ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПРЕДОСТАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
Официальная копия данного перечня последовательностей предоставлена в электронном виде с помощью EFS-Web как перечень последовательностей в формате ASCII с названием файла “7768WOPCT_ST25.txt”, созданном 18 апреля 2018 года и имеющем размер 51 килобайт, и подана одновременно с описанием. Перечень последовательностей, содержащийся в данном документе в формате ASCII, является частью описания и включен в данный документ посредством ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Раскрытые в данном документе варианты осуществления относятся к области молекулярной биологии растений, в частности к конструкциям ДНК для повышения урожайности растения. Раскрытые в данном документе варианты осуществления более конкретно относятся к растениям, генам, клеткам, семенам, частям растений, ДНК, переработанному растительному продукту и конструкциям маиса, относящимся к объекту DP-202216-6 маиса, а также к способам и композициям на их основе.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Кукуруза является агрономически важной сельскохозяйственной культурой и служит источником пищи и кормов для животных, человека и путей применения в промышленности. Повышение урожайности зерна может быть достигнуто в растениях маиса множеством способов, включая экспрессию трансгена для повышения урожайности зерна в дополнение к улучшенной селекции. На производительность трансгена в растении, в том числе агрономические параметры, могут влиять различные факторы, такие как применение элементов экспрессии, в том числе промоторных/регуляторных элементов, геномное расположение последовательности вставки, количество копий вставленного трансгена и генетические факторы (идиоплазма) и факторы окружающей среды, такие как почва, температура, свет и влажность. Идентификация конструкций, испытание ортологов и объектов трансформации, которые приводят к повышению урожайности зерна растения маиса на коммерчески значимом уровне в поле, являются результатом существенных и значительных конструкторских усилий по усовершенствованию продукта. Соответственно, было бы желательно иметь растения маиса, у которых проявляется повышенная урожайность зерна.
КРАТКОЕ ОПИСАНИЕ
Семя кукурузы включает объект DP-202216-6, где указанное семя содержит молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12, 13, 14 и их комбинации, где репрезентативный образец семени объекта DP-202216-6 кукурузы депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653. В некоторых вариантах осуществления в данном документе описано растение кукурузы или его часть, выращенные из семян PTA-124653.
Растение маиса, стабильно трансформированное рекомбинантной полинуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, которая на по меньшей мере 90%, 93% 95%, 97%, 98% или 99% идентична SEQ ID NO: 1, где растение маиса демонстрирует повышенную урожайность зерна по сравнению с контрольным растением маиса, не содержащим рекомбинантный полинуклеотид. В некоторых вариантах осуществления рекомбинантный полинуклеотид функционально связан со слабым гетерологичным конститутивным регуляторным элементом. В некоторых вариантах осуществления урожайность зерна составляет по меньшей мере приблизительно три бушеля/акр по сравнению с контрольным растением маиса, где растение маиса и контрольное растение маиса выращивают в поле при нормальных условиях выращивания сельскохозяйственных культур. В некоторых вариантах осуществления урожайность зерна в поле находится в диапазоне от приблизительно 2 до приблизительно 8 бушелей/акр по сравнению с контрольной популяцией растений маиса при выращивании с плотностью популяции от приблизительно 20000 до приблизительно 50000 растений на акр. В некоторых вариантах осуществления слабый гетерологичный конститутивный регуляторный элемент представляет собой промотор GOS2 маиса. В некоторых вариантах осуществления аминокислотная последовательность на по меньшей мере 95% идентична SEQ ID NO: 1, и растение маиса содержит полинуклеотид, кодирующий полипептид, который обеспечивает переносимость гербицидов, и полинуклеотид, который кодирует полипептид или последовательность РНК, которые обеспечивают устойчивость к одному или нескольким насекомым-вредителям. Семя маиса, полученное из растения маиса, описанного в данном документе, демонстрируют характеристики улучшения урожайности. В одном варианте осуществления регуляторный элемент предусматривает гетерологичный интронный элемент.
Рекомбинантная полинуклеотидная конструкция предусматривает полинуклеотид, кодирующий полипептид, содержащий аминокислотную последовательность, которая на по меньшей мере 90%, 93% 95%, 97%, 98% или 99% идентична SEQ ID NO: 1, где полинуклеотид функционально связан с гетерологичным регуляторным элементом. В некоторых вариантах осуществления аминокислотная последовательность на основе SEQ ID NO: 1 может иметь одну или несколько вариаций, в том числе вставку, делецию или замену.
Способ повышения урожайности зерна растения маиса, при этом способ включает экспрессию полинуклеотидной последовательности, кодирующей полипептид, который на по меньшей мере 90%, 93% 95%, 97%, 98% или 99% идентичен SEQ ID NO: 1, где полинуклеотид функционально связан с гетерологичной регуляторной последовательностью; и выращивание растения маиса в поле с повышением урожайности зерна по сравнению с контрольным растением маиса, не содержащим полинуклеотид, функционально связанный с гетерологичной регуляторной последовательностью.
Способ получения семени, при этом способ включает следующее:
(а) скрещивание первого растения со вторым растением, где по меньшей мере одно из первого растения и второго растения содержит конструкцию рекомбинантной ДНК, где конструкция рекомбинантной ДНК содержит полинуклеотид, функционально связанный с по меньшей мере одним регуляторным элементом, где полинуклеотид кодирует белок MADS, имеющий аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности, с использованием способа выравнивания Clustal V или Clustal W, по сравнению с SEQ ID NO: 1; и
(b) отбор семени, полученного в результате скрещивания на стадии (а), где семя содержит конструкцию рекомбинантной ДНК.
Растение, выращенное из семени, полученного описанным в данном документе способом, при этом растение демонстрирует повышенную урожайность по сравнению с контрольным растением, не содержащим конструкцию рекомбинантной ДНК.
В некоторых вариантах осуществления способ отбора растения, демонстрирующего повышенную урожайность, включает:
(a) получение растения, где растение содержит в своем геноме конструкцию рекомбинантной ДНК, содержащую полинуклеотид, функционально связанный с по меньшей мере одним регуляторным элементом, где указанный полинуклеотид кодирует белок MADS, имеющий аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности, с использованием способа выравнивания Clustal V или Clustal W, по сравнению с SEQ ID NO: 1;
(b) выращивание растения в поле в условиях, при которых экспрессируется полинуклеотид; и
(c) отбор растения из части, которая демонстрирует повышенную урожайность по сравнению с контрольным растением, не содержащим конструкцию рекомбинантной ДНК.
В некоторых вариантах осуществления растение выбрано из группы, состоящей из маиса, сои, подсолнечника, сорго, канолы, пшеницы, люцерны, хлопчатника, риса, ячменя, проса, сахарного тростника и проса прутьевидного. В одном из вариантов осуществления аминокислотная последовательность белка MADS содержит последовательность, которая на по меньшей мере 99% идентична SEQ ID NO: 1.
Рекомбинантный полинуклеотид включает полинуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая на по меньшей мере 95% идентична SEQ ID NO: 1, где рекомбинантный полинуклеотид содержит гетерологичный регуляторный элемент. В некоторых вариантах осуществления растение или семя включает в себя рекомбинантный полинуклеотид, описанный в данном документе.
Растение маиса, которое демонстрирует повышенную экспрессию эндогенного полинуклеотида, кодирующего полипептид, содержащий последовательность, которая на по меньшей мере 95% идентична SEQ ID NO: 1, где повышенная экспрессия обусловлена гетерологичным регуляторным элементом. В некоторых вариантах осуществления гетерологичный регуляторный элемент представляет собой энхансерный элемент, полученный из растения. В некоторых вариантах осуществления гетерологичный регуляторный элемент представляет собой слабый конститутивный промоторный элемент. В некоторых вариантах осуществления растение маиса является инбредным или гибридным растением.
В некоторых вариантах осуществления растение маиса включает в себя второй полипептид, который обеспечивает переносимость гербицидов, и третий полипептид, который обеспечивает устойчивость к насекомым.
Конструкция рекомбинантной ДНК, содержащая кассету экспрессии, где кассета экспрессии включает в функциональной связи промотор gos2 маиса; интрон гена убиквитина 1 маиса (ubiZM1); ген MADS-бокса маиса, кодирующий белок ZMM28 маиса; терминатор pinII; промотор гена убиквитина 1 маиса (ubiZM1); 5’UTR гена убиквитина 1 маиса (ubiZM1); интрон гена убиквитина 1 маиса (ubiZM1); ген mo-pat и терминатор pinII. В некоторых вариантах осуществления растение включает в себя описанную в данном документе конструкцию ДНК, и растение представляет собой растение кукурузы. В некоторых вариантах осуществления растение включает в себя последовательность, которая на по меньшей мере 95% идентична полинуклеотидной последовательности, изложенной под SEQ ID NO: 6.
Растение, семя, клетка кукурузы или их часть включает в себя объект DP-202216-6, где объект содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 7 и SEQ ID NO: 8. В некоторых вариантах осуществления объект содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 9 и SEQ ID NO: 10. В некоторых вариантах осуществления объект содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 11 и SEQ ID NO: 12. В некоторых вариантах осуществления объект содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 13 или SEQ ID NO: 14. В некоторых вариантах осуществления часть растения выбрана из группы, состоящей из околоплодника, пыльцы, семяпочки, цветка, зерна, побега, корня, стебля, шелка, метелки, початка и ткани листа.
Растение, семя, клетка кукурузы или их часть включает в себя объект DP-202216-6, где репрезентативный образец семени указанного объекта кукурузы депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653. В некоторых вариантах осуществления часть растения выбрана из группы, состоящей из околоплодника, пыльцы, семяпочки, цветка, зерна, побега, корня, стебля, шелка, метелки, початка и ткани листа.
Выделенная молекула нуклеиновой кислоты включает в себя нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12, и в некоторых вариантах осуществления ампликон включает последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12 и их полноразмерных комплементарных последовательностей. В некоторых вариантах осуществления ампликон имеет размер менее чем приблизительно 500 п.н., 1 т.н., 1,5 т.н., 2,0 т.н., 3,0 т.н., 5,0 т.н. и 10 т.н.
Биологический образец, полученный из растения, ткани или семени объекта DP-202216-6 кукурузы, где указанный образец содержит нуклеотидную последовательность, которая представляет собой последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11 и 12, или является комплементарной ей, где указанная нуклеотидная последовательность обнаруживается в указанном образце с применением способа амплификации нуклеиновой кислоты или гибридизации нуклеиновой кислоты, где репрезентативный образец указанного семени объекта DP-202216-6 кукурузы депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653. В некоторых вариантах осуществления биологический образец предусматривает растение, ткань или части семени, околоплодника семени трансгенного объекта DP-202216-6 кукурузы. В некоторых вариантах осуществления биологический образец представляет собой образец ДНК, экстрагированный из трансгенного объекта DP-202216-6 растения кукурузы, и где указанный образец ДНК содержит одну или несколько нуклеотидных последовательностей, выбранных из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12 и их комплементарных последовательностей. В некоторых вариантах осуществления биологический образец выбран из группы, состоящей из кукурузной муки, кукурузной крупы, кукурузного сиропа, кукурузного масла, кукурузного крахмала и злаковых продуктов, изготовленных так, что они полностью или частично содержат сопутствующие продукты из кукурузы, где указанный биологический образец содержит обнаруживаемое количество указанной нуклеотидной последовательности.
Экстракт, полученный из растения, ткани или семени объекта DP-202216-6 кукурузы и содержащий нуклеотидную последовательность, которая представляет собой последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11 и 12, или является комплементарной ей, где репрезентативный образец указанного семени объекта DP-202216-6 кукурузы депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653. В некоторых вариантах осуществления указанная нуклеотидная последовательность обнаруживается в указанном экстракте с применением способа амплификации нуклеиновой кислоты или гибридизации нуклеиновой кислоты. В некоторых вариантах осуществления композиция выбрана из группы, состоящей из кукурузной муки, кукурузной крупы, кукурузного сиропа, кукурузного масла, кукурузного крахмала и злаковых продуктов, изготовленных так, что они полностью или частично содержат сопутствующие продукты из кукурузы, где указанная композиция содержит обнаруживаемое количество указанной нуклеотидной последовательности.
Способ получения гибридных семян кукурузы, при этом способ включает:
a) скрещивание половым путем первой инбредной линии кукурузы, содержащей нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11 и 12, и второй инбредной линии, имеющей отличный генотип;
b) выращивание потомства от указанного скрещивания и
c) сбор гибридного семени, полученного таким образом.
В некоторых вариантах осуществления первая инбредная линия кукурузы является родителем женского пола или первая инбредная линия кукурузы является родителем мужского пола.
Способ получения растения кукурузы, которое демонстрирует повышенную урожайность зерна в поле, при этом способ включает:
a) скрещивание половым путем первого родительского растения кукурузы со вторым родительским растением кукурузы, где указанное первое или второе родительское растение кукурузы содержит ДНК объекта DP-202216-6, в результате чего получают множество растений-потомков первого поколения;
b) самоопыление растения-потомка первого поколения с получением таким образом множества растений-потомков второго поколения; и
c) отбор из растений-потомков второго поколения, которые содержат объект DP-202216-6, растения, которое демонстрирует повышенную урожайность зерна в поле по сравнению с контрольным растением кукурузы, не содержащим объект DP-202216-6.
В некоторых вариантах осуществления объект DP-202216-6 содержит конструкцию рекомбинантной ДНК, где объект DP-202216-6 кодирует полипептид, который на по меньшей мере 99% идентичен SEQ ID NO: 1.
Способ получения гибридных семян кукурузы, включающий:
a) скрещивание половым путем первой инбредной линии кукурузы, содержащей описанную в данном документе конструкцию ДНК, со второй инбредной линией, не содержащей конструкцию ДНК; и
b) сбор гибридного семени, полученного таким образом.
В некоторых вариантах осуществления стадия возвратного скрещивания включает возвратное скрещивание растения-потомка второго поколения, которое содержит объект DP-202216-6 кукурузы, с родительским растением, в котором отсутствует объект DP-202216-6 кукурузы, в результате чего получают растение-потомка, полученное путем возвратного скрещивания, которое демонстрирует повышенную урожайность зерна по сравнению с контрольным растением кукурузы, не содержащим объект DP-202216-6.
Способ получения растения кукурузы, которое демонстрирует повышенную урожайность зерна, при этом указанный способ включает:
a) скрещивание половым путем первого родительского растения кукурузы со вторым родительским растением кукурузы, где указанное первое или второе родительское растение кукурузы является растением объекта DP-202216-6 кукурузы, в результате чего получают множество растений-потомков первого поколения;
b) отбор растения-потомка первого поколения, которое демонстрирует повышенную урожайность зерна;
c) возвратное скрещивание растения-потомка первого поколения из стадии (b) с родительским растением, в котором отсутствует объект DP-202216-6 кукурузы, в результате чего получают множество растений-потомков, полученных путем возвратного скрещивания; и
d) отбор из растений-потомков, полученных путем возвратного скрещивания, растения, растения, которое демонстрирует повышенную урожайность зерна;
где выбранное растение-потомок, полученное путем возвратного скрещивания, из стадии (d) содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11 и 12.
В некоторых вариантах осуществления растения первого родительского растения кукурузы являются родителями женского пола или родителями мужского пола. Гибридное семя получают описанными в данном документе способами.
Способ определения зиготности ДНК растения кукурузы, содержащего объект DP-202216-6 кукурузы, в биологическом образце, включающий:
a) приведение указанного образца в контакт с первой парой молекул ДНК и второй отличающейся парой молекул таким образом, что: (i) при использовании в реакции амплификации нуклеиновой кислоты, предусматривающей ДНК объекта DP-202216-6 кукурузы, образуется первый ампликон, который является диагностическим признаком объекта DP-202216-6 кукурузы, и (ii) при использовании в реакции амплификации нуклеиновой кислоты, предусматривающей геномную ДНК кукурузы, отличную от ДНК DP-202216-6, образуется второй ампликон, который является диагностическим признаком геномной ДНК кукурузы, отличной от ДНК DP-202216-6;
b) проведение реакции амплификации нуклеиновой кислоты и
c) обнаружение первого и второго ампликонов, полученных таким образом, где обнаружение присутствия первого и второго ампликонов указывает на то, что указанный образец является гетерозиготным в отношении ДНК объекта DP-202216-6 кукурузы, где обнаружение первого ампликона указывает на то, что указанный образец является гомозиготным в отношении ДНК объекта DP-202216-6 кукурузы.
В некоторых вариантах осуществления первая пара молекул ДНК включает пары праймеров, которые амплифицируют фрагмент ДНК, который содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12 и их обратных комплементарных последовательностей. В некоторых вариантах осуществления первая и вторая пара молекул ДНК содержат обнаруживаемую метку. В некоторых вариантах осуществления обнаруживаемая метка представляет собой флуоресцентную метку. В некоторых вариантах осуществления обнаруживаемая метка ковалентно связана с одной или несколькими молекулами праймера. В некоторых вариантах осуществления пара праймеров содержит SEQ ID NO: 15 и 16.
Способ обнаружения присутствия молекулы нуклеиновой кислоты, которая является уникальной для объекта DP-202216-6 или дает возможность отличить его, в образце, при этом способ включает:
a) приведение образца в контакт с парой праймеров или зондом, который при использовании в реакции амплификации нуклеиновой кислоты с геномной ДНК из объекта DP-202216-6 образует молекулу нуклеиновой кислоты, которая является диагностическим признаком объекта DP-202216-6;
b) проведение реакции амплификации нуклеиновой кислоты с получением таким образом молекулы нуклеиновой кислоты, которая является диагностическим признаком объекта DP-202216-6; и
c) обнаружение молекулы нуклеиновой кислоты, которая является диагностическим признаком объекта DP-202216-6.
В некоторых вариантах осуществления молекула нуклеиновой кислоты, которая является диагностическим признаком объекта DP-202216-6, представляет собой ампликон, образуемый при цепной реакции амплификации нуклеиновой кислоты. В некоторых вариантах осуществления зонд содержит обнаруживаемую метку. В некоторых вариантах осуществления обнаруживаемая метка представляет собой флуоресцентную метку. В некоторых вариантах осуществления обнаруживаемая метка ковалентно связана с зондом.
Множество полинуклеотидных праймеров, содержащих один или несколько полинуклеотидов длиной по меньшей мере 10 смежных оснований, которые нацелены на ДНК-матрицу объекта DP-202216-6 в образце, чтобы получить ампликон, являющийся диагностическим признаком объекта DP-202216-6, в результате применения способа амплификации посредством полимеразной цепной реакции. В некоторых вариантах осуществления полинуклеотидные праймеры характеризуются тем, что:
a) первый полинуклеотидный праймер содержит по меньшей мере 10 смежных нуклеотидов из нуклеотидной последовательности, выбранной из группы, состоящей из нуклеотидов 1-425 в SEQ ID NO: 31, нуклеотидов 1-417 в SEQ ID NO: 32 и их комплементарных последовательностей; и
b) второй полинуклеотидный праймер содержит по меньшей мере 10 смежных нуклеотидов из нуклеотидов SEQ ID NO: 6 или ее комплементарных последовательностей.
В некоторых вариантах осуществления полинуклеотидные праймеры характеризуются тем, что:
a) первый полинуклеотидный праймер содержит полинуклеотидную последовательность, содержащую SEQ ID NO: 15 и ее комплементарные последовательности; и
b) второй полинуклеотидный праймер содержит полинуклеотидную последовательность, содержащую SEQ ID NO: 16 и ее комплементарные последовательности.
В некоторых вариантах осуществления указанный первый праймер и указанный второй праймер составляют по меньшей мере 18 нуклеотидов.
Способ обнаружения присутствия ДНК, соответствующей объекту DP-202216-6, в образце, при этом способ включает:
a) приведение образца, содержащего ДНК маиса, в контакт с полинуклеотидным зондом, который гибридизируется в жестких условиях гибридизации с ДНК из объекта DP-202216-6 маиса и не гибридизируется в указанных жестких условиях гибридизации с ДНК растения маиса, не относящегося DP-202216-6;
b) подвергание образца и зонда жестким условиям гибридизации и
c) обнаружение гибридизации зонда с ДНК; где обнаружение гибридизации указывает на присутствие объекта DP-202216-6.
Набор для обнаружения нуклеиновой кислоты, которая является уникальной для объекта DP-202216-6, включает по меньшей мере одну молекулу нуклеиновой кислоты достаточной длины из смежных полинуклеотидов для функционирования в качестве праймера или зонда в способе обнаружения нуклеиновой кислоты, и которая после амплификации или гибридизации с последовательностью целевой нуклеиновой кислоты в образце с последующим обнаружением ампликона или продукта гибридизации с целевой последовательностью является диагностическим признаком присутствия последовательности нуклеиновой кислоты, уникальной для объекта DP-202216-6, в образце. В некоторых вариантах осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность из SEQ ID NO: 7 или 8.
В некоторых вариантах осуществления молекула нуклеиновой кислоты представляет собой пару праймеров, содержащую пару полинуклеотидных последовательностей, при этом каждая из них содержит по меньшей мере 10 смежных оснований, где пара праймеров амплифицирует последовательность соединения объекта DP-202216-6, при этом соединение предусматривает полинуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12, 13, 14, 31 и 32 и их комплементарных последовательностей.
Товарный продукт, полученный из трансгенного растения кукурузы, содержащего объект DP-202216-6 и содержащего молекулу рекомбинантной ДНК, содержащую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 8, 9, 10, 11, 12, 13, 14, 31 и 32 и их полных комплементарных последовательностей, где обнаружение указанной молекулы рекомбинантной ДНК в образце, полученном из указанного товарного продукта, является определяющим в том, что указанный товарный продукт был получен из указанного трансгенного растения кукурузы, содержащего объект DP-202216-6. В некоторых вариантах осуществления товарный продукт выбран из группы, состоящей из цельных или переработанных семян, корма для животных, масла, крупы, муки, хлопьев, отрубей, биомассы и топливных продуктов.
Способ получения товарного продукта, при этом способ включает: (a) получение растения кукурузы или его части, содержащих трансгенный объект DP-202216-6 кукурузы; и (b) изготовление товарного продукта кукурузы из растения кукурузы или его части.
Антитело, полученное для нацеливания на полипептид, вырабатываемый объектом DP-202216-6, где полипептид вырабатывается с помощью гетерологичного регуляторного элемента и содержит аминокислотную последовательность, которая на по меньшей мере 99% идентична SEQ ID NO: 1. В некоторых вариантах осуществления антитело представляет собой моноклональное антитело и содержит обнаруживаемую метку.
Способ повышения урожайности зерна популяции растений маиса в поле, при этом способ включает выращивание популяции растений маиса, содержащих объект DP-202216-6, в поле и таким образом обеспечивает повышение урожайности зерна популяции растений маиса по сравнению с контрольным растением, не содержащим объект DP-202216-6. В некоторых вариантах осуществления популяцию растений маиса выращивают в условиях абиотического стресса. В некоторых вариантах осуществления абиотический стресс связан с низким содержанием азота. В некоторых вариантах осуществления при выращивании в условиях низкого содержания азота популяция растений маиса, содержащих объект DP-202216-6, демонстрирует стабильность урожайности по сравнению с контрольной популяцией растений, выращенных в условиях низкого содержания азота. В некоторых вариантах осуществления низкое содержание азота представляет собой уменьшение количества азота на от приблизительно 25% до приблизительно 75% от обычно применяемого для выращивания гибридных растений кукурузы в поле. В некоторых вариантах осуществления уменьшение количества азота, вносимого в поле, составляет от приблизительно 5% до приблизительно 10%, 20%, 30%, 40%, 50%, 60% или 70% по сравнению с обычным внесением азота.
В соответствии с некоторыми вариантами осуществления предусмотрены композиции и способы для идентификации нового растения кукурузы, обозначенного DP-202216-6 (номер депонирования в ATCC PTA-124653). Способы основаны на праймерах или зондах, которые специфически распознают 5’- и/или 3’-фланкирующую последовательность DP-202216-6. Предусмотрены молекулы ДНК, содержащие последовательности праймеров, которые при использовании в реакции ПЦР будут образовывать ампликоны, уникальные для трансгенного объекта DP-202216-6. В одном варианте осуществления рассматриваются растение кукурузы и семя, содержащие эти молекулы. Кроме того, предусматриваются наборы, использующие эти последовательности праймеров для идентификации объекта DP-202216-6.
Дополнительные варианты осуществления относятся к конкретной фланкирующей последовательности DP-202216-6, которая описана в данном документе, которая может быть использована для разработки специфических способов идентификации DP-202216-6 в биологических образцах. Более конкретно, настоящее изобретение относится к 5’- и/или 3’-фланкирующим областям DP-202216-6, которые можно использовать для разработки специфических праймеров и зондов. Дополнительные варианты осуществления относятся к способам идентификации присутствия DP-202216-6 в биологических образцах на основе применения таких специфических праймеров или зондов.
Согласно другому варианту осуществления предусмотрены способы обнаружения присутствия ДНК, соответствующей объекту DP-202216-6 кукурузы, в образце. Такие способы включают: (a) приведение образца, содержащего ДНК, в контакт с набором ДНК-праймеров, который при использовании в реакции амплификации нуклеиновой кислоты с геномной ДНК, экстрагированной из объекта DP-202216-6 кукурузы, образует ампликон, который является диагностическим признаком объекта DP-202216-6 кукурузы, соответственно; (b) проведение реакции амплификации нуклеиновой кислоты с получением таким образом ампликона и (c) обнаружение ампликона. В некоторых аспектах набор праймеров содержит SEQ ID NO: 15 и/или 16, полинуклеотид, который обнаруживает по меньшей мере одну последовательность соединения, выбранную из группы, состоящей из SEQ ID NO: 7-12 и их комбинации.
Согласно другому варианту осуществления способы обнаружения присутствия молекулы ДНК, соответствующей объекту DP-202216-6, в образце, при этом такие способы включают: (a) приведение образца, содержащего ДНК, экстрагированную из растения кукурузы, в контакт с молекулой ДНК-зонда, которая гибридизируется в жестких условиях гибридизации с ДНК, экстрагированной из объекта DP-202216-6 кукурузы, и не гибридизируется в жестких условиях гибридизации с ДНК контрольного растения кукурузы; (b) подвергание образца и зонда жестким условиям гибридизации и (c) обнаружение продукта гибридизации зонда с ДНК. Более конкретно, способ обнаружения присутствия молекулы ДНК, соответствующей объекту DP-202216-6, в образце, при этом такие способы предусматривают (а) приведение образца, содержащего ДНК, экстрагированную из растения кукурузы, в контакт с молекулой ДНК-зонда, которая состоит из последовательностей, уникальных для данного объекта, например последовательностей соединения, где указанная молекула ДНК-зонда гибридизируется в жестких условиях гибридизации с ДНК, экстрагированной из объекта DP-202216-6 кукурузы, и не гибридизируется в жестких условиях гибридизации с ДНК контрольного растения кукурузы; (b) подвергание образца и зонда жестким условиям гибридизации и (c) обнаружение продукта гибридизации зонда с ДНК.
Кроме того, предусматриваются набор и способы для идентификации объекта DP-202216-6 в биологическом образце, которые обнаруживают специфическую область DP-202216-6.
Предусматриваются молекулы ДНК, которые содержат по меньшей мере одну последовательность соединения DP-202216-6; где последовательность соединения охватывает соединение между гетерологичной ДНК, вставленной в геном, и ДНК из клетки кукурузы, фланкирующей сайт вставки, т.е. фланкирующей ДНК, и является диагностическим признаком объекта DP-202216-6.
В соответствии с другим вариантом осуществления способы получения растения кукурузы включают стадии: (a) скрещивания половым путем первой родительской линии кукурузы, содержащей раскрытые в данном документе кассеты экспрессии, которые повышают урожайность, и второй родительской линии кукурузы, в которой отсутствуют такие конструкции, в результате чего получают множество растений-потомков; и (b) отбора растения-потомка, которое проявляет повышение урожайности. Такие способы могут необязательно включать дополнительную стадию возвратного скрещивания растения-потомка со второй родительской линией кукурузы для получения гомозиготного растения кукурузы, которое демонстрирует повышение урожайности.
Другой вариант осуществления дополнительно относится к набору для обнаружения ДНК для идентификации объекта DP-202216-6 маиса в биологических образцах. Набор включает первый праймер или зонд, который специфически амплифицирует или обнаруживает 5'- или 3'-фланкирующую область DP-202216-6, и второй праймер или зонд, который специфически амплифицирует или обнаруживает последовательность во вставленной ДНК DP-202216-6, соответственно, или во фланкирующей ДНК, для применения в протоколе идентификации при помощи ПЦР. Дополнительный вариант осуществления относится к набору для идентификации объекта DP-202216-6 в биологических образцах, при этом набор содержит специфический зонд, имеющий последовательность, которая соответствует или является комплементарной последовательности, обладающей от 80% до 100% идентичности последовательности с конкретной областью объекта DP-202216-6. Последовательность зонда соответствует конкретной области, предусматривающей часть 5’- или 3’-фланкирующей области объекта DP-202216-6. В некоторых вариантах осуществления первый или второй праймер или соответствующий зонд содержит SEQ ID NO: 15, 16, 17, 18, 19, 20 и их обратные комплементарные последовательности.
Способы и наборы, охватываемые вариантами осуществления, раскрытыми в данном документе, можно использовать для различных целей, таких как без ограничения следующие: для идентификации объекта DP-202216-6 в растениях, растительном материале или в таких продуктах, как без ограничения пищевые продукты или кормовые продукты (свежие или переработанные), содержащие растительный материал или полученные из него; дополнительно или в качестве альтернативы, способы и наборы можно использовать для идентификации трансгенного растительного материала с целью разделения трансгенного и нетрансгенного материала; дополнительно или в качестве альтернативы, способы и наборы можно использовать для определения качества растительного материала, содержащего объект DP-202216-6 маиса. Наборы также могут содержать реагенты и материалы, необходимые для выполнения способа обнаружения.
Дополнительный вариант осуществления относится к растению маиса DP-202216-6 или его частям, в том числе без ограничения пыльце, семяпочкам, околоплоднику, вегетативным клеткам, ядрам клеток пыльцы и ядрам оотидов растения DP-202216-6 кукурузы и потомству, полученному от него. В другом варианте осуществления включены специфические ампликоны, полученные из растения маиса и семени DP-202216-6.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг. 1. изображена схематическая диаграмма области T-ДНК, которая интегрирована в геном растения маиса для получения объекта DP-202216-6. Размер Т-ДНК составляет 7470 п.н.
На фиг. 2A показана последовательность белка SEQ ID NO: 1, аминокислотная последовательность, которая включает MADS-бокс, промежуточный (сплошное подчеркивание), K-бокс (пунктирная линия) и С-концевой домены.
На фиг. 2B показан филогенетический анализ ZMM28 (GRMZM2G147716_P01) с другими членами клады AP1-FUL из репрезентативных видов растений. Клада, содержащая ZMM28, выделена областью с точками.
На фиг. 2C показана относительная экспрессия гена zmm28 в ткани листа дикого типа (WT) с фоновым генотипом (контрольным) для различных стадий роста.
На фиг. 2D показана относительная экспрессия гена zmm28 в ткани листа объекта DP202216 (незаштрихованные столбики) и дикого типа (WT) с фоновым генотипом (заштрихованные столбики) для различных стадий роста. Планки погрешностей представляют стандартную ошибку. Общая (нативная и трансгенная) экспрессия zmm28 значительно выше в трансгенных растениях, чем в контроле на всех стадиях роста (р<0,05).
На фиг. 2E показана относительная экспрессия гена zmm28 в ткани листа объекта 18 в ходе различных вегетативных (V) и репродуктивных (R) стадий.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящее изобретение можно лучше понять исходя из следующего подробного описания, а также прилагаемых графических материалов и перечня последовательностей, которые образуют часть данной заявки.
Описания последовательностей обобщают прилагаемый перечень последовательностей, который включен в данный документ посредством ссылки. Перечень последовательностей содержит однобуквенные коды для обозначений нуклеотидных последовательностей и однобуквенные и трехбуквенные коды для аминокислот, которые определены в соответствии со стандартами IUPAC-IUB, описанными в Nucleic Acids Research 13:3021-3030 (1985) и в Biochemical Journal 219(2):345-373 (1984).
Таблица 1. Описание перечня последовательностей
ПОДРОБНОЕ ОПИСАНИЕ
Используемая в данном документе форма единственного числа включает ссылки на множественное число, если в контексте явно не указано иное. Так, например, ссылка на "клетку" включает множество таких клеток, и ссылка на "белок" включает ссылку на один или несколько белков или их эквивалентов и т.д. Все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается средним специалистом в данной области, к которой относится настоящее изобретение, если явно не указано иное.
Композиции по настоящему изобретению включают репрезентативный образец семян, который был депонирован как патентный депонированный материал № PTA-124653, и растения, клетки растений и полученное из них семя. Заявитель(-и) депонировал(-и) по меньшей мере 2500 семян объекта DP-202216-6 маиса (патентный депонированный материал № PTA-124653) в Американскую коллекцию типовых культур (ATCC), Манассас, штат Вирджиния 20110-2209, США, 12 января 2018 года. Эти депонированные материалы будут поддерживаться согласно условиям Будапештского договора о международном признании депонированого материала микроорганизмов для целей патентной процедуры. Семена, депонированные в АТСС 12 января 2018 года, были взяты из депонированного материала репрезентативных образцов, поддерживаемого в Pioneer Hi-Bred International, Inc., 7250 NW 62nd Avenue, Джонстон, штат Айова 50131-1000, США. Доступ к этому депонированному материалу ATCC будет предоставлен во время нахождения заявки на рассмотрении комиссара по патентам и товарным знакам, и лиц, определенных комиссаром, как обладающие правом на это по первому требованию в соответствии с применимыми законами и правилами. После выдачи патента этот депонированный материал семян объекта DP-202216-6 маиса предназначен для удовлетворения всех необходимых требований 37 C.F.R. §§1.801-1.809, и будет храниться в депозитарии ATCC в течение 30 лет, или 5 лет после последнего запроса, или в течение срока действия патента, в зависимости от того, какой срок дольше, и будет заменен, если он станет нежизнеспособным в течение этого периода. Неправомочное размножение семян запрещено. Семена могут регулироваться одним или несколькими применимыми национальными, государственными или другими местными инструкциями и постановлениями, налагаемыми одним или несколькими компетентными государственными органами.
Используемый в данном документе термин “кукуруза” означает Zea mays или маис и включает все сорта растений, которые могут быть выведены из кукурузы, в том числе виды дикого маиса.
Используемое в данном документе выражение “устойчивый к воздействию насекомых” и “воздействие на насекомых-вредителей” относится к осуществлению изменений питания, роста и/или поведения насекомого на любой стадии развития, в том числе без ограничения к: уничтожению насекомого; задержке роста; снижению способности к репродукции; подавлению поедания и т.п.
Используемые в данном документе термины “пестицидная активность” и “инсектицидная активность” используются как синонимы и относятся к активности организма или вещества (такого как, например, белок), которую можно измерить по многочисленным параметрам, в том числе без ограничения по смертности вредителя, потере веса вредителем, привлечению вредителя, отпугиванию вредителя и другим изменениям поведения и физических характеристик вредителя после поедания и/или воздействия организма или вещества в течение соответствующего периода времени. Например, “пестицидные белки” представляют собой белки, которые проявляют пестицидную активность сами по себе или в комбинации с другими белками.
Используемая в данном документе “вставленная ДНК” относится к гетерологичной ДНК в кассетах экспрессии, используемых для трансформации растительного материала, в то время как “фланкирующая ДНК” может происходить либо из геномной ДНК, естественно присутствующей в организме, таком как растение, либо из чужеродной (гетерологичной) ДНК, введенной посредством способа трансформации, которая является посторонней по отношению к исходной молекуле вставленной ДНК, например фрагменты, связанные с объектом трансформации. “Фланкирующая область” или “фланкирующая последовательность”, используемая в данном документе, относится к последовательности размером по меньшей мере 20 п.н., для некоторых вариантов осуществления по меньшей мере 50 п.н. и до 5000 п.н., которая расположена либо непосредственно в выше исходной чужеродной молекулы вставленной ДНК и примыкает к ней, либо непосредственно ниже исходной чужеродной молекулы вставленной ДНК и примыкает к ней. Процедуры трансформации чужеродной ДНК приводят к получению трансформантов, содержащих различные фланкирующие области, характерные и уникальные для каждого трансформанта. Когда рекомбинантную ДНК вводят в растение путем традиционного скрещивания, ее фланкирующие области обычно не изменяются. Трансформанты также будут содержать уникальные соединения между частью гетерологичной вставленной ДНК и геномной ДНК, или двумя (2) частями геномной ДНК, или двумя (2) частями гетерологичной ДНК. "Соединение" представляет собой точку, где соединяются два (2) определенных фрагмента ДНК. Например, соединение существует там, где вставленная ДНК присоединяется к фланкирующей ДНК. Точка соединения также существует в трансформированном организме, где два (2) фрагмента ДНК соединяются вместе способом, измененным по сравнению с тем, который обнаруживается в нативном организме. “Соединение ДНК” относится к ДНК, которая содержит точку соединения. Две последовательности соединения, изложенные в настоящем изобретении, представляют собой точку соединения между геномной ДНК маиса и 5’- и 3’-концом вставки, которая указана в одной из SEQ ID NO: 7-14, 31-32 (см. таблицу 1 и прилагаемый перечень последовательностей для описания).
В одном варианте осуществления последовательности соединения объекта DP-202216-6, например одна или несколько из SEQ ID NO: 7-14, 31-32, могут предусматривать полиморфизмы (например, SNP) или мутации, которые могут возникать спонтанно в эндогенной геномной области последовательности соединения. Они могут предусматривать вставку, делецию или замену одного или нескольких нуклеотидов в последовательности соединения. Полинуклеотидные последовательности, которые составляют по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99% от одной или более последовательностей соединения, представленных одной из SEQ ID NO: 7-14, 31-32, раскрыты в данном документе.
Используемое в данном документе выражение “гетерологичный” по отношению к последовательности нуклеиновой кислоты означает, что последовательность нуклеиновой кислоты происходит из другого вида или, если она происходит из того же вида, в значительной степени модифицирована по составу и/или местоположению в геноме по сравнению с ее нативной формой в результате преднамеренного вмешательства человека. Например, промотор, функционально связанный с гетерологичной нуклеотидной последовательностью, может принадлежать к виду, отличному от вида, из которого произошла нуклеотидная последовательность, или, если из того же вида, промотор не обнаруживается в природе функционально связанным с данной нуклеотидной последовательностью. Гетерологичный белок может происходить из другого вида, или, если он происходит из одного и того же вида, он является в значительной степени модифицированным по сравнению с его первоначальной формой путем преднамеренного вмешательства человека.
Термин “регуляторный элемент” относится к молекуле нуклеиновой кислоты, обладающей регуляторной активностью по отношению к гену, т.е. молекуле нуклеиновой кислоты, которая обладает способностью влиять на транскрипционный и/или трансляционный профиль экспрессии функционально связанного транскрибируемого полинуклеотида. Термин “регуляторная активность по отношению к гену”, таким образом, относится к способности влиять на экспрессию функционально связанной транскрибируемой полинуклеотидной молекулы посредством влияния на транскрипцию и/или трансляцию этой функционально связанной транскрибируемой полинуклеотидной молекулы. Регуляторная активность по отношению к гену может быть положительной и/или отрицательной, и при этом эффект может быть охарактеризован по его временным свойствам, пространственным свойствам, свойствам, связанным с развитием, свойствам, связанным с тканями, свойствам, связанным с окружающей средой, физиологическим свойствам, патологическим свойствам, свойствам, связанным с клеточным циклом и/или чувствительностью к химическим веществам, равно как и по количественным или качественным показателям.
“Промотор” означает нуклеотидную последовательность, способную контролировать экспрессию кодирующей последовательности или функциональной РНК. Как правило, кодирующая последовательность расположена в 3'-направлении от промоторной последовательности. Последовательность промотора содержит проксимальные и более дистальные расположенные выше элементы; последние элементы часто называют энхансерами. Соответственно, “энхансер” представляет собой нуклеотидную последовательность, которая может стимулировать активность промотора и может представлять собой изначально природный элемент промотора или гетерологичный элемент, вставленный для повышения уровня или тканевой специфичности промотора. Промоторы могут быть получены как целое из нативного гена или могут состоять из различных элементов, полученных из различных промоторов, встречающихся в природе, или даже содержать синтетические нуклеотидные сегменты. Специалисты в данной области понимают, что разные регуляторные элементы могут управлять экспрессией гена в разных тканях или типах клеток, или на разных стадиях развития, или в ответ на разные условия окружающей среды. Промоторы, которые вызывают экспрессию фрагмента нуклеиновой кислоты в большинстве типов клеток в большинстве случаев, обычно называют “конститутивными промоторами”.
“Лидерная последовательность трансляции” относится к нуклеотидной последовательности, расположенной между последовательностью промотора гена и кодирующей последовательностью. Лидерная последовательность трансляции присутствует в полностью процессированной mRNA выше последовательности инициации трансляции. Лидерная последовательность трансляции может влиять на множество параметров, в том числе процессирование первичного транскрипта до mRNA, стабильность и/или эффективность трансляции mRNA. Были описаны примеры лидерных последовательностей трансляции (Turner and Foster (1995) Mol. Biotechnol. 3:225-236).
Термин “3’-некодирующие последовательности” относится к нуклеотидным последовательностям, расположенным ниже кодирующей последовательности и включающим последовательности распознавания полиаденилирования и другие последовательности, которые кодируют регуляторные сигналы, способные повлиять на процессирование mRNA или экспрессию генов. Сигнал полиаденилирования обычно характеризуют по влиянию добавления участков полиадениловой кислоты к 3’-концу предшественника mRNA. Применение различных 3’-некодирующих последовательностей проиллюстрировано в Ingelbrecht et al. (1989) Plant Cell 1:671-680.
Конструкция ДНК представляет собой совокупность связанных вместе молекул ДНК, которые обеспечивают одну или несколько кассет экспрессии. Конструкция ДНК может представлять собой плазмиду, способную к саморепликации в бактериальной клетке и содержащую сайты рестрикции различных эндонуклеазных ферментов, которые полезны для введения молекул ДНК, обеспечивающих функциональные генетические элементы, т. е., среди прочего, промоторов, интронов, лидерных последовательностей, кодирующих последовательностей, 3’-концевых областей; или конструкция ДНК может представлять собой линейную сборку молекул ДНК, такую как кассета экспрессии. Кассета экспрессии, содержащаяся в конструкции ДНК, содержит необходимые генетические элементы для обеспечения транскрипции информационной РНК. Кассета экспрессии может быть сконструирована для экспрессии в прокариотических клетках или эукариотических клетках. Кассеты экспрессии согласно вариантам осуществления сконструированы для экспрессии в растительных клетках.
Раскрытые в данном документе молекулы ДНК предусмотрены в кассетах экспрессии для экспрессии в представляющем интерес организме. Кассета будет включать 5’- и 3’-регуляторные последовательности, функционально связанные с кодирующей последовательностью. “Функционально связанный” означает, что последовательности нуклеиновой кислоты, которые связаны, являются смежными, и в случае, когда необходимо соединить два белок-кодирующих области, они являются смежными и находятся в одной рамке считывания. “Функционально связанный” предназначено для обозначения функциональной связи между промотором и второй последовательностью, где промоторная последовательность инициирует и опосредует транскрипцию последовательности ДНК, соответствующей второй последовательности. Кассета может дополнительно содержать по меньшей мере один дополнительный ген, подлежащий введению в организм путем котрансформации. В качестве альтернативы, дополнительный(-ые) ген(-ы) может(могут) предусматриваться в нескольких кассетах экспрессии или нескольких конструкциях ДНК.
В направлении транскрипции 5’ - 3’ кассета экспрессии может включать: область инициации транскрипции и трансляции, кодирующую область и область терминации транскрипции и трансляции, функционирующие в организме, служащем хозяином. Область инициации транскрипции (т.е. промотор) может быть нативным или аналогичным или чужеродным или гетерологичным по отношению к организму-хозяину. Кроме того, промотор может представлять собой природную последовательность или, в качестве альтернативы, синтетическую последовательность. Кассеты экспрессии могут дополнительно содержать 5’-лидерные последовательности в конструкции кассеты экспрессии. Такие лидерные последовательности могут способствовать усилению трансляции.
Трансгенный “объект” образуется путем трансформации растительных клеток гетерологичной(-ыим) конструкцией(-ями) ДНК, включающей(-ими) кассету экспрессии нуклеиновой кислоты, которая содержит трансген, представляющий интерес, регенерации популяции растений, полученных в результате вставки трансгена в геном растения, и отбора конкретного растения, характеризующегося вставкой в конкретное местоположение в геноме. Объект характеризуют фенотипически по экспрессии трансгена. На генетическом уровне объект представляет собой часть набора генов растения. Термин “объект” также относится к потомству, полученному путем полового ауткросса между трансформантом и другой разновидностью, которая включает гетерологичную ДНК. Даже после повторных возвратных скрещиваний с реккурентным родителем вставленная ДНК и фланкирующая ДНК из трансформированного родителя присутствуют у потомства скрещивания в том же местоположении в хромосоме. Термин “объект” также относится к ДНК из исходного трансформанта, содержащей вставленную ДНК и фланкирующую последовательность, непосредственно прилегающую ко вставленной ДНК, которая, как ожидается, будет перенесена потомству, которое получает вставленную ДНК, в том числе трансген, представляющий интерес, в результате полового скрещивания родительской линии, которая содержит вставленную ДНК (например, исходный трансформант и потомство, полученное в результате самоопыления), и родительской линии, которая не содержит вставленную ДНК.
Растение кукурузы, содержащее объект DP-202216-6, может быть выведено путем первого скрещивания половым путем первого родительского растения кукурузы, состоящего из растения кукурузы, выращенного из растения объекта DP-202216-6 кукурузы, и его потомства, полученного в результате трансформации кассетами экспрессии по вариантам осуществления, которые повышают урожайность по сравнению с контрольным растением, и второго родительского растения кукурузы, которое не содержит таких конструкций, в результате получая множество растений-потомков первого поколения; и затем отбора растения-потомка первого поколения, демонстрирующего повышение урожайности; и самоопыления растения-потомка первого поколения с получением таким образом множества растений-потомков второго поколения; а затем отбора из растений-потомков второго поколения растения с повышенной урожайностью.
Используемый в данном документе термин "растение" предусматривает ссылку на целые растения, части растений, органы растений (например, листья, стебли, корни и т.д.), семена, растительные клетки и потомство таковых. В некоторых вариантах осуществления части трансгенных растений предусматривают, например, растительные клетки, протопласты, ткани, каллус, зародыши, а также цветы, стебли, плоды, листья и корни, происходящие от трансгенных растений или их потомства, ранее трансформированных раскрытой в данном документе молекулой ДНК, и, следовательно, состоящие по меньшей мере частично из трансгенных клеток.
Как используется в данном документе, выражение "растительная клетка" включает, без ограничения, семена, суспензионные культуры, зародыши, меристематические области, каллюсную ткань, листья, корни, побеги, гаметофиты, спорофиты, пыльцу и микроспоры. Класс растений, которые можно применять, обычно настолько же широк, как и класс высших растений, поддающихся методикам трансформации, включающий как однодольные, так и двудольные растения.
В настоящем изобретении предусмотрен товарный продукт, который получен из растения кукурузы, содержащего объект DP-202216-6. Используемый в данном документе "товарный продукт" обычно относится к любой композиции или материалу, который включает материал, полученный из растения, семени, растительной клетки или части растения, содержащей объект DP-202216-6, или который является результатом их переработки. Товарные продукты могут быть жизнеспособными (например, семена) или нежизнеспособными (например, кукурузная крупа). Нежизнеспособные товарные продукты включают без ограничения нежизнеспособные семена и зерно; переработанные семена, части семян и части растений; высушенную растительную ткань, замороженную растительную ткань и переработанную растительную ткань; семена и части растений, переработанные в корма для животных, предназначенные для потребления наземными и/или водными животными, масло, крупу, муку, хлопья, отруби, клетчатку, молоко, сыр, бумагу, сливки, вино, этанол и любые другие продукты питания для потребления человеком и виды биомассы и топливные продукты. К жизнеспособным товарным продуктам относятся без ограничения семена и растительные клетки. Таким образом, растение, содержащее объект DP-202216-6, можно использовать для изготовления любого подходящего товарного продукта, получаемого из растения кукурузы. Такой товарный продукт, полученный из растений, содержащих объект DP-202216-6, может содержать обнаруживаемое количество специфической и уникальной ДНК, соответствующей объекту DP-202216-6, и, в частности, может содержать обнаруживаемое количество полинуклеотида, имеющего нуклеотидную последовательность из по меньшей мере 15 последовательных нуклеотидов из SEQ ID NO: 9-14, по меньшей мере 20 последовательных нуклеотидов из SEQ ID NO: 9-14 и 31-32, по меньшей мере 30 последовательных нуклеотидов из SEQ ID NO: 9-14 и 31-32. Для товарного продукта можно использовать любой стандартный способ обнаружения полинуклеотидных молекул, в том числе способы обнаружения, раскрытые в данном документе.
“Трансформация” относится к переносу фрагмента нуклеиновой кислоты в геном организма-хозяина, что приводит к генетически стабильному наследованию. Организмы-хозяева, содержащие трансформированные фрагменты нуклеиновой кислоты, называются “трансгенными” организмами. Примеры способов трансформации растений включают трансформацию, опосредованную Agrobacterium (De Blaere et al. (1987) Meth. Enzymol. 143:277), и техники трансформации с ускорением частиц или “генной пушкой” (Klein et al. (1987) Nature (London) 327:70-73; патент США № 4945050, включенные в данный документ посредством ссылки). Дополнительные способы трансформации раскрыты ниже.
Используемый в данном документе термин "потомство" обозначает потомство любого поколения родительского растения, которое содержит объект DP-202216-6 кукурузы.
Выделенные полинуклеотиды, раскрытые в данном документе, могут быть включены в рекомбинантные конструкции, обычно конструкции ДНК, которые способны вводиться в клетку-хозяина и реплицироваться в ней. Такая конструкция может быть вектором, который включает систему репликации и последовательности, способные к транскрипции и трансляции последовательности, кодирующей полипептид, в данной клетке-хозяине. Ряд векторов, подходящих для стабильной трансфекции растительных клеток или для создания трансгенных растений, был описан, например, в Pouwels et al., (1985; Supp. 1987) Cloning Vectors: A Laboratory Manual, Weissbach and Weissbach (1989) Methods for Plant Molecular Biology, (Academic Press, New York) и Flevin et al., (1990) Plant Molecular Biology Manual, (Kluwer Academic Publishers). Как правило, векторы экспрессии в растениях включают, например, один или несколько клонированных генов растения под транскрипционным контролем 5’- и 3’-регуляторных последовательностей и доминантный селектируемый маркер. Такие векторы экспрессии в растениях также могут содержать промоторную регуляторную область (например, регуляторную область, контролирующую индуцируемую или конститутивную, регулируемую факторами окружающей среды или стадиями развития или клеточно- или тканеспецифическую экспрессию), старт-сайт для инициации транскрипции, сайт связывания рибосом, сигнальную последовательность процессинга РНК, сайт терминации транскрипции и/или сигнал полиаденилирования.
В процессе введения вставки в геном растительных клеток нередко возникают некоторые делеции или другие изменения вставки и/или геномных фланкирующих последовательностей. Таким образом, соответствующий сегмент плазмидной последовательности, представленной в данном документе, может содержать некоторые незначительные вариации. То же самое верно и для фланкирующих последовательностей, представленных в данном документе. Таким образом, растение, содержащее полинуклеотид, имеющий некоторый диапазон идентичности с рассматриваемыми фланкирующими и/или вставляемыми последовательностями, находится в пределах объема настоящего изобретения. Идентичность последовательности по настоящему изобретению может представлять собой полинуклеотидную последовательность, обладающую по меньшей мере 65% идентичностью последовательности, для некоторых вариантов осуществления по меньшей мере 70% идентичностью последовательности, для некоторых вариантов осуществления по меньшей мере 75% идентичностью последовательности, для некоторых вариантов осуществления по меньшей мере 80% идентичностью и для некоторых вариантов осуществления по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% идентичностью последовательности с последовательностью, приведенной в качестве примера или описанной в данном документе. Гибридизация и условия гибридизации, как предусмотрено в данном документе, также могут быть использованы для определения таких растений и полинуклеотидных последовательностей согласно настоящему изобретению. Последовательность, которая включает фланкирующие последовательности плюс полную вставляемую последовательность, может быть подтверждена со ссылкой на депонированные семена.
“Зонд” представляет собой выделенную нуклеиновую кислоту, к которой присоединена обычная обнаруживаемая метка или репортерная молекула, например радиоактивный изотоп, лиганд, хемилюминесцентное средство или фермент. Такой зонд является комплементарным цепи целевой нуклеиновой кислоты, например цепи выделенной ДНК из объекта DP-202216-6 кукурузы, будь то из растения кукурузы или из образца, который включает ДНК из объекта. Зонды могут включать не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, но также полиамиды и другие материалы для зондов, которые специфически связываются с последовательностью целевой ДНК и могут использоваться для обнаружения присутствия этой последовательности целевой ДНК. Иллюстративный зонд для обнаружения объекта DP-202216-6 содержит SEQ ID NO: 17. Кроме того, любой меченый зонд, который связывается или демонстрирует комплементарность при условиях высокой жесткости с одной или несколькими последовательностями соединения, например 5'- и/или 3'-последовательностями соединения вставленной ДНК, смежной с геномной ДНК объекта DP-202216-6 маиса, содержащими последовательность, которая на по меньшей мере 99% идентична SEQ ID NO: 7-14, 31 и 32, подходит для применения в качестве зондов.
“Праймеры” представляют собой выделенные нуклеиновые кислоты, которые подвергают отжигу с комплементарной цепью целевой ДНК путем гибридизации нуклеиновых кислот с образованием гибрида праймера и цепи целевой ДНК, которые затем удлиняют вдоль цепи целевой ДНК с помощью полимеразы, например ДНК-полимеразы. Пары праймеров относятся к их применению для амплификации целевой последовательности нуклеиновой кислоты, например с помощью ПЦР или других традиционных способов амплификации нуклеиновых кислот. “ПЦР” или “полимеразная цепная реакция” представляет собой способ, используемый для амплификации определенных сегментов ДНК (см. патенты США №№ 4683195 и 4800159; включенные в данный документ посредством ссылки).
Зонды и праймеры имеют достаточную длину нуклеотидов для связывания с целевой последовательностью ДНК, особенно в условиях гибридизации или условиях реакции, определенных оператором. Эта длина может быть любой длиной, достаточной для использования в выбранном способе обнаружения. Обычно используют длину 11 нуклеотидов или более, 18 нуклеотидов или более и 22 нуклеотида или более. Такие зонды и праймеры специфически гибридизируются с целевой последовательностью в условиях гибридизации высокой жесткости. Зонды и праймеры в соответствии с вариантами осуществления могут иметь полное сходство последовательностей ДНК смежных нуклеотидов с целевой последовательностью, хотя зонды, отличающиеся от целевой последовательности ДНК и сохраняющие способность к гибридизации с целевыми последовательностями ДНК, можно сконструировать обычными способами. Зонды можно использовать в качестве праймеров, но обычно они предназначены для связывания с целевой ДНК или РНК и не используются в процессе амплификации.
Специфические праймеры можно использовать для амплификации встраиваемого фрагмента с целью получения ампликона, который можно использовать в качестве “специфического зонда” для идентификации объекта DP-202216-6 в биологических образцах. Когда зонд гибридизируется с нуклеиновыми кислотами биологического образца в условиях, которые дают возможность связывания зонда с образцом, это связывание может быть обнаружено и, таким образом, позволяет указать на присутствие объекта DP-202216-6 в биологическом образце. Такая идентификация связанного зонда описана в уровне техники. В одном варианте осуществления настоящего изобретения специфический зонд представляет собой последовательность, которая в оптимизированных условиях специфически гибридизируется с областью в пределах 5’- или 3’-фланкирующей области объекта, а также включает часть чужеродной ДНК, смежную с ней. Специфический зонд может содержать последовательность, которая на по меньшей мере 80%, от 80 до 85%, от 85 до 90%, от 90 до 95% и от 95 до 100% идентична (или комплементарна) конкретной области объекта.
Зонды и праймеры (и ампликоны) обычно имеют длину 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, или 500 полинуклеотидов или более. Такие зонды и праймеры специфически гибридизируются с целевой последовательностью в условиях гибридизации высокой жесткости. В некоторых вариантах осуществления зонды и праймеры имеют полное сходство последовательности с целевой последовательностью, хотя зонды, отличающиеся от целевой последовательности и сохраняющие способность гибридизироваться с целевыми последовательностями, можно сконструировать обычными способами.
Способы получения и применения зондов и праймеров описаны, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 1989 (далее, “Sambrook et al., 1989”); Ausubel et al. eds., Current Protocols in Molecular Biology, Greene Publishing and Wiley-Interscience, New York, 1995 (с периодическими обновлениями) (далее, “Ausubel et al., 1995”) и Innis et al., PCR Protocols: A Guide to Methods and Applications, Academic Press: San Diego, 1990. Пары праймеров для ПЦР могут быть получены из известной последовательности, например, с помощью компьютерных программ, предназначенных для этой цели, таких как инструмент анализа праймеров для ПЦР в Vector NTI версии 6 (Informax Inc., Бетесда, штат Мэриленд, США); PrimerSelect (DNASTAR Inc., Мэдисон, штат Висконсин, США) и Primer (Version 0.5©, 1991, Whitehead Institute for Biomedical Research, Кембридж, штат Массачусетс, США). Кроме того, последовательность можно сканировать визуально и идентифицировать праймеры вручную с использованием руководящих принципов, известных специалисту в данной области.
Используемый в данном документе “набор” относится к набору реагентов для целей выполнения вариантов осуществления способа по настоящему изобретению, более конкретно, идентификации объекта DP-202216-6 в биологических образцах. Набор может использоваться, и его компоненты могут быть специально откорректированы, для целей контроля качества (например, чистоты партий семян), обнаружения объекта DP-202216-6 в растительном материале или в материале, содержащем растительный материал, или полученном из него, например таком как без ограничения, продукты питания или корма. Используемый в данном документе “растительный материал” относится к материалу, полученному или происходящему из растения.
Праймеры и зонды, основанные на фланкирующих последовательностях ДНК и вставляемых последовательностях, описанных в данном документе, можно использовать для подтверждения (и, если необходимо, для исправления) раскрытых последовательностей общепринятыми способами, например путем повторного клонирования и секвенирования таких последовательностей. Зонды и праймеры нуклеиновых кислот гибридизируются в жестких условиях с целевой последовательностью ДНК. Любой традиционный способ гибридизации или амплификации нуклеиновых кислот можно использовать для идентификации присутствия ДНК из трансгенного объекта в образце. Молекулы нуклеиновых кислот или их фрагменты способны специфически гибридизироваться с другими молекулами нуклеиновых кислот при определенных обстоятельствах.
Говорят, что молекула нуклеиновой кислоты является “комплементтарной последовательностью” другой молекулы нуклеиновой кислоты, если они демонстрируют полную комплементарность или минимальную комплементарность. Как используется в данном документе, говорят, что молекулы проявляют “полную комплементарность”, когда каждый нуклеотид одной из молекул комплементарен нуклеотиду другой. Две молекулы называются “минимально комплементарными”, если они могут гибридизироваться друг с другом с достаточной стабильностью, чтобы позволить им оставаться отожженными друг с другом по меньшей мере в обычных условиях “низкой жесткости”. Аналогичным образом, говорят, что молекулы являются “комплементарными”, если они могут гибридизироваться друг с другом с достаточной стабильностью, чтобы позволить им оставаться отожженными друг с другом в обычных условиях “высокой жесткости”. Обычные условия жесткости описываются в Sambrook et al., 1989 и в Haymes et al., в Nucleic Acid Hybridization, a Practical Approach, IRL Press, Washington, D.C. (1985), отклонения от полной комплементарности, следовательно, допустимы, если такие отклонения не полностью исключают способность молекул образовывать двухцепочечную структуру. Для того чтобы молекула нуклеиновой кислоты служила праймером или зондом, она должна быть достаточно комплементарной по последовательности, чтобы иметь возможность образовывать стабильную двухцепочечную структуру при конкретных используемых концентрациях растворителя и соли.
В реакциях гибридизации специфичность обычно зависит от отмывок после гибридизации, при этом релевантными факторами являются ионная сила и температура конечного раствора для отмывки. Термическая точка плавления (Tm) представляет собой температуру (при определенной ионной силе и pH), при которой 50% комплементарной целевой последовательности гибридизируется с идеально совпадающим зондом. Для гибридов ДНК-ДНК Tm может быть аппроксимирована из уравнения Meinkoth and Wahl (1984) Anal. Biochem. 138:267-284: Tm=81,5 °C+16,6 (log M) + 0,41 (%GC) - 0,61 (% форм.) - 500/л; где М представляет собой молярность одновалентных катионов, % GC представляет собой процент гуанозиновых и цитозиновых нуклеотидов в ДНК, % форм. представляет собой процент формамида в гибридизационном растворе, и L представляет собой длину гибрида в парах оснований. Tm снижают на приблизительно 1°C для каждого 1% несовпадения; таким образом, Tm, условия гибридизации и/или отмывки можно скорректировать для гибридизации с последовательностями с необходимой идентичностью. Например, если осуществляют поиск последовательностей с идентичностью >90%, то Tm можно снизить на 10 C. Обычно, жесткие условия выбирают таким образом, чтобы температура была приблизительно на 5 C ниже, чем Tm для конкретной последовательности и комплементарной ей последовательности при определенной ионной силе и pH. Тем не менее при условиях высокой жесткости можно проводить гибридизацию и/или отмывку при температуре на 1, 2, 3 или 4°С ниже, чем Tm; при условиях умеренной жесткости можно проводить гибридизацию и/или отмывку при температуре на 6, 7, 8, 9 или 10°C ниже, чем Tm; при условиях низкой жесткости можно проводить гибридизацию и/или отмывку при температуре на 11, 12, 13, 14, 15 или 20°С ниже, чем Tm.
При использовании уравнения, композиций для гибридизации и отмывки и необходимой Tm специалистам в данной области будет понятно, что вариации жесткости растворов для гибридизации и/или отмывки по определению описаны. Если требуемая степень ошибочного спаривания приводит к Tm меньше 45°C (водный раствор) или 32°C (раствор формамида), предпочтительно увеличивать концентрацию SSC так, чтобы можно было применять более высокую температуру. Исчерпывающее пособие по гибридизации нуклеиновых кислот находится в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 (Elsevier, New York), и Ausubel et al., eds. (1995), и Sambrook et al. (1989).
Принцип гибридизационного анализа состоит в том, что одноцепочечная молекула ДНК или РНК с известной последовательностью (например, зонд) может соединяться в пару со второй молекулой ДНК или РНК, которая содержит комплементарную последовательность (мишень), со стабильностью гибридизации в зависимости от степени спаривания оснований, которая происходит в тестируемых условиях. Соответствующие условия жесткости для гибридизации ДНК, включая, например, 6x хлорид натрия/цитрат натрия (SSC) при приблизительно 45°C с последующей промывкой 2,0×SSC при 50°C, известны специалистам в данной области или могут быть найдены в Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6. Например, концентрация солей на стадии отмывки может быть выбрана от низкой жесткости приблизительно 2,0×SSC при 50°C до высокой жесткости приблизительно 0,2×SSC при 50°C или до 0,1×SSC или 0,2×SSC при 55°C или 65°C. Кроме того, температура на стадии отмывки может быть повышена от условий низкой жесткости при комнатной температуре, приблизительно 22°C, до условий высокой жесткости приблизительно при 65°C. Можно изменять как температуру, так и концентрацию солей, или либо температура, либо концентрация солей может оставаться постоянной, в то время как другая переменная (например, время) изменяется. В одном варианте осуществления нуклеиновая кислота по настоящему изобретению будет специфически гибридизироваться с одной или несколькими молекулами нуклеиновой кислоты, указанными в SEQ ID NO: 6-14, или их комплементарными последовательностями или фрагментами в условиях высокой жесткости. Гибридизацию зонда с целевой молекулой ДНК можно обнаружить способами, известными специалистам в данной области. Они могут включать без ограничения флуоресцентные метки, радиоактивные метки, метки на основе антител и хемилюминесцентные метки.
В некоторых вариантах осуществления комплементарная последовательность имеет ту же длину, что и молекула нуклеиновой кислоты, с которой она гибридизируется. В некоторых вариантах осуществления комплементарная последовательность на 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов длиннее или короче молекулы нуклеиновой кислоты, с которой она гибридизируется. В некоторых вариантах осуществления комплементарная последовательность на 1%, 2%, 3%, 4% или 5% длиннее или короче молекулы нуклеиновой кислоты, с которой она гибридизируется. В некоторых вариантах осуществления комплементарная последовательность комплементарна нуклеотид-к-нуклеотиду, что означает, что нет ошибочно спаренных нуклеотидов (каждый А спарен с Т, а каждый G спарен с С). В некоторых вариантах осуществления комплементарная последовательность содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или менее ошибочных спариваний. В некоторых вариантах осуществления комплементарная последовательность содержит 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9% или 10% или менее ошибочных спариваний.
"Процентная (%) идентичность последовательности" относительно эталонной последовательности (рассматриваемой) определяется как процентная доля аминокислотных остатков или нуклеотидов в кандидатной последовательности (запрашиваемой), которые идентичны соответствующим аминокислотным остаткам или нуклеотидам эталонной последовательности после выравнивания последовательностей и внесения гэпов, при необходимости, для достижения максимальной процентной идентичности последовательности и без учета каких-либо консервативных аминокислотных замен как части идентичности последовательности. Выравнивание в целях определения процентной идентичности последовательности может быть достигнуто различными способами, которые известны специалисту в данной области, например с использованием общедоступного компьютерного программного обеспечения, такого как BLAST, BLAST-2. Специалисты в данной области могут определить подходящие параметры для выравнивания последовательностей, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Процентная идентичность двух последовательностей является функцией количества идентичных положений, имеющихся в обеих последовательностях (например, процентная идентичность запрашиваемой последовательности = количество идентичных положений между запрашиваемой последовательностью и рассматриваемой последовательностью/общее количество положений в запрашиваемой последовательности × 100). Например, способ множественного выравнивания последовательностей Clustal W описан в Thompson J, Higgins D and Gibson T (1994). Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting." Nucleic Acids Research, Vol. 22: pp. 4673-80. Другим способом является Clustal V, описанный в Higgins DG and Sharp PM (1989). "Fast and sensitive multiple sequence alignments on a microcomputer." CABIOS, Vol. 5, No. 2: pp. 151-153.
Что касается амплификации последовательности целевой нуклеиновой кислоты (например, с помощью ПЦР) с использованием конкретной пары праймеров для амплификации, жесткие условия позволяют паре праймеров гибридизироваться только с последовательностью целевой нуклеиновой кислоты, с которой праймер, имеющий соответствующую последовательность дикого типа (или ее комплементарных последовательностей), будет связываться с образованием уникального продукта амплификации, ампликона, в реакции термической амплификации ДНК.
Используемые в данном документе “амплифицированная ДНК” или “ампликон” относятся к нуклеиновой кислоте, полученной в результате амплификации целевой последовательности нуклеиновой кислоты, которая является частью матрицы нуклеиновой кислоты. Например, в одном варианте осуществления такая амплифицированная ДНК или ампликоны могут содержать последовательность нуклеиновой кислоты, которая специфична для объектов, раскрытых в данном документе, например DP-202216-6. ДНК, выделенную из образца ткани растения, можно подвергнуть способу амплификации нуклеиновой кислоты с использованием пары ДНК-праймеров, которая включает первый праймер, полученный из фланкирующей последовательности, смежной с сайтом вставки вставленной гетерологичной ДНК, и второй праймер, полученный из вставленной гетерологичной ДНК, для получения ампликона, который является диагностическим признаком присутствия ДНК объекта, например DP-202216-6. В качестве альтернативы, второй праймер может быть получен из фланкирующей геномной последовательности. Ампликон может быть любой подходящей длины и иметь последовательность нуклеиновой кислоты, которая также является диагностическим признаком объекта. В качестве альтернативы, пары праймеров могут быть получены из фланкирующей последовательности с обеих сторон вставленной ДНК, чтобы получить ампликон, который включает всю вставленную нуклеотидную последовательность, а также последовательность, фланкирующую вставку. Праймер или пара праймеров, полученные из фланкирующей геномной последовательности, могут быть расположены на расстоянии от вставленной последовательности ДНК, это расстояние может варьироваться от одной пары нуклеотидных оснований до пределов реакции амплификации, например 10000 или приблизительно 20000 п.н.
Амплификация нуклеиновой кислоты может выполняться любым из различных способов амплификации нуклеиновой кислоты, известных в уровне техники, в том числе ПЦР. В уровне техники известно множество способов амплификации, которые описаны, среди прочего, в патентах США №№ 4683195 и 4683202 и в Innis et al., (1990) выше. Способы ПЦР-амплификации были разработаны для амплификации геномной ДНК размером до 22 т.н. и до 42 т.н. ДНК бактериофага (Cheng et al., Proc. Natl. Acad. Sci. USA 91:5695-5699, 1994). Эти способы, а также другие способы, известные в уровне техники амплификации ДНК, можно использовать при применении вариантов осуществления настоящего изобретения. Понятно, что некоторые параметры в конкретном протоколе ПЦР могут нуждаться в корректировке для конкретных лабораторных условий и могут быть немного изменены, но все же позволяют получить аналогичные результаты. Эти настройки будут очевидны специалисту в данной области.
Ампликон, полученный этими способами, может быть обнаружен множеством методик, в том числе без ограничения Genetic Bit Analysis (Nikiforov, et al. Nucleic Acid Res. 22:4167-4175, 1994), где конструируется олигонуклеотид ДНК, который перекрывает как соседнюю фланкирующую последовательность ДНК, так и вставленную последовательность ДНК. Олигонуклеотид иммобилизуют в лунках микропланшета. После ПЦР представляющей интерес области (с использованием одного праймера во вставленной последовательности и одного в соседней фланкирующей последовательности) одноцепочечный ПЦР-продукт может гибридизироваться с иммобилизованным олигонуклеотидом и служить в качестве матрицы для реакции удлинения на одно основание с использованием ДНК-полимеразы и меченых ddNTP, специфичных для ожидаемого следующего основания. Считывание может быть на основе флуоресценции или ELISA. Сигнал указывает на присутствие вставленной/фланкирующей последовательности из-за успешных амплификации, гибридизации и удлинения на одно основание.
Другим способом обнаружения является методика пиросеквенирования, описанная в Winge (2000) Innov. Pharma. Tech. 00:18-24. В этом способе конструируется олигонуклеотид, который перекрывает соединение смежной ДНК и вставленной ДНК. Олигонуклеотид гибридизируют с одноцепочечным ПЦР-продуктом из представляющей интерес области (один праймер во вставленной последовательности и один во фланкирующей последовательности) и инкубируют в присутствии ДНК-полимеразы, АТФ, сульфурилазы, люциферазы, апиразы, аденозин-5’-фосфосульфата и люциферина. dNTP добавляют по отдельности, и встраивание дает световой сигнал, который измеряют. Световой сигнал указывает на присутствие трансгенной вставленной/фланкирующей последовательности благодаря успешным амплификации, гибридизации и удлинению на одно или несколько оснований.
Поляризация флуоресценции, как описано Chen et al., (1999) Genome Res. 9:492-498, также представляет собой способ, который можно применять для обнаружения ампликона. При применении этого способа конструируется олигонуклеотид, который перекрывает соединение фланкирующей и вставленной ДНК. Олигонуклеотид гибридизуют с одноцепочечным ПЦР-продуктом из представляющей интерес области (один праймер во вставленной ДНК и один во фланкирующей последовательности ДНК) и инкубируют в присутствии ДНК-полимеразы и флуоресцентно меченного ddNTP. Удлинение на одно основание приводит к включению ddNTP. Включение можно измерить как изменение поляризации с помощью флуорометра. Изменение поляризации указывает на присутствие трансгенной вставленной/фланкирующей последовательности благодаря успешным амплификации, гибридизации и удлинению на одно основание.
TaqMan® (PE Applied Biosystems, Фостер-Сити, штат Калифорния, США) представляет собой количественную реакцию амплификации (qPCR) для обнаружения и количественной оценки присутствия последовательности ДНК, и она доступна для приобретения. Коротко, зонды TaqMan сконструированы таким образом, что они подвергаются отжигу в области ДНК, амплифицированной определенным набором праймеров, и включают флуорофорный (FRET) олигонуклеотидный зонд, который перекрывает соединение фланкирующей ДНК и вставленной ДНК. FRET-зонд и ПЦР-праймеры (один праймер в последовательности вставленной ДНК и один во фланкирующей геномной последовательности) подвергают термоциклированию в присутствии термостабильной полимеразы и dNTP. Гибридизация FRET-зонда приводит к расщеплению и отделению флуоресцентного фрагмента от гасящего фрагмента на FRET-зонде. Флуоресцентный сигнал указывает на присутствие и/или количество фланкирующей/трансгенной вставленной последовательности благодаря успешной амплификации и гибридизации.
Молекулярные маяки были описаны для применения при обнаружении последовательности, как описано в Tyangi et al. (1996) Nature Biotech. 14:303-308. Коротко, конструируют олигонуклеотидный FRET-зонд, который перекрывает соединение фланкирующей ДНК и вставленной ДНК. Уникальная структура FRET-зонда приводит к тому, что он содержит вторичную структуру, которая удерживает флуоресцентный и гасящий фрагменты в непосредственной близости. FRET-зонд и ПЦР-праймеры (один праймер в последовательности вставленной ДНК и один во фланкирующей последовательности) подвергают термоциклированию в присутствии термостабильной полимеразы и dNTP. После успешной ПЦР-амплификации гибридизация FRET-зонда с целевой последовательностью приводит к удалению вторичной структуры зонда и пространственному разделению флуоресцентных и гасящих фрагментов. Получают флуоресцентный сигнал. Флуоресцентный сигнал указывает на наличие фланкирующей/трансгенной вставленной последовательности благодаря успешной амплификации и гибридизации.
Термин "аллель" относится к альтернативной форме гена, при которой два гена могут отличаться по последовательностям ДНК. Такие различия могут быть результатом по меньшей мере одной мутации (например, делеции, вставки и/или замены) в последовательности нуклеиновой кислоты. Аллели могут приводить к модифицированным mRNA или полипептидам, структура или функция которых могут быть изменены, а могут и не изменяться. У любого данного гена может не быть ни одной аллельной формы, может быть одна или много аллельных форм. Каждый из этих типов изменений может происходить отдельно или в комбинации с другими один или несколько раз в заданной последовательности.
Реакция гибридизации с использованием зонда, специфичного для последовательности, находящейся в ампликоне, является еще одним способом, применяемым для обнаружения ампликона, полученного в результате реакции ПЦР.
Термин "зиготность" обычно относится к сходству аллелей гена или признака в организме (например, в растении). Если оба аллеля одинаковы, то организм гомозиготен по аллелю. Если два аллеля различны, то организм гетерозиготен по гену или признаку. Если один аллель отсутствует, то организм гемизиготен. Если оба аллеля отсутствуют, то организм нуль-зиготен. Например, растение является гомозиготным по представляющему интерес признаку, если вставленная ДНК вместе с последовательностью соединения присутствует в одном и том же месте на каждой хромосоме из пары хромосом (оба аллеля). Например, растение маиса, имеющее объект DP-202216-6 в одном и том же месте на обеих копиях хромосомы. Аналогичным образом, растение считается гетерозиготным, если вставка трансгена вместе с последовательностью соединения (например, объекта DP-202216-6) присутствует только на одной хромосоме из пары хромосом (только один аллель). Растение дикого типа считается “нулевым” по сравнению с ДНК трансгенного объекта.
Термин "метка" при использовании в данном документе относится к обнаруживаемому соединению или композиции, которые прямо или косвенно конъюгированы с зондом с получением "меченого" зонда. Метка может быть обнаружена сама по себе (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать химическое изменение субстратного соединения или композиции, которая поддается обнаружению (например, авидин-биотин).
Используемый в данном документе термин “линия” представляет собой группу растений, которые демонстрируют незначительные генетические вариации между особями по меньшей мере по одному признаку или вообще не имеют их. Такие линии могут быть созданы несколькими поколениями самоопыления и отбора или вегетативным размножением от одного родителя с использованием методик культивирования тканей или клеток.
Используемые в данном документе термины “культивар” и “сорт” являются синонимами и относятся к линии, которая используется для коммерческого производства. “Стабильность” или “стабильный” означает, что по отношению к данному компоненту этот компонент поддерживается от поколения к поколению и, для некоторых вариантов осуществления, по меньшей мере три поколения практически на одном уровне, например для некоторых вариантов осуществления ± 15%, для некоторых вариантов осуществления ± 10%, в большинстве для некоторых вариантов осуществления ± 5%. На стабильность могут влиять температура, местоположение, стресс и время посадки.
“Агрономически элитный” означает, что линия обладает желательными агрономическими характеристиками, такими как зрелость, устойчивость к болезням, устойчивость к полеганию, высота початка, высота растения и т.п., в дополнение к повышению урожайности благодаря рассматриваемому(-ым) объекту(-ам).
В некоторых вариантах осуществления объект маиса DP-202216-6 может дополнительно содержать пакет дополнительных признаков. Растения, содержащие "пакеты" из последовательностей полинуклеотидов, можно получить с помощью традиционных способов разведения или посредством способов генной инженерии либо с применением и того, и другого. Эти способы включают без ограничения разведение отдельных линий, каждая из которых содержит полинуклеотид, представляющий интерес, трансформацию трансгенного растения, содержащего ген, раскрытый в данном документе, дополнительным геном и котрансформацию генами одной растительной клетки. Применяемый в данном документе термин “пакетированный” включает состояние, при котором в одном растении присутствуют несколько признаков (т.е. оба признака внедрены в ядерный геном, один признак внедрен в ядерный геном и один признак внедрен в геном пластиды или оба признака внедрены в геном пластиды). Дополнительные признаки могут включать, например, засухоустойчивость и признаки переносимости другого вида абиотического стресса. Такие признаки могут быть введены путем скрещивания с растениями маиса, содержащими другие рекомбинантные объекты, или с растениями маиса, содержащими нативные варианты или варианты с отредактированным геномом.
В некоторых вариантах осуществления объект DP-202216-6 маиса может быть пакетирован с одним или несколькими дополнительными признаками, влияющими на затраты (например, устойчивость к гербицидам, устойчивость к грибам, устойчивость к вирусам, переносимость стрессов, устойчивость к заболеваниям, мужская стерильность, сила стебля и т.п.), или признаками, влияющими на выход (например, повышенная урожайность, модифицированные разновидности крахмала, улучшенный профиль масел, сбалансированные аминокислоты, высокое содержание лизина или метионина, повышенная усвояемость, улучшенное качество волокон, засухоустойчивость и т.п.). В дополнительном варианте осуществления объект DP-202216-6 маиса может быть объединен с одним или несколькими дополнительными инсектицидными токсинами Bt или другими инсектицидными белками, отличными от Bt.
В некоторых вариантах осуществления растения кукурузы, содержащие объект DP-202216-6, можно скрещивать с растениями кукурузы, содержащими другие объекты кукурузы или их комбинацию, и оценивать полученные свойства растений-потомков. Например, растения кукурузы, содержащие объект DP-202216-6, можно скрещивать или объединять с растениями кукурузы, включая одну или несколько следующих комбинаций: MON810; DAS-59122-7; MIR604; MON89034; MON863; MON87411; MON87403; MON87427; MON-00603-6 (NK603); MON-87460-4; MON-88017-3; LY038; TC1507; 5307; DAS-06275-8; BT176; BT11; MIR162; GA21; MZDT09Y; SYN-05307-1; DP-004114-3 и DAS-40278-9.
Следующие примеры представлены для иллюстрации, а не для ограничения. Как описано в данном документе, объект DP-202216-6 также называется “Объектом 16”, “E16”, “объектом 16” или “объектом 16-6”, и все они относятся к одному и тому же объекту маиса DP-202216-6. Белок, кодируемый геном MADS-бокса маиса ZmM28 в плазмиде PHP40099, или объект DP-202216-6, также упоминается как белок AG099, а соответствующая последовательность ДНК - как ген AG099 или ДНК AG099.
ПРИМЕРЫ
Пример 1
Характеристики растений с AG099 по уровням урожайности, обработкам, гибридам и плотности популяции
Для оценки урожайности элитных гибридов маиса с объектами AG099 была проведена серия испытаний урожайности зерна с 1-го по 4-й годы на элитных гибридах кукурузы в нескольких местоположениях проведения испытаний. Всего приблизительно восемьдесят шесть местоположений, содержащих примерно тридцать уникальных гибридов со сроком созревания от 105 до 112 дней, использовали для оценки характеристик объектов AG099 по сравнению с контролем дикого типа. Центры проведения испытаний были созданы в таких местоположениях, как штаты Айова, Иллинойс, Миссури, Небраска, Индиана, Канзас, Техас, Калифорния, Висконсин, Южная Дакота и Миннесота. На местоположениях удалось достичь различных уровней урожайности в диапазоне от 70 бушелей/акр, вызванных сильной засухой, до 250 бушелей/акр при оптимальных условиях выращивания. Типы почв состояли из разнообразных "высоких" песков, супесей, илистых суглинков, суглинков и некоторых типов глин.
Во всех местоположениях вводные образцы, содержащие конструкцию AG099 (объекты 18 и объекты 16), сравнивали непосредственно с диким типом в отношении фонового генотипа каждого конкретного гибрида (таблица 2). От двух до четырех повторов проектов дробных делянок создавали в каждом местоположении, с основной делянкой гибрида и вводным образцом в виде вспомогательной делянки (WT, объект 18, объект 16).
Смешанную модель дисперсионного анализа выполняли с использованием ASREML, где BLUE (наилучшие линейные объективные оценки) получали для каждого объекта и дикого типа для всех гибридов в данном местоположении. Для проверки значимых различий проводили попарные сопоставления BLUE этих объектов с BLUE дикого типа.
Таблица 2. Сравнение двух объектов AG099 в нескольких фоновых генотипах гибридов с контролем дикого типа, усредненное по множеству местоположений в течение 4-го года многолетнего испытания.
По всем местоположениям проведения испытаний и гибридам Объект 16 и Объект 18 продемонстрировали средний прирост на 4,1 бушеля/акр и 3,5 бушеля/акр соответственно. Более того, при усреднении по всем гибридам в условиях окружающей среды, Объект 16 улучшил урожайность по сравнению с контролем дикого типа в 83% этих условий окружающей среды (~ 5,7 бушелей/акр), а Объект 18 улучшил урожайность в 78% условий окружающей среды.
Таким образом, повышенная экспрессия гена, кодирующего AG099 (SEQ ID NO: 1), как показано на примере Объекта 16, приводит к появлению растений с повышенной урожайностью зерна в нескольких фоновых генотипах гибридов кукурузы, в нескольких местоположениях, в разных условиях окружающей среды при проведении испытаний и повторяется в течение нескольких лет проведения испытаний.
Таблица 3. Повышение урожайности по сравнению с контролем и % получения выгоды в различных условиях окружающей среды при получении урожая для объектов AG099
Несколько центров проведения испытаний урожайности были созданы в штатах Айова, Иллинойс, Миссури, Небраска, Индиана, Висконсин и Миннесота для оценки объектов AG099 в высокоурожайных районах, характерных для кукурузного пояса Среднего Запада США. В этих местоположениях были установлены значения урожайности более 160 бушелей/акр и часто превышали 180 бушелей/акр, что отображает большую часть наиболее продуктивных регионов выращивания кукурузы в Соединенных Штатах Америки. Чтобы оценить реакцию AG099 на стресс, вызванный засухой, были созданы дополнительные центры в штате Канзас, Техас и Калифорния, в которых можно было отдельно регулировать количество воды, вносимой на испытательных делянках в течение вегетационного периода. Управляемые стрессовые условия варьировались от тяжелого стресса с менее 120 бушелей/акр до очень легкого стресса чуть ниже 160 бушелей/акр.
Из результатов, показанных в таблице 3, видно, что Объект 16 имел 91% получения выгоды и продемонстрировал повышение урожайности на 5 бушелей/акр по сравнению с диким типом в зоне с урожайностью менее 120 бушелей/акр. В зоне с урожайностью 120-160 бушелей/акр Объект 16 имел 91% получения выгоды и повышение урожайности приблизительно на 5,3 бушелей/акр по сравнению с контролем дикого типа. В зоне с умеренно высокой урожайностью (160-200 бушелей/акр) Объект 16 показал существенно более высокий % получения выгоды (88%) по сравнению с Объектом 18 и повышение урожайности на 5 бушелей/акр по сравнению с контролем дикого типа. В зоне проведения испытаний с самой высокой урожайностью - зоне с более 200 бушелями/акр, Объект 16 дал приблизительно на 3 бушеля/акр больше, чем контроль дикого типа, и имел 77% получения выгоды в различных местоположениях.
Для тестирования объектов, содержащих AG099, и полученного в результате повышения урожайности в популяциях с различной посадкой, проводили эксперимент на 4 году в шести уникальных местоположениях (таблица 4). Экспериментальные виды обработки включали популяции с посадкой 36000, 40000, 44000 и 48000 растений на акр. В каждой популяции объект DP-202216-6 кукурузы и контроль дикого типа оценивали в фоновых генотипах 12 различных гибридов. Урожайность объекта DP-202216-6 кукурузы в пределах одной плотности и по разным плотностям измеряли путем вычисления разницы в урожайности (BLUE) Объекта 16 и урожайности дикого типа.
Таблица 4. Уровень урожайности объекта DP-202216-6 (E16) кукурузы при различных значениях плотности популяции.
N = общее количество сравнений = номер места x фоновый генотип гибрида x количество повторений.
Объект DP-202216-6 для всех гибридов привел к повышению урожайности зерна по сравнению с диким типом во всех испытанных популяциях посадки. Повышение урожайности по сравнению с контролем дикого типа составило приблизительно 5,0 бушелей/акр при самой высокой испытанной плотности популяции растений, составлявшей 48000 растений на акр. Это на 2,1 бушеля/акр больше, чем достигнутое повышение при испытании самой низкой плотности популяции с 36000 растений на акр.
Пример 2
Характеристики вторичных признаков растений маиса, содержащих объект DP-202216-6
Данные о вторичных агрономических признаках были взяты для обоих объектов (Объекты 16 и 18) в поле за четырехлетний период. Статистически значимое увеличение высоты растений и початков наблюдалось в обоих объектах при усреднении по всем годам и оцененным гибридам. Оба объекта увеличили высоту растений на 0,7 дюйма. Объект 16 и объект 18 показали увеличенную высоту початков по сравнению с контролем дикого типа на 2,0 и 1,4 дюйма соответственно. Для объекта 16 не наблюдалось значительных различий ни в раннем, ни в позднем полегании, однако растения объекта 18 показали повышенное раннее полегание на 4,8% по сравнению с контролем. Ломкость у объекта 18 была больше, чем у контроля дикого типа, тогда как объект 16 не показал статистически значимой разницы для ломкости. Для объекта 16 не наблюдалось разницы в натурной массе, однако у объекта 18 натурная масса снизилась на 0,5 фунта/бушель по сравнению с контролем. У обоих объектов влажность зерна немного увеличилась по сравнению с контролем дикого типа, на 0,2% и 0,3% для объектов 16 и 18 соответственно. Плесневение початков кукурузы из-за фузариоза было значительно снижено по сравнению с контролем дикого типа на 1 и 0,6 балла по шкале от одного до десяти для объектов 16 и 18 соответственно. Результаты показаны в таблице 5 ниже.
Таблица 5. Сравнение вторичных признаков в гибридах маиса для объектов 16 и 18 по сравнению с контролем.
Единицы степени роста до шелка (GDUSLK). Измерение регистрирует общее накопленное количество единиц степени роста, когда у 50% растений на делянке полностью показался шелк. Эквивалент одного дня составляет приблизительно 2,5 единицы степени роста (GDU) для этого набора данных.
Единицы степени роста до осыпания (GDUSHD). Измерение регистрирует общее накопленное количество единиц степени роста, когда 50% растений на делянке имеют метелки, которые сбрасывают пыльцу. Эквивалент одного дня для этого набора данных составляет примерно 2,5 единицы степени роста.
Высота початка (EARHT). Измерение от земли до точки крепления наиболее развитого початка на растении. Высота початка измеряется в дюймах.
Высота растения (PLTHT). Измерение от земли до основания флагового листа. Высота растения измеряется в дюймах.
Влажность (MST). Измерение влажности зерна в процентах при уборке урожая.
Урожайность. Зарегистрированная масса зерна, собранного с каждой делянки. Расчеты зарегистрированной урожайности в бушелях/акр сделали с учетом измеренной влажности для каждой делянки.
Инбредные испытания размещали в восьми местоположениях с двумя повторениями перечня вводных образцов в каждом местоположении. Оба повторения высаживали в виде гнездового плана, где объекты AG099 и дикий тип создавали вложенную структуру на основе инбредного фонового генотипа. Агрономические данные и наблюдения собирали в ходе инбредных испытаний и анализировали для сравнения с вводным образцом дикого типа (WT) или с версией того же генотипа без признака AG099. В среднем и у разных инбредов присутствие гена AG099 как части объекта DP-202216-6 не показало каких-либо значимых агрономических характеристик в рамках инбредных оценок.
Для оценки данных по гибридам использовали схему модели со смешанными эффектами при проведении анализа нескольких местоположений. В анализе нескольких местоположений главный эффект строения конструкции рассматривается как фиксированный эффект. Факторы для месторасположения, фонового генотипа, тестера, объекта, фонового генотипа от строения конструкции, тестера от строения конструкции, тестера от объекта, местоположения от фонового генотипа, местоположения от строения конструкции, местоположения от тестера, местоположения от фонового генотипа от строения конструкции, местоположения от тестера от строения конструкции, местоположения от объекта, местоположения от тестера от объекта считаются случайными эффектами. Пространственные эффекты, в том числе область и делянка в пределах местоположений, рассматривали как случайные эффекты для удаления постороннего пространственного шума. Гетерогенный остаток задавали с авторегрессионной корреляцией как AR1*AR1 для каждого местоположения. Производили оценку строения конструкции и прогноз объекта для каждого фонового генотипа. Проводили T-тесты для сравнения строения конструкции/объекта с WT. Различие считалось статистически значимым, если значение P данного различия составляло менее 0,05. Анализ урожайности осуществляли с помощью ASREML (VSN International Ltd; Best Linear Unbiased Prediction; Cullis, B. Ret al (1998) Biometrics 54: 1-18, Gilmour, A. R. et al (2009); ASReml User Guide 3.0, Gilmour, A.R., et al (1995) Biometrics 51: 1440-50).
Для оценки данных по инбредам использовали схему модели со смешанными эффектами при проведении анализа нескольких местоположений. В анализе нескольких местоположений главный эффект строения конструкции рассматривается как фиксированный эффект. Факторы для местоположения, фонового генотипа, объекта, фонового генотипа от строения конструкции, местоположения от фонового генотипа, местоположения от строения конструкции, местоположения от фонового генотипа от строения конструкции, местоположения от объекта и повтора в местоположении считаются случайными эффектами. Пространственные эффекты, в том числе область и делянка в пределах местоположений, рассматривали как случайные эффекты для удаления постороннего пространственного шума. Гетерогенный остаток задавали с авторегрессионной корреляцией как AR1*AR1 для каждого местоположения. Производили оценку строения конструкции и прогноз объекта для каждого фонового генотипа. Проводили T-тесты для сравнения строения конструкции/объекта с WT. Различие считалось статистически значимым, если значение P данного различия составляло менее 0,05. Анализ урожайности осуществляли с помощью ASREML (VSN International Ltd; Best Linear Unbiased Prediction; Cullis, B. Ret al (1998) Biometrics 54: 1-18, Gilmour, A. R. et al (2009); ASReml User Guide 3.0, Gilmour, A.R., et al (1995) Biometrics 51: 1440-50).
Пример 3
Повышение урожайности зерна маиса в поле
Трансгенные растения маиса, экспрессирующие рекомбинантную полинуклеотидную последовательность маиса, кодирующую полипептид (SEQ ID NO: 1), проходили полевые испытания. Трансгенные растения маиса продемонстрировали эффективность в отношении повышенной урожайности и стабильности урожайности. Трансформированные растения маиса, содержащие рекомбинантный полинуклеотид, кодирующий SEQ ID NO: 1, затем преобразовали в элитных инбредов и провели внутривидовое скрещивание для серии испытаний урожайности зерна в течение трех лет. Несколько гибридных платформ оценивали в нескольких уникальных условиях окружающей среды, которые включали различные уровни засухи и недостатка азота, а также условия окружающей среды, нацеленные на оптимальные уровни урожайности, где уровни урожайности варьировались от 80 бушелей/акр до 250 бушелей/акр. Для этих полевых испытаний использовали среднюю плотность посадки от приблизительно 30000 до приблизительно 36000 растений/акр. Плотность посадки большинства испытательных делянок составляла от приблизительно 34000 до приблизительно 35000 растений на акр. Эффективность трансгена продемонстрировали в сравнении с изогенным контролем дикого типа во всех испытаниях. Эффективность в отношении урожайности и стабильности урожайности продемонстрировали в сравнении с контролем во всех категориях классификации условий окружающей среды, а также по общей разнице с контролем в течение трех лет испытаний (ниже). В этом примере рекомбинантный полинуклеотид, кодирующий SEQ ID NO: 1, экспрессировали под управлением умеренно конститутивного промотора, например последовательности промотора GOS2 маиса. Трансгенные растения маиса, содержащие рекомбинантный полинуклеотид, кодирующий SEQ ID NO: 1, продемонстрировали повышение урожайности приблизительно на 3,4 бушеля/акр по сравнению с контрольными растениями маиса, не содержащими рекомбинантный ген AG099, на основании многолетних мультигибридных полевых испытаний в нескольких местоположениях при Р <0,05. Смешанную модель дисперсионного анализа проводили с использованием ASREML.
Для оценки эффекта ZmGos2, функционально связанного с полинуклеотидом, кодирующим SEQ ID NO: 1, на урожайность зерна, проводили полевые испытания в широком диапазоне условий окружающей среды в течение четырех лет. За четырехлетний период оценили в общей сложности 40 фоновых генотипов уникальных гибридов в сочетании с объектами DP-202216-6 и DP382118. Для каждого фонового генотипа гибрида проводили внутривидовое скрещивание инбредной конверсии, которая была гомозиготной по соответствующему объекту, с выбранным тестером для получения семени F1. Для одного и того же фонового генотипа гибрида проводили внутривидовое скрещивание рекуррентного родителя конверсии с тем же тестером для получения семени F1 контроля. Все последующие сравнения проводили между гетерозиготным трансгенным гибридом F1 и контролем того же самого фонового генотипа гибрида.
В течение четырехлетнего периода экспериментальные вводные образцы оценивались в различных условиях окружающей среды в центрах проведения испытаний, расположенных в Вудленде, штат Калифорния; Плейнвью, штат Техас; Гарден-Сити, штат Канзас; Йорк, штат Небраска; Юнион-Сити, штат Теннесси; Джонстон, штат Айова; Адель, штат Айова; Марион, штат Айова; Ридлин, штат Айова; Риснор, штат Айова; Майами, штат Миссури; Сикестон, штат Миссури; Сайота, штат Иллинойс; Сан-Хосе, штат Иллинойс; Буда, штат Иллинойс; Принстон, штат Иллинойс; Гумбольдт, штат Иллинойс; Сеймур, штат Иллинойс; Уиндфолл, штат Индиана; Волга, штат Южная Дакота; Джейнсвилл, штат Висконсин; Манкато, штат Миннесота и Вилуко и Буэн, Чили. Все центры проведения испытаний были созданы и управлялись с целью достижения оптимальных уровней урожайности. Пятьдесят шесть уникальных центров проведения испытаний предоставили данные достаточного качества за четырехлетний период. Средние уровни урожайности в этих центрах проведения испытаний колебались от 10900 кг/га до 19570 кг/га.
Экспериментальные проекты представляли собой дробные делянки с фоновым генотипом гибрида в виде основной делянки и объекта или контроля в виде вспомогательной делянки. В каждом центре проведения испытаний создавали от двух до трех повторов, причем основные делянки рандомизировали в пределах повтора, а вспомогательные делянки рандомизировали в пределах основной делянки. Экспериментальные вводные образцы выращивали на четырехрядных делянках длиной от 4,4 м до 5,3 м с проходом 0,5 м между ними. Целые центры проведения испытаний и отдельные делянки низкого качества исключили из процедур сбора и анализа данных в соответствии со стандартной процедурой. Массу и влажность зерна для каждого экспериментального вводного образца измеряли, собирая два центральных ряда четырехрядной делянки с использованием исследовательского комбайна для небольших делянок. Урожайность стандартизировали в рамках эксперимента путем коррекции массы собранного зерна на каждой делянке до пятнадцати процентов влажности. Смешанную модель дисперсионного анализа проводили с использованием ASREML с учетом случайных полевых и пространственных компонентов, а также фиксированных компонентов объекта или контроля. BLUE (наилучшие линейные объективные оценки) получали для каждого объекта и контроля для всех фоновых генотипов гибридов для каждого местоположения. Попарные сопоставления BLUE объекта с BLUE контроля проводили для проверки значимых различий (BLUE DIFF) при P<0,05 как для DP-202216-6, так и для Объекта 18.
Эти результаты демонстрируют, что экспрессия рекомбинантного полинуклеотида, кодирующего SEQ ID NO: 1, повышает урожайность зерна маиса в полевых условиях.
Пример 4
Ген AG099, строение конструкции для получения объекта DP-202216-6
Маис (Zea mays L.) трансформировали путем трансформации, опосредованной Agrobacterium, с помощью вектора/плазмиды для трансформации растений. Область Т-ДНК этой плазмиды схематично представлена на фиг. 1, и последовательность представлена как SEQ ID NO: 5, а вставка представлена как SEQ ID NO: 6. Сводный обзор генетических элементов и их положений на Т-ДНК приведен в таблицах 6-7.
Т-ДНК трансформирующей конструкции содержит две генные кассеты. Первая кассета (кассета гена AG099) содержит AG099, кодирующий белок AG099. Белок из 251 остатка, полученный путем экспрессии последовательности AG099, имеет примерную молекулярную массу 28 кДа. Экспрессия гена AG099 контролируется промотором гена gos2 фактора инициации трансляции из Zea mays (zm-gos2) вместе с областью интрона 1 гена убиквитина 1 маиса (ubiZM1). Транскрипция кассеты гена AG099 завершается присутствием терминаторной последовательности из гена ингибитора протеиназы II (pinII) из Solanum tuberosum.
Вторая генная кассета (кассета гена mo-pat) содержит ген фосфинотрицинацетилтрансферазы (mo-pat) из Streptomyces viridochromogenes. Ген mo-pat экспрессирует фермент фосфинотрицинацетилтрансферазу (PAT), который придает переносимость фосфинотрицина. Белок PAT состоит из 183 аминокислот в длину и имеет примерную молекулярную массу 21 кДа. Экспрессия гена mo-pat контролируется промотором ubiZM1, 5'-UTR и интроном в сочетании со второй копией терминатора pinII.
Таблица 6. Описание генетических элементов в области Т-ДНК плазмиды PHP40099
(Положение пар оснований)
5’-некодирующая область в п.н. 2749-2808 (длиной 60 п. н.); кодирующая последовательность в п.н. 2809-3564 (длиной 756 п. н.); 3’- некодирующая область в п.н. 3565-3605 (длиной 41 п. н.)
Таблица 7. Описание генетических элементов в области Т-ДНК плазмиды PHP40099 (продолжение)
Пример 5
Расщепление DP-202216-6 маиса в двух поколениях
Отдельные поколения (T2 и F1*1) маиса DP-202216-6 выращивали в 4-дюймовых горшках, объединенных на платформах, содержащих по 15 горшков, с использованием типичных условий тепличного производства. Для каждого поколения посадили до 165 семян. После прорастания, но перед взятием образцов листьев, растения маиса прореживали до 100 здоровых растений. Когда растения находились примерно на стадии роста V3 (т.е. когда становится виден узел третьего листа), образцы, взятые путем пробивания листа, собирали у 100 растений. Образцы листьев анализировали с использованием анализа ПЦР в реальном времени в отношении наличия или отсутствия объекта DP-2∅2216-6 и генов AG099 и mo-pat.
ПЦР-амплификация уникальных областей внутри введенных генетических элементов может отличить тестируемые растения от их генетически не модифицированных аналогов и может использоваться для скрининга в отношении наличия встроенной области Т-ДНК плазмиды, содержащей AG099. Для обнаружения генов AG099 и mo-pat, содержащихся во вставке Т-ДНК, а также геномного 5’-соединения, охватывающего сайт вставки маиса DP-202216-6, области от 76 п.н. до 105 п.н. амплифицировали при помощи праймеров и зондов, специфичных для каждой уникальной последовательности. Кроме того, ампликон эндогенного эталонного гена длиной 79 п.н., High Mobility Group A (hmg-A), использовали в дуплексе с каждым анализом как для качественной, так и для количественной оценки каждого анализа и для демонстрации наличия достаточного качества и количества ДНК в реакции ПЦР. Данные для hmg-A использовали при расчетах относительно оценки. Данные сравнивали с характеристиками подтвержденного отрицательного геномного контроля. Результаты ПЦР оценивали на предмет правильного расщепления. Ожидается, что популяция поколения T2 будет расщепляться в соотношении 3:1, а популяция поколения F1*1, как ожидается, будет расщепляться в соотношении 1:1, в соответствии с менделевскими правилами наследования. Растение с положительными результатами ПЦР для соответствующего анализа считалось положительным растением, а растение с отрицательными результатами ПЦР считалось отрицательным растением. Общее количество положительных и отрицательных растений для каждого поколения представлено в таблице 8. Результаты ПЦР для всех поколений показывают, что Т-ДНК AG099 встраивалась в хромосому с получением объекта DP-202216-6 в геноме маиса.
Таблица 8. Сводный обзор результатов расщепления DP202216 маиса в двух поколениях
b Степени свободы=1. Значение хи-квадрат более 3,84 (значение P менее 0,05) указывает на значимую разницу.
Пример 6
Определение числа копий/зиготности Объекта 16
ПЦР-анализ числа копий и анализ секвенирования следующего поколения (NGS) использовали для демонстрации того, что в маисе DP-202216-6 (Объект 16) произошла единственная вставка, и что Т-ДНК стабильно передается из поколения в поколение, и определяли последовательности соединений с использованием анализа секвенирования. ПЦР-анализ числа копий проводили для двух поколений (T2 и F1*1), а применение NGS, называемого Southern-by-Sequencing (SbS), проводили на поколении T1 маиса DP202216.
Экстракты геномной ДНК из тканей листа отдельных растений получали из маиса DP202216, которую получали путем трансформации линии маиса плазмидой PHP40099. Для анализа SbS использовали восемь растений маиса DP-202216-6 поколения Т1. Кроме того, для анализа SbS получали геномную ДНК из контрольной линии маиса (использованной для трансформации).
Анализ Southern-by-Sequencing
В SbS используется захват последовательности на основе зондов, методики секвенирования следующего поколения (NGS) и биоинформационные процедуры для выделения, секвенирования и идентификации вставленной ДНК в геноме маиса. Путем составления большого количества уникальных считываний секвенирования и сравнения их с трансформационной плазмидой в биоинформационном анализе идентифицируются уникальные соединения, обусловленные вставленной ДНК, и они используются для определения числа вставок в геном растения. Восемь растений маиса DP202216 поколения T1 проанализировали с помощью SbS для определения числа копий вставки в каждом растении. Шесть растений содержали вставку ДНК DP202216, как показал специфический для объекта анализ ПЦР; оставшиеся два растения оказались отрицательными в отношении вставки согласно тому же анализу. Кроме того, аналогичным образом с помощью SbS анализировали контрольную ДНК маиса.
Зонды захвата, используемые для отбора последовательностей плазмиды PHP40099, конструировали и синтезировали коммерчески доступным способом. Серию уникальных последовательностей, охватывающих последовательность плазмиды, использовали для создания биотинилированных олигонуклеотидов (70-74 п.н.) в качестве зондов захвата.
Библиотеки секвенирования следующего поколения конструировали с использованием геномной ДНК из отдельных растений маиса DP202216 и контрольной линии маиса. Геномную ДНК случайным образом разрезали до длины примерно 400 п.н. и к концам лигировали адаптеры для секвенирования. SbS выполняли на растениях DP202216 T1, по сути, как описано в Zastrow-Hayes, et al. Southern-by-Sequencing: A Robust Screening Approach for Molecular Characterization of Genetically Modified Crops. The Plant Genome 8: 1-15 (2015), включенной в данный документ посредством ссылки в той степени, в которой способы, описанные в этой ссылке, применимы к анализу, раскрытому в данном документе. Коротко, библиотеки секвенирования от каждого растения гибридизировали с зондами захвата посредством двух раундов гибридизации для обогащения целевых последовательностей. После NGS на коммерчески доступной платформе HiSeq 2500 (Illumina, Сан-Диего, штат Калифорния, США) считывания секвенирования поступали в биоинформационный конвейер для обрезки и контроля качества. Считывания сравнивали как с геномом маиса, так и с конструкцией для трансформации, и считывания, которые содержали как геномную, так и плазмидную последовательность, идентифицировали как считывания соединений. Выравнивание считываний соединений с конструкцией для трансформации показало границы вставленной ДНК относительно ожидаемой вставки.
Результаты Southern-by-Sequencing
Интеграцию и число копий вставки определяли в маисе DP202216-6, полученной с конструкцией PHP40099. Т-ДНК из PHP40099, которую переносили в объект маиса DP-2∅2216-6, представлена на фиг. 1. SbS использовали для оценки вставки в маис DP-2∅2216-6. В SbS использовали зонды захвата к трансформационной плазмиде для выделения геномной ДНК, которая гибридизировалась с последовательностями зондов. Захваченную ДНК затем секвенировали с использованием процедуры NGS и анализировали с использованием биоинформационных инструментов. Во время анализа считывания последовательностей, которые показали частичную идентичность последовательности плазмидной ДНК, в то время как остальная часть считывания не соответствовала смежной плазмидной последовательности, идентифицировали как соединения между встроенной ДНК и геномной ДНК или между вставками двух последовательностей плазмидной ДНК, которые не были смежными в исходной плазмиде. Для каждого соединения получали несколько считываний последовательностей, и эти считывания компилировали в консенсусную последовательность для соединения. Уникальное соединение определили как соединение, в котором последовательность, полученная из плазмиды, и соседняя последовательность были одинаковыми, хотя общая длина множества считываний для этого соединения варьировалась из-за процесса секвенирования. Число уникальных соединений было связано с числом вставок плазмиды, присутствующих в геноме (например, ожидалось, что одна вставка Т-ДНК будет иметь два уникальных соединения). Обнаружение дополнительных уникальных соединений помимо двух, ожидаемых для одной вставки (если таковая имеется), указывает на присутствие дополнительных плазмидных вставок. Две или более вставки могут быть либо генетически связаны, либо не связаны друг с другом. Для трансформированных линий, содержащих более одной вставки, анализ нескольких растений из одного поколения предоставит информацию о статусе расщепления нескольких вставок, поскольку тесно связанные вставки будут обнаружены в одном и том же отдельном растении, в то время как несвязанные вставки будут расщепляться в популяции случайным образом. SbS также предоставил информацию о последовательности относительно геномного контекста вставки, которую можно использовать для идентификации местоположения в хромосоме, если была доступна достаточная последовательность нетрансформированного генома растения. Каждое из растений, которые были положительными в отношении вставки ДНК DP202216, приводило к одним и тем же двум уникальным соединениям, которые были одинаковыми для всех шести растений. 5’-Соединение для всех растений начиналось с 23 п.н. Т-ДНК PHP40099, а заканчивалась вставка в 3’-соединении на 7458 п.н. Т-ДНК (фиг. 1). Как сообщалось ранее, делеции концов правого пограничного района и левого пограничного района могут часто возникать при трансформации, опосредованной Agrobacterium. Эти местоположения были идентичными у всех шести растений, что указывает на то, что ДНК-вставка DP-202216-6 является последовательной и стабильной для поколения T1 маиса DP-202216-6. Определили результаты SbS для каждого растения, включая число уникальных считываний в 5’- и 3’-соединениях для каждого растения (таблица 9). Никаких других соединений между последовательностями PHP40099 и геномом маиса, обнаруженными в этих шести растениях, не было, что указывает на то, что результаты SbS показали отсутствие дополнительных плазмидных вставок в маисе DP-202216-6. Кроме того, не было выявлено соединений между несмежными областями Т-ДНК PHP40099, что указывает на отсутствие обнаруживаемых перестроек или усечений во вставленной ДНК, кроме усечений правого и левого пограничных районов, указанных выше. В то время как охват эндогенных элементов маиса в их естественном контексте был обнаружен в двух отрицательных растениях, никаких соединений между последовательностями PHP40099 и геномом маиса не было обнаружено ни у двух отрицательных (для объекта DP-202216-6) растений из расщепляющейся популяции поколения T1, ни в контрольной линии, указывая на то, что, как и ожидалось, эти растения не содержали каких-либо вставок, происходящих от PHP40099. Кроме того, не было соединений между геномными последовательностями маиса и каркасной последовательностью PHP40099 ни в одном из проанализированных растений, свидетельствуя о том, что каркасные последовательности плазмиды не были включены в маис DP202216.
Анализ SbS поколения T1 маиса DP-202216-6 показал, что существует одна копия Т-ДНК PHP40099 в маисе DP-202216-6 и что в ее геноме нет дополнительных вставок.
Таблица 9. Анализ SbS растений маиса DP202216
b Уникальные считывания подтверждают расположение 5’-геномного соединения ДНК-вставки DP202216 в 23 п.н. Т-ДНК PHP40099. Множество идентичных считываний NGS конденсировали в каждое уникальное считывание.
c Уникальные считывания подтверждают местоположение 3’-геномного соединения ДНК-вставки DP202216 в 7458 п.н. Т-ДНК PHP40099. Множество идентичных считываний NGS конденсировали в каждое уникальное считывание.
Пример 7
Специфические для объекта способы обнаружения, праймеры и зонды
Для обнаружения генов AG099 и mo-pat, содержащихся в объекте маиса DP-2∅2216-6, а также геномного соединения, охватывающего сайт вставки маиса DP-202216-6, области от приблизительно 76 п.н. до 105 п.н. амплифицировали при помощи праймеров и зондов Taqman®в, специфических для каждой уникальной последовательности. Кроме того, область эндогенного эталонного гена длиной 79 п.н., High Mobility Group A (hmg-A, номер доступа в GenBank AF171874.1) утвердили для использования в дуплексе с каждым анализом как для качественной, так и для количественной оценки каждого анализа и для демонстрации наличия достаточного качества и количества ДНК в реакции ПЦР. Данные для hmg-A использовали при расчетах относительно оценки. Данные сравнивали с характеристиками аттестованного положительного калибратора или калибратора числа копий, а также отрицательных геномных контролей.
Реакция ПЦР в реальном времени включает 5’-нуклеазную активность активируемой нагреванием ДНК-полимеразы. Два праймера и один зонд подвергали отжигу с целевой ДНК с зондом, который содержал 5’-флуоресцентный репортерный краситель и 3’-гасящий краситель. В каждом цикле ПЦР репортерный краситель отщепляется от подвергнутого отжигу зонда полимеразой, испуская флуоресцентный сигнал, который усиливается с каждым последующим циклом. Цикл, при котором интенсивность излучения ампликона образца поднимается выше порога обнаружения, называется значением CT. Когда не происходило амплификации, прибор не вычислял CT и присваивал значение CT 40,00.
Образцы определяли как положительные или отрицательные для конкретного представляющего интерес гена с применением приведенных ниже критериев.
Положительный:
CT эндогенного гена <35
CT представляющего интерес гена (GOI) <35
ΔCT (дельта CT) (CT эндогенного гена - CT GOI) >-5
Отрицательный:
CT эндогенного гена <35
CT представляющего интерес гена (GOI) >35
ΔCT (CT эндогенного гена - CT GOI) < и >-5
Если нужно было определить число копий тестовых образцов, калибраторы числа копий (образцы, о которых известно, что они содержат определенные копии представляющего интерес гена, например 1 или 2 копии) использовали в качестве контролей как для эндогенного, так и для представляющего интерес гена. ΔCT рассчитывали для тестовых образцов и калибраторов числа копий, как описано выше. ΔΔCT (дельта дельта CT) использовали для статистического расчета числа копий (ΔΔCT=ΔCT калибратора числа копий - ΔCT образца GOI). Допуски алгоритма использовали для заявления количества копий для каждого тестового образца. Число копий, равное 1, применяли к выборке, дающей аналогичное среднее значение ΔCT по сравнению с калибраторами с одной копией, или когда ΔΔCT0,7-1,0 получили с калибратором с 2 копиями. Аналогичным образом, число копий 2 применяли к выборке, дающей ΔΔCT в диапазоне 0,7-1,0 по сравнению с калибраторами для одной копии; и число копий 3 применяли к выборке, дающей ΔΔCT примерно 0,5 по сравнению с популяцией с 2 копиями. Статистический алгоритм также применял вероятности присвоения каждого потенциального числа копий на основе присвоенных значений ΔΔCT после анализа. Любые значения ΔΔCT, выходящие за пределы ожидаемых диапазонов, приведут к результатам числа копий с более низкой степенью вероятностей, тогда как значения ΔΔCT в пределах ожидаемых диапазонов будут давать результаты с высокой степенью вероятностей.
Выделение ДНК
Образцы геномной ДНК, выделенные из ткани листа 200 растений, представляющих поколения T2 и F1 маиса DP202216, экстрагировали с использованием щелочного буфера, состоящего из гидроксида натрия, дигидрата динатриевой соли этилендиаминтетрауксусной кислоты (Na2-EDTA) и Трис. Примерно 3 нг матричной ДНК использовали на реакцию, независимо от общего реакционного объема.
Подробная информация о составе и получении реакционных смесей
Каждый анализ, подтверждающий сайт вставки маиса DP202216 и гены AG099 и mo-PAT, содержащиеся в объекте DP-2∅2216-6, объединяли с эндогенным эталонным анализом hmg-A. Реакционные смеси включали все соответствующие компоненты для подтверждения как представляющего интерес гена, так и эндогенного гена в ПЦР-реакции. Применяли базовый мастер-микс, мастер-микс Bioline SensiFast™ Probe Lo-ROX (коммерчески доступный) с 30% бычьим сывороточным альбумином (BSA), включенным в качестве добавки. Отдельные концентрации каждого праймера варьировались в зависимости от реакции от 300 нМ до 900 нМ в зависимости от оптимальной концентрации, установленной во время валидации анализа. Отдельные концентрации каждого зонда на реакцию составляли 80 нМ. Контрольные образцы для анализов включали контроли без матрицы (NTC), которые состояли из воды или буфера Трис-EDTA (ТЕ) (10 мМ Трис pH 8,0, 1 мМ EDTA), а также калибратор числа копий и отрицательные контроли, все из которых валидировали для каждого проведенного анализа. Температуры отжига и количество циклов, используемое во время анализа ПЦР, приведены в таблице 10. Праймер и зонды, используемые для анализа ПЦР, представлены в таблицах 11 и 12. Составы мастер-миксов для каждого анализа ПЦР представлены в таблицах 13-15.
Параметры ПЦР
Параметры ПЦР, используемые во время анализа ПЦР, перечислены ниже.
Таблица 10. Температуры отжига и циклы, используемые во время ПЦР-реакции
(°C)
(секунды)
b Если термоциклирование выполняли с использованием Roche LightCycler®480, стадии 2a и 2b выполняли за 45 циклов.
Праймеры и зонды
Праймеры и зонд, используемые для каждого проведенного анализа, перечислены ниже.
Таблица 11. Праймеры и зонды для анализа ПЦР генов AG099 и mo-PAT и сайта вставки маиса DP202216
(п. н.)
CCATCTGAGGTCTGCACTCTCACCGGTAGTACAGCACAAACAACACACTCAAACACTGATAGTTTAAACTGAAGGCGGGAAACGACAATCTGATCATGAGCGGAG (SEQ ID NO: 24)
GGATGCTCCGCACTGTCAAGTAACAGGTGAGGTCTTCCCAGTGTAGTTTTGCAGCTGATCTCGAAAGGGTGGGCGCGCCGACCCAGCTTTCTT (SEQ ID NO: 25)
CATCGTGAACCACTACATCGAGACCTCCACCGTGAACTTCCGCACCGAGCCGCAGACCCCGCAGGAGTGGATCGAC (SEQ ID NO: 26)
Таблица 12. Праймеры и зонды для анализа ПЦР эндогенного эталонного гена hmg-A
TTGGACTAGAAATCTCGTGCTGATTAATTGTTTTACGCGTGCGTTTGTGTGGATTGTAGGACAAGGCTCCCTATGTAGC (SEQ ID NO: 30)
Подготовка мастер-микса
Компоненты и концентрации, обеспечивающие каждый мастер-микс, перечислены ниже.
Таблица 13. Мастер-микс для обеспечения объединенного анализа: DP-2∅2216-6 и hmg-Aa
b Концентрация раствора бычьего сывороточного альбумина в реакционной смеси в качестве реагента составляла 0,3%; концентрация в расчете на исходную составляла 0,08%.
c N_A эквивалентно выражению "неприменимо".
Таблица 14. Мастер-микс для обеспечения объединенного анализа: AG099 и hmg-Aa
b Концентрация раствора бычьего сывороточного альбумина в реакционной смеси в качестве реагента составляла 0,3%; концентрация в расчете на исходную составляла 0,08%.
c N_A эквивалентно выражению "неприменимо".
Таблица 15. Мастер-микс для обеспечения объединенного анализа: mo-PAT и hmg-Aa
b Концентрация раствора бычьего сывороточного альбумина в реакционной смеси в качестве реагента составляла 0,3%; концентрация в расчете на исходную составляла 0,08%.
c N_A эквивалентно выражению "неприменимо".
Анализ ПЦР
Образцы геномной ДНК, выделенные из собранных образцов листьев растений маиса DP202216, вместе с калибратором числа копий, отрицательным контролем и контролем NTC, подвергали амплификации qPCR с использованием мастер-микса Lo-ROX зонда SensiFast™ (Bioline, Лондон, Великобритания) в присутствии пары праймеров и зондов, специфических для генов mo-PAT и AG099, и сайта вставки, специфического для маиса DP202216, которые позволяют однозначно идентифицировать вставку Т-ДНК PHP40099 в маис DP202216. Для анализа и мониторинга качества ДНК hmg-A маиса включали в дуплексе с каждой реакцией в качестве эндогенного контроля. Каждую реакцию qPCR устанавливали в общем объеме 3 мкл (маиса DP202216 и AG099) или 6 мкл (mo-PAT) с 3 нг выделенной геномной ДНК.
Результаты
Результаты анализов числа копий qPCR для нескольких поколений указывают на стабильную интеграцию и расщепление одной копии генов в Т-ДНК плазмиды PHP40099 с продемонстрированным переносом в последующие поколения.
Продукты ПЦР от 76 п.н. до 105 п.н., представляющие сайт вставки/соединение для маиса DP202216, а также гены AG099 и mo-PAT в Т-ДНК плазмиды PHP40099, амплифицировали и наблюдали в образцах листьев маиса DP202216, а также восьми геномных контролей калибраторов числа копий, но они отсутствовали в каждом из восьми отрицательных геномных контролей и восьми контролей NTC. Каждый анализ выполняли по меньшей мере четыре раза, каждый раз получая одинаковые результаты. Для каждого образца и всех контролей рассчитывали значения CT, значения ΔCT и число копий.
Используя эндогенный эталонный ген hmg-A маиса, продукт ПЦР из 79 п.н. амплифицировали и наблюдали в образцах листьев маиса DP202216, а также в восьми калибраторах числа копий и в восьми отрицательных геномных контролях. Амплификацию эндогенного гена не наблюдали в восьми протестированных контролях без матрицы (NTC). Анализ выполняли по меньшей мере четыре раза, каждый раз получая одинаковые результаты. Для каждого образца и всех контролей рассчитывали значения CT, значения ΔCT и число копий (если применимо).
Чувствительность специфических для конструкции анализов ПЦР для маиса DP202216
Для оценки чувствительности анализов ПЦР, специфических для конструкции, ДНК маиса DP202216 разбавляли контрольной геномной ДНК маиса, в результате чего получали тестируемые образцы, содержащие различные количества ДНК DP202216 маиса (5 нг, 1 нг, 100 пг, 50 пг, 20 пг, 10 пг, 5 пг, 1 пг, 0,5 пг, 0,1 пг) в общих 5 нг ДНК маиса. Эти различные количества ДНК маиса DP202216 соответствуют 100%, 20%, 2%, 1%, 0,4%, 0,2%, 0,1%, 0,01% и 0,002% ДНК маиса DP202216 в общей геномной ДНК маиса соответственно. Различные количества ДНК DP202216 маиса подвергали амплификации ПЦР в реальном времени для генов AG099 и mo-PAT. На основании этих анализов предел обнаружения (LOD) определили как приблизительно 5 пг ДНК маиса DP202216 в 5 нг общей ДНК для mo-PAT, или 0,1%, и 10 пг ДНК маиса DP202216 в 5 нг общей ДНК для AG099, или 0,2%. Определенная чувствительность каждого описанного анализа достаточна для многих скрининговых путей применения. Каждую концентрацию тестировали в общей сложности четыре раза, каждый раз получая одинаковые результаты. Анализ ПЦР в режиме реального времени для маиса DP202216 с использованием анализов, специфичных для объектов и конструкций, подтверждает стабильную интеграцию и расщепление одной копии Т-ДНК плазмиды PHP40099 в тестируемых образцах листьев, что продемонстрировано с помощью количественного определения объекта DP-2∅2216-6 и генов AG099 и mo-PAT в маисе DP-202216-6. Эти результаты были воспроизведены во всех проведенных повторных анализах qPCR. Анализ эндогенного эталонного гена маиса для обнаружения hmg-A амплифицировали, как ожидалось, во всех тестовых образцах, отрицательных контролях и не обнаруживали в образцах NTC. Чувствительность способов ПЦР в предоставленных условиях демонстрирует, что эти анализы могут обнаруживать приблизительно 5 пг, или 0,1%, ДНК маиса DP202216 в общих 5 нг геномной ДНК маиса для mo-PAT и примерно 10 пг, или 0,2%, ДНК маиса DP202216 в общих 5 нг геномной ДНК маиса для AG099.
Пример 8
Расчеты экспрессии и концентрации белка AG099
Выделение белка
Аликвоты обработанных образцов ткани листа отвешивали в пробирки объемом 1,2 мл до целевой массы 10 мг. Каждый образец, анализируемый на концентрацию белка AG099, экстрагировали в охлажденном 0,25% ASB-14 в фосфатно-солевом буфере, содержащем полисорбат 20 (PBST), и каждый образец, анализируемый на концентрацию белка PAT, экстрагировали в 0,6 мл охлажденного PBST. После центрифугирования супернатанты удаляли, разбавляли 0,25% ASB-14 в PBST (AG099) или PBST (PAT) и анализировали.
Определение концентрации белка AG099
В способе ELISA AG099 использовали ELISA, произведенный Pioneer Hi-Bred International, Inc. для определения концентрации белка AG099 в образцах. Стандарты (обычно анализируемые в лунках с тремя повторами) и разбавленные образцы (обычно анализируемые в лунках с двумя повторами) инкубировали в планшете, предварительно покрытом антителом к AG099. После инкубации несвязанные вещества смывали с планшета. Другое антитело к AG099, конъюгированное с ферментом пероксидазой хрена (HRP), добавляли в планшет и инкубировали. Несвязанные вещества смывали с планшета. Обнаружение связанного комплекса AG099-антитело осуществляли путем добавления субстрата, который вырабатывал окрашенный продукт в присутствии HRP. Реакцию останавливали кислым раствором и определяли оптическую плотность (OD) каждой лунки с помощью планшет-ридера.
Определение концентрации белка PAT
В способе ELISA PAT использовали набор ELISA, произведенный EnviroLogix Inc. для определения концентрации белка PAT в образцах. Стандарты (обычно анализируемые в лунках с тремя повторами) и разбавленные образцы (обычно анализируемые в лунках с двумя повторами) совместно инкубировали с антителом, специфическим для PAT, конъюгированным с ферментом HRP, в планшете, предварительно покрытом другим специфическим для PAT антителом. После инкубации несвязанные вещества смывали с планшета. Обнаружение связанного комплекса PAT-антитело осуществляли путем добавления субстрата, который вырабатывал окрашенный продукт в присутствии HRP. Реакцию останавливали кислым раствором и определяли OD каждой лунки с помощью планшет-ридера.
Расчеты для определения концентрации белков
Программное обеспечение для обработки данных микропланшетов SoftMax Pro GxP (Molecular Devices) использовали для выполнения расчетов, необходимых для преобразования значений OD, полученных для каждого набора лунок с образцами, в значение концентрации белка.
Стандартная кривая была включена в каждый планшет для ELISA. Уравнение для стандартной кривой получали с помощью программного обеспечения, которое использовало квадратичную аппроксимацию для соотнесения значений OD, полученных для каждого набора лунок со стандартом, с соответствующей концентрацией стандарта (нг/мл).
Уравнение квадратичной регрессии применялось следующим образом: y=Cx2+Bx+A,
где x=известная стандартная концентрация, и y=соответствующее значение абсорбции (OD).
Интерполяцию концентрации образца (нг/мл) выполняли путем решения для x в приведенном выше уравнении с использованием значений для A, B и C, которые были определены для стандартной кривой.
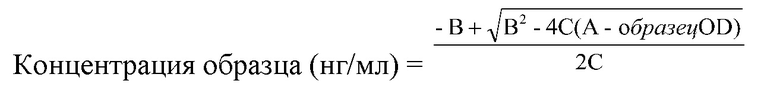
Например, для заданных параметров кривой A=0,0476, B=0,4556, C= -0,01910 и в образце
OD=1,438
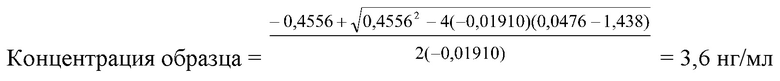
Значения концентрации образца скорректировали с учетом коэффициента разбавления, выраженного как 1:N, путем умножения интерполированной концентрации на N.
Скорректированная концентрация=интерполированная концентрация образца x коэффициент разбавления
Например, учитывая интерполированную концентрацию 3,6 нг/мл и коэффициент разбавления 1:20
Скорректированная концентрация = 3,6 нг/мл x 20 = 72 нг/мл
Скорректированные значения концентрации образца, полученные с помощью программного обеспечения SoftMax Pro GxP, преобразовывали из нг/мл в нг/мг массы образца следующим образом:

Например, концентрация образца=72 нг/мл, объем буфера для экстракции=0,60 мл и целевая масса образца=10 мг
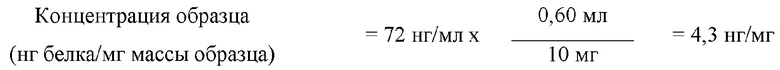
Фиксируемый нижний предел количественного определения (LLOQ) анализа в нг/мл рассчитывали следующим образом:
Фиксируемый LLOQ анализа (нг/мл) = (самая низкая концентрация стандарта - 10%) × минимальное разведение
Например, самый низкий стандарт=0,50 нг/мл и минимальное разведение=10
Фиксируемый LLOQ анализа (нг/мл) = (0,50 нг/мг - (0,50×0,10)) × 10=4,5 нг/мл
LLOQ в нг/мг массы образца рассчитывали следующим образом:

Например, фиксируемый LLOQ анализа=4,5 нг/мл, объем буфера для экстракции=0,60 мл и целевая масса образца=10 мг

Средние значения, стандартные отклонения и диапазоны для белков AG099 и PAT для маиса DP202216 с 1 копией и маиса DP202216 с нулевой копией представлены в таблицах 16 и 17 соответственно.
Таблица 16. Концентрации экспрессированного белка AG099 в образцах листьев
(Стадия роста)
Общее количество фиксируемых образцов
Таблица 17. Концентрации экспрессированного белка PAT в образцах листьев
(Стадия роста)
Общее количество фиксируемых образцов
Пример 9
Трансформация маиса с помощью трансформации Agrobacterium и регенерация трансгенных растений, содержащих объект DP-202216-6
Объект DP-202216-6 маиса получали путем опосредованной Agrobacterium трансформации плазмидой PHP40099. В этом протоколе для создания трансгенных растений маиса использовали сконструированные Agrobacterium tumefaciens и незрелые зародыши трансформируемой линии маиса с использованием moPAT в качестве селектируемого маркера. Коротко, штамм Agrobacterium tumefaciens (JTLBA4404), содержащий описанную выше плазмиду, получили с использованием стандартных условий роста, включая инкубирование бактерий в темноте при 28°C перед использованием для агроинфекции. Незрелые зародыши собирали из початков трансформируемой линии маиса приблизительно через 8-11 дней после опыления, при этом размер зародыша составлял приблизительно от 1,3 до 1,8 мм в длину. Культуру клеток Agrobacterium использовали для заражения выделенных незрелых зародышей путем добавления культуры клеток Agrobacterium в пробирки, содержащие зародыши, и культивирования зародышей щитком вверх в среде для совместного культивирования при 21°C в течение приблизительно 3 дней в темноте. Через 3 дня такие культивируемые зародыши переносили в питательную среду, содержащую карбенициллин для контроля роста Agrobacterium, и культивировали при 28°C в темноте в течение приблизительно 7 дней с последующим переносом в другую питательную среду и культивацией при 28°C в темноте в течение приблизительно 21 дня. Каллус зародыша переносили в питательную среду, содержащую биалафос, и культивировали дополнительно при 28°C в темноте с переносом в свежую среду с 14-дневным интервалом. Каллус переносили в среду для созревания зародышей приблизительно на 14 дней при 28°C в темноте. Зрелые зародыши переносили в среду для проращивания и помещали на свет при 28°C приблизительно на 7 дней. После развития побегов и корней проростки переносили в пробирки для роста на свету при 28°C в течение приблизительно 7 дней. После развития побегов проростки отправляли в теплицу для получения семян Т1 и дальнейшего анализа.
Пример 10
Экспрессия трансгена в DP-2∅2216-6 маиса
По сравнению с экспрессией zmm28 в контрольных растениях в объекте 16 экспрессия zmm28 наблюдается на ранних стадиях роста (V2-V5) в листе из-за присутствия трансгенного zmm28, а затем имеет профиль экспрессии, аналогичный профилю экспрессии, наблюдаемому для контрольной маиса от V6 до старения (фиг. 2d и e). Экспрессия zmm28 также обнаруживается в корне, стебле, апикальной меристеме побегов (SAM), метелке, початке и ядре объектов маиса. Концентрации белка ZMM28 в листе в этих двух объектах соответствуют временному профилю, аналогичному таковому для уровней РНК, и возникают на уровнях ниже ppm. Однако нет сильной корреляционной связи между уровнем транскрипта и экспрессией белка в корне, как измерено в этом исследовании.
Эксперименты по субклеточной локализации трансгенного белка ZmGos2-zmm28 с N-концевой или С-концевой меткой зеленого флуоресцентного белка Aequorea coerulescens (AcGFP1) указали на локализацию в ядре в независимых стабильно трансформированных объектах маиса. Этот результат был подтвержден во временных анализах, включающих ядерный маркерный белок, гистон H2B (GRMZM2G401147), слитый с mKate2.
Пример 11
Продленная и повышенная экспрессия AG099 и агрономическая характеристика
В объектах ZmGos2-zmm28 провели серию морфометрических измерений. Продленная и повышенная экспрессия гена zmm28 приводила к наблюдаемому раннему росту проростков, биомассы листьев и общей площади листьев. Количество образцов, протестированных для V2 - V8, составляет 96 и приблизительно 48 для V10. Сухая масса листьев на стадии V8 из приблизительно 48 образцов. Общая площадь листьев на стадии R1 была из 24 образцов. Скорость обмена углерода в листе на стадии V11 измеряли на 60 образцах. Скорость фотосинтетического переноса электронов на V11 определяли на приблизительно 60 образцах; поглощение N на приблизительно 31 для контроля и 29 для DP202216 и DP382118. Содержание ассимилированного N в листьях и корнях, n=19 для контроля и DP382118 и n=20 для DP202216.
Высота растений DP202216 и DP382118 была значительно больше, чем высота контрольных растений от V2 до V7, усредненная по всем протестированным гибридам. Сухая масса листа увеличилась на 11% и 22% для DP202216 и DP382118 соответственно на стадии V8. Более того, оба объекта ZmGos2-zmm28 имели большую общую площадь листьев, чем WT, увеличиваясь в среднем на 4% на стадии R1.
Модулирование экспрессии zmm28 в маисе приводило к улучшению определенных измеренных вегетативных фенотипов. К ним относилось увеличение ранней мощности проростков, измеряемое как увеличение высоты растения и биомассы листьев, а также увеличение общей площади листьев на стадии R1. Высоту растений измеряли от поверхности почвы до узла самого молодого полностью распустившегося листа. Для измерения сухой массы листьев все листья, собранные с каждого отдельного растения, помещали в бумажный пакет и сушили при 70°C в течение 72 часов или до полного высыхания. Образцы взвешивали после выдерживания при комнатной температуре в течение 1 часа. Общую площадь листьев измеряли у растений, выращенных на полевых делянках. Коротко, растения собирали с полевой делянки путем срезания стебля на поверхности почвы на стадии роста R1, затем вырезали все листья отдельного растения и измеряли площадь каждого листа с помощью измерителя площади листа Li-3100C (Li-Cor, Линкольн, штат Небраска, США).
Пример 12
Повышенный фотосинтез и активность ферментов фотосинтеза
Фотосинтез определяли, чтобы увидеть, был ли изменен этот характерный признак в трансгенных объектах. Фотосинтез, выраженный как скорость обмена CO2 (CER) и скорость фотосинтетического переноса электронов (ETR), измеряли у выращенных в полевых горшках растений DP202216 и DP382118 с двумя фоновыми генотипами гибридов вместе с их контролями на стадии роста V11. CER и ETR в DP-202216-6 увеличивались на 10% и 8% соответственно; тогда как CER и ETR в DP382118 увеличивались на 10% и 9% соответственно. Повышенная и продленная экспрессия zmm28 в маисе приводила к увеличению площади зеленых листьев как на вегетативной, так и на репродуктивной стадиях, и к увеличению скорости фотосинтеза на площадь листа. Это подтверждается наблюдаемым увеличением как ETR, так и ферментативной активности ключевых фотосинтетических ферментов цикла C4.
Продленная и повышенная экспрессия zmm28 приводила к увеличенным значениям активности фотосинтетических ферментов C4 и активности нитратредуктазы, но не активности глутаминсинтазы. Исследования газообмена показали увеличение скорости фиксации CO2 как у DP202216, так и у DP382118. Чтобы определить, изменились ли значения активности релевантных для фотосинтеза ферментов в этих объектах, исследовали значения специфической активности основных фотосинтетических ферментов C4 на двух стадиях роста, V4 и V11. Эти две стадии роста выбирали потому, что в V4 экспрессируется только трансгенный zmm28 (как измерено), в то время как в V11 экспрессируются как нативный, так и трансгенный zmm28 (поддается обнаружению). Статистически значимое увеличение значений активности наблюдали в одном или обоих трансгенных объектах для PEPC и NADP-MDH в V4 и для PEPC, NADP-MDH и PPDK в V11. Наиболее устойчивый эффект наблюдали для NADP-MDH, причем оба объекта в V11 и объект DP382118 в V4 характеризовались значимым увеличением специфической ферментативной активности. Это увеличение значений активности фотосинтетических ферментов C4 согласуется с увеличением скорости фиксации CO2.
Нитратредуктаза (NR) катализирует скорость-лимитирующую стадию ассимиляции нитрата, инициируя восстановление нитрата до органических форм, и этот фермент хорошо известен как ключевой регулятор ассимиляции и усвоения азота. Специфическую активность NR оценивали в тканях листьев и корней объектов DP202216, DP382118 маиса и контрольных растений на стадиях роста V4 и V11. Из результатов видно, что ферментативная активность NR была значительно увеличена в DP202216 и DP382118 в ткани листьев на обеих стадиях роста. Увеличение активности NR согласуется с результатами поглощения и ассимиляции N. Однако не было значительных различий в активности NR в ткани корня между двумя объектами и контрольными линиями.
Специфическую активность другого ключевого фермента ассимиляции N-глутаминсинтетазы (GS) - также исследовали в тех же образцах ткани, которые использовали для анализа NR. Значения активности GS не показали значительных различий между DP202216 и DP382118 и контролями ни в листе, ни в корне. Это может указывать на то, что для повышенной ассимиляции N, измеренной в DP202216 и DP382118, не требуется дополнительной активности GS, учитывая продленную и повышенную экспрессию zmm28 в трансгенных растениях. Из результатов видно, что растения ZmGos2-zmm28 демонстрируют усиленный метаболизм азота, а также повышенную скорость поглощения и емкость ассимиляции азота.
Пример 13
Повышенное поглощение и ассимиляция N
Улучшение использования азота (N) является характерным признаком повышения урожайности сельскохозяйственных культур. Было исследовано, может ли продленная и повышенная экспрессия zmm28 улучшить поглощение N. DP202216, DP382118 и контрольные растения выращивали гидропонно в вегетационной камере и анализировали на стадии V8. Из результатов видно, что поглощение N увеличивалось на 16% в DP202216 и на 18% в DP382118 по сравнению с контролем (P <0,05). Кроме того, ассимиляция азота, измеряемая как количество ассимилированного N в тканях листа и корня на стадии роста R1, была значительно выше у DP202216 и DP382118. На стадии R1 DP202216 и DP382118 характеризовались увеличением ассимилированного N по сравнению с контролем на 10% и 12% соответственно в листе и на 23% и 17% соответственно в корне (P <0,05).
Для ответа в приведенных в данном документе примерах отдельные статистические анализы проводили с использованием программного обеспечения SAS, версия 9.4 (SAS Institute Inc., Кэри, штат Северная Каролина, США) или ASReml 3.0 (VSN International, Хемел Хемпстед, Великобритания, 2009). Линейные смешанные модели аппроксимировали под проект каждого эксперимента, оценивали средние значения для объектов (известные как эмпирические наилучшие линейные объективные оценки) и рассчитывали 95% доверительные интервалы для этих оценок.
Статистические допущения линейных смешанных моделей (т.е. нормальность, независимость и однородную дисперсию остаточной ошибки) оценивали с использованием графиков стьюдентизированных условных остатков. Для нитратредуктазы проводили логарифмическое преобразование до анализа, чтобы исправить отклонения от допущений. Результаты для нитратредуктазы обратно преобразовывали в исходную шкалу данных перед отчетом. Допущения выполняли для всех остальных ответов. Каждый из трансгенных объектов сравнивали с соответствующим контролем, используя двусторонний t-критерий различия между оценками. В экспериментах с несколькими гибридами, когда взаимодействие объекта и гибрида признавали значимым, сравнения проводили в пределах каждого гибрида. Аппроксимированные степени свободы для статистических тестов получали с применением способа Кенварда-Роджера.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pioneer Hi-Bred International, Inc.
<120> ГЕНЫ, КОНСТРУКЦИИ И ОБЪЕКТ DP-202216-6 МАИСА
<130> 7768-US-PSP
<160> 32
<170> PatentIn версия 3.5
<210> 1
<211> 251
<212> БЕЛОК
<213> Zea mays
<400> 1
Met Gly Arg Gly Pro Val Gln Leu Arg Arg Ile Glu Asn Lys Ile Asn
1. 5 10 15
Arg Gln Val Thr Phe Ser Lys Arg Arg Asn Gly Leu Leu Lys Lys Ala
20 25 30
His Glu Ile Ser Val Leu Cys Asp Ala Glu Val Ala Leu Ile Val Phe
35 40 45
Ser Thr Lys Gly Lys Leu Tyr Glu Tyr Ser Ser His Ser Ser Met Glu
50 55 60
Gly Ile Leu Glu Arg Tyr Gln Arg Tyr Ser Phe Glu Glu Arg Ala Val
65 70 75 80
Leu Asn Pro Ser Ile Glu Asp Gln Ala Asn Trp Gly Asp Glu Tyr Val
85 90 95
Arg Leu Lys Ser Lys Leu Asp Ala Leu Gln Lys Ser Gln Arg Gln Leu
100 105 110
Leu Gly Glu Gln Leu Ser Ser Leu Thr Ile Lys Glu Leu Gln Gln Leu
115 120 125
Glu Gln Gln Leu Asp Ser Ser Leu Lys His Ile Arg Ser Arg Lys Asn
130 135 140
Gln Leu Met Phe Asp Ser Ile Ser Ala Leu Gln Lys Lys Glu Lys Ala
145 150 155 160
Leu Thr Asp Gln Asn Gly Val Leu Gln Lys Phe Met Glu Ala Glu Lys
165 170 175
Glu Lys Asn Lys Ala Leu Met Asn Ala Gln Leu Arg Glu Gln Gln Asn
180 185 190
Gly Ala Ser Thr Ser Ser Pro Ser Leu Ser Pro Pro Ile Val Pro Asp
195 200 205
Ser Met Pro Thr Leu Asn Ile Gly Pro Cys Gln His Arg Gly Ala Ala
210 215 220
Glu Ser Glu Ser Glu Pro Ser Pro Ala Pro Ala Gln Ala Asn Arg Gly
225 230 235 240
Asn Leu Pro Pro Trp Met Leu Arg Thr Val Lys
245 250
<210> 2
<211> 857
<212> ДНК
<213> Zea mays
<400> 2
ccccatcacc attgctgcga cgagagtgag cgggagaggg taggtggcga ggcggcggag 60
atggggcggg ggccggtgca gctgcgccgg atcgagaaca agatcaaccg ccaggtgacc 120
ttctccaagc gccggaacgg gctgctgaag aaggcccacg agatctccgt gctctgcgac 180
gcagaggtcg cgctcatcgt cttctccact aaggggaagc tctacgagta ctctagccat 240
tccagcatgg aaggcattct tgagcgttac cagcgttact catttgaaga aagggcagta 300
cttaacccaa gtattgaaga ccaggcaaat tggggagatg aatatgtccg gttaaaatcc 360
aaacttgatg cacttcagaa gagtcaaagg cagctgttag gagaacaatt gagttcactg 420
accataaaag aactccagca actggagcaa caactggaca gttctttgaa gcatattagg 480
tcaagaaaga atcagctcat gttcgattca atttccgcgc ttcagaaaaa ggagaaagca 540
cttacagatc aaaacggtgt cctgcaaaag ttcatggagg cagagaagga gaaaaacaag 600
gctttgatga acgcgcagct ccgggagcag caaaatggag catcaacaag ctccccatca 660
ctttcaccac caatagttcc agattccatg ccaactctaa atatagggcc atgtcaacat 720
agaggggcag cagaatctga gtctgaaccg tctcctgctc ctgcacaagc aaacaggggc 780
aacctgccac catggatgct ccgcactgtc aagtaacagg tgaggtcttc ccagtgtagt 840
tttgcagctg atctcga 857
<210> 3
<211> 857
<212> ДНК
<213> Zea mays
<400> 3
taattattgg ctgtaggatt ctaaacagag cctaaatagc tggaatagct ctagccctca 60
atccaaacta atgatatcta tacttatgca actctaaatt tttattctaa aagtaatatt 120
tcatttttgt caacgagatt ctctactcta ttccacaatc ttttgaagca atatttacct 180
taaatctgta ctctatacca ataatcatat attctattat ttatttttat ctctctccta 240
aggagcatcc ccctatgtct gcatggcccc cgcctcgggt cccaatctct tgctctgcta 300
gtagcacaga agaaaacact agaaatgact tgcttgactt agagtatcag ataaacatca 360
tgtttactta actttaattt gtatcggttt ctactatttt tataatattt ttgtctctat 420
agatactacg tgcaacagta taatcaacct agtttaatcc agagcgaagg attttttact 480
aagtacgtga ctccatatgc acagcgttcc ttttatggtt cctcactggg cacagcataa 540
acgaaccctg tccaatgttt tcagcgcgaa caaacagaaa ttccatcagc gaacaaacaa 600
catacatgcg agatgaaaat aaataataaa aaaagctccg tctcgatagg ccggcacgaa 660
tcgagagcct ccatagccag ttttttccat cggaacggcg gttcgcgcac ctaattatat 720
gcaccacacg cctataaagc caaccaaccc gtcggagggg cgcaagccag acagaagaca 780
gcccgtcagc ccctctcgtt tttcatccgc cttcgcctcc aaccgcgtgc gctccacgcc 840
tcctccagga aagcgag 857
<210> 4
<211> 1013
<212> ДНК
<213> Zea mays
<400> 4
gtacgccgct cgtcctcccc cccccccctc tctaccttct ctagatcggc gttccggtcc 60
atgcatggtt agggcccggt agttctactt ctgttcatgt ttgtgttaga tccgtgtttg 120
tgttagatcc gtgctgctag cgttcgtaca cggatgcgac ctgtacgtca gacacgttct 180
gattgctaac ttgccagtgt ttctctttgg ggaatcctgg gatggctcta gccgttccgc 240
agacgggatc gatttcatga ttttttttgt ttcgttgcat agggtttggt ttgccctttt 300
cctttatttc aatatatgcc gtgcacttgt ttgtcgggtc atcttttcat gctttttttt 360
gtcttggttg tgatgatgtg gtctggttgg gcggtcgttc tagatcggag tagaattctg 420
tttcaaacta cctggtggat ttattaattt tggatctgta tgtgtgtgcc atacatattc 480
atagttacga attgaagatg atggatggaa atatcgatct aggataggta tacatgttga 540
tgcgggtttt actgatgcat atacagagat gctttttgtt cgcttggttg tgatgatgtg 600
gtgtggttgg gcggtcgttc attcgttcta gatcggagta gaatactgtt tcaaactacc 660
tggtgtattt attaattttg gaactgtatg tgtgtgtcat acatcttcat agttacgagt 720
ttaagatgga tggaaatatc gatctaggat aggtatacat gttgatgtgg gttttactga 780
tgcatataca tgatggcata tgcagcatct attcatatgc tctaaccttg agtacctatc 840
tattataata aacaagtatg ttttataatt attttgatct tgatatactt ggatgatggc 900
atatgcagca gctatatgtg gattttttta gccctgcctt catacgctat ttatttgctt 960
ggtactgttt cttttgtcga tgctcaccct gttgtttggt gttacttctg cag 1013
<210> 5
<211> 7470
<212> ДНК
<213> Искусственная последовательность
<220>
<223> T-ДНК из плазмиды
<400> 5
gtttacccgc caatatatcc tgtcaaacac tgatagttta aactgaaggc gggaaacgac 60
aatctgatca tgagcggaga attaagggag tcacgttatg acccccgccg atgacgcggg 120
acaagccgtt ttacgtttgg aactgacaga accgcaacgt tgaaggagcc actcagcaag 180
ctggtacgat tgtaatacga ctcactatag ggcgaattga gcgctgttta aacgctcttc 240
aactggaaga gcggttacta ccggctggat ggcggggcct tgatcgtgca ccgccggcgt 300
ccggactaac taactagtcg agctagttac cctatgaggt gacatgaagc gctcacggtt 360
actatgacgg ttagcttcac gactgttggt ggcagtagcg tacgacttag ctatagttcc 420
ggacttaccc ttaagataac ttcgtatagc atacattata cgaagttatg ggcccaccgg 480
tggtaccgag ctcgtttaaa cgctcttcaa ctggaagagc ggttaccaga gctggtcacc 540
tttgtccacc aagatggaac tggcgcgcct cattaattaa gtcagcggcc gctctagttg 600
aagacacgtt catgtcttca tcgtaagaag acactcagta gtcttcggcc agaatggcca 660
tctggattca gcaggcctag aaggccattt atcgctatca actttgtata gaaaagttgg 720
gccgaattcg agctcggtac ggccagaatg gccctggtaa ttattggctg taggattcta 780
aacagagcct aaatagctgg aatagctcta gccctcaatc caaactaatg atatctatac 840
ttatgcaact ctaaattttt attctaaaag taatatttca tttttgtcaa cgagattctc 900
tactctattc cacaatcttt tgaagcaata tttaccttaa atctgtactc tataccaata 960
atcatatatt ctattattta tttttatctc tctcctaagg agcatccccc tatgtctgca 1020
tggcccccgc ctcgggtccc aatctcttgc tctgctagta gcacagaaga aaacactaga 1080
aatgacttgc ttgacttaga gtatcagata aacatcatgt ttacttaact ttaatttgta 1140
tcggtttcta ctatttttat aatatttttg tctctataga tactacgtgc aacagtataa 1200
tcaacctagt ttaatccaga gcgaaggatt ttttactaag tacgtgactc catatgcaca 1260
gcgttccttt tatggttcct cactgggcac agcataaacg aaccctgtcc aatgttttca 1320
gcgcgaacaa acagaaattc catcagcgaa caaacaacat acatgcgaga tgaaaataaa 1380
taataaaaaa agctccgtct cgataggccg gcacgaatcg agagcctcca tagccagttt 1440
tttccatcgg aacggcggtt cgcgcaccta attatatgca ccacacgcct ataaagccaa 1500
ccaacccgtc ggaggggcgc aagccagaca gaagacagcc cgtcagcccc tctcgttttt 1560
catccgcctt cgcctccaac cgcgtgcgct ccacgcctcc tccaggaaag cgaggatctc 1620
ccccaaatcc acccgtcggc acctccgctt caaggtacgc cgctcgtcct cccccccccc 1680
cctctctacc ttctctagat cggcgttccg gtccatgcat ggttagggcc cggtagttct 1740
acttctgttc atgtttgtgt tagatccgtg tttgtgttag atccgtgctg ctagcgttcg 1800
tacacggatg cgacctgtac gtcagacacg ttctgattgc taacttgcca gtgtttctct 1860
ttggggaatc ctgggatggc tctagccgtt ccgcagacgg gatcgatttc atgatttttt 1920
ttgtttcgtt gcatagggtt tggtttgccc ttttccttta tttcaatata tgccgtgcac 1980
ttgtttgtcg ggtcatcttt tcatgctttt ttttgtcttg gttgtgatga tgtggtctgg 2040
ttgggcggtc gttctagatc ggagtagaat tctgtttcaa actacctggt ggatttatta 2100
attttggatc tgtatgtgtg tgccatacat attcatagtt acgaattgaa gatgatggat 2160
ggaaatatcg atctaggata ggtatacatg ttgatgcggg ttttactgat gcatatacag 2220
agatgctttt tgttcgcttg gttgtgatga tgtggtgtgg ttgggcggtc gttcattcgt 2280
tctagatcgg agtagaatac tgtttcaaac tacctggtgt atttattaat tttggaactg 2340
tatgtgtgtg tcatacatct tcatagttac gagtttaaga tggatggaaa tatcgatcta 2400
ggataggtat acatgttgat gtgggtttta ctgatgcata tacatgatgg catatgcagc 2460
atctattcat atgctctaac cttgagtacc tatctattat aataaacaag tatgttttat 2520
aattattttg atcttgatat acttggatga tggcatatgc agcagctata tgtggatttt 2580
tttagccctg ccttcatacg ctatttattt gcttggtact gtttcttttg tcgatgctca 2640
ccctgttgtt tggtgttact tctgcaggtc gactctagga tccaagcttg gcctagaagg 2700
ccagcttcaa gtttgtacaa aaaagcaggc tccgcggccg cccccttccc ccatcaccat 2760
tgctgcgacg agagtgagcg ggagagggta ggtggcgagg cggcggagat ggggcggggg 2820
ccggtgcagc tgcgccggat cgagaacaag atcaaccgcc aggtgacctt ctccaagcgc 2880
cggaacgggc tgctgaagaa ggcccacgag atctccgtgc tctgcgacgc agaggtcgcg 2940
ctcatcgtct tctccactaa ggggaagctc tacgagtact ctagccattc cagcatggaa 3000
ggcattcttg agcgttacca gcgttactca tttgaagaaa gggcagtact taacccaagt 3060
attgaagacc aggcaaattg gggagatgaa tatgtccggt taaaatccaa acttgatgca 3120
cttcagaaga gtcaaaggca gctgttagga gaacaattga gttcactgac cataaaagaa 3180
ctccagcaac tggagcaaca actggacagt tctttgaagc atattaggtc aagaaagaat 3240
cagctcatgt tcgattcaat ttccgcgctt cagaaaaagg agaaagcact tacagatcaa 3300
aacggtgtcc tgcaaaagtt catggaggca gagaaggaga aaaacaaggc tttgatgaac 3360
gcgcagctcc gggagcagca aaatggagca tcaacaagct ccccatcact ttcaccacca 3420
atagttccag attccatgcc aactctaaat atagggccat gtcaacatag aggggcagca 3480
gaatctgagt ctgaaccgtc tcctgctcct gcacaagcaa acaggggcaa cctgccacca 3540
tggatgctcc gcactgtcaa gtaacaggtg aggtcttccc agtgtagttt tgcagctgat 3600
ctcgaaaggg tgggcgcgcc gacccagctt tcttgtacaa agtggccgtt aacggatcca 3660
gacttgtcca tcttctggat tggccaactt aattaatgta tgaaataaaa ggatgcacac 3720
atagtgacat gctaatcact ataatgtggg catcaaagtt gtgtgttatg tgtaattact 3780
agttatctga ataaaagaga aagagatcat ccatatttct tatcctaaat gaatgtcacg 3840
tgtctttata attctttgat gaaccagatg catttcatta accaaatcca tatacatata 3900
aatattaatc atatataatt aatatcaatt gggttagcaa aacaaatcta gtctaggtgt 3960
gttttgcgaa ttgcggcaag cttgcggccg ccccgggcaa ctttattata caaagttgat 4020
agataaatcc tgaggatctg gtcttcctaa ggacccggga tatcggaccg attaaacttt 4080
aattcggtcc gataacttcg tatagcatac attatacgaa gttatacctg gtggcgccgc 4140
tagcctgcag tgcagcgtga cccggtcgtg cccctctcta gagataatga gcattgcatg 4200
tctaagttat aaaaaattac cacatatttt ttttgtcaca cttgtttgaa gtgcagttta 4260
tctatcttta tacatatatt taaactttac tctacgaata atataatcta tagtactaca 4320
ataatatcag tgttttagag aatcatataa atgaacagtt agacatggtc taaaggacaa 4380
ttgagtattt tgacaacagg actctacagt tttatctttt tagtgtgcat gtgttctcct 4440
ttttttttgc aaatagcttc acctatataa tacttcatcc attttattag tacatccatt 4500
tagggtttag ggttaatggt ttttatagac taattttttt agtacatcta ttttattcta 4560
ttttagcctc taaattaaga aaactaaaac tctattttag tttttttatt taataattta 4620
gatataaaat agaataaaat aaagtgacta aaaattaaac aaataccctt taagaaatta 4680
aaaaaactaa ggaaacattt ttcttgtttc gagtagataa tgccagcctg ttaaacgccg 4740
tcgacgagtc taacggacac caaccagcga accagcagcg tcgcgtcggg ccaagcgaag 4800
cagacggcac ggcatctctg tcgctgcctc tggacccctc tcgagagttc cgctccaccg 4860
ttggacttgc tccgctgtcg gcatccagaa attgcgtggc ggagcggcag acgtgagccg 4920
gcacggcagg cggcctcctc ctcctctcac ggcaccggca gctacggggg attcctttcc 4980
caccgctcct tcgctttccc ttcctcgccc gccgtaataa atagacaccc cctccacacc 5040
ctctttcccc aacctcgtgt tgttcggagc gcacacacac acaaccagat ctcccccaaa 5100
tccacccgtc ggcacctccg cttcaaggta cgccgctcgt cctccccccc ccccctctct 5160
accttctcta gatcggcgtt ccggtccatg catggttagg gcccggtagt tctacttctg 5220
ttcatgtttg tgttagatcc gtgtttgtgt tagatccgtg ctgctagcgt tcgtacacgg 5280
atgcgacctg tacgtcagac acgttctgat tgctaacttg ccagtgtttc tctttgggga 5340
atcctgggat ggctctagcc gttccgcaga cgggatcgat ttcatgattt tttttgtttc 5400
gttgcatagg gtttggtttg cccttttcct ttatttcaat atatgccgtg cacttgtttg 5460
tcgggtcatc ttttcatgct tttttttgtc ttggttgtga tgatgtggtc tggttgggcg 5520
gtcgttctag atcggagtag aattctgttt caaactacct ggtggattta ttaattttgg 5580
atctgtatgt gtgtgccata catattcata gttacgaatt gaagatgatg gatggaaata 5640
tcgatctagg ataggtatac atgttgatgc gggttttact gatgcatata cagagatgct 5700
ttttgttcgc ttggttgtga tgatgtggtg tggttgggcg gtcgttcatt cgttctagat 5760
cggagtagaa tactgtttca aactacctgg tgtatttatt aattttggaa ctgtatgtgt 5820
gtgtcataca tcttcatagt tacgagttta agatggatgg aaatatcgat ctaggatagg 5880
tatacatgtt gatgtgggtt ttactgatgc atatacatga tggcatatgc agcatctatt 5940
catatgctct aaccttgagt acctatctat tataataaac aagtatgttt tataattatt 6000
ttgatcttga tatacttgga tgatggcata tgcagcagct atatgtggat ttttttagcc 6060
ctgccttcat acgctattta tttgcttggt actgtttctt ttgtcgatgc tcaccctgtt 6120
gtttggtgtt acttctgcag gtcgactcta gaggatcaat tcgctagcga agttcctatt 6180
ccgaagttcc tattctctag aaagtatagg aacttcagat ccaccgggat ccacacgaca 6240
ccatgtcccc cgagcgccgc cccgtcgaga tccgcccggc caccgccgcc gacatggccg 6300
ccgtgtgcga catcgtgaac cactacatcg agacctccac cgtgaacttc cgcaccgagc 6360
cgcagacccc gcaggagtgg atcgacgacc tggagcgcct ccaggaccgc tacccgtggc 6420
tcgtggccga ggtggagggc gtggtggccg gcatcgccta cgccggcccg tggaaggccc 6480
gcaacgccta cgactggacc gtggagtcca ccgtgtacgt gtcccaccgc caccagcgcc 6540
tcggcctcgg ctccaccctc tacacccacc tcctcaagag catggaggcc cagggcttca 6600
agtccgtggt ggccgtgatc ggcctcccga acgacccgtc cgtgcgcctc cacgaggccc 6660
tcggctacac cgcccgcggc accctccgcg ccgccggcta caagcacggc ggctggcacg 6720
acgtcggctt ctggcagcgc gacttcgagc tgccggcccc gccgcgcccg gtgcgcccgg 6780
tgacgcagat ctgagtcgaa acctagactt gtccatcttc tggattggcc aacttaatta 6840
atgtatgaaa taaaaggatg cacacatagt gacatgctaa tcactataat gtgggcatca 6900
aagttgtgtg ttatgtgtaa ttactagtta tctgaataaa agagaaagag atcatccata 6960
tttcttatcc taaatgaatg tcacgtgtct ttataattct ttgatgaacc agatgcattt 7020
cattaaccaa atccatatac atataaatat taatcatata taattaatat caattgggtt 7080
agcaaaacaa atctagtcta ggtgtgtttt gcgaatgcgg ccctagcgta tacgaagttc 7140
ctattccgaa gttcctattc tccagaaagt ataggaactt ctgtacacct gagctgattc 7200
cgatgacttc gtaggttcct agctcaagcc gctcgtgtcc aagcgtcact tacgattagc 7260
taatgattac ggcatctagg accgactagc taactaacta gtacgtagaa ttaattcatt 7320
ccgattaatc gtggcctctt gctcttcagg atgaagagct atgtttaaac gtgcaagcgc 7380
tactagacaa ttcagtacat taaaaacgtc cgcaatgtgt tattaagttg tctaagcgtc 7440
aatttgttta caccacaata tatcctgcca 7470
<210> 6
<211> 7436
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ВСТАВЛЕННАЯ ДНК
<400> 6
tcaaacactg atagtttaaa ctgaaggcgg gaaacgacaa tctgatcatg agcggagaat 60
taagggagtc acgttatgac ccccgccgat gacgcgggac aagccgtttt acgtttggaa 120
ctgacagaac cgcaacgttg aaggagccac tcagcaagct ggtacgattg taatacgact 180
cactataggg cgaattgagc gctgtttaaa cgctcttcaa ctggaagagc ggttactacc 240
ggctggatgg cggggccttg atcgtgcacc gccggcgtcc ggactaacta actagtcgag 300
ctagttaccc tatgaggtga catgaagcgc tcacggttac tatgacggtt agcttcacga 360
ctgttggtgg cagtagcgta cgacttagct atagttccgg acttaccctt aagataactt 420
cgtatagcat acattatacg aagttatggg cccaccggtg gtaccgagct cgtttaaacg 480
ctcttcaact ggaagagcgg ttaccagagc tggtcacctt tgtccaccaa gatggaactg 540
gcgcgcctca ttaattaagt cagcggccgc tctagttgaa gacacgttca tgtcttcatc 600
gtaagaagac actcagtagt cttcggccag aatggccatc tggattcagc aggcctagaa 660
ggccatttat cgctatcaac tttgtataga aaagttgggc cgaattcgag ctcggtacgg 720
ccagaatggc cctggtaatt attggctgta ggattctaaa cagagcctaa atagctggaa 780
tagctctagc cctcaatcca aactaatgat atctatactt atgcaactct aaatttttat 840
tctaaaagta atatttcatt tttgtcaacg agattctcta ctctattcca caatcttttg 900
aagcaatatt taccttaaat ctgtactcta taccaataat catatattct attatttatt 960
tttatctctc tcctaaggag catcccccta tgtctgcatg gcccccgcct cgggtcccaa 1020
tctcttgctc tgctagtagc acagaagaaa acactagaaa tgacttgctt gacttagagt 1080
atcagataaa catcatgttt acttaacttt aatttgtatc ggtttctact atttttataa 1140
tatttttgtc tctatagata ctacgtgcaa cagtataatc aacctagttt aatccagagc 1200
gaaggatttt ttactaagta cgtgactcca tatgcacagc gttcctttta tggttcctca 1260
ctgggcacag cataaacgaa ccctgtccaa tgttttcagc gcgaacaaac agaaattcca 1320
tcagcgaaca aacaacatac atgcgagatg aaaataaata ataaaaaaag ctccgtctcg 1380
ataggccggc acgaatcgag agcctccata gccagttttt tccatcggaa cggcggttcg 1440
cgcacctaat tatatgcacc acacgcctat aaagccaacc aacccgtcgg aggggcgcaa 1500
gccagacaga agacagcccg tcagcccctc tcgtttttca tccgccttcg cctccaaccg 1560
cgtgcgctcc acgcctcctc caggaaagcg aggatctccc ccaaatccac ccgtcggcac 1620
ctccgcttca aggtacgccg ctcgtcctcc cccccccccc tctctacctt ctctagatcg 1680
gcgttccggt ccatgcatgg ttagggcccg gtagttctac ttctgttcat gtttgtgtta 1740
gatccgtgtt tgtgttagat ccgtgctgct agcgttcgta cacggatgcg acctgtacgt 1800
cagacacgtt ctgattgcta acttgccagt gtttctcttt ggggaatcct gggatggctc 1860
tagccgttcc gcagacggga tcgatttcat gatttttttt gtttcgttgc atagggtttg 1920
gtttgccctt ttcctttatt tcaatatatg ccgtgcactt gtttgtcggg tcatcttttc 1980
atgctttttt ttgtcttggt tgtgatgatg tggtctggtt gggcggtcgt tctagatcgg 2040
agtagaattc tgtttcaaac tacctggtgg atttattaat tttggatctg tatgtgtgtg 2100
ccatacatat tcatagttac gaattgaaga tgatggatgg aaatatcgat ctaggatagg 2160
tatacatgtt gatgcgggtt ttactgatgc atatacagag atgctttttg ttcgcttggt 2220
tgtgatgatg tggtgtggtt gggcggtcgt tcattcgttc tagatcggag tagaatactg 2280
tttcaaacta cctggtgtat ttattaattt tggaactgta tgtgtgtgtc atacatcttc 2340
atagttacga gtttaagatg gatggaaata tcgatctagg ataggtatac atgttgatgt 2400
gggttttact gatgcatata catgatggca tatgcagcat ctattcatat gctctaacct 2460
tgagtaccta tctattataa taaacaagta tgttttataa ttattttgat cttgatatac 2520
ttggatgatg gcatatgcag cagctatatg tggatttttt tagccctgcc ttcatacgct 2580
atttatttgc ttggtactgt ttcttttgtc gatgctcacc ctgttgtttg gtgttacttc 2640
tgcaggtcga ctctaggatc caagcttggc ctagaaggcc agcttcaagt ttgtacaaaa 2700
aagcaggctc cgcggccgcc cccttccccc atcaccattg ctgcgacgag agtgagcggg 2760
agagggtagg tggcgaggcg gcggagatgg ggcgggggcc ggtgcagctg cgccggatcg 2820
agaacaagat caaccgccag gtgaccttct ccaagcgccg gaacgggctg ctgaagaagg 2880
cccacgagat ctccgtgctc tgcgacgcag aggtcgcgct catcgtcttc tccactaagg 2940
ggaagctcta cgagtactct agccattcca gcatggaagg cattcttgag cgttaccagc 3000
gttactcatt tgaagaaagg gcagtactta acccaagtat tgaagaccag gcaaattggg 3060
gagatgaata tgtccggtta aaatccaaac ttgatgcact tcagaagagt caaaggcagc 3120
tgttaggaga acaattgagt tcactgacca taaaagaact ccagcaactg gagcaacaac 3180
tggacagttc tttgaagcat attaggtcaa gaaagaatca gctcatgttc gattcaattt 3240
ccgcgcttca gaaaaaggag aaagcactta cagatcaaaa cggtgtcctg caaaagttca 3300
tggaggcaga gaaggagaaa aacaaggctt tgatgaacgc gcagctccgg gagcagcaaa 3360
atggagcatc aacaagctcc ccatcacttt caccaccaat agttccagat tccatgccaa 3420
ctctaaatat agggccatgt caacatagag gggcagcaga atctgagtct gaaccgtctc 3480
ctgctcctgc acaagcaaac aggggcaacc tgccaccatg gatgctccgc actgtcaagt 3540
aacaggtgag gtcttcccag tgtagttttg cagctgatct cgaaagggtg ggcgcgccga 3600
cccagctttc ttgtacaaag tggccgttaa cggatccaga cttgtccatc ttctggattg 3660
gccaacttaa ttaatgtatg aaataaaagg atgcacacat agtgacatgc taatcactat 3720
aatgtgggca tcaaagttgt gtgttatgtg taattactag ttatctgaat aaaagagaaa 3780
gagatcatcc atatttctta tcctaaatga atgtcacgtg tctttataat tctttgatga 3840
accagatgca tttcattaac caaatccata tacatataaa tattaatcat atataattaa 3900
tatcaattgg gttagcaaaa caaatctagt ctaggtgtgt tttgcgaatt gcggcaagct 3960
tgcggccgcc ccgggcaact ttattataca aagttgatag ataaatcctg aggatctggt 4020
cttcctaagg acccgggata tcggaccgat taaactttaa ttcggtccga taacttcgta 4080
tagcatacat tatacgaagt tatacctggt ggcgccgcta gcctgcagtg cagcgtgacc 4140
cggtcgtgcc cctctctaga gataatgagc attgcatgtc taagttataa aaaattacca 4200
catatttttt ttgtcacact tgtttgaagt gcagtttatc tatctttata catatattta 4260
aactttactc tacgaataat ataatctata gtactacaat aatatcagtg ttttagagaa 4320
tcatataaat gaacagttag acatggtcta aaggacaatt gagtattttg acaacaggac 4380
tctacagttt tatcttttta gtgtgcatgt gttctccttt ttttttgcaa atagcttcac 4440
ctatataata cttcatccat tttattagta catccattta gggtttaggg ttaatggttt 4500
ttatagacta atttttttag tacatctatt ttattctatt ttagcctcta aattaagaaa 4560
actaaaactc tattttagtt tttttattta ataatttaga tataaaatag aataaaataa 4620
agtgactaaa aattaaacaa atacccttta agaaattaaa aaaactaagg aaacattttt 4680
cttgtttcga gtagataatg ccagcctgtt aaacgccgtc gacgagtcta acggacacca 4740
accagcgaac cagcagcgtc gcgtcgggcc aagcgaagca gacggcacgg catctctgtc 4800
gctgcctctg gacccctctc gagagttccg ctccaccgtt ggacttgctc cgctgtcggc 4860
atccagaaat tgcgtggcgg agcggcagac gtgagccggc acggcaggcg gcctcctcct 4920
cctctcacgg caccggcagc tacgggggat tcctttccca ccgctccttc gctttccctt 4980
cctcgcccgc cgtaataaat agacaccccc tccacaccct ctttccccaa cctcgtgttg 5040
ttcggagcgc acacacacac aaccagatct cccccaaatc cacccgtcgg cacctccgct 5100
tcaaggtacg ccgctcgtcc tccccccccc ccctctctac cttctctaga tcggcgttcc 5160
ggtccatgca tggttagggc ccggtagttc tacttctgtt catgtttgtg ttagatccgt 5220
gtttgtgtta gatccgtgct gctagcgttc gtacacggat gcgacctgta cgtcagacac 5280
gttctgattg ctaacttgcc agtgtttctc tttggggaat cctgggatgg ctctagccgt 5340
tccgcagacg ggatcgattt catgattttt tttgtttcgt tgcatagggt ttggtttgcc 5400
cttttccttt atttcaatat atgccgtgca cttgtttgtc gggtcatctt ttcatgcttt 5460
tttttgtctt ggttgtgatg atgtggtctg gttgggcggt cgttctagat cggagtagaa 5520
ttctgtttca aactacctgg tggatttatt aattttggat ctgtatgtgt gtgccataca 5580
tattcatagt tacgaattga agatgatgga tggaaatatc gatctaggat aggtatacat 5640
gttgatgcgg gttttactga tgcatataca gagatgcttt ttgttcgctt ggttgtgatg 5700
atgtggtgtg gttgggcggt cgttcattcg ttctagatcg gagtagaata ctgtttcaaa 5760
ctacctggtg tatttattaa ttttggaact gtatgtgtgt gtcatacatc ttcatagtta 5820
cgagtttaag atggatggaa atatcgatct aggataggta tacatgttga tgtgggtttt 5880
actgatgcat atacatgatg gcatatgcag catctattca tatgctctaa ccttgagtac 5940
ctatctatta taataaacaa gtatgtttta taattatttt gatcttgata tacttggatg 6000
atggcatatg cagcagctat atgtggattt ttttagccct gccttcatac gctatttatt 6060
tgcttggtac tgtttctttt gtcgatgctc accctgttgt ttggtgttac ttctgcaggt 6120
cgactctaga ggatcaattc gctagcgaag ttcctattcc gaagttccta ttctctagaa 6180
agtataggaa cttcagatcc accgggatcc acacgacacc atgtcccccg agcgccgccc 6240
cgtcgagatc cgcccggcca ccgccgccga catggccgcc gtgtgcgaca tcgtgaacca 6300
ctacatcgag acctccaccg tgaacttccg caccgagccg cagaccccgc aggagtggat 6360
cgacgacctg gagcgcctcc aggaccgcta cccgtggctc gtggccgagg tggagggcgt 6420
ggtggccggc atcgcctacg ccggcccgtg gaaggcccgc aacgcctacg actggaccgt 6480
ggagtccacc gtgtacgtgt cccaccgcca ccagcgcctc ggcctcggct ccaccctcta 6540
cacccacctc ctcaagagca tggaggccca gggcttcaag tccgtggtgg ccgtgatcgg 6600
cctcccgaac gacccgtccg tgcgcctcca cgaggccctc ggctacaccg cccgcggcac 6660
cctccgcgcc gccggctaca agcacggcgg ctggcacgac gtcggcttct ggcagcgcga 6720
cttcgagctg ccggccccgc cgcgcccggt gcgcccggtg acgcagatct gagtcgaaac 6780
ctagacttgt ccatcttctg gattggccaa cttaattaat gtatgaaata aaaggatgca 6840
cacatagtga catgctaatc actataatgt gggcatcaaa gttgtgtgtt atgtgtaatt 6900
actagttatc tgaataaaag agaaagagat catccatatt tcttatccta aatgaatgtc 6960
acgtgtcttt ataattcttt gatgaaccag atgcatttca ttaaccaaat ccatatacat 7020
ataaatatta atcatatata attaatatca attgggttag caaaacaaat ctagtctagg 7080
tgtgttttgc gaatgcggcc ctagcgtata cgaagttcct attccgaagt tcctattctc 7140
cagaaagtat aggaacttct gtacacctga gctgattccg atgacttcgt aggttcctag 7200
ctcaagccgc tcgtgtccaa gcgtcactta cgattagcta atgattacgg catctaggac 7260
cgactagcta actaactagt acgtagaatt aattcattcc gattaatcgt ggcctcttgc 7320
tcttcaggat gaagagctat gtttaaacgt gcaagcgcta ctagacaatt cagtacatta 7380
aaaacgtccg caatgtgtta ttaagttgtc taagcgtcaa tttgtttaca ccacaa 7436
<210> 7
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПОСЛЕДОВАТЕЛЬНОСТЬ СОЕДИНЕНИЯ
<400> 7
cacactcaaa 10
<210> 8
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> СОЕДИНЕНИЕ
<400> 8
cacaaagtag 10
<210> 9
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> СОЕДИНЕНИЕ
<400> 9
aacaacacac tcaaacactg 20
<210> 10
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> СОЕДИНЕНИЕ
<400> 10
tacaccacaa agtagacaag 20
<210> 11
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> СОЕДИНЕНИЕ
<400> 11
gcacaaacaa cacactcaaa cactgatagt 30
<210> 12
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> СОЕДИНЕНИЕ
<400> 12
ttgtttacac cacaaagtag acaagttgtg 30
<210> 13
<211> 7456
<212> ДНК
<213> Искусственная последовательность
<220>
<223> СОЕДИНЕНИЕ + ВСТАВКА
<400> 13
aacaacacac tcaaacactg atagtttaaa ctgaaggcgg gaaacgacaa tctgatcatg 60
agcggagaat taagggagtc acgttatgac ccccgccgat gacgcgggac aagccgtttt 120
acgtttggaa ctgacagaac cgcaacgttg aaggagccac tcagcaagct ggtacgattg 180
taatacgact cactataggg cgaattgagc gctgtttaaa cgctcttcaa ctggaagagc 240
ggttactacc ggctggatgg cggggccttg atcgtgcacc gccggcgtcc ggactaacta 300
actagtcgag ctagttaccc tatgaggtga catgaagcgc tcacggttac tatgacggtt 360
agcttcacga ctgttggtgg cagtagcgta cgacttagct atagttccgg acttaccctt 420
aagataactt cgtatagcat acattatacg aagttatggg cccaccggtg gtaccgagct 480
cgtttaaacg ctcttcaact ggaagagcgg ttaccagagc tggtcacctt tgtccaccaa 540
gatggaactg gcgcgcctca ttaattaagt cagcggccgc tctagttgaa gacacgttca 600
tgtcttcatc gtaagaagac actcagtagt cttcggccag aatggccatc tggattcagc 660
aggcctagaa ggccatttat cgctatcaac tttgtataga aaagttgggc cgaattcgag 720
ctcggtacgg ccagaatggc cctggtaatt attggctgta ggattctaaa cagagcctaa 780
atagctggaa tagctctagc cctcaatcca aactaatgat atctatactt atgcaactct 840
aaatttttat tctaaaagta atatttcatt tttgtcaacg agattctcta ctctattcca 900
caatcttttg aagcaatatt taccttaaat ctgtactcta taccaataat catatattct 960
attatttatt tttatctctc tcctaaggag catcccccta tgtctgcatg gcccccgcct 1020
cgggtcccaa tctcttgctc tgctagtagc acagaagaaa acactagaaa tgacttgctt 1080
gacttagagt atcagataaa catcatgttt acttaacttt aatttgtatc ggtttctact 1140
atttttataa tatttttgtc tctatagata ctacgtgcaa cagtataatc aacctagttt 1200
aatccagagc gaaggatttt ttactaagta cgtgactcca tatgcacagc gttcctttta 1260
tggttcctca ctgggcacag cataaacgaa ccctgtccaa tgttttcagc gcgaacaaac 1320
agaaattcca tcagcgaaca aacaacatac atgcgagatg aaaataaata ataaaaaaag 1380
ctccgtctcg ataggccggc acgaatcgag agcctccata gccagttttt tccatcggaa 1440
cggcggttcg cgcacctaat tatatgcacc acacgcctat aaagccaacc aacccgtcgg 1500
aggggcgcaa gccagacaga agacagcccg tcagcccctc tcgtttttca tccgccttcg 1560
cctccaaccg cgtgcgctcc acgcctcctc caggaaagcg aggatctccc ccaaatccac 1620
ccgtcggcac ctccgcttca aggtacgccg ctcgtcctcc cccccccccc tctctacctt 1680
ctctagatcg gcgttccggt ccatgcatgg ttagggcccg gtagttctac ttctgttcat 1740
gtttgtgtta gatccgtgtt tgtgttagat ccgtgctgct agcgttcgta cacggatgcg 1800
acctgtacgt cagacacgtt ctgattgcta acttgccagt gtttctcttt ggggaatcct 1860
gggatggctc tagccgttcc gcagacggga tcgatttcat gatttttttt gtttcgttgc 1920
atagggtttg gtttgccctt ttcctttatt tcaatatatg ccgtgcactt gtttgtcggg 1980
tcatcttttc atgctttttt ttgtcttggt tgtgatgatg tggtctggtt gggcggtcgt 2040
tctagatcgg agtagaattc tgtttcaaac tacctggtgg atttattaat tttggatctg 2100
tatgtgtgtg ccatacatat tcatagttac gaattgaaga tgatggatgg aaatatcgat 2160
ctaggatagg tatacatgtt gatgcgggtt ttactgatgc atatacagag atgctttttg 2220
ttcgcttggt tgtgatgatg tggtgtggtt gggcggtcgt tcattcgttc tagatcggag 2280
tagaatactg tttcaaacta cctggtgtat ttattaattt tggaactgta tgtgtgtgtc 2340
atacatcttc atagttacga gtttaagatg gatggaaata tcgatctagg ataggtatac 2400
atgttgatgt gggttttact gatgcatata catgatggca tatgcagcat ctattcatat 2460
gctctaacct tgagtaccta tctattataa taaacaagta tgttttataa ttattttgat 2520
cttgatatac ttggatgatg gcatatgcag cagctatatg tggatttttt tagccctgcc 2580
ttcatacgct atttatttgc ttggtactgt ttcttttgtc gatgctcacc ctgttgtttg 2640
gtgttacttc tgcaggtcga ctctaggatc caagcttggc ctagaaggcc agcttcaagt 2700
ttgtacaaaa aagcaggctc cgcggccgcc cccttccccc atcaccattg ctgcgacgag 2760
agtgagcggg agagggtagg tggcgaggcg gcggagatgg ggcgggggcc ggtgcagctg 2820
cgccggatcg agaacaagat caaccgccag gtgaccttct ccaagcgccg gaacgggctg 2880
ctgaagaagg cccacgagat ctccgtgctc tgcgacgcag aggtcgcgct catcgtcttc 2940
tccactaagg ggaagctcta cgagtactct agccattcca gcatggaagg cattcttgag 3000
cgttaccagc gttactcatt tgaagaaagg gcagtactta acccaagtat tgaagaccag 3060
gcaaattggg gagatgaata tgtccggtta aaatccaaac ttgatgcact tcagaagagt 3120
caaaggcagc tgttaggaga acaattgagt tcactgacca taaaagaact ccagcaactg 3180
gagcaacaac tggacagttc tttgaagcat attaggtcaa gaaagaatca gctcatgttc 3240
gattcaattt ccgcgcttca gaaaaaggag aaagcactta cagatcaaaa cggtgtcctg 3300
caaaagttca tggaggcaga gaaggagaaa aacaaggctt tgatgaacgc gcagctccgg 3360
gagcagcaaa atggagcatc aacaagctcc ccatcacttt caccaccaat agttccagat 3420
tccatgccaa ctctaaatat agggccatgt caacatagag gggcagcaga atctgagtct 3480
gaaccgtctc ctgctcctgc acaagcaaac aggggcaacc tgccaccatg gatgctccgc 3540
actgtcaagt aacaggtgag gtcttcccag tgtagttttg cagctgatct cgaaagggtg 3600
ggcgcgccga cccagctttc ttgtacaaag tggccgttaa cggatccaga cttgtccatc 3660
ttctggattg gccaacttaa ttaatgtatg aaataaaagg atgcacacat agtgacatgc 3720
taatcactat aatgtgggca tcaaagttgt gtgttatgtg taattactag ttatctgaat 3780
aaaagagaaa gagatcatcc atatttctta tcctaaatga atgtcacgtg tctttataat 3840
tctttgatga accagatgca tttcattaac caaatccata tacatataaa tattaatcat 3900
atataattaa tatcaattgg gttagcaaaa caaatctagt ctaggtgtgt tttgcgaatt 3960
gcggcaagct tgcggccgcc ccgggcaact ttattataca aagttgatag ataaatcctg 4020
aggatctggt cttcctaagg acccgggata tcggaccgat taaactttaa ttcggtccga 4080
taacttcgta tagcatacat tatacgaagt tatacctggt ggcgccgcta gcctgcagtg 4140
cagcgtgacc cggtcgtgcc cctctctaga gataatgagc attgcatgtc taagttataa 4200
aaaattacca catatttttt ttgtcacact tgtttgaagt gcagtttatc tatctttata 4260
catatattta aactttactc tacgaataat ataatctata gtactacaat aatatcagtg 4320
ttttagagaa tcatataaat gaacagttag acatggtcta aaggacaatt gagtattttg 4380
acaacaggac tctacagttt tatcttttta gtgtgcatgt gttctccttt ttttttgcaa 4440
atagcttcac ctatataata cttcatccat tttattagta catccattta gggtttaggg 4500
ttaatggttt ttatagacta atttttttag tacatctatt ttattctatt ttagcctcta 4560
aattaagaaa actaaaactc tattttagtt tttttattta ataatttaga tataaaatag 4620
aataaaataa agtgactaaa aattaaacaa atacccttta agaaattaaa aaaactaagg 4680
aaacattttt cttgtttcga gtagataatg ccagcctgtt aaacgccgtc gacgagtcta 4740
acggacacca accagcgaac cagcagcgtc gcgtcgggcc aagcgaagca gacggcacgg 4800
catctctgtc gctgcctctg gacccctctc gagagttccg ctccaccgtt ggacttgctc 4860
cgctgtcggc atccagaaat tgcgtggcgg agcggcagac gtgagccggc acggcaggcg 4920
gcctcctcct cctctcacgg caccggcagc tacgggggat tcctttccca ccgctccttc 4980
gctttccctt cctcgcccgc cgtaataaat agacaccccc tccacaccct ctttccccaa 5040
cctcgtgttg ttcggagcgc acacacacac aaccagatct cccccaaatc cacccgtcgg 5100
cacctccgct tcaaggtacg ccgctcgtcc tccccccccc ccctctctac cttctctaga 5160
tcggcgttcc ggtccatgca tggttagggc ccggtagttc tacttctgtt catgtttgtg 5220
ttagatccgt gtttgtgtta gatccgtgct gctagcgttc gtacacggat gcgacctgta 5280
cgtcagacac gttctgattg ctaacttgcc agtgtttctc tttggggaat cctgggatgg 5340
ctctagccgt tccgcagacg ggatcgattt catgattttt tttgtttcgt tgcatagggt 5400
ttggtttgcc cttttccttt atttcaatat atgccgtgca cttgtttgtc gggtcatctt 5460
ttcatgcttt tttttgtctt ggttgtgatg atgtggtctg gttgggcggt cgttctagat 5520
cggagtagaa ttctgtttca aactacctgg tggatttatt aattttggat ctgtatgtgt 5580
gtgccataca tattcatagt tacgaattga agatgatgga tggaaatatc gatctaggat 5640
aggtatacat gttgatgcgg gttttactga tgcatataca gagatgcttt ttgttcgctt 5700
ggttgtgatg atgtggtgtg gttgggcggt cgttcattcg ttctagatcg gagtagaata 5760
ctgtttcaaa ctacctggtg tatttattaa ttttggaact gtatgtgtgt gtcatacatc 5820
ttcatagtta cgagtttaag atggatggaa atatcgatct aggataggta tacatgttga 5880
tgtgggtttt actgatgcat atacatgatg gcatatgcag catctattca tatgctctaa 5940
ccttgagtac ctatctatta taataaacaa gtatgtttta taattatttt gatcttgata 6000
tacttggatg atggcatatg cagcagctat atgtggattt ttttagccct gccttcatac 6060
gctatttatt tgcttggtac tgtttctttt gtcgatgctc accctgttgt ttggtgttac 6120
ttctgcaggt cgactctaga ggatcaattc gctagcgaag ttcctattcc gaagttccta 6180
ttctctagaa agtataggaa cttcagatcc accgggatcc acacgacacc atgtcccccg 6240
agcgccgccc cgtcgagatc cgcccggcca ccgccgccga catggccgcc gtgtgcgaca 6300
tcgtgaacca ctacatcgag acctccaccg tgaacttccg caccgagccg cagaccccgc 6360
aggagtggat cgacgacctg gagcgcctcc aggaccgcta cccgtggctc gtggccgagg 6420
tggagggcgt ggtggccggc atcgcctacg ccggcccgtg gaaggcccgc aacgcctacg 6480
actggaccgt ggagtccacc gtgtacgtgt cccaccgcca ccagcgcctc ggcctcggct 6540
ccaccctcta cacccacctc ctcaagagca tggaggccca gggcttcaag tccgtggtgg 6600
ccgtgatcgg cctcccgaac gacccgtccg tgcgcctcca cgaggccctc ggctacaccg 6660
cccgcggcac cctccgcgcc gccggctaca agcacggcgg ctggcacgac gtcggcttct 6720
ggcagcgcga cttcgagctg ccggccccgc cgcgcccggt gcgcccggtg acgcagatct 6780
gagtcgaaac ctagacttgt ccatcttctg gattggccaa cttaattaat gtatgaaata 6840
aaaggatgca cacatagtga catgctaatc actataatgt gggcatcaaa gttgtgtgtt 6900
atgtgtaatt actagttatc tgaataaaag agaaagagat catccatatt tcttatccta 6960
aatgaatgtc acgtgtcttt ataattcttt gatgaaccag atgcatttca ttaaccaaat 7020
ccatatacat ataaatatta atcatatata attaatatca attgggttag caaaacaaat 7080
ctagtctagg tgtgttttgc gaatgcggcc ctagcgtata cgaagttcct attccgaagt 7140
tcctattctc cagaaagtat aggaacttct gtacacctga gctgattccg atgacttcgt 7200
aggttcctag ctcaagccgc tcgtgtccaa gcgtcactta cgattagcta atgattacgg 7260
catctaggac cgactagcta actaactagt acgtagaatt aattcattcc gattaatcgt 7320
ggcctcttgc tcttcaggat gaagagctat gtttaaacgt gcaagcgcta ctagacaatt 7380
cagtacatta aaaacgtccg caatgtgtta ttaagttgtc taagcgtcaa tttgtttaca 7440
ccacaaagta gacaag 7456
<210> 14
<211> 7476
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ВСТАВКА + СОЕДИНЕНИЕ
<400> 14
gtacagcaca aacaacacac tcaaacactg atagtttaaa ctgaaggcgg gaaacgacaa 60
tctgatcatg agcggagaat taagggagtc acgttatgac ccccgccgat gacgcgggac 120
aagccgtttt acgtttggaa ctgacagaac cgcaacgttg aaggagccac tcagcaagct 180
ggtacgattg taatacgact cactataggg cgaattgagc gctgtttaaa cgctcttcaa 240
ctggaagagc ggttactacc ggctggatgg cggggccttg atcgtgcacc gccggcgtcc 300
ggactaacta actagtcgag ctagttaccc tatgaggtga catgaagcgc tcacggttac 360
tatgacggtt agcttcacga ctgttggtgg cagtagcgta cgacttagct atagttccgg 420
acttaccctt aagataactt cgtatagcat acattatacg aagttatggg cccaccggtg 480
gtaccgagct cgtttaaacg ctcttcaact ggaagagcgg ttaccagagc tggtcacctt 540
tgtccaccaa gatggaactg gcgcgcctca ttaattaagt cagcggccgc tctagttgaa 600
gacacgttca tgtcttcatc gtaagaagac actcagtagt cttcggccag aatggccatc 660
tggattcagc aggcctagaa ggccatttat cgctatcaac tttgtataga aaagttgggc 720
cgaattcgag ctcggtacgg ccagaatggc cctggtaatt attggctgta ggattctaaa 780
cagagcctaa atagctggaa tagctctagc cctcaatcca aactaatgat atctatactt 840
atgcaactct aaatttttat tctaaaagta atatttcatt tttgtcaacg agattctcta 900
ctctattcca caatcttttg aagcaatatt taccttaaat ctgtactcta taccaataat 960
catatattct attatttatt tttatctctc tcctaaggag catcccccta tgtctgcatg 1020
gcccccgcct cgggtcccaa tctcttgctc tgctagtagc acagaagaaa acactagaaa 1080
tgacttgctt gacttagagt atcagataaa catcatgttt acttaacttt aatttgtatc 1140
ggtttctact atttttataa tatttttgtc tctatagata ctacgtgcaa cagtataatc 1200
aacctagttt aatccagagc gaaggatttt ttactaagta cgtgactcca tatgcacagc 1260
gttcctttta tggttcctca ctgggcacag cataaacgaa ccctgtccaa tgttttcagc 1320
gcgaacaaac agaaattcca tcagcgaaca aacaacatac atgcgagatg aaaataaata 1380
ataaaaaaag ctccgtctcg ataggccggc acgaatcgag agcctccata gccagttttt 1440
tccatcggaa cggcggttcg cgcacctaat tatatgcacc acacgcctat aaagccaacc 1500
aacccgtcgg aggggcgcaa gccagacaga agacagcccg tcagcccctc tcgtttttca 1560
tccgccttcg cctccaaccg cgtgcgctcc acgcctcctc caggaaagcg aggatctccc 1620
ccaaatccac ccgtcggcac ctccgcttca aggtacgccg ctcgtcctcc cccccccccc 1680
tctctacctt ctctagatcg gcgttccggt ccatgcatgg ttagggcccg gtagttctac 1740
ttctgttcat gtttgtgtta gatccgtgtt tgtgttagat ccgtgctgct agcgttcgta 1800
cacggatgcg acctgtacgt cagacacgtt ctgattgcta acttgccagt gtttctcttt 1860
ggggaatcct gggatggctc tagccgttcc gcagacggga tcgatttcat gatttttttt 1920
gtttcgttgc atagggtttg gtttgccctt ttcctttatt tcaatatatg ccgtgcactt 1980
gtttgtcggg tcatcttttc atgctttttt ttgtcttggt tgtgatgatg tggtctggtt 2040
gggcggtcgt tctagatcgg agtagaattc tgtttcaaac tacctggtgg atttattaat 2100
tttggatctg tatgtgtgtg ccatacatat tcatagttac gaattgaaga tgatggatgg 2160
aaatatcgat ctaggatagg tatacatgtt gatgcgggtt ttactgatgc atatacagag 2220
atgctttttg ttcgcttggt tgtgatgatg tggtgtggtt gggcggtcgt tcattcgttc 2280
tagatcggag tagaatactg tttcaaacta cctggtgtat ttattaattt tggaactgta 2340
tgtgtgtgtc atacatcttc atagttacga gtttaagatg gatggaaata tcgatctagg 2400
ataggtatac atgttgatgt gggttttact gatgcatata catgatggca tatgcagcat 2460
ctattcatat gctctaacct tgagtaccta tctattataa taaacaagta tgttttataa 2520
ttattttgat cttgatatac ttggatgatg gcatatgcag cagctatatg tggatttttt 2580
tagccctgcc ttcatacgct atttatttgc ttggtactgt ttcttttgtc gatgctcacc 2640
ctgttgtttg gtgttacttc tgcaggtcga ctctaggatc caagcttggc ctagaaggcc 2700
agcttcaagt ttgtacaaaa aagcaggctc cgcggccgcc cccttccccc atcaccattg 2760
ctgcgacgag agtgagcggg agagggtagg tggcgaggcg gcggagatgg ggcgggggcc 2820
ggtgcagctg cgccggatcg agaacaagat caaccgccag gtgaccttct ccaagcgccg 2880
gaacgggctg ctgaagaagg cccacgagat ctccgtgctc tgcgacgcag aggtcgcgct 2940
catcgtcttc tccactaagg ggaagctcta cgagtactct agccattcca gcatggaagg 3000
cattcttgag cgttaccagc gttactcatt tgaagaaagg gcagtactta acccaagtat 3060
tgaagaccag gcaaattggg gagatgaata tgtccggtta aaatccaaac ttgatgcact 3120
tcagaagagt caaaggcagc tgttaggaga acaattgagt tcactgacca taaaagaact 3180
ccagcaactg gagcaacaac tggacagttc tttgaagcat attaggtcaa gaaagaatca 3240
gctcatgttc gattcaattt ccgcgcttca gaaaaaggag aaagcactta cagatcaaaa 3300
cggtgtcctg caaaagttca tggaggcaga gaaggagaaa aacaaggctt tgatgaacgc 3360
gcagctccgg gagcagcaaa atggagcatc aacaagctcc ccatcacttt caccaccaat 3420
agttccagat tccatgccaa ctctaaatat agggccatgt caacatagag gggcagcaga 3480
atctgagtct gaaccgtctc ctgctcctgc acaagcaaac aggggcaacc tgccaccatg 3540
gatgctccgc actgtcaagt aacaggtgag gtcttcccag tgtagttttg cagctgatct 3600
cgaaagggtg ggcgcgccga cccagctttc ttgtacaaag tggccgttaa cggatccaga 3660
cttgtccatc ttctggattg gccaacttaa ttaatgtatg aaataaaagg atgcacacat 3720
agtgacatgc taatcactat aatgtgggca tcaaagttgt gtgttatgtg taattactag 3780
ttatctgaat aaaagagaaa gagatcatcc atatttctta tcctaaatga atgtcacgtg 3840
tctttataat tctttgatga accagatgca tttcattaac caaatccata tacatataaa 3900
tattaatcat atataattaa tatcaattgg gttagcaaaa caaatctagt ctaggtgtgt 3960
tttgcgaatt gcggcaagct tgcggccgcc ccgggcaact ttattataca aagttgatag 4020
ataaatcctg aggatctggt cttcctaagg acccgggata tcggaccgat taaactttaa 4080
ttcggtccga taacttcgta tagcatacat tatacgaagt tatacctggt ggcgccgcta 4140
gcctgcagtg cagcgtgacc cggtcgtgcc cctctctaga gataatgagc attgcatgtc 4200
taagttataa aaaattacca catatttttt ttgtcacact tgtttgaagt gcagtttatc 4260
tatctttata catatattta aactttactc tacgaataat ataatctata gtactacaat 4320
aatatcagtg ttttagagaa tcatataaat gaacagttag acatggtcta aaggacaatt 4380
gagtattttg acaacaggac tctacagttt tatcttttta gtgtgcatgt gttctccttt 4440
ttttttgcaa atagcttcac ctatataata cttcatccat tttattagta catccattta 4500
gggtttaggg ttaatggttt ttatagacta atttttttag tacatctatt ttattctatt 4560
ttagcctcta aattaagaaa actaaaactc tattttagtt tttttattta ataatttaga 4620
tataaaatag aataaaataa agtgactaaa aattaaacaa atacccttta agaaattaaa 4680
aaaactaagg aaacattttt cttgtttcga gtagataatg ccagcctgtt aaacgccgtc 4740
gacgagtcta acggacacca accagcgaac cagcagcgtc gcgtcgggcc aagcgaagca 4800
gacggcacgg catctctgtc gctgcctctg gacccctctc gagagttccg ctccaccgtt 4860
ggacttgctc cgctgtcggc atccagaaat tgcgtggcgg agcggcagac gtgagccggc 4920
acggcaggcg gcctcctcct cctctcacgg caccggcagc tacgggggat tcctttccca 4980
ccgctccttc gctttccctt cctcgcccgc cgtaataaat agacaccccc tccacaccct 5040
ctttccccaa cctcgtgttg ttcggagcgc acacacacac aaccagatct cccccaaatc 5100
cacccgtcgg cacctccgct tcaaggtacg ccgctcgtcc tccccccccc ccctctctac 5160
cttctctaga tcggcgttcc ggtccatgca tggttagggc ccggtagttc tacttctgtt 5220
catgtttgtg ttagatccgt gtttgtgtta gatccgtgct gctagcgttc gtacacggat 5280
gcgacctgta cgtcagacac gttctgattg ctaacttgcc agtgtttctc tttggggaat 5340
cctgggatgg ctctagccgt tccgcagacg ggatcgattt catgattttt tttgtttcgt 5400
tgcatagggt ttggtttgcc cttttccttt atttcaatat atgccgtgca cttgtttgtc 5460
gggtcatctt ttcatgcttt tttttgtctt ggttgtgatg atgtggtctg gttgggcggt 5520
cgttctagat cggagtagaa ttctgtttca aactacctgg tggatttatt aattttggat 5580
ctgtatgtgt gtgccataca tattcatagt tacgaattga agatgatgga tggaaatatc 5640
gatctaggat aggtatacat gttgatgcgg gttttactga tgcatataca gagatgcttt 5700
ttgttcgctt ggttgtgatg atgtggtgtg gttgggcggt cgttcattcg ttctagatcg 5760
gagtagaata ctgtttcaaa ctacctggtg tatttattaa ttttggaact gtatgtgtgt 5820
gtcatacatc ttcatagtta cgagtttaag atggatggaa atatcgatct aggataggta 5880
tacatgttga tgtgggtttt actgatgcat atacatgatg gcatatgcag catctattca 5940
tatgctctaa ccttgagtac ctatctatta taataaacaa gtatgtttta taattatttt 6000
gatcttgata tacttggatg atggcatatg cagcagctat atgtggattt ttttagccct 6060
gccttcatac gctatttatt tgcttggtac tgtttctttt gtcgatgctc accctgttgt 6120
ttggtgttac ttctgcaggt cgactctaga ggatcaattc gctagcgaag ttcctattcc 6180
gaagttccta ttctctagaa agtataggaa cttcagatcc accgggatcc acacgacacc 6240
atgtcccccg agcgccgccc cgtcgagatc cgcccggcca ccgccgccga catggccgcc 6300
gtgtgcgaca tcgtgaacca ctacatcgag acctccaccg tgaacttccg caccgagccg 6360
cagaccccgc aggagtggat cgacgacctg gagcgcctcc aggaccgcta cccgtggctc 6420
gtggccgagg tggagggcgt ggtggccggc atcgcctacg ccggcccgtg gaaggcccgc 6480
aacgcctacg actggaccgt ggagtccacc gtgtacgtgt cccaccgcca ccagcgcctc 6540
ggcctcggct ccaccctcta cacccacctc ctcaagagca tggaggccca gggcttcaag 6600
tccgtggtgg ccgtgatcgg cctcccgaac gacccgtccg tgcgcctcca cgaggccctc 6660
ggctacaccg cccgcggcac cctccgcgcc gccggctaca agcacggcgg ctggcacgac 6720
gtcggcttct ggcagcgcga cttcgagctg ccggccccgc cgcgcccggt gcgcccggtg 6780
acgcagatct gagtcgaaac ctagacttgt ccatcttctg gattggccaa cttaattaat 6840
gtatgaaata aaaggatgca cacatagtga catgctaatc actataatgt gggcatcaaa 6900
gttgtgtgtt atgtgtaatt actagttatc tgaataaaag agaaagagat catccatatt 6960
tcttatccta aatgaatgtc acgtgtcttt ataattcttt gatgaaccag atgcatttca 7020
ttaaccaaat ccatatacat ataaatatta atcatatata attaatatca attgggttag 7080
caaaacaaat ctagtctagg tgtgttttgc gaatgcggcc ctagcgtata cgaagttcct 7140
attccgaagt tcctattctc cagaaagtat aggaacttct gtacacctga gctgattccg 7200
atgacttcgt aggttcctag ctcaagccgc tcgtgtccaa gcgtcactta cgattagcta 7260
atgattacgg catctaggac cgactagcta actaactagt acgtagaatt aattcattcc 7320
gattaatcgt ggcctcttgc tcttcaggat gaagagctat gtttaaacgt gcaagcgcta 7380
ctagacaatt cagtacatta aaaacgtccg caatgtgtta ttaagttgtc taagcgtcaa 7440
tttgtttaca ccacaaagta gacaagttgt gttcaa 7476
<210> 15
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 15
ccatctgagg tctgcactct cac 23
<210> 16
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ОЛИГОНУКЛЕОТИД
<400> 16
ctccgctcat gatcagattg tc 22
<210> 17
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> зонд
<400> 17
aacacactca aacac 15
<210> 18
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 18
ggatgctccg cactgtcaa 19
<210> 19
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 19
aagaaagctg ggtcggcg 18
<210> 20
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> зонд
<400> 20
tctcgaaagg gtgg 14
<210> 21
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 21
catcgtgaac cactacatcg agac 24
<210> 22
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 22
gtcgatccac tcctgcgg 18
<210> 23
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> зонд
<400> 23
accgtgaact tccgcaccga gc 22
<210> 24
<211> 105
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ампликон
<400> 24
ccatctgagg tctgcactct caccggtagt acagcacaaa caacacactc aaacactgat 60
agtttaaact gaaggcggga aacgacaatc tgatcatgag cggag 105
<210> 25
<211> 93
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ампликон
<400> 25
ggatgctccg cactgtcaag taacaggtga ggtcttccca gtgtagtttt gcagctgatc 60
tcgaaagggt gggcgcgccg acccagcttt ctt 93
<210> 26
<211> 76
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ампликон
<400> 26
catcgtgaac cactacatcg agacctccac cgtgaacttc cgcaccgagc cgcagacccc 60
gcaggagtgg atcgac 76
<210> 27
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигомер
<400> 27
ttggactaga aatctcgtgc tga 23
<210> 28
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигомер
<400> 28
gctacatagg gagccttgtc ct 22
<210> 29
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> зонд
<400> 29
gcgtttgtgt ggattg 16
<210> 30
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ампликон
<400> 30
ttggactaga aatctcgtgc tgattaattg ttttacgcgt gcgtttgtgt ggattgtagg 60
acaaggctcc ctatgtagc 79
<210> 31
<211> 435
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Соединение
<400> 31
tacttatatg ggctctgccg ttggagatga tgggcaaggc cttccgagga gccgagatga 60
ggatttaggc cttccattga gttgcgctcg ggccgtcctc ggtggcacga accagtgggc 120
caacggggcc aacgggccta actaatcgtg tctgatggat ggtcctccgt tccagagaaa 180
tcagacctgc tggctgtggg cctgtggctg aggagccggc agtgatccat cgagtgctac 240
tgttttcttt tttcagaaca gcgtgcagca aaggctcact gtgcagttta aacacctaaa 300
agacccgaca agtcggtgtt ggtactgagg acgaacccag accgaatgat tgcctcgccc 360
tcaccaccta tttcttccca tctgaggtct gcactctcac cggtagtaca gcacaaacaa 420
cacactcaaa cactg 435
<210> 32
<211> 417
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Соединение
<400> 32
tacaccacaa agtagacaag ttgtgttcaa accgcggatc agaaacaaat aaagcgaccc 60
cgcatgcacg tcaggtacca acccaaatcg ggaccctccc cccggctccg gccggtgtct 120
cccaagttaa acggccgccc gcgccaataa ttaccaccac accccggtag agagatggag 180
tagaagaaga aaaaaaatca tccgccaagc cgcgatccgc gaccgcagcg catcgacggc 240
gcacccccct cccctccggt cgtcggtcca cagcagacag gccgcgtcat tgcagattac 300
tgcaccaccg catcgcatca tcgcagcggc ccgcatgaac gaacacgagc ccccattttc 360
cgcgcgggtt gccgagccgg cagcccagcg cggacacggg gagctagcgg cggtcag 417
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ КУКУРУЗЫ DBN9501 И СПОСОБ ЕГО ВЫЯВЛЕНИЯ | 2020 |
|
RU2815214C2 |
| СПОСОБЫ БОРЬБЫ С СОРНЯКАМИ С ИСПОЛЬЗОВАНИЕМ РАСТЕНИЙ AAD-1 И ПРЕДПОСЕВНОГО И/ИЛИ ДОВСХОДОВОГО ВНЕСЕНИЯ ГЕРБИЦИДОВ | 2012 |
|
RU2610664C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| РАСТЕНИЕ КУКУРУЗЫ DBN9858, УСТОЙЧИВОЕ К ГЕРБИЦИДУ, И НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СПОСОБ ДЛЯ ЕЕ ВЫЯВЛЕНИЯ | 2016 |
|
RU2712730C2 |
| УСТОЙЧИВЫЙ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ И ТОЛЕРАНТНЫЙ К ГЕРБИЦИДАМ СЕЛЕКЦИОННЫЙ ГИБРИД ТРАНСФОРМАНТА СОИ pDAB9582.814.19.1 и pDAB4468.04.16.1 | 2012 |
|
RU2627161C2 |
| РАСТЕНИЯ-ТРАНСФОРМАНТЫ КУКУРУЗЫ PV-ZMIR13 (MON863) И КОМПОЗИЦИИ И СПОСОБЫ ИХ ОБНАРУЖЕНИЯ | 2003 |
|
RU2352638C2 |
| ДЕТЕКЦИЯ AAD-1 ОБЪЕКТА DAS-40278-9 | 2010 |
|
RU2577143C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ СОИ DBN8002 И СПОСОБ ЕЕ ВЫЯВЛЕНИЯ | 2019 |
|
RU2818368C2 |
| СОБЫТИЕ DAS-40278-9 AAD-1, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ КУКУРУЗЫ И ИХ СОБЫТИЕ-СПЕЦИФИЧЕСКАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2614120C2 |
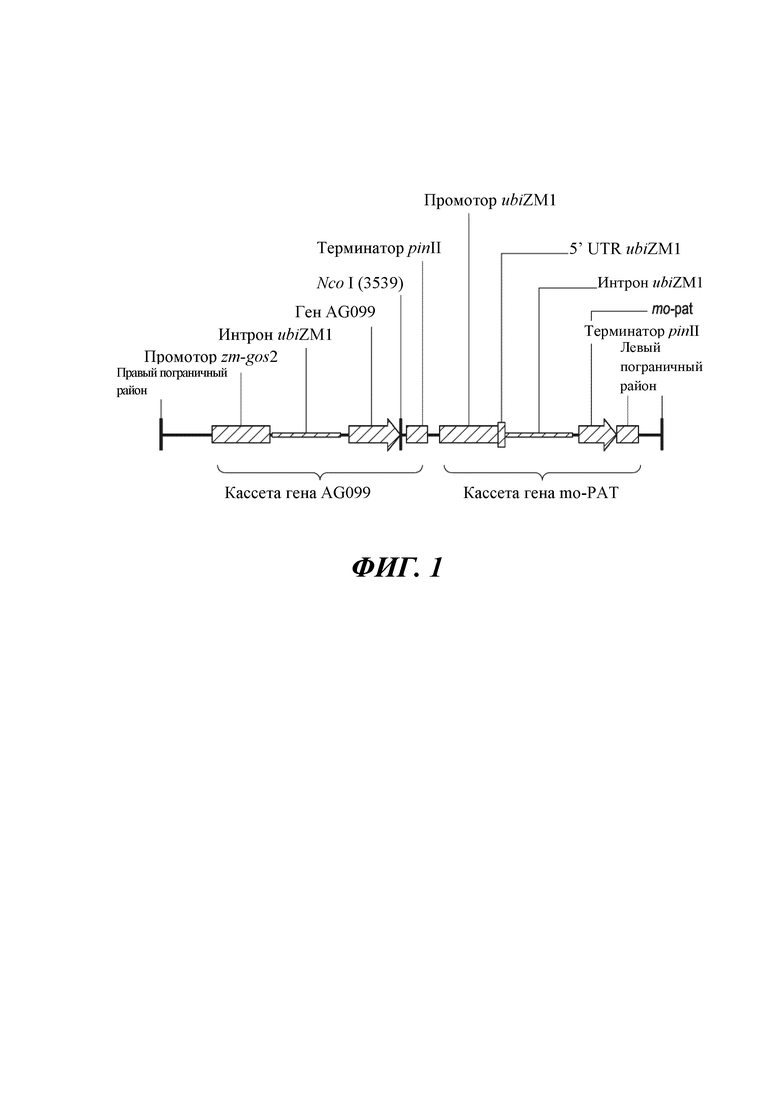


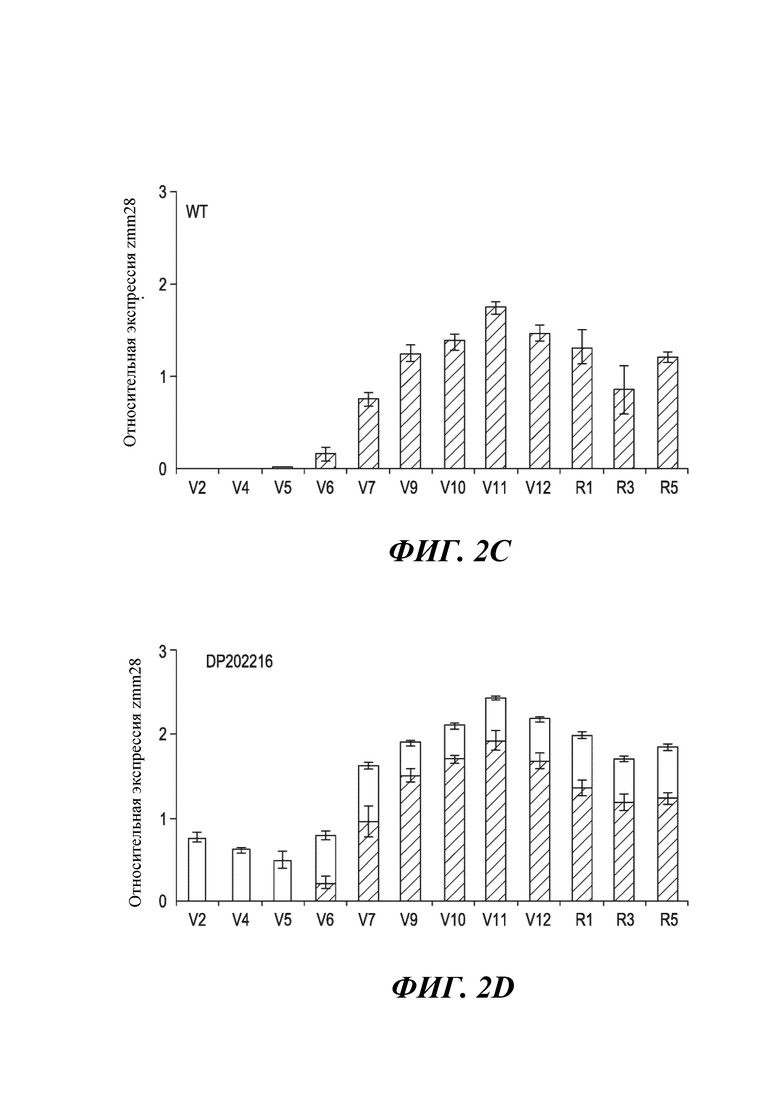
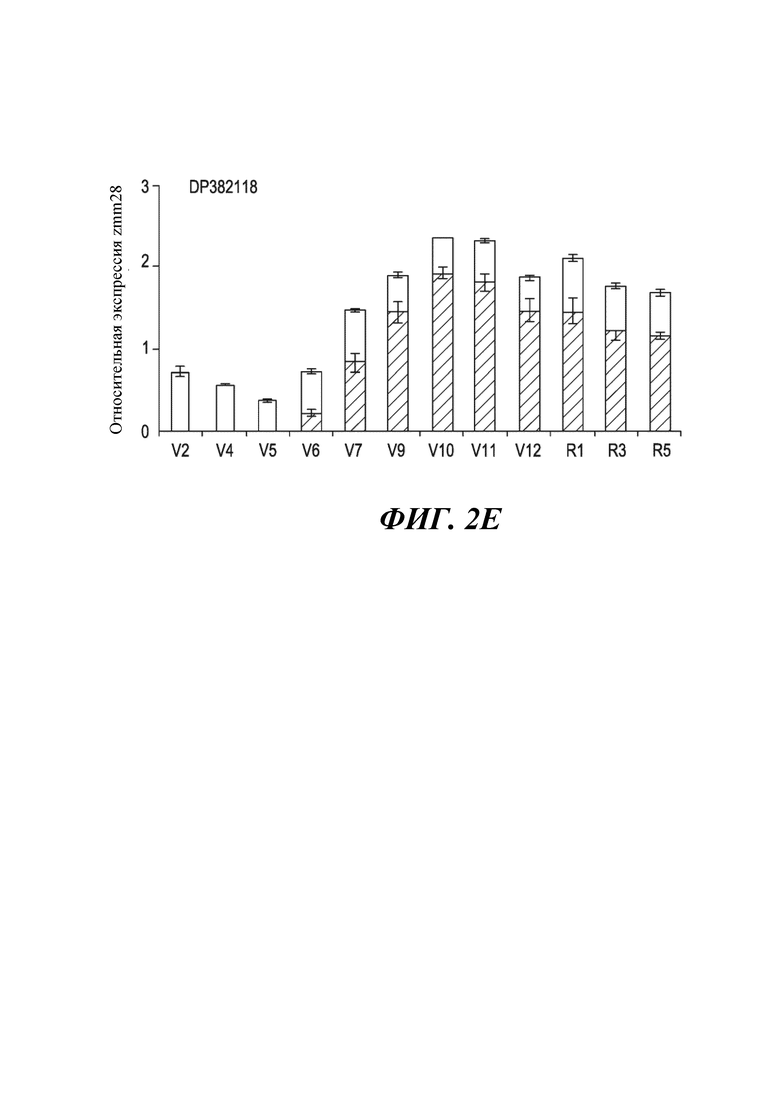
Изобретение относится к области биохимии, в частности к трансгенному растению кукурузы. Также раскрыты трансгенное семя кукурузы, биологический образец. Также раскрыт способ получения товарного продукта. Изобретение позволяет эффективно получать трансгенное растение кукурузы, которое проявляет повышенную активность ферментов фотосинтеза. 4 н. и 4 з.п. ф-лы, 17 табл., 13 пр., 6 ил.
1. Трансгенное растение кукурузы, содержащее объект кукурузы, где объект содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 14, в которой последовательности соединения представлены SEQ ID NO: 9 и 10, где репрезентативный образец семени указанного объекта кукурузы был депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653, и при этом трансгенное растение кукурузы проявляет повышенную активность ферментов фотосинтеза по сравнению с контрольным растением кукурузы, не содержащим объект.
2. Трансгенное семя кукурузы для получения трансгенного растения кукурузы, которое проявляет повышенную активность ферментов фотосинтеза по сравнению с контрольным растением кукурузы, не содержащим объект кукурузы, где указанное трансгенное семя кукурузы содержит объект кукурузы, где указанное семя содержит молекулу ДНК, изложенную под SEQ ID NO: 14, в которой последовательности соединения представлены SEQ ID NO: 9 и 10, и где репрезентативный образец семени указанного объекта кукурузы депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653.
3. Биологический образец, полученный из объекта трансгенного растения кукурузы по п. 1, или ткани или семени трансгенного растения кукурузы, для получения трансгенного растения кукурузы, которое проявляет повышенную активность ферментов фотосинтеза по сравнению с контрольным растением кукурузы, не содержащим объект кукурузы, где указанный образец содержит объект кукурузы, включающий нуклеотидную последовательность, изложенную под SEQ ID NO: 14, в которой последовательности соединения представлены SEQ ID NO: 9 и 10, или является комплементарной ей, где указанная нуклеотидная последовательность обнаруживается в указанном образце с применением способа амплификации нуклеиновой кислоты или гибридизации нуклеиновой кислоты, где репрезентативный образец семени указанного объекта кукурузы депонирован в Американской коллекции типовых культур (АТСС) под номером доступа PTA-124653.
4. Биологический образец по п. 3, где биологический образец предусматривает растение, ткань или части семени, околоплодника семени трансгенного объекта кукурузы.
5. Биологический образец по п. 3, где биологический образец представляет собой образец ДНК, экстрагированный из трансгенного растения, содержащего объект кукурузы, и где указанный образец ДНК содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 14, в которой последовательности соединения представлены SEQ ID NO: 9 и 10.
6. Биологический образец по п. 3, где указанный биологический образец выбран из группы, состоящей из кукурузной муки, кукурузной крупы, кукурузного сиропа, кукурузного масла, кукурузного крахмала и злаковых продуктов, изготовленных так, что они полностью или частично содержат сопутствующие продукты из кукурузы, где указанный биологический образец содержит обнаруживаемое количество указанной нуклеотидной последовательности.
7. Способ получения товарного продукта, содержащего молекулу рекомбинантной ДНК, содержащую нуклеотидную последовательность, изложенную под SEQ ID NO: 14, в которой последовательности соединения представлены SEQ ID NO: 9 и 10, при этом способ включает: (a) получение растения кукурузы, содержащего трансгенный объект кукурузы; и (b) изготовление товарного продукта кукурузы из растения кукурузы.
8. Способ по п. 7, в котором указанный товарный продукт выбран из группы, состоящей из цельных или переработанных семян, корма для животных, масла, крупы, муки, хлопьев, отрубей, биомассы и топливных продуктов.
| WO 2016144688 A1, 15.09.2016 | |||
| WO 2011075590 A1, 23.06.2011 | |||
| ВАРИАНТ КУКУРУЗЫ MIR604 | 2005 |
|
RU2351122C2 |

