ферментов от 6 до 50 раз. Данная химера является одной из самых больших среди гибридных белков, она состоит из 1200 аминокислотных остатков и включается 4 домена с разной каталитической активностью. Более подробный обзор по получению химерных ферментов с формиатдегидрогеназой и их применением можно найти в обзоре (Тишков В.И., Пометун А.А., Савин С.С. Формиатдегидрогеназа: от регенерации NAD(P)H до мишени в биопленках патогенов, компонента высокоэффективных гибридных биокатализаторов и фиксации СО2 из атмосферы. Вестник Московского университета. Серия 2. Химия. 2023, т. 64, № 4, с. 289-311. doi: 10.3103/S0027131423040077). Примеры и особенности использования формиатдегидрогеназы в тонком органическом синтезе оптически активных соединений рассмотрены в обзоре (Tishkov V.I., Pometun A.A., Stepashkina A.V., Fedorchuk V.V., Zarubina S.A., Kargov I.S., Atroshenko D.L., Parshin P.D., Shelomov M.D., Kovalevski R.P., Boiko K.M., Eldarov M.A., D'Oronzo E., Facheris S., Secundo F., Savin S.S. Rational design of practically important enzymes. Moscow University Chemistry Bulletin, 2018, Vol. 73, No. 2, p. 1-6. DOI 10.3103/S0027131418020153).
Формиатдегидрогеназа найдена в бактериях, дрожжах, микроскопических грибах, а также растениях. Впервые формиатдегидрогеназная активность в растениях была обнаружена в 1921 г. (Thunberg T. Sur la presence de certains ferments oxydants dans les grains de Phaseolus vulgaris. Arch. int. Physiol., 1921, v. 18, pp. 601-606). В растениях ФДГ - это фермент стресса, синтез которого резко возрастает при таких стрессовых воздействиях, как засуха, повышенная доза ультрафиолета, резкие скачки температуры, нехватка кислорода и т.п. (Hourton-Cabassa C., Ambard-Bretteville F., Moreau F., Davy de V., Remy R., and Francs-Small C.C. Stress Induction of Mitochondrial Formate Dehydrogenase in Potato Leaves. Plant Physiol, 1998, v. 116, pp. 627-635; Suzuki K., Itai R., Suzuki K., Nakanishi H., Nishizawa N.K., Yoshimura E., and Mori S. Formate dehydrogenase, an enzyme of anaerobic metabolism, is induced by iron deficiency in barley roots. Plant Physiol, 1998, v. 116, pp. 725-732; Thompson P., Bowsher C.G., and Tobin A.K. Heterogeneity of mitochondrial protein biogenesis during primary leaf development in barley. Plant Physiol, 1998, v.118, p.1089-1099; Andreadeli A., Flemetakis E., Axarli I., Dimou M., Udvardi M.K., Katinakis P., and Labrou N.E. Cloning and characterization of Lotus japonicus formate dehydrogenase: a possible correlation with hypoxia. Biochim.Biophys.Acta, 2009, v. 1794, pp. 976-984; David P., des Francs-Small C.C., Sevignac M., Thareau V., Macadre C., Langin T., and Geffroy V. Three highly similar formate dehydrogenase genes located in the vicinity of the B4 resistance gene cluster are differentially expressed under biotic and abiotic stresses in Phaseolus vulgaris. Theor.Appl Genet., 2010, v. 121, pp. 87-103). Подробный обзор по физиологической роли, получению и свойствам ФДГ из растений был опубликован в (Алексеева А.А., Савин С.С., Тишков В.И. НАД+-зависимая формиатдегидрогеназа растений. Acta Naturae, 2011, т.3, № 4 (11), с. 40-56). Были клонирован ген и получены мутантные формы рекомбинантной формиатдегидрогеназы из сои (Kargov, I.S., Kleymenov, S.Y., Savin, S.S., Tishkov, V.I., Alekseeva, A.A. Improvement of the soy formate dehydrogenase properties by rational design. Protein Engineering, Design and Selection, 2015, v. 28, N6, p. 171-178. doi: 10.1093/protein/gzv007; Alekseeva A.A., Kargov I.S., Kleimenov S.Yu., Savin S.S., Tishkov V.I. Additivity of the stabilization effect of single amino acid substitutions in triple mutants of recombinant formate dehydrogenase from the soybean Glycine max. Acta Naturae, 2015, v. 7, No 3 (26), p. 55-64. https://actanaturae.ru/2075-8251/article/view/10478/282. doi: 10.32607/20758251-2015-7-3-55-64). Полученные мутантные формы формиатдегидрогеназы из сои предполагается использовать для создания сортов сои с повышенной устойчивостью к стрессовым воздействиям без создания ген-модифицированных растений.
Гены формиатдегидрогеназ найдены в геномах практически всех патогенных бактерий и дрожжей. В случае такого опасного патогена как Staphylococcus aureus (SauFDH) было показано, что биосинтез фермента увеличивается в 20 раз при переходе бактерий из состояния планктона в состояние биопленок (Resch, A., Rosenstein, R., Nerz, C. and Gotz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl.Environ.Microbiol., 2005, v. 71, p. 2663-2676). Поскольку данный фермент критически важен для выживания стафилококков в биопленках, то высокоселективные ингибиторы SauFDH могут быть использованы в качестве антибактериального препарата именно против биопленок S.aureus. Ген ФДГ из S.aureus был клонирован и экспрессирован в клетках E.coli. Оказалось, что SauFDH имеет самую высокую каталитическую активность среди описанных формиатдегидрогеназ (Пометун А.А., Бойко К.М., Юрченко Т.С., Николаева А.Ю., Каргов И.С., Атрошенко Д.Л., Савин С.С., Попов В.О., Тишков В.И. Высокоактивная рекомбинантная формиатдегидрогеназа патогенных бактерий Staphylococcus aureus: получение и кристаллизация. Биохимия, 2020, т. 85, №6, с. 807-816, DOI: 10.1134/S0006297920060061). Однако применение SauFDH проблематично из-за высоких значений констант Михаэлиса.
В настоящее время формиатдегидрогеназа очень активно изучается как биокатализатор фиксации углекислого газа из атмосферы. Данное направление является одним из самых популярных в современной зеленой химии. Каждый год публикуется более ста статей на эту тему с участием формиатдегидрогеназы. Разрабатываются методы и подходы с использованием нанотехнологий и сопряжения формиатдегидрогеназной реакции восстановления СО2 до формиата с использованием электрохимии, фотохимии и др. (см. обзоры Sokol, K. P., Robinson, W.E., Oliveira, A.R., Warnan, J., Nowaczyk, M.M., Ruff, A., Pereira, I.A.C., & Reisner, E. Photoreduction of CO2 with a Formate Dehydrogenase Driven by Photosystem II Using a Semi-artificial Z-Scheme Architecture. J. Am. Chem. Soc. 2018, v. 140, p. 16418-16422, doi:10.1021/jacs.8b10247; Schlager, S., Dibenedetto, A., Aresta, M., Apaydin, D.H., Dumitru, L.M., Neugebauer, H., & Sariciftci, N.S. Biocatalytic and Bioelectrocatalytic Approaches for the Reduction of Carbon Dioxide using Enzymes. Energy Technol. (Weinh.) 2017, v. 5, p. 812-821, doi:10.1002/ente.201600610; Cakar, M. M., Ruupunen, J., Mangas-Sanchez, J., Birmingham, W. R., Yildirim, D., Turunen, O., Turner, N. J., Valjakka, J., & Binay, B. . Engineered formate dehydrogenase from Chaetomium thermophilum, a promising enzymatic solution for biotechnical CO2 fixation. Biotechnol. Lett. 2020, v. 42, p. 2251-2262, doi:10.1007/s10529-020-02937-7 [doi];10.1007/s10529-020-02937-7; Chen, Y., Li, P., Noh, H., Kung, C. W., Buru, C. T., Wang, X., Zhang, X., & Farha, O. K. (2019). Stabilization of Formate Dehydrogenase in a Metal-Organic Framework for Bioelectrocatalytic Reduction of CO2. Angew. Chem. Int. Ed Engl. 58, 7682-7686, doi:10.1002/anie.201901981.
В метилотрофных микроорганизмах при их росте на метаноле ФДГ снабжает клетку энергией в виде НАДH. Это свойство используется для улучшения метаболизма рекомбинантных штаммов. В рекомбинантный штамм вводят ген формиатдегидрогеназы, который экспрессирует ФДГ, а в культуральную среду добавляют формиат. Дополнительное снабжение клетки энергией в виде NADH с помощью ФДГ имеет явные преимущества. Во-первых, формиат-ион легко проникает в клетку, во-вторых, продуктом реакции является углекислый газ, который также легко выводить из клетки и, в-третьих, формиат-ион является ингибитором клеточного дыхания, что обеспечивает дополнительное селективное давление для сохранения в ходе культивирования только тех клеток, в которых имеется ФДГ. В литературе можно найти большое количество примеров работ по метаболитической инженерии рекомбинантных штаммов с использованием ФДГ. Полный список публикацй представлен в обзоре (Тишков В.И., Пометун А.А., Савин С.С. Формиатдегидрогеназа: от регенерации NAD(P)H до мишени в биопленках патогенов, компонента высокоэффективных гибридных биокатализаторов и фиксации СО2 из атмосферы. Вестник Московского университета. Серия 2. Химия. 2023, т. 64, № 4, с. 289-311. doi: 10.55959/MSU0579-9384-2-2023-64-4-289-311).
Формиатдегидрогеназы специфичны к формиату и большинство из них высокоспецифичны к коферменту НАД+. Это свойство используется для определения формиата и кофермента НАД+ в различных сложных биологических образцах, включая жидкости и ткани млекопитающих и человека. Метод определения формиата с помощью формиатдегидрогеназы был предложен еще в 1964 году (Johnson, P.A., Jones-Mortimer, M.C., Quayle, J.R. (1964). Use of a purified bacterial formate dehydrogenase for the micro-estimation of formate. Biochim. Biophys. Acta 89, 351-353, doi:10.1016/0926-6569(64)90225-1. Фирма Roshe более 37 лет выпускает набор для определения формиата в пищевых продуктах, однако используемый фермент в растворе инактивируется в течение 2 недель даже при 4°С. Более важным с точки зрения медицинской диагностики является определение уровня НАД+ в крови и тканях. НАД+ является важнейшим коферментом в организме и нарушение гомеостаза приводит к снижению концентрации кофермента в клетке. Поэтому контроль за уровнем НАД+ может быть использован для ранней диагностики различных заболеваний. Было показано, что формиатдегидрогеназа из дрожжей Hansenula polymorpha позволяет с высокой точностью и селективностью определять концентрацию НАД+ в экстрактах из мозга крыс (Artiukhov A.V., Pometun A.A., Zubanova S.A., Tishkov V.I., Bunik V.I. Advantages of formate dehydrogenase reaction for efficient NAD+ quantification in biological samples. Analytical Biochemistry, 2020, v. 603, art 113797. DOI: 10.1016/j.ab.2020.113797). Также было показано достоверное изменение концентрации НАД+ в крови пациентов различного возраста и пола в норме и патологии (сердечно-сосудистые и нейродегенеративные заболевания) (Balashova N.V., Zavileyskiy L., Artiukhov A., Shaposhnikov L.A., Sidorova O.P., Tishkov V.I., Tramonti A., Pometun A.A., Bunik V.I Efficient assay and marker significance of NAD+ in the human blood. Frontiers in Medicine, 2022, v. 9, paper 886485. DOI: 10.3389/fmed.2022.886485). Для такого типа анализа формиатдегидрогеназы должны иметь низкие значения констант Михаэлиса как по формиату, так и по НАД+. Этим требованиям удовлетворят только ФДГ из дрожжей и растений. Однако последние проигрывают дрожжевым формиатдегидрогеназам по каталитической активности и стабильности. Среди ФДГ из дрожжей наиболее изученным и коммерчески доступным является фермент из дрожжей Candida boidinii (CboFDH), однако CboFDH) имеет недостаточную стабильность и довольно быстро инактивируется при хранении в буфере при 4°С. Поэтому CboFDH хранят или в виде лиофилизованного препарата, или в 50% при -20°С. В 2014 году был клонирован и экспрессирован синтетический ген ФДГ из термотолератных дрожжей Ogataea parapolymorpha DL-1 (альтернативное название штамма Hansenula polymorpha DL-1) (HpFDH) (Yu, S., Zhu, L., Zhou, C., An, T., Zhang, T., Jiang, B., & Mu, W. Promising properties of a formate dehydrogenase from a methanol-assimilating yeast Ogataea parapolymorpha DL-1 in His-tagged form. Appl. Microbiol. Biotechnol, 2014, v. 98 (4), p. 1621-1630, https://doi.org/10.1007/s00253-013-4996-5). Для быстрой очистки и выделения фермента на С-конец была добавлена последовательность из 6 остатков гистидина (HpFDH_HisC). HpFDH_HisC значительно превосходил по стабильности ФДГ из Candida boidinii и на тот момент была самой стабильной ФДГ среди эукариот, однако HpFDH_HisC имел высокую Км по НАД+ (84 против 37 мкМ у CboFDH) и более низкую каталитическую константу (2,3 против 3,7 с-1). Клонирование и экспрессия гена HpFDH без His-tag на С-конце в клетках E.coli показало, что такой фермент имеет низкие значения Км по формиату и НАД+ (1,3 мМ и 14 мкМ соответственно), а каталитическая константа была намного выше (4,5 с-1) (Tishkov, V.I., Pometun, A.A., Stepashkina, A.V., Fedorchuk, V.V., Zarubina, S.A., Kargov, I.S., Atroshenko, D.L., Parshin, P.D., Kovalevski, R.P., Boiko, K.M., Eldarov, M.A., D’Oronzo, E., Facheris, S., Secundo, F., and Savin, S.S. Rational design of practically important enzymes, Moscow Univ. Chem. Bull., 2018, 73, 16, doi: 10.3103/S0027131418020153). По сравнению с ферментом дикого типа в ген HpFDH на N-конце был введен дополнительный триплет, кодирующий остаток глицина. Дополнительный остаток глицина был добавлен в последовательность фермента для снижения вероятности деградации HpFDH при культивировании в клетках E.coli (HpFDH_G). Именно этот фермент, был использован при анализе НАД+ в экстрактах мозга крыс и в крови здоровых и больных пациентов. Позднее был получен вариант HpFDH дикого типа без дополнительного остатка глицина (Пометун А.А., Шапошников Л.А., Зубанова С.А., Ковалевский Р.П., Атрошенко Д.Л., Пометун Е.В., Савин С.С., Тишков В.И. NAD+-зависимая формиатдегидрогеназа из термотолерантных дрожжей Ogataea parapolymorpha: свойства и белковая инженерия N-концевой последовательности. Биохимия, 2023, т. 88, №9, C. 1667-1680. DOI: 10.31857/S0320972523090178). Оказалось, что этот дополнительный остаток не влияет на термостабильность и каталитические свойства. Однако, несмотря на более высокую активность и стабильность HpFDH по сравнению с CboFDH, вопрос повышения стабильности HpFDH остается актуальным. Для практического применения также необходим вариант фермента, позволяющий получать его в высокоочищенном состоянии быстро и с минимальными затратами. Желательно, чтобы была всего одна стадия очистки. Для этого на N- или С-конец белков и ферментов вводят последовательность из шести остатков гистидина (His-tag), что позволяет проводить очистку в одну стадию с помощью метал-хелатной хроматографии. В литературе описаны формиатдегидрогеназы, имеющие His-tag на N- или С-конце (Yu, S., Zhu, L., Zhou, C., An, T., Zhang, T., Jiang, B., & Mu, W. (2014). Promising properties of a formate dehydrogenase from a methanol-assimilating yeast Ogataea parapolymorpha DL-1 in His-tagged form. Appl. Microbiol. Biotechnol., 98 (4), 1621-1630. doi.org/10.1007/s00253-013-4996-5; Esen H., Alpdağtaş S., Çakar M.M. Binay B. / Tailoring of recombinant FDH: effect of histidine tag location on solubility and catalytic properties of Chaetomium thermophilum formate dehydrogenase (CtFDH) / Prepar. Bioch. Biotechnol. 2019. V. 49:5. P. 529-534. DOI: 10.1080/10826068.2019.1599394 Шапошников Л.А., Савин С.С., Атрошенко Д.Л., Чубарь Т.А., Пометун Е.В., Тишков В.И., Пометун А.А. (2023) Инженерия N-концевой последовательности формиатдегидрогеназы из сои Glycine max. Вестн. Моск. ун-та. Сер. 2. Химия. 64 (4), 377-390. doi: 10.55959/MSU0579-9384-2-2023-64-3-377-390). У ФДГ из бактерий Pseudomonas sp.101 и сои введение His-tag на N-конец не приводило к изменению активности и стабильности. В случае ФДГ из Chaetomium thermophilum фермент с His-tag на С-конце имел немного лучшие кинетические характеристики по сравнению с вариантом His-tag на N-конце/ Фермент без His-tag в данной работе не получали. В случае HpFDH_HisC наличие His-tag на С-конце приводит к значительному ухудшению Км по НАД+ и снижению каталитической константы по сравнению с ферментом дикого типа HpFDH и вариантом с остатком глицина на N-конце HpFDH_G. Из вышесказанного следует, что влияние введения His-tag для каждой ФДГ имеет индивидуальный характер и предсказать на каком конце аминокислотной последовательности следует вводить метку нельзя.
Анализ технического уровня свидетельствует, что наиболее близким прототипом к заявляемому объекту по кинетическим свойствам являются плазмиды, кодирующие варианты HpFDH и HpFDH_G. Однако продуцируемые ферменты нуждаются в увеличении термостабильности и не позволяют провести очистку в одну стадию. В плане возможности очистки наиболее близким прототипом являются плазмида и штамм-продуцент E.coli, продуцирующие фермент HpFDH_HisC c His-tag на С-конце. Недостатком данного прототипа являются очень высокое по сравнению с HpFDH без His-tag значения Км по НАД+ (84 и 13 мкМ соответственно) и более низкое значение каталитической константы (2,3 с-1 против 4,4 с-1). Кроме того, для экспрессии используется плазмида, созданная на основе вектора pET23b(+). Данная система экспрессии E.coli BL21(DE3) - pET23b(+) имеет существенные недостатки. Плазмида pET23b(+), используемая для создания экспрессионного вектора, в качестве маркера селективного отбора клеток с плазмидой содержит ген устойчивости к ампициллину. Известно, что при культивировании рекомбинантного штамма по достижении поглощения культуры клеток на 600 нм величины А600 1 ед. поглощения и более, образующаяся бета-лактамаза полностью разрушает ампициллин и селективное действие антибиотика прекращается. Во-вторых, система экспрессии E.coli BL21(DE3) - pET23b(+) не обеспечивает строгого уровня контроля синтеза целевого белка, т.е. в этой системе идет хоть небольшая, но постоянная базовая экспрессии без всякой индукции. Если белок токсичен, то это приводит к преждевременной гибели клеток. Если нет, то в клетках происходит индукция протеаз, которые могут преждевременно деградировать целевой фермент в процессе культивирования.
Таким образом, техническая проблема, решаемая посредством заявляемого изобретения, заключается в необходимости преодоления недостатков, присущих аналогам и прототипу за счет создания рекомбинантного штамма E.coli - продуцента формиатдегидрогеназы, который одновременно обладает повышенной стабильностью по сравнению с известными штаммами и дает возможность проводить очистку в одну стадию.
Раскрытие изобретения
Техническим результатом заявляемой группы изобретений является разработка плазмиды и штамма, модифицированного данной плазмидой, которые позволяют получить высокие выходы нетоксичного высокоочищенного фермента (не менее 90%), обладающего повышенной термостабильностью (при 63°С термостабильность целевого фермента в 3 раза выше, чем термостабильность фермента дикого типа). Также заявляемые плазмида и штамма позволяют при упрощении и сокращении сроков синтеза получить нетоксичный высокоочищенны фермент - формиатдегидрогеназы HpFDH_HisA.
Заявляемая плазмида содержит ген формиатдегидрогеназы метилотрофных дрожжей Hansenula polymorpha DL-1, которая содержит на N-конце дополнительную последовательность из 6 остатков гистидина и одного остатка аланина. Наличие дополнительного остатка аланина обеспечивает повышенную термостабильность, а последовательность, а последовательность His-tag позволяет в одну стадию быстро получить препараты высокоочищенного фермента. За счет наличия в плазмиде гена белка-репрессора laqI осуществляется строгий контроль процесса экспрессии. Отсутствие преждевременной гибели клеток свидетельствует о нетоксичности экспрессируемого фермента. Высокий уровень экспрессии достигается за счет использования высокоэффективного промотора РНК-полимеразы фага Т7.
Штамм продуцент E.coli BL21(DE3)/pHpFDH_HisA содержит на хромосоме ген РНК-полимеразы фага Т7, что позволяет использовать полученную плазмиду pHpFDH_HisA для экспрессии целевого белка. Кроме того, исходный штамм E.coli BL21(DE3), который использовали для получения целевого штамма-продуцента мутантен по основной протеазе клеток E.coli (lonA-), что значительно предотвращает деградацию целевого фермента в процессе культивирования штамма-продуцента. Заявляемый штамм продуцент E.coli BL21(DE3)/pHpFDH_HisA является суперпродуцентом HpFDH_HisA. Даже при культивировании в качалочных колбах выход целевого фермента составляет более 1 г с литра среды.
Технический результат достигается рекомбинантной плазмидой pHpFDH_HisA размером 6359 п.о., содержащей модифицированный ген hpfdh_hisa формиатдегидрогеназы из дрожжей Hansenula polymorpha DL-1 под контролем T7 промотора. обеспечивающей синтез формиатдегидрогеназы из дрожжей H. polymorpha DL-1 (HpFDH_HisA) в клетках E.coli. Плазмида pHpFDH_HisA получена путем клонирования по сайтам NdeI/HindIII фрагмента размером 1113 п.о. (последовательность SEQ ID №3) с геном, кодирующим HpFDH_HisA (1110 п.о.) в плазмиду pET24a(+), расщепленную рестриктазами NdeI и HindIII.
Также технический результат достигается рекомбинантным штаммом Escherichia coli BL21(DE3)/pHpFDH_HisA - продуцентом мутантной формиатдегидрогеназы HpFDH_HisA, полученным путем трансформации штамма Escherichia. coli BL21(DE3) заявляемой плазмидной ДНК, задепонированный во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под номером В-14559.
Для обеспечения эффективной экспрессии HpFDH_HisA в клетках E.coli ген фермента клонируется в плазмидный вектор под контроль сильного промотора РНК-полимеразы фага Т7. В используемом для клонирования векторе pET24a(+) имеется ген Kan, который в качестве селективного маркера обеспечивает устойчивость к канамицину, а не к ампициллину как в pET23b(+). Кроме того, исходная плазмида pET24a(+) содержит ген белка-репрессора lac-промотора lacI, который предотвращает неспецифическую базовую экспрессию целевого белка. Многие гены ферментов из различных микроорганизмов в качестве стоп кодона содержат триплет TAG, который при экспрессии в клетках E.coli менее эффективен, чем стоп кодон TAA. Этот вариант наблюдается и в природном гене hpfdh. В связи с этим, клонируемая в вектор pET24a(+) нуклеотидная последовательность с геном hpfdh_hisa содержит замену триплета стоп кодона TAG на два триплета стоп кодона TAATAA.
Краткое описание чертежей
Изобретение поясняется следующими чертежами.
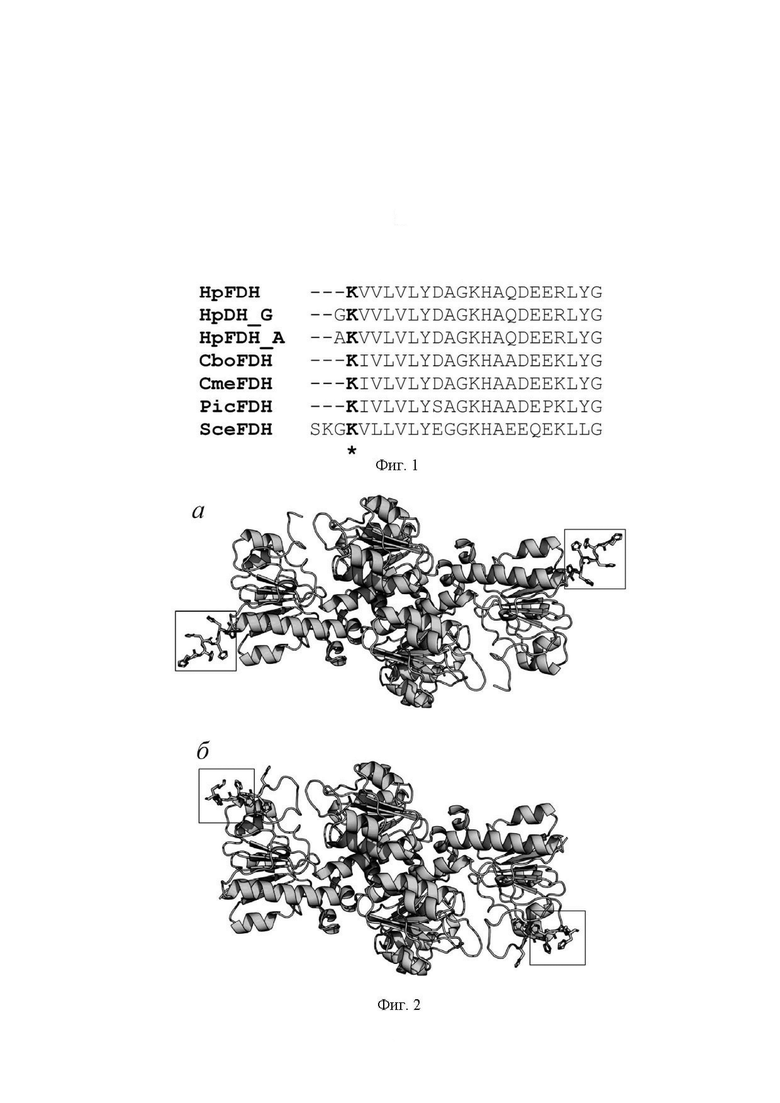
Фиг. 1 Сравнение N-концевых участков аминокислотных последовательностей формиатдегидрогеназ из метилотрофных дрожжей Hansenula polymorpha (HpFDH), Candida boidinii (CboFDH), Candida methylica (CmrFDH) и пекарских дрожжей Saccharomyces cerevisiae (SceFDH). Также приведены N-концевые последовательнсти вариантов HpFDH с дополнительными остатками глицина и аланина на N-конце фермента (HpFDH_G и HpFDH_A соответственно).
Фиг. 2. Модельные структуры HpFDH с дополнительными последовательностями из 6 остатков гистидина на N- и С-концах аминокислотной последовательности (фиг. а и б соответственно). Структура получена моделированием с использованием алгоритма AlphaFold2.
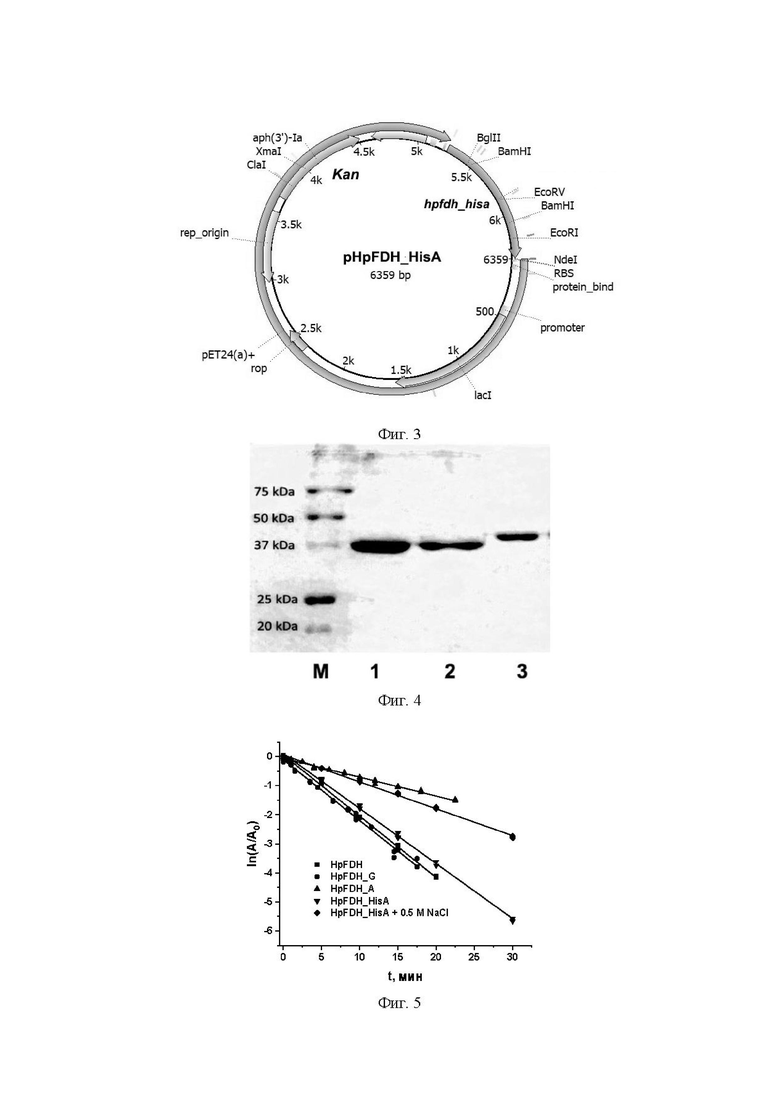
Фиг. 3. Физическая и генетическая карта плазмидного вектора HpFDH_HisA. Обозначены положения индикаторных сайтов рестрикции, рибосом-связывающий участок, ген устойчивости к канамицину (Kan) и ген lac-репрессора (lacI).
Фиг. 4. Аналитический SDS-Na электрофорез в 12% полиакриламидном геле очищенных препаратов HpFDH. М - маркеры молекулярной массы, 1 - HpFDH A, 2 - HpFDH, 3 - HpFDH_HisA.
Фиг 5. Зависимости остаточной активности от времени для различных форм HpFDH при 64°C в полулогарифмических координатах (0,1 М натрий-фосфатный буфер, pH 7,0).
Фиг 6. Фрагмент NdeI-HindIII размером 1113 п.о. (соответствует SEQ ID № 3).
Осуществление изобретения
С учетом требуемых свойств HpFDH по повышению стабильности и проведения очистки в одну стадию и необходимости создания системы экспрессии (комбинации "экспрессирующий вектор"/"штамм-реципиент") этого фермента достижение поставленных целей было реализовано за счет того, что:
1) в результате сравнения N-концевых участков аминокислотных последовательностей формиатдегидрогеназ из метилотрофных дрожжей было решено ввести на N-конц HpFDH остаток аланина, что должно согласно правилу N-конца Варшавского (N-end rule) снизить вероятность деградации фермента при его получении в клетках E.coli. Для обеспечения очистки целевого фермента в одну стадию была добавлена последовательность из 6 остатков гистидина. Компьютерное моделирование показало, что наименьшее влияние His-tag на свойства целевой HpFDH при его размещении на N-конце. Полученный ген был обозначен как hpfdh_hisa.
2) сконструирован экспрессионный вектор, содержащий ген hpfdh_hisa, который должен обеспечивать экспрессию этого гена в клетках E.coli. В качестве базового вектора для клонирования использовали плазмиду pET24a(+), которая обеспечивает длительное сохранение экспрессионного вектора в клетке в процессе культивирования за счет наличия гена устойчивости к канамицину и которая не допускает неспецифической базовой экспрессии целевого гена за счет наличия в плазмидном векторе гена белка- ингибитора lac- промотора lacI.
3) путем трансформации полученным вектором получается рекомбинантный штамм E.coli - продуцент HpFDH_HisA. Проводится культивирование полученного штамма-продуцента для проверки его способности экспрессировать активный рекомбинантный фермент. После культивирования проводится проверка эффективности очистки HpFDH_HisA в одну стадию с помощью метал-хелатной хроматографии.
4) изучаются каталитические свойства и термостабильность полученной HpFDH_HisA.
I. Выбор изменений в гене фермента проводится в двух направлениях:
1) Сравнение и анализ типа аминокислотных остатков на N-конце формиатдегидрогеназ метилотрофных дрожжей показали, что все последовательности начинаются с остатка лизина. Исключение составляет только ФДГ из пекарских дрожжей S.serevisiae.. На основании «правила N-конца Варшавского» было решено ввести на N-конец аминокислотный остаток, позволяющий повысить стабильность фермента против деградации при культивировании штамма-продуцента.
2) для практического применения фермента необходимо иметь быструю и высокоэффективную методику очистки. Поэтому было решено ввести дополнительную последовательность из шести остатков гистидина. Для выбора положения для введения His-tag - на N- или на С- конце, была построены модели белковой глобулы с шестью остатками гистидина на N- и С-концах. Анализ структур показал, что более оптимальным является His-tag на N-конце.
II. Конструирование экспрессирующего вектора включает три этапа:
1) анализ нуклеотидной последовательности гена, кодирующего HpFDH_HisA, показал, что стоп кодоном в этом гене является триплет TAG, в то время как в клетках E.coli в качестве стоп кодона более эффективен триплет TAA. Поэтому для обеспечения эффективной терминации трансляции гена hpfdh_hisa в клетках E.coli было принято решение при конструировании вектора заменить в гене hpfdh_hisa стоп кодон TAG на два триплета ТААТАА.
2) получение модифицированного гена hpfdh_hisa с последовательностью His6Ala на N- конце и двойным триплетом ТААТАА стоп кодонов проводили с помощью полимеразной цепной реакции с использованием прямого и обратного праймеров (SEQ ID №1 и SEQ ID №2). Для клонирования в экспрессирующий вектор в прямой и обратный праймеры были введены соответственно сайты рестрикции NdeI и HindIII.
3) экспрессирующий вектор pHpFDH_HisA был получен путем лигирования продукта ПЦР с модифицированным геном hpfdh_hisa в плазмидный в вектор pET24a(+) после их обработки рестриктазами Ndei и HindIII. Клонированная последовательность указана как SEQ ID №3. Сконструированный вектор pHpFDH_HisA (Фиг 3) содержит ген устойчивости к канамицину и ген белка-репрессора lac-промотора lacI.
III. Конструирование рекомбинантного штамма E.coli - продуцента HpFDH_HisA.
Рекомбинантный штамм E.coli BL21(DE3)/pHpFDH_HisA - продуцент HpFDH_HisA был получен трансформацией компетентных клеток E.coli BL21(DE3) экспрессирующей плазмидой pHpFDH_HisA. Сконструированный штамм Escherichia coli BL21(DE3)/pHpFDH_HisA оказался суперпродуцентом рекомбинантной HpFDH_HisA с выходом более 1 г целевого фермента с литра среды.
Штамм E.coli BL21(DE3)/pHpFDH_HisA депонирован во Всероссийской коллекции промышленных микроорганизмов как E. coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559.
Заявляемый штамм E. coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки
Клетки прямые, палочковидные, подвижные, грамотрицателиные, неспорообразующие. При выращивании в течение 24-48 ч при температуре 30-37°С на агаризованных средах (LB, 2YT), содержащих канамицин (25 мкг/мл), колонии гладкие, круглые, блестящие, край ровный.
Состав среды LB, мас.%:
Состав среды 2YT, мас.%:
Физиолого-биохимические признаки
Штамм растет при температуре от 15 до 37°С (оптимум 30-37°С). В качестве источника азота использует органический азот в виде триптона, аминокислот.
Штамм E.coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559 синтезирует рекомбинантную HpFDH_HisA после нндукции 0,1 мМ ИПТГ (изопропил-β-D-тиогалактозидом).
Генотипические признаки
Штамм E.coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559 устойчив к канамицину.
Штамм E.coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559 содержит на плазмиде pHpFDH_HisA модифицированный ген мутантной формиатдегидрогеназыт (hpFDH_HisA) из H.polymorpha DL-1 под контролем T7 промотора.
Условия хранения:
1. В ночную культуру, выращенную на жидкой среде 2YT с канамицином (25 мгк/мл), добавляют стерильный глицерин (до 15%), расфасовывают по 1,5 мл в пластиковые микропробирки и хранят в замороженном виде при -70°С.
2. Хранят в лиофилизированном виде в ампулах при +4°C.
3. Штамм может поддерживаться регулярными пересевами (1 раз в 3-4 недели) на агаризованной среде LB или 2YT, содержащей канамицин 25 мкг/мл.
IV. Одностадийная очистка HpFDH_HisA
Биомассу клеток E.coli после завершения культивирования отделяют от культуральной среды с помощью центрифугирования при 5000 об/мин в течение 15 мин при 4°С. Осадок ресуспендировали в буфере 0,05М Tris-HCl, 0,5М NaCl, 20 мМ имидазол, pH 7,5 в виде 10% (вес/вес) суспензии, подвергали двум циклам заморозки-разморозки, и затем клетки разрушали с использованием ультразвукового дезинтегратора «Branson Sonifier 250» (Германия) при постоянном охлаждении. Осадок удаляли центрифугированием на центрифуге «Eppendorf 5804R» (11 000 об/мин, 30 мин), наносили на колонку объёмом 1 мл HisTrap FF Ni Sepharose 6 Fast Flow (Cytiva, США), уравновешенной раствором, содержащим 0,05М Tris-HCl, 0,5М NaCl, 20 мМ имидазол, pH 7,5 (раствор А). После нанесения фермента колонку промывали раствором А в течение 30 минут до исчезновения поглощения на 280 нм. Фермент элюировали с колонки восходящим линейным градиентом раствора Б, содержащего 0,05М Tris-HCl, 0,5М NaCl, 500 мМ имидазол, pH 7,5. Начальная точка градиента - 0% раствора Б, конечная точка градиента - 60% раствора Б. Время прохождения градиента - 10 мл за 10 мин. Во время проведения хроматографии собирали фракции по 2 мл и измеряли поглощение на 280 нм (A280) и ферментативную активность (А340). Фракции с максимальным соотношением (A340/A280) объёмом около 2 мл для обессоливания наносили на колонку 1×10 см (объём 10 мл) с Sephadex G25 (Pharmacia Fine Chemicals, Швеция). Для обессоливания использовали раствор, содержащий 0,05 М Tris-HCl, pH 7,5. Чистоту полученных препаратов анализировали с помощью аналитического электрофореза в 12% полиакриламидном геле в присутствии 0,1% додецилсульфата натрия на приборе Miniran II (BioRad) согласно инструкции фирмы производителя
V. Каталитические параметры и термостабильность
Величины констант Михаэлиса по НАД+ и формиату и максимальную скорость реакции определяли из зависимости скорости реакции от концентрации (0,4-6 Км) одного субстрата при постоянной насыщающей (>20Км) концентрации второго субстрата. Обработку кинетической зависимости проводили методом нелинейной регрессии с использованием программы OriginPro 8.5. Значение каталитической константы определяли из уравнения Vmax = kcat*[E]. Концентрацию фермента определяли по формуле Калькара (С(г/л)= 1,45*А280-0,74*А260).
Термостабильность фермента измеряли в 0,1 М натрий-фосфатном буфере, pН 7,0 при нескольких температурах. В диапазоне 60-65°С. Для каждого эксперимента готовили серию из пластиковых пробирок объемом 0,5 мл по 100 мкл раствора фермента (0,2 мг/мл) в каждой. Пробирки помещали в предварительно прогретый до необходимой температуры водный термостат (точность термостатирования ±0,1°С). В определенные моменты времени отбирали по одной пробирке и переносили в лед на 5 мин, после чего пробирку центрифугировали в течение 3 мин при 12000 об/мин на центрифуге «Eppendorf Centrfuge5430». Из пробирки отбирали пробу 25 мкл и определяли остаточную активность. Константу скорости термоинактивации kin определяли как тангенс угла наклона прямой из графика зависимости натурального логарифма величины остаточной активности от времени (полулогарифмические координаты ln(А/A0) - t) методом линейной регрессии, используя программу Origin Pro 8.5.
Таким образом, были получены:
- рекомбинантная плазмида pHpFDH_HisA размером 6359 п.о., содержащая модифицированный ген hpfdh_hisa формиатдегидрогеназы из дрожжей H.polymorpha DL-1 под контролем T7 промотора. обеспечивающая синтез мутантной формиатдегидрогеназы с повышенной стабильностью из дрожжей H.polymorpha DL-1 (HpFDH_HisA) в клетках E. coli. Плазмида pHpFDH_HisA получена путем клонирования по сайтам NdeI/HindIII фрагмента размером 1113 п.о. (последовательность SEQ ID №3, фиг. 6) с геном, кодирующим HpFDH_HisA (1110 п.о.) в плазмиду pET24a(+), расщепленную рестриктазами NdeI и HindIII;
- рекомбинантный штамм Escherichia coli BL21(DE3)/pHpFDH_HisA - суперпродуцент мутантной формиатдегидрогеназы с повышенной стабильностью HpFDH_HisA, полученный путем трансформации штамма Escherichia. coli BL21(DE3) плазмидной ДНК.
Изобретение поясняется следующими примерами.
Пример 1. Введение дополнительных остатков в аминокислотную последовательность HpFDH
Анализ N-концевых участков аминокислотных последовательностей природных формиатдегидрогеназ из метилотрофных дрожжей свидетельствует, что во всех ферментах первым остатком является остаток лизина (фиг. 1). Исключение составляет формиатдегидрогеназа из пекарских дрожжей S.cerevisiae.
Согласно правилу N-конца Варшавского (Varshavsky ФA.The N-end rule pathway and regulation by proteolysis. Protein Sci. 2011 Aug; 20 (8): 1298-1345. doi: 10.1002/pro.666), белки и ферменты с остаткаи Lys и Arg наиболее подвержены деградации протеазами клетки. В работе (Klyushnichenko, V., Tishkov, V., Kula, M.-R. Rapid SDS-gel capillary electrophoresis for the analysis of recombinant NADP+-dependent formate dehydrogenase during expression in E.coli cells and purification. J.Biotechnol. - 1997, v. 8, N3, p. 87-195) было показано, что ФДГ из дрожжей C.boidinii (CboFDH) при экспрессии в клетках E.coli действительно подвергается достаточно быстрой деградации, в то время как ФДГ из бактерий Pseudomonas sp.101 (PseFDH) стабильна к действию протеаз штамма-хозяина. Для повышения устойчивости фермента к действию протеаз клетки рекомендуется на N-конец добавить остаток глицина. Ген CboFDH с дополнительным остатком глицина на N-конце описан в патенте (US2004115691, 2004-06-17), однако влияние дополнительного остатка Gly на свойства фермента не исследовали. Дополнительный остаток глицина был введен на N-конец HpFDH и было показано, что он не влияет ни на каталитические свойства, ни на термостабильность (Pometun A.A., Shaposhnikov L.A., Zubanova S.A., Kovalevskii R.P. Atroshenko D.L., Pometun E.V., Savin S.S., Tishkov V.I. NAD+-Dependent Formate Dehydrogenase from Themotolerant Yeast Ogataea parapolymorpha: Properties and Protein Engineering of the N-Terminal Sequence/ Biochemistry (Moscow), 2023, Vol. 88, No. 9, P. 1378-1389. DOI: 10.1134/S0006297923090171). PseFDH проявляет высокую стабильность к действию протеаз E.coli и содержит на N-конце остаток аланина, Поэтому было решено получить HpFDH с дополнительным остатком аланина на N-конце (HpFDH_A, Фиг. 1).
Второй модификацией гена hpfdh было введение нуклеотидной последовательности, кодирующей шесть остатков гистидина для очистки фермента в одну стадию с помощью метал-хелатной хроматографии. Для выбора оптимального положения (N- или С-конец) с помощью алгоритма AlphaFold2 были построены трехмерные модели фермента с дополнительными шестью остатками гистидина на N- и С-конце (Фиг. 2). Данные моделирования однозначно показывают, что His-tag на N-конце (Фиг. 2а) расположен от активного центра намного дальше, чем His-tag на С-конце (Фиг. 2б).
Результаты анализа N-концевой части аминокислотной последовательности и данные моделирования трехмерной структуры показали, что для создания штамма-продуцента формиатдегидрогеназ HpFDH с улучшенными свойствами в начало гена следует ввести нуклеотидную последовательность, кодирующую шесть остатков гистидина и один остаток аланина, Новый фермент получил обозначение HpFDH_HisA.
Пример 2. Конструирование экспрессионного вектора pHpFDH_HisA
Экспрессионный вектор pHpFDH_HisA получали клонированием в плазмиду pET24a(+) фрагмента ДНК с модифицированным геном hpfdh_hisa. Фрагмент ДНК с модифицированным геном hpfdh_hisa получали с помощью полимеразной цепной реакции (ПЦР). В качестве матрицы использовали плазмидну pHpFDH с геном фермента дикого типа, получение которой описано в (Pometun A.A., Shaposhnikov L.A., Zubanova S.A., Kovalevskii R.P. Atroshenko D.L Pometun E.V., Savin S.S., Tishkov V.I. NAD+-Dependent Formate Dehydrogenase from Themotolerant Yeast Ogataea parapolymorpha: Properties and Protein Engineering of the N-Terminal Sequence/ Biochemistry (Moscow), 2023, Vol. 88, No. 9, P. 1378-1389. DOI: 10.1134/S0006297923090171). Для проведения ПЦР использовали прямой
и обратный праймеры
В прямой праймер был введен сайт рестрикции NdeI CATATG (указан полужирным шрифтом). В обратном праймере для клонирования был введен сайт рестрикции HindIII AAGCTT, а вместо стоп-кодона TAG введен тандем стоп-кодонов TAATAA (выделено подчеркиванием).
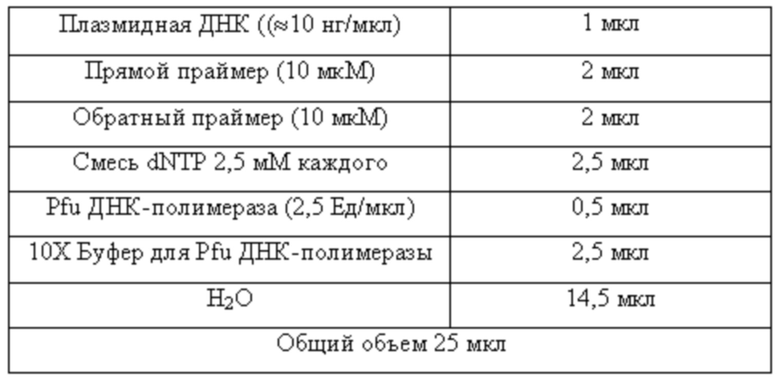
ПЦР проводили в тонкостенной пробирке объемом 0,5 мл. Состав реакционной смеси приведен в таблице 1.
Таблица 1
Состав реакционной смеси для проведения полимеразной цепной реакции
Перед началом ПЦР реакционную смесь без ДНК-полимеразы инкубировали 5 мин при 95°С, затем в пробирку добавляли фермент и проводили 35 циклов по следующей программе: 1-я стадия - 95°С, 30 с; 2-стадия - 62°С, 60 с и 3-я стадия - 72°С, 2 мин. После этого реакционную смесь выдерживали еще 10 мин при 72°С.
Продукт ПЦР очищали с помощью электрофореза в 1% агарозном геле. Вырезали фрагмент размером 1150 п.о и выделяли ДНК из геля с помощью набора для выделения и очистки ДНК из агарозного геля «PureLink Quick Gel Extraction Kit» (ThermoFisher Scientific).
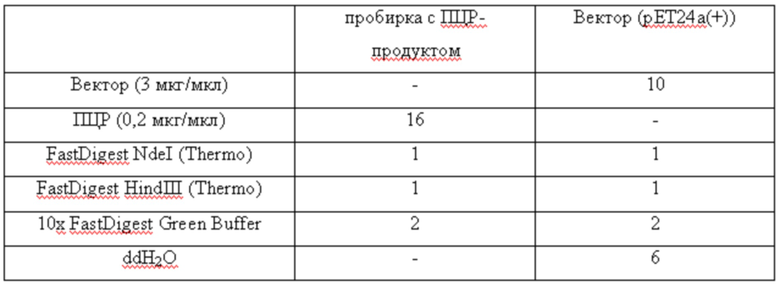
Очищенный ПЦР фрагмент ДНК и плазмиду pET24a(+) обрабатывают рестриктазами NdeI и HindIII. Условия проведения реакций представлены в таблице 2.
Таблица 2
Условия расщепления продукта ПЦР и плазмиды pET24a(+)
Реакцию проводят в пробирке объемом 0,5 мл в течение 1,5 часов при 37°С. Продукты обработки рестриктазами очищали с помощью электрофореза в 1% агарозном геле. Затем ДНК экстрагировали из геля и очищали с помощью набора для выделения и очистки ДНК из агарозного геля «PureLink Quick Gel Extraction Kit» (ThermoFisher Scientific).
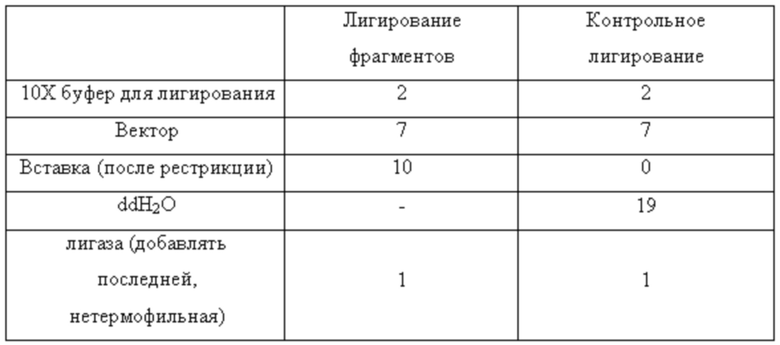
Очищенные продукты рестрикции лигировали при следующих условиях (Таблица 3).
Таблица 3
Условия лигирования продуктов рестрикции ПЦР и плазмиды pET24a(+)
Реакцию лигирования проводили в течение 3 часов при 22°С. Затем по стандартной методике [Sambrook J, Maniatis T, Fritsch EF. 1989. Molecular cloning a laboratory manual Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory] продуктом реакции лигирования трансформировали компетентные клетки E.coli DH5α (50 мкл компетентных клеток + 7 мкл лигазной смеси), После теплового шока клетки инкубировали в 1 мл среды LB при 37°С и затем высевали на агаризованную среду LB, содержащую 25 мкг/мкл канамицина, и инкубировали в течение ночи при 37°С.
Для выделения плазмид с чашки брали единичную колонию, инкубировали в 4 мл среды LB, содержащую 25 мкг/мкл канамицина, и инкубировали в течение ночи при 37°С. Полученные клетки осаждали на центрифуге при 5000 об/мин, 4°С 5 мин и из них выделяли плазмидную ДНК с помощью набора для выделения плазмидной ДНК «PureLink™ HiPure Plasmid Miniprep Kit». Выделение плазмид проводили в трех повторах.
Секвенирование полученных плазмид с помощью стандартных секвенирующих праймеров T7for и T7rev (Novagen) показало, что во всех плазмидах клонирована последовательность ДНК, соответствующая последовательности SEQ ID №3.
Полученная плазмида получила название pHpFDH_HisA.
Пример 3. Получение рекомбинантного штамма E.coli-продуцента HpFDH_HisA Полученной плазмидой pHpFDH_HisA трансформировали штамм E. coli BL21(DE3) ]F- ompT gal dcm lon hsdSB(rB-mB-) λ(DE3 [lacI lacUV5-T7p07 ind1 sam7 nin5]) [malB+]K-12(λS). Отбор клонов проводили на агаризованной среде LB, содержащую 25 мкг/мкл канамицина.
Для проверки эффективности продуцирования HpFDH_HisA полученным штаммом с чашки отбирали отдельные колонии и выращивали в 4 мл среды 2YT, содержащей 25 мкг/мкл канамицина, в течение ночи при 30°С. Затем отбирали 1 мл культуральной жидкости и переносили в колбу с отбойниками объемом 1 л со 100 мл среды 2YT, содержащей 25 мкг/мкл канамицина. Клетки культивировали при 30°С до поглощения на 600 нм А600 1 ед. поглощения, добавляли индуктор ИПТГ до концентрации 0,1 мМ. Через 2 часа после индукции в колбы добавляли раствор формиата натрия (20 г/л) до конечной концентрации 1 г/л и продолжали культивирование ещё 12-14 часов (20°С, 120 об/мин). Через 16 часов клетки осаждали на центрифуге Eppendorf 5804R в течение 30 мин при 6000 об/мин, 4°С. Полученный осадок ресуспендировали в 0,05М Tris-HCl, 0,5М NaCl, 20 мМ имидазол, pH 7,5 (концентрация клеток 10% по массе) и подвергали заморозке на -20°С. Культивирование проводили в трех повторах. Выход биомассы составил 14,6±2 г/л среды, выход по активности 6500±500 Ед активности с литра среды, что составляет почти 1,1 г целевого фермента.
Для очистки фермента суспензию клеток 20 мл подвергали заморозке - разморозке и затем обрабатывали ультразвуком при постоянном охлаждении 4°С - 4 цикла 1 мин озвучивания + 4 минуты охлаждения. Клеточный дебрис осаждали центрифугированием при 15000 об/мин в течение 20 мин при 4°С. Полученный раствор фермента в буфере 0,05М Tris-HCl, 0,5М NaCl, 20 мМ имидазол, pH 7,5 наносили на колонку объёмом 1 мл HisTrap FF Ni Sepharose 6 Fast Flow (Cytiva, США), уравновешенной раствором, содержащим 0,05М Tris-HCl, 0,5М NaCl, 20 мМ имидазол, pH 7,5 (раствор А). После нанесения фермента колонку промывали раствором А в течение 30 минут до исчезновения поглощения на 280 нм. Элюцию фермента с колонки проводили как описано выше. Выход по активности после очистки составил 90%. Результаты анализа чистоты полученного препарата HpFDH_HisA с помощью аналитического электрофореза в 12% полиакриламидном геле в присутствии додецилсульфата натрия представлены на Фиг. 3 (дорожка 3). Для сравнения приведены результаты анализа препаратов HpFDH дикого типа и с остатком аланина на N-конце HpFDH_A (дорожки 1 и 2 соответственно). Эти ферменты очищали по классической методике, включающей дезинтеграцию клеток ультразвуком, фракционирование сульфатом аммония, гидрофобную хроматографию на Phenyl Sepharose Fast Flow и гель фильтрацию через Сефакрил S-200. В общем, такая методика занимает не менее трех дней. Как видно из Фиг. 3 чистота препаратов HpFDH_HisA такая же, как и ферментов, очищенных по классической методике с несколькими стадиями хроматографии, по препарат HpFDH_HisA был получен всего за 2 часа. На Фиг. 3 также видно, что HpFDH_HisA имеет большую молекулярную массу, чем HpFDH и HрFDH_A за счет дополнительной последовательности His6Ala.
Таким образом, полученный рекомбинантный штамм E.coli BL21(DE3)/pHpFDH_HisA является суперпродуцентом рекомбинантной HpFDH_HisA. Полученный рекомбинантный штамм Escherichia coli BL21(DE3)/pHpFDH_HisA зарегистрирован во Всероссийской коллекции промышленных микроорганизмов как Escherichia coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559. Введене на N-конец фермента позволило получать с помощью метал-хелатной хроматографии высокоочищенные препараты в течение 2 часов вместо 3 дней при очистке с помощью класической методики.
Пример 4. Каталитические свойства и термостабильность HpFDH_HisA
В таблице 4 приведены значения каталитических параметров для HpFDH_HisA в присутствии и в отсутствие NaCl. Определение каталитических параметров в присутствии хлорида натрия обусловлено тем, что NaCl является компонентом физиологического раствора, который часто используется для манипуляций с биологическими образцами.
Таблица 4
Каталитические свойства различных формиатдегидрогеназ*
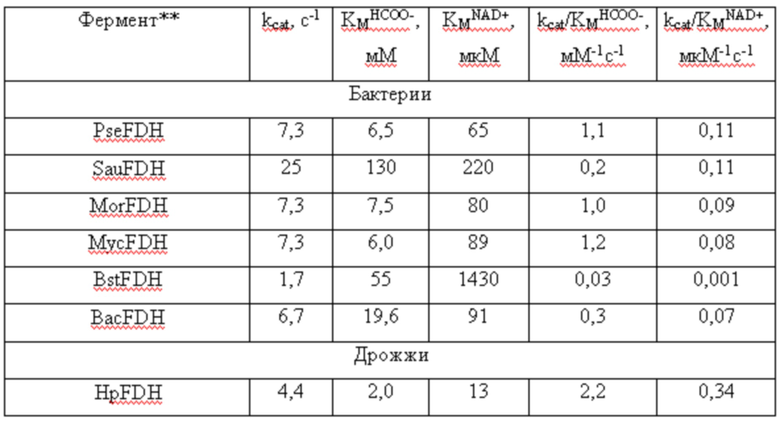
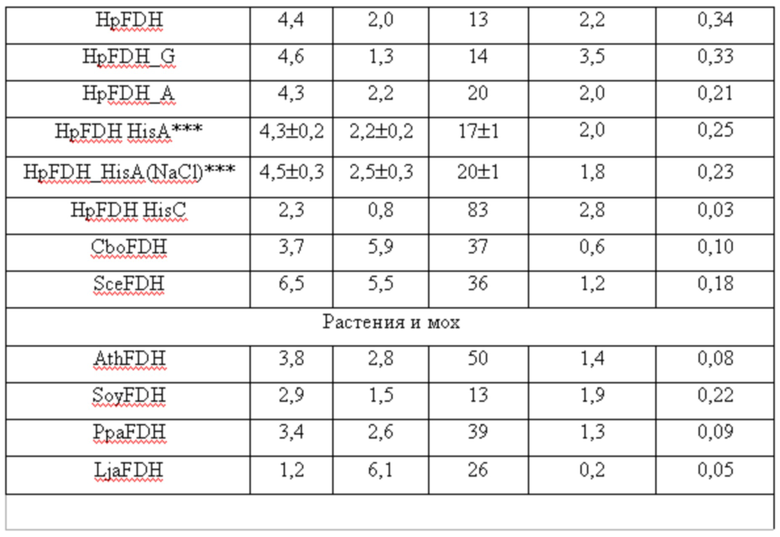
*литературные данные для ферментов за исключением HpFDH_HisA (Pometun A.A., Shaposhnikov L.A., Zubanova S.A., Kovalevskii R.P. Atroshenko D.L .., Pometun E.V., Savin S.S., Tishkov V.I. NAD+-Dependent Formate Dehydrogenase from Themotolerant Yeast Ogataea parapolymorpha: Properties and Protein Engineering of the N-Terminal Sequence/ Biochemistry (Moscow), 2023, Vol. 88, No. 9, P. 1378-1389. DOI: 10.1134/S0006297923090171).
**Сокращения: PseFDH - ФДГ из бактерий Pseudomonas sp 101, SauFDH - ФДГ из золотистого стафилококка S.aureus, MorFDH - ФДГ из бактерий Moraxella sp. C2, MycFDH - ФДГ из бактерий Mycobacterium vaccae, BstFDH - ФДГ из бактерий Burkholderia stabilis, BacFDH - ФДГ из бактерий Bacillus sp. F1, CboFDH - ФДГ из дрожжей C.boidinii, AthFDH - ФДГ из модельного растения Arabidopsis thaliana, SoyFDH - ФДГ из сои Glycine max, PpaFDH - ФДГ из мха Physcomitrella patens, LjaFDH - ФДГ из Lotus japonicus.
***Результаты являются средним из трех измерений.
Из данных, приведенных в таблице 4, следует:
1. Все варианты HpFDH имеют самую высокую каталитическую эффективность как с формиатом, так и с НАД+. Именно поэтому данный фермент является самым лучшим для определения формиата и кофермента НАД+ в сложных биологических образцах.
2. Наличие на N-конце HpFDH дополнительных остатков не влияет на каталитические свойства этого фермента. Кроме того, каталитические параметры продуцируемой заявляемым штаммом-продуцентом рекомбинантной HpFDH_HisA не изменяются в присутствии высокой концентрации соли, что исключает влияние на точность и чувствительность определения формиата и НАД+ компонентов буферных растворов, используемых при подготовки биологических образцов.
На Фиг. 5. приведены зависимости остаточной активности от времени для HpFDH_HisA при 64°С. Для сравнения приведены данные по термоинактивации других форм фермента - HpFDH дикого типа и с дополнительными на N-конце остатком глицина (HpFDH_G) и аланина (HpFDH_A). Данные на фиг. 5 свидетельствуют, что введение на N-конец остатка глицина не влияет на термостабильность, но в случае введения на N-конец остатка аланина стабильность фермента при 64°С возрастает в 3 раза. Эффект для точечной замены большой и, самое главное, неожиданный и абсолютно непредсказуемый. Согласно данным моделирования этот остаток аланина расположен на поверхности белковой глобулы вдали от активного центра и не входит ни в один из структурных элементов (альфа-спирали ли бета-листа). Разумных предположений о механизме стабилизации просто нет. Тем не менее, повышение термостабильности существенно улучшает эксплуатационные качества фермента. Добавление в HpFDH_A последовательности из 6 остатков гистидина в стандартных для измерения стабильности ФДГ условиях (0,1 М фосфатный буфер, рН 7,0) нивелирует эффект стабилизации. Однако в присутствии 0,5 М хлорида натрия термостабильность HpFDH_HisA пи 64°С восстанавливается почти до уровня стабильности HpFDH_A (константы скорости термоинактивации 0,09 и 0,07 с-1 соответственно по сравнению с 0,21 с-1 для HpFDH и HpFDH_G). При 65°С HpFDH_A и HpFDH_HisA в пределах ошибки эксперимента имеют одинаковые значения константы скорости инактивации (0,135 и 0,141 с-1 соответственно).
Таким образом, можно сделать вывод, что созданный нами рекомбинантный штамм Escherichia coli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559 - суперпродуцент HpFDH_HisA, позволяет получить фермент, который по совокупности свойств имеет гораздо больший биотехнологический и биоаналитический потенциал по сравнению с другими вариантами формиатдегидрогеназ, в том числе и вариантами HpFDH. При равных каталитических свойствах и термостабильности с HpFDH_A (самая термостабильная среди ФДГ эукариот) HpFDH_HisA превосходит HpFDH_A по скорости, простоте и соответственно по стоимости получения.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Плазмида
pHpFDH_HisA.xml" softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-12-27">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate></FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>105-2023</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное образовательное учреждение высшего образования
"Московский государственный университет имени
М.В.Ломоносова" (МГУ)</ApplicantName>
<ApplicantNameLatin>Federal state budgetary education institution of
higher education "Lomonosov Moscow State University"
(MSU)</ApplicantNameLatin>
<InventionTitle languageCode="ru">РЕКОМБИНАНТНАЯ ПЛАЗМИДА
pHpFDH_HisA, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ В КЛЕТКАХ Escherichia coli
ФОРМИАТДЕГИДРОГЕНАЗЫ МЕТИЛОТРОФНЫХ ДРОЖЖЕЙ Hansenula polymorpha
(HpFDH_HisA) И РЕКОМБИНАНТНЫЙ ШТАММ E.сoli BL21(DE3)/pHpFDH_HisA ВКПМ
В-14559 - ПРОДУЦЕНТ HpFDH_HisA</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>55</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..55</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>caaggagatatacatatgcaccaccaccaccaccacgccaaggttgtac
tagttc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>37</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..37</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cagctccttaagcttttattatttgtcggcaccgtac</INSDSeq_se
quence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>1113</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1113</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgcaccaccaccaccaccacgccaaggttgtactagttctctacgacg
caggaaaacacgcccaagacgaggaaagactctacggttgcaccgaaaacgcccttggtatcagggactg
gcttgagaagcagggccacgagctcgttgtcaccagtgacaaggagggcgagaattctgtgctcgagaag
aacatcccggacgcagatgtcatcatctccactcctttccacccggcctacatcactaaggaaagaattg
acaaggccaagaagctcaagttgctggtggttgccggagtgggatccgaccacatcgaccttgactacat
caaccagtccggcagagacatttctgtgctggaggtgaccggttcgaacgtggtttccgttgccgagcac
gtcgtgatgacgatgctggtgctggtcagaaactttgttcctgcccacgagcaaatcatctctggcggct
ggaacgtggccgagattgccaaggactccttcgatatcgagggtaaggtcattgccaccatcggagcagg
cagaatcggctaccgtgtgttggagagacttgtcgccttcaaccctaaggagctgctctactacgactac
cagtcgctgtccagagaggccgaggagaaagtcggagcccgcagagttcacgacatcaaggagctggttg
cccaggccgacattgtcacaatcaactgtccactgcacgccggctcgaagggcctggtcaacgcagagtt
gctcaagcacttcaagaagggtgcctggctcgttaacaccgccagaggtgccatttgtgtggccgaggac
gttgcagcagccgtcaagagcggacagctcagaggatacggtggagatgtgtggtacccacagccagctc
caaaggaccacccatggagatctatggccaacaagtacggtgctggtaatgccatgactccgcactactc
gggctctgtcattgacgcccaggtcagatacgcccagggtaccaagaacatcctggagtcattcttcact
cagaagttcgactacagacctcaggacatcattctgctgaacggtaagtacaagaccaagtcgtacggtg
ccgacaaataataa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ РАСТИТЕЛЬНАЯ ФОРМИАТДЕГИДРОГЕНАЗА (ВАРИАНТЫ) | 2014 |
|
RU2557299C1 |
| МУТАНТНАЯ ФОРМИАТДЕГИДРОГЕНАЗА (ВАРИАНТЫ) | 2013 |
|
RU2545966C1 |
| МУТАНТНАЯ ФОРМИАТДЕГИДРОГЕНАЗА (ВАРИАНТЫ) | 2012 |
|
RU2522819C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pNhaDAAO, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ ОБЫЧНЫХ И N-ЗАМЕЩЕННЫХ D-АМИНОКИСЛОТ (NhaDAAO) АРХЕЙ Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO) В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 - ПРОДУЦЕНТ NhaDAAO | 2023 |
|
RU2829362C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pNhyDAAO, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ D-АМИНОКИСЛОТ БАКТЕРИЙ Natronosporangium hydrolyticum ACPA39 (NhyDAAO) В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pNhyDAAO ВКПМ В-14337 - ПРОДУЦЕНТ NhyDAAO | 2022 |
|
RU2803295C1 |
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ФОРМИАТДЕГИДРОГЕНАЗА | 2006 |
|
RU2312897C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pETTvDAO2, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ D-АМИНОКИСЛОТ (DAO) ДРОЖЖЕЙ Trigonopsis variabilis В КЛЕТКАХ Escherichia coli И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli C41(DE3)/pETTvDAO2 - ПРОДУЦЕНТ DAO | 2006 |
|
RU2310687C1 |
| РЕКОМБИНАНТНАЯ ДНК, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНЫЙ ГИБРИДНЫЙ БЕЛОК ОКСИДАЗЫ D-АМИНОКИСЛОТ С ХИТИНСВЯЗЫВАЮЩИМ ДОМЕНОМ (DAOcbd) РЕКОМБИНАНТНАЯ ПЛАЗМИДА pVR1, ОБЕСПЕЧИВАЮЩАЯ ЕГО СИНТЕЗ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli С 41(DE3)/pVR1 - ПРОДУЦЕНТ DAOcbd | 2006 |
|
RU2310688C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ ИЗ Xanthomonas rubrilineans И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ НА ОСНОВЕ ЭТОГО ШТАММА | 2012 |
|
RU2499830C1 |
| Рекомбинатная плазмида pET21a-ProChym, обеспечивающая синтез химерного белка прохимозина В Bos taurus, и штамм Escherichia coli BL21(DE3)pLysE pET21a-ProChym - продуцент химерного белка прохимозина В Bos Taurus | 2017 |
|
RU2670071C1 |

Изобретение относится к области биотехнологии, в частности к генетической инженерии. Была сконструирована рекомбинантная плазмидная ДНК pHpFDH_HisA, содержащая модифицированный ген hpfdh_hisa, кодирующий мутантную `НАД+-зависимую формиатдегидрогеназу (КФ 1.2.1.2, HpFDH_HisA) c дополнительной на N-конце фермента последовательностью из шести остатков гистидина и одного остатка аланина. Введение на N-конец фермента дополнительной последовательности His6Ala приводит к повышению термостабильности и позволяет в одну стадию получать препараты высокоочищенного фермента. Для обеспечения высокой эффективности экспрессии в целевом штамме-продуценте в модифицированном гене HpFDH_HisA в качестве стоп кодона используется тандем триплетов TAATAA вместо триплета TGA в исходном гене. Полученная плазмида позволяет проводить экспрессию включенного в нее гена hpfdh_hisa в клетках Escherichia coli. В результате трансформации штамма E.coli BL21(DE3) предложенной плазмидой получен новый рекомбинантный штамм-продуцент E.coli BL21(DE3))/pHpFDH_HisA ВКПМ В-14559, обеспечивающий суперэкспрессию HpFDH_HisA в активной форме. 2 н.п. ф-лы, 6 ил., 4 табл., 4 пр.
1 Рекомбинантный вектор pHpFDH_HisA для экспрессии мутантной формиатдегидрогеназы из дрожжей Hansenula polymorpha (HpFDH_HisA), содержащий NdeI-HindIII фрагмент размером 5246 п.о. вектора pET24a(+) и NdeI-HindIII фрагмент размером 1113 п.о., представляющий собой нуклеотидную последовательность, соответствующую SEQ ID № 3.
2. Рекомбинантный штамм Escherichia coli BL21(DE3)/pHpFDH_HisA – продуцент мутантной HpFDH_HisA с повышенной стабильностью, полученный путем трансформации штамма Escherichia. coli BL21(DE3) плазмидной ДНК по п.1, задепонированный во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под номером В-14559.
| EP 1900821 A1, 19.03.2008 | |||
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ФОРМИАТДЕГИДРОГЕНАЗА | 2006 |
|
RU2312897C1 |
| МУТАНТНАЯ ФОРМИАТДЕГИДРОГЕНАЗА (ВАРИАНТЫ) | 2013 |
|
RU2545966C1 |
| KR 101564315 B1, 29.10.2015. | |||

