Изобретение относится к области генной и белковой инженерии и может быть использовано в процессах синтеза оптически активных соединений или физиологически активных веществ, детекции формиата, а также для получения стрессоустойчивых растений.
Получены три новые мутантные формы растительной формиатдегидрогеназы (ФДГ), обладающие улучшенными свойствами по сравнению с белком дикого типа. В первом из предложенных мутантных белков природный остаток фенилаланина в положении, соответствующем положению 290 в последовательности формиатдегидрогеназы из сои Glycine max, заменен на остаток аспарагина. Во второй мутантной форме фермента этот же природный остаток фенилаланина 290 заменен на остаток аспарагиновой кислоты, а в третьей - на остаток серина. Данные замены могут быть использованы в комбинации с другими мутациями, обеспечивающими приобретение формиатдегидрогеназами новых или улучшенных свойств.
Данное изобретение относится к новому белку, в частности к мутантным формиатдегидрогеназам из растений, которые обладают улучшенными свойствами по сравнению с соответствующим ферментом дикого типа, к ДНК, кодирующей мутантные формы формиатдегидрогеназы, к применению данных ферментов в системах синтеза оптически активных соединений или физиологически активных веществ, для детекции формиата, а также для получения стрессоустойчивых растений.
NAD+-зависимая формиатдегидрогеназа катализирует окисление формиата до углекислого газа при сопряженном восстановлении NAD+ до NADH.

ФДГ обладает высокой специфичностью к формиату, поэтому этот фермент успешно применяют для определения формиата в сложных смесях.
Реакция окисления формиат-иона является необратимой, продуктом является углекислый газ, который можно легко вывести из сферы реакции. Также ФДГ обладает широким рН-оптимумом активности. Поэтому формиатдегидрогеназа может быть эффективно использована для регенерации NADH в процессах синтеза оптически активных соединений с использованием дегидрогеназ. В качестве примера можно привести промышленный процесс получения терт-L-лейцина из триметилпирувата с использованием лейциндегидрогеназы с системой регенерации NADH на основе формиатдегидрогеназы из дрожжей Candida boidinii (Bommarius A.S., Schwarm M., Stingl K., Kottenhahn M., Huthmacher K. and Drauz K., Synthesis and Use of Enantiomerically Pure tert. - Leucine, Tetrahedron Asymmetry 1995, v.6, pp.2851-2888).
В настоящее время часто применяют способ, в котором процесс синтеза и регенерацию осуществляют непосредственно к бактериальной клетке. Так, ген ФДГ из дрожжей Candida boidini был экспрессирован в E.coli и использован в процессах синтеза ксилитола из ксилозы и S-1-(2-хлорфенил) этанола из о-хлорацетофенона (Mädje К., Schmölzer К., Nidetzky В., Kratzer R. Host cell and expression engineering for development of an E.coli ketoreductase catalyst: Enhancement of formate dehydrogenase activity for regeneration of NADH. Microb Cell Fact. 2012, v.11, pp.11-17).
Формиатдегидрогеназа найдена в бактериях, дрожжах, микроскопических грибах, а также растениях. Впервые формиатдегидрогеназная активность в растениях была обнаружена в 1921 г. (Thunberg Т. Sur la presence de certains ferments oxydants dans les grains de phaseolus vulgaris. Arch. int. Physiol., 1921, v.18, pp.601-606). В растениях ФДГ - это фермент стресса, синтез которого резко возрастает при таких стрессовых воздействиях, как засуха, повышенная доза ультрафиолета, резкие скачки температуры, нехватка кислорода и т.п. (Hourton-Cabassa С., Ambard-Bretteville F., Moreau F., Davy de V., Remy R., and Francs-Small С.С. Stress Induction of Mitochondrial Formate Dehydrogenase in Potato Leaves. Plant Physiol, 1998, v.116, pp.627-635; Suzuki K., Itai R., Suzuki K., Nakanishi H., Nishizawa N.K., Yoshimura E., and Mori S. Formate dehydrogenase, an enzyme of anaerobic metabolism, is induced by iron deficiency in barley roots. Plant Physiol, 1998, v.116, pp.725-732; Thompson P., Bowshеr C.G., and Tobin A.K. Heterogeneity of mitochondrial protein biogenesis during primary leaf development in barley. Plant Physiol, 1998, v.118, p.1089-1099; Andreadеli A., Flemetakis R., Axarli I., Dimou M., Udvardi M.K., Katinakis P., and Labrou N.E. Cloning and characterization of Lotus japonicus formate dehydrogenase: a possible correlation with hypoxia. Biochim. Biophys. Acta, 2009, v.1794, pp.976-984; David P., des Francs-Small C.C., Sevignac M., Thareau V., Macadre C., Langin Т., and Geffroy V. Three highly similar formate dehydrogenase genes located in the vicinity of the B4 resistance gene cluster are differentially expressed under biotic and abiotic stresses in Phaseolus vulgaris. Theor. Appl Genet., 2010, v.121, pp.87-103). Подробный обзор по физиологической роли, получению и свойствам ФДГ из растений был опубликован в 2011 году (Алексеева А.А., Савин С.С., Тишков В.И. NAD+-зависимая формиатдегидрогеназа растений. Acta Naturae, 2011, т.3, №4(11), с.40-56).
Многие из генов, кодирующих данные ферменты, были клонированы и секвенированы, поэтому их можно получать с использованием технологии рекомбинантных ДНК. Последовательность кДНК, кодирующую формиатдегидрогеназу, встраивают в генно-инженерный вектор (плазмида, фаг и т.д.), который используют для трансформации микроорганизмов, например бактерий E.coli. Полученный рекомбинантный штамм используют для экспрессии желаемого ферментного продукта.
Для растительных формиатдегидрогеназ характерны низкие значения констант Михаэлиса по сравнению с ферментами из других источников. Одной из наиболее перспективных для практического применения формиатдегидрогеназ из растений является формиатдегидрогеназа из сои Glycine max (SoyFDH). Этот фермент был получен в активной и растворимой форме и он имеет самые низкие значения констант Михаэлиса по сравнению со всеми изученными формиатегидрогеназами на настоящий момент (Садыхов Э.Г., Серов А.Е., Ясный И.Е., Войнова Н.С., Алексеева А.А., Петров А.С., Тишков В.И. NAD+-зависимые формиатдегидрогеназы из Arabidopsis thaliana и сои: экспрессия в клетках E.coli и кинетические свойства рекомбинантных ферментов. Вестн. Моск. Ун-та. Сер.2. Химия, 2006, т.47, N1, c.31-34).
Основными недостатками известных природных и рекомбинантных формиатдегидрогеназ из растений дикого типа является невысокая температурная стабильность этих ферментов. Таким образом, для успешного применения формиатдегидрогеназы из сои на практике необходимо повысить стабильность этого фермента. Одним из подходов для решения поставленной задачи является мутагенез аминокислотных остатков целевого белка.
Для бактериальных и дрожжевых ферментов в литературе описано много примеров успешного улучшения свойств методами белковой инженерии. Для повышения термостабильности формиатдегидрогеназы из бактерий Pseudomonas sp.101 (PseFDH) были использованы подходы, основанные на гидрофобизации α-спиралей (замены остатков Ser в α-спиралях на Ala) (Rojkova A.M., Galkin A.G., Kulakova L.B., Serov A.E., Savitsky P.A., Fedorchuk V.V., and Tishkov V.I. Bacterial formate dehydrogenase. Increasing the еnzyme thermal stability by hydrophobization of alpha-helices. FEBS Lett., 1999, v.445, pp.183-188), и снятия конформационных напряжений в полипептидной цепи (замены объемных Asn и Tyr на Gly) (Serov A.E. and Tishkov V.I. Role of Pro residues in stability of prokaryotic and euacaryotic formate dehydrogenases. Вестн. Моск. Ун-та. Сер. 2. Химия, 2002, v.43, pp.345-349; Serov A.E., Odintzeva E.R., Uporov I.V., and Tishkov V.I. Use of Ramachandran plot for increasing thermal stability of bacterial formate dehydrogenase. Biochemistry (Mosc.), 2005, v.70, pp.804-808). Поиск положений для мутагенеза был основан на данных рентгеноструктурного анализа и сравнении аминокислотных последовательностей ФДГ из разных источников. В результате единичной замены эффект стабилизации был небольшим - от 10 до 50%, но практически во всех случаях - аддитивным (Tishkov, V.I., Popov, V.O. Protein engineering of formate dehydrogenase. Biomolеcular Engineering 2006, v.23, N2-3, p.89-110. Мутации, повышающие операционную и температурную стабильность, были объединены в мутанте PseFDH GAV. В результате такого объединения сродство к NAD+ увеличилось в 2 раза, а температурная стабильность - в 2,5 раза, а операционная - более, чем в 1000 раз по сравнению с PseFDH дикого типа. Получение мутантных PseFDH с улучшенной термостабильностью позволило ввести в процесс очистки рекомбинантного фермента стадию термообработки. Нагревание бесклеточного экстракта при 60°С в течение 20-30 мин приводит к повышению чистоты препарата PseFDH GAV с 50 до 80-85% без потери ферментативной активности (Tishkov V.I. and Popov V.O. Protein engineering of formate dehydrogenase. Biomol. Eng., 2006, v.23, pp.89-110).
Также в литературе имеются данные по увеличению стабильности ФДГ из дрожжей С. boidini. В работе (Slusarczyk Н., Felber S., Kula M.R., and Pohl M. Novel mutants of formate dehydrogenase from Candida boidinii. US Patent Application Publication, 2003, US 2003/0157664) получен химически более стабильный мутант CboFDH SM с заменой Cys23Ser, который был гораздо устойчивее фермента дикого типа в растворах 150 мМ H2O2 и 50 мкМ CuSO4, однако его термостабилытость уменьшилась в 6,7 раз (уменьшение Тm на 5°С). Для компенсации потери стабильности использовали метод направленной эволюции. Были найдены 4 мутации, дающие наибольший эффект стабилизации (Glu151Asp, Arg178Ser, Lys306Arg, Thr315Asn), которые повысили термостабильность мутанта CboFDH_SM в 36 раз (увеличение Тm на 10°С), а по сравнению с ферментом дикого типа - в 5,7 раз.
Техническая задача, решаемая посредством предлагаемого технического решения, состоит в получении мутантных форм формиатдегидрогеназы из сои Glycine max с улучшенными свойствами по сравнению с ферментом дикого типа.
Технический результат, получаемый при реализации предлагаемого технического решения, состоит в улучшении свойств полученной мутантной формы формиатдегидрогеназы из сои Glycine max по отношению к свойствам фермента дикого типа.
Для достижения указанного технического результата предложено использовать рекомбинантную растительную формиатдегидрогеназу, в которой в качестве основы использована аминокислотная последовательность формиатдегидрогеназы из сои Glycine max дикого типа. Полная последовательность кДНК этого фермента (код доступа в базе GeneBank ВТ094321.1) содержит в начале нуклеотидную последовательность, кодирующую сигнальный пептид, необходимый для транспорта белка-предшественника формиатдегидрогеназы из цитоплазмы в митохондрии клеток растений. После попадания про-белка внутрь митохондрии этот пептид отщепляется с образованием активной формиатдегидрогеназы. В связи с отсутствием в бактериях митохондрии, вышеописанная посттрансляционная модификация белка-предшественника формиатдегидрогеназы невозможна. Поэтому для экспрессии в клетках Е.соli из последовательности кДНК формиатдегидрогеназы сои была удалена последовательность, кодирующая сигнальный пептид (31 аминокислотный остаток). В результате синтезируется активный рекомбинантный фермент дикого типа без дополнительной стадии посттрансляционной модификации (Алексеева А.А, Савин С.С., Тишков В.И. NAD+-зaвиcимaя формиатдегидрогеназа растений. Acta Naturae, 2011, т.3, №4(11), с.40-56).
В дальнейшем получение и преимущества полученной мутантной формиатдегидрогеназы будут рассмотрены с использованием примеров получения и реализации этого фермента.
Пример 1.
Получение мутантной формиатдегидрогеназы, в которой остаток Phe290 заменен на остаток Asn.
Направленный мутагенез проводили с использованием полимеразной цепной реакции (ПЦР). В качестве матрицы для проведения направленного мутагенеза использовали плазмиду pSoyFDH. Плазмида pSoyFDH получена из коммерчески доступной плазмиды рЕТ21а (Novagen, США), в которую по сайтам рестрикции NdeI и EcoRI встроена кДНК формиатдегидрогеназы сои Glycine max без участка, кодирующего сигнальный пептид.
Для введения мутации Phe290Asn в ген формиатдегидрогеназы с использованием ПЦР применяли универсальные праймеры Т7 for и Т7 rev:
T7For 5' - TAATACGACTCACTATAGGG - 3'
T7 rev 5' - GCTAGTTATTGCTCAGCGG - 3',
а также прямой и обратный праймеры, несущие требуемую замену в триплете, кодирующем в гене soyfdh остаток Asn в положении 290:
SoyF290Nfor 5' - TGATGTTTGG AAC CCACAGCCAGCTCCAAA - 3'
SoyF290Nrev 5' - GGCTGTGG GTT CCAAACATCACCACTATAACCT -3'
Полужирным шрифтом с подчеркиванием выделены нуклеотидные замены, обеспечивающие мутацию Phe290Asn.
Реакцию ПЦР проводили в тонкостенной пластиковой пробирке объемом 0,5 мл («SSI», США). Для предотвращения испарения реакционной смеси и ее конденсации на крышке в пробирку добавляли 30 мкл минерального масла. Пробирку прогревали 5 мин при 95°С и затем проводили реакцию ПЦР по следующей программе: 1-я стадия - 95°С, 60 с; 2-стадия - 56°С, 60 с и 3-я стадия - 72°С, 2 мин, всего 25-35 циклов. После этого реакционную смесь выдерживали еще 10 мин при 72°С. Температуру на второй стадии выбирали на 3-5°С ниже Тm температуры плавления дуплексов, образуемых праймерами.
Реакционная смесь для проведения ПЦР имела следующий состав (таблица 1):
На первом этапе проводили две полимеразные цепные реакции с использованием следующих пар праймеров: 1) T7For+SoyF290Nrev и 2) SoyF290Nfor+T7Rev. В качестве матрицы использовали плазмиду pSoyFDH. Полученные продукты ПЦР, фрагмент 1 и фрагмент 2, очищали с использованием препаративного электрофореза в агарозном геле с применением предназначенных для этого наборов, например GeneJet Gel Extraction Kit (Fermentas, Литва). Затем проводили третью, объединяющую ПЦР с праймерами T7For и T7Rev, где в качестве ДПК-матрицы использовали полученные ранее фрагменты 1 и 2.
Продукт третьей ПЦР очищали аналогично фрагментам 1 и 2 и обрабатывали эндонуклеазами рестрикции KpnI и EcoRI в течение двух часов при 37°С, затем инкубировали рестрикционную смесь для инактивации рестриктаз в течение 20 минут при 65°С (использовали рестриктазы и буферные растворы фирмы Fermentas (Литва)). Затем полученные фрагменты ДНК очищали от коротких фрагментов препаративным электрофорезом в агарозном геле с последующей экстракцией нужной полосы из агарозы с использованием специального набора, например, GeneJet Gel Extraction Kit (Fermentas, Литва), и лигировали с фрагментом исходной плазмиды pSoyFDH, из которой расщеплением по тем же сайтам рестрикции KpnI и EcoRI был удален фрагмент гена soyfdh из G.max дикого типа.
Полученной после лигирования реакционной смесью трансформировали клетки Е.coli TG1 (генотип supE hsdΔ5 thi Δ(lасproAB) F'[traD36 proAB+ lacIq lacZΔM15]) и выращивали в течение ночи при 37°С на твердой агаризованной среде LВ, содержащей ампициллин в концентрации 100 мкг/мл (плазмида pSoyFDH содержит ген устойчивости к ампициллину).
Выросшие колонии инокулировали в 6-8 мл среды 2YT, содержащей 150 мкг/мл ампициллина, и растили при 37°С в течение 8-10 часов при аэрировании. Плазмидную ДНК выделяли из полученной культуры клеток с использованием предназначенного для этого набора, например GeneJet Plasmid Miniprep Kit (Fermentas, Литва). Для контроля введения требуемых мутаций проводили секвенирование плазмидной ДНК в Центре коллективного пользования «Геном» (Институт молекулярной биологии им. В.А.Энгельгардта РАН). В результате из 5 выделенных плазмид все 5 содержали только требуемую замену Phe290Asn. Таким образом, была получена плазмида pSoyFDH_F290N, которая обеспечивала получение повой формиатдегидрогеназы SoyFDH Phe290Asn с заменой Phe290Asn.
Для получения мутантного фермента клетки E.coli BL21 (DE3) pLysS Codon Plus (генотип E.coli В F- ompT hsdS(rB- mB-) dcm+ Tetr gal λDE3)pLysS endA Hte [argU proL Camr]) трансформировали плазмидой pSoyFDFI_F290N, высевали на чашки Петри с агаризованной средой, содержащей анпициллин (100 мкг/мл) и хлорамфеникол (25 мкг/мл) и проводили их культивирование (см. ниже).
Пример 2.
Получение мутантной растительной формиатдегидрогеназы, и которой остаток Phe290 заменен на остаток Asp.
Данный пример идентичен примеру 1 за исключением того, что к качестве прямого и обратного праймеров, обеспечивающих введение замены Phe290Asp, использовали следующие дезоксирибоолигопуклеотиды:
Soy_F290Dfor 5' - TGATGTTTGG GAC CCACAGCCAGCTCCAAA - 3'
Soy_F290Drev 5' - GCTGTGG GTC CCAAACATCACCACTATAACCT - 3'
Полужирным шрифтом с подчеркиванием выделены нуклеотидные замены, обеспечивающие мутацию Phe290Asp.
Плазмида pSoyFDH с нуклеотидными заменами, обеспечивающими в гене фермента мутацию Phe290Asp, получила название pSoyFDH_F290D.
Для получения мутантного фермента клетки E.coli BL21 (DE3) pLysS Codon Plus (генотип Е.coli В F- ompT hsdS(rB- mВ-) dcm+ Tetr gal λ(DE3)pLysS endA Hte [argU proL Camr]) трансформировали плазмидой pSoyFDH_F290D, высевали на чашки Петри с агаризованной средой, содержащей ампициллин (100 мкг/мл) и хлорамфеникол (25 мкг/мл) и проводили их культивирование (см. ниже).
Пример 3.
Получение мутантной растительной формиатдегидрогеназы, в которой остаток Phe290 заменен на остаток Ser.
Данный пример идентичен примеру 1 за исключением того, что в качестве прямого и обратного праймеров, обеспечивающих введение замены Phe290Ser, использовали следующие дезоксирибоолигонуклеотиды:
Soy_F290SFor 5' - GATGTTTGG TCC CCACAGCCAGCTCCAAA - 3'
Soy_F290SRev 5' - GCTGTGG GGA CCAAACATCACCACTATAACCT - 3'
Полужирным шрифтом с подчеркиванием выделены нуклеотидные замены, обеспечивающие мутацию Phe290Ser.
Плазмида pSoyFDH с нуклеотидными заменами, обеспечивающими в гене фермента мутацию Phe290Ser, получила название pSoyFDH_F290S.
Для получения мутантного фермента клетки E.coli BL 21 (DE3) pLysS Codon Plus (генотип E.coli В F- ompT hsdS(rB- mB-) dcm+ Tetr gal λ(DE3)pLysS endA Hte [argU proL Camr]) трансформировали плазмидой pSoyFDH_F290S, высевали на чашки Петри с агаризованной средой, содержащей ампициллин (100 мкг/мл) и хлорамфеникол (25 мкг/мл) и проводили их культивирование (см. ниже).
Пример 4.
Изучение температурной стабильности мутантных форм формиатдегидрогеназы SoyFDH с заменами F290N, F290D, F290S и фермента дикого типа.
Для точной оценки эффекта изменения температурной стабильности за счет введенных замен мутантный фермент SoyFDH Phe290Asn и формиатдегидрогеназу дикого типа выделяли в высокоочищенном состоянии и определяли зависимость остаточной активности от времени инкубации при 56°С.
Методика получения высокоочищенных препаратов была одинакова как для мутантных SoyFDH Phe290Asn, SoyFDH Phe290Asp, SoyFDH Phe290Ser, так и для фермента дикого типа.
Экспрессию SoyFDH дикого типа и ее мутантов проводили к клетках E.coli BL21 (DE3) CodonPlus/pFysS. Для приготовления посевного материала с чашки отбирали единичную колонию и культивировали в течение 7-9 ч при 30°С и 180 об/мин до достижения величины поглощения на длине волны 600 нм А600≈0,6-0,8 в 5 мл среды 2YT (дрожжевой экстракт 10 г/л, бактотриптон 16 г/л, хлорид натрия 5 г/л, рН 7.0) в присутствии 150 мкг/мл ампициллина и 25 мкг/мл хлорамфеникола. Затем содержимое пробирок переносили в конические качалочные колбы с отбойниками объемом 1 л, содержащими 200 мл среды 2YT и 150 мкг/мл ампициллина и клетки культивировали при 30°С и 80-90 об/мин до достижения величины поглощения на 600 нм, А600=0,6-0,8. Далее проводили индукцию клеток, добавляя в среду для культивирования раствор лактозы (300 г/л) до конечной концентрации индуктора 20 г/л. После индукции температуру культивирования снижали до 20°С и клетки культивировали в течение 17 ч при 120 об/мин. Полученную биомассу осаждали на центрифуге Beckman J-21 (США) при 7500 об/мин в течение 20 мин при 4°С и после удаления культуральной жидкости клетки ресуспендировали в 0,1 М натрий-фосфатном буфере, рH 8,0 в соотношении 1:4 (масс.). Полученную суспензию замораживали и хранили при -20°С.
Для выделения мутантных SoyFDH Phe290Asn, SoyFDH Phe290Asp, SoyFDH Phe290Ser, и фермента дикого типа 20% суспензию клеток в 0,1 М натрий-фосфатном буфере, рH 8,0 подвергали двум циклам заморозки-разморозки, и затем клетки разрушали с использованием ультразвукового дезинтегратора «Branson Sonifier 250» (Германия) при постоянном охлаждении. Осадок удаляли центрифугированием на центрифуге «Eppendorf 5804 R»(11000 об/мин, 30 мин).
Для очистки использовали модифицированную методику, разработанную для получения рекомбинантной ФДГ из бактерий Pseudomonas sp.101 (Rojkova A.M., Galkin A.G., Kulakova L.B., Serov A.E., Savitsky P.A., Fedorchuk V.V., and Tishkov V.I. Bacterial formate dehydrogenase. Increasing the enzyme thermal stability by hydrophobization of alpha-helices. FEES Lett., 1999, v.445, pp.183-188). Процедура очистки фермента включала высаживание балластных белков сульфатом аммония (45% от насыщения), осаждение целевого белка при концентрации сульфата аммония 85% от насыщения и его последующее перерастворение в растворе, содержащем 45%) сульфат аммония в 0,1 М фосфатном буфере, рН 7,0 (буфера А). Полученный супернатант использовали для гидрофобной хроматографии на Phcnyl Sepharose Fast Flow (Pharmacia Biotech) в нисходящем градиенте 45-0% концентрации сульфата аммония в буфере А и обессоливание на колонке с Сефадекс G-25 в том же буфере. Контроль чистоты полученных препаратов осуществлялся с помощью аналитического электрофореза в 12%) полиакриламидном геле в присутствии 0,1% додецильсульфата натрия на приборе для электрофореза MiniProtcan II фирмы «BioRad».
Активность ФДГ определяли спектрофотометрически по накоплению NADH на длине волны 340 им (ε340=6220 М-1 см-1) на спектрофотометре «Schimadzu UV 1800 PC» при 30°С в 0,1 М натрий-фосфатном буфере, рН 7,0. Концентрация формиата натрия и NAD+ в кювете составляла 0,6 М и 0,2 мг/мл соответственно.
Термостабильность фермента измеряли в 0,1 М натрий-фосфатном буфере, рН 7,0. Для каждого эксперимента готовили серию из пластиковых пробирок объемом 0,5 мл по 100 мкл раствора фермента (0,2 мг/мл) в каждой. Пробирки помещали в предварительно прогретый до необходимой температуры водный термостат (52-56°С, точность термостатирования ±0,1°С). В определенные моменты времени отбирали по одной пробирке и переносили в лед на 5 мин, после чего пробирку центрифугировали в течение 3 мин при 12000 об/мин на центрифуге «Eppendorf 5415D». Остаточную активность ФДГ измеряли, как описано выше. Константу скорости термоинактивации kin определяли как тангенс угла наклона прямой из графика зависимости натурального логарифма величины остаточной активности от времени (полулогарифмические координаты ln(A/A0)-t) методом линейной регрессии, используя программу «Origin Pro 8.5».
В качестве параметра, характеризующего термостабильность, использовали константу скорости процесса термоинактивации. Результаты представлены в таблице 2. Как следует из таблицы 2, введение мутаций Phe290Asn, Phe290Asp, Phe290Ser, повышает стабильность формиатдегидрогеназы из сои при температуре 56°С в 10, 42 и 3,6 раза соответственно.
Пример 5.
Определение кинетических и каталитических параметров мутантных форм формиатдегидрогеназы из сои, содержащих аминокислотные замены Phe290Asn, Phe290Asp и Phe290Ser.
Константы Михаэлиса по NAD+ и формиату определяли из зависимостей активности фермента от концентрации (0,4-6 КМ) соответствующего субстрата. Концентрация второго субстрата была насыщающей (>30 КМ). Точную концентрацию исходного раствора NAD+ определяли спектрофотометрически на длине волны 260 нм (ε260=17800 М-1 см-1). Раствор формиата натрия с заданной концентрацией готовили, растворяя нужное количество субстрата к 0,1 М натрий-фосфатном буфере, рH 7,0. Объем раствора доводили в мерной колбе. Значения КМ были рассчитаны из экспериментальных кривых с применением метода нелинейной регрессии, используя программу Origin Pro 8.5.
Для нахождения каталитических констант готовили несколько проб исследуемого фермента с различной концентрацией. Для каждой пробы определяли значения максимальной скорости и концентрацию активных центров. Концентрацию активных центров определяли из зависимостей тушения собственной флуоресценции фермента в комплексе с NAD+ при добавлении азид-иона как описано в (Романова Е.Г., Алексеева А.А., Пометун Е.В., Тишков В.И. Определение концентрации активных центров и каталитической константы рекомбинантной формиатдегидрогеназы из сои Glycine max. Вестник Московского Университета, Сер. 2: Химия, 2010, т.51, №3, с.156-159). Измерения проводили в 0,1 М натрий-фосфатном буфере, рH 7,0 на спектрофлуориметре Cary Eclipse (“Varian” США). Далее строили график зависимости максимальной скорости от концентрации активных центров и значение каталитической константы определяли как тангенс угла наклона прямой методом линейной регрессии, используя программу Origin Pro 8.5.
Результаты для полученных мутантных форм SoyFDH Phe290Asn, SoyFDH Phe290Asp, SoyFDH Phe290Ser и белка дикого типа представлены в таблице 3. Как следует из таблицы 3, замена фенилаланина на аспарагин в 290 положении не влияет на КМ но NAD+ и каталитическую константу.
Как видно из таблицы 3, замена Phe290Asp привела к увеличению каталитической константы на 76% и, как следствие, к увеличению каталитической эффективности с NAD+ в 2 раза.
Замена Phe290Ser привела к увеличению каталитической константы на 41%, а также к уменьшению константы Михаэлиса по NAD+ на 46%. Также из таблицы 3 следует, что для фермента с заменой Phe290Ser происходит увеличение каталитической эффективности с NAD+ в 2,5 раза. Все три замены в 290 положении приводят к увеличению КМ по формиату.
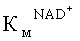
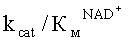
(мкМ*с)-1
| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ ФОРМИАТДЕГИДРОГЕНАЗА (ВАРИАНТЫ) | 2013 |
|
RU2545966C1 |
| МУТАНТНАЯ РАСТИТЕЛЬНАЯ ФОРМИАТДЕГИДРОГЕНАЗА (ВАРИАНТЫ) | 2014 |
|
RU2557299C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pHpFDH_HisA, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ В КЛЕТКАХ Escherichia coli ФОРМИАТДЕГИДРОГЕНАЗЫ МЕТИЛОТРОФНЫХ ДРОЖЖЕЙ Hansenula polymorpha (HpFDH_HisA) И РЕКОМБИНАНТНЫЙ ШТАММ E.сoli BL21(DE3)/pHpFDH_HisA ВКПМ В-14559 - ПРОДУЦЕНТ HpFDH_HisA | 2023 |
|
RU2829762C1 |
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ФОРМИАТДЕГИДРОГЕНАЗА | 2006 |
|
RU2312897C1 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ | 2009 |
|
RU2412998C1 |
| МУТАНТНЫЕ ОКСИДАЗЫ D-АМИНОКИСЛОТ | 2007 |
|
RU2362806C2 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ (ВАРИАНТЫ) | 2012 |
|
RU2507262C1 |
| ГЕНЕТИЧЕСКИ СКОНСТРУИРОВАННАЯ ЗАВИСИМАЯ ОТ ПИРРОЛОХИНОЛИНХИНОНА ГЛЮКОЗОДЕГИДРОГЕНАЗА, СОДЕРЖАЩАЯ ИНСЕРЦИЮ АМИНОКИСЛОТЫ | 2005 |
|
RU2336306C1 |
| МУТАНТНАЯ ФОРМА ПЕРОКСИДАЗЫ ХРЕНА | 2012 |
|
RU2474613C1 |
| Способ получения 2,4-дигидроксимасляной кислоты | 2011 |
|
RU2626531C2 |
Изобретение относится к области генной и белковой инженерии и представляет собой три новые мутантные формы растительной формиатдегидрогеназы, обладающие улучшенными свойствами по сравнению с белком дикого типа. В первом из предложенных мутантных белков природный остаток фенилаланина в положении, соответствующем положению 290 в последовательности формиатдегидрогеназы из сои Glycine max, заменен на остаток аспарагина. Во второй мутантной форме фермента этот же природный остаток фенилаланина 290 заменен на остаток аспарагиновой кислоты, а в третьей - на остаток серина. Изобретение позволяет получить улучшенные варианты формиатдегидрогеназы с повышенной термостабильностью. 3 н.п. ф-лы, 3 табл., 5 пр.
1. Мутантная формиатдегидрогеназа с улучшенными свойствами, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности формиатдегидрогеназы из Glycine max дикого типа, в которой остаток фенилаланина в положении 290 заменен остатком аспарагина.
2. Мутантная формиатдегидрогеназа с улучшенными свойствами, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности формиатдегидрогеназы из Glycine max дикого типа, в которой остаток фенилаланина в положении 290 заменен остатком аспарагиновой кислоты.
3. Мутантная формиатдегидрогеназа с улучшенными свойствами, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности формиатдегидрогеназы из Glycine max дикого типа, в которой остаток фенилаланина в положении 290 заменен остатком серина.
| А.А.АЛЕКСЕЕВА Рекомбинантная формиатдегидрогеназа из сои Glycine max: белковая инженерия и структурные исследования | |||
| Диссертация на соискание ученой степени кандидата химических наук, Москва, 2011, стр | |||
| Кровля из глиняных обожженных плит с арматурой из проволочной сетки | 1921 |
|
SU120A1 |
| Способ получения алкилянтарных кислот | 1951 |
|
SU94321A1 |

