ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По этой заявке испрашивается приоритет в соответствии со статьей Свода законов США § 119 (c) временной заявки США, серийный № 62/607101, поданной 18 декабря 2017 года.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к конструкциям векторов и/или нуклеиновых кислот, которые кодируют один или более антигенов нейраминидазы (NA) вируса гриппа A. Настоящее изобретение дополнительно относится к вакцинам против вируса гриппа А, содержащим эти конструкции векторов и/или нуклеиновых кислот. Настоящее изобретение также относится к способам получения и применения вакцин по отдельности или в комбинации с другими защитными средствами.
УРОВЕНЬ ТЕХНИКИ
Вирусы гриппа А (IAV) создают значительную нагрузку на здоровье людей и животных во всем мире. Член семейства Orthomyyxoviridae, IAV подразделяется на различные подтипы на основе гликопротеинов вирусной поверхности, гемагглютинина (HA) и нейраминидазы (NA). IAV заражает домашнюю птицу, свиней, лошадей, кошек, собак, морских млекопитающих (например, китов), летучих мышей и людей. Дикие водоплавающие птицы и прибрежные птицы (утки, гуси, лебеди и чайки) являются естественными резервуарами, и они могут быть заражены шестнадцатью различными подтипами HA и девятью различными подтипами NA [Webster et al., Microbiol Rev 56: 152-179 (1992)]. Недавно у летучих мышей были идентифицированы два новых подтипа, H17N10 и H18N11 [Tong et al., Proc. Natl. Акад. Sci. 109 (11): 4269-4274 (2012) и Mehle, Viruses 6 (9): 3438-3449 (2014)]. Межвидовая передача вируса чаще всего происходит между дикой и домашней водоплавающей птицей и между свиньями и человеком [Nelson and Vincent, 23 (3) Trends in Microbiol. 142-153 (2015)]. Млекопитающие, инфицированные IAV, демонстрируют острое респираторное заболевание, приводящее к воздушно-капельной передаче вируса. У водоплавающей и домашней птицы вирус может размножаться как в дыхательном, так и в кишечном тракте и распространяться через фекально-оральный путь.
Один из таких IAV, вирус свиного гриппа А (IAV-S), представляет собой серьезный респираторный патоген домашних свиней, который оказался экономически дорогостоящим, особенно для животноводства, во всем мире [Holtkamp et al., The American Association of Swine Veterinarians Annual Meeting. (2007)]. Кроме того, передача вирусов гриппа свиного происхождения людям хорошо задокументирована и представляет собой серьезную угрозу для здравоохранения, что, таким образом, обеспечивает еще больший стимул для борьбы с вирусом гриппа А в стаде свиней [Krueger and Grey, Curr Top Microbiol Immunol 370: 201-225 (2013)].
В ответ на эту проблему многие свиноводы в настоящее время вакцинируют своих свиней против IAV-S, используя коммерчески доступные вакцины. Однако контролировать IAV-S с помощью общепринятных вакцин сложно, поскольку многие разнообразные штаммы IAV-S совместно циркулируют в полевых условиях и продолжают развиваться [Gao et al., J Gen Virol 98 (8): 2001-2010 (2017)]. Разнообразие и изменчивость IAV-S обусловлены генетической структурой вируса. Как и другие вирусы гриппа А, IAV-S имеет гены, закодированные в восьми сегментах РНК, и механизм геномной репликации, который приводит к частым мутациям. Эти генетические характеристики позволяют IAV-S быстро адаптироваться, в том числе избегать существующих нейтрализующих антител, вызванных воздействием предыдущих штаммов. Таким образом, инактивированные вакцины против вируса IAV-S, которые являются коммерчески доступными на рынке США, несмотря на то, что в их состав входит до пяти различных штаммов IAV-S, оказались неполноценными из-за новых штаммов, возникающих как следствие непрерывного антигенного дрейфа.
Классификация вирусов гриппа А начинается с подтипов HA и NA, двух основных гликопротеинов на поверхности вируса. Белок HA опосредует прикрепление и слияние вируса с клетками-хозяевами. Антитела, которые связываются с HA, блокируют прикрепление вируса, вирусную репликацию, а также снижают или даже предотвращают заболевание. Нейраминидаза представляет собой фермент, который функционирует на заключительной стадии цикла репликации вируса гриппа, отщепляя недавно сформированные вирусные частицы от клетки-хозяина, тем самым позволяя новому потомству вируса распространяться и заражать другие клетки. Антитела, которые связываются с NA, ограничивают ее ферментативную активность и тем самым могут снижать уровень распространения вируса и тяжесть заболеваемости у определенных моделей на животных.
В то время как человеческий грипп A, как правило, имеет один или два доминирующих штамма, циркулирующих повсеместно в течение сезона указанного гриппа, многие другие штаммы IAV-S циркулируют одновременно, причем эти штаммы различаются в разных географических регионах. Штаммы IAV-S имеют антигенную вариабельность, но в основном содержат подтип H1 или H3 HA и подтип N1 или N2 NA. В пределах каждого подтипа HA и NA IAV-S существует дополнительное филогенетическое разнообразие. В популяции свиней в США имеются четыре преобладающих филогенетических кластера H1 (гамма, дельта 1, дельта 2, пандемический), два преобладающих кластера H3 (кластер IV и человекоподобный), два преобладающих кластера N1 (классический, пандемический), и два преобладающих кластера N2 (N2-1998 и N2-2002). [См., Anderson et al., Flu and other Respiratory Viruses 7 (Suppl. 4); 42-51 (2013); и Anderson et al., mSphere 1(6) e00275-16:1-14 (2016)]. В Европе есть три основных линии H1 (евразийский, подобный птичьему H1, Scotland/410440/1994-подобный H1 и пандемический 2009 подобный H1), одна основная линия H3 (Gent/1/1984-подобный H3), две основные линии N1 (евразийский, подобный птичьему N1, пандемический 2009 подобный N1), две основные линии N2 (Gent/1/1984-подобный N2, Scotland/410440/1994-подобный N2) и две второстепенные линии N2 (Italy/4675/2003 подобный N2, подобный человеческому сезонному N2) [Watson et al., J. Virol., 89: 9920-9931 (2015); DOI: 10,1128/JVI.00840-15].
Ряд стратегий на основе векторов использовался в течение многих лет для вакцин с целью защиты от определенных патогенов. Одна из таких векторных стратегий включает использование репликонов частиц РНК, полученных из альфа-вируса (RP) [Vander Veen, et al. Anim Health Res Rev. 13 (1): 1-9. (2012) doi: 10, 1017/S1466252312000011; Kamrud et al., J Gen Virol., 91 (Pt 7): 1723-1727 (2010)], которые были разработаны из нескольких различных альфа-вирусов, включая вирус венесуэльского энцефалита лошадей (VEE) [Pushko et al., Virology 239: 389 -401 (1997)], Sindbis (SIN) [Bredenbeek et al., Journal of Virology 67: 6439-6446 (1993)], и вирус леса Семлики (SFV) [Liljestrom and Garoff, Biotechnology (NY) 9: 1356-1361 (1991)]. Вакцины RP доставляют репликоны альфавирусной РНК, дефектные по размножению, в клетки-хозяева, что в результате приводит к экспрессии желаемого антигенного трансгена/трансгенов in vivo [Pushko et al., Virology 239 (2): 389-401 (1997)]. RP имеют привлекательные профили безопасности и эффективности по сравнению с некоторыми традиционными составами вакцин [Vander Veen, et al. Anim Health Res Rev. 13 (1): 1-9. (2012)]. Платформа RP была использована для кодирования патогенных антигенов и является основой для нескольких вакцин для свиней и птицы, лицензированных Министерством сельского хозяйства США.
Как указано выше, коммерческие вакцины против IAV-S, доступные для использования у домашних свиней, часто не защищают стада, поскольку антигены не соответствуют всем современным штаммам, циркулирующим в полевых условиях [Lee et al., Can J Vet Res 71 (3): 207-12 (2007); Vincent et al., Vaccine 28 (15): 2782-2787 (2010)]. Обычная платформа инактивированных вирусных антигенов ограничена регулирующими и производственными факторами, которые ограничивают способность к быстрому обновлению вакцинных штаммов в ответ на продолжающийся антигенный дрейф вируса. При использовании обычных инактивированных вакцин против вируса IAV-S выбор вирусных штаммов основан на свойствах антигенов HA. Серологическая оценка иммунных ответов аналогичным образом сфокусирована на измерении антител к этому HA. Тот же акцент на антиген HA также верен и для коммерческих инактивированных вакцинах против вируса гриппа, выпускаемых для людей, а также других видов животных, восприимчивых к гриппу (например, птицы, лошади, собаки). Вакцины IAV-S, которые индуцируют титры антител, ингибирующих HA (HI), защищают свиней от экспериментальной инфекции антигенно схожим штаммом [Kyriakis et al., Vet Microbiol 144 (1-2): 67-74 (2010)]. Однако относительно быстрый генетический дрейф генов HA позволяет появиться новым штаммам, которые функционально не ингибируются индуцируемыми вакциной антителами к HA.
Клинические исследования показали статистически значимую корреляцию между титрами антител, специфичных к NA, и сниженной заболеваемостью гриппом у людей [Memoli et al., MBio 7 (2): e00417-16. (2016); Monto et al., J Infect Dis 212 (8): 1191-1199 (2015); Murphy et al., N Engl. J Med 286 (25): 1329-32 (1972)]. Различные исследования, проведенные на инбредных мышах, показали, что NA-специфический иммунитет может защищать от контрольного заражения [Brett and Johansson, Virology 339 (2): 273-80 (2005); Johansson and Kilbourne, Proc Natl Acad Sci USA 91 (6): 2358-2361 (1994); Kilbourne et al., J Infect Dis 189 (3): 459-461 (2004)]. Hessel et al. [PLoS One 5 (8), e12217 (2010)] показали, что вакцина, содержащая рекомбинантный поксвирусный вектор, кодирующей NA IAV-S, может частично индуцировать защиту мыши от вирусного заражения, хотя поксвирусный вектор с NA был менее эффективен, чем эквивалентная вакцина, кодирующая соответствующий ген HA. Однако важно отметить, что использование мышей в качестве модели на животных для гриппа нельзя считать надежным показателем эффективности вакцины, поскольку в отличие от свиней, птиц и людей, мыши не являются естественным носителем вирусов гриппа А [Bodewes et al., Expert Rev Vaccines 9 (1): 59-72 (2010); Tripp и Tompkins, Curr Top Microbiol Immunol 333: 397-412 (2009); Vander Veen et al., Vaccine 30 (11): 1944-1950 (2012)]. Фактически, вирусные штаммы должны быть в первую очередь приспособлены к росту на мышах, потому что этот вид не подвержен риску заражения вирусом гриппа А. Таким образом, патогенез и заболеваемость представлены по потере массы и смертности, которые отличаются от параметров инфекции, используемых для природного хозяина.
Аналогичные исследования проводили на курах, естественных видах хозяев для гриппа, с использованием вакцину с вирусным вектором с NA против контрольной инфекции. Иммунитет к NA у кур лишь частично защищал от инфекций и был значительно менее устойчивым, чем иммунитет индуцированный эквивалентной вакциной с вектором с против HA [Nayak et al., J Virol 84 (5): 2408-2420 (2010); Pavlova et al., Vaccine 27 (5): 773-785 (2009); Sylte et al., Vaccine 25 (19): 3763-72 (2007)]. Аналогичные результаты были получены у другого природно восприимчивого хозяина гриппа, хорька, которого вакцинировали растворимой рекомбинантной NA и/или белками HA с последующим заражением вирусом [Bosch et al., J Virol 84 (19): 10366-103674 (2010)]. Взятые вместе, эти данные подразумевают, что иммунитет только к NA может играть дополнительную и/или дополняющую роль к более важному иммунитету к HA. Фактически, похоже, что в отсутствие вакцины против вируса гриппа А гемагглютинин не достаточно эффективен для защиты от инфекции гриппа А или от болезни, вызванной вирусом гриппа А.
В одном опубликованном исследовании была проведена проверка вакцины на основе вектора NA, аттенуированной вирусом псевдобешенства (PrV), против вирусного контрольного заражения у свиней [Klingbeil et al., Virus Res 199: 20-30 (2015)]. Примечательно, что включение PrV-NA в PrV-HA не обеспечило существенного улучшения по сравнению с защитой, индуцированной PrV-HA отдельно. Данные этого исследования все же показали, что вакцина PrV-NA, вводимая отдельно или совместно с вакциной PrV-HA, индуцирует сывороточные антитела к белку NA. Однако, несмотря на выработку антител, свиньи, вакцинированные NA, показали очень скромное снижение величины вирусной репликации (статистически значимо только на одном суточном образце: 4 суток после заражения). Как и ожидалось, свиньи, вакцинированные исключительно вектором PrV-HA, индуцировали ответы антител к HV, но эти ответы коррелировали с более выраженным снижением выделения вируса (статистически значимо с 2-6 суток после заражения). Фактически до сих пор не было опубликованных исследований, которые бы указывали на то, что вакцина к IAV-S, содержащая и/или кодирующая антиген нейраминидазы вируса гриппа A в отсутствие соответствующего антигена гемагглютинина, может адекватно защитить свинью от инфекции IAV-S.
Цитирование любой ссылки в настоящем описании не должно быть истолковано как признание того, что такая ссылка доступна как «известный уровень техники» для настоящей заявки.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, настоящее изобретение относится к конструкциям вектора и/или нуклеиновых кислот, которые кодируют один или более антигенов нейраминидазы (NA) вируса гриппа А. Конструкции этих векторов и/или нуклеиновых кислот можно использовать в иммуногенных композициях, содержащих эти векторы. Иммуногенные композиции по настоящему изобретению можно использовать в вакцинах, которые помогают защитить вакцинированного индивидуума (например, человека, домашнее животное или домашний скот) от вируса гриппа А, например, помогают в профилактике инфекции вирусом гриппа А. В конкретных вариантах осуществления иммуногенные композиции и вакцины по настоящему изобретению содержат один или более NA, которые происходят из вируса гриппа А, и не содержат ни гемагглютинина (HA) вируса гриппа или его антигенного фрагмента, ни нуклеотидную последовательность, кодирующую HA вируса гриппа А или его антигенный фрагмент.
В определенных вариантах осуществления предлагаются конструкции векторов и/или нуклеиновых кислот, которые кодируют один или более антигенов NA вируса человеческого гриппа А. В других вариантах осуществления конструкции векторов и/или нуклеиновых кислот кодируют один или более антигенов NA вируса собачьего гриппа А. В других вариантах осуществления конструкции векторов и/или нуклеиновых кислот кодируют один или более антигенов NA вируса лошадиного гриппа А. В еще одних вариантах осуществления конструкции векторов и/или нуклеиновых кислот кодируют один или более антигенов NA вируса птичьего гриппа А. В других вариантах осуществления конструкции векторов и/или нуклеиновых кислот кодируют один или более антигенов NA вируса бычьего гриппа А. В конкретных вариантах осуществления конструкции векторов и/или нуклеиновых кислот кодируют от двух до четырех антигенов NA вирусов гриппа.
В конкретных вариантах осуществления вектор представляет собой частицу репликона РНК альфавируса, которая кодирует один или более антигенов, которые происходят от вируса гриппа А. В конкретных вариантах осуществления частица репликона РНК альфавируса кодирует одну или более NA вирусов гриппа А. В родственных вариантах осуществления частица репликона РНК альфавируса кодирует один или более антигенных фрагментов одной или нескольких NA вирусов гриппа А. Настоящее изобретение также относится к другим конструкциям векторов и/или нуклеиновых кислот, которые кодируют один или более антигенов, происходящих из вируса свиного гриппа А (IAV-S). В конкретных вариантах осуществления конструкции векторов и/или нуклеиновых кислот также могут кодировать один или более антигенов нейраминидазы (NA) IAV-S. В родственных вариантах осуществления конструкции векторов и/или нуклеиновых кислот могут кодировать один или более антигенных фрагментов из одной или нескольких NA IAV-S.
В одном важном аспекте настоящее изобретение относится к конструкциям векторов и/или нуклеиновых кислот, которые кодируют один или более антигенов нейраминидазы (NA) вирусов гриппа А (IAV-S). Такие конструкции векторов и/или нуклеиновых кислот можно использовать в иммуногенных композициях, содержащих эти векторы. Иммуногенные композиции по настоящему изобретению можно использовать в вакцинах, которые помогают защитить вакцинированного свиного индивидуума (например, свиноматку и/или поросенка) от IAV-S, например, помогают в профилактике заражения вирусом гриппа свиней. В конкретных вариантах осуществления, иммуногенные композиции и вакцины, содержащие одну или более NA, которые происходят из IAV-S, по настоящему изобретению, не содержат ни HA IAV-S, ни его антигенный фрагмент, ни нуклеотидную последовательность, которая кодирует HA IAV-S или его антигенный фрагмент. Настоящее изобретение дополнительно относится к комбинированным вакцинам для вызывания защитного иммунитета против IAV-S и других заболеваний, например, других инфекционных заболеваний свиней. Также изобретение относится к способам получения и применения иммуногенных композиций и вакцин по настоящему изобретению.
В более конкретных вариантах осуществления вектор представляет собой частицу репликона РНК альфавируса, которая кодирует один или более антигенов, происходящих из IAV-S. В конкретных вариантах осуществления частица репликона РНК альфавируса кодирует один или более антигенных фрагментов одной или нескольких NA IAV-S. В родственных вариантах осуществления частица репликона РНК альфавируса кодирует один или более антигенных фрагментов одной или нескольких NA, которые происходят из IAV-S. В конкретных вариантах осуществления частицы репликона РНК альфавируса, кодирующие одну или более NA, происходящих из IAV-S по настоящему изобретению, не содержат ни HA IAV-S, ни его антигенный фрагмент, не кодируют нуклеотидную последовательность, которая кодирует HA IAV-S или его антигенный фрагмент.
В некоторых вариантах осуществления NA IAV-S происходит из филогенетического кластера N1-классический. В других вариантах осуществления частица репликона РНК альфавируса кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N1-классический. В других вариантах осуществления NA IAV-S происходит из филогенетического кластера N2-1998. В других вариантах осуществления частица репликона РНК альфавируса кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N2-1998. В других вариантах осуществления NA IAV-S происходит из филогенетического кластера N1-пандемический. В других вариантах осуществления частица репликона РНК альфавируса кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N1-пандемический. В других вариантах осуществления NA IAV-S происходит из филогенетического кластера N2-2002. В других вариантах осуществления частица репликона РНК альфавируса кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N2-2002.
В аналогичном варианте осуществления, NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - N1-пандемической (EU) линии. В аналогичном варианте осуществления, NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - N1-евразийской линии Avian.
В аналогичном варианте осуществления, NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - линии N2-Gent/1984. В другом аналогичном варианте осуществления, NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - линии N2-Italy/2003. В еще одном аналогичном варианте осуществления NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - линии N2-Scotland/1994. В некоторых вариантах осуществления этого типа NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - линии N2-Scotland/1994 (клада 1). В других вариантах осуществления этого типа NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - линии N2-Scotland/1994 (клада 2). В других вариантах осуществления этого типа NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - A/Swine/Italy/4675/2003, подобного N2. В других вариантах осуществления этого типа NA IAV-S происходит из - и/или частица репликона РНК альфавируса кодирует (антигенный фрагмент) NA из - человеческого гриппа, сезонного типа, подобного N2.
В конкретных вариантах осуществления NA IAV-S, происходящая из филогенетического кластера N1-классический, содержит аминокислотную последовательность, которая имеет 95% идентичности, 97% идентичности, 98% идентичности или более с аминокислотной последовательностью SEQ ID NO: 2. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 2. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 1. В других вариантах осуществления NA IAV-S, происходящая из филогенетического кластера N1-пандемический, содержит аминокислотную последовательность, которая имеет 95% идентичности, 97% идентичности, 98% идентичности или более с аминокислотной последовательностью SEQ ID NO: 4. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO : 4. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 3. В других вариантах осуществления, NA IAV-S, происходящая из филогенетического кластера N2-1998, содержит аминокислотную последовательность, которая имеет 92% идентичности, 94% идентичности, 97% идентичности или более с аминокислотной последовательностью SEQ ID NO: 6. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 6. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 5. В других вариантах осуществления, NA IAV-S, происходящая из филогенетического кластера N2-2002, содержит аминокислотную последовательность, которая имеет 92% идентичности, 94% идентичности, 97% идентичности или более с аминокислотной последовательностью SEQ ID NO: 8. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 8. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 7.
В конкретных вариантах осуществления NA IAV-S, происходящая из N1-пандемической (EU) линии, содержит аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более, с аминокислотной последовательностью SEQ ID NO: 12. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 12. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 11. В родственных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 23. В других вариантах осуществления NA IAV-S, происходящая из N1-евразийской линии Avian, содержит аминокислотную последовательность, которая имеет 85% идентичности, 90% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 14. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 14. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 13. В других вариантах осуществления, NA IAV-S, происходящая от линии N2-Gent/1984 содержит аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 16. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 16. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 15. В родственных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 24. В других вариантах осуществления, NA IAV-S, происходящая из линии N2-Italy/2003, содержит аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 18. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 18. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 17. В родственных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 25. В других вариантах осуществления NA IAV-S, происходящая из линии N2-Scotland/1994 (клада 1), включает в себя аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 20. В более конкретных вариантах осуществления этого типа NA включает аминокислотную последовательность SEQ ID NO: 20. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 19. В родственных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 26. В других вариантах осуществления NA IAV-S, происходящая из линии N2-Scotland/1994 (клада 2), содержит аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 22. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 22. В еще более конкретных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 21. В родственных вариантах осуществления этого типа NA кодируется нуклеотидной последовательностью SEQ ID NO: 27.
В еще более конкретных вариантах осуществления частица репликона РНК альфавируса представляет собой частицу репликона альфавируса венесуэльского конского энцефалита (VEE). В еще более конкретных вариантах осуществления частица репликона РНК альфавируса VEE является частицей репликона РНК альфавируса VEE ТС-83. В других вариантах осуществления частица репликона РНК альфавируса является частицей репликона альфа-вируса Синдбиса (SIN). В других вариантах осуществления частица репликона РНК альфавируса является частицей репликона РНК альфавируса леса Семлики (SFV). В альтернативном варианте осуществления, вектор с «голой» ДНК содержит конструкцию нуклеиновой кислоты, которая кодирует один или более антигенов, происходящих из патогена свиньи. В конкретных вариантах осуществления этого типа вектор с «голой» ДНК содержит конструкцию нуклеиновой кислоты, которая кодирует NA, происходящую из IAV-S, или ее антигенный фрагмент. В других вариантах осуществления вектор с «голой» РНК содержит конструкцию нуклеиновой кислоты, которая кодирует один или более антигенов, происходящих из патогена свиньи. В конкретных вариантах осуществления этого типа вектор с «голой» содержит конструкцию нуклеиновой кислоты, которая кодирует NA, происходящую из IAV-S, или ее антигенный фрагмент.
Настоящее изобретение относится к частицам репликона РНК альфавируса, которые кодируют два или более антигенов IAV-S или их антигенные фрагменты. В конкретных вариантах осуществления частицы репликона РНК альфавируса, кодирующие одну или более NA, которые происходят из IAV-S по настоящему изобретению, не содержат ни HA IAV-S, ни его антигенный фрагмент, ни кодируют нуклеотидную последовательность, которая кодирует HA IAV-S или его антигенный фрагмент.
В конкретных вариантах осуществления частицы репликона РНК альфавируса кодируют от двух до четырех или более антигенов NA, которые происходят из IAV-S, или их антигенные фрагменты. В родственных вариантах осуществления частицы репликона РНК альфавируса кодируют от двух до четырех или более антигенов NA, которые происходят из разных филогенетических кластеров IAV-S, или их антигенные фрагменты. В некоторых вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент. В других вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент. В других вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N1-пандемический, или ее антигенный фрагмент.
В некоторых вариантах осуществления частицы репликона РНК альфавируса кодируют NA, происходящую из линии N1-пандемической (EU), или ее антигенный фрагмент, и NA, происходящую из линии N1-евразийской Avian, или ее антигенный фрагмент. В других вариантах осуществления частицы репликона РНК альфавируса кодируют NA, происходящую из линии N2-Gent/1984, или ее антигенный фрагмент, и NA, происходящую из линии N2-Italy/2003, или ее антигенный фрагмент. В других вариантах осуществления частицы репликона РНК альфавируса кодируют NA, происходящую из линии N2-Gent/1984, или ее антигенный фрагмент, и NA, происходящую из линии N2-Scotland/1994 (клада 1), или ее антигенный фрагмент. В других вариантах осуществления частицы репликона РНК альфавируса кодируют NA, происходящую из линии N2-Gent/1984, или ее антигенный фрагмент, и NA, происходящую из линии N2-Scotland/1994 (клада 2), или ее антигенный фрагмент. В других вариантах осуществления частицы репликона РНК альфавируса кодируют NA, происходящую из линии N2-Italy/2003, или ее антигенный фрагмент, и NA, происходящую из линии N2-Scotland/1994 (клада 1), или ее антигенный фрагмент. В других вариантах осуществления частицы репликона РНК альфавируса кодируют NA, происходящую из линии N2-Italy/2003, или ее антигенный фрагмент, и NA, происходящую из линии N2-Scotland/1994 (клада 2) или ее антигенный фрагмент. В других вариантах осуществления NA, происходящая из одной из европейских линий N1, или ее антигенный фрагмент кодируется частицей репликона РНК альфавируса, наряду с NA, происходящей из одной из европейских линий N2, или ее антигенным фрагментом. Аналогично настоящее изобретение относится к частицам репликона РНК альфавируса, кодирующим три, четыре или более NA, причем каждая NA происходит из разных европейских линий. Кроме того, настоящее изобретение относится к частицам репликона РНК альфавируса, кодирующим две, три, четыре или более NA, причем каждая NA происходит из разных европейских линий и/или филогенетических кластеров.
Таким образом, настоящее изобретение дополнительно относится к частицам репликона РНК альфавируса, которые кодируют три или четыре антигена IAV-S или их фрагменты. В конкретных вариантах осуществления иммуногенные композиции содержат частицы репликона РНК альфавируса, которые кодируют три или четыре антигена NA, происходящей из IAV-S, или их антигенные фрагменты. В родственных вариантах осуществления иммуногенные композиции содержат частицы репликона РНК альфавируса, которые кодируют три или четыре антигена NA, которые происходят из разных филогенетических кластеров IAV-S, или их антигенные фрагменты. В некоторых вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент. В других вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N1-пандемический, или ее антигенный фрагмент. В других вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N1-пандемический, или ее антигенный фрагмент. В других вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N1-пандемический, или ее антигенный фрагмент.
В конкретных вариантах осуществления частицы репликона РНК альфавируса кодируют четыре или более антигенов NA IAV-S или их антигенных фрагментов. В более конкретных вариантах осуществления частицы репликона РНК альфавируса кодируют четыре или более антигенов NA, которые происходят из разных филогенетических кластеров IAV-S, или их антигенных фрагментов. В еще более конкретных вариантах осуществления частица репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N1-пандемический, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент.
В аналогичных вариантах осуществления, как указано выше, частица репликона РНК альфавируса кодирует три, четыре и более антигенов или антигенных фрагментов NA IAV-S, происходящих из N1-пандемической (EU) линии IAV, N1-евразийская линии Avian, линии NA, линии N2-Gent/1984, линии N2-Italy/2003, линии N2-Scotland/1994 (клада 1) и/или линии N2-Scotland/1994 (клада 2).
Как указано выше, все частицы репликона РНК альфавируса по настоящему изобретению могут быть компонентами иммуногенных композиций и/или вакцин. Таким образом, иммуногенная композиция и/или вакцина по настоящему изобретению может содержать одну или более частиц репликона РНК альфавируса по настоящему изобретению. В некоторых вариантах осуществления иммуногенная композиция и/или вакцина содержит только один набор идентичных частиц репликона РНК альфавируса, которые могут кодировать одну или более NA, происходящих из одного или нескольких филогенетических кластеров и/или одной или нескольких линий, как подробно описано выше.
В более конкретном варианте осуществления этого типа иммуногенная композиция и/или вакцина содержит частицы репликона РНК, которые кодируют как NA (или ее антигенный фрагмент), происходящий из N1-филогенетического кластера, так и NA (или ее антигенный фрагмент), происходящий из N2-филогенетического кластера. В частности, иммуногенная композиция и/или вакцина содержит репликон РНК, где первая NA представляет собой N1-филогенетический кластер, выбранный из группы N1-классического и N1-пандемического филогенетического кластера, а вторая NA представляет собой N2-филогенетический кластер, выбранный из группы, состоящей из филогенетического кластера N2-1998 и N2-2002.
В дополнительных конкретных вариантах осуществления этого типа иммуногенная композиция и/или вакцина включает репликон РНК, где первая NA имеет N1-линию, выбранную из группы, состоящей из N1-пандемической (EU) и N1-евразийской линии Avian, и вторая NA имеет N2-филогенетическое происхождение, выбранное из группы, состоящей из линий N2-Gent/1984, N2-Italy/2003, N2-Scotland 1994 (клада 1) и линии N2-Scotland 1994 (клада 2).
Настоящее изобретение дополнительно относится к иммуногенным композициям и/или вакцинам, которые содержат два или более набора таких частиц репликона РНК альфавируса. В конкретных вариантах осуществления этого типа один набор частиц репликона РНК альфавируса кодирует одну или более NA, происходящих из одного филогенетического кластера и/или линии, тогда как другой набор частиц репликона РНК альфавируса кодирует одну или более NA, происходящих из другого филогенетического кластера и/или линии.
В конкретных вариантах осуществления этого типа первый набор частиц репликона РНК альфавируса кодирует один или более антигенов NA, которая происходит из N1-филогенетического кластера и/или его линии, или ее антигенные фрагменты, и второй набор частиц репликона РНК альфавируса кодирует один или более антигенов NA, которая происходят из N2-филогенетического кластера и/или линии, или ее антигенные фрагменты. В частности, в первом наборе антиген NA (или его фрагмент) представляет собой N1-филогенетический кластер, выбранный из групп, состоящих из N1-классического и N1-пандемического филогенетического кластера, а во втором наборе - антиген NA (или его антигенный фрагмент) представляет собой N2-филогенетический кластер, выбранный из группы, состоящей из филогенетического кластера N2-1998 и N2-2002.
В аналогичном параллельном варианте осуществления в первом наборе антиген NA (или его фрагмент) относится к N1-филогенетической линии, выбранной из группы, состоящей из N1-пандемической (EU) и N1-евразийской птичьей линии, а во втором - антиген NA (или его антигенный фрагмент) принадлежит к линии N2, выбранной из группы, состоящей из линии N2-Gent/1984, N2-Italy/2003, N2-Scotland 1994 (клада 1), и N2-Scotland 1994 (клада 2).
В других вариантах осуществления иммуногенная композиция и/или вакцина включает один набор частиц репликона РНК альфавируса, которые кодируют первый антиген, другой набор частиц репликона РНК альфавируса, которые кодируют второй антиген, и третий набор частиц репликона РНК альфавируса, которые кодируют третий антиген. Более конкретно, в иммуногенной композиции и/или вакцине антигены представляют собой либо антигены NA филогенетического кластера, выбранные из групп, состоящих из N1-классического, N1-пандемического-, N2-1998- и N2-2002 филогенетического кластера, либо они являются антигенами NA из линии, выбранной из группы, состоящей из N1-пандемической (EU), N1-евразийской Avian, N2-Gent/1984, N2-Italy/2003, N2-Scotland 1994 (клада 1), и N2-Scotland 1994 (клада 2).
В конкретных вариантах осуществления этого типа первый набор частиц репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент; второй набор частиц репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент; и третий набор частиц репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент.
В других вариантах осуществления иммуногенная композиция и/или вакцина содержит один набор частиц репликона РНК альфавируса, которые кодируют первый антиген, другой набор частиц репликона РНК альфавируса, которые кодируют второй антиген, третий набор частиц репликона РНК альфавируса, которые кодируют третий антиген, и четвертый набор частиц репликона РНК альфавируса, которые кодируют четвертый антиген.
Более конкретно, в иммуногенной композиции и/или вакцине антигены представляют собой либо антигены NA филогенетического кластера, выбранные из групп, состоящих из N1-классического филогенетического кластера, N1-пандемического филогенетического кластера, филогенетического кластера N2-1998 и филогенетического кластера N2-2002, или они являются антигенами NA линии, выбранной из группы, состоящей из линии N1-пандемической (EU), линии N1-евразийской Avian, линии N2-Gent/1984, линии N2-Italy/2003, линии N2-Scotland 1994 (клада 1), и линии N2-Scotland 1994 (клада 2).
В конкретных вариантах осуществления этого типа первый набор частиц репликона РНК альфавируса кодирует NA, происходящую из классического филогенетического кластера N1, или ее антигенный фрагмент; второй набор частиц репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент; третий набор частиц репликона РНК альфавируса кодирует NA, происходящую из филогенетического кластера N2 1998, или ее антигенный фрагмент; и четвертый набор частиц репликона РНК альфавируса кодирует NA, происходящую из пандемического филогенетического кластера N1, или ее антигенный фрагмент.
Таким образом, иммуногенные композиции/вакцины содержат множество наборов (например, 2-10) частиц репликона РНК альфавируса. В конкретных вариантах осуществления этого типа один или более наборов содержат частицы репликона РНК альфавируса, которые кодируют одну или более NA IAV-S, происходящих из одного или нескольких филогенетических кластеров (или их антигенные фрагменты) или линий (или их антигенные фрагменты). В конкретных вариантах осуществления этого типа иммуногенная композиция содержит один или более наборов частиц репликона РНК альфавируса, которые кодируют одну или более NA IAV-S, происходящих из одного или нескольких филогенетических кластеров (или их антигенные фрагменты) или линий (или их антигенные фрагменты), в сочетании с одним или несколькими наборами частиц репликона РНК альфавируса, которые кодируют один или более антигенов или их антигенных фрагментов, не относящихся к IAV-S. В еще более конкретных вариантах осуществления, один или более наборов частиц репликона РНК альфавируса кодируют как одну, так и несколько NA IAV-S, происходящих из одного или нескольких филогенетических кластеров (или их антигенные фрагменты), или линий (или их антигенные фрагменты) и кодируют один или более антигенов или их антигенных фрагментов, не относящихся к IAV-S. В конкретных вариантах осуществления этого типа, частица репликона РНК альфавируса кодирует NA, происходящую из классического филогенетического кластера N1 или ееантигенный фрагмент, и/или NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент, наряду от одного до трех антигенов или их антигенных фрагментов, не относящихся к IAV-S.
В конкретных вариантах осуществления антиген, не относящийся к IAV-S, происходит от вируса репродуктивного и респираторного синдрома свиней (PRRS). В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от цирковируса свиней (PCV). В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от трансмиссивного вируса гастроэнтерита (TGE). В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от вируса псевдобешенства свиньи (PPRV). В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от парвовируса свиней (PPV). В других вариантах осуществления антиген, не относящийся к IAV-S,происходит от ротавируса свиньи (PRV). В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от эпидемической диареи свиней (PED). В других вариантах осуществления, один или более антигенов, не относящихся к IAV-S, происходят из одного или нескольких серотипов Pasteurella multocida. В других вариантах осуществления один или более антигенов, не относящихся к IAV-S, происходят от одного или нескольких серотипов Salmonella ssp. В других вариантах осуществления, один или более антигенов, не относящихся к IAV-S, происходят от одного или нескольких серотипов Escherichia coli. В других вариантах осуществления один или более антигенов, не относящихся к IAV-S, происходят от одного или нескольких серотипов Haemophilus parasuis. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от Lawsonia intracellularis. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от Mycoplasma ssp (например, Mycoplasma hyopneumoniae). В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от Bordetella bronchiseptica. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от Erysipelas ssp. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит из Campylobacter ssp. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит от Actinobacillus pleuropneumoniae. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит из Clostridium perfringens. В других вариантах осуществления антиген, не относящийся к IAV-S, происходит из Clostridium difficile.
В более конкретных вариантах осуществления частицы репликона РНК альфавируса представляют собой частицы репликона РНК альфавируса венесуэльского конского энцефалита (VEE). В еще более конкретных вариантах осуществления частицы репликона РНК альфавируса VEE представляют собой частицы репликона РНК альфавируса VEE ТНК-83.
Настоящее изобретение дополнительно относится к комбинированным иммуногенным композициям и/или вакцинам (мультивалентным вакцинам), которые включают частицы репликона РНК альфавируса, которые кодируют одну или более NA или ее антигенных фрагментов, происходящих из IAV-S, как указано выше, и дополнительно содержат один или более модифицированных живых/аттенуированных или убитых свиных патогенов. В конкретных вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых вирусов PRRS. В других вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых PCV. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых TGE. В других вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых PPRV. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых PPV. В других вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых PRV. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых PED. В других вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых серотипов Pasteurella multocida. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых серотипов Salmonella ssp. В других вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых серотипов Escherichia coli. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых Haemophilus parasuis. В других вариантах осуществления иммуногенные композиции содержат один или более модифицированных живых/аттенуированных и/или убитых Lawsonia intracellularis. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых Mycoplasma ssp. (например, Mycoplasma hyopneumoniae). В других вариантах осуществления иммуногенные композиции также содержат один или более модифицированных живых/аттенуированных и/или убитых Bordetella bronchiseptica. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых Erysipelas ssp. В других вариантах осуществления иммуногенные композиции включают один или более модифицированных живых/аттенуированных и/или убитых Campylobacter ssp. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых Actinobacillus pleuropneumoniae. В других вариантах осуществления иммуногенные композиции включают один или более модифицированных живых/аттенуированных и/или убитых Clostridium perfringens. В других вариантах осуществления иммуногенные композиции дополнительно содержат один или более модифицированных живых/аттенуированных и/или убитых Clostridium difficile.
В конкретных вариантах осуществления конструкция нуклеиновой кислоты по настоящему изобретению кодирует один или более антигенов NA, происходящих из филогенетических кластеров IAV-S, или их антигенные фрагменты. Таким образом, хотя частицы репликона РНК альфавируса конкретно приведены ниже в качестве примера, следует понимать, что настоящее изобретение дополнительно включает другие конструкции векторов и/или нуклеиновых кислот.
Таким образом, в некоторых вариантах осуществления конструкция нуклеиновой кислоты кодирует NA IAV-S, которая происходит из филогенетического кластера N1-классический. В других вариантах осуществления конструкция нуклеиновой кислоты кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N1-классический. В других вариантах осуществления NA IAV-S происходит из филогенетического кластера N2-1998. В других вариантах осуществления частица репликона РНК альфавируса кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N2-1998. В других вариантах осуществления NA IAV-S происходит из филогенетического кластера N1-пандемический. В других вариантах осуществления частица репликона РНК альфавируса кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N1-пандемический. В других вариантах осуществления NA IAV-S происходит из филогенетического кластера N2-2002. В еще одном варианте осуществления конструкция нуклеиновой кислоты кодирует антигенный фрагмент NA, который происходит из филогенетического кластера N2-2002.
В аналогичном варианте осуществления NA IAV-S происходит от NA из N1-пандемической (EU) линии. В аналогичном варианте осуществления NA IAV-S происходит от NA из N1-евразийской линии Avian. В аналогичном варианте осуществления NA IAV-S происходит от NA из линии N2-Gent/1984. В другом аналогичном варианте осуществления NA IAV-S происходит из линии N2-Italy/2003. В еще одном аналогичном варианте осуществления NA IAV-S происходит из линии N2-Scotland/1994. В некоторых вариантах осуществления этого типа NA IAV-S происходит из линии N2-Scotland/1994 (клада 1). В других вариантах осуществления этого типа NA IAV-S происходит из линии N2-Scotland/1994 (клада 2).
В конкретных вариантах осуществления NA IAV-S, происходящая из филогенетического кластера N1-классический, содержит аминокислотную последовательность, которая имеет 95% идентичности, 97% идентичности, 98% идентичности или более с аминокислотной последовательностью SEQ ID NO: 2. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 2. В других вариантах осуществления NA IAV-S, происходящая из филогенетического кластера N1-пандемический, содержит аминокислотную последовательность, которая имеет 95% идентичности, 97% идентичности, 98% идентичности или больше с аминокислотной последовательностью SEQ ID NO: 4. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 4. В других вариантах осуществления NA IAV-S, происходящая из филогенетического кластера N2-1998, содержит аминокислотную последовательность, которая имеет 92% идентичности, 94% идентичности, 97% идентичности или более с аминокислотной последовательностью SEQ ID NO: 6. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 6. В других вариантах осуществления NA IAV-S, происходящая из филогенетического кластера N2-2002, содержит аминокислотную последовательность, которая имеет 92% идентичности, 94% идентичности, 97% идентичности или более с аминокислотной последовательностью SEQ ID NO: 8. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 8.
В конкретных вариантах осуществления NA IAV-S, происходящая из N1-пандемической (EU) линии, включает аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более, с аминокислотной последовательностью SEQ ID NO: 12. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 12. В других вариантах осуществления NA IAV-S, происходящая из N1-евразийской линии Avian, содержит аминокислотную последовательность, которая имеет 85% идентичности, 90% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 14. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 14. В других вариантах осуществления NA IAV-S, происходящая из линии N2-Gent/1984 включает аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 16. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 16. В других вариантах осуществления NA IAV-S, происходящая из линии N2-Italy/2003, содержит аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 18. В более конкретных вариантах осуществления этого типа NA содержит аминокислотную последовательность SEQ ID NO: 18. В других вариантах осуществления NA IAV-S, происходящая из линии N2-Scotland/1994 (клада 1), включает аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 20. В более конкретных вариантах осуществления этого типа, NA содержит аминокислотную последовательность SEQ ID NO: 20. В других вариантах осуществления, NA IAV-S, происходящая от линии N2-Scotland /1994 (клада 2) включает аминокислотную последовательность, которая имеет 90% идентичности, 92% идентичности, 95% идентичности или более с аминокислотной последовательностью SEQ ID NO: 22. В более конкретных вариантах осуществления этого типа, NA содержит аминокислотную последовательность SEQ ID NO: 22.
Настоящее изобретение относится к конструкциям нуклеиновой кислоты, которые кодируют два или более антигенов IAV-S или их антигенных фрагментов. В конкретных вариантах осуществления конструкции нуклеиновой кислоты кодируют от двух до четырех или более антигенов NA, которые происходят из IAV-S, или их антигенных фрагментов. В родственных вариантах осуществления конструкции нуклеиновой кислоты кодируют от двух до четырех или более антигенов NA, которые происходят из разных филогенетических кластеров IAV-S, или их антигенных фрагментов. В некоторых вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент. В еще более конкретных вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из филогенетического кластера N2-2002, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N2-1998, или ее антигенный фрагмент, NA, происходящую из филогенетического кластера N1-пандемический, или ее антигенный фрагмент, и NA, происходящую из филогенетического кластера N1-классический, или ее антигенный фрагмент.
В некоторых вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из линии N1-пандемической (ЕС), или ее антигенный фрагмент, и NA, происходящую из линии N1-евразийской Avian, или ее антигенный фрагмент. В других вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из линии N2-Gent/1984, или ее антигенный фрагмент, и NA, происходящую из линии N2-Italy/2003, или ее антигенный фрагмент. В других вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из линии N2-Gent/1984, или ее антигенный фрагмент, и NA, происходящую из линии N2 Scotland/1994 (клада 1), или ее антигенный фрагмент. В других вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из линии N2-Gent/1984, или ее антигенный фрагмент, и NA, происходящую из линии N2-Scotland/1994 (клада 2), или ее антигенный фрагмент. В других вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из линии N2-Italy/2003, или ее антигенный фрагмент, и NA, происходящую из линии N2-Scotland/1994 (клада 1), или ее антигенный фрагмент. В других вариантах осуществления конструкции нуклеиновой кислоты кодируют NA, происходящую из линии N2-Italy/2003, или ее антигенный фрагмент, и NA, происходящую из линии N2 Scotland/1994 (клада 2), или ее антигенный фрагмент. В других вариантах осуществления NA, происходящая из одной из европейских линий N1, или ее антигенный фрагмент кодируется в той же конструкции нуклеиновой кислоты, что и NA, происходящая из одной из европейских линий N2 или ее антигенный фрагмент. Аналогично, настоящее изобретение относится к конструкциям нуклеиновой кислоты, кодирующим три, четыре или более NA, причем каждая NA происходит из разных европейских линий. Кроме того, настоящее изобретение относится к конструкциям нуклеиновой кислоты, кодирующим две, три, четыре и более NA, причем каждая NA происходит из разных европейских линий и/или филогенетических кластеров.
Таким образом, настоящее изобретение относится ко всем частицам репликона РНК альфавируса по настоящему изобретению, векторам с «голой» ДНК, которые содержат конструкции нуклеиновой кислоты по настоящему изобретению, векторам с «голой» РНК, которые содержат конструкции нуклеиновой кислоты по настоящему изобретению, к конструкциям нуклеиновой кислоты по настоящему изобретению, включая синтетическую мРНК, и репликоны РНК, а также ко всем иммуногенным композициям и/или вакцинам, которые содержат конструкции нуклеиновой кислоты (например, синтетическую мРНК, репликоны РНК), частицы репликона РНК альфавируса, векторы с «голой» РНК и/или «голой» ДНК по настоящему изобретению.
Настоящее изобретение дополнительно включает вакцины (поливалентные) вакцины, включающие иммуногенные композиции по настоящему изобретению. В конкретных вариантах осуществления вакцины представляют собой неадъювантную вакцину. В других вариантах осуществления вакцины содержат адъювант. В конкретных вариантах осуществления адъювант представляет собой биоразлагаемое масло. В конкретных вариантах осуществления этого типа биоразлагаемое масло представляет собой dl-α-токоферил ацетат (ацетат витамина E). В других вариантах осуществления адъювант содержит эмульсию «масло-в-воде» с 2,5%-50% (об./об.) минерального масла. В конкретных вариантах осуществления адъювант включает эмульсию «масло-в-воде» с 2,5% (об./об.) минерального масла. В родственных вариантах осуществления адъювант включает эмульсию «масло-в-воде» с 5% (об./об.) минерального масла. В других вариантах осуществления адъювант содержит эмульсию «масло-в-воде» с 12,5% (об./об.) минерального масла. В других вариантах осуществления адъювант содержит эмульсию «масло-в-воде» с 25% (об./об.) минерального масла. В других вариантах осуществления адъювант включает эмульсию «масло-в-воде» с 50% (об./об.) минерального масла. В более конкретных вариантах осуществления адъювант включает смесь биоразлагаемого масла с адъювантом из минерального масла. В конкретных вариантах осуществления биоразлагаемое масло представляет собой dl-α-токоферил ацетат, а минеральное масло представляет собой парафиновое масло. В более конкретных вариантах осуществления биоразлагаемое масло представляет собой dl-α-токоферил ацетат, а минеральное масло представляет собой легкое парафиновое масло.
В некоторых вариантах осуществления вакцина помогает в предотвращении заболеваемости, вызванной IAV-S. В родственных вариантах осуществления антитела индуцируются у свиного индивидуума, когда свинью иммунизируют вакциной. В некоторых вариантах осуществления свинья является свиноматкой. В родственных вариантах осуществления вакцина обеспечивает защитные материнские антитела для потомства вакцинированной свиноматки. В других вариантах осуществления свиным индивидуумом является поросенок. В конкретных вариантах осуществления этого типа вакцину вводят поросенку уже в возрасте трех суток. В конкретных вариантах осуществления вакцину вводят в виде бустерной вакцины. В некоторых вариантах осуществления вакцину вводят в виде однократной дозы вакцины. В конкретных вариантах осуществления этого типа вакцину вводят в виде бустерной вакцины. В других вариантах осуществления вакцину вводят в виде многодозовой вакцины. В конкретных вариантах осуществления этого типа вакцину вводят в виде двухдозной вакцины.
Настоящее изобретение также относится к способу иммунизации свиньи (например, свиноматки или поросенка) против свиного патогена, например, IAV-S, включающему введение свинье иммунологически эффективного количества вакцины или поливалентной вакцины по настоящему изобретению. В конкретных вариантах осуществления вакцину вводят путем внутримышечной инъекции. В альтернативных вариантах осуществления вакцину вводят путем подкожной инъекции. В других вариантах осуществления вакцину вводят путем внутривенной инъекции. В других вариантах осуществления вакцину вводят путем интрадермальной инъекции. В других вариантах осуществления вакцину вводят через пероральное введение. В других вариантах осуществления вакцину вводят через интраназальное введение.
Таким образом, вакцины и поливалентные вакцины можно вводить в виде праймерной вакцины и/или в виде бустерной вакцины. В конкретных вариантах осуществления вакцину по настоящему изобретению вводят в виде одноразовой вакцины (одна доза), не требующей последующего введения. В некоторых вариантах осуществления в случае введения как праймерной вакцины, так и бустерной вакцины, праймерную вакцину и бустерную вакцину можно вводить одинаковым путем. В некоторых вариантах осуществления этого типа праймерную вакцину и бустерную вакцину вводят интрадермальным путем. В других вариантах осуществления этого типа праймерную вакцину и бустерную вакцину вводят внутримышечно. В альтернативных вариантах осуществления в случае введения как праймерной вакцины, так и бустерной вакцины, введение праймерной вакцины можно проводить одним путем, а бустерной вакцины - другим. В некоторых вариантах осуществления этого типа, праймерную вакцину можно вводить путем интрадермальной инъекции, а бустерную вакцину можно вводить перорально. В родственных вариантах осуществления этого типа праймерную вакцину можно вводить путем внутримышечной инъекции, а бустерную вакцину можно вводить перорально. В других вариантах осуществления этого типа праймерную вакцину можно вводить путем внутримышечной инъекции, а бустерную вакцину можно вводить путем интрадермальной инъекции. В других вариантах осуществления этого типа праймерную вакцину можно вводить путем внутримышечной инъекции, а бустерную вакцину можно вводить путем внутримышечного введения.
Изобретение дополнительно относится к способу иммунизации свиньи (например, свиноматки или поросенка) против iav-s, включающему инъекцию свиньи иммунологически эффективным количеством вышеописанных вакцин по изобретению. В конкретных вариантах осуществления вакцины могут включать приблизительно от 1×104 до приблизительно 1×1010 rp или выше, например. В более конкретных вариантах осуществления вакцины могут включать приблизительно от 1×105 до приблизительно 1×109 rp. В еще более конкретных вариантах осуществления вакцины могут включать приблизительно от 1×106 до приблизительно 1×108 rp.
В конкретных вариантах осуществления вакцины по настоящему изобретению вводят в дозах от 0,05 до 3 мл. В более конкретных вариантах осуществления вводимая доза составляет от 0,1 мл до 2 мл. В еще более конкретных вариантах осуществления введенная доза составляет от 0,2 мл до 1,5 мл. В еще более конкретных вариантах осуществления введенная доза составляет от 0,3 до 1,0 мл. В еще более конкретных вариантах осуществления введенная доза составляет от 0,4 мл до 0,8 мл.
Эти и другие аспекты настоящего изобретения будут лучше поняты при ссылках на следующие фигуры и подробное описание.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фигуры 1A-1D показывают ответы антител, ингибирующих сывороточную нейраминидазу (NI), специфичные для всех четырех штаммов NA из вакцинных композиций, описанных в примере 1 ниже. Образцы сыворотки собирают до первичной вакцинации (возраст 3 недели), до второй вакцинации (возраст 6 недель) и до заражения (возраст 9 недель).
Фигуры 2A и 2B изображают оценки поражения легкого у свиней после введения вакцинной композиции, описанной в примере 1 ниже, и заражения вирусной инфекцией (A) H1-гамма-N1-классический или (B) H1-дельта1-N2-2002A.
Фигуры 3A и 3B показывают вирусные титры в назальных мазках свиней после введения вакцинной композиции, описанной в примере 1 ниже, и заражения вирусной инфекцией (A) H1-гамма-N1-классический или (B) H1-дельта1-N2-2002A.
Фигуры 4A и 4B показывают вирусные титры в образцах жидкости бронхоальвеолярного лаважа свиней после введения вакцинных композиций, описанных в примере 1 ниже, и заражения вирусной инфекцией (A) H1-гамма-N1-классический или (B) H1-дельта1-N2-2002A.
Фигура 5 показывает ответы антител, ингибирующих сывороточную нейраминидазу (NI), специфичные для N1-классического штамма из вакцинных композиций, описанных в примере 2. Образцы сыворотки собирали до первой вакцинации (возраст 3 недели), до второй вакцинации (возраст 7 недель), и перед заражением (возраст 10 недель).
На фигуре 6 представлены оценки поражения легкого у свиней после введения вакцинной композиции, описанной в примере 2, и после заражения вирусом H1-гамма-N1-классический.
Фигура 7 показывает процент макроскопического поражения легкого на пятые сутки после заражения вирусом H1N2.
Фигура 8 изображает назальное выделение возбудителя после заражения вирусом H1N2.
На фигуре 9 представлен титр ингибирования нейроаминидазы (NI) для вакцинной фракции N22002 после вакцинации. Примечание: DPV=сутки после вакцинации.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к вакцинам и иммуногенным композициям, которые включают иммунологически эффективное количество одной или нескольких частиц репликона РНК альфавируса, которые кодируют одну или более нейраминидаз (NA) из одного или нескольких серотипов вирусов гриппа А. В одном из аспектов настоящего изобретения вакцины и иммуногенные композиции не содержат ни гемагглютинин (HA) или его антигенный фрагмент, ни нуклеотидную последовательность, которая кодирует HA или его антигенный фрагмент. Одно из преимуществ вакцины против вируса гриппа А, основанной на множестве NA и отсутствии HA, заключается в том, что на NA меньше влияет антигенный дрейф, чем на HA, что позволяет соответствующим вакцинам содержать антигены из меньшего количества изолятов гриппа. Это экономит производственные затраты, а также увеличивает период времени, необходимый для обновления данной вакцины против гриппа. Кроме того, поскольку в настоящее время реализуемые на рынке вакцины против вируса гриппа А основаны на инактивированных вирусах гриппа, которые генерируют гораздо более высокие титры антител против HA, чем антител против NA у животных-индивидуумов, отрицательное влияние материнских антител на иммунизацию потомства животных-индивидуумов должно быть значительно уменьшено для вакцины на основе NA.
В одном важном аспекте настоящего изобретения предлагаются вакцины и иммуногенные композиции, которые включают иммунологически эффективное количество одной или нескольких частиц репликона РНК альфавируса, которые кодируют одну или более нейраминидаз (NA) из одного или нескольких филогенетических кластеров и/или линий вируса свиного вируса А (IAV-S). В одном из аспектов настоящего изобретения вакцины и иммуногенные композиции не содержат ни гемагглютинин (HA) IAV-S, или его антигенный фрагмент, ни нуклеотидную последовательность, которая кодирует HA IAV-S или его антигенный фрагмент.
Как указано выше, вакцина против IAV-S, на основе нескольких NA и отсутствия HA, позволяет соответствующим вакцинам содержать антигены из меньшего количества изолятов IAV-S. Это экономит производственные затраты, а также увеличивает период времени, необходимый для обновления данной вакцины IAV-S. Кроме того, поскольку имеющиеся в настоящее время на рынке вакцины против IAV-S основаны на инактивированных вирусах гриппа, которые генерируют гораздо более высокие титры антител к HA, чем антител против NA у животных, отрицательное влияние материнских антител на иммунизацию поросят должно быть значительно снижено для вакцин, основанных на нейраминидазе.
Для более полного понимания изобретения предоставлены следующие определения.
Использование терминов единственного числа для удобства описания никоим образом не предназначено для ограничения. Таким образом, например, ссылка на композицию, содержащую «полипептид», включает ссылку на один или более таких полипептидов. Кроме того, ссылка на «частицу репликона РНК альфавируса» включает ссылку на множество таких частиц репликона РНК альфавируса, если не указано иначе.
Как применяют в настоящем документе термин «приблизительно» применяется взаимозаменяемо с термином «примерно» и означает, что значение находится в пределах пятидесяти процентов от указанного значения, т.е. композиция, содержащая «приблизительно» 1×108 частиц репликона РНК альфавируса на миллилитр, содержит от 5×107 до 1,5×108 частиц репликона РНК альфавируса на миллилитр.
Как применяют в настоящем документе, термины «свинья» или «свиной» используют взаимозаменяемо, и они включают все домашние виды свиней, если не указано иначе.
Как применяют в настоящем документе, «филогенетический кластер» представляет собой набор нейраминидаз вируса гриппа, которые были сгруппированы (на одной ветви) в филогенетическое дерево или эволюционное дерево, которое восходит к аналогичному (гомологичному) предку (см. Примеры 5). Для нейраминидаз IAV-S (NA), обнаруженных в США, существует два преобладающих филогенетических кластера N1, N1-пандемический и N1-классический, и два преобладающих филогенетических кластера N2, N2-1998 и N2-2002. Филогенетический кластер N1-классический содержит NA, сгруппированные вместе с NA от классического свиного гриппа H1N1. Филогенетический кластер вируса N1-пандемический содержит NA, сгруппированные вместе с NA, полученной из вируса пандемического гриппа H1N1. Филогенетический кластер N2-1998 содержит NA, сгруппированные вместе с NA от вируса гриппа человека H3N2, который попал в свиней в 1998 году, тогда как филогенетический кластер N2-2002 содержит NA, сгруппированные вместе с NA от вируса гриппа человека H3N2, который попал в свиней в 2002. [См., Anderson et al., Influenza and other Respiratory Viruses 7 (Suppl. 4): 42-51 (2013)]. В Примере 5 представлена методология определения филогенетического кластера. Для филогенетических кластеров США NA IAV-S N1-классического, N1-пандемического, N2-1998, N2-2002 соответствующие репрезентативные нейраминидазы имеют следующие соответствующие аминокислотные последовательности SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, и SEQ ID NO: 8.
Как применяют в настоящем документе, «линия» представляет собой набор нейраминидаз вируса гриппа, которые были сгруппированы (на одной ветви) в эволюционном дереве, которое связано с похожим (гомологичным) предком (см. Пример 6 ниже). Эти группировки были сделаны для европейских нейраминидаз и аналогичны, но не эквивалентны, филогенетическим кластерам для вирусов США. Определения линий можно получать с помощью легкодоступного программного обеспечения, т.е. MEGA X, как описано в примере 6 ниже. Для европейскийх линий NA IAV-S N1-пандемической (EU), N1-евразийской Avian, N2-Gent/1984, N2-Italy/4675/2003, N2-Scotland/1994 клады 1 или N2-Scotland/1994 клады 2 соответствующие репрезентативные нейраминидазы имеют следующие соответствующие аминокислотные последовательности SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, и 22.
Как применяют в настоящем документе, термин «репликон» относится к модифицированной РНК вирусного генома, в котором отсутствуют один или более элементов (например, кодирующие элементы для структурных белков), которые, если бы они присутствовали, обеспечили бы успешное распространение родительского вируса в клеточных культурах. или животных-хозяевах. В подходящем клеточном контексте репликон будет самореплицироваться и может продуцировать один или более субгеномных видов РНК.
Как применяют в настоящем документе, термин «частица репликона РНК альфавируса», сокращенно «RP», представляет собой репликон РНК, полученной из альфавируса, упакованный в структурные белки, например, капсид и гликопротеины, которые также происходят от альфавируса, например, как описано Pushko et al., [Virology 239 (2): 389-401 (1997)]. RP не может размножаться в клеточных культурах или животных-хозяевах (без вспомогательной плазмиды или аналогичного компонента), потому что репликон не кодирует структурные компоненты альфавируса (например, капсид и гликопротеины).
Термин «не относящийся к IAV-S» применяют для модификации терминов, таких как патоген, и/или антиген (или иммуноген), для обозначения того, что соответствующий патоген, и/или антиген (или иммуноген) не является ни патогеном IAV-S, ни антигеном IAV-S (или иммуногеном) и того, что белок-антиген, не относящийся к IAV-S (или иммуноген), не происходит из IAV-S.
Термины «происхождение от», «происходит от» и «происходящий от» используют взаимозаменяемо в отношении данного белкового антигена и патогена, или штамма того патогена, который его естественным образом кодирует, и, как применяют в настоящем документе, означает, что немодифицированная и/или усеченная белковая последовательность этого данного белкового антигена кодируется этим патогеном или штаммом этого патогена. Кодирующая последовательность в пределах конструкции нуклеиновой кислоты по настоящему изобретению для белкового антигена, происходящая из патогена, возможно, была подвергнута генетической манипуляции для получения модификации и/или укорочения аминокислотной последовательности экспрессируемого белкового антигена относительно соответствующей последовательности этого белкового антигена в патогене или штамме патогена (включая естественно ослабленные штаммы), из которого он происходит.
Как применяют в настоящем документе, термины «защита», или «обеспечение защиты», или «вызов защитного иммунитета», «помощь в профилактике заболеваний» и «средства защиты» не требуют полной защиты от каких-либо признаков инфекция. Например, «средства защиты» могут означать, что защита достаточна для того, чтобы после заражения симптомы основной инфекции по меньшей мере уменьшались, и/или чтобы одна или более основных клеточных, физиологических или биохимических причин или механизмов, вызывающих симптомы, были уменьшены и/или устранены. Следует понимать, что «уменьшены», как используется в данном контексте, означает по отношению к состоянию инфекции, включая молекулярное состояние инфекции, а не только физиологическое состояние инфекции.
Как применяют в настоящем документе, «вакцина» представляет собой композицию, которая подходит для применения у животных, например, свиней (включая, в некоторых вариантах осуществления людей, хотя в других вариантах осуществления она конкретно не предназначена для людей), содержащую один или более антигенов, как правило. в сочетании с фармацевтически приемлемым носителем, таким как жидкость, содержащая воду, которая при введении животному вызывает иммунный ответ, достаточно сильный, чтобы минимально помочь в защите от заболевания, возникающего в результате инфекции микроорганизмом дикого типа, т.е. достаточно сильный для помощи в профилактике заболевания и/или профилактике, улучшении или излечении заболевания.
Как применяют в настоящем документе, поливалентная вакцина представляет собой вакцину, которая включает два или более различных антигена. В конкретном варианте осуществления этого типа поливалентная вакцина стимулирует иммунную систему реципиента против двух или более различных патогенов.
Термины «адъювант» и «иммуностимулятор» используются взаимозаменяемо в настоящем документе, и определяются как одно или несколько веществ, вызывающих стимуляцию иммунной системы. В этом контексте адъювант применяют для усиления иммунного ответа к одному или нескольким антигенам/изолятам вакцины. Таким образом, «адъюванты» представляют собой агенты, которые неспецифически увеличивают иммунный ответ на определенный антиген, таким образом, уменьшая количество антигена, необходимого в любой данной вакцине, и/или частоту инъекций, необходимых для генерации адекватного иммунного ответа на интересующий антиген. В этом контексте адъювант применяют для усиления иммунного ответа к одному или нескольким антигенам/изолятам вакцины.
Как применяют в настоящем документе, «неадъювантная вакцина» представляет собой вакцину или поливалентную вакцину, которая не содержит адъюванта.
Как применяют в настоящем документе термин «фармацевтически приемлемый» испоользуют в качестве прилагательного, означающего, что модифицированное существительное подходит для применения в фармацевтическом препарате. Когда его применяют, например, для описания эксципиента в фармацевтической вакцине, он характеризует эксципиент как совместимый с другими ингредиентами композиции и не являющийся неблагоприятно вредным для предполагаемого животного-реципиента, например, свиньи.
«Парентеральное введение» включает подкожные инъекции, подслизистые инъекции, внутривенные инъекции, внутримышечные инъекции, интрадермальные инъекции и инфузию.
Как применяют в настоящем документе термин «антигенный фрагмент» в отношении конкретного белка (например, белкового антигена) представляет собой фрагмент этого белка, который является антигенным, то есть способен специфически взаимодействовать с антигенраспознающей молекулой иммунной системы, такой как иммуноглобулин (антитело) или Т-клеточный антигенный рецептор. Например, антигенный фрагмент нейраминидазы IAV-S (NA) является антигенным фрагментом белка NA. Предпочтительно, антигенный фрагмент по настоящему изобретению является иммунодоминантным для распознавания антителом и/или Т-клеточным рецептором. В конкретных вариантах осуществления антигенный фрагмент в отношении данного белкового антигена представляет собой фрагмент этого белка, который сохраняет, по меньшей мере, 25% антигенности полноразмерного белка. В предпочтительных вариантах осуществления антигенный фрагмент сохраняет, по меньшей мере, 50% антигенности полноразмерного белка. В более предпочтительных вариантах осуществления антигенный фрагмент сохраняет, по меньшей мере, 75% антигенности полноразмерного белка. Антигенные фрагменты могут быть небольшими, до 20 аминокислот, или, в другом крайнем случае, быть крупными фрагментами, в которых отсутствует только одна аминокислота из полноразмерного белка. В конкретных вариантах осуществления антигенный фрагмент содержит от 25 до 150 аминокислотных остатков. В других вариантах осуществления антигенный фрагмент содержит от 50 до 250 аминокислотных остатков.
Как применяют в настоящем документе, одна аминокислотная последовательность на 100% «идентична» или имеет 100% «идентичности» со второй аминокислотной последовательностью, когда аминокислотные остатки обеих последовательностей идентичны. Таким образом, аминокислотная последовательность на 50% «идентична» второй последовательности, когда 50% аминокислотных остатков двух аминокислотных последовательностей идентичны. Сравнение последовательностей проводят со смежным блоком аминокислотных остатков, составляющих данный белок, например, белок, или часть полипептида, который сравнивается. В конкретном варианте осуществления учитываются выбранные вставки или делеции, которые в противном случае могли бы изменить соответствие между двумя аминокислотными последовательностями.
Как применяют в настоящем документе, процент идентичности нуклеотидной и аминокислотной последовательности можно определять с использованием С, MacVector (MacVector, Inc. Cary, NC 27519), Вектор NTI (Informax, Inc. MD), Oxford Molecular Group PLC (1996) и алгоритма Clustal W с выравниванием параметров по умолчанию, и с параметрами по умолчанию для идентичности. Эти коммерчески доступные программы также можно использовать для определения последовательности сходства с использованием одинаковых или аналогичных параметров по умолчанию. Альтернативно, можно использовать расширенный поиск Blast в условиях фильтра по умолчанию, например, с помощью программы накопления GCG (Genetics Computer Group, Program Manual for the GCG Package, Version 7, Madison, Wisconsin) с использованием параметров по умолчанию.
Как применяют в настоящем документе, термин «инактивированный» микроорганизм применяется взаимозаменяемо с термином «убитый» микроорганизм. Для целей по настоящему изобретению «инактивированный» микроорганизм представляет собой организм, который способен вызывать иммунный ответ у животного, но не способен заражать животное. Антиген по настоящему изобретению (например, инактивированная Mycoplasma hyopneumoniae) может быть инактивирован агентом, выбранным из группы, состоящей из бинарного этиленимина, формалина, бета-пропиолактона, тимеросала или нагревания. В конкретном варианте осуществления инактивированные изоляты Mycoplasma hyopneumoniae в сочетании с RP по настоящему изобретению инактивируются бинарным этиленимином.
Настоящее изобретение также относится к вакцинам против множества свиных патогенов. Например, кодирующая последовательность белкового антигена или его антигенного фрагмента, или комбинация таких кодирующих последовательностей белковых антигенов, полезная в вакцине для свиней, может быть присоединена к частице репликона РНК альфавируса (RP) и/или объединена в том же RP, что и последовательность, кодирующая в вакцине NA, происходящую из IAV-S. Примеры патогенных микроорганизмов, из которых могут происходить один или более белковых антигенов или антигенных фрагментов, включают вирус репродуктивного и респираторного синдрома свиней (PRRS), цирковирус свиней (PCV), вирус трансмиссивного гастроэнтерита (TGE), вирус псевдобешенства свиней (PPRV), парвовирус свиней (PPV), свиной ротавирус (PRV), вирус эпидемической диареи свиней (PED), Pasteurella multocida с несколькими серотипами, Salmonella ssp., Escherichia coli, например, (серотипы K99, K88, 987P или F41), Haemophilus parasuis, Lawsonia intracellularis, Mycoplasma ssp. (например, Mycoplasma hyopneumoniae), Bordetella bronchiseptica, Erysipelas ssp., Campylobacter ssp., Actinobacillus pleuropneumoniae, Clostridium perfringens и Clostridium difficile.
Кроме того, настоящее изобретение относится к вакцинам, содержащим один или более RP по настоящему изобретению в комбинации с одним или несколькими другими векторами, кодирующими несколько из этих антигенов свиньи (например, бакуловирусный вектор, кодирующий белок ORF-2 из цирковируса свиньи-2, (PCV-2) и/или цирковирус свиньи-3 (PCV-3) и/или инактивированные токсоиды, происходящие из одного или нескольких из этих патогенных микроорганизмов свиньи. Кроме того, такие вакцины могут включать любую частицу репликона РНК альфавируса, которая кодирует NA, происходящую из IAV-S, в вакцине по настоящему изобретению вместе с одним или несколькими убитыми и/или модифицированными (ослабленными) живыми изолятами вируса свиньи и/или бактериями свиньи. Все такие поливалентные вакцины включены в настоящее изобретение.
Таким образом, одну или более частиц репликона РНК альфавируса (RP), которые кодируют одну или более NA, происходящих из IAV-S, можно добавлять вместе с одним или несколькими другими векторами, кодирующими один или более свиных антигенов и/или с одним или несколькми убитыми и/или модифицированными(аттенуированными) изолятами живого вируса, такими как один или более убитых или модифицированных живых штаммов IAS-V, один или более убитых и/или модифицированных живых вирусов PRRS, один или более убитых и/или модифицированных живых PCV, один или более убитых и/или модифицированных живых TGE, один или более убитых и/или модифицированных живых PPRV, один или более убитых и/или модифицированных живых PPV, один или более убитых и/или модифицированных живых PRV и один или более убитых и/или модифицированных живых PED. Кроме того, одну или более частиц репликона РНК альфавируса (RP), которые кодируют один или более NA, происходящих из IAV-S, можно добавлять вместе с одним или несколькими другими векторами, кодирующими один или более свиных антигенов и/или добавлять вместе с одной или несколькими убитыми и/или модифицированными (аттенуированными) живыми бактериями, которые также могут инфицировать свиней, в том числе, с одной или несколькими убитыми и/или модифицированными живыми Pasteurella multocida (из одного или нескольких множественных серотипов), Salmonella ssp., Escherichia coli (из одного или нескольких множественных серотипов), Haemophilus parasuis, Lawsonia intracellularis, Mycoplasma ssp. (например, Mycoplasma hyopneumoniae), Bordetella bronchiseptica, Erysipelas ssp., Campylobacter ssp., Actinobacillus pleuropneumoniae, Clostridium perfringens и Clostridium difficile.
Частицы репликона РНК альфавируса по настоящему изобретению могут быть лиофилизированы и повторно регидратированы стерильной водой. С другой стороны, когда частицы репликона РНК альфавируса хранятся раздельно, но предназначены для смешивания с другими компонентами вакцины до введения, частицы репликона РНК альфавируса могут храниться в стабилизирующем растворе этих компонентов, например, в растворе с высоким содержанием сахарозы.
В одном из аспектов вакцина по настоящему изобретению может содержать адъювант. В определенных аспектах осуществления адъювант представляет собой биоразлагаемое масло. В определенных составах биоразлагаемое масло представляет собой dl-α-токоферил ацетат (ацетат витамина E). В других составах адъювант содержит эмульсию «масло-в-воде», составляющую от 2,5% до 50% (об./об.) минерального масла. В более конкретных составах адъювант содержит эмульсию «масло-в-воде», которая содержит от 5 до 25% минерального масла. В родственных составах адъювант представляет собой смесь двух компонентов. Первый компонент состоит из капель минерального масла с приблизительным средним (объемным весом) размером около 1 мкм, который стабилизирован полисорбатом 80 (полиоксиэтилен (20) сорбитан моноолеат) в воде. Первый компонент может составлять 25 массовых процентов минерального масла и 1 массовый процент полисорбата 80 с остатком воды. Второй компонент может состоять из капель биоразлагаемого dl-α-токоферил ацетата с приблизительным средним (объемным весом) размером 400 нм, который также стабилизируется полисорбатом 80. Конкретные составы включают 15 массовых процентов dl-α-токоферил ацетата и 6 массовых процентов полисорбат 80, с остатком воды. В конкретных вариантах осуществления адъювант представляет собой X-SOLVE™ (который представляет собой комбинацию двухкомпонентных адъювантов: DILUVAC FORTETM, который основан на dl-α-токоферил ацетате, и MICROSOLTM, который основан на легком парафиновом масле [см. например, US 8597662] В родственных составах адъювант содержит капли масла субмикронного размера и капли биоразлагаемого масла, причем капли биоразлагаемого масла имеют средний размер, который отличается от среднего размера капель минерального масла [см. например, US 9084768].
Вакцину по настоящему изобретению можно легко вводить любым стандартным путем, включая внутривенную, внутримышечную, подкожную, пероральную, интраназальную, интрадермальную и/или интраперитонеальную вакцинацию. Специалисты в данной области оценят, что вакцинная композиция сформулирована соответствующим образом для каждого типа животного-реципиента и пути введения.
Таким образом, настоящее изобретение также относится к способу иммунизации свиньи против IAV-S и/или других патогенов свиньи. Один из таких способов включает введение свинье инъекции с иммунологически низким количеством вакцины по настоящему изобретению, таким образом что свинья производит соответствующие антитела к IAV-S.
Также следует понимать, что это изобретение не ограничено конкретными конфигурациями, этапами процесса и материалами, описываемыми в настоящем документе, поскольку такие конфигурации, этапы процесса и материалы могут несколько отличаться. Также следует понимать, что терминологию, используемую в настоящем документе, применяют только для описания конкретных вариантов осуществления, и она не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения и ее эквивалентами.
ТАБЛИЦА ПОСЛЕДОВАТЕЛЬНОСТЕЙ1
1Нуклеотидные последовательности представлены только в форме ДНК, хотя следует понимать, что, когда последовательность включена в конструкцию РНК, подразумевается соответствующая последовательность РНК (с урацилом, «u», заменяющим тимидин, «t»).
ПОСЛЕДОВАТЕЛЬНОСТИ
N1-классический (SEQ ID NO: 1)
atgaatactaatcaaaggatcattaccattgggacagtctgcatgattgtcggtatcatctctcttttgcttcagattggaaacattgtctcactttggattagccattcaattcagaccggatgggagaatcacactgagatgtgcaatcaaagtgtcattacttatgtcaataacacttgggtgaaccggacttatgtgaacattagcaatatcaagattgcaactattcaagatgtgactagtatcattttggccggaaattctagtctttgcccggtgtcagggtgggctgtctacagcaaagacaatagcattaggattggatcaaaaggggacattttcgtcattagagagcctttcatctcatgctcacaattggagtgccggaccttctttctgacccaaggggcattgctgaatgacaaacattcaaatggtaccgtcaaggacaggagtccttatagaaccctgatgagctgccctatcggtgaggccccttcgccatacaactcacggttcgaatctgtcgcatggtcagcatctgcttgtcatgatggaatgggatggcttacaatcgggatcagtggaccggataatggtgctgtcgcagtcttgaaatacaacggaatcattacagatacaatcaaatcttggaggaacaagattcttagaactcaagaatcagagtgtgtctgtatgaacggatcatgttttacagtcttgacagatggcccaagcaatggacaagcctcttacaaaatctttaaggtggaaaaaggaaagattatcaagtcgattgagctggatgcccccaattaccactatgaagaatgctcttgttatccagatactggcaaagtcatgtgtgtctgccgggacaattggcacgcctcaaaccggccatgggtgtcgttcaatcagaatcttgactatcaaattggatacatttgctctggagtctttggtgataaccctagatccaatgatgggaagggcaattgtggcccggtcctttctaatggagcaaatggagtgaaaggtttctcatatcggtatggaaatggtgtgtggattggtcggaccaagtcaatcaactctcggtcgggttttgagatgatttgggatccgaatggatggactgagacagattcatcattctcgatgaagcaggacattatcgctcttaatgattggtctggatactcgggatcttttgtccaacatccggagcttactggtatgaattgcattaggccttgtttctgggtggaattgatcagagggcaacccaaggaaagcactatctgggctagcggttccagcatctcattctgtggcgtcaattcggaaaccgcttcctggtcttggccagacggagctgatctgccattcaccattgacaag
N1-классический (SEQ ID NO: 2)
MNTNQRIITIGTVCMIVGIISLLLQIGNIVSLWISHSIQTGWENHTEMCNQSVITYVNNT
WVNRTYVNISNIKIATIQDVTSIILAGNSSLCPVSGWAVYSKDNSIRIGSKGDIFVIREP
FISCSQLECRTFFLTQGALLNDKHSNGTVKDRSPYRTLMSCPIGEAPSPYNSRFESVAWS
ASACHDGMGWLTIGISGPDNGAVAVLKYNGIITDTIKSWRNKILRTQESECVCMNGSCFT
VLTDGPSNGQASYKIFKVEKGKIIKSIELDAPNYHYEECSCYPDTGKVMCVCRDNWHASN
RPWVSFNQNLDYQIGYICSGVFGDNPRSNDGKGNCGPVLSNGANGVKGFSYRYGNGVWIG
RTKSINSRSGFEMIWDPNGWTETDSSFSMKQDIIALNDWSGYSGSFVQHPELTGMNCIRP
CFWVELIRGQPKESTIWASGSSISFCGVNSETASWSWPDGADLPFTIDK
N1-пандемический (SEQ ID NO: 3)
atgaatcctaaccaaaagatcattaccattggttcggtctgtatgacaattggaatggctaacctgatccttcaaattggaaacattatctcaatctgggtcagccactcaattcaaattggaaatcaatcgcagattgaaacatgcaaccaaagcgtcattacttacgaaaacaacacttgggtgaaccagacctacgtgaacatcagcaacaccaacttcgctgctggacagtccgtggtttccgtgaaactggcgggcaactcctctctctgccctgtgagcggatgggctatctactccaaagacaactcagtcagaatcggttccaagggggatgtgtttgtcataagggaaccattcatctcatgctctccccttgaatgcagaaccttcttcttgactcaaggggccttgctaaatgacaaacattccaacggaaccattaaagacaggagcccatatcggaccctgatgagctgtcctatcggtgaagtcccctcgccatacaactcaagatttgagtcagtcgcttggtcagcatccgcttgtcatgatggcatcaattggctcaccattggaatttctggcccagacagtggggcagtggctgtgctgaagtacaatggcattataacagacactatcaagtcgtggaggaacaacatattgagaactcaagagtctgaatgtgcatgtgtgaatggttcttgctttaccatcatgaccgatggaccatccgatggacaggcctcctacaagatcttcagaatcgaaaagggaaagatcgtcaaatcagtcgaaatgaatgcccctaactaccactatgaggaatgctcctgttatcctgattcctccgaaatcacgtgcgtgtgcagggataactggcatggctccaatcggccgtgggtgtctttcaaccagaatctggaatatcagatcggatacatttgctccggggtgttcggagacaatccgcgccctaatgataagacaggctcgtgtggtccagtctcgtctaacggagccaacggagtcaaaggattttcattcaaatacggcaatggagtgtggatagggagaactaagagcatttcctcaagaaaaggtttcgagatgatttgggatccgaatggatggactgggactgacaacaagttctcaatcaagcaagacatcgtgggaatcagcgagtggtcaggatatagcgggtcctttgtgcagcaccccgaactgaccgggctggattgtattagaccttgcttctgggtcgaactcatcagagggcgacccgaagagaacacaatctggactagcgggagcagcatctccttttgtggtgtgaactcggacactgtgggttggtcttggccagacggtgctgagttgccttttaccattgacaag
N1-пандемический (SEQ ID NO: 4)
MNPNQKIITIGSVCMTIGMANLILQIGNIISIWVSHSIQIGNQSQIETCNQSVITYENNT
WVNQTYVNISNTNFAAGQSVVSVKLAGNSSLCPVSGWAIYSKDNSVRIGSKGDVFVIREP
FISCSPLECRTFFLTQGALLNDKHSNGTIKDRSPYRTLMSCPIGEVPSPYNSRFESVAWS
ASACHDGINWLTIGISGPDSGAVAVLKYNGIITDTIKSWRNNILRTQESECACVNGSCFT
IMTDGPSDGQASYKIFRIEKGKIVKSVEMNAPNYHYEECSCYPDSSEITCVCRDNWHGSN
RPWVSFNQNLEYQIGYICSGVFGDNPRPNDKTGSCGPVSSNGANGVKGFSFKYGNGVWIG
RTKSISSRKGFEMIWDPNGWTGTDNKFSIKQDIVGISEWSGYSGSFVQHPELTGLDCIRP
CFWVELIRGRPEENTIWTSGSSISFCGVNSDTVGWSWPDGAELPFTIDK
N2-1998A (SEQ ID NO: 5)
atgaatccaaaccaaaagataatcacaattggctctgtttctctcactattgccacaatgtgcctccttatgcaaattgccatcctgattactaatgtcacattgcactccaatcagtacgaatgcaactaccccccaaacaaccaagtgatactgtgtgaaccaactatcatcaaaagaaacattactgagattgtgtatctggccaacaccaccatagagaaggaaatctgccccaagctggcagaatacagaaactggtcgaagccgcaatgtaaaattacagggtttgcacctttttccaaggacaattcgattaggctttctgcgggtggcgacatttgggtaacgagagaaccttatgtgtcatgcgatcctgataagtgttaccagtttgcccttggacaaggaacaacgctcaacaacagacattcaaacgacaccgtgcatgataggaccccttatcgaaccctattgatgaatgagttgggtattccattccatttggggaccaaacaagtgtgcatcgcatggtccagctcatcctgccatgatggacgggcttggcttcatgtctgtattactgggcatgacaacaatgcaactgccagcatcatttacaatggacgccttgtcgatagtattggttcatggtccaaaagaatcctcaggacccaggagtcggaatgcgtgtgcatcaatggaacttgtaccgtggtcatgactgatgggtccgcttcaggaatagctgacactaaaatcctgttcattgaagaggggaaaatcgtgcacattagcccactgctggggtccgctcagcacgtggaggagtgctcctgctatccccgatacccaggtgtcagatgcatctgtagagacaactggaaaggctccaacagacctgtcgtggatattaatgtgaaggattatagcattgtgtcctcctacgtgtgctccggactggtgggagacacccccagaaaagacgacagatccagctccagcaattgtctgaatcctaacaacgagaaaggggagcatggagtgaaaggctgggcctttgatgatggaaatgacgtgtggatggggaggacaatcaacgagacattacgctcaggttatgaaaccttcaaagtcattgaaggctggtccaaacctaattccaaattgcagataaatcgccaagtcattgttgaaagagatgataggtccggttattctggaattttctctgtcgaaggaaagagctgtatcaatcggtgtttttacgtggagctgatcagaggaaggaaacaggaaactgcagtgtggtggacgtcaaattccattgtggtgttttgtggcacctcaggtacctatggaaccggctcatggcctgatggggcggacatcaatctcatgcctgtg
N2-1998A (SEQ ID NO: 6)
MNPNQKIITIGSVSLTIATMCLLMQIAILITNVTLHSNQYECNYPPNNQVILCEPTIIKR
NITEIVYLANTTIEKEICPKLAEYRNWSKPQCKITGFAPFSKDNSIRLSAGGDIWVTREP
YVSCDPDKCYQFALGQGTTLNNRHSNDTVHDRTPYRTLLMNELGIPFHLGTKQVCIAWSS
SSCHDGRAWLHVCITGHDNNATASIIYNGRLVDSIGSWSKRILRTQESECVCINGTCTVV
MTDGSASGIADTKILFIEEGKIVHISPLLGSAQHVEECSCYPRYPGVRCICRDNWKGSNR
PVVDINVKDYSIVSSYVCSGLVGDTPRKDDRSSSSNCLNPNNEKGEHGVKGWAFDDGNDV
WMGRTINETLRSGYETFKVIEGWSKPNSKLQINRQVIVERDDRSGYSGIFSVEGKSCINR
CFYVELIRGRKQETAVWWTSNSIVVFCGTSGTYGTGSWPDGADINLMPV
N2-2002A (SEQ ID NO: 7)
atgaatccaaatcaaaagatcattactattggatcagtctcactcatcattgccacaatttgtttccttatgcaaattgcaatccttgtcactactgtcacattgcatttcaagcagcatgactacaactcccccccaaacaaccaagctactctgtgtgaaccaacaatcattgaacggaaaacaactgaaattgtgtatcttactaacaccaccattgagaaagaagtctgccccaaacttgcagagtaccggaactggtcaaagcctcaatgtaacattactggatttgcaccattttcgaaagacaattctattcggttgtctgctggtggggacatctgggtgactagggaaccttatgtgtcatgcgatcctgacaagtgttaccaatttgcccttggacagggtacaactcttaacaacggacattcgaataacacagtccatgataggaccccgtatcggacccttcttatgaatgagcttggtgtcccttttcatcttggaaccagacaagtgtgcatggcttggtctagctcatcttgtcacgatgggaaagcatggctgcatgtctgtgtcactggaaatgataacaatgctactgctagcttcatctacaatggtaggcttgtggattctattggttcgtggtcgaaaaacattctccggacccaagagtcagaatgcgtctgtatcaatggaacatgtactgtcgtcatgactgatggatccgctagtggaaaagcagataccaaaatcttgttcgtcgaagaggggaagatcgtccatatcagcactctgttgggatctgcacagcacgtcgaggaatgctcctgttatcctaggtttccgggagtccggtgtgtctgccgggacaactggaaaggatctaatagacccatcgtcgacatcaatgtcaagaattacagcattgtctcttcgtatgtctgcagtggacttgtcggtgatactcccagagagagcgactcagtctcctcatcttattgcttggatccgaacaatgagaagggtggtcatggggtgaaagggtgggcctttgatgatggtaatgacgtgtggatgggaagaacaatcaacgagactttgcgcttgggatatgaaaccttcaaagtcattgaaggctggtccacagctaactccaagtcacagacaaatagacaagtgattgtcgaaaaaggagacaggtcaggatattctgggattttctcagtcgagggaaagaactgcatcaataggtgcttctatgtggagttgattagaggacggaaagaggagacaaaagtctggtggaccagtaactcaattgtcgtgttttgtggcacctcagggacttatggtactggctcttggccggatggtgctgacatcaatctcatgccaatt
N2-2002A (SEQ ID NO: 8)
MNPNQKIITIGSVSLIIATICFLMQIAILVTTVTLHFKQHDYNSPPNNQATLCEPTIIER
KTTEIVYLTNTTIEKEVCPKLAEYRNWSKPQCNITGFAPFSKDNSIRLSAGGDIWVTREP
YVSCDPDKCYQFALGQGTTLNNGHSNNTVHDRTPYRTLLMNELGVPFHLGTRQVCMAWSS
SSCHDGKAWLHVCVTGNDNNATASFIYNGRLVDSIGSWSKNILRTQESECVCINGTCTVV
MTDGSASGKADTKILFVEEGKIVHISTLLGSAQHVEECSCYPRFPGVRCVCRDNWKGSNR
PIVDINVKNYSIVSSYVCSGLVGDTPRESDSVSSSYCLDPNNEKGGHGVKGWAFDDGNDV
WMGRTINETLRLGYETFKVIEGWSTANSKSQTNRQVIVEKGDRSGYSGIFSVEGKNCINR
CFYVELIRGRKEETKVWWTSNSIVVFCGTSGTYGTGSWPDGADINLMPI
N1-пандемический EU (SEQ ID NO: 11)
atgaatccaaaccaaaagataataaccattggttcggtctgtatgacaattggaatggctaacttaatattacaaattggaaacataatctcaatatggattagccactcaattcaacttgggaatcaaagtcagattgaaacatgcaatcaaagcgtcattacttatgaaaacaacacttgggtaaatcagacatatgttaacatcagcaacaccaactttgctgctggacagtcagtggtttccgcgaaattagcgggcaattcctccctctgccctgttagtggatgggctatatacagtaaagacaacagtgtaagaatcggttccaagggggatgtgtttgtcataagggaaccattcatatcatgctcccccttagaatgcagaaccttcttcttgactcaaggggccttgctaaatgacaaacattccaatggaaccattaaagataggagcccatatcgaaccctgatgagctgtcctattggtgaagttccctctccatacaactcaagatttgagtcggtcgcttggtcagcaagtgcttgtcacgatggcatcaattggctaacaatcggaatttctggcccagacagtggggcagtggctgtattaaagtacaatggcataataacagacactatcaagagttggaaaaacaatatattgagaacacaagagtctgaatgtgcatgtgtaaatggttcttgctttaccataatgaccgatggaccaagtgatggacaggcctcatacaagatcttcagaatagaaaagggaaagatagtcaaatcagtcgaaatgaatgcccctaattatcactatgaggaatgctcctgttatcctgattctagtgaaatcacatgtgtgtgcagggataactggcatggctcgaatcgaccgtgggtgtctttcaaccagaatctggaatatcagataggatacatatgcagtgggattttcggagacaatccacgccctaatgataagacaggcagttgtggtccagtatcgtctaatggagcaaatggagtaaaaggattttcattcaaatatggcaatggtgtttggatagggagaactaaaagcattagttcaagaaaaggttttgagatgatttgggatccaaatggatggactgggacagacaaaaacttctcaataaagcaagatatcataggaataaatgagtggtcaggatacagcgggagttttgttcagcatccagaactaacagggctgaattgtataagaccttgcttctgggttgaactaatcagagggcgacccaaagagaacacaatctggactagcgggagcagcatatccttttgtggtgtaaacagtgacactgtgggttggtcttggccagacggtgctgagttgccatttaccattgacaagtaa
N1-пандемический EU (SEQ ID NO: 12)
MNPNQKIITIGSVCMTIGMANLILQIGNIISIWISHSIQLGNQSQIETCNQSVITYENNTWVNQTYVNISNTNFAAGQSVVSAKLAGNSSLCPVSGWAIYSKDNSVRIGSKGDVFVIREPFISCSPLECRTFFLTQGALLNDKHSNGTIKDRSPYRTLMSCPIGEVPSPYNSRFESVAWSASACHDGINWLTIGISGPDSGAVAVLKYNGIITDTIKSWKNNILRTQESECACVNGSCFTIMTDGPSDGQASYKIFRIEKGKIVKSVEMNAPNYHYEECSCYPDSSEITCVCRDNWHGSNRPWVSFNQNLEYQIGYICSGIFGDNPRPNDKTGSCGPVSSNGANGVKGFSFKYGNGVWIGRTKSISSRKGFEMIWDPNGWTGTDKNFSIKQDIIGINEWSGYSGSFVQHPELTGLNCIRPCFWVELIRGRPKENTIWTSGSSISFCGVNSDTVGWSWPDGAELPFTIDK
N1-Евразийский Avian (SEQ ID NO: 13)
atgaacccaaatcagaagataataatcattagttcaatctgtatgacaaatggaattgctagcttgatattacaaattgggaacataatatcaatatggattagccattcaattcaaattgagaacccaaaccagaccgaccatgcaatcaaagcgttattatttacgaaaacaacacatgggtaaatcaaacgtatgttaacatcagcaacaataattttgttgttgaacagacagtggtttcaatgaaattagcgggcagttcttctctctgccctgttagtggatgggctatatacagtaaagataacagtgtaagaatcggttccaaaggggatgtgtttgtcataagagagccattcatctcatgctcccatttggaatgtagaaccttcttcttaactcaaggggccctactgaatgataaacattctaatggaaccgttaaagacagaagcccctatcgaaccctgatgagctgtcctattggtgaagtcccctctccatacaactcaaaatttgagtcagttgcttggtcagcaagtgcttgccatgatggcaccagttggttgacaattgggatttctggtccagacaatggagcagtggctgtgttgaaatacaatgacataataacagacactatcaagagttggaaaaacaacatattgagaacacaagaatctgaatgtgcatgtttgaatggttcttgctttactgtaatgaccgatggaccaagtaatgggcaggcctcatacaagatcttcaaaatagaaaaggggaaagtagtcaaatcagtcgagttgaatgctcctaattatcactatgaggaatgttcctgttatcctgattctggtgaaatcatatgtgtatgcagggacaattggcatggctcgaatcgaccatgggtgtctttcaatcagaatctggagtatcagataggatacatatgcagtggggttctcggagacaatccgcgccctaatgatagaacaggcagttgtggtccagtatcatctcatggagcaaatggggtaaaagggttttcgtttaaatacggcaatggaatttggatagggagaactaaaagcactattacaaggagtggttttgagatgatttgggacccaaacggatggactggaacagacaataatttctcagtgaagcaagatatcgtaggaataactaactggtcaggatacgcgggagttttgtccaacatccagaattaaccggattggattgtattagaccttgcttctgggttgaactaatcagagggagacccaaagagaacacaatctggactagcggaagcagcatatccttttgtggtgtaaatagtgacactgtgggttggtcttggccagacggtgctgagttgccatttaccattgacaagtaa
N1-Евразийский Avian (SEQ ID NO: 14)
MNPNQKIIIISSICMTNGIASLILQIGNIISIWISHSIQIENPNQTEPCNQSVIIYENNTWVNQTYVNISNNNFVVEQTVVSMKLAGSSSLCPVSGWAIYSKDNSVRIGSKGDVFVIREPFISCSHLECRTFFLTQGALLNDKHSNGTVKDRSPYRTLMSCPIGEVPSPYNSKFESVAWSASACHDGTSWLTIGISGPDNGAVAVLKYNDIITDTIKSWKNNILRTQESECACLNGSCFTVMTDGPSNGQASYKIFKIEKGKVVKSVELNAPNYHYEECSCYPDSGEIICVCRDNWHGSNRPWVSFNQNLEYQIGYICSGVLGDNPRPNDRTGSCGPVSSHGANGVKGFSFKYGNGIWIGRTKSTITRSGFEMIWDPNGWTGTDNNFSVKQDIVGITNWSGYSGSFVQHPELTGLDCIRPCFWVELIRGRPKENTIWTSGSSISFCGVNSDTVGWSWPDGAELPFTIDK
N2-Gent/1984 (SEQ ID NO: 15)
atgaatccaaatcaaaagataataacaattggttctgtttctctcactattacaacaatgtgcctcttcttgcagattgccatcctagtaactactataacattgcatttcaagcaatatgaatgcgattcccctgcaaacaaccaagtaataccgtgtgaaccaataataatagaaaaaaacataacaaaaatagtgtatttgaccaataccaccatagagaaagaggtatgcccaaaattaggggaatacaggaattggtcaaaaccacaatgcaagatcacaggatttgcacctttttctaaggacaattcaattcggctctctgcgggtggggccatttgggtcacgagagaaccttatgtgtcatgcgaccctaacaagtgttatcaatttgcattaggacagggaaccacattagataacagacattcaaatgacacaatacatgatagaaccccttttagaaccctgttgatgagtgaattaggtgttccatttcatttgggaaccagacaagtatgcatagcatggtccagttcaagttgtcacgatgggaaagcttggttgcatgtttgtgtcactgggcatgataaaaatgcaactgctagtttcatttatgacggaaagcttgtagacagcatcagttcatggtccaaaaacatactccggactcaggaatcagaatgtgtttgtatcgatggaatctgtacagtggtgatgactgatggaagtgcttcagggaaagctgatactaagatactatttattgaaaaagggaagatcattcatattagtccattgttgggaagtgctcagcatgtagaagaatgttcctgttaccctagataccctgatgtcaggtgtatttgcagggataactggaaaggttcaaataggcccatcgtagacataagaatgaaaaattatagcattggttccagttatatgtgctcaggacttgttggcgacacacccaggaacaatgatgggtctagtaatagcaattgtcggaatcccaataatgaaagaggaaatcatggagtgaaaggttgggcctttgatgatggaaatgacacatggatgggaagaactatcagcaaggactcacgcttaggttacgaaaccttcaaagttgttggtggttggtcccaacccaattccaaatcccagataaatagacaagttattgttgacagcgataatagatcaggttactctggtattttctctgttgaggggaaagattgcattaataggtgtttttatgtggaactaataagaggaaggagacaggaaactagagtgtggtggacttcgaacagtattgttgtgttctgtggcacttctggcacctatgggtcaggctcatggcccgatggagcaaacatcaatttcatgcctgtataa
N2-Gent/1984 (SEQ ID NO: 16)
MNPNQKIITIGSVSLTITTMCLFLQIAILVTTITLHFKQYECDSPANNQVIPCEPIIIEKNITKIVYLTNTTIEKEVCPKLGEYRNWSKPQCKITGFAPFSKDNSIRLSAGGAIWVTREPYVSCDPNKCYQFALGQGTTLDNRHSNDTIHDRTPFRTLLMSELGVPFHLGTRQVCIAWSSSSCHDGKAWLHVCVTGHDKNATASFIYDGKLVDSISSWSKNILRTQESECVCIDGICTVVMTDGSASGKADTKILFIEKGKIIHISPLLGSAQHVEECSCYPRYPDVRCICRDNWKGSNRPIVDIRMKNYSIGSSYMCSGLVGDTPRNNDGSSNSNCRNPNNERGNHGVKGWAFDDGNDTWMGRTISKDSRLGYETFKVVGGWSQPNSKSQINRQVIVDSDNRSGYSGIFSVEGKDCINRCFYVELIRGRRQETRVWWTSNSIVVFCGTSGTYGSGSWPDGANINFMPV
N2-Italy/4675/2003 (SEQ ID NO: 17)
atgaattcaaatcaaaagataataacaattggctctgtttctctcactattgccacactatgcctccttatgcaaattgctatcatggtaactactgtaacatttcatttcaagcagtatgaatacaactcccccccgaacaaccaagtaatgttgtgtgaaccaacgataattgaaagaaacataacagagacagtgtacatgaccaacaccaccatagtgaaagaaatatgccccaaactagcggaatacagaaattggtcaaaaccgcaatgcaaaattacaggatttgcacctttttcaaaggacaactcaattcggctttccgctggtggggacatctgggtgacaagagaaccttatgtgtcatgcgatcctaacaagtgttatcaatttgcccttgggcagggaacaacgttaaacaacaggcattcaaatgacacagtacatgatagaaccccttaccgaaccctgttgatgaatgaattgggtgttccatttcatttaggaaccaagcaggtttgcatagcttggtccagttcaagttgtcatgatggaaaagcatggttgcatgtttgtgtaactgggcatgatgaaaatgcaactgccagtttcatttacaacgagagacttgtagatagtattggttcatggtccaagaaaatcctcagaacccaggagtcggaatgcgtttgcataaatgggacttgtacagtggtgatgacagatgggagtgcttcaggtagagctgatactaaaatactattcattgaggaggggaaaatcgttcatgttagccaactgacaggaagtgctcagcatgtagaggagtgctcctgttatccccggtatcctggtgtcagatgtgtttgcagagataattggaaaggctccaataggcccattgtagatataaatgtaaaggatcatagcattgtttccagttatgtgtgctcaggacttgtcggagacacacccagaaaaaacgacagctctagcagtagtaactgcctgaatcctaacaatgaagaagggggtcatggggtgaaaggctgggcctttgatgatgaaaatgacttgtggatgggaagaacgatcagcgaaaagttacgattaggttatgaaaccttcaaggtcattgaaggctggtccaagcctaattccaaattgcagataaataggcaagtaatagttgacaaagataatagatccggttattctggtattttctctgttgaaagtaaaagttgcatcaatcggtgcttttatgtggagttgataagaggaaggaaacaggaaaatgaagtatggtggacctcaaacagcattgttgtattttgtggcacctcaggtacatatggaacaggctcatggcctgatggggcagacatcaatctcatgcctatatga
N2-Italy/4675/2003 (SEQ ID NO: 18)
MNSNQKIITIGSVSLTIATLCLLMQIAIMVTTVTFHFKQYEYNSPPNNQVMLCEPTIIERNITETVYMTNTTIVKEICPKLAEYRNWSKPQCKITGFAPFSKDNSIRLSAGGDIWVTREPYVSCDPNKCYQFALGQGTTLNNRHSNDTVHDRTPYRTLLMNELGVPFHLGTKQVCIAWSSSSCHDGKAWLHVCVTGHDENATASFIYNERLVDSIGSWSKKILRTQESECVCINGTCTVVMTDGSASGRADTKILFIEEGKIVHVSQLTGSAQHVEECSCYPRYPGVRCVCRDNWKGSNRPIVDINVKDHSIVSSYVCSGLVGDTPRKNDSSSSSNCLNPNNEEGGHGVKGWAFDDENDLWMGRTISEKLRLGYETFKVIEGWSKPNSKLQINRQVIVDKDNRSGYSGIFSVESKSCINRCFYVELIRGRKQENEVWWTSNSIVVFCGTSGTYGTGSWPDGADINLMPI
N2-Scotland/1994 (клада 1) (SEQ ID NO: 19)
atgaatccaaatcagaagataataacaattggctctgtttctctcgtcattgcaacattatgcttcttaatgcagatggccatcctaataactactgtaaaattacatttcaaacaatatgagtgcggcttccctgcgaacaaccaagtaataacatgtgagccaacagtaatagaaaggaacacaacagagatagtgtacttaactaacaccaccatagagaaagaaacatgccacaaaacagtggaatacaggaattggtcaaagcctcaatgcaaaataacaggctttgcacctttttccaaggacaattcaattcgactttctgctggtggggacatatgggtgacgagggaaccttacgtgtcatgcgagcctggcaaatgttatcagtttgcactcgggcaagggaccacactagacaataaacattcaaacgatacaatacatgacagaaccccctatcgaactctattgatgaatgaattgggtgtcccatttcatttagggacaagacaagtgtgtattgcatggtccagctcaagttgttatgatgggaaagcatggttgcatgtctgtatcactggacatgataaaaatgcaactgccagtttcatttacgatggtagacttgtagatagcattggttcatggtctaaaaatatacttagaacccaggaatcagaatgcgtttgcatcaatggggtctgtacagtagtaatgactgatggaagtgcttcgggaagagctgatactaaaatactattcattgaagaagggaaaattgttcatattagcccattagcggggagtgcacagcatgtggaggagtgctcctgttatccccgatatcctggcgtaaggtgtatctgcagagacaactggaaaggctctaacagacccgttgtggatataaatatagaagattatagcattgattccagttatgtgtgttcagggcttgttggcgacacacccagaatcaatgacggatccagtagtagctactgccgtgatcctaacaacgaaaaaggaaatcacggagtgaagggctgggcttttgacgatggaaatgatgtgtggatgggaagaacgatcaacgaagattcacgctcaggttatgaaacattcaaagtcattggtggttggtccactcctaattccaaattgcagataaataggcaagtaatagttgatagcaacaataggtcaggttattctggtgttttctccgttgaaggcaaaagctgcatcaatagatgtttctacgtggagttgataagaggaagaaggtcagaagcgcgagtatggtggacctcaaacagtattgttgtattttgtggcacttcaggtacctatggaacaggctcatggcctgatggagcagacatcaacctcatgcctatatga
N2-Scotland/1994 (клада 1) (SEQ ID NO: 20)
MNPNQKIITIGSVSLVIATLCFLMQMAILITTVKLHFKQYECGFPANNQVITCEPTVIERNTTEIVYLTNTTIEKETCHKTVEYRNWSKPQCKITGFAPFSKDNSIRLSAGGDIWVTREPYVSCEPGKCYQFALGQGTTLDNKHSNDTIHDRTPYRTLLMNELGVPFHLGTRQVCIAWSSSSCYDGKAWLHVCITGHDKNATASFIYDGRLVDSIGSWSKNILRTQESECVCINGVCTVVMTDGSASGRADTKILFIEEGKIVHISPLAGSAQHVEECSCYPRYPGVRCICRDNWKGSNRPVVDINIEDYSIDSSYVCSGLVGDTPRINDGSSSSYCRDPNNEKGNHGVKGWAFDDGNDVWMGRTINEDSRSGYETFKVIGGWSTPNSKLQINRQVIVDSNNRSGYSGVFSVEGKSCINRCFYVELIRGRRSEARVWWTSNSIVVFCGTSGTYGTGSWPDGADINLMPI
N2-Scotland/1994 (клада 2) (SEQ ID NO: 21)
atgaatccaaaccagaaaataataacgatcggttctgtctccttgatcattgcaacaatgtgctttttcatgcaagttgccattctggtaactactgtaacattgcatttcaggcagtgcgaatgcaactcctccgcaaccaaccaaataatgccatgtaaaccaacaaaaatagagagaaacataactgaaattgtgtacttaaccaataccaccataaaaacagaggtatgccccaaactagtgaaatacagggattgggcaaaaccacaatgtagaatcacagggtttgcacctttttccaaggacaattcgattcggctttctgccggtggggccatttgggtaacgagagaaccctatgtatcatgcgatcttagcaagtgttaccagtttgcgctcggacaggggactacactagacaacagacattcaaatgacacaatacatgatagaactccttatcggaccctattgatgaatgaattgggtgttccatttcatttaggaaccaggcaagtgtgtatagcttggtccagttcaagttgtcacgatggaaaagcatggctgcatgtttgtgtcactgggtatgataaaaatgctactgctagcctcatttatgacggaaggcttgtggacagcatcggttcatggtcccaaaacatcctccggacccaggaatcggaatgtgtttgtataaatggtacttgcacagtggtaatgactgatgggagtgcttcaggaaaagctgataccagaatactatttattgaagaagggaagattattcacattagtccattgacaggaagtgcacagcatgttgaagagtgttcttgttatcctcgataccccggtgtaagatgtgtttgtagagacaactggaagggctctaacagacccgtcgtggatataaatgtaaaagattataaaattaactccagttatgtatgctcaggccttgttggcgatacacccagaaacaacgatagatctagcaatagcaactgccaaaatcctaacaaccagagagggaatcatggagtgaagggctgggcctttgacgatggaaatgacatatggatgggaagaaccatcagcaatgattcacgtttaggttatgaaactttcaaagttattggtggttggtccaaacccaactccaaagttcagacaaataggcaagtcatagttgacagcgataatagatcaggttattctggcgttttctctgttgaaggcaaaagctgcatcaataggtgcttttatgtagagctaataagaggaaggagacaggaagctagagtatggtggacttcgaacagtattgttgtgttttgtggtacttcgggtacatatggttcaggctcatggcctgatggggctgacatcaatcttatgcctatataa
N2-Scotland/1994 (клада 2) (SEQ ID NO: 22)
MNPNQKIITIGSVSLIIATMCFFMQVAILVTTVTLHFRQCECNSSATNQIMPCKPTKIERNITEIVYLTNTTIKTEVCPKLVKYRDWAKPQCRITGFAPFSKDNSIRLSAGGAIWVTREPYVSCDLSKCYQFALGQGTTLDNRHSNDTIHDRTPYRTLLMNELGVPFHLGTRQVCIAWSSSSCHDGKAWLHVCVTGYDKNATASLIYDGRLVDSIGSWSQNILRTQESECVCINGTCTVVMTDGSASGKADTRILFIEEGKIIHISPLTGSAQHVEECSCYPRYPGVRCVCRDNWKGSNRPVVDINVKDYKINSSYVCSGLVGDTPRNNDRSSNSNCQNPNNQRGNHGVKGWAFDDGNDIWMGRTISNDSRLGYETFKVIGGWSKPNSKVQTNRQVIVDSDNRSGYSGVFSVEGKSCINRCFYVELIRGRRQEARVWWTSNSIVVFCGTSGTYGSGSWPDGADINLMPI
Следующие примеры служат для обеспечения дальнейшего понимания изобретения, но никоим образом не предназначены для ограничения эффективного объема изобретения.
ПРИМЕРЫ
ПРИМЕР 1
ЭФФЕКТИВНОСТЬ НЕАДЪЮВАНТНОЙ ПОЛИВАЛЕНТНОЙ ВАКЦИНЫ NA-RP, СОДЕРЖАЩЕЙ ЧЕТЫРЕ ОТДЕЛЬНЫХ КОНСТРУКЦИИ NA-RP, У ОТНЯТЫХ ОТ ГРУДИ ПОРОСЯТ
ВВЕДЕНИЕ
РНК-вирусы использовали в качестве векторов-носителей для введения вакцинных антигенов, которые были генетически встроены в их геномы. Однако их использование до настоящего времени было ограничено прежде всего включением вирусных антигенов в РНК-вирус, а затем внедрением вируса в хозяина-реципиента. Результатом является индукция защитных антител против встроенных вирусных антигенов. Частицы репликона РНК альфавируса были использованы для кодирования патогенных антигенов. Такие платформы репликонов альфавируса были разработаны из нескольких различных альфавирусов, включая вирус венесуэльского энцефалита лошадей (VEE) [Pushko et al., Virology 239: 389-401 (1997)], вируса синдбиса (SIN) [Bredenbeek et al., Journal of Virology 67: 6439-6446 (1993), содержание которых настоящим полностью включено в указанный документ], и вируса леса Семлики (SFV) [Liljestrom and Garoff, Biotechnology (NY) 9: 1356-1611 (1991), содержание которых настоящим включено в указанный документ во всей их полноте]. Кроме того, частицы репликона РНК альфавируса являются основой для нескольких лицензированных USDA вакцин для свиней и птицы. Эти вакцины включают вакцину против эпидемической диареи свиней, частицы РНК (код продукта 19U5.P1), вакцину против свиного гриппа, РНК (код продукта 19A5.D0), вакцину против птичьего гриппа, РНК (код продукта 19O5.D0) и рецептурный продукт, частицу РНК (Код продукта 9PP0,00).
Было проведено исследование для определения эффективности и иммуногенности двух уровней доз четырехфакторной неадъювантной вакцины NA-RP. Неадъювантная вакцина включала четыре конструкции RP, каждая из которых по отдельности кодировала один отдельный белок NA современного изолята США IAV-S. Вместе эти гены NA представляют два филогенетических кластера N1 и два кластера N2, преобладающих в популяции свиней в США. Неадъювантную вакцину вводили путем двух внутримышечных (в/м) вакцинаций (1 мл на дозу), отнятым от груди свиньям, серонегативных по отношению к вакцинной фракции и контрольному штамму. Эффективность четырехкомпонентной вакцины NA-RP была затем протестирована против гетерологичных инфекций N1 (вирус H1N1) и N2 (вирус H1N2).
МАТЕРИАЛЫ И СПОСОБЫ
Конструирование антигенов NA-RP:
Векторы с репликоном VEE, разработанные, чтобы экспрессировать гены нейраминидазы (NA), конструировали как описано ранее [см., США 9441247 B2; содержание которого включено в настоящий документ со следующими модификациями. Вектор «pVEK» с репликоном, полученным из TC-83 [раскрыт и описан в США 9441247 B2], расщепляли ферментами рестрикции AscI и PacI. ДНК-плазмиду, содержащую оптимизированную по кодонам последовательность открытой рамки считывания генов N1 или N2 (Таблица 1) с 5'-фланкирующей последовательностью (5'-GGCGCGCCGCACC-3') [SEQ ID NO: 9] и 3 'фланкирующей последовательностью (5' TTAATTAA-3 ') [SEQ ID NO: 10] аналогичным образом расщепляли рестрикционными ферментами AscI и PacI. Затем кассету с синтетическим геном лигировали в расщепленный вектор pVEK, и полученные клоны были переименованы в «pVHV-N1-пандемический», «pVHV-N1-классический», «pVHV-N2-2002» и «pVHV-N2-1998». Номенклатура вектора «pVHV» была выбрана для ссылки на векторы с репликоном, полученные из pVEK, содержащие трансгенные кассеты, клонированные через сайты AscI и PacI в участок множественного клонирования pVEK.
Получение частиц репликона РНК ТС-83 (RP) проводили в соответствии с ранее описанными способами [U.S. 9441247 В2 и США 8460913 В2; содержание которых включено в настоящий документ в качестве ссылки]. В кратком изложении, ДНК-вектор с репликоном pVHV и хелперные ДНК-плазмиды были линеаризованы с помощью фермента рестрикции NotI перед транскрипцией in vitro с использованием MegaScript РНК-полимераза T7 и аналога «кэпирования» (Promega, Madison, WI). Важно отметить, что у вспомогательной РНК, используемой в производстве, отсутствует субгеномная промоторная последовательность VEE, как описано ранее [Kamrud et al., J Gen Virol. 91 (Pt 7): 1723-1727 (2010)]. Очищенную РНК для репликона и вспомогательных компонентов объединяли и смешивали с суспензией клеток Vero, электропорировали в кюветах 4 мм и возвращали в среды OptiPro SFM для культивирования клеток (Thermo Fisher, Waltham, MA). После инкубации в течение ночи очищенные частицы репликона РНК альфавируса, полученные в фосфатно-солевом буфере с 5% сахарозой (масс./об.) и 1% сыворотки свиней, пропускают через мембранный фильтр 0,22 мкм и разливают в аликвоты для хранения. Титр функциональной RP определяли путем иммунофлуоресцентного анализа на монослоях инфицированных клеток Vero. Партии RP были идентифицированы в соответствии с геном, кодируемым упакованным репликоном.
ТАБЛИЦА 1
Гены N1 и N2, кодируемые в конструкциях NA-RP
Вирусы: Вирусы для заражения получали из Национальной лаборатории ветеринарных служб Министерства сельского хозяйства США.
A/swine/Illinois/A01554351/2015 (H1N1) обладает геном HA из кластера H1-гамма и геном NA из кластера N1-классический.
A/swine/Iowa/A02076654/2015 (H1N2) обладает геном HA из кластера H1-дельта1 и геном NA из кластера N2-2002A.
Вирусы выращивали в клеточной культуре MDCK. Конфлюэнтные клетки инфицировали в течение приблизительно 48 часов, пока цитопатическое действие не проявилось в более чем 70% клеточного монослоя. При сборе клеток супернатант удаляли из сосудов и осветляли путем центрифугирования перед хранением вируса при -60°С или ниже.
Животные: поросят, отнятых от груди, отбирали из стада с высоким уровнем здоровья на основе серологического скрининга, чтобы подтвердить отсутствие ранее существовавших антител HI или NI к вакцине и контрольным штаммам. Эти животные были смесью самцов и самок, примерно трехнедельного возраста на момент первичной вакцинации.
Вакцинация и заражение: группы лечения описаны в таблице 2 ниже. Четырех-компонентную вакцину NA-RP формулируют на два уровня дозы: 106 копий на каждую RP/дозу (низкая доза) и 107 на каждую RP/дозу (высокая доза) на основе количественного анализа на основе иммунофлуоресценции для количественного определения функциональных RP. Антигены NA-RP формулируют в стабилизаторе, состоящем из 1% сыворотки свиней и 5% сахарозы. Вакцина плацебо состояла из того же стабилизатора, но без антигена. Неадъювантные вакцины вводили свиньям в возрасте 3 и 7 недель в/м в дозе 1 мл. Уровни дозы подвергали обратному титрованию с помощью теста ИФА на оставшемся вакцинном материале после вакцинации. Образцы сыворотки собирали в сутки первой вакцинации, сутки второй вакцинации и сутки заражения.
ТАБЛИЦА 2
Группы лечения
(1 мл)
Вирус для заражения вводили свиньям через 3 недели после второй вакцинации. Материалы для заражения вирусами H1N1 (H1-гамма-N1-классический) и H1N2 (H1-дельта-1b-N2-2002A) были сформулированы для целевых доз 106,5 TCID50/свинью (объем 6 мл). Материал для заражения вводили внутритрахеально. Дозу вируса для заражения подтверждали путем обратного титрования оставшегося материала для заражения. У свиней собирали назальные мазки в -1, 1, 3 и 5 сутки после заражения.
Вскрытие производили через 5 суток после заражения (DPC). Свиней подвергали эвтаназии путем передозировки барбитурата в 5 DPC под наблюдением лицензированного ветеринарного врача. Собирали легкие и наблюдали с целью документирования площади поверхности каждой доли, пораженной макроскопическими поражениями, в результате чего получали всесторонний процент оценки повреждения легких. Бронхоальвеолярный лаваж и назальные мазки собирали у всех свиней, чтобы измерить титры вируса. Срезы легких собирали для анализа микроскопических повреждений.
Анализ иммунного ответа: IAV-S-специфические антитела в образцах сыворотки свиней определяли по HI и NI тестам. Сыворотки инактивировали нагреванием в течение 30-60 минут при 56°C. Для HI сыворотки также обрабатывали ферментом, разрушающим рецептор, и/или каолином, и абсорбировали на эритроциты индейки для удаления неспецифических агглютининов. Тест HI проводили как описано ранее [Kitikoon et al., Methods Mol Biol 1161: 295-301 (2014)] с использованием эритроцитов от индейки. Тест NI проводили с незначительными модификациями способом, описанным ранее [Sandbulte и Eichelberger, Methods Mol Biol 1161: 337-45 (2014)]. В кратком изложении, 2-кратные серийные разведения сыворотки смешивали с экспрессированным белковым антигеном в равных объемах на покрытых фетуином 96-луночных планшетах и инкубировали в течение ночи при 37°C. Конъюгат пероксидазы хрена с арахисовым агглютинином (PNA-HRP) добавляли в течение 2 часов при комнатной температуре, чтобы связать фетуиновые молекулы, лишенные сиаловой кислоты. Сигнал получали при помощи субстрата TMB, а результаты считывали при 650 нм. Среднее значение оптической плотности (OD) отрицательного контроля (без добавления антигена NA) вычитали из всех лунок. Затем значения OD тестируемых образцов были нормализованы по шкале 0-100%, где среднее значение OD для лунок положительного контроля (содержащих NA-антиген, но не сыворотку) определяли как 100%. Титр NI антитела определяли как наибольшее разведение образца, которое ингибировало ≥ 50% активности нейраминидазы.
Патологическое исследование легких: Макроскопические поражения, наблюдаемые на внешней стороне всех долей легких (хорошо разграниченные уплотнения от пурпурного до сливового цвета), были зафиксированы на сетках диаграмм легкого спереди и сзади. Комплексные баллы (процент поражения легкого) для каждой свиньи рассчитывали в зависимости от количества сеток с поражениями.
Выделение вируса: Назальные мазки и жидкость бронхоальвеолярного лаважа 10-кратно серийно разбавляли заражающей средой [минимальная поддерживающая среда Дульбеко (DMEM) с добавлением 0,3% бычьего сывороточного альбумина, фракция V; 2 мМ L-глутамина; 25 мкг/мл гентамицина; 2 мкг/мл трипсина IX], и 100 мкл каждого разведения добавляли в четыре лунки с конфлюэнтными клетками MDCK в 96-луночном планшете. Планшеты инкубировали при 37°С с 5% СО2 и наблюдали через 72 часа на наличие инфекционного вируса по тестам гемагглютинации супернатантов из каждой лунки. Титры IAV-S рассчитывали по способу Спирмена-Карбера и выражали в виде log10 TCID50 на мл.
РЕЗУЛЬТАТЫ
Иммунные ответы свиней, вакцинированных поливалентной NA-RP, см. фигура 1А-1D.
Четырех-компонентная вакцина NA-RP, высокие и низкие дозы вакцины (107 и 106 RP копий/дозу соответственно) индуцировали функциональные титры NI антитела против всех четырех компонентов NA.
Титры NI, индуцированные высокой дозой вакцины, были выше приблизительно в 1-2 раза.
Свиньи в группе вакцины плацебо остались серонегативными.
Эффективность четырехкомпонентной вакцины NA-RP против заражения инфекциями:
Поражения легкого: см., фигуры 2A-2B.
Вирусная инфекция H1N1 вызвала более 13% поражений легких в контрольной группе свиней с плацебо. Обе группы лечения, иммунизированные высокой дозой и низкой дозой, имели значительно более низкие оценки поражения легкого на 5 сутки после заражения H1N1, чем иммунологически наивная группа с плацебо (р <0,05).
Защита в отношении поражений легкого была надежной независимо от уровня дозы вакцины.
Вирусная инфекция H1N2 вызвала только 3,2% поражений легких в контрольной группе свиней с плацебо. Обе группы лечения имели более низкие оценки поражения легких на 5 сутки после заражения H1N2, чем иммунологически наивная группа с плацебо, со средними показателями поражения легкого ниже 1,1% и 1,2%, но различия между вакцинированными группами и группой плацебо не были статистически значимыми.
Назальное выделение вируса: см., фигуры 3A-3B.
Выделение вируса заражения H1N1 было максимальным на 3 сутки после заражения (DPC) для всех трех групп лечения.
На 5 DPC наблюдалось значительное снижение титра вируса в обеих группах, получавших четырехкомпонентные RP с высокой и низкой дозой, по сравнению с контролем с плацебо (p <0,05).
Назальное выделение вируса заражения H1N2 было более отсроченным, при этом средние титры в группе с плацебо не превышали нижний предел детекции до 5 DPC. На 5 DPC для вакцинированных групп с обеими уровнями доз значительно снизились вирусные титры H1N2 (p <0,05).
Вирус в легких: см., фигуры 4A-4B.
Титр вируса H1N1 в жидкости бронхоальвеолярного лаважа (BALF), собранной на 5 DPC, был более чем на 2 log больше в группе вакцины с плацебо, чем в любой из групп, вакцинированных четырехкомпонентным (4-компонентным) NA-RP, где не было обнаружено положительных титров. Это представляло статистически значимое отличие для обеих вакцинированных групп (р <0,05).
Вирусные титры в группах, зараженных H1N2, показали снижение на 2-3 log между обеими вакцинированными группами и группой, получавшей вакцину с плацебо, и были статистически значимыми (р <0,05.
Краткие выводы
Инфекция H1N1 была более надежной моделью заражения, с высокими показателями поражения легких, сопровождаемыми назальными и легочными вирусными титрами. Группы, вакцинированные четырехкомпонентными NA-RP, продемонстрировали статистически значимую защиту от вируса H1N1 в отношении тяжести повреждения легкого, выделения вируса и титра вируса в BALF. Отдельные животные в группе вакцины с низкой дозой 4-компонентной вакцины NA-RP, у которой развились более слабые титры NI в сыворотке (<80), также были защищены. Вирусная инфекция H1N2 была менее надежной моделью, поскольку поражение легких было низким в группе плацебо. Однако даже в этой группе существовала значительная защита вакцинированных групп в отношении назального выделения и титра вируса в легких. Поливалентные вакцинные композиции NA-RP, не содержащие адъювантов, являются иммуногенными и демонстрируют эффективность против заражения антигенно сходными штаммами NA.
ПРИМЕР 2
ЭФФЕКТИВНОСТЬ АДЪЮВАНТНОЙ ЧЕТЫРЕХКОМПОНЕНТНОЙ ВАКЦИНЫ NA-RP ПРОТИВ ЗАРАЖЕНИЯ H1N1 У СВИНЕЙ, ОТНЯТЫХ ОТ ГРУДИ, С АНТИТЕЛОМ К N1
Во время первичной вакцинации
Было проведено исследование с целью определения эффективности и иммуногенности четырехкомпонентной вакцины NA-RP с адъювантом. Адъювантная вакцина включала четыре конструкции RP, каждая из которых по отдельности кодировала один, различный ген NA современного изолята IAV-S в США. Вместе эти гены NA представляют два филогенетических кластера N1 (N1-классический и N1-пандемический) и два кластера N2 (N2-1998 и N2-2002). Адъювантную вакцину вводили двумя внутримышечными (в/м) вакцинациями (1 мл на дозу) отлученным от груди поросятам, серопозитивным к N1-классическому антигену во время их первичной вакцинации. Эффективность адъювантной четырехкомпонентной вакцины NA-RP тестировали против гетерологичной инфекции N1 (вирус H1N1).
МАТЕРИАЛЫ И СПОСОБЫ
Конструкция антигенов NA-RP: репликация дефектных частиц РНК (RP), содержащих репликон РНК альфавируса, экспрессирующий ген нейраминидазы (NA), получали, как описано в пример 1.
ТАБЛИЦА 3
Гены N1 и N2, кодируемые в конструкциях NA-RP
Вирусы: Вирусы для заражения получали из Национальной лаборатории ветеринарных служб Министерства сельского хозяйства США. A/swine/Illinois/A01554351/2015 (H1N1) обладает геном HA из кластера H1-гамма и геном NA из кластера N1-классический. Вирусы выращивали в клеточной культуре MDCK. Конфлюэнтные клетки инфицировали в течение приблизительно 48 часов, пока цитопатическое действие не проявилось в более чем 70% клеточного монослоя. При сборе клеток супернатант удаляли из сосудов и осветляли путем центрифугирования перед хранением вируса при -0°С или ниже.
Животные: поросят, отнятых от груди, отбирали из стада с высоким уровнем здоровья на основе серологического скрининга, чтобы подтвердить отсутствие ранее существовавших антител HI или NI к вакцине и контрольным штаммам. Эти животные были смесью самцов и самок, примерно трехнедельного возраста на момент первичной вакцинации.
Вакцинация и заражение: группы лечения описаны в таблице 4 ниже. Четырехкомпонентную вакцину NA-RP формулируют по 106 копий на каждую RP/дозу на основе количественного анализа на основе иммунофлуоресценции для количественного определения функциональных RP. Антигены NA-RP формулируют в стабилизаторе, состоящем из 1% сыворотки свиней и 5% сахарозы. Вакцина плацебо состояла из того же стабилизатора, но без антигена. Вакцины смешивали с адъювантом Xsolve (1:1 об./об., 1 мл доза) перед в/м введением свиньям в возрасте 3 и 7 недель. Уровни дозы подвергали обратному титрованию с помощью теста ИФА на оставшемся вакцинном материале после вакцинации. Образцы сыворотки собирали в сутки первой вакцинации, сутки второй вакцинации и сутки заражения.
За сутки до первой вакцинации всех свиней взвешивали, свиней в группах, позитивных по антителу к N1-классическому антигену инъецировали подкожно 2 мл/кг дозы гипериммунной сыворотки с N1-классическим (NI титр 1:2560 к N1-классический антигену).
ТАБЛИЦА 4
Группы лечения
(n=10 свиней на группу)
(с Xsolve®50)
Вирус для заражения вводили свиньям через 3 недели после второй вакцинации. Материалы для заражения вирусом H1N1 (H1-гамма-N1-классический) были сформулированы для целевых доз 106,5 TCID50/свинью (объем 6 мл). Материал для заражения вводили внутритрахеально. Дозу вируса для заражения подтверждали путем обратного титрования оставшегося материала для заражения. У свиней собирали назальные мазки в -1, 1, 3 и 5 сутки после заражения.
Вскрытие производили через 5 суток после заражения. Свиней подвергали эвтаназии путем передозировки барбитурата в 5 DPC под наблюдением лицензированного ветеринарного врача. Собирали легкие и наблюдали с целью документирования площади поверхности каждой доли, пораженной макроскопическими поражениями, в результате чего получали всесторонний процент оценки повреждения легких. Бронхоальвеолярный лаваж и назальные мазки собирали у всех свиней, чтобы измерить титры вируса. Срезы легких собирали для анализа микроскопических повреждений.
Анализ иммунного ответа: IAV-S-специфические антитела в образцах сыворотки свиней определяли по NI тесту. Сыворотку инактивировали нагреванием в течение 30-60 минут при 56°C. Тест NI проводили с незначительными модификациями способом, описанным ранее [Sandbulte и Eichelberger, Methods Mol Biol 1161: 337-45 (2014)]. В кратком изложении, 2-кратные серийные разведения сыворотки смешивали с экспрессированным белковым антигеном в равных объемах на покрытых фетуином 96-луночных планшетах и инкубировали в течение ночи при 37°C. Конъюгат пероксидазы хрена с арахисовым агглютинином (PNA-HRP) добавляли в течение 2 часов при комнатной температуре, чтобы связать фетуиновые молекулы, лишенные сиаловой кислоты. Сигнал получали ри помощи субстрата TMB, а результаты считывали при 650 нм. Среднее значение оптической плотности (OD) отрицательного контроля (без добавления антигена NA) вычитали из всех лунок. Затем значения OD тестируемых образцов были нормализованы по шкале 0-100%, где среднее значение OD для лунок положительного контроля (содержащих NA-антиген, но не сыворотку) определяли как 100%. Титр NI антитела определяли как наибольшее разведение образца, которое ингибировало ≥ 50% активности нейраминидазы.
Патологическое исследование легких: Макроскопические поражения, наблюдаемые на внешней стороне всех долей легких (хорошо разграниченные уплотнения от пурпурного до сливового цвета), были зафиксированы на сетках диаграмм легкого спереди и сзади. Комплексные баллы (процент поражения легкого) для каждой свиньи рассчитывали в зависимости от количества сеток с поражениями.
Выделение вируса: Назальные мазки и жидкость бронхоальвеолярного лаважа 10-кратно серийно разбавляли заражающей средой [минимальная поддерживающая среда Дульбеко (DMEM) с добавлением 0,3% бычьего сывороточного альбумина, фракция V; 2 мМ L-глутамина; 25 мкг/мл гентамицина; 2 мкг/мл трипсина IX], и 100 мкл каждого разведения добавляли в четыре лунки с конфлюэнтными клетками MDCK в 96-луночном планшете. Планшеты инкубировали при 37°С с 5% СО2 и наблюдали через 72 часа на наличие инфекционного вируса по тестам гемагглютинации супернатантов из каждой лунки. Титры IAV-S рассчитывали по способу Спирмена-Карбера и выражали в виде log10 TCID50 на мл.
РЕЗУЛЬТАТЫ
Иммунные ответы свиней, вакцинированных поливалентными NA-RP (см., фигура 5)
Свиньи, которым была пассивно перенесена N1-классическая гипериммунной сывороткой за один день до получения первичной вакцинации, имели титр 40-80 на момент вакцинации.
После двух прививок 4-компонентной вакциной NA-RP с Xsolve 50 свиньи, которые были негативными по антителу к N1-классическому антигену, индуцировали приблизительно в 3 раза более высокий титр выше титр NI по сравнению с N1-классическим антигеном по сравнению с свиньями, позитивными по антителу к N1-классическому антигену в время их первичной вакцинации.
Титр NI свиней в группе, негативной по N1-классическому антигену/вакцинированной плацебо, оставался серонегативным.
Эффективность четырехкомпонентной вакцины NA-RP против заражения инфекциями, поражения легкого (см. фигура 6).
4-компонентная вакцинация NA-RP была очень эффективной в уменьшении поражения легкого при вакцинации свиней без антитела к N1-классическому антигену во время их первичной вакцинации. Процент поражения легкого был значительно снижен по сравнению с обеими группами, вакцинированными плацебо.
Свиньи, вакцинированные в присутствии титра NI к N1-классическому антигену, также имели значительно уменьшенное поражение легкого по сравнению с обеими группами, вакцинированными плацебо.
Краткие выводы
Четырехкомпонентная адъювантная вакцинация NA-RP у свиней, серопозитивных и серонегативных по N1-классическому антигену, продемонстрировала статистически значимую защиту от вируса H1N1 в отношении тяжести поражения легкого по сравнению с группами, вакцинированными плацебо. Уровень N1-классического титра был снижен, но все же был положительным, когда свиньи были вакцинированы в присутствии пассивно перенесенных анти-N1-классических антител. Кроме того, четырехкомпонентная вакцина NA-RP, используемая с адъювантом Xsolve, неожиданно оказалась высокоиммуногенной по отношению к N1-классическому антигену при вакцинации свиней, серонегативных по N1-классическому антигену.
ПРИМЕР 3
ОЦЕНКА ЭФФЕКТИВНОСТИ 4-КОМПОНЕНТНОЙ НЕЙРАМИНИДАЗНОЙ (NA) ВАКЦИНЫ, СФОРМУЛИРОВАННОЙ С КОНСТРУКЦИЯМИ RP С ДВОЙНЫМ ГЕНОМ NA
Вакцина в этом исследовании состояла из двух конструкций частиц РНК, каждая из которых кодирует два гена NA из современного изолята IAV-S из США. Одна конструкция имеет два филогенетических кластера N1 (N1-классический и N1-пандемический), а другая имеет два кластера N2 (N2-1998 и N2-2002). Вакцины формулируют в разных дозах без адъюванта. Задача состояла в том, чтобы определить эффективность и иммуногенность 4-компонентных вакцин NA-RP, сформулированных в различных дозах, против инфекции вирусом H1N2 (N2-2002) у отнятых от груди свиней, серонегативных по отношению к фракциям вакцины.
Материалы и Способы
Конструкция NA-RP антигенов с двойным геном (DG) и состав вакцины
Каждая конструкция состоит из открытой рамки считывания неструктурного белка альфавируса, субгеномного промотора, за которым следует одна последовательность гена гликопротеина NA, промежуточной последовательности, второго субгеномного промотора, за которым следует второй ген гликопротеина NA, и, наконец, 3'-нетранслируемой области альфавируса. DG NA, включенный в единую конструкцию RP, был получен из циркулирующих в настоящее время изолятов вируса гриппа свиней (SIV) в США и использовался для синтеза генов NA в плазмидном векторе pVHV (см. Таблицу 5). Плазмидный вектор pVHV представляет собой производное невирулентного штамма человеческой вакцины для вируса венесуэльского лошадиного энцефалита (TC-83). Завершенные плазмиды с репликонами были проверены на состав последовательности и транскрибированы в РНК in vitro. Каждый репликон РНК NA вводили путем электропорации в клетки Vero вместе со вспомогательной РНК, кодирующий капсидный хелпер VEE и гликопротеиновую последовательность, вызывающую упаковку репликона РНК NA в RP. Мультивалентный иммунофлуоресценционный анализ использовали для для количественной оценки функциональных N1- и N2-специфических RP и определения дозы вакцины. Тестовый материал серийно разбавляли, добавляли в монослой клеток Vero в 48-луночных планшетах и инкубировали при 37°С в течение 18-24 часов. Клетки фиксировали и окрашивали первичным антителом к N1 или N2 с последующим вторичным антителом, конъюгированным с FITC. Считали отдельные антиген-положительные клетки и рассчитывали титр к единицам RP/мл.
ТАБЛИЦА 5
Гены N1 и N2, кодируемые в конструкциях NA-RP
(номер доступа GENBANK) и положение гена
3': A/swine/IA/A01410307/2014 (H1N1)
3': A/swine/IL/A01475495/2014 (H1N2)
Вакцины формулируют для получения четырех различных уровней дозы (см., таблица 6) в стабилизаторе, состоящем из consisting мононатриевого глутамината, сахарозы, PVP, сульфата натрия и HEPES. Вакцина-плацебо состояла из фосфатно-солевого буфера (PBS) без каких-либо конструкций RP.
ТАБЛИЦА 6
Дозы вакцины
Вирус
Вирус для заражения, A/swine/IL/A01475495/2014 (H1N2), получали из Национальной лаборатории ветеринарных служб Министерства сельского хозяйства США. Изолят H1N2 обладает геном HA из кластера H1-дельта1 и геном NA из кластера N2-2002. Вирус выращивали в клеточной культуре MDCK. Конфлюэнтные клетки инфицировали в течение приблизительно 48 часов, пока цитопатическое действие не проявлялось в более чем 70% клеточного монослоя. При сборе клеток супернатант удаляли из сосудов и осветляли путем центрифугирования перед хранением вируса при -60°С или ниже.
Животные
Поросят, отнятых от груди, отбирали из стада с высоким уровнем здоровья на основе серологического скрининга, чтобы подтвердить отсутствие ранее существовавших антител, ингибирующих нейраминидазу (NI), к вакцине и контрольным штаммам. Эти животные были смесью самцов и самок, примерно трехнедельного возраста на момент первичной вакцинации.
Вакцинация, заражение и сбор образцов; патологическое исследование легких; выделение вируса и тест на ингибирование нейроаминидазы (NI).
Проводили, как описано выше.
Результаты
Эффективность 4-компонентной NA-DG-RP вакцины против заражения
Макроскопическое поражение легкого показано на фигуре 7. Заражение H1N2 привело в результате в среднем к 16,7% поражений легких у свиней с контролем плацебо. Вакцинированные свиньи независимо от уровня дозы имели в среднем 3,3% поражения легких или менее, что было достоверно ниже, чем в группе плацебо (р <0,0001).
Назальное выделение вируса (см., фигура 8)
Все свиньи были негативны по отношению к вирусу в момент времени перед заражением. Ни одна из вакцинированных свиней не выделяла вирус на 1 DPC, в то время как одна из свиней в группе плацебо выделила 102,5 TCID50/мл вируса. НА 3 DPC свиньи, вакцинированные первыми двумя сформулированными наивысшими дозами (вакцина 1 и 2), имели значительно меньшее количество вируса в назальной полости по сравнению со свиньями с плацебо. На 5 DPC только свиньи, вакцинированные самой высокой дозой вакцины (вакцина 1), значительно снизили выделение вируса по сравнению с группой с плацебо.
Иммунный ответ на вакцинацию
Титры ингибирования нейраминидазы (NI) показаны на фигуре 9. Вакцины, сформулированные с различными уровнями доз, индуцировали сероконверсию у всех вакцинированных свиней (титр N2-2002> 40), и пик титра был на 14 суток после второй вакцинации. Средний титр, обнаруженный на 14 DPV2 и -1DPC, достоверно не различался между вакцинированными группами. Однако оказалось, что уровень титра NI, который обнаруживается, имеет тенденцию к связи с дозой вакцины (более высокая доза, более высокий титр NI). Свиньи в группе плацебо оставались серонегативными на всем протяжении сбора образцов в разные моменты времени.
Краткие выводы
Инфекция H1N2 вызывала высокие оценки повреждения легких, сопровождающиеся высоким назальным выделением вируса. Защита в отношении поражения легких была надежной независимо от уровня тестируемой дозы вакцины (самая низкая доза N2-DG-RP составляла 2,5×105 RP/мл). Для конструкции N2-DG-RP требуется минимальный уровень 2,5×107 RP/мл, чтобы значительно уменьшить выделение вируса после первых 5 суток после заражения. Все свиньи имели положительный титр N22002 через две недели после второй вакцинации. Выявленные уровни антител проявляются в зависимости от дозы вакцины, причем более высокие уровни наблюдаются у свиней, получающих более высокую дозу вакцины.
ПРИМЕР 4
ОЦЕНКА ЭФФЕКТИВНОСТИ ВАКЦИНЫ С ДВУХКОМПОНЕНТНОЙ НЕЙРАМИНИДАЗОЙ (NA), СФОРМУЛИРОВАННОЙ С КОНСТРУКЦИЯМИ RP С ОДНИМ ГЕНОМ NA
Было проведено исследование для определения эффективности и иммуногенности пяти различных двухвалентных вакцин с NA-RP с адъювантом, каждая из которых по отдельности кодировала один отдельный ген NA современного изолята IAV-S в EU. Вместе эти гены NA представляют собой две филогенетические линии N1 и три N2 линии N2 (см. Таблицу 7 ниже). Адъювантную вакцину вводили свиньям двцумя внутримышечными (в/м) вакцинациями в возрасте 5 и 8 недель (1 мл на дозу; 5×106 RP/дозу). Эффективность пяти адъювантных вакцин NA-RP была индивидуально проверена против их соответствующей заражающей инфекции, как указано в таблице 8 ниже.
МАТЕРИАЛЫ И СПОСОБЫ
Конструкция антигенов NA-RP:
Репликацию дефектных частиц РНК (RP), содержащих репликон РНК альфавируса, экспрессирующий ген нейраминидазы (NA), получали как описано в примере 1.
Эксперименты по серологическому ответу:
Группы лечения описаны в таблице 8 ниже. Вакцины NA-RP формулируют с уровнем дозы 5×106 RP/дозу в объеме 1 мл с адъювантом (XSolve50). Вакцины вводили в/м в дозе 1 мл свиньям в возрасте 5 и 8 недель. Образцы сыворотки крови собирали через две недели после последней вакцинации для количественной оценки титров NI антител против гетерологичного штамма гриппа, принадлежащего к соответствующей линии, и гомологичного белка NA.
Вакцинация и заражение:
Девять свиней на группу были иммунизированы дважды внутримышечным путем в возрасте 5 и 8 недель с помощью вакцины RP, кодирующей антигены NA из двух разных линий, с адъювантом X-Solve50. Группа плацебо состояла из семи свиней каждая и была вакцинирована RP, кодирующим посторонний антиген, с X-solve50. Через две недели после второй вакцинации собирали образцы крови у всех свиней для количественного определения титров антител, ингибирующих нейраминидазу (NI) против гетерологичного штамма гриппа, принадлежего к соответствующим линиям, и средние титры NI представлены в таблице. После сбора образцов крови свиньи были заражены гетерологичным штаммом гриппа A, подобранным по линии происхождения к одному из компонентов вакцины. Свиней умерщвляли через 3 дня после заражения, собирали шесть образцов легких от каждой свиньи (по одному от каждой доли легкого) и определяли вирусную нагрузку, используя объединенные гомогенаты легких. Значения представляют собой среднее геометрическое значение титра вируса (log10 TCID50/мл). Данные анализируют с помощью U-критерия Манна-Уитни*. Значения значимо отличаются (P <0,05) от соответствующей невакцинированной группы.
ТАБЛИЦА 7
Гены N1 и N2 кодируемые в конструкциях европейских NA-RP
(H1N1)
ALX30323
AKJ81669
ALX30429
(H1N2)
ALX30277
(H1N2)
AKJ82042
ALX29925
а) Нуклеотидные и аминокислотные послеовательности
ТАБЛИЦА 8
Группы лечения и результаты заражения после вакцинации
Результаты
Результаты этого эксперимента представлены в таблице 8. Все пять вакцин NA-RP индуцировали функциональные титры антител против NI у свиньи против гетерологичного штамма гриппа, принадлежащего к соответствующей линии и гомологичному белку NA.
Собирали образцы легкого у каждой свиньи и объединяли гомогенаты легких. Указанные значения представляют собой среднее геометрическое значение титра вируса (log 10 TCID50/мл). Как видно из таблицы 8, значения вакцинированных свиней значимо отличаются (P <0,05) от соответствующих невакцинированных групп, что указывает на успех вакцин NA-RP в отношении вирусной нагрузки при заражении гетерологичными штаммами вируса.
ПРИМЕР 5
ОПРЕДЕЛЕНИЕ ФИЛОГЕНЕТИЧЕСКИХ КЛАСТЕРОВ ДЛЯ США
Определение кластера нейраминидазы (NA) для США
Кластер США N1:
(1) Идентичность аминокиллот в пределах кластеров N1
а. N1пандемический или N-пандемический составляет ~ 97%-100% (средний % различий составляет 3,12%)
б. N1классический или N1-классический составляет ~ 95%-100% (средний % различий составляет 4,91%)
(2) Идентичность аминокислот между кластерами N1 составляет ~ 82% (средний % различий составляет 18,26%)
Кластер США N2:
(1) Идентичность аминокиллот в пределах кластеров N2
а. N21998 или N2-1998 составляет ~ 94%-100% (средний % различий составляет 5,94%)
б. N22002 или N2-2002 составляет ~ 94% -100% (средний % различий составляет 6,37%)
(2) Идентичность аминокислот между кластерами N2 составляет ~ 88% (средний % различий составляет 11,84%)
Способы:
Полноразмерные не избыточные аминокислотные последовательности из N1 США от вирусов H1N1 и H3N1 (всего около 1550 основных) и N2 США от вирусов H1N2 и H3N2 (всего около 2700 основных), собранные с 2000 года по настоящее время, были загружены из Influenza Research Database. Процент идентичности аминокислот внутри кластеров и между кластерами для данных США N1 и N2 анализировали с использованием версии MEGA 7.0.7 [см., Kumar et al., Bioinformatics 28: 2685-2686 (2012)]. Два выравнивания последовательностей с использованием MAFFT версии 7.294b Katoh et al. Nucleic Acids Res. 30:3059-3066 (2002); Katoh et al., Molecular Biology and Evolution 30:772-780 (2013] и два отдельных филогенетических дерева N1 и N2 с максимальным правдоподобием получали в версии FastTree 2.1.8 [Price et al., PLoS ONE, 5 (3) 2010: e9490]. В зависимости от филогении N1 и N2, N1 и N2 были отнесены к одной из четырех ранее описанных антигенных линий NA, N1классический, N1пандемический, N21998 и N22002 [см., Nelson et al., J Virol. 86 (16): 8872-8878 (2012); Anderson et al., Influenza Other Respir Viruses Suppl 4: 42-51 (2013)]. Конкретные кластеры были помечены, а метрики разнообразия были дополнительно рассчитаны с использованием командной строки MEGA с настройками по умолчанию и расчетом p-расстояния для разнообразия внутри кластера и между кластерами с гамма-распределенной вариацией скорости (альфа 0,5) и частичной делецией, установленной на 95%.
ПРИМЕР 6
ОПРЕДЕЛЕНИЕ ЛИНИй ДЛЯ ЕВРОПЫ
Введение в часть EU-NA :
Подобно Северной Америке, три подтипа вируса гриппа А H1N1, H3N2 и H1N2 одновременно циркулируют среди свиней в европейских популяциях свиней. Однако линии генов HA и NA значительно различаются в пределах каждого подтипа [см., Kuntz-Simon и Madec Zoonoses Public Health., 56 (6-7): 310-325 (2009)]. Гены NA европейских изолятов свиного гриппа принадлежат к четырем основным и двум второстепенным линиям, как установлено Watson et al., [J. Virol., 89: 9920-9931 (2015); DOI: 10,1128/JVI.00840-15]. Основные линии включают:
1. A (H1N1) Пандемическая 2009, подобная N1,
2. Евразийскиая Avian,подобная N1,
3. A/swine/Gent/1/1984, подобная N2 (Gent/84), и
4. A/swine/Scotland/ 410440/1994, подобная N2 (Scot/94).
В отличие от других европейских основных линий, сегменты NA линии Scotland/94 включают четыре основные группы вирусов, циркулирующих в европейской популяции свиней. Малые европейские линии гриппа NA включают:
1. A/Swine/Italy/4675/2003, подобная N2 и
2. Человеческая сезонная, подобная N2.
Описание способа, применяемого для анализа сегментов генов EU NA-N1 и N2:
Аминокислотные последовательности полноразмерного гена NA европейских изолятов свиней, которые собирали в период между 2005 и 2018 годами, были получены из NCBI Influenza Virus Resource [см., Ref.32 Watson et al., J. Virol., 89: 9920-9931 (2015) ); DOI: 10,1128/JVI.00840-15]. Последовательности N1 и N2 были выровнены с помощью выравнивателя Muscle (codon) с настройками по умолчанию, представленными в MEGA X [Kumar et al., Molecular Biology and Evolution 35: 1547-1549 (2018)]. Эволюционная история была выведена с помощью метода максимального правдоподобия, основанного на модели Уилана и Голдмана [Molecular Biology and Evolution 18: 691-699 (2001)]. Способ Bootstrap с 250 повторениями применяли для теста филогении. Дискретное распределение Гамма применяли для моделирования эволюционных различий в скорости среди сайтов [5 категорий (+G, параметр=0,7258)]. Модель изменения скорости позволила некоторым сайтам быть эволюционно неизменными {[+I], 38,08% сайтов}. Анализ включал 244 аминокислотных последовательности. Все положения с охватом сайтов менее 95% были исключены. То есть, для каждого положения допускалось менее 5% пропусков по выравниванию, отсутствующих данных и неоднозначных оснований. Всего в окончательном наборе данных было 469 положений. Эволюционный анализ проводили в MEGA X [Kumar et al., Molecular Biology and Evolution 35: 1547-1549 (2018)]. Изолятам N1 назначали либо Евразийскую Avian-N1-подобную, либо пандемическую 2009-N1-подобную линию, и изолятам N2 назначали Scotland/410440/1994-N2-подобную линию (например, клада 1 или клада 2), Gent/1/1984-N2-подобную линию, Italy/4675/2003-N2-подобную линию, или человеческую сезонную N2-подобную линию, как описано ранее [Watson et al., J. Virol., 89: 9920-9931 (2015)]. Генетическую идентичность внутри и между линиями рассчитывали, используя MEGA X. Затем репрезентативный штамм от каждой из вышеупомянутых линий выбирали кандидатом для вакцины.
Следующие целевые последовательности для соответствующих линий и/или клад имеют полученный диапазон изменчивости:
A/swine/Italy/179057/2015 (H1N1) последовательность с 10% изменчивостью
A/swine/Italy/28762-3/2013 (H1N1) последовательность с 15% изменчивостью
A/swine/Italy/248147-8/2015 (H3N2) последовательность с 10% изменчивостью
A/swine/Italy/129452/2015(H1N2) последовательность с 10% изменчивостью
A/swine/England/61470/2013 (H1N2) последовательность с 10% изменчивостью
A/swine/Italy/246087/2014 (H1N2) последовательность с 10% изменчивостью.
Настоящее изобретение не должно быть ограничено по объему конкретными вариантами осуществления, описываемыми в настоящем документе. Фактически, различные модификации по изобретению в дополнение к этим, описываемым в настоящем документе, станут очевидны специалистам в данной области из приведенного выше описания. Такие модификации предназначены для включения в объем прилагаемой формулы изобретения.
Кроме того, следует понимать, что все базовые размеры или размеры аминокислот, а также молекулярные массы или значения молекулярной массы, приведенные для нуклеиновых кислот или полипептидов, являются приблизительными и приведены для описания.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Intervet, Inc.
Intervet International, BV
Mogler, Mark A.
Kitikoon, Pravina
Puttamreddy, Supraja
Strait, Erin
Segers, Ruud A.M.
Nagaraj, Basav
<120> ВАКЦИНА ПРОТИВ ВИРУСА СВИНОГО ГРИППА А
<130> 24554WO
<150> US 62/607,101
<151> 2017-12-18
<160> 27
<170> PatentIn version 3.5
<210> 1
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 1
atgaatacta atcaaaggat cattaccatt gggacagtct gcatgattgt cggtatcatc 60
tctcttttgc ttcagattgg aaacattgtc tcactttgga ttagccattc aattcagacc 120
ggatgggaga atcacactga gatgtgcaat caaagtgtca ttacttatgt caataacact 180
tgggtgaacc ggacttatgt gaacattagc aatatcaaga ttgcaactat tcaagatgtg 240
actagtatca ttttggccgg aaattctagt ctttgcccgg tgtcagggtg ggctgtctac 300
agcaaagaca atagcattag gattggatca aaaggggaca ttttcgtcat tagagagcct 360
ttcatctcat gctcacaatt ggagtgccgg accttctttc tgacccaagg ggcattgctg 420
aatgacaaac attcaaatgg taccgtcaag gacaggagtc cttatagaac cctgatgagc 480
tgccctatcg gtgaggcccc ttcgccatac aactcacggt tcgaatctgt cgcatggtca 540
gcatctgctt gtcatgatgg aatgggatgg cttacaatcg ggatcagtgg accggataat 600
ggtgctgtcg cagtcttgaa atacaacgga atcattacag atacaatcaa atcttggagg 660
aacaagattc ttagaactca agaatcagag tgtgtctgta tgaacggatc atgttttaca 720
gtcttgacag atggcccaag caatggacaa gcctcttaca aaatctttaa ggtggaaaaa 780
ggaaagatta tcaagtcgat tgagctggat gcccccaatt accactatga agaatgctct 840
tgttatccag atactggcaa agtcatgtgt gtctgccggg acaattggca cgcctcaaac 900
cggccatggg tgtcgttcaa tcagaatctt gactatcaaa ttggatacat ttgctctgga 960
gtctttggtg ataaccctag atccaatgat gggaagggca attgtggccc ggtcctttct 1020
aatggagcaa atggagtgaa aggtttctca tatcggtatg gaaatggtgt gtggattggt 1080
cggaccaagt caatcaactc tcggtcgggt tttgagatga tttgggatcc gaatggatgg 1140
actgagacag attcatcatt ctcgatgaag caggacatta tcgctcttaa tgattggtct 1200
ggatactcgg gatcttttgt ccaacatccg gagcttactg gtatgaattg cattaggcct 1260
tgtttctggg tggaattgat cagagggcaa cccaaggaaa gcactatctg ggctagcggt 1320
tccagcatct cattctgtgg cgtcaattcg gaaaccgctt cctggtcttg gccagacgga 1380
gctgatctgc cattcaccat tgacaag 1407
<210> 2
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 2
Met Asn Thr Asn Gln Arg Ile Ile Thr Ile Gly Thr Val Cys Met Ile
1 5 10 15
Val Gly Ile Ile Ser Leu Leu Leu Gln Ile Gly Asn Ile Val Ser Leu
20 25 30
Trp Ile Ser His Ser Ile Gln Thr Gly Trp Glu Asn His Thr Glu Met
35 40 45
Cys Asn Gln Ser Val Ile Thr Tyr Val Asn Asn Thr Trp Val Asn Arg
50 55 60
Thr Tyr Val Asn Ile Ser Asn Ile Lys Ile Ala Thr Ile Gln Asp Val
65 70 75 80
Thr Ser Ile Ile Leu Ala Gly Asn Ser Ser Leu Cys Pro Val Ser Gly
85 90 95
Trp Ala Val Tyr Ser Lys Asp Asn Ser Ile Arg Ile Gly Ser Lys Gly
100 105 110
Asp Ile Phe Val Ile Arg Glu Pro Phe Ile Ser Cys Ser Gln Leu Glu
115 120 125
Cys Arg Thr Phe Phe Leu Thr Gln Gly Ala Leu Leu Asn Asp Lys His
130 135 140
Ser Asn Gly Thr Val Lys Asp Arg Ser Pro Tyr Arg Thr Leu Met Ser
145 150 155 160
Cys Pro Ile Gly Glu Ala Pro Ser Pro Tyr Asn Ser Arg Phe Glu Ser
165 170 175
Val Ala Trp Ser Ala Ser Ala Cys His Asp Gly Met Gly Trp Leu Thr
180 185 190
Ile Gly Ile Ser Gly Pro Asp Asn Gly Ala Val Ala Val Leu Lys Tyr
195 200 205
Asn Gly Ile Ile Thr Asp Thr Ile Lys Ser Trp Arg Asn Lys Ile Leu
210 215 220
Arg Thr Gln Glu Ser Glu Cys Val Cys Met Asn Gly Ser Cys Phe Thr
225 230 235 240
Val Leu Thr Asp Gly Pro Ser Asn Gly Gln Ala Ser Tyr Lys Ile Phe
245 250 255
Lys Val Glu Lys Gly Lys Ile Ile Lys Ser Ile Glu Leu Asp Ala Pro
260 265 270
Asn Tyr His Tyr Glu Glu Cys Ser Cys Tyr Pro Asp Thr Gly Lys Val
275 280 285
Met Cys Val Cys Arg Asp Asn Trp His Ala Ser Asn Arg Pro Trp Val
290 295 300
Ser Phe Asn Gln Asn Leu Asp Tyr Gln Ile Gly Tyr Ile Cys Ser Gly
305 310 315 320
Val Phe Gly Asp Asn Pro Arg Ser Asn Asp Gly Lys Gly Asn Cys Gly
325 330 335
Pro Val Leu Ser Asn Gly Ala Asn Gly Val Lys Gly Phe Ser Tyr Arg
340 345 350
Tyr Gly Asn Gly Val Trp Ile Gly Arg Thr Lys Ser Ile Asn Ser Arg
355 360 365
Ser Gly Phe Glu Met Ile Trp Asp Pro Asn Gly Trp Thr Glu Thr Asp
370 375 380
Ser Ser Phe Ser Met Lys Gln Asp Ile Ile Ala Leu Asn Asp Trp Ser
385 390 395 400
Gly Tyr Ser Gly Ser Phe Val Gln His Pro Glu Leu Thr Gly Met Asn
405 410 415
Cys Ile Arg Pro Cys Phe Trp Val Glu Leu Ile Arg Gly Gln Pro Lys
420 425 430
Glu Ser Thr Ile Trp Ala Ser Gly Ser Ser Ile Ser Phe Cys Gly Val
435 440 445
Asn Ser Glu Thr Ala Ser Trp Ser Trp Pro Asp Gly Ala Asp Leu Pro
450 455 460
Phe Thr Ile Asp Lys
465
<210> 3
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 3
atgaatccta accaaaagat cattaccatt ggttcggtct gtatgacaat tggaatggct 60
aacctgatcc ttcaaattgg aaacattatc tcaatctggg tcagccactc aattcaaatt 120
ggaaatcaat cgcagattga aacatgcaac caaagcgtca ttacttacga aaacaacact 180
tgggtgaacc agacctacgt gaacatcagc aacaccaact tcgctgctgg acagtccgtg 240
gtttccgtga aactggcggg caactcctct ctctgccctg tgagcggatg ggctatctac 300
tccaaagaca actcagtcag aatcggttcc aagggggatg tgtttgtcat aagggaacca 360
ttcatctcat gctctcccct tgaatgcaga accttcttct tgactcaagg ggccttgcta 420
aatgacaaac attccaacgg aaccattaaa gacaggagcc catatcggac cctgatgagc 480
tgtcctatcg gtgaagtccc ctcgccatac aactcaagat ttgagtcagt cgcttggtca 540
gcatccgctt gtcatgatgg catcaattgg ctcaccattg gaatttctgg cccagacagt 600
ggggcagtgg ctgtgctgaa gtacaatggc attataacag acactatcaa gtcgtggagg 660
aacaacatat tgagaactca agagtctgaa tgtgcatgtg tgaatggttc ttgctttacc 720
atcatgaccg atggaccatc cgatggacag gcctcctaca agatcttcag aatcgaaaag 780
ggaaagatcg tcaaatcagt cgaaatgaat gcccctaact accactatga ggaatgctcc 840
tgttatcctg attcctccga aatcacgtgc gtgtgcaggg ataactggca tggctccaat 900
cggccgtggg tgtctttcaa ccagaatctg gaatatcaga tcggatacat ttgctccggg 960
gtgttcggag acaatccgcg ccctaatgat aagacaggct cgtgtggtcc agtctcgtct 1020
aacggagcca acggagtcaa aggattttca ttcaaatacg gcaatggagt gtggataggg 1080
agaactaaga gcatttcctc aagaaaaggt ttcgagatga tttgggatcc gaatggatgg 1140
actgggactg acaacaagtt ctcaatcaag caagacatcg tgggaatcag cgagtggtca 1200
ggatatagcg ggtcctttgt gcagcacccc gaactgaccg ggctggattg tattagacct 1260
tgcttctggg tcgaactcat cagagggcga cccgaagaga acacaatctg gactagcggg 1320
agcagcatct ccttttgtgg tgtgaactcg gacactgtgg gttggtcttg gccagacggt 1380
gctgagttgc cttttaccat tgacaag 1407
<210> 4
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 4
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Cys Met Thr
1 5 10 15
Ile Gly Met Ala Asn Leu Ile Leu Gln Ile Gly Asn Ile Ile Ser Ile
20 25 30
Trp Val Ser His Ser Ile Gln Ile Gly Asn Gln Ser Gln Ile Glu Thr
35 40 45
Cys Asn Gln Ser Val Ile Thr Tyr Glu Asn Asn Thr Trp Val Asn Gln
50 55 60
Thr Tyr Val Asn Ile Ser Asn Thr Asn Phe Ala Ala Gly Gln Ser Val
65 70 75 80
Val Ser Val Lys Leu Ala Gly Asn Ser Ser Leu Cys Pro Val Ser Gly
85 90 95
Trp Ala Ile Tyr Ser Lys Asp Asn Ser Val Arg Ile Gly Ser Lys Gly
100 105 110
Asp Val Phe Val Ile Arg Glu Pro Phe Ile Ser Cys Ser Pro Leu Glu
115 120 125
Cys Arg Thr Phe Phe Leu Thr Gln Gly Ala Leu Leu Asn Asp Lys His
130 135 140
Ser Asn Gly Thr Ile Lys Asp Arg Ser Pro Tyr Arg Thr Leu Met Ser
145 150 155 160
Cys Pro Ile Gly Glu Val Pro Ser Pro Tyr Asn Ser Arg Phe Glu Ser
165 170 175
Val Ala Trp Ser Ala Ser Ala Cys His Asp Gly Ile Asn Trp Leu Thr
180 185 190
Ile Gly Ile Ser Gly Pro Asp Ser Gly Ala Val Ala Val Leu Lys Tyr
195 200 205
Asn Gly Ile Ile Thr Asp Thr Ile Lys Ser Trp Arg Asn Asn Ile Leu
210 215 220
Arg Thr Gln Glu Ser Glu Cys Ala Cys Val Asn Gly Ser Cys Phe Thr
225 230 235 240
Ile Met Thr Asp Gly Pro Ser Asp Gly Gln Ala Ser Tyr Lys Ile Phe
245 250 255
Arg Ile Glu Lys Gly Lys Ile Val Lys Ser Val Glu Met Asn Ala Pro
260 265 270
Asn Tyr His Tyr Glu Glu Cys Ser Cys Tyr Pro Asp Ser Ser Glu Ile
275 280 285
Thr Cys Val Cys Arg Asp Asn Trp His Gly Ser Asn Arg Pro Trp Val
290 295 300
Ser Phe Asn Gln Asn Leu Glu Tyr Gln Ile Gly Tyr Ile Cys Ser Gly
305 310 315 320
Val Phe Gly Asp Asn Pro Arg Pro Asn Asp Lys Thr Gly Ser Cys Gly
325 330 335
Pro Val Ser Ser Asn Gly Ala Asn Gly Val Lys Gly Phe Ser Phe Lys
340 345 350
Tyr Gly Asn Gly Val Trp Ile Gly Arg Thr Lys Ser Ile Ser Ser Arg
355 360 365
Lys Gly Phe Glu Met Ile Trp Asp Pro Asn Gly Trp Thr Gly Thr Asp
370 375 380
Asn Lys Phe Ser Ile Lys Gln Asp Ile Val Gly Ile Ser Glu Trp Ser
385 390 395 400
Gly Tyr Ser Gly Ser Phe Val Gln His Pro Glu Leu Thr Gly Leu Asp
405 410 415
Cys Ile Arg Pro Cys Phe Trp Val Glu Leu Ile Arg Gly Arg Pro Glu
420 425 430
Glu Asn Thr Ile Trp Thr Ser Gly Ser Ser Ile Ser Phe Cys Gly Val
435 440 445
Asn Ser Asp Thr Val Gly Trp Ser Trp Pro Asp Gly Ala Glu Leu Pro
450 455 460
Phe Thr Ile Asp Lys
465
<210> 5
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 5
atgaatccaa accaaaagat aatcacaatt ggctctgttt ctctcactat tgccacaatg 60
tgcctcctta tgcaaattgc catcctgatt actaatgtca cattgcactc caatcagtac 120
gaatgcaact accccccaaa caaccaagtg atactgtgtg aaccaactat catcaaaaga 180
aacattactg agattgtgta tctggccaac accaccatag agaaggaaat ctgccccaag 240
ctggcagaat acagaaactg gtcgaagccg caatgtaaaa ttacagggtt tgcacctttt 300
tccaaggaca attcgattag gctttctgcg ggtggcgaca tttgggtaac gagagaacct 360
tatgtgtcat gcgatcctga taagtgttac cagtttgccc ttggacaagg aacaacgctc 420
aacaacagac attcaaacga caccgtgcat gataggaccc cttatcgaac cctattgatg 480
aatgagttgg gtattccatt ccatttgggg accaaacaag tgtgcatcgc atggtccagc 540
tcatcctgcc atgatggacg ggcttggctt catgtctgta ttactgggca tgacaacaat 600
gcaactgcca gcatcattta caatggacgc cttgtcgata gtattggttc atggtccaaa 660
agaatcctca ggacccagga gtcggaatgc gtgtgcatca atggaacttg taccgtggtc 720
atgactgatg ggtccgcttc aggaatagct gacactaaaa tcctgttcat tgaagagggg 780
aaaatcgtgc acattagccc actgctgggg tccgctcagc acgtggagga gtgctcctgc 840
tatccccgat acccaggtgt cagatgcatc tgtagagaca actggaaagg ctccaacaga 900
cctgtcgtgg atattaatgt gaaggattat agcattgtgt cctcctacgt gtgctccgga 960
ctggtgggag acacccccag aaaagacgac agatccagct ccagcaattg tctgaatcct 1020
aacaacgaga aaggggagca tggagtgaaa ggctgggcct ttgatgatgg aaatgacgtg 1080
tggatgggga ggacaatcaa cgagacatta cgctcaggtt atgaaacctt caaagtcatt 1140
gaaggctggt ccaaacctaa ttccaaattg cagataaatc gccaagtcat tgttgaaaga 1200
gatgataggt ccggttattc tggaattttc tctgtcgaag gaaagagctg tatcaatcgg 1260
tgtttttacg tggagctgat cagaggaagg aaacaggaaa ctgcagtgtg gtggacgtca 1320
aattccattg tggtgttttg tggcacctca ggtacctatg gaaccggctc atggcctgat 1380
ggggcggaca tcaatctcat gcctgtg 1407
<210> 6
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 6
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Thr
1 5 10 15
Ile Ala Thr Met Cys Leu Leu Met Gln Ile Ala Ile Leu Ile Thr Asn
20 25 30
Val Thr Leu His Ser Asn Gln Tyr Glu Cys Asn Tyr Pro Pro Asn Asn
35 40 45
Gln Val Ile Leu Cys Glu Pro Thr Ile Ile Lys Arg Asn Ile Thr Glu
50 55 60
Ile Val Tyr Leu Ala Asn Thr Thr Ile Glu Lys Glu Ile Cys Pro Lys
65 70 75 80
Leu Ala Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Lys Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Asp Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Asp Pro Asp Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asn Asn Arg His
130 135 140
Ser Asn Asp Thr Val His Asp Arg Thr Pro Tyr Arg Thr Leu Leu Met
145 150 155 160
Asn Glu Leu Gly Ile Pro Phe His Leu Gly Thr Lys Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys His Asp Gly Arg Ala Trp Leu His Val
180 185 190
Cys Ile Thr Gly His Asp Asn Asn Ala Thr Ala Ser Ile Ile Tyr Asn
195 200 205
Gly Arg Leu Val Asp Ser Ile Gly Ser Trp Ser Lys Arg Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asn Gly Thr Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Ile Ala Asp Thr Lys Ile Leu Phe
245 250 255
Ile Glu Glu Gly Lys Ile Val His Ile Ser Pro Leu Leu Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Gly Val Arg
275 280 285
Cys Ile Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Val Val Asp
290 295 300
Ile Asn Val Lys Asp Tyr Ser Ile Val Ser Ser Tyr Val Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Lys Asp Asp Arg Ser Ser Ser Ser Asn
325 330 335
Cys Leu Asn Pro Asn Asn Glu Lys Gly Glu His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Val Trp Met Gly Arg Thr Ile Asn Glu
355 360 365
Thr Leu Arg Ser Gly Tyr Glu Thr Phe Lys Val Ile Glu Gly Trp Ser
370 375 380
Lys Pro Asn Ser Lys Leu Gln Ile Asn Arg Gln Val Ile Val Glu Arg
385 390 395 400
Asp Asp Arg Ser Gly Tyr Ser Gly Ile Phe Ser Val Glu Gly Lys Ser
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Lys Gln
420 425 430
Glu Thr Ala Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Thr Gly Ser Trp Pro Asp Gly Ala Asp Ile
450 455 460
Asn Leu Met Pro Val
465
<210> 7
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 7
atgaatccaa atcaaaagat cattactatt ggatcagtct cactcatcat tgccacaatt 60
tgtttcctta tgcaaattgc aatccttgtc actactgtca cattgcattt caagcagcat 120
gactacaact cccccccaaa caaccaagct actctgtgtg aaccaacaat cattgaacgg 180
aaaacaactg aaattgtgta tcttactaac accaccattg agaaagaagt ctgccccaaa 240
cttgcagagt accggaactg gtcaaagcct caatgtaaca ttactggatt tgcaccattt 300
tcgaaagaca attctattcg gttgtctgct ggtggggaca tctgggtgac tagggaacct 360
tatgtgtcat gcgatcctga caagtgttac caatttgccc ttggacaggg tacaactctt 420
aacaacggac attcgaataa cacagtccat gataggaccc cgtatcggac ccttcttatg 480
aatgagcttg gtgtcccttt tcatcttgga accagacaag tgtgcatggc ttggtctagc 540
tcatcttgtc acgatgggaa agcatggctg catgtctgtg tcactggaaa tgataacaat 600
gctactgcta gcttcatcta caatggtagg cttgtggatt ctattggttc gtggtcgaaa 660
aacattctcc ggacccaaga gtcagaatgc gtctgtatca atggaacatg tactgtcgtc 720
atgactgatg gatccgctag tggaaaagca gataccaaaa tcttgttcgt cgaagagggg 780
aagatcgtcc atatcagcac tctgttggga tctgcacagc acgtcgagga atgctcctgt 840
tatcctaggt ttccgggagt ccggtgtgtc tgccgggaca actggaaagg atctaataga 900
cccatcgtcg acatcaatgt caagaattac agcattgtct cttcgtatgt ctgcagtgga 960
cttgtcggtg atactcccag agagagcgac tcagtctcct catcttattg cttggatccg 1020
aacaatgaga agggtggtca tggggtgaaa gggtgggcct ttgatgatgg taatgacgtg 1080
tggatgggaa gaacaatcaa cgagactttg cgcttgggat atgaaacctt caaagtcatt 1140
gaaggctggt ccacagctaa ctccaagtca cagacaaata gacaagtgat tgtcgaaaaa 1200
ggagacaggt caggatattc tgggattttc tcagtcgagg gaaagaactg catcaatagg 1260
tgcttctatg tggagttgat tagaggacgg aaagaggaga caaaagtctg gtggaccagt 1320
aactcaattg tcgtgttttg tggcacctca gggacttatg gtactggctc ttggccggat 1380
ggtgctgaca tcaatctcat gccaatt 1407
<210> 8
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 8
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Ile
1 5 10 15
Ile Ala Thr Ile Cys Phe Leu Met Gln Ile Ala Ile Leu Val Thr Thr
20 25 30
Val Thr Leu His Phe Lys Gln His Asp Tyr Asn Ser Pro Pro Asn Asn
35 40 45
Gln Ala Thr Leu Cys Glu Pro Thr Ile Ile Glu Arg Lys Thr Thr Glu
50 55 60
Ile Val Tyr Leu Thr Asn Thr Thr Ile Glu Lys Glu Val Cys Pro Lys
65 70 75 80
Leu Ala Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Asn Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Asp Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Asp Pro Asp Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asn Asn Gly His
130 135 140
Ser Asn Asn Thr Val His Asp Arg Thr Pro Tyr Arg Thr Leu Leu Met
145 150 155 160
Asn Glu Leu Gly Val Pro Phe His Leu Gly Thr Arg Gln Val Cys Met
165 170 175
Ala Trp Ser Ser Ser Ser Cys His Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Val Thr Gly Asn Asp Asn Asn Ala Thr Ala Ser Phe Ile Tyr Asn
195 200 205
Gly Arg Leu Val Asp Ser Ile Gly Ser Trp Ser Lys Asn Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asn Gly Thr Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Lys Ala Asp Thr Lys Ile Leu Phe
245 250 255
Val Glu Glu Gly Lys Ile Val His Ile Ser Thr Leu Leu Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Phe Pro Gly Val Arg
275 280 285
Cys Val Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Ile Val Asp
290 295 300
Ile Asn Val Lys Asn Tyr Ser Ile Val Ser Ser Tyr Val Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Glu Ser Asp Ser Val Ser Ser Ser Tyr
325 330 335
Cys Leu Asp Pro Asn Asn Glu Lys Gly Gly His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Val Trp Met Gly Arg Thr Ile Asn Glu
355 360 365
Thr Leu Arg Leu Gly Tyr Glu Thr Phe Lys Val Ile Glu Gly Trp Ser
370 375 380
Thr Ala Asn Ser Lys Ser Gln Thr Asn Arg Gln Val Ile Val Glu Lys
385 390 395 400
Gly Asp Arg Ser Gly Tyr Ser Gly Ile Phe Ser Val Glu Gly Lys Asn
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Lys Glu
420 425 430
Glu Thr Lys Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Thr Gly Ser Trp Pro Asp Gly Ala Asp Ile
450 455 460
Asn Leu Met Pro Ile
465
<210> 9
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' фланкирующая последовательность
<400> 9
ggcgcgccgc acc 13
<210> 10
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' фланкирующая последовательность
<400> 10
ttaattaa 8
<210> 11
<211> 1410
<212> ДНК
<213> вирус свиного гриппа
<400> 11
atgaatccaa accaaaagat aataaccatt ggttcggtct gtatgacaat tggaatggct 60
aacttaatat tacaaattgg aaacataatc tcaatatgga ttagccactc aattcaactt 120
gggaatcaaa gtcagattga aacatgcaat caaagcgtca ttacttatga aaacaacact 180
tgggtaaatc agacatatgt taacatcagc aacaccaact ttgctgctgg acagtcagtg 240
gtttccgcga aattagcggg caattcctcc ctctgccctg ttagtggatg ggctatatac 300
agtaaagaca acagtgtaag aatcggttcc aagggggatg tgtttgtcat aagggaacca 360
ttcatatcat gctccccctt agaatgcaga accttcttct tgactcaagg ggccttgcta 420
aatgacaaac attccaatgg aaccattaaa gataggagcc catatcgaac cctgatgagc 480
tgtcctattg gtgaagttcc ctctccatac aactcaagat ttgagtcggt cgcttggtca 540
gcaagtgctt gtcacgatgg catcaattgg ctaacaatcg gaatttctgg cccagacagt 600
ggggcagtgg ctgtattaaa gtacaatggc ataataacag acactatcaa gagttggaaa 660
aacaatatat tgagaacaca agagtctgaa tgtgcatgtg taaatggttc ttgctttacc 720
ataatgaccg atggaccaag tgatggacag gcctcataca agatcttcag aatagaaaag 780
ggaaagatag tcaaatcagt cgaaatgaat gcccctaatt atcactatga ggaatgctcc 840
tgttatcctg attctagtga aatcacatgt gtgtgcaggg ataactggca tggctcgaat 900
cgaccgtggg tgtctttcaa ccagaatctg gaatatcaga taggatacat atgcagtggg 960
attttcggag acaatccacg ccctaatgat aagacaggca gttgtggtcc agtatcgtct 1020
aatggagcaa atggagtaaa aggattttca ttcaaatatg gcaatggtgt ttggataggg 1080
agaactaaaa gcattagttc aagaaaaggt tttgagatga tttgggatcc aaatggatgg 1140
actgggacag acaaaaactt ctcaataaag caagatatca taggaataaa tgagtggtca 1200
ggatacagcg ggagttttgt tcagcatcca gaactaacag ggctgaattg tataagacct 1260
tgcttctggg ttgaactaat cagagggcga cccaaagaga acacaatctg gactagcggg 1320
agcagcatat ccttttgtgg tgtaaacagt gacactgtgg gttggtcttg gccagacggt 1380
gctgagttgc catttaccat tgacaagtaa 1410
<210> 12
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 12
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Cys Met Thr
1 5 10 15
Ile Gly Met Ala Asn Leu Ile Leu Gln Ile Gly Asn Ile Ile Ser Ile
20 25 30
Trp Ile Ser His Ser Ile Gln Leu Gly Asn Gln Ser Gln Ile Glu Thr
35 40 45
Cys Asn Gln Ser Val Ile Thr Tyr Glu Asn Asn Thr Trp Val Asn Gln
50 55 60
Thr Tyr Val Asn Ile Ser Asn Thr Asn Phe Ala Ala Gly Gln Ser Val
65 70 75 80
Val Ser Ala Lys Leu Ala Gly Asn Ser Ser Leu Cys Pro Val Ser Gly
85 90 95
Trp Ala Ile Tyr Ser Lys Asp Asn Ser Val Arg Ile Gly Ser Lys Gly
100 105 110
Asp Val Phe Val Ile Arg Glu Pro Phe Ile Ser Cys Ser Pro Leu Glu
115 120 125
Cys Arg Thr Phe Phe Leu Thr Gln Gly Ala Leu Leu Asn Asp Lys His
130 135 140
Ser Asn Gly Thr Ile Lys Asp Arg Ser Pro Tyr Arg Thr Leu Met Ser
145 150 155 160
Cys Pro Ile Gly Glu Val Pro Ser Pro Tyr Asn Ser Arg Phe Glu Ser
165 170 175
Val Ala Trp Ser Ala Ser Ala Cys His Asp Gly Ile Asn Trp Leu Thr
180 185 190
Ile Gly Ile Ser Gly Pro Asp Ser Gly Ala Val Ala Val Leu Lys Tyr
195 200 205
Asn Gly Ile Ile Thr Asp Thr Ile Lys Ser Trp Lys Asn Asn Ile Leu
210 215 220
Arg Thr Gln Glu Ser Glu Cys Ala Cys Val Asn Gly Ser Cys Phe Thr
225 230 235 240
Ile Met Thr Asp Gly Pro Ser Asp Gly Gln Ala Ser Tyr Lys Ile Phe
245 250 255
Arg Ile Glu Lys Gly Lys Ile Val Lys Ser Val Glu Met Asn Ala Pro
260 265 270
Asn Tyr His Tyr Glu Glu Cys Ser Cys Tyr Pro Asp Ser Ser Glu Ile
275 280 285
Thr Cys Val Cys Arg Asp Asn Trp His Gly Ser Asn Arg Pro Trp Val
290 295 300
Ser Phe Asn Gln Asn Leu Glu Tyr Gln Ile Gly Tyr Ile Cys Ser Gly
305 310 315 320
Ile Phe Gly Asp Asn Pro Arg Pro Asn Asp Lys Thr Gly Ser Cys Gly
325 330 335
Pro Val Ser Ser Asn Gly Ala Asn Gly Val Lys Gly Phe Ser Phe Lys
340 345 350
Tyr Gly Asn Gly Val Trp Ile Gly Arg Thr Lys Ser Ile Ser Ser Arg
355 360 365
Lys Gly Phe Glu Met Ile Trp Asp Pro Asn Gly Trp Thr Gly Thr Asp
370 375 380
Lys Asn Phe Ser Ile Lys Gln Asp Ile Ile Gly Ile Asn Glu Trp Ser
385 390 395 400
Gly Tyr Ser Gly Ser Phe Val Gln His Pro Glu Leu Thr Gly Leu Asn
405 410 415
Cys Ile Arg Pro Cys Phe Trp Val Glu Leu Ile Arg Gly Arg Pro Lys
420 425 430
Glu Asn Thr Ile Trp Thr Ser Gly Ser Ser Ile Ser Phe Cys Gly Val
435 440 445
Asn Ser Asp Thr Val Gly Trp Ser Trp Pro Asp Gly Ala Glu Leu Pro
450 455 460
Phe Thr Ile Asp Lys
465
<210> 13
<211> 1408
<212> ДНК
<213> вирус свиного гриппа
<400> 13
atgaacccaa atcagaagat aataatcatt agttcaatct gtatgacaaa tggaattgct 60
agcttgatat tacaaattgg gaacataata tcaatatgga ttagccattc aattcaaatt 120
gagaacccaa accagaccga ccatgcaatc aaagcgttat tatttacgaa aacaacacat 180
gggtaaatca aacgtatgtt aacatcagca acaataattt tgttgttgaa cagacagtgg 240
tttcaatgaa attagcgggc agttcttctc tctgccctgt tagtggatgg gctatataca 300
gtaaagataa cagtgtaaga atcggttcca aaggggatgt gtttgtcata agagagccat 360
tcatctcatg ctcccatttg gaatgtagaa ccttcttctt aactcaaggg gccctactga 420
atgataaaca ttctaatgga accgttaaag acagaagccc ctatcgaacc ctgatgagct 480
gtcctattgg tgaagtcccc tctccataca actcaaaatt tgagtcagtt gcttggtcag 540
caagtgcttg ccatgatggc accagttggt tgacaattgg gatttctggt ccagacaatg 600
gagcagtggc tgtgttgaaa tacaatgaca taataacaga cactatcaag agttggaaaa 660
acaacatatt gagaacacaa gaatctgaat gtgcatgttt gaatggttct tgctttactg 720
taatgaccga tggaccaagt aatgggcagg cctcatacaa gatcttcaaa atagaaaagg 780
ggaaagtagt caaatcagtc gagttgaatg ctcctaatta tcactatgag gaatgttcct 840
gttatcctga ttctggtgaa atcatatgtg tatgcaggga caattggcat ggctcgaatc 900
gaccatgggt gtctttcaat cagaatctgg agtatcagat aggatacata tgcagtgggg 960
ttctcggaga caatccgcgc cctaatgata gaacaggcag ttgtggtcca gtatcatctc 1020
atggagcaaa tggggtaaaa gggttttcgt ttaaatacgg caatggaatt tggataggga 1080
gaactaaaag cactattaca aggagtggtt ttgagatgat ttgggaccca aacggatgga 1140
ctggaacaga caataatttc tcagtgaagc aagatatcgt aggaataact aactggtcag 1200
gatacgcggg agttttgtcc aacatccaga attaaccgga ttggattgta ttagaccttg 1260
cttctgggtt gaactaatca gagggagacc caaagagaac acaatctgga ctagcggaag 1320
cagcatatcc ttttgtggtg taaatagtga cactgtgggt tggtcttggc cagacggtgc 1380
tgagttgcca tttaccattg acaagtaa 1408
<210> 14
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 14
Met Asn Pro Asn Gln Lys Ile Ile Ile Ile Ser Ser Ile Cys Met Thr
1 5 10 15
Asn Gly Ile Ala Ser Leu Ile Leu Gln Ile Gly Asn Ile Ile Ser Ile
20 25 30
Trp Ile Ser His Ser Ile Gln Ile Glu Asn Pro Asn Gln Thr Glu Pro
35 40 45
Cys Asn Gln Ser Val Ile Ile Tyr Glu Asn Asn Thr Trp Val Asn Gln
50 55 60
Thr Tyr Val Asn Ile Ser Asn Asn Asn Phe Val Val Glu Gln Thr Val
65 70 75 80
Val Ser Met Lys Leu Ala Gly Ser Ser Ser Leu Cys Pro Val Ser Gly
85 90 95
Trp Ala Ile Tyr Ser Lys Asp Asn Ser Val Arg Ile Gly Ser Lys Gly
100 105 110
Asp Val Phe Val Ile Arg Glu Pro Phe Ile Ser Cys Ser His Leu Glu
115 120 125
Cys Arg Thr Phe Phe Leu Thr Gln Gly Ala Leu Leu Asn Asp Lys His
130 135 140
Ser Asn Gly Thr Val Lys Asp Arg Ser Pro Tyr Arg Thr Leu Met Ser
145 150 155 160
Cys Pro Ile Gly Glu Val Pro Ser Pro Tyr Asn Ser Lys Phe Glu Ser
165 170 175
Val Ala Trp Ser Ala Ser Ala Cys His Asp Gly Thr Ser Trp Leu Thr
180 185 190
Ile Gly Ile Ser Gly Pro Asp Asn Gly Ala Val Ala Val Leu Lys Tyr
195 200 205
Asn Asp Ile Ile Thr Asp Thr Ile Lys Ser Trp Lys Asn Asn Ile Leu
210 215 220
Arg Thr Gln Glu Ser Glu Cys Ala Cys Leu Asn Gly Ser Cys Phe Thr
225 230 235 240
Val Met Thr Asp Gly Pro Ser Asn Gly Gln Ala Ser Tyr Lys Ile Phe
245 250 255
Lys Ile Glu Lys Gly Lys Val Val Lys Ser Val Glu Leu Asn Ala Pro
260 265 270
Asn Tyr His Tyr Glu Glu Cys Ser Cys Tyr Pro Asp Ser Gly Glu Ile
275 280 285
Ile Cys Val Cys Arg Asp Asn Trp His Gly Ser Asn Arg Pro Trp Val
290 295 300
Ser Phe Asn Gln Asn Leu Glu Tyr Gln Ile Gly Tyr Ile Cys Ser Gly
305 310 315 320
Val Leu Gly Asp Asn Pro Arg Pro Asn Asp Arg Thr Gly Ser Cys Gly
325 330 335
Pro Val Ser Ser His Gly Ala Asn Gly Val Lys Gly Phe Ser Phe Lys
340 345 350
Tyr Gly Asn Gly Ile Trp Ile Gly Arg Thr Lys Ser Thr Ile Thr Arg
355 360 365
Ser Gly Phe Glu Met Ile Trp Asp Pro Asn Gly Trp Thr Gly Thr Asp
370 375 380
Asn Asn Phe Ser Val Lys Gln Asp Ile Val Gly Ile Thr Asn Trp Ser
385 390 395 400
Gly Tyr Ser Gly Ser Phe Val Gln His Pro Glu Leu Thr Gly Leu Asp
405 410 415
Cys Ile Arg Pro Cys Phe Trp Val Glu Leu Ile Arg Gly Arg Pro Lys
420 425 430
Glu Asn Thr Ile Trp Thr Ser Gly Ser Ser Ile Ser Phe Cys Gly Val
435 440 445
Asn Ser Asp Thr Val Gly Trp Ser Trp Pro Asp Gly Ala Glu Leu Pro
450 455 460
Phe Thr Ile Asp Lys
465
<210> 15
<211> 1410
<212> ДНК
<213> вирус свиного гриппа
<400> 15
atgaatccaa atcaaaagat aataacaatt ggttctgttt ctctcactat tacaacaatg 60
tgcctcttct tgcagattgc catcctagta actactataa cattgcattt caagcaatat 120
gaatgcgatt cccctgcaaa caaccaagta ataccgtgtg aaccaataat aatagaaaaa 180
aacataacaa aaatagtgta tttgaccaat accaccatag agaaagaggt atgcccaaaa 240
ttaggggaat acaggaattg gtcaaaacca caatgcaaga tcacaggatt tgcacctttt 300
tctaaggaca attcaattcg gctctctgcg ggtggggcca tttgggtcac gagagaacct 360
tatgtgtcat gcgaccctaa caagtgttat caatttgcat taggacaggg aaccacatta 420
gataacagac attcaaatga cacaatacat gatagaaccc cttttagaac cctgttgatg 480
agtgaattag gtgttccatt tcatttggga accagacaag tatgcatagc atggtccagt 540
tcaagttgtc acgatgggaa agcttggttg catgtttgtg tcactgggca tgataaaaat 600
gcaactgcta gtttcattta tgacggaaag cttgtagaca gcatcagttc atggtccaaa 660
aacatactcc ggactcagga atcagaatgt gtttgtatcg atggaatctg tacagtggtg 720
atgactgatg gaagtgcttc agggaaagct gatactaaga tactatttat tgaaaaaggg 780
aagatcattc atattagtcc attgttggga agtgctcagc atgtagaaga atgttcctgt 840
taccctagat accctgatgt caggtgtatt tgcagggata actggaaagg ttcaaatagg 900
cccatcgtag acataagaat gaaaaattat agcattggtt ccagttatat gtgctcagga 960
cttgttggcg acacacccag gaacaatgat gggtctagta atagcaattg tcggaatccc 1020
aataatgaaa gaggaaatca tggagtgaaa ggttgggcct ttgatgatgg aaatgacaca 1080
tggatgggaa gaactatcag caaggactca cgcttaggtt acgaaacctt caaagttgtt 1140
ggtggttggt cccaacccaa ttccaaatcc cagataaata gacaagttat tgttgacagc 1200
gataatagat caggttactc tggtattttc tctgttgagg ggaaagattg cattaatagg 1260
tgtttttatg tggaactaat aagaggaagg agacaggaaa ctagagtgtg gtggacttcg 1320
aacagtattg ttgtgttctg tggcacttct ggcacctatg ggtcaggctc atggcccgat 1380
ggagcaaaca tcaatttcat gcctgtataa 1410
<210> 16
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 16
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Thr
1 5 10 15
Ile Thr Thr Met Cys Leu Phe Leu Gln Ile Ala Ile Leu Val Thr Thr
20 25 30
Ile Thr Leu His Phe Lys Gln Tyr Glu Cys Asp Ser Pro Ala Asn Asn
35 40 45
Gln Val Ile Pro Cys Glu Pro Ile Ile Ile Glu Lys Asn Ile Thr Lys
50 55 60
Ile Val Tyr Leu Thr Asn Thr Thr Ile Glu Lys Glu Val Cys Pro Lys
65 70 75 80
Leu Gly Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Lys Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Ala Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Asp Pro Asn Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asp Asn Arg His
130 135 140
Ser Asn Asp Thr Ile His Asp Arg Thr Pro Phe Arg Thr Leu Leu Met
145 150 155 160
Ser Glu Leu Gly Val Pro Phe His Leu Gly Thr Arg Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys His Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Val Thr Gly His Asp Lys Asn Ala Thr Ala Ser Phe Ile Tyr Asp
195 200 205
Gly Lys Leu Val Asp Ser Ile Ser Ser Trp Ser Lys Asn Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asp Gly Ile Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Lys Ala Asp Thr Lys Ile Leu Phe
245 250 255
Ile Glu Lys Gly Lys Ile Ile His Ile Ser Pro Leu Leu Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Asp Val Arg
275 280 285
Cys Ile Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Ile Val Asp
290 295 300
Ile Arg Met Lys Asn Tyr Ser Ile Gly Ser Ser Tyr Met Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Asn Asn Asp Gly Ser Ser Asn Ser Asn
325 330 335
Cys Arg Asn Pro Asn Asn Glu Arg Gly Asn His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Thr Trp Met Gly Arg Thr Ile Ser Lys
355 360 365
Asp Ser Arg Leu Gly Tyr Glu Thr Phe Lys Val Val Gly Gly Trp Ser
370 375 380
Gln Pro Asn Ser Lys Ser Gln Ile Asn Arg Gln Val Ile Val Asp Ser
385 390 395 400
Asp Asn Arg Ser Gly Tyr Ser Gly Ile Phe Ser Val Glu Gly Lys Asp
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Arg Gln
420 425 430
Glu Thr Arg Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Ser Gly Ser Trp Pro Asp Gly Ala Asn Ile
450 455 460
Asn Phe Met Pro Val
465
<210> 17
<211> 1410
<212> ДНК
<213> вирус свиного гриппа
<400> 17
atgaattcaa atcaaaagat aataacaatt ggctctgttt ctctcactat tgccacacta 60
tgcctcctta tgcaaattgc tatcatggta actactgtaa catttcattt caagcagtat 120
gaatacaact cccccccgaa caaccaagta atgttgtgtg aaccaacgat aattgaaaga 180
aacataacag agacagtgta catgaccaac accaccatag tgaaagaaat atgccccaaa 240
ctagcggaat acagaaattg gtcaaaaccg caatgcaaaa ttacaggatt tgcacctttt 300
tcaaaggaca actcaattcg gctttccgct ggtggggaca tctgggtgac aagagaacct 360
tatgtgtcat gcgatcctaa caagtgttat caatttgccc ttgggcaggg aacaacgtta 420
aacaacaggc attcaaatga cacagtacat gatagaaccc cttaccgaac cctgttgatg 480
aatgaattgg gtgttccatt tcatttagga accaagcagg tttgcatagc ttggtccagt 540
tcaagttgtc atgatggaaa agcatggttg catgtttgtg taactgggca tgatgaaaat 600
gcaactgcca gtttcattta caacgagaga cttgtagata gtattggttc atggtccaag 660
aaaatcctca gaacccagga gtcggaatgc gtttgcataa atgggacttg tacagtggtg 720
atgacagatg ggagtgcttc aggtagagct gatactaaaa tactattcat tgaggagggg 780
aaaatcgttc atgttagcca actgacagga agtgctcagc atgtagagga gtgctcctgt 840
tatccccggt atcctggtgt cagatgtgtt tgcagagata attggaaagg ctccaatagg 900
cccattgtag atataaatgt aaaggatcat agcattgttt ccagttatgt gtgctcagga 960
cttgtcggag acacacccag aaaaaacgac agctctagca gtagtaactg cctgaatcct 1020
aacaatgaag aagggggtca tggggtgaaa ggctgggcct ttgatgatga aaatgacttg 1080
tggatgggaa gaacgatcag cgaaaagtta cgattaggtt atgaaacctt caaggtcatt 1140
gaaggctggt ccaagcctaa ttccaaattg cagataaata ggcaagtaat agttgacaaa 1200
gataatagat ccggttattc tggtattttc tctgttgaaa gtaaaagttg catcaatcgg 1260
tgcttttatg tggagttgat aagaggaagg aaacaggaaa atgaagtatg gtggacctca 1320
aacagcattg ttgtattttg tggcacctca ggtacatatg gaacaggctc atggcctgat 1380
ggggcagaca tcaatctcat gcctatatga 1410
<210> 18
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 18
Met Asn Ser Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Thr
1 5 10 15
Ile Ala Thr Leu Cys Leu Leu Met Gln Ile Ala Ile Met Val Thr Thr
20 25 30
Val Thr Phe His Phe Lys Gln Tyr Glu Tyr Asn Ser Pro Pro Asn Asn
35 40 45
Gln Val Met Leu Cys Glu Pro Thr Ile Ile Glu Arg Asn Ile Thr Glu
50 55 60
Thr Val Tyr Met Thr Asn Thr Thr Ile Val Lys Glu Ile Cys Pro Lys
65 70 75 80
Leu Ala Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Lys Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Asp Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Asp Pro Asn Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asn Asn Arg His
130 135 140
Ser Asn Asp Thr Val His Asp Arg Thr Pro Tyr Arg Thr Leu Leu Met
145 150 155 160
Asn Glu Leu Gly Val Pro Phe His Leu Gly Thr Lys Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys His Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Val Thr Gly His Asp Glu Asn Ala Thr Ala Ser Phe Ile Tyr Asn
195 200 205
Glu Arg Leu Val Asp Ser Ile Gly Ser Trp Ser Lys Lys Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asn Gly Thr Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Arg Ala Asp Thr Lys Ile Leu Phe
245 250 255
Ile Glu Glu Gly Lys Ile Val His Val Ser Gln Leu Thr Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Gly Val Arg
275 280 285
Cys Val Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Ile Val Asp
290 295 300
Ile Asn Val Lys Asp His Ser Ile Val Ser Ser Tyr Val Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Lys Asn Asp Ser Ser Ser Ser Ser Asn
325 330 335
Cys Leu Asn Pro Asn Asn Glu Glu Gly Gly His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Glu Asn Asp Leu Trp Met Gly Arg Thr Ile Ser Glu
355 360 365
Lys Leu Arg Leu Gly Tyr Glu Thr Phe Lys Val Ile Glu Gly Trp Ser
370 375 380
Lys Pro Asn Ser Lys Leu Gln Ile Asn Arg Gln Val Ile Val Asp Lys
385 390 395 400
Asp Asn Arg Ser Gly Tyr Ser Gly Ile Phe Ser Val Glu Ser Lys Ser
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Lys Gln
420 425 430
Glu Asn Glu Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Thr Gly Ser Trp Pro Asp Gly Ala Asp Ile
450 455 460
Asn Leu Met Pro Ile
465
<210> 19
<211> 1410
<212> ДНК
<213> вирус свиного гриппа
<400> 19
atgaatccaa atcagaagat aataacaatt ggctctgttt ctctcgtcat tgcaacatta 60
tgcttcttaa tgcagatggc catcctaata actactgtaa aattacattt caaacaatat 120
gagtgcggct tccctgcgaa caaccaagta ataacatgtg agccaacagt aatagaaagg 180
aacacaacag agatagtgta cttaactaac accaccatag agaaagaaac atgccacaaa 240
acagtggaat acaggaattg gtcaaagcct caatgcaaaa taacaggctt tgcacctttt 300
tccaaggaca attcaattcg actttctgct ggtggggaca tatgggtgac gagggaacct 360
tacgtgtcat gcgagcctgg caaatgttat cagtttgcac tcgggcaagg gaccacacta 420
gacaataaac attcaaacga tacaatacat gacagaaccc cctatcgaac tctattgatg 480
aatgaattgg gtgtcccatt tcatttaggg acaagacaag tgtgtattgc atggtccagc 540
tcaagttgtt atgatgggaa agcatggttg catgtctgta tcactggaca tgataaaaat 600
gcaactgcca gtttcattta cgatggtaga cttgtagata gcattggttc atggtctaaa 660
aatatactta gaacccagga atcagaatgc gtttgcatca atggggtctg tacagtagta 720
atgactgatg gaagtgcttc gggaagagct gatactaaaa tactattcat tgaagaaggg 780
aaaattgttc atattagccc attagcgggg agtgcacagc atgtggagga gtgctcctgt 840
tatccccgat atcctggcgt aaggtgtatc tgcagagaca actggaaagg ctctaacaga 900
cccgttgtgg atataaatat agaagattat agcattgatt ccagttatgt gtgttcaggg 960
cttgttggcg acacacccag aatcaatgac ggatccagta gtagctactg ccgtgatcct 1020
aacaacgaaa aaggaaatca cggagtgaag ggctgggctt ttgacgatgg aaatgatgtg 1080
tggatgggaa gaacgatcaa cgaagattca cgctcaggtt atgaaacatt caaagtcatt 1140
ggtggttggt ccactcctaa ttccaaattg cagataaata ggcaagtaat agttgatagc 1200
aacaataggt caggttattc tggtgttttc tccgttgaag gcaaaagctg catcaataga 1260
tgtttctacg tggagttgat aagaggaaga aggtcagaag cgcgagtatg gtggacctca 1320
aacagtattg ttgtattttg tggcacttca ggtacctatg gaacaggctc atggcctgat 1380
ggagcagaca tcaacctcat gcctatatga 1410
<210> 20
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 20
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Val
1 5 10 15
Ile Ala Thr Leu Cys Phe Leu Met Gln Met Ala Ile Leu Ile Thr Thr
20 25 30
Val Lys Leu His Phe Lys Gln Tyr Glu Cys Gly Phe Pro Ala Asn Asn
35 40 45
Gln Val Ile Thr Cys Glu Pro Thr Val Ile Glu Arg Asn Thr Thr Glu
50 55 60
Ile Val Tyr Leu Thr Asn Thr Thr Ile Glu Lys Glu Thr Cys His Lys
65 70 75 80
Thr Val Glu Tyr Arg Asn Trp Ser Lys Pro Gln Cys Lys Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Asp Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Glu Pro Gly Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asp Asn Lys His
130 135 140
Ser Asn Asp Thr Ile His Asp Arg Thr Pro Tyr Arg Thr Leu Leu Met
145 150 155 160
Asn Glu Leu Gly Val Pro Phe His Leu Gly Thr Arg Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys Tyr Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Ile Thr Gly His Asp Lys Asn Ala Thr Ala Ser Phe Ile Tyr Asp
195 200 205
Gly Arg Leu Val Asp Ser Ile Gly Ser Trp Ser Lys Asn Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asn Gly Val Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Arg Ala Asp Thr Lys Ile Leu Phe
245 250 255
Ile Glu Glu Gly Lys Ile Val His Ile Ser Pro Leu Ala Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Gly Val Arg
275 280 285
Cys Ile Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Val Val Asp
290 295 300
Ile Asn Ile Glu Asp Tyr Ser Ile Asp Ser Ser Tyr Val Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Ile Asn Asp Gly Ser Ser Ser Ser Tyr
325 330 335
Cys Arg Asp Pro Asn Asn Glu Lys Gly Asn His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Val Trp Met Gly Arg Thr Ile Asn Glu
355 360 365
Asp Ser Arg Ser Gly Tyr Glu Thr Phe Lys Val Ile Gly Gly Trp Ser
370 375 380
Thr Pro Asn Ser Lys Leu Gln Ile Asn Arg Gln Val Ile Val Asp Ser
385 390 395 400
Asn Asn Arg Ser Gly Tyr Ser Gly Val Phe Ser Val Glu Gly Lys Ser
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Arg Ser
420 425 430
Glu Ala Arg Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Thr Gly Ser Trp Pro Asp Gly Ala Asp Ile
450 455 460
Asn Leu Met Pro Ile
465
<210> 21
<211> 1410
<212> ДНК
<213> вирус свиного гриппа
<400> 21
atgaatccaa accagaaaat aataacgatc ggttctgtct ccttgatcat tgcaacaatg 60
tgctttttca tgcaagttgc cattctggta actactgtaa cattgcattt caggcagtgc 120
gaatgcaact cctccgcaac caaccaaata atgccatgta aaccaacaaa aatagagaga 180
aacataactg aaattgtgta cttaaccaat accaccataa aaacagaggt atgccccaaa 240
ctagtgaaat acagggattg ggcaaaacca caatgtagaa tcacagggtt tgcacctttt 300
tccaaggaca attcgattcg gctttctgcc ggtggggcca tttgggtaac gagagaaccc 360
tatgtatcat gcgatcttag caagtgttac cagtttgcgc tcggacaggg gactacacta 420
gacaacagac attcaaatga cacaatacat gatagaactc cttatcggac cctattgatg 480
aatgaattgg gtgttccatt tcatttagga accaggcaag tgtgtatagc ttggtccagt 540
tcaagttgtc acgatggaaa agcatggctg catgtttgtg tcactgggta tgataaaaat 600
gctactgcta gcctcattta tgacggaagg cttgtggaca gcatcggttc atggtcccaa 660
aacatcctcc ggacccagga atcggaatgt gtttgtataa atggtacttg cacagtggta 720
atgactgatg ggagtgcttc aggaaaagct gataccagaa tactatttat tgaagaaggg 780
aagattattc acattagtcc attgacagga agtgcacagc atgttgaaga gtgttcttgt 840
tatcctcgat accccggtgt aagatgtgtt tgtagagaca actggaaggg ctctaacaga 900
cccgtcgtgg atataaatgt aaaagattat aaaattaact ccagttatgt atgctcaggc 960
cttgttggcg atacacccag aaacaacgat agatctagca atagcaactg ccaaaatcct 1020
aacaaccaga gagggaatca tggagtgaag ggctgggcct ttgacgatgg aaatgacata 1080
tggatgggaa gaaccatcag caatgattca cgtttaggtt atgaaacttt caaagttatt 1140
ggtggttggt ccaaacccaa ctccaaagtt cagacaaata ggcaagtcat agttgacagc 1200
gataatagat caggttattc tggcgttttc tctgttgaag gcaaaagctg catcaatagg 1260
tgcttttatg tagagctaat aagaggaagg agacaggaag ctagagtatg gtggacttcg 1320
aacagtattg ttgtgttttg tggtacttcg ggtacatatg gttcaggctc atggcctgat 1380
ggggctgaca tcaatcttat gcctatataa 1410
<210> 22
<211> 469
<212> Белок
<213> вирус свиного гриппа
<400> 22
Met Asn Pro Asn Gln Lys Ile Ile Thr Ile Gly Ser Val Ser Leu Ile
1 5 10 15
Ile Ala Thr Met Cys Phe Phe Met Gln Val Ala Ile Leu Val Thr Thr
20 25 30
Val Thr Leu His Phe Arg Gln Cys Glu Cys Asn Ser Ser Ala Thr Asn
35 40 45
Gln Ile Met Pro Cys Lys Pro Thr Lys Ile Glu Arg Asn Ile Thr Glu
50 55 60
Ile Val Tyr Leu Thr Asn Thr Thr Ile Lys Thr Glu Val Cys Pro Lys
65 70 75 80
Leu Val Lys Tyr Arg Asp Trp Ala Lys Pro Gln Cys Arg Ile Thr Gly
85 90 95
Phe Ala Pro Phe Ser Lys Asp Asn Ser Ile Arg Leu Ser Ala Gly Gly
100 105 110
Ala Ile Trp Val Thr Arg Glu Pro Tyr Val Ser Cys Asp Leu Ser Lys
115 120 125
Cys Tyr Gln Phe Ala Leu Gly Gln Gly Thr Thr Leu Asp Asn Arg His
130 135 140
Ser Asn Asp Thr Ile His Asp Arg Thr Pro Tyr Arg Thr Leu Leu Met
145 150 155 160
Asn Glu Leu Gly Val Pro Phe His Leu Gly Thr Arg Gln Val Cys Ile
165 170 175
Ala Trp Ser Ser Ser Ser Cys His Asp Gly Lys Ala Trp Leu His Val
180 185 190
Cys Val Thr Gly Tyr Asp Lys Asn Ala Thr Ala Ser Leu Ile Tyr Asp
195 200 205
Gly Arg Leu Val Asp Ser Ile Gly Ser Trp Ser Gln Asn Ile Leu Arg
210 215 220
Thr Gln Glu Ser Glu Cys Val Cys Ile Asn Gly Thr Cys Thr Val Val
225 230 235 240
Met Thr Asp Gly Ser Ala Ser Gly Lys Ala Asp Thr Arg Ile Leu Phe
245 250 255
Ile Glu Glu Gly Lys Ile Ile His Ile Ser Pro Leu Thr Gly Ser Ala
260 265 270
Gln His Val Glu Glu Cys Ser Cys Tyr Pro Arg Tyr Pro Gly Val Arg
275 280 285
Cys Val Cys Arg Asp Asn Trp Lys Gly Ser Asn Arg Pro Val Val Asp
290 295 300
Ile Asn Val Lys Asp Tyr Lys Ile Asn Ser Ser Tyr Val Cys Ser Gly
305 310 315 320
Leu Val Gly Asp Thr Pro Arg Asn Asn Asp Arg Ser Ser Asn Ser Asn
325 330 335
Cys Gln Asn Pro Asn Asn Gln Arg Gly Asn His Gly Val Lys Gly Trp
340 345 350
Ala Phe Asp Asp Gly Asn Asp Ile Trp Met Gly Arg Thr Ile Ser Asn
355 360 365
Asp Ser Arg Leu Gly Tyr Glu Thr Phe Lys Val Ile Gly Gly Trp Ser
370 375 380
Lys Pro Asn Ser Lys Val Gln Thr Asn Arg Gln Val Ile Val Asp Ser
385 390 395 400
Asp Asn Arg Ser Gly Tyr Ser Gly Val Phe Ser Val Glu Gly Lys Ser
405 410 415
Cys Ile Asn Arg Cys Phe Tyr Val Glu Leu Ile Arg Gly Arg Arg Gln
420 425 430
Glu Ala Arg Val Trp Trp Thr Ser Asn Ser Ile Val Val Phe Cys Gly
435 440 445
Thr Ser Gly Thr Tyr Gly Ser Gly Ser Trp Pro Asp Gly Ala Asp Ile
450 455 460
Asn Leu Met Pro Ile
465
<210> 23
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 23
atgaacccta accagaagat tattaccatc ggatccgtgt gcatgaccat cggaatggcc 60
aacctgatcc tgcaaatcgg aaacatcatc tccatctgga ttagccatag cattcagctt 120
ggaaaccaga gccagatcga gacttgtaac cagtccgtga tcacctacga gaacaacacc 180
tgggtcaacc agacttacgt caatatcagc aacaccaact tcgctgcggg gcagtccgtg 240
gtgtccgcaa agctggccgg caacagctcc ctgtgccccg tgtccggatg ggcgatctac 300
tccaaggata acagcgtgcg catcgggtcc aaaggggacg tgttcgtcat ccgcgaacca 360
ttcatttctt gctccccctt ggaatgtcgg accttcttcc tgacccaagg ggcgttgctc 420
aacgacaagc acagcaacgg aaccatcaaa gatcggtcgc cctaccgcac tctgatgtcg 480
tgccctatcg gcgaagtgcc atccccctac aactcacgct tcgagtccgt ggcctggtcc 540
gcttccgcct gccacgatgg aatcaactgg ctcacaatcg gcatctccgg cccggactcg 600
ggagccgtgg ccgtgctgaa gtacaatggt attattactg acactatcaa gtcgtggaag 660
aacaatattc tccggactca agaatctgaa tgcgcctgcg tgaacggttc ctgcttcact 720
atcatgaccg acggcccttc cgatggacag gcctcataca agatcttccg gatcgagaag 780
ggaaagatcg tgaagtccgt cgagatgaac gcaccgaact accattatga ggaatgctcg 840
tgctacccgg actcctcgga aattacttgc gtgtgccgcg acaattggca cgggtccaac 900
aggccctggg tgtccttcaa ccaaaacctg gagtaccaga tcggttacat ctgctccggg 960
atttttggag acaaccctag acctaacgac aagaccggct catgcggacc tgtgtcctcc 1020
aacggagcca acggcgtgaa gggattctcg ttcaaatatg ggaacggcgt ctggataggt 1080
cggaccaagt ccatctcgtc acggaagggc tttgaaatga tttgggaccc gaacggttgg 1140
accggaaccg acaagaactt cagcatcaag caggacatta tcggcattaa cgagtggagc 1200
ggatactcgg gcagcttcgt ccagcacccg gaacttacgg gcctcaattg tattaggccc 1260
tgtttttggg tcgagctgat tagagggcgc cccaaggaaa acaccatctg gaccagcggc 1320
tccagcatct cattctgcgg agtgaactcc gacaccgtgg gctggtcgtg gcccgacggt 1380
gccgagctgc cgttcaccat cgataaatga 1410
<210> 24
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 24
atgaatccta accaaaagat catcaccatc ggctccgtca gcctcactat taccaccatg 60
tgcctgttcc ttcaaattgc catcctcgtg accacgatca ccctgcactt caagcagtac 120
gagtgcgact ccccggccaa caaccaggtc atcccgtgtg aaccgattat cattgagaag 180
aacatcacaa agattgtgta cctgactaac accaccatcg agaaggaagt ctgcccaaaa 240
ttgggagagt accggaactg gtccaagcct caatgcaaaa tcactggatt tgctccgttc 300
agcaaggata actcaatcag actgagcgcg ggtggcgcca tctgggtcac cagggaacct 360
tatgtgtcgt gcgatcccaa caagtgctac cagtttgcgc tgggccaggg caccaccctc 420
gacaaccgcc actccaacga caccatccat gatcgcaccc ctttccgaac cctgctgatg 480
agcgagcttg gcgtgccctt ccacctcgga acgcgccaag tctgtattgc ctggagctcg 540
tcctcctgcc acgatggaaa ggcctggctg catgtctgtg tgactggtca cgacaagaac 600
gccaccgcta gcttcatcta tgacggaaaa ttggtggatt ccatctcctc ctggtctaag 660
aacattctgc ggacccagga atccgaatgc gtgtgcatcg atggcatctg cactgtggtc 720
atgactgacg gatccgcaag cggaaaggcg gacactaaga tcctgtttat cgaaaaggga 780
aagatcattc atatctcgcc gctgctgggg tccgcccagc acgtggaaga gtgcagctgt 840
tacccccggt accccgacgt gcggtgcatt tgtcgggaca actggaaggg tagcaacagg 900
cccatcgtgg acattcgcat gaagaattac tcaatcggat cctcctacat gtgctcaggg 960
ctcgtgggag acacccccag gaacaacgac ggttcctcaa actccaactg tcggaacccg 1020
aacaatgagc gcgggaatca cggagtgaag ggatgggcat tcgacgacgg aaacgacacc 1080
tggatgggtc ggactatcag caaggactcc cgcctgggtt acgaaacttt caaggtcgtg 1140
gggggctggt cgcagccaaa ctcgaagtcg cagattaaca gacaggtcat cgtggactcc 1200
gacaataggt ccggctactc cggaatcttc tcggtcgagg ggaaggattg cattaaccgg 1260
tgcttctacg tggagctgat cagaggcaga cgccaggaaa cccgcgtgtg gtggacctcc 1320
aactcaatcg tggtgttctg cggaacatcc ggcacttacg gcagcggatc ctggcctgat 1380
ggcgccaaca taaacttcat gcccgtgtga 1410
<210> 25
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 25
atgaacccca accaaaagat cattacaatc ggatcagtgt cacttgtgat cgccaccttg 60
tgtttcctca tgcaaatggc cattctgatc accactgtga agctgcactt caaacagtat 120
gagtgcggat tcccggcgaa caatcaggtc atcacctgtg aacctaccgt gatcgagcgg 180
aacaccaccg agatagtgta cctgaccaat acgactatcg aaaaggaaac ttgccataag 240
accgtcgagt accgaaactg gtcgaagccg cagtgcaaga tcactgggtt cgcaccgttc 300
agcaaggata acagcatccg gctgtccgcc gggggagaca tttgggtcac ccgcgaaccg 360
tacgtgtcgt gcgagcccgg aaagtgctac cagttcgccc tgggccaggg taccaccctg 420
gataacaagc acagcaacga taccatccac gataggaccc cctaccggac tctgctgatg 480
aacgaactgg gcgtgccttt tcacctcggc acgagacaag tctgcattgc ctggtcctca 540
tcgtcctgct acgacggaaa ggcctggctg cacgtctgca tcaccggcca tgacaaaaac 600
gccaccgcgt ccttcatcta cgacggtcgc ctggtggact ccattggatc ctggagcaag 660
aacattctgc ggacccagga atcggaatgc gtgtgtatca acggagtgtg caccgtcgtg 720
atgactgatg gctctgcttc cggccgggcc gacaccaaga tcttgtttat cgaagaaggg 780
aagattgtcc acatttcccc gcttgctgga tccgcgcagc acgtggaaga gtgcagctgt 840
tacccccgct accctggcgt gcggtgcatt tgtcgggaca actggaaggg atccaatagg 900
cccgtggtgg acatcaacat cgaagattac tcgattgact ccagctacgt gtgctccggc 960
ctcgtcgggg acactccccg catcaacgac gggtcgagct cctcctattg ccgggatcca 1020
aacaacgaga aggggaacca cggcgtgaag ggctgggctt tcgacgacgg aaacgacgtc 1080
tggatgggta gaactatcaa cgaggattcc cggagcggat acgagacttt caaggtcatc 1140
ggtggttggt caacccctaa tagcaaactc cagattaaca gacaggtcat cgtggactcg 1200
aacaatcgga gcggctactc cggagtgttc tccgtggaag gaaagtcgtg catcaaccgg 1260
tgcttctacg tggagctgat taggggtcgc cgcagcgaag cccgcgtgtg gtggacttcc 1320
aactcaatcg tggtgttctg cggcacctcc gggacttacg gaaccggatc ctggccggat 1380
ggcgccgaca ttaacctgat gccaatttga 1410
<210> 26
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 26
atgaacccca accaaaagat cattacaatc ggatcagtgt cacttgtgat cgccaccttg 60
tgtttcctca tgcaaatggc cattctgatc accactgtga agctgcactt caaacagtat 120
gagtgcggat tcccggcgaa caatcaggtc atcacctgtg aacctaccgt gatcgagcgg 180
aacaccaccg agatagtgta cctgaccaat acgactatcg aaaaggaaac ttgccataag 240
accgtcgagt accgaaactg gtcgaagccg cagtgcaaga tcactgggtt cgcaccgttc 300
agcaaggata acagcatccg gctgtccgcc gggggagaca tttgggtcac ccgcgaaccg 360
tacgtgtcgt gcgagcccgg aaagtgctac cagttcgccc tgggccaggg taccaccctg 420
gataacaagc acagcaacga taccatccac gataggaccc cctaccggac tctgctgatg 480
aacgaactgg gcgtgccttt tcacctcggc acgagacaag tctgcattgc ctggtcctca 540
tcgtcctgct acgacggaaa ggcctggctg cacgtctgca tcaccggcca tgacaaaaac 600
gccaccgcgt ccttcatcta cgacggtcgc ctggtggact ccattggatc ctggagcaag 660
aacattctgc ggacccagga atcggaatgc gtgtgtatca acggagtgtg caccgtcgtg 720
atgactgatg gctctgcttc cggccgggcc gacaccaaga tcttgtttat cgaagaaggg 780
aagattgtcc acatttcccc gcttgctgga tccgcgcagc acgtggaaga gtgcagctgt 840
tacccccgct accctggcgt gcggtgcatt tgtcgggaca actggaaggg atccaatagg 900
cccgtggtgg acatcaacat cgaagattac tcgattgact ccagctacgt gtgctccggc 960
ctcgtcgggg acactccccg catcaacgac gggtcgagct cctcctattg ccgggatcca 1020
aacaacgaga aggggaacca cggcgtgaag ggctgggctt tcgacgacgg aaacgacgtc 1080
tggatgggta gaactatcaa cgaggattcc cggagcggat acgagacttt caaggtcatc 1140
ggtggttggt caacccctaa tagcaaactc cagattaaca gacaggtcat cgtggactcg 1200
aacaatcgga gcggctactc cggagtgttc tccgtggaag gaaagtcgtg catcaaccgg 1260
tgcttctacg tggagctgat taggggtcgc cgcagcgaag cccgcgtgtg gtggacttcc 1320
aactcaatcg tggtgttctg cggcacctcc gggacttacg gaaccggatc ctggccggat 1380
ggcgccgaca ttaacctgat gccaatttga 1410
<210> 27
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность вируса свиного гриппа
<400> 27
atgaacccca accagaagat tatcaccatc ggatccgtgt ctctcatcat cgccaccatg 60
tgcttcttta tgcaagtggc cattctcgtc accactgtga ccctgcactt tcggcagtgc 120
gaatgcaact cctccgctac taaccaaatt atgccgtgca agcctactaa gatcgaacgg 180
aacatcaccg agatcgtgta cttgacgaac accaccatca agaccgaagt ctgcccgaag 240
cttgtgaaat atcgggactg ggcgaagcca caatgtcgca tcaccggttt cgccccgttc 300
agcaaagata acagcattcg gctgtccgcc gggggcgcga tctgggtcac cagggagccg 360
tacgtgtcgt gtgatctgtc caagtgctac caattcgcac tgggacaggg taccaccctc 420
gacaaccgcc actccaacga taccatacat gaccggaccc cttaccgcac tctcttgatg 480
aacgagctgg gagtgccctt ccacctgggt accagacaag tctgcattgc ctggtcctca 540
tcctcctgcc atgacgggaa ggcctggctg catgtctgcg tgactggcta cgataagaac 600
gcgacggcct ccctgatcta cgacggtcgc ctggtggact ccattgggtc gtggagccag 660
aatatcctgc ggactcagga gtcagaatgc gtgtgcatca acggcacttg caccgtggtc 720
atgactgacg gatccgcctc ggggaaggcc gacaccagga tcctgttcat tgaggaggga 780
aagattatcc acatctcgcc tctgactgga tccgcccagc acgtggaaga gtgttcctgc 840
taccctcgat accccggcgt gcgctgtgtg tgtcgggata actggaaggg aagcaaccgg 900
cccgtggtgg acattaacgt gaaggactac aagattaact catcatacgt gtgctccgga 960
ctcgtgggcg atacaccaag aaacaacgac cgcagcagca acagcaattg tcagaacccc 1020
aacaaccagc gcggcaacca cggcgtgaag ggctgggcat tcgacgacgg aaatgacatc 1080
tggatgggga ggactatctc caacgattca cgcctgggct acgaaacctt caaggtcatc 1140
ggcggctggt caaaaccgaa ctccaaggtc cagaccaaca gacaggtcat cgtggattcg 1200
gacaatagaa gcggatacag cggagtgttc agcgtggagg gaaagtcgtg catcaaccgc 1260
tgcttctacg tggaactgat caggggtcgg agacaggaag cgcgcgtgtg gtggacttcg 1320
aactcgattg tggtgttctg cggtacctcc ggaacctatg ggagcggatc ctggccggac 1380
ggcgctgaca ttaaccttat gccgatctga 1410
<---
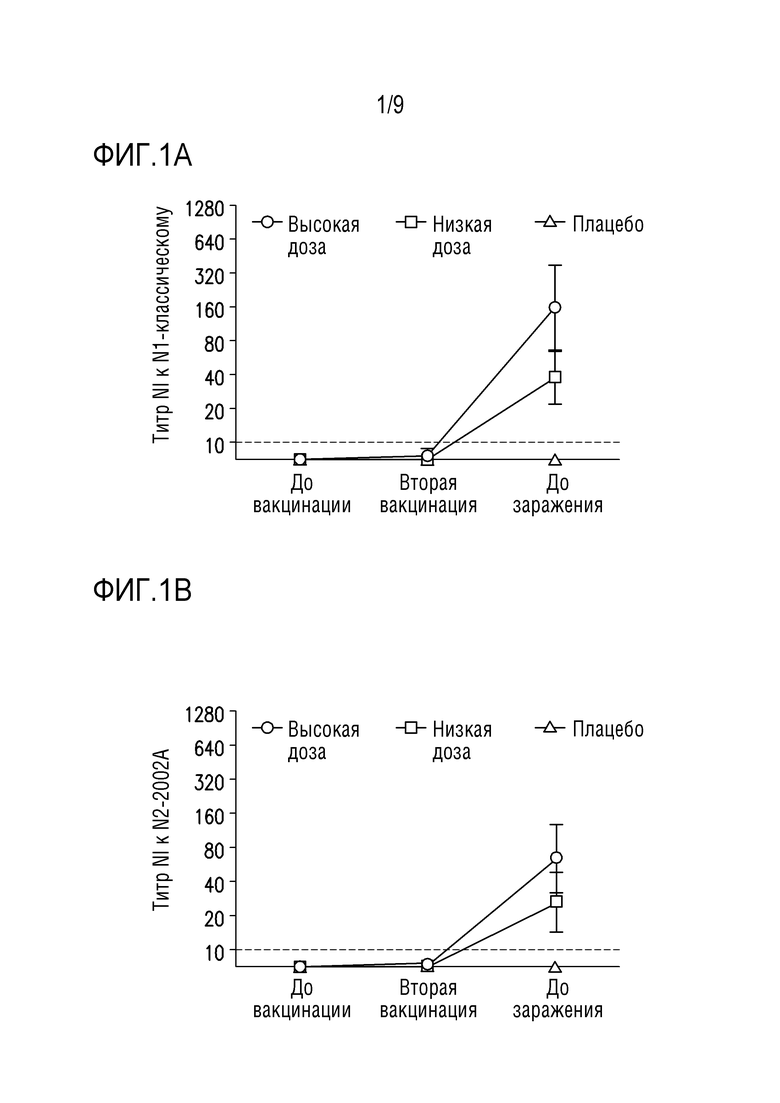
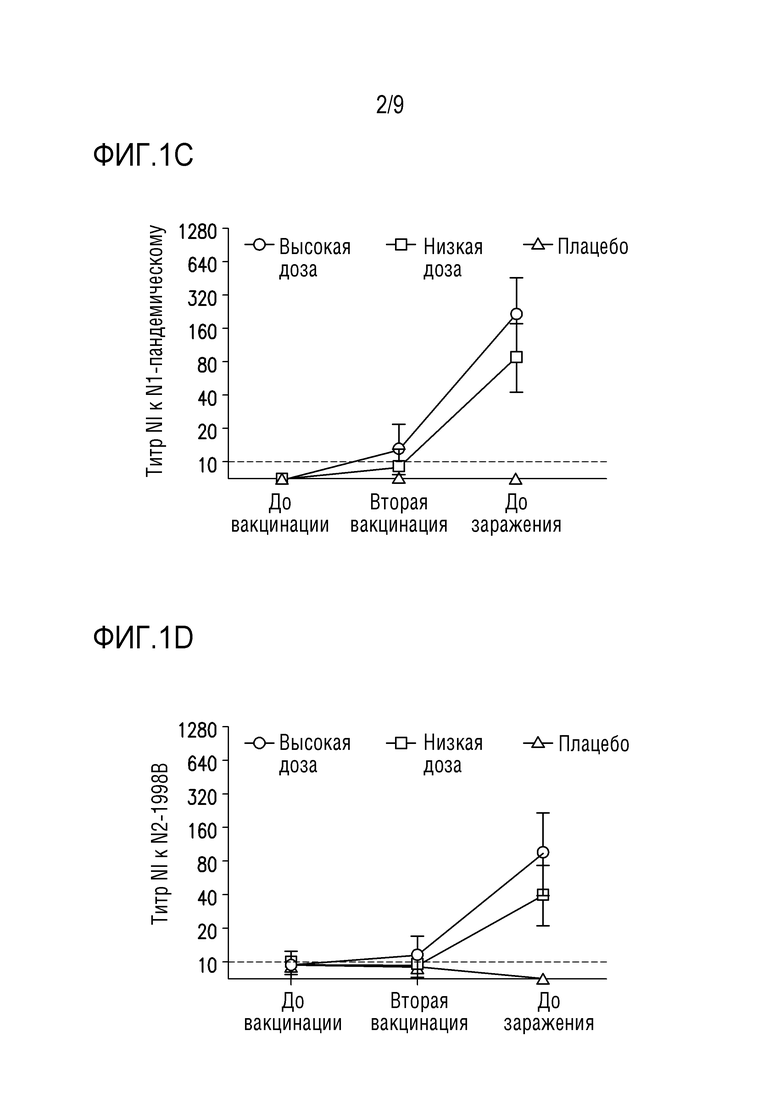
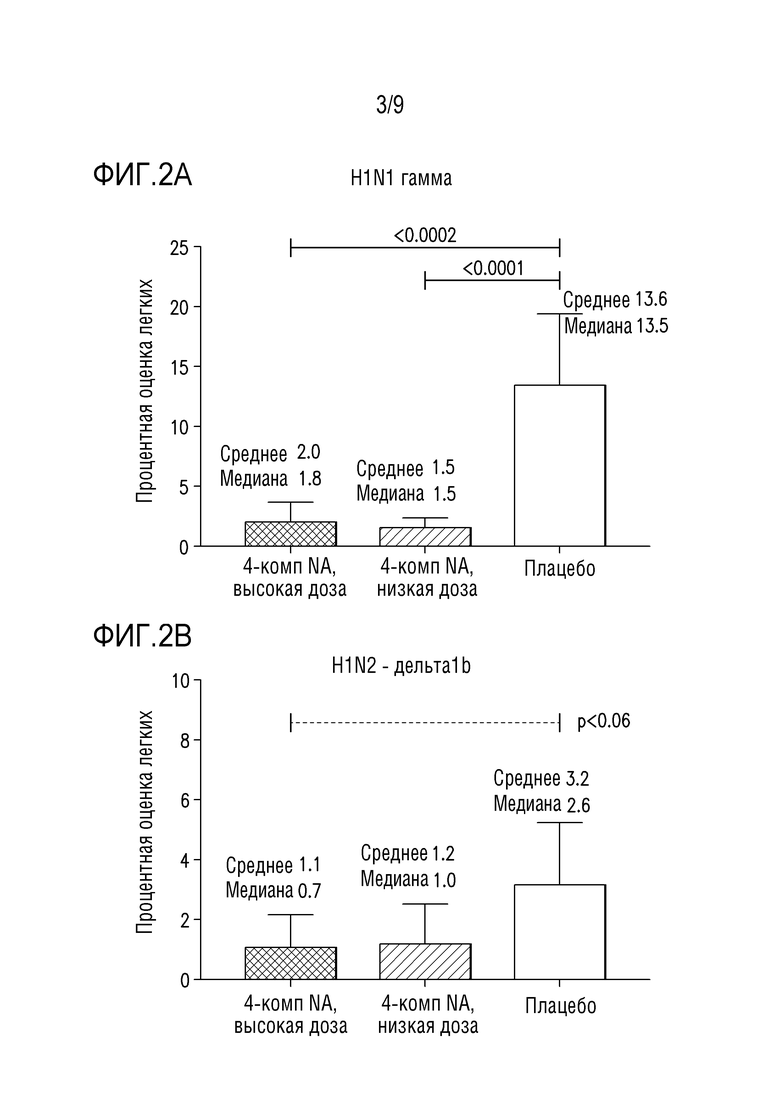

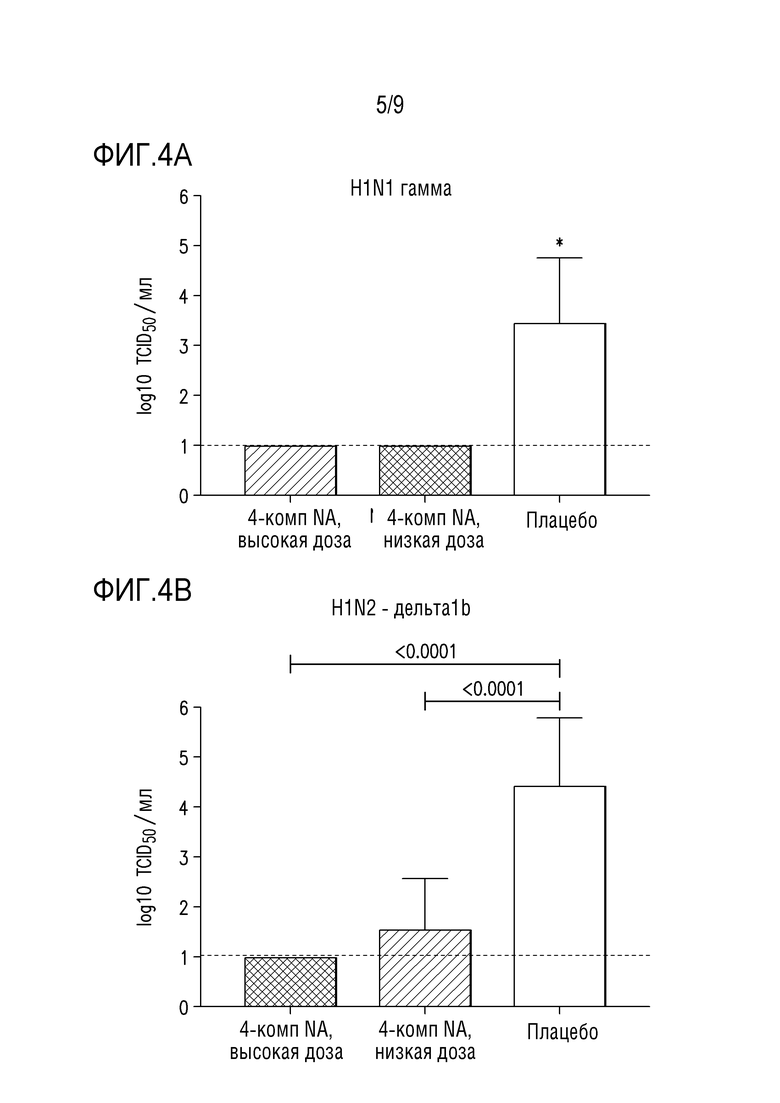
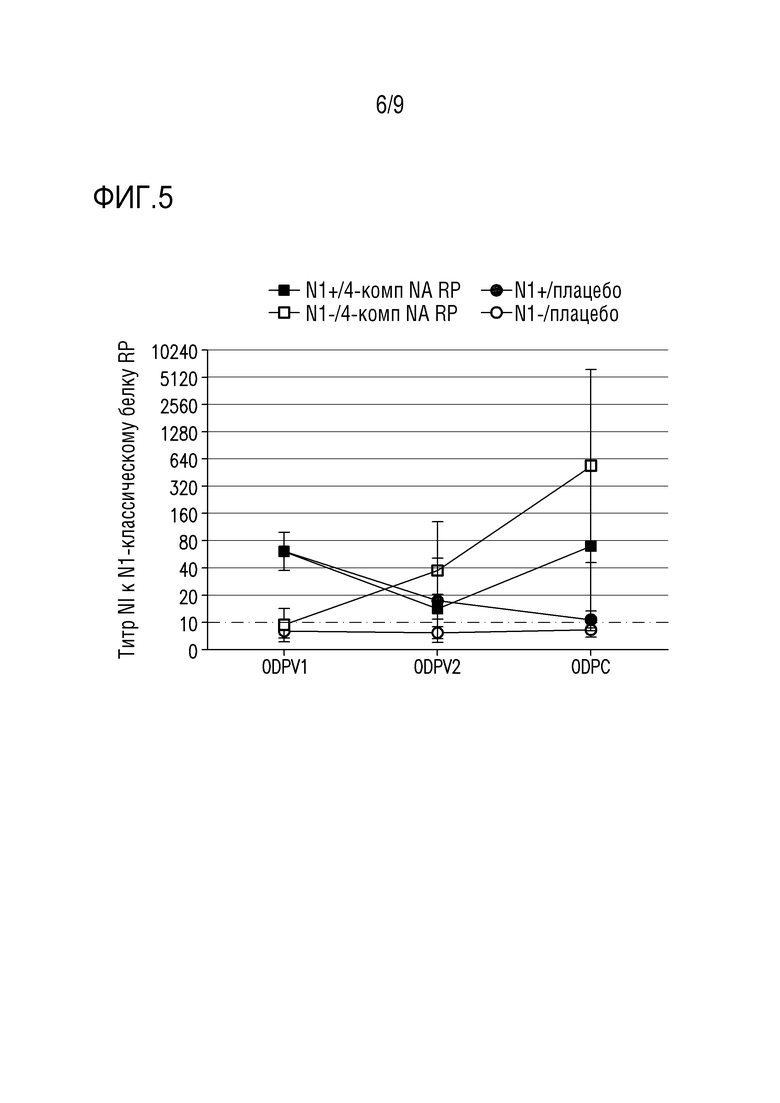

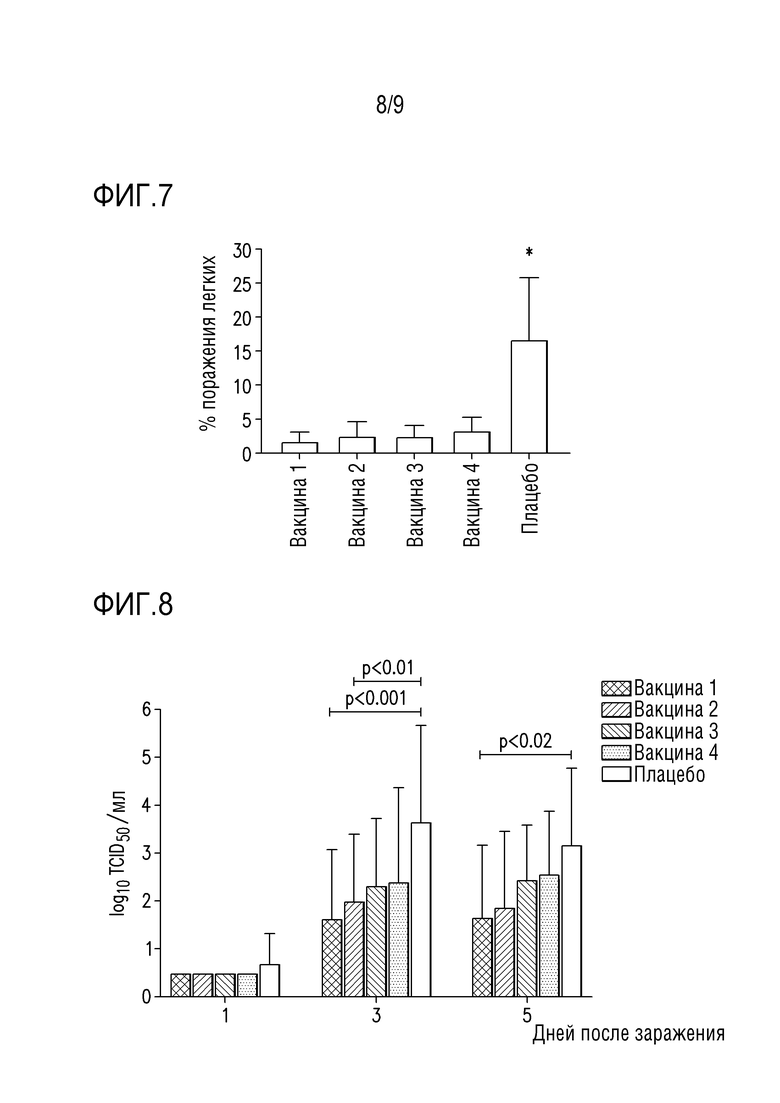
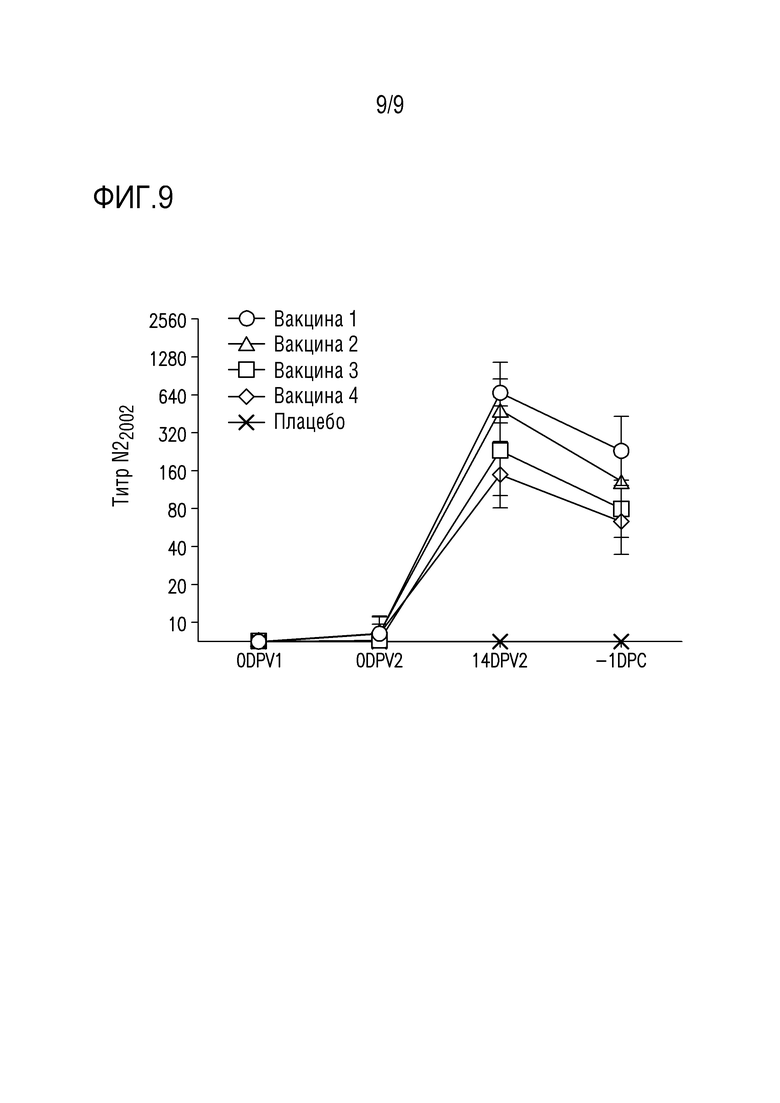
Настоящее изобретение относится к биотехнологии и ветеринарии. Представлена вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, содержащая две или более частицы репликона РНК(RP), каждая из которых независимо кодирует одну или более нейраминидаз (NA), и фармацевтически приемлемый носитель, где первая частица репликона РНК альфавируса кодирует первую нейраминидазу (NA), и вторая частица репликона РНК альфавируса кодирует вторую NA, или где частица репликона РНК альфавируса кодирует две и более NA, при условии, что иммуногенная композиция не содержит ни гемагглютинин (HA) вируса свиного гриппа А (IAV-S), ни нуклеотидную последовательность, кодирующую гемагглютинин (HA) IAV-S; и где первая NA и вторая NA содержат аминокислотные последовательности, которые имеют идентичность 98% или менее. Представлены также способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества заявленной вакцины; применение вакцины в помощи профилактики заболевания, вызываемого вирусом свиного гриппа А, у свиней. Вакцина к IAV-S, содержащая и/или кодирующая антиген нейраминидазы вируса гриппа A в отсутствие соответствующего антигена гемагглютинина, может адекватно защитить свинью от инфекции IAV-S. 3 н. и 13 з.п. ф-лы, 8 табл., 15 ил., 6 пр.
1. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, содержащая две или более частицы репликона РНК(RP), каждая из которых независимо кодирует одну или более нейраминидаз (NA), и фармацевтически приемлемый носитель,
где первая частица репликона РНК альфавируса кодирует первую нейраминидазу (NA) и вторая частица репликона РНК альфавируса кодирует вторую NA, или
где частица репликона РНК альфавируса кодирует две и более NA,
при условии, что иммуногенная композиция не содержит ни гемагглютинин (HA) вируса свиного гриппа А (IAV-S), ни нуклеотидную последовательность, кодирующую гемагглютинин (HA) IAV-S; и где первая NA и вторая NA содержат аминокислотные последовательности, которые имеют идентичность 98% или менее.
2. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.1, где первая NA происходит из IAV-S из первого филогенетического кластера, а вторая NA происходит из IAV-S из второго филогенетического кластера, где первый филогенетический кластер и второй филогенетический кластер различны и где первый филогенетический кластер и второй филогенетический кластер независимо выбраны из группы, состоящей из кластера N1-классического, кластера N1-пандемического, кластера N2-1998 и кластера N2-2002.
3. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.2, где первый филогенетический кластер выбран из группы, состоящей из кластера N1-классического и кластера N1-пандемического, и где второй филогенетический кластер выбран из группы, состоящей из кластера N2-1998 и кластера N2-2002.
4. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.3, где первый филогенетический кластер представляет собой кластер N1-классический, а второй филогенетический кластер представляет собой кластер N2-2002.
5. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по любому из пп.2-4, дополнительно содержащая третью частицу репликона РНК альфавируса, которая кодирует третью NA; где третья NA происходит из IAV-S из третьего филогенетического кластера и где третий филогенетический кластер отличается от первого филогенетического кластера и второго филогенетического кластера.
6. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.5, дополнительно содержащая четвертую частицу репликона РНК альфавируса, которая кодирует четвертую NA, где четвертая NA происходит из IAV-S из четвертого филогенетического кластера и где четвертый филогенетический кластер отличается от первого филогенетического кластера, второго филогенетического кластера и третьего филогенетического кластера.
7. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.6, где первый филогенетический кластер представляет собой кластер N1-классический, второй филогенетический кластер представляет собой кластер N2-2002, третий филогенетический кластер представляет собой кластер N1-пандемический, а четвертый филогенетический кластер представляет собой кластер N2-1998.
8. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.1, где частица репликона РНК альфавируса кодирует две и более NA и где первая NA происходит из IAV-S из первой линии, а вторая NA происходит из IAV-S из второй линии; где первая линия и вторая линия отличаются и где первая линия и вторая линия независимо выбраны из группы, состоящей из линии N1-пандемической (EU), линии N1-Евразийской Avian, линии N2-Gent/1984, линии N2-Italy/4675/2003, линии N2-Scotland /1994 (клада 1) и линии N2- Scotland/1994 (клада 2).
9. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.8, где первая линия выбрана из группы, состоящей из линии N1-пандемической (EU) и линии N1-Евразийской Avian; и где вторая линия выбрана из группы, состоящей из линии N2-Gent/1984, линии N2-Italy/4675/2003, линии N2-Scotland/1994 (клада 1) и линии N2-Scotland/1994 (клада 2).
10. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.2, где частица репликона РНК альфавируса кодирует две или более NA и где первый филогенетический кластер представляет собой кластер N1-классический, а второй филогенетический кластер представляет собой кластер N1-пандемический.
11. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.10, дополнительно содержащая вторую частицу репликона РНК альфавируса, которая кодирует третью NA и четвертую NA; где третья NA происходит из IAV-S из кластера N2-1998, а четвертая NA происходит из IAV-S из кластера N2-2002.
12. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по любому из пп.1-11, где каждая частица репликона РНК альфавируса представляет собой частицу репликона РНК венесуэльского конского энцефалита (VEEV).
13. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по любому из пп.1-12, которая включает адъювант, выбранный из группы, состоящей из биоразлагаемого масла, эмульсии «масло-в-воде» с 2,5-50% (об./об.) минерального масла и биоразлагаемого масла, смешанного с эмульсией «масло-в-воде» с 2,5-50% (об./об.) минерального масла.
14. Вакцина для помощи в профилактике заболевания, вызванного вирусом гриппа, по п.13, где биоразлагаемое масло представляет собой dl-α-токоферил ацетат, а минеральное масло представляет собой парафиновое масло.
15. Способ иммунизации свиньи против вируса свиного гриппа А, включающий введение свинье иммунологически эффективного количества любой из вакцин по пп.1-14.
16. Применение вакцины по любому из пп.1-14 для помощи в профилактике заболевания, вызываемого вирусом свиного гриппа А, у свиней.
| WO 2006078294 A2, 27.07.2006 | |||
| MARTHA I, et al., Evolution of novel resassortant A/H3N2 Influenza viruses in North American Swine and Humans, Journal of virology, vol.86, no.16, 15.08.2012, p.8872-8878, abstract, figure 4, p 8877, column 2, paragraph 3 | |||
| WO 2008033966 A2, 20.03.2008 | |||
| КУЗНЕЦОВ В.И | |||
| и др | |||
| Методические рекомендации по диагностике и |

