Данная заявка испрашивает приоритет по предварительной заявке на патент США №62/950 624, поданной 19 декабря 2019 г., содержание которой в полном объеме включено в данный документ посредством ссылки.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способам оценки способности противораковых терапевтических средств вызывать побочные эффекты со стороны кожи. Настоящее изобретение также относится к способам оценки эффективности композиции в снижении воздействия противораковых терапевтических средств на кожу.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Афатиниб, EGFRi второго поколения, предназначенный для преодоления резистентности после терапии препаратами первого поколения, был разработан для необратимого ингибирования сигнальных путей как EGFR, так и HER2 (Li et al. 2008). См. структуру ниже.
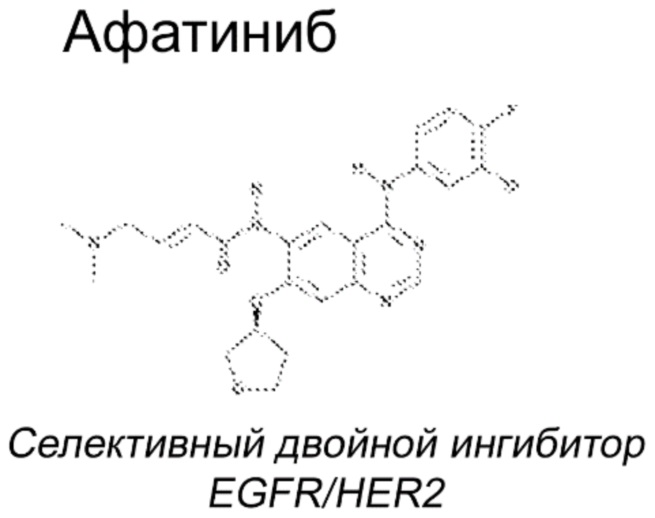
Даже несмотря на то, что механизм индуцированного лекарственным средством ингибирования EGFR в значительной степени известен, механизм, приводящий к появлению CARR, остается неясным.
Потенциальным путем выживаемости дифференцирующихся кератиноцитов является сигнальный путь, который сочетает в себе фосфоинозитид-3-киназы (PI3K), семейство внутриклеточных преобразователей сигналов с липидкиназной активностью (Whitman et al., 1988), и нижерасположенные серин-треонинкиназные Akt-эффекторы. Комплекс PI3K/AKT вызывает различные клеточные реакции, включающие рост, пролиферацию, дифференцировку и выживание клеток, контролируя антиапоптотический механизм (Calautti et al. 2005; Vivanco и Sawyers 2002). Нисходящая регуляция пути PI3K/AKT опосредована несколькими белками, в том числе супрессором опухоли - гомологом фосфатазы и тензина (PTEN) (Ali et al. 2018) и каспазой 3 (Jänicke et al. 1998), которые управляют дифференцировкой кератиноцитов и возможным ороговением, а не их апоптозом (Lippens et al. 2005). Другие белки, в том числе ингибитор циклинзависимой киназы (CDK) p21Cip1/WAF1, замедляют рост клеток и способствуют дифференцировке (Missero et al. 1996).
Витамин D3 (VD3) или холекальциферол является фактором, который часто упускают из виду, несмотря на его критически важную роль в развитии эпидермиса. VD3 представляет собой секостероидный гормон, синтезируемый в кератиноцитах в результате UVB-индуцированного фотолиза посредством синтеза 7-дегидрохолестерина, что приводит к образованию его предшественника (пре-VD3). Последний затем ферментативно гидроксилируется до кальцидиола с помощью цитохрома P450 (CYP) и, более конкретно, CYP2R1 и CYP27A1. Следует отметить, что пре-VD3 плохо регулирует CYP2R1, в отличие от CYP27A1, который сильно активируется пре-VD3 (Cheng et al. 2004; Mehlig et al. 2015). Продемонстрировано, что VD3 оказывает лияние на мышечную, скелетную и иммунную физиологию и может способствовать апоптозу и дифференцировке эпителия (Shaurova et al. 2020). CYP27A1 также участвует в разщеплении холестерина в желчных кислотах как классическим, так и кислотным путями (Norlin et al. 2003).
Развитие поляризованных иммунных ответов (включая баланс Th1/Th2) регулируется активностью субпопуляций CD4+Th-клеток и их цитокиновыми продуктами. Цитокины, родственные с Th2, включая интерлейкины (IL) 4, IL5 и IL13, приводят к продукции IgE через эозинофильный ответ, противодействуя Th1-опосредованному микробному действию (Howell et al. 2008). Аллергическое воспаление кожи приводит к атопическому дерматиту (AD), хроническому заболеванию, характеризующемуся интенсивным зудом, сухостью и эритемой в локализованных очагах поражения (Bieber 2008). Экспрессия цитокинов Th2 способствует снижению экспрессии белков S100, которые участвуют в регуляции пролиферации, дифференцировки, апоптоза, гомеостаза Ca2+, энергетического метаболизма, воспаления и миграции/инвазии посредством взаимодействия с различными белками-мишенями, включая ферменты, субъединицы цитоскелета, рецепторы, факторы транскрипции и нуклеиновые кислоты (Howell et al. 2008).
Ранее авторы сообщали, что EGFRi способствуют дифференцировке кератиноцитов и снижают пролиферацию (Joly-Tonetti et al. 2020). Однако молекулярные механизмы, ассоциированные с CADR, включая кожную сыпь или более тяжелые реакции, такие как синдром «рука-нога» или синдром Стивена Джонсона (SJS), остаются неясными. Афатиниб, препарат второго поколения EGFRi, нарушает структуру кожного барьера, уменьшая толщину эпидермиса, снижая пролиферацию кератиноцитов и повышая экспрессию маркеров дифференцировки, таких как инволюкрин, филагрин и десмоглеин-1 (Joly-Tonetti et al. 2020).
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Фосфопротеомные и транскриптомные анализы были проводили на тканях реконструированного эпидермиса человека (RHE), подвергшихся воздействию терапевтически значимой концентрации афатиниба, для выявления молекулярных сигнатур, ассоциированных с CADR.
После воздействия препарата наблюдалась активация пути PI3K/AKT, ассоциированная с повышенной экспрессией семейств генов, участвующих в дифференцировке кератиноцитов, старении, окислительном стрессе и изменениях эпидермальных иммунных маркеров.
Полученные результаты демонстрируют, что афатиниб может влиять на метаболизм витамина D3 (VD3), действуя через CYP27A1 и CYP24A1, регулируя концентрацию кальция посредством пути PI3K/AKT. Следовательно, кератиноциты базального слоя переключаются с пролиферирующей на продифференцирующую программу, характеризующуюся активацией биомаркеров, ассоцииронных с повышенной кератинизацией, ороговением, реакцией Th2 и снижением врожденного иммунитета. Такие эффекты могут повышать восприимчивость кожи к проникновению через кожу раздражителей и патогенов.
В совокупности эти результаты демонстрируют молекулярный механизм EGFR-индуцированных CADR. Эти пути обеспечивают новые биомаркеры для оценки эффективности композиции в снижении нежелательных реакций терапевтических средств, включая терапевтические средства для лечения рака.
ОПИСАНИЕ ФИГУР
На Фиг. 1 продемонстрировано функциональное нарушение в кератиноцитах RHE после воздействия афатиниба по сравнению с контролем.
Была рассмотрена обобщенная линейная модель, учитывающая взаимодействие между временем и лечением (афатиниб в дозе 100 нМ по сравнению с контролем).
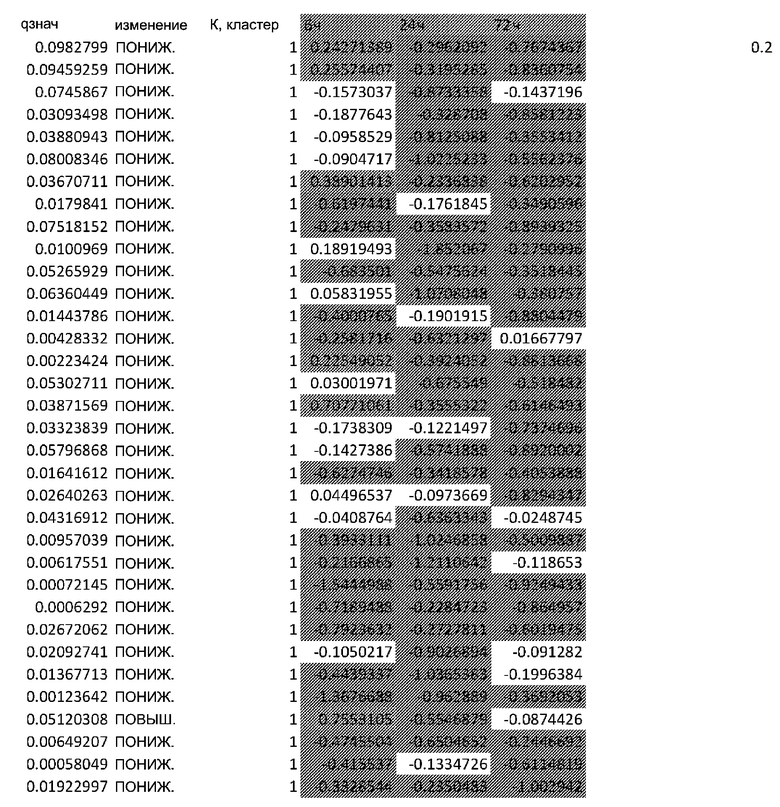
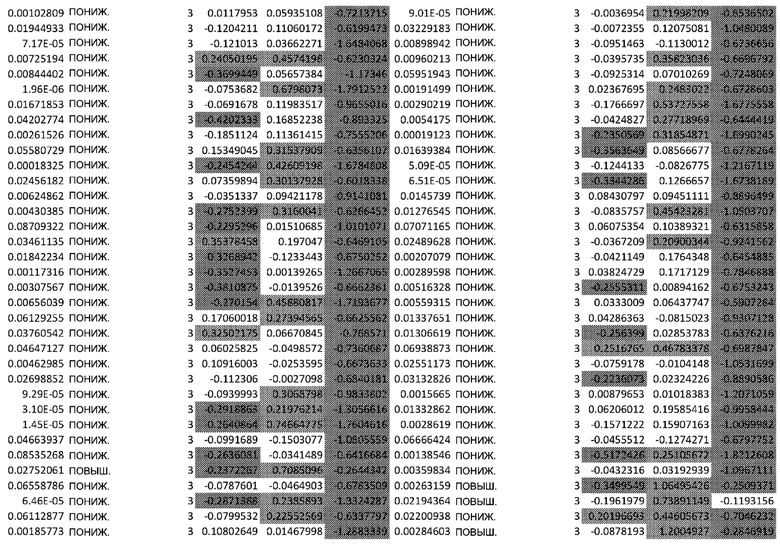
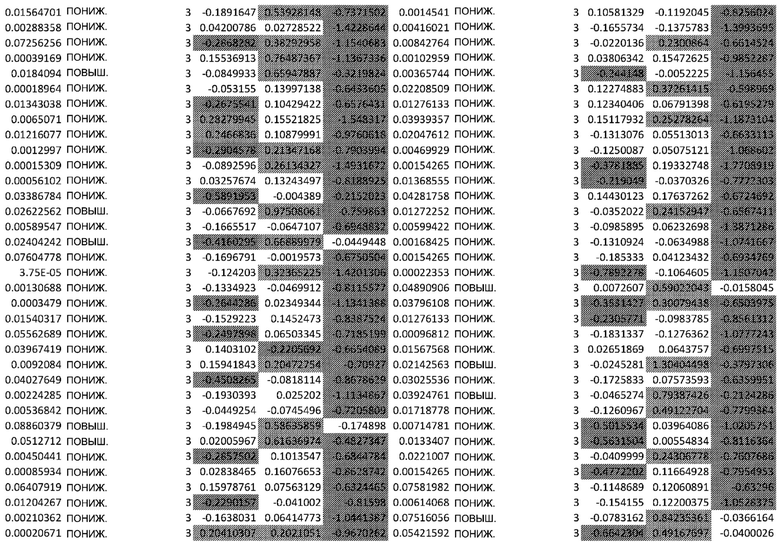
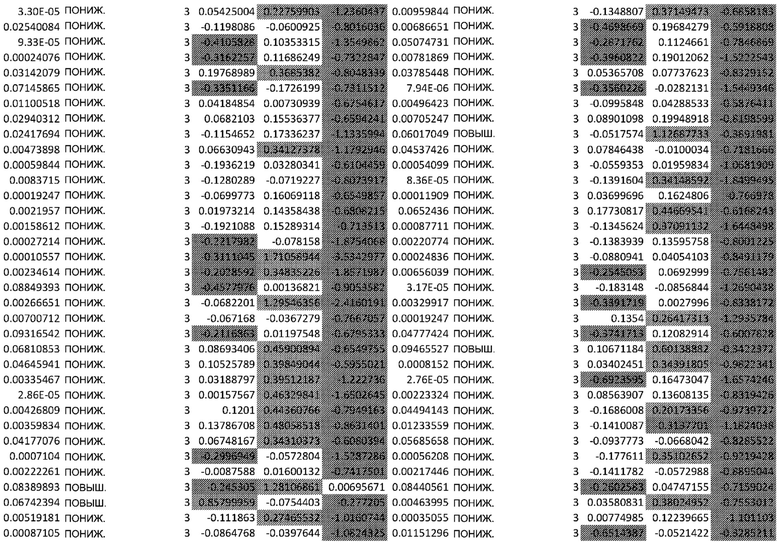
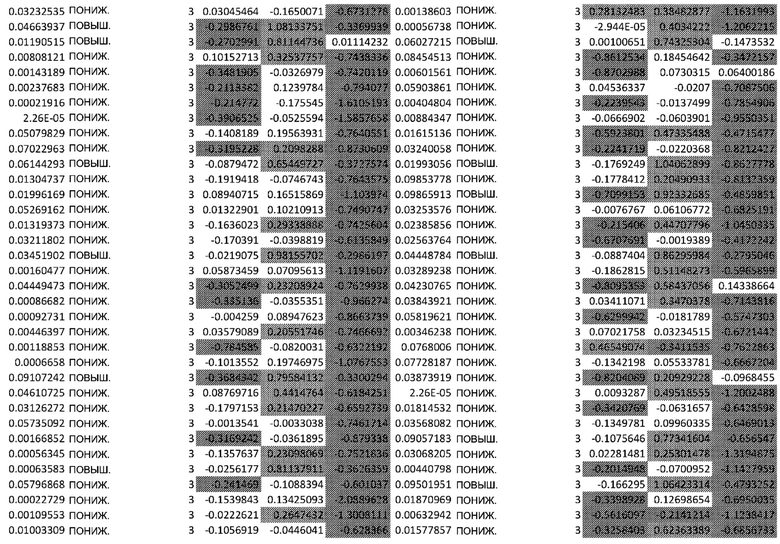
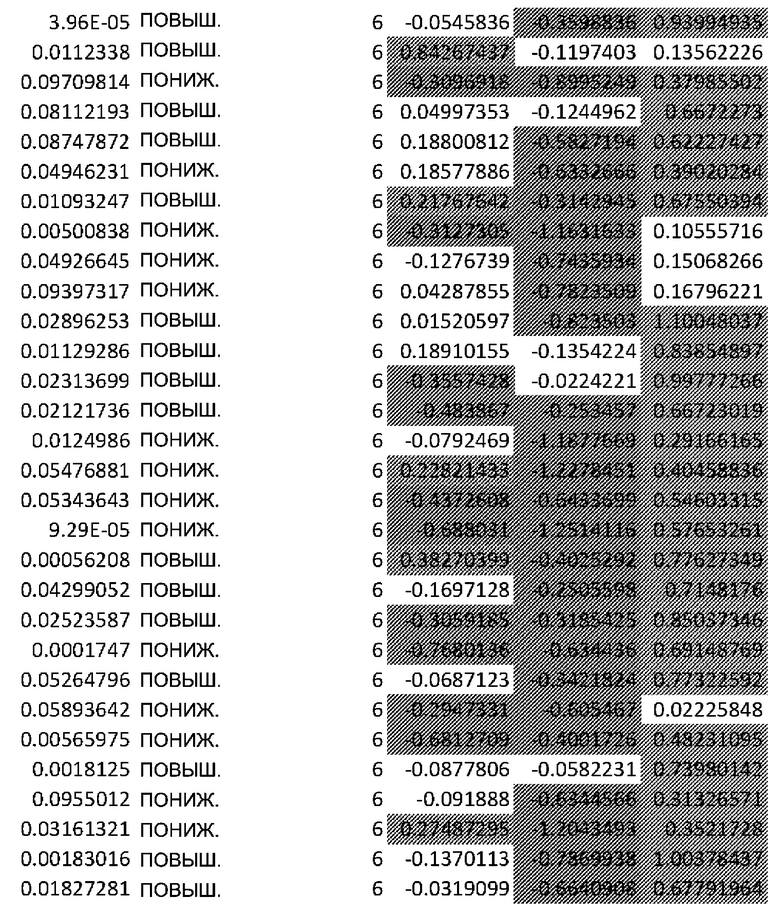
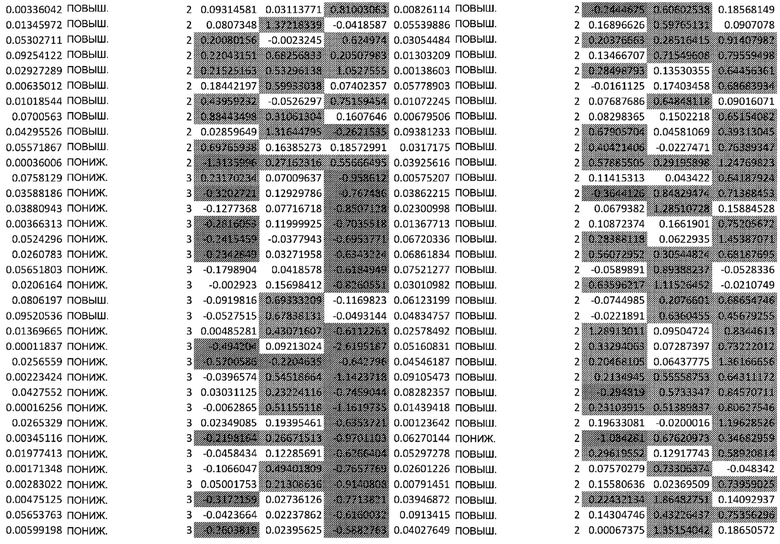
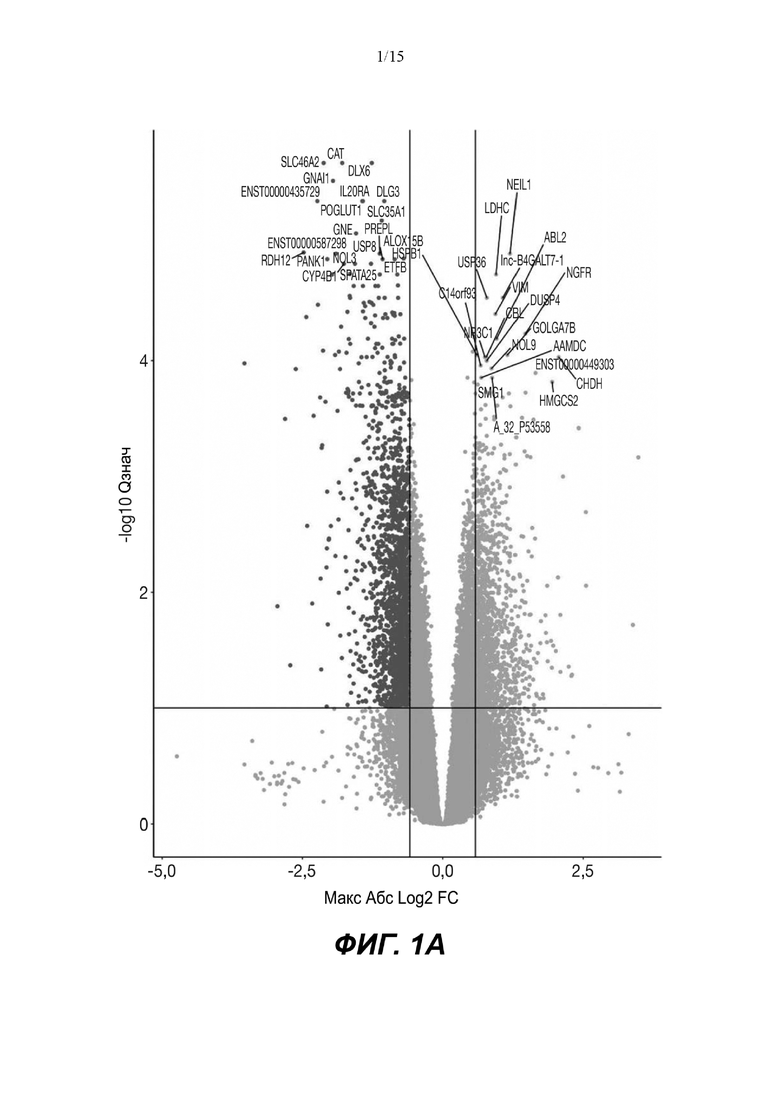
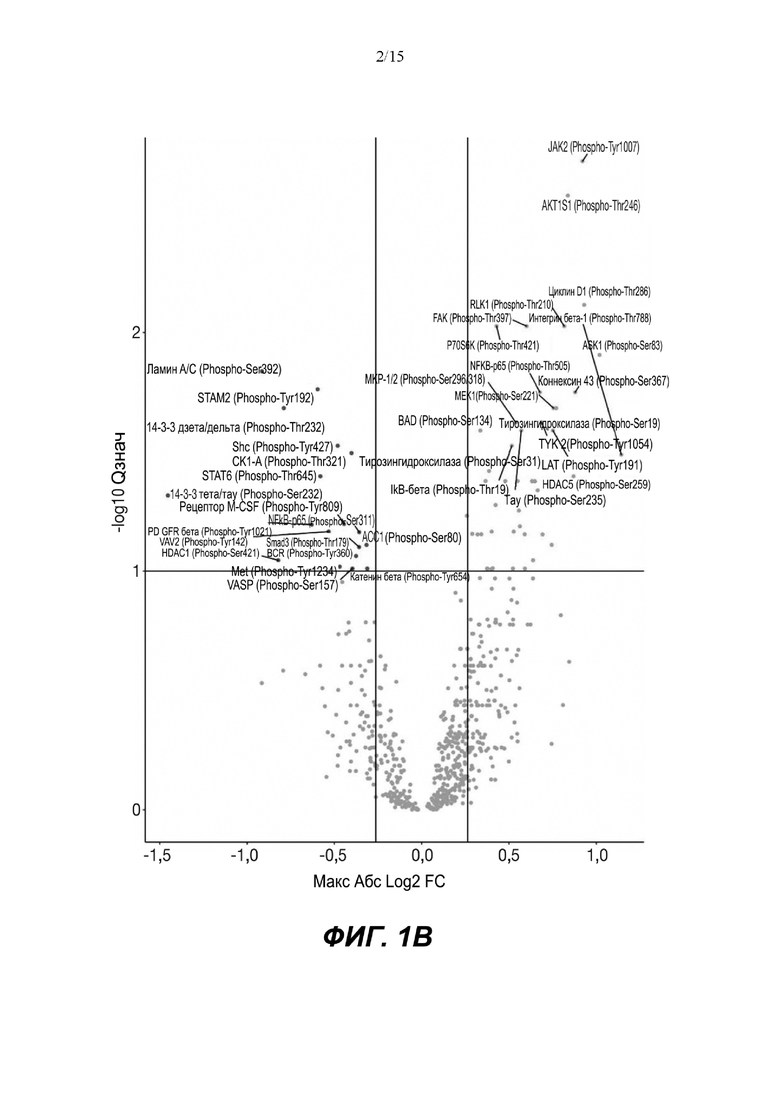
(A) Вулканический график, изображающий результаты дифференциального анализа белков, выполненного на матрице фосфорилирования. С применением отсечки 1,2-кратного изменения было идентифицировано 62 белка с изменением их статуса фосфорилирования, в том числе 44 белка со значительным повышением (зеленый цвет) и 18 белков со значительным снижением (красный цвет) их статуса фосфорилирования. В зависимости от времени, после 20 минут воздействия афатиниба статус фосфорилирования 6 белков повышался, а 3 белков - снижался. Через 24 ч повышенный статус фосфорилирования наблюдался для 18 белков и сниженный статус фосфорилирования - для 23 белков. В момент времени 72 ч статус фосфорилирования белков был снижен для 6 белков и повышен для 6 других.
(B) Вулканический график, изображающий результаты дифференциального анализа экспрессии генов (отсечка: 50%). В момент времени 6 ч экспрессия 170 генов была сниженной, а экспрессия 201 гена была повышенной. В момент времени 24 ч 335 генов были ингибированы, а 281 ген был активирован. В момент времени 72 ч экспрессия гена отмечалась повышенной для 2888 генов и сниженнной - для 1728 генов.
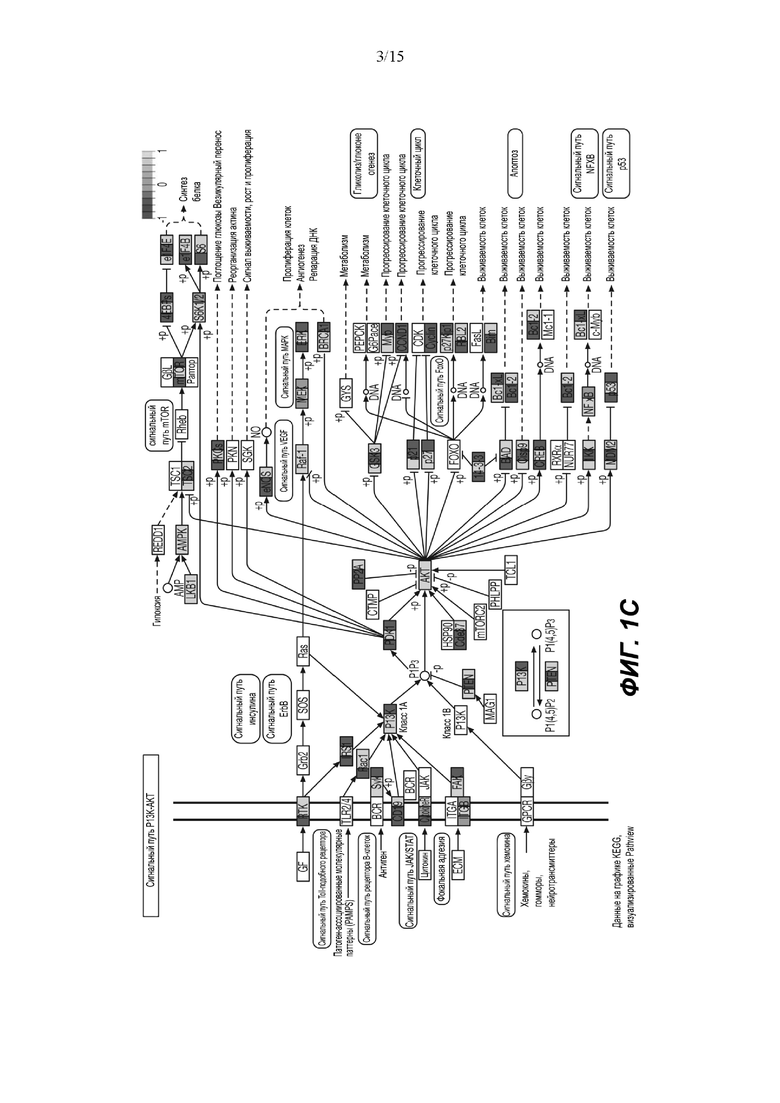
(C) Анализ обогащения набора выявил значительное обогащение пути PI3K-AKT через 24 часа воздействия афатиниба. Белки с повышенным статусом фосфорилирования показаны зеленым, а белки с пониженным статусом фосфорилирования показаны красным.
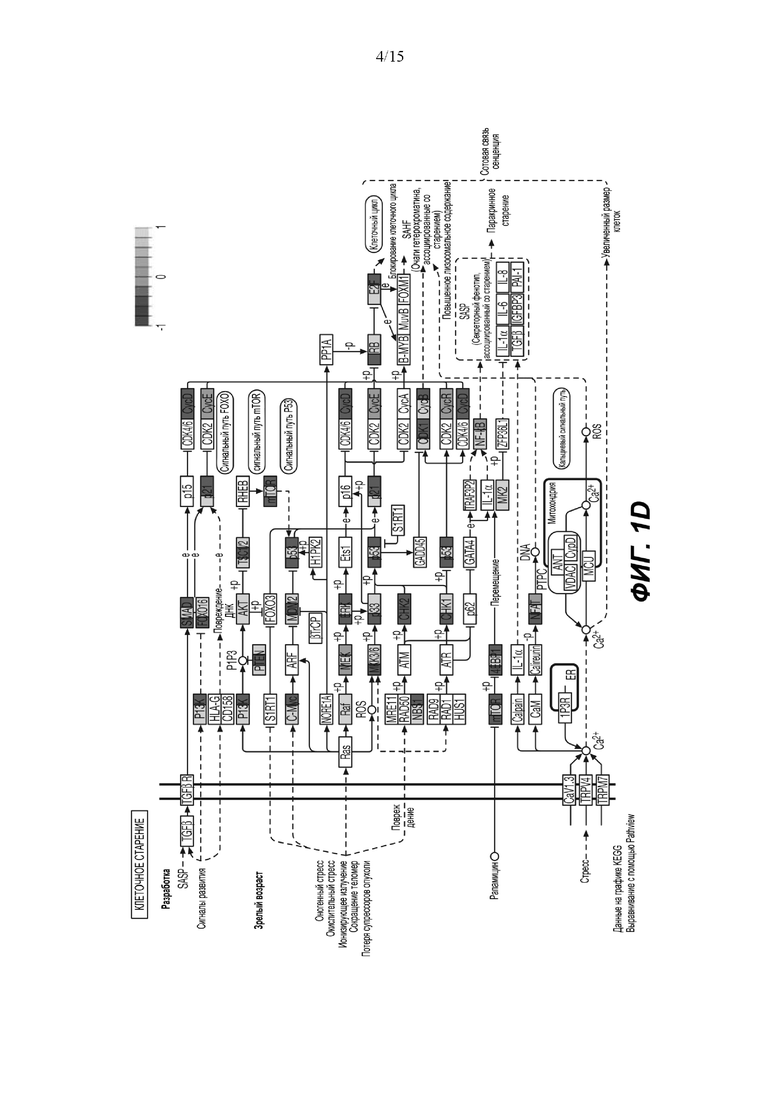
(D) Путь, ассоциированный с клеточным старением, включающий необратимую остановку роста, сопровождаемую фенотипической модификацией, продемонстрировал значительное обогащение, основанное на изменении статуса фосфорилирования белка. Путь старения находится ниже пути PI3K/AKT.
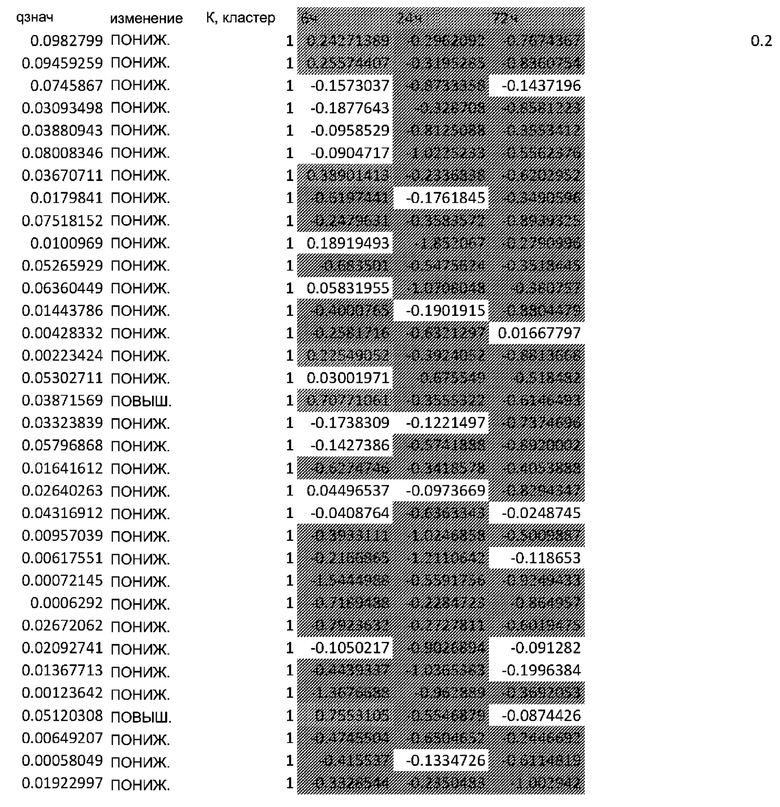
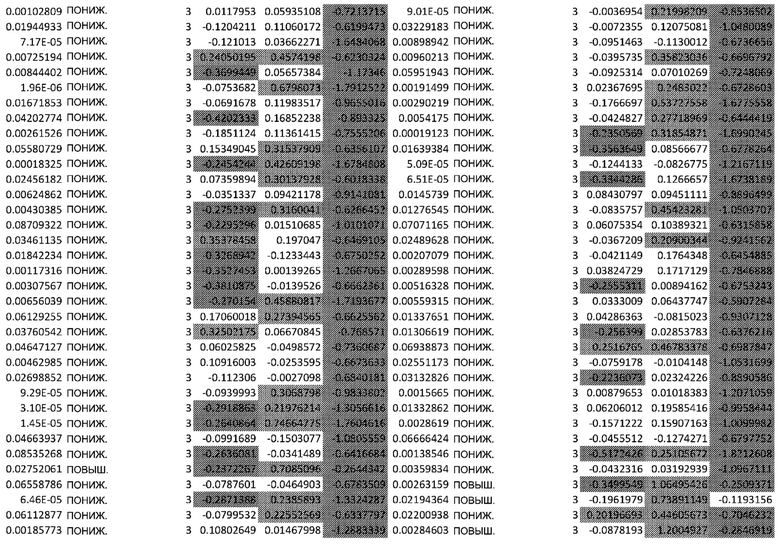
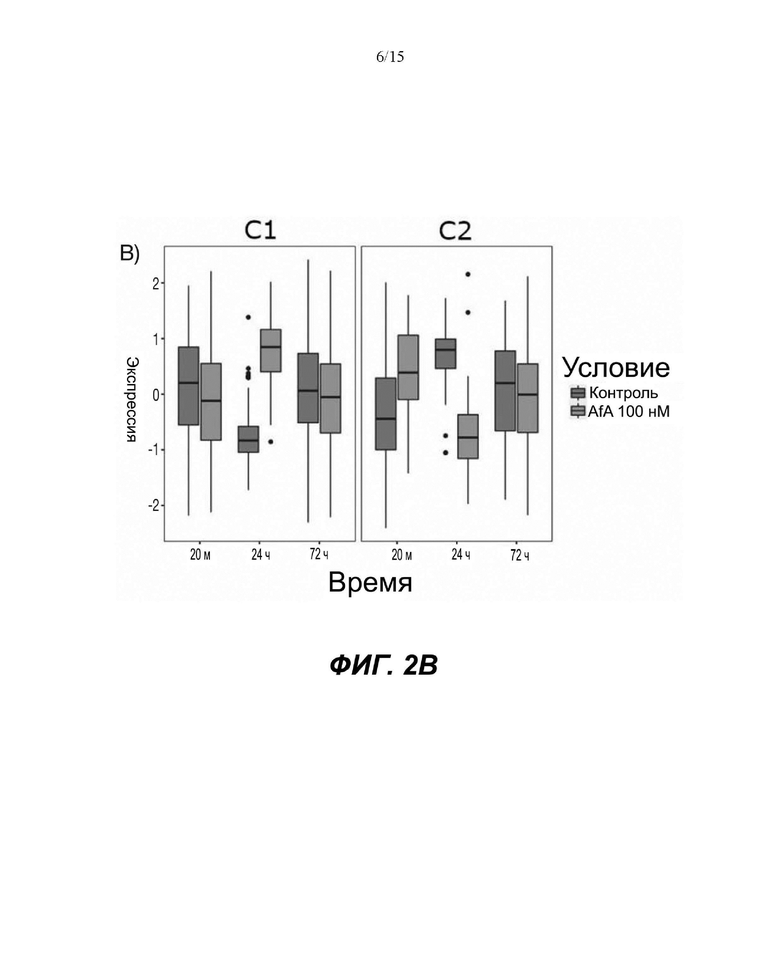
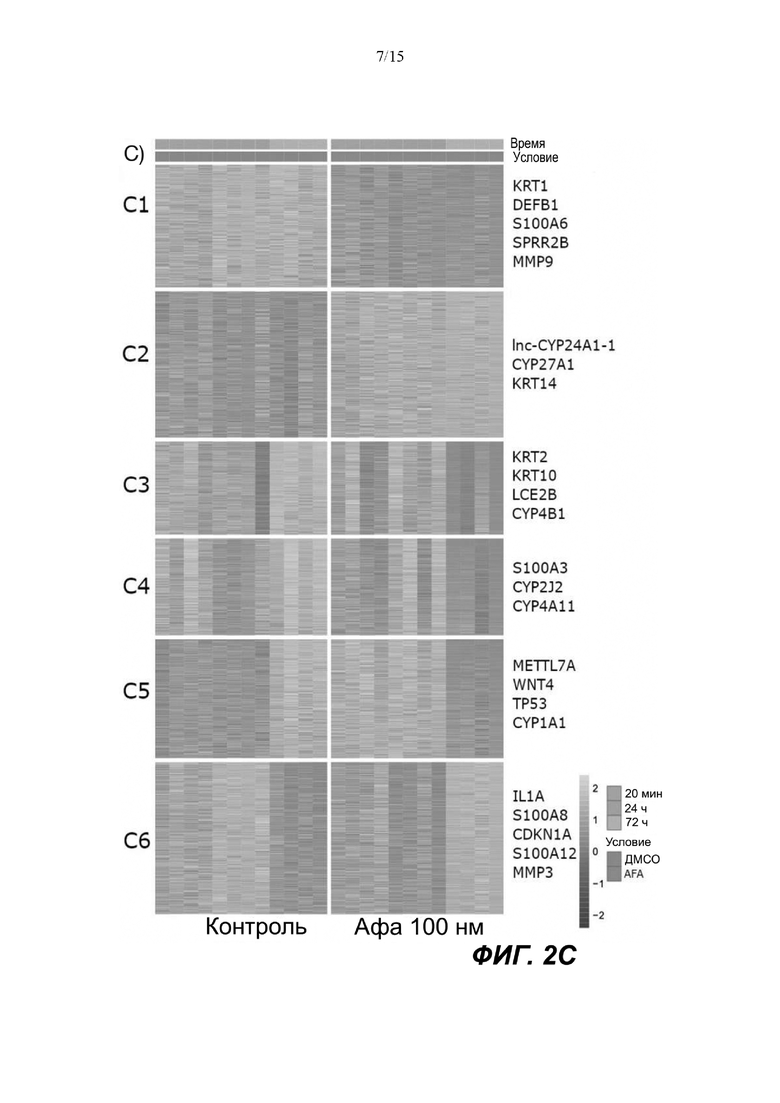
Фигуры: Кластеризация демонстрирует функциональное влияние афатиниба на дифференцировку кератиноцитов, окислительный стресс и врожденный иммунный ответ.
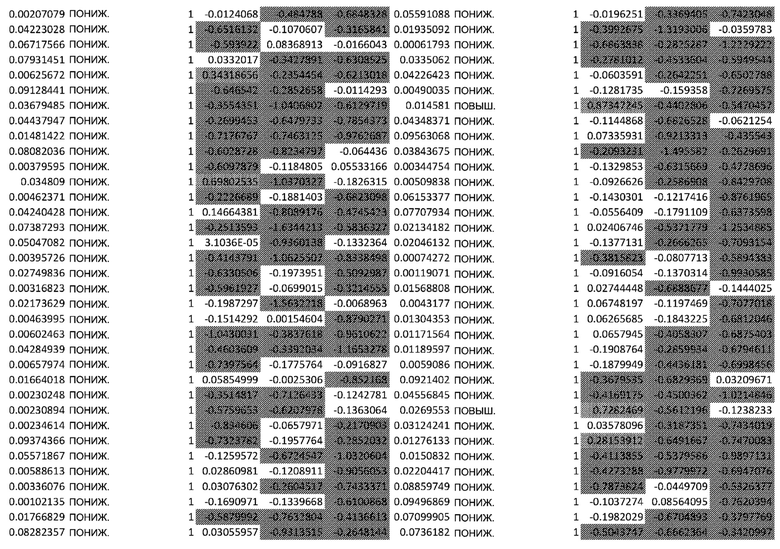
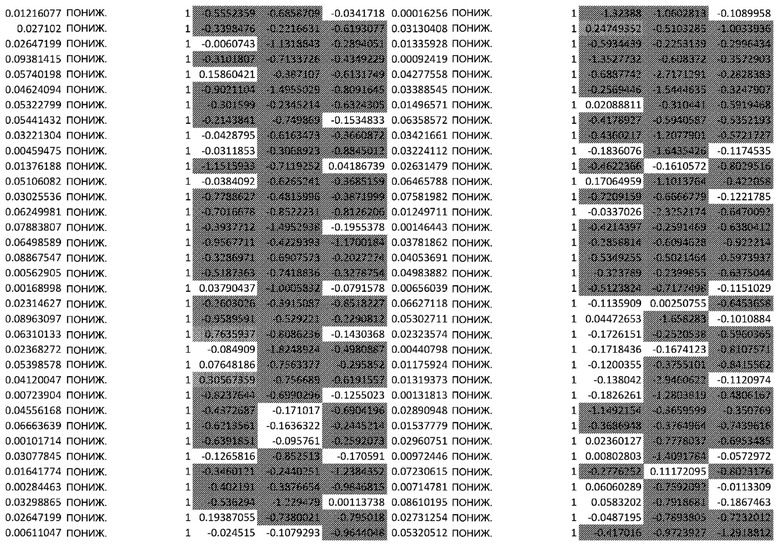
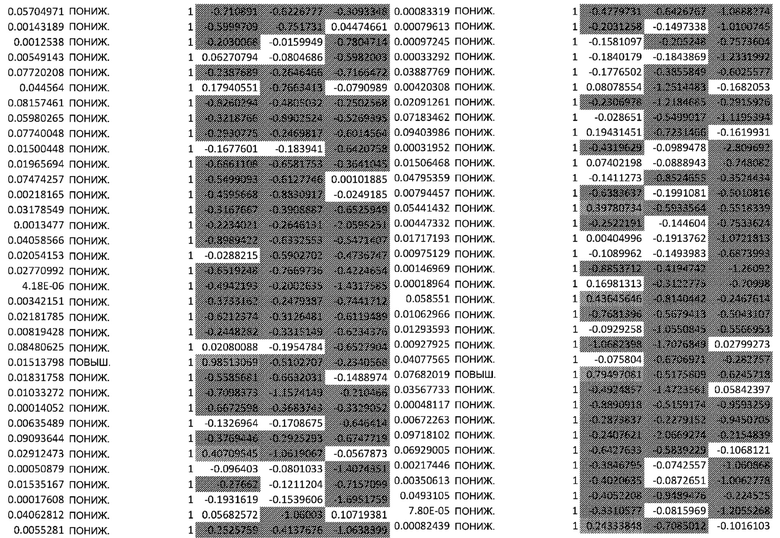
(A) Тепловая карта и Фигура 2B представляют собой диаграммы, изображающие результаты кластеризации k-средних на основе статуса фосфорилирования в динамике. Данные на тепловой карте выражены в виде Z-показателя строки. На диаграммах центральные линии изображают медиану, нижние и верхние края прямоугольников соответствуют первому и третьему квартилям. Верхний ус простирается от края прямоугольника до наибольшего значения вплоть до 1,5-кратного межквартильного размаха (IQR) от края, а нижний ус простирается от края прямоугольника до наименьшего значения при 1,5-кратном IQR края
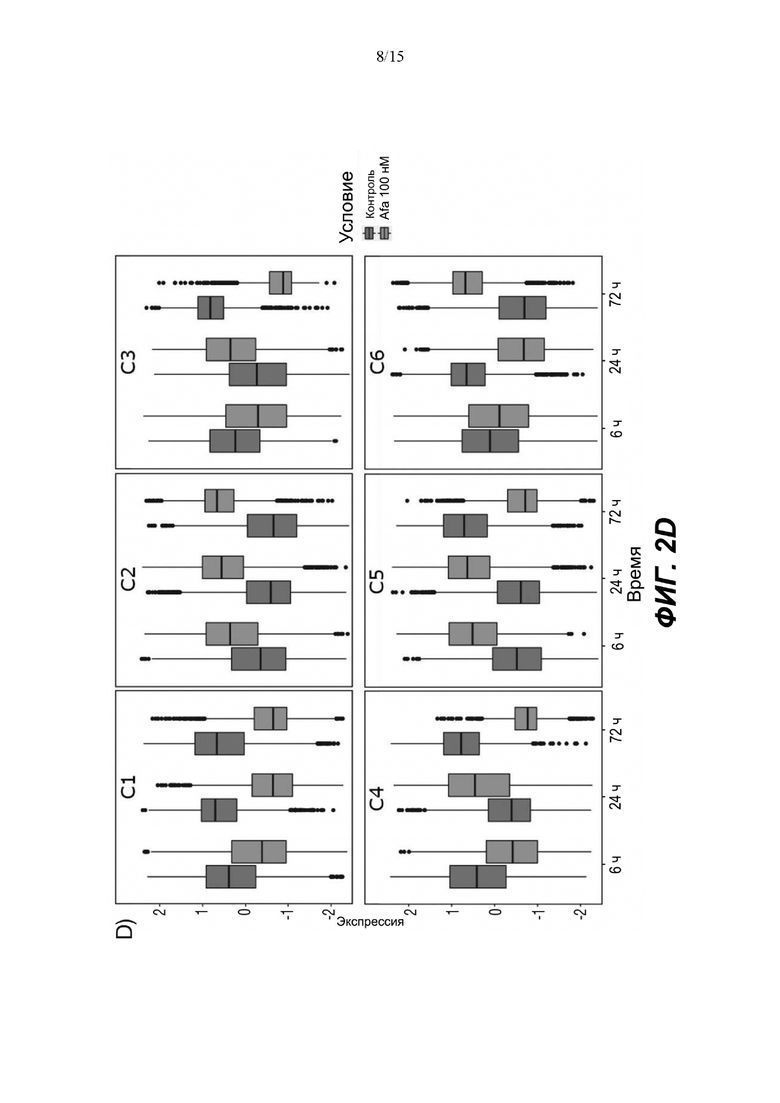
(C) Тепловая карта и диаграммы (D), изображающие результаты кластеризации k-средних на основе экспрессии генов в динамике. Гены были разделены на 6 кластеров, чтобы классифицировать изменение экспрессии генов с учетом лечения по сравнению с контролем. Данные на тепловой карте выражены в виде Z-показателя строки. На диаграммах центральные линии изображают медиану, нижние и верхние края прямоугольников соответствуют первому и третьему квартилям. Верхний ус простирается от края прямоугольника до наибольшего значения вплоть до 1,5-кратного межквартильного размаха (IQR) от края, а нижний ус простирается от края прямоугольника до наименьшего значения при 1,5-кратном IQR края
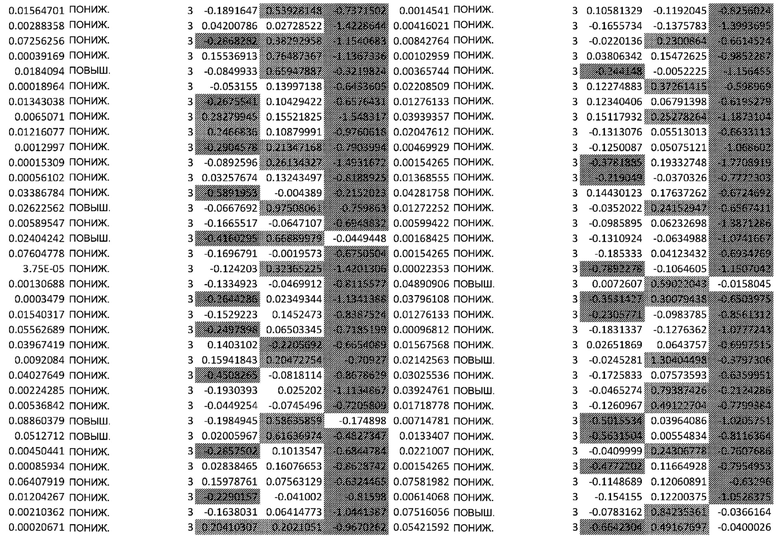
Фиг. 3: Анализ сверхрепрезентативности, основанный на KEGG, подтверждает функциональное влияние афатиниба на дифференцировку кератиноцитов, окислительный стресс и врожденный иммунный ответ.
Карты функциональной сверхрепрезентативности, изображающие наборы генов KEGG, ассоциированные с каждым кластером фосфорилирования (A) и транскриптомным кластером (B). Точки имеют цветовую кодировку в соответствии с скорректированным значением p FDR на основе гипергеометрического распределения. Размер пропорционален проценту генов в наборе генов, принадлежащих кластеру.
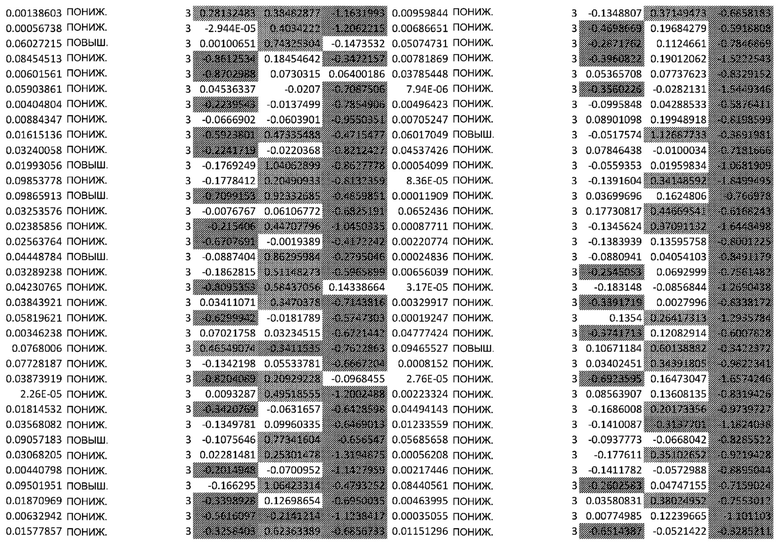
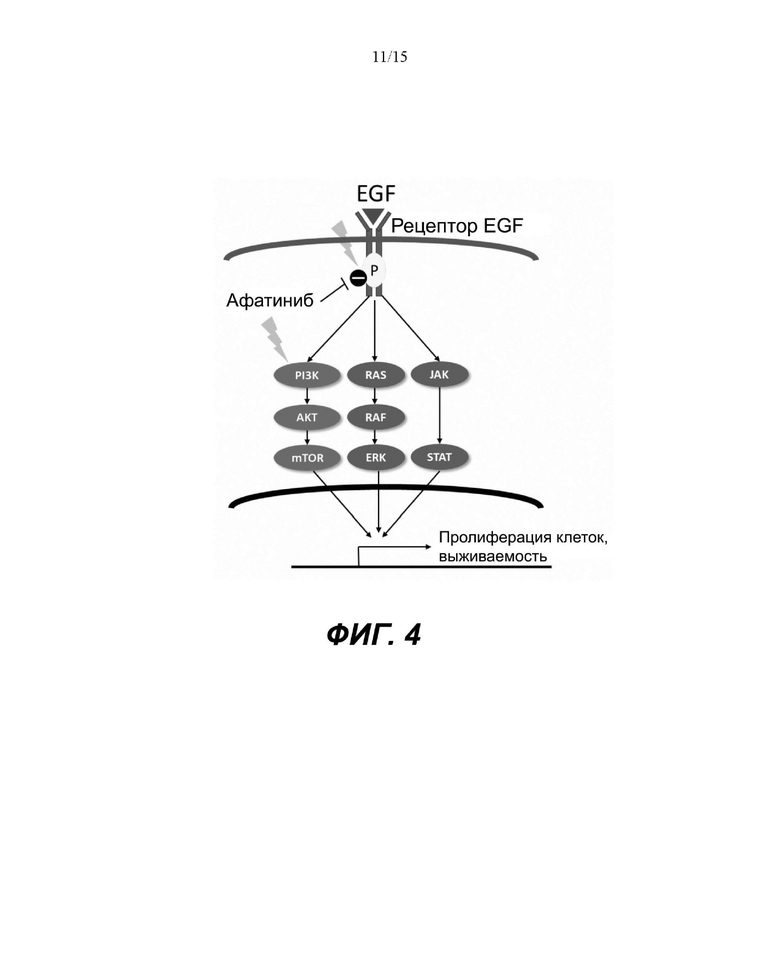
Фиг. 4: Сигнальный путь EGFR и его зависимости. Афатиниб ингибирует EGFR, но активирует путь PI3K/Akt.
Активация пути EGFR инициирует 3 разных пути: PI3K/Akt, RAS/RAF/ERK и STAT. При воздействии афатиниба на RHE кератиноциты, альтернативно усиливается путь PI3K/AKT. Следовательно, гены, расположенные ниже пути PI3K/AKT, участвующие в пролиферации, выживании и старении, подверглись воздействию, что привело к нарушению барьерной функции кожи.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Приведенные примеры демонстрируют, что статус фосфорилирования белка и экспрессия генов в кератиноцитах RHE динамически изменяются после воздействия EGFRi.
Микроматрицу фосфоантител применяли для выявления различий в статусе фосфорилирования белков из тканей RHE после воздействия 100 нМ афатиниба в течение 20 минут, 24 часов и 72 часов. В этом анализе было скринировано 1318 белков, включая 615 фосфопротеинов, и в каждый момент времени регистрировали изменения в их статусе фосфорилирования (дополнительная таблица S1). В общей сложности 62 фосфопротеина изменили свой статус фосфорилирования более чем на 20% (Фиг.1А). После воздействия афатиниба в течение 20 минут, 24 часов и 72 часов количество белков с повышенным статусом фосфорилирования по сравнению с контролем составило соответственно 6, 18 и 6, а количество белков с пониженным статусом фосфорилирования по сравнению с контролем составило соответственно 3, 23 и 6..
Неконтролируемую кластеризацию выполняли для классификации белков на кластеры (C) в зависимости от уровня изменения их статуса фосфорилирования и момента времени, когда указанное явление было обнаружено. Такая кластеризация обеспечивает более точное определение биологических функций, взаимодействий или путей, которые являются обогащенными. После определения оптимального количества кластеров, белки группировали в два кластера: первый (C1), представляющий белки с пониженным уровнями фосфорилирования в момент времени 24 ч, и второй (C2), представляющий белки с повышенным уровнем фосфорилирования в момент времени 24 ч (Фиг.2A и 2B).
Транскрипционный анализ RHE кератиноцитов, обработанных афатинибом, оценивали путем скрининга экспрессии 36 ООО генов и длинной некодирующей РНК (lncРНК) (дополнительная таблица S2). Отсечка на уровне 50% в кратности изменения экспресии выявила нарушение регуляции 2182 генов в разные моменты времени. После 6-часового воздействия 100 нМ афатиниба 170 генов были ингибированы по сравнению с контролем, а 201 ген был активирован. После 24-часового воздействия экспрессия 335 генов была сниженной, а экспрессия 281 гена - повышенной. После 72-часового воздействия экспрессия 2888 генов была сниженной, а экспрессия 1728 - повышенной (Фиг. 1B).
Неконтролируемую кластеризацию выполняли для транскриптомных данных, чтобы классифицировать гены по кластерам в зависимости от изменений уровней их экспрессии после воздействия афатиниба и момента времени, когда указанное явление было обнаружено. После определения оптимального количества кластеров данные были сгруппированы следующим образом (Фиг. 2C и 2D):
C1, представляющие ранние (20 минут) ингибированные гены, которые сохраняют свой статус во всех трех протестированных моментах времени; C2, представляющие ранние (20 минут) активированные гены, которые сохраняют свой статус во всех трех протестированных моментах времени; C3, представляющие временно активированные гены в момент времени 24 ч; C4, представляющие временно активированные гены в момент времени 24 ч с более высокой амплитудой по сравнению с C3; C5, представляющие ранние активированные гены с последующим ингибированием в момент времени 72 ч, и C6, представляющие ингибированные гены в момент времени 24 ч с последующим активированием в момент времени 72 ч.
Анализ матрицы фосфорилирования белков демонстрирует активацию пути PI3K/AKT с последствиями для клеточного старения.
Основываясь на изменениях статуса фосфорилирования, анализ обогащения набора (SEA) идентифицирует путь PI3K/AKT как основной обогащенный путь (Фиг.1C) и, более конкретно, путь старения (Фиг.1D). Белки, расположенные выше PI3K, с повышенным фосфорилированием включали RAC1 и FAK. Белки, расположенные ниже PI3K: p21 (CDKN1) участвует в развитии клеточного цикла, BAD - в процессе апоптоза, и p53 - в выживании клеток. Белки со сниженным фосфорилированием в основном являются ингибиторами, такие как PTEN, GSK3 и BCL2, и, следовательно, способствуют активации этого пути. Представляет интерес тот факт, что значительное количество регулируемых белков, расположенных ниже пути PI3K/AKT, участвует в клеточном старении путем активации FOXO3, семейства генов циклина (CYCB, CYCD, CYCE), а также пути mTOR и дефосфорилирования 4EBP1. Функциональный тест сверхрепрезентативности продемонстрировал повышенную активность белков, связанных с фосфорилированием AKT (Фиг. 3А).
Результаты транскриптомного анализа подтверждают активацию пути PI3K/AKT, на что указывает активация генов, связанных с этим путем (C6, Фиг. 3B).
Анализ матрицы экспрессии генов подчеркивает, что CYP является потенциальным индуктором окислительного стресса для дифференцировки кератиноцитов.
Как фосфорилирование белков, так и матрицы экспрессии генов продемонстрировали, что после воздействия афатиниба были затронуты семейства генов, участвующих в клеточном стрессе, окислительном стрессе и старении (Фиг. 3). Например, фосфорилирование FOXO3 снижалось вследствие активации пути PI3K/AKT. Фактор транскрипции FOXO3 влияет на способность клеток регулировать апоптоз и запрограммированную гибель клеток.
Афатиниб динамически влиял на экспрессию генов, связанных с окислительным стрессом. Большая панель металлотионеина (MT1E, MT1L, MT1HL1, MT1X, MT1B, MT1A) уменьшилась в момент времени 24 ч и увеличилась в момент времени 72 ч (Фиг. 3B). Семейства генов, участвующих в дыхательном транспорте электронов и терминации митохондриальной трансляции, были временно активированы в момент времени 24 ч (Фиг. 3В). В частности, экспрессия антиоксидантных ферментов PRDX2 и PRDX3 снижалась, а экспрессия SIRT4 увеличивалась в момент времени 72 ч (дополнительная таблица S2). Следует отметить, что экспрессия генов, относящихся к метаболизму жирных кислот, ассоциированных с окислительным стрессом, сначала повышалась в момент времени 24 ч и затем снижалась в момент времени 72 ч (Фиг. 3B).
Представляет интерес тот факт, что были также идентифицированы изменения экспрессии CYP в моменты времени 24 ч и 72 ч. Экспрессия CYP была повышена (lnc-CYP24A1-1, CYP27A1, CYP4F12), а экспрессия других CYP была снижена (CYP4B1, CYP3A7, CYP3A5, CYP2J2, CYP4A11, CYP1A1, CYP4F22) (дополнительная таблица S2). Экспрессия CYP27A1 может быть ассоциирована с транскриптомными данными, включающими биосинтез холестерина и метаболизм жирных кислот в кластере C1.
Помимо генов, связанных с окислительным стрессом, тест функциональной сверхрепрезентативности (Фиг.3) продемонстрировал обогащение путей кератинизации и ороговения, ассоциированное с повышением экспрессии генов KRT2, KRT9, KRT13, KRT15 и поздних генов ороговевающей оболочки (LCE2A, LCE1B, LCE2B, LCE1C, LCE2D) в момент времени 24 ч.
Воздействие афатиниба нарушало экспрессию маркеров врожденного иммунитета и усиливало выработку цитокинов, связанных с ответом Th2.
Путь PI3K/AKT непосредственно влияет как на врожденные, так и на адаптивный эпидермальный иммунитет.Анализ транскриптомных данных продемонстрировал, что гены, участвующие в врожденном иммунитете, отрицательно регулируются (дополнительная таблица S2), включая гены, относящиеся к семейству S100 (S100A2, S100A6, S100A7, S100A8, S100A9, S100A10, S100A12), семейству малых белков 2B, богатых пролином (SPRR2B), и семейству β-дефенсина B1 (DEFB1) (Фиг. 3A).
Затрагивалось семейство генов, ассоциированных с ответом Th2, о чем свидетельствует сверхрепрезентативность генов, связанных с передачей сигналов IL4 и IL13 (C2). Представляет интерес то, что эти результаты демонстрируют кинетику последствий после активации PI3K, которые включают первую активацию передачи сигналов TCR в момент времени 24 часа, а затем поляризацию Th2 в момент времени 72 часа.
С другой стороны, пути воспаления, ассоциированные с ответом Th1, почти не активировались. Транскриптомные данные не демонстрировали какого-либо повышения экспрессии генов, связанных с воспалением. Анализ фосфорилирования белка и матрица экспрессии генов продемонстрировали снижение воспалительного ответа через семейства TLR, NLR и IL1.
Обсуждение
Большинство клинически зарегистрированных кожных симптомов у пациентов, получавших низкомолекулярный EGFRi, включают сыпь (63% пациентов), ксероз (30%) и гранулемы (30%). Не сообщалось о связи между симптомами CADR и специфическими EGFRi, за исключением более частого появления гранулем у пациентов, получавших афатиниб (Annunziata et al. 2019). Эти данные подтверждают пригодность афатиниба в качестве всестороннего модельного препарата для изучения CADR.
Помимо лечения низкомолекулярными препаратами, существуют иммунотерапевтические препараты, нацеленные на внеклеточную часть EGFR. Хотя моноклональные антитела (mAB) проявляют меньше нецелевых эффектов, они все же могут вызывать тяжелые кожные высыпания (Tischer et al. 2017), которые обычно ассоциированы с активацией компонентов иммунной системы. Сосредоточив внимание на прямом влиянии онкологической терапии на физиологию эпидермиса и ее последствия на барьерную функцию кожи, в предложенной модели авторы намеренно убрали эффекты, связанные со сверхактивацией иммунной системы.
Более того, чтобы лучше сосредоточиться на эффектах афатиниба, помимо его известного действия как EGFRi, экзогенный EGF не был включен в модель. Присутствие EGF, ассоциированного с афатинибом, могло бы определить последствия приема препарата после EGFR.
Поддержание кератиноцитов в пролиферативном фенотипе требует особых условий культивирования клеток. Любая модификация клеточного метаболизма быстро вызывает дифференцировку кератиноцитов (Bakondi et al. 2003; Tsuchisaka et al. 2014; Vessey et al. 1995; Zhang et al. 2002). Воздействие афатиниба может влиять на формирование эпидермиса посредством одного или нескольких изменений метаболизма кератиноцитов.
Путь PI3K-AKT способствует старению кератиноцитов и подавляет пролиферацию кератиноцитов.
После воздействия афатиниба на кератиноциты наблюдалось значительное увеличение активности, связанной с путем PI3K/AKT. Это было неожиданно, поскольку этот путь находится ниже EGFR (Фиг.4) и, как ожидается, отрицательно регулируется посредством EGFRi.
Хотя путь PI3K/AKT активировался, путь RAF/RAS/ERK1/2, который является еще одним независимым путем после активации EGFR, не был положительно активирован после лечения афатинибом в момент времени 24 ч (Фиг.4) (Wee и Wang 2017). Фосфорилирование Raf1 (Phospho-Ser338), eIF4E (Phospho-Ser209) было снижено, а фосфорилирование Raf1 (Phospho-Tyr341), MEK1 (Phospho-Ser221), MEK1 (Phospho-Ser217) не было затронуто (дополнительная таблица S1). Эти результаты продемонстрировали ожидаемый эффект афатиниба после EGFR. Авторы предполагают, что активация PI3K/AKT была следствием альтернативного механизма.
Путь PI3K/AKT влияет на экспрессию генов кератиноцитов, связанную с ростом клеток, дифференцировкой клеток, старением и апоптозом, а также усилением окислительного стресса, приводящего к дифференцировке. Афатиниб влияет на формирование кожного барьера, вызывая воспаление и снижая врожденный иммунный ответ (Calautti et al. 2005; Janes et al. 2009). Результаты, полученные авторами, продемоснтрировали, что путь старения (Фиг.1D) был специфически активирован за счет участия белков PUK, C-Myc, p53 и mTOR. Известно, что повышенная активность в регуляции этих белков напрямую вызывает остановку клеточного цикла и способствует старению (Demideno et al. 2010; Iglesias-Barbolome et al. 2012).
Ранее авторы сообщили, что экспрессия маркеров эпидермальной дифференцировки повышалась после воздействия афатиниба (Joly-Tonetti et al. 2020). Представляет интерес тот факт, что путь PI3K/AKT регулирует увеличение филаггрина, лорикрина и кадгерин-ассоциированного катенина (Calautti et al. 2005), что подтверждает предыдущие наблюдения авторов.
Более того, активация пути PI3K/AKT способствует уменьшению прикрепления клеток, ведущему к апоптозу (Janes et al. 2009), и приводит к повышению экспрессии маркера дифференцировки и, следовательно, вызывает дифференцировку кератиноцитов. Экспрессия генов, ассоциированныхъ с терминальной дифференцировкой, временно повышалась в момент времени 24 ч и снижалась в момент времени 72 ч после воздействия афатиниба. Эти результаты указывают на то, что афатиниб вызывает быструю индукцию дифференцировки кератиноцитов и это может быть следствием повышенной активации PI3K.
Таким образом, активация пути PI3K/AKT способствует дифференцировке кератиноцитов и подавляет пролиферацию кератиноцитов. В некоторых случаях индукция апоптоза может быть связана с более тяжелыми состояниями, такими как синдром «рука-нога».
Афатиниб индуцирует клеточный окислительный стресс, который запускает дифференцировку кератиноцитов
При воздействии афатиниба характер экспрессии нескольких белков, расположенных ниже от AKT, был изменен, включая снижение экспрессии FOXO3A, белка, который, как известно, напрямую подавляется AKT (Brunet et al. 1999), и, следовательно, повышение экспрессии циклина D1 (CDKN1), который связан с окислительным стрессом (Marinkovic et al. 2007).
Белок р53 играет центральную роль в регуляции жизненного цикла кератиноцитов: он контролирует скорость клеточной пролиферации и способствует дифференцировке, а не апоптозу. Активный p53 запускает экспрессию хорошо известных мишеней, способствующих старению. Белок p21 контролирует пути, связанные с возрастными изменениями и старением клетки (Hergig et al. 2008). В связи с этим повышенные уровни p53 способны ограничивать окислительное повреждение и участвовать в активности, способствующей апоптозу и старению (Rufini et al. 2013). Тем не менее, микроматричные результаты фосфо-антител, полученные авторами, не демонстрируют увеличения активности каспазы 3 (дополнительная таблица S1) в любой момент времени, что также подтверждает предыдущие результаты авторов (Joly-Tonetti et al. 2020), согласно которым CADR от легкой до умеренной степени не ассоциируются с апоптозом кератиноцитов. Однако при более тяжелых состояниях, таких как TEN, активация p53 может быть причиной более серьезных симптомов (Ido Y. 2015).
EGFRi влияет на метаболизм витамина D3 в кератиноцитах
В присутствии EGF, афатиниб необратимо ингибирует внутриклеточную передачу сигналов EGFR путем ковалентного связывания с Cys797 EGFR, Cys805 HER2 и Cys803 ErbB-4 (Solca et al. 2012). Нецелевая активность препарата уже была продемонстрирована за счет неспецифического ковалентного связывания (например, с комплексами CDK), приводящего к нарушению клеточных процессов (Klaeger et al. 2017). Эти результаты предполагают, что ингибиторы ковалентных киназ могут перекрестно реагировать, либо специфически, либо неспецифически, с белками вне их кинома. Такая активность усложняет присвоение биологических функций киназам в химико-биологических экспериментах и может привести к непредвиденной токсичности, вызывая апоптоз и/или старение (Lanning et al. 2014). Было продемонстрировано, что нецелевое связывание TKi усиливает метаболизм кальция и способствует дифференцировке кератиноцитов (Kroschwald et al. 2018).
Параллельная активация метаболизма витамина D3 (VD3) может, следовательно, влиять на уровень внеклеточного кальция, вызывая дифференцировку кератиноцитов (Teichert и Bikle 2011). Недавнее исследование (Shaurova et al. 2020) продемонстрировало прямое влияние EGFRi на 1,25-дигидроксивитамин D3 (1,25(OH)2D3) (1,25(OH)2VD3), метаболит VD3, участвующий в дифференцировке эпителия.
Полученные авторами результаты демонстрируют, что после воздействия афатиниба изменялась экспрессия нескольких CYP. К ним относятся CYP27A1, участвующие в метаболизме VD3 и расщеплении холестерина до желчных кислот как классическим, так и кислотным путем. Предыдущее исследование (Kroschwald et al. 2018) продемонстрировало, что TKi напрямую модулирует активность CYP24A1. Как следствие, происходит изменение метаболизма кальция, которое может повлиять на дифференцировку кератиноцитов (Elsholz et al., 2014).
Более того, низкие уровни ключевого фермента, катаболизирующего витамин D, 24-гидроксилазы, коррелируют с экспрессией гена CYP24A1 (Shaurova et al. 2020). Представляет интерес тот факт, что полученные авторами результаты демонстрируют увеличение lnc-CYP24A1 и, следовательно, предполагают снижение экспрессии CYP24A1. Было продемонстрировано, что оба этих CYP индуцируют модификацию метаболизма кальция (Dinour et al. 2013). Кроме того, 1,25(OH)2D3 индуцировал реакции транскрипции, включающие повышенные уровни mРНК CYP24A1, хорошо охарактеризованной непосредственной мишени рецептора витамина D (VDR) (Shavurova et al. 2020). В совокупности эти результаты продемонстрировали повышенную экспрессию 1,25(OH)2D3, ассоциированную с активацией CYP24A1, обе из которых способствуют дифференцировке эпителия.
Известно, что 1,25(OH)2D3 индуцирует транскрипцию множественных молекул клеточной адгезии (Pálmer et al., 2001) и подтверждает полученные авторами предыдущие результаты, демонстрирующие, что афатиниб влияет на размер и объем эпидермиса и увеличивает экспрессию маркеров дифференцировки (инволюкрина, десмоглеина и филаггрина) (Joly-Tonetti et al. 2020). Известно, что в отношении окислительного стресса, VD3 является мощным индуктором металлотионеина (МТ), способного захватывать вредные окислители, такие как супероксидные и гидроксильные радикалы (Nzengen et al. 2008) (Фиг.3B). МТ может действовать как поглотитель радикалов в опосредованной кислородом активности CYP.
Интересно то, что исследования продемонстрировали синергизм нарушения регуляции пути PI3K/AKT с антипролиферативной передачей сигналов VD3, вызывающий старение клеток (Axanova et al. 2010; da Silva Teixeira et al. 2020), и подтвердили то, что как вовлечение VD3, так и путь старения клеток являются потенциальными механизмами CADR.
Известно, что TKi в целом и афатиниб в частности вызывают фотосенсибилизацию (Dai et al. 2017). Весьма вероятно, что воздействие солнечного UV-излучения может взаимодействовать с такой терапией, влияя на метаболизм VD3 и вызывая повышенную фоточувствительность кожи.
Иммунные изменения после лечения EGFRi ухудшают защиту кожного барьера
Механизмы, посредством которых терапия TKi влияет на иммунный ответ, до сих пор плохо изучены. В этой работе, анализы экспрессии белков и генов продемонстрировали вовлечение Th2-ответа в результате комбинированного повышения экспрессии связанных генов IL-4 и IL-13, а также снижение как Th1-ответа, так и врожденного иммунного ответа через белки S100 и инфламмасому (Howell et al. 2008). Представляет интерес тот факт, что такие же изменения наблюдаются и при атопическом дерматите. (Howell et al. 2008). Более того, нарушение регуляции метаболизма VD3 негативно влияет на иммунный ответ в кератиноцитах (Kroschwald et al. 2018).
Нарушение функции эпидермиса, ассоциированное с нарушением врожденного иммунного ответа, повышает восприимчивость лиц к рецидивирующим бактериальным и вирусным инфекциям кожи, которые часто наблюдаются в клинической практике. Применение тетрациклина для компенсации нарушенного врожденного иммунного ответа после воздействия афатиниба продемонстрировало улучшение симптомов CADR (Arrieta et al. 2015).
Эти новые маркеры могут быть полезны при разработке более эффективных стратегий предотвращения поражения кожи в результате применения противораковых препаратов.
В заключение, эта работа демонстрирует, что путь PI3K/AKT играет ключевую роль в патогенезе CADR, быстро стимулируя дифференцировку кератиноцитов. Этот механизм может объяснить проявление симптомов CADR. Эти результаты также указывают на то, что VD3 играет важную роль в CADR. Известно, что TKi влияет на метаболизм VD3. Нарушение регуляции пути VD3 влияет на дифференцировку кератиноцитов за счет изменений активности внутриклеточного кальция, повышенной экспрессии дифференцировочного белка, снижения врожденного ответа и сверхэкспрессии CYP21A1. В совокупности эти результаты дают новое молекулярное понимание механизмов CADR, которые могут быть полезны в клинической диагностике, а также для оценки эффективности протокола лечения CADR.
Материалы и методы
Приготовление лекарственного средства
Концентрацию афатиниба, применяемую в экспериментах, выбирали на субцитотоксическом уровне 100 нМ, как сообщалось ранее (Joly-Tonetti et al. 2020). Афатиниб приобретали у компании Caymanchem (Анн-Арбор, штат Мичиган, США) и готовили из 10 мМ исходного раствора, растворенного в DMSO (Sigma, Сен-Кантен-Фаллавье, Франция). Следовательно, конечная концентрация DMSO составляла 0,001% для 100 нМ. Контроль состоял из того же объема DMSO, что и раствор афатиниба.
Реконструированная модель эпидермиса человека
Крупные 4 см² RHE приобретали у компании Episkin (Лион, Франция) и культивировали в течение 24 ч без EGF перед воздействием афатиниба. Обработку RHE проводили в среде Epilife без EGF и без белков (MEPI500CA, Thermo Fisher Scientific, Уолтем, штат Массачусетс, США). Короткое, промежуточное и длительное время воздействия лекарственного средства были выбраны соответственно как 20 минут, 24 часа, 72 часа для анализа фосфорилирования белка и 6 часов, 24 часа и 72 часа для анализа профиля гена. Для всех моментов времени и условий выполняли четыре повтора.
Микроматрицы антител для фосфорилирования белка и профилирования экспрессии
Матрица антител Phospho Explorer (Full Moon BioSystems, Inc., Саннивейл, штат Калифорния, США) была изготовлена компанией TebuBio (Ле-Перре-ан-Ивлин, Франция) с применением 1330 дублирующих пятен, соответствующих 1318 белкам (фосфорилированным и нефосфорилированным); были проанализированы белки «домашнего хозяйства» (бета-актин, GAPDH), отрицательные контроли (n=4), пустые пятна (n=4) и положительные маркеры (n=2). Вкратце, RHE промывали холодным PBS и хранили непосредственно при -80°C Образцы обрабатывали согласно протоколу производителя. Образцы объемом 60 мкг инкубировали с биотином в течение 2 часов с последующей инкубацией в течение 30 минут в реагенте Stop (Full Moon BioSystems, Саннивейл, штат Калифорния, США). Мембраны, содержащие отпечатанные антитела, блокировали в течение 40 мин блокирующим реагентом при перемешивании, затем промывали и инкубировали в камере сопряжения в течение 2 ч. После удаления образца и трех последовательных промывок этап детектирования выполняли после добавления 30 мкл Cy3-стрептавидина (1 мг/мл) в течение 20 мин инкубации при комнатной температуре в темноте. Затем образцы промывали, сушили центрифугированием и сканировали на микроматричном сканере (Innopsys Innoscan 710).
Профилирование экспрессии генов
Транскриптомный анализ выполнялся компанией Genex (Лонжюмо, Франция). RHE удаляли с опорной поликарбонатной мембраны и немедленно переносили в RLT-буфер, содержащий 1% β-меркаптоэтанола. Поликарбонатные мембраны соскребали для удаления прикрепленных оставшихся базальных клеток и переносили в соответствующие флаконы, содержащие RHE. Образцы хранили при температуре -80°C.
Общую РНК, включая miРНК, выделяли с помощью универсального набора AllPrep DNA/RNA/miRNA от компании Qiagen (Хильден, Германия). Качество РНК оценивали с помощью системы Experion (Biorad, Марн-ла-Кокетт, Франция). Все показатели качества РНК были выше 7. Количество 100 нг общей РНК транскрибировали и окрашивали цианином 3 (Cy3) с помощью набора RNA Low Input Quick Amp Labeling Kit One Color (Agilent Technologies, Лез-Юлис, Франция) в соответствии с инструкциями производителя. Вся специфическая активность составляла выше 6 пмоль Cy3/мкг cРНК, а выход - выше 1,65 мкг.Равное количество (600 нг) cРНК, меченной Cy3, фрагментировали и затем гибридизовали в течение 17 часов при 65°C на микроматрице SurePrint 8x60K v3 человека (Agilent Technologies). Затем микроматрицы промывали и сканировали в соответствии с инструкциями производителя с помощью сканера G2505C (Agilent Technologies). Данные экспрессии генов дополнительно обрабатывали с помощью программного обеспечения Feature Extraction (версия 10.7).
Обработка данных
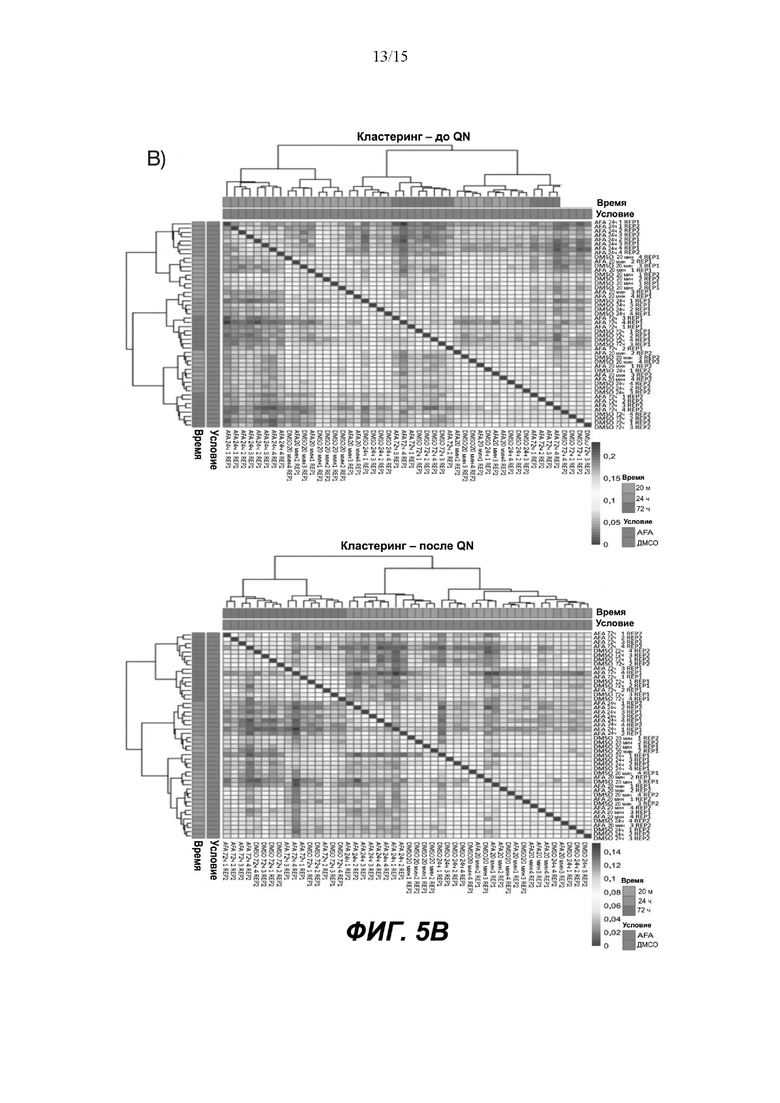
Как для фосфорилирования белков, так и для данных матрицы транскриптомов выполняли коррекцию фона с последующей квантильной нормализацией (QN) с применением каркаса RMA, реализованного в пакете limma. Эта процедура направлена на нормализацию медианной экспрессии между матрицами (дополнительные Фигуры 5A-5D).
Проверку качества выполняли на предмет объективной оценки влияния QN и фильтрации с помощью графической визуализации данных до и после нормализации, ассоциированной с блочной диаграммой, неконтролируемой кластеризацией, анализом основных компонентов и многомерным масштабированием (дополнительные Фигуры 5A-5D).
Вулканические графики были определены на основе значительной дифференциальной экспрессии -log10 преобразованных значений q после применения 10% ложного обнаружения с последующим пост-анализом Бенджамини-Хохберга (ось Y). По оси X показаны различия между двумя группами (афатиниб по сравнению с контролем). Значения фосфорилирования белков и экспрессии генов были преобразованы в log2 для обеспечения нормального распределения.
Чтобы идентифицировать сайты фосфорилирования и гены, на которые воздействует афатиниб в любой момент времени, был проведен дифференциальный анализ, основанный на общей линейной модели, с учетом времени и взаимодействия лечения с применением пакета limma.
Затем был проведен анализ обогащения наборов генов и белков (SEA) для выявления классов генов или белков, которые являются сверхрепрезентативными в наборах генов, которые регулируются как на повышение, так и на понижение, и могут иметь связь с фенотипами заболеваний. Для определения основного пораженного пути выполняли анализ путей с применением базы данных «Bioto энциклопедия генов и геномов» (Bioto Encyclopedia of Genes и Genomes, KEGG) на основе SEA со значительно измененными сайтами фосфорилирования и экспрессией генов с p<0,05.
Доступность данных
Данные микроматриц являются общедоступными из базы данных экспрессии генов Omnibus (https://www.ncbi.nlm.nih.gov/ge/).
Следует понимать, что хотя различные аспекты настоящего описания были проиллюстрированы и описаны с помощью примеров, заявляемое в настоящем документе изобретение ими не ограничивается, но может быть реализовано иными различными способами в соответствии с объемом формулы изобретения, включенной в настоящую и/или любую родственную настоящей заявку на патент.
KO
Ортологи KEGG
QN
Нормализация количества
Ссылки
Ali R, Brown W, Purdy SC, Davisson VJ, Wendt MK. Biased signaling downstream of epidermal growth factor receptor regulates proliferative versus apoptotic response to ligand. Cell Death Dis. Springer US; 2018;9(10) Available from: http://dx.doi.org/10.1038/s41419-018-1034-7
Annunziata MC, Ferrillo M, Cinelli E, Panariello L, Rocco D, Fabbrocini G. Retrospective analysis of skin toxicity in patients under anti-EGFR tyrosine kinase inhibitors: Our experience in lung cancer. Open Access Maced. J. Med. Sci. 2019;7(6):973-7
Arrieta O, Vega-González MT, López-Macías D, Martínez-Hernández JN, Bacon-Fonseca L, Macedo-Pérez EO, et al. Randomized, open-label trial evaluating the preventive effect of tetracycline on afatinib induced-skin toxicities in non-small cell lung cancer patients. Lung Cancer. Elsevier Ireland Ltd; 2015;88(3):282-8
Axanova LS, Chen YQ, McCoy T, Sui G, Cramer SD. 1,25-dihydroxyvitamin D3 and PI3K/AKT inhibitors synergistically inhibit growth and induce senescence in prostate cancer cells. Prostate. NIH Public Access; 2010;70(15):1658-71
Bakondi E, Gönczi M, Szabó É, Bai P, Pacher P, Gergely P, et al. Role of intracellular calcium mobilization and cell-density-dependent signaling in oxidative-stress-induced cytotoxicity in HaCaT keratinocytes. J. Invest. Dermatol. Blackwell Publishing Inc.; 2003;121(1):88-95
Bieber T. Atopic dermatitis. N. Engl. J. Med. Massachussetts Medical Society; 2008. p.1483
Calautti E, Li J, Saoncella S, Brissette JL, Goetinck PF. Phosphoinositide 3-kinase signaling to Akt promotes keratinocyte differentiation versus death. J. Biol. Chem. 2005;280(38):32856-65
Cheng JB, Levine MA, Bell NH, Mangelsdorf DJ, Russell DW. Genetic evidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase. Proc. Natl. Acad. Sci. U. S. A. National Academy of Sciences; 2004;101(20):7711-5
Dai J, Belum VR, Wu S, Sibaud V, Lacouture ME. Pigmentary changes in patients treated with targeted anticancer agents: A systematic review and meta-analysis. J. Am. Acad. Dermatol. Elsevier Inc; 2017;77(5):902-910.e2 Available from: http://dx.doi.org/10.1016/j.jaad.2017.06.044
Demidenko ZN, Korotchkina LG, Gudkov A V., Blagosklonny M V. Paradoxical suppression of cellular senescence by p53. Proc. Natl. Acad. Sci. U. S. A. National Academy of Sciences; 2010;107(21):9660-4
Dinour D, Beckerman P, Ganon L, Tordjman K, Eisenstein Z, Holtzman EJ. Loss-of-function mutations of CYP24A1, the vitamin D 24-hydroxylase gene, cause long-standing hypercalciuric nephrolithiasis and nephrocalcinosis. J. Urol. J Urol; 2013;190(2):552-7
Eckert RL, Efimova T, Dashti SR, Balasubramanian S, Deucher A, Crish JF, et al. Keratinocyte survival, differentiation, and death: Many roads lead to mitogen-activated protein kinase. J. Investig. Dermatology Symp.Proc. Blackwell Publishing Inc.; 2002. p.36-40
Elsholz F, Harteneck C, Muller W, Friedland K. Calcium - A central regulator of keratinocyte differentiation in health and disease. Eur. J. Dermatology. 2014. p.650-61
Herbig U, Wei W, Dutriaux A, Jobling WA, Sedivy JM. Real-time imaging of transcriptional activation in live cells reveals rapid up-regulation of the cyclin-dependent kinase inhibitor gene CDKN1A in replicative cellular senescence. Aging Cell. Wiley-Blackwell; 2008;2(6):295-304 Available from: http://doi.wiley.com/10.1046/j.1474-9728.2003.00067.x
Howell MD, Fairchild HR, Kim BE, Bin L, Boguniewicz M, Redzic JS, et al. Th2 cytokines act on S100/A11 to downregulate keratinocyte differentiation. J. Invest. Dermatol. Elsevier
Masson SAS; 2008;128(9):2248-58 Available from: http://dx.doi.org/10.1038/jid.2008.74
Iglesias-Bartolome R, Patel V, Cotrim A, Leelahavanichkul K, Molinolo AA, Mitchell JB, et al. MTOR inhibition prevents epithelial stem cell senescence and protects from radiationinduced mucositis. Cell Stem Cell. NIH Public Access; 2012;11(3):401-14
Janes SM, Ofstad TA, Campbell DH, Eddaoudi A, Warnes G, Davies D, et al. PI3-kinasedependent activation of apoptotic machinery occurs on commitment of epidermal keratinocytes to terminal differentiation. Cell Res. 2009;19(3):328-39
Jänicke RU, Sprengart ML, Wati MR, Porter AG. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. American Society for Biochemistry and Molecular Biology; 1998;273(16):9357-60
Joly-Tonetti N, Ondet T, Monshouwer M, Stamatas GN. EGFR inhibitors switch keratinocytes from a proliferative to a differentiative phenotype affecting epidermal development and barrier function. (Under review 2020).
Klaeger S, Heinzlmeir S, Wilhelm M, Polzer H, Vick B, Koenig P-A, et al. The target landscape of clinical kinase drugs. Science. 2017;358(6367) Available from: http://www.ncbi.nlm.nih.gov/pubmed/29191878
Kroschwald L, Suttorp M, Tabeatauer J, Zimmermann N, Günther C, Bauer A. Off-target effect of imatinib and nilotinib on human Vitamin D3 metabolism. Mol. Med. Rep.Spandidos Publications; 2018;17(1):1382-8
Lacouture ME. Mechanisms of cutaneous toxicities to EGFR inhibitors. Nat. Rev. Cancer. 2006;6(10):803-12
Lanning BR, Whitby LR, Dix MM, Douhan J, Gilbert AM, Hett EC, et al. A road map to evaluate the proteome-wide selectivity of covalent kinase inhibitors. Nat. Chem. Biol. 2014;10(9):760-7
Li D, Ambrogio L, Shimamura T, Kubo S, Takahashi M, Chirieac LR, et al. BIBW2992, an irreversible EGFR/HER2 inhibitor highly effective in preclinical lung cancer models. Oncogene. NIH Public Access; 2008;27(34):4702-11
Lippens S, Denecker G, Ovaer P, Vandenabeele P, Declercq W. Death penalty for keratinocytes: Apoptosis versus cornification. Cell Death Differ. Nature Publishing Group; 2005;12(2):1497-508
Marinkovic D, Zhang X, Yalcin S, Luciano JP, Brugnara C, Huber T, et al. Foxo3 is required for the regulation of oxidative stress in erythropoiesis. J. Clin. Invest. American Society for Clinical Investigation; 2007;117(8):2133-44
Mehlig LM, Garve C, Tauer JT, Suttorp M, Bauer A. Inhibitory effects of imatinib on vitamin D3 synthesis in human keratinocytes. Mol. Med. Rep.Spandidos Publications; 2015;11(4):3143-7
Missero C, Di Cunto F, Kiyokawa H, Koff A, Dotto GP. The absence of p21(Cip1/WAF1) alters keratinocyte growth and differentiation and promotes ras-tumor progression. Genes Dev. Cold Spring Harbor Laboratory Press; 1996;10(23):3065-75
Norlin M, Von Bahr S, Björkhem I, Wikvall K. On the substrate specificity of human CYP27A1: Implications for bile acid and cholestanol formation. J. Lipid Res. 2003;44(8):1515-22 Nzengue Y, Steiman R, Garrel C, Lefèbvre E, Guiraud P. Oxidative stress and DNA damage induced by cadmium in the human keratinocyte HaCaT cell line: Role of glutathione in the resistance to cadmium. Toxicology. Elsevier; 2008;243(1-2):193-206
Pálmer HG, González-Sancho JM, Espada J, Berciano MT, Puig I, Baulida J, et al. Vitamin D3 promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of β-catenin signaling. J. Cell Biol. J Cell Biol; 2001;154(2):369-87
Rufini A, Tucci P, Celardo I, Melino G. Senescence and aging: The critical roles of p53. Oncogene. Nature Publishing Group; 2013;32(43):5129-43 Available from: www.nature.com/onc
Shaurova T, Dy GK, Battaglia S, Hutson A, Zhang L, Zhang Y, et al. Vitamin d3 metabolites demonstrate prognostic value in EGFR-mutant lung adenocarcinoma and can be deployed to oppose acquired therapeutic resistance. Cancers (Basel). 2020;12(3) Available from: www.mdpi.com/journal/cancers
da Silva Teixeira S, Harrison K, Uzodike M, Rajapakshe K, Coarfa C, He Y, et al. Vitamin D actions in neurons require the PI3K pathway for both enhancing insulin signaling and rapid depolarizing effects. J. Steroid Biochem. Mol. Biol. Elsevier Ltd; 2020;200:105690
Solca F, Dahl G, Zoephel A, Bader G, Sanderson M, Klein C, et al. Target binding properties and cellular activity of afatinib (BIBW 2992), an irreversible ErbB family blocker. J. Pharmacol. Exp.Ther. American Society for Pharmacology and Experimental Therapeutics; 2012;343(2):342-50
Teichert A, Bikle DD. Regulation of keratinocyte differentiation by vitamin D and its relationship to squamous cell carcinoma. Signal. Pathways Squamous Cancer. Springer NewYork; 2011. p.283-303
Tischer B, Huber R, Kraemer M, Lacouture ME. Dermatologic events from EGFR inhibitors: the issue of the missing patient voice. Support. Care Cancer. Supportive Care in Cancer; 2017;25(2):651-60 Available from: http://dx.doi.org/10.1007/s00520-016-3419-4
Tsuchisaka A, Furumura M, Hashimoto T. Cytokine regulation during epidermal differentiation and barrier formation. J. Invest. Dermatol. Nature Publishing Group; 2014. p.1194-6
Vessey DA, Lee KH, Boyer TD. Differentiation-induced enhancement of the ability of cultured human keratinocytes to suppress oxidative stress. J. Invest. Dermatol. Nature Publishing Group; 1995;104(3):355-8
Vivanco I, Sawyers CL. The phosphatidylinositol 3-kinase-akt pathway in human cancer. Nat. Rev. Cancer. 2002;2(7):489-501
Wee P, Wang Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers (Basel). MDPI AG; 2017. p.52
Whitman M, Downes CP, Keeler M, Keller T, Cantley L. Type I phosphatidylinositol kinase makes a novel inositol phospholipid, phosphatidylinositol-3-phosphate. Nature. Nature; 1988;332(6165):644-6
Zhang T, Woods TL, Elder JT. Differential responses of S100A2 to oxidative stress and increased intracellular calcium in normal, immortalized, and malignant human keratinocytes. J. Invest. Dermatol. Blackwell Publishing Inc.; 2002;119(5):1196-201
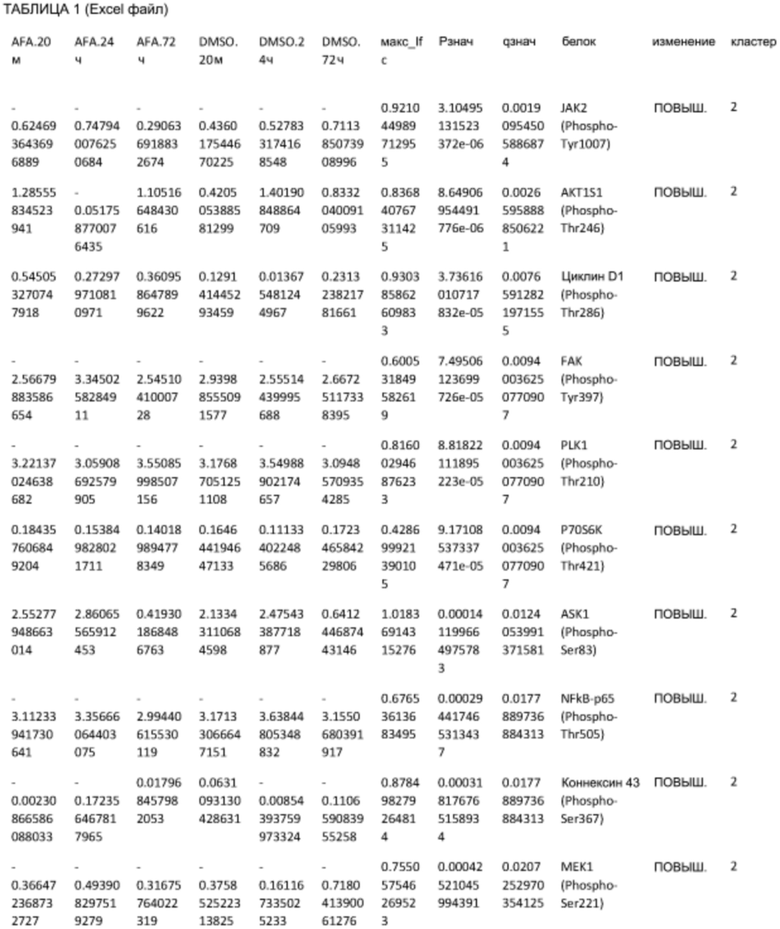
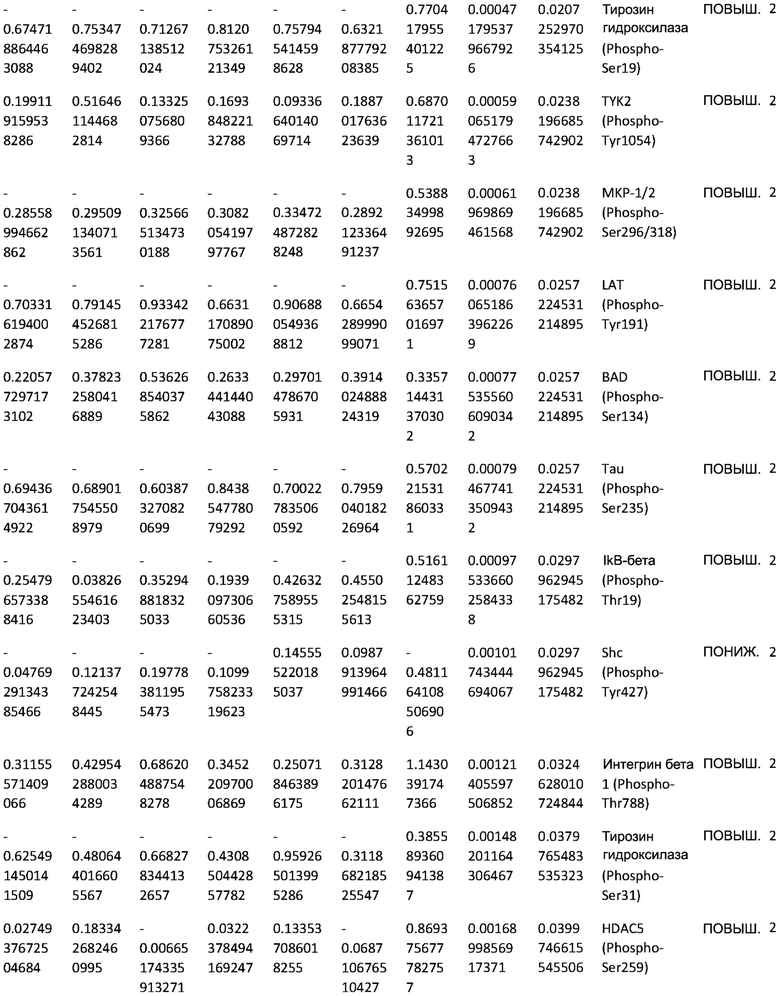
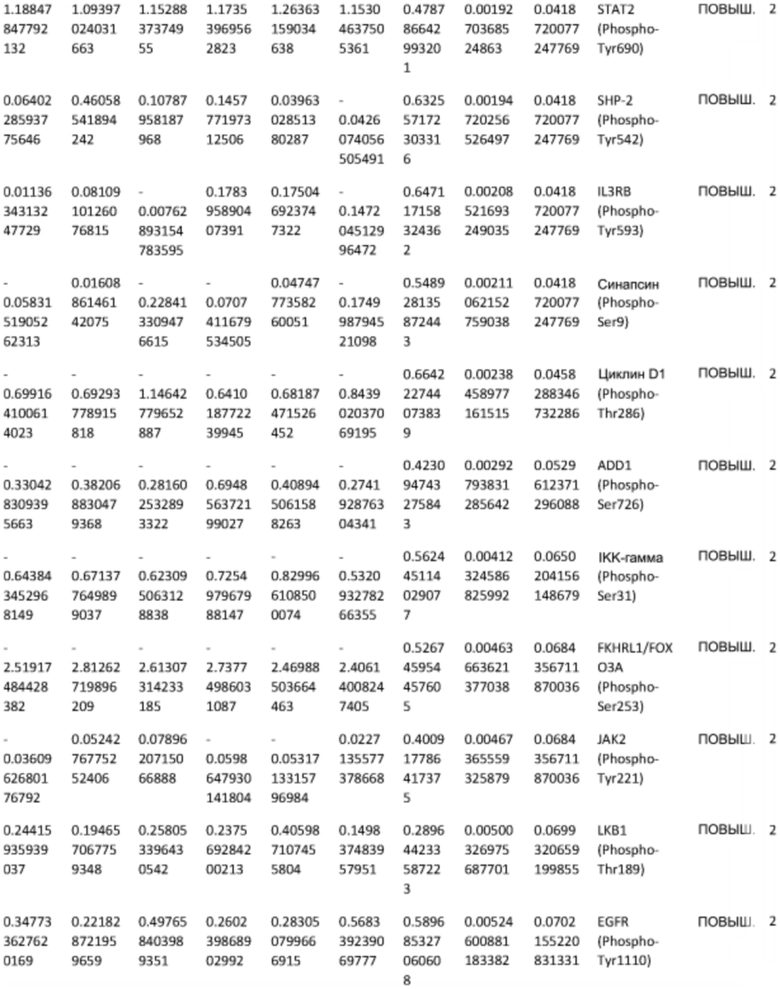
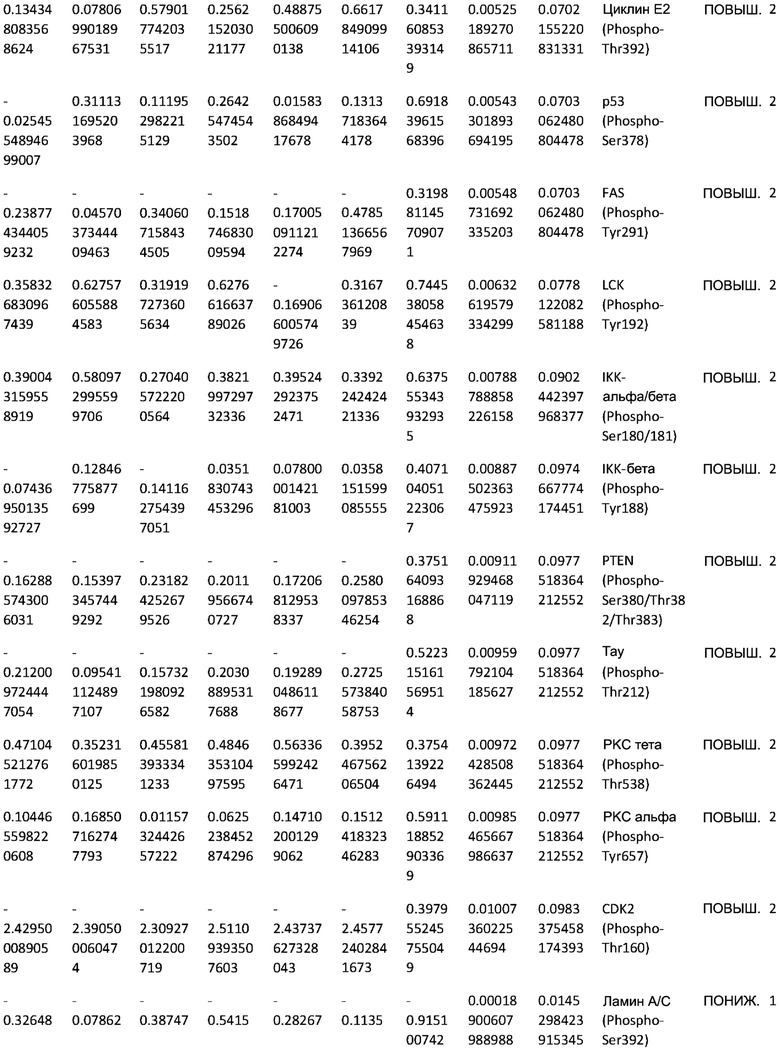
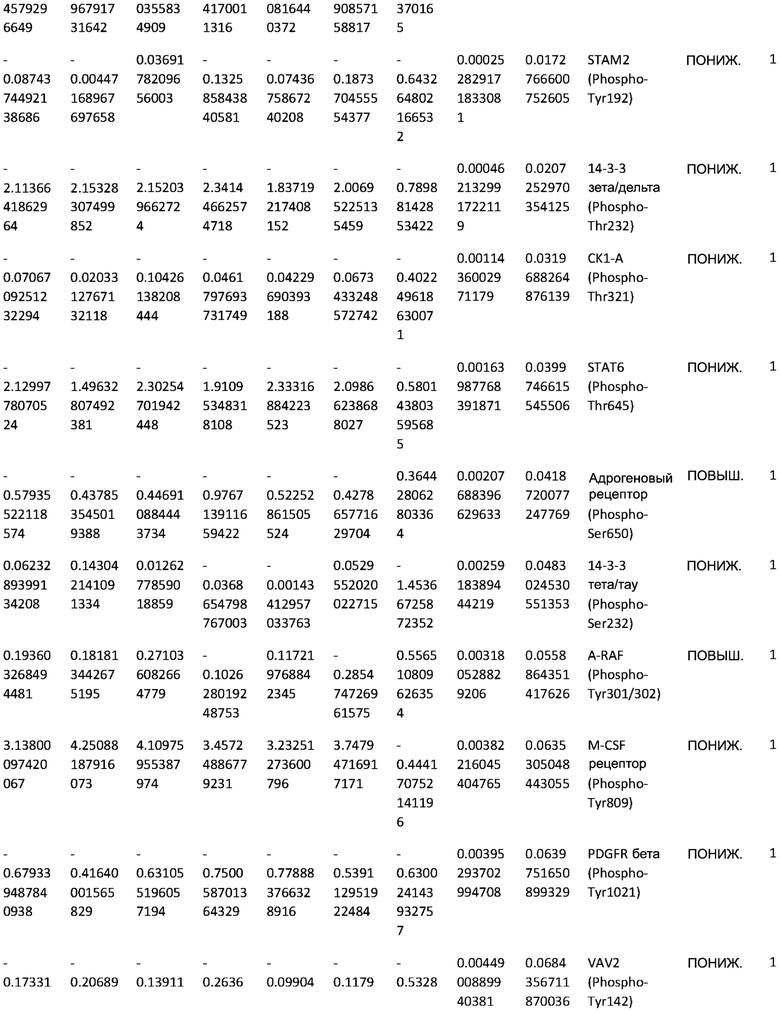
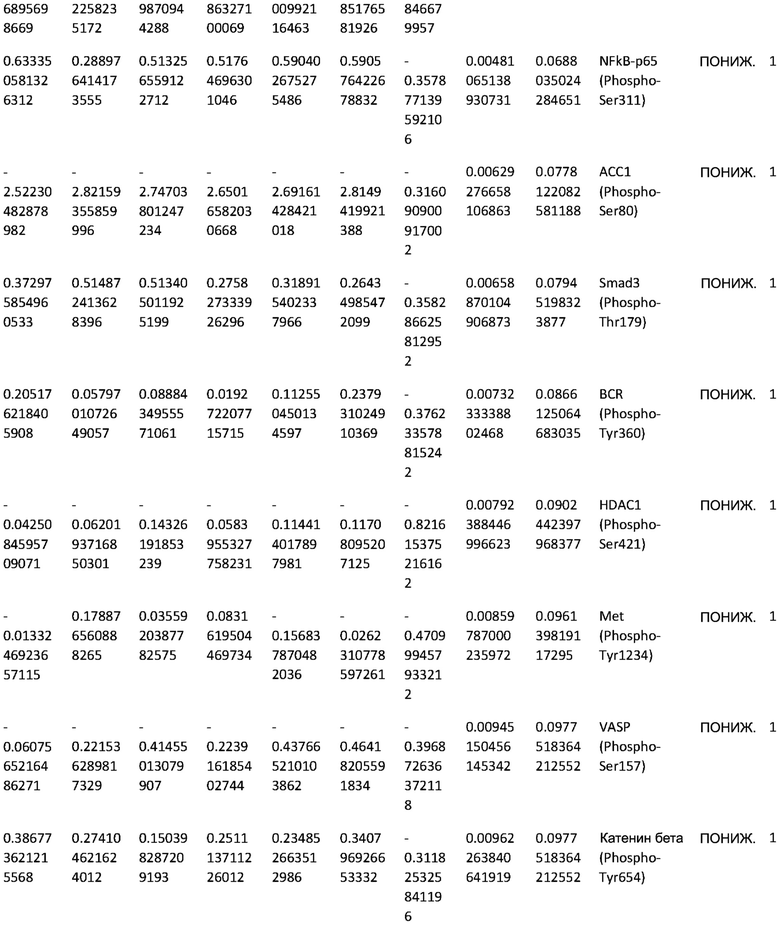
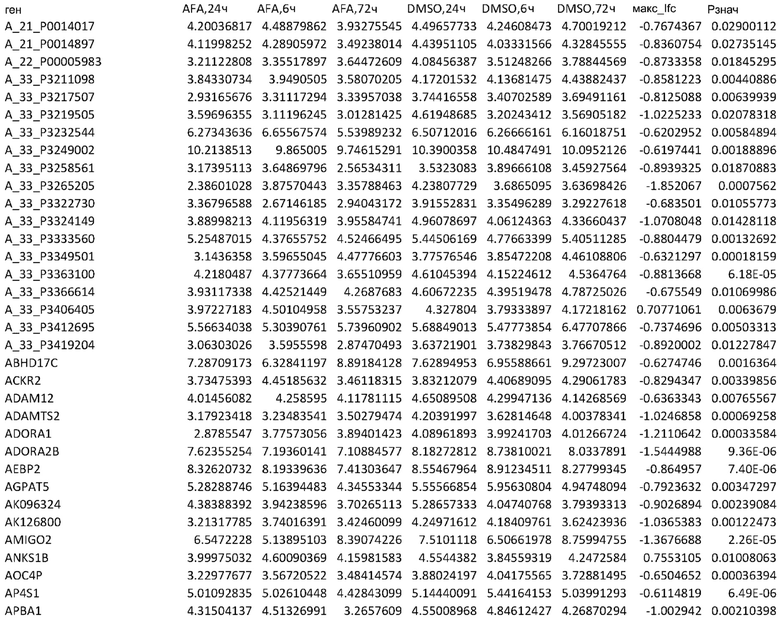
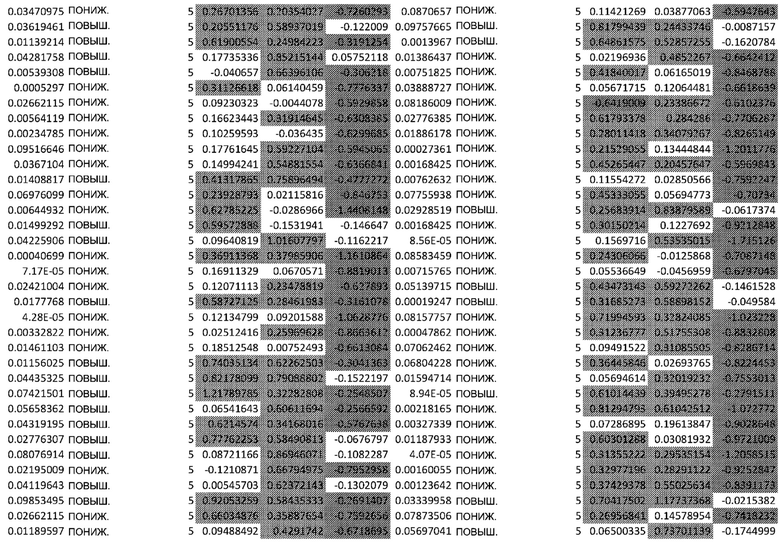
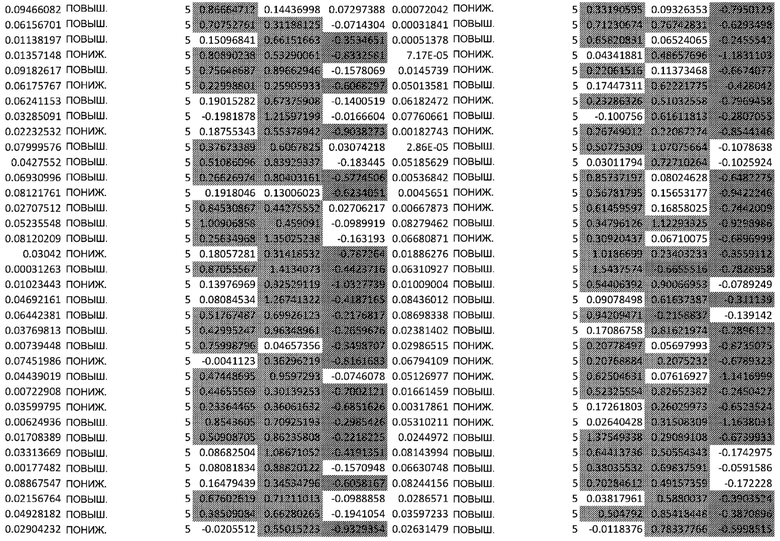
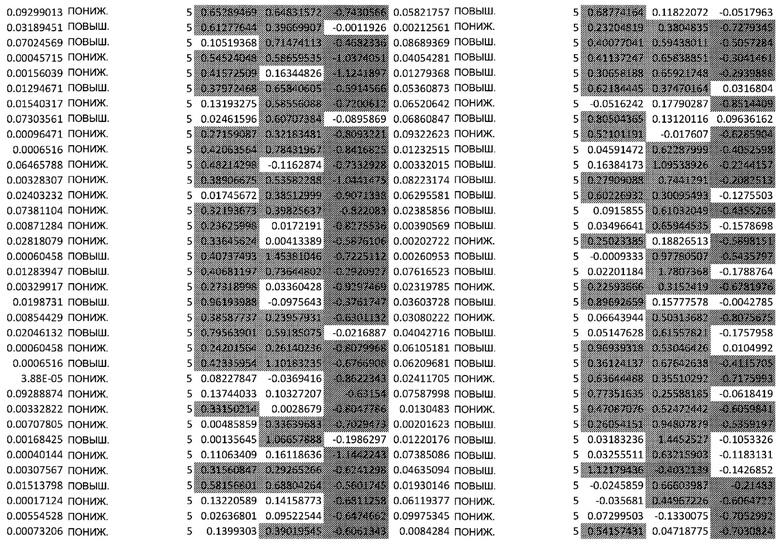
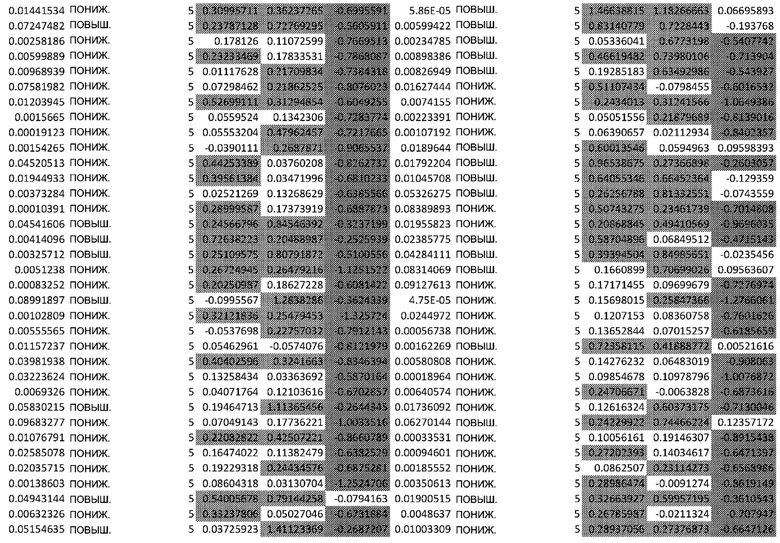
ТАБЛИЦА2 (Excel файл)







































































































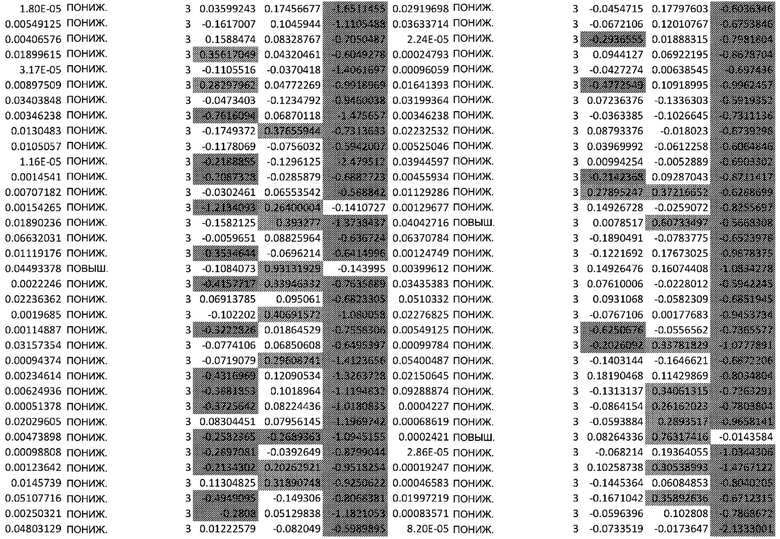
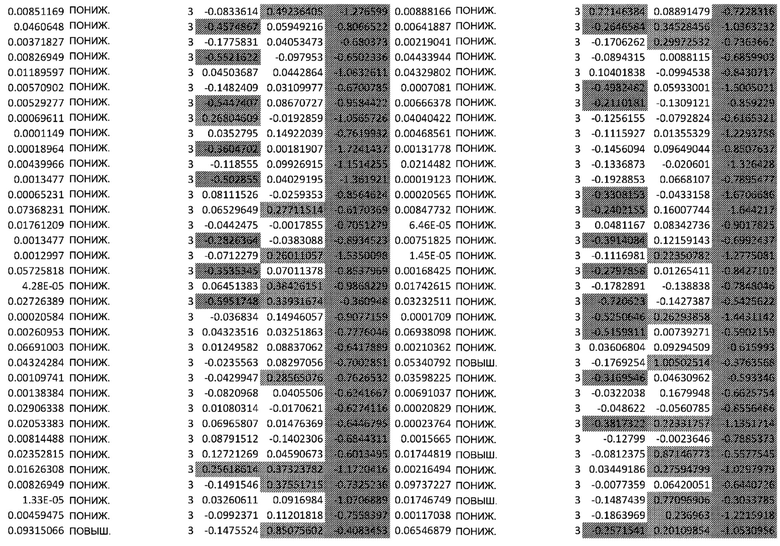
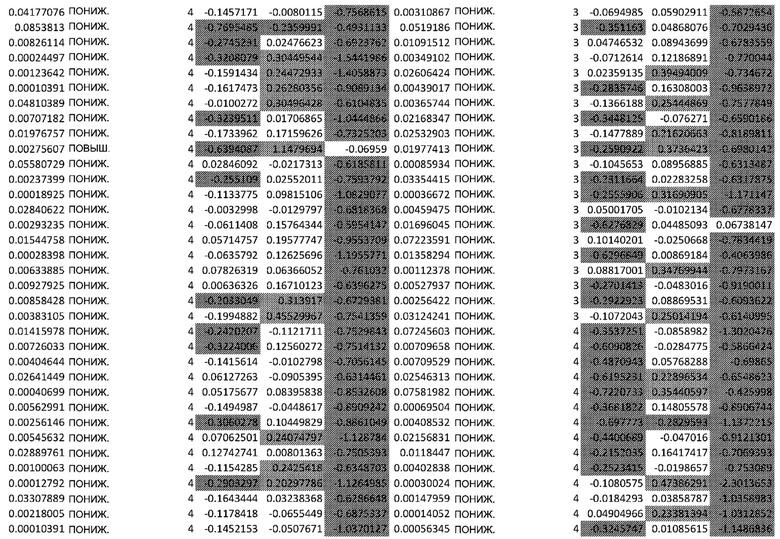
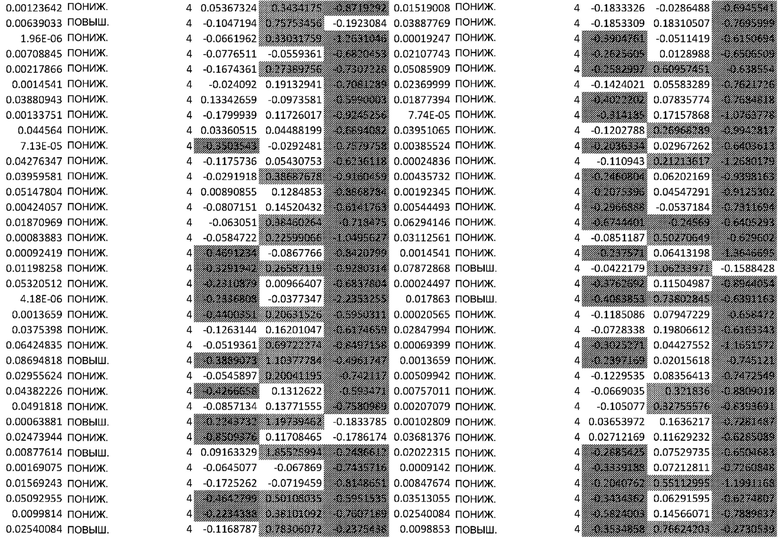
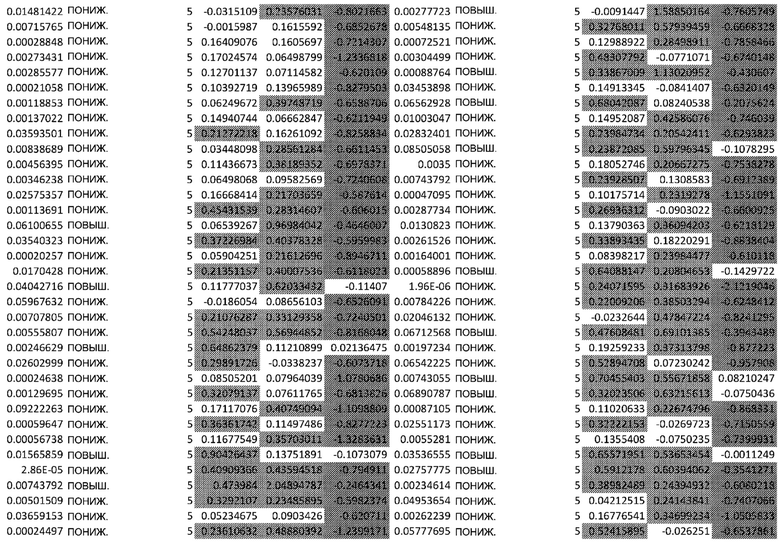
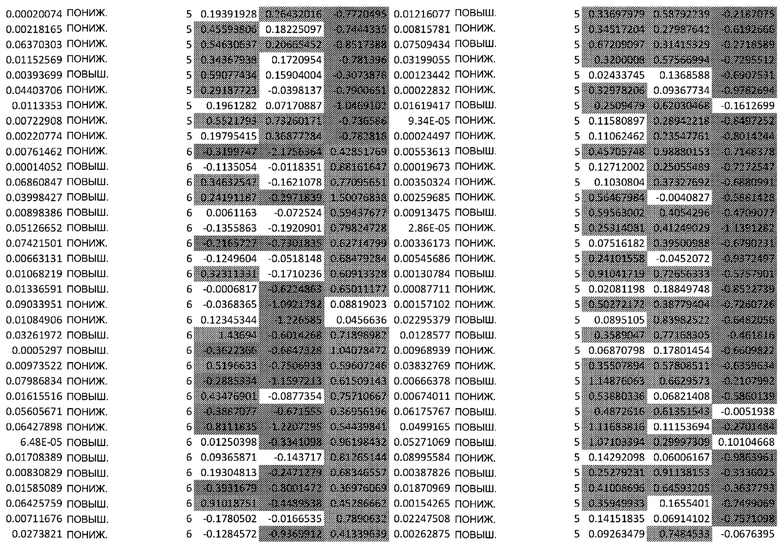
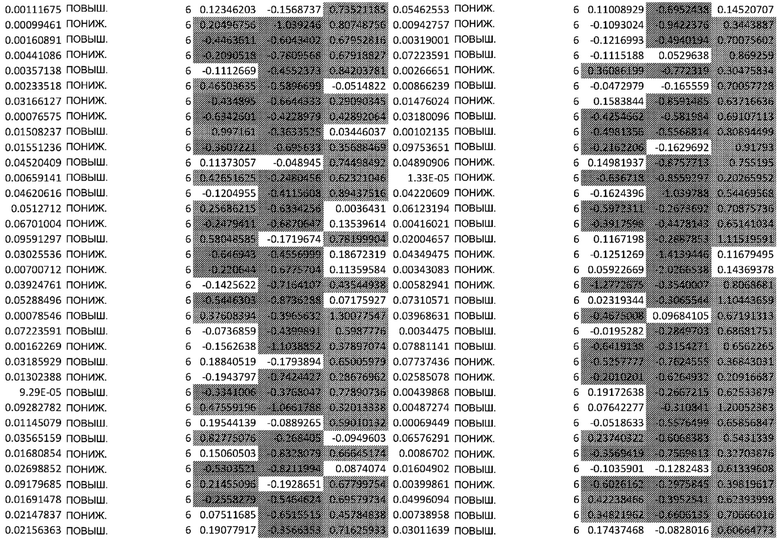
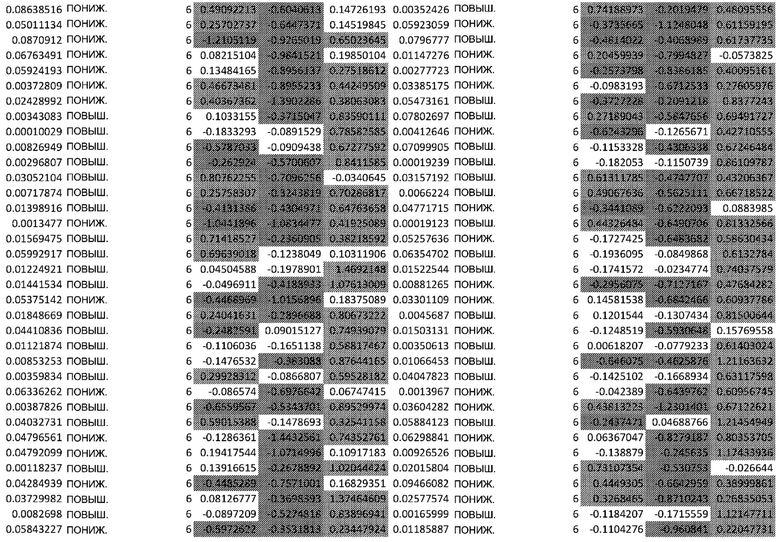







































































































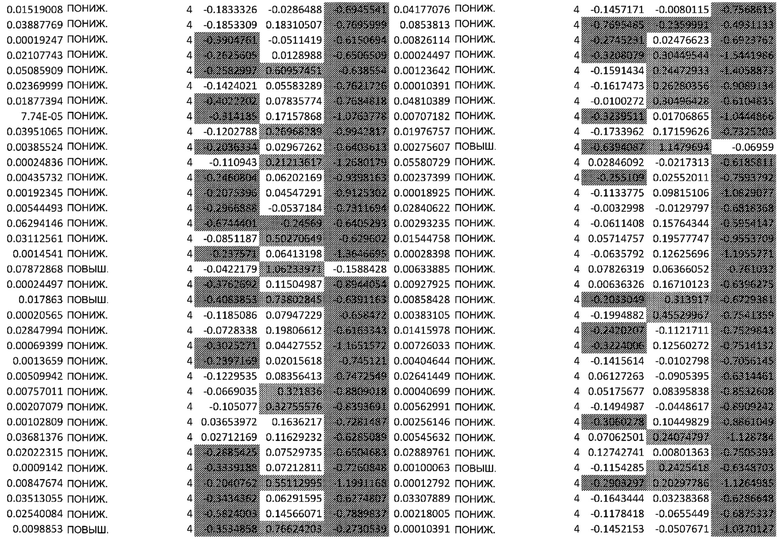
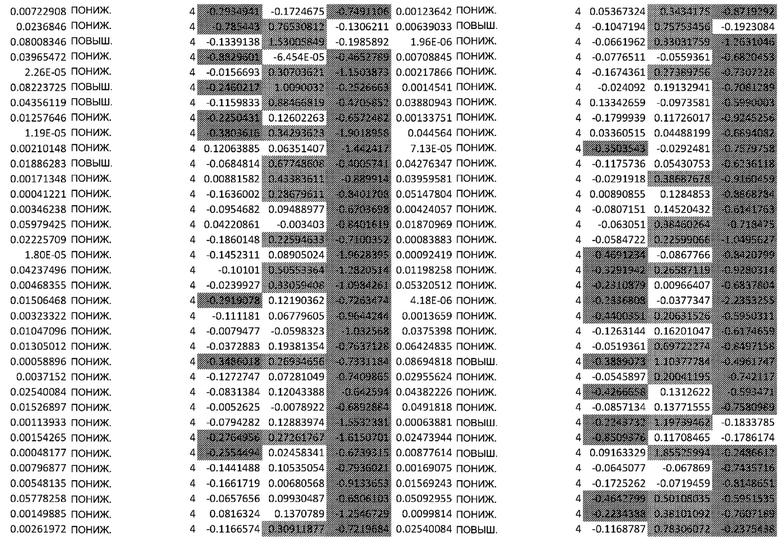
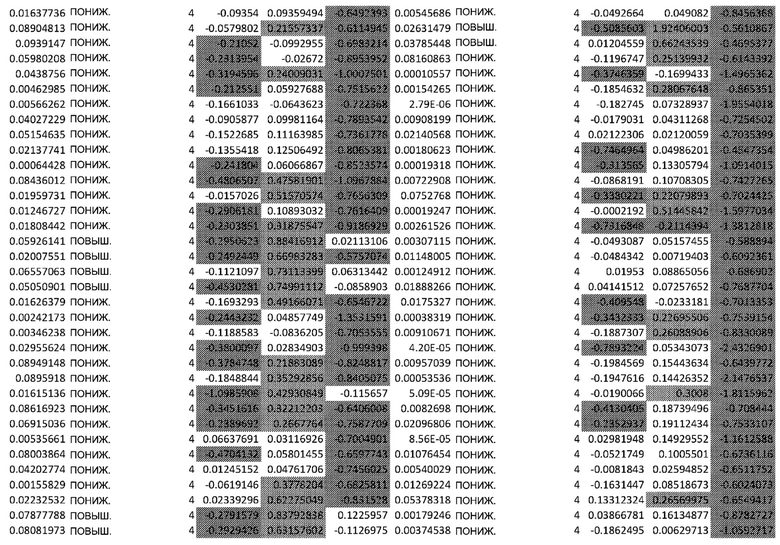
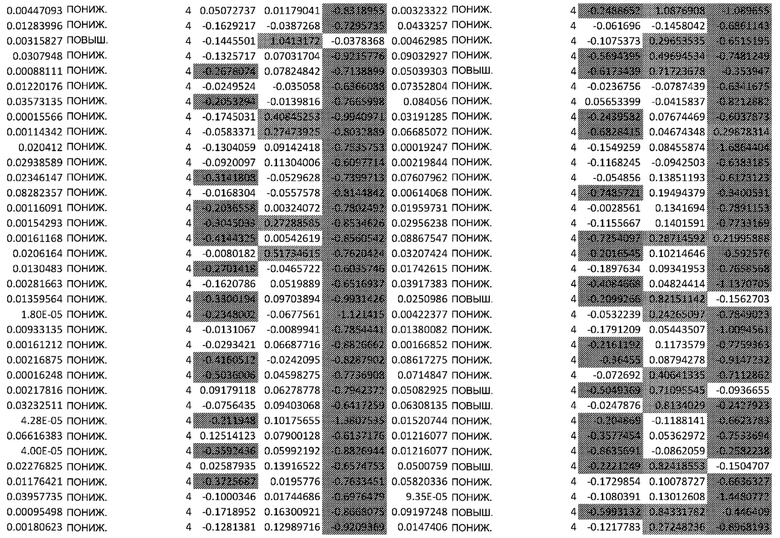

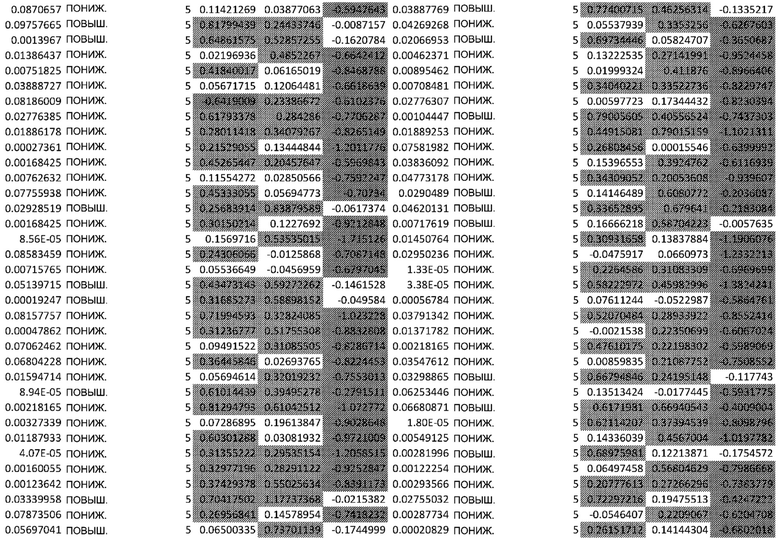
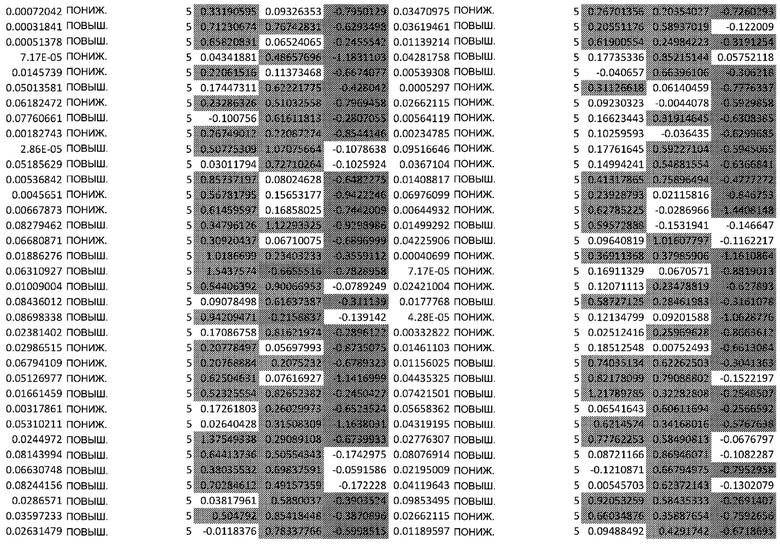
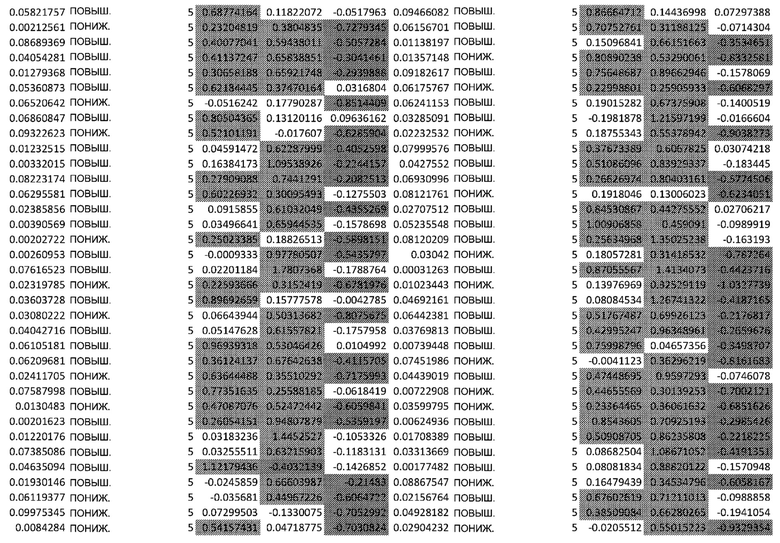
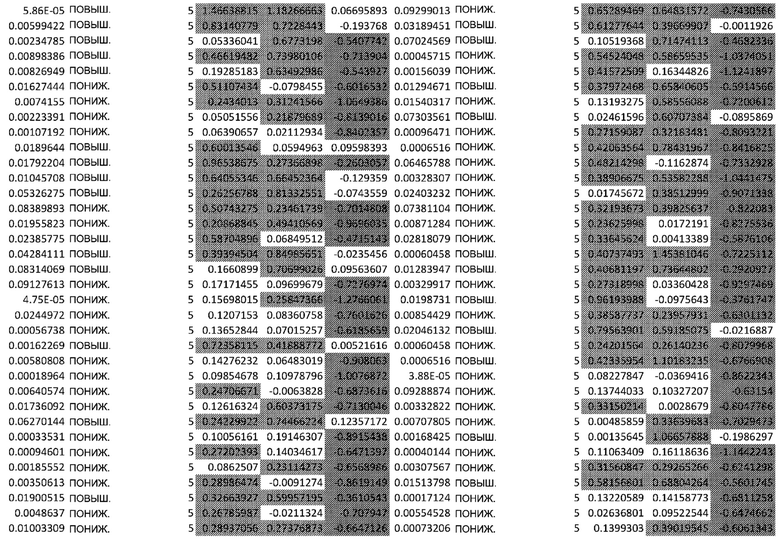







Изобретение относится к биотехнологии и медицине, в частности к способу оценки потенциала противораковых терапевтических средств вызывать побочные эффекты со стороны кожи. Способ включает использование трехмерной модели эпидермиса, демонстрирующей дифференцирующиеся кератиноциты в реконструированном роговом слое, при этом модель содержит противораковое терапевтическое средство в количестве, эффективном для имитации хронического воздействия лекарственного средства. Также предложено применение указанной модели для тестирования композиции для определения того, может ли композиция предотвращать эффект, который оказывает указанное противораковое терапевтическое средство на структуру и функцию кожного барьера. Способ позволяет оценивать способность противораковых терапевтических средств вызывать побочные эффекты со стороны кожи, а также эффективность композиций в снижении воздействия противораковых терапевтических средств на кожу. 2 н. и 4 з.п. ф-лы, 5 ил., 2 табл.
1. Способ in vitro оценки воздействия противоракового терапевтического средства на кожу, включающий:
использование трехмерной модели эпидермиса in vitro или ex vivo, демонстрирующей дифференцирующиеся кератиноциты в реконструированном роговом слое;
при этом указанная модель содержит противораковое терапевтическое средство в количестве, эффективном для имитации хронического воздействия лекарственного средства;
при этом указанная модель функционально воспроизводит нарушение барьера вследствие лечения указанным противораковым терапевтическим средством;
при этом указанное нарушение барьера определяется путем измерения полученного биологического процесса; и
при этом биологический процесс представляет собой статус фосфорилирования одного или большего количества белков или представляет собой экспрессию генов; и
оценку воздействия противоракового терапевтического средства на кожу.
2. Способ по п. 1, отличающийся тем, что указанное фосфорилирование белка регулируется с повышением.
3. Способ по п. 1, отличающийся тем, что указанное фосфорилирование белка регулируется с понижением.
4. Способ по п. 1, отличающийся тем, что указанная экспрессия гена регулируется с повышением.
5. Способ по п. 1, отличающийся тем, что указанная экспрессия гена регулируется с понижением.
6. Применение трехмерной модели эпидермиса in vitro или ex vivo, демонстрирующей дифференцирующиеся кератиноциты в реконструированном роговом слое;
при этом указанная модель содержит противораковое терапевтическое средство в количестве, эффективном для имитации хронического воздействия лекарственного средства;
при этом указанная модель функционально воспроизводит нарушение барьера вследствие лечения указанным противораковым терапевтическим средством;
при этом указанное нарушение барьера определяется путем измерения полученного биологического процесса; и
при этом биологический процесс представляет собой статус фосфорилирования одного или большего количества белков или представляет собой экспрессию генов;
для тестирования композиции для определения того, может ли указанная композиция предотвращать эффект, который оказывает указанное противораковое терапевтическое средство на структуру и функцию кожного барьера.
| JOLY-TONETTI et al., 202 Cutaneous adverse drug reactions of oncology treatments, Journal of Investigative Dermatology, 09.2019, vol | |||
| Способ подпочвенного орошения с применением труб | 1921 |
|
SU139A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Трансляция, предназначенная для телефонирования быстропеременными токами | 1921 |
|
SU249A1 |
| DESHAYES N | |||
| et al., 3D In vitro model of the re-epithelialization phase in the wound-healing process, Exp Dermatol, 2018, vol | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Способ получения сульфокислот из нефтяных дестиллатов, минеральных масел, парафина или церезина, обработанных серною кислотою | 1912 |
|
SU460A1 |
| ROBERT C | |||
| et al., RAF inhibition | |||

