Область техники, к которой относится изобретение
Изобретение относится к области сельскохозяйственной биотехнологии, в частности к овцеводству, и может быть использовано для оценки генетического потенциала ягнят на основе исследования генов, влияющих на технологические параметры мяса, с использованием генетических маркеров, влияющих на мясную продуктивность, также изобретение может быть использовано для проведения селекции овец, в раннем возрасте, основанной на определении перспективных генов (соматотропина, кальпастатина и дифференциального фактора роста), влияющих на мясные качества.
Уровень техники
Развитие молекулярно-генетических методов анализа, основанных на полимеразной цепной реакции (ПЦР), позволило разработать новые маркерные системы, обеспечивающие определение генотипов животных непосредственно на уровне генетического материала клетки (на уровне ДНК) независимо от пола и возраста, сократить время анализа и повысить его точность.
Разработка методов более эффективного использования генофонда имеющихся специализированных мясных пород овец с целью повышения уровня и качества мясной продуктивности, генетического контроля и управления селекционным процессом, улучшающих экономические показатели отрасли, является важнейшей задачей на современном этапе развития овцеводства.
Актуальным в решении задач интенсификации овцеводческой отрасли является внедрение современных методов генной диагностики - определение и выявление генов-маркеров хозяйственно ценных признаков. Применение разработанного способа позволит повысить рентабельность овцеводства за счет улучшенных методов производства, хранения, эффективной переработки баранины.
Известен способ по оценке генетического потенциала овец в раннем возрасте, который учитывает живую массу, продуктивность, конституционно-экстерьерные показатели (В.А. Мороз и др. «Методические рекомендации по раннему прогнозированию, отбору и выращиванию высокопродуктивных баранов-производителей тонкорунных и полутонкорунных пород». - Ставрополь, 2001. - 29 с.).
Недостатком данного способа является невысокая точность оценки генетического потенциала овец, так как учитываются только зоотехнические параметры.
Известен способ оценки, прогноза продуктивности сельскохозяйственных животных в раннем возрасте, который основан на учете биохимических, генетических параметров (Л.Н. Чижова и др. «Способ оценки прогноза продуктивности сельскохозяйственных животных в раннем возрасте на основе биохимических тест-систем, генетических маркеров». - Ставрополь, 2010. - 40 с.).
Недостатком данного способа является то, что в качестве критерия оценки продуктивности молодняка используют только биохимические, генетические параметры крови, что не обеспечивает объективность прогноза генетического потенциала молодняка.
Известен способ селекции овец романовской породы по адаптационной способности к промышленной технологии, включающий отбор и разведение "в себе" наиболее приспособленных овец, при этом у овец дополнительно определяют генетическую структуру по локусам арилэстеразы Es 1 и альбумина Al и для воспроизводства отбирают животных-носителей аллелей Es 1B, AlA и AlC (патент №2044482, 1995 г.).
Недостатком данного способа является невысокое качество селекции, так как учитывается только продуктивность новорожденных ягнят на основе особенностей плаценты, а также то, что в качестве критерия оценки продуктивности молодняка используют только полиморфизм белков и ферментов сыворотки крови, что менее информативно по сравнению с полиморфизмом ДНК.
Наиболее близким по технической сущности и достигаемому положительному эффекту и принятый авторами за прототип является способ оценки высокой мясной продуктивности овец сальской породы, включающий выделение ДНК, амплификацию фрагмента гена GH с использованием праймеров 5-GGAGGCAGGAAGGGATGAA-3' и 5'-CCAAGGGAGGGAGAGACAGA-3', рестрикцию амплифицированного фрагмента гена GH эндонуклеазой НаеШ, определение генотипов и отбор животных с генотипом AB/GH (патент №2662679, 2018 г.).
Однако данный метод предусматривает оценку мясной продуктивности овец только по результатам молекулярно-генетических тестов на основании одного белка соматотропина (GH), хотя оценка комбинированных полиморфных вариантов ДНК в генах, ассоциированных с высокой продуктивностью, может быть более информативной.
Раскрытие изобретения
Задачей предлагаемого изобретения является разработка способа оценки генетического потенциала ягнят (провести анализ ассоциативных связей аллельного спектра с показателями роста и убойными качествами), на основе исследования генов-маркеров: GH, CAST и GDF9, влияющих на технологические параметры мяса.
Технический результат, который может быть получен с помощью предлагаемого способа, сводится к возможности проводить прижизненный отбор животных с повышенной мясной продуктивностью, повысить численность популяции специализированных мясных овец, увеличить производство мраморной баранины.
Технический результат достигается с помощью способа оценки генетического потенциала ягнят на основе исследования генов, влияющий амплификацию фрагмента гена с использованием праймеров GH 277 п.н. -F:5'- ggaggcaggaagggatgaa - 3' и R:5'- ccaagggagggagagacaga - 3', CAST 622 п.н. - F:5'- tggggcccaatgacgccatcgatg и R:5'- ggtggagcacttctgatcacc - 3), GDF9 (462 п.н.) - F:5' - gaagactggtatggggaaatg - 3' и R:5'- ccaatctgctcctacacacct - 3%, при чем в качестве биоматериала у животных берут кровь из яремной вены, для проведения полимеразной цепной реакции применяют наборы «GenePakPCRCore», генотипирование овец осуществляют по комплексу генов GH, CAST и GDF9, для этого на программируемом четырехканальном термоциклере «Терцик» проводят амплификацию фрагмента ДНК в объеме 20 мкл - 17 мкл реакционная смесь+3 мкл ДНК; 15 мкл реакционная смесь+5 мкл ДНК - с использованием праймеров: GH 277 п.н. - F:5'-ggaggcaggaagggatgaa - 3' и R:5'- ccaagggagggagagacaga - 3', CAST 622 п.н. -F:5'- tggggcccaatgacgccatcgatg и R:5'- ggtggagcacttctgatcacc - 3), GDF9 (462 п.н.) - F:5' - gaagactggtatggggaaatg - 3' и R:5'- ccaatctgctcctacacacct - 3', после чего проводят рестрикционный анализ полученных амплификатов, при помощи эндонуклеаз рестрикции НаеШ, MspI, BstHHl, после окончания электрофореза в 2-4,0% агарозном геле, окрашенным бромистым этидием, который помещают на фильтр трансиллюминатора, излучающего свет в ультрафиолетовом диапазоне, производят визуализацию числа и длин фрагментов рестрикции, при этом в качестве маркера молекулярных масс используют стандартный набор М 50 «GenePakDNAMarkers» И устанавливают наличие и частоту аллелей и генотипов, а также комплексных генотипов, при чем подсчет частоты встречаемости генотипов определялся по формуле:
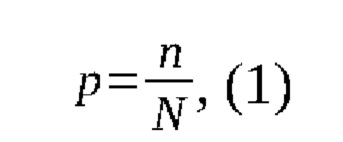
где р - частота определенного генотипа,
n - количество животных, имеющих определенный генотип,
N - общее число животных,
а расчет частоты встречаемости аллелей осуществлялся по формуле:
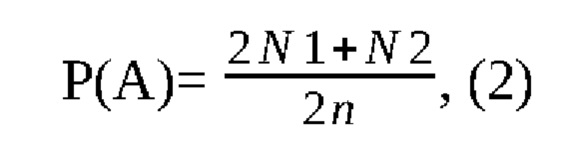
где: Р - частота встречаемости аллели;
А - аллель;
N1 - число гомозигот по исследуемому аллелю;
N2 - число гетерозигот;
n - объем выборки животных.
Ген гормона роста GH (соматотропин, соматотропный гормон, GH), расположенный на 5-й хромосоме, включает пять экзонов и четыре интрона. Суперэкспрессия гена GH распространяется на весь организм, в том числе на клеточном уровне. Основной эффект анаболический и метаболический. В результате активации биосинтеза цепи ДНК-РНК-белок и регуляции скорости протекания процесса, наблюдается мобилизация расщепления липидов, распаду высших жирных кислот и глюкозы в тканях организма. Это приводит к тому, что у животных происходит увеличение интенсивности развития, в том числе за счет роста скелета. Гормон роста - белок с молекулярной массой около 22000 дальтон, его полипептидная цепь состоит из 191 аминокислотного остатка.
Ген кальпастатина (CAST), у овец ген (ID: 443364) локализован на 5-й хромосоме, состоит из 29 экзонов и имеет общий размер 89553 п.н. Действие гена широкое, регулирует образование связки кальпаин-кальпастатин, действие которое весьма влиятельное: регуляция процессов жизнеобеспечения в организме на клеточном уровне, передача нервных импульсов в синапсах, секреции, дифференциации клеток, и других. Его полиморфизм определяют методом ПЦР-ПДРФ в первом интроне. Наличие полиморфизма гена CAST/MspI позволяет использовать его в качестве маркера продуктивности овец.
Ген дифференциального фактора роста (GDF9) является составляющим суперсемейства трансформирующего фактора роста бета (TGFb). Исследования по изучению роли GDF9 в фолликулогенезе показали, что он выполняет важную роль в поддержании нормального фолликулогенеза яичников у овец и является ооцит специфическим фактором роста и играет ключевую роль в росте и дифференцировке клеток гранулезы, а также в формировании клеток теки и, соответственно, влияет на фертильность большинства видов млекопитающих. У овец ген GDF9 расположен на 5 хромосоме, последовательность составляет 2491 п.н., состоит из двух экзонов (первый экзон содержит 397 п.н., кодирующих 134 аминокислоты, второй экзон - 968 п.н., кодирующих 322 аминокислоты), разделенных одним интроном (1126 п.н.). Пропептид представлен 453 аминокислотами, зрелый пептид состоит из 135 аминокислот.
Проведенные ранние исследования ДНК-генотипирования, данные Колосова Ю.А. и соавторов (2017 г.), по сравнению показателем роста и развития у ремонтного молодняка овец сальской породы в зависимости от генотипов по гену кальпастатин (CAST), показали, что животные - носители гетерозиготного CASTMN генотипа имели живую массу при рождении больше, чем животные сверстники гомозиготного варианта CASTMM. Также было установлено, что наличие у баранчиков этой породы гетерозиготного GHAB генотипа оказывает положительное влияние на темпы роста. По результатам исследования гена дифференциального фактора роста (GDF9), проведенным Gorlov I.F. и соавторами (2021 г.), известно, что у овец сальской и волгоградской породы были отмечены более высокие показатели плодовитости у носителей гетерозиготного GDF9AG генотипа по сравнению с особями носителями гомозиготного GDF9GG генотипа.
В связи с этим, выявлено, что аллель N гена CAST - ассоциируется с нежностью мяса; аллель В гена GH - с приростом живой массы; а аллель А гена GDF9 - с воспроизводительными качествами и энергией роста животных.
Способ осуществляется следующим образом: выделяются бараны производители с желательным генотипом, закладывается линия, в которой отбираются животные-потомки, выполняется выделение ДНК, амплификация фрагмента генов GH, CAST и GDF9 методом ПЦР-ПДРФ (полиморфизм длин рестрикционных фрагментов) с использованием праймеров: GH (277 п.н.) -(F:5 - ggaggcaggaagggatgaa - 3' и R:5 - ccaagggagggagagacaga - 3'); CAST (622 п.н.) - (F:5'- tggggcccaatgacgccatcgatg и R:5'- ggtggagcacttctgatcacc-3'); GDF9 (462 п.н.) - (F:5' - gaagactggtatggggaaatg - 3' и R:5'-ccaatctgctcctacacacct - 3') рестрикция амплифицированного фрагмента гена GH эндонуклеазой HaeIII, CAST эндонуклеазой MspI, гена GDF9 эндонуклеазой рестрикции BstHHl. Формируются группы животньж по желательным генотипам, создаются детализированные нормы кормления, в настоящих исследованиях используется кормовая добавка животного происхождения перерабатывающих отраслей АПК «ЛактуВет». Проводится анализ ассоциативных связей аллельного спектра с показателями роста и продуктивности ягнят разных генотипов. В возрасте 6 месяцев проводится убой животных, анализируются полученные гистологические и морфометрические данные.
Таким образом, представленный способ позволяет более эффективно проводить оценку мясной продуктивности овец в период постэмбрионального онтогенеза и повысить рентабельность овцеводства.
Краткое описание чертежей и иных материалов
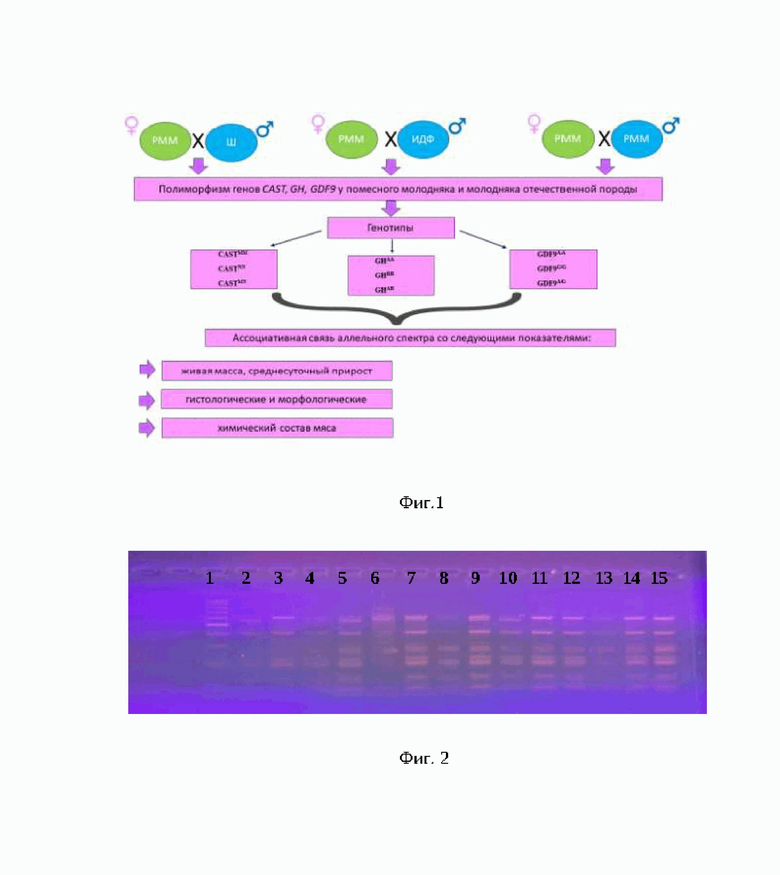
На фиг. 1 - схема опыта.
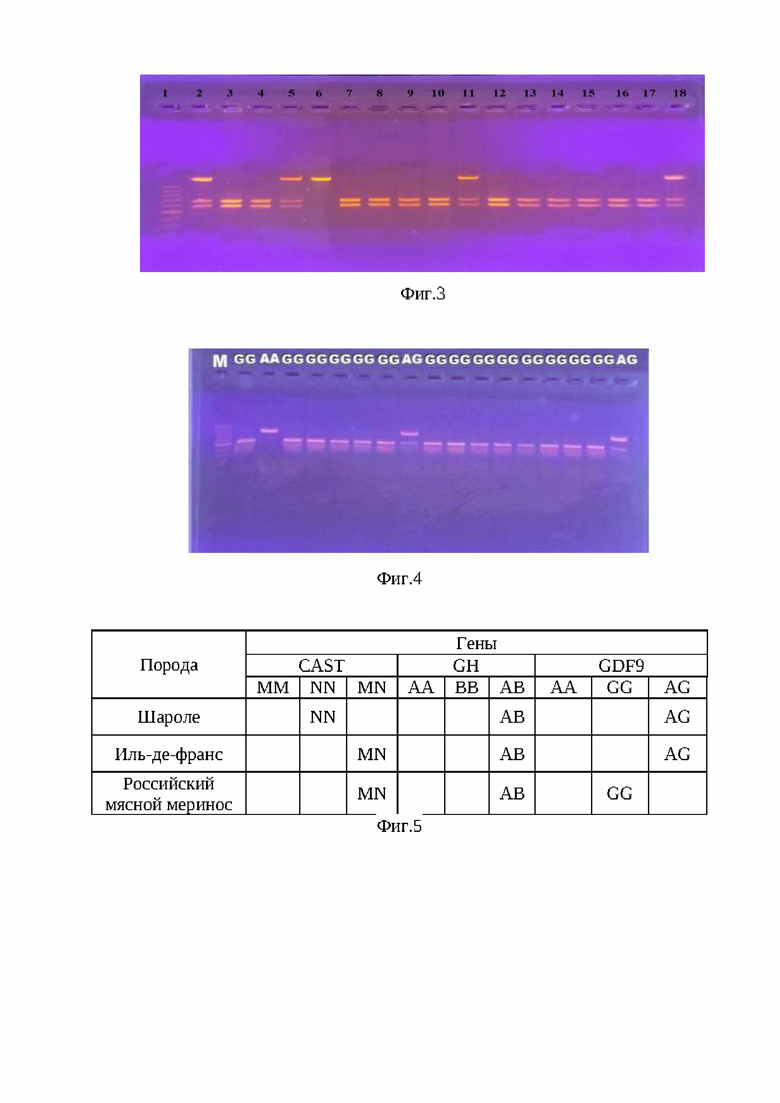
На фиг. 2 - электрофореграммы результата ПЦР-ПДРФ гена GH в 4% агарозном геле. Обозначения: 1 - ДНК-маркер 50 bp (IsoGeneLab); 3, 11, 12, 13, 14, 15 - генотип АА (277; 202; 110; 100; 94; 68; 49; 22; 8 и 4 п.н); 4, 5, 6, 7, 8, 9, 10 - генотип АВ (277; 256; 202; 110; 100; 94; 68; 49; 22; 8 и 4 п.н). Дорожка 2 - генотип ВВ (256; 202; 110; 94; 68; 49; 22; 8 и 4 п.н.).
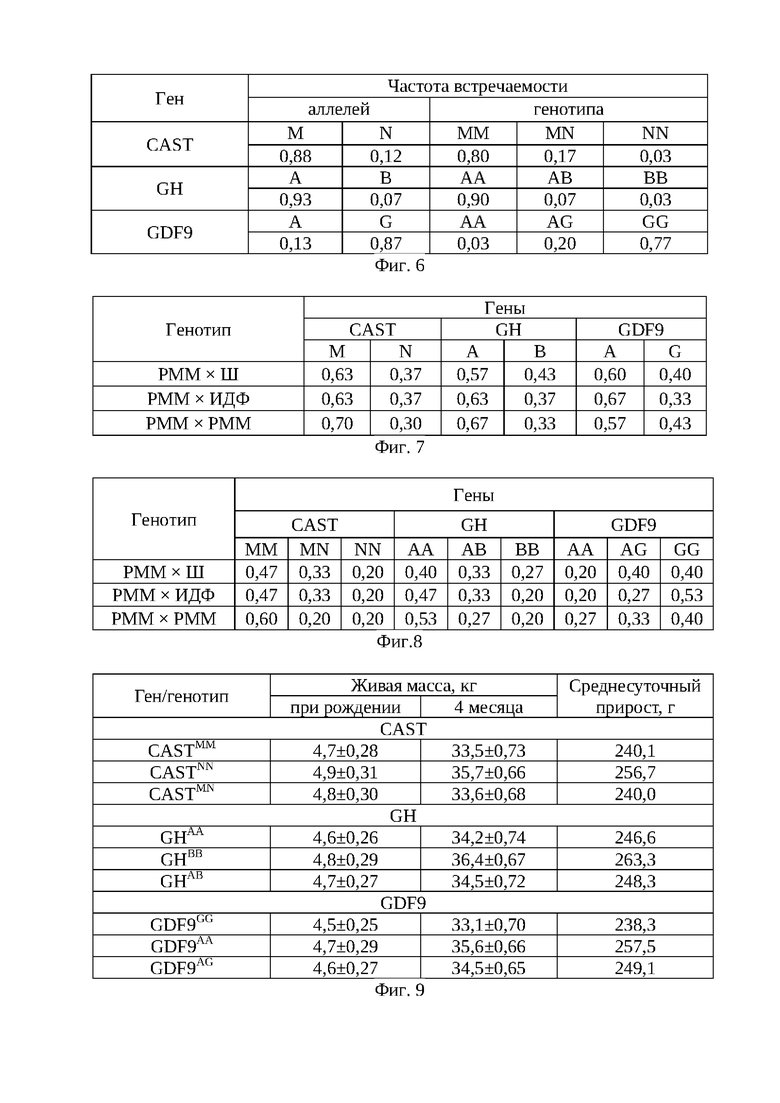
На фиг. 3 - электрофреграммы фрагментов рестрикции гена CAST в 1,8% агарозном геле. Обозначения: 1 - ДНК-маркер 50 bp (IsoGeneLab); 2, 5, 11, 18 - генотип MN (622; 336; 286 п.н.) 3, 4, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 - генотип ММ (336; 286 п.н.). Дорожка 6 - генотип NN (622 п.н.).
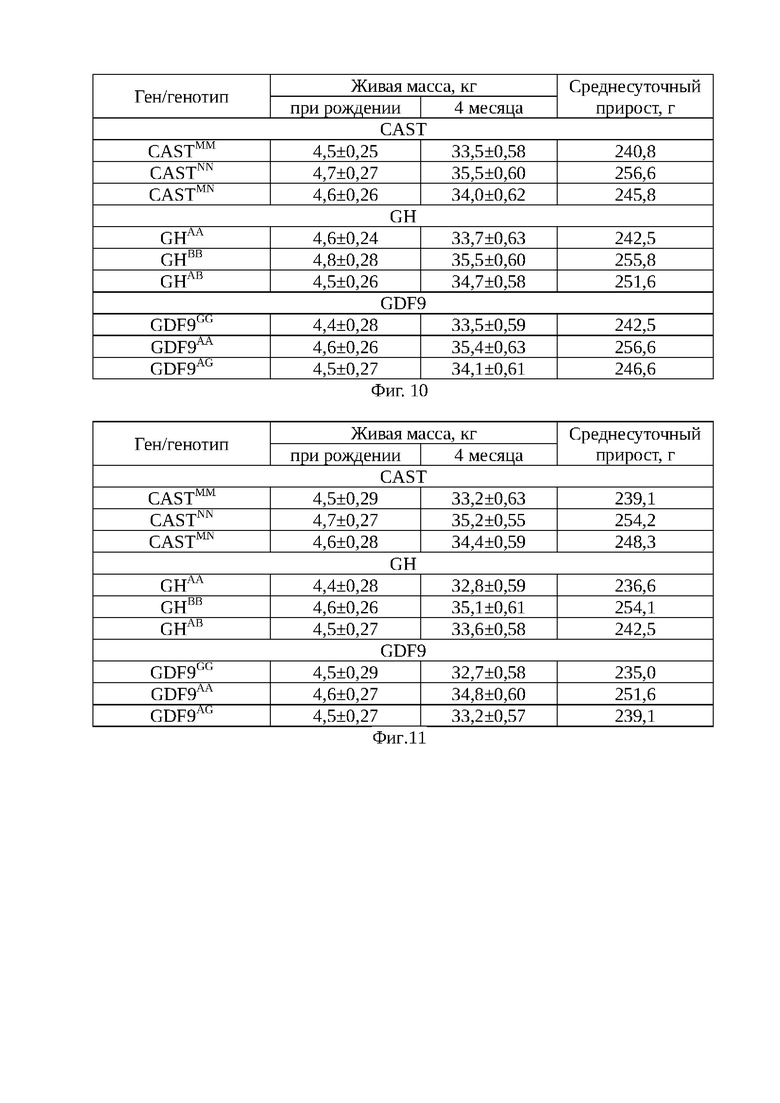
На фиг. 4 - электрофореграммы результата ПЦР-ПДРФ гена дифференциального фактора роста (GDF9) в 2% агарозном геле. Обозначения: 1 - ДНК-маркер 50 bp (IsoGeneLab); 9, 18 - генотип AG (410, 254, 117 п.н). 2, 4, 5, 6, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17 - генотип GG (254, 117 п.н.). 3, - генотип АА (410 п.н.).
На фиг. 5 - полиморфизм генов CAST, GH, GDF9 у баранов-производителей разных пород (n=3).
На фиг. 6 - аллельный спектр генов CAST, GH, GDF9 у овцематок породы российский мясной меринос (n=100).
На фиг. 7 - аллельный спектр генов GH; CAST; GDF9 помесного молодняка пород шароле (n=15), иль-де-франс (n=15) и чистопородного РММ (n=15).
На фиг. 8 - полиморфизм генов CAST, GH, GDF9 у баранчиков разных генотипов.
На фиг. 9 - динамика роста помесных баранчиков (РММ×III).
На фиг. 10 - динамика роста помесных баранчиков (РММ×ИДФ).
На фиг. 11 - динамика роста чистопородных баранчиков породы РММ.
На фиг. 12 - микроструктурный анализ m. longissimus dorsi 6 мес.баранчиков разных пород CASTNN и CASTMM генотипов.
На фиг. 13 - гистосрез мышечной ткани длиннейшей мышцы спины (1 - диаметр мышечных волокон, 2 - жировая ткань, 3 - соединительная ткань) баранчиков 6 мес.породы Шаролле CASTNN генотипа (окраска гематоксилин-эозин, увеличение ×500 раз).
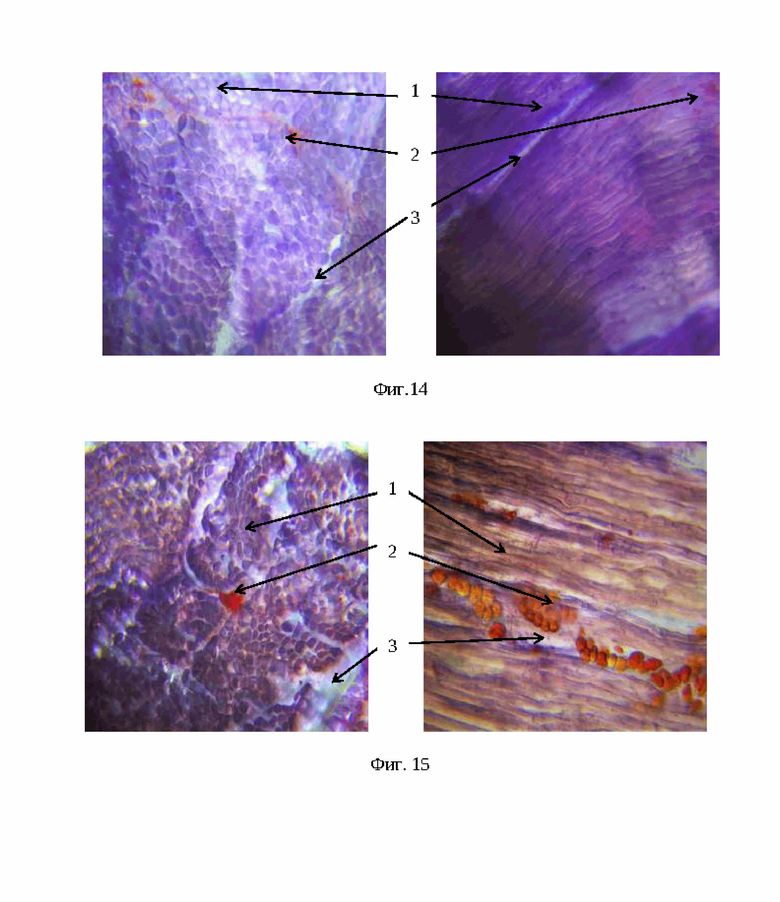
На фиг. 14 - гистосрез мышечной ткани длиннейшей мышцы спины (1 - диаметр мышечных волокон, 2 - жировая ткань, 3 - соединительная ткань) баранчиков 6 мес.породы Иль-де-франс CASTNN генотипа (окраска гематоксилин-эозин, увеличение × 500 раз).
На фиг. 15 - гистосрез мышечной ткани длиннейшей мышцы спины (1 - диаметр мышечных волокон, 2 - жировая ткань, 3 - соединительная ткань) баранчиков 6 мес.породы РММ CASTNN генотипа (окраска гематоксилин-эозин, увеличение × 500 раз).
Осуществление изобретения
Материалом исследований служили помесные баранчики, полученные от овцематок отечественной селекции и импортных баранов-производителей. Базой для выполнения экспериментальной части послужило производственно-экспериментальное подразделение Всероссийского научно-исследовательского института овцеводства и козоводства - ФГБНУ «Северо-Кавказский ФНАЦ», расположенное в Шпаковском районе Ставропольского края.
Примеры конкретного выполнения способа оценки генетического потенциала ягнят на основе исследования генов, влияющих на технологические параметры мяса.
Способ оценки генетического потенциала ягнят осуществляют следующим образом. Молекулярно-генетическое тестирование животных проводят методом ПЦР-ПДРФ, в результате которого получены сведения о комплексных аллельных вариантах и генотипах. В качестве биоматериала для проведения ДНК-генотипирования у животных использовалась кровь, взятая из яремной вены. Для проведения полимеразной цепной реакции (ПЦР) применяют наборы «GenePakPCRCore», (IsoGeneLab, Москва). Генотипирование овец осуществляют по комплексу генов GH, CAST и GDF9. На программируемом четырехканальном термоциклере «Терцик» фирмы «ДНК-технология» (Россия) проводят амплификацию фрагмента ДНК в объеме 20 мкл (17 мкл реакционная смесь+3 мкл ДНК; 15 мкл реакционная смесь+5 мкл ДНК) с использованием праймеров: GH (277 п.н.) - (F:5'-ggaggcaggaagggatgaa - 3' и R:5'- ccaagggagggagagacaga - 3'); CAST (622 п.н.) - (F:5'- tggggcccaatgacgccatcgatg и R:5'- ggtggagcacttctgatcacc - 3'); GDF9 (462 п.н.) - (F:5' - gaagactggtatggggaaatg - 3' и R:5'- ccaatctgctcctacacacct -3'). Рестрикционный анализ полученных амплификатов, проводят при помощи эндонуклеаз рестрикции HaeIII, MspI, BstHH1, после окончания электрофореза в 2-4,0% агарозном геле, окрашенным бромистым этидием, помещают на фильтр трансиллюминатора, излучающего свет в ультрафиолетовом диапазоне, производят визуализацию числа и длин фрагментов рестрикции. В качестве маркера молекулярных масс используют стандартный набор М 50 «GenePakDNAMarkers» (IsoGeneLab) (Фиг. 2-4).
По результатам молекулярно-генетического анализа устанавливают наличие и частоту аллелей и генотипов, а также комплексных генотипов.
Подсчет частоты встречаемости генотипов определялся по формуле:
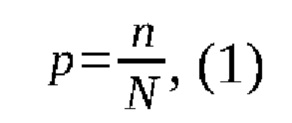
где р - частота определенного генотипа;
n - количество животных, имеющих определенный генотип;
N - общее число животных.
Расчет частоты встречаемости аллелей осуществлялся по формуле:
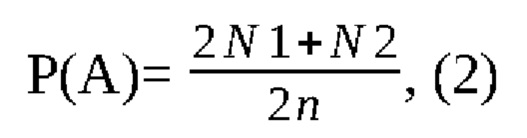
где: Р - частота встречаемости аллели;
А - аллель;
N1 - число гомозигот по исследуемому аллелю;
N2 - число гетерозигот;
n - объем выборки животных.
Для проведения исследований были выбраны бараны-производители пород шароле (n=3), иль-де-франс (n=3), российский мясной меринос (n=3) и овцематки породы российский мясной меринос (n=100), а также полученный молодняк первого поколения (F1) (n=45).
Методом ДНК-генотипирования у баранов-производителей импортных пород установлено присутствие желательных аллелей в генах кальпастатина (CAST), гормона роста (GH), дифференциального фактора роста (GDF9), маркирующих высокую продуктивность (Фиг. 5). При изучение аллельного профиля генов GH, CAST, GDF9 было выявлено, что баран-производитель породы шароле является носителем 4-х желательных аллелей по трем маркерным генам (CASTNN/GHAB/GDF9AG), породы иль-де-франс - носитель 3-х аллелей в 3-х генах (CASTMN/GHAB/GDF9AG), а также отмечено присутствие двух маркерных аллелей по генам кальпастатина и гормона роста (CASTMN/GHAB/GDF9GG) у российского мясного мериноса.
Следующим этапом работы является изучение полиморфизма исследуемых генов у овцематок породы российский мясной меринос. Анализируя полученные результаты ДНК-генотипирования, методом ПЦР-ПДРФ, исследуемого поголовья овцематок породы РММ, установлена раза значительная разница в 7,3 по превосходству (0,88) частоты встречаемости аллеля CASTM гена кальпастатин над селекционно-значимым аллелем CASTN (0,12), что, в свою очередь, обусловило присутствие носителей желательных вариантов гомозиготного CASTNN и гетерозиготного CASTMN генотипа в изучаемой выборке животных, составившей 3,0 и 17,0% (Фиг. 6). Анализ полученных результатов показал, что полиморфизм гена гормона роста (GH) в исследуемой выборке овцематок российского мясного мериноса представлен двумя аллелями GHA и GHB с очень низкой (0,07) частотой встречаемости аллеля GHB, но высокой (0,93) аллеля GHA. В данной выборке наблюдается низкая 3,0 и 7,0% частота встречаемости животных-носителей селекционно-значимого гомозиготного GHBB и гетерозиготного GHAB генотипов. Частота встречаемости аллеля GDF9A в гене дифференциального фактора роста в выборке овцематок данной породы составила 0,13, аллеля GDF9G - 0,87, что нашло отражение в наличии вариантов гомо- и гетерозиготных генотипов. Так частота встречаемости животных-носителей гомозиготного CDF9AA генотипа составила 3,0%, гетерозиготного GDF9AG 20,0%. Метод генотипирования позволяет изучить наследственность на уровне ДНК, что важно в условиях крупномасштабной селекции, когда от одного производителя получают множество тысяч потомков. В результате проведения скрещивания баранов-производителей разных пород (шароле, иль-де-франс, российского мясного мериноса) с овцематками породы российский мясной меринос был получен помесный и чистопородный молодняк первого поколения (F1). У полученного ремонтного молодняка (F1) изучены особенности полиморфизма генов GH, CAST, GDF9.
Анализ результатов выявил, что частота встречаемости селекционно-значимого аллеля CASTN гена кальпастатина в выборках помесных баранчиков, полученных от овцематок породы российский мясной меринос и баранов-производителей пород шароле и иль-де-франс, была одинаковой (0,37), у чистопородных баранчиков породы российский мясной меринос отмечена приблизительно такая же (0,30); частота встречаемости CASTM аллеля у помесных баранчиков от производителей импортной селекции была равновеликой, составив 0,63, баранчиков российский мясной меринос - 0,70 соответственно. Молекулярно-генетическими исследованиями гена GH анализируемого помесного поголовья (РММ×Ш и РММ×ИДФ) выявлена приблизительно одинаковая частота встречаемости (0,43 и 0,37) желательного аллеля GHB, а в выборке чистопородных животных (РММ × РММ) она была низкой (0,33). Распределение необходимого аллеля GDF9A гена дифференциального фактора роста у помесного молодняка (РММ×Ш, РММ×ИДФ) и чистопородного (РММ × РММ) варьировалось в пределах от минимальных значений (0,57) до максимальных (0,67). Что касается полиморфного состояния гена CAST у помесных баранчиков (РММ×Ш, РММ×ИДФ), то частота встречаемости желательных (CASTNN и CASTMN) генотипов составила 20,0 и 33,0%. Анализом полученных данных установлено, что частота встречаемости гомозиготного и гетерозиготного (GHBB и GHAB) желательных генотипов гена GH исследованного помесного молодняка (РММ×Ш и РММ×ИДФ) составила: у РММ×Ш - 27,0 и 33,0%; РММ×ИДФ - 20,0 и 33,0%; а чистопородного РММ × РММ - 20,0 и 27,0%. Частота встречаемости гомозиготного GDF9AA генотипа у помесных баранчиков (РММ×Ш и РММ×ИДФ) была одинаковой, составив 20,0%, у гетерозиготного GDF9AG генотипа варьировалась в пределах от 27,0 до 40,0%. У чистопородных ягнят породы РММ частота встречаемости селекционно-значимого гомозиготного СDF9AА генотипа составила 27,0%, а частота встречаемости гетерозиготного GDF9AG генотипа достигла 33,0% (Фиг. 7-8).
Были отработаны рецептуры комбикормов - стартеров для молодняка овец в возрасте от рождения до 4-х месяцев с включением 3,0% пребиотика «ЛактуВет». Разработанные рецептуры комбикормов - стартеров при скармливании молодняку овец оказали положительное влияние на продуктивность. У животных опытных групп живая масса к отбивке составила 34,8 - 36,4 кг, что на 6,4% больше, чем у сверстников контрольной группы. У опытного поголовья среднесуточный прирост живой массы составил 251-263 г, что выше, чем у животных контрольной группы на 6,3 -6,8%.
Влияние комплексных генотипов генов GH, CAST, GDF9 на скорость роста учитывают у баранчиков по следующим показателям: живая масса при рождении и в возрасте 4 месяца (кг), среднесуточный прирост от рождения до 4 месяцев (г). Выявлено, величина живой массы в возрасте 4-х месяцев у помесного поколения баранчиков (РММ×Ш) носителей желательных гомозиготных CASTNN, GHBB и CDF9AA генотипов, по сравнению со сверстниками носителями вариантов CASTMM, GHAA и GDF9GG была выше на 6,5; 6,4 и 7,5% соответственно. Определено, среднесуточный прирост у животных носителей желательных генотипов преобладал на 6,9; 6,7 и 8,5%, в сравнении с группой сверстников. Установлено, живая масса была выше на 5,9; 5,3 и 5,6% у помесных ягнят (РММ×ИДФ) носителей гомозиготных CASTNN, GHBB и CDF9AA генотипов по сравнению с животными аналогами GHAA, CASTMM, GDF9GG. Выявлено, среднесуточный прирост в группе животных носителей селекционно-значимых генотипов был выше на 6,5; 5,4 и 5,8%, в сравнении с животными аналогами. Установлено, у чистопородного ремонтного молодняка (РММ) носителей селекционно-значимых CASTNN, GHBB и GDF9AA генотипов преимущество по величине живой массы на 6,0; 7,0 и 6,4% в сравнении со сверстниками GH^, CASTMM, GDF9GG вариантов генотипа. Выявлено, среднесуточный прост у животных носителей желательных генотипов был выше на 6,3; 7,3 и 7,0%, в сравнении с группой сверстников, имеющих нежелательный генотип (Фиг. 9-11).
Объективные сведения о качестве мяса, его химическом составе, биологической ценности позволил получить контрольный убой животных. Микроструктурный анализ мышечных волокон длиннейшей мышцы спины дает объективную характеристику качества мяса на гистологическом уровне. Нежность мяса обусловлена породными особенностями, возрастом, упитанностью, физическими нагрузкам. Гистогенез мышечной ткани включает в себя: количество мышечных волокон, их диаметр, балльную оценку мраморности, процентное содержание соединительной ткани. У животных мясных пород жир откладывается между мышцами, образуя на разрезе мышечной ткани «мраморность». Мраморность мяса -наличие жировых внутримышечных прослоек, придающих мясу вид мрамора (процентное соотношение мышечной, жировой и соединительной ткани). Оценку «мраморности» мяса определяли по запатентованной шкале гистологической оценки «мраморности» длиннейшего мускула спины у животных (Дмитрик И.И. и др., 2010 г.). В проведенном эксперименте выявлено, группа животных пород (РММ×Ш); (РММ×РММ); (РММ×ИДФ) носителей нежелательного генотипа CASTMM по количеству мышечных волокон и оценки «мраморности» уступали животным с селекционно-значимым генотипом CASTNN на 2,68; 1,37; 2,57% соответственно. Диаметр мышечного волокна у животных нежелательного генотипа CASTMM был меньше на 2,73; 2,93; 3,38% по сравнению с вариантами аналогами CASTNN соответственно. По содержанию соединительной ткани у животных с генотипом CASTMM обнаружено превосходство на 5% (Фиг. 12-15).
Таким образом, микроструктурный анализ мышечной ткани баранчиков с важным для селекции генотипом CASTNN выявил лучшие вкусовые и потребительские качества.
Предлагаемое изобретение по сравнению с прототипом и другими известными техническими решениями имеет следующие преимущества:
проводить прижизненный отбор животных с повышенными показателями мясной продуктивности на основе оценки аллельного состояния перспективных генов-маркеров;
повысить численность популяции специализированных мясных овец;
увеличить производство мраморной баранины.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="СПОСОБ ОЦЕНКИ
ГЕНЕТИЧЕСКОГО ПОТЕНЦИАЛА ЯГНЯТ НА ОСНОВЕ ИССЛЕДОВАНИЯ ГЕНОВ,
ВЛИЯЮЩИХ НА ТЕХНОЛОГИЧЕСКИЕ ПАРАМЕТРЫ МЯСА.xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2024-02-28">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023120551</ApplicationNumberText>
<FilingDate>2023-08-06</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>W23044809</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023120552</ApplicationNumberText>
<FilingDate>2023-08-06</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное научное учреждение «Северо-Кавказский федеральный научный
аграрный центр»</ApplicantName>
<ApplicantNameLatin>Federal State Budgetary Scientific Institutione
"Septentrionalis Caucasi Federal Scientific Agrariam
Centrum"</ApplicantNameLatin>
<InventionTitle languageCode="ru">СПОСОБ ОЦЕНКИ ГЕНЕТИЧЕСКОГО
ПОТЕНЦИАЛА ЯГНЯТ НА ОСНОВЕ ИССЛЕДОВАНИЯ ГЕНОВ, ВЛИЯЮЩИХ НА
ТЕХНОЛОГИЧЕСКИЕ ПАРАМЕТРЫ МЯСА</InventionTitle>
<SequenceTotalQuantity>6</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Ovis aries</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggaggcaggaagggatgaa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>20</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..20</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Ovis aries</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ccaagggagggagagacaga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>24</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..24</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Ovis aries</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tggggcccaatgacgccatcgatg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Ovis aries</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggtggagcacttctgatcacc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q10">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Ovis aries</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gaagactggtatggggaaatg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q12">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Ovis aries</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ccaatctgctcctacacacct</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ оценки генетического потенциала овец породы манычский меринос на основе молекулярно-генетических маркеров | 2021 |
|
RU2776044C1 |
| Способ подбора грубошерстных овцематок для улучшения мясной продуктивности у помесей с заводскими породами | 2022 |
|
RU2819439C1 |
| Способ оценки высокой мясной продуктивности овец сальской породы | 2016 |
|
RU2662679C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТОНКОРУННЫХ ПЛЕМЕННЫХ ПОМЕСНЫХ ОВЕЦ | 2002 |
|
RU2229225C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТОНКОРУННЫХ ПЛЕМЕННЫХ ПОМЕСНЫХ ОВЕЦ (ВАРИАНТЫ) | 2002 |
|
RU2229803C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТОНКОРУННЫХ ПЛЕМЕННЫХ ПОМЕСНЫХ ОВЕЦ (ВАРИАНТЫ) | 2002 |
|
RU2229802C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТОНКОРУННЫХ ПЛЕМЕННЫХ ПОМЕСНЫХ ОВЕЦ | 2002 |
|
RU2229224C2 |
| СПОСОБ ГЕНЕТИЧЕСКОГО ПРОГНОЗИРОВАНИЯ КАЧЕСТВЕННЫХ ПОКАЗАТЕЛЕЙ ГОВЯДИНЫ ПО АДДИТИВНОМУ ДЕЙСТВИЮ НЕКОТОРЫХ SNP-МАРКЕРОВ | 2020 |
|
RU2751959C1 |
| СПОСОБ ПОДБОРА РОДИТЕЛЬСКИХ ПАР В ТОНКОРУННОМ ОВЦЕВОДСТВЕ | 1999 |
|
RU2148316C1 |
| СПОСОБ ПОТОЧНОГО ПРОИЗВОДСТВА МОЛОДОЙ БАРАНИНЫ | 2012 |
|
RU2528865C2 |

Изобретение относится к области сельскохозяйственной биотехнологии, в частности к овцеводству, и может быть использовано для оценки генетического потенциала баранчиков на ранней стадии постэмбрионального онтогенеза. Предложен способ оценки генетического потенциала ягнят по комплексу генов GH, CAST и GDF9. Способ включает взятие у животных крови из ярёмной вены, амплификацию фрагмента ДНК в объеме 20 мкл: 17 мкл реакционная смесь+3 мкл ДНК; 15 мкл реакционная смесь+5 мкл ДНК на четырехканальном программируемом термоциклере «Терцик» с использованием для проведения полимеразной цепной реакции набора «GenePakPCRCore», праймеров GH 277 п.н. - F:5'- (SEQ ID NO: 1) - 3' и R:5'- (SEQ ID NO: 2) - 3', CAST 622 п.н. - F:5'- (SEQ ID NO: 3) и R:5'- (SEQ ID NO: 4) - 3), GDF9 (462 п.н.) - F:5' - (SEQ ID NO: 5) - 3' и R:5'- (SEQ ID NO: 6) - 3'; число и длину фрагментов рестрикции определяют в 2,0-4,0% агарозном геле методом гель-электрофореза при УФ-свете в присутствии бромистого этидия. Генетический потенциал ягнят включает скорость роста ягнят, среднесуточный прирост, вкусовые качества и мраморность мяса, и его оценивают положительно в отношении ягнят по носительству гомозиготных CASTNN, GHBB и CDF9AA генотипов. Способ позволяет проводить прижизненный отбор животных с повышенной мясной продуктивностью, повысить численность популяции специализированных мясных овец, увеличить производство мраморной баранины. 15 ил., 1 пр.
Способ оценки генетического потенциала ягнят по комплексу генов GH, CAST и GDF9, включающий взятие у животных крови из ярёмной вены, амплификацию фрагмента ДНК в объеме 20 мкл: 17 мкл реакционная смесь+3 мкл ДНК; 15 мкл реакционная смесь+5 мкл ДНК на четырехканальном программируемом термоциклере «Терцик» с использованием для проведения полимеразной цепной реакции набора «GenePakPCRCore», праймеров GH 277 п.н. - F:5'- (SEQ ID NO: 1) - 3' и R:5'- (SEQ ID NO: 2) - 3', CAST 622 п.н. - F:5'- (SEQ ID NO: 3) и R:5'- (SEQ ID NO: 4) - 3), GDF9 (462 п.н.) - F:5' - (SEQ ID NO: 5) - 3' и R:5'- (SEQ ID NO: 6) - 3'; число и длину фрагментов рестрикции определяют в 2,0-4,0% агарозном геле методом гель-электрофореза при УФ-свете в присутствии бромистого этидия, отличающийся тем, что генетический потенциал ягнят включает скорость роста ягнят, среднесуточный прирост, вкусовые качества и мраморность мяса, и его оценивают положительно в отношении ягнят по носительству гомозиготных CASTNN, GHBB и CDF9AA генотипов.
| Способ оценки генетического потенциала овец породы манычский меринос на основе молекулярно-генетических маркеров | 2021 |
|
RU2776044C1 |
| Алимсолтан А | |||
| Оздемиров и др., Полиморфизм генов CAST, GH, GDF9 овец дагестанской горной породы, Юг России: экология, развитие | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| C | |||
| Машина для изготовления проволочных гвоздей | 1922 |
|
SU39A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| N.V | |||
| Shirokova et al., Genetic structure of the herd by genes GDF9, GH, CAST in merino sheep of the North Caucasus region of | |||

