Настоящее изобретение относится к конструкции нуклеиновой кислоты, кодирующей в указанном порядке первый антиген, гемагглютинин (HA), свиного гриппа A (IAV-S) линии Scot/94 и второй антиген, гемагглютинин (HA), свиного гриппа A (IAV-S) линии, подобной евразийскому штамму птичьего происхождения (EA), и к конструкции нуклеиновой кислоты, кодирующей в указанном порядке первый антиген HA IAV-S линии Gent/84 и второй антиген HA IAV-S линии pandemic09 (pdm09). В других вариантах осуществления настоящее изобретение относится к частицам РНК-репликона, содержащим одну или обе конструкции нуклеиновых кислот, к иммуногенной композиции, содержащей частицы репликона, такой как вакцина, которая может быть использована против инфекции, вызванной вирусом гриппа А. Также представлены способы получения вакцины и применение вакцины.
УРОВЕНЬ ТЕХНИКИ
Вирусы гриппа А (IAV) создают значительную нагрузку на здоровье людей и животных во всем мире. IAV подразделяется на разные подтипы на основе поверхностных гликопротеинов, гемагглютинина (HA) и нейраминидазы (NA), вируса. IAV поражает домашнюю птицу, свиней, лошадей, кошек, собак, морских млекопитающих (например, китов), летучих мышей и людей. Дикие водоплавающие и береговые виды (утки, гуси, лебеди и чайки) являются естественными резервуарами и могут быть инфицированы 16 разными подтипами HA и 9 разными подтипами NA [Webster et al., Microbiol Rev 56:152-179 (1992)].
Вирус гриппа А у свиней (IAV-S) является серьезным респираторным патогеном домашних свиней, который признан во всем мире как наносящий существенный экономический ущерб, особенно в сфере индустриального животноводства [Holtkamp et al., The American Association of Swine Veterinarians Annual Meeting (2007)]. Он характеризуется внезапным началом респираторного заболевания и обычно сопровождается анорексией, вялостью и лихорадкой. В дополнение к клиническим осложнениям у продуктивных животных, связанным с IAV-S, имеются сообщения об участии свиней в передаче вируса гриппа людям [Myers KP, Olsen CW, Gray GC. Clin Infect Dis 2007; 44(8):1084-8, Krueger and Gray, Curr Top Microbiol Immunol 370: 201-225 (2013)], что представляет собой серьезную угрозу для здоровья населения и создает еще более высокий стимул для борьбы с IAV в стадах свиней.
В ответ на эту проблему многие свиноводы в настоящее время вакцинируют своих свиней против IAV-S, используя имеющиеся в продаже вакцины. Однако контролировать IAV-S с помощью обычных вакцин сложно, поскольку в полевых условиях одновременно циркулирует множество разных штаммов IAV-S, которые продолжают эволюционировать [Gao et al., J Gen Virol 98(8):2001-2010 (2017)]. Разнообразие и мутабельность IAV-S обусловлены генетической структурой вируса. Как и другие вирусы гриппа А, IAV-S имеет гены, закодированные в восьми сегментах РНК, и механизм репликации генома, который приводит к частым мутациям. Эти генетические характеристики позволяют IAV-S быстро адаптироваться, в том числе ускользать от существующих нейтрализующих антител, продуцированных в результате воздействия предыдущих штаммов. Следовательно, в связи с появлением новых штаммов, возникающих в результате постоянного антигенного дрейфа и/или изменчивости, инактивированные вирусные вакцины против IAV-S, коммерчески доступные на рынке США, оказались неадекватными, несмотря на то что они содержат до пяти разных штаммов IAV-S.
Классификация вирусов гриппа А начинается с субтипирования HA и NA, двух основных гликопротеинов на поверхности вируса. Белок HA опосредует прикрепление и слияние вируса с клетками-хозяевами. Нейраминидаза представляет собой фермент, который действует на заключительной стадии цикла репликации вируса гриппа путем отщепления вновь образованных вирусных частиц от клетки-хозяина, тем самым обеспечивая распространение нового потомства вируса и инфицирование других клеток. Недавние исследования показали, что иммунитет к NA может играть только дополнительную и/или комплементарную роль по отношению к более важному иммунитету к HA [Nayak et al., J Virol 84(5):2408-2420 (2010); Pavlova et al., Vaccine 27(5):773-785 (2009); Sylte et al., Vaccine 25(19):3763-72 (2007)]. Действительно, оказывается, что в отсутствие антигена, гемагглютинина, вакцина на основе нейраминидазы вируса гриппа А недостаточно эффективна ни для защиты от заражения гриппом А, ни для защиты от заболевания, вызванного вирусом гриппа А.
Тогда как во время определенного сезона гриппа человеческий грипп А обычно имеет 1 или 2 доминирующих штамма, циркулирующих во всем мире, количество одновременно циркулирующих штаммов IAV-S намного больше, при этом эти штаммы различаются в зависимости от географического региона. Аналогично, штаммы IAV-S также антигенно вариабельны, но в основном содержат подтип H1 или H3 HA и подтип N1 или N2 NA. Внутри каждого подтипа HA и NA IAV-S существует дополнительное филогенетическое разнообразие.
В популяции свиней в США преобладают четыре филогенетических кластера H1 (гамма, дельта1, дельта2, пандемический), два кластера H3 (кластер IV и человекоподобный), два кластера N1 (классический, пандемический) и два кластера N2 (N2-1998 и N2-2002). [См. Anderson et al., Influenza and other Respiratory Viruses 7 (Suppl. 4); 42-51 (2013); и Anderson et al., mSphere 1(6) e00275-16:1-14 (2016)].
В Европе существуют три основные линии H1 (H1, подобная евразийскому штамму птичьего происхождения, H1, подобная Scotland/410440/1994, и H1, подобная Pandemic_2009), одна основная линия H3 (H3, подобная Gent/1/1984), две основные линии N1 (N1, подобная евразийскому штамму птичьего происхождения, и N1, подобная Pandemic_2009), две основные линии N2 (N2, подобная Gent/1/1984, и N2, подобная Scotland/410440/1994) и две второстепенные линии N2 (N2, подобная Italy/4675/2003, и N2, подобная вирусу человеческого сезонного гриппа) [Watson et al., J. Virol., 89:9920-9931 (2015); doi:10.1128/ОВИ.00840-15].
Вакцинация против IAV-S является наилучшим вариантом для снижения клинических осложнений у свиней, а также снижения возможности дополнительной реассортации вируса и его зоонозного распространения от свиней к человеку. До недавнего времени единственными вакцинами, доступными для широкого применения, были инактивированные вакцины, приготовленные из вирусов гриппа, выращенных в яйцах с эмбрионами, но их поставки в значительной степени ограничены из-за нехватки яиц, свободных от конкретных патогенов, и полностью признана необходимость в новых подходах к вакцинам против гриппа.
В случае обычных инактивированных вирусных вакцин против IAV-S выбор вирусных штаммов основан на свойствах антигена HA. Вакцины IAV-S, которые индуцируют титры антител, ингибирующих НА (HI), защищают свиней от экспериментального заражения антигенно сходным штаммом [Kyriakis et al., Vet Microbiol 144(1-2):67-74 (2010)]. Однако относительно быстрый генетический дрейф генов HA способствует появлению новых штаммов, которые функционально не ингибируются антителами HA, индуцированными вакциной.
Как следствие, имеющиеся в продаже вакцины часто не защищают от новых и появляющихся подтипов/кластеров вирусов и обеспечивают лишь ограниченную защиту от гетеросубтипического заражения, поскольку антигены не соответствуют всем современным штаммам, циркулирующим в полевых условиях [Lee et al., Can J. Vet Res 71(3):207-12(2007); Vincent et al., Vaccine 28(15):2782-2787 (2010)]. Таким образом, такие вакцины необходимо периодически обновлять, чтобы они соответствовали циркулирующим в настоящее время штаммам.
Следовательно, в данной области техники существует потребность в разработке новых вакцин против IAV-S, которые являются безопасными, эффективными и могут быть быстро изменены для достижения антигенного соответствия появляющемуся штамму.
Поскольку большинство вирусов, таких как вирусы гриппа, имеют относительно простую структуру, использование одного антигена из их антигенного профиля иногда может быть достаточным для формирования защитного иммунного ответа. Такую субъединичную вакцину можно получить путем экстракции специфического антигена из вируса или его культуры или путем рекомбинантной экспрессии такого специфического антигена. Альтернативно, антиген вируса может быть доставлен целевому животному и экспрессирован в его организме с помощью живого рекомбинантного микроорганизма-носителя, который действует в качестве вектора. Векторы могут быть живыми ослабленными или неживыми. На протяжении многих лет для вакцин применялся ряд стратегий, основанных на векторах, с целью защиты от определенных патогенов.
Разновидностью использования вакцин на основе вирусного вектора является использование вакцин на основе частиц репликона [RP; см. Lundstrom, 2014, Vaccines, vol. 6, р. 2392-2415]. Такие частицы являются вирусоподобными частицами, но содержат дефектный вирусный геном и, как правило, гетерологичный ген. Эти частицы репликона обычно содержат РНК, упакованную в частицы (т.е. они инкапсулированы), благодаря чему они способны проникать в клетку-хозяина целевого животного и выполнять один цикл амплификации вирусного генома без способности образовывать новые частицы. Частица репликона не распространяется из инфицированной клетки, поскольку в ней отсутствует(ют) необходимая(ые) последовательность(и), кодирующая(ие) структурный белок. Как таковые, эти частицы похожи на вирус дикого типа (например, с точки зрения тропизма) больше, чем другие вакцины на основе репликона, такие как вакцины с голой РНК или вакцины, содержащие РНК, сгенерированную из ДНК-плазмиды.
Геном RP обычно экспрессирует гетерологичный ген, кодирующий иммунозащитный антиген. Наиболее широко используемыми и наиболее изученными являются частицы РНК-репликона альфавируса [Vander Veen et al., 2012, Anim. Health. Res. Rev., vol. 13, p. 1-9; and: Kamrud et al., 2010, J. Gen. Virol., vol. 91, p. 1723-1727], которые поэтому предпочтительны из практических соображений и которые были получены из вирусных геномов путем замены генов структурных белков гетерологичными генами. Полученные РНК, называемые репликонами, способны управлять собственной репликацией и экспрессировать высокие уровни гетерологичного гена при введении в цитоплазму клеток-хозяев. Поскольку в этих репликонах отсутствуют гены структурных белков альфавируса, они не способны образовывать вирионы и распространяться на соседние клетки. Однако репликоны могут быть эффективно упакованы в частицы репликона (RP) вируса путем введения их в клетки, в которых структурные белки находятся в транс-форме [Pushko et al., 1997, Virology, vol. 239, с. 389-401].
Кроме того, считается, что RP альфавируса являются несколько более сильными иммуностимулирующими средствами, чем другие RP, известные в данной области и основаны на других вирусах, таких как буньявирус. Для разработки RP-вакцин было использовано несколько видов альфавирусов, например: вирус венесуэльского энцефалита лошадей (VEEV) [Pushko et al., 1997, Virology, vol. 239, p. 389-401], вирус Синдбис [Bredenbeek et al., 1993, J. Virol., vol. 67, p. 6439-6446] и вирус леса Семлики [Liljestrom & Garoff, 1991, Biotechnology (NY), vol. 9, p. 1356-1361].
RP-вакцины могут вызывать слизистые и системные иммунные реакции после иммунизации целевого животного [Davis et al., 2002, IUBMB Life, vol. 53, p. 209-211]. RP-вакцины на основе VEE также являются основой нескольких вакцин, лицензированных Министерством сельского хозяйства США (USDA), которые включают: вакцину против эпидемической диареи свиней, РНК (код продукта 19U5.P1), вакцину против свиного гриппа, РНК (код продукта 19A5.D0), вакцину против птичьего гриппа, РНК (код продукта 19O5.D0) и рецептурный продукт, частица РНК (код продукта 9PP0.00).
Поскольку векторной системой на основе RP можно легко манипулировать на молекулярном уровне, это обеспечивает возможность быстрого производства вакцин, что позволяет реагировать на появляющиеся подтипы вируса.
Таким образом, существует постоянная потребность в новых вакцинах, которые обеспечивают широкий спектр защиты от циркулирующих штаммов IAV-S, в частности, обеспечивают широкий спектр защиты от большинства или всех четырех основных штаммов IAV-S, циркулирующих в Европе: EurAsianAvian H1N1, Gent84 H3N2, Scot/94 H1N2 и Pandemic_2009 H1N1, которые могут быть быстро адаптированы для ответа на появляющиеся подтипы вируса и антигенный дрейф.
Однако векторная система на основе RP, такая как платформа на основе репликона альфавируса, не позволяет встраивать любое требуемое количество антигенов, например, встраивать все гены NA и HA четырех основных циркулирующих штаммов IAV-S в вектор на основе репликона, чтобы добиться самой широкой защиты. Платформы на основе альфавирусного вектора обычно представляют собой трехкомпонентную систему, состоящую из РНК, содержащей неструктурные гены с ассоциированным с ними сигналом упаковки, и структурных белков, удаленных и замененных последовательностями гетерологичных генов. Две хелперные РНК содержат структурные белки вируса без сигнала упаковки. Количество РНК, которое может быть упаковано при использовании этих трехкомпонентных систем, ограничено объемом вирусного капсида [Nanda K. et al., Vol. 390(2), 2009, 368-373]. Это ограничение, присущее векторной системе на основе RP, не позволяет удовлетворять постоянный спрос на вакцину, способную обеспечить широкий спектр защиты от большинства или всех циркулирующих штаммов IAV-S.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В первом аспекте настоящего изобретения неожиданно было обнаружено, что в случае введения более одного антигена, гемагглютинина, вируса свиного гриппа А (HA IAV-S) положение генов, кодирующих антигены HA, в вирусном геноме частицы РНК-репликона оказывает сильное влияние на уровень индуцированного иммунитета.
Соответственно, настоящее изобретение относится к конструкциям нуклеиновых кислот, которые кодируют комбинацию двух антигенов НА IAV-S из разных линий в определенном порядке. Эти конструкции нуклеиновых кислот можно использовать в частицах РНК-репликона. Эти частицы РНК-репликона по настоящему изобретению могут быть использованы в иммуногенных композициях для получения вакцин для применения для профилактики возникновения заболевания, вызываемого вирусом свиного гриппа А (IAV-S), у вакцинированного субъекта (например, человека, животного-компаньона или домашнего скота, в частности свиньи).
В первом варианте осуществления настоящего изобретения конструкция нуклеиновой кислоты содержит комбинацию антигена HA IAV-S линии Scot/94 и линии, подобной евразийскому штамму птичьего происхождения (EA), где HA IAV-S линии Scot/94 расположен первым (в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты), и HA IAV-S линии ЕА расположен вторым. Термин «в направлении от 5' к 3'-концу», также известный как «в нисходящем направлении», хорошо известен в данной области. Вместе с терминами «в таком порядке» он служит для обозначения относительной ориентации, которую должны иметь указанные за ним элементы по отношению друг к другу, чтобы они могли функционировать с аппаратом экспрессии генов клетки-хозяина, т.е. в которой RP по изобретению, содержащая конструкцию нуклеиновой кислоты, может быть реплицирована и экспрессирована. Как будет понятно специалисту в данной области техники, в данном случае это направление относится к цепи нуклеиновой кислоты генома, которая является «кодирующей цепью». Гены могут присутствовать в последовательном порядке в направлении от 5' к 3'-клнцу, т.е. в конструкции нет промежуточных генов для экспрессии в белки. В этом случае конструкция нуклеиновой кислоты обычно содержит в порядке от 5' к 3'-концу открытую рамку считывания неструктурного белка остова вируса, субгеномный промотор, за которым следуют последовательность гена первого антигена HA, интерстициальная последовательность, последовательность второго субгеномного промотора, за которой следуют ген второго антигена HA и, наконец, 3'-нетранслируемая область остова вируса.
Таким образом, настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген, гемагглютинин (HA), IAV-S линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
Во втором варианте осуществления настоящего изобретения конструкция нуклеиновой кислоты содержит комбинацию антигена НА IAV-S линии Gent/84 и линии pdm09, где НА IAV-S линии Gent/84 расположен первым (в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты), и HA IAV-S линии pdm09 расположен вторым. Таким образом, настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген HA IAV-S линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09).
Во втором аспекте настоящего изобретения неожиданно было обнаружено, что гемагглютинин вируса свиного гриппа А (HA IAV-S) определенных штаммов четырех основных циркулирующих линий IAV-S может обеспечивать улучшенный иммунитет против IAV-S по сравнению с другими штаммами. В частности, было обнаружено, что специфические комбинации HA IAV-S могут обеспечивать улучшенный иммунитет. Таким образом, такие комбинации HA IAV-S могут быть успешно использованы в конструкциях нуклеиновых кислот, которые могут быть включены в частицы РНК-репликона. Эти частицы РНК-репликона можно использовать в качестве иммуногенных композиций для получения вакцин, которые способствуют защите вакцинированного субъекта (например, человека, животного-компаньона или домашнего скота, в частности свиней) от IAV-S, например, позволяют предотвратить заражение вирусом IAV-S.
Таким образом, настоящее изобретение также относится к конструкциям нуклеиновых кислот, кодирующим комбинацию двух антигенов HA IAV-S конкретных штаммов, определенных в настоящем описании.
В первом варианте осуществления настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген HA IAV-S линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), из штамма A/swine/Italy/3033-1/2015 (H1N2), и
вторая последовательность нуклеиновой кислоты кодирует второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA), из штамма A/swine/Italy/28762-3/2013 (H1N1).
Во втором варианте осуществления настоящее изобретение относится к конструкции нуклеиновой кислоты для применения для профилактики или лечения у субъекта заболевания, вызванного вирусом свиного гриппа А, где конструкция нуклеиновой кислоты содержит первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген, гемагглютинин (HA), вируса свиного гриппа A (IAV-S) линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), из штамма A/swine/Italy/240849/2015 (H3N2), и
вторая последовательность нуклеиновой кислоты кодирует второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09) из штамма A/swine/England/373/2010 (H1N1).
В другом важном варианте осуществления предлагается частица РНК-репликона, содержащая конструкцию нуклеиновой кислоты по настоящему изобретению. Таким образом, частица РНК-репликона может содержать конструкцию нуклеиновой кислоты согласно первому или согласно второму варианту осуществления.
Настоящее изобретение включает любые комбинации вариантов осуществления первого и второго аспектов, представленных в данном описании. Таким образом, настоящее изобретение также относится к конструкциям нуклеиновых кислот, в которых антигены HA IAV-S расположены в определенном порядке, как определено в первом аспекте, и в которых антигены IAV-S получены из конкретных штаммов, как определено во втором аспекте.
В другом важном аспекте настоящее изобретение относится к частице РНК-репликона, содержащей конструкции нуклеиновых кислот, представленные в настоящем описании.
В другом важном аспекте настоящее изобретение относится к иммуногенной композиции, содержащей частицу РНК-репликона, представленной в настоящем описании.
В другом важном аспекте настоящее изобретение относится к иммуногенной композиции, содержащей комбинацию частиц РНК-репликона, где комбинация содержит первую частицу РНК-репликона, содержащую конструкцию нуклеиновой кислоты в соответствии с первым вариантом осуществления, и вторую частицу РНК-репликона, содержащую конструкцию нуклеиновой кислоты в соответствии со вторым вариантом осуществления.
Еще один вариант осуществления настоящего изобретения относится к вакцине, содержащей представленную в настоящем описании иммуногенную композицию.
В другом важном варианте осуществления вакцина по настоящему изобретению может быть использована для профилактики или лечения у субъекта заболевания, вызванного вирусом свиного гриппа А.
В другом важном варианте осуществления настоящее изобретение относится к способу иммунизации свиней против вируса свиного гриппа А, включающему введение свиньям иммунологически эффективного количества вакцины по настоящему изобретению.
В третьем аспекте неожиданно было обнаружено, что комбинация двух частиц РНК-репликона, каждая из которых содержит конструкцию нуклеиновой кислоты, кодирующую первый и второй антигены НА IAV-S разных линий, может обеспечить улучшенный иммунитет против IAV-S.
Таким образом, настоящее изобретение также относится к иммуногенной композиции, содержащей первую и вторую частицы РНК-репликона, где первая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены HA IAV-S, где
первый антиген HA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
второй антиген HA относится к линии A(H1N1)pdm09 (pdm09),
вторая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
четвертый антиген HA относится к линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
Настоящее изобретение включает любые комбинации вариантов осуществления третьего аспекта с вариантами осуществления первого и второго аспектов, представленных в настоящем описании. Таким образом, настоящее изобретение также относится к частицам репликона, описанным в третьем аспекте, где конструкции нуклеиновых кислот кодируют антигены HA IAV-S, которые расположены в определенном порядке, как определено в первом аспекте, и/или где антигены IAV-S получены из конкретных штаммов, как определено во втором аспекте.
В четвертом аспекте неожиданно было обнаружено, что конструкция нуклеиновой кислоты, содержащая конкретную комбинацию антигенов, нейраминидаз (NA), IAV-S трех разных линий, представленных в настоящем описании, может быть использована для обеспечения иммунитета против всех четырех основных циркулирующих линий IAV-S.
Таким образом, настоящее изобретение также относится к конструкции нуклеиновой кислоты, содержащей первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены NA IAV-S, где
первый антиген NA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94),
второй антиген NA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
третий антиген NA выбирают из линии A(H1N1)pdm09 (pdm09) или линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
В другом важном варианте осуществления настоящее изобретение относится к частице РНК-репликона, содержащей конструкцию нуклеиновой кислоты, представленную в четвертом аспекте.
В другом важном варианте осуществления настоящее изобретение относится к иммуногенной композиции, содержащей частицу РНК-репликона, описанную в четвертом аспекте.
Еще один вариант осуществления настоящего изобретения относится к вакцине, содержащей иммуногенную композицию, представленную в четвертом аспекте.
В другом важном варианте осуществления вакцина, описанная в четвертом аспекте, может быть использована для профилактики или лечения у субъекта заболевания, вызванного вирусом свиного гриппа А.
В другом важном варианте осуществления настоящее изобретение относится к способу иммунизации свиней против вируса свиного гриппа А, где способ включает введение свиньям иммунологически эффективного количества вакцины, описанной в четвертом аспекте.
В другом важном аспекте настоящее изобретение относится к иммуногенной композиции, содержащей комбинацию частиц РНК-репликона, где комбинация включает первую и вторую частицы РНК-репликона в соответствии с третьим аспектом, и третью частицу РНК-репликона, содержащую конструкцию нуклеиновой кислоты в соответствии с четвертым аспектом.
Настоящее изобретение включает любые комбинации вариантов осуществления четвертого аспекта с вариантами осуществления первого, второго и/или третьего аспектов, представленных в настоящем описании. Таким образом, настоящее изобретение также относится к частицам репликона, описанным в третьем аспекте, где конструкции нуклеиновых кислот кодируют антигены HA IAV-S, которые расположены в определенном порядке, как определено в первом аспекте, и/или где антигены IAV-S получены из конкретных штаммов, как определено во втором аспекте, в комбинации с частицами репликона, описанными в четвертом аспекте.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фиг. 1: Титры антител ингибирования гемагглютинации (HI), индуцированные частицей одногенной РНК, кодирующей один антиген HA IAV-S линии EurAsianAvian.
Фиг. 2: HI-титры антител, индуцированные частицей одногенной РНК, кодирующей один антиген HA IAV-S линии Scot1994.
Фиг. 3: HI-титры антител, индуцированные частицей одногенной РНК, кодирующей один антиген HA IAV-S линии Pdm2009.
Фиг. 4: HI-титры антител, индуцированные частицей одногенной РНК, кодирующей один антиген HA IAV-S линии Gent1984.
Фиг. 5: HI-титры антител, индуцированные частицей двухгенной РНК, кодирующей один антиген HA IAV-S линии EurAsianAvian (EUHA1-2, EUHA1-3 и EUHA1-5) и другой антиген HA из штаммов IAV-S линии Scot1994 (EUHA1-15 или EUHA1-17) в разных комбинациях.
Фиг. 6: HI-титры антител, индуцированные частицей двухгенной РНК, кодирующей один антиген HA линии Pandemic (EUHA1-11) и другой антиген линии Gent1984 (EUHA3-4) или один антиген HA линии Scot1994 (EUH1-15, EUHA1-17) и другой антиген HA IAS из штаммов линии EurAsianAvian (EUHA1-3 и EUHA1-5) в двух разных положениях.
Фиг. 7: Титры антител ингибирования нейраминидазы (NI), индуцированные частицей одногенной РНК, кодирующей один антиген NA IAV-S линии EurAsianAvian (EA).
Фиг. 8: Титры (NI) антител, индуцированные частицей одногенной РНК, кодирующей один антиген NA IAV-S линии Pdm09.
Фиг. 9: Титры (NI) антител, индуцированные частицей одногенной РНК, кодирующей один антиген NA IAV-S линии Scot/94.
Фиг. 10: Титры (NI) антител, индуцированные частицей одногенной РНК, кодирующей один антиген NA IAV-S линии Gent/84.
Фиг. 11: NI-титры антител, индуцированные частицами двухгенной РНК, кодирующими один антиген NA IAS линии EurAsianAvian (EUNA1-2) и другой антиген NA линии Gent1984 (EUNA2-7), или частицами трехгенной РНК, кодирующими один антиген NA из каждой из линии EurAsianAvian (EUNA1-2), линии Gent1984 (EUNA2-7) и штаммов линии Scot1994 (EUHNA2-6) в разных положениях.
Фиг. 12: Титры анти-NI антител, индуцированные частицами двухгенной РНК, кодирующими один антиген NA линии EurAsianAvian (EUNA1-2) и другой антиген NA IAV-S линии Gent1984 (EUNA2-7), или частицами трехгенной РНК, кодирующими один антиген NA из каждой из линии EurAsianAvian (EUNA1-2), штаммов линии Gent1984 (EUNA2-7) и линии Scot1994 (EUHNA2-6) или лини Pdm09 (EUNA1-4) в разных положениях.
Значение терминов:
Для более полного понимания изобретения ниже приведены следующие определения.
Конструкция нуклеиновой кислоты представляет собой искусственно сконструированный сегмент нуклеиновой кислоты (например, ДНК, РНК, мРНК), обычно предназначенный для трансплантации в целевую клетку.
Использование терминов в единственном числе для удобства описания никоим образом не предназначено для ограничения. Таким образом, например, ссылка на композицию, содержащую «полипептид», включает ссылку на один или более таких полипептидов. Кроме того, ссылка на «частицу РНК-репликона альфавируса» включает ссылку на множество таких частиц РНК-репликона альфавируса, если не указано иное.
В контексте настоящего описания термин «приблизительно» используется взаимозаменяемо с термином «примерно» и означает, что значение находится в пределах пятидесяти процентов указанного значения, т.е. композиция, содержащая «приблизительно» 1×108 частиц РНК-репликона альфавируса на миллилитр, содержит от 5×108 до 1,5×108 частиц РНК-репликона альфавируса на миллилитр.
В контексте настоящего описания термины «поросенок», «свинья» или «представитель свинообразных» используются взаимозаменяемо и включают все виды одомашненных свиней, если не указано иное.
В контексте настоящего описания термин «филогенетический кластер» представляет собой набор антигенов вируса гриппа, таких как гемагглютинины (HA) или нейраминидазы (NA), которые сгруппированы вместе (на одной и той же ветви) в филогенетическом дереве или эволюционном дереве, которое восходит корнями к подобному (гомологичному) предку. Для нейраминидаз и гемагглютининов IAV-S, обнаруженных в США, преобладающие филогенетические кластеры описаны в: [Anderson et al., Influenza and other Respiratory Viruses 7 (Suppl. 4):42-51 (2013)].
В контексте настоящего описания термин «линия» представляет собой набор гемагглютининов вируса гриппа, которые сгруппированы вместе (на одной ветви) в эволюционном древе, которое восходит корнями к подобному (гомологичному) предку. Эти группы определены для европейских гемагглютининов и нейраминидазы и аналогичны, но не эквивалентны, филогенетическим кластерам для вирусов США. Определения линий можно получить путем филогенетического анализа рассматриваемых последовательностей HA или NA с предварительно установленными эталонными последовательностями с помощью легкодоступного программного обеспечения, например, Clustal Omega [Sievers F., et al., (2011) Mol. Syst. Biol. 7:539] или с помощью доступного в Интернете инструмента аннотации для последовательностей H1 HA [Anderson TK, et al., mSphere, 2016; 1(6):e00275-16].
Для гемагглютинина (HA) IAV-S, обнаруженного в Европе, существуют четыре преобладающие линии, описанные в [Watson et al., J. Virol. 89:9920-9931 (2015)], которые соответствуют трем филогенетическим ветвям (кладам) H1 HA, описанным Anderson et al., mSphere 1(6):e00275-16 (2016), и одной кладе H3 HA [Anderson et al., не опубликовано]. Заражение европейских свиней исключительно вирусами линии CS происходило до 1979 года, когда у свиней в Бельгии и Германии был выделен вирус птичьего гриппа H1N1, названный «свиным H1N1, аналогичным евразийскому штамму птичьего происхождения» (EA), который генетически отличался от подтипа CS. Линия EA продолжает циркулировать среди европейских свиней и с момента своего появления подверглась реассортации с сезонными человеческими вирусами, что привело к совместной циркуляции трех различающихся линий вируса в Европе: (i) H1avN1, аналогичной евразийскому штамму птичьего происхождения (EA или клада 1C.2.); (ii) H3N2, аналогичной A/swine/Gent/1/1984 (Gent/84 или клада 3.1970.1); и (iii) H1huN2, аналогичной A/swine/Scotland/410440/1994 (Scot/94 или клада 1B.1). С апреля 2009 года среди населения распространяется новый вирус IAV H1N1, названный (iv) A(H1N1)pdm09 или кладой 1A.3.3.2 свиного происхождения. Таким образом, в контексте настоящего изобретения эти четыре линии упоминаются как «EA», «Gent/84», «Scot/94» и «pdm09».
В контексте настоящего описания термин «репликон» относится к модифицированному вирусному РНК-геному, в котором отсутствует один или более элементов (например, последовательностей, кодирующих структурные белки), которые, в случае их присутствия, обеспечили бы успешное размножение родительского вируса в клеточных культурах или животных-хозяевах. В подходящих клеточных контекстах репликон будет амплифицироваться и может продуцировать один или более видов субгеномной РНК.
В контексте настоящего описания термин «частица РНК-репликона», сокращенно «RP», представляет собой РНК-репликон, упакованный в структурные белки, например, капсид и гликопротеины, которые могут быть получены из альфавируса, например, представляет собой частицу РНК-репликона альфавируса, как описано Pushko et al., [Virology 239(2):389-401 (1997)], но также может быть вирусом Синдбис [Bredenbeek et al., 1993, J. of Virol., vol. 67, p. 6439-6446] и вирусом леса Семлики [Liljestrom & Garoff, 1991, Biotechnology (NY), vol. 9, p. 1356-1361]. RP не может размножаться в клеточных культурах или в организме животных-хозяев (без хелперной плазмиды или аналогичного компонента), поскольку репликон не кодирует структурные компоненты альфавируса (например, капсид и гликопротеины). Предпочтительно, РНК-RP по настоящему изобретению представляет собой РНК-RP альфавируса.
Термин «не-IAV-S» используется для модификации таких терминов, как патоген и/или антиген (или иммуноген) для обозначения того, что соответствующий патоген и/или антиген (или иммуноген) не является ни патогеном IAV-S, ни антигеном (или иммуногеном) IAV-S и что не-IAV-S белковый антиген (или иммуноген) не происходит от IAV-S.
Термин «происходить(происходит) от» используется в настоящем описании для обозначения того, что немодифицированная и/или укороченная аминокислотная последовательность рассматриваемого белкового антигена кодируется этим патогеном или штаммом этого патогена. Кодирующая последовательность в конструкции нуклеиновой кислоты по настоящему изобретению для белкового антигена, происходящего от патогена, могла быть подвергнута генетическим манипуляциям, что привело к модификации и/или укорочению аминокислотной последовательности экспрессированного белкового антигена относительно соответствующей последовательности этого белкового антигена в патогене или штамме патогена (включая естественно аттенуированные штаммы), от которого она происходит.
В контексте настоящего описания термины «лечение» или «лечащий», «профилактика» или «предупреждение», «защита» или «обеспечение защиты» или «индуцирование защитного иммунитета к», «способствовать предотвращению заболевания» и «способствовать защите» не требуют полной защиты от каких-либо признаков инфекции. Например, «для применения в целях профилактики» может означать, что обеспечиваемая защита достаточна для по меньшей мере уменьшения симптомов основной инфекции после заражения и/или для уменьшения и/или устранения одной или более основных клеточных, физиологических или биохимических причин или механизмов, вызывающих эти симптомы. Понятно, что «уменьшение», как используется в этом контексте, относится к состоянию инфекции, включая молекулярное состояние инфекции, а не только физиологическое состояние инфекции. Следовательно, термины «профилактика заболевания» или «лечение» охватывают профилактическое лечение инфекции, вызванной вирусом, или расстройства, возникающего в результате инфекции.
В контексте настоящего описания термин «вакцина» представляет собой композицию, подходящую для применения по отношению к животному, например свинье (включая, в некоторых вариантах осуществления, людей, тогда как в других вариантах осуществления композиция специально не предназначена для людей), содержащую один или более антигенов, обычно в комбинации с фармацевтически приемлемым носителем, таким как содержащая воду жидкость, которая при введении животному индуцирует иммунный ответ, достаточно сильный, чтобы по меньшей мере способствовать защите от заболевания, возникающего в результате инфекции микроорганизмом дикого типа, т.е. достаточно сильный, чтобы способствовать предотвращению заболевания и/или обеспечить профилактику, облегчение или излечение заболевания.
В контексте настоящего описания термин «поливалентная вакцина» представляет собой вакцину, содержащую два или более разных антигена. В конкретном варианте осуществления этого типа поливалентная вакцина стимулирует иммунную систему реципиента против двух или более разных патогенов.
Термины «адъювант» и «иммуностимулятор» используются в настоящем описании взаимозаменяемо и означают одно или более веществ, вызывающих стимуляцию иммунной системы. В этом контексте адъювант используется для усиления иммунного ответа на один или более вакцинных антигенов/изолятов. Соответственно, «адъюванты» представляют собой агенты, которые неспецифически усиливают иммунный ответ на конкретный антиген, тем самым уменьшая количество антигена, необходимого в любой рассматриваемой вакцине, и/или частоту инъекций, необходимых для формирования адекватного иммунного ответа на представляющий интерес антиген. В этом контексте адъювант используется для усиления иммунного ответа на один или более вакцинных антигенов/изолятов.
В контексте настоящего описания термин «неадъювантная вакцина» представляет собой вакцину или поливалентную вакцину, не содержащую адъювант.
В контексте настоящего описания термин «фармацевтически приемлемый» используется в качестве прилагательного для обозначения того, что модифицированное им существительное подходит для использования в фармацевтическом продукте. Когда этот термин используется, например, для описания вспомогательного вещества в фармацевтической вакцине, он характеризует вспомогательное вещество как совместимое с другими ингредиентами композиции и не оказывающее вредного воздействия на организм предполагаемого животного-реципиента, например, свиньи.
«Парентеральное введение» включает подкожные инъекции, подслизистые инъекции, внутривенные инъекции, внутримышечные инъекции, внутрикожные инъекции и инфузию.
Антигены IAV-S, гемагглютинин и нейраминидаза, могут относиться к полному, т.е. полноразмерному белку, указанному в последовательностях, определенных в настоящем описании, или могут относиться к его антигенному фрагменту, который в равной степени может быть подходящим для индукции адекватного иммунологического ответа, о чем хорошо известно в области вакцин против гриппа (см., например, исследовательскую статью в PLOS ONE «An Influenza A/H1N1/2009 Hemagglutinin Vaccine Produced in Escherichia coli», José M. Aguilar-Yáñez et al.July 22, 2010; https://doi.org/10.1371/journal.pone.0011694; Vaccines (Basel) «Optimal Use of Vaccines for Control of Influenza A Virus in Swine», Matthew R. Sandbulte et al. 2015 Marc 3(1) 22-73).
В общем случае, антигенный фрагмент определенного белка (например, белкового антигена) представляет собой фрагмент этого белка, который является антигенным, т.е. способным специфически взаимодействовать с молекулой распознавания антигена иммунной системы, такой как иммуноглобулин (антитело) или Т-клеточный антигенный рецептор. Например, антигенный фрагмент гемагглютинина (HA) IAV-S представляет собой фрагмент белка HA, который является антигенным, т.е. выполняет функцию иммуногенного эпитопа. Антигенный фрагмент по настоящему изобретению предпочтительно является иммунодоминантным для распознавания антителом и/или Т-клеточным рецептором. В конкретных вариантах осуществления антигенный фрагмент по отношению к данному белковому антигену представляет собой фрагмент этого белка, который сохраняет по меньшей мере 25% антигенности (т.е. способности индуцировать соответствующие антитела, определенной с помощью анализа ингибирования HI или NI, как описано ниже) полноразмерного белка. В предпочтительных вариантах осуществления антигенный фрагмент сохраняет по меньшей мере 50% антигенности полноразмерного белка. В более предпочтительных вариантах осуществления антигенный фрагмент сохраняет по меньшей мере 75% антигенности полноразмерного белка. Антигенные фрагменты могут состоять всего из 20 аминокислот или, с другой стороны, могут представлять собой большие фрагменты, в которых отсутствует всего одна аминокислота полноразмерного белка. В конкретных вариантах осуществления антигенный фрагмент содержит от 25 до 150 аминокислотных остатков. В других вариантах осуществления антигенный фрагмент содержит от 50 до 250 аминокислотных остатков.
В контексте настоящего описания одна аминокислотная последовательность является на 100% «идентичной» или имеет 100% «идентичность» второй аминокислотной последовательности, когда аминокислотные остатки обеих последовательностей являются идентичными. Соответственно, аминокислотная последовательность на 50% «идентична» второй аминокислотной последовательности, когда идентичны 50% аминокислотных остатков двух аминокислотных последовательностей. Сравнение последовательностей выполняют на непрерывном блоке аминокислотных остатков, содержащихся в рассматриваемом белке, например, белке или части сравниваемого полипептида. В конкретном варианте осуществления учитываются выбранные делеции или вставки, которые в противном случае могли бы изменить соответствие между двумя аминокислотными последовательностями.
В контексте настоящего описания процент идентичности нуклеотидной и аминокислотной последовательностей может быть определен с помощью сетевой программы Clustal Omega, программы множественного выравнивания последовательностей с параметрами по умолчанию [Sievers and Higgins, Protein Sci. 2018 янв; 27(1):135-1452018]. Процент идентичности представляет собой единую числовую оценку, определяемую для каждой пары выровненных последовательностей. Он определяет количество идентичных остатков («совпадений») по отношению к длине выравнивания. Помимо Clustal Omega, другими программами, которые можно использовать для определения процента идентичности нуклеотидной и аминокислотной последовательностей, являются C, MacVector (MacVector, Inc. Cary, NC 27519), Vector NTI (Informax, Inc. MD), Oxford Molecular Group. PLC (1996) и алгоритм Clustal W с параметрами выравнивания по умолчанию и параметрами по умолчанию для идентичности. В качестве альтернативы можно использовать расширенный поиск Blast в условиях фильтра по умолчанию, например, с помощью программы GCG PileUp (Genetics Computer Group, Program Manual for the GCG Package, Version 7, Madison, Wisconsin) с параметрами по умолчанию.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Первый вариант осуществления первого аспекта настоящего изобретения относится к первой конструкции нуклеиновой кислоты, которая объединяет в определенном порядке по меньшей мере первую и вторую последовательности нуклеиновых кислот, кодирующие антигены, гемагглютинины (HA). Первый антиген HA, кодируемый первой последовательностью нуклеиновой кислоты в направлении от 5' к 3'-концу конструкции нуклеиновой кислоты, относится к линии Scot/94. Второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты в направлении от 5' к 3'-концу конструкции нуклеиновой кислоты, относится к линии EA.
Первый антиген HA линии Scot/94 может происходить от любого штамма, такому как штамм A/swine/Italy/3033-1/2015 (H1N2) или A/swine/France/35-140041 (H1N2). В предпочтительном варианте осуществления первый антиген НА линии Scot/94 происходит от штамма A/swine/Italy/3033-1/2015 (H1N2).
Более предпочтительно, первый антиген HA содержит и еще более предпочтительно состоит из аминокислотной последовательности в соответствии с SEQ ID NO: 3 или аминокислотной последовательности, идентичной на по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 89%, по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Более предпочтительно, первый антиген HA состоит из аминокислотной последовательности SEQ ID NO: 3 или аминокислотной последовательности, идентичной на по меньшей мере 90%, предпочтительно на по меньшей мере 93%, более предпочтительно на по меньшей мере 95%, 96%, 97%, 98% или 99%.
Второй антиген HA линии EA может происходить из любого штамма, например, штамма A/swine/Denmark/101048-2/2011 (H1N1), A/swine/Italy/28762-3/2013 (H1N1) или A/swine/France/44-120070/2012 (H1N1). В предпочтительном варианте осуществления второй антиген HA линии EA происходит от штамма A/swine/Italy/28762-3/2013 (H1N1).
Более предпочтительно, второй антиген HA содержит и еще более предпочтительно состоит из аминокислотной последовательности в соответствии с SEQ ID NO: 6 или аминокислотной последовательности, идентичной на по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Более предпочтительно второй антиген HA состоит из аминокислотной последовательности SEQ ID NO: 6 или аминокислотной последовательности, идентичной на по меньшей мере 93%, предпочтительно на по меньшей мере 95%, еще более предпочтительно на по меньшей мере 96%, 97%, 98% или 99%.
Второй вариант осуществления настоящего изобретения относится ко второй конструкции нуклеиновой кислоты, которая объединяет в определенном порядке по меньшей мере первую и вторую последовательности нуклеиновых кислот, кодирующие антигены, гемагглютинины (HA). Первый антиген HA, кодируемый первой последовательностью нуклеиновой кислоты в направлении от 5' к 3'-концу конструкции нуклеиновой кислоты, относится к линии Gent/84. Второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты в направлении от 5' к 3'-концу конструкции нуклеиновой кислоты, относится к линии pdm09.
Первый антиген НА линии Gent/84 может происходить от любого штамма, например, штамма A/swine/Italy/240849/2015 (H3N2). В предпочтительном варианте осуществления первый антиген НА линии Gent/84 происходит от штамма A/swine/Italy/240849/2015 (H3N2).
Более предпочтительно, первый антиген HA содержит и еще более предпочтительно состоит из аминокислотной последовательности в соответствии с SEQ ID NO: 9 или аминокислотной последовательности, идентичной на по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Более предпочтительно, первый антиген HA состоит из аминокислотной последовательности SEQ ID NO: 9 или аминокислотной последовательности, идентичной на по меньшей мере 93%, предпочтительно на по меньшей мере 95%, еще более предпочтительно на по меньшей мере 96%, 97%, 98% или 99%.
Второй антиген HA линии pdm09 может происходить от любого штамма, такого как штамм A/swine/England/373/2010 (H1N1). В предпочтительном варианте осуществления второй HA линии EA происходит от штамма A/swine/England/373/2010 (H1N1).
Более предпочтительно, второй антиген HA содержит и еще более предпочтительно состоит из аминокислотной последовательности в соответствии с SEQ ID NO: 12 или аминокислотной последовательности, идентичной на по меньшей мере 95%, 96%, 97%, 98% или 99%%. Более предпочтительно, первый антиген HA состоит из аминокислотной последовательности SEQ ID NO: 12 или аминокислотной последовательности, идентичной на по меньшей мере 95%, предпочтительно на по меньшей мере 96%, еще более предпочтительно на по меньшей мере 97%, 98% или 99%.
В первом варианте осуществления второго аспекта предоставлена конструкция нуклеиновой кислоты, содержащая первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген HA IAV-S линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), из штамма A/swine/Italy/3033-1/2015 (H1N2), и
вторая последовательность нуклеиновой кислоты кодирует второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA), из штамма A/swine/Italy/28762-3/2013 (H1N1).
Предпочтительно аминокислотная последовательность первого антигена HA IAV-S линии Scot/94 из штамма A/swine/Italy/3033-1/2015 (H1N2) содержит и более предпочтительно состоит из последовательности SEQ ID NO: 3 или аминокислотной последовательности, идентичной на по меньшей мере 85%, предпочтительно на по меньшей мере 90%. Идентичность аминокислотной последовательности также предпочтительно составляет по меньшей мере 91%, 92%, более предпочтительно по меньшей мере 93%, 94%, 95%, 96%, 97%, 98% или даже 99% или более.
Предпочтительно аминокислотная последовательность второго антигена HA IAV-S линии EA из штамма A/swine/Italy/28762-3/2013 (H1N1) содержит и более предпочтительно состоит из последовательности SEQ ID NO: 6 или аминокислотной последовательности, идентичной на по меньшей мере 90%, предпочтительно на по меньшей мере 93%. Идентичность аминокислотной последовательности также предпочтительно составляет по меньшей мере 94%, 95%, более предпочтительно по меньшей мере 96%, 97%, 98% или даже 99% или более.
Во втором варианте осуществления второго аспекта представлена конструкция нуклеиновой кислоты для применения для профилактики или лечения у субъекта заболевания, вызванного вирусом свиного гриппа А, где конструкция нуклеиновой кислоты содержит первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген, гемагглютинин (HA), вируса свиного гриппа A (IAV-S) линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), из штамма A/swine/Italy/240849/2015 (H3N2), и
вторая последовательность нуклеиновой кислоты кодирует второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09) из штамма A/swine/England/373/2010 (H1N1).
Предпочтительно аминокислотная последовательность первого антигена HA IAV-S линии Gent/84 из штамма A/swine/Italy/240849/2015 (H3N2) содержит и более предпочтительно состоит из последовательности SEQ ID NO: 9 или аминокислотной последовательности, идентичной на по меньшей мере 90%, предпочтительно на по меньшей мере 95%. Идентичность аминокислотной последовательности предпочтительно составляет по меньшей мере 96%, 97%, более предпочтительно по меньшей мере 98% или даже 99% или более.
Предпочтительно аминокислотная последовательность второго антигена HA IAV-S линии pdm09 из штамма A/swine/England/373/2010 (H1N1) содержит и более предпочтительно состоит из последовательности SEQ ID NO: 12 или аминокислотной последовательности, идентичной на по меньшей мере 90%, предпочтительно на по меньшей мере 95%. Идентичность аминокислотной последовательности предпочтительно составляет по меньшей мере 96%, 97%, более предпочтительно по меньшей мере 98% или даже 99% или более.
Конструкции нуклеиновых кислот согласно первому и/или второму варианту осуществления первого и/или второго аспекта могут быть включены в экспрессионную кассету, включающую последовательности нуклеиновых кислот, кодирующие антигены, гемагглютинины (HA), как описано выше, в виде гетерологичных генов вместе с последовательностями нуклеиновых кислот, контролирующими транскрипцию и/или экспрессию, такими как последовательности субгеномного промотора альфавируса и т.д., которые подходят для экспрессии антигенов HA. Такие экспрессионные кассеты могут быть получены хорошо известными способами путем включения гетерологичных последовательностей нуклеиновых кислот, кодирующих антигены HA, в вектор, такой как ДНК-вектор или РНК-вектор. Вектор может представлять собой остов вирусного репликона, такой как частица РНК-репликона (RP), и предпочтительно представляет собой частицу РНК-репликона альфавируса.
Таким образом, в первом и втором аспектах настоящего изобретения дополнительно представлена РНК-RP, предпочтительно РНК-RP альфавируса, содержащая нуклеотидную конструкцию в соответствии с первым вариантом осуществления. Кроме того, настоящее изобретение дополнительно относится к РНК, предпочтительно РНК-RP альфавируса, содержащей нуклеотидную конструкцию в соответствии со вторым вариантом осуществления.
«Частица РНК-репликона альфавируса (RP)» хорошо известна как вектор «непередаваемой», «одноцикловой» или «лишенной способности к размножению» вирусоподобной частицы. Геном может кодировать один или более гетерологичных генов с своего субгеномного промотора 26S. RP может реплицироваться в целевой клетке без образования потомства и, таким образом, доставлять и экспрессировать гетерологичный(ые) антиген(ы) для иммунной системы целевого животного. РНК-RP альфавируса может быть основана на штамме TC-83 вакцины против венесуэльского энцефалита лошадей (VEEV), опасного для человека.
Системы экспрессии на основе RP для гетерологичной экспрессии антигенов доступны в данной области и включают, например, коммерчески доступные платформы на основе векторов RP для производства вакцин, такие как Alphavaccine Platform System на основе вируса VEE, и технологии SEQUIVITYТМ, доступные от компании MSD/Merck Animal Health, США. Таким образом, в еще одном предпочтительном варианте осуществления частица РНК-репликона представляет собой частицу РНК-репликона на основе альфавируса венесуэльского энцефалита лошадей (VEE).
Например, ген(ы) антигена HA вируса затем может(могут) быть экспрессирован(ы) с субгеномного промотора (26S-Alphavirus), и транскрибированная РНК-репликона может быть упакована в RP путем экспрессии структурных белков в пакующих клеточных линиях или посредством ко-трансфекции в подходящие клетки-хозяева РНК-репликона с одной или более «хелперными» РНК, кодирующими структурные белки. Генерация частиц РНК-репликона TC-83 VEE описана, например, в US 9441247 и US 8460913. Вкратце, гены HA или NA синтезируют de novo (ДНК2.0) с использованием последовательности штаммов SIV. Два гена HA или три гена NA клонируют в плазмиду вектора репликона с помощью кассет мононаправленной экспрессии в тандеме, и последовательность подтверждают на отсутствие мутаций, возникших в процессе клонирования. РНК генерируют путем транскрипции in vitro линеаризованной плазмидной ДНК репликона с помощью РНК-полимеразы Т7, как описано ранее [Kamrud et al., Virology. 2007;360(2):376-387]. RP генерируют посредством совместной электропорации РНК-репликона для HA или NA и хелперной РНК структурных генов в клетки Vero с последующей сборкой частиц [Hooper et al., Vaccine. 2009; 28(2):494-511].
Общие молекулярно-биологические методы, включающие клонирование, трансфекцию, рекомбинацию, отбор и амплификацию, например, очень подробно описаны в стандартных учебниках, таких как Sambrook & Russell: «Molecular cloning: a laboratory manual» [2001, Cold Spring Harbour Laboratory Press; ISBN: 0879695773; Ausubel et al., в: Current Protocols in Molecular Biology, J. Wiley and Sons Inc., NY, 2003, ISBN: 047150338X; C. Dieffenbach & G. Dveksler: «PCR primers: a laboratory manual», CSHL Press, ISBN 0879696540; и «PCR protocols», J. Bartlett and D. Stirling, Humana Press, ISBN: 0896036421].
Конструкции нуклеиновых кислот по настоящему изобретению можно использовать в иммуногенных композициях, содержащих конструкции нуклеиновых кислот. Предпочтительно иммуногенные композиции содержат одну или более частиц репликона, содержащих конструкции нуклеиновых кислот по настоящему изобретению. Таким образом, частицы репликона по настоящему изобретению можно использовать в иммуногенных композициях, таких как вакцины, включающие частицы репликона. Иммуногенные композиции или вакцины могут состоять из частиц репликона или могут содержать частицы репликона в комбинации с дополнительными компонентами, такими как носители или адъюванты. Иммуногенные композиции по настоящему изобретению могут быть использованы в вакцинах для профилактики у субъекта заболевания, вызываемого вирусом свиного гриппа А (IAV-S).
Таким образом, в первом и/или втором аспектах настоящее изобретение дополнительно относится к иммуногенной композиции, содержащей или состоящей из РНК-RP, содержащей конструкцию нуклеиновой кислоты в соответствии с первым вариантом осуществления. Альтернативно, настоящее изобретение дополнительно относится к иммуногенной композиции, содержащей или состоящей из РНК-RP, содержащей конструкцию нуклеиновой кислоты в соответствии со вторым вариантом осуществления.
В предпочтительном варианте осуществления первого и/или второго аспектов настоящее изобретение относится к иммуногенной композиции, содержащей первую РНК-RP, содержащую нуклеотидную конструкцию в соответствии с первым вариантом осуществления, и вторую РНК-RP, содержащую нуклеотидную конструкцию в соответствии со вторым вариантом осуществления. В настоящем изобретении можно показать, что иммуногенная композиция, содержащая комбинацию частиц репликона в соответствии с первым и вторым вариантами осуществления обеспечивает широкую защиту от существующих линий IAV-S, и, таким образом, такую иммуногенную композицию можно успешно использовать в качестве вакцины, которая способствует защите, т.е. способствует профилактике или лечению вакцинированного субъекта, такого как свинья (например, свиноматка или поросенок), от инфекции IAV-S.
Следовательно, в предпочтительном варианте осуществления настоящее изобретение относится к иммуногенной композиции, такой как вакцина, содержащей первую и вторую частицы РНК-репликона, где,
(i) первая частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены, гемагглютинины (HA), вируса свиного гриппа A (IAV-S), где
первый антиген НА относится к линии Gent/84, и
второй антиген HA относится к линии pdm09,
(ii) вторая частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии Scot/94, и
четвертый антиген HA относится к линии EA.
В особенно предпочтительном варианте осуществления настоящее изобретение относится к иммуногенной композиции, такой как вакцина, которая содержит первую и вторую частицы РНК-репликона, где:
(i) первая частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержит первую конструкцию нуклеиновой кислоты, содержащую в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген HA IAV-S линии Scot/94, и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии EA,
и где иммуногенная композиция дополнительно содержит:
(ii) вторую частицу РНК-репликона, предпочтительно частицу РНК-репликона альфавируса, содержащую вторую конструкцию нуклеиновой кислоты, содержащую в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
третью последовательность нуклеиновой кислоты, кодирующую третий антиген HA IAV-S линии Gent/84, и
четвертую последовательность нуклеиновой кислоты, кодирующую четвертый антиген HA IAV-S линии pdm09.
В особенно предпочтительном варианте осуществления настоящее изобретение относится к иммуногенной композиции, такой как вакцина, которая содержит первую и вторую частицы РНК-репликона, где:
(i) первая частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержит первую конструкцию нуклеиновой кислоты, содержащую в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген HA IAV-S линии Scot/94, и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии EA,
и где иммуногенная композиция дополнительно содержит:
(ii) вторую частицу РНК-репликона, предпочтительно частицу РНК-репликона альфавируса, содержащую вторую конструкцию нуклеиновой кислоты, содержащую в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
третью последовательность нуклеиновой кислоты, кодирующую третий антиген HA IAV-S линии Gent/84, и
четвертую последовательность нуклеиновой кислоты, кодирующую четвертый антиген HA IAV-S линии pdm09.
Таким образом, в третьем аспекте настоящее изобретение относится к иммуногенной композиции, такой как вакцина, содержащей первую и вторую частицы РНК-репликона, где:
(i) первая частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты первый и второй антигены, гемагглютинины (HA), вируса свиного гриппа А (IAV-S), где
первый антиген НА, кодируемый первой последовательностью нуклеиновой кислоты, относится к линии Gent/84 из штамма A/swine/Italy/240849/2015 (H3N2) предпочтительно с SEQ ID NO: 9 или аминокислотной последовательностью, идентичной ей на по меньшей мере 90%, и
второй антиген HA, кодируемый первой последовательностью нуклеиновой кислоты, относится к линии pdm09 из штамма A/swine/England/373/2010 (H1N1) предпочтительно с SEQ ID NO: 12 или аминокислотной последовательностью, идентичной ей на по меньшей мере 95%.
(ii) вторая частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты третий и четвертый антигены HA IAV-S, где
третий антиген HA, кодируемый третьей последовательностью нуклеиновой кислоты, относится к линии Scot/94 из штамма A/swine/Italy/3033-1/2015 (H1N2) предпочтительно с SEQ ID NO: 3 или аминокислотной последовательностью, идентичной ей на по меньшей мере 85%, и
четвертый антиген HA, кодируемый четвертой последовательностью нуклеиновой кислоты, относится к линии EA из штамма A/swine/Italy/28762-3/2013 (H1N1) предпочтительно с SEQ ID NO: 6 или аминокислотной последовательностью, идентичной ей на по меньшей мере 90%.
Конструкции нуклеиновых кислот, иммуногенные композиции и частицы репликона третьего аспекта являются такими, как описано выше для первого и второго аспектов настоящего изобретения. Таким образом, настоящее изобретение дополнительно включает любые комбинации вариантов осуществления третьего аспекта с вариантами осуществления первого и второго аспектов, как представлено в настоящем описании. Таким образом, настоящее изобретение дополнительно относится к частицам репликона, описанным в третьем аспекте, где конструкции нуклеиновых кислот кодируют антигены HA IAV-S, расположенные в определенном порядке, как определено в первом аспекте, и/или где антигены IAV-S получены из конкретных штаммов, как определено во втором аспекте.
Иммуногенная композиция может быть адаптирована для одновременного или последовательного введения первой и второй частиц РНК-репликона, как описано выше, т.е. для одновременного или последовательного введения РНК-RP, содержащих конструкции нуклеиновых кислот, согласно первому и второму вариантам осуществления. Предпочтительно иммуногенная композиция адаптирована для одновременного введения первой и второй частиц РНК-репликона. Таким образом, в предпочтительном варианте осуществления иммуногенная композиция содержит первую и вторую частицы РНК-репликона в стандартной лекарственной форме.
В еще одном предпочтительном варианте осуществления иммуногенная композиция может содержать одну или более дополнительных частиц РНК-репликона. Такие дополнительные РНК-RP могут содержать конструкции нуклеиновых кислот, кодирующие один или более дополнительных антигенов. Например, дополнительные РНК-RP могут содержать конструкции нуклеиновых кислот, кодирующие один или более антигенов, нейраминидаз (NA), IAV-S. В конкретных вариантах осуществления конструкции нуклеиновых кислот кодируют два или три, предпочтительно три, антигена NA IAV-S или их иммуногенные фрагменты.
В особенно предпочтительном варианте осуществления дополнительные РНК-RP содержат конструкцию нуклеиновой кислоты, содержащую первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены NA IAV-S, где
первый антиген NA относится к линии Scot/94,
второй антиген NA относится к линии Gent/84, и
третий антиген NA выбирают из линии pdm09 или линии EA.
Соответственно, в четвертом аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены NA IAV-S, где
первый антиген NA, кодируемый первой последовательностью нуклеиновой кислоты, относится к линии Scot/94,
второй антиген NA, кодируемый второй последовательностью нуклеиновой кислоты, относится к линии Gent/84, и
третий антиген NA, кодируемый третьей последовательностью нуклеиновой кислоты, выбирают из линии pdm09 или линии EA.
Предпочтительно аминокислотная последовательность первого антигена NA IAV-S линии Scot/94 происходит от штамма A/swine/England/61470/2013 (H1N2). Аминокислотная последовательность первого антигена NA предпочтительно содержит и еще более предпочтительно состоит из последовательности SEQ ID NO: 15 или аминокислотной последовательности, идентичной на по меньшей мере 90%. Идентичность аминокислотной последовательности предпочтительно составляет по меньшей мере 96%, 97%, более предпочтительно по меньшей мере 98% или даже 99% или более.
Предпочтительно аминокислотная последовательность второго NA антигена IAV-S линии Gent/84 происходит от штамма A/swine/Italy/248147-8/2015 (H3N2). Аминокислотная последовательность второго антигена NA предпочтительно содержит и еще более предпочтительно состоит из последовательности SEQ ID NO: 18 или аминокислотной последовательности, идентичной на по меньшей мере 90%. Идентичность аминокислотной последовательности предпочтительно составляет по меньшей мере 96%, 97%, более предпочтительно по меньшей мере 98% или даже 99% или более.
Предпочтительно аминокислотная последовательность третьего антигена NA IAV-S линии pdm09 происходит от штамма A/swine/England/373/2010 (H1N1) или A/swine/Italy/179057/2015 (H1N1), предпочтительно от штамма A/swine/Italy/179057/2015 (H1N1). Аминокислотная последовательность третьего антигена NA предпочтительно содержит и еще более предпочтительно состоит из последовательности SEQ ID NO: 21 или аминокислотной последовательности, идентичной на по меньшей мере 90%. Идентичность аминокислотной последовательности предпочтительно составляет по меньшей мере 96%, 97%, более предпочтительно по меньшей мере 98% или даже 99% или более.
Альтернативно аминокислотная последовательность третьего антигена NA IAV-S линии EA происходит от штамма A/swine/Italy/28762-3/2013 (H1N1). Аминокислотная последовательность третьего антигена NA предпочтительно содержит и еще более предпочтительно состоит из последовательности SEQ ID NO: 24 или аминокислотной последовательности, идентичной на по меньшей мере 90%. Идентичность аминокислотной последовательности предпочтительно составляет по меньшей мере 96%, 97%, более предпочтительно по меньшей мере 98% или даже 99% или более.
Кроме того, предложена частица РНК-репликона, предпочтительно частица РНК-репликона альфавируса, содержащая конструкцию нуклеиновой кислоты, содержащую первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены, нейраминидазы (NA), вируса свиного гриппа А (IAV-S), где
первый антиген NA относится к линии Scot/94,
второй антиген NA относится к линии Gent/84, и
третий антиген NA выбирают из линии pdm09 или линии EA.
Частицу репликона, содержащую конструкцию нуклеиновой кислоты в соответствии с четвертым аспектом, можно использовать отдельно или в комбинации с частицей репликона в соответствии с первым, вторым и/или третьим аспектами настоящего изобретения, как представлено в настоящем описании, и предпочтительно использовать в комбинации с частицей репликона, содержащей антигены, гемагглютинины, описанные в первом, втором и/или третьем аспектах настоящего изобретения.
Частица репликона в соответствии с этим четвертым аспектом не имеет конкретных ограничений и предпочтительно представляет собой частицу репликона, такую как частица репликона альфавируса, наиболее предпочтительно частицу РНК-репликона альфавируса вируса венесуэльского энцефалита лошадей (VEEV), как описано в первом, втором и/или третьем аспектах.
В еще одном предпочтительном варианте осуществления настоящее изобретение относится к иммуногенной композиции, такой как вакцина, содержащей по меньшей мере первую, вторую и третью частицы РНК-репликона, где
первая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены HA IAV-S, где
первый антиген HA относится к линии Scot/94, и
второй антиген HA относится к линии EA,
вторая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии Gent/84, и
четвертый антиген HA относится к линии pdm09, и
третья частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены NA IAV-S, где
первый антиген NA относится к линии Scot/94,
второй антиген NA относится к линии Gent/84, и
третий антиген NA выбирают из линии pdm09 или линии EA.
Описанная выше иммуногенная композиция, такая как вакцина, может быть успешно использована в качестве вакцины, которая способствует защите вакцинированного субъекта, такого как свинья (например, свиноматка или поросенок), от инфекции IAV-S.
Иммуногенная композиция может быть адаптирована для одновременного или последовательного введения первой, второй и третьей частиц РНК-репликона, как описано выше, т.е. для одновременного или последовательного введения РНК-RP, содержащих конструкции нуклеиновых кислот в соответствии с первым, вторым и/или третьим аспектом в комбинации с конструкцией нуклеиновой кислоты в соответствии с четвертым аспектом. Предпочтительно иммуногенная композиция адаптирована для одновременного введения первой, второй и третьей частиц РНК-репликона. Таким образом, в предпочтительном варианте осуществления иммуногенная композиция содержит первую, вторую и третью частицы РНК-репликона в стандартной лекарственной форме.
Настоящее изобретение также относится к вакцинам против множества патогенов свиней. Например, кодирующая последовательность белкового антигена или его антигенного фрагмента, или комбинация таких кодирующих последовательностей белковых антигенов, используемых в свиной вакцине, может быть добавлена к частице РНК-репликона (RP) и/или объединена в вакцине в одной RP в виде одной частицы, кодирующей HA или NA, полученные из IAV-S, как описано в настоящем документе. Примеры патогенов, от которых может происходить один или более белковых антигенов или их антигенных фрагментов, включают вирус репродуктивно-респираторного синдрома свиней (PRRS), цирковирус свиней (PCV), вирус трансмиссивного гастроэнтерита (TGE), вирус псевдобешенства свиней (PPRV), парвовирус свиней (PPV), ротавирус свиней (PRV), вирус эпидемической диареи свиней (PED), Pasteurella multocida нескольких серотипов, Salmonella ssp., Escherichia coli, например, (серотипов K99, K88, 987P или F41), Haemophilus parasuis, Lawsonia intracellularis, Mycoplasma ssp. (например, Mycoplasma hyopneumoniae), Bordetella bronchiseptica, Erysipelas ssp., Campylobacter ssp., Actinobacillus pleuropneumoniae, Clostridium perfringens и Clostridium difficile.
Кроме того, настоящее изобретение относится к вакцинам, содержащим одну или более RP по настоящему изобретению в комбинации с одним или более другими векторами, кодирующими один или более из этих антигенов свиней (например, бакуловирусный вектор, кодирующий белок ORF-2 из цирковируса-2 свиней (PCV-2) и/или цирковируса-3 свиней (PCV-3)) и/или инактивированные анатоксины, происходящие от одного или более этих патогенов свиней. Более того, такие вакцины включают любую частицу РНК-репликона, кодирующую НА и/или NA, происходящие от IAV-S, в вакцине по настоящему изобретению вместе с одним или более изолятами убитых и/или модифицированных (аттенуированных) живых вирусов свиней и/или бактерий свиней.
Соответственно, одна или более РНК-RP, кодирующие один или более HA и/или NA, происходящие от IAV-S, могут быть добавлены вместе с одним или более другими векторами, кодирующими один или более антигенов свиней, и/или одним или более изолятами убитых и/или модифицированных (аттенуированных) живых вирусов, таких как один или более убитых или модифицированных живых штаммов IAS-V, один или более убитых и/или модифицированных живых вирусов РРСС, один или более убитых и/или модифицированных живых PCV, один или более убитых и/или модифицированных живых TGE, один или более убитых и/или модифицированных живых вирусов PPRV, один или более убитых и/или модифицированных живых PPV, один или более убитых и/или модифицированных живых PRV и один или более убитых и/или модифицированных живых PED. Кроме того, одну или более частиц РНК-репликона альфавируса (RP), которые кодируют один или более HA или NA, происходящих от IAV-S, можно добавлять вместе с одним или более другими векторами, кодирующими один или более антигенов свиней, и/или добавлять вместе с одной или более убитыми и/или модифицированными (аттенуированными) живыми бактериями, которые также способны инфицировать свиней, включая одну или более убитых и/или модифицированных живых Pasteurella multocida (одного или более из множества серотипов), Salmonella ssp., Escherichia coli (одного или более из множества серотипов), Haemophilus parasuis, Lawsonia intracellularis, Mycoplasma ssp. (например, Mycoplasma hyopneumoniae), Bordetella bronchiseptica, Erysipelas ssp., Campylobacter ssp., Actinobacillus pleuropneumoniae, Clostridium perfringens и Clostridium difficile.
Соответственно, настоящее изобретение также включает все частицы РНК-репликона по настоящему изобретению, векторы «голой» ДНК, которые содержат конструкции нуклеиновых кислот по настоящему изобретению, векторы «голой» РНК, которые содержат конструкции нуклеиновых кислот по настоящему изобретению, конструкции нуклеиновых кислот по настоящему изобретению, включая синтетическую матричную РНК и РНК-репликоны, а также все иммуногенные композиции и/или вакцины, которые содержат конструкции нуклеиновых кислот (например, синтетическую матричную РНК, РНК-репликоны), частицы РНК-репликона альфавируса, векторы «голой» РНК и/или векторы «голой» ДНК по настоящему изобретению.
Иммуногенная композиция по настоящему изобретению может быть использована в качестве вакцины, которая может представлять собой неадъювантную вакцину или адъювантную вакцину. Таким образом, настоящее изобретение дополнительно включает вакцины (поливалентные) вакцины, содержащие иммуногенные композиции по настоящему изобретению. В конкретных вариантах осуществления вакцины представляют собой неадъювантную вакцину. В других вариантах осуществления вакцины содержат адъювант. Адъюванты, подходящие для использования в вакцине по настоящему изобретению, не имеют конкретных ограничений и могут включать один или более адъювантов, выбранных из группы, состоящей из биоразлагаемого масла, эмульсии масло-в-воде с содержанием минерального масла 2,5-50% (об./об.) и биоразлагаемого масла, смешанного с эмульсией масло-в-воде с содержанием минерального масла 2,5-50% (об./об.).
В конкретных вариантах осуществления адъювант представляет собой биоразлагаемое масло. В конкретных вариантах осуществления этого типа биоразлагаемое масло представляет собой dl-α-токоферилацетат (ацетат витамина Е). В других вариантах осуществления адъювант представляет собой эмульсию масло-в-воде с содержанием минерального масла 2,5%-50% (об./об.). В конкретных вариантах осуществления адъювант представляет собой эмульсию масло-в-воде с содержанием минерального масла 2,5% (об./об.). В родственных вариантах осуществления адъювант представляет собой эмульсию масло-в-воде с содержанием минерального масла 5% (об./об.). В других вариантах осуществления адъювант представляет собой эмульсию масло-в-воде с содержанием минерального масла 12,5% (об./об.). В других вариантах осуществления адъювант представляет собой эмульсию масло-в-воде с содержанием минерального масла 25% (об./об.). В других вариантах осуществления адъювант представляет собой эмульсию масло-в-воде с содержанием минерального масла 50% (об./об.). В более конкретных вариантах осуществления адъювант содержит смесь биоразлагаемого масла с адъювантом на основе минерального масла. В конкретных вариантах осуществления биоразлагаемое масло представляет собой dl-α-токоферилацетат, и минеральное масло представляет собой жидкий парафин. В более конкретных вариантах осуществления биоразлагаемое масло представляет собой dl-α-токоферилацетат, и минеральное масло представляет собой легкий жидкий парафин.
В родственных составах адъювант представляет собой смесь двух компонентов. Первый компонент состоит из капель минерального масла с приблизительным средним (объемно-взвешенным) размером примерно 1 мкм, которые стабилизированы полисорбатом 80 (полиоксиэтилен(20)сорбитанмоноолеат) в воде. Первый компонент может содержать 25 мас.% минерального масла и 1 мас.% полисорбата 80, а остальное - вода. Второй компонент может состоять из капель биоразлагаемого dl-α-токоферилацетата с приблизительным средним (объемно-взвешенным) размером 400 нм, который также стабилизирован полисорбатом 80. Конкретные составы содержат 15 мас.% dl-α-токоферилацетата и 6 мас.% полисорбата 80, остальное - вода. В конкретных вариантах осуществления адъювант представляет собой X-SOLVETM (который представляет собой комбинацию двухкомпонентных адъювантов: DILUVAC FORTETM на основе dl-α-токоферилацетата и MICROSOLTM на основе легкого жидкого парафина [см., например, US 8,597,662].
В некоторых вариантах осуществления вакцина способствует предотвращению развития заболевания, вызванного IAV-S. В родственных вариантах осуществления индукция формирования антител в организме свиньи происходит при ее иммунизации вакциной. В некоторых вариантах осуществления свиньи представляют собой свиноматок. В родственных вариантах осуществления вакцина обеспечивает защитные материнские антитела потомству вакцинированной свиноматки. В других вариантах осуществления свинья представляет собой поросенка. В конкретных вариантах осуществления этого типа вакцину вводят поросенку уже в возрасте 3 дней. В конкретных вариантах осуществления вакцину вводят в виде бустер-вакцины. В некоторых вариантах осуществления вакцину вводят в виде однодозовой вакцины. В конкретных вариантах осуществления этого типа вакцину вводят в виде бустер-вакцины. В других вариантах осуществления вакцину вводят в виде многодозовой вакцины. В конкретных вариантах осуществления этого типа вакцину вводят в виде двухдозовой вакцины.
Настоящее изобретение также относится к способам иммунизации свиньи (например, свиноматки или поросенка) против патогена свиньи, например, IAV-S, включающим введение свинье иммунологически эффективного количества вакцины или поливалентной вакцины по настоящему изобретению. В конкретных вариантах осуществления вакцину вводят посредством внутримышечной инъекции. В альтернативных вариантах осуществления вакцину вводят посредством подкожной инъекции. В других вариантах осуществления вакцину вводят посредством внутривенной инъекции. В других вариантах осуществления вакцину вводят посредством внутрикожной инъекции. В других вариантах осуществления вакцину вводят посредством перорального введения. В других вариантах осуществления вакцину вводят интраназально. Предпочтительным способом является внутрикожное введение. Другим предпочтительным способом является внутримышечное введение.
Соответственно, вакцины и поливалентные вакцины по настоящему изобретению можно вводить в виде прайм-вакцины и/или бустер-вакцины. В конкретных вариантах осуществления вакцину по настоящему изобретению вводят в виде одноразовой вакцины (одна доза) без последующих введений. В некоторых вариантах осуществления, в случае введения как прайм-вакцины, так и бустер-вакцины, прайм-вакцину и бустер-вакцину можно вводить одинаковым способом введения.
В некоторых вариантах осуществления этого типа как прайм-вакцину, так и бустер-вакцину вводят путем внутрикожной инъекции. В других вариантах осуществления этого типа как прайм-вакцину, так и бустер-вакцину вводят путем внутримышечной инъекции. В альтернативных вариантах осуществления, в случае введения как прайм-вакцины, так и бустер-вакцины, введение прайм-вакцины можно осуществлять одним путем, а бустер-вакцины - другим путем. В некоторых вариантах осуществления этого типа прайм-вакцину можно вводить путем внутрикожной инъекции, а бустер-вакцину можно вводить перорально. В родственных вариантах осуществления этого типа прайм-вакцину можно вводить путем внутримышечной инъекции, а бустер-вакцину можно вводить перорально. В других вариантах осуществления этого типа прайм-вакцину можно вводить путем внутримышечной инъекции, а бустер-вакцину можно вводить путем внутрикожной инъекции. В других вариантах осуществления этого типа прайм-вакцину можно вводить путем внутрикожной инъекции, а бустер-вакцину можно вводить путем внутримышечной инъекции. Квалифицированному специалисту понятно, что состав вакцинной композиции готовят соответствующим образом для каждого типа животного-реципиента и пути введения.
Настоящее изобретение также относится к способу иммунизации свиней против IAV-S, включающему введение свиньям иммунологически эффективного количества вакцины по настоящему изобретению. Способ предпочтительно включает внутрикожное введение вакцины. Изобретение дополнительно относится к способу иммунизации свиньи (например, свиноматки или поросенка) против IAV-S, включающему введение свинье иммунологически эффективного количества вышеописанных вакцин по изобретению таким образом, чтобы в организме свиньи продуцировались соответствующие антитела к IAV-S. В конкретных вариантах осуществления вакцины могут включать, например, от примерно 1×104 до примерно 1×1010 RP или более. В более конкретных вариантах осуществления вакцины могут включать от примерно 1×105 до примерно 1×109 RP. В еще более конкретных вариантах осуществления вакцины могут включать от примерно 1×106 до примерно 1×108 RP.
В конкретных вариантах осуществления вакцины по настоящему изобретению вводят в дозах от 0,05 мл до 3 мл. В более конкретных вариантах осуществления вводимая доза составляет от 0,1 мл до 2 мл. В еще более конкретных вариантах осуществления вводимая доза составляет от 0,2 мл до 1,5 мл. В еще более конкретных вариантах осуществления вводимая доза составляет от 0,3 до 1,0 мл. В еще более конкретных вариантах осуществления вводимая доза составляет от 0,4 мл до 0,8 мл.
Таким образом, в первом аспекте настоящего изобретения предусмотрены следующие варианты осуществления:
[1] Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А (IAV-S), где конструкция нуклеиновой кислоты содержит в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген, гемагглютинин (HA), IAV-S линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
[2] Конструкция нуклеиновой кислоты для применения, как определено в [1], где первый антиген HA происходит от штамма A/swine/Italy/3033-1/2015 (H1N2).
[3] Конструкция нуклеиновой кислоты для применения, как определено в [1] или [2], где первый антиген HA, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, идентичную ей на по меньшей мере 85%.
[4] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[3], где второй антиген HA происходит от штамма A/swine/Italy/28762-3/2013 (H1N1).
[5] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[4], где второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[6] Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А (IAV-S), где конструкция нуклеиновой кислоты содержит в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген HA IAV-S линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09).
[7] Конструкция нуклеиновой кислоты для применения, как определено в [6], где первый антиген HA происходит от штамма A/swine/Italy/240849/2015 (H3N2).
[8] Конструкция нуклеиновой кислоты для применения, как определено в [6] или [7], где первый антиген HA, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 9 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[9] Конструкция нуклеиновой кислоты для применения, как определено в любом из пунктов [6]-[8], где второй антиген HA происходит от штамма A/swine/England/373/2010 (H1N1).
[10] Конструкция нуклеиновой кислоты для применения, как определено в любом из [6]-[9], где второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 12 или аминокислотную последовательность, идентичную ей на по меньшей мере 95%.
[11] Частица РНК-репликона, содержащая конструкцию нуклеиновой кислоты, как определено в любом из [1]-[5].
[12] Частица РНК-репликона, содержащая конструкцию нуклеиновой кислоты, как определено в любом из [6]-[10].
[13] Частица РНК-репликона, как определено в [15] или [16], которая представляет собой частицу РНК-репликона альфавируса.
[14] Частица РНК-репликона, как определено в [13], которая представляет собой частицу РНК-репликона альфавируса венесуэльского энцефалита лошадей (VEE).
[15] Иммуногенная композиция, содержащая частицу РНК-репликона по любому из [11]-[14].
[16] Иммуногенная композиция, как определено в [15], содержащая частицу РНК-репликона, как определено в [11] и [12].
[17] Иммуногенная композиция, как определено в [16], которая адаптирована для одновременного введения частиц РНК-репликона альфавируса, как определено в [11] и [12].
[18] Вакцина, содержащая иммуногенную композицию, как определено в любом из [15]-[17].
[19] Вакцина, как определено в [18], которая представляет собой неадъювантную вакцину.
[20] Вакцина, как определено в [18], которая содержит адъювант, выбранный из группы, состоящей из биоразлагаемого масла, эмульсии масло-в-воде с содержанием минерального масла 2,5-50% (об./об.) и биоразлагаемого масла, смешанного с эмульсией масло-в-воде с содержанием минерального масла 2,5-50% (об./об.).
[21] Вакцина, как определено в любом из [18]-[20], для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А.
[22] Способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества вакцины по любому из [18]-[20].
[23] Конструкция нуклеиновой кислоты, содержащая в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген HA вируса свиного гриппа A (IAV-S) линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
[24] Конструкция нуклеиновой кислоты, содержащая в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген HA IAV-S линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09).
Во втором аспекте настоящего изобретения предусмотрены следующие варианты осуществления:
[1] Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А, где конструкция нуклеиновой кислоты содержит первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген HA IAV-S линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), из штамма A/swine/Italy/3033-1/2015 (H1N2), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA), из штамма A/swine/Italy/28762-3/2013 (H1N1).
[2] Конструкция нуклеиновой кислоты для применения, как определено в [1], где первый антиген HA, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, идентичную ей на по меньшей мере 85%.
[3] Конструкция нуклеиновой кислоты для применения, как определено в [1] или [2], где второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[4] Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А, где конструкция нуклеиновой кислоты содержит первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген, гемагглютинин (HA), вируса свиного гриппа A (IAV-S) линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), из штамма A/swine/Italy/240849/2015 (H3N2), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09) из штамма A/swine/England/373/2010 (H1N1).
[5] Конструкция нуклеиновой кислоты для применения, как определено в [4], где первый антиген НА, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 9 или аминокислотную последовательность, идентичную ей на по меньшей мере 95%.
[6] Конструкция нуклеиновой кислоты для применения, как определено в [4] или [5], где второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 12 или аминокислотную последовательность, идентичную ей на по меньшей мере 95%.
[7] Частица РНК-репликона, содержащая нуклеотидную конструкцию, как определено в любом из [1]-[3].
[8] Частица РНК-репликона, содержащая нуклеотидную конструкцию, как определено в любом из [4]-[6].
[9] Частица РНК-репликона, как определено в [7] или [8], которая представляет собой частицу РНК-репликона альфавируса.
[10] Частица РНК-репликона, как определено в [9], которая представляет собой частицу РНК-репликона альфавируса венесуэльского энцефалита лошадей (VEEV).
[11] Иммуногенная композиция, содержащая частицу РНК-репликона, как определено в любом из [7]-[10].
[12] Иммуногенная композиция, как определено в [11], содержащая частицу РНК-репликона, как определено в [7] и [8].
[13] Вакцина, содержащая иммуногенную композицию, как определено в [12].
[14] Вакцина, как определено в [13], которая представляет собой неадъювантную вакцину.
[15] Вакцина, как определено в [13], которая содержит адъювант, выбранный из группы, состоящей из биоразлагаемого масла, эмульсии масло-в-воде с содержанием минерального масла 2,5-50% (об./об.) и биоразлагаемого масла, смешанного с эмульсией масло-в-воде с содержанием минерального масла 2,5-50% (об./об.).
[16] Вакцина, как определено в любом из [13]-[15], для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А.
[17] Способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества вакцины, как определено в любом из [14]-[16].
[18] Конструкция нуклеиновой кислоты, содержащая первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген, гемагглютинин (HA), вируса свиного гриппа A (IAV-S) линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), из штамма A/swine/Italy/240849/2015 (H3N2), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии A(H1N1)pdm09 (pdm09) из штамма A/swine/England/373/2010 (H1N1).
[19] Конструкция нуклеиновой кислоты, содержащая первую и вторую последовательности нуклеиновых кислот, где:
первая последовательность нуклеиновой кислоты кодирует первый антиген HA IAV-S линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), из штамма A/swine/Italy/3033-1/2015 (H1N2), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген HA IAV-S линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA), из штамма A/swine/Italy/28762-3/2013 (H1N1).
В третьем аспекте настоящего изобретения предусмотрены следующие варианты осуществления:
[1] Иммуногенная композиция для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А, содержащая первую и вторую частицы РНК-репликона, где:
первая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены, гемагглютинины (НА), вируса свиного гриппа А (IAV-S), где
первый антиген HA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
второй антиген HA относится к линии A(H1N1)pdm09 (pdm09),
вторая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
четвертый антиген HA относится к линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
[2] Иммуногенная композиция для применения, как определено в [1], где первый антиген HA происходит от штамма A/swine/Italy/240849/2015 (H3N2).
[3] Иммуногенная композиция для применения, как определено в [1] или [2], где первый антиген HA, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 9 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[4] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[3], где второй антиген НА происходит от штамма A/swine/England/373/2010 (H1N1).
[5] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[4], где второй антиген HA, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 12 или аминокислотную последовательность кислоты, идентичную ей на по меньшей мере 95%.
[6] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[5], где третий антиген НА происходит от штамма A/swine/Italy/3033-1/2015 (H1N2).
[7] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[6], где третий антиген HA, кодируемый третьей последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, идентичную на по меньшей мере 85%.
[8] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[7], где четвертый антиген НА происходит от штамма A/swine/Italy/28762-3/2013 (H1N1).
[9] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[8], где четвертый антиген НА, кодируемый четвертой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность, идентичную на по меньшей мере 90%.
[10] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[9], которая адаптирована для одновременного введения первой и второй частиц РНК-репликона.
[11] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[10], дополнительно содержащая третью частицу РНК-репликона, где:
третья частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены, нейраминидазы (NA), IAV-S, где
первый антиген NA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94),
второй антиген NA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
третий антиген NA выбирают из линии A(H1N1)pdm09 (pdm09) или линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
[12] Иммуногенная композиция для применения, как определено в любом из предшествующих [1]-[11], где частица РНК-репликона представляет собой частицу РНК-репликона альфавируса.
[13] Иммуногенная композиция для применения, как определено в [12], которая представляет собой частицу РНК-репликона альфавируса венесуэльского энцефалита лошадей (VEEV).
[14] Вакцина, содержащая иммуногенную композицию, как определено в любом из предшествующих [1]-[13].
[15] Вакцина, как определено в [14], которая представляет собой неадъювантную вакцину.
[16] Вакцина, как определено в [14], которая содержит адъювант, выбранный из группы, состоящей из биоразлагаемого масла, эмульсии масло-в-воде с содержанием минерального масла 2,5-50% (об./об.) и биоразлагаемого масла, смешанного с эмульсией масло-в-воде с содержанием минерального масла 2,5-50% (об./об.).
[17] Вакцина, определенная в любом из [14]-[16], для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А.
[18] Способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества вакцины, как определено в любом из [14]-[16].
[19] Иммуногенная композиция, содержащая первую и вторую частицы РНК-репликона, где:
первая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены, гемагглютинины (НА), вируса свиного гриппа А (IAV-S), где
первый антиген HA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
второй антиген HA относится к линии A(H1N1)pdm09 (pdm09),
вторая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
четвертый антиген HA относится к линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
[20] Иммуногенная композиция, содержащая первую, вторую и третью частицы РНК-репликона, где:
первая частица репликона РНК содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены, гемагглютинины (НА), вируса свиного гриппа А (IAV-S), где
первый антиген HA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
второй антиген HA относится к линии A(H1N1)pdm09 (pdm09),
вторая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
четвертый антиген HA относится к линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA),
третья частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены, нейраминидазы (NA), IAV-S, где
первый антиген NA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94),
второй антиген NA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
третий антиген NA выбирают из линии A(H1N1)pdm09 (pdm09) или линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
В четвертом аспекте настоящего изобретения предусмотрены следующие варианты осуществления:
[1] Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А, где конструкция нуклеиновой кислоты содержит первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены, нейраминидазы (NA), вируса свиного гриппа А (IAV-S), где
первый антиген NA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94),
второй антиген NA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
третий антиген NA выбирают из линии A(H1N1)pdm09 (pdm09) или линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
[2] Конструкция нуклеиновой кислоты для применения, как определено в [1], где первый антиген NA происходит от штамма A/swine/England/61470/2013 (H1N2).
[3] Конструкция нуклеиновой кислоты для применения, как определено в [1] или [2], где первый антиген NA, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 15 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[4] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[3], где второй антиген NA происходит от штамма A/swine/Italy/248147-8/2015 (H3N2).
[5] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[4], где второй антиген NA, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 18 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[6] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[5], где третий антиген NA происходит от штамма A/swine/England/373/2010 (H1N1) или A/swine/Italy/179057/2015 (H1N1).
[7] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[6], где третий антиген NA происходит от штамма A/swine/Italy/28762-3/2013 (H1N1).
[8] Конструкция нуклеиновой кислоты для применения, как определено в любом из [1]-[7], где третий антиген NA, кодируемый третьей последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 24 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
[9] Частица РНК-репликона, содержащая конструкцию нуклеиновой кислоты, определенную в любом из [1]-[8].
[10] Частица РНК-репликона, как определено в [9], которая представляет собой частицу РНК-репликона альфавируса.
[11] Частица РНК-репликона, как определено в [9] или [10], которая представляет собой частицу РНК-репликона альфавируса венесуэльского энцефалита лошадей (VEEV).
[12] Иммуногенная композиция, содержащая частицу РНК-репликона, как определено в любом из [9]-[11].
[13] Иммуногенная композиция, содержащая первую, вторую и третью частицы РНК-репликона, где:
первая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую первую и вторую последовательности нуклеиновых кислот, кодирующие первый и второй антигены, гемагглютинины (НА), вируса свиного гриппа А (IAV-S), где
первый антиген HA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
второй антиген HA относится к линии A(H1N1)pdm09 (pdm09),
вторая частица РНК-репликона содержит конструкцию нуклеиновой кислоты, содержащую третью и четвертую последовательности нуклеиновых кислот, кодирующие третий и четвертый антигены HA IAV-S, где
третий антиген HA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94), и
четвертый антиген HA относится к линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA), и
третья частица РНК-репликона представляет собой частицу РНК-репликона, как определено в любом из [9]-[11].
[14] Вакцина, содержащая иммуногенную композицию, как определено в [12] или [13].
[15] Вакцина, как определено в [14], которая представляет собой неадъювантную вакцину.
[16] Вакцина, как определено в [14], которая содержит адъювант, выбранный из группы, состоящей из биоразлагаемого масла, эмульсии масло-в-воде с содержанием минерального масла 2,5-50% (об./об.) и биоразлагаемого масла, смешанного с эмульсией масло-в-воде с содержанием минерального масла 2,5-50% (об./об.).
[17] Вакцина, определенная в любом из [14]-[16], для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А.
[18] Способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества вакцины, как определено в любом из [14]-[16].
[19] Конструкция нуклеиновой кислоты, содержащая первую, вторую и третью последовательности нуклеиновых кислот, кодирующие первый, второй и третий антигены, нейраминидазы (NA), вируса свиного гриппа A (IAV-S), где
первый антиген NA относится к линии H1huN2, подобной A/swine/Scotland/410440/1994 (Scot/94),
второй антиген NA относится к линии H3N2, подобной A/swine/Gent/1/1984 (Gent/84), и
третий антиген NA выбирают из линии A(H1N1)pdm09 (pdm09) или линии H1avN1, подобной евразийскому штамму птичьего происхождения (EA).
Приведенные ниже примеры служат для дальнейшего понимания изобретения, и никоим образом не предназначены для ограничения эффективного объема изобретения.
ПРИМЕРЫ
МАТЕРИАЛЫ И МЕТОДЫ
Приготовление вакцины на основе РНК-RP альфавируса
Получение частиц репликона с одним геном HA или NA (RP)
Векторы на основе репликона VEE, предназначенные для экспрессии генов гемагглютинина (НА) или нейраминидазы (НА), конструировали, как описано ранее [см. патенты США 9441247 В2; содержание которого включено в настоящее описание посредством ссылки] со следующими изменениями. Вектор на основе репликона pVEK, полученный из ТС-83 [раскрытый и описанный в US 9441247 B2], расщепляли рестрикционными ферментами Ascl и Pad. ДНК-плазмиду, содержащую кодон-оптимизированную последовательность открытой рамки считывания генов HA или NA (таблица 1a и b) с 5'-фланкирующей последовательностью (5'-GGCGCGCCGCACC-3') и 3'-фланкирующей последовательностью (5'-TTAATTAA-3'), расщепляли аналогичным образом с помощью ферментов рестрикции Ascl и Pad. Затем синтетическую генную кассету лигировали в расщепленный вектор pVEK, и полученные клоны переименовывали в код RP, соответствующий pVHV. Для обозначения векторов репликона, происходящего от pVEK, содержащих трансгенные кассеты, клонированные через сайты Ascl и Pad в сайт pVEK множественного клонирования, была выбрана номенклатура вектора «pVHV».
Частицы РНК-репликона ТС-83 (RP) получали в соответствии с ранее описанными способами [U.S. 9,441,247 В2 и 8,460,913 В2 США; содержание которых настоящим включено в описание посредством ссылки]. Вкратце, ДНК вектора репликона pVHV и хелперные ДНК-плазмиды линеаризовали с помощью фермента рестрикции Not1 перед транскрипцией in vitro с помощью РНК-полимеразы MegaScript T7 и аналога кэпа (Promega, Madison, Wl). Важно отметить, что хелперные РНК, использованные при получении, не содержали последовательности субгеномного промотора VEE, как описано ранее [Kamrud et al., J Gen Virol. 91 (Pt 7):1723-1727 (2010)]. Очищенную РНК для репликона и хелперные компоненты объединяли и смешивали с суспензией клеток Vero, подвергали электропорации в кюветах диаметром 4 мм и возвращали в среду OptiPro SFM для культивирования клеток (Thermo Fisher, Waltham, MA). После инкубации в течение ночи частицы РНК-репликона альфавируса очищали, готовили содержащий их состав в забуференном фосфатом солевом растворе с 5% сахарозой (мас./об.) и 1% свиной сывороткой, который пропускали через мембранный фильтр с размером пор 0,22 микрона и распределяли на аликвоты для хранения. Титр функциональных RP определяли с помощью иммунофлуоресцентного анализа в инфицированных монослоях клеток Vero. Партии RP идентифицировали в соответствии с геном, кодируемым упакованным репликоном (таблицы 1a и b).
Генерация частиц репликона (RP) с несколькими генами HA или NA
Векторы на основе репликона VEE, используемые для экспрессии генов HA или NA, конструировали, как описано ранее [см. патент США 9441247 B2; содержание которого включено в настоящее описание посредством ссылки] со следующими изменениями. Вектор на основе репликона pVEK, полученный из ТС-83 [раскрытый и описанный в US 9441247 B2], расщепляли ферментами рестрикции AscI и PacI. Для двухгенных конструкций HA и NA выбранные последовательности открытой рамки считывания оптимизировали по кодонам и синтезировали с фланкирующими сайтами AscI и PacI. Кроме того, интерстициальная последовательность между двумя синтетическими открытыми рамками считывания HA или NA состояла из 47 нуклеотидов некодирующей гетерологичной последовательности и второй копии промотора субгеномной (сг)РНК нативного TC-83 и последовательности 5'-нетранслируемой области сгРНК. Эти двухгенные конструкции обозначали как «pVDG», чтобы их можно было отличить от родительского вектора с последовательностью с одним промотором сгРНК. В случае конструкций с тремя генами NA конструкции на основе pVDG, содержащие два гена NA, дополнительно модифицировали следующим образом. Третью выбранную открытую рамку считывания NA оптимизировали по кодонам и синтезировали с фланкирующими сайтами PacI и SphI для направленного клонирования в вектор pVDG ниже двух существующих генов NA. Новая синтетическая конструкция также содержала 50 нуклеотидов гетерологичной некодирующей последовательности и третью копию промотора сгРНК нативного TC-83 и последовательность 5'-нетранслируемой области сгРНК на 5'-конце третьей последовательности гена NA. 3'-область третьей последовательности гена NA состояла из 3'-нетранслируемой области ТС-83 до соответствующего сайта SphI родительского вектора pVDG. Векторы с тремя генами обозначали как «pVTG», чтобы их можно было отличить от родственных векторов pVEK, pVHV и pVDG.
Выбранные последовательности HA (таблица 1a: EUHA1-3, EUHA1-2, EUHA1-5, EUHA1-15, EUHA1-17, EUHA1-8, EUHA1-11 и HA3-4) или NA (таблица 1b: EUNA1-2, Гены EUN1-4, EUN2-6 и EUN2-7) из ПРИМЕРОВ 1 и 3 использовали для синтеза генов мульти-HA или NA в плазмидном векторе pVDG или pVTG, как описано выше.
Частицы РНК-репликона ТС-83 (RP) получали в соответствии с ранее описанными способами [U.S. 9,441,247 B2 и 8,460,913 B2 США; содержание которых включено в настоящее описание посредством ссылки]. Вкратце, ДНК вектора на основе репликона pVDG или pVTG и хелперные ДНК-плазмиды линеаризовали с помощью ферментов рестрикции NotI перед транскрипцией in vitro с помощью РНК-полимеразы MegaScript T7 и аналога кэпа. Важно отметить, что хелперные РНК, использованные при получении, не содержали последовательности субгеномного промотора VEE, как описано ранее [Kamrud et al., J Gen Virol. 91(Pt 7):1723-1727 (2010)]. Очищенную РНК для репликона и хелперные компоненты объединяли и смешивали с суспензией клеток Vero, подвергали электропорации в кюветах диаметром 4 мм и возвращали в бессывороточную культуральную среду. После инкубации в течение ночи частицы РНК-репликона альфавируса очищали от клеток и среды путем пропускания суспензии через глубинный фильтр, промывки фосфатно-солевым буфером, содержащим 5% сахарозы (мас./об.), и, наконец, элюирования оставшихся RP 200 мМ буферным раствором, содержащим Na2SO4+5% сахарозы (мас./об.). В качестве альтернативы клетки и среду центрифугировали в присутствии приготовленной смолы Cellufine Sulfate®, промывали фосфатно-солевым буфером, содержащим 5% сахарозы (мас./об.), и элюировали 200 мМ буферным раствором, содержащим Na2SO4+5% сахарозы (мас./об.). Элюированные RP пропускали через 0,22-микронный мембранный фильтр и распределяли на аликвоты для хранения. Титр функциональных RP определяли с помощью иммунофлуоресцентного анализа в инфицированных монослоях клеток Vero.
Конструировали следующие частицы репликона, которые использовали в экспериментах:
Таблица 1а. Частицы РНК, исходные штаммы HA, линии, клады и номера доступа в GenBank (№ доступа)
Таблица 1b. Частицы РНК, исходные штаммы NA, линии и номер доступа в GenBank (№ доступа)
Примечание. Информация о кладах недоступна для антигенов NA.
Если в примерах или на фигурах не указано иное, для анализов HI использовали следующие штаммы и линии:
Таблица 1с. Штаммы SIV, использованные в качестве источника антигена HA для анализов HI на фиг. 1, 2, 3 и 4
Таблица 1d. Штаммы SIV, использованные в качестве источника антигена HA для анализа HI на фиг. 5 и 6
Таблица 1e. Штамм SIV или источник экспрессированного антигена NA для анализа NI на фиг. 7, 8, 9 и 10
* Лизаты клеток Vero, экспрессирующих соответствующий антиген NA, использовали в качестве источника антигена NA
Таблица 1f. Источник антигена NA для анализа NI на фиг. 11 и 12
*Лизаты клеток Vero, экспрессирующих соответствующий антиген NA, использовали в качестве источника антигена NA.
Общий план исследования
Каждую из приблизительно пятинедельных здоровых свиней (по 3 свиньи на одну вакцину), которые были серонегативными или имели низкий уровень анти-SIV антител, вакцинировали внутримышечно один раз вакцинами, содержащими 5-10×106 частиц РНК, кодирующих один или более генов HA или NA, и адъювант Xsolve50. Соответствующую вакцинацию повторяли в возрасте примерно 8 недель, и образцы крови собирали в возрасте примерно 9 недель и использовали либо для анализа ингибирования гемагглютинации (HI), либо для анализа ингибирования нейраминидазы (NI) для количественного определения уровней антиген-специфических антител.
Анализ ингибирования гемагглютинации (HI):
Все образцы сыворотки инактивировали нагреванием при 56°C в течение 30 мин, затем обрабатывали 0,25% периодатом, затем 0,75% глицерином и адсорбировали 2,6% куриными эритроцитами для удаления неспецифических агглютининов. Для титрования HI антител серийные разведения предварительно обработанной сыворотки инкубировали в течение 1 часа с 8 единицами гемагглютинации штаммов SIV, перечисленных в таблице 1c или 1d в качестве антигенов HA. Затем смесь инкубировали с 0,2% куриными эритроцитами в течение 1 часа при комнатной температуре, и планшеты анализировали на ингибирование агглютинации. Обратную величину от максимального разведения сыворотки, которое полностью ингибировало агглютинацию эритроцитов, принимали за HI-титр и выражали в виде логарифмических значений по основанию 2.
Анализ ингибирования нейраминидазы (NA) в сыворотке (NI):
В качестве источника антигенов NA использовали штаммы SIV лизатов клеток Vero, экспрессирующих антигены NA, подвергнутые электропорации РНК-репликонами, кодирующими гены соответствующих NA (таблицы 1e и 1f). Выполняли количественное определение ферментативной активности этих NA по отщеплению сиаловой кислоты от фетуина в 96-луночных планшетах во время инкубации в течение ночи при 37°C. Затем добавляли конъюгат агглютинина арахиса и пероксидазы хрена (PNA-HRP) на 2 часа при комнатной температуре для связывания молекул фетуина, лишенных сиаловой кислоты. Сигнал получали с субстратом 3,3',5,5'-тетраметилбензидина (ТМВ) и считывали при 450 нм. Тестовые антигены титровали для определения разведения, которое может дать 70% максимального сигнала. Равные объемы антигена NA добавляли к серийным разведениям сыворотки в лунках, покрытых фетуином, во время инкубации в течение ночи при 37°C. Значения оптической плотности (OD) нормализовали к значениям, полученным из лунок положительного контроля, не содержащих сыворотку. Титры ингибирования нейраминидазы определяли в виде обратной величины от интерполированного разведения сыворотки, имеющего значение экстинкции, равное 50% ингибирования по сравнению с контролем, и выражали в логарифмических значениях по основанию 2.
Корреляция титров антител к нейраминидазе и гемагглютинину с индуцированной вакциной защитой против SIV-A описана в: [Hobson D. et al., JHyg (Lond) 70, 767-777 (1972); Ohmit SE, et al., J Infect. Dis 204, 1879-1885 (2011); Walz L, et al., J Virol. 2018; 92(17):01006-18. (2018)]. Таким образом, результаты серологии в анализах ингибирования Hi и Ni, описанные в приведенных ниже примерах, указывают на предотвращение возникновения заболевания, вызванного SIV-A.
ПРИМЕР 1: Титры антител ингибирования гемагглютинации (HI), индуцированные RP, кодирующими отдельные антигены HA
Для определения защиты и перекрестной защиты, обусловленной РНК-RP альфавируса, кодирующими отдельные антигены НА каждого из штаммов EurAsianAvian (EA), Gent/84, Scot/94 и pdm09, выполняли следующее исследование:
Пятинедельных свиней (по 3 в группе) вакцинировали соответствующей частицей РНК с адъювантом XSolve50 по схеме первичной бустер-вакцинации с интервалом примерно в 3 недели. Сыворотки собирали через одну-две недели после бустер-вакцинации для определения титров антител к антигену гриппа, ингибирующих гемагглютинацию, что коррелирует с защитой от гриппа. Анализ HI позволяет измерить максимальное разведение сыворотки, которое предотвращает индуцированную вирусом гриппа гемагглютинацию эритроцитов. Обратную величину от этого разведения определяли в виде HI-титра, выраженного Log по основанию 2. Приведенные значения являются средними по 3 животным. Предел обнаружения для этого анализа равен 4 (пунктирная линия на чертежах), и, следовательно, титр, ниже 4 на чертежах, указан равным 3.
Результаты экспериментов по оценке HI показаны на фиг. 1-4. Можно сделать следующие выводы:
• Фиг. 1: RP штамма EUHA1-3 линии EA показали самые высокие HI-титры антиген-специфических антител почти ко всем протестированным антигенам EA IAS, за которыми следуют EUH1-5 и EUH1-2. Кроме того, можно было наблюдать перекрестно-реактивные титры антител к некоторым антигенам НА Scot/94 и pdm09. Ни один из протестированных штаммов не показал перекрестно-реактивные титры антител к антигенам Gent/84 IAS (все титры HI ниже 4).
• Фиг. 2: RP штамма EUHA1-15 продемонстрировали самые высокие HI-титры антиген-специфических антител почти ко всем протестированным антигенам Scot/94, за которыми следует EUH1-17, и, таким образом, показали лучшие результаты для антигенов Scot/94 кладов 2 и 3. RP штамма EUHA1-8 показали самые высокие HI-титры антиген-специфических антител к протестированным антигенам Scot/94 клады 1. Кроме того, можно было наблюдать перекрестно-реактивные титры антител к некоторым антигенам HA IAS штаммов EA и pdm09. Ни один из протестированных штаммов не показал перекрестно-реактивных титров антител к антигенам Gent/84 IAS (все титры HI ниже 4).
• Фиг. 3: RP штамма EUHA1-11 линии Pdm09 показали самые высокие HI-титры антиген-специфических антител почти ко всем антигенам pdm09. Кроме того, можно было наблюдать перекрестно-реактивные титры антител к большинству антигенов EA и Scot/94HA. Ни один из протестированных штаммов не показал перекрестно-реактивные титры антител к антигенам Gent/84 IAS (все титры HI ниже 4).
• Фиг. 4: RP штамма EUHA3-4 линии Gent/84 показали самые высокие HI-титры антиген-специфических антител ко всем протестированным антигенам Gent/84. Значимые перекрестно-реактивные титры антител к антигенам HA линий EA, Scot/94 и pdm09 не наблюдались.
ПРИМЕР 2: Титры антител ингибирования гемагглютинации (HI), индуцированные RP, кодирующими два антигена НА
Для определения серологической эффективности РНК-RP альфавируса, кодирующей два антигена HA, в следующих комбинациях:
1) антигены НА линии pdm09 и Gent/84 или
2) антигены НА EA и штаммов Scot/94,
выполняли исследование по схеме, описанной в примере 1.
Результаты анализа HI показаны на фиг. 5. Можно сделать следующие выводы:
Было обнаружено, что не все протестированные комбинации вызывают сильную серологическую реакцию. Кроме того, неожиданно было обнаружено, что порядок расположения генов в вирусном геноме частицы репликона имеет решающее значение для индуцирования серологического ответа.
• Комбинация антигенов HA линий Pdm09 и Gent/84: только комбинация, в которой Gent/84 расположен первым, а pdm09 расположен вторым в вирусном геноме частицы репликона, индуцировала сильный серологический ответ. Альтернативно, когда Pdm09 расположен первым, а Gent/84 расположен вторым в вирусном геноме частицы репликона, то можно наблюдать только существенно более низкий серологический ответ к антигену HA Gent/84 и очень слабый серологический ответ к антигену HA Pdm09.
• Комбинация антигенов HA линий EA и Scot/94: не все протестированные комбинации индуцировали сильный серологический ответ. Комбинация штаммов EUHA1-17 Scot/94 и EUHA1-3 EA показала наилучший серологический ответ (наивысшие HI-титры против антигенов IAS обеих линий).
Кроме того, сильный серологический ответ индуцировала только комбинация, в которой Scot/94 расположен первым, а ЕА расположен вторым в РНК репликона частицы репликона. Альтернативно, когда EA расположен первым, а Scot/94 расположен вторым в РНК репликона частицы репликона, значимый серологический ответ к антигенам HA EA не наблюдался.
• Среди различных протестированных комбинаций комбинация штаммов EUHA3-4+EUHA1-11 и EUHA1-17+EUHA1-3 индуцирует наилучший иммунитет, измеренный в виде HI-титра. Таким образом, эти комбинации предпочтительно использовать в составе, объединяющем две частицы репликона, т.е. для объединения первой частицы РНК-репликона, кодирующей штаммы EUHA3-4+EUHA1-11 в указанном порядке, со второй частицей РНК-репликона, кодирующей штаммы EUHA1-17+EUHA1-3 в указанном порядке.
В результате неожиданно было обнаружено, что положение гена НА в частице РНК-репликона и/или специфическая комбинация антигенов НА определяет уровень индуцированного иммунитета, измеряемый в виде HI-титра.
ПРИМЕР 3: Титры антител ингибирования нейраминидазы (NI), индуцированные RP, кодирующими отдельные антигены NA
Для определения серологической эффективности РНК-RP альфавируса, кодирующих отдельные антигены NA каждого из штаммов EurAsianAvian (EA), Gent/84, Scot/94 и pdm09, выполняли следующее исследование:
Пятинедельных свиней (по 3 в группе) вакцинировали соответствующей частицей РНК-репликона с адъювантом XSolve50 по схеме первичной бустер-вакцинации с интервалом примерно в 3 недели. Сыворотки собирали через одну-две недели после бустер-вакцинации для определения титров антител к антиген-специфичному ингибированию нейраминидазы (NI) гриппа. NI-титры измеряли с помощью анализа на основе лектина (агглютинина арахиса), как описано выше, обратную величину от самого высокого разведения сыворотки, которое ингибирует активность NA на по меньшей мере 50% по сравнению с контрольными лунками, определяли в виде NI-титра. Предел обнаружения для этого анализа был равен 2 (пунктирная линия на чертежах).
Результаты экспериментов по оценке NI показаны на фиг. 7-10. Можно сделать следующие выводы:
• Фиг. 7: RP штамма EUNA1-2 линии EA показали самые высокие NI-титры антиген-специфических антител почти ко всем протестированным антигенам EA IAS. Кроме того, можно было наблюдать перекрестную реакцию против некоторых антигенов NA Scot/94, pdm09 и Gent/84.
• Фиг. 8: RP штамма EUNA1-4 показали самые высокие NI-титры антиген-специфических антител к большинству протестированных антигенов pdm09, хотя уровень наблюдаемых NI-титров был ниже по сравнению с NI-титрами, достигнутыми с RP линии EA. Кроме того, можно было наблюдать титр перекрестно-реактивных антител к антигенам NA IAS EA, Scot/94 и Gent/84. Различия в титрах, измеренных между тестируемыми штаммами, были низкими.
• Фиг. 9: RP штамма EUNA2-6 линии Scot/94 показали самые высокие NI-титры антиген-специфических антител ко всем протестированным антигенам Scot/94. Кроме того, для штамма EUNA2-6 можно было наблюдать высокий уровень перекрестной реактивности к антигенам NA EA, pdm09 и Gent/84.
• Фиг. 10: RP штамма EUNA2-7 линии Gent/84 показали высокие NI-титры антиген-специфических антител ко всем протестированным антигенам Gent/84, а также показали значимую перекрестную защиту против антигенов NA EA, Scot/94 и pdm09.
ПРИМЕР 4: NI-титры антител, индуцированные RP, кодирующими два или три антигена NA
Для определения серологической эффективности конструировали РНК-RP альфавируса, кодирующие два или три антигена NA, и получали RP, кодирующие антигены NA из перечисленных ниже линий:
1) антигены NA линий EA и Gent/84 или
2) антигены NA линий EA, Gent/84 и Scot/94,
и исследование проводили по схеме, описанной в примере 3.
Результаты экспериментов по оценке NI показаны на фиг. 11. Можно сделать следующие выводы:
Было обнаружено, что все протестированные комбинации вызывают серологический ответ независимо от порядка расположения генов. Таким образом, неожиданно было обнаружено, что в противоположность наблюдениям в случае антигенов HA (см. пример 2 выше) порядок генов NA в вирусном геноме частицы репликона не является критическим для индукции серологического ответа.
ПРИМЕР 5: NI-титры антител, индуцированные RP, кодирующими два и три антигена NA
Результаты, показанные на фиг. 7-10, демонстрируют, что комбинация штамма линии EA, линии Gent/84 со штаммом линии Scot/94 должна обеспечивать наилучшую защиту от IAS с наилучшей защитой и перекрестной защитой от всех четырех линий. Таким образом, лучшим кандидатом для проверки такой перекрестной защиты является комбинация штамма EUNA2-6 линии Scot/94 со штаммом EUNA2-7 линии Gent/84, которую затем можно комбинировать со штаммом либо линии EA, таким как штамм EUNA1-2, либо линии pdm09, таким как штамм EUNA1-4. Как следствие, эти комбинации штаммов оценивали на их серологический ответ.
Таким образом, для определения защиты, обеспечиваемой РНК-RP альфавируса, кодирующими два и три антигена NA, использовали следующие комбинации:
1) антигены NA линий EA и Gent/84
2) антигены NA линий Scot/94, Gent/84 и EA или
3) антигены NA линий Scot/94, Gent/84 и pdm09,
и исследование выполняли по схеме, описанной в примере 3.
Результаты показаны на фиг. 12:
• В отличие от результатов, наблюдаемых для антигенов HA, комбинации антигенов NA только трех линий достаточно, чтобы вызвать серологический ответ против всех четырех линий IAS.
• Слабый серологический ответ против всех четырех линий IAS уже может быть достигнут при комбинации антигенов NA только из двух линий, независимо от порядка расположения генов в частице РНК-репликона.
• Наивысший серологический ответ может быть достигнут при комбинации антигенов NA Scot/94 и Gent/84, дополнительно объединенных с антигенами NA линий pdm09 или EA.
Таблица 2: Список SEQ ID NO
ПРИМЕР 6: Оценка эффективности поливалентной вакцины против IAV-S
Выполняли исследование для определения иммуногенности и эффективности поливалентной вакцины против IAV-S, содержащей две RP, кодирующие два HA (RP EUSIV-T8, кодирующая антигены EUHA1-17 и EUHA1-3, и RP EUSIV-K, кодирующая антигены EUH3-4 и EUH1-11, таблицы 1a и 2) и одну конструкцию с тремя NA (EUSIV-R, кодирующая антигены EUN2-6, EUN1-2 и EUN2-5, таблицы 1b и 2). Адъювантную вакцину вводили 5 свиньям путем двукратной внутримышечной (IM) вакцинации в возрасте 5 и 8 недель (2 мл на дозу; 3×5×106 RP/доза, вакцины). Равное количество непривитых животных получали забуференный фосфатом физиологический раствор с адъювантом. Иммуногенность вакцины измеряли путем количественного определения HI- и NI-титров в образцах сыворотки, собранных до экспериментального заражения в возрасте 10 недель. Эффективность вакцины тестировали против заражения Gent/84 [A/swine/Belgium/113/2013 (H3N2)] интратрахеальным путем в возрасте 10 недель (32-й день исследования). Измеряли эффективность вакцины против лихорадки, вызванной инфекцией IAV-S, т.е. повышения ректальной температуры, и поражений легких через 3 дня после заражения.
Результаты этого эксперимента показаны на фиг. 13A, B, C и D. Поливалентная вакцина против IAV-S индуцировала функциональные HI-титры против гетерологичных штаммов IAV-S, принадлежащих ко всем четырем линиям (фиг. 13A), и NI-титры против гомологичного антигена NA всех трех линий (фиг. 13B). Кроме того, поливалентная вакцина против IAV-S защищала свиней от индуцированной экспериментальной инфекции, повышения ректальной температуры, лихорадки (фиг. 13С) и поражений у свиней (фиг. 13D). Эти результаты демонстрируют, что протестированная поливалентная вакцина против IAV-S оказалась как иммуногенной, так и эффективной.
ПРИМЕР 7: Оценка эффективности вакцины после внутрикожного введения
Выполняли исследование для определения серологической эффективности поливалентной вакцины против IAV-S, содержащей две RP с двумя HA (RP EUSIV-T8, кодирующая антигены EUHA1-17 и EUHA1-3, и RP EUSIV-K, кодирующая антигены EUH3-4 и EUH1-11, таблицы 1a и 2) и одну конструкцию с тремя NA (EUSIV-R, кодирующая антигены EUN2-6, EUN1-2 и EUN2-5, таблицы 1b и 2). Адъювантную вакцину вводили 3 свиньям путем двукратной внутрикожной (ID) вакцинации с помощью безыгольного инъектора IDAL® в возрасте 5 и 8 недель (200 мкл на дозу; 3×3×106 RP/доза, вакцины). Равное количество непривитых животных получали забуференный фосфатом физиологический раствор с адъювантом. Иммуногенность вакцины измеряли путем количественного определения HI- и NI-титров в образцах сыворотки, собранных в возрасте 10 недель.
Результаты этого эксперимента показаны на фиг. 14 А и В. Поливалентная вакцина против IAV-S индуцировала функциональные HI-титры против гетерологичных штаммов IAV-S, принадлежащих к трем из четырех протестированных линий (фиг. 14A), и NI-титры против двух из трех протестированных гомологичных антигенов NA (фиг. 14B). Эти результаты показывают, что внутрикожное применение поливалентной вакцины против IAV-S также эффективно.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ИНТЕРВЕТ ИНТЕРНЭШНЛ Б.В.
<120> Вакцина против вируса свиного гриппа А
<130> 24970 EPPD
<160> 25
<170> BiSSAP 1.3.6
<210> 1
<211> 1695
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KU323133.1 Вирус гриппа A (A/swine/Italy/3033-1/2015(H1N2))
ген гемагглютинина (НА) сегмента 4, полная кодирующая
последовательность
<400> 1
atgaaagcta aactactaat cctgttgtgt gcactttcaa ctacagatgc agacacaata 60
tgtataggat accatgctaa caattcaacc gacaccgttg acacagtcct agaaaagaat 120
gtgacagtaa cacactctgt caacctcctt gaggacagtc acaatgggaa actatgtaaa 180
ctgaagggag tggccccact gcaattggga aaatgcagca tcgcaggatg gatactaggg 240
aacccagagt gtgaatcact gctttctaag aaatcatggt cctatattgc agaagcacca 300
aatgctgaga atggaatatg ttacccaggg cacttctccg actacgaaga gctgagggag 360
caattaagtt cagtatcttc attcgagagg ttcgaaatat ttcctaagga gagttcatgg 420
cccaaacaca gcataggagc aactgcatca tgctccaaac aagggaaaag cagtttttat 480
acaaatctgc tatggctaac tgaaaaaaat gggtcttatc caaatttgac catgtcctat 540
atgaacgaca aagagagaga agtccttgtg ctatggggag tccatcatcc gtccaacata 600
gaggaccaaa gagcaatcta tcgcaaagaa actgcttatg tttctgtaat gtcatcacac 660
tacaacaaga ggttcacccc agagattgca aaaagaccca aagtaaggaa tcaagagggg 720
agaatcaact actactggac actgctggaa cccagggaca caataatatt tgaagcaaat 780
gggaatttga tagcaccatg gtatgctttt gcattgaata gaggctttga gtcaggaatc 840
attgtctcaa atgcatcgat ggatgagtgt aacgcgaagt gtcaaacgcc ccgaggagcg 900
ataaacaata gcctgccttt ccagaatgta cacccaatca caatagggga atgccccaag 960
tatgtaaaga gtacaaaatt aaagatggct acaggactta gaaacattcc atccatccaa 1020
tccagaggtt tgtttggagc cattgctggt ttcattgaag ggggatggac cggaatgata 1080
gatggatggt atggttatca tcatcagaat gaacagggtt ctggctatgc tgcagatcaa 1140
aaaagcacac aaaatgccat taatgggatt acaaacaagg tgaactctgt tatcgataaa 1200
atgaacactc aatttacagc tgtaggtaaa gaattcaaca gattagaaaa aagaatggaa 1260
aacctgaata agaaggttga tgacggtttt cttgacattt ggacgtataa tgcagaattg 1320
ttggttctgc tggagaatga gaggactttg gattttcatg attcaaatgt gaagagctta 1380
tatgaaggag taaaggccca attgaagaat aatgccaaag aagtaggaaa cgggtgtttt 1440
gaattctacc acaagtgtga caacgaatgc atggacagtg taaaaaatgg aacttatgac 1500
tatccaagat actcagagga atcaaaatta aacagagaac aaattgatgg agtggaattg 1560
aaatcaatgg gagtctatca gatcctggcg atatactcaa ctgccgccag ttcattggtg 1620
cttttagtca ccctgggggc aatcagtttt tggatgtgtt ctaatggttc tttgcagtgc 1680
agaatatgta tctga 1695
<210> 2
<211> 1695
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUH1-17 Оптимизированная последовательность свиньи
<400> 2
atgaaagcca agctcctcat tctcctttgt gccctgtcca cgaccgacgc tgatactatc 60
tgcatcggct accatgccaa caactccact gataccgtgg acaccgtgct ggaaaagaac 120
gtgaccgtga ctcactcggt gaacctcctt gaggacagcc acaacgggaa gctctgcaag 180
ctgaagggcg tggcacccct acaactggga aagtgttcca tcgcgggctg gattctggga 240
aaccccgagt gcgaatccct tctgtcgaaa aagtcatggt cctacattgc cgaggccccg 300
aatgctgaga acggcatttg ctacccggga cacttcagcg attacgaaga actgcgggaa 360
cagctgtcga gcgtgtcaag tttcgagagg ttcgagatct tcccgaagga aagctcatgg 420
ccgaagcaca gcatcggcgc caccgcctcg tgttcgaaac agggaaagtc ctcgttctac 480
actaacctcc tgtggctgac cgagaagaat gggtcctatc ctaacctgac catgtcgtac 540
atgaacgaca aggaacggga agtgctcgtg ttgtgggggg tgcaccatcc atcaaatatc 600
gaggaccaga gggccatcta ccgcaaagaa actgcctacg tgtccgtgat gtctagccac 660
tacaacaagc gcttcacgcc tgaaatcgcc aaaagaccca aggtccgcaa ccaagagggc 720
agaatcaatt actactggac cctcctggag ccccgggaca ccataatctt tgaagcaaat 780
ggaaacctca ttgcgccttg gtacgccttc gccctgaacc ggggtttcga gtccggaatt 840
atcgtcagca acgcctccat ggacgagtgt aacgcgaagt gccagacccc gcgcggcgct 900
atcaacaaca gcctgccttt ccaaaacgtg cacccaatca ccatcggcga atgccccaaa 960
tacgtgaagt ccactaagct gaagatggca acaggactgc ggaacatccc gtcaattcag 1020
tcccgagggt tgttcggggc aatcgccggg ttcattgagg gcggatggac cggaatgatt 1080
gacggttggt acggatacca ccaccagaac gaacagggtt ccggttatgc ggcggatcag 1140
aagtccaccc agaacgccat caacggcatc accaacaagg tcaactccgt catcgacaag 1200
atgaacaccc agttcactgc cgtgggaaag gagttcaacc ggctggagaa gcggatggaa 1260
aatctgaaca agaaagtcga cgacggcttc ctggacattt ggacctacaa cgccgaactg 1320
ctggtgctgc tggaaaacga aagaactctc gactttcatg attcgaacgt gaagtcactg 1380
tacgaaggag tgaaggccca gctcaagaac aacgccaagg aagtcggaaa cggttgcttc 1440
gagttttacc ataagtgcga caacgagtgc atggatagcg tgaagaacgg aacctacgac 1500
tacccccggt attccgaaga gtcgaagctc aacagagaac agatcgatgg cgtggagctg 1560
aagtccatgg gagtgtacca gatcctggcg atctactcca ctgcggctag ctccctggtg 1620
ctgttggtca ccttgggggc tatctctttc tggatgtgct ccaacggttc cctgcaatgc 1680
cgcatttgca tttga 1695
<210> 3
<211> 564
<212> Белок
<213> Вирус гриппа A
<220>
<223> >ALX30160.1 гемагглютинин [Вирус гриппа A
(A/swine/Italy/3033-1/2015(H1N2))]
<400> 3
Met Lys Ala Lys Leu Leu Ile Leu Leu Cys Ala Leu Ser Thr Thr Asp
1 5 10 15
Ala Asp Thr Ile Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr
20 25 30
Val Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn
35 40 45
Leu Leu Glu Asp Ser His Asn Gly Lys Leu Cys Lys Leu Lys Gly Val
50 55 60
Ala Pro Leu Gln Leu Gly Lys Cys Ser Ile Ala Gly Trp Ile Leu Gly
65 70 75 80
Asn Pro Glu Cys Glu Ser Leu Leu Ser Lys Lys Ser Trp Ser Tyr Ile
85 90 95
Ala Glu Ala Pro Asn Ala Glu Asn Gly Ile Cys Tyr Pro Gly His Phe
100 105 110
Ser Asp Tyr Glu Glu Leu Arg Glu Gln Leu Ser Ser Val Ser Ser Phe
115 120 125
Glu Arg Phe Glu Ile Phe Pro Lys Glu Ser Ser Trp Pro Lys His Ser
130 135 140
Ile Gly Ala Thr Ala Ser Cys Ser Lys Gln Gly Lys Ser Ser Phe Tyr
145 150 155 160
Thr Asn Leu Leu Trp Leu Thr Glu Lys Asn Gly Ser Tyr Pro Asn Leu
165 170 175
Thr Met Ser Tyr Met Asn Asp Lys Glu Arg Glu Val Leu Val Leu Trp
180 185 190
Gly Val His His Pro Ser Asn Ile Glu Asp Gln Arg Ala Ile Tyr Arg
195 200 205
Lys Glu Thr Ala Tyr Val Ser Val Met Ser Ser His Tyr Asn Lys Arg
210 215 220
Phe Thr Pro Glu Ile Ala Lys Arg Pro Lys Val Arg Asn Gln Glu Gly
225 230 235 240
Arg Ile Asn Tyr Tyr Trp Thr Leu Leu Glu Pro Arg Asp Thr Ile Ile
245 250 255
Phe Glu Ala Asn Gly Asn Leu Ile Ala Pro Trp Tyr Ala Phe Ala Leu
260 265 270
Asn Arg Gly Phe Glu Ser Gly Ile Ile Val Ser Asn Ala Ser Met Asp
275 280 285
Glu Cys Asn Ala Lys Cys Gln Thr Pro Arg Gly Ala Ile Asn Asn Ser
290 295 300
Leu Pro Phe Gln Asn Val His Pro Ile Thr Ile Gly Glu Cys Pro Lys
305 310 315 320
Tyr Val Lys Ser Thr Lys Leu Lys Met Ala Thr Gly Leu Arg Asn Ile
325 330 335
Pro Ser Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile
340 345 350
Glu Gly Gly Trp Thr Gly Met Ile Asp Gly Trp Tyr Gly Tyr His His
355 360 365
Gln Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Gln Lys Ser Thr Gln
370 375 380
Asn Ala Ile Asn Gly Ile Thr Asn Lys Val Asn Ser Val Ile Asp Lys
385 390 395 400
Met Asn Thr Gln Phe Thr Ala Val Gly Lys Glu Phe Asn Arg Leu Glu
405 410 415
Lys Arg Met Glu Asn Leu Asn Lys Lys Val Asp Asp Gly Phe Leu Asp
420 425 430
Ile Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Leu Glu Asn Glu Arg
435 440 445
Thr Leu Asp Phe His Asp Ser Asn Val Lys Ser Leu Tyr Glu Gly Val
450 455 460
Lys Ala Gln Leu Lys Asn Asn Ala Lys Glu Val Gly Asn Gly Cys Phe
465 470 475 480
Glu Phe Tyr His Lys Cys Asp Asn Glu Cys Met Asp Ser Val Lys Asn
485 490 495
Gly Thr Tyr Asp Tyr Pro Arg Tyr Ser Glu Glu Ser Lys Leu Asn Arg
500 505 510
Glu Gln Ile Asp Gly Val Glu Leu Lys Ser Met Gly Val Tyr Gln Ile
515 520 525
Leu Ala Ile Tyr Ser Thr Ala Ala Ser Ser Leu Val Leu Leu Val Thr
530 535 540
Leu Gly Ala Ile Ser Phe Trp Met Cys Ser Asn Gly Ser Leu Gln Cys
545 550 555 560
Arg Ile Cys Ile
<210> 4
<211> 1701
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KR700530.1 Вирус гриппа A (A/swine/Italy/28762-3/2013(H1N1))
ген гемагглютинина (НА) сегмента 4, полная кодирующая
последовательность
<400> 4
atgaaagcaa aattatttgt attattctgt gcatttactg cactgaaagc tgacaccatt 60
tgtgtaggct atcatgctaa caattccaca gacactgtcg acacgatact ggaaaagaat 120
gttactgtta cccactcagt taatttacta gaaaacagcc acaatggaaa actctgcagc 180
ctgaatggaa aggctccgtt gcaactggag aactgcaacg tagcaggatg gatccttggt 240
aatccagaat gtgactttct gctcacagcg aattcatggt cctacataat agagacttca 300
aattcaaaaa atggaacatg ctatcctgga gaattcactg attatgagga actaagggag 360
caactgagta cagtttcttc atttgaaaga tttgaaattt tcccaaaagc aacctcatgg 420
ccaaatcatg agacaaccaa aggtaccaca gttgcatgct cccactctgg agccaacagt 480
ttttatcgga acttgctatg gatagtaaag aaaggaaact cctatcccaa gctcagcaag 540
tcatacacaa acaacaaagg gaaagaagtg cttgtaatct ggggagtgca tcacccccct 600
acagacagtg accaacaaac cctctatcag aataatcaca catatgtttc agttggatca 660
tcaaaatact accaaaggtt cacaccagaa atagtagcca gacctaaagt cagagagcaa 720
gcgggcagaa tgaattatta ttggatacta ttagatcaag gagacaccat aacatttgaa 780
gccacgggga atttaatagc accatggcat gcatttgcat tgaataaggg ctctagctct 840
ggaattatga tgtcagatgc tcatgttcaa aattgcacta caaagtgcca aactcctcat 900
ggggcattga aaagcaatct tccttttcaa aatgtacatc ccatcactat tggagagtgc 960
cccaaatatg ttaaaagcac ccaactaaga atggcaacag gattaaggaa cataccatct 1020
atccaatcca gaggactttt tggggcaatt gccggattta ttgaaggagg atggacagga 1080
atgatagatg gatggtatgg atatcaccat caaaatgagc aaggatctgg ttatgcagca 1140
gataagaaaa gcacacaaat tgcaattgat gagatcagca acaaagtaaa ctcagtaatt 1200
gagaaaatga acattcaatt cacttcagtg ggcaaggagt tcaataattt agagaaaagg 1260
attgagaatt tgaataaaaa ggtcgatgat ggatttttgg atgtgtggac atataacgct 1320
gagttactca ttttgctcga gaacgaaaga accctagatt tccatgactt taacgtaaga 1380
aatttgtatg aaaaggtcaa atcacaactg agaaacaatg ccaaggaaat cggtaatggt 1440
tgttttgagt tctatcacag atgtgacaat gaatgcatgg aaagtgtaaa gaatggcaca 1500
tataattatc ccagatattc agaagaatcc aaattaaata gagagaaaat agacggtgtg 1560
aaactagaat cattaggagt acatcagatt ttggcaatct actccacagt cgccagttcc 1620
ctggtattgc tagtctccct gggggcaatc agtttctgga tgtgctctaa tggatcattg 1680
caatgcagaa tatgcatttg a 1701
<210> 5
<211> 1701
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUH1-3 Оптимизированная последовательность свиньи
<400> 5
atgaaagcca agctgtttgt ccttttctgc gccttcaccg cccttaaagc cgacaccatc 60
tgtgtgggct accatgctaa caactccacc gatacggtcg acactattct ggagaagaac 120
gtgaccgtga cccactccgt gaatctgctg gagaatagcc acaacggaaa gctgtgcagc 180
ctgaacggaa aggccccgct gcaactggag aattgtaacg tggccggatg gatcctgggc 240
aaccctgaat gcgacttcct gctgaccgcc aattcctggt cctacatcat cgagacttcc 300
aactccaaga acggaacgtg ctaccccggg gagttcactg attacgaaga actgcgcgaa 360
cagctgtcaa ctgtgtcctc gttcgagcgg ttcgaaatct tcccgaaagc cacctcgtgg 420
cccaatcatg aaaccaccaa gggaaccact gtggcctgct cccactctgg ggcgaactca 480
ttctatcgca acttgctgtg gatcgtcaag aagggaaaca gttaccctaa gctgtccaag 540
tcctacacta acaacaaggg aaaggaagtc ctcgtgatct ggggcgtgca tcacccaccc 600
accgactccg atcagcaaac actgtaccag aacaaccaca cctacgtgtc cgtgggatca 660
tccaagtact accagagatt caccccggaa atcgtcgcgc ggcccaaggt ccgcgaacaa 720
gccggccgga tgaactacta ctggatcctg ctggaccagg gcgacactat cactttcgag 780
gcgaccggga acctcatcgc cccttggcac gcattcgcac tgaacaaggg atcgagctcc 840
gggattatga tgagcgatgc tcatgtgcaa aactgtacta ccaagtgcca gacccctcac 900
ggtgccctga agtccaattt gcccttccaa aatgtgcacc ccattaccat tggcgaatgc 960
ccgaaatacg tgaagtcgac ccaactccgg atggccaccg gccttcggaa catcccatcg 1020
atacagtcga ggggtctttt cggagcgatt gcgggattca tcgagggcgg ctggaccgga 1080
atgattgacg gttggtacgg ataccaccac cagaacgaac agggctccgg atacgccgcc 1140
gacaaaaagt cgactcagat cgcaattgac gagattagca acaaggtcaa ctccgtgatc 1200
gagaagatga atatccagtt tactagcgtg ggaaaggagt tcaacaacct ggagaagagg 1260
atcgagaacc tgaacaagaa agtggacgat ggattcctcg acgtgtggac atacaacgcc 1320
gagctcctca ttttgctgga gaacgaacgg accctcgact tccatgactt caacgtgcgc 1380
aacctgtacg aaaaggtcaa gagccagctg agaaacaacg ccaaggaaat tggcaacggt 1440
tgctttgagt tttaccaccg ctgcgacaac gaatgcatgg agagcgtgaa gaacgggacc 1500
tacaactatc cgagatactc agaggaatca aagctgaacc gcgaaaagat cgatggcgtg 1560
aagctcgaat cactcggcgt gcaccagatc ctggccatct attccactgt ggctagctcc 1620
ctcgtgctgc tggtgtccct gggtgctatt tccttctgga tgtgcagcaa cgggtcgctc 1680
cagtgccgga tctgtatctg a 1701
<210> 6
<211> 566
<212> Белок
<213> Вирус гриппа A
<220>
<223> >AKJ81667.1 гемагглютинин [Вирус гриппа A
(A/swine/Italy/28762-3/2013(H1N1))]
<400> 6
Met Lys Ala Lys Leu Phe Val Leu Phe Cys Ala Phe Thr Ala Leu Lys
1 5 10 15
Ala Asp Thr Ile Cys Val Gly Tyr His Ala Asn Asn Ser Thr Asp Thr
20 25 30
Val Asp Thr Ile Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn
35 40 45
Leu Leu Glu Asn Ser His Asn Gly Lys Leu Cys Ser Leu Asn Gly Lys
50 55 60
Ala Pro Leu Gln Leu Glu Asn Cys Asn Val Ala Gly Trp Ile Leu Gly
65 70 75 80
Asn Pro Glu Cys Asp Phe Leu Leu Thr Ala Asn Ser Trp Ser Tyr Ile
85 90 95
Ile Glu Thr Ser Asn Ser Lys Asn Gly Thr Cys Tyr Pro Gly Glu Phe
100 105 110
Thr Asp Tyr Glu Glu Leu Arg Glu Gln Leu Ser Thr Val Ser Ser Phe
115 120 125
Glu Arg Phe Glu Ile Phe Pro Lys Ala Thr Ser Trp Pro Asn His Glu
130 135 140
Thr Thr Lys Gly Thr Thr Val Ala Cys Ser His Ser Gly Ala Asn Ser
145 150 155 160
Phe Tyr Arg Asn Leu Leu Trp Ile Val Lys Lys Gly Asn Ser Tyr Pro
165 170 175
Lys Leu Ser Lys Ser Tyr Thr Asn Asn Lys Gly Lys Glu Val Leu Val
180 185 190
Ile Trp Gly Val His His Pro Pro Thr Asp Ser Asp Gln Gln Thr Leu
195 200 205
Tyr Gln Asn Asn His Thr Tyr Val Ser Val Gly Ser Ser Lys Tyr Tyr
210 215 220
Gln Arg Phe Thr Pro Glu Ile Val Ala Arg Pro Lys Val Arg Glu Gln
225 230 235 240
Ala Gly Arg Met Asn Tyr Tyr Trp Ile Leu Leu Asp Gln Gly Asp Thr
245 250 255
Ile Thr Phe Glu Ala Thr Gly Asn Leu Ile Ala Pro Trp His Ala Phe
260 265 270
Ala Leu Asn Lys Gly Ser Ser Ser Gly Ile Met Met Ser Asp Ala His
275 280 285
Val Gln Asn Cys Thr Thr Lys Cys Gln Thr Pro His Gly Ala Leu Lys
290 295 300
Ser Asn Leu Pro Phe Gln Asn Val His Pro Ile Thr Ile Gly Glu Cys
305 310 315 320
Pro Lys Tyr Val Lys Ser Thr Gln Leu Arg Met Ala Thr Gly Leu Arg
325 330 335
Asn Ile Pro Ser Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly
340 345 350
Phe Ile Glu Gly Gly Trp Thr Gly Met Ile Asp Gly Trp Tyr Gly Tyr
355 360 365
His His Gln Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Lys Lys Ser
370 375 380
Thr Gln Ile Ala Ile Asp Glu Ile Ser Asn Lys Val Asn Ser Val Ile
385 390 395 400
Glu Lys Met Asn Ile Gln Phe Thr Ser Val Gly Lys Glu Phe Asn Asn
405 410 415
Leu Glu Lys Arg Ile Glu Asn Leu Asn Lys Lys Val Asp Asp Gly Phe
420 425 430
Leu Asp Val Trp Thr Tyr Asn Ala Glu Leu Leu Ile Leu Leu Glu Asn
435 440 445
Glu Arg Thr Leu Asp Phe His Asp Phe Asn Val Arg Asn Leu Tyr Glu
450 455 460
Lys Val Lys Ser Gln Leu Arg Asn Asn Ala Lys Glu Ile Gly Asn Gly
465 470 475 480
Cys Phe Glu Phe Tyr His Arg Cys Asp Asn Glu Cys Met Glu Ser Val
485 490 495
Lys Asn Gly Thr Tyr Asn Tyr Pro Arg Tyr Ser Glu Glu Ser Lys Leu
500 505 510
Asn Arg Glu Lys Ile Asp Gly Val Lys Leu Glu Ser Leu Gly Val His
515 520 525
Gln Ile Leu Ala Ile Tyr Ser Thr Val Ala Ser Ser Leu Val Leu Leu
530 535 540
Val Ser Leu Gly Ala Ile Ser Phe Trp Met Cys Ser Asn Gly Ser Leu
545 550 555 560
Gln Cys Arg Ile Cys Ile
565
<210> 7
<211> 1701
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KU323308.1 Вирус гриппа A (A/swine/Italy/240849/2015(H3N2))
ген гемагглютинина (НА) сегмента 4, полная кодирующая
последовательность
<400> 7
atgaagactg tcattgcttt aagctacgtc ttttgcctgg tttttggcca agattttcca 60
ggaaagggca acaacacagc aacactgtgc ctagggcatc atgcagtgcc aaatgggaca 120
ttagtgaaaa caatcacaga tgatcaaatc gaagtaacca atgccacaga actggtccaa 180
aatttctcaa tgggtaaaat atgcaagaat cctcatcgaa ttcttgatgg ggcaaattgt 240
acattggtag actctctatt gggggaccct cactgtgatg gttttcaaaa tgagaaatgg 300
gacctcttca ttgaacgcag cagggctttc agcaactgct acccttatga tgtgccggag 360
tatacctctc ttaggtcact aattgcctca tcaggcacct tggaatttac caatgaaaat 420
ttcaattgga ctggagttac ccaaaatggg ggaagcagtg cttgcaagag ggggcccaac 480
aacagctttt tcagtagatt gaactggttg tacaaatcag gaaacacata tccgatgcta 540
aatgttacta tgccaaacag tgatgatttc gacaaattgt acatttgggg agttcatcat 600
ccgagcaccg atagagaaca gaccaacctg tacattcaag catcagggaa aataatagta 660
tccacaaaga gaagtcaaca aaccataatc ccgaatatcg ggtctagacc ctgggtaagg 720
ggtttatcta gtaggataag catctactgg acaatagtta aaccaggaga catcctgata 780
atcaacagca atggaaacct aattgctcct cggggttact tcaagataca gactgggaaa 840
agttcagtaa tgaagtcaga tgcacctatt ggcacctgca attcagaatg cattactcca 900
aacggaagca tacccaatga taaacccttt caaaacgtaa acaggatcac atatggggca 960
tgccctcatt acatcaagca aaacactctg aagctagcaa caggaatgcg gaatatacca 1020
gaaagacaaa ccagaggtat attcggtgca atagcagggt tcatagagaa tggttgggaa 1080
ggaatggtga acggttggta cggtttcaga caccaaaatt ctgagggcat aggacaggca 1140
gcagatctta aaagtaccca agcagctatc aaccaaatca atggaaaact gaatagagta 1200
atcgagaaaa caaacgagaa attccatcaa atcgaaaagg agttctcaga agtagaagga 1260
aggatccaag accttgaaag atatgttgaa gatactaaga tagatctttg gtcttacaac 1320
gcggaacttc ttgtcgcact agagaatcaa cacacaattg acctgactga ctcagaaatg 1380
aacaaactct ttgaaaaaac aaggaaacaa ctaagggaaa atgctgagga catgggaaat 1440
ggttgcttca aaatatacca caaatgtgac aactcttgca tggagtcaat cagaaatggg 1500
acttacgacc ataatgaata cagagacgaa gcagtaaaca atcgattcca aatcaaaagt 1560
gttgagctga agtcaggata caaagactgg atcctgtgga tttcctttgc catatcatgc 1620
tttttgcttt gcgctatttg gatgggattc gtaatatggg cctgccagaa aggcaacatt 1680
agatgcaata tttgcatctg a 1701
<210> 8
<211> 1701
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUH3-4_Оптимизированная последовательность свиньи
<400> 8
atgaaaaccg tcattgcgtt gagctatgtg ttttgtctcg tgtttggaca agacttcccg 60
ggaaagggca acaacaccgc caccctgtgc ctggggcacc atgccgtgcc caacggaact 120
ctcgtcaaga ccatcaccga cgaccagatc gaggtcacca atgcgaccga gttggtgcag 180
aatttctcca tgggaaagat ctgcaagaac ccgcaccgga ttttggatgg agccaactgc 240
acactggtgg actccctgct gggtgatcct cattgcgacg gattccaaaa cgagaagtgg 300
gatctgttca tcgagagatc ccgggccttc agcaactgct acccgtacga cgtgccagag 360
tacactagct tgcggtccct gatcgcctcc agcggtaccc tggagttcac caacgagaac 420
ttcaactgga ccggcgtgac gcagaacggg ggttcctcag cgtgcaagcg gggaccgaac 480
aactcgttct tctcccggct taactggctg tacaagagcg gcaacacata ccccatgctg 540
aacgtgacca tgcccaactc tgacgacttt gataagctgt acatctgggg agtgcaccac 600
ccatcgactg accgcgaaca gaccaacctg tacatccaag cctccggcaa aatcattgtg 660
tccaccaaga gatcgcagca gaccatcatt ccgaacattg gcagtcggcc atgggtcaga 720
ggactgtcgt ccagaatcag catctactgg accattgtga agcccggcga catcctcatc 780
ataaactcga acggcaacct gatcgcccca cggggatatt tcaagatcca gaccggaaag 840
agctccgtga tgaagtccga tgccccgatt gggacttgca actccgagtg catcaccccc 900
aacggctcca tccccaacga caaacccttc caaaacgtga accgcatcac ttacggagcc 960
tgcccgcatt acatcaagca gaacacgctc aagctcgcca ctgggatgcg caacattcct 1020
gagcgccaaa cccggggaat cttcggagcc attgcgggct tcatcgaaaa cggctgggaa 1080
gggatggtca acggttggta cggttttaga caccaaaatt ccgaaggaat cggacaggct 1140
gccgatctca aatcaaccca ggccgccatc aaccagatca acgggaagct gaacagagtg 1200
atcgaaaaga ccaacgaaaa gttccatcag attgagaagg agttttccga agtcgaggga 1260
cggattcaag acctggagcg ctacgtggag gacaccaaga ttgatctgtg gagctacaac 1320
gcagagctcc tggtcgcgct ggaaaatcag cacaccattg acttgaccga ctcggagatg 1380
aacaagctgt ttgaaaagac caggaagcag ctgcgggaaa acgccgaaga tatggggaac 1440
ggatgcttca aaatctacca taagtgcgac aatagctgca tggaaagcat taggaacggg 1500
acttacgacc ataacgagta cagagatgaa gcagtgaaca acagattcca gatcaagtcc 1560
gtggaactca agagcggtta caaggactgg atcctctgga tttcgttcgc tatttcgtgc 1620
ttcctgctgt gcgccatctg gatgggattc gtcatctggg cctgccagaa gggcaacatc 1680
cggtgcaaca tttgtatttg a 1701
<210> 9
<211> 566
<212> Белок
<213> Вирус гриппа A
<220>
<223> >ALX30415.1 гемагглютинин [Вирус гриппа A
(A/swine/Italy/240849/2015(H3N2))] EUH3-4
<400> 9
Met Lys Thr Val Ile Ala Leu Ser Tyr Val Phe Cys Leu Val Phe Gly
1 5 10 15
Gln Asp Phe Pro Gly Lys Gly Asn Asn Thr Ala Thr Leu Cys Leu Gly
20 25 30
His His Ala Val Pro Asn Gly Thr Leu Val Lys Thr Ile Thr Asp Asp
35 40 45
Gln Ile Glu Val Thr Asn Ala Thr Glu Leu Val Gln Asn Phe Ser Met
50 55 60
Gly Lys Ile Cys Lys Asn Pro His Arg Ile Leu Asp Gly Ala Asn Cys
65 70 75 80
Thr Leu Val Asp Ser Leu Leu Gly Asp Pro His Cys Asp Gly Phe Gln
85 90 95
Asn Glu Lys Trp Asp Leu Phe Ile Glu Arg Ser Arg Ala Phe Ser Asn
100 105 110
Cys Tyr Pro Tyr Asp Val Pro Glu Tyr Thr Ser Leu Arg Ser Leu Ile
115 120 125
Ala Ser Ser Gly Thr Leu Glu Phe Thr Asn Glu Asn Phe Asn Trp Thr
130 135 140
Gly Val Thr Gln Asn Gly Gly Ser Ser Ala Cys Lys Arg Gly Pro Asn
145 150 155 160
Asn Ser Phe Phe Ser Arg Leu Asn Trp Leu Tyr Lys Ser Gly Asn Thr
165 170 175
Tyr Pro Met Leu Asn Val Thr Met Pro Asn Ser Asp Asp Phe Asp Lys
180 185 190
Leu Tyr Ile Trp Gly Val His His Pro Ser Thr Asp Arg Glu Gln Thr
195 200 205
Asn Leu Tyr Ile Gln Ala Ser Gly Lys Ile Ile Val Ser Thr Lys Arg
210 215 220
Ser Gln Gln Thr Ile Ile Pro Asn Ile Gly Ser Arg Pro Trp Val Arg
225 230 235 240
Gly Leu Ser Ser Arg Ile Ser Ile Tyr Trp Thr Ile Val Lys Pro Gly
245 250 255
Asp Ile Leu Ile Ile Asn Ser Asn Gly Asn Leu Ile Ala Pro Arg Gly
260 265 270
Tyr Phe Lys Ile Gln Thr Gly Lys Ser Ser Val Met Lys Ser Asp Ala
275 280 285
Pro Ile Gly Thr Cys Asn Ser Glu Cys Ile Thr Pro Asn Gly Ser Ile
290 295 300
Pro Asn Asp Lys Pro Phe Gln Asn Val Asn Arg Ile Thr Tyr Gly Ala
305 310 315 320
Cys Pro His Tyr Ile Lys Gln Asn Thr Leu Lys Leu Ala Thr Gly Met
325 330 335
Arg Asn Ile Pro Glu Arg Gln Thr Arg Gly Ile Phe Gly Ala Ile Ala
340 345 350
Gly Phe Ile Glu Asn Gly Trp Glu Gly Met Val Asn Gly Trp Tyr Gly
355 360 365
Phe Arg His Gln Asn Ser Glu Gly Ile Gly Gln Ala Ala Asp Leu Lys
370 375 380
Ser Thr Gln Ala Ala Ile Asn Gln Ile Asn Gly Lys Leu Asn Arg Val
385 390 395 400
Ile Glu Lys Thr Asn Glu Lys Phe His Gln Ile Glu Lys Glu Phe Ser
405 410 415
Glu Val Glu Gly Arg Ile Gln Asp Leu Glu Arg Tyr Val Glu Asp Thr
420 425 430
Lys Ile Asp Leu Trp Ser Tyr Asn Ala Glu Leu Leu Val Ala Leu Glu
435 440 445
Asn Gln His Thr Ile Asp Leu Thr Asp Ser Glu Met Asn Lys Leu Phe
450 455 460
Glu Lys Thr Arg Lys Gln Leu Arg Glu Asn Ala Glu Asp Met Gly Asn
465 470 475 480
Gly Cys Phe Lys Ile Tyr His Lys Cys Asp Asn Ser Cys Met Glu Ser
485 490 495
Ile Arg Asn Gly Thr Tyr Asp His Asn Glu Tyr Arg Asp Glu Ala Val
500 505 510
Asn Asn Arg Phe Gln Ile Lys Ser Val Glu Leu Lys Ser Gly Tyr Lys
515 520 525
Asp Trp Ile Leu Trp Ile Ser Phe Ala Ile Ser Cys Phe Leu Leu Cys
530 535 540
Ala Ile Trp Met Gly Phe Val Ile Trp Ala Cys Gln Lys Gly Asn Ile
545 550 555 560
Arg Cys Asn Ile Cys Ile
565
<210> 10
<211> 1701
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >CY116127.1 Вирус гриппа A (A/swine/England/373/2010(H1N1))
ген гемагглютинина (HA), полная кодирующая последовательность
<400> 10
atgaaggcaa tactagtagt tctgctacat acatttgcaa ccgcaaatgc agacacatta 60
tgtataggtt atcatgcgaa caattcaaca gacactgtag acacagtact agaaaagaat 120
gtaacagtaa cacactctgt taaccttcta gaagacaagc ataacgggaa actatgcaaa 180
ctaagagggg tagccccatt gcatttgggt aaatgtaaca ttgctggctg gatcctggga 240
aatccagagt gtgaatcact ctccacagca agctcatggt cctacattgt ggaaacatct 300
agttcagaca atggaacgtg ttacccagga gatttcatcg attatgagga gctaagagag 360
caattgagct cagtgtcatc atttgaaagg tttgagatat tccccaagac aagttcatgg 420
cccaatcatg actcgaacaa aggtgtaacg gcagcatgtc ctcatgctgg agcaaaaagc 480
ttctacaaaa atttaatatg gctagttaaa aaaggaaatt catacccaaa gctcagcaaa 540
tcctacatta atgataaagg gaaagaagtc ctcgtgctat ggggcattca ccatccatct 600
actagtgctg accaacaaag tctctatcag aatgcagatg catatgtttt tgtggggaca 660
tcaagataca gcaagaagtt caagccggaa atagcaataa gacccaaagt gagggatcaa 720
gaagggagaa tgaactatta ctggacacta gtagagccgg gagacaaaat aacattcgaa 780
gcaactggaa atctagtggt accgagatat gcattcgcaa tggaaagaaa tgctggatct 840
ggtattatca tttcagatac accagtccac gattgcaata caacttgtca gacacccaag 900
ggtgctataa acaccagcct cccatttcag aatatacatc cgatcacaat tggaaaatgt 960
ccaaaatatg taaaaagcac aaaattgaga ctggccacag gattgaggaa cgtcccgtct 1020
attcaatcta gaggcctatt tggggccatt gccggtttca ttgaaggggg gtggacaggg 1080
atggtagatg gatggtacgg ttatcaccat caaaatgagc aggggtcagg atatgcagcc 1140
gacctgaaga gcacacagca tgccattgac gagattacta acaaagtaaa ttctgttatt 1200
gaaaagatga atacacagtt cacagcagta ggtaaagagt tcaaccacct ggaaaaaaga 1260
atagagaatt taaataaaaa agttgatgat ggtttcctgg acatttggac ttacaatgcc 1320
gaactgttgg ttctattgga aaatgaaaga actttggact accacgattc aaatgtgaag 1380
aacttatatg aaaaggtaag aagccagtta aaaaacaatg ccaaggaaat tggaaacggc 1440
tgctttgaat tttaccacaa atgcgataac acgtgcatgg aaagtgtcaa aaatgggact 1500
tatgactacc caaaatactc agaggaagca aaattaaaca gagaagaaat agatggggta 1560
aagctagaat caacaaggat ttaccagatt ttggcgatct attcaactgt cgccagttca 1620
ttggtactgg tagtctccct gggggcaatc agtttctgga tgtgctctaa tgggtctcta 1680
cagtgtagaa tatgtattta a 1701
<210> 11
<211> 1701
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUH1-11 Оптимизированная последовательность свиньи
<400> 11
atgaaggcta ttctggtggt cctgctccac acttttgcca ccgcgaacgc agataccctc 60
tgcatcggat accacgccaa caattccacc gatactgtgg ataccgtgct ggaaaagaac 120
gtgaccgtga ctcactcagt gaacctcctg gaagataagc acaacggaaa gctttgcaag 180
cttcggggag tggctccgtt gcatctggga aagtgcaaca tcgccggatg gatcctgggc 240
aaccccgagt gcgagagcct gtccaccgcc tcctcctggt cttacattgt ggaaacctcc 300
tcgtccgata acggcacatg ctatccgggg gacttcattg actacgaaga actgagggag 360
caactcagct ccgtgagctc ctttgagcgc ttcgaaatct tccctaagac ctcatcctgg 420
cccaaccacg actccaacaa gggagtgact gcagcgtgtc cgcacgctgg agccaagtca 480
ttctacaaga acctgatctg gctggtcaaa aagggaaact cgtaccccaa gctgtctaag 540
tcctacatca atgacaaggg gaaggaagtg ctggtgctgt ggggaatcca ccacccttcc 600
acctccgccg accagcagtc gctgtaccag aacgctgatg cgtacgtgtt cgtcggcact 660
agccgctact ccaagaagtt caagccagaa attgccatcc ggcccaaagt ccgcgatcag 720
gaggggcgga tgaactacta ctggactctc gtggaacccg gggataagat cacgttcgaa 780
gctaccggca acctcgtggt gcctcgctac gccttcgcaa tggagaggaa tgctggaagc 840
ggtatcatca tttcggacac tcctgtgcat gactgcaata cgacttgtca gactccgaag 900
ggagcgatca acacttccct cccgttccaa aacatccacc ccattacaat tggcaaatgt 960
cctaaatacg tgaagtcgac caagctccgg cttgcaactg ggctgcgcaa tgtgcctagc 1020
attcagtcaa ggggcctatt tggggccatc gccggtttca tcgagggagg atggaccgga 1080
atggtggacg gttggtatgg ctaccaccac caaaacgaac agggctccgg ttacgccgcc 1140
gacctgaagt ccacccagca cgccattgac gaaatcacca acaaggtcaa ctccgtcatc 1200
gagaagatga acacccagtt cactgccgtg ggaaaggagt tcaaccatct ggagaagaga 1260
attgagaacc tcaataagaa agtggacgac ggcttcctgg acatctggac ctacaacgcc 1320
gagctgctgg tcctgctgga aaacgagagg accctggact accacgactc gaacgtgaag 1380
aacctgtacg agaaggtccg atcacaactg aagaacaacg ccaaggaaat cggaaatggg 1440
tgcttcgagt tctaccacaa atgcgacaat acttgcatgg aatccgtgaa gaatggcacc 1500
tacgattacc ctaagtattc cgaggaagcg aagctgaacc gggaagagat tgacggcgtg 1560
aagctcgaat caacccggat ctaccagatc cttgcgatct actccaccgt ggccagctca 1620
ctcgtgctcg tggtgtccct gggtgccatc tccttctgga tgtgtagcaa cggcagcctc 1680
cagtgccgca tctgtatctg a 1701
<210> 12
<211> 566
<212> Белок
<213> Вирус гриппа A
<220>
<223> >AFR76205.1 гемагглютинин [Вирус гриппа A
(A/swine/England/373/2010(H1N1) EUH1-11
<400> 12
Met Lys Ala Ile Leu Val Val Leu Leu His Thr Phe Ala Thr Ala Asn
1 5 10 15
Ala Asp Thr Leu Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr
20 25 30
Val Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn
35 40 45
Leu Leu Glu Asp Lys His Asn Gly Lys Leu Cys Lys Leu Arg Gly Val
50 55 60
Ala Pro Leu His Leu Gly Lys Cys Asn Ile Ala Gly Trp Ile Leu Gly
65 70 75 80
Asn Pro Glu Cys Glu Ser Leu Ser Thr Ala Ser Ser Trp Ser Tyr Ile
85 90 95
Val Glu Thr Ser Ser Ser Asp Asn Gly Thr Cys Tyr Pro Gly Asp Phe
100 105 110
Ile Asp Tyr Glu Glu Leu Arg Glu Gln Leu Ser Ser Val Ser Ser Phe
115 120 125
Glu Arg Phe Glu Ile Phe Pro Lys Thr Ser Ser Trp Pro Asn His Asp
130 135 140
Ser Asn Lys Gly Val Thr Ala Ala Cys Pro His Ala Gly Ala Lys Ser
145 150 155 160
Phe Tyr Lys Asn Leu Ile Trp Leu Val Lys Lys Gly Asn Ser Tyr Pro
165 170 175
Lys Leu Ser Lys Ser Tyr Ile Asn Asp Lys Gly Lys Glu Val Leu Val
180 185 190
Leu Trp Gly Ile His His Pro Ser Thr Ser Ala Asp Gln Gln Ser Leu
195 200 205
Tyr Gln Asn Ala Asp Ala Tyr Val Phe Val Gly Thr Ser Arg Tyr Ser
210 215 220
Lys Lys Phe Lys Pro Glu Ile Ala Ile Arg Pro Lys Val Arg Asp Gln
225 230 235 240
Glu Gly Arg Met Asn Tyr Tyr Trp Thr Leu Val Glu Pro Gly Asp Lys
245 250 255
Ile Thr Phe Glu Ala Thr Gly Asn Leu Val Val Pro Arg Tyr Ala Phe
260 265 270
Ala Met Glu Arg Asn Ala Gly Ser Gly Ile Ile Ile Ser Asp Thr Pro
275 280 285
Val His Asp Cys Asn Thr Thr Cys Gln Thr Pro Lys Gly Ala Ile Asn
290 295 300
Thr Ser Leu Pro Phe Gln Asn Ile His Pro Ile Thr Ile Gly Lys Cys
305 310 315 320
Pro Lys Tyr Val Lys Ser Thr Lys Leu Arg Leu Ala Thr Gly Leu Arg
325 330 335
Asn Val Pro Ser Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly
340 345 350
Phe Ile Glu Gly Gly Trp Thr Gly Met Val Asp Gly Trp Tyr Gly Tyr
355 360 365
His His Gln Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Leu Lys Ser
370 375 380
Thr Gln His Ala Ile Asp Glu Ile Thr Asn Lys Val Asn Ser Val Ile
385 390 395 400
Glu Lys Met Asn Thr Gln Phe Thr Ala Val Gly Lys Glu Phe Asn His
405 410 415
Leu Glu Lys Arg Ile Glu Asn Leu Asn Lys Lys Val Asp Asp Gly Phe
420 425 430
Leu Asp Ile Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Leu Glu Asn
435 440 445
Glu Arg Thr Leu Asp Tyr His Asp Ser Asn Val Lys Asn Leu Tyr Glu
450 455 460
Lys Val Arg Ser Gln Leu Lys Asn Asn Ala Lys Glu Ile Gly Asn Gly
465 470 475 480
Cys Phe Glu Phe Tyr His Lys Cys Asp Asn Thr Cys Met Glu Ser Val
485 490 495
Lys Asn Gly Thr Tyr Asp Tyr Pro Lys Tyr Ser Glu Glu Ala Lys Leu
500 505 510
Asn Arg Glu Glu Ile Asp Gly Val Lys Leu Glu Ser Thr Arg Ile Tyr
515 520 525
Gln Ile Leu Ala Ile Tyr Ser Thr Val Ala Ser Ser Leu Val Leu Val
530 535 540
Val Ser Leu Gly Ala Ile Ser Phe Trp Met Cys Ser Asn Gly Ser Leu
545 550 555 560
Gln Cys Arg Ile Cys Ile
565
<210> 13
<211> 1410
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KR700793.1 Вирус гриппа A (A/swine/England/61470/2013(H1N2))
ген нейраминидазы (NА) сегмента 6, полная кодирующая
последовательность
<400> 13
atgaatccaa atcagaagat aataacaatt ggctctgttt ctctcgtcat tgcaacatta 60
tgcttcttaa tgcagatggc catcctaata actactgtaa aattacattt caaacaatat 120
gagtgcggct tccctgcgaa caaccaagta ataacatgtg agccaacagt aatagaaagg 180
aacacaacag agatagtgta cttaactaac accaccatag agaaagaaac atgccacaaa 240
acagtggaat acaggaattg gtcaaagcct caatgcaaaa taacaggctt tgcacctttt 300
tccaaggaca attcaattcg actttctgct ggtggggaca tatgggtgac gagggaacct 360
tacgtgtcat gcgagcctgg caaatgttat cagtttgcac tcgggcaagg gaccacacta 420
gacaataaac attcaaacga tacaatacat gacagaaccc cctatcgaac tctattgatg 480
aatgaattgg gtgtcccatt tcatttaggg acaagacaag tgtgtattgc atggtccagc 540
tcaagttgtt atgatgggaa agcatggttg catgtctgta tcactggaca tgataaaaat 600
gcaactgcca gtttcattta cgatggtaga cttgtagata gcattggttc atggtctaaa 660
aatatactta gaacccagga atcagaatgc gtttgcatca atggggtctg tacagtagta 720
atgactgatg gaagtgcttc gggaagagct gatactaaaa tactattcat tgaagaaggg 780
aaaattgttc atattagccc attagcgggg agtgcacagc atgtggagga gtgctcctgt 840
tatccccgat atcctggcgt aaggtgtatc tgcagagaca actggaaagg ctctaacaga 900
cccgttgtgg atataaatat agaagattat agcattgatt ccagttatgt gtgttcaggg 960
cttgttggcg acacacccag aatcaatgac ggatccagta gtagctactg ccgtgatcct 1020
aacaacgaaa aaggaaatca cggagtgaag ggctgggctt ttgacgatgg aaatgatgtg 1080
tggatgggaa gaacgatcaa cgaagattca cgctcaggtt atgaaacatt caaagtcatt 1140
ggtggttggt ccactcctaa ttccaaattg cagataaata ggcaagtaat agttgatagc 1200
aacaataggt caggttattc tggtgttttc tccgttgaag gcaaaagctg catcaataga 1260
tgtttctacg tggagttgat aagaggaaga aggtcagaag cgcgagtatg gtggacctca 1320
aacagtattg ttgtattttg tggcacttca ggtacctatg gaacaggctc atggcctgat 1380
ggagcagaca tcaacctcat gcctatatga 1410
<210> 14
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUN2-6 Оптимизированная последовательность свиньи
<400> 14
atgaatccga accagaaaat tatcaccatt gggtccgtca gccttgtgat tgccactctg 60
tgctttctga tgcaaatggc catcctcatt accactgtga agctgcattt caagcagtac 120
gaatgcggct tccccgcgaa caaccaggtc atcacttgcg agccaaccgt gattgagcgg 180
aacactaccg aaattgtata cctcacgaac actaccattg aaaaggagac ttgccacaag 240
accgtggaat accggaactg gtccaagcct cagtgcaaga ttaccgggtt cgccccattc 300
tccaaggaca atagcatccg cctttcggcc ggaggagata tttgggtgac gcgcgaaccc 360
tacgtcagct gcgaacccgg gaagtgttac cagttcgcac tgggccaggg aaccaccctg 420
gataataagc acagcaacga caccatccac gaccggactc cctaccgcac cctcctgatg 480
aatgagctcg gcgtgccgtt ccacctgggt accagacagg tctgcatcgc ctggtcgtcc 540
tcctcatgct acgatgggaa ggcttggctc cacgtgtgca taaccggtca tgacaagaac 600
gccacagcct cattcatcta cgacggacgg ctagtggact ccatcggctc ctggtccaag 660
aacattctga ggacccagga atccgaatgc gtctgtatca acggagtgtg caccgtggtg 720
atgaccgatg gctccgcaag cggaagggcc gatactaaga tcctgttcat cgaggaggga 780
aagattgtgc acatctctcc tctcgcggga tcagcccagc acgtggaaga atgttcctgt 840
taccctcgct acccgggagt gcgctgtatt tgccgggaca actggaaagg ctccaacagg 900
cctgtggtgg acattaatat cgaggactac tccatcgata gctcctacgt gtgctccggt 960
ctggtcgggg acactccaag aatcaacgat ggaagctcca gcagctactg ccgggacccc 1020
aataacgaaa aggggaacca cggcgtgaag ggttgggcgt tcgacgatgg aaatgacgtc 1080
tggatgggac ggactatcaa cgaggactca cggagcggat acgaaacctt taaggtcatc 1140
ggcggatggt cgaccccgaa cagcaagctg cagatcaacc gccaagtgat cgtggactcc 1200
aacaaccgct cggggtatag cggggtgttc tccgtggagg gaaagtcctg catcaaccgg 1260
tgcttctacg tcgaactgat cagaggccgg agatcagaag cccgcgtgtg gtggacctcc 1320
aactccattg tggtgttttg cggcacttcc gggacctatg gcactggatc ctggcccgat 1380
ggagcagaca tcaacctgat gccaatctga 1410
<210> 15
<211> 469
<212> Белок
<213> Вирус гриппа A
<220>
<223> >AKJ82042.1 нейраминидаза [Вирус гриппа A
(A/swine/England/61470/2013(H1N2))] EUN2-6
<400> 15
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Val
1 5 10 15
Ile Ala Thr Leu Cys Phe Leu Met Gln Met Ala Ile Leu Ile Thr Thr
20 25 30
Val Lys Leu His Phe Lys Gln Tyr Glu Cys Gly Phe Pro Ala Asn Asn
35 40 45
Gln Val Ile Thr Cys Glu Pro Thr Val Ile Glu Arg Asn Thr Thr Glu
50 55 60
Ile Val Tyr Leu Thr Asn Thr Thr Ile Glu Lys Glu Thr Cys His Lys
65 70 75 80
Thr Val Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Lys Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Asp Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Glu Pro Gly Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asp Asn Lys His
130 135 140
Ser Asn Asp Thr Ile His Asp Arg Thr Pro Tyr Arg Thr Leu Leu Met
145 150 155 160
Asn Glu Leu Gly Val Pro Phe His Leu Gly Thr Arg Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys Tyr Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Ile Thr Gly His Asp Lys Asn Ala Thr Ala Ser Phe Ile Tyr Asp
195 200 205
Gly Arg Leu Val Asp Ser Ile Gly Ser Trp Ser Lys Asn Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asn Gly Val Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Arg Ala Asp Thr Lys Ile Leu Phe
245 250 255
Ile Glu Glu Gly Lys Ile Val His Ile Ser Pro Leu Ala Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Gly Val Arg
275 280 285
Cys Ile Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Val Val Asp
290 295 300
Ile Asn Ile Glu Asp Tyr Ser Ile Asp Ser Ser Tyr Val Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Ile Asn Asp Gly Ser Ser Ser Ser Tyr
325 330 335
Cys Arg Asp Pro Asn Asn Glu Lys Gly Asn His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Val Trp Met Gly Arg Thr Ile Asn Glu
355 360 365
Asp Ser Arg Ser Gly Tyr Glu Thr Phe Lys Val Ile Gly Gly Trp Ser
370 375 380
Thr Pro Asn Ser Lys Leu Gln Ile Asn Arg Gln Val Ile Val Asp Ser
385 390 395 400
Asn Asn Arg Ser Gly Tyr Ser Gly Val Phe Ser Val Glu Gly Lys Ser
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Arg Ser
420 425 430
Glu Ala Arg Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Thr Gly Ser Trp Pro Asp Gly Ala Asp Ile
450 455 460
Asn Leu Met Pro Ile
465
<210> 16
<211> 1410
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KU323318.1 Вирус гриппа A (A/swine/Italy/248147-8/2015(H3N2))
ген нейраминидазы (NА) сегмента 6, полная кодирующая
последовательность
<400> 16
atgaatccaa atcaaaagat aataacaatt ggttctgttt ctctcactat tacaacaatg 60
tgcctcttct tgcagattgc catcctagta actactataa cattgcattt caagcaatat 120
gaatgcgatt cccctgcaaa caaccaagta ataccgtgtg aaccaataat aatagaaaaa 180
aacataacaa aaatagtgta tttgaccaat accaccatag agaaagaggt atgcccaaaa 240
ttaggggaat acaggaattg gtcaaaacca caatgcaaga tcacaggatt tgcacctttt 300
tctaaggaca attcaattcg gctctctgcg ggtggggcca tttgggtcac gagagaacct 360
tatgtgtcat gcgaccctaa caagtgttat caatttgcat taggacaggg aaccacatta 420
gataacagac attcaaatga cacaatacat gatagaaccc cttttagaac cctgttgatg 480
agtgaattag gtgttccatt tcatttggga accagacaag tatgcatagc atggtccagt 540
tcaagttgtc acgatgggaa agcttggttg catgtttgtg tcactgggca tgataaaaat 600
gcaactgcta gtttcattta tgacggaaag cttgtagaca gcatcagttc atggtccaaa 660
aacatactcc ggactcagga atcagaatgt gtttgtatcg atggaatctg tacagtggtg 720
atgactgatg gaagtgcttc agggaaagct gatactaaga tactatttat tgaaaaaggg 780
aagatcattc atattagtcc attgttggga agtgctcagc atgtagaaga atgttcctgt 840
taccctagat accctgatgt caggtgtatt tgcagggata actggaaagg ttcaaatagg 900
cccatcgtag acataagaat gaaaaattat agcattggtt ccagttatat gtgctcagga 960
cttgttggcg acacacccag gaacaatgat gggtctagta atagcaattg tcggaatccc 1020
aataatgaaa gaggaaatca tggagtgaaa ggttgggcct ttgatgatgg aaatgacaca 1080
tggatgggaa gaactatcag caaggactca cgcttaggtt acgaaacctt caaagttgtt 1140
ggtggttggt cccaacccaa ttccaaatcc cagataaata gacaagttat tgttgacagc 1200
gataatagat caggttactc tggtattttc tctgttgagg ggaaagattg cattaatagg 1260
tgtttttatg tggaactaat aagaggaagg agacaggaaa ctagagtgtg gtggacttcg 1320
aacagtattg ttgtgttctg tggcacttct ggcacctatg ggtcaggctc atggcccgat 1380
ggagcaaaca tcaatttcat gcctgtataa 1410
<210> 17
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUN2-7 Оптимизированная последовательность свиньи
<400> 17
atgaacccta atcagaagat catcacaatc ggctccgtgt ccctcaccat caccactatg 60
tgcctcttcc ttcaaatcgc cattctggtc accaccatca ccctgcattt caagcagtac 120
gaatgcgact caccagcgaa caatcaggtc atcccgtgtg aaccgatcat tatcgagaag 180
aacatcacta agattgtgta tttgaccaac accaccattg agaaggaagt ctgcccaaag 240
ctcggggaat accgcaactg gagcaagccc cagtgcaaga ttaccggctt cgccccgttc 300
tcaaaagaca actccatccg gctgtctgcg ggaggagcca tctgggtcac gagagaaccg 360
tacgtgtcgt gcgaccccaa caagtgctac cagttcgccc tgggacaagg gaccaccctg 420
gacaaccggc actccaacga cactatacat gataggactc ccttccggac ccttctcatg 480
agcgaactgg gggtcccctt ccatctcgga accagacaag tctgcattgc gtggagctcc 540
tcctcctgcc acgacggaaa agcttggctc cacgtctgcg tgacgggcca cgacaagaac 600
gccactgcct ccttcatcta cgatgggaag cttgtggact ccatctcgtc atggagcaag 660
aacatcctgc ggactcagga gtcagaatgc gtgtgcatcg acggcatttg caccgtcgtg 720
atgactgacg gttcggccag cggaaaggcc gacaccaaga tcctgttcat cgagaaggga 780
aagatcattc acatctcccc acttctgggt tccgcgcaac acgtggaaga atgctcctgc 840
tacccccgct accccgatgt ccggtgcatc tgccgggaca actggaaggg ttccaaccgg 900
ccaatcgtgg acatccgcat gaagaactat tccatcgggt cctcttacat gtgctcgggt 960
ctggtcggag atacaccgcg caacaacgac gggtcctcaa acagcaactg ccggaaccct 1020
aacaacgaga ggggcaacca cggcgtgaaa ggatgggctt tcgacgacgg aaatgacact 1080
tggatgggga gaaccattag caaggatagc aggcttggct acgaaacctt caaggtcgtg 1140
ggcggttggt cgcagccgaa ctcgaagtcg cagatcaaca gacaggtcat cgtggactcg 1200
gataacagat cggggtactc agggatcttc tccgtggagg gaaaggactg catcaataga 1260
tgtttctacg tcgagctgat tcgaggccgg agacaggaaa cccgcgtatg gtggacgtcc 1320
aactcaatcg tcgtgttttg cggtacctcc ggaacttacg gctcgggatc atggcccgat 1380
ggagccaata ttaacttcat gcctgtgtga 1410
<210> 18
<211> 469
<212> Белок
<213> Вирус гриппа A
<220>
<223> >ALX30429.1 нейраминидаза [Вирус гриппа A
(A/swine/Italy/248147-8/2015(H3N2))] EUN2-7
<400> 18
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Thr
1 5 10 15
Ile Thr Thr Met Cys Leu Phe Leu Gln Ile Ala Ile Leu Val Thr Thr
20 25 30
Ile Thr Leu His Phe Lys Gln Tyr Glu Cys Asp Ser Pro Ala Asn Asn
35 40 45
Gln Val Ile Pro Cys Glu Pro Ile Ile Ile Glu Lys Asn Ile Thr Lys
50 55 60
Ile Val Tyr Leu Thr Asn Thr Thr Ile Glu Lys Glu Val Cys Pro Lys
65 70 75 80
Leu Gly Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Lys Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Ala Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Asp Pro Asn Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asp Asn Arg His
130 135 140
Ser Asn Asp Thr Ile His Asp Arg Thr Pro Phe Arg Thr Leu Leu Met
145 150 155 160
Ser Glu Leu Gly Val Pro Phe His Leu Gly Thr Arg Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys His Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Val Thr Gly His Asp Lys Asn Ala Thr Ala Ser Phe Ile Tyr Asp
195 200 205
Gly Lys Leu Val Asp Ser Ile Ser Ser Trp Ser Lys Asn Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asp Gly Ile Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Lys Ala Asp Thr Lys Ile Leu Phe
245 250 255
Ile Glu Lys Gly Lys Ile Ile His Ile Ser Pro Leu Leu Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Asp Val Arg
275 280 285
Cys Ile Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Ile Val Asp
290 295 300
Ile Arg Met Lys Asn Tyr Ser Ile Gly Ser Ser Tyr Met Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Asn Asn Asp Gly Ser Ser Asn Ser Asn
325 330 335
Cys Arg Asn Pro Asn Asn Glu Arg Gly Asn His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Thr Trp Met Gly Arg Thr Ile Ser Lys
355 360 365
Asp Ser Arg Leu Gly Tyr Glu Thr Phe Lys Val Val Gly Gly Trp Ser
370 375 380
Gln Pro Asn Ser Lys Ser Gln Ile Asn Arg Gln Val Ile Val Asp Ser
385 390 395 400
Asp Asn Arg Ser Gly Tyr Ser Gly Ile Phe Ser Val Glu Gly Lys Asp
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Arg Gln
420 425 430
Glu Thr Arg Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Ser Gly Ser Trp Pro Asp Gly Ala Asn Ile
450 455 460
Asn Phe Met Pro Val
465
<210> 19
<211> 1410
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KU323247.1 Вирус гриппа A (A/swine/Italy/179057/2015(H1N1))
ген нейраминидазы (NА) сегмента 6, полная кодирующая
последовательность
<400> 19
atgaatccaa accaaaagat aataaccatt ggttcggtct gtatgacaat tggaatggct 60
aacttaatat tacaaattgg aaacataatc tcaatatgga ttagccactc aattcaactt 120
gggaatcaaa gtcagattga aacatgcaat caaagcgtca ttacttatga aaacaacact 180
tgggtaaatc agacatatgt taacatcagc aacaccaact ttgctgctgg acagtcagtg 240
gtttccgcga aattagcggg caattcctcc ctctgccctg ttagtggatg ggctatatac 300
agtaaagaca acagtgtaag aatcggttcc aagggggatg tgtttgtcat aagggaacca 360
ttcatatcat gctccccctt agaatgcaga accttcttct tgactcaagg ggccttgcta 420
aatgacaaac attccaatgg aaccattaaa gataggagcc catatcgaac cctgatgagc 480
tgtcctattg gtgaagttcc ctctccatac aactcaagat ttgagtcggt cgcttggtca 540
gcaagtgctt gtcacgatgg catcaattgg ctaacaatcg gaatttctgg cccagacagt 600
ggggcagtgg ctgtattaaa gtacaatggc ataataacag acactatcaa gagttggaaa 660
aacaatatat tgagaacaca agagtctgaa tgtgcatgtg taaatggttc ttgctttacc 720
ataatgaccg atggaccaag tgatggacag gcctcataca agatcttcag aatagaaaag 780
ggaaagatag tcaaatcagt cgaaatgaat gcccctaatt atcactatga ggaatgctcc 840
tgttatcctg attctagtga aatcacatgt gtgtgcaggg ataactggca tggctcgaat 900
cgaccgtggg tgtctttcaa ccagaatctg gaatatcaga taggatacat atgcagtggg 960
attttcggag acaatccacg ccctaatgat aagacaggca gttgtggtcc agtatcgtct 1020
aatggagcaa atggagtaaa aggattttca ttcaaatatg gcaatggtgt ttggataggg 1080
agaactaaaa gcattagttc aagaaaaggt tttgagatga tttgggatcc aaatggatgg 1140
actgggacag acaaaaactt ctcaataaag caagatatca taggaataaa tgagtggtca 1200
ggatacagcg ggagttttgt tcagcatcca gaactaacag ggctgaattg tataagacct 1260
tgcttctggg ttgaactaat cagagggcga cccaaagaga acacaatctg gactagcggg 1320
agcagcatat ccttttgtgg tgtaaacagt gacactgtgg gttggtcttg gccagacggt 1380
gctgagttgc catttaccat tgacaagtaa 1410
<210> 20
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUN1-4 Оптимизированная последовательность свиньи
<400> 20
atgaacccta accagaagat tattaccatc ggatccgtgt gcatgaccat cggaatggcc 60
aacctgatcc tgcaaatcgg aaacatcatc tccatctgga ttagccatag cattcagctt 120
ggaaaccaga gccagatcga gacttgtaac cagtccgtga tcacctacga gaacaacacc 180
tgggtcaacc agacttacgt caatatcagc aacaccaact tcgctgcggg gcagtccgtg 240
gtgtccgcaa agctggccgg caacagctcc ctgtgccccg tgtccggatg ggcgatctac 300
tccaaggata acagcgtgcg catcgggtcc aaaggggacg tgttcgtcat ccgcgaacca 360
ttcatttctt gctccccctt ggaatgtcgg accttcttcc tgacccaagg ggcgttgctc 420
aacgacaagc acagcaacgg aaccatcaaa gatcggtcgc cctaccgcac tctgatgtcg 480
tgccctatcg gcgaagtgcc atccccctac aactcacgct tcgagtccgt ggcctggtcc 540
gcttccgcct gccacgatgg aatcaactgg ctcacaatcg gcatctccgg cccggactcg 600
ggagccgtgg ccgtgctgaa gtacaatggt attattactg acactatcaa gtcgtggaag 660
aacaatattc tccggactca agaatctgaa tgcgcctgcg tgaacggttc ctgcttcact 720
atcatgaccg acggcccttc cgatggacag gcctcataca agatcttccg gatcgagaag 780
ggaaagatcg tgaagtccgt cgagatgaac gcaccgaact accattatga ggaatgctcg 840
tgctacccgg actcctcgga aattacttgc gtgtgccgcg acaattggca cgggtccaac 900
aggccctggg tgtccttcaa ccaaaacctg gagtaccaga tcggttacat ctgctccggg 960
atttttggag acaaccctag acctaacgac aagaccggct catgcggacc tgtgtcctcc 1020
aacggagcca acggcgtgaa gggattctcg ttcaaatatg ggaacggcgt ctggataggt 1080
cggaccaagt ccatctcgtc acggaagggc tttgaaatga tttgggaccc gaacggttgg 1140
accggaaccg acaagaactt cagcatcaag caggacatta tcggcattaa cgagtggagc 1200
ggatactcgg gcagcttcgt ccagcacccg gaacttacgg gcctcaattg tattaggccc 1260
tgtttttggg tcgagctgat tagagggcgc cccaaggaaa acaccatctg gaccagcggc 1320
tccagcatct cattctgcgg agtgaactcc gacaccgtgg gctggtcgtg gcccgacggt 1380
gccgagctgc cgttcaccat cgataaatga 1410
<210> 21
<211> 469
<212> Белок
<213> Вирус гриппа A
<220>
<223> >ALX30323.1 нейраминидаза [Вирус гриппа A
(A/swine/Italy/179057/2015(H1N1))] EUN1-4
<400> 21
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Cys Met Thr
1 5 10 15
Ile Gly Met Ala Asn Leu Ile Leu Gln Ile Gly Asn Ile Ile Ser Ile
20 25 30
Trp Ile Ser His Ser Ile Gln Leu Gly Asn Gln Ser Gln Ile Glu Thr
35 40 45
Cys Asn Gln Ser Val Ile Thr Tyr Glu Asn Asn Thr Trp Val Asn Gln
50 55 60
Thr Tyr Val Asn Ile Ser Asn Thr Asn Phe Ala Ala Gly Gln Ser Val
65 70 75 80
Val Ser Ala Lys Leu Ala Gly Asn Ser Ser Leu Cys Pro Val Ser Gly
85 90 95
Trp Ala Ile Tyr Ser Lys Asp Asn Ser Val Arg Ile Gly Ser Lys Gly
100 105 110
Asp Val Phe Val Ile Arg Glu Pro Phe Ile Ser Cys Ser Pro Leu Glu
115 120 125
Cys Arg Thr Phe Phe Leu Thr Gln Gly Ala Leu Leu Asn Asp Lys His
130 135 140
Ser Asn Gly Thr Ile Lys Asp Arg Ser Pro Tyr Arg Thr Leu Met Ser
145 150 155 160
Cys Pro Ile Gly Glu Val Pro Ser Pro Tyr Asn Ser Arg Phe Glu Ser
165 170 175
Val Ala Trp Ser Ala Ser Ala Cys His Asp Gly Ile Asn Trp Leu Thr
180 185 190
Ile Gly Ile Ser Gly Pro Asp Ser Gly Ala Val Ala Val Leu Lys Tyr
195 200 205
Asn Gly Ile Ile Thr Asp Thr Ile Lys Ser Trp Lys Asn Asn Ile Leu
210 215 220
Arg Thr Gln Glu Ser Glu Cys Ala Cys Val Asn Gly Ser Cys Phe Thr
225 230 235 240
Ile Met Thr Asp Gly Pro Ser Asp Gly Gln Ala Ser Tyr Lys Ile Phe
245 250 255
Arg Ile Glu Lys Gly Lys Ile Val Lys Ser Val Glu Met Asn Ala Pro
260 265 270
Asn Tyr His Tyr Glu Glu Cys Ser Cys Tyr Pro Asp Ser Ser Glu Ile
275 280 285
Thr Cys Val Cys Arg Asp Asn Trp His Gly Ser Asn Arg Pro Trp Val
290 295 300
Ser Phe Asn Gln Asn Leu Glu Tyr Gln Ile Gly Tyr Ile Cys Ser Gly
305 310 315 320
Ile Phe Gly Asp Asn Pro Arg Pro Asn Asp Lys Thr Gly Ser Cys Gly
325 330 335
Pro Val Ser Ser Asn Gly Ala Asn Gly Val Lys Gly Phe Ser Phe Lys
340 345 350
Tyr Gly Asn Gly Val Trp Ile Gly Arg Thr Lys Ser Ile Ser Ser Arg
355 360 365
Lys Gly Phe Glu Met Ile Trp Asp Pro Asn Gly Trp Thr Gly Thr Asp
370 375 380
Lys Asn Phe Ser Ile Lys Gln Asp Ile Ile Gly Ile Asn Glu Trp Ser
385 390 395 400
Gly Tyr Ser Gly Ser Phe Val Gln His Pro Glu Leu Thr Gly Leu Asn
405 410 415
Cys Ile Arg Pro Cys Phe Trp Val Glu Leu Ile Arg Gly Arg Pro Lys
420 425 430
Glu Asn Thr Ile Trp Thr Ser Gly Ser Ser Ile Ser Phe Cys Gly Val
435 440 445
Asn Ser Asp Thr Val Gly Trp Ser Trp Pro Asp Gly Ala Glu Leu Pro
450 455 460
Phe Thr Ile Asp Lys
465
<210> 22
<211> 1410
<212> ДНК
<213> Вирус гриппа A
<220>
<223> >KR700532.1 Вирус гриппа A (A/swine/Italy/28762-3/2013(H1N1))
ген нейраминидазы (NА) сегмента 6, полная кодирующая
последовательность
<400> 22
atgaacccaa atcagaagat aataatcatt agttcaatct gtatgacaaa tggaattgct 60
agcttgatat tacaaattgg gaacataata tcaatatgga ttagccattc aattcaaatt 120
gagaacccaa accagaccga accatgcaat caaagcgtta ttatttacga aaacaacaca 180
tgggtaaatc aaacgtatgt taacatcagc aacaataatt ttgttgttga acagacagtg 240
gtttcaatga aattagcggg cagttcttct ctctgccctg ttagtggatg ggctatatac 300
agtaaagata acagtgtaag aatcggttcc aaaggggatg tgtttgtcat aagagagcca 360
ttcatctcat gctcccattt ggaatgtaga accttcttct taactcaagg ggccctactg 420
aatgataaac attctaatgg aaccgttaaa gacagaagcc cctatcgaac cctgatgagc 480
tgtcctattg gtgaagtccc ctctccatac aactcaaaat ttgagtcagt tgcttggtca 540
gcaagtgctt gccatgatgg caccagttgg ttgacaattg ggatttctgg tccagacaat 600
ggagcagtgg ctgtgttgaa atacaatgac ataataacag acactatcaa gagttggaaa 660
aacaacatat tgagaacaca agaatctgaa tgtgcatgtt tgaatggttc ttgctttact 720
gtaatgaccg atggaccaag taatgggcag gcctcataca agatcttcaa aatagaaaag 780
gggaaagtag tcaaatcagt cgagttgaat gctcctaatt atcactatga ggaatgttcc 840
tgttatcctg attctggtga aatcatatgt gtatgcaggg acaattggca tggctcgaat 900
cgaccatggg tgtctttcaa tcagaatctg gagtatcaga taggatacat atgcagtggg 960
gttctcggag acaatccgcg ccctaatgat agaacaggca gttgtggtcc agtatcatct 1020
catggagcaa atggggtaaa agggttttcg tttaaatacg gcaatggaat ttggataggg 1080
agaactaaaa gcactattac aaggagtggt tttgagatga tttgggaccc aaacggatgg 1140
actggaacag acaataattt ctcagtgaag caagatatcg taggaataac taactggtca 1200
ggatacagcg ggagttttgt ccaacatcca gaattaaccg gattggattg tattagacct 1260
tgcttctggg ttgaactaat cagagggaga cccaaagaga acacaatctg gactagcgga 1320
agcagcatat ccttttgtgg tgtaaatagt gacactgtgg gttggtcttg gccagacggt 1380
gctgagttgc catttaccat tgacaagtaa 1410
<210> 23
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >EUN1-2 Оптимизированная последовательность свиньи
<400> 23
atgaatccga atcagaagat cattatcatc tcctccattt gtatgactaa cggaatcgca 60
tctctgatct tgcaaattgg gaacatcatt agcatctgga tctcccactc gatccaaatc 120
gagaacccga accagaccga gccctgcaac cagagcgtga tcatatacga aaacaatacc 180
tgggtgaacc agacctatgt caatatctcg aacaacaact tcgtggtgga gcagactgtc 240
gtgtccatga agctcgccgg gtccagctcg ttgtgccctg tctccggctg ggctatctac 300
agcaaagaca acagcgtcag aatcggcagc aagggagatg tgttcgtgat ccgcgagcct 360
ttcatctcct gctcacacct ggaatgccgg acctttttcc tgacacaagg cgccctgctc 420
aatgacaagc actcgaacgg gaccgtgaag gacagaagcc cctacagaac cctgatgtcc 480
tgtccgattg gagaagtgcc ctccccgtat aactcaaagt tcgaatccgt ggcttggtca 540
gcatccgcgt gccatgacgg gaccagctgg ctgactatcg gaatctccgg tcctgacaac 600
ggcgcggtcg ccgtgctcaa gtacaacgac attatcactg acaccatcaa gtcctggaaa 660
aacaacatcc tccggaccca agagtccgag tgcgcctgcc tgaacggttc gtgtttcacc 720
gtgatgaccg atggaccaag caacggacaa gcctcgtaca agattttcaa gatcgaaaag 780
ggaaaagtgg tcaagtccgt ggagctgaac gcccctaact accactacga agagtgttca 840
tgctaccctg actccggcga aattatctgc gtgtgtcggg ataactggca cggatccaac 900
cgcccctggg tcagcttcaa ccagaacctg gaatatcaga tcggctacat ttgctccggc 960
gtgctggggg acaacccgag gccgaatgac cgcactggtt cctgcggacc cgtgtcatcc 1020
cacggggcga acggagtgaa gggtttctct ttcaaatacg gaaacggcat ctggattggc 1080
agaaccaaga gcactatcac ccggagcgga tttgagatga tctgggaccc caacggatgg 1140
accggtaccg ataacaactt ttcggtcaag caggatattg tgggcatcac taactggtcg 1200
ggatactccg gctcgttcgt gcagcatccc gagctgactg gcctggactg cattcgcccg 1260
tgcttctggg tggaacttat caggggtcgg cctaaggaga acactatctg gacctccgga 1320
tcgtcgattt ccttctgcgg cgtgaactca gataccgtgg gatggtcctg gccggacggg 1380
gccgagcttc cattcacgat tgataagtga 1410
<210> 24
<211> 469
<212> Белок
<213> Вирус гриппа A
<220>
<223> >AKJ81669.1 нейраминидаза [Вирус гриппа A
(A/swine/Italy/28762-3/2013(H1N1))] EUN1-2
<400> 24
Met Asn Pro Asn Gln Lys Ile Ile Ile Ile Ser Ser Ile Cys Met Thr
1 5 10 15
Asn Gly Ile Ala Ser Leu Ile Leu Gln Ile Gly Asn Ile Ile Ser Ile
20 25 30
Trp Ile Ser His Ser Ile Gln Ile Glu Asn Pro Asn Gln Thr Glu Pro
35 40 45
Cys Asn Gln Ser Val Ile Ile Tyr Glu Asn Asn Thr Trp Val Asn Gln
50 55 60
Thr Tyr Val Asn Ile Ser Asn Asn Asn Phe Val Val Glu Gln Thr Val
65 70 75 80
Val Ser Met Lys Leu Ala Gly Ser Ser Ser Leu Cys Pro Val Ser Gly
85 90 95
Trp Ala Ile Tyr Ser Lys Asp Asn Ser Val Arg Ile Gly Ser Lys Gly
100 105 110
Asp Val Phe Val Ile Arg Glu Pro Phe Ile Ser Cys Ser His Leu Glu
115 120 125
Cys Arg Thr Phe Phe Leu Thr Gln Gly Ala Leu Leu Asn Asp Lys His
130 135 140
Ser Asn Gly Thr Val Lys Asp Arg Ser Pro Tyr Arg Thr Leu Met Ser
145 150 155 160
Cys Pro Ile Gly Glu Val Pro Ser Pro Tyr Asn Ser Lys Phe Glu Ser
165 170 175
Val Ala Trp Ser Ala Ser Ala Cys His Asp Gly Thr Ser Trp Leu Thr
180 185 190
Ile Gly Ile Ser Gly Pro Asp Asn Gly Ala Val Ala Val Leu Lys Tyr
195 200 205
Asn Asp Ile Ile Thr Asp Thr Ile Lys Ser Trp Lys Asn Asn Ile Leu
210 215 220
Arg Thr Gln Glu Ser Glu Cys Ala Cys Leu Asn Gly Ser Cys Phe Thr
225 230 235 240
Val Met Thr Asp Gly Pro Ser Asn Gly Gln Ala Ser Tyr Lys Ile Phe
245 250 255
Lys Ile Glu Lys Gly Lys Val Val Lys Ser Val Glu Leu Asn Ala Pro
260 265 270
Asn Tyr His Tyr Glu Glu Cys Ser Cys Tyr Pro Asp Ser Gly Glu Ile
275 280 285
Ile Cys Val Cys Arg Asp Asn Trp His Gly Ser Asn Arg Pro Trp Val
290 295 300
Ser Phe Asn Gln Asn Leu Glu Tyr Gln Ile Gly Tyr Ile Cys Ser Gly
305 310 315 320
Val Leu Gly Asp Asn Pro Arg Pro Asn Asp Arg Thr Gly Ser Cys Gly
325 330 335
Pro Val Ser Ser His Gly Ala Asn Gly Val Lys Gly Phe Ser Phe Lys
340 345 350
Tyr Gly Asn Gly Ile Trp Ile Gly Arg Thr Lys Ser Thr Ile Thr Arg
355 360 365
Ser Gly Phe Glu Met Ile Trp Asp Pro Asn Gly Trp Thr Gly Thr Asp
370 375 380
Asn Asn Phe Ser Val Lys Gln Asp Ile Val Gly Ile Thr Asn Trp Ser
385 390 395 400
Gly Tyr Ser Gly Ser Phe Val Gln His Pro Glu Leu Thr Gly Leu Asp
405 410 415
Cys Ile Arg Pro Cys Phe Trp Val Glu Leu Ile Arg Gly Arg Pro Lys
420 425 430
Glu Asn Thr Ile Trp Thr Ser Gly Ser Ser Ile Ser Phe Cys Gly Val
435 440 445
Asn Ser Asp Thr Val Gly Trp Ser Trp Pro Asp Gly Ala Glu Leu Pro
450 455 460
Phe Thr Ile Asp Lys
465
<210> 25
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5` фланкирующая последовательность открытой рамки считывания
<400> 25
ggcgcgccgc acc 13
<---
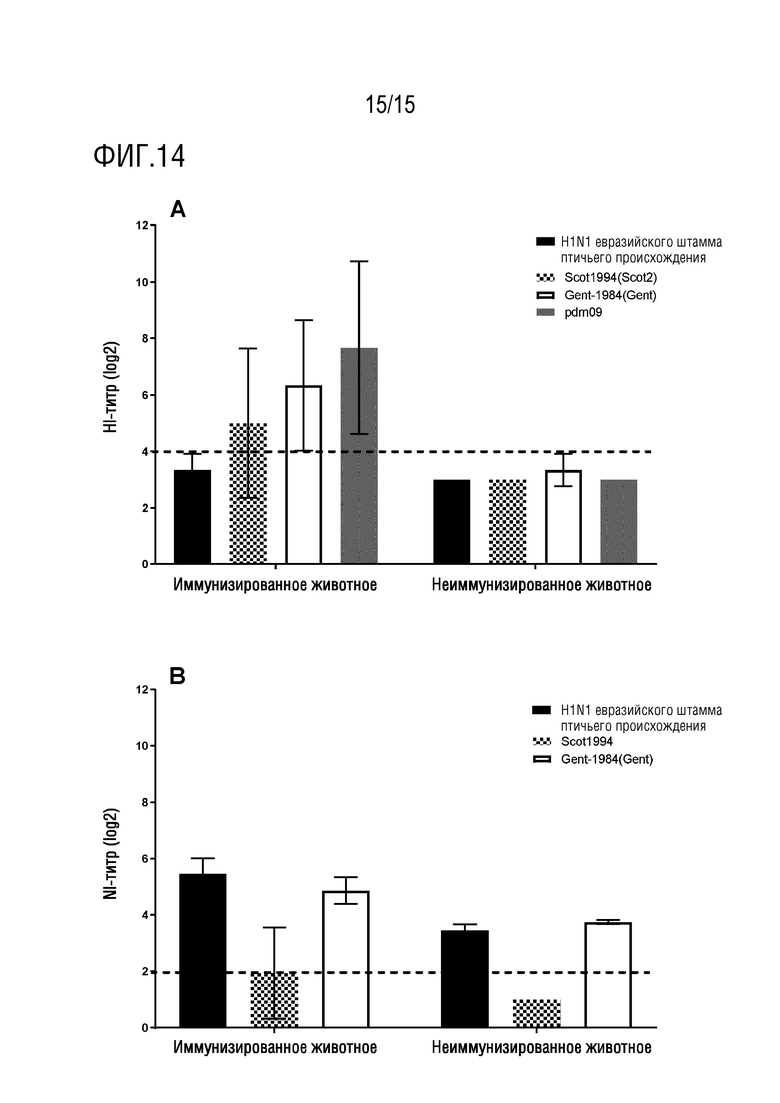
Настоящее изобретение относится к биотехнологии, вирусологии и медицине. Предложена конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа A (IAV-S). Конструкция нуклеиновой кислоты содержит в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты: первую последовательность нуклеиновой кислоты, кодирующую первый антиген гемагглютинина (НА) IAV-S из штамма A/swine/England/9953/2012(H1N2); A/swine/Italy/186822/2011(H1N2); A/swine/Italy/3033-1/2015(H1N2); A/swine/France/22-130212/2013(H1N2); A/swine/France/35-140041/2014(H1N2) или A/swine/Denmark/10802-1/2012(H1N2), и вторую последовательность нуклеиновой кислоты, кодирующую второй антиген НА IAV-S из штамма A/swine/Denmark/10-1048-2/2011(H1N1); A/swine/Italy/28762-3/2013(H1N1); A/swine/France/29-120326/2012(H1N1); A/swine/France/44-120070/2012(H1N1); или A/swine/France/56-140048-2014(H1N1). Предложена также конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа A (IAV-S), которая содержит в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты: первую последовательность нуклеиновой кислоты, кодирующую первый антиген НА IAV-S из штамма A/swine/Netherlands/Ysselsteyn-CVI8864A/2012(H3N2), A/swine/Belgium/Glabbeek-284/2012(H3N2), A/swine/Spain/33936/2012(H3N2) или A/swine/Italy/240849/2015(H3N2), и вторую последовательность нуклеиновой кислоты, кодирующую второй антиген НА IAV-S линии A(H1N1)pdm09 (pdm09). Предложена частица РНК-репликона альфавируса, используемая в профилактике заболевания, вызванного вирусом свиного гриппа A (IAV-S) у субъекта, содержащая указанную конструкцию нуклеиновой кислоты; иммуногенная композиция против вируса свиного гриппа A (IAV-S), содержащая указанную частицу РНК-репликона альфавируса и фармацевтически приемлемый адъювант; вакцина против вируса свиного гриппа A (IAV-S), содержащая указанную иммуногенную композицию и способ иммунизации свиньи против вируса свиного гриппа А. Изобретение позволяет расширить ассортимент вакцин против инфекций, вызванных вирусом гриппа А. 7 н. и 12 з.п. ф-лы, 14 ил., 2 табл., 7 пр.
1. Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа A (IAV-S), где конструкция нуклеиновой кислоты содержит в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген гемагглютинина (НА) IAV-S из штамма A/swine/England/9953/2012(H1N2); A/swine/Italy/186822/2011(H1N2); A/swine/Italy/3033-1/2015(H1N2); A/swine/France/22-130212/2013(H1N2); A/swine/France/35-140041/2014(H1N2) или A/swine/Denmark/10802-1/2012(H1N2), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген НА IAV-S из штамма A/swine/Denmark/10-1048-2/2011(H1N1); A/swine/Italy/28762-3/2013(H1N1); A/swine/France/29-120326/2012(H1N1); A/swine/France/44-120070/2012(H1N1); или A/swine/France/56-140048-2014(H1N1).
2. Конструкция нуклеиновой кислоты для применения по п. 1, в которой первый антиген НА происходит от штамма A/swine/Italy/3033-1/2015 (H1N2).
3. Конструкция нуклеиновой кислоты для применения по п. 1 или 2, где первый антиген НА, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, идентичную ей на по меньшей мере 85%.
4. Конструкция нуклеиновой кислоты для применения по любому из пп. 1-3, где второй антиген НА происходит от штамма A/swine/Italy/28762-3/2013 (H1N1).
5. Конструкция нуклеиновой кислоты для применения по любому из пп. 1-4, где второй антиген НА, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
6. Конструкция нуклеиновой кислоты для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа A (IAV-S), где конструкция нуклеиновой кислоты содержит в порядке от 5' к 3'-концу последовательности нуклеиновой кислоты:
первую последовательность нуклеиновой кислоты, кодирующую первый антиген НА IAV-S из штамма A/swine/Netherlands/Ysselsteyn-CVI8864A/2012(H3N2), A/swine/Belgium/Glabbeek-284/2012(H3N2), A/swine/Spain/33936/2012(H3N2) или A/swine/Italy/240849/2015(H3N2), и
вторую последовательность нуклеиновой кислоты, кодирующую второй антиген НА IAV-S линии A(H1N1)pdm09 (pdm09).
7. Конструкция нуклеиновой кислоты для применения по п. 6, где первый антиген НА происходит от штамма A/swine/Italy/240849/2015 (H3N2).
8. Конструкция нуклеиновой кислоты для применения по п. 6 или 7, где первый антиген НА, кодируемый первой последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 9 или аминокислотную последовательность, идентичную ей на по меньшей мере 90%.
9. Конструкция нуклеиновой кислоты для применения по любому из пп. 6-8, где второй антиген НА происходит от штамма A/swine/England/373/2010 (H1N1).
10. Конструкция нуклеиновой кислоты для применения по любому из пп. 6-9, где второй антиген НА, кодируемый второй последовательностью нуклеиновой кислоты, содержит аминокислотную последовательность SEQ ID NO: 12 или аминокислотную последовательность, идентичную ей на по меньшей мере 95%.
11. Частица РНК-репликона альфавируса, используемая в профилактике заболевания, вызванного вирусом свиного гриппа A (IAV-S) у субъекта, содержащая конструкцию нуклеиновой кислоты по любому из пп. 1-5.
12. Частица РНК-репликона альфавируса, используемая в профилактике заболевания, вызванного вирусом свиного гриппа A (IAV-S) у субъекта, содержащая конструкцию нуклеиновой кислоты по любому из пп. 6-10.
13. Частица РНК-репликона альфавируса по п. 11 или 12, представляющая собой частицу РНК-репликона альфавируса венесуэльского энцефалита лошадей (VEE).
14. Иммуногенная композиция против вируса свиного гриппа A (IAV-S), содержащая частицу РНК-репликона альфавируса по любому из пп. 11-13 или обе, и фармацевтически приемлемый адъювант.
15. Иммуногенная композиция по п. 14, адаптированная для одновременного введения частиц РНК-репликона альфавируса по пп. 11 и 12.
16. Вакцина против вируса свиного гриппа A (IAV-S), содержащая иммуногенную композицию по п. 14 или 15.
17. Вакцина по п. 16, которая содержит адъювант, выбранный из группы, состоящей из биоразлагаемого масла, эмульсии масло-в-воде с содержанием минерального масла 2,5-50% (об./об.) и биоразлагаемого масла, смешанного с эмульсией масло-в-воде с содержанием минерального масла 2,5-50% (об./об.).
18. Вакцина по п. 16 или 17 для применения для профилактики у субъекта заболевания, вызванного вирусом свиного гриппа А.
19. Способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества вакцины по п. 16 или 17.
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| RYAN L VANDER VEEN et al., Safety, immunogenicity, and efficacy of an alphavirus replicon-based swine influenza virus hemagglutinin vaccine, VACCINE, ELSEVIER, AMSTERDAM, NL, v | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Приспособление к банкаброшу для предупреждения неровностей в пряже | 1925 |
|
SU1944A1 |
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| ПОЛИВАЛЕНТНАЯ ВАКЦИНА ПРОТИВ ГРИППА | 2018 |
|
RU2706191C1 |
| АНТИГЕНЫ ВИРУСА ГРИППА H1N1 С ШИРОКИМ СПЕКТРОМ АКТИВНОСТИ, ОПТИМИЗИРОВАННЫЕ С ПРИМЕНЕНИЕМ ВЫЧИСЛИТЕЛЬНЫХ СРЕДСТВ | 2012 |
|
RU2612900C2 |
