Все документы, процитированные в настоящем описании, включены в него в полном объеме в качестве ссылок.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области сред и способов для культивирования клеток, в частности, сред для культивирования и способов дифференцировки клеток-предшественников, например, эпителиальных стволовых клеток человека.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Существует огромный интерес к средам для культивирования и способам дифференцировки клеток-предшественников. Клетки-предшественники и их дифференцированное потомство можно использовать в клеточных анализах, скрининге лекарственных средств и анализах токсичности. Клетки-предшественники и их дифференцированное потомство также являются перспективными для клеточной терапии, например, для регенеративной медицины для лечения поврежденной ткани. Кроме того, эффективные среды для культивирования клеток важны для получения и поддержания популяций клеток в исследовательских целях.
Аргентофильные клетки (EEC) являются редкими, гормон-секретирующими клетками, которые можно получать из Lgr5 стволовых клеток (Koo and Clevers (2014) Gastroenterology 147:289-302). Чаще всего подтипы различают с учетом секретируемых ими гормонов, и они включают соматостатин+ (Sst) D-клетки, желудочный ингибиторный белок+ (Gip) K-клетки, секретин+ (Sct) S-клетки, холецистокинин (Cck) I-клетки, глюкагон-подобный пептид 1+ (GLP-1) L-клетки, нейротензин+ (Nts) N-клетки и серотонин-продуцирующие энтерохромаффинные клетки (Gunawardene et al. (2011) International journal of experimental pathology 92:219-231). Однако одна EEC может экспрессировать множество гормонов в разной степени, что подчеркивает высокий уровень гетерогенности (Egerod et al. (2012) Nature cell biology 14:1099-1104). Хотя считают, что EEC играют ключевую роль в контроле различных аспектов функционирования кишечника и метаболизма организма, их небольшое количество представляет собой препятствие для их глубокого изучения и использования.
Описаны способы дифференцировки клеток-предшественников, полученных из некоторых тканей (например, поджелудочной железы, толстого кишечника, крипт кишечника и желудка) (см. WO 2010/090513, WO 2012/014076, WO 2012/168930 и WO 2015/173425). Известны способы индукции дифференцировки энтероцитов, бокаловидных клеток и клеток Панета. Существует потребность в улучшенных средах для культивирования и способах, приводящих к более высокой эффективности дифференцировки клеток-предшественников в направлении EEC.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к способу дифференцировки клеток-предшественников, включающему:
культивирование клеток в дифференцировочной среде, содержащей минимальную среду и дополнительно содержащей один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt.
Изобретение дополнительно относится к дифференцировочной среде, содержащей минимальную среду и дополнительно содержащей один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt.
Изобретение дополнительно относится к способу дифференцировки клеток-предшественников кишечника для получения популяции клеток кишечника, обогащенных аргентофильными клетками, включающему:
культивирование клеток-предшественников кишечника в дифференцировочной среде по изобретению.
Изобретение дополнительно относится к способу культивирования эпителиальных стволовых клеток, предпочтительно, для получения органоида, включающему:
культивирование одной или нескольких эпителиальных стволовых клеток, приведенных в контакт с внеклеточным матриксом, в присутствие среды для выращивания; и
культивирование одной или нескольких выращенных эпителиальных стволовых клеток в дифференцировочной среде по изобретению.
Изобретение дополнительно относится к способу культивирования эпителиальных стволовых клеток кишечника, предпочтительно, для получения дифференцированного органоида кишечника, включающему:
культивирование одной или нескольких эпителиальных стволовых клеток кишечника, приведенных в контакт с внеклеточным матриксом в присутствие среды для выращивания; предпочтительно, где среда для выращивания содержит минимальную среду и дополнительно содержит: лиганд рецепторной тирозинкиназы (например, EGF), ингибитор BMP (например, Noggin) и агонист Wnt (например, R-спондин) и, необязательно, вальпроевую кислоту и ингибитор GSK-3 (например, CHIR99021); а затем
культивирование одной или нескольких выращенных эпителиальных стволовых клеток кишечника, приведенных в контакт с внеклеточным матриксом, в присутствие дифференцировочной среды по изобретению.
Изобретение дополнительно относится к органоиду, получаемому или полученному способом по изобретению.
Изобретение дополнительно относится к органоиду, в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток.
Изобретение дополнительно относится к органоиду по изобретению или клетке, полученной из указанного органоида, для применения в медицине.
Изобретение дополнительно относится к применению органоида по изобретению или клетки, полученной из указанного органоида, в скрининге для обнаружения лекарственных средств; анализе токсичности; диагностике; исследовании эмбриологии тканей, линий клеток и путей дифференцировки; исследовании для идентификации химических и/или нейронных сигналов, приводящих к высвобождению соответствующих гормонов; исследованиях экспрессии генов, включая экспрессию рекомбинантных генов; исследовании механизмов, участвующих в повреждении и репарации ткани; исследовании воспалительных и инфекционных заболеваний; исследованиях патогенетических механизмов или исследованиях механизмов трансформации клеток и этиологии злокачественных новообразований.
Изобретение дополнительно относится к фармацевтическому составу, содержащему один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt.
Способ скрининга терапевтического или профилактического фармацевтического лекарственного средства или косметического средства, включающий:
приведение дифференцированного органоида по изобретению в контакт с молекулой-кандидатом (или библиотекой молекул-кандидатов),
оценку указанного органоида на какие-либо эффекты (например, любое изменение клетки, такое как снижение или утрата пролиферации, морфологическое изменение и/или гибель клеток) или изменение органоида (например, размера или подвижности органоида);
идентификацию молекулы-кандидата, вызывающей указанные эффекты, в качестве потенциального лекарственного средства или косметического средства; и необязательно
получение указанной молекулы-кандидата в качестве фармацевтического или косметического средства.
Изобретение дополнительно относится к способу индукции состояния покоя у Lgr5+ стволовой клетки, где указанный способ включает:
обработку клетки одним или несколькими ингибиторами пути EGFR.
Изобретение дополнительно относится к популяции покоящихся стволовых клеток, полученной способом индукции состояния покоя Lgr5+ стволовых клеток-предшественников по изобретению, где клетки экспрессируют Lgr5 и Lef1 и не экспрессируют KI67 и маркер M-фазы фосфогистон H4.
Изобретение дополнительно относится к способу получения популяции клеток, обогащенной EEC, включающему культивирование популяции клеток в дифференцировочной среде по изобретению.
Изобретение дополнительно относится к способу получения популяции клеток, обогащенных GLP1-секретирующими EEC, включающему культивирование популяции клеток в дифференцировочной среде по изобретению, где дифференцировочная среда содержит ингибитор BMP.
Изобретение дополнительно относится к способу получения популяции клеток, обогащенных секретин-секретирующими EEC, включающему культивирование популяции клеток в дифференцировочной среде по изобретению, где дифференцировочная среда содержит активатор пути BMP.
Изобретение дополнительно относится к ингибитору BMP для применения в способе лечения или профилактики сахарного диабета или ассоциированного заболевания или нарушения, где способ включает введение терапевтически эффективного количества ингибитора BMP нуждающемуся в этом индивидууму.
Изобретение дополнительно относится к активатору BMP для применения в способе лечения гиперхлоргидрии или ожирения, где способ включает введение терапевтически эффективного количества активатора BMP нуждающемуся в этом индивидууму.
ПОДРОБНОЕ ОПИСАНИЕ
Способы дифференцировки клеток-предшественников из различных тканей ранее описаны в WO 2010/090513, WO 2012/014076, WO 2012/168930 и WO 2015/173425. Описаны способы стимуляции дифференцировки клетки-предшественника кишечника в направлении энтероцита, бокаловидной клетки или клетки Панета. Они представлены на фигуре 11. Однако ранее не описаны способы и среды для усиления дифференцировки клетки-предшественника кишечника в направлении EEC. Авторы настоящего изобретения неожиданно обнаруживали, что комбинация ингибитора Wnt, ингибитора пути EGFR и ингибитора Notch в дифференцировочной среде усиливает дифференцировку клеток-предшественников в направлении EEC (см. пример 2). EEC составляют менее 1% клеток эпителия кишечника in vivo. Однако с помощью способов и сред для дифференцировки по изобретению можно получать органоиды, в которых ~50% клеток являются EEC.
Авторы настоящего изобретения также неожиданно обнаруживали, что посредством модуляции пути передачи сигнала BMP можно модулировать фенотип EEC, в частности, в том, что касается уровней экспрессии и секреции гормонов (см. пример 5). Авторы настоящего изобретения показали, что конкретные фенотипы EEC можно получать посредством активации или ингибирования передачи сигнала BMP. Авторы настоящего изобретения также показали, что эти способы могут быть эффективными in vivo, таким образом, приводя к многообещающему новому применению активаторов и ингибиторов BMP в терапии.
Авторы настоящего изобретения также неожиданно обнаруживали, что наличие ингибитора пути EGFR в среде для культивирования стимулирует состояние покоя Lgr5+ стволовых клеток (см. пример 1). В частности, авторы настоящего изобретения обнаружили новое состояние покоя, при котором сохраняется потенциал стволовых клеток, но приостанавливается пролиферация. Ранее не было известно, что потенциал стволовых клеток и пролиферацию можно разделять.
Ингибиторы Wnt
Дифференцировочная среда по изобретению содержит ингибитор Wnt. Можно использовать любой подходящий ингибитор Wnt.
Путь передачи сигнала Wnt при активации, как правило, предотвращает деградацию β-катенина и повышает опосредованную β-катенином передачу сигнала. Этот путь определяется серией событий, происходящих при активации комплекса рецептора Wnt на поверхности клеток, содержащего рецептор Frizzled и LRP5/6, как правило, под действием внеклеточной сигнальной молекулы, такой как член семейства Wnt. Это приводит к активации белков семейства Dishevelled, ингибирующих деструктивный комплекс белков, осуществляющих деградацию внутриклеточного β-катенина. Деструктивный комплекс образован структурными компонентами, включающими APC и аксин, к которым рекрутируются казеинкиназы CK1α, δ и ε и GSK-3. Считают, что деструктивный комплекс фосфорилирует β-катенин и экспонирует его для убиквитинлигазы β-TrCP. Затем убиквитинилирование β-катенина приводит к его деградации в протеасоме.
Основная эффекторная функция β-катенина осуществляется в ядре, где он регулирует транскрипцию посредством взаимодействия с различными факторами транскрипции, включая факторы транскрипции семейства TCF/LEF (например Tcf-1, Tcf-3, Tcf-4 и Lef1).
Путь Wnt подвергается значительной регуляции. Например, передача сигнала Wnt повышается, когда R-спондин связывается со своими рецепторами (Lgr4, Lgr5 и/или Lgr6). Однако, показано, что две трансмембранные убиквитинлигазы E3, Rnf43 и Znrf3, удаляют рецепторы R-спондина (например, Lgr4, Lgr5 и/или Lgr6) с поверхности клеток (см., например, de Lau et al. 2016). R-спондины являются Wnt-усиливающими молекулами позвоночных. Кроме того, связывание белков семейства Dishevelled с рецептором Frizzled может ингибироваться белками семейства Dapper (например, Dapper1 и Dapper3). Кроме того, считают, что активность деструктивного комплекса частично регулируется статусом фосфорилирования APC, аксина и GSK-3. Например, дефосфорилирование APC или аксина фосфатазами (например, серин/треониновыми фосфатазами, такими как PP1, PP2C или PP2A) может ингибировать деградацию β-катенина. Кроме того, фосфорилирование GSK-3 киназами (например, p38 MAPK, PKA, PKB, PKC, p90RSK или p70S6K) может ингибировать активность GSK-3 и, таким образом, ингибировать деградацию β-катенина.
Считают, что стабильность деструктивного комплекса частично регулируется двумя PARP, танкиразами 1 и 2. Поли-АДФ-рибозилирование аксина и ауто-поли-АДФ-рибозилирование этими танкиразами может способствовать деолигомеризации деструктивного комплекса.
В ядре белки семейства Dishevelled могут образовывать комплекс с гистондеацетилазой SIRT1, поддерживающей транскрипцию генов-мишеней Wnt.
Белком, который, как считают, является ключевым в секреции Wnt, является многопроходный мембранный белок поркупин (Porc), утрата которого приводит к накоплению Wnt в эндоплазматическом ретикулуме.
Путь передачи сигнала Wnt можно ингибировать на многих уровнях, подробный обзор ингибиторов Wnt представлен в Voronkov and Krauss, (2013), Current Pharmaceutical Design 19:634-664.
Ингибитор Wnt определяют как средство, ингибирующее TCF/LEF-опосредованную транскрипцию в клетке или популяции клеток. Таким образом, ингибиторы Wnt, пригодные для использования в изобретении, включают:
(1) ингибиторы секреции Wnt (например, ингибиторы Porc, такие как LGK974, IWP-1 или IWP-2),
(2) конкурентные и неконкурентные ингибиторы взаимодействия между Wnt или R-спондином и их соответствующими рецепторами (например, OMP-18R5, OMP54F28),
(3) факторы, способствующие деградации компонентов комплекса рецептора Wnt, такие как LRP (например, никлозамид), и факторы, способствующие деградации рецепторов R-спондина, такие как Znrf3 и/или Rnf43, или факторы, активирующие Znrf3 и/или Rnf43,
(4) ингибиторы белков семейства Dishevelled, такие как ингибиторы, снижающие связывание белков семейства Dishevelled с рецепторами Frizzled и/или компонентами деструктивного комплекса (например, белки семейства Dapper, FJ9, сулиндак, 3289-8625, J01-017a, NSC668036), или ингибиторы, отрицательно регулирующие экспрессию белков семейства Dishevelled (например, никлозамид),
(5) факторы, способствующие активности деструктивного комплекса, включая (a) ингибиторы фосфатаз (например, PP1, PP2A и/или PP2C), дефосфорилирующих компоненты деструктивного комплекса, такие как аксин и/или APC (например, окадаиковая кислота или таутомицин), и (b) ингибиторы киназ (например, p38 MAPK, PKA, PKB, PKC, p90RSK или p70S6K), фосфорилирующих GSK-3 (например, SB239063, SB203580 или Rp-8-Br-cAMP),
(6) ингибиторы деолигомеризации деструктивного комплекса, такие как ингибиторы танкираз 1 и/или 2 (например, XAV939, IWR1, JW74, JW55, 2-[4-(4-фторфенил)пиперазин-1-ил]-6-метилпиримидин-4(3H)-он или PJ34), и
(7) ингибиторы экспрессии гена-мишени β-катенина, включая ингибиторы транскрипционного комплекса β-катенин : TCF/Lef, такие как ингибиторы, разрушающие комплекс β-катенин : TCF 4 (например, iCRT3, CGP049090, PKF118310, PKF115-584, ZTM000990, PNU-74654, BC21, iCRT5, iCRT14 или FH535), и ингибиторы гистондеацетилазы SIRT1 (например, камбинол).
Дифференцировочная среда по изобретению содержит ингибитор Wnt. Можно использовать любой подходящий ингибитор Wnt, как описано в (1)-(7) выше. Например, в одном из предпочтительных вариантов осуществления ингибитор Wnt является ингибитором секреции Wnt, таким как ингибитор Porc, например, выбранный из IWP-2, IWP-1 и LGK974. В другом предпочтительном варианте осуществления ингибитор Wnt является ингибитором экспрессии гена-мишени β-катенина, например, ингибитором транскрипционного комплекса β-катенин : TCF/Lef или ингибитором гистондеацетилазы SIRT1 (например, камбинолом). В некоторых вариантах осуществления ингибитором транскрипционного комплекса β-катенина : TCF/Lef является ингибитор, разрушающий комплекс β-катенин : TCF-4, например, ингибитор, выбранный из iCRT3, CGP049090, PKF118310, PKF115-584, ZTM000990, PNU-74654, BC21, iCRT5, iCRT14 и FH535.
В некоторых вариантах осуществления ингибитор Wnt выбран из IWP-2, OMP-18R5, OMP54F28, LGK974, 3289-8625, FJ9, NSC 668036, IWR1 и XAV939.
В некоторых вариантах осуществления ингибитор Wnt выбран из iCRT3, PFK115-584, CGP049090, iCRT5, iCRT14 и FH535.
В некоторых вариантах осуществления ингибитор Wnt является одним из соединений, приведенных в таблице 1 ниже.
Таблица 1 - Ингибиторы Wnt
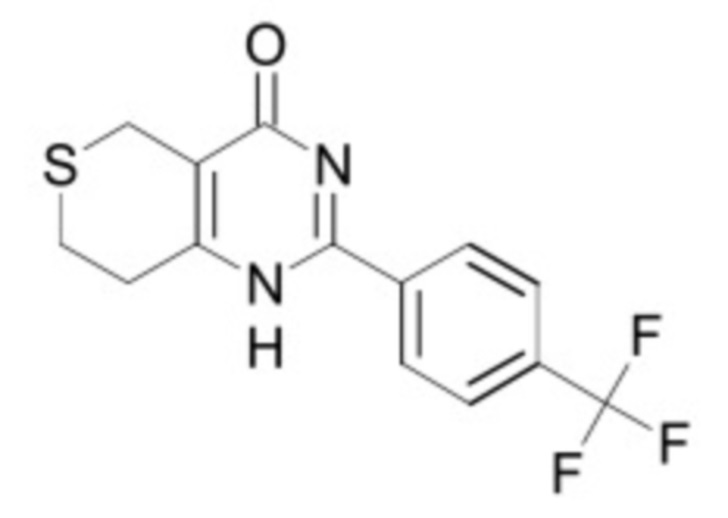
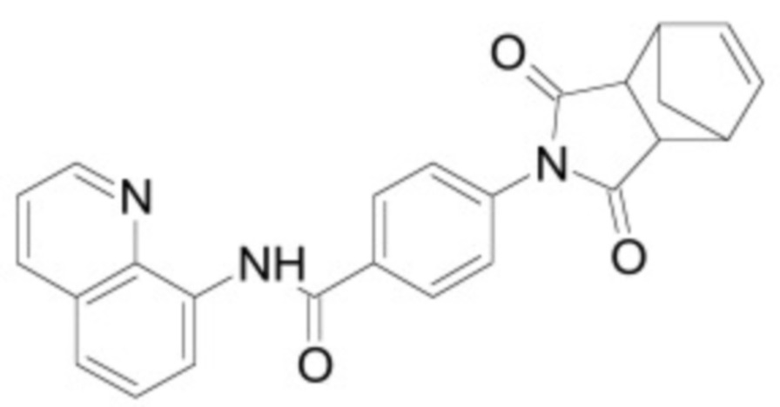
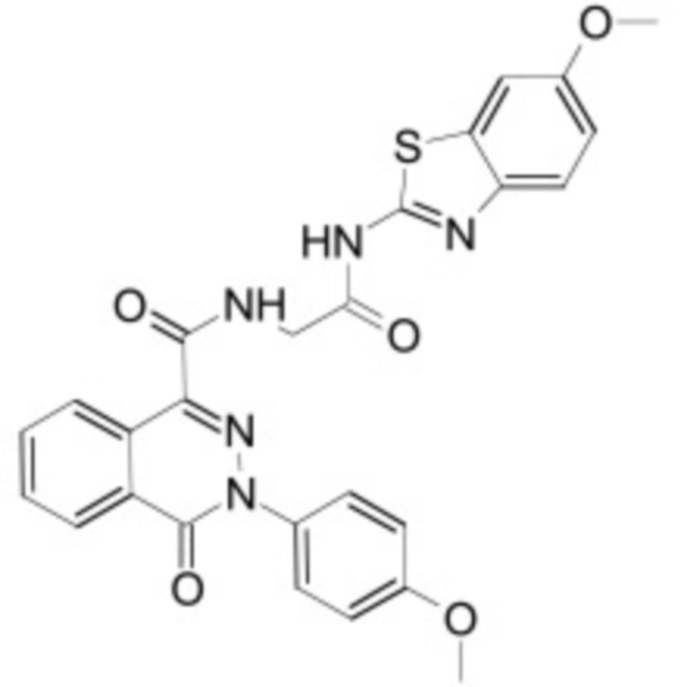
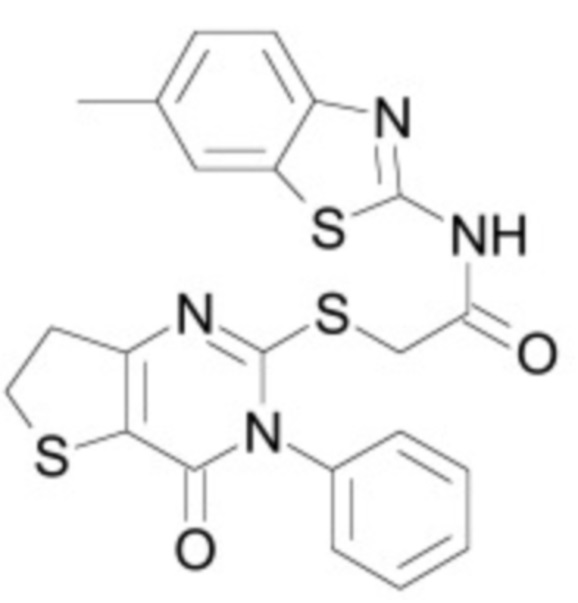
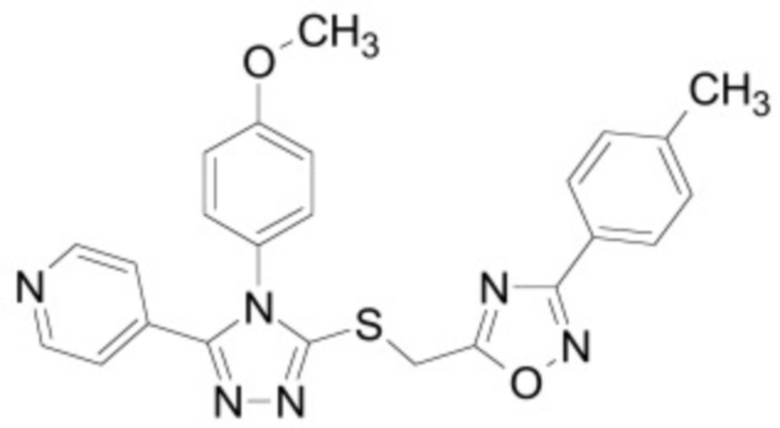
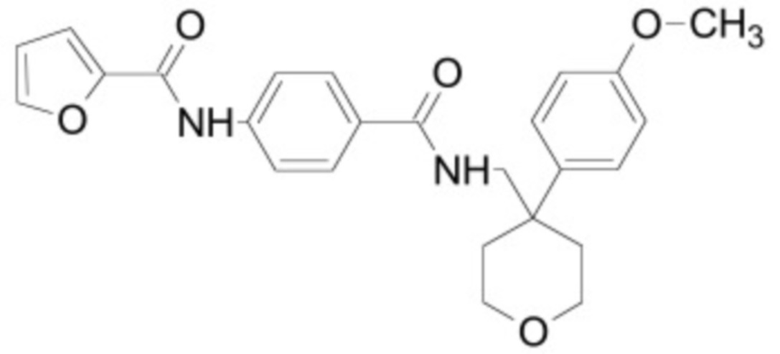
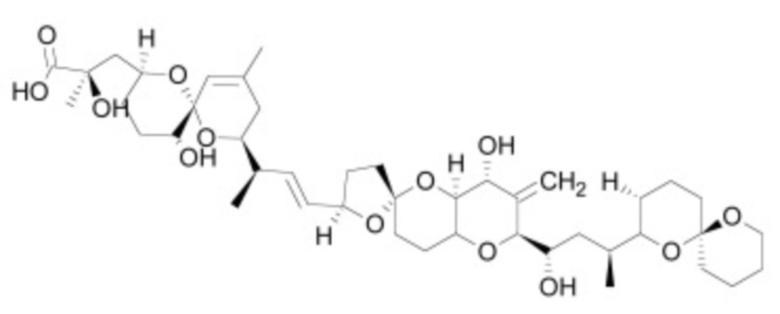
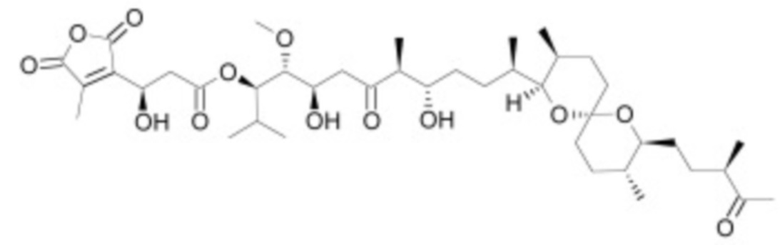
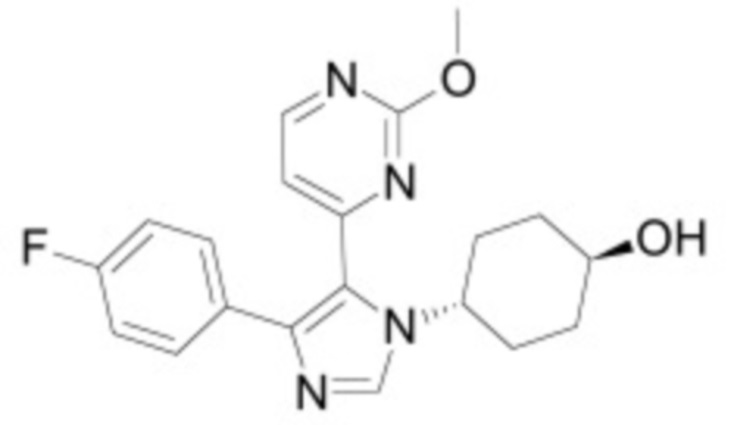
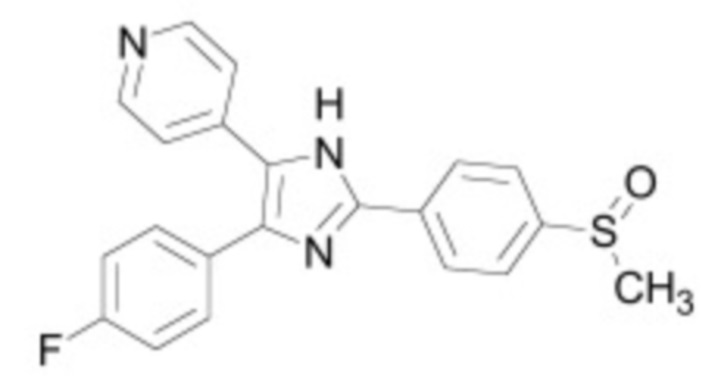
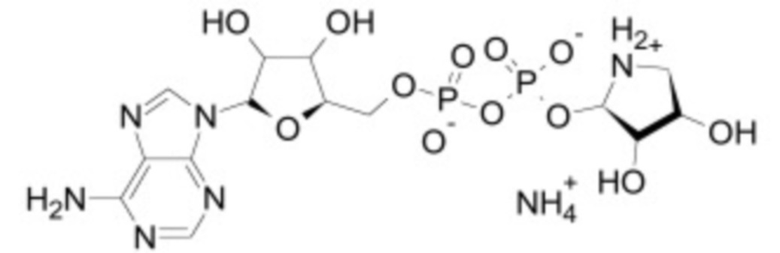
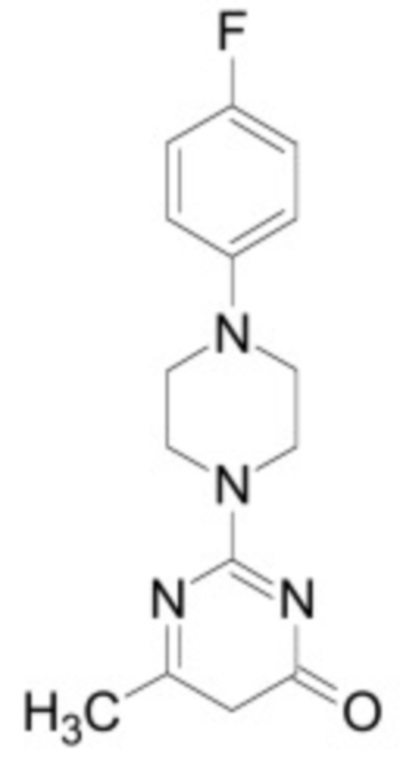
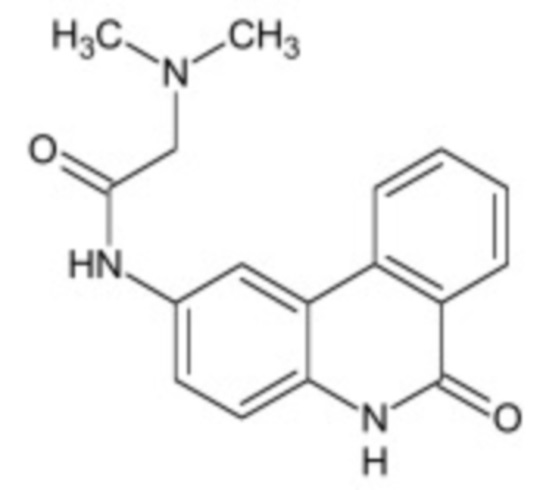
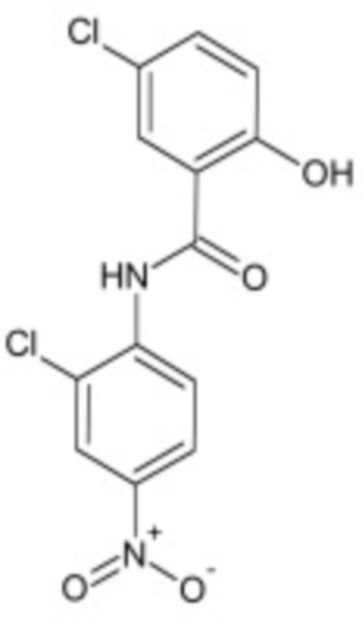
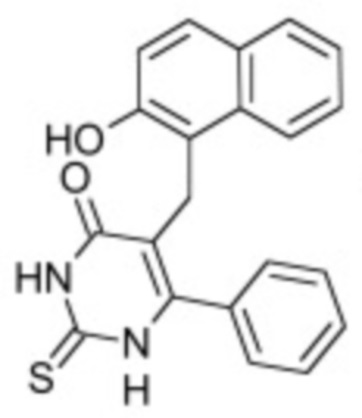
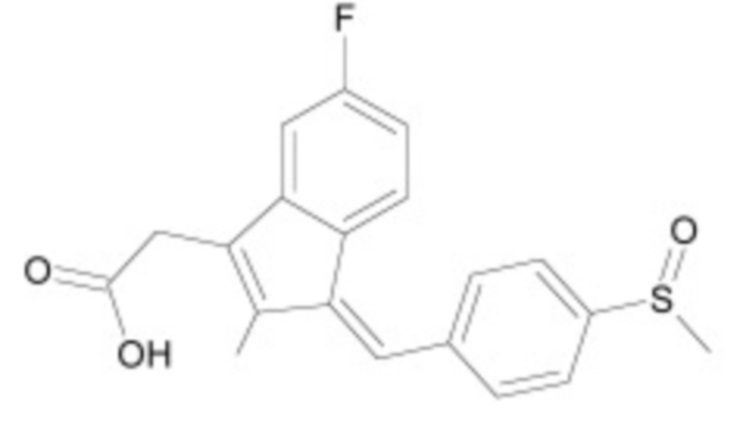
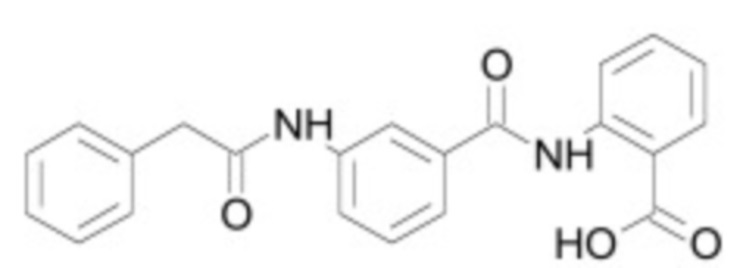
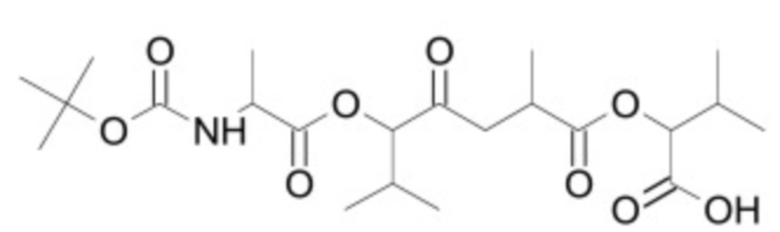
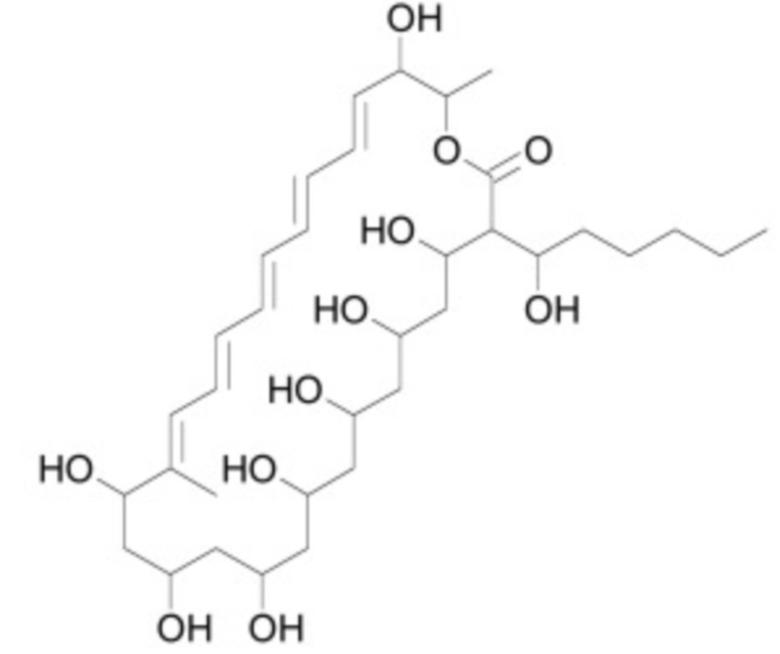
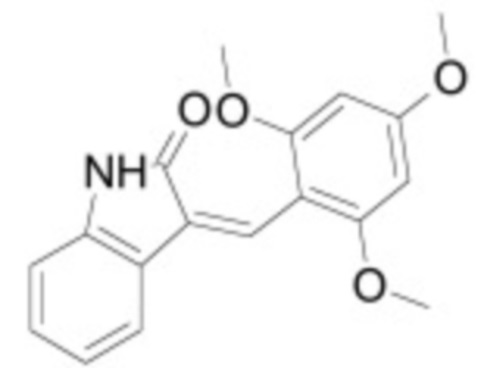
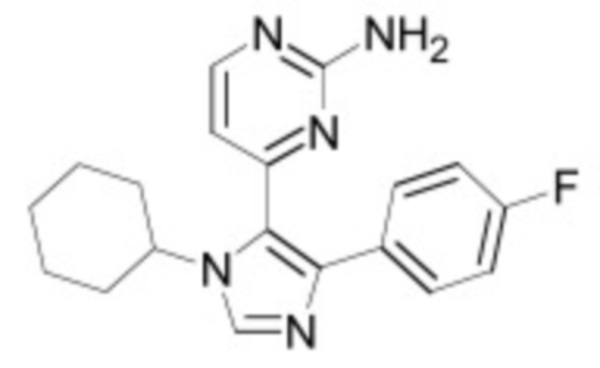
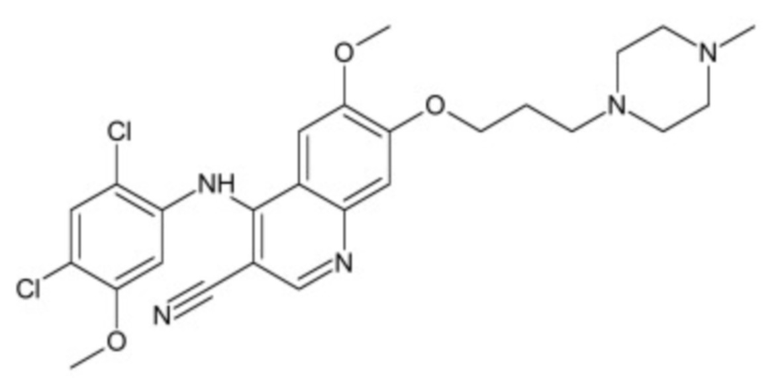
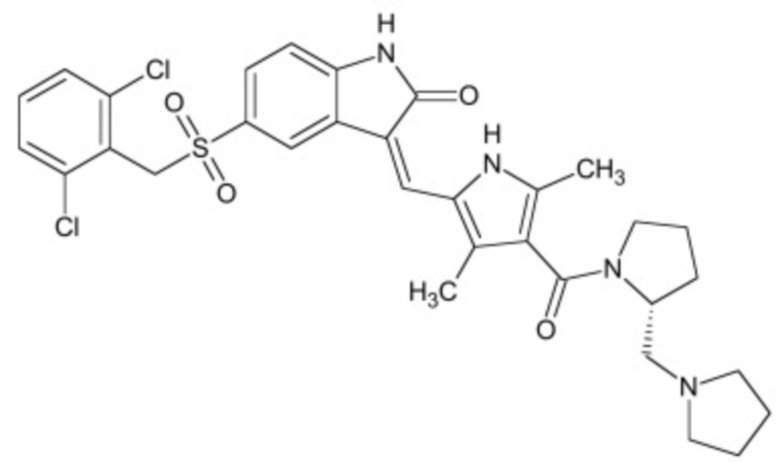
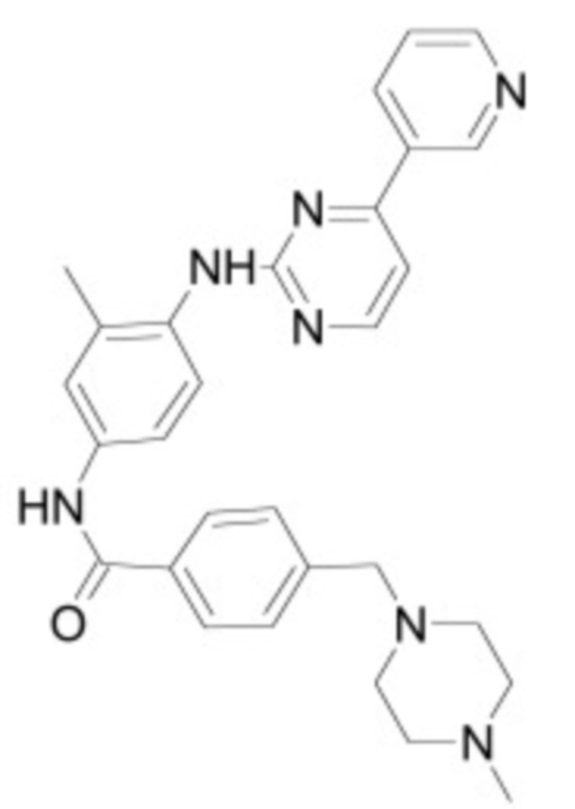
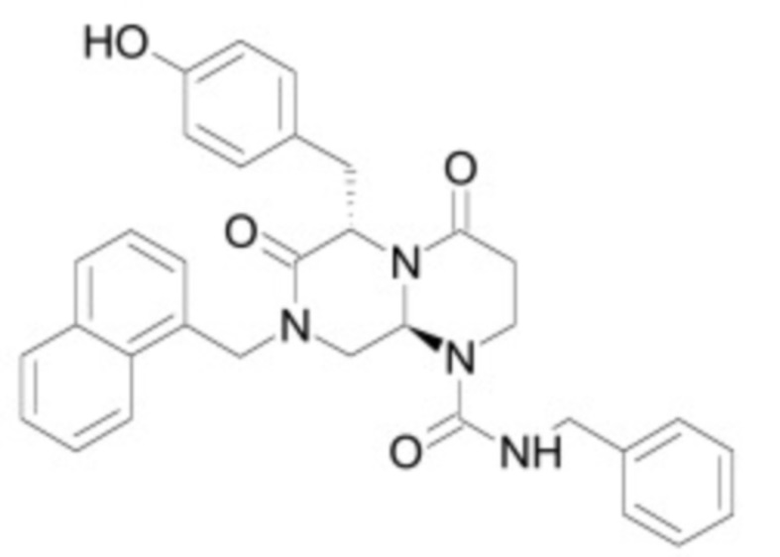
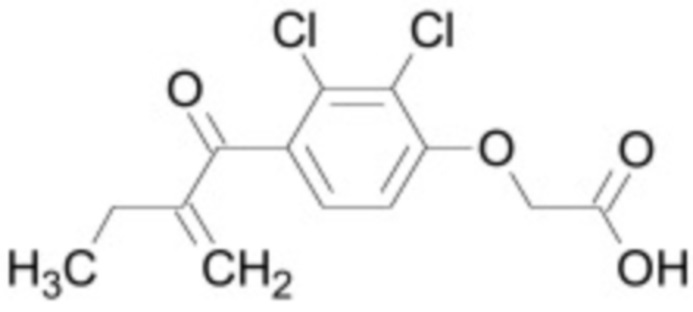
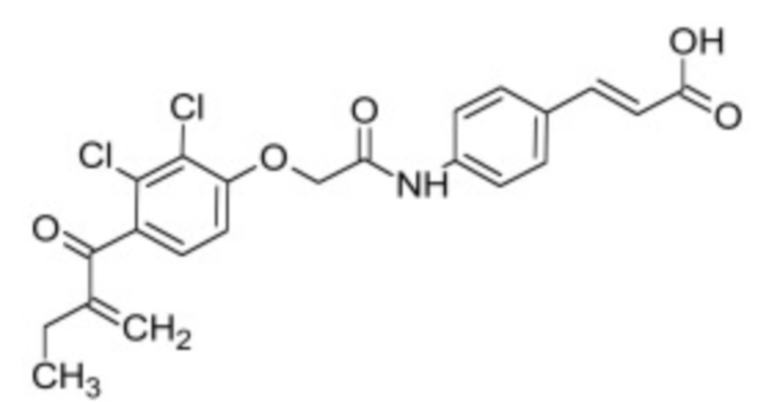
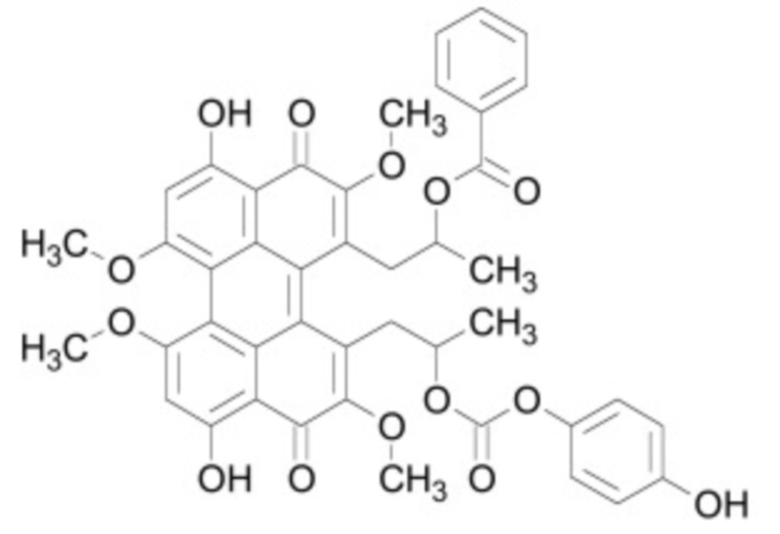
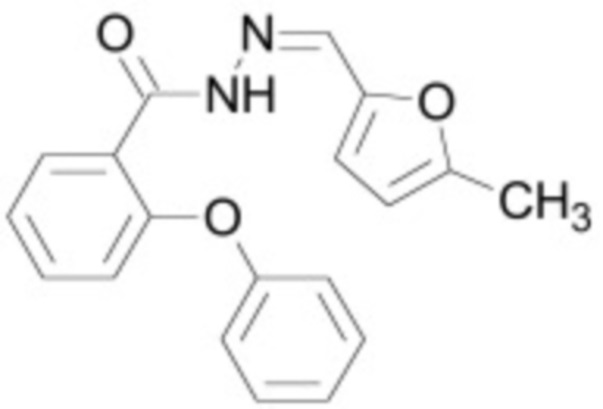
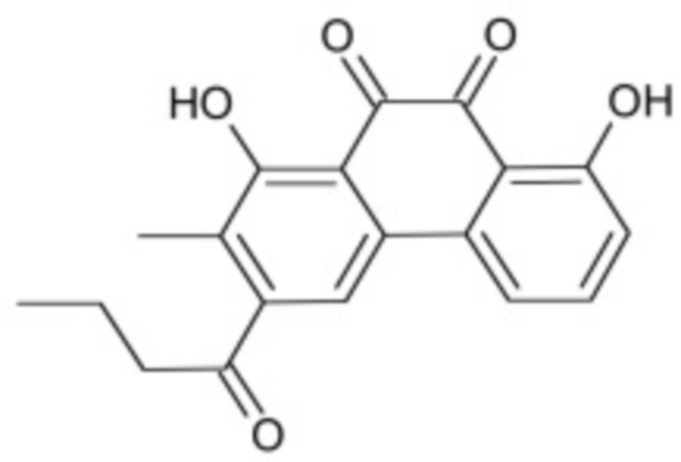
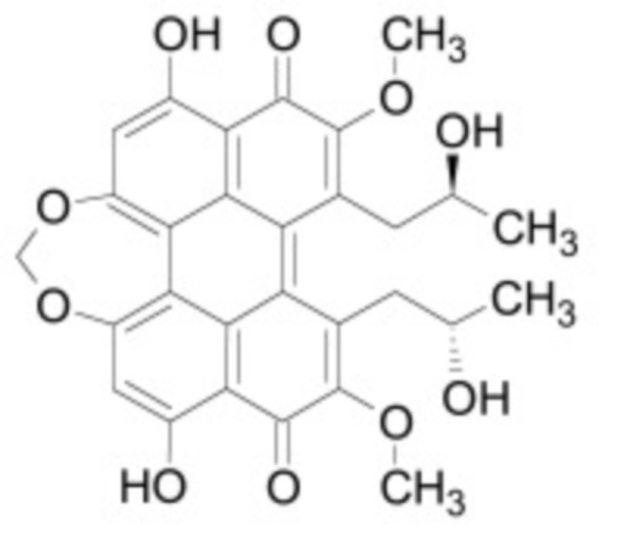
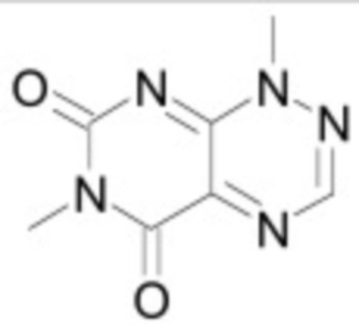
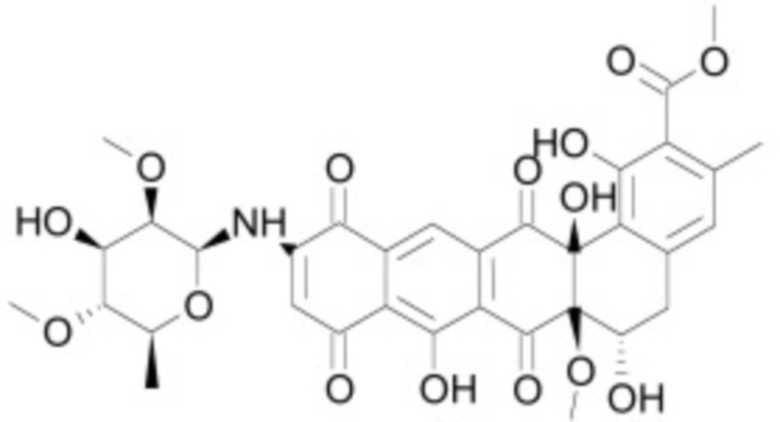
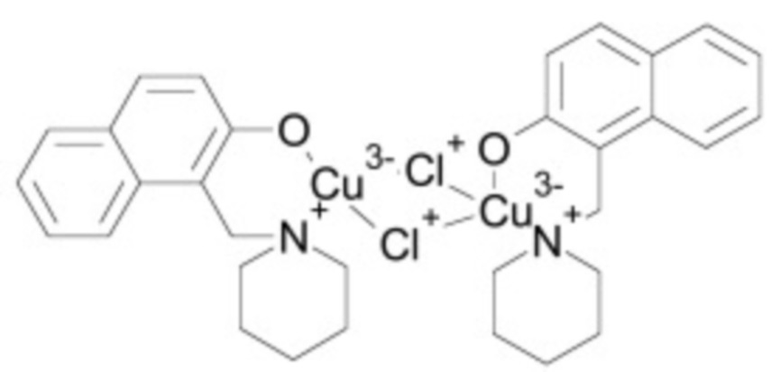
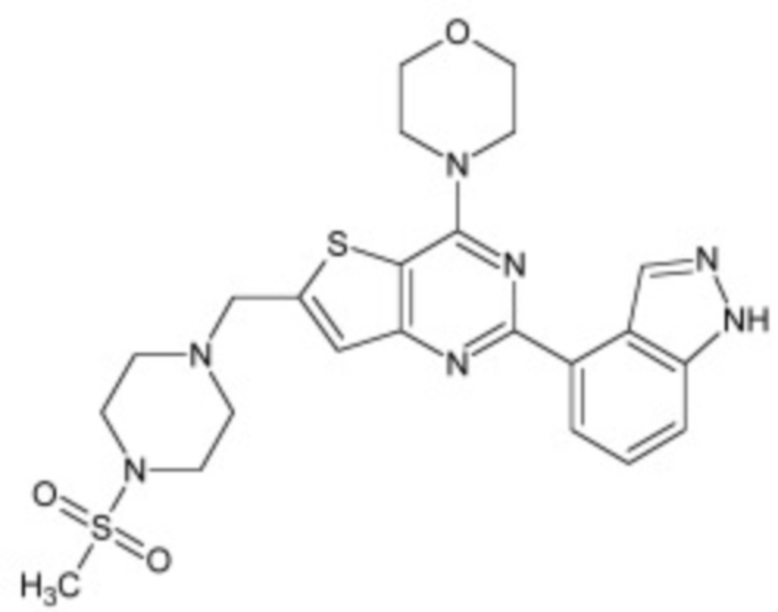
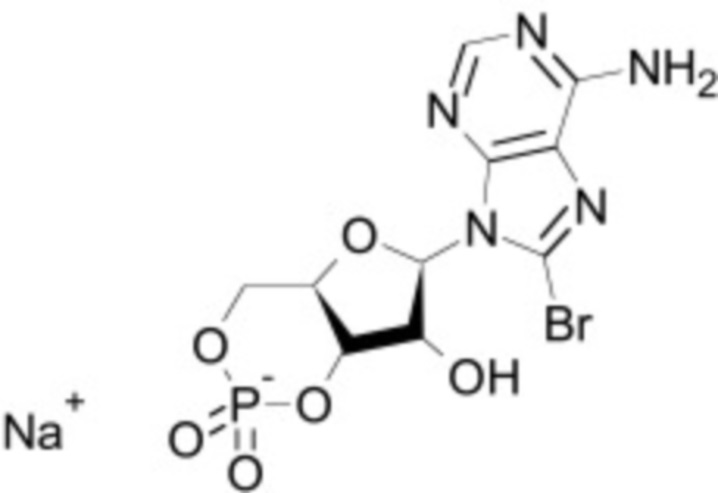
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит один или несколько любых ингибиторов Wnt, приведенных в таблице 1.
Ингибитор Wnt, предпочтительно, добавляют в среды в количестве, эффективном для ингибирования активности Wnt в клетке по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - 100% относительно уровня указанной активности Wnt в отсутствие указанной молекулы, как оценивают с использованием того же типа клеток. Как известно специалисту в этой области, активность Wnt можно определять посредством измерения транскрипционной активности Wnt, например, с помощью люцеферазных репортерных конструкций pTOPFLASH и pFOPFLASH Tcf (Korinek et al., (1997), Science 275:1784-1787). Таким образом, специалист в этой области легко может идентифицировать ингибиторы Wnt с использованием анализа, известного в этой области.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит ингибитор Wnt в концентрации 0,01-150 мкМ, 0,1-150 мкМ, 0,5-100 мкМ, 0,1-100 мкМ, 0,5-50 мкМ, 1-100 мкМ или 10-80 мкМ, 1-20 мкМ или 1-5 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 в концентрации 0,01-150 мкМ, 0,1-100 мкМ, 0,5-50 мкМ, 1-20 мкМ или 1-5 мкМ. Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 в концентрации приблизительно 1,5 мкМ.
В некоторых вариантах осуществления дифференцировочная среда не содержит агонист Wnt, связывающий и активирующий комплекс рецептора Wnt, включающий любой и все белки семейства Wnt и R-спондин.
В других вариантах осуществления дифференцировочная среда дополнительно содержит агонист Wnt, такой как R-спондин 1-4 или его биологически активный фрагмент или вариант. Как описано выше, R-спондины усиливают передачу сигнала Wnt через рецепторы на поверхности клеток. Авторы настоящего изобретения показали, что удаление R-спондина из дифференцировочной среды EEC снижает эффективность дифференцировки EEC (см. пример 5). Предполагают, что некоторая передача сигнала Wnt может потребоваться для направления клеток в сторону секреторного пути (а не поглощающего). Таким образом, в некоторых вариантах осуществления дифференцировочная среда содержит агонист Wnt (в частности, R-спондин) и ингибитор Wnt. Например, в некоторых вариантах осуществления дифференцировочная среда содержит R-спондин и ингибитор Porc, такой как IWP-2. В некоторых вариантах осуществления R-спондин используют в конечной концентрации от 1 до 1000 нг/мл, от 50 до 1000 нг/мл или от 100 до 1000 нг/мл. В некоторых вариантах осуществления R-спондин используют в конечной концентрации от 0,1 до 100 мкг/мл, от 0,1 до 50 мкг/мл, от 0,1 до 20 мкг/мл, от 0,1 до 10 мкг/мл, от 0,1 до 5 мкг/мл, от 0,5 до 100 мкг/мл, от 0,5 до 50 мкг/мл, от 0,5 до 20 мкг/мл, от 0,5 до 10 мкг/мл, от 0,5 до 5 мкг/мл, от 1 до 10 мкг/мл или от 1 до 5 мкг/мл. В некоторых вариантах осуществления R-спондин используют в конечной концентрации по меньшей мере 1 нг/мл, по меньшей мере 50 нг/мл, по меньшей мере 100 нг/мл, по меньшей мере 500 нг/мл или по меньшей мере 1 мкг/мл. В некоторых вариантах осуществления R-спондин используют в конечной концентрации приблизительно 100 нг/мл. В некоторых вариантах осуществления R-спондин используют в конечной концентрации приблизительно 1 мкг/мл.
Ингибиторы пути EGFR
Дифференцировочная среда по изобретению содержит ингибитор пути EGFR. Можно использовать любой подходящий ингибитор, как определено в настоящем описании.
Рецептор эпидермального фактора роста (EGFR), также известный как ErbB1 или HER1, является рецептором поверхности клетки для членов семейства эпидермального фактора роста (EGF) внеклеточных белковых лигандов. EGFR принадлежит к семейству рецепторов HER, содержащему четыре родственных белка (EGFR(HER1/ErbB1), ErbB2(HER2), ErbB3(HER3) и ErbB4(HER4)). Известно, что рецепторы HER активируются посредством связывания с различными лигандами, включая EGF, TGFA, гепарин-связывающий EGF-подобный фактор роста, амфирегулин, бетацеллюлин и эпирегулин. После связывания лиганда с внеклеточным доменом рецептора, рецептор образует функционально активные димеры (EGFR-EGFR (гомодимер) или EGFR-HER2, EGFR-HER3, EGFR-HER4 (гетеродимер)). Димеризация индуцирует активацию тирозинкиназного домена, что приводит к аутофосфорилированию рецептора по множеству остатков тирозина. Это приводит к рекрутированию множества адаптерных белков (таких как SHC, GRB2) и активации серии внутриклеточных каскадов передачи сигнала для осуществления транскрипции гена.
Пути, опосредующие нисходящие эффекты EGFR, хорошо исследованы и идентифицированы три основных пути передачи сигнала. Первый путь включает путь RAS-RAF-MAPK, где фосфорилированный EGFR рекрутирует фактор обмена гуаниновых нуклеотидов через адаптерные белки GRB2 и Shc, активируя RAS, а затем стимулируя RAF и путь киназы MAP для осуществления пролиферации клеток, опухолевой инвазии и метастазирования. Активированный RAS активирует протеинкиназную активность киназы RAF. Киназа RAF фосфорилирует и активирует MEK (также известный как MAP2K или MAPKK), фосфорилирующий и активирующий киназу MAP (также известный как ERK, внеклеточная сигнал-регулирующая киназа). Второй путь включает путь PI3K/AKT, активирующий основной клеточный сигнал выживания и антиапоптотический сигнал посредством активации ядерных факторов транскрипции, таких как NFKB. Третий путь включает путь JAK/STAT, также вовлеченный в активацию транскрипции генов, ассоциированных с выживанием клетки. Активация EGFR также может приводить к фосфорилированию PLCG и последующему гидролизу фосфатидилинозитол-4,5-бифосфата (PIP2) в инозитол-1,4,5-трифосфат (IP3) и диацилглицерин (DAG), что приводит к активации протеинкиназы C (PRKC) и CAMK.
Доступны ингибиторы EGFR, такие как моноклональные антитела против EGFR и низкомолекулярные ингибиторы тирозинкиназ EGFR. Некоторые антитела против EGFR, такие как цетуксимаб и панитумумаб, связываются с внеклеточным доменом мономера EGFR и конкурируют за связывание с рецептором с эндогенными лигандами; таким образом, они блокируют индуцируемую лигандом активацию рецептора. Некоторые низкомолекулярные ингибиторы EGFR, такие как эрлотиниб, гефитиниб и лапатиниб, конкурируют с АТФ за связывание с киназным доменом EGFR, что, в свою очередь, ингибирует аутофосфорилирование EGFR и нисходящую передачу сигнала.
Можно использовать один или несколько ингибиторов пути EGFR, например, 2, 3, 4 или более.
Ингибитор пути EGFR, предпочтительно, добавляют в среды в количестве, эффективном для ингибирования активности пути EGFR в клетке по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - 100% относительно уровня активности указанного пути EGFR в отсутствие указанной молекулы, как оценивают с использованием того же типа клеток. Как известно специалисту в этой области, активность пути EGFR можно измерять множеством способов. Например, анализ для мониторинга активности EGFR и чувствительности ингибитора описывают в Ghosh et al., (2013), Assay and Drug Development Technologies 11(1):44-51. Этот конкретный анализ включает пептидные субстраты, ковалентно иммобилизованные на магнитных частицах. После киназных реакций частицы промывают и определяют фосфорилирование пептидов посредством хемифлуоресценции с использованием HRP-конъюгированного первичного антитела против фосфорилированного тирозина. Измеренная интенсивность флуоресценции прямо пропорциональна фосфорилированию субстрата, которое, в свою очередь, пропорционально киназной активности EGFR. Этот анализ также можно использовать для скрининга на ингибиторы других киназ пути EGFR (например, RAS, RAF, MEK или ERK). Альтернативный способ анализа киназной активности включает детекцию встраивания концевого фосфата из P32-меченого АТФ. Таким образом, специалист в этой области легко может идентифицировать новые ингибиторы пути EGFR с использованием анализа, известного в этой области.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором EGFR, ингибирующим киназную активность EGFR по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - на 100%.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором RAS, ингибирующим киназную активность RAS по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - на 100%.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором RAF, ингибирующим киназную активность RAF по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - на 100%.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором MEK, ингибирующим киназную активность MEK по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - на 100%.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором ERK, ингибирующим киназную активность ERK по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - на 100%.
В некоторых вариантах осуществления EGF присутствует в дифференцировочной среде в концентрации менее 1 мМ. В предпочтительном варианте осуществления EGF отсутствует в дифференцировочной среде.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором EGFR, таким как гефитиниб (Santa Cruz Biotechnology), AG-18, AG-490 (тирфостин B42), AG-1478 (тирфостин AG-1478), AZ5104, AZD3759, бригатиниб, эрлотиниб, цетуксимаб, CL-387785 (EKI-785), CNX-2006, икотиниб, нецитумумаб, осимертиниб (AZD9291), OSI-420, PD153035 HCl, PD168393, пелитиниб (EKB-569), роцилетиниб (CO-1686, AVL-301), TAK-285, тирфостин 9, вандетаниб, WHI-P154, WZ3146, WZ4002, WZ8040, панитумумаб, залутумумаб, нимотузумаб или матузумаб. В некоторых вариантах осуществления ингибитор EGFR связывается с внеклеточным доменом мономера EGFR и конкурирует с EGF за связывание с рецептором. В некоторых вариантах осуществления ингибитор EGFR конкурирует с АТФ за связывание с киназным доменом EGFR. Можно использовать один или несколько ингибиторов EGFR, например, 2, 3, 4 или более.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором EGFR и ErbB-2, таким как афатиниб (Selleckchem), афатиниба дималеат, AC480 (BMS-599626), AEE788 (NVP-AEE788), AST-1306, канертиниб, CUDC-101, дакомитиниб, лапатиниб, нератиниб, позиотиниб (HM781-36B), сапитиниб (AZD8931) или варлитиниб. Можно использовать один или несколько ингибиторов EGFR и ErbB-2, например, 2, 3, 4 или более.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором пути RAS-RAF-MAPK. В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором пути PI3K/AKT. В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором пути JAK/STAT.
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором RAF, таким как GW5074, ZM 336372, NVP-BHG712, TAK-632, дарафениб (GSK2118436), сорафениб, сорафениба тозилат, PLX-4720, AZ 628, CEP-32496 или вемурафениб (PLX4032, RG7204).
В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором MEK, таким как PD0325901 (Sigma Aldrich). В некоторых вариантах осуществления ингибитор пути EGFR является ингибитором ERK, таким как SCH772984 (Selleckchem).
В некоторых вариантах осуществления ингибитор пути EGFR используют в концентрации 0,01-200 мкМ, 0,01-100 мкМ, 0,1-50 мкМ, 0,1-20 мкМ, 1-100 мкМ, 1-50 мкМ, 1-30 мкМ, 5-100 мкМ, 5-50 мкМ или 5-20 мкМ. Например, в некоторых вариантах осуществления дифференцировочная среда содержит: (i) гефитиниб в концентрации приблизительно 5 мкМ, (ii) афатиниб в концентрации приблизительно 10 мкМ, (iii) PD0325901 в концентрации приблизительно 5 мкМ или (iv) SCH772984 в концентрации приблизительно 10 мкМ.
Ингибитор Notch
Дифференцировочная среда содержит ингибитор Notch. Можно использовать любой подходящий ингибитор Notch.
Notch является трансмембранным поверхностным рецептором, который может активироваться посредством множественного протеолитического расщепления, в том числе расщепления комплексом белков с протеазной активностью, обозначенного как гамма-секретаза. Гамма-секретаза является протеазой, осуществляющей свою расщепляющую активность в мембране. Гамма-секретаза является многокомпонентным ферментом и состоит из по меньшей мере четырех разных белков, а именно, пресенилинов (пресенилин 1 или 2), никастрина, PEN-2 и APH-I. Пресенилин представляет собой каталитический центр гамма-секретазы. После связывания лиганда рецептор Notch подвергается конформационному изменению, позволяющему эктодомену отщепляться под действием протеазы ADAM, являющейся металлопротеазой. Сразу после этого действует гамма-секретазный комплекс, что приводит к высвобождению внутриклеточного домена Notch (NICD). NICD транслоцируется в ядро, где он взаимодействует с CSL (C-промотор-связывающим фактором/рекомбинантный белком Jκ, связывающимся с сигнальной последовательностью/супрессором Hairless/Lagl). Связывание NICD преобразует CSL из транскрипционного репрессора в активатор, что приводит к экспрессии генов-мишеней Notch.
В некоторых вариантах осуществления ингибитор Notch является ингибитором, способным снижать опосредованную лигандом активацию Notch (например, посредством доминантного отрицательного лиганда Notch, или доминантного отрицательного Notch, или антитела, способного, по меньшей мере частично, блокировать взаимодействие между лигандом Notch и Notch), или ингибитором протеаз ADAM.
В некоторых вариантах осуществления ингибитор Notch является ингибитором гамма-секретазы, например DAPT, дибензазепином (DBZ), бензодиазепином (BZ) или LY-411575. Можно использовать один или несколько ингибиторов Notch, например, 2, 3, 4 или более.
В некоторых вариантах осуществления ингибитор Notch (например, DAPT) используют в концентрации 0,001-200 мМ, 0,01-100 мМ, 0,1-50 мМ, 0,1-20 мМ, 0,5-10 мМ или 0,5-5 мМ. Например, в некоторых вариантах осуществления дифференцировочная среда содержит DAPT в концентрации приблизительно 1 мМ.
Минимальная среда
Дифференцировочная среда, используемая в способе по изобретению, содержит минимальную среду. Минимальная среда является любой подходящей минимальной средой для клеток животного или человека с учетом ограничений, предусмотренных в настоящем описании.
Минимальные среды для культивирования клеток животного или человека, как правило, содержат большое количество ингредиентов, необходимых для поддержания культивируемых клеток. Специалист в этой области легко может составлять подходящие комбинации ингредиентов с учетом следующего описания. Минимальная среда для применения в изобретении, как правило, будет содержать раствор питательных веществ, содержащий стандартные ингредиенты для культивирования клеток, такие как аминокислоты, витамины, липидные добавки, неорганические соли, углеродные источники энергии и буфер, как более подробно описано в литературе и настоящем описании выше. В некоторых вариантах осуществления среду для культивирования дополнительно дополняют одним или несколькими стандартными ингредиентами для культивирования клеток, например, выбранных из аминокислот, витаминов, липидных добавок, неорганических солей, углеродного источника энергии и буфера.
Специалисту в этой области с использованием общих знаний будут известны типы сред для культивирования, которые можно использовать в качестве минимальной среды в дифференцировочной среде по изобретению. Потенциально подходящие среды для культивирования клеток доступны в коммерческих источниках и включают, в качестве неограничивающих примеров, модифицированная по способу Дульбекко среда Игла (DMEM), минимальная поддерживающая среда (MEM), Knockout-DMEM (KO-DMEM), минимальная поддерживающая среда Глазго (G-MEM), минимальная среда Игла (BME), DMEM/F12 Хэма, продвинутая DMEM/F12 Хэма, среда Дульбекко в модификации Искова и минимальная поддерживающая среда (MEM), F-10 Хэма, F-12 Хэма, среда 199 и среды RPMI 1640.
Например, минимальную среду можно выбирать из DMEM/F12 и RPMI 1640, дополненных глутамином, инсулином, пенициллином/стрептомицином и трансферрином. В дополнительном предпочтительном варианте осуществления используют продвинутую DMEM/F12 или продвинутую RPMI, оптимизированную для бессывороточного культивирования и уже включающую инсулин. В этом случае, указанную продвинутую среду DMEM/F12 или продвинутую среду RPMI, предпочтительно, дополняют глутамином и пенициллином/стрептомицином. Продвинутая DMEM/F12 (Invitrogen), дополненная N2 и B27, также является предпочтительной. Предпочтительно, минимальная среда является продвинутой DMEM/F12. Более предпочтительно, минимальная среда содержит продвинутую DMEM/F12, глутамин и B27.
В некоторых вариантах осуществления минимальная среда содержит продвинутую DMEM/F12, HEPES, пенициллин/стрептомицин, глутамин, N-ацетилцистеин и B27.
В некоторых вариантах осуществления минимальная среда для культивирования содержит или состоит из продвинутой DMEM/F12, дополненной пенициллином/стрептомицином, 10 мМ HEPES, глютамакс, B27 (все от Life Technologies, Carlsbad, CA) и приблизительно 1 мМ N-ацетилцистеина (Sigma).
Кроме того, предпочтительно, указанную минимальную среду для культивирования дополняют очищенным, природным, полусинтетическим и/или синтетическим фактором роста, и она не содержит неопределенный компонент, такой как эмбриональная телячья сыворотка. Различные составы заменителей сыворотки являются коммерчески доступными и известны специалисту в этой области. Если используют заменитель сыворотки, его можно использовать общепринятыми способами в количестве от приблизительно 1% до приблизительно 30% по объему среды.
Дифференцировочная среда, используемая в изобретении, может содержать сыворотку или может являться бессывороточной и/или не содержащей заменитель сыворотки, как описано где-либо в настоящем описании. Среды для культивирования и препараты клеток, предпочтительно, соответствуют требованиям GMP и стандартам FDA для биологических продуктов и для обеспечения соответствия продукции.
Дифференцировочную среду по изобретению, как правило, будут составлять в деионизированной, дистиллированной воде. Дифференцировочную среду по изобретению, как правило, будут стерилизовать перед использованием для предотвращения контаминации, например, посредством ультрафиолетового излучения, нагревания, облучения или фильтрации. Дифференцировочную среду можно замораживать (например, при -20°C или -80°C) для хранения или транспортировки. Среда может содержать один или несколько антибиотиков для предотвращения контаминации. Среда может иметь содержание эндотоксина менее 0,1 единиц эндотоксина на мл или менее 0,05 единиц эндотоксина на мл. В этой области известны способы определения содержания эндотоксина в средах для культивирования.
Предпочтительная минимальная среда для культивирования является определенной синтетической средой, забуференной при pH 7,4 (предпочтительно - pH 7,2-7,6 или по меньшей мере 7,2 и не выше 7,6) буфером на основе карбоната, в то время как клетки культивируют в атмосфере, содержащей от 5% до 10% CO2, или по меньшей мере 5% и не более 10% CO2, предпочтительно - 5% CO2.
В некоторых вариантах осуществления дифференцировочная среда содержит минимальную среду, готовую для использования в способе дифференцировки. В других вариантах осуществления дифференцировочная среда не содержит минимальную среду, например, в виде добавки для среды для культивирования, и минимальную среду (или другие компоненты) можно добавлять перед использованием в способе дифференцировки. Таким образом, настоящее изобретение относится к любой дифференцировочной среде, представленной в настоящем описании, где минимальная среда отсутствует, или где минимальная среда является лишь необязательным компонентом.
Дополнительные факторы
Ранее показано, что ряд факторов усиливает дифференцировку клеток-предшественников в некоторых контекстах. В некоторых вариантах осуществления один или несколько из этих факторов включают в дифференцировочную среду по изобретению. Эти факторы включают, в качестве неограничивающих примеров, ингибиторы p38, ингибиторы TGF-бета, гастрин, глюкокортикоиды, лиганды рецепторной тирозинкиназы, активаторы пути BMP, активаторы Hedgehog, ингибиторы Hedgehog, модуляторы передачи сигнала mTOR, ингибиторы GSK-3, агонисты CHIR99021, стимуляторы AP-1, агонисты мускариновых ацетилхолиновых рецепторов, карбахоловые агонисты и активаторы пути цАМФ. В некоторых вариантах осуществления эти факторы выбраны из ингибиторов p38, ингибиторов TGF-бета, гастрина, глюкокортикоидов, лигандов рецепторной тирозинкиназы, активаторов пути BMP, ингибиторов пути BMP, активаторов Hedgehog, ингибиторов Hedgehog, модуляторов передачи сигнала mTOR, ингибиторов GSK-3, агонистов CHIR99021, стимуляторов AP-1, агонистов мускариновых ацетилхолиновых рецепторов, карбахоловых агонистов и активаторов пути цАМФ. В некоторых вариантах осуществления эти факторы выбраны из гастрина, глюкокортикоидов, лигандов рецепторной тирозинкиназы, активаторов пути BMP, ингибиторов пути BMP, активаторов Hedgehog, ингибиторов Hedgehog, модуляторов передачи сигнала mTOR, ингибиторов GSK-3, агонистов CHIR99021, стимуляторов AP-1, агонистов мускариновых ацетилхолиновых рецепторов, карбахоловых агонистов и активаторов пути цАМФ.
Ингибитор p38
В некоторых вариантах осуществления изобретения дифференцировочная среда дополнительно содержит ингибитор p38, являющийся любым ингибитором, который прямо или косвенно отрицательно регулирует передачу сигнала p38. В некоторых вариантах осуществления ингибитор по изобретению связывается с p38 и снижает его активность (GI №1432). Протеинкиназы p38 являются частью семейства митоген-активируемых протеинкиназ (MAPK). MAPK являются серин/треонин-специфическими протеинкиназами, отвечающими на внеклеточные стимулы, такие как внешний стресс и воспалительные цитокины, и регулирует различные активности клеток, такие как экспрессия генов, митоз, дифференцировка, пролиферация и выживание клеток/апоптоз. p38 MAPK существуют в виде изоформ α, β, β2, γ и δ. Ингибитор p38 является средством, связывающимся и снижающим активность по меньшей мере одной изоформы p38. Известны различные способы определения того, является вещество ингибитором p38, и их можно использовать в комбинации с изобретением. Примеры включают фосфо-специфическую детекцию фосфорилирования Thr180/Tyr182, представляющего собой хорошо известную меру клеточной активации и ингибирования p38, с помощью антител; биохимические анализы с использованием рекомбинантных киназ; анализы секреции фактора некроза опухоли альфа (ФНО) и высокопроизводительную скрининговую платформу DiscoverRx для ингибиторов p38 (см. http://www.discoverx.com/kinases/litrature/biochemical/collaterals/DRx_poster_p38%20KBA.pdf). Также существует несколько наборов для анализа активности p38 (например, Millipore, Sigma-Aldrich).
В этой области известны различные ингибиторы p38. В некоторых вариантах осуществления ингибитор, прямо или косвенно отрицательно регулирующий передачу сигнала p38, выбран из группы, состоящей из SB-202190, SB-203580, VX-702, VX-745, PD-169316, RO-4402257 и BIRB-796.
В одном из вариантов осуществления ингибитор p38 по изобретению связывается и снижает активность его мишени более чем на 10%; более чем на 30%; более чем на 60%; более чем на 80%; более чем на 90%; более чем на 95%; или более чем на 99% по сравнению контролем, что оценивают с помощью клеточного анализа. Примеры клеточных анализов для измерения ингибирования мишени хорошо известны в этой области, как описано выше.
SB-203580 можно добавлять в дифференцировочную среду в концентрации от 50 нМ до 100 мкМ, или от 100 нМ до 50 мкМ, или от 1 мкМ до 50 мкМ. Например, SB-203580 можно добавлять в среду для культивирования в количестве приблизительно 30 мкМ.
Ингибитор TGF-бета
В некоторых вариантах осуществления дифференцировочная среда дополнительно содержит ингибитор TGF-бета.
Передача сигнала TGF-бета вовлечена во множество функций клеток, включая рост клеток, направление дифференцировки клеток и апоптоз. Передача сигнала, как правило, начинается со связывания лиганда суперсемейства TGF-бета с рецептором типа II, рекрутирующим и фосфорилирующим рецептор типа I. Затем рецептор типа I фосфорилирует SMAD, действующие в качестве факторов транскрипции в ядре и регулирующие экспрессию гена-мишени.
Ранее показано, что ингибитор пути передачи сигнала TGF-бета участвует в стимуляции дифференцировки клеток-предшественников. Например, добавление TGF-бета к эксплантатам печени облегчает дифференцировку клеток желчных протоков in vitro (Clotman et al., (2005), Genes Dev. 19(16):1849-54). Кроме того, ранее показано, что включение ингибитора TGF-бета в дифференцировочную среду может ингибировать судьбу клеток желчных протоков и запускать дифференцировку клеток в сторону более гепатоцитарного фенотипа (см. WO 2012/168930). В частности, обнаружено, что включение ингибитора TGF-бета (такого как A83-01) в дифференцировочную среду повышает экспрессию маркеров зрелых гепатоцитов и повышает количество гепатоцит-подобных клеток.
Лиганды суперсемейства TGF-бета включает морфогенетические белки костей (BMP), факторы роста и факторы дифференцировки (GDF), антимюллеров гормон (AMH), активин, Nodal и TGF-бета. В основном, Smad2 и Smad3 фосфорилируются ALK4, 5 и 7 рецепторами в пути TGF-бета/активина. В отличие от этого, Smad1, Smad5 и Smad8 фосфорилируются как часть пути морфогенетического белка кости (BMP). Хотя в некоторых точках эти пути пересекаются, в контексте настоящего изобретения "ингибитор TGF-бета" или "ингибитор передачи сигнала TGF-бета", предпочтительно, является ингибитором пути TGF-бета, действующим через Smad2 и Smad3 и/или через ALK4, ALK5 или ALK7. Таким образом, в некоторых вариантах осуществления ингибитор TGF-бета не является ингибитором BMP, т.е. ингибитор TGF-бета не является Noggin. В некоторых вариантах осуществления ингибитор BMP добавляют в среду для культивирования в дополнение к ингибитору TGF-бета. Таким образом, ингибитором TGF-бета может являться любое средство, снижающее активность пути передачи сигнала TGF-бета, предпочтительно, пути передачи сигнала, действующего через Smad2 и/или Smad3, более предпочтительно, пути передачи сигнала, действующего через ALK4, ALK5 или ALK7.
Существует множество способов нарушения пути передачи сигнала TGF-бета, известных в этой области и которые можно использовать в комбинации с настоящим изобретением. Например, передачу сигнала TGF-бета можно нарушать посредством: ингибирования экспрессии TGF-бета с помощью стратегии малых интерферирующих РНК; ингибирования фурина (TGF-бета-активирующей протеазы); ингибирования пути с помощью физиологических ингибиторов; нейтрализации TGF-бета с использованием моноклонального антитела; ингибирования с помощью низкомолекулярных ингибиторов киназы 1 рецептора TGF-бета (также известной как киназа, подобная рецептору активина, ALK5), ALK4, ALK6, ALK7 или других TGF-бета-родственных рецепторных киназ; ингибирования передачи сигнала Smad 2 и Smad 3, например, посредством гиперэкспрессии их физиологического ингибитора, Smad 7, или с использованием тиоредоксина в качестве якоря для Smad, препятствующего активации Smad (Fuchs, O. Inhibition of TGF-Signalling for the Treatment of Tumor Metastasis and Fibrotic Diseases. Current Signal Transduction Therapy, Volume 6, Number 1, January 2011, pp. 29-43(15)).
Известны различные способы определения того, является ли вещество ингибитором TGF-бета, и их можно использовать в комбинации с изобретением. Например, можно использовать клеточный анализ, в котором клетки стабильно трансфицируют с использованием репортерной конструкции, содержащей промотор PAI-1 человека или участки связывания Smad, регулирующей люциферазный репортерный ген. Ингибирование активности люциферазы относительно контрольных групп можно использовать в качестве меры активности соединения (De Gouville et al., (2005), Br J Pharmacol. 145(2): 166-177). Таким образом, специалист в этой области легко может идентифицировать новые ингибиторы TGF-бета.
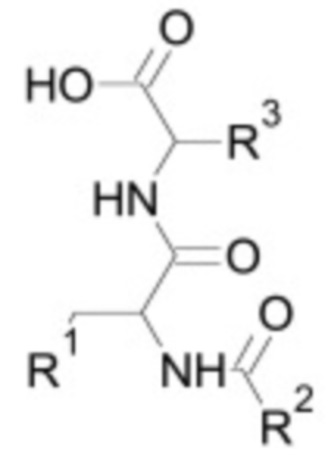
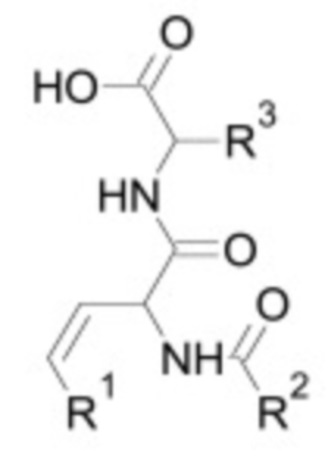
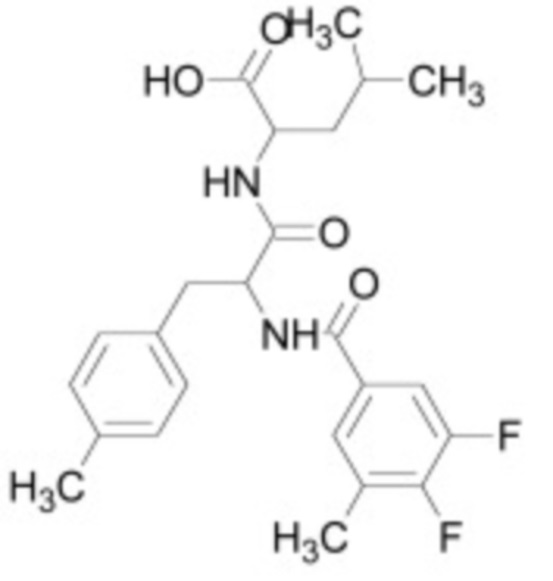
Ингибитор TGF-бета по настоящему изобретению может являться белком, пептидом, низкомолекулярным соединением, малой интерферирующей РНК, антисмысловым олигонуклеотидом, аптамером или антителом. Ингибитор может являться природным или синтетическим. В одном из вариантов осуществления ингибитор TGF-бета является ингибитором ALK4, ALK5 и/или ALK7. Например, ингибитор TGF-бета может связываться и напрямую ингибировать ALK4, ALK5 и/или ALK7. Неограничивающие примеры предпочтительных низкомолекулярных ингибиторов TGF-бета, которые можно использовать в контексте настоящего изобретения, включают низкомолекулярные ингибиторы, приведенные в таблице 2 ниже.
Таблица 2: Низкомолекулярные ингибиторы TGF-бета, воздействующие на рецепторные киназы
В некоторых вариантах осуществления ингибитор TGF-бета является низкомолекулярным ингибитором, необязательно, выбранным из группы, состоящей из: A83-01, SB-431542, SB-505124, SB-525334, LY 364947, SD-208 и SJN 2511.
В некоторых вариантах осуществления в дифференцировочной среде присутствует не более одного ингибитора TGF-бета. В других вариантах осуществления в дифференцировочной среде присутствуют несколько ингибиторов TGF-бета, например, 2, 3, 4 или более. В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит один или несколько из любых ингибиторов, приведенных в таблице 2. Дифференцировочная среда может содержать любую комбинацию одного ингибитора с другим указанным ингибитором. Например, среда может содержать SB-525334 или SD-208 или A83-01; или SD-208 и A83-01. Специалисту в этой области будет понятно, что существует ряд других низкомолекулярных ингибиторов, созданных, главным образом, для воздействия на другие киназы, но в высоких концентрациях они также могут ингибировать киназы рецепторов TGF-бета. Например, SB-203580 является ингибитором MAP-киназы p38, который, как считают, в высоких концентрациях (например, приблизительно 10 мкМ или более) ингибирует ALK5. Любой такой ингибитор, ингибирующий путь передачи сигнала TGF-бета, также можно использовать в контексте настоящего изобретения.
В некоторых вариантах осуществления ингибитор TGF-бета (например, A83-01) присутствует в дифференцировочной среде в количестве по меньшей мере 1 нМ, например, по меньшей мере 5 нМ, по меньшей мере 50 нМ, по меньшей мере 100 нМ, по меньшей мере 300 нМ, по меньшей мере 450 нМ или, по меньшей мере 475 нМ. Например, ингибитор TGF-бета (например, A83-01) присутствует в дифференцировочной среде в количестве 1 нМ-200 мкМ, 10 нМ-200 мкМ, 100 нМ-200 мкМ, 1 мкМ-200 мкМ, 10 нМ-100 мкМ, 50 нМ-100 мкМ, 50 нМ-10 мкМ, 100 нМ-1 мкМ, 200 нМ-800 нМ, 350-650 нМ или приблизительно 500 нМ. Таким образом, в некоторых вариантах осуществления дифференцировочная среда содержит A83-01 в концентрации приблизительно 500 нМ.
В некоторых вариантах осуществления дифференцировочная среда не содержит ингибитор TGF-бета. В некоторых вариантах осуществления дифференцировочная среда не содержит A83-01.
Гастрин
В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит гастрин. В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит гастрин в концентрации 0,01-500 нМ, 0,1-100 нМ, 1-100 нМ, 1-20 нМ или 5-15 нМ. Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит гастрин в концентрации приблизительно 10 нМ.
Глюкокортикоид
В некоторых вариантах осуществления дифференцировочная среда дополнительно содержит глюкокортикоид.
Глюкокортикоиды представляют собой класс кортикостероидов, являющихся классом стероидных гормонов. Глюкокортикоиды являются кортикостероидами, связывающимися с рецептором глюкокортикоидов. Кортизол является наиболее важным глюкокортикоидом человека. Гидрокортизон является синтетической версией кортизола. Существует множество других синтетических глюкокортикоидов с родственными структурами (например, преднизон, преднизолон, метилпреднизолон, дексаметазон, бетаметазон, триамцинолон, беклометазон и флудрокортизона ацетат). Глюкокортикоиды имеют различную активность активации рецепторов глюкокортикоидов. Ее называют глюкокортикоидной активностью и, как правило, измеряют по сравнению с кортизолом. Обзор биохимии, фармакологии и механизма действия различных глюкокортикоидов приведен, например, в Cecil Textbook of Medicine (1988), стр. 128-130, и The Science и Practice of Pharmacy 20th Edition (2000), стр. 1363-1370.
В некоторых вариантах осуществления дифференцировочная среда содержит глюкокортикоид. Можно использовать любой подходящий глюкокортикоид. В некоторых вариантах осуществления глюкокортикоид выбран из одного или нескольких из следующего: кортизола, кортизона, гидрокортизона ацетата, гидрокортизона гидрохлорида, гидрокортизона валерата, преднизона, преднизолона, метилпреднизолона, дексаметазона, бетаметазона, бетаметазона дипропионата, бетаметазона валерата, триамцинолона, триамцинолона ацетонида, беклометазона, беклометазона дипропионата, флудрокортизона, флудрокортизона ацетата, флутиказона, флутиказона ацетонида, флутиказона пропионата, флунизолида, будезонида, клобетазола, клобетазола пропионата, дифлуразона, дифлуразона диацетата, галобетазола, галобетазола пропионата, амцинонида, дезоксиметазона, флуоцинонида, флуоцинонида ацетонида, галцинонида, мометазона, мометазона фуроата, флуандренолида, предникарбата, алклометазона, алклометазона дипропионата, дезонида, флуцинолона, флуцинолона ацетонида, прамоксина и прамоксина гидрохлорида.
Дексаметазон является одним из наиболее активных глюкокортикоидов и предпочтительным глюкокортикоидом для использования в дифференцировочной среде по изобретению. В некоторых вариантах осуществления глюкокортикоид является любым глюкокортикоидом с той же или более высокой глюкокортикоидной активностью относительно дексаметазона.
Бетаметазон и флудрокортизона ацетат также являются очень активными глюкокортикоидами. Таким образом, бетаметазон является предпочтительным глюкокортикоидом для использования в дифференцировочной среде по изобретению. В некоторых вариантах осуществления глюкокортикоид является любым глюкокортикоидом с той же или более высокой глюкокортикоидной активностью относительно бетаметазона. Таким образом, флудрокортизона ацетат является предпочтительным глюкокортикоидом для использования в дифференцировочной среде по изобретению. В некоторых вариантах осуществления глюкокортикоид является любым глюкокортикоидом с той же или более высокой глюкокортикоидной активностью относительно флудрокортизон ацетат.
В некоторых вариантах осуществления глюкокортикоид является любым глюкокортикоидом с той же или более высокой глюкокортикоидной активностью относительно кортизола.
Список примеров глюкокортикоидов для использования в дифференцировочной среде по изобретению представлен ниже. Представленная активность относится к пероральной дозе.
В некоторых вариантах осуществления глюкокортикоид (например, дексаметазон) используют в концентрации 0,01-150 мкМ, 0,1-15 мкМ, 0,5-10 мкМ или 1-5 мкМ. В предпочтительном варианте осуществления глюкокортикоид является дексаметазоном. Например, в некоторых вариантах осуществления дифференцировочная среда содержит дексаметазон в концентрации приблизительно 3 мкМ.
В некоторых вариантах осуществления дифференцировочная среда не содержит глюкокортикоид.
Лиганды рецепторных тирозинкиназ
В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит один или несколько лигандов рецепторных тирозинкиназ.
Рецепторные тирозинкиназы (RTK) являются высокоаффинными рецепторами поверхности клетки для полипептидных факторов роста, цитокинов и гормонов. RTK являются ключевыми регуляторами поддержания, роста и развития клеток, а также играют критическую роль в развитии и прогрессировании многих типов злокачественных новообразований. В контексте изобретения лиганд рецепторных тирозинкиназ является любым лигандом, активирующим RTK. Многие лиганды рецепторных тирозинкиназ являются митогенными факторами роста. Таким образом, в некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ в дифференцировочной среде включают один или несколько митогенных факторов роста.
Существует приблизительно 20 различных известных классов RTK, включая RTK класса I (семейство рецепторов EGF) (семейство ErbB), RTK класса II (семейство рецепторов инсулина), RTK класса III (семейство рецепторов PDGF), RTK класса IV (семейство рецепторов FGF), RTK класса V (семейство рецепторов VEGF), RTK класса VI (семейство рецепторов HGF), RTK класса VII (семейство рецепторов Trk), RTK класса VIII (семейство рецепторов Eph), RTK класса IX (семейство рецепторов AXL), RTK класса X (семейство рецепторов LTK), RTK класса XI (семейство рецепторов TIE), RTK класса XII (семейство рецепторов ROR), RTK класса XIII (семейство рецепторов DDR), RTK класса XIV (семейство рецепторов RET), RTK класса XV (семейство рецепторов KLG), RTK класса XVI (семейство рецепторов RYK), RTK класса XVII (семейство рецепторов MuSK). В некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ включают лиганды для одного, или нескольких, или всех из этих 20 классов RTK.
В некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ включают лиганд для RTK класса IV (семейства рецепторов FGF). В некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ включают лиганд для RTK класса VI (семейства рецепторов HGF). В некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ включают лиганд для RTK класса IV (семейства рецепторов FGF) и лиганд для RTK класса VI (семейства рецепторов HGF).
Таким образом, в некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ в дифференцировочной среде выбраны из группы, состоящей из фактора роста фибробластов (FGF) и фактора роста гепатоцитов (HGF). В некоторых вариантах осуществления один или несколько лигандов рецепторных тирозинкиназ включают FGF и HGF. В некоторых вариантах осуществления только один лиганд рецепторных тирозинкиназ включен в дифференцировочную среду, например, где рецепторная тирозинкиназа выбрана из FGF и HGF.
Предпочтительно, FGF является FGF, способным связываться с FGFR2 или FGFR4 и, предпочтительно, является FGF19.
В дифференцировочных средах по изобретению можно использовать три или более, например, 3, 4, 5 или более, лигандов рецепторных тирозинкиназ.
Как описано выше, авторы настоящего изобретения обнаружили, что ингибирование пути EGFR вовлечено в дифференцировку аргентофильных клеток. Таким образом, предпочтительно, один или несколько лигандов рецепторных тирозинкиназ не включают лиганд, активирующий EGFR (например, EGF). В некоторых вариантах осуществления дифференцировочная среда содержит менее 1 мМ EGF.
Активатор пути BMP
Авторы настоящего изобретения неожиданно обнаружили, что включение активатора пути BMP в EEC-дифференцировочную среду повышает присутствие секретин-секретирующих клеток в популяции дифференцированных EEC (см. фигуры 13 и 14). Таким образом, в некоторых вариантах осуществления дифференцировочная среда дополнительно содержит активатор пути BMP. В некоторых вариантах осуществления дифференцировочная среда не содержит ингибитор пути BMP (например, Noggin). В некоторых вариантах осуществления дифференцировочная среда дополнительно содержит активатор пути BMP и не содержит ингибитор пути BMP (например, Noggin).
Способы идентификации подходящих активаторов BMP известны в этой области. Подходящий анализ для измерения активности BMP описан в Zilberberg et al., BMC Cell Biology 2007 8:41.
В некоторых вариантах осуществления активатор пути BMP выбран из BMP7, BMP4 и BMP2. BMP7 является предпочтительным. BMP7 индуцирует фосфорилирование SMAD1 и SMAD5. Таким образом, в некоторых вариантах осуществления активатор пути BMP является любым соединением, способным индуцировать фосфорилирование SMAD1 и SMAD5. Кроме того, если упомянут BMP7, можно использовать любое соединение, индуцирующее фосфорилирование SMAD1 или SMAD5, вместо BMP7.
В некоторых вариантах осуществления активатор пути BMP, такой как BMP4 или BMP7, присутствует в дифференцировочной среде в количестве по меньшей мере 0,01 нг/мл, по меньшей мере 0,1 нг/мл, по меньшей мере 1 нг/мл, по меньшей мере 10 нг/мл, по меньшей мере 20 нг/мл, по меньшей мере 25 нг/мл, по меньшей мере 100 нг/мл, по меньшей мере 500 нг/мл, по меньшей мере 1 мкг/мл, по меньшей мере 10 мкг/мл или по меньшей мере 50 мкг/мл. В некоторых вариантах осуществления активатор пути BMP, такой как BMP4 или BMP7, присутствует в дифференцировочной среде в количестве от приблизительно 0,01 нг/мл до приблизительно 500 нг/мл, от приблизительно 1 нг/мл до приблизительно 500 нг/мл, от приблизительно 10 нг/мл до приблизительно 500 нг/мл, от приблизительно 20 нг/мл до приблизительно 500 нг/мл. В некоторых вариантах осуществления активатор пути BMP, такой как BMP4 или BMP7, присутствует в дифференцировочной среде в количестве от приблизительно 0,01 нг/мл до приблизительно 200 нг/мл, от приблизительно 0,1 нг/мл до приблизительно 100 нг/мл, от приблизительно 1 нг/мл до приблизительно 100 нг/мл, от приблизительно 10 нг/мл до приблизительно 100 нг/мл, от приблизительно 10 нг/мл до приблизительно 50 нг/мл, от приблизительно 15 нг/мл до приблизительно 30 нг/мл. В некоторых вариантах осуществления активатор пути BMP, такой как BMP4 или BMP7, присутствует в дифференцировочной среде в количестве приблизительно 25 нг/мл. В некоторых вариантах осуществления BMP4 присутствует в дифференцировочной среде от приблизительно 0,1 мкг/мл до приблизительно 50 мкг/мл, от приблизительно 1 до приблизительно 50 мкг/мл, от приблизительно 5 мкг/мл до приблизительно 25 мкг/мл или от приблизительно 5 мкг/мл до приблизительно 15 мкг/мл. В некоторых вариантах осуществления BMP4 присутствует в дифференцировочной среде в количестве приблизительно 10 мкг/мл.
В некоторых вариантах осуществления дифференцировочная среда не содержит активатор пути BMP.
Изобретение также относится к способу повышения секреции секретина в популяции клеток, содержащей EEC, где способ включает приведение популяции клеток в контакт с активатором BMP.
Изобретение также относится к способу снижения секреции GLP-1 в популяции клеток, содержащей EEC, где способ включает приведение популяции клеток в контакт с активатором BMP.
Изобретение также относится к способу повышения экспрессии Pyy и/или Nts в популяции клеток, содержащей EEC, где способ включает приведение популяции клеток в контакт с активатором BMP.
Ингибитор BMP
Авторы настоящего изобретения также неожиданно обнаружили, что включение ингибитора передачи сигнала BMP в EEC-дифференцировочную среду стимулирует гормональную сигнатуру крипт в популяции дифференцированных EEC (см. пример 5). Гормональная сигнатура крипт отличается высокой экспрессией GLP-1 и отсутствием секреции секретина. Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит ингибитор BMP. В этих вариантах осуществления дифференцировочная среда, предпочтительно, не содержит активатор BMP.
BMP являются небольшими сигнальными молекулами, связывающимися с двумя классами рецепторов морфогенетического белка кости на поверхности клетки (BMPR-I и BMPRII). Класс рецепторов BMPR-I состоит из трех типов рецепторов, подобной рецептору активина киназы-2 (ALK-2 или ActR-IA), ALK-3 (BMPR-IA) и ALK-6 (BMPR-IB). Класс рецепторов BMPR-II состоит из трех типов рецепторов, BMPR-II, ActR-IIA и ActR-IIB. Связывание BMP приводит к образованию гетеротетрамерных комплексов, содержащих два рецептора типа I и два рецептора типа II. В дополнение к внеклеточному связывающему домену, каждый рецептор BMP содержит внутриклеточный серин/треониновый киназный домен. После связывания BMP, конститутивно активные рецепторные киназы типа II фосфорилируют киназные домены рецептора типа I, которые, в свою очередь, фосфорилируют BMP-чувствительные SMAD 1, 5, и 8, которые могут проникать в ядро клетки и функционировать в качестве факторов транскрипции. Фосфорилирование этих конкретных SMAD приводит к различным клеточным эффектам, включая регуляцию роста и дифференцировку. Ингибитор BMP является любым ингибитором, приводящим к значительному снижению передачи сигнала через эти пути. Например, ингибитор BMP может нарушать взаимодействие BMP с рецептором BMP; связываться с рецептором BMP и ингибировать активацию нисходящей передачи сигнала; ингибировать фосфорилирование Smad 1, Smad 5 или Smad 8; ингибировать транслокацию Smad 1, Smad 5 или Smad 8 в ядро; ингибировать SMAD 1-, SMAD 5- или SMAD 8-опосредованную транскрипцию генов-мишеней или ингибировать экспрессию, фолдинг или секрецию BMP. В некоторых вариантах осуществления ингибитор BMP снижает передачу сигнала через рецептор класса BMPR-I. В некоторых вариантах осуществления ингибитор BMP снижает передачу сигнала через рецептор класса BMPR-II. В некоторых вариантах осуществления ингибитор BMP снижает передачу сигнала через SMAD 1/5/8. Ингибирование может быть прямым или косвенным.
В этой области известно множество ингибиторов BMP, например, как описано в Cuny, et al., (2008) Structure-activity relationship study of bone morphogenetic protein (BMP) signaling inhibitors. Bioorg Med Chem Lett 18: 4388-4392. Любые из этих ингибиторов BMP пригодны для использования в способах по изобретению. В этой области известны способы идентификации подходящих ингибиторов BMP. Подходящий анализ описывают в Zilberberg et al., BMC Cell Biology 2007 8:41. Специалист в этой области может определять другой подходящий анализ на ингибитор BMP (в частности, ингибитор BMP, ингибирующий фосфорилирование Smad 1, 5 или 8 через ALK2 и ALK3) с использованием клеточного анализа ELISA Cytobot, описанного в Cuny, et al., (2008) Structure-activity relationship study of bone morphogenetic protein (BMP) signaling inhibitors. Bioorg Med Chem Lett 18: 4388-4392.
В некоторых вариантах осуществления ингибитор BMP выбран из Noggin, хордина, фоллистатина, гремлина, tsg (белка скрученной гаструляции), sog (белка короткой гаструляции), дорсоморфина и LDN193189. В некоторых вариантах осуществления ингибитор BMP выбран из:
a. дорсоморфина или LDN193189 или его аналога или варианта; и/или
b. Noggin, склеростина, хордина, CTGF, фоллистатина, гремлина, tsg, sog или их аналога или варианта.
В некоторых предпочтительных вариантах осуществления ингибитором BMP является Noggin. Noggin особенно подходит для способов культивирования in vitro. В других предпочтительных вариантах осуществления ингибитором BMP является LDN193189. LDN193189 особенно подходит для способов лечения in vivo, т.к. он является перорально доступным и, таким образом, подходит для перорального введения (см. дополнительные комментарии в отношении способов лечения ниже).
В некоторых вариантах осуществления Noggin включают в дифференцировочную среду в конечной концентрации от 1 до 1000 нг/мл, от 10 до 1000 нг/мл, от 100 до 1000 нг/мл, от 1 до 500 нг/мл, от 1 до 200 нг/мл, от 1 до 100 нг/мл, от 10 до 500 нг/мл, от 20 до 500 нг/мл, от 10 до 200 нг/мл, от 20 до 200 нг/мл, от 50 до 500 нг/мл или от 50 до 200 нг/мл. В некоторых вариантах осуществления Noggin включают в дифференцировочную среду в конечной концентрации приблизительно 100 нг/мл.
В некоторых вариантах осуществления LDN193189 включают в дифференцировочную среду в конечной концентрации от 1 нМ до 10 мкМ, от 5 нМ до 10 мкМ, от 10 нМ до 10 мкМ, от 100 нМ до 10 мкМ, от 1 мкМ до 10 мкМ или от 1 мкМ до 5 мкМ. В некоторых вариантах осуществления LDN193189 включают в дифференцировочную среду в конечной концентрации по меньшей мере 1 нМ, по меньшей мере 5 нМ, по меньшей мере 10 нМ, по меньшей мере 100 нМ, по меньшей мере 1 мкМ или приблизительно 10 мкМ.
Изобретение также относится к способу повышения секреции GLP-1 в популяции клеток, содержащей EEC, включающему приведение популяции клеток в контакт с ингибитором BMP. Изобретение также относится к способу снижения секреции секретина в популяции клеток, содержащей EEC, включающему приведение популяции клеток в контакт с ингибитором BMP.
Изобретение также относится к способу повышения экспрессии Tac1 в популяции клеток, содержащей EEC, включающему приведение популяции клеток в контакт с ингибитором BMP.
Изобретение также относится к способу повышения экспрессии Gcg в популяции клеток, содержащей EEC, включающему приведение популяции клеток в контакт с ингибитором BMP.
Активаторы и ингибиторы Hedgehog
Авторы настоящего изобретения неожиданно обнаружили, что включение ингибитор передачи сигнала Hedgehog в EEC-дифференцировочную среду в некоторых контекстах может повышать присутствие GLP1- и CCK-секретирующих клеток в популяции дифференцированных EEC (см. фигура 14). Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит один или несколько ингибиторов Hedgehog. Один или несколько ингибиторов Hedgehog могут являться любым подходящим ингибитором, снижающим передачу сигнала Hedgehog в клетке. Эта среда подходит для получения популяции EEC, обогащенной GLP1- и CCK-секретирующими клетками. Альтернативно, популяцию EEC, в которой снижено количество GLP1- и CCK-секретирующих клеток, можно получать с использованием дифференцировочной среды по изобретению, дополнительно содержащей активатор Hedgehog. Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит один или несколько активаторов Hedgehog. Один или несколько активаторов Hedgehog могут являться любым подходящим активатором, повышающим передачу сигнала Hedgehog в клетке.
Лиганды Hedgehog (Hh) млекопитающих включают Sonic hedgehog (Shh), Indian hedgehog (Ihh) и Desert hedgehog (Dhh). Лиганды Hh, как правило, синтезируются в виде белков-предшественников, подвергающихся аутокаталитическому расщеплению и сопутствующей холестероловой модификации на карбокси-конце и пальмитоилированию на амино-конце, что приводит к образованию секретируемого, дважды липидированного белка. Лиганды Hh высвобождаются с поверхности клетки при комбинированном воздействии Dispatched и Scube2, а затем транспортируются через множество клеток посредством взаимодействий с белками LRP2 поверхности клетки и глипиканового семейства гепарансульфатных протеогликанов (GPC1-6).
Белки Hh инициируют передачу сигнала посредством связывания с каноническим рецептором Patched (PTCH1) и корецепторами GAS1, CDON и BOC. Связывание Hh с PTCH1 приводит к дерепрессии GPCR-подобного белка Smoothened (SMO), что приводит к накоплению SMO в ресничках и фосфорилированию его цитоплазматического хвоста. SMO опосредует нисходящую передачу сигнала, включающую диссоциацию белков GLI (транскрипционных эффекторов пути Hh) от белка из семейства кинезинов, Kif7, и ключевого внутриклеточного регулятора пути Hh SUFU.
В некоторых вариантах осуществления один или несколько ингибиторов Hedgehog включают ингибитор SMO. В некоторых вариантах осуществления ингибитор SMO является циклопамином или конкурентным ингибитором циклопамина (например, висмогенибом, саридегибом или циклопамином). В других вариантах осуществления ингибитор SMO не является конкурентным ингибитором циклопамина (например, итраконазолом).
В некоторых вариантах осуществления один или несколько ингибиторов Hedgehog включают антитело против PTCH1, ингибирующее связывание Hh с PTCH1 (см., например, Nakamura et al. (2007) Anticancer Research 27:3743-3748).
В некоторых вариантах осуществления один или несколько активаторов Hedgehog включают активатор SMO. В некоторых вариантах осуществления активатор SMO является SAG (Hh-Ag1.3) или пурморфамином.
Ингибиторы Hedgehog и активаторы Hedgehog можно идентифицировать известными в этой области способами, например, с использованием способа RT-ПЦР для определения уровней мРНК Gli1 и Ptch1 (см., например, Nakamura et al. (2007) Anticancer Research 27:3743-3748). Gli1 и Ptch1 являются генами-мишенями транс-активации Gli1, и, таким образом, их можно использовать в качестве маркеров активности пути передачи сигнала Hh. Например, супрессированная экспрессия Gli1 и Ptch1 является показателем сниженной активности пути передачи сигнала Hh.
В одном из вариантов осуществления ингибитор Hedgehog по изобретению снижает активность пути передачи сигнала Hedgehog более чем на 10%; более чем на 30%; более чем на 60%; более чем на 80%; более чем на 90%; более чем на 95% или более чем на 99% по сравнению контролем, как оценивают с помощью анализа RT-ПЦР. Примеры анализов RT-ПЦР для измерения ингибирования хорошо известны в этой области, как описано выше.
В одном из вариантов осуществления активатор Hedgehog по изобретению повышает активность пути передачи сигнала Hedgehog более чем на 10%; более чем на 30%; более чем на 60%; более чем на 80%; более чем на 90%; более чем на 95% или более чем на 99% по сравнению контролем, как оценивают с помощью анализа RT-ПЦР. Примеры анализов RT-ПЦР для измерения активации хорошо известны в этой области, как описано выше.
В некоторых вариантах осуществления ингибитор Hedgehog используют в концентрации 0,01-200 мкМ, 0,01-100 мкМ, 0,05-100 мкМ, 0,1-50 мкМ, 0,1-20 мкМ, 1-100 мкМ, 1-50 мкМ, 1-30 мкМ, 1-10 мкМ, 5-100 мкМ, 5-50 мкМ или 5-20 мкМ. В некоторых вариантах осуществления ингибитор Hedgehog (например, висмогениб) присутствует в концентрации приблизительно 5 мкМ.
В некоторых вариантах осуществления активатор Hedgehog используют в концентрации 0,01-200 мкМ, 0,01-100 мкМ, 0,05-100 мкМ, 0,1-50 мкМ, 0,1-20 мкМ, 1-100 мкМ, 1-50 мкМ, 1-30 мкМ, 1-10 мкМ, 5-100 мкМ, 5-50 мкМ или 5-20 мкМ. В некоторых вариантах осуществления активатор Hedgehog (например, SAG (Hh-Ag1.3) или пурморфамин) присутствует в концентрации приблизительно 5 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит два, три, четыре или более ингибитора Hedgehog.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит два, три, четыре или более активатора Hedgehog.
В некоторых вариантах осуществления дифференцировочная среда не содержит ингибитор Hedgehog.
Модуляторы передачи сигнала mTOR
Авторы настоящего изобретения неожиданно обнаружили, что включая модулятор передачи сигнала, mTOR может влиять на относительные соотношения подтипов EEC (например, активаторы mTOR могут способствовать дифференцировке в направлении GLU-экспрессирующих EEC). В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит один или несколько модуляторов передачи сигнала mTOR. В некоторых вариантах осуществления один или несколько модуляторов передачи сигнала mTOR включают ингибитор передачи сигнала mTOR. Один или несколько ингибиторов mTOR могут являться любым подходящим ингибитором, снижающим передачу сигнала mTOR в клетке. В некоторых вариантах осуществления один или несколько модуляторов передачи сигнала mTOR включают активатор передачи сигнала mTOR (например, MHY1485). Один или несколько активаторов mTOR могут являться любым подходящим активатором, повышающим передачу сигнала mTOR в клетке.
Мишень рапамицина в клетках млекопитающих (mTOR) является атипичной серин/треониновой киназой, присутствующей в двух отдельных комплексах. Первый, комплекс mTOR 1 (mTORC1), состоит из mTOR, Raptor, GβL и DEPTOR и ингибируется рапамицином. Он является основным регулятором роста, распознающим и интегрирующим различные питательные сигналы и сигналы из окружающей среды, включая факторы роста, уровни энергии, клеточный стресс и аминокислоты. Он объединяет эти сигналы для стимуляции роста клеток посредством фосфорилирования субстратов, потенцирующего анаболические процессы, такие как трансляция мРНК и синтез липидов, или ограничивающего катаболические процессы, такие как аутофагия. Небольшая ГТФаза Rheb в своем ГТФ-связанном состоянии является необходимым и мощным стимулятором активности киназы mTORC1, отрицательно регулируемой GAP, гетеродимером туберозного склероза TSC1/2. Большинство вышележащих входных сигналов направляются через Akt и TSC1/2 для регуляции состояния нагрузки нуклеотидами Rheb. И наоборот, аминокислоты передают сигнал на mTORC1 независимо от оси PI3K/Akt для стимуляции транслокации mTORC1 на поверхность липосом, где он активируется после контакта с Rheb. Этот процесс опосредуется координированным действием множества комплексов, а именно v-АТФазой, Ragulator, ГТФаз Rag и GATOR1/2. Второй комплекс, комплекс mTOR 2 (mTORC2), состоит из mTOR, Rictor, GβL, Sin1, PRR5/Protor-1 и DEPTOR. mTORC2 способствует выживанию клеток посредством активации Akt, регулирует динамику цитоскелета посредством активации PKCα и контролирует ионный транспорт и рост посредством фосфорилирования SGK1.
В некоторых вариантах осуществления активатором передачи сигнала mTOR является MHY1485.
В некоторых вариантах осуществления ингибитором передачи сигнала mTOR является рапамицин или аналог рапамицина (например, рапамицин, дефоролимус (AP23573), эверолимус (RAD001) и темсиролимус (CCI-779)). В некоторых вариантах осуществления ингибитор передачи сигнала mTOR является АТФ-конкурентным ингибитором киназы mTOR (например, MLN0128, pp242 или AZD8055).
Ингибиторы mTOR и активаторы mTOR можно идентифицировать известными в этой области способами, например, с использованием анализа киназы mTOR на основе ELISA (например, с использованием набора для анализа активности mTOR K-LISA™, представляющего собой анализ активности на основе ELISA для детекции фосфорилирования слитого белка p70S6K-GST (специфического субстрата mTOR) в присутствие АТФ).
В одном из вариантов осуществления ингибитор mTOR по изобретению снижает активность пути передачи сигнала mTOR более чем на 10%; более чем на 30%; более чем на 60%; более чем на 80%; более чем на 90%; более чем на 95% или более чем на 99% по сравнению контролем, как оценивают с помощью анализа на основе ELISA. Примеры анализов на основе ELISA для измерения ингибирования хорошо известны в этой области, как описано выше.
В одном из вариантов осуществления активатор mTOR по изобретению повышает активность пути передачи сигнала mTOR более чем на 10%; более чем на 30%; более чем на 60%; более чем на 80%; более чем на 90%; более чем на 95% или более чем на 99% по сравнению контролем, как оценивают с помощью анализа на основе ELISA. Примеры анализов на основе ELISA для измерения активации хорошо известны в этой области, как описано выше.
В некоторых вариантах осуществления ингибитор mTOR используют в концентрации 0,01-200 мкМ, 0,01-100 мкМ, 0,05-100 мкМ, 0,1-50 мкМ, 0,1-20 мкМ, 1-100 мкМ, 1-50 мкМ, 1-30 мкМ, 1-10 мкМ, 5-100 мкМ, 5-50 мкМ или 5-20 мкМ. В некоторых вариантах осуществления ингибитор mTOR (например, рапамицин, дефоролимус (AP23573), эверолимус (RAD001) или темсиролимус (CCI-779)) присутствует в концентрации приблизительно 5 мкМ.
В некоторых вариантах осуществления активатор mTOR используют в концентрации 0,01-200 мкМ, 0,01-100 мкМ, 0,05-100 мкМ, 0,1-50 мкМ, 0,1-20 мкМ, 1-100 мкМ, 1-50 мкМ, 1-30 мкМ, 1-10 мкМ, 5-100 мкМ, 5-50 мкМ или 5-20 мкМ. В некоторых вариантах осуществления активатор mTOR (например, MHY1485) присутствует в концентрации приблизительно 5 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит два, три, четыре или более ингибитора mTOR.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит два, три, четыре или более активатора mTOR.
Ингибитор GSK-3
Ранее обнаружено, что включение ингибитора GSK-3 (например, CHIR99021) также улучшает дифференцировку эпителиальных стволовых клеток (см. GB 1603569.3). Как описано выше, GSK-3 является компонентом деструктивного комплекса β-катенина. Ингибирующая активность GSK-3 приводит к снижению деструкции β-катенина, и, таким образом, ингибиторы GSK-3 являются агонистами Wnt. Усиленная дифференцировка, вызванная включением ингибитора GSK-3 в дифференцировочную среду, значительно повышается при добавлении ингибитора Wnt (iCRT3), ингибирующего TCF/LEF-опосредованную транскрипцию, нисходящую от деструктивного комплекса. Например, наблюдают значительное улучшение экспрессии маркеров гепатоцитов (альбумина и Cyp3A4), когда и агонист Wnt (CHIR99021) и ингибитор Wnt (iCRT3) включены в дифференцировочную среду для клеток-предшественников печени. Не желая быть связанными какой-либо теорией, авторы настоящего изобретения полагают, что ингибиторы GSK-3 могут стимулировать и ингибировать дифференцировку эпителиальных стволовых клеток (при этом стимуляция сильнее ингибирования). Считают, что ингибирование дифференцировки ингибиторами GSK-3 происходит посредством стимуляции деструкции β-катенина. И наоборот, считают, что стимуляция дифференцировки ингибиторами GSK-3 происходит с помощью другого эффекторного механизма. Таким образом, если в дифференцировочную среду включают ингибитор GSK-3, ингибитор Wnt должен быть ингибитором Wnt, ингибирующим TCF/LEF-опосредованную транскрипцию, нисходящую от деструктивного комплекса, в дифференцировочной среде, содержащей ингибитор GSK-3, препятствующим ингибирующим дифференцировку эффектам ингибитора GSK-3. Ингибиторы Wnt, действующие ниже деструктивного комплекса, включают ингибиторы Wnt в классе (7), упомянутом выше, т.е. ингибиторы экспрессии гена-мишени β-катенина, включая ингибиторы транскрипционного комплекса β-катенин : TCF/Lef, такие как ингибиторы, нарушающие комплекс β-катенин : TCF-4 (например, iCRT3, CGP049090, PKF118310, PKF115 584, ZTM000990, PNU 74654, BC21, iCRT5, iCRT14 или FH535) и ингибиторы гистондеацетилазы SIRT1 (например, камбинол).
Таким образом, эффекты стимуляции дифференцировки ингибиторов GSK-3 остаются незатронутыми и дополняют эффекты стимуляции дифференцировки ингибитора Wnt.
Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит ингибитор GSK-3. Можно использовать любой подходящий ингибитор GSK-3. Ингибитор GSK-3 определяют как средство, снижающее активность киназы GSK-3.
У людей обнаружены две разные изоформы GSK-3 (GSK-3α и GSK-3β). Известны ингибиторы, ингибирующие одну или обе эти изоформы. Таким образом, в некоторых вариантах осуществления ингибитор GSK-3 ингибирует GSK-3α и GSK-3β. В некоторых вариантах осуществления ингибитор GSK-3 ингибирует GSK-3α, но не ингибирует GSK-3β. В некоторых вариантах осуществления ингибитор GSK-3 ингибирует GSK-3β, но не ингибирует GSK-3α.
В некоторых вариантах осуществления дифференцировочная среда содержит несколько ингибиторов GSK-3 (например, два, три, четыре, пять или более ингибиторов GSK-3).
В некоторых вариантах осуществления ингибитором GSK-3 является CHIR99021.
В некоторых вариантах осуществления ингибитор GSK-3 добавляют в среды в количестве, эффективном для ингибирования активности GSK-3 в клетке по меньшей мере на 10%, более предпочтительно - по меньшей мере на 20%, более предпочтительно - по меньшей мере на 30%, более предпочтительно - по меньшей мере на 50%, более предпочтительно - по меньшей мере на 70%, более предпочтительно - по меньшей мере на 90%, более предпочтительно - на 100% относительно уровня указанной активности GSK-3 в отсутствие указанной молекулы, как оценивают с использованием того же типа клеток. Как известно специалисту в этой области, активность GSK-3 можно измерять посредством мониторинга фосфорилирования специфических субстратов GSK-3, например, β-катенина (см., например, Cole et al., (2008), Methods Mol Biol. 468:45-65). Таким образом, специалист в этой области легко может идентифицировать новые ингибиторы GSK-3 с использованием анализа, известного в этой области.
В некоторых вариантах осуществления ингибитор GSK-3 выбран из одного или нескольких из следующего: CHIR99201, 6-BIO, дибромокантареллина, химениалдезина, индирубинов, меридианинов, CT98014, CT98023, CT99021, TWS119, SB-216763, SB-41528, AR-A014418, AZD-1080, алстерпауллона, казпауллона, кенпауллона, алоизинов, манзамина A, палинурина, трикантина, TDZD-8, NP00111, NP031115, тидеглусиба, HMK-32 и L803-mts.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит ингибитор GSK-3 в концентрации 0,001-500 мкМ, 0,01-150 мкМ, 0,1-100 мкМ, 1-100 мкМ, 0,5-50 мкМ, 1-50 мкМ, 1-20 мкМ или 1-5 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит CHIR99021 в концентрации 0,01-150 мкМ, 0,1-100 мкМ, 1-100 мкМ, 0,5-50 мкМ, 1-50 мкМ, 1-20 мкМ или 1-5 мкМ. Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит CHIR99021 в концентрации приблизительно 3 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит ингибитор GSK-3 и ингибитор Wnt, действующий ниже деструктивного комплекса, такой как ингибиторы экспрессии гена-мишени β-катенина, как описано выше. Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит ингибитор GSK-3 и ингибитор экспрессии гена-мишени β-катенина. Ингибиторы экспрессии гена-мишени β-катенина включают ингибиторы транскрипционного комплекса β-катенин : TCF/Lef, такие как ингибиторы, нарушающие комплекс β-катенин : TCF-4 (например, iCRT3, CGP049090, PKF118310, PKF115-584, ZTM000990, PNU-74654 или BC21), и ингибиторы гистондеацетилазы SIRT1 (например, камбинол).
Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит ингибитор GSK-3 (например, в концентрации 0,001-500 мкМ, 0,01-150 мкМ, 0,1-100 мкМ, 0,5-50 мкМ, 1-20 мкМ или 1-5 мкМ) и ингибитор экспрессии гена-мишени β-катенина (например, в концентрации 0,001-500 мкМ, 0,01-150 мкМ, 0,1-100 мкМ, 0,5-50 мкМ, 1-500 мкМ, 1-150 мкМ, 1-100 мкМ, 1-50 мкМ, 1-20 мкМ или 1-5 мкМ). В некоторых вариантах осуществления ингибитор экспрессии гена-мишени β-катенина является ингибитором транскрипционного комплекса β-катенин : TCF/Lef.
Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит CHIR99021 (например, в концентрации 0,01-150 мкМ, 0,1-100 мкМ, 0,5-50 мкМ, 1-20 мкМ или 1-5 мкМ) и iCRT3 (например, в концентрации 0,05-150 мкМ, 0,1-150 мкМ, 1-150 мкМ, 0,5-100 мкМ, 1-100 мкМ или 10-80 мкМ). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит CHIR99021 в концентрации приблизительно 3 мкМ и iCRT3 в концентрации приблизительно 50 мкМ.
В некоторых вариантах осуществления дифференцировочная среда не содержит ингибитор GSK-3.
Агонисты CHIR99021
Как описано выше, ранее обнаружено, что включение агониста Wnt CHIR99021 в дифференцировочную среду способствует дифференцировке. Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит CHIR99021 или агонист CHIR99021. Термин "агонист CHIR99021" в настоящем описании определяют как средство, имеющее общие один или несколько видов биологической активности с CHIR99021.
Стимуляторы AP-1
Комплекс активирующего белка 1 (AP-1) является фактором транскрипции, являющимся гетеродимером, состоящим из белков, принадлежащих к семейству белков Fos, семействам Jun, ATF и JDP. Этот фактор транскрипции регулирует экспрессию генов в ответ на множество стимулов, включая цитокины, факторы роста, стресс и бактериальные и вирусные инфекции.
Ранее обнаружено, что органоиды печени человека имеют сниженную экспрессию компонентов комплекса AP-1 по сравнению с первичными гепатоцитами. Учитывая это наблюдение, предполагают, что стимуляция образования комплекса AP-1 будет способствовать дифференцировке эпителиальных стволовых клеток в органоидах печени в направлении гепатоцитов. Авторы настоящего изобретения обнаружили, что включение стимулятора AP-1 карбахола (2[(аминокарбонил)окси]-N,N,N-триметилэтанаминия хлорида) в дифференцировочную среду повышало экспрессию маркеров гепатоцитов (включая Cyp3A4 и альбумин).
Ранее тестируемый стимулятор AP-1 является агонистом мускариновых ацетилхолиновых рецепторов. Существует пять подтипов мускариновых ацетилхолиновых рецепторов (M1-M5). Ими являются сопряженные с G-белком рецепторы, обнаруживаемые в клеточных мембранах различных типов клеток (например, нейронов).
Полагают, что стимулятор AP-1 или агонист мускариновых ацетилхолиновых рецепторов также может усиливать дифференцировку эпителиальных стволовых клеток из иных тканей, чем печень (например, кишечника, желудка, поджелудочной железы или легких).
Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит стимулятор AP-1. В некоторых вариантах осуществления стимулятор AP-1 является агонистом мускариновых ацетилхолиновых рецепторов. Можно использовать любой подходящий агонист мускариновых ацетилхолиновых рецепторов. В некоторых вариантах осуществления агонист мускариновых ацетилхолиновых рецепторов является агонистом мускариновых ацетилхолиновых рецепторов M3 (например, ацетилхолином, бетанехолом, карбахолом, оксотреморином или пилокарпином).
В некоторых вариантах осуществления агонист мускариновых ацетилхолиновых рецепторов выбран из одного или нескольких из следующего: ацетилхолина, бетанехола, карбахола, оксотреморина, L-689,660 (1-азабицикло[2.2.2]октана, 3-(6-хлорпиразинил)малеата), пилокарпина, мускарина, McN-A 343 (4-[[[(3-хлорфенил)амино]карбонил]окси]-N,N,N-триметил-2-бутин-1-аминия хлорида), 77-LH-218-1 (1-[3-(4-бутил-1-пиперидинил)пропил]-3,4-дигидро-2(1H)-хинолинонп) и метахолина.
В некоторых вариантах осуществления агонистом мускариновых ацетилхолиновых рецепторов является карбахол.
В некоторых вариантах осуществления стимулятор AP-1 (например, агонист мускариновых ацетилхолиновых рецепторов) присутствует в дифференцировочной среде по изобретению в концентрации 0,001-500 мкМ, 0,01-500 мкМ, 0,01-250 мкМ, 0,01-150 мкМ, 0,1-500 мкМ, 0,1-100 мкМ, 0,5-500 мкМ, 0,5-100 мкМ, 0,5-50 мкМ, 1-500 мкМ, 1-300 мкМ, 1-200 мкМ, 1-20 мкМ, 1-5 мкМ, 10-300 мкМ, 50-300 мкМ, 50-200 мкМ или 50-150 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит карбахол в концентрации 0,001-500 мкМ, 0,001-300 мкМ, 0,01-500 мкМ, 0,01-300 мкМ, 0,1-500 мкМ, 0,1-300 мкМ, 1-500 мкМ, 10-500 мкМ, 100-500 мкМ, 1-300 мкМ, 10-300 мкМ, 50-300 мкМ, 50-200 мкМ или 50-150 мкМ. Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит карбахол в концентрации 100 мкМ.
Агонисты мускариновых ацетилхолиновых рецепторов
Как описано выше, ранее обнаружено, что включение агониста мускариновых ацетилхолиновых рецепторов карбахола в дифференцировочной среде способствует дифференцировке. Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит агонист мускариновых ацетилхолиновых рецепторов.
Можно использовать любой подходящий агонист мускариновых ацетилхолиновых рецепторов. В некоторых вариантах осуществления агонист мускариновых ацетилхолиновых рецепторов является агонистом мускариновых ацетилхолиновых рецепторов M3 (например, ацетилхолином, бетанехолом, карбахолом, оксотреморином или пилокарпином).
В некоторых вариантах осуществления агонист мускариновых ацетилхолиновых рецепторов выбран из одного или нескольких из следующего: ацетилхолина, бетанехола, карбахола, оксотреморина, L-689,660 (1-азабицикло[2.2.2]октана, 3-(6-хлорпиразинил)малеата), пилокарпина, мускарина, McN-A 343 (4-[[[(3-хлорфенил)амино]карбонил]окси]-N,N,N-триметил-2-бутин-1-аминия хлорида), 77-LH-218-1 (1-[3-(4-бутил-1-пиперидинил)пропил]-3,4-дигидро-2(1H)-хинолинона) и метахолина.
В некоторых вариантах осуществления агонистом мускариновых ацетилхолиновых рецепторов является карбахол.
В некоторых вариантах осуществления стимулятор AP-1 (например, агонист мускариновых ацетилхолиновых рецепторов) присутствует в дифференцировочной среде по изобретению в концентрации 0,001-500 мкМ, 0,01-500 мкМ, 0,01-250 мкМ, 0,01-150 мкМ, 0,1-500 мкМ, 0,1-100 мкМ, 0,5-500 мкМ, 0,5-100 мкМ, 0,5-50 мкМ, 1-500 мкМ, 1-300 мкМ, 1-200 мкМ, 1-20 мкМ,1-5 мкМ, 10-300 мкМ, 50-300 мкМ, 50-200 мкМ или 50-150 мкМ.
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит карбахол в концентрации 0,001-500 мкМ, 0,001-300 мкМ, 0,01-500 мкМ, 0,01-300 мкМ, 0,1-500 мкМ, 0,1-300 мкМ, 1-500 мкМ, 10-500 мкМ, 100-500 мкМ, 1-300 мкМ, 10-300 мкМ, 50-300 мкМ, 50-200 мкМ или 50-150 мкМ. Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит карбахол в концентрации 100 мкМ.
Агонист карбахола
Как описано выше, ранее обнаружено, что включение агониста мускариновых ацетилхолиновых рецепторов карбахола в дифференцировочной среде способствует дифференцировке. Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит карбахол или агонист карбахола. Термин "агонист карбахола" определяют в настоящем описании как средство, обладающее общими одним или несколькими видами биологической активности с карбахолом.
Активаторы пути цАМФ
В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит активатор пути цАМФ. Активатор пути цАМФ может являться любым подходящим активатором, повышающим уровни цАМФ в клетке. Путь цАМФ включает активацию многих типов сопряженных с G-белком рецепторов гормонов и нейромедиаторов. Связывание гормона или нейромедиатора с его мембраносвязанным рецептором индуцирует конформационное изменение в рецепторе, приводящее к активации α-субъединицы G-белка. Активированная субъединица G-белка стимулирует, а неактивированная субъединица G-белка ингибирует аденилатциклазу. Стимуляция аденилатциклазы приводит к катализу преобразования цитоплазматического АТФ в цАМФ, таким образом, повышая уровни цАМФ в клетке. Таким образом, активатор пути цАМФ, например, может являться активатором аденилатциклазы. Примеры подходящих активаторов аденилатциклазы включают форсколин, аналог форсколина и холерный токсин. В некоторых вариантах осуществления активатором пути цАМФ является форсколин. В некоторых вариантах осуществления активатор пути цАМФ не является холерным токсином. В некоторых вариантах осуществления активатор пути цАМФ может являться аналогом цАМФ, например, 8-бром-цАМФ. 8-бром-цАМФ является проникающим в клетку аналогом, имеющим более высокую устойчивость к гидролизу фосфодиэстеразами, чем цАМФ. В некоторых вариантах осуществления активатором пути цАМФ является NKH477 (например, Tocris кат. №1603).
Активаторы пути цАМФ можно идентифицировать известными в этой области способами, например, с использованием конкурентного иммунологического анализа, посредством которого измеряют уровни цАМФ. Набор для флуоресцентного анализа цикло-АМФ CatchPoint® (Molecular Devices LLC) является примером коммерчески доступного набора для осуществления такого иммунологического анализа. цАМФ в образце или стандарте конкурирует с меченым пероксидазой хрена (HRP) конъюгатом цАМФ за участки связывания на антителах против цАМФ. В отсутствие цАМФ, большая часть конъюгата HRP-цАМФ связана с антителом. Увеличивающиеся концентрации цАМФ конкурентно снижают количество связанного конъюгата, таким образом, снижая измеряемую активность HRP. Активатор пути цАМФ будет приводить к повышению уровней цАМФ и снижению измеряемой активности HRP по сравнению контролем.
В некоторых вариантах осуществления активатор пути цАМФ используют в концентрации от приблизительно 10 нМ до приблизительно 500 мкМ, от приблизительно 10 нМ до приблизительно 100 мкМ, от приблизительно 1 мкМ до приблизительно 50 мкМ, от приблизительно 1 мкМ до приблизительно 25 мкМ, от приблизительно 5 мкМ до приблизительно 1000 мкМ, от приблизительно 5 мкМ до приблизительно 500 мкМ, от приблизительно 5 мкМ до приблизительно 100 мкМ, от приблизительно 5 мкМ до приблизительно 50 мкМ, от приблизительно 5 мкМ до приблизительно 25 мкМ, от приблизительно 10 мкМ до приблизительно 1000 мкМ, от приблизительно 10 мкМ до приблизительно 500 мкМ, от приблизительно 10 мкМ до приблизительно 100 мкМ, от приблизительно 10 мкМ до приблизительно 50 мкМ, от приблизительно 10 мкМ до приблизительно 25 мкМ или от приблизительно 20 мкМ. В некоторых вариантах осуществления активатор пути цАМФ используют в концентрации по меньшей мере 10 нМ, 20 нМ, 50 нМ, 100 нМ, 200 нМ, 500 нМ, 1 мкМ, по меньшей мере 2 мкМ, по меньшей мере 5 мкМ, по меньшей мере 10 мкМ, по меньшей мере 20 мкМ, по меньшей мере 30 мкМ, по меньшей мере 50 мкМ или по меньшей мере 100 мкМ.
Выбранная концентрация может зависеть от используемого активатора пути цАМФ, и ее может определять специалист в этой области в зависимости от активности активатора пути цАМФ. Например, NKH477, как правило, является более активным, чем 8-BR-цАМФ и форсколин. Более активный активатор пути цАМФ можно использовать в меньших концентрациях с тем же эффектом.
Например, в некоторых вариантах осуществления NKH477 можно использовать в концентрации от приблизительно 100 нМ до приблизительно 10 мкМ или в концентрации приблизительно 100 нМ, приблизительно 1 мкМ или приблизительно 10 мкМ. В некоторых вариантах осуществления 8-BR-цАМФ или форсколин можно использовать в концентрации от приблизительно 1 мкМ до приблизительно 100 мкМ, или в концентрации приблизительно 1 мкМ, приблизительно 10 мкМ или приблизительно 100 мкМ.
В некоторых вариантах осуществления холерный токсин можно использовать в концентрации от приблизительно 1 нг/мл до приблизительно 500 нг/мл, от приблизительно 10 нг/мл до приблизительно 100 нг/мл, от приблизительно 50 нг/мл до приблизительно 100 нг/мл или приблизительно 10 нг/мл, приблизительно 20 нг/мл, приблизительно 30 нг/мл, приблизительно 40 нг/мл, приблизительно 50 нг/мл, приблизительно 60 нг/мл, приблизительно 70 нг/мл, приблизительно 80 нг/мл, приблизительно 90 нг/мл, приблизительно 100 нг/мл, приблизительно 200 нг/мл, приблизительно 300 нг/мл, приблизительно 400 нг/мл или приблизительно 500 нг/мл.
Добавки
Дифференцировочную среду по изобретению, предпочтительно, дополняют одним или несколькими (например, 1, 2, 3 или всеми) соединениями, выбранными из группы, состоящей из B27, N-ацетилцистеина и N2. Таким образом, в некоторых вариантах осуществления дифференцировочная среда дополнительно содержит один или несколько компонентов, выбранных из группы, состоящей из: B27, N2 и N-ацетилцистеина. Например, в некоторых вариантах осуществления дифференцировочная среда дополнительно содержит B27, N-ацетилцистеин и N2.
Считают, что B27 (Invitrogen), N-ацетилцистеин (Sigma) и N2 (Invitrogen) и никотинамид (Sigma) контролируют пролиферацию клеток и способствуют стабильности ДНК.
В некоторых вариантах осуществления N-ацетилцистеин присутствует в дифференцировочной среде в концентрации 0,1-200 мМ, 0,1-100 мМ, 0,1-50 мМ, 0,1-10 мМ, 0,1-5 мМ, 0,5-200 мМ, 0,5-100 мМ, 0,5-50 мМ, 0,5-10 мМ, 0,5-5 мМ, 1-100 мМ, 1-50 мМ, 1-10 мМ, 1-5 мМ. В некоторых вариантах осуществления N-ацетилцистеин присутствует в дифференцировочной среде в концентрации приблизительно 1,25 мМ.
В некоторых вариантах осуществления добавка B27 является "добавкой B27 без витамина A" (также обозначаемой в настоящем описании как "B27 без витамина A"; доступной в Invitrogen, Carlsbad, CA; www.invitrogen.com; кат. №12587010; и PAA Laboratories GmbH, Pasching, Austria; www.paa.com; кат. № F01-002; Brewer et al., (1993), J Neurosci Res. 35(5):567-76). В некоторых вариантах осуществления добавку B27 можно заменять составом-дженериком, содержащим один или несколько из компонентов, выбранных из следующего списка: биотин, холестерин, линолевая кислота, линоленовая кислота, прогестерон, путресцин, ретинил ацетат, селенит натрия, трийодтиронин (T3), DL-альфа-токоферол (витамин E), альбумин, инсулин и трансферрин.
Добавку B27 от PAA Laboratories GmbH поставляют в виде жидкого 50-кратного концентрата, содержащего, среди прочего, ингредиенты биотин, холестерин, линолевую кислоту, линоленовую кислоту, прогестерон, путресцин, ретинол, ретинил ацетат, селенит натрия, трийодтиронин (T3), DL-альфа-токоферол (витамин E), альбумин, инсулин и трансферрин. Из этих ингредиентов, по меньшей мере, линоленовая кислота, ретинол, ретинил ацетат и трийодтиронин (T3) являются агонистами ядерных рецепторов гормонов. Добавку B27 можно добавлять в дифференцировочную среду в виде концентрата или разводить перед добавлением в дифференцировочную среде. Ее можно использовать в 1-кратной конечной концентрации или в других конечных концентрациях (например, от 0,1 до 4-кратной концентрации, от 0,1 до 2-кратной концентрации, от 0,5 до 2-кратной концентрации, от 1 до 4-кратной концентрации или от 1 до 2-кратной концентрации). Использование добавки B27 является удобным способом включения биотина, холестерина, линолевой кислоты, линоленовой кислоты, прогестерона, путресцина, ретинола, ретинил ацетата, селенита натрия, трийодтиронина (T3), DL-альфа-токоферола (витамин E), альбумина, инсулина и трансферрина в дифференцировочную среду по изобретению. Также предполагают, что некоторые или все компоненты можно добавлять в дифференцировочную среду раздельно вместо использования добавки B27. Таким образом, дифференцировочная среда может содержать некоторые или все из этих компонентов.
В некоторых вариантах осуществления ретиноевая кислота отсутствует в добавке B27, используемой в дифференцировочной среде, и/или отсутствует в дифференцировочной среде.
"Добавка N2" (также обозначаемая в настоящем описании как "N2") доступна в Invitrogen, Carlsbad, CA; www.invitrogen.com; кат. №17502-048; и PAA Laboratories GmbH, Pasching, Austria; www.paa.com; кат. № F005-004; Bottenstein & Sato, PNAS, 76(1):514-517, 1979. Добавку N2 от PAA Laboratories GmbH поставляют в виде жидкого 100-кратного концентрата, содержащего 500 мкг/мл трансферрина человека, 500 мкг/мл инсулина быка, 0,63 мкг/мл прогестерона, 1611 мкг/мл путресцина и 0,52 мкг/мл селенита натрия. Добавку N2 можно добавлять в дифференцировочную среду в виде концентрата или разводить перед добавлением в дифференцировочную среду. Ее можно использовать в 1-кратной конечной концентрации или в других конечных концентрациях (например, от 0,1- до 4-кратной концентрации, от 0,1- до 2-кратной концентрации, от 0,5- до 2-кратной концентрация, от 1- до 4-кратной концентрации или от 1- до 2-кратной концентрации). Использование добавки N2 является удобным способом включения трансферрина, инсулина, прогестерона, путресцина и селенита натрия в дифференцировочную среду по изобретению. Также предполагают, что некоторые или все из этих компонентов можно добавлять в дифференцировочную среду раздельно вместо использования добавки N2. Таким образом, дифференцировочная среда может содержать некоторые или все из этих компонентов.
В некоторых вариантах осуществления, в которых среда содержит B27, она также не содержит N2. Таким образом, варианты осуществления настоящего изобретения, при желании, можно адаптировать для исключения N2, если присутствует B27.
В некоторых вариантах осуществления N2 не присутствует в дифференцировочной среде.
В некоторых вариантах осуществления, в которых среда содержит N2, она также не содержит B27. Таким образом, варианты осуществления настоящего изобретения, при желании, можно адаптировать для исключения B27, если присутствует N2.
В некоторых вариантах осуществления B27 не присутствует в дифференцировочной среде.
В некоторых вариантах осуществления дифференцировочную среду дополняют B27 и/или N2.
В некоторых вариантах осуществления минимальную среду дополняют 150-250 нг/мл N-ацетилцистеина; предпочтительно, минимальную среду дополняют приблизительно 200 нг/мл N-ацетилцистеина.
Можно использовать любой подходящий pH. Например, pH среды может находиться в диапазоне приблизительно от 7,0 до 7,8, в диапазоне приблизительно от 7,2 до 7,6, или приблизительно 7,4. pH можно поддерживать с использованием буфера. Специалист в этой области легко может выбирать подходящий буфер. Буферы, которые можно использовать, включают карбонатные буферы (например, NaHCO3), и фосфаты (например, NaH2PO4). Эти буферы, как правило, используют в количестве от приблизительно 50 до приблизительно 500 мг/л. Также можно использовать другие буферы, такие как N-[2-гидроксиэтил]-пиперазин-N'-[2-этансульфоновая кислота] (HEPES) и 3-[N-морфолино]-пропансульфоновая кислота (MOPS), как правило, в количестве от приблизительно 1000 до приблизительно 10000 мг/л. В некоторых вариантах осуществления буфер выбран из одного или нескольких из следующего списка: фосфатный буфер (например, KH2PO4, K2HPO4, Na2HPO4, NaCl, NaH2PO4) ацетатный буфер (например, HOAc или NaOAc), цитратный буфер (например, лимонная кислота или Na-цитрат), или буфер Трис (например, Трис, Трис-HCl) или органический буфер. В некоторых вариантах осуществления органический буфер является цвиттерионным буфером, таким как буфер Гуда, например, выбранный из HEPES, MOPS, MES, ADA, PIPES, ACES, MOPSO, хлорида холамина, BES, TES, DIPSO, ацетамидоглицина, TAPSO, POPSO, HEPPSO, HEPPS, трицина, глицинамида, бицина, TAPS, AMPSO, CABS, CHES, CAPS и CAPSO. Предпочтительным буфером является HEPES, например, в концентрации 0,1-100 мМ, 0,1-50 мМ, 0,5-50 мМ, 1-50 мМ, 1-20 мМ или 5-15 мМ. В некоторых вариантах осуществления HEPES добавляют в среду для культивирования в количестве приблизительно 10 мМ. Дифференцировочная среда также может содержать индикатор pH, такой как феноловый красный, для легкого мониторинга pH среды (например, в количестве от приблизительно 5 до приблизительно 50 мг/л).
Дифференцировочная среда для использования в изобретении может содержать одну или несколько аминокислот. Специалисту в этой области понятны соответствующие типы и количества аминокислот для использования в дифференцировочных средах. Аминокислоты, которые могут присутствовать, включают L-аланин, L-аргинин, L-аспарагин, L-аспарагиновая кислота, L-цистеин, L-цистин, L-глутаминовую кислоту, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин, L-валин и их комбинации. Некоторые дифференцировочные среды будут содержать все из этих аминокислот. Как правило, каждая аминокислота, при ее наличии, присутствует в количестве от приблизительно 0,001 до приблизительно 1 г/л среды (как правило, в количестве от приблизительно 0,01 до приблизительно 0,15 г/л), за исключением L-глутамина, присутствующего в количестве от приблизительно 0,05 до приблизительно 1 г/л (как правило, от приблизительно 0,1 до приблизительно 0,75 г/л). Аминокислоты могут иметь синтетическое происхождение.
Дифференцировочная среда для использования в изобретении может содержать один или несколько витаминов. Специалисту в этой области понятны соответствующие типы и количества витаминов для использования в дифференцировочных средах. Витамины, которые могут присутствовать, включают тиамин (витамин B1), рибофлавин (витамин B2), ниацин (витамин B3), D-пантотенат кальция (витамин B5), пиридоксаль/пиридоксамин/пиридоксин (витамин B6), фолиевую кислоту (витамин B9), цианокобаламин (витамин B12), аскорбиновую кислоту (витамин C), кальциферол (витамин D2), DL-альфа-токоферол (витамин E), биотин (витамин H) и менадион (витамин K).
Дифференцировочная среда для использования в изобретении может содержать одну или несколько неорганических солей. Специалисту в этой области понятны соответствующие типы и количества неорганических солей для использования в дифференцировочных средах. Неорганические соли, как правило, включают в дифференцировочные среды для поддержания осмотического баланса клеток и для регуляции мембранного потенциала. Неорганические соли, которые могут присутствовать, включают соли кальция, меди, железа, магния, калия, натрия, цинка. Соли, как правило, используют в форме хлоридов, фосфатов, сульфатов, нитратов и бикарбонатов. Конкретные соли, которые можно использовать, включают CaCl2, CuSO4-5H2O, Fe(NO3)-9H2O, FeSO4-7H2O, MgCl, MgSO4, KCl, NaHCO3, NaCl, Na2HPO4, Na2HPO4-H2O и ZnSO4-7H2O.
Осмолярность среды может находиться в диапазоне от приблизительно 200 до приблизительно 400 мОсм/кг, в диапазоне от приблизительно 290 до приблизительно 350 мОсм/кг или в диапазоне от приблизительно 280 до приблизительно 310 мОсм/кг. Осмолярность среды может составлять менее приблизительно 300 мОсм/кг (например, приблизительно 280 мОсм/кг).
Дифференцировочная среда для использования в изобретении может содержать углеродный источник энергии в форме одного или нескольких сахаров. Специалисту в этой области понятны соответствующие типы и количества сахаров для использования в дифференцировочных средах. Сахара, которые могут присутствовать, включают глюкозу, галактозу, мальтозу и фруктозу. Сахар, предпочтительно, является глюкозой, в частности, D-глюкозой (декстрозой). Углеродный источник энергии, как правило, будет присутствовать в количестве от приблизительно 1 и приблизительно 10 г/л.
Дифференцировочная среда по изобретению может содержать сыворотку. Можно использовать сыворотку, полученную из любого подходящего источника, включая эмбриональную телячью сыворотку (FBS), сыворотку козы или сыворотку человека. Предпочтительно, используют сыворотку человека. Сыворотку можно использовать в количестве от приблизительно 1% до приблизительно 30% по объему среды общепринятыми способами.
В других вариантах осуществления дифференцировочная среда по изобретению может содержать заменитель сыворотки. Различные составы заменителей сыворотки коммерчески доступны и известны специалисту в этой области. Если используют заменитель сыворотки, его можно использовать в количестве от приблизительно 1% до приблизительно 30% по объему среды общепринятыми способами.
В других вариантах осуществления дифференцировочная среда по изобретению может не содержать сыворотку и/или заменитель сыворотки. Бессывороточная среда является средой, несодержащей сыворотку животного любого типа. Бессывороточные среды могут быть предпочтительными во избежание возможной ксено-контаминации стволовых клеток. Среда без заменителя сыворотки является средой, недополненной каким-либо коммерческим составом заменителя сыворотки.
В предпочтительном варианте осуществления дифференцировочную среду дополняют очищенным, природным, полусинтетическим и/или синтетическим фактором роста, и она не содержит неопределенный компонент, такой как эмбриональная телячья сыворотка. Например, добавки, такие как B27 (Invitrogen), N-ацетилцистеин (Sigma) и N2 (Invitrogen), стимулируют пролиферацию некоторых клеток. В некоторых вариантах осуществления дифференцировочную среду дополняют одной или несколькими из этих добавок, например, одной, любыми двумя или всеми тремя из этих добавок.
Дифференцировочная среда для использования в изобретении может содержать один или несколько микроэлементов, таких как ионы бария, брома, кобальта, йода, марганца, хрома, меди, никеля, селена, ванадия, титана, германия, молибдена, кремния, железа, фтора, серебра, рубидия, олова, циркония, кадмия, цинка и/или алюминия.
Среда может содержать восстановитель, такой как бета-меркаптоэтанол, в концентрации приблизительно 0,1 мМ.
Дифференцировочная среда по изобретению может содержать одно или несколько дополнительных средств, таких как питательные вещества или факторы роста, как описано, улучшающие культивирование стволовых клеток, такие как холестерин/трансферрин/альбумин/инсулин/прогестерон, путресцин, селенит/другие факторы.
Композиции и сосуды
Изобретение также относится к композиции или сосуду для культивирования клеток, содержащим клетки и/или органоиды по любому из аспектов изобретения, описанных выше, и дифференцировочную среду по любому из аспектов изобретения, описанных выше. Например, такая композиция или сосуд для культивирования клеток может содержать любое количество клеток или органоидов, культивируемых способом по изобретению, в дифференцировочной среде, как описано выше.
В дополнительном аспекте изобретение относится к герметично запаянному сосуду, содержащему дифференцировочную среду по изобретению. Герметично запаянные сосуды могут быть предпочтительными для транспортировки или хранения дифференцировочных сред для предотвращения контаминации. Сосуд может являться любым подходящим сосудом, таким как колба, планшет, бутыль, банка, сосуд или мешок.
Пример дифференцировочных сред по изобретению
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch и EGFR ингибитор. В некоторых вариантах осуществления дифференцировочная среда дополнительно содержит минимальную среду.
Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch и EGFR ингибитор (например, гефитиниб). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ) и гефитиниб (например, в концентрации приблизительно 5 мкМ).
Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch и ингибитор EGFR и ErbB2 (например, афатиниб). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ) и афатиниб (например, в концентрации приблизительно 10 мкМ).
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch и ингибитор пути RAS-RAF-MAPK (например, ингибитор MEK).
Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ) и ингибитор MEK, такой как PD0325901 (например, в концентрации приблизительно 1,5 мкМ).
Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ) и ингибитор ERK, такой как SCH772984 (например, в концентрации приблизительно 10 мкМ).
В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит активатор пути BMP (например, BMP4). Как указано выше, авторы настоящего изобретения неожиданно обнаружили, что включение активатора пути BMP может стимулировать дифференцировку секретин-продуцирующих клеток и ингибировать дифференцировку энтерохромаффинных клеток. В других вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит ингибитор BMP (например, Noggin). Как указано выше, авторы настоящего изобретения неожиданно обнаруживали, что включение ингибитора BMP может стимулировать дифференцировку GLP-1-продуцирующих клеток и ингибировать дифференцировку секретин-продуцирующих клеток.
Таким образом, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt (например, IWP2), ингибитор Notch (например, DAPT), ингибитор EGFR (например, гефитиниб) и активатор пути BMP (например, BMP4). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), гефитиниб (например, в концентрации приблизительно 5 мкМ) и BMP4 (например, в концентрации приблизительно 10 мкг/мл). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch, ингибитор EGFR и ErbB2 (например, афатиниб) и активатор пути BMP (например, BMP4). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), афатиниб (например, в концентрации приблизительно 10 мкМ) и BMP4 (например, в концентрации приблизительно 10 мкг/мл).
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch, ингибитор пути RAS-RAF-MAPK (например, ингибитор MEK) и активатор пути BMP, такой как BMP4 (например, в концентрации приблизительно 10 мкг/мл). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), ингибитор MEK, такой как PD0325901 (например, в концентрации приблизительно 1,5 мкМ), и активатор пути BMP, такой как BMP4 (например, в концентрации приблизительно 10 мкг/мл). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), ингибитор ERK, такой как SCH772984 (например, в концентрации приблизительно 10 мкМ) и активатор пути BMP, такой как BMP4 (например, в концентрации приблизительно 10 мкг/мл).
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt (например, IWP2), ингибитор Notch (например, DAPT), ингибитор EGFR (например, гефитиниб) и ингибитор BMP (например, Noggin). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), гефитиниб (например, в концентрации приблизительно 5 мкМ) и Noggin (например, в концентрации приблизительно 100 нг/мл). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch, ингибитор EGFR и ErbB2 (например, афатиниб) и ингибитор BMP (например, Noggin). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), афатиниб (например, в концентрации приблизительно 10 мкМ) и Noggin (например, в концентрации приблизительно 100 нг/мл).
В некоторых вариантах осуществления дифференцировочная среда по изобретению содержит: ингибитор Wnt, ингибитор Notch, ингибитор пути RAS-RAF-MAPK (например, ингибитор MEK) и ингибитор BMP, такой как Noggin (например, в концентрации приблизительно 100 нг/мл). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), ингибитор MEK, такой как PD0325901 (например, в концентрации приблизительно 1,5 мкМ), и ингибитор BMP, такой как Noggin (например, в концентрации приблизительно 100 нг/мл). Например, в некоторых вариантах осуществления дифференцировочная среда по изобретению содержит IWP-2 (например, в концентрации приблизительно 1,5 мкМ), DAPT (например, в концентрации приблизительно 1 мМ), ингибитор ERK, такой как SCH772984 (например, в концентрации приблизительно 10 мкМ), и ингибитор BMP, такой как Noggin (например, в концентрации приблизительно 100 нг/мл).
В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит активатор mTOR, такой как MHY1485 (например, в концентрации приблизительно 5 мкМ).
В некоторых вариантах осуществления дифференцировочная среда по изобретению дополнительно содержит ингибитор Hedgehog, такой как ингибитор SMO (например, в концентрации приблизительно 5 мкМ). В некоторых вариантах осуществления ингибитором SMO является висмогениб.
В предпочтительных вариантах осуществления дифференцировочная среда дополнительно содержит R-спондин (например, в концентрации приблизительно 1 мкг/мл).
В предпочтительных вариантах осуществления концентрация EGF в среде составляет менее 1 мМ.
Внеклеточный матрикс
В некоторых вариантах осуществления способы дифференцировки клетки включают культивирование клеток, приведенных в контакт с внеклеточным матриксом (ECM). Можно использовать любой подходящий ECM. Клетки, предпочтительно, культивируют в микроокружении, имитирующем, по меньшей мере, частично клеточную нишу, в которой указанные клетки находятся в природе. Клеточная ниша частично определяется клетками и ECM, секретируемым клетками в указанной нише. Клеточную нишу можно имитировать посредством культивирования указанных клеток в присутствие биоматериалов или синтетических материалов, обеспечивающих взаимодействие с белками клеточной мембраны, такими как интегрины. Таким образом, ECM, как представлено в настоящем описании, является любым биоматериалом или синтетическим материалом или их комбинацией, имитирующими клеточную нишу in vivo, например, посредством взаимодействия с белками клеточной мембраны, такими как интегрины.
В предпочтительном способе по изобретению клетки культивируют в контакте с ECM. Термин "в контакте" означает физический, или механический, или химический контакт, что означает, что для отделения указанного полученного органоида или популяции эпителиальных клеток от указанного внеклеточного матрикса необходимо приложить силу. В некоторых вариантах осуществления ECM является трехмерным матриксом. В некотором варианте осуществления клетки заключают в ECM. В некоторых вариантах осуществления клетки прикреплены к ECM. Среда для культивирования по изобретению может диффундировать в трехмерный ECM.
В других вариантах осуществления ECM находится в суспензии, т.е. клетки приведены в контакт с ECM в суспензионной системе. В некоторых вариантах осуществления ECM находится в суспензии в концентрации по меньшей мере 1%, по меньшей мере 2% или по меньшей мере 3%. В некоторых вариантах осуществления ECM находится в суспензии в концентрации от 1% до приблизительно 10% или от 1% до приблизительно 5%. Суспензионный способ может обладать преимуществами в качестве способов большего масштаба.
Один из типов ECM секретируется эпителиальными клетками, эндотелиальными клетками, париетальными эндодермально-подобными клетками (например, париетальные эндодермально-подобные клетки Энгельберта-Холма-Сворма, описанные в Hayashi et al., (2004), Matrix Biology 23:47-62) и клетки соединительной ткани. Этот ECM содержит различные полисахариды, воду, эластин и гликопротеины, где гликопротеины содержат коллаген, энтактин (нидоген), фибронектин и ламинин. Таким образом, в некоторых вариантах осуществления ECM для использования в способы по изобретению содержит один или несколько из компонентов, выбранных из следующего списка: полисахариды, эластин и гликопротеины, например, где гликопротеины содержат коллаген, энтактин (нидоген), фибронектин и/или ламинин. Например, в некоторых вариантах осуществления коллаген используют в качестве ECM. Известны различные типы ECM, содержащие различные композиции, включая различные типы гликопротеинов и/или различные комбинации гликопротеинов.
ECM можно получать посредством культивирования ECM-продуцирующих клеток, таких как, например, эпителиальные клетки, эндотелиальные клетки, париетальные эндодермально-подобные клетки или фибробласты, в контейнере перед удалением этих клеток и добавления выделенных фрагментов ткани или выделенных эпителиальных клеток. Примерами продуцирующих внеклеточный матрикс клеток являются хондроциты, продуцирующие, в основном, коллаген и протеогликаны, фибробласты, продуцирующие, в основном, коллаген типа IV, ламинин, интерстициальные проколлагены и фибронектин, и миофибробласты толстого кишечника, продуцирующие, в основном, коллагены (типа I, III, и V), хондроитинсульфатные протеогликаны, гиалуроновую кислоту, фибронектин и тенасцин-C. Они являются "природно продуцируемыми ECM". Природно продуцируемые ECM можно получать из коммерческих источников. Примеры коммерчески доступных внеклеточных матриксов включают: белки внеклеточного матрикса (Invitrogen) и препараты базальной мембраны из клеток Энгельберта-Холма-Сворма (EHS) саркомы мыши (например, экстракт базальной мембраны Cultrex® (Trevigen, Inc.) или Matrigel™ (BD Biosciences)).
Таким образом, в некоторых вариантах осуществления ECM является природно продуцируемым ECM. В некоторых вариантах осуществления ECM является ламинин-содержащим ECM, таким как Matrigel™ (BD Biosciences). В некоторых вариантах осуществления ECM является Matrigel™ (BD Biosciences), содержащим ламинин, энтактин и коллаген IV. В некоторых вариантах осуществления ECM содержит ламинин, энтактин, коллаген IV и гепарансульфат (например, экстракт базальной мембраны Cultrex® типа 2 (Trevigen, Inc.)). В некоторых вариантах осуществления ECM содержит по меньшей мере один гликопротеин, такой как коллаген и/или ламинин. Предпочтительный ECM для использования в способе по изобретению содержит коллаген и ламинин. Дополнительный предпочтительный ECM содержит ламинин, энтактин и коллаген IV. При желании, можно использовать смеси природно продуцируемых или синтетических материалов ECM.
В другом варианте осуществления ECM может являться синтетическим ECM. Например, можно использовать синтетический ECM, такой как ProNectin (Sigma Z378666). В дополнительном примере ECM может являться пластиком, например полиэфиром, или гидрогелем. В некоторых вариантах осуществления синтетический матрикс можно покрывать биоматериалом, например, одним или несколькими гликопротеинами, такими как коллаген или ламинин.
Трехмерный ECM поддерживает культивирование трехмерных эпителиальных органоидов. Материал внеклеточного матрикса, как правило, будет представлять собой каплю на дне чашки, в которой суспендируют клетки. Как правило, когда матрикс отверждается при 37°C, добавляют среду, и она диффундирует в ECM. Клетки в среде прикрепляются к ECM посредством взаимодействия с его поверхностными структурами, например, взаимодействия с интегринами.
Среду для культивирования и/или клетки можно помещать, заключать или смешивать с ECM.
Например, в некоторых вариантах осуществления одну клетку, популяцию клеток или фрагмент ткани заключают в Matrigel™, необязательно, с низким содержанием факторов роста и/или без фенолового красного.
В некоторых вариантах осуществления среду для культивирования помещают сверху ECM. Затем среду для культивирования при необходимости, можно удалять и пополнять. В некоторых вариантах осуществления среду для культивирования пополняют каждые 1, 2, 3, 4, 5, 6 или 7 дней. Если компоненты "добавляют" или "удаляют" из сред, это в некоторых вариантах осуществления может означать, что сами среды удаляют из ECM, а затем новые среды, содержащие "добавляемый" компонент или исключающие "удаляемый" компонент, помещают на ECM.
Клетки-предшественники и стволовые клетки для культивирования и получения указанных клеток
Способы дифференцировки применимы для клеток-предшественников. Клетки-предшественники в настоящем описании определяют как любые клетки, имеющие дифференцировочный потенциал. Таким образом, термин "клетки-предшественники" включает стволовые клетки, включая, в качестве неограничивающих примеров, стволовые клетки взрослых, эмбриональные стволовые клетки и клетки iPS. Клетки-предшественники могут являться, например, первичными стволовыми клетками или выращенными стволовыми клетками или частично дифференцированными стволовыми клетками. В некоторых вариантах осуществления клетки-предшественники являются первичными клетками, что означает, что их получают непосредственно из живой ткани. В других вариантах осуществления клетки-предшественники являются вторичными клетками, т.е. культивируемыми и/или пассируемыми клетками. В некоторых вариантах осуществления клетки-предшественники являются выращиваемыми клетками. Термин "выращенный" означает, что клетки культивируют in vitro в среде для культивирования, способствующей экспансии (например, пролиферации) клеток вместо дифференцировки.
В предпочтительном варианте осуществления клетки-предшественники являются клетками-предшественниками взрослого, т.е. их получают из ткани взрослого. В некоторых вариантах осуществления клетки-предшественники взрослого являются стволовыми клетками взрослого (например, эпителиальными стволовыми клетками взрослого). В этом контексте термин "взрослый" включает новорожденного младенца или ребенка, но исключает эмбрион или плод. В некоторых вариантах осуществления клетки-предшественники не получают из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток, например, эмбриональных стволовых клеток человека или линий эмбриональных стволовых клеток человека.
В предпочтительном варианте осуществления клетки-предшественники являются эпителиальными клетками-предшественниками. Например, в предпочтительном варианте осуществления клетки-предшественники получают из эпителиальной ткани, более предпочтительно - эпителиальной ткани взрослого. Эпителиальные ткани включают печень, поджелудочную железу, кишечник, желудок, предстательную железу, легкие, молочную железу, яичник, слюнную железу, волосяной фолликул, кожу, пищевод, ухо, мочевой пузырь или щитовидную железу. Таким образом, в некоторых вариантах осуществления клетки-предшественники получают из печени, поджелудочной железы, кишечника, желудка, предстательной железы, легких, молочной железы, яичника, слюнной железы, волосяного фолликула, кожи, пищевода, уха, мочевого пузыря или щитовидной железы. В некоторых вариантах осуществления клетки-предшественники получают из поджелудочной железы, желудка, легких или кишечника. В предпочтительном варианте осуществления клетки-предшественники получают из кишечника.
В предпочтительном варианте осуществления клетки-предшественники являются клетками млекопитающих. Например, в предпочтительном варианте осуществления клетки получают из ткани млекопитающего. Например, в некоторых вариантах осуществления клетки-предшественники являются клетками человека. В некоторых вариантах осуществления клетки-предшественники получают из лабораторного животного (например, мыши, кролика, крысы, морской свинки), животного-компаньона (например, собаки, кошки, лошади) или сельскохозяйственного животного (например, коровы, свиньи, овцы, козы).
Первичные клетки представляют собой лучшие экспериментальные модели для исследований in vivo. В предпочтительном варианте осуществления изобретения клетки-предшественники являются первичными клетками-предшественниками (например, первичными эпителиальными стволовыми клетками) (или их получают в культурах клеток из первичных клеток-предшественников). Культуры первичных клеток можно пассировать для получения культур вторичных клеток. За исключением злокачественных клеток, общепринятые культуры вторичных эпителиальных клеток имеют ограниченный срок эксплуатации. После конкретного количества удвоений популяции (например, 50-100 поколений) клетки подвергаются процессу старения и прекращают делиться. Клетки из вторичных культур можно иммортализовать так, чтобы они становились стабильными линиями клеток. Иммортализация может происходить спонтанно, или ее можно индуцировать с помощью вируса или химического вещества. Иммортализованные линии клеток также известны как трансформированные клетки. В предпочтительном варианте осуществления изобретения клетки получают из выращенных культур эпителиальных стволовых клеток, предпочтительно, выращенных органоидов, выращенных и/или пассированных без иммортализации или трансформации. В некоторых вариантах осуществления эти выращенные эпителиальные культуры или органоиды могут являться генетически гетерогенными (в отличие от общепринятых линий клеток). Таким образом, в некоторых вариантах осуществления клетки-предшественники не являются иммортализованными или трансформированными клетками, или их не получают из линии иммортализованных клеток или линии трансформированных клеток.
Клетки-предшественники можно получать любым подходящим способом, например, как описано в WO 2010/090513, WO 2012/014076, WO 2012/168930 или WO 2015/173425. В некоторых вариантах осуществления клетки выделяют посредством расщепления коллагеназой, например, как описано в Dorell et al., 2008, (Hepatology. 2008 Oct;48(4):1282-91. Surface markers for the murine oval cell response. Dorrell C, Erker L, Lanxon-Cookson KM, Abraham SL, Victoroff T, Ro S, Canaday PS, Streeter PR, Grompe M). В некоторых вариантах осуществления коллагеназа расщепление осуществляют на биоптате ткани. В некоторых вариантах осуществления расщепление коллагеназой и аккутазой используют для получения клеток-предшественников для использования в изобретении.
В некоторых вариантах осуществления клетка-предшественник является эпителиальной стволовой клеткой, экспрессирующей Lgr5. Органоид, предпочтительно, получают с использованием клетки из ткани взрослого, предпочтительно - эпителиальной стволовой клетки из ткани взрослого, более предпочтительно - эпителиальной стволовой клетки, экспрессирующей Lgr5.
В наиболее предпочтительном варианте осуществления клетки-предшественники являются эпителиальными стволовыми клетками взрослого, экспрессирующими Lgr5, или содержат их.
В некоторых вариантах осуществления клетки-предшественники являются нормальными клетками, что означает, что клетки имеют нормальный кариотип, генотип и/или фенотип. В альтернативных вариантах осуществления клетки-предшественники являются клетками заболевания, что означает, что они имеют кариотип, генотип и/или фенотип заболевания. Например, в некоторых вариантах осуществления клетки-предшественники являются злокачественными клетками. Таким образом, предполагают, что, например, эпителиальные стволовые клетки могут являться Lgr5-положительными злокачественными стволовыми клетками. Таким образом, клетки, при необходимости, можно получать из опухоли. В альтернативных вариантах осуществления клетки-предшественники являются пораженными клетками-предшественниками, например, клетками-предшественниками, инфицированными внутриклеточными патогенами (например, бактериями, вирусами или паразитами).
Примеры способов
Изобретение относится к способу дифференцировки клеток-предшественников, где указанный способ включает культивирование клеток в дифференцировочной среде, представленной в настоящем описании. В предпочтительном варианте осуществления клетки культивируют в контакте с ECM, как представлено в настоящем описании.
Изобретение также относится к способу культивирования клеток-предшественников, где указанный способ включает культивирование клеток в среде для выращивания и последующее культивирование клеток в дифференцировочной среде, представленной в настоящем описании. В некоторых вариантах осуществления клетки культивируют в контакте с ECM, как представлено в настоящем описании, на стадиях выращивания и/или дифференцировки. В некоторых вариантах осуществления среду для выращивания удаляют перед культивированием клеток в дифференцировочной среде, например, посредством повторных промывок или разделения культуры клеток.
Изобретение относится к способу культивирования одной эпителиальной стволовой клетки, популяции эпителиальных стволовых клеток или выделенного фрагмента ткани, предпочтительно, для получения органоида, включающему:
культивирование эпителиальной стволовой клетки, популяции эпителиальных стволовых клеток или выделенного фрагмента ткани в среде для выращивания для получения выращенной популяции клеток;
необязательно, индуцирование состояния покоя выращенной популяции клеток (например, посредством обработки одним или несколькими ингибиторами пути EGFR); и
культивирование выращенной популяции клеток в дифференцировочной среде.
Изобретение также относится к способу дифференцировки одной клетки-предшественника или популяции клеток-предшественников, включающему:
культивирование клетки-предшественника или популяции клеток-предшественников в дифференцировочной среде.
Дифференцировочная среда может являться любой дифференцировочной средой, представленной в настоящем описании. Например, в некоторых вариантах осуществления дифференцировочная среда содержит: ингибитор Wnt (например, IWP-2), ингибитор Notch (например, DAPT) и ингибитор пути EGFR (например, один или несколько из гефитиниба, афатиниба, PD0325901 и SCH772984). Эта дифференцировочная среда особенно пригодна для использования в способе получения популяции клеток, обогащенных EEC. Маркеры, характерные для EEC, включают Chga, Chgb, Tac1, Tph1, Gip, Fabp5, Ghrl, Pyy, Nts, Neurod1, Sst, Sct, холецистокинин, глюкагон и/или проглюкагон. Дополнительные типы клеток EEC и их характеристики описаны в Grun et al. (2015) Nature 525:251-255. См. также таблицу 2 ниже. Таким образом, в некоторых вариантах осуществления способ дифференцировки по изобретению приводит к получению популяции клеток, экспрессирующих один или несколько маркеров, выбранных из Chga, Chgb, Tac1, Tph1, Gip, Fabp5, Ghrl, Pyy, Nts, Neurod1, Sst, Sct, холецистокинина, глюкагона и/или проглюкагона. В некоторых вариантах осуществления EEC, полученные способами по изобретению, экспрессируют серотонин.
В некоторых вариантах осуществления дифференцировочная среда содержит: ингибитор Wnt (например, IWP-2), ингибитор Notch (например, DAPT) и ингибитор пути EGFR (например, один или несколько из гефитиниба, афатиниба, PD0325901 и SCH772984) и ингибитор BMP (например, Noggin или LDN193189). Эта дифференцировочная среда особенно пригодна для использования в способе получения популяции клеток, обогащенных GLP1-секретирующими EEC. В некоторых вариантах осуществления этот способ приводит к получению популяции клеток, положительных по одному или нескольким маркерам, выбранным из Tac1, GLP1 и Chg; и отрицательных по одному или нескольким маркерам, выбранным из секретина, pyy и nts. В некоторых вариантах осуществления популяция клеток экспрессирует серотонин.
В некоторых вариантах осуществления дифференцировочная среда содержит: ингибитор Wnt (например, IWP-2), ингибитор Notch (например, DAPT) и ингибитор пути EGFR (например, один или несколько из гефитиниба, афатиниба, PD0325901 и SCH772984) и активатор BMP (например BMP4). Эта дифференцировочная среда особенно пригодна для использования в способе получения популяции клеток, обогащенных секретин-секретирующими EEC. В некоторых вариантах осуществления этот способ приводит к получению популяции клеток, отрицательных по одному или нескольким маркерам, выбранным из Tac1, GLP1 и Chg; и положительных по одному или нескольким маркерам, выбранным из секретина, pyy и nts. В некоторых вариантах осуществления популяция клеток экспрессирует серотонин.
Среда для выращивания может являться любой подходящей средой для выращивания эпителиальных стволовых клеток или клеток-предшественников, предпочтительно, подходящей средой для выращивания эпителиальных стволовых клеток (например, как описано в WO 2010/090513, WO 2012/014076, WO 2012/168930 или WO 2015/173425).
В некоторых вариантах осуществления среда для выращивания содержит один или несколько лигандов рецепторных тирозинкиназ (например, EGF, HGF и/или FGF10), никотинамид и один или несколько агонистов Wnt (например, кондиционированная R-спондином среда и/или кондиционированная Wnt среда).
В некоторых вариантах осуществления среда для выращивания содержит один или несколько лигандов рецепторных тирозинкиназ (например, EGF, HGF и/или FGF10), один или несколько агонистов Wnt (например, кондиционированная R-спондином среда и/или кондиционированная Wnt среда) и один или несколько ингибиторов TGF-бета (например, A83-01).
В некоторых вариантах осуществления среда для выращивания содержит один или несколько лигандов рецепторных тирозинкиназ (например, EGF, HGF и/или FGF10), никотинамид, один или несколько агонистов Wnt (например, кондиционированная R-спондином среда и/или кондиционированная Wnt среда) и один или несколько ингибиторов TGF-бета (например, A83-01).
В любом из этих вариантов осуществления среда для выращивания может дополнительно содержать активатор пути цАМФ (например, форсколин), гастрин и/или ингибитор BMP (например, Noggin).
Например, в некоторых вариантах осуществления (например, в предпочтительном варианте осуществления в случае кишечника) среда для выращивания содержит (i) EGF (например, в количестве приблизительно от 10 до 50 нг/мл); (ii) кондиционированную Noggin среду (например, в количестве приблизительно от 50 до 100 нг/мл или приблизительно 5% конечного объема); (iii) кондиционированную R-спондином среду (например, в количестве приблизительно 1 мкг/мл или приблизительно 5% конечного объема). В некоторых вариантах осуществления среда для выращивания дополнительно содержит N-ацетилцистеин (например, в количестве приблизительно 1 мМ) и 1-кратный B27.
В некоторых вариантах осуществления среда для выращивания дополнительно содержит вальпроевую кислоту (например, в количестве приблизительно 1 мМ) и ингибитор GSK-3 (например, в количестве приблизительно 3 мкМ, такой как CHIR99021 в количестве приблизительно 3 мкМ). Обнаружено, что включение вальпроевой кислоты и ингибитора GSK-3 приводит к получению популяции клеток, обогащенных стволовыми клетками.
Предпочтительная среда для выращивания органоидов человека содержит (i) EGF (например, в количестве приблизительно от 10 до 50 нг/мл); (ii) кондиционированную Noggin среду (например, в количестве приблизительно от 50 до 100 нг/мл или приблизительно 5% конечного объема); (iii) кондиционированную R-спондином среду (например, в количестве приблизительно 1 мкг/мл или приблизительно 5% конечного объема); (iv) ингибитор p38 (например, SB-203580 в концентрации приблизительно 30 мкМ); (v) ингибитор TGF-β (например, A83-01 в концентрации приблизительно 500 нМ); и (vi) никотинамид (например, в концентрации приблизительно 10 мМ).
Предпочтительно, клетки выращивают для получения одного или нескольких (например, по меньшей мере 2, 3, 4, 5, 6, 10, 15, 20 или более 20) органоидов перед дифференцировкой.
В некоторых вариантах осуществления клетки исходно культивируют в среде для выращивания, представленной в настоящем описании и после успешного получения органоидов среду для выращивания заменяют дифференцировочной средой, представленной в настоящем описании. Таким образом, в некоторых вариантах осуществления после одного или нескольких (например, после двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти или более) пассажей среду для выращивания заменяют дифференцировочной средой. В некоторых вариантах осуществления пассаж осуществляют еженедельно. В некоторых вариантах осуществления через два, три, четыре, пять, шесть, семь, восемь, девять, десять или более дней среду для выращивания заменяют дифференцировочной средой. В предпочтительном варианте осуществления среду для выращивания заменяют дифференцировочной средой через пять или более дней.
В некоторых вариантах осуществления перед дифференцировкой индуцируют состояние покоя клеток.
В некоторых вариантах осуществления перед дифференцировкой эпителиальную стволовую клетку, популяцию эпителиальных стволовых клеток или выделенный фрагмент ткани промывают и высевают во внеклеточный матрикс (например, матригель). В некоторых вариантах осуществления промывку осуществляют с использованием минимальной среды или PBS. Не желая быть связанными какой-либо теорией, авторы настоящего изобретения полагают, что эта стадия промывки является предпочтительной для дифференцировки, т.к. посредством нее из эпителиальной стволовой клетки, популяции эпителиальных стволовых клеток или выделенного фрагмента ткани удаляют факторы стволовых клеток.
В некоторых вариантах осуществления эпителиальные стволовые клетки, популяцию эпителиальных стволовых клеток или выделенный фрагмент ткани культивируют в дифференцировочной среде, содержащей ингибитор BMP (например, Noggin) и R-спондин и несодержащей EGF, никотинамид, ингибитор TGFβ или кондиционированную Wnt среду, перед дифференцировкой в дифференцировочной среде по изобретению. В некоторых вариантах осуществления культивирование в первой дифференцировочной среде осуществляют в течение приблизительно одного дня.
Как будет очевидно специалисту в этой области, предпочтительные способы культивирования по изобретению являются предпочтительными, т.к. не требуются фидерные клетки. Слои фидерных клеток часто используют для поддержания культуры стволовых клеток и для ингибирования их дифференцировки. Слой фидерных клеток, как правило, является монослоем клеток, сокультивируемым с интересующими клетками и обеспечивающим поверхность, подходящую для их роста. Слой фидерных клеток обеспечивает среду, в которой могут расти интересующие клетки. Фидерные клетки зачастую являются митотически инактивированными (например, посредством облучения или обработки митомицином C) для предотвращения их пролиферации. Использование фидерных клеток является нежелательным, т.к. это осложняет пассирование клеток (при каждом пассаже клетки необходимо отделять от фидерных клеток, и при каждом пассаже необходимы новые фидерные клетки). Использование фидерных клеток также может приводить к контаминации желаемых клеток фидерными клетками. Очевидно, что это проблематично для какого-либо медицинского применения, и а при исследованиях осложняет анализ результатов любых экспериментов, осуществленных с использованием клеток. Как указано в настоящем описании, среды для культивирования по изобретению являются особенно предпочтительными, т.к. их можно использовать для культивирования клеток без контакта с фидерными клетками, т.е. в случае способов по изобретению не требуется слой фидерных клеток для поддержания клеток, рост которых они должны поддерживать.
Таким образом, композиции по изобретению могут являться композициями без фидерных клеток. Композицию общепринято считают несодержащей фидерные клетки, если клетки в композиции культивируют в течение по меньшей мере одного пассажа в отсутствие слоя фидерных клеток. Композиция без фидерных клеток по изобретению, как правило, будет содержать менее приблизительно 5%, менее приблизительно 4%, менее приблизительно 3%, менее приблизительно 2%, менее приблизительно 1% фидерных клеток (что выражено как % от общего количества клеток в композиции) или, предпочтительно, вообще не будет содержать фидерные клетки.
В дополнительном аспекте изобретение относится к способу получения популяции дифференцированных клеток или органоида, где способ включает культивирование клеток-предшественников в дифференцировочной среде по изобретению. Предпочтительно, способ включает культивирование клеток-предшественников в дифференцировочной среде по изобретению способом дифференцировки, как представлено в настоящем описании.
В некоторых вариантах осуществления способ включает получение органоида/популяции дифференцированных клеток из одной эпителиальной стволовой клетки. В другом варианте осуществления способ включает получение органоида/популяции дифференцированных клеток из популяции эпителиальных стволовых клеток или фрагмента эпителиальной ткани.
В конкретном варианте осуществления изобретение относится к способу получения дифференцированного органоида, где способ включает культивирование эпителиальных клеток-предшественников (например, эпителиальных стволовых клеток, необязательно, экспрессирующих Lgr5) в дифференцировочной среде по изобретению, предпочтительно, где эпителиальные клетки-предшественники приводят в контакт с ECM, предпочтительно, трехмерным ECM.
В некоторых вариантах осуществления способ включает культивирование клеток-предшественников в среде для выращивания в течение периода времени, например, от 3 дней до 10 недель, от 1 до 10 недель, от 1 до 4 недель или от 10 дней до 3 недель, а затем пассирование клеток (например, диссоциацию клеток до плотности отдельных клеток, высевание одной или нескольких клеток в соотношении 1 клетка на контейнер (например, на лунку)), продолжение выращивания клеток с использованием среды для выращивания в течение периода времени, например, от 3 дней до 10 недель, от 1 до 10 недель, от 1 до 4 недель или от 10 дней до 3 недель и повторение стадий пассирования и выращивания по меньшей мере один раза, по меньшей мере два раза, по меньшей мере три раза, по меньшей мере четыре раза, по меньшей мере пять раз, по меньшей мере шесть раз, по меньшей мере семь раз, по меньшей мере восемь раз, по меньшей мере девять раз, по меньшей мере десять раз, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз, по меньшей мере тринадцать раз или по меньшей мере четырнадцать раз перед дифференцировкой клеток.
В некоторых вариантах осуществления способ включает культивирование клеток-предшественников в дифференцировочной среде в течение по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14 дней. В некоторых вариантах осуществления способ включает культивирование клеток-предшественников в дифференцировочной среде в течение от приблизительно 1 до приблизительно 20 дней, от приблизительно 1 до приблизительно 10 дней или от приблизительно 1 до приблизительно 5 дней.
После дифференцировки, способ может дополнительно включать получение и/или выделение одной или нескольких дифференцированных клеток или дифференцированного органоида. Например, после культивирования клеток-предшественников полезным может быть удаление одной или нескольких клеток и/или одного или нескольких органоидов, культивируемых в среде для культивирования, из среды для культивирования для использования в дальнейшем. Для выбора клеток и их различения от других типов клеток можно использовать любой из ряда физических способов разделения, известных в этой области. Такие физические способы могут включать FACS и различные иммуноаффинные способы в зависимости от маркеров, специфически экспрессируемыми клетками по изобретению.
В одном из вариантов осуществления клетки можно выделять посредством FACS с использованием антитела, например, против одного из этих маркеров. Как будет очевидно специалисту в этой области, этого можно достигать с помощью флуоресцентно меченого антитела или флуоресцентно меченого вторичного антитела со специфичностью связывания в отношении первичного антитела. Неограничивающие примеры подходящих флуоресцентных меток включают FITC, Alexa Fluor® 488, GFP, CFSE, CFDA-SE, DyLight 488, PE, PerCP, PE-Alexa Fluor® 700, PE-Cy5 (TRI-COLOR®), PE-Cy5,5, PI, PE-Alexa Fluor® 750 и PE-Cy7. Этот список представлен исключительно в качестве примера и не предназначен для ограничения.
Альтернативно, клетки можно выделять посредство иммуноаффинной очистки, являющейся способом разделения, хорошо известным в этой области. Этот способ основан на иммобилизации антител на колонке для очистки. Затем образец клеток нагружают на колонку, позволяя соответствующим клеткам связаться с антителами и, таким образом, с колонкой. После стадии промывки клетки элюируют из колонки с использованием конкурентного вещества, предпочтительно связывающегося с иммобилизованным антителом и позволяющего клеткам высвобождаться из колонки.
Специалисту в этой области очевидно, что иммуноаффинная очистка с использованием иммобилизованного антитела будет приводить к получению очищенной популяции клеток. Однако в некоторых вариантах осуществления предпочтительной может являться очистка популяции клеток посредством осуществления дополнительного раунда иммуноаффинной очистки с использованием одного или нескольких из других идентифицируемых маркеров и аликвоты выделенных клонов для определения экспрессии других важных внутриклеточных маркеров.
Специалисту в этой области очевидно, что последовательные стадии очистки не обязательно должны включать одинаковый физический способ разделения.
Дифференцированные клетки и органоиды
Изобретение дополнительно относится к дифференцированному органоиду или популяции одной или нескольких дифференцированных клеток.
Дифференцированный органоид является трехмерной структурой, содержащей дифференцированные эпителиальные типы клеток. Дифференцированный органоид, как правило, является самоорганизующимся, что означает, что трехмерное распределение клеток в органоиде происходит спонтанно с дифференцировкой клеток. В некоторых вариантах осуществления дифференцированный органоид получают из эпителиальных стволовых клеток, необязательно, экспрессирующих Lgr5.
В одном из вариантов осуществления изобретение относится к дифференцированному органоиду или популяции одной или нескольких дифференцированных клеток, получаемых или полученных способом по изобретению, например, включающим культивирование клеток-предшественников в дифференцировочной среде по изобретению, предпочтительно, где клетки-предшественники приводят в контакт с трехмерным ECM.
"Популяция" клеток представляет собой любое количество клеток более 1, но предпочтительно - по меньшей мере 10 клеток, по меньшей мере 50 клеток, по меньшей мере 100 клеток, по меньшей мере 500 клеток, по меньшей мере 1×103 клеток, по меньшей мере 1×104 клеток, по меньшей мере 1×105 клеток, по меньшей мере 1×106 клеток, по меньшей мере 1×107 клеток, по меньшей мере 1×108 клеток или по меньшей мере 1×109 клеток.
Дифференцированный органоид по настоящему изобретению может содержать популяцию клеток из по меньшей мере 10 клеток, по меньшей мере 50 клеток, по меньшей мере 100 клеток, по меньшей мере 500 клеток, по меньшей мере 1×103 клеток, по меньшей мере 1×104 клеток, по меньшей мере 1×105 клеток, по меньшей мере 1×106 клеток, по меньшей мере 1×107 клеток или более. В некоторых вариантах осуществления каждый органоид содержит приблизительно от 1×103 до 5×103 клеток; как правило, 10-20 органоидов можно выращивать вместе в одной лунке, например, 24-луночного планшета.
Специалисту в этой области очевидно, что органоид по изобретению не является природным фрагментом ткани и/или не содержит кровеносный сосуд.
Органоиды по изобретению, например, отличаются от природной ткани, т.к. они содержат только эпителиальные типы клеток (и не мезенхимальные клетки или другие структурные типы клеток). Таким образом, в некоторых вариантах осуществления дифференцированный органоид по изобретению содержит только эпителиальные клетки. В некоторых вариантах осуществления дифференцированные органоиды не содержат неэпителиальные клетки. Например, в конкретном варианте осуществления дифференцированные органоиды не содержат мезенхимальные клетки.
Дифференцировочная среда, представленная в настоящем описании, предпочтительно, индуцирует или способствует специфической дифференцировке клеток в течение по меньшей мере пяти дней культивирования. Дифференцировку можно измерять посредством детекции наличия специфического маркера, ассоциированного с конкретной линией ткани, например, аргентофильной линией, как определено в настоящем описании. Дифференцировку можно измерять посредством детекции наличия специфического маркера, ассоциированного с линией ткани, например, аргентофильной линий, как определено в настоящем описании. В зависимости от маркера, экспрессию указанного маркера можно анализировать посредством RT-ПЦР или иммуногистохимии через по меньшей мере 5, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или более дней культивирования в дифференцировочной среде, как определено в настоящем описании.
Термин "экспрессируемый" используют для описания наличия маркера в клетке. Для того чтобы считаться экспрессируемым, маркер должен присутствовать на определимом уровне. Термин "определимый уровень" означает, что маркер можно определять с использованием одного из стандартных лабораторных способов, таких как ПЦР, блоттинг или анализ FACS. Ген считают экспрессируемым клеткой из популяции по изобретению, если экспрессию можно обосновано определить после 30 циклов ПЦР, что соответствует уровню экспрессии в клетке по меньшей мере приблизительно 100 копий на клетку. Термины "экспрессирует" и "экспрессия" имеют соответствующие значения. На уровне экспрессии ниже этого порогового значения маркер считают неэкспрессируемым. Сравнение между уровнем экспрессии маркера в клетке по изобретению и уровнем экспрессии того же маркера в другой клетке, такой как, например, эмбриональная стволовая клетка, предпочтительно, можно осуществлять посредством сравнения двух типов клеток, выделенных из одного вида. Предпочтительно, этот вид представляет собой млекопитающего и, более предпочтительно, представляет собой человека. Такое сравнение можно осуществлять с использованием эксперимента посредством полимеразной цепной реакции с обратной транскрипцией (RT-ПЦР).
Дифференцированный органоид по изобретению или популяция дифференцированных клеток по изобретению, предпочтительно, содержит по меньшей мере 50% жизнеспособных клеток, более предпочтительно - по меньшей мере 60% жизнеспособных клеток, более предпочтительно - по меньшей мере 70% жизнеспособных клеток, более предпочтительно - по меньшей мере 80% жизнеспособных клеток, более предпочтительно - по меньшей мере 90% жизнеспособных клеток. Жизнеспособность клеток можно анализировать с использованием окрашивания хехстом или окрашивания йодидом пропидия при FACS. Жизнеспособные клетки, предпочтительно, обладают соответствующими функциями и характеристиками in vivo. Например, жизнеспособные аргентофильные клетки, предпочтительно, обладают функциями или характеристиками аргентофильных клеток.
В предпочтительных вариантах осуществления органоид является органоидом кишечника. Это означает, что органоид получают из клеток кишечника.
В предпочтительных вариантах осуществления популяцию дифференцированных клеток по изобретению получают из клеток кишечника.
Авторы настоящего изобретения показали, что органоиды кишечника, полученные способами по изобретению, являются улучшенными, т.к. более высокая доля клеток в этих органоидах представляет собой аргентофильные клетки, чем в дифференцированных органоидах кишечника, описанных ранее в Grün et al. (2015) Nature 525(7568):251-5.
Некоторые из типов аргентофильных клеток, обнаруживаемых в желудочно-кишечном тракте млекопитающего, приведены в таблице 2 ниже.
Таблица 2: Аргентофильные клетки желудочно-кишечного тракта млекопитающего
Маркеры, характерные для EEC, включают Chga, Chgb, Tac1, Tph1, Gip, Fabp5, Ghrl, Pyy, Nts, Neurod1, Sst, Sct, холецистокинин, глюкагон и/или проглюкагон.
Дополнительные типы клеток EEC и их характеристики описаны в Grun et al., (2015), Nature 525:251-255.
В некоторых вариантах осуществления дифференцированный органоид по изобретению является дифференцированным органоидом кишечника, в котором по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток (например, Chga, Chgb, Tac1, Tph1, Gip, Fabp5, Ghrl, Pyy, Nts, Neurod1, Sst, Sct, холецистокинин, глюкагон и/или проглюкагон). В некоторых вариантах осуществления экспрессию мРНК измеряют посредством анализа секвенирования РНК отдельных клеток. В некоторых вариантах осуществления дифференцированный органоид по изобретению является дифференцированным органоидом кишечника, в котором по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркер аргентофильных клеток, характерный для конкретного типа аргентофильных клеток, в частности, типа клеток, описанного в таблице 2. Например, в некоторых вариантах осуществления маркером аргентофильных клеток, характерным для конкретного типа аргентофильных клеток, является Gcg и/или GLP-1. Gcg является геном, экспрессирующим препроглюкагон, из которого образуется GLP-1. В других вариантах осуществления маркером аргентофильных клеток, характерным для конкретного типа аргентофильных клеток, является Sct.
В некоторых вариантах осуществления дифференцированный органоид по изобретению является дифференцированным органоидом кишечника, в котором по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют один или несколько (например, 2, 3, 4 или более) продуктов, указанных в таблице 2.
В некоторых вариантах осуществления дифференцированный органоид по изобретению является дифференцированным органоидом кишечника, в котором менее 50%, менее 40%, менее 30%, менее 20%, менее 10% или менее 1% клеток экспрессируют маркеры бокаловидных клеток или клеток Панета (например, Lyz1, Defa6, Agr2, Gob5, Muc2, Ttf3 и/или Defa24). В некоторых вариантах осуществления экспрессию мРНК измеряют посредством анализа секвенирования РНК отдельных клеток.
В некоторых вариантах осуществления дифференцированный органоид по изобретению является дифференцированным органоидом кишечника, в котором менее 50%, менее 40%, менее 30%, менее 20%, менее 10% или менее 1% клеток экспрессируют маркеры энтероцитов (например, Aldob, Apoa1 и/или Alpi). В некоторых вариантах осуществления экспрессию мРНК измеряют посредством анализа секвенирования РНК отдельных клеток.
В некоторых вариантах осуществления дифференцированный органоид по изобретению является дифференцированным органоидом кишечника, в котором менее 50%, менее 40%, менее 30%, менее 20%, менее 10% или менее 1% клеток экспрессируют маркеры пучковых клеток (например, Dclk1 и/или Trpm5). В некоторых вариантах осуществления экспрессию мРНК измеряют посредством анализа секвенирования РНК отдельных клеток.
В некоторых вариантах осуществления дифференцированный органоид имеет пузырную структуру с центральным просветом. В некоторых вариантах осуществления центральный просвет окружен эпителиальным монослоем.
Изобретение также относится к дифференцированному органоиду или дифференцированной популяции аргентофильных клеток по изобретению в дифференцировочной среде по изобретению.
В одном из вариантов осуществления изобретение относится к органоиду в дифференцировочной среде, например, как представлено в настоящем описании.
В варианте осуществления дифференцированный органоид является органоидом, все еще культивируемым способом по изобретению и, таким образом, приведенным в контакт с внеклеточным матриксом. Предпочтительно, дифференцированный органоид заключен в немезенхимальный внеклеточный матрикс.
Органоид или популяция клеток-предшественников может происходить из ткани любого млекопитающего, но предпочтительно - из человека. В некоторых вариантах осуществления он происходит из мыши, кролика, крысы, морской свинки или другого не являющегося человеком млекопитающего.
Таблица 3 - Различия между дифференцированными органоидами кишечника и первичной тканью кишечника
Применение дифференцированных органоидов
Аналогично, изобретение относится к применению органоидов, представленных в настоящем описании, и клеток, полученных из органоидов. Когда в этом разделе упоминают органоиды, имеют в виду дифференцированные органоиды по изобретению. Специалисту в этой области также следует понимать, что применение также применимо к популяции дифференцированных клеток, полученных и/или получаемых способами по изобретению. Изобретение также относится к такому применению популяции дифференцированных клеток, полученных и/или получаемых способами по изобретению.
Например, изобретение относится к применению дифференцированного органоида или клетки, полученной из указанного органоида, в скрининге для обнаружения лекарственных средств; анализе токсичности; исследовании эмбриологии ткани, линий клеток и путей дифференцировки; исследовании для идентификации химических и/или нейронных сигналов, приводящих к высвобождению соответствующих гормонов; исследованиях экспрессии генов, включающих экспрессию рекомбинантных генов; исследовании механизмов, вовлеченных в повреждение и репарацию ткани; исследованиях воспалительных и инфекционных заболеваний; исследованиях патогенетических механизмов или исследованиях механизмов трансформации клеток и этиологии злокачественных новообразований.
Изобретение также относится к органоиду по изобретению или клетке, полученной из указанного органоида, для применения в медицине.
Изобретение также относится к органоиду по изобретению или клетке, полученной из указанного органоида, для применения в лечении нарушения, состояния или заболевания.
Изобретение также относится к органоиду по изобретению или клетке, полученной из указанного органоида, для применения в регенеративной медицине, например, где применение включает трансплантацию органоида или клетки пациенту.
Изобретение относится к применению органоида по изобретению или клетки, полученной из указанного органоида, в скрининге лекарственных средств, валидации мишени (лекарственного средства), обнаружении мишени (лекарственного средства), токсикологическом исследовании и токсикологическом скрининге, персонализированной медицине, регенеративной медицине и/или в качестве моделей клеток/органов ex vivo, таких как модели заболевания.
Считают, что клетки и органоиды, культивируемые с использованием сред и способов по изобретению, точно соответствуют условиям in vivo. Это верно и для популяций дифференцированных клеток и органоидов, выращенных из нормальной ткани, и для популяций дифференцированных клеток и органоидов, выращенных из пораженной ткани. Таким образом, помимо получения нормальных моделей клеток/органов ex vivo, органоиды по изобретению можно использовать в качестве моделей заболеваний ex vivo.
Органоиды по изобретению также можно использовать для культивирования патогена и, таким образом, их можно использовать в качестве моделей инфекций ex vivo. Примеры патогенов, которые можно культивировать с использованием органоида по изобретению, включают вирусы, бактерии, прионы или грибы, вызывающие заболевание у животного-хозяина. Таким образом, органоид по изобретению можно использовать в качестве модели заболевания, представляющей состояние инфекции. В некоторых вариантах осуществления изобретения, органоиды можно использовать при разработке и/или получении вакцин.
Заболевания, которые можно исследовать с помощью органоидов по изобретению, таким образом, включают генетические заболевания, метаболические заболевания, патогенетические заболевания, воспалительные заболевания и т.д., включая, в качестве неограничивающих примеров: диабет (такой как диабет типа I или типа II), кистозный фиброз, карциномы, аденокарциномы, аденомы, нейроэндокринные опухоли желудочно-кишечного тракта или поджелудочной железы, воспалительное заболевание кишечника (такое как болезнь Крона).
Общепринято, линии клеток, а с недавнего времени и клетки iPS, используют в качестве моделей клеток/органов и/или заболеваний ex vivo (например, см. Robinton et al., Nature 481, 295, 2012). Однако эти способы обладают рядом недостатков. Например, линии клеток нельзя получать из всех пациентов (только из конкретных биоптатов получают удачные линии клеток) и, таким образом, линии клеток нельзя использовать в персонализированной диагностике и медицине. Для клеток iPS, как правило, необходим некоторый уровень генетических манипуляций для перепрограммирования клеток в конкретном направлении. Альтернативно, их подвергают воздействию условий культивирования, влияющих на целостность кариотипа, и, таким образом, время культивирования необходимо делать минимальным (это также актуально для эмбриональных стволовых клеток человека). Это означает, что клетки iPS не могут точно соответствовать условиям in vivo, но вместо этого являются попыткой имитировать свойства клеток in vivo. Линии клеток и клетки iPS также страдают генетической нестабильностью.
В отличие от этого, органоиды по изобретению представляют собой генетически стабильную платформу, точно соответствующую условиям in vivo. В некоторых вариантах осуществления органоиды по изобретению содержат полностью дифференцированные типы клеток, присутствующие в соответствующих условиях in vivo. В других вариантах осуществления органоиды по изобретению можно подвергать дополнительной дифференцировке для получения полностью дифференцированных типов клеток, присутствующих in vivo. Таким образом, органоиды по изобретению можно использовать для достижения механистического понимания различных заболеваний и терапевтических средств, для осуществления скрининга лекарственных средств in vitro, для оценки потенциальных терапевтических средств, для идентификации возможных мишеней (например, белков) для разработки новых терапевтических средств (лекарственных средств) и/или для исследования генетической репарации, сопряженной с клеточной заместительной терапией.
Органоиды по изобретению можно замораживать и размораживать и помещать в культуру без утраты их генетической целостности или фенотипических характеристик и способности к пролиферации. Таким образом, органоиды легко можно хранить и транспортировать. Таким образом, в некоторых вариантах осуществления изобретение относится к замороженному органоиду.
По этой причине органоиды или дифференцированные популяции клеток по изобретению могут представлять собой инструмент для скрининга лекарственных средств, валидации мишени, обнаружения мишени, токсикологического исследования и токсикологического скрининга и персонализированной медицины.
Таким образом, в дополнительном аспекте изобретение относится к применению органоида или клетки, полученной из указанного органоида по изобретению, в скрининге для обнаружения лекарственных средств, анализа токсичности или в медицине, например, регенеративной медицине. Например, любой из органоидов кишечника можно использовать в скрининге для обнаружения лекарственных средств, анализа токсичности или в медицине, например, регенеративной медицине.
Слизистые вакцины
Дополнительным важным применением органоидов является разработка слизистых вакцин. Слизистые вакцины являются вакцинами, вводимыми через слизистую оболочку. Она может являться любой слизистой поверхностью, такой как слизистая оболочка носа, ротовой полости или прямой кишки. Их можно вводить с помощью ингалятора, спрея или других внешних средств. Они обладают несколькими очевидными преимуществами по сравнению с инъекциями, например, то, что для введения вакцины не требуется медицинский персонал, что может быть важным, например, в развивающихся странах.
В кишечнике M-клетки (или "микроскладчатые клетки") являются клетками, обнаруживаемыми в фолликул-ассоциированном эпителии пейеровых бляшек подвздошной кишки. Они транспортируют организмы и частицы из просвета кишечника к иммунным клеткам через эпителиальный барьер и, таким образом, важны для стимуляции слизистого иммунитета. Они обладают уникальной способностью к захвату антигена из просвета тонкого кишечника посредством эндоцитоза или фагоцитоза, а затем доставляют его посредством трансцитоза в дендритные клетки (антигенпрезентирующие клетки) и лимфоциты (а именно T-клетки), локализованные в уникальной карманообразной структуре на их базолатеральной стороне. M-клетки представляют собой тип аргентофильных клеток. Как описано выше, авторы настоящего изобретения обнаружили улучшенный способ дифференцировки клеток-предшественников в аргентофильные клетки. Таким образом, дифференцированные органоиды по изобретению обогащены аргентофильными клетками, такими как M-клетки.
В некоторых случаях органоиды могут развиваться в M-клетки при стимуляции лигандом RANK (например, см. фигуру 49 из WO 2012/169830). Таким образом, в некоторых вариантах осуществления дифференцировочная среда дополнительно содержит лиганд RANK.
Эффективность слизистых вакцин можно значительно повышать, если они направлены на M-клетки. Таким образом, популяцию дифференцированных клеток или органоид по изобретению можно использовать для тестирования способности M-клеток захватывать патогены или антигены и презентировать их иммунной системе. Таким образом, в некоторых вариантах осуществления изобретение относится к применению органоида по изобретению в скрининге лекарственных средств, например, в разработке и/или получении вакцин. Например, в некоторых вариантах осуществления органоид можно использовать для разработки или получения вакцин против вирусных, бактериальных, грибковых или других паразитарных инфекций, в качестве неограничивающих примеров, холеры, респираторно-синцитиального вируса (RSV), ротавируса и ВИЧ. В конкретном варианте осуществления изобретение относится к органоидам, дифференцированным в среде для культивирования по изобретению, для применения в разработке слизистых вакцин.
Скрининг лекарственных средств
Предпочтительно, в целях высокой производительности, указанный органоид по изобретению культивируют в многолуночных планшетах, таких как, например, 96-луночные планшеты или 384-луночные планшеты. Для идентификации молекулы, влияющей на указанные органоиды, используют библиотеки молекул. Предпочтительные библиотеки включают библиотеки фрагментов антител, библиотеки пептидного фагового дисплея, пептидные библиотеки (например, LOPAP™, Sigma Aldrich), липидные библиотеки (BioMol), библиотеки синтетических соединений (например, LOP AC™, Sigma Aldrich) или библиотеки природных соединений (Specs, TimTec). Кроме того, можно использовать генетические библиотеки, индуцирующие или репрессирующие экспрессию одного или нескольких генов в потомстве стволовых клеток. Эти генетические библиотеки включают библиотеки кДНК, библиотеки антисмысловых нуклеотидов и библиотеки миРНК или других некодирующих РНК. Клетки, предпочтительно, подвергают воздействию множества концентраций тестового средства в течение конкретного периода времени. В конце периода воздействия культуры подвергают оценке. Термин "влияющий" используют для охвата любого изменения в клетке, включая, в качестве неограничивающих примеров, снижение или утрату пролиферации, морфологическое изменение и гибель клеток. Указанный органоид по изобретению также можно использовать для идентификации лекарственных средств, специфически воздействующих на эпителиальные клетки карциномы, но не на указанный органоид по изобретению.
Возможность получения применимого органоида по изобретению за короткий период времени (дни) свидетельствует о том, что органоиды будут очень полезными для тестирования ответов отдельного пациента на конкретные лекарственные средства и адаптации лечения в соответствии с такой отвечаемостью. В некоторых вариантах осуществления, в которых органоид получают из биоптата из пациента, органоид культивируют в течение менее 21 дня, например, менее 14 дней, менее 13 дней, менее 12 дней, менее 11 дней, менее 10 дней, менее 9 дней, менее 8 дней, менее 7 дней (и т.д.).
Органоиды также являются полезными для целей более широкого обнаружения лекарственных средств (например, см. WO 2013/093812, где описывают скрининг лекарственных средств от кистозного фиброза или холеры). Таким образом, в некоторых вариантах осуществления органоиды по изобретению можно использовать для скрининга лекарственных средств от кистозного фиброза. Однако специалисту в этой области будет очевидно, что органоиды по изобретению будут применимы в качестве инструментов для скрининга лекарственных средств для инфекционных, воспалительных и неопластических патологий желудочно-кишечного тракта человека, других заболеваний желудочно-кишечного тракта и инфекционных, воспалительных и неопластических патологий и других заболеваний других тканей, представленных в настоящем описании, таких как поджелудочная железа, желудок или легкие. В некоторых вариантах осуществления органоиды по изобретению можно использовать для скрининга лекарственных средств от злокачественных новообразований.
В некоторых вариантах осуществления органоиды по изобретению можно использовать для тестирования библиотек химических веществ, антител, природных продуктов (экстрактов растений) и т.д. на пригодность для применения в качестве лекарственных средств, косметических средств и/или профилактических средств. Например, в некоторых вариантах осуществления клеточные биоптаты интересующего пациента, такие как опухолевые клетки пациента со злокачественным новообразованием, можно культивировать с использованием сред для культивирования и способов по изобретению, а затем обрабатывать химическим соединением или библиотекой химических веществ. Можно определять, с помощью каких соединений эффективно модифицируют, уничтожают и/или лечат клетки пациента. Это позволяет тестировать отвечаемость конкретного пациента на конкретное лекарственное средство, что, таким образом, позволяет адаптировать лечение для конкретного пациента. Таким образом, это делает возможным подход персонализированной медицины.
Таким образом, дополнительным преимуществом использования органоидов для идентификации лекарственных средств является то, что также можно осуществлять скрининг нормальных органоидов (органоидов, полученных из здоровой ткани) для проверки того, какие лекарственные средства и соединения имеют минимальный эффект в отношении здоровой ткани. Это делает возможным скрининг на лекарственные средства с минимальной нецелевой активностью или нежелательными побочными эффектами.
Таким образом, можно осуществлять скрининг лекарственных средств для любого количества заболеваний. Например, органоиды по изобретению можно использовать для скрининга лекарственных средств от диабета, кистозного фиброза, карцином, аденокарцином, аденом, нейроэндокринных опухолей желудочно-кишечного тракта или поджелудочной железы, воспалительного заболевания кишечника (такого как болезнь Крона) и т.д. Параметры тестирования зависят от интересующего заболевания. Например, при скрининге лекарственных средств от злокачественных новообразований конечной целью, как правило, является гибель злокачественных клеток. В случае кистозного фиброза интерес представляет измерение экспансии органоидов в ответ на лекарственные средства и стимулы CFTR. В других вариантах осуществления можно оценивать метаболизм или экспрессию генов для исследования эффектов соединений и лекарственных средств, подвергнутых скринингу, в отношении интересующих клеток или органоидов.
Таким образом, изобретение относится к способу скрининга терапевтического или профилактического фармацевтического лекарственного средства или косметического средства, включающему:
приведение популяции дифференцированных клеток или органоида в контакт с молекулой-кандидатом (или библиотекой молекул-кандидатов),
оценку указанной популяций дифференцированных клеток или органоида на любые эффекты (например, любое изменение в клетке, такое как снижение или утрата пролиферации, морфологическое изменение и/или гибель клеток) или изменение в органоиде (например, размер или подвижность органоида);
идентификацию молекулы-кандидата, вызывающей указанные эффекты, в качестве потенциального лекарственного средства или косметического средства; и, необязательно,
получение указанной молекулы-кандидата в качестве фармацевтического или косметического средства.
В некоторых вариантах осуществления изобретение относится к способу получения фармацевтического или косметического средства, включающему:
приведение популяции дифференцированных клеток или органоида в контакт с молекулой-кандидатом (или библиотекой молекулы-кандидатов),
оценку указанной популяций дифференцированных клеток или органоида на любые эффекты (например, любое изменение в клетке, такое как снижение или утрата пролиферации, морфологическое изменение и/или гибель клеток) или изменение в органоиде (например, размер или подвижность органоида);
идентификацию молекулы-кандидата, вызывающей указанные эффекты, в качестве потенциального лекарственного средства или косметического средства; и, необязательно,
получение указанной молекулы-кандидата в качестве фармацевтического или косметического средства.
В некоторых вариантах осуществления для повышения производительности скрининга используют компьютеризированные или роботизированные способы культивирования и сбора данных. В некоторых вариантах осуществления органоид получают из биоптата пациента. В некоторых вариантах осуществления молекулу-кандидата, вызывающую желаемый эффект в отношении культивируемой дифференцированной популяции клеток (например, органоида), вводят указанному пациенту.
Таким образом, в одном из аспектов изобретение относится к способу лечения пациента, включающему:
(a) получение биоптата из интересующей пораженной ткани пациента;
(b) культивирование биоптата для получения органоида;
(c) скрининг подходящего лекарственного средства с использованием способа скрининга по изобретению; и
(d) лечение указанного пациента с использованием лекарственного средства, полученного на стадии (c).
В некоторых вариантах осуществления лекарственное средство или косметическое средство используют для лечения, профилактики или облегчения симптомов генетических заболеваний, метаболических заболеваний, патогенетических заболеваний, воспалительных заболеваний и т.д., включая, в качестве неограничивающих примеров: кистозный фиброз, воспалительное заболевание кишечника (такое как болезнь Крона), карциному, аденому, аденокарциному, рак толстого кишечника, диабет (такой как диабет типа I или типа II), нейроэндокринные опухоли желудочно-кишечного тракта или поджелудочной железы и т.д.
В некоторых вариантах осуществления изобретение относится к способам скрининга лекарственных средств для регенеративной медицины.
Обнаружение мишеней
В некоторых вариантах осуществления органоиды по изобретению можно использовать для обнаружения мишеней. Клетки органоидов, происходящие из здоровой или пораженной ткани, можно использовать для идентификации мишеней. Органоиды по изобретению можно использовать для обнаружения мишеней лекарственных средств от кистозного фиброза, воспалительного заболевания кишечника (такого как болезнь Крона), карциномы, аденомы, аденокарциномы, рака толстого кишечника, диабета (такого как диабет типа I или типа II), нейроэндокринных опухолей желудочно-кишечного тракта или поджелудочной железы и т.д. Считают, что органоиды, культивируемые с использованием сред и способов по изобретению, точно соответствуют условиям in vivo. По этой причине они могут представлять собой инструмент для обнаружения новых (молекулярных) мишеней при конкретных заболеваниях.
Для поиска новой мишени лекарственного средства можно использовать библиотеку соединений (таких как миРНК) для трансдукции клеток и инактивации конкретных генов. В некоторых вариантах осуществления клетки трансдуцируют с использованием миРНК для ингибирования функции (большой) группы генов. Для определения того, подходит ли мишень для исследования, можно использовать любой функциональный показатель для группы генов или конкретной клеточной функции. Специфический для заболевания показатель можно определять с использованием анализов, хорошо известных в этой области. Например, анализируют пролиферацию клеток для тестирования генов, ассоциированных со злокачественным новообразованием. Например, для определения изменений активности Wnt, вызванной ингибированием миРНК, можно использовать анализ Topflash, как представлено в настоящем описании. Если происходит снижения роста или гибель клеток, можно идентифицировать соответствующие связанные с миРНК гены известными в этой области способами. Эти гены являются возможными мишенями для ингибирования роста этих клеток. После идентификации необходимо определять специфичность идентифицированной мишени для исследуемого клеточного процесса способами, хорошо известными в этой области. Используя эти способы, можно идентифицировать новые молекулы в качестве возможных мишеней лекарственных средств для терапии.
Скрининги для валидации мишеней и лекарственных средств
Специфичные для пациента органоиды, полученные мз пораженной и/или нормальной ткани, можно использовать для валидации мишеней молекул, идентифицированных при высокопроизводительном скрининге. То же актуально для валидации соединений, идентифицированных в качестве терапевтических лекарственных средств при высокопроизводительном скрининге. Использование первичного материала пациента, подвергнутого дифференцировке в системе культивирования органоидом, может быть полезным для тестирования на ложноположительные результаты и т.д. при высокопроизводительных исследованиях обнаружения лекарственных средств на линиях клеток.
В некоторых вариантах осуществления органоид по изобретению можно использовать для валидации соединений, идентифицированных в качестве возможных лекарственных средств или косметических средств при высокопроизводительном скрининге.
Культивирование патогенов
Кроме того, органоид по изобретению можно использовать для культивирования патогена, такого как норовирус, для которого в настоящее время отсутствует подходящая культура ткани или модель на животных.
Регенеративная медицина и трансплантация
Изобретение относится к применению органоидов в регенеративной медицине и/или трансплантации. Изобретение также относится к способам лечения, где способ включает трансплантацию органоида животному или человеку.
Органоиды по изобретению, такие как органоиды желудка, органоиды кишечника или органоиды поджелудочной железы, применимы в регенеративной медицине, например, в восстановлении эпителия кишечника после облучения и/или хирургического вмешательства, восстановлении эпителия кишечника у пациентов, страдающих воспалительным заболеванием кишечника, таким как болезнь Крона и язвенный колит, и восстановлении эпителия кишечника у пациентов, страдающих синдромом короткой кишки. Кроме того, их можно использовать в восстановлении эпителия кишечника у пациентов с наследственными заболеваними тонкого кишечника/толстого кишечника. Культуры, содержащие органоиды поджелудочной железы, также применимы в регенеративной медицине, например, в качестве имплантатов после резекции поджелудочной железы или ее части и для лечения диабета, такого как диабет типа I и диабет типа II.
В альтернативном варианте осуществления органоиды или клетки, выделенные из органоидов, перепрограммируют в направлении родственной ткани, например, клеток поджелудочной железы, включая бета-клетки поджелудочной железы. Способы культивирования по настоящему изобретению позволят анализировать факторы, трансдифференцирующие близкородственную клетку-предшественника в клетку поджелудочной железы, включая бета-клетку поджелудочной железы или гепатоцит.
Специалисту в этой области будет очевидно, что в способе, направленном на восстановление поврежденной или пораженной ткани, дополнительно можно использовать генную терапию. Например, можно использовать аденовирусное или ретровирусное средство доставки гена для доставки генетической информации, подобной ДНК и/или РНК, в стволовые клетки. Специалист в этой области может заменять или восстанавливать конкретные гены, на которые направлена генная терапия. Например, можно встраивать нормальный ген в неспецифическое место в геноме для замены нефункционального гена. В другом примере, аномальную последовательность гена можно заменять нормальной последовательностью гена посредством гомологичной рекомбинации. Альтернативно, с помощью селективной обратной мутации можно возвращать ген к его нормальному функционированию. Дополнительным примером является изменение регуляции (степени, до которой ген активируют или ингибируют) конкретного гена. Предпочтительно, клетки органоида или клетки, полученные из органоида, обрабатывают ex vivo с помощью подхода генной терапии, а затем переносят в организм млекопитающего, предпочтительно, человека, нуждающегося в лечении.
Т.к. небольшие биоптаты от взрослых доноров можно выращивать без очевидного ограничения или генетического повреждения, эту технологию можно использовать для получения трансплантируемого эпителия для регенеративных целей. Тот факт, что органоиды можно замораживать, размораживать и помещать в культуру без утраты их трехмерной структуры и целостности и без значительной гибели клеток, дополнительно свидетельствует в пользу применимости органоидов для трансплантации. Кроме того, в некоторых вариантах осуществления органоиды, заключенные или приведенные в контакт с ECM, можно трансплантировать млекопитающему, предпочтительно, человеку. В другом варианте осуществления органоиды и ECM можно трансплантировать млекопитающему, предпочтительно, человеку, одновременно.
Специалисту в этой области будет понятно, что ECM можно использовать в качестве трехмерного каркаса для получения ткане-подобных структур, содержащих выращенные популяции клеток или органоиды по изобретению. Такие структуры затем можно трансплантировать пациенту способами, хорошо известными в этой области. Каркас ECM можно получать синтетически с использованием белков ECM, таких как коллаген и/или ламинин, или, альтернативно, каркас ECM можно получать посредством "децеллюляризации" выделенного органа или фрагмента ткани для получения каркаса, состоящего из ECM (например, см. Macchiarini et al. The Lancet, Volume 372, Issue 9655, Pages 2023-2030, 2008). В некоторых вариантах осуществления каркас ECM можно получать посредством децеллюляризации органа или фрагмента ткани, где, необязательно, указанный орган или фрагмент ткани получают из кишечника, поджелудочной железы, легких или желудка.
Изобретение относится к органоиду по изобретению или клеткам, полученным из указанного органоида, для применения в трансплантации млекопитающему, предпочтительно, человеку. Изобретение также относится к способу лечения пациента, нуждающегося в трансплантации, включающему трансплантацию органоида по изобретению или клеток, полученных из указанного органоида, указанному пациенту, где указанный пациент представляет собой млекопитающего, предпочтительно, человека. В некоторых вариантах осуществления органоид дополнительно подвергают дифференцировке перед трансплантацией указанному пациенту.
Например, небольшой биоптат получают из взрослого донора, выращивают способом экспансии, а затем подвергают дифференцировке по изобретению. Таким образом, технологию, представленную в настоящем описании, можно использовать для получения трансплантируемого эпителия для регенеративных целей.
Изобретение относится к способу лечения нарушения с недостаточностью инсулина, такого как диабет, у пациента или пациента, имеющего дисфункцию поджелудочной железы, включающему трансплантацию пациенту органоида поджелудочной железы по изобретению или клеток из органоида поджелудочной железы по изобретению.
В некоторых вариантах осуществления клетки или органоид не экспрессируют или секретируют инсулин после трансплантации пациенту, но дифференцируются в организме пациента таким образом, что они секретируют инсулин. Например, способность секретировать инсулин может быть неопределимой сразу после трансплантации, но может иметь место приблизительно через один месяц после трансплантации, например, через 6 недель, 2 месяца или 3 месяца после трансплантации.
Пациент, предпочтительно, является человеком, но альтернативно может являться не являющимся человеком млекопитающим, таким как кошка, собака, лошадь, корова, свинья, овца, кролик или мышь.
Таким образом, в объем изобретения входят способы лечения человека или не являющегося человеком животного посредством клеточной терапии. Такая клеточная терапия включает использование стволовых клеток или органоидов по изобретению в отношении пациента любыми соответствующими способами. В частности, такие способы лечения включают регенерацию поврежденной ткани. По изобретению, пациента можно лечить аллогенными или аутологичными стволовыми клетками или органоидами. "Аутологичные" клетки являются клетками, происходящими из того же организма, в который его повторно вводят для клеточной терапии, например, чтобы сделать возможной регенерацию ткани. Однако клетки не обязательно выделять из той же ткани, что и ткань, в которую их вводят. При использовании аутологичных клеток не требуется совпадение с пациентом для преодоления проблем отторжения. "Аллогенные" клетки являются клетками, происходящими из индивидуума, отличающегося от индивидуума, которому вводят клетки для клеточной терапии, например, чтобы сделать возможной регенерацию ткани, хотя и принадлежащего тому же виду. Но все же может требоваться некоторая степень совпадения пациента для предотвращения проблем отторжения. Таким образом, в некоторых вариантах осуществления трансплантация включает аутологичные клетки. В некоторых вариантах осуществления трансплантация включает аллогенные клетки.
Как правило, клетки или органоиды по изобретению вводят в организм пациента посредством инъекции или имплантации. Как правило, клетки будут инъецировать непосредственно в ткань, в которой они должны действовать. Альтернативно, клетки будут инъецировать через воротную вену. Шприц, содержащий клетки по изобретению и фармацевтически приемлемый носитель, входит в объем изобретения. Катетер, прикрепленный к шприцу, содержащему клетки по изобретению и фармацевтически приемлемый носитель, входит в объем изобретения.
Специалист в этой области может выбирать подходящий способ и путь введения в зависимости от трансплантируемого материала (т.е. популяции клеток, отдельных клеток в суспензии клеток, органоидов или фрагментов органоидов), а также органа, подвергаемого лечению.
Как указано выше, органоид или клетки по изобретению можно использовать в регенерации ткани. Для достижения этой функции клетки можно инъецировать или имплантировать непосредственно в поврежденную ткань, где они могут делиться и, в конечном итоге, дифференцироваться в необходимый тип клеток в соответствии с их локализацией в организме. Альтернативно, органоид можно инъецировать или имплантировать непосредственно в поврежденную ткань. Ткани, восприимчивые к лечению, включают все поврежденные ткани, в частности, включая ткани, которые могут быть повреждены в результате заболевания, повреждения, травмы, аутоиммунной реакции или вирусной или бактериальной инфекции. В некоторых вариантах осуществления изобретения клетки или органоиды по изобретению используют для регенерации толстого кишечника, тонкого кишечника, легких, поджелудочной железы или желудочно-кишечного тракта.
Например, в одном из вариантов осуществления клетки или органоиды по изобретению инъецируют пациенту с использованием шприца Гамильтона.
Специалисту в этой области будут известны подходящие дозы клеток или органоидов по изобретению для конкретного состояния, подвергаемого лечению.
В одном из вариантов осуществления органоиды или клетки по изобретению в растворе, микросферах или микрочастицах в различных композициях будут вводить в артерию, снабжающую ткань или часть поврежденного органа, нуждающегося в регенерации. Как правило, такое введение будут осуществлять с использованием катетера. Катетер может являться катетером из широкого спектра балонных катетеров, используемых для ангиопластики и/или доставки клеток, или катетером, созданным с конкретной целью доставки клеток в конкретное место в организме. Для конкретного применения клетки или органоиды можно инкапсулировать в микросферы, полученные из ряда различных биодергадируемых соединений, диаметром приблизительно 15 мкм. Этим способом можно интраваскулярно вводить клетки или органоиды так, чтобы они оставались в месте повреждения и не проходили через капиллярную сеть и в системный кровоток при первом прохождении. Удержание в артериальной части капиллярной сети также может облегчать их транслокацию во внесосудистом пространстве.
В другом варианте осуществления органоиды или клетки можно ретроградно инъецировать в сосудистое русло через вену для доставки в весь организм или место в конкретную вену, дренирующую ткань или часть тела, для которой предназначены клетки или органоиды. В этом варианте осуществления можно использовать многие из препаратов, описанных выше.
В другом варианте осуществления клетки или органоиды по изобретению можно имплантировать в поврежденную ткань прикрепленными к биосовместимому имплантату. В этом варианте осуществления клетки можно прикреплять к биосовместимому имплантату in vitro перед имплантацией пациенту. Как будет очевидно специалисту в этой области, для прикрепления клеток к имплантату перед имплантацией можно использовать любой из ряда адгезивов. Исключительно в качестве примера, такие адгезивы могут включать фибрин, один или несколько членов семейства интегринов, один или несколько членов семейства кадгеринов, один или несколько членов семейства селектинов, одну или несколько молекул клеточной адгезии (CAM), один или несколько членов семейства иммуноглобулинов и один или несколько искусственных адгезивов. Этот список представлен исключительно в качестве примера и не предназначен для ограничения. Специалисту в этой области будет очевидно, что можно использовать любую комбинацию одного или нескольких адгезивов.
В другом варианте осуществления органоиды или клетки по изобретению можно заключать в матрикс перед трансплантацией матрикса пациенту. Как правило, матрикс будут имплантировать в поврежденную ткань пациента. Примеры матриксов включают матриксы на основе коллагена, матриксы на основе фибрина, матриксы на основе ламинина, матриксы на основе фибронектина и искусственные матриксы. Этот список представлен исключительно в качестве примера и не предназначен для ограничения.
В дополнительном варианте осуществления органоиды или клетки по изобретению можно имплантировать или инъецировать пациенту вместе с компонентом, образующим матрикс. Это может позволить клеткам образовывать матрикс после инъекции или имплантации обеспечивать, чтобы клетки или органоиды оставались в соответствующем месте в организме пациента. Примеры компонентов, образующих матрикс, включают фибриновый клей, жидкий алкил, мономеры цианоакрилата, пластификаторы, полисахариды, такие как декстран, этиленоксид-содержащие олигомеры, блок-сополимеры, такие как полоксамер и плюроники, неионные поверхностно-активные средства, такие как Tween и Triton'8', и искусственные компоненты, образующие матрикс. Этот список представлен исключительно в качестве примера и не предназначен для ограничения. Специалисту в этой области будет очевидно, что можно использовать любую комбинацию одного или нескольких компонентов, образующих матрикс.
В дополнительном варианте осуществления органоиды или клетки по изобретению могут содержаться в микросфере. В этом варианте осуществления клетки можно инкапсулировать в центре микросферы. В этом варианте осуществления клетки можно заключать в материал матрикса микросферы. Материал матрикса может включать любой подходящий биодеградируемый полимер, включая, в качестве неограничивающих примеров, альгинаты, полиэтиленгликоль (PLGA) и полиуретаны. Этот список представлен исключительно в качестве примера и не предназначен для ограничения.
В дополнительном варианте осуществления клетки или органоиды по изобретению можно прикреплять к медицинскому устройству, предназначенному для имплантации. Примеры таких медицинских устройств включают стенты, стержни, шовный материал, сплиты, протезы суставов, искусственную кожу и штифты. Этот список представлен исключительно в качестве примера и не предназначен для ограничения. Специалисту в этой области будет очевидно, что клетки можно прикреплять к медицинскому устройству множеством способов. Например, клетки или органоиды можно прикреплять к медицинскому устройству с использованием фибрина, одного или нескольких членов семейства интегринов, одного или нескольких членов семейства кадгеринов, одного или нескольких членов семейства селектинов, одной или нескольких молекул клеточной адгезии (CAM), одного или нескольких членов семейства иммуноглобулинов и одного или нескольких искусственных адгезивов. Этот список представлен исключительно в качестве примера и не предназначен для ограничения. Специалисту в этой области будет очевидно, что можно использовать любую комбинацию одного или нескольких адгезивов.
Органоид или популяция дифференцированных клеток, полученные способом по изобретению, имеют различное применение. Например, изобретение относится к применению органоида или популяции дифференцированных клеток, как представлено в настоящем описании, в скрининге для обнаружения лекарственных средств; анализе токсичности; исследовании эмбриологии ткани, линий клеток и путей дифференцировки; исследовании для идентификации химических и/или нейронных сигналов, приводящих к высвобождению соответствующих гормонов; исследованиях экспрессии генов, включая экспрессию рекомбинантных генов; исследованиях механизмов, вовлеченных в повреждение и репарацию ткани; исследованиях воспалительных и инфекционных заболеваний; исследованиях патогенетических механизмов или исследованиях механизмов трансформации клеток и этиологии злокачественных новообразований.
В одном из аспектов изобретение относится к применению органоида или популяции дифференцированных клеток, как представлено в настоящем описании, в скрининге для обнаружения лекарственных средств, анализе токсичности или регенеративной медицине. Аналогично, изобретение относится к применению потомства органоидов по изобретению для этих целей.
Анализы токсичности могут являться анализами in vitro с использованием органоида, или его части, или клетки, полученной из органоида. Такое потомство и органоиды легко культивировать, и они больше напоминают первичные эпителиальные клетки, чем, например, линии эпителиальных клеток, такие как Caco-2 (ATCC HTB-37), I-407 (ATCC CCL6) и XBF (ATCC CRL 8808), в настоящее время используемые в анализах токсичности. Предполагают, что результаты анализов токсичности, полученные с использованием органоидов, больше напоминают результаты, полученные на пациентах. Клеточный анализ токсичности используют для определения органоспецифической цитотоксичности. Соединения, тестируемые в указанном тесте, включают химиотерапевтические средства против злокачественных новообразований, химические вещества из окружающей среды, пищевые добавки и потенциальные токсины. Клетки подвергают воздействию множества концентраций тестируемого средства в течение конкретного периода времени. Диапазоны концентраций тестируемых средств в анализе определяют при предварительном анализе с использованием воздействия в течение пяти дней и логарифмических разведений наибольшей растворимой концентрации. В конце периода воздействия культуры оценивают на ингибирование роста. Данные анализируют для определения концентрации, при которой происходит ингибирование на 50 процентов (TC50).
Например, в этом аспекте изобретения соединение-кандидата можно приводить в контакт с клеткой или органоидом, как представлено в настоящем описании, и подвергать мониторингу любое изменение клеток или активности клеток.
В целях высокой производительности указанные органоиды культивируют в многолуночных планшетах, таких как, например, 96-луночные планшеты или 384-луночные планшеты. Для идентификации молекулы, влияющей на указанные органоиды, используют библиотеки молекул. Предпочтительные библиотеки включают библиотеки фрагментов антител, библиотеки пептидного фагового дисплея, пептидные библиотеки (например, LOPAP™, Sigma Aldrich), липидные библиотеки (BioMol), библиотеки синтетических соединений (например, LOP AC™, Sigma Aldrich) или библиотеки природных соединений (Specs, TimTec). Кроме того, можно использовать генетические библиотеки, индуцирующие или репрессирующие экспрессию одного или нескольких генов в потомстве клеток аденомы. Эти генетические библиотеки включают библиотеки кДНК, библиотеки антисмысловых нуклеотидов и библиотеки миРНК или других некодирующих РНК. Клетки, предпочтительно, подвергают воздействию множества концентраций тестируемого средства в течение конкретного периода времени. В конце периода воздействия культуры подвергают оценке. Термин "влияющий" используют для охвата любого изменения в клетке, включая, в качестве неограничивающих примеров, снижение или утрату пролиферации, морфологическое изменение и гибель клеток. Указанные органоиды также можно использовать для идентификации лекарственных средств, специфически воздействующих на эпителиальные клетки карциномы, но не на указанные органоиды.
Кроме того, органоиды по изобретению могут заменять использование линий клеток, таких как клетки Caco-2, в скрининге для обнаружения лекарственных средств и анализах токсичности потенциальных новых лекарственных средств, или известных лекарственных средств, или известных или новых пищевых добавок.
Кроме того, такие органоиды можно использовать для культивирования патогена.
Изобретение дополнительно относится к дифференцированному органоиду по изобретению или популяции дифференцированных клеток по изобретению для применения в терапии. Изобретение также относится к дифференцировке органоида по изобретению или клетки, полученной из указанного органоида, для применения в лечении заболевания или состояния, как представлено в настоящем описании.
Аналогично, изобретение относится к способу лечения заболевания или состояния, как представлено в настоящем описании, включающему введение одного или нескольких органоидов по изобретению или клетки, полученной из указанного органоида.
Авторы настоящего изобретения также продемонстрировали успешную трансплантацию органоидов у мышей с иммунодефицитом (см. пример 7 в WO 2012/014076) с использованием трансплантированных клеток, полученных из органоида печени, образующих холангиоциты и гепатоциты in vivo. Таким образом, в одном из вариантов осуществления изобретение относится к органоидам или клеткам, полученным из органоида, по изобретению для трансплантации человеку или животным.
Преимуществом органоидов по изобретению является то, что их можно замораживать, а позднее размораживать без утраты функции. Это делает возможным создание банков клеток, простое хранение и доступность для срочного использования. Это может быть полезным, например, в получении "готового к использованию" продукта, например, в случае печени, который можно использовать для лечения острого токсического поражения печени. Органоиды также можно выращивать из клеток или фрагментов ткани, полученных из небольших биоптатов от живых доноров, что минимизирует какие-либо этические проблемы, связанные с лечением. Донор даже может являться пациентом, подлежащим лечению, которое может снизить какие-либо отрицательные побочные эффекты, ассоциированные с трансплантацией чужеродных клеток и органов, и снижать потребность в иммуносупрессорных лекарственных средствах.
Фармацевтические составы
В некоторых вариантах осуществления изобретение также относится к фармацевтическому составу, содержащему компоненты дифференцировочной среды, представленной в настоящем описании, и фармацевтически приемлемый дилюент и/или эксципиент. Например, изобретение относится к фармацевтическому составу, содержащему один или несколько ингибиторов Wnt (например, IWP-2), один или несколько ингибиторов пути EGFR (например, один или несколько из гефитиниба, афатиниба, PD0325901 и SCH772984), один или несколько ингибиторов Notch (например, DAPT) и фармацевтически приемлемый дилюент и/или эксципиент. В предпочтительном варианте осуществления фармацевтический состав не содержит минимальную среду. В некоторых вариантах осуществления фармацевтический состав не содержит внеклеточный матрикс. Предполагают, что такие составы могут подходить для стимуляции дифференцировки стволовых клеток in vivo, например, для регенеративной терапии. Такие составы можно вводить in situ (например, в участок повреждения ткани) или системно. Альтернативно, составы можно составлять таким образом, что они подходят для введения любыми способами введения, известными в этой области, например, внутривенного, подкожного, внутримышечного, слизистого, интрадермального, внутрикожного, перорального и глазного введения. Таким образом, фармацевтический состав может находиться в любой форме, подходящей для такого введения, например, таблетке, инфузионной жидкости, капсуле, сиропе и т.д.
В некоторых вариантах осуществления изобретение относится к фармацевтическому составу, содержащему один или несколько ингибиторов Wnt (например, IWP-2), один или несколько ингибиторов пути EGFR (например, один или несколько из гефитиниба, афатиниба, PD0325901 и SCH772984), один или несколько ингибиторов Notch (например, DAPT), один или несколько ингибиторов BMP (например, дорсоморфин или LDN193189) и фармацевтически приемлемый дилюент и/или эксципиент.
В некоторых вариантах осуществления изобретение относится к фармацевтическому составу, содержащему один или несколько ингибиторов Wnt (например, IWP-2), один или несколько ингибиторов пути EGFR (например, один или несколько из гефитиниба, афатиниба, PD0325901 и SCH772984), один или несколько ингибиторов Notch (например, DAPT), один или несколько активаторов BMP (например, BMP4, BMP7 или BMP2) и фармацевтически приемлемый дилюент и/или эксципиент.
Способы лечения
Модуляция уровней гормонов in vivo
Изобретение также относится к способам лечения, включающим один или несколько компонентов дифференцировочной среды и/или фармацевтической композиции по изобретению. В частности, предполагают, что EEC in vivo можно направлять в сторону конкретных фенотипов EEC, экспрессирующих конкретные гормоны, и, таким образом, в некоторых вариантах осуществления способы по изобретению можно использовать для модуляции уровней гормонов in vivo.
Например, авторы настоящего изобретения показали, что активаторы BMP способствуют секреции секретина (и супрессируют секрецию GLP-1) в EEC (см. пример 5). Таким образом, предполагают, что активатор BMP или фармацевтическая композиция, содержащая активатор BMP (с другими компонентами дифференцировочной среды, представленной в настоящем описании, или без них), могут быть полезными для медицинского применения, включающего повышенные уровни секретина или супрессированные уровни GLP-1. Секретин ассоциирован с нейтрализацией pH желудка посредством ингибирования секреции желудочного сока (Afroze et al., Ann Transl Med. 2013 Oct; 1(3): 29) и подавления аппетита (Cheng et al., Neuropsychopharmacology. 2011 Jan; 36(2): 459-471). Таким образом, повышение уровней секретина in vivo может быть применимым механизмом для лечения гиперхлоргидрии (избытка желудочного сока) или ожирения.
Таким образом, изобретение относится к способу лечения гиперхлоргидрии или ожирения, где способ включает введение активатора BMP нуждающемуся в этом индивидууму. Изобретение также относится к активатору BMP для применения в способе лечения гиперхлоргидрии или ожирения, где способ включает введение терапевтически эффективного количества активатора BMP нуждающемуся в этом индивидууму. Изобретение также относится к применению активатора BMP для производства лекарственного средства для лечения гиперхлоргидрии или ожирения, где способ включает введение терапевтически эффективного количества активатора BMP нуждающемуся в этом индивидууму. Примеры подходящих активаторов BMP известны в этой области и представлены в настоящем описании выше.
Авторы настоящего изобретения также показали, что ингибиторы BMP стимулируют секрецию GLP-1 (и супрессируют секрецию секретина) в EEC (см. пример 5). Таким образом, предполагают, что ингибитор BMP или фармацевтическая композиция, содержащая ингибитор BMP (с другими компонентами дифференцировочной среды, представленной в настоящем описании, или без них), могут быть полезными для медицинского применения, включающего повышенные уровни GLP-1 или супрессированные уровни секретина.
Например, GLP-1 (глюкагон-подобный пептид-1) является эндогенным инкретином и играет значительную роль в гомеостазе глюкозы (Manandhar & Ahn J Med Chem. 2015 Feb 12; 58(3): 1020-1037). Он связывается с рецептором GLP-1 (GLP-1R), принадлежащим к семейству класса B сопряженных с G-белком рецепторов (GPCR), и активирует его, влияя на его регуляторные функции. Активация рецептора на β-клетках приводит к быстрому повышению уровней цАМФ и внутриклеточного кальция с последующим экзоцитозом инсулина глюкозо-зависимым образом. Хотя GLP-1R в α-клетках составляет <0,2% от GLP-1R в β-клетках, GLP-1 ингибирует секрецию глюкагона на 50% посредством модуляции активности кальциевого канала. Показано, что терапия GLP-1 потенцирует секрецию инсулина у здоровых индивидуумов и пациентов с диабетом. В отличие от других лекарственных средств против диабета, инсулинотропный эффект GLP-1 является самограничивающимся, т.к. он снижается, когда уровень глюкозы в плазме снижается до нормального диапазона, что снижает риск гипогликемии. Кроме того, GLP-1 регулирует постпрандиальное повышение глюкозы с помощью нескольких других механизмов, включая стимуляцию транскрипции гена инсулина, стимуляцию пролиферацию и неогенез β-клеток поджелудочной железы, ингибирование апоптоза β-клеток и блокирование высвобождения глюкагона. Он также предотвращает опорожнение желудка и способствует насыщению, что приводит к снижению массы тела. Предполагают, что терапия GLP-1 также имеет кардиопротективный эффект. Однако эндогенный GLP-1 имеет очень небольшое время полужизни вследствие быстрой метаболической деградации протеазами, подобными дипептидилпептидазе IV (DPP-IV) и нейтральной эндопептидазе 24.11 (NEP 24.11). Это ограничивает его использование в качестве терапевтического средства. Существуют агонисты GLP (такие как ингибиторы DPP-IV), которые, как считают, стабилизируют GLP-1. Предполагаемым недостатком ингибиторов DPP-IV является то, что GLP1 просто стабилизируется, но не контролируется эндогенным потреблением пищи. Таким образом, существует потребность в альтернативных средствах для альтернативной и улучшенной терапии для лечения сахарного диабета или заболеваний и нарушений, ассоциированных с ним.
Авторы настоящего изобретения предположили, что введение ингибитора BMP in vivo может повышать секрецию GLP-1 в EEC и, таким образом, действует в качестве терапевтического средства для лечения сахарного диабета и ассоциированных заболеваний и нарушений. Авторы настоящего изобретения показали, что введение ингибитора BMP мышам повышало секрецию GLP-1 (см. пример 6). Повышение количества клеток GLP1 является благоприятным для терапии, т.к. для этих клеток все равно требуется потребление пищи для высвобождения их GLP1. Таким образом, пик GLP1 будет более высоким в то время, когда индивидууму необходимы повышенные уровни инсулина. Это также является особенно благоприятным для пациентов с небольшим количеством клеток GLP1.
Таким образом, изобретение относится к способу лечения сахарного диабета или заболевания или нарушения, ассоциированного с ним, где способ включает введение терапевтически эффективного количества ингибитора BMP нуждающемуся в этом индивидууму. Изобретение также относится к ингибитору BMP для применения в способе лечения сахарного диабета или заболевания или нарушения, ассоциированного с ним, где способ включает введение терапевтически эффективного количества ингибитора BMP нуждающемуся в этом индивидууму. Изобретение также относится к применению ингибитора BMP для производства лекарственного средства для лечения сахарного диабета или заболевания или нарушения, ассоциированного с ним, где способ включает введение терапевтически эффективного количества ингибитора BMP нуждающемуся в этом индивидууму. Примеры подходящих ингибиторов BMP известны в этой области и представлены в настоящем описании выше.
Термин "индивидуум" может относиться к человеку или любому не являющемуся человеком животному (такому как любое из мыши, крысы, кролика, собаки, кошки, крупного рогатого скота, свиньи, овцы, лошади или примата). В предпочтительных вариантах осуществления индивидуум представляет собой млекопитающего, более предпочтительно - человека. Индивидуум может являться "пациентом", что обозначает человека, представленного поставщику медицинских услуг для диагностики или лечения заболевания. Индивидуум может страдать заболеванием или нарушением или иметь предрасположенность к нему, но может проявлять или не проявлять симптомы заболевания или нарушения.
Термин "терапевтически эффективное количество" относится к количеству терапевтического средства, достаточному для лечения, диагностики, профилактики и/или задержки дебюта симптомов заболевания, нарушения и/или состояния при введении индивидууму, страдающему заболеванием, нарушением и/или состоянием или имеющему предрасположенность к нему. Специалисту в этой области следует понимать, что терапевтически эффективное количество, как правило, вводят с помощью схемы дозирования, включающей по меньшей мере одну однократную дозу.
В рамках изобретения термин "лечение" относится к любому способу, используемому для частичного или полного облегчения, снижения, ингибирования, профилактики, задержки дебюта, снижения тяжести и/или снижения частоты одного или нескольких симптомов или признаков конкретного заболевания, нарушения и/или состояния. Лечение можно проводить в отношении индивидуума, не проявляющего признаки заболевания и/или проявляющего лишь ранние признаки заболевания с целью снижения риска развития патологии, ассоциированной с заболеванием.
"Сахарный диабет" может являться диабетом типа I и диабетом типа II. Альтернативно, он может являться гестационным диабетом. Термин "сахарный диабет" также включает пациентов, нечувствительных к инсулину, но у которых все равно могут наблюдать преддиабет. "Ассоциированные заболевания и нарушения" включают, в качестве неограничивающих примеров, гипергликемию, ожирение, целиакию, заболевание щитовидной железы, синдром поликистоза яичников, несахарный диабет, липоидный некробиоз, мастопатию, мышечные нарушения и заболевания зубов.
Например, изобретение относится к следующим пронумерованным вариантам осуществления.
1. Ингибитор BMP для применения в способе лечения или профилактики сахарного диабета или ассоциированного заболевания или нарушения, где способ включает введение терапевтически эффективного количества ингибитора BMP нуждающемуся в этом индивидууму.
2. Ингибитор BMP для применения в способе лечения или профилактики сахарного диабета или ассоциированного заболевания или нарушения посредством повышения секреции GLP-1 аргентофильными клетками (для повышения уровней инсулина и, таким образом, снижения уровней глюкозы в плазме), где способ включает введение терапевтически эффективного количества ингибитора BMP нуждающемуся в этом индивидууму.
3. Ингибитор BMP для применения по варианту осуществления 1 или варианту осуществления 2, где ингибитор BMP может
a. нарушать взаимодействие BMP с рецептором BMP;
b. связываться с рецептором BMP и ингибировать активацию нисходящей передачи сигнала;
c. ингибировать фосфорилирование Smad 1, Smad 5 или Smad 8;
d. ингибировать транслокацию Smad 1, Smad 5 или Smad 8 в ядро;
e. ингибировать опосредованную SMAD 1, SMAD 5 или SMAD 8 транскрипцию генов-мишеней; или
f. ингибировать экспрессию, фолдинг или секрецию BMP.
4. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где ингибитор BMP ингибирует фосфорилирование Smad1, Smad5 или Smad8 и является замещенным производным пиразоло[1,5-a]пиримидина, например, формулы I:
 Формула I
Формула I
где
X и Y независимо выбраны из CR15 и N;
Z выбран из CR3 и N;
Ar выбран из замещенного или незамещенного арила и гетероарила;
L1 отсутствует или выбран из замещенного или незамещенного алкила и гетероалкила;
A и B в каждом случае независимо выбраны из CR16 и N;
E и F являются CR5, и оба случая включения R5 вместе с E и F приводят к замещенному или незамещенному 5- или 6-членному циклоалкильному, гетероциклоалкильному, арильному или гетероарильному кольцу;
R3 выбран из H и замещенного или незамещенного алкила, циклоалкила, галогена, ациламиногруппы, карбамата, цианогруппы, сульфонила, сульфоксидогруппы, сульфамоила или сульфонамидогруппы;
R4 выбран из H и замещенного или незамещенного алкенила, алкинила, циклоалкила, гетероциклила, арила, гетероарила, ацила, карбоксила, сложного эфира, гидроксила, алкоксила, алкилтиогруппы, ацилоксигруппы, аминогруппы, ациламиногруппы, карбамата, амидогруппы, амидиногруппы, сульфонила, сульфоксидогруппы, сульфамоила или сульфонамидогруппы;
R15 в каждом случае независимо выбран из H и замещенного или незамещенного алкила, циклоалкила, гетероциклила, циклоалкилалкила, гетероциклилалкила, галогена, ациламиногруппы, карбамата, цианогруппы, сульфонила, сульфоксидогруппы, сульфамоила или сульфонамидогруппы;
R16 в каждом случае независимо отсутствует или выбран из H и замещенного или незамещенного алкила, алкенила, алкинила, аралкила, циклоалкила, гетероциклила, арила, гетероарила, гетероаралкила, циклоалкилалкила, гетероциклилалкила, галогена, ацила, карбоксила, сложного эфира, гидроксила, алкоксила, алкилтиогруппы, ацилоксигруппы, аминогруппы, ациламиногруппы, карбамата, амидогруппы, амидиногруппы, цианогруппы, сульфонила, сульфоксидогруппы, сульфамоила или сульфонамидогруппы,
или его фармацевтически приемлемой соли или сложному эфиру.
5. Ингибитор BMP для применения по варианту осуществления 4, где:
a. каждый из A и B является CH;
b. каждый из E и F является CR5, и вместе с атомами, к которым R5 присоединены в обоих случаях, образуют 6-членное кольцо;
c. E и F вместе представляют собой группу:
где R40 отсутствует или представляет собой один из 1-4 заместителей, выбранных из замещенного или незамещеннго алкила, циклоалкила, галогена, ациламиногруппы, карбамата, цианогруппы, сульфонила, сульфоксидогруппы, сульфамоила или сульфонамидогруппы;
d. L1 имеет структуру
где
Q выбран из CR10R11, NR12, O, S, S(O) и SO2; и
R10 и R11 в каждом случае независимо выбраны из H и замещенного или незамещенного алкила, циклоалкила, гетероциклила, циклоалкилалкила, гетероциклилалкила, аминогруппы, ациламиногруппы, карбамата, амидогруппы, амидиногруппы, цианогруппы, сульфонила, сульфоксидогруппы, сульфамоила или сульфонамидогруппы;
R12 выбран из H и замещенного или незамещеннго алкила, циклоалкила, гетероциклила, гетероциклилалкила, аминогруппы, ациламиногруппы, карбамата, амидогруппы, амидиногруппы, сульфонила, сульфамоила или сульфонамидогруппы; и
n является целым числом от 0 до 4;
e. R4 выбран из:
где
W отсутствует или является C(R21)2, O или NR21;
R20 отсутствует или выбран из замещенного или незамещенного алкила, аралкила, циклоалкила, гетероциклила, арила, гетероарила, гетероаралкила, циклоалкилалкила, гетероциклилалкила, ацила, сульфонила, сульфоксидогруппы, сульфамоила и сульфонамидогруппы; и
R21 в каждом случае независимо выбран из H и замещенного или незамещенного алкила, аралкила, циклоалкила, гетероциклила, арила, гетероарила, гетероаралкила, циклоалкилалкила, гетероциклилалкила, ацила, сульфонила, сульфамоила или сульфонамидогруппы; и/или
f. Ar является 6-членным арильным или гетероарильным кольцом, необязательно, где L1 располагается в пара-положении Ar относительно бициклического ядра.
6. Ингибитор BMP для применения по вариантам осуществления 4 или 5, где терапевтически эффективное количество составляет
a. по меньшей мере 0,1 мг/кг, по меньшей мере 0,2 мг/кг, по меньшей мере 0,5 мг/кг, по меньшей мере 1,0 мг/кг, по меньшей мере 2 мг/кг, по меньшей мере 5 мг/кг, по меньшей мере 10 мг/кг, по меньшей мере 20 мг/кг, по меньшей мере 30 мг/кг или приблизительно 35 мг/кг;
b. от 0,1 мг/кг до 50 мг/кг, от 0,1 мг/кг до 30 мг/кг, от 0,1 мг/кг до 10 мг/кг, от 0,1 мг/кг до 1 мг/кг, от 1 мг/кг до 50 мг/кг, от 1 мг/кг до 30 мг/кг, от 1 мг/кг до 10 мг/кг; и/или
c. где терапевтически эффективное количество вводят один, два или три раза в сутки.
7. Ингибитор BMP для применения по любому из вариантов осуществления 1-3, где ингибитор BMP выбран из:
a. дорсоморфина, или LDN193189, или его аналога или варианта; и/или
b. Noggin, склеростина, хордина, CTGF, фоллистатина, гремлина, tsg, sog или его аналога или варианта.
8. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где ингибитор BMP находится в форме фармацевтически приемлемой соли.
9. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где индивидуум представляет собой млекопитающего, предпочтительно человека, кошку или собаку.
10. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где индивидуум является человеком.
11. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где ингибитор BMP вводят
a. перорально, местно или посредством инъекции, предпочтительно перорально, и/или
b. системно или местно.
12. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где ингибитор BMP вводят в комбинации с одним или несколькими дополнительными терапевтическими средствами против диабета, например, сульфонилкарбамидом, бигуанидом, метформином, ингибитором альфа-глюкозидазы, тиазолидиндионом, меглитинидом, ингибитором дипептидилпептидазы-4 или другим инкретиновым миметиком, аналогом амилина или глюкозурическим средством.
13. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где ингибитор BMP вводят в комбинации с агонистом рецептором GLP-1, например, выбранным из: эксенатида, лираглутида, таспоглутида, ликсисенатида.
14. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где ингибитор BMP вводят в комбинации с инсулином или его биологически активным аналогом.
15. Ингибитор BMP для применения по любому из вариантов осуществления 12-14, где комбинацию вводят в виде одной композиции или двух отдельных композиций.
16. Ингибитор BMP для применения по любому из вариантов осуществления 12-15, где комбинацию вводят одновременно или последовательно.
17. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где сахарный диабет является диабетом типа 1, диабетом типа 2, гестационным диабетом или нечувствительностью к инсулину, предпочтительно - диабетом типа 2.
18. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где индивидуум имеет аномально низкие уровни GLP-1.
19. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где способом лечат диабет посредством повышения количества аргентофильных клеток, экспрессирующих гормоны, характерные для крипт, где гормоны, характерные для крипт, включают GLP-1, нейрокинин A, субстанцию P и глюкагон.
20. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где способ приводит к повышению уровней гормона GLP-1 в кровотоке/кишечнике/поджелудочной железе по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% относительно уровня до введения ингибитора BMP тому же пациенту.
21. Ингибитор BMP для применения по любому из предшествующих вариантов осуществления, где способ приводит к уровню глюкозы в плазме натощак у индивидуума менее 10,0 ммоль/л, менее 9,0 ммоль/л, менее 8,0 ммоль/л, менее 7,0 ммоль/л, менее 6,9 ммоль/л, менее 6,8 ммоль/л, менее 6,7 ммоль/л, менее 6,5 ммоль/л, менее 6,4 ммоль/л, менее 6,3 ммоль/л, менее 6,2 ммоль/л, менее 6,1 ммоль/л или менее 6,0 ммоль/л.
Клеточная терапия
В объем изобретения также входят способы лечения человека или не являющегося человеком животного посредством клеточной терапии. В настоящем описании термин "животное" означает любых млекопитающих. Пациент может находиться на любой стадии развития, включая эмбриональную стадию. Например, пациент может быть взрослым, или терапия может предназначаться для педиатрического применения (например, новорожденных, детей или подростков). Такая клеточная терапия включает введение клеток или органоидов, полученных по изобретению, пациенту любым подходящим способом. В частности, такие способы лечения включают регенерацию поврежденной ткани. В рамках изобретения термин "введение" относится к общепризнанным формам введения, таким как внутривенное введение или инъекция, а также к введению посредством трансплантации, например, трансплантации посредством хирургического вмешательства, пересадки или трансплантации сконструированных популяций клеток, полученных из клеток или органоидов по настоящему изобретению. В случае клеток, системное введение индивидууму можно осуществлять, например, посредством инфузии в верхнюю брыжеечную артерию, чревную артерию, подключичную вену через грудной проток, инфузии в сердце через верхнюю полую вену или инфузии в брюшную полость с последующей миграцией клеток через поддиафрагмальные лимфатические сосуды, или непосредственно в участки кишечника посредством инфузии в артерии кишечника (например, в верхнюю или нижнюю брыжеечные артерии).
В некоторых вариантах осуществления вводят от 104 до 1013 клеток на 100 кг массы тела на инфузию. Предпочтительно, можно внутривенно инфузировать приблизительно от 1-5×104 до 1-5×107 клеток на 100 кг массы тела. Более предпочтительно, можно внутривенно инфузировать приблизительно от 1×104 до 10×106 клеток на 100 кг массы тела. В некоторых вариантах осуществления осуществляют одно введение клеток или органоидов. В других вариантах осуществления используют многократные введения. Многократные введения можно осуществлять по исходной схеме лечения, например, в течение 3-7 последовательных дней, а затем повторять в другое время.
Также органоид можно получать из одной клетки, экспрессирующей Lgr5, как представлено в настоящем описании. Эту одну клетку можно модифицировать посредством встраивания конструкции нуклеиновой кислоты, как определено в настоящем описании, например, для коррекции генетического дефекта или мутации. Также можно специфически устранять экспрессию, при желании, например, с использованием миРНК. Потенциальными полипептидами, подлежащими экспрессии, могут являться любые полипептиды, являющиеся недостаточными при метаболических заболеваниях, включая, например, недостаточность полипептида при метаболическом заболевании печени, такого как AAT (альфа-антитрипсин). Для выявления физиологической основы, авторы настоящего изобретения также могли экспрессировать или инактивировать гены, вовлеченные в путь Wnt, EGF, FGF, BMP или Notch.
Специалисту в этой области будет очевидно, что в способе, предназначенном для восстановления поврежденной или пораженной ткани, дополнительно можно использовать генную терапию. Например, можно использовать аденовирусное или ретровирусное средство доставки генов для доставки генетической информации, подобной ДНК и/или РНК, в стволовые клетки. Специалист в этой области может заменять или восстанавливать конкретные гены, на которые воздействуют при генной терапии. Например, нормальный ген можно встраивать в неспецифический участок генома для замены нефункционального гена. В другом примере аномальную последовательность гена можно заменять нормальной последовательностью гена посредством гомологичной рекомбинации. Альтернативно, с помощью избирательной обратной мутации можно возвращать ген к его нормальной функции. Дополнительным примером является изменение регуляции (степени "включения" и "выключения" гена) конкретного гена. Предпочтительно, стволовые клетки обрабатывают ex vivo с помощью подхода генной терапии, а затем переносят в организм млекопитающего, предпочтительно человека, нуждающегося в лечении. Например, полученные из органоида клетки можно генетически модифицировать в культуре перед трансплантацией пациентам.
Таким образом, в некоторых вариантах осуществления органоид или популяция эпителиальных стволовых клеток предназначены для применения в медицине, например, для лечения нарушения, состояния или заболевания и/или для применения в регенеративной медицине.
В одном из предпочтительных вариантов осуществления, например, если органоид предназначен для применения в регенеративной медицине, способ можно начинать с эпителиальных клеток или фрагмента ткани, где клетки или фрагмент ткани являются аутологичными или аллогенными. Некоторая степень совпадения с пациентом все равно может потребоваться для предотвращения проблемы отторжения. Способы минимизации отторжения ткани будут известны специалистам в этой области.
В вариантах осуществления, в которых органоиды и/или клетки трансплантируют пациенту, предпочтительным может являться введение клеток в каркас. Таким образом, изобретение относится к каркасу, содержащему один или несколько органоидов по изобретению или клеток, полученных из указанных органоидов. Каркас представляет собой двумерную или трехмерную сеть. Подходящие синтетические материалы для такого каркаса содержат полимеры, выбранные из пористых твердых веществ, нановолокон и гидрогелей, таких как, например, пептиды, включая самособирающиеся пептиды, гидрогели, состоящие из фосфата полиэтиленгликоля, фумарата полиэтиленгликоля, полиакриламида, полигидроксиэтилметакрилата, ацетата полицеллюлозы и/или их сополимеров (см., например, Saha et al. (2007) Curr Opin Chem Biol. 11(4): 381-387; Saha et al. (2008) Biophysical Journal 95:4426-4438; Little et al. (2008) Chem. Rev 108:1787-1796). Как известно специалисту в этой области, механические свойства, такие как, например, эластичность каркаса, влияют на пролиферацию, дифференцировку и миграцию стволовых клеток. Предпочтительный каркас содержит биодеградируемые сополимеры, заменяемые природными компонентами после трансплантации индивидууму, например, для стимуляции регенерации ткани и/или заживления ран. Кроме того, предпочтительно, чтобы указанный каркас, по существу, не индуцировал иммуногенный ответ после трансплантации индивидууму. Указанный каркас дополняют природными, полусинтетическими или синтетическими лигандами, обеспечивающими сигналы, необходимые для пролиферации, и/или дифференцировки, и/или миграции стволовых клеток. В предпочтительном варианте осуществления указанные лиганды содержат определенные аминокислотные фрагменты. Примеры указанных синтетических полимеров включают поверхностно-активное вещество блок-сополимер Pluronic® F127 (BASF) и Ethisorb® (Johnson и Johnson). В некоторых вариантах осуществления клетки культивируют в каркасе. В других вариантах осуществления их культивируют, а затем добавляют в каркас.
При применении по настоящему изобретению можно использовать один органоид или несколько органоидов, например, 2, 3, 4, 5, 10, 15, 20, 30, 50, 100, 200 или более органоидов. Предпочтительно, способы по настоящему изобретению позволяют получать значительное количество органоидов и эпителиальных стволовых клеток за короткий период времени, т.к. это приводит к экспоненциальному росту, таким образом, обеспечивая доступность достаточного количества клеток для интересующего применения. Когда в настоящем описании упоминают "способ лечения", например, включающий органоиды или клетки, полученные из органоидов по изобретению, это также в равной степени относится к органоидам или клеткам "для применения в лечении" и органоидам или клеткам "для применения в производстве лекарственного средства".
Искусственные органы
В некоторых вариантах осуществления изобретение относится к применению дифференцированного органоида или клеток, полученных из дифференцированного органоида, в искусственном органе. Искусственный орган можно трансплантировать in vivo способами, описанными где-либо. Альтернативно, искусственный орган может находиться ex vivo. В некоторых вариантах осуществления искусственный орган ex vivo можно соединять с пациентом, например, через кровеносную сеть. Например, искусственный орган, содержащий дифференцированный органоид, можно использовать в качестве части аппарата для диализа. Таким образом, дифференцированный органоид можно использовать для поддержки пациента с пораженной или поврежденной эпителиальной тканью.
Применение органоидов и популяций клеток кишечника
Как представлено в примерах, среды и способы дифференцировки по изобретению повышают дифференцировку клеток-предшественников, полученных из желудка, в направлении аргентофильных клеток.
Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду кишечника или клетке, полученной из органоида кишечника для применения в медицине, например, для применения в лечении желудочного нарушения, состояния или заболевания или для применения в регенеративной медицине.
Изобретение дополнительно относится к органоиду кишечника или клетке, полученной из органоида кишечника, для применения в лечении диабета (такого как диабет типа I или типа II), кистозного фиброза и воспалительного заболевания кишечника (такого как болезнь Крона), при этом лечение, необязательно, включает трансплантацию органоида или клеток, полученных из органоида желудка нуждающемуся в этом пациенту.
Применение органоидов и популяции клеток желудка
Способы культивирования клеток желудка описаны в WO 2010/090513. Предполагают, что среды и способы дифференцировки по изобретению будут повышать дифференцировку клеток-предшественников, полученных из желудка, в направлении аргентофильных клеток.
Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду желудка или клетке, полученной из органоида желудка, для применения в медицине, например, для применения в лечении желудочного нарушения, состояния или заболевания или для применения в регенеративной медицине.
Изобретение дополнительно относится к органоиду желудка или клетке, полученной из органоидов желудка, для применения в лечении гастрита, атрофического гастрита, пилоростеноза, рака желудка или пептической язвы, при этом лечение, необязательно, включает трансплантацию органоида или клеток, полученных из органоида желудка, нуждающемуся в этом пациенту.
Применение органоидов и популяций клеток поджелудочной железы
Способы культивирования клеток поджелудочной железы описаны в WO 2010/090513. Предполагают, что среды и способы дифференцировки по изобретению будут повышать дифференцировку эпителиальных стволовых клеток, полученных из поджелудочной железы, в направлении аргентофильных клеток.
Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду поджелудочной железы или клетке, полученной из органоида поджелудочной железы, для применения в медицине, например, для применения в лечении нарушения, состояния или заболевания поджелудочной железы или для применения в регенеративной медицине.
Изобретение дополнительно относится к органоиду поджелудочной железы или клетке, полученной из органоидов поджелудочной железы, для применения в лечении диабета (например, диабета типа I или типа II), панкреатита, рака поджелудочной железы или кистозного фиброза, при этом лечение, необязательно, включает трансплантацию органоида или клеток, полученных из органоида поджелудочной железы, нуждающемуся в этом пациенту. В некоторых вариантах осуществления трансплантируемые клетки являются инсулин-секретирующими клетками. В других вариантах осуществления клетки являются клетками-предшественниками, созревающими после трансплантации в инсулин-секретирующие клетки.
Применение органоидов и популяций клеток легких
Способы культивирования клеток легких описаны в WO 2016/083613. Предполагают, что среды и способы дифференцировки по изобретению будут повышать дифференцировку эпителиальных стволовых клеток, полученных из легкого, в направлении аргентофильных клеток.
Таким образом, в некоторых вариантах осуществления изобретение относится к органоиду легкого или клетке, полученной из органоида легкого, для применения в медицине, например, для применения в лечения нарушения, состояния или заболевания легких или для применения в регенеративной медицине.
Изобретение дополнительно относится к органоиду легкого или клетке, полученной из органоидов легкого, для применения в лечении мелкоклеточного рака легких или немелкоклеточного рака легких (например, аденокарциномы, плоскоклеточной карциномы или крупноклеточной карциномы), интерстициальной болезни легких, пневмонии (например, организующейся пневмонии), туберкулеза, кистозного фиброза, бронхита, легочного фиброза, саркоидоза, гиперплазии типа II, хронической обструктивной болезни легких, эмфиземы, астмы, отека легких, острого респираторного дистресс-синдрома, одышки, бронхоэктазии, хантавирусного легочного синдрома, ближневосточного респираторного синдрома (MERS), тяжелого острого респираторного синдрома (SARS) или пневмокониоза. Изобретение дополнительно относится к органоиду легкого или клетке, полученной из органоидов легкого, для применения в лечении патогенного заболевания, вызванного патогеном, таким как аденовирус, коронавирус (например, SARS-CoV или MERS-CoV), метапневмовирус человека, вирус гриппа, вирус парагриппа, респираторно-синцитиальный вирус, риновирус, хантавирус, энтеровирус (например, энтеровирус D68 (EV-D68)), Bordetella pertussis, Chlamydophila pneumoniae, Corynebacterium diphtheria, Coxiella burnetii, Haemophilus influenzae, Legionella pneumophila, Moraxella catarrhalis, Mycobacterium tuberculosis, Mycoplasma pneumoniae, Staphylococcus aureus, Streptococcus pneumoniae или Streptococcus pyogenes.
Покоящиеся Lgr5+ стволовые клетки
Покоящиеся Lgr5+ стволовые клетки существуют in vivo, но ранее их не получали in vitro. Авторы настоящего изобретения неожиданно обнаружили, что ингибирование пути EGFR может индуцировать состояние покоя Lgr5+ стволовых клеток in vitro.
Таким образом, изобретение относится к способу индукции состояния покоя у Lgr5+ стволовой клетки, включающему:
обработку клетки одним или несколькими ингибиторами пути EGFR.
В некоторых вариантах осуществления способ является способом in vitro.
Изобретение дополнительно относится к популяции покоящихся стволовых клеток, полученных способом индукции состояние покоя Lgr5+ стволовых клеток-предшественников по изобретению, где клетки экспрессируют Lgr5 и Lef1 и не экспрессируют KI67 и маркер M-фазы фосфогистон H4.
Изобретение дополнительно относится к культуре in vitro, содержащей популяцию покоящихся стволовых клеток по изобретению.
В некоторых вариантах осуществления состояние покоя поддерживают в течение по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9 или по меньшей мере 10 дней. Таким образом, в некоторых вариантах осуществления состояние покоя поддерживают в течение по меньшей мере 7-10 дней.
Популяция покоящихся стволовых клеток по изобретению обладает множеством преимуществ и применений. Например, популяцию покоящихся стволовых клеток можно хранить (например, в морозильной камере), и она будет повторно расти более эффективно, чем популяция непокоящихся стволовых клеток. В некоторых вариантах осуществления популяцию покоящихся стволовых клеток по изобретению используют для исследования клеточного цикла стволовых клеток или для идентификации молекул, индуцирующих вход стволовых клеток в клеточный цикл.
Сокращения
β-TrCP: белок, содержащий β-трансдуциновый повтор
BME: Экстракт базальной мембраны
Cck: Холецистокинин
CHGA: Хромогранин A
DAPI: 4',6-диамидино-2-фенилиндол
EdU: 5-этинил-2'-дезоксиуридин
EEC: Аргентофильная клетка
GIP: Желудочный ингибиторный белок
GLP-1: Глюкагон-подобный белок 1
GSK-3: Киназа гликогенсинтазы 3
IDMI: Ингибитор IWP2, DAPT и MEK
LGR: Богатый лейциновыми повторами, сопряженный с G-белком рецептор
LRP: Белок, родственный рецептору липопротеинов низкой плотности
Nts: Нейротензин
PP1: Протеинфосфатаза 1
PP2A: Протеинфосфатаза 2A
PP2C: Протеинфосфатаза 2C
Sct: Секретин
Sst: Соматостатин.
Определения
В рамках изобретения глагол "содержать" и его спряжения используют в неограничивающем смысле для обозначения включения объектов после этого слова, но объекты, не упомянутые конкретно, не исключены. Кроме того, глагол "состоять" можно заменять, при необходимости, термином "состоит, по существу, из", означающим, что продукт, как определено в настоящем описании, может содержать дополнительные компоненты, иные, чем конкретно указанные компоненты, указанные дополнительные компоненты не изменяют уникальные характеристики изобретения. Кроме того, способ, как определено в настоящем описании, может включать дополнительные стадии, иные, чем конкретно указанные стадии, указанные дополнительные стадии не изменяют уникальные характеристики изобретения. Кроме того, упоминание элемента в единственном числе не исключает вероятности наличия нескольких элементов, если контекст четко не указывает на наличие только одного из элементов. Таким образом, термин в единственном числе, как правило, означает "по меньшей мере один".
В рамках изобретения термин "приблизительно" означает, что присутствующее значение может варьироваться на ±10%. Значение также можно читать как точное значение и, таким образом, опускать термин "приблизительно". Например, термин "приблизительно 100" включает 90-110, а также 100.
Все патентные и другие ссылки, процитированные в настоящем описании, включены, таким образом, в настоящее описание в качестве ссылок в полном объеме.
Следующие примеры предложены только в иллюстративных целях и не предназначены для ограничения объема настоящего изобретения каким-либо образом.
Термин "кишечник" включает толстый и тонкий кишечник.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. Ингибирование EGFR индуцирует выход из клеточного цикла в органоидах кишечника. (A) Схема эксперимента. Органоиды обрабатывали EGFR (или ингибиторами MEK/ERK) или DMSO через неделю после помещения в BME. Образцы собирали через 1 (d1), 2(d2), 4 (d4) или 7 (d7) день после обработки. Действия повторяли со стадии обработки для экспериментов по пересеву. (B) Органоиды кишечника через 4 дня в контроле (ENR) или при ингибировании EGFR (EGFRi). Флуоресценция Lgr5GFPiresCreER повышается после обработки EGFRi. Канал RFP используют для отображения фона. На нижних панелях представлены светлопольные изображения. (C) Анализ клеточного цикла в органоидах кишечника. EdU вводили за 1 ч. до окончания эксперимента. Контрольные (ENR) органоиды непрерывно включают EdU (верхние панели) и экспрессируют KI67 (нижние панели), в то время как обработанные EGFRi органоиды выходят из клеточного цикла с течением времени. (D) Количественный анализ C. (E) Анализ HOECHST содержания ДНК в контрольных или обработанных EGFRi органоидах. Левый пик соответствует обработанным EGFRi органоидам, а правый пик соответствует контрольным органоидам в среде ENR. Столбики справа соответствуют количественному анализу для 3 независимых экспериментов. Верхние полосы на столбиках соответствуют клеткам в фазе G2/M, средние полосы на столбиках соответствуют клеткам в фазе S, и нижние полосы на столбиках соответствуют клеткам в фазе G0/G1. (F) Анализ клеточного цикла органоидов кишечника после повторного введения EGF в среду для культивирования. (G)Lgr5GFPiresCreER+ клетки выходят из клеточного цикла через 4 дня после обработки EGFRi. Окрашивание на фосфогистон H3 (pH3) используют для визуализации фазы M. На графике внизу представлен количественный анализ. DAPI используют для визуализации ядер. Масштаб = 50 мкм.
Фигура 2. Индуцируемый передачей сигнала EGFR выход из клеточного цикла опосредован путем передачи сигнала MAPK. (A) Гистологический анализ фосфорилирования ERK после ингибирования EGFR. Быстрая утрата pERK является постепенной в течение 24 ч., но остается низкой через 48 часов (верхние панели). Этот период совпадает с выходом органоидов из клеточного цикла (окрашивание KI67, нижние панели). (B) Отдельное ингибирование Mek или Erk, а также одновременное ингибирование EGFR и ErbB-2 с использованием афатиниба приводит к результатам, аналогичным гефитиниб-индуцируемому ингибированию EGFR. EdU добавляют в среду для культивирования за 1 час до окончания эксперимента. На средних панелях представлена экспрессия эндогенного GFP с аллеля Lgr5GFPiresCreER. На нижних панелях представлены светлопольные изображения. DAPI используют для визуализации ядер. Масштаб = 50 мкм.
Фигура 3. Отслеживание клеточной линии свидетельствует о том, что qLgr5+ клетки являются стволовыми клетками.(A) Схема эксперимента. Органоиды повторно высевали в BME в ENR (контроль) или среду с EGFRi через 7 дней после диссоциации. Через 4 дня после обработки индуцировали рекомбинацию с использованием 4'OH-тамоксифена (T) в течение 16 часов и восстанавливали передачу сигнала EGF. Органоиды собирали через 4 или 12 дней после индукции тамоксифеном или повторно высевали в ENR на 2 раунда. (B) Рекомбинированные (YFP+) клетки образовывали CHGA+ EEC, LYZ+ клетки Панета (левые панели) или пучковые клетки, идентифицированные по апикальному актину и плотным пучкам ацетилированного тубулина (правые панели). Отслеживали активные (верхние панели) и покоящиеся (нижние панели) Lgr5+ клетки. (C) Количественный анализ B. Нижняя полоса на каждом столбике соответствует CBC, полоса непосредственно над нижней полосой соответствует EEC, следующая полоса выше соответствует клеткам Панета, следующая полоса выше соответствует пучковым клеткам, и верхняя полоса на каждом столбике соответствует покоящимся клеткам. (D) Рекомбинированные (идентифицированные посредством окрашивания X-Gal) активные (левая верхняя панель) и покоящиеся (левая нижняя панель) Lgr5+ клетки образовывали целые органоиды, демонстрирующие мультипотентность. Рекомбинированные Dclk1+ клетки (правые панели) не росли в каких-либо условиях. Масштаб = 50 мкм.
Фигура 4. Посредством секвенирования РНК идентифицируют ключевые молекулярные различия между qLgr5+ и aLgr5+ стволовыми клетками. (A) Иерархическая кластеризация целого транскрипта из сортированных Lgr5+ клеток с использованием Lgr5GFPDTR (Lgr5DTR), Lgr5GFPiresCreER (Lgr5GFP) клеток и пучковых клеток с использованием аллеля Dclk1GFPiresCreER(Dclk1) в контрольных (ENR) условиях или условиях ингибирования EGFR (EGFRi). Контрольные органоиды добавляли в качестве референса. (B) Анализ основных компонентов (PCA). (C) Вулканная диаграмма, на которой представлено сравнение активных и покоящихся Lgr5 клеток. На оси X показано скорректированное значение p (значение q, -log10), и на оси y представлено кратное изменение (log2). Серыми точками показаны гены с уровнем ложноположительных результатов (FDR) менее 0,01, черными точками показаны гены, не изменившиеся значимо. (D) На тепловой карте показаны гены, дифференциально регулируемые посредством ингибирования EGFR в Lgr5+ клетках. Цветом показаны значения z для каждого ряда (гена). (E) На коробчатых диаграммах представлены нормализованные значения экспрессии маркерных генов. (F) На тепловой карте представлена кластеризация способом k-средних коэффициента корреляции Пирсона полного транскриптома отдельных активных и покоящихся Lgr5+ клеток.
Фигура 5. Получение культуры EEC высокой чистоты.(A) Анализ маркеров EEC (CHGA) и клеток Панета (LYZ). Органоиды в течение 4 дней обрабатывали ингибитором Notch DAPT (D), ингибитором секреции Wnt IWP-2 (I), гефитинибом (EGFRi) или их комбинацией. DMSO используют в качестве контроля. (B) Ингибирование передачи сигнала Mek (Meki) вместе с путями передачи сигнала Wnt и Notch схожим образом повышает количество клеток EEC. Темно-серыми и светло-серыми V-образными стрелками на левой половине фигуре указаны области высокой экспрессии SCT и GIP, соответственно. Темно-серыми и светло-серыми V-образными стрелками на левой половине фигуры указаны области высокой экспрессии CCK и SST, соответственно. Значительно повышали количества клеток, положительных по желудочному ингибиторному белку (GIP), секретину (SCT), соматостатину (SST) и холецистокинину (CCK). (C) Анализ qPCR экспрессии связанных с EEC маркерных генов в органоидах. EI: EGFRi и MI: Meki. Масштаб = 50 мкм. Планками погрешностей указано стандартное отклонение.
Фигура 6. При профилировании транскриптома отдельных клеток выявили гетерогенность среди индуцированных EEC. (A) На тепловой карте показана кластеризация способом k-средних коэффициента корреляции Пирсона полного транскриптома отдельных живых клеток органоида из экспериментов с использованием Meki и EGFRi. Числами указаны кластеры. Цветами закодирован коэффициент корреляции Пирсона полного клеточного транскриптома. (B) На карте t-SNE показаны отдельные клетки и маркерные гены, экспрессируемые группой клеток. (C) На картах t-SNE показаны log2-трансформированные количества транскриптов с цветовой кодировкой для соответствующих генов. (D) На тепловой карте показаны количества транскриптов с цветовой кодировкой для соответствующих генов в кластерах 2, 5, 6 и 7.
Фигура 7. Роль нишевых путей передачи в пролиферации Lgr5+ клеток. (A) На графиках FACS показана эндогенная флуоресценция Lgr5GFPDTR (верхние панели, ось x) и иммуноокрашивание KI67efluor660 (нижние панели, ось x). Канал PL3 используют (ось y) для различения фона. Гейтами показаны положительные клетки относительно контролей дикого типа. (B) Светлопольные (верхние) и флуоресцентные (нижние) изображения контрольных (ENR) и EGFR-ингибированных (EGFRi) органоидов с использованием аллелей Lgr5GFPiresCreER и Rosa-TOPCFP. Канал RFP используют для различения фона. (C) Анализ маркерных генов Lgr5GFPiresCreER+ клеток через 4 дня после обработки EGFRi. Темно-серыми и светло-серыми V-образными стрелками на левом изображении указаны области высокой экспрессии CHGA и LYZ, соответственно. На среднем и правом изображениях светло-серыми V-образными стрелками указаны области высокой экспрессии фаллоидина. (D) Анализ qPCR экспрессии маркерных генов в органоидах, обработанных EGFRi в течение 4 дней. (E) Относительное количество клеток, положительных по маркерному белку, после повторной индукции передачи сигнала EGF в течение 1 (d1), 3 (d3) или 5 (d5) дней по сравнению с соответствующими контролями (обработанными DMSO, столько же дней в культуре). (F) Вход органоидов в клеточный цикл после введения EGF после повторных циклов удаления EGF (NR) или обработки EGFRi (2xNR+EGFRi). (G) Количественный анализ F. Масштаб = 50 мкм. Планками погрешностей указано стандартное отклонение. Ключ: ENR=EGF, Noggin и R-спондин 1; -R = удаление R-спондина 1; -N = удаление Noggin; EGFRi = ингибирование передачи сигнала EGFR (с использованием гефитиниба с сопутствующим удалением EGF из среды для культивирования).
Фигура 8. Клеточная композиция органоидов после обработки EGFRi. (A) Окрашивание альциановым синим, на PAS и муцин 2 (MUC2) с использование срезов контрольных (ENR) или обработанных EGFRi органоидов в парафине. (B) Трехмерная реконструкция целых органоидов, окрашенных на маркеры клеток Панета (LYZ) и EEC (CHGA). Оба типа клеток являлись более концентрированными в обработанных EGFRi органоидах (нижняя панель) по сравнению с контролями. На графиках справа представлен количественный анализ. (C) Количества пучковых клеток, визуализированных с использованием эндогенной флуоресценции аллеля Dclk1GFPiresCreER, повышались после обработки EGFRi (правая панель) по сравнению с контролями (левая панель). Флуоресценция GFP не перекрывалась с маркерами EEC (CHGA) или клеток Панета (LYZ). Масштаб = 50 мкм.
Фигура 9. Термины генной онтологии (GO) и анализ отдельных клеток на сигнатуры Lgr5+клеток. (A) Анализ терминов GO (с использованием Revigo) генов, отрицательно регулируемых после обработки EGFRi в клетках Lgr5. Термины, связанные с клеточным циклом и делением, а также биосинтезом низкомолекулярных соединений (все из них связаны с прохождением клеточного цикла), были значимо выше в активных клетках по сравнению с покоящимися Lgr5+ клетками. На оси x указано значение p (-log10), на оси y указан размер выборки терминов GO. (B) На карте t-SNE показано распределение секвенированных Lgr5+ клеток. (C) На картах t-SNE показаны log2-трансформированные уровни транскриптов с цветной кодировкой для мРНК Lyz и Dclk1, используемых для идентификации клеток Панета и пучковых клеток, соответственно.
Фигура 10. Исходный анализ секвенирования отдельных клеток индуцированных органоидов EEC. (A) На карте t-SNE показано распределение кластеров, идентифицированных посредством RaceID. (B) На карте t-SNE представлено сравнение распределения клеток, полученных в экспериментах с использованием IDEGFRi и IDMeki. (C) На картах t-SNE показаны log2-трансформированные уровни транскриптов с цветной кодировкой для мРНК Apoa1, используемой для идентификации энтероцитов.
Судьба клеток-предшественников кишечника. Клетка-предшественник кишечника может дифференцироваться в ряд типов клеток, таких как энтероцит, бокаловидная клетка, клетка Панета или аргентофильная клетка. Известно, что активация Notch, сопряженная с ингибированием Wnt, способствует дифференцировке энтероцитов. Также известно, что ингибирование Notch, сопряженное с активацией Wnt, может способствовать дифференцировке клеток Панета. Кроме того, известно, что ингибирование Wnt и Notch может способствовать дифференцировке бокаловидных клеток. Однако отсутствует понимание того, как повысить дифференцировку аргентофильных клеток.
Фигура 11. Композиция культуры EEC. Обработка дифференцировочной средой IDMI приводила к дифференцировке, в основном, смещенной в сторону энтерохромаффинных клеток. Абсолютное повышение EEC на органоид, в основном, соответствует энтерохромаффинным клеткам. GLP1-, CCK-, NTS-, STT- и GIP-продуцирующие клетки присутствуют в меньшей степени.
Фигура 12. Результаты qPCR, где представлены уровни матричной РНК при различных способах дифференцировки EEC. На диаграммах представлена кратная экспрессия мРНК различных маркеров EEC относительно контроля. Активация пути BMP селективно повышает матричную РНК секретина за счет TAC1 (маркера энтерохромаффинных клеток). BMP4 использовали для активации пути BMP, и он присутствовал в концентрации 10 мкг/мл. Основные условия: IDMI (IWP2, DAPT и ингибитор MEK). MHY1485 является активатором mTOR, тестируемым в концентрации 5 мкМ. Висмогениб является ингибитором Hedgehog (в особенности, ингибитором Smoothened), тестируемым в концентрации 5 мкМ.
Фигура 13. Окрашивание на секретин и GIP при различных способах дифференцировки аргентофильных клеток. Активация пути BMP в значительной степени повышает количество S-клеток.
Фигура 14. Окрашивание на серотонин при различных способах дифференцировки аргентофильных клеток. Активация пути BMP снижает количество энтерохромаффинных клеток.
Фигура 15. Результаты qPCR, в которых представлено log2-изменение уровней матричной РНК различных маркеров EEC в способе дифференцировки EEC по сравнению со стандартной дифференцировочной средой. Дифференцировочная среда EEC значительно повышала количество EEC в органоидах человека, но не клеток Панета.
Фигура 16. Экспрессия CHGA при различных способах дифференцировки аргентофильных клеток.
Фигура 17. Тройное ингибирование Notch, Wnt и MEK приводит к повышению культур, обогащенных EEC. Получали 30-80% CHGA+ клеток на органоид.
Фигура 18. Экспрессия гормонов в культуре EEC. Все различные подтипы EEC присутствуют в культурах IDMI и экспрессируют один гормон. Некоторые клетки являются CHGA-, но положительными на гормон.
Фигура 19. Секреция гормонов EEC в культуре. 2-дневную среду для культивирования собирали для получения результатов анализа на -форсколин. Затем культуры индуцировали с использованием форсколина в течение 1 ч. и собирали среду для получения результатов анализа на +форсколин.
Фигура 20. Дифференциальная локализация Tac1-, GLP1- и секретин-положительных иммунореактивных клеток в крипте кишечника и ворсинке A-C). Двойные PYY-GLP1-положительные клетки локализовались в крипте, в то время как L-клетки в ворсинке утрачивали экспрессию GLP1 D-E). Энтерохромаффинные клетки в крипте, экспрессирующие серотонин, коэкспрессируют Tac1. Экспрессия Tac1 в ворсинке утрачена, при этом энтерохромаффинные клетки начинают коэкспрессировать секретин.
Фигура 21. Передача сигнала BMP является движущей силой в случае гормональной сигнатуры ворсинки. A) Среда для дифференцировки аргентофильных клеток (ингибирование Wnt, Notch и MAPK), в которой передача сигнала BMP ингибируется в присутствие Noggin, генерирует гормональную сигнатуру, напоминающую таковую в крипте: секретин отсутствует, и серотонин+ EEC всегда коэкспрессируют Tac1. Исключение Noggin и введение BMP4 в эту дифференцировочную смесь, определенную как EEC с высоким BMP, значительно снижает количество GLP1, а также Tac1. В этой среде появляются отдельные серотонин+ аргентофильные клетки, а также секретин+ клетки. B) Количественный анализ A) C) Светлопольное изображение небольших органоидов кишечника в дифференцировочной среде EEC с низким или высоким BMP. Репортер GCG-Venus использовали для отслеживания GLP1-положительных клеток. Хотя при разных способах дифференцировки получают морфологически неразличимые органоиды, экспрессия GLP1 значительно снижается на фоне активации BMP.
Фигура 22. Ингибирование передачи сигнала BMP in vivo вызывает экспансию GLP1+ компартмента и супрессирует экспрессию секретина. A) 60-часовая обработка LDN193189 мышей через желудочный зонд вызывает повышение количества GLP1. GLP1 широко экспрессируется L-клетками в ворсинке, не всегда коэкспрессирующей PYY. B) Секретин значительно снижен в ворсинке мышей с ингибированием BMPR по сравнению с контролем.
Примеры
Материалы и способы
Культивирование органоидов
Основную среду для культивирования (продвинутую модифицированную по способу Дульбекко среду Игла/F12, дополненную пенициллином/стрептомицином, 10 мМ HEPES, глютамаксом, B27 [Life Technologies, Carlsbad, CA] и 1 мМ N-ацетилцистеином [Sigma]) дополняли 50 нг/мл рекомбинантного эпидермального фактора роста мыши (EGF; Peprotech, Hamburg, Germany), R-спондином 1 (кондиционированная среда, 5% конечного объема) и Noggin (кондиционированная среда, 5% конечного объема), дав ей название среда "ENR". Кондиционированные среды получали с использованием клеток HEK293T, стабильно трансфицированных с использованием Rspo1-Fc HA мыши (предоставленного Calvin Kuo, Stanford University), или клеток HEK293T после транзиторной трансфекции с использованием экспрессирующего вектора Noggin-Fc мыши. Продвинутую модифицированную по способу Дульбекко среду Игла/F12, дополненную пенициллином/стрептомицином и глютамакс, кондиционировали в течение 1 недели. Клетки высевали в BME (Trevigen). Для ингибирования передачи сигнала EGF клетки обрабатывали гефитинибом (5 мкМ; Santa Cruz Biotechnology) и удаляли EGF из среды. Секрецию Wnt ингибировали с использованием IWP-2 (1,5 мкМ; Sigma Aldrich) и секрецию Notch ингибировали с использованием DAPT (1 мМ, Sigma Aldrich). Все обработки осуществляли в день 5 после пассажа органоидов. Для экспериментов по повторной активации EGFR органоиды повторно высевали в свежий BME и среду ENR для обеспечения вымывания ингибитора EGFR. Для индукции активности Cre-ERT органоиды обрабатывали O/N 4-гидрокситамоксифеном (1 мкМ). Все контрольные органоиды обрабатывали схожими концентрациями соединения-растворителя, диметилсульфоксида (DMSO). При обработке клетки визуализировали с использованием микроскопа EVOS (Electron Microscopy Sciences).
Для индукции аргентофильной дифференцировки клетки культивировали в ENR или ENR с вальпроевой кислотой и CHIR99021 (Yin et al. (2014) Nature methods 11:106-112). Через 5 дней культивирования среду удаляли и промывали органоиды PBS. Смесь для дифференцировки EEC включала: IWP2 (1,5 мкМ; Sigma Aldrich), DAPT (1 мМ, Sigma Aldrich) и ингибитор MEK PD0325901 (5 мкМ; Sigma Aldrich).
Иммуноокрашивание
Целые органоиды собирали посредством осторожного растворения матригеля в ледяном PBS, а затем фиксировали в течение ночи при 4°C в 4% параформальдегиде. Затем органоиды пермеабилизовали и блокировали в PBS, содержащем 0,5% Triton X-100 и 2% нормальной сыворотки отца (Jackson ImunoResearch) в течение 30 минут при комнатной температуре. Органоиды инкубировали в течение 2 часов при комнатной температуре в блокирующем буфере, содержащем первичные антитела. Используемыми первичными антителами являлись антитело кролика против лизоцима (1:500; DAKO), антитело козы против хромогранина A (1:500; Santa Cruz), антитело мыши против Ki67 (1:250; BD Pharmingen), антитело кролика против фосфогистона 3 (pH3 Ser10, 1:1000; Millipore), антитело мыши против цитокератина 20 (1:1000; Dako), антитело козы против холецистокинина (sc-21617,1:100; Santa Cruz), антитело кролика против нейротензина (sc-20806, 1:100; Santa Cruz), антитело козы против секретина (sc-26630,1:100; Santa Cruz), антитело козы против соматостатина (sc-7819, 1:100; Santa Cruz), антитело кролика против желудочного ингибиторного белка (ab22624-50, 1:500, Abcam), антитело кролика против глюкагон-подобного пептида 1 (ab22625, 1;500, Abcam) и антитело мыши против ацетилированного тубулина (1:100; Santa Cruz). Органоиды инкубировали с соответствующими вторичными конъюгированными с Alexa488, 568 и 647 антителами против кролика, против козы и против мыши (1:1000; Molecular Probes) в блокирующем буфере, содержащем DAPI (1;1000, Invitrogen), или с фаллоидином с техасским красным (1:1000; Life technologies). Включение EdU визуализировали с использованием набора Click-iT Assay Kit (Thermo Fisher) после 1-часовой предварительной инкубации с EdU (10 мкМ). Окрашивание LacZ осуществляли, как описано ранее (Barker et al. (2007) Nature 449(7165):1003-7). Срезы заключали в Vectashield (Vector Labs) и визуализировали с использованием конфокального микроскопа Sp5 и Sp8 (Leica). Анализ изображений осуществляли с использованием программного обеспечения ImageJ.
Сортировка FACS
Для анализа FACS экспрессии LGR5 и KI67 Lgr5GFPDTR органоиды сначала подвергали диссоциации на отдельные клетки посредством механического разрушения после 15 минут обработки трипсином при 37°C (TrypLE Express; Life Technologies, Carlsbad, CA). Отдельные клетки фиксировали на льду с использованием 4% параформальдегида в течение 30 минут и промывали 3 раза в PBS. Клетки пермеабилизовали в PBS, содержащем 0,5% Triton X-100, в течение 30 минут и окрашивали с использованием конъюгированного с eFluor-660 антитела крысы против KI67 (1:1000; eBioscience) в течение 30 минут на льду. Для анализа клеточного цикла клетки окрашивали в 1 мкг/мл хехст 33342 (ThermoFisher). Затем окрашенные клетки анализировали с помощью BD FACS Calibur (BD Biosciences).
Для анализа экспрессии органоиды подвергали диссоциации, а затем незамедлительно сортировали с помощью BD FACS Aria (BD Biosciences). Клетки сортировали как отдельные клетки в тризоле в 96-луночном планшете или как совокупность в тризоле в пробирках Eppendorf.
Выделение РНК
Для секвенирования РНК клетки сортировали в тризоле (Life Technologies) и выделяли тотальную РНК по инструкциям производителя со следующими изменениями. РНК осаждали в течение ночи при -20°C с использованием 2 мкг гликогена (Life Technologies). Не использовали дополнительную стадию выделения РНК для клеток, сортированных в 384-луночные планшеты. Для количественного анализа ПЦР РНК выделяли из органоидов с использованием набора RNAeasy (QIAGEN) по инструкциям производителя.
Количественная ПЦР
Анализ ПЦР осуществляли с использованием систем SYBR-Green и Bio-Rad, как описано (Munoz et al.(2012) The EMBO Journal 31:3079-3091). Реакции ПЦР осуществляли в трех параллелях с использованием стандартной кривой для каждого праймера. Изменения экспрессии вычисляли с использованием программного обеспечения CFX manager (Bio-Rad). Праймеры конструировали с использованием инструмента для дизайна праймеров NCBI.
Секвенирование отдельных клеток и совокупности клеток
Образцы РНК получали с использованием модификации способа CEL-seq, описанного ранее (Grun et al. (2015) Nature 525:251-255; Hashimshony et al. (2012) Cell reports 2:666-673). В раствор тризола добавляли добавку ERCC. Осадок РНК растворяли в смеси праймеров и инкубировали в течение 2 минут при 70°C. Клетки, сортированные в 384-луночные планшеты, лизировали напрямую при 65°C в течение 5 минут. Библиотеки кДНК секвенировали с помощью Illumina NextSeq500 с использованием секвенирования спаренных концов размером 75 п.н.
Анализ данных секвенирования РНК. Риды при секвенировании спаренных концов количественно анализировали, как описано ранее (Grun et al. (2015) Nature 525:251-255) со следующими исключениями. Исключали риды, не выровненные или выровненные с множеством участков. В случае анализа секвенирования совокупности игнорировали UMI; вместо этого количество ридов для каждого транскрипта определяли по количеству ридов, картированных в этом транскрипте уникальным образом. Это количество делили на общее количество ридов, картированных по всем транскриптам, и умножали на миллион для получения количества ридов на миллион (RPM). RPM использовали вместо RPKM, т.к. CEL-seq позволяет секвенировать только 3'-концы. Дифференциальную экспрессию генов оценивали с использованием пакета DESeq на платформе R (Anders and Huber (2010) Genome biology 11:R106). Используемые пороговые значения представляли собой скорректированное значение p <0,1 и FDR < 0,1 и по меньшей мере 2-кратное отличие от сравниваемой популяции. Для предотвращения негативного влияния образцов без ридов на анализ соотношений, все риды 0 преобразовывали в риды 0,1 перед вычислением соотношений и log2-преобразованием. Анализ генной онтологии осуществляли с использованием программного обеспечения Revigo (Supek et al. (2011) PloS one 6:e21800) и Gorilla (Eden et al. (2009) BMC bioinformatics 10:48). Данные секвенирования отдельных клеток анализировали, как описано ранее (Grun et al. (2015) Nature 525:251-255).
Пример 1
Для понимания того, как Lgr5+ стволовые клетки удерживаются в клеточном цикле, авторы настоящего изобретения воздействовали на ключевые пути передачи сигнала, активные в нише крипты. С помощью анализа проточной цитометрии Lgr5GFPDTR органоидов с использованием антител против KI67, маркера циклирующих клеток во всех фазах клеточного цикла, подтверждали, что большинство Lgr5+ клеток находятся в цикле (фигура 7). Авторы настоящего изобретения ингибировали передачу сигнала Wnt двумя независимыми способами: i) обработка IWP-2 ингибирует секрецию Wnt3 клетками Панета, и ii) удаление R-спондина 1 из культуральной среды приводит к утрате рецепторов Frizzled с поверхности клеток. Удаление R-спондина 1 незамедлительно вызывает утрату экспрессии Lgr5GFP (фигура 7A). Обработка IWP вызывает более слабое ингибирование Wnt, зависящее от разведения лигандов посредством пролиферации. Экспрессия Lgr5GFP постепенно подвергалась отрицательной регуляции, в то время как стволовые клетки дифференцировались. Кроме того, остальные Lgr5GFP клетки сохраняли экспрессию KI67 (63,5±2,8% по сравнению с 94,4±2,1% в контроле). Удаление ингибитора BMP Noggin или добавление ингибитора Notch DAPT индуцировало быстрое снижение количества GFP+ клеток, но не влияло на пролиферацию оставшихся Lgr5 клеток (82,3±1,4% при удалении Noggin, 45,1±10% при использовании DAPT) (фигура 7). Затем авторы настоящего изобретения ингибировали передачу сигнала EGFR с использованием гефитиниба с сопутствующим удалением EGF из среды для культивирования (обработка EGFRi, фигура 7). Хотя экспрессия Lgr5GFP сохранялась, Lgr5 клетки в конечном итоге утрачивали экспрессию KI67 (13,1±1,0%), что свидетельствует о выходе из клеточного цикла.
Авторы настоящего изобретения дополнительно анализировали ранние события, ассоциированные с ингибированием EGFR (фигура 1A). Несмотря на обширный апоптоз компартментов "ворсинки" органоида, бугорки, напоминающие структуры крипты, содержащие Lgr5+ (Lgr5GFPiresCreER) клетки, сохранялись в течение периода до недели (фигура 1B). Авторы настоящего изобретения подтверждали эти результаты с использованием аллеля Lgr5GFPDTR (Tian et al.(2011) Nature 478:255-259), подтверждая, что CBC выживают в отсутствие передачи сигнала EGF (фигура 7). После 4 дней обработки EGFRi Lgr5+ клетки составляли 44,4±0,8% (13,6±6,5% в контроле) органоидов (фигура 7). С помощью репортерного аллеля TCFCFP (RosaTCF-CFP) Wnt (Serup et al. (2012) Disease models & mechanisms 5:956-966) подтверждали, что сигналы Wnt в непролиферирующих клетках Lgr5 оставались высокими (фигура 7). Белок KI67 сохранялся в течение первых 24 ч., но утрачивалась, начиная с 48 ч. (фигуры 1C и 1D). Используя короткий импульс EdU в качестве меры клеток с репликацией ДНК, авторы настоящего изобретения обнаружили, что EGFRi приводил к быстрой остановке репликации ДНК уже через 24 ч, сохраняющейся по меньшей мере неделю (фигуры 1C и 1D). В соответствии с выходом из S-фазы и, в конечном итоге, из клеточного цикла, мечение содержания ДНК в обработанных EGFRi органоидах с использованием окрашивания ДНК хехстом подтверждало, что все клетки находились в фазе G0/G1 (фигура 1E). Через 4 дня после обработки EGFRi восстановление передачи сигнала EGF индуцировало быстрый вход в клеточный цикл в пределах 24 ч. (KI67+) и прогрессирование до S-фазы в пределах 48 ч. (EdU+) (фигуры 1F и 7). Даже после второго цикла обработки EGFRi покоящиеся Lgr5 клетки повторно входили в клеточный цикл (фигура 7).
Затем авторы настоящего изобретения дополнительно улучшали анализ неделящихся, индуцированных EGFRi Lgr5+ клеток. В них отсутствует маркер клеточного цикла KI67 и маркер M-фазы pH3, и они не включают EdU, исключая сохранение редких делящихся клеток при обработке EGFRi (фигура 1G). Отсутствие лизоцима (LYZ) и хромогранина A (CHGA) означает, что Lgr5+ клетки не являлись дифференцированными клетками Панета или аргентофильными клетками (EEC), соответственно (фигура 7). Пучковые клетки (M-клетки кишечника) являются редкими механосенсорными клетками, также участвующими в ответе на паразитарную инвазию (Howitt et al. (2016) Science 351:1329-1333). Их можно отличить по типичным апикальным пучкам актина, помеченным ацетилированным тубулином и фаллоидином (Hofer and Drenckhahn (1996) Histochemistry and cell biology 105:405-412). Подавляющее большинство Lgr5+ клеток не демонстрирует морфологию пучковых клеток, и, аналогично, пучковые клетки, главным образом, являлись Lgr5- (фигура 7). С помощью количественного анализа ПЦР подтверждали, что Lgr5 клетки не дифференцировались в энтероциты, клетки Панета, EEC, бокаловидные клетки или пучковые клетки (фигура 7).
При окрашивании на Muc2 (муцин 2), а также на PAS и с использованием альцианового синего для визуализации структур слизистой оболочки выявляли значительное снижение количество бокаловидных клеток после обработки EGFRi (фигура S2). Большинство делящихся предшественников TA образовывали зрелые энтероциты. Хотя авторы настоящего изобретения обнаруживали повышение количества LYZ+ клеток Панета и CHGA+ аргентофильных клеток на бугорок через 4 дня после обработки EGFRi, общее количество (на органоид) любого типа клеток не изменялось значительно (фигура 8). Общее количество пучковых клеток после обработки EGFRi повышалось (фигура S2). Авторы настоящего изобретения подтверждали эти результаты с использованием аллеля Dclk1GFPiresCreER (Nakanishi et al. (2013), подтверждая, что обработка EGFRi повышала абсолютное количество Dclk1+ пучковых клеток в 3,2 раза (11,3±6,6 в ENR, 35,8±8,8 в EGFRi) (фигура 9). В кратком изложении, хотя ингибирование EGFR влияло на композицию типов клеток в органоидах, оно приводит Lgr5+ клетки в состояние покоя без индукции дифференцировки в одну из линий клеток кишечника.
Передача сигнала MAPK является основной нижележащей мишенью пути передачи сигнала EGFR и регулирует прохождение клеточного цикла. Киназа MAPK (Mek) фосфорилирует MAPK (Erk), что приводит к ее ядерной локализации и активации. Обработка EGFRi снижала фосфорилирование ERK уже через 1 ч (фигура 2A). Однако авторы настоящего изобретения наблюдали постепенное восстановление уровней фосфо-ERK (pERK) в течение 48 ч. несмотря на продолжающееся состояние покоя (фигура 2A). Авторы настоящего изобретения исследовали вопрос о том, необходима ли передача сигнала Mek/Erk для прохождения клеточного цикла стволовых клеток кишечника, используя низкомолекулярные ингибиторы Mek (PD0325901; Meki) или Erk (SCH772984; Erki). Оба ингибитора индуцировали состояние покоя Lgr5+ клеток, что означает, что путь ERK ниже EGFR регулирует пролиферацию Lgr5+ клеток (фигура 2B). Использование афатиниба, ингибирующего EGFR и ErbB2, приводило к схожим результатам (фигура 2B). Авторы настоящего изобретения делали вывод о том, что ингибирование EGFR посредством передачи сигнала MAPK является достаточным для индукции обратимого состояния покоя у стволовых клеток органоида кишечника.
Для тестирования того, сохраняют ли покоящиеся Lgr5 клетки потенциал стволовых клеток, авторы настоящего изобретения использовали органоиды Lgr5GFPiresCreERRosa26YFP/LacZ и Lgr5GFPiresCreERRosa26tdTomato для отслеживания судьбы Lgr5+ клеток (фигура 3A). Индукция CreER с использованием 4-OH-тамоксифена (Tmx) приводила к быстрой рекомбинации, которую можно визуализировать с помощью флуоресценции YFP (или tdTomato) или экспрессии LacZ (фигура 3B и 3C). Меченые делящиеся Lgr5 клетки образовывали покоящиеся Lgr5 клетки после обработки EGFRi, что свидетельствует о том, что последние из них фактически получали из активных Lgr5 клеток (фигура 3B). Кроме того, фракция меченых клеток Панета и EEC повышалась после обработки EGFRi (фигуры 3B и 3C). В необработанных EGFRi контролях рекомбинировавшие клетки образовывали целые органоиды после пассажа, что свидетельствует об эффективном мечении стволовых клеток (фигура 3D). После пассажа и повторной активации передачи сигнала EGF потомство рекомбинировавших покоящихся Lgr5 клеток сохранялось в течение нескольких пассажей и образовывало органоиды (фигура 3D). Эти результаты свидетельствуют о том, что покоящиеся Lgr5 клетки, индуцируемые после обработки EGFRi, имели свойства настоящих стволовых клеток.
Пучковые клетки являются неделящимися клетками и могут проявляться свойства, подобные свойствам стволовых клеток, участвуя в регенерации ткани после регенерации и росте опухоли (Nakanishi et al. (2013) Nature Genetics 45:98-103). Авторы настоящего изобретения отслеживали судьбу пучковых клеток с использованием органоидов, полученных из модели мыши Dclk1GFPiresCreERRosaYFP/LacZ. И в нормальных, и в обработанных EGFRi органоидах меченые клетки оставались в виде отдельных клеток с течением времени (фигура 3C). При пассаже в присутствие EGF меченые клетки утрачивались и не участвовали в образовании органоидов (фигура 3C), что свидетельствует против того, что Dclk1+ пучковые клетки имеют потенциал стволовых клеток в условиях органоида.
Для лучшего понимания молекулярных характеристик покоящихся Lgr5 клеток авторы настоящего изобретения осуществляли секвенирование РНК с использованием выделенных посредством FACS активных (DMSO-контроль) и покоящихся (обработка EGFRi, d4) Lgr5+ стволовых клеток. Авторы настоящего изобретения включали в свое исследование аллели Lgr5GFPiresCreER (n=2) и Lgr5GFPDTR (n=2). Авторы настоящего изобретения также включали сортированные Dclk1GFP пучковые клетки и совокупные органоиды для сравнения. При иерархической кластеризации и анализе основных компонентов (PCA) определяли, что Lgr5+ клетки были больше похожи друг на друга, чем на целые органоиды или пучковые клетки (фигуры 4A и 4B). В соответствии с различиями в их клеточном цикле, активные и покоящиеся Lgr5+ стволовые клетки кластеризовали раздельно (фигуры 4A и 4B). При анализе дифференциальной экспрессия генов с использованием активных и покоящихся Lgr5 клеток выявили 533 дифференциально регулируемых генов, 290 из которых были обогащены в покоящихся Lgr5 клетках (FDR <0,01 фигура 5C). Транскрипционные эффекторы пути ERK (Etv4 [в 7,7 раз, скорректированное p <0,001] и Etv5 [в 7,7 раз, скорректированное p<0,001]) подвергались отрицательной регуляции в покоящихся Lgr5+ стволовых клетках, что подтверждает эффективное ингибирование Erk (фигуры 4C и 4D). Аналогично, несколько ассоциированных с клеточным циклом генов, таких как Ccnb1 (в 2,1 раз, скорректированное p<0,005) и Ccnb2 (в 1,9 раз, скорректированное p<0,05), были снижены, что соответствует аресту клеточного уцикла в фазе G0 (фигуры 4C и S9). Посредством анализа GO генов, подвергающихся отрицательной регуляции после обработки EGFRi, подтверждали очевидную утрату ассоциированных с клеточным циклом генов (фигура S9). Одним из определяющих транскриптов в покоящихся Lgr5 клетках является Lef1 (неопределяемый в активных Lgr5, скорректированное p<0,001), компонент пути передачи сигнала Wnt. В соответствии с экспрессией используемого репортера, авторы настоящего изобретения наблюдали значительное повышение некоторых хорошо известных генов-мишеней Wnt, включая Rnf43 (в 2,3 раз, скорректированное p<0,005) и Lgr5 (в 2 раза, скорректированное p<0,05)(фигура 4D). Эти результаты подтверждали с помощью количественного анализа ПЦР в независимых экспериментах (фигура 9). Авторы настоящего изобретения также отмечали значительное повышение членов семейства факторов транскрипции AP-1 (Junb, Fos, Fosb) в покоящихся Lgr5 клетках (фигура 4D). В соответствии с гистологическим анализом, специфическая для клеток Панета, энтероцитов и бокаловидных клеток экспрессия генов оставалась неизменной. Chga, экспрессируемый EEC и их предшественниками, в покоящихся клетках повышался в 7,3 раза по сравнению с активными Lgr5+ стволовыми клетками, хотя высокое изменение низкоуровневой экспрессии препятствует дальнейшим выводам (значение p=0,019 и скорректированное p=0,28). Аналогично, хотя Dclk1 (в 6 раз, скорректированное p<0,05) и некоторые другие маркеры пучковых клеток повышались после обработки EGFRi, их уровни были значительно ниже в покоящихся Lgr5 клетках, чем в пучковых клетках (фигура 4D). Глобальные изменения экспрессии генов должны быть общими для всех покоящихся Lgr5 стволовых клеток или должны быть результатом изменений в конкретной субпопуляции. Авторы настоящего изобретения недавно показали, что отдельные активные Lgr5 клетки скорее являются гомогенными (Grun et al. (2015) Nature 525:251-255). Авторы настоящего изобретения осуществляли секвенирование с использованием отдельных клеток всего 192 очищенных посредством FACS отдельных активных или покоящихся Lgr5 клеток из контрольных или обработанных EGFRi органоидов. Используя RaceID (Grun et al. (2015) Nature 525:251-255), авторы настоящего изобретения идентифицировали отдельную выделяющуюся популяцию (кластер 1), содержащую клетки из контрольных и обработанных EGFRi клеток (фигура 4F), содержащих активные и покоящиеся Lgr5+ клетки (фигура 9). Минорные кластеры 2 (четыре клетки) и 4 (одна клетка) экспрессировали гены, ассоциированные с клетками Панета (Lyz1) и пучковыми клетками (Dclk1), в то время как кластер 3 (одна клетка) напоминал кластер 1 (фигура 9). Таким образом, популяция покоящихся Lgr5 клеток является гомогенной и, несмотря на некоторые значительные изменения экспрессии генов, очень напоминают покоящиеся Lgr5 клетки.
Пример 2
Состояние покоя в комбинации с повышенной экспрессией Chga и Lgr5 (как описано в примере 1) напоминает удерживающих метку, секреторных предшественников, описанных Winton с соавт. (Buczacki et al. (2013) Nature 495:65-69). Эти клетки эффективно дифференцируются в EEC, что позволяет предполагать, что выход Lgr5 клеток из клеточного цикла может облегчать образование EEC. Гормоны, экспрессируемые EEC, регулируют широкий спектр физиологических ответов, таких как опорожнение желудка, высвобождение ферментов поджелудочной железы, настроение и толерантность к глюкозе. Их также используют для определения подтипов (см. введение). Сопряженные с G-белком вкусовые рецепторы идентифицированы в качестве регуляторов секреции гормонов (Janssen and Depoortere (2013) TEM 24:92-100). EEC могут иметь непосредственный контакт с просветом и ощущать его содержимое с помощью микроворсинок. Другие EEC, так называемые клетки закрытого типа, не образуют выстилку в просвете и нуждаются в других источниках стимуляции (Janssen and Depoortere (2013) TEM 24:92-100). Длина их базального отростка также варьируется, и он может образовывать синаптические контакты с нейронами кишечника для связи с нервной системой.
Для разработки способа дифференцировки EEC авторы настоящего изобретения использовали покоящиеся Lgr5 клетки в качестве отправной точки и модулировали пути передачи сигнала Notch и Wnt, участвующие в секреторной дифференцировке. Ингибирование передачи сигнала Notch посредством обработки DAPT (D) повышало секреторную дифференцировку, что приводило к значительному повышению количества LYZ+ клеток Панета (фигура 5A). Ингибирование секреции Wnt с использованием IWP-2 (I) в комбинации с DAPT прекращало дифференцировку клеток Панета и индуцировала EEC и бокаловидные клетки (фигура 5A). Обработка EGFRi ингибировала дифференцировку бокаловидных клеток, но не клеток Панета и EEC (фигура 5A). Комбинированное ингибирование путей EGFR/WNT/Notch (IDEGFRi) приводило к значительному повышению EEC, одновременно ингибируя дифференцировку энтероцитов, бокаловидных клеток и клеток Панета (фигура 5A). Аналогично, ингибирование Mek вместе с путями передачи сигнала Wnt и Notch (IDMeki) повышало количество CHGA+ клеток. Различные подтипы EEC редки в нормальных культурах органоидов кишечника (фигура 5B). Обработка IDMeki приводила к устойчивому повышению количества этих типов клеток EEC. Для подтверждения этих результатов авторы настоящего изобретения использовали qPCR. Экспрессия пан-EEC маркера Chga была в 25 раз выше в обработанных IDEGFRi органоидах и в 100 раз выше в обработанных IDMeki органоидах (фигура 5C). Удивительно, что экспрессия мРНК Sst (в 55 раз), Gip (в 14 раз), Sct (в 5 раз), холецистокинина (в 15 раз) и глюкагона (Gcg/проглюкагона, в 4 раза) подвергалась положительной регуляции после обработки IDEGFRi, аналогично IDMeki (фигура 5C). Nts являлся единственным анализируемым гормоном, экспрессируемым на уровне контроля. Таким образом, используя этот способ, авторы настоящего изобретения успешно получали большие количества EEC и различных подтипов, экспрессирующих панель гормонов, регулирующих физиологию млекопитающих (Egerod et al. (2012) Endocrinology 153:5782-5795).
Для определения клеточной композиции индуцированных органоидов EEC и степени гетерогенности экспрессии гормонов отдельными ECC авторы настоящего изобретения осуществляли секвенирование РНК с использованием отдельных клеток. Авторы настоящего изобретения сортировали живые отдельные клетки (без дополнительных маркеров) из обработанных IDEGFRi и IDMeki органоидов. Среди 289 клеток, прошедших фильтрацию, авторы настоящего изобретения идентифицировали кластер из 94 клеток как энтероцитов, обогащенных Aldob (в 4,9 раз, скорректированное p<0,001), Apoa1 (в 12,6 раз, скорректированное p<0,001) и Alpi (в 5,6 раз, скорректированное p<0,001, фигура 10). Их исключали из дальнейшего анализа для лучшей характеризации оставшихся популяций. Клетки, полученные из обработанных IDEGFRi или IDMeki органоидов, распределялись в пространстве t-SNE схожим образом, и их анализировали совместно (фигура S4).
Используя RaceID, авторы настоящего изобретения идентифицировали 12 отдельных кластеров клеток (фигура 6A). С помощью кластеризации способом k-средних коэффициента корреляции Пирсона транскриптомов клеток выявляли четкое разделение между кластерами, а также возможную гетерогенность в кластерах (например, 7 и 8, фигура 6A). При анализе дифференциальной экспрессии генов выявляли сигнатуры генов для каждого кластера, используемые авторами настоящего изобретения для классификации типов клеток (таблица S1).
Наиболее выдающимися кластерами являлись 3 (53 клетки) и 4 (35 клеток), экспрессирующие пан-EEC маркеры Chga и Chgb (фигура 6C). В случае экспрессии Chga и Reg4 наблюдали градиент, при этом она была выше в кластере 4. Продукцию гормонов в этих кластерах с высоким Chgb определяли по экспрессии Tac1 и Tph1, оба из которых являются маркерами энтерохромаффинных клеток. Tac1 кодирует гормон субстанцию P, в то время как Tph1 кодирует скорость-лимитирующий фермент при синтезе серотонина (Egerod et al. (2012) Endocrinology 153:5782-5795; Grun et al. (2015) Nature 525:251-255). Оба из них могут действовать в качестве нейромедиаторов, возбуждающих связанные нейроны кишечника (Latorre et al. (2016) Neurogastroenterology and motility: the official journal of the European Gastrointestinal Motility Society 28(5):620-630).
Другие кластеры имели относительно низкие уровни транскриптов Chga и Chgb, но включали клетки, экспрессирующие гормоны (фигура 6C). Для кластера 2 (21 клетка) была характерна экспрессия Gip (в 74 раза), экспрессируемого K-клетками. Этот кластер также был богат Fabp5 (в 12,6 раз), что соответствует его роли в секреции Gip (Shibue et al. (2015) American journal of physiology, endocrinology and metabolism 308:E583-591). Члены кластера 5 (9 клеток) экспрессировали очень высокие уровни Sst (в 182 раза), что позволяет идентифицировать их как D-клетки (фигура 6C). Экспрессию грелина (Ghrl) наблюдали в нескольких кластерах, но наиболее высокой она была в кластере 6 (в 19 раз, 3 клетки). Авторы настоящего изобретения также отмечали, что Islet1 (Isl-1, в 9,7 раза) в этих клетках коэкспрессировался с Ghrl. Islet1 играет важную роль в определении судьбы клетки, и его утрата приводит к нарушению гомеостаза глюкозы (Terry et al. (2014) American journal of physiology, gastrointestinal and liver physiology 307:G979-991). Клетки в кластере 7 (18 клеток) в значительной степени экспрессировали Cck (в 55,7 раз), и подгруппы этого кластера также были богаты другими гормонами, такими как Gcg (в 28,2 раз), Ghrl (в 5,3 раз) и Pyy (в 11,4 раз). Соответственно, сообщают, что I-клетки коэкспрессируют Cck с другими гормонами с различными уровнями (Egerod et al. (2012) Endocrinology 153:5782-5795).
Одним из ранних индукторов дифференцировки EEC является нейрогенин-3 (Neurog3), за которым следует Neurod1. Экспрессия Neurog3 (в 5,2 раза) была наиболее высокой в кластере 9 (6 клеток) и в некоторых клетках кластера 3, наиболее схожих с кластером 9. Фактически, все кластеры EEC экспрессировали Neurod1. Учитывая временную экспрессию этих факторов транскрипции, авторы настоящего изобретения предполагали, что кластер 9 представляет собой предшественников EEC, затем с помощью Neurod1 образующих панель EEC. Кластер 1 (18 клеток) был обогащен генами, ассоциированными с бокаловидными клетками и клетками Панета, такими как Agr2 (в 33 раз), Muc2 (в 26 раз), Ttf3 (в 23 раз) и Defa24 (в 28 раз). Даже после фильтрации некоторые оставшиеся энтероцит-подобные клетки, экспрессирующие Aldob и Mt1/2, были видимыми (клетки кластера 8, 7). Эти кластеры могли образоваться до индукции EEC, т.к. авторы настоящего изобретения не наблюдали повышения их количества после ингибирования EGFR или Mek. Учитывая экспрессию Dclk1 и Trpm5, кластер 10 (15 клеток) идентифицировали как пучковые клетки (фигуры 6B и 6C). Всего 145/289 анализируемых клеток (50% всех клеток) являлись EEC или их предшественниками, что подтверждает высокую эффективность способа индукции по изобретению.
Т.к. в одной клетке может коэкспрессироваться множество гормонов, авторы настоящего изобретения дополнительно рассматривали гетерогенность экспрессии гормонов на уровне отдельных клеток (фигура 6D). Авторы настоящего изобретения сфокусировались на кластерах 2, 5, 6 и 7, экспрессирующих множество гормонов. Наблюдали 4 основные группы с учетом экспрессии Gip, Sst, Cck и грелина. Cck+ клетки дополнительно разделяли на кластеры Gcg+Ghrl+, Gcg+Ghrl-, Gcg-Ghrl+ и Gcg-Ghrl-, при этом некоторые также коэкспрессировали низкие уровни Sst и/или Gip. Транскриптомы Sst+ клеток были более гомогенными, при этом клетки коэкспрессировали низкие уровни Gip и Cck, в то время как одна клетка коэкспрессировала только Ghrl. Авторы настоящего изобретения ранее сообщали о частичном перекрывании между Cck+ и Tac1+ клетками. Соответственно, некоторые из Tac1+ клеток в кластерах 3 и 4 экспрессировали низкие уровни Cck (фигуры 6B и 6C).
В целом, анализ отдельных клеток, проведенный авторами настоящего изобретения, свидетельствовал о том, что способ по изобретению приводил к обогащению EEC до ~50% культуры с учетом экспрессии маркерных генов. Кроме того, авторы настоящего изобретения получали панель подтипов EEC, включая некоторые редкие клетки с комплексными профилями экспрессии гормонов.
Пример 3
Для получения органоидов использовали дифференцировочную среду для EEC мыши, содержащую ингибиторы передачи сигнала Wnt, Notch и MAPK. Эти органоиды содержали смесь всех подтипов EEC. Энтерохромаффинные клетки, продуцирующие серотонин, являлись наиболее многочисленным типом клеток в этих культурах. Исследовали механизм, посредством которого образуются различные подтипы EEC. Это сделано с расчетом на расширение применения культуры и для понимания развития этих клеток. При первом скрининге путей передачи сигнала, которые могут контролировать определение подтипа EEC, обнаруживали, что модуляции путей BMP, Hedgehog и mTOR влияют на относительные соотношения подтипов EEC. По-видимому, секретин-продуцирующие S-клетки являются особенно редкими при ингибировании Wnt, Notch и MAPK. Как правило, S-клетки располагаются в проксимальных отделах двенадцатиперстной кишки, том же месте, откуда выделяли органоиды в этом исследовании. Активация пути BMP посредством удаления Noggin из дифференцировочной среды для EEC и добавление BMP4 (10 мкг/мл) в среду для культивирования изменяли относительный избыток подтипов EEC. После активации пути BMP наблюдали значительное повышение количества S-клеток (с учетом матричной РНК, в 400 раз относительно контроля), а также уровней секретина на клетку (с учетом IHC). Это повышение S-клеток, по-видимому, происходит за счет количества энтерохромаффинных клеток, что позволяет предполагать потенциальное перекрывание путей развития. См. дальнейшее обсуждение роли передачи сигнала BMP в примере 5.
Пример 4
Органоиды тонкого кишечника (SI) человека сначала культивировали в среде для выращивания (как описано в Sato et al. (2011) Gastroenterology 141(5):1762-72) для повышения количества стволовых клеток, а затем высевали в дифференцировочную среду (среду для выращивания без Wnt-кондиционированной среды, ингибитора TGFβ, ингибитора p38 и никотинамида) для направления дифференцировки. Повторный посев включал разрушение матригеля (без диссоциации органоидов) в холодной среде, промывку органоидов минимальной средой для культивирования (также можно использовать PBS) и последующее высевание в свежий матригель (без диссоциации органоидов). Затем органоиды культивировали в дифференцировочной среде в течение 1 дня. На следующий день использовали способ EEC-специфической аргентофильной дифференцировки. Он включал добавление EEC-дифференцировочной среды на 5 дней, являвшейся такой же, как стандартная дифференцировочная среда (среда для выращивания без Wnt-кондиционированной среды, ингибитора TGFβ, ингибитора p38 и никотинамида) с добавлением 1,5 мкМ IWP2, 10 мМ DAPT, 100 нМ MEKi PD0325901. Для дифференцировки этих клеток человека в направлении EEC в этом эксперименте требуются гораздо более низкие уровни ингибиторов MAPK, чем для дифференцировки клеток мыши в направлении EEC в примере 3 (100-500 нМ по сравнению с 1-5 мкМ в системе мыши), вероятно, по причине отсутствия клеток Панета, продуцирующих EGF в системе человека.
Обсуждение
Авторы настоящего изобретения идентифицировали передачу сигнала EGF как необходимый регулятор пролиферации Lgr5 стволовых клеток в органоидах. В условиях, в которых передача сигнала Wnt остается нетронутой, но передача сигнала EGF заблокирована, активно делящиеся Lgr5 стволовые клетки преобразуются в покоящиеся Lgr5 клетки, сохраняющие экспрессию различных генов-мишеней Wnt. Это состояние клеток можно поддерживать в течение периода до недели. Кроме того, простое восстановление передачи сигнала EGF преобразует покоящиеся клетки обратно в состояние нормальных активных стволовых клеток. При анализе дифференциальной экспрессии выявили утрату хорошо известных индуцирующих пролиферацию факторов транскрипции, таких как Ets-подобные факторы Etv 4 и 5, что позволяет предполагать, что они играют ключевую роль в делении стволовых клеток.
Таким образом, химическое ингибирование передачи сигнала EGFR приводило к аресту Lgr5+ стволовых клеток, не влияя на их потенциал стволовых клеток. Авторы настоящего изобретения считают, что передача сигнала Wnt на высоком уровне поддерживает этот потенциал стволовых клеток при индуцированном состоянии покоя. В органоидах, а также криптах Lgr5+ клетки всегда являются непосредственными соседями Wnt3-секретирующих клеток Панета (Sato et al. (2011) Nature 469:415-418). В этих условиях Wnt3 не диффундирует на расстояния, а воздействует напрямую на Lgr5 стволовые клетки (Farin et al. (2016) Nature 530:340-343). Покоящиеся Lgr5 стволовые клетки остаются вблизи клеток Панета в обработанных EGFRi органоидах и, таким образом, подвергаются воздействию высоких локальных сигналов Wnt.
Фактически, при использовании 3 независимых аллелей генов-мишеней Wnt, а также анализа экспрессии генов подтверждали повышение передачи сигнала Wnt после ингибирования EGFR. В целом, результаты, полученные авторами настоящего изобретения, свидетельствуют о том, что для сохранения судьбы стволовых клеток необходим Wnt, но не EGF, в то время как пролиферация стволовых клеток зависит от комбинации Wnt и EGF.
Предыдущие исследования показали наличие покоящихся клеток вблизи зоны дифференцировки в положении "+4" с потенциалом стволовых клеток (Sangiorgi and Capecchi (2008) Nature Genetics 40:915-920; Takeda et al. (2011) Science 334:1420-1424; Yan et al. (2012) Proceedings of the National Academy of Sciences of the United States of America 109:466-471). Авторы настоящего изобретения сообщали о существовании Dll1+ предшественников, образующих исключительно секреторные клетки в этом положении (van Es et al. (2012) Nature cell biology 14:1099-1104). Используя анализ удержания гистоновой метки, группа под руководством Doug Winton идентифицировала схожую популяцию с секреторным дифференцировочным потенциалом. Эти удерживающие метку клетки обладают общей сигнатурой с CBC, включая экспрессию Lgr5, но экспрессируют значительные уровни некоторых из генов секреторной линии клеток, таких как Chga (Buczacki et al. (2013) Nature 495:65-69). В совокупности, эти секреторные предшественники соответствуют транзиторным состояниям, при этом они могут дедифференцироваться в стволовые клетки, когда в этом возникает потребность, и, таким образом, их можно рассматривать в качестве факультативных стволовых клеток (Buczacki et al. (2013) Nature 495:65-69; van Es et al. (2012) Nature cell biology 14:1099-1104). Похожая ситуация имеет место в случае избыточных предшественников энтероцитов в крипте (Tetteh et al. (2016) Cell Stem Cell 18:203-213).
Неожиданно, авторы настоящего изобретения отмечали небольшое отклонение покоящихся Lgr5 клеток в сторону экспрессии маркеров EEC, таких как Chga, что делает их напоминающими Lgr5+ удерживающие метку клетки, идентифицированные Doug Winton. EEC составляют менее 1% клеток эпителия кишечника, при этом вместе они образуют наиболее крупный эндокринный орган организма человека в терминах количества клеток (Latorre et al. (2016) Neurogastroenterology and motility: the official journal of the European Gastrointestinal Motility Society 28(5):620-630). Предполагают, что EEC действуют как сенсоры содержимого просвета, такого как питательные вещества и микроорганизмы, и регулируют физиологические ответы, подобные чувствительности к глюкозе, опорожнение желудка, настроение и аппетит, посредством секреции гормонов (Janssen and Depoortere (2013) TEM 24:92-100). Функциональному исследованию EEC мешает отсутствие надежных систем in vitro для получения этих клеток в больших количеств. Авторы настоящего изобретения ранее сообщали о направленной дифференцировке стволовых клеток в секреторные клетки посредством ингибирования Notch (van Es et al. (2005) Nature 435:959-963). Для образования клеток Панета необходима активная передача сигнала Wnt в комбинации с ингибированием Notch (van Es et al. (2012) Nature cell biology 14:1099-1104; van Es et al. (2005) Nature 435:959-963), и комбинированное ингибирование передачи сигнала Wnt и Notch, в основном, приводит к образованию бокаловидных клеток (van Es et al. (2005) Nature 435:959-963; Yin et al. (2014) Nature Methods 11:106-112). При комбинировании ингибирования пути EGFR с блокадой Wnt и Notch из культуры первичных клеток эффективно образуются EEC различных типов. Показано, что для образования бокаловидных клеток важна передача сигнала EGFR (Heuberger et al. (2014) Proceedings of the National Academy of Sciences of the United States of America 111:3472-3477). Авторы настоящего изобретения считают, что одновременное ингибирование судьбы энтероцитов, клеток Панета и бокаловидных клеток посредством ингибирования передачи сигнала Notch, Wnt и EGFR, соответственно, является ключом к получению EEC.
Можно различать до 20 подтипов EEC с учетом продукции конкретных пептидов. Эти клетки присутствуют в разных отделах ЖКТ от проксимальных до дистальных в разных количествах. Обнаружено, что конкретные EEC экспрессируют несколько функционально родственных гормонов на уровнях транскрипта и белка (Egerod et al. (2012) Endocrinology 153:5782-5795; Grun et al. (2015) Nature 525:251-255), хотя некоторые из них могут представлять собой промежуточную стадию развития. Авторы настоящего изобретения демонстрировали одновременное получение нескольких основных подтипов EEC посредством профилирования транскриптома отдельных клеток. Подтверждая приведенное в своей предыдущей работе (Grun et al. (2015) Nature 525:251-255), авторы настоящего изобретения определяли популяции EEC с высоким и низким Chga. Первые из них обогащены Tac1/Tph1-экспрессирующими EEC, при этом некоторые клетки экспрессировали Cck на низком уровне. Последние из них очень отличаются по экспрессии гормонов. Клетки с общим схожим профилированием транскриптомов кластеризовались совместно, даже при том, что их можно подразделять на несколько классов с учетом экспрессии гормонов. Широкое распределение уровней экспрессии гормонов позволяет предполагать, что разделение подтипов EEC не является четко выраженным. Заманчиво предположить, что их экспрессия, по меньшей мере, частично является случайной и даже может иметь временную динамику.
Пример 5
Аргентофильные клетки изменяют экспрессию гормонов при миграции из крипты в ворсинку
Множество гормонов аргентофильных клеток являются дифференциально обогащенными в крипте и ворсинке кишечника. L-клетки коэкспрессируют GLP1 и PYY в крипте, но, чаще всего, в них отсутствует экспрессия GLP1 в ворсинке (фигура 20A-C). Энтерохромаффинные клетки продуцируют серотонин по всей оси крипта-ворсинка, но селективно коэкспрессируют Tac1 в крипте и секретин в ворсинке (фигура 20D-E). Это позволяет предполагать, что EEC могут переключать экспрессию гормонов при миграции из крипты в ворсинку, и что существует потенциальная взаимосвязь линий клеток между EEC крипты и ворсинки. Однако существует альтернативное объяснение этого наблюдения, состоящее в том, что клетки, иммунореактивные в отношении секретина или PYY в ворсинке, получают из неродственного предшественника в крипте, отрицательного по серотонину и Tac1 или GLP1. Для демонстрации того, что такая трансдифференцировка может происходить в течение времени жизни EEC, авторы настоящего изобретения анализировали кишечник мышей Tac1-Cre/Rosa26tdTomato. С помощью этого репортера точно метят клетки в крипте, коэкспрессирующие Tac1 и серотонин. Кроме того, отслеживают серотонин+ клетки и большинство секретин+ клеток в ворсинке, хотя они отрицательны по Tac1. Из Tac1/серотонин+ клетки-предшественника не получают другие гормоны, включая CCK. Эти данные позволяют предполагать, что отдельные EEC могут экспрессировать различные наборы гормонов в крипте и ворсинке. Большинство секретин-продуцирующих клеток являются частью энтерохромаффинной линии. Способность EEC переключать экспрессию гормонов означает, что может существовать меньше несвязанных путей дифференцировки EEC, чем предполагали ранее, но скорее изменения типов гормонов, продуцируемых ограниченным количество клеток, приводят к избытку состояний EEC.
Передача сигнала BMP индуцирует сигнатуру гормонов ворсинки в аргентофильных клетках
Затем авторы настоящего изобретения исследовали то, являются ли гормональные переключения, возникающие при миграции из крипты в ворсинку, частью процесса созревания EEC по умолчанию, или это просто отражает различные сигналы ниши, воздействию которых подвергаются EEC. Известно, что существует множество морфогенов в крипте и ворсинке, включая возрастающие уровни BMP и Hedgehog и снижающиеся уровни сигналы Wnt. Способ дифференцировки аргентофильных клеток в системе органоида кишечника мыши, описанной в примерах выше, главным образом, основан на тройном ингибировании путей передачи сигнала Wnt, MAPK и Notch. Авторы настоящего изобретения использовали эту дифференцировочную среду и добавляли фактор стволовых клеток Noggin в смесь для дифференцировки, таким образом, создавая среду с низким BMP. Неожиданно, авторы настоящего изобретения наблюдали, что все энтерохромаффинные клетки в этой культуре всегда коэкспрессировали серотонин и Tac1, в то время как экспрессия секретина отсутствовала. Таким образом, эта культура отражает сигнатуру гормонов EEC, напоминающую состояние крипты, что означает, что сигналы ниши доминируют в течение регулируемого во времени созревания EEC.
Для исследования того, могут ли сигналы ниши фактически управлять сигнатурами гормонов EEC, авторы настоящего изобретения использовали описанную выше систему дифференцировки EEC в качестве отправной точки и модулировали путь передачи сигнала с использованием этой смеси. Висмогениб добавляли для ингибирования передачи сигнала Hedgehog, Noggin удаляли для активации передачи сигнала BMPR1a/BMPR2 и ингибитор ALK5/4/7 A83 или TGF-бета 1 добавляли для модуляции различных путей рецептора семейства TGF-бета. Хотя Wnt уже ограничен в смеси для дифференцировки EEC с помощью ингибитора поркупина IWP2, авторы настоящего изобретения вдобавок удаляли R-спондин для достижения более выраженного и быстрого ингибирования Wnt. Авторы настоящего изобретения использовали уровни транскриптов Sct и Gcg в качестве заместителей сигнатур гормонов ворсинки и крипты, соответственно, в то время как ChgA включали в качестве транскрипта, имеющего постоянную экспрессию в крипте и ворсинке. Ингибирование передачи сигнала Hedgehog не модулирует любой из анализируемых гормонов, что соответствует его доминирующей роли в мезенхиме кишечника. A83 и TGF-бета 1 ингибировали экспрессию всех гормонов EEC. Аналогично, удаление R-спондина нарушало дифференцировку EEC в отношении всех анализируемых гормонов. Теоретически, это связано с быстрой дифференцировкой стволовых клеток в абсорбирующую линию, когда сигналы Wnt ингибировались, а не следовали в секреторном направлении при утрате сигналов Notch. В конечном итоге, удаление Noggin при дифференцировке EEC индуцировало экспрессию Sct, одновременно репрессируя транскрипцию Gcg, что соответствует переключению с профиля экспрессии крипты на профиль экспрессии ворсинки. Авторы настоящего изобретения добавляли экзогенный BMP4 в эту смесь для дифференцировки EEC для дополнительного усиления передачи сигнала BMP и назвали эту комбинацию средой "EEC с высоким BMP" по сравнению с "EEC с низким BMP", как авторы настоящего изобретения определяли ранее. Стимуляция органоидов с использованием этой дифференцировочной среды "EEC с высоким BMP" индуцировало продукцию клеток, иммунореактивных в отношении секретина, а также энтерохромаффинных клеток, экспрессирующих серотонин без Tac1. При использовании этой схемы количество GLP1+ клеток значительно уменьшалось (фигура 21A-C) без очевидных морфологических изменений органоидов. Органоиды, полученные из различных частей кишечника, сохраняли свою регионарную идентичность в терминах сигнатур гормонов с обогащением GIP в проксимальных отделах SI и Pyy, Nts и Gcg в дистальных отделах SI. Другие гормоны или маркеры EEC, демонстрирующие гомогенное распределение от крипты к ворсинке, включая ChgA, Tph1, CCK и GIP двенадцатиперстной кишки, подвергались лишь небольшому влиянию различных уровней BMP. Авторы настоящего изобретения наблюдали положительную регуляцию Pyy и Nts, больше всего экспрессировавшихся в ворсинке. Эффекты BMP приведены в таблице ниже. В совокупности, эти данные свидетельствуют о роли сигналов BMP в индукции гормонов, ассоциированных с EEC в ворсинке.
Пример 6
LDN193189 (Selleckchem, кат. № S2618) в количестве 35 мг/кг вводили мышам с помощью желудочного зонда при обработке в течение 60 ч. LDN193189 растворяли в лимонной кислоте при pH 3,1 и желудочный зонд вводили дважды в сутки. Это приводило к экспансии GLP1-экспрессирующих клеток в ворсинке кишечника по сравнению с контрольными мышами. Экспрессия секретина в клетках ворсинки кишечника значительно снижалась по сравнению с контрольными мышами (см. фигура 22). Используемая доза была высокой, и предполагают, что более низкие дозы также будут эффективными. Предполагают, что эти результаты можно экстраполировать на людей. LDN193189 является селективным ингибитором передачи сигнала BMP, ингибирующим транскрипционную активность рецепторов BMP типа I ALK2 и ALK3 с IC50 5 нМ и 30 нМ в клетках C2C12, соответственно, проявляющим 200-кратную селективность в отношении BMP по сравнению с TGF-β. Предполагают, что другие ингибиторы передачи сигнала BMP, такие как ингибиторы, представленные в настоящем описании, также будут иметь схожие эффекты in vivo. Делали вывод о том, что ингибиторы BMP (и, наоборот, активаторы BMP) обладают потенциалом для применения в терапии, когда желательна модуляция GLP1 и/или секретина in vivo. Неограничивающие примеры включают лечение диабета, ожирения, гипохлоргидрии или гиперхлоргидрии.
Структура LDN193189:
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> KONINKLIJKE NEDERLANDSE AKADEMIE VAN WETENSCHAPPEN
<120> УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ
<130> P068547WO
<141> 2017-06-20
<150> GB 1610748.4
<151> 2016-06-20
<160> 28
<170> SeqWin2010, version 1.0
<210> 1
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 1
ccagctgcct gtagtgtcaa 20
<210> 2
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 2
tcttgaggct cgccttgatg 20
<210> 3
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 3
gccaagagcc atgtgactat c 21
<210> 4
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 4
cagagctggt actttggtgt tc 22
<210> 5
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 5
acccgccagt ctcctacatc 20
<210> 6
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 6
gcatctaggc gcagggattg 20
<210> 7
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 7
gctgtgcaag ctgaaggg 18
<210> 8
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 8
gctgtgcaag ctgaaggg 18
<210> 9
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 9
cagctcgtcc actctttccg 20
<210> 10
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 10
cctctcgtct ccttggaggg 20
<210> 11
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 11
ggaatggatg gctaccgtgg 20
<210> 12
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 12
catgccaccc atgctcgaat 20
<210> 13
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 13
actaaggtgg cctacctcca a 21
<210> 14
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 14
ggaggtgaca gtcaaggtga ga 22
<210> 15
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 15
aggatccatc tgtcctttgg 20
<210> 16
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 16
acgttgtatg tcttggacag 20
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 17
gaccccaaga cactcagacg 20
<210> 18
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 18
ttttctgtgt cctgctcgct 20
<210> 19
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 19
cttcccagaa gaagtcgcca 20
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 20
gtgactggca cgagatgttg 20
<210> 21
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 21
gaagagcggc gtatgtctgt 20
<210> 22
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 22
ccagaaggag ctttgcgga 19
<210> 23
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 23
gacctgcgac tagactgacc 20
<210> 24
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 24
ccagttcctg tttcccggtg 20
<210> 25
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 25
aactgttggc taggggacac 20
<210> 26
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 26
tgatgaaagt cccctctgcg 20
<210> 27
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 27
tgctgaccat cttccagctc 20
<210> 28
<211> 20
<212> ДНК
<213> праймер
<400> 28
gaatgtaggg ccttctgggt 20
<---
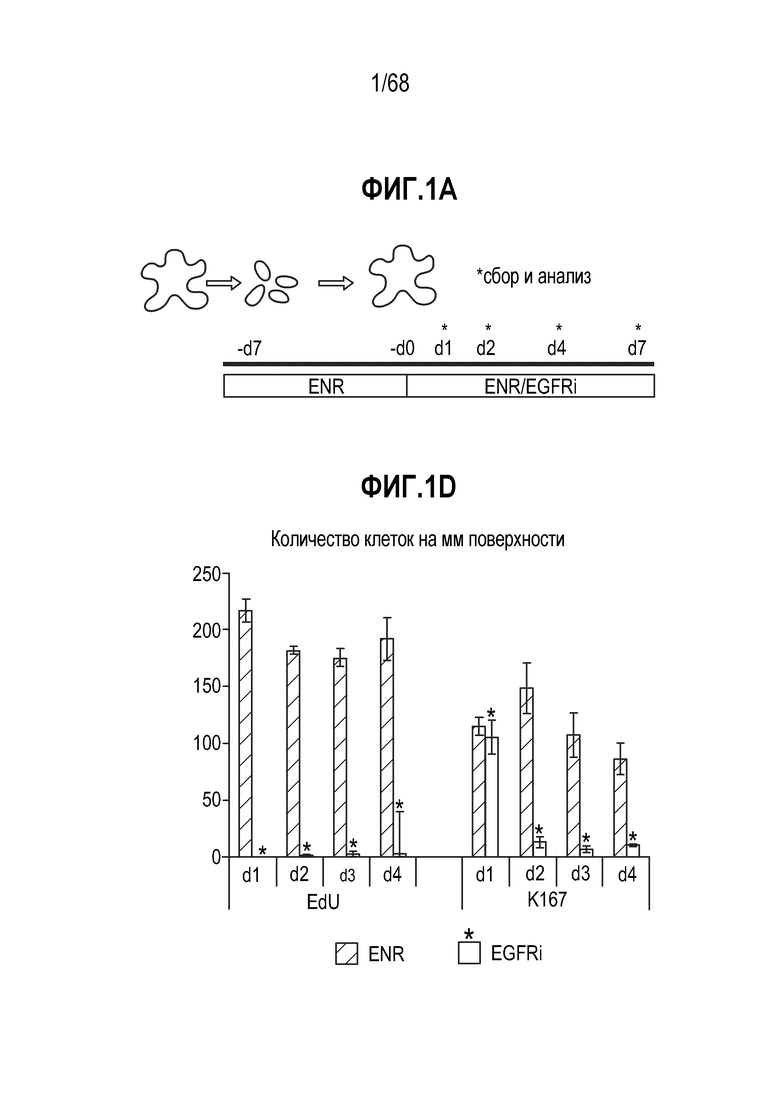
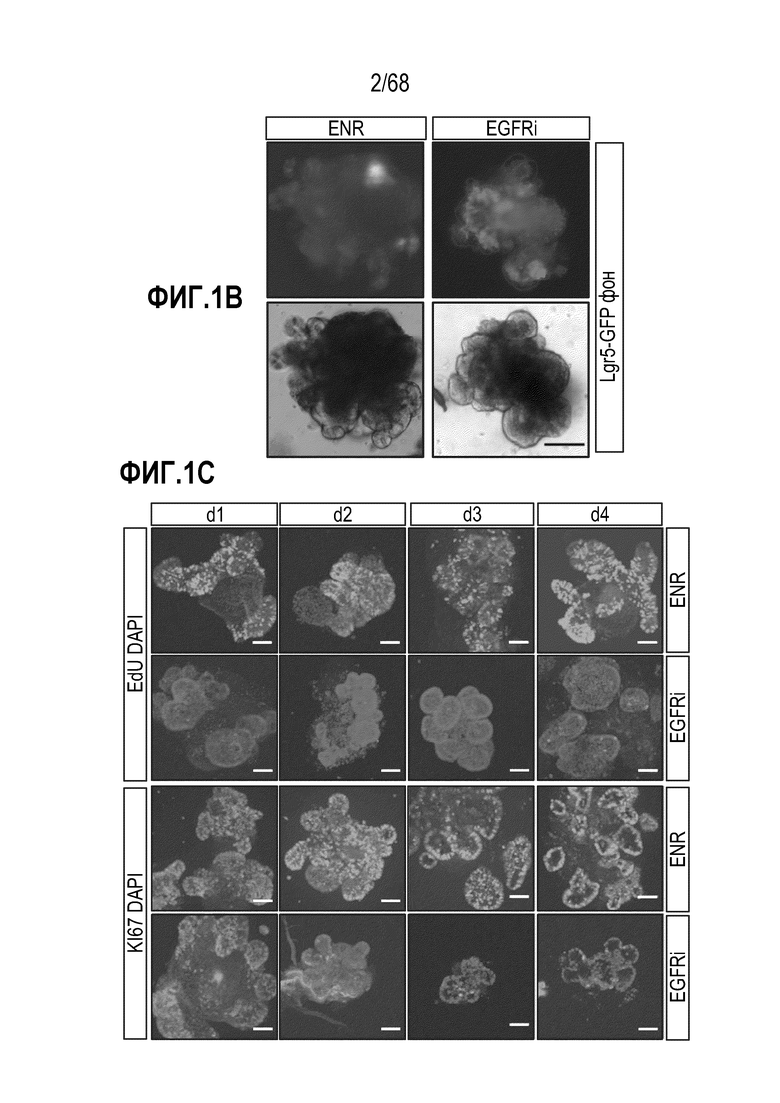
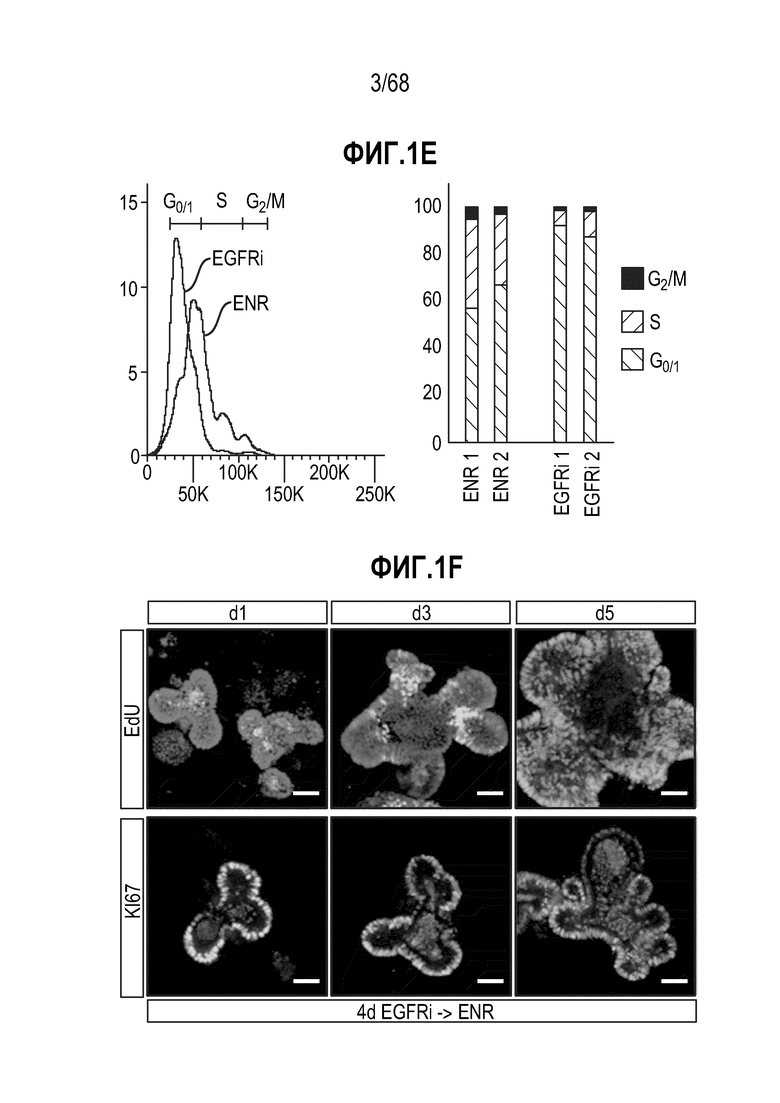
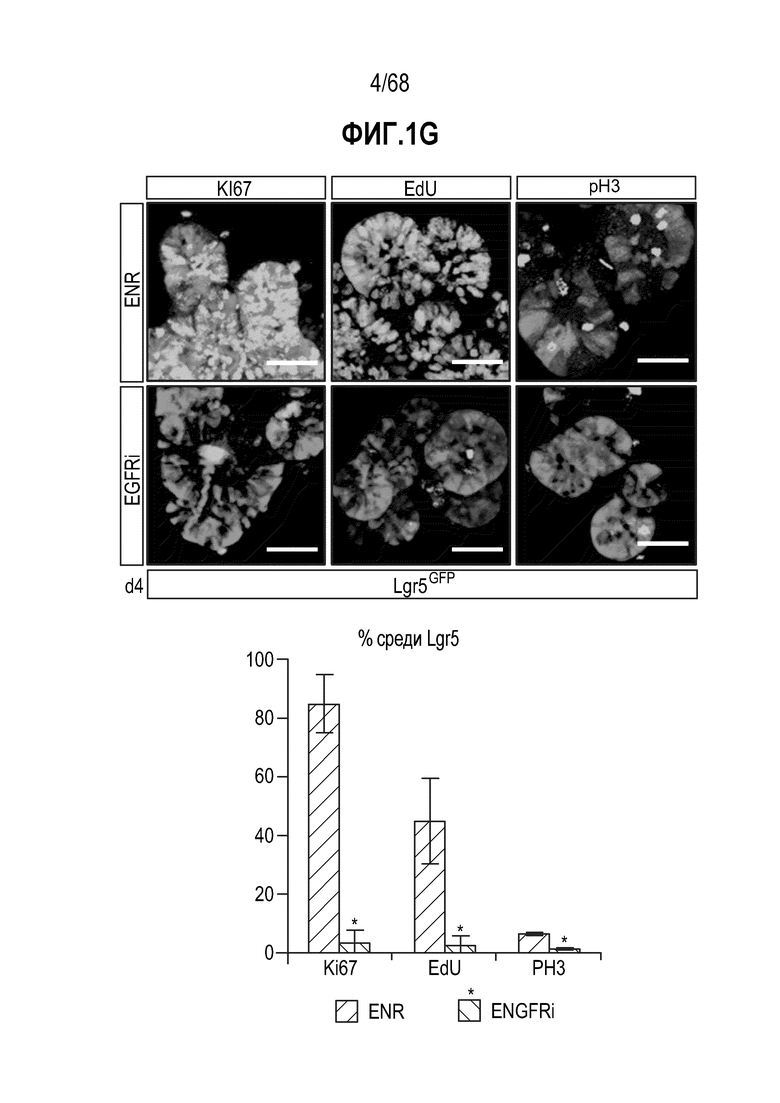
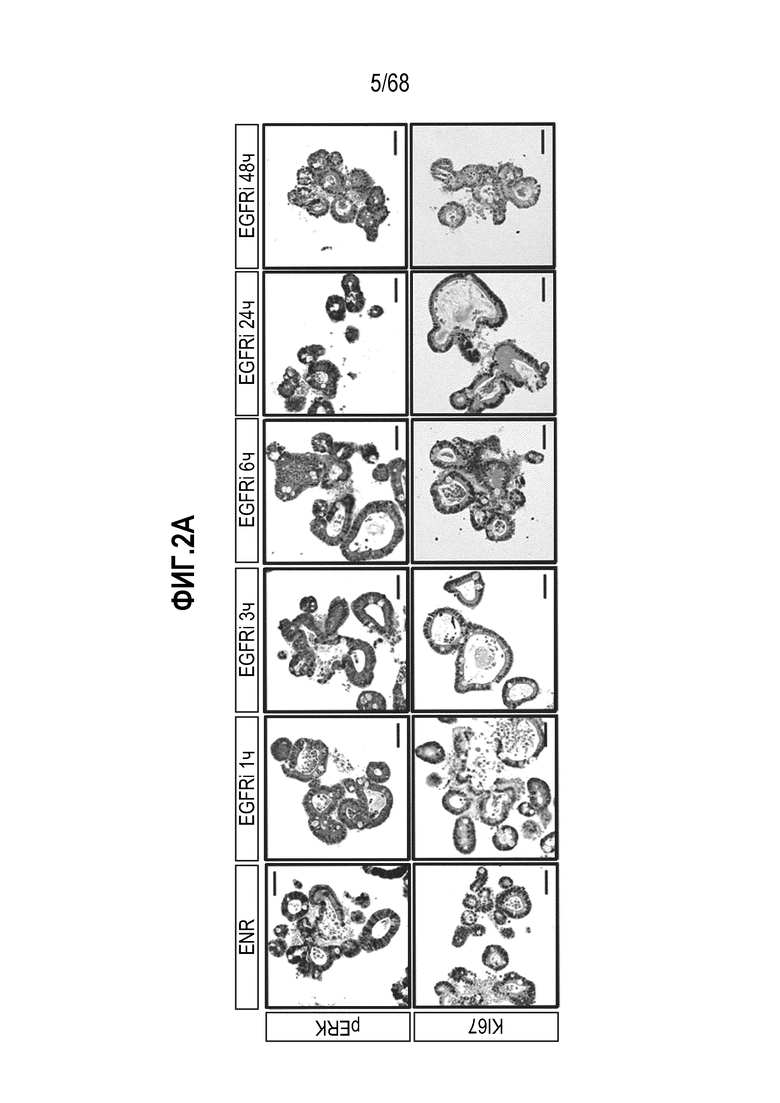
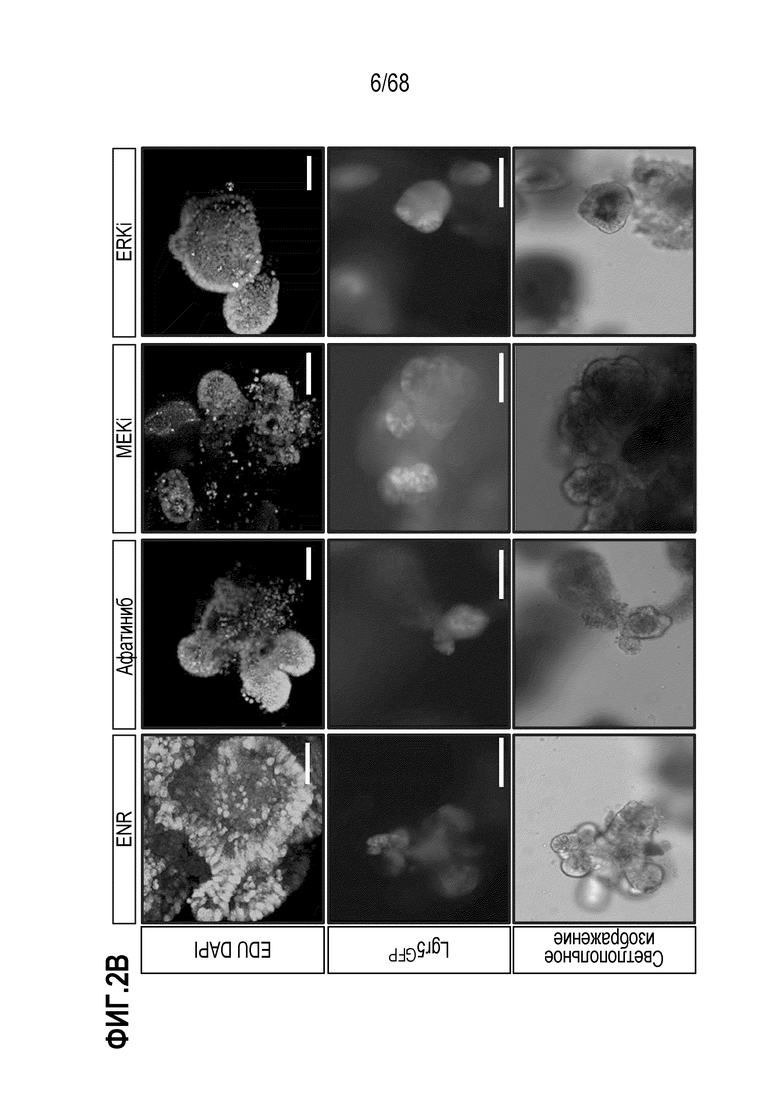
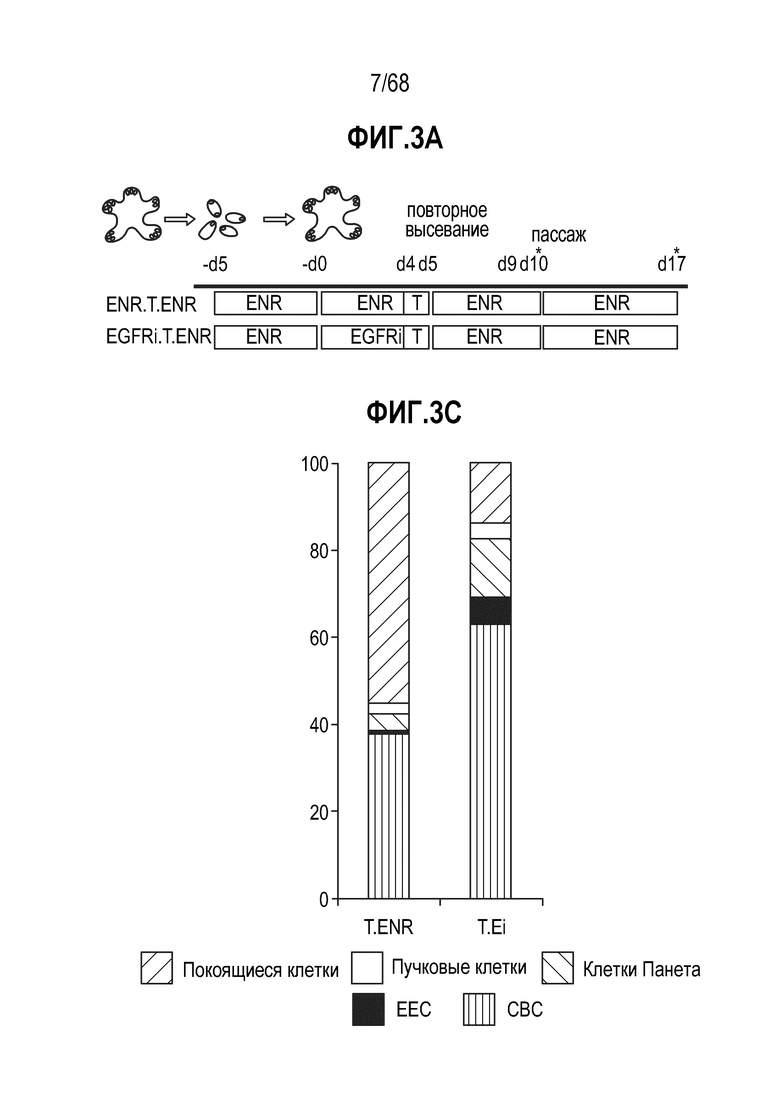
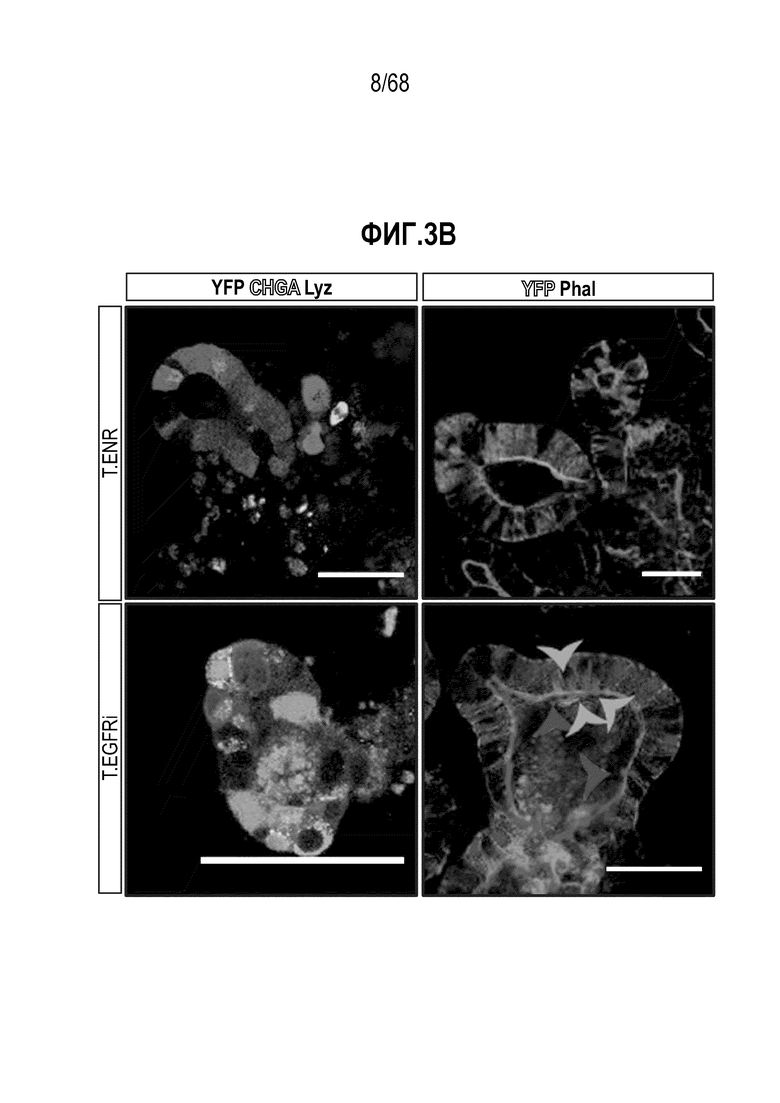
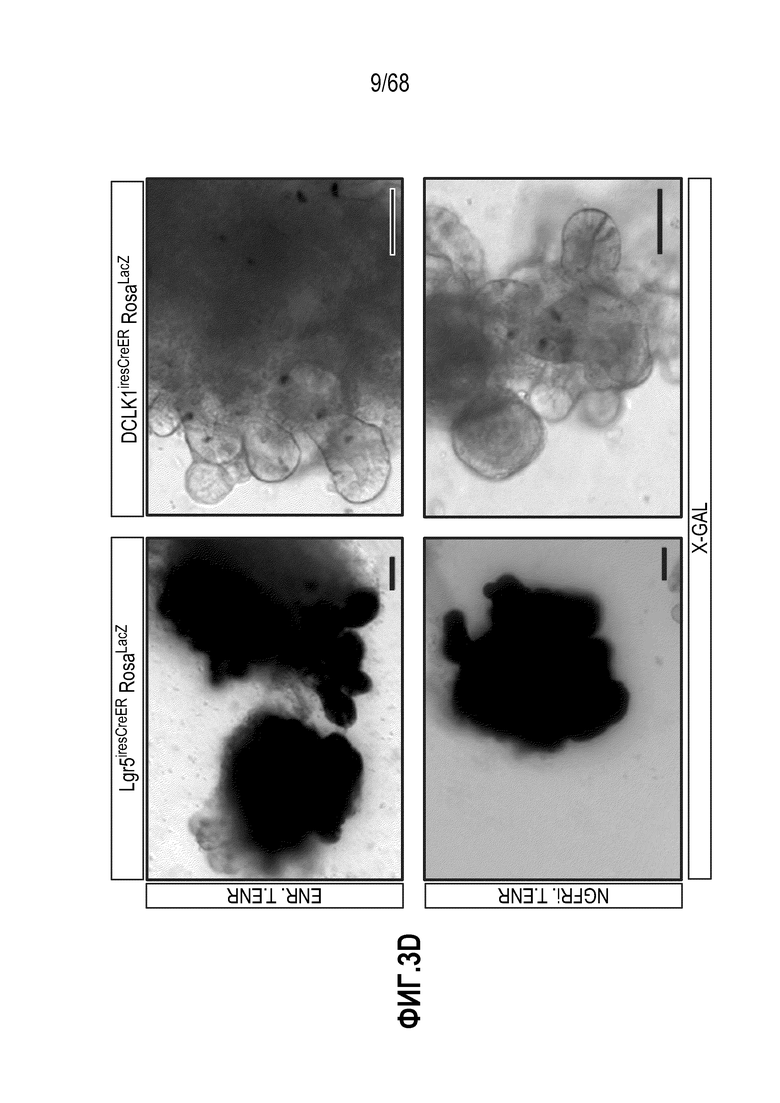
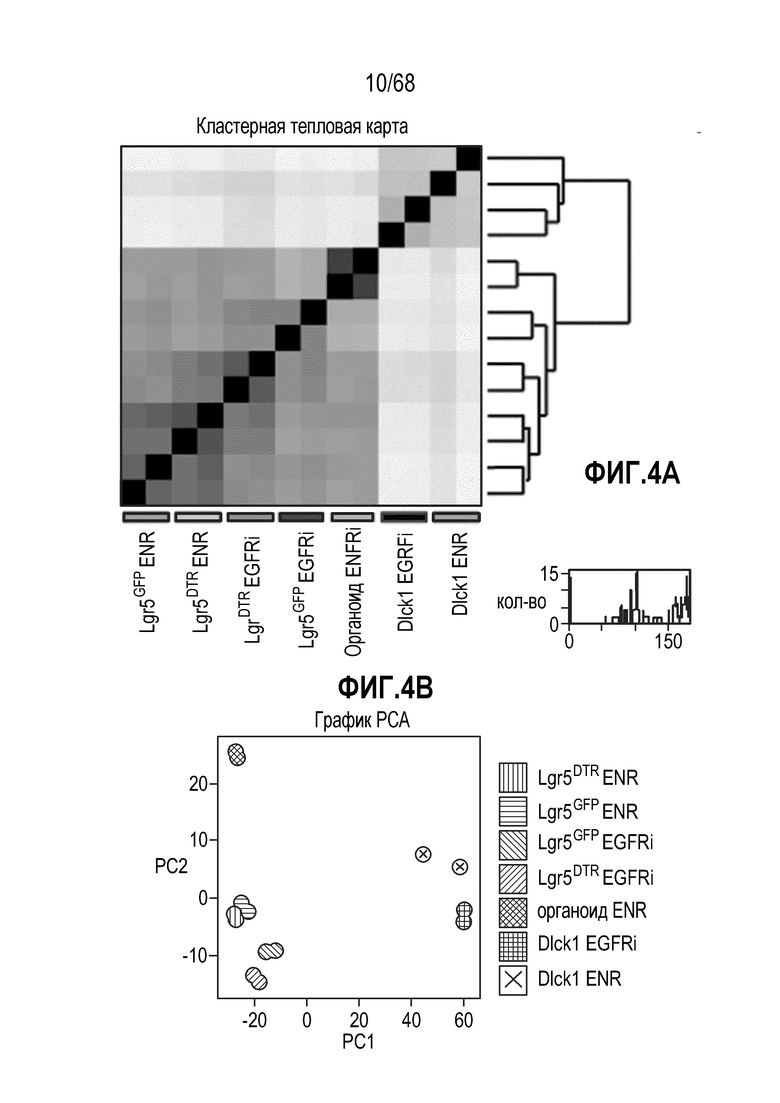
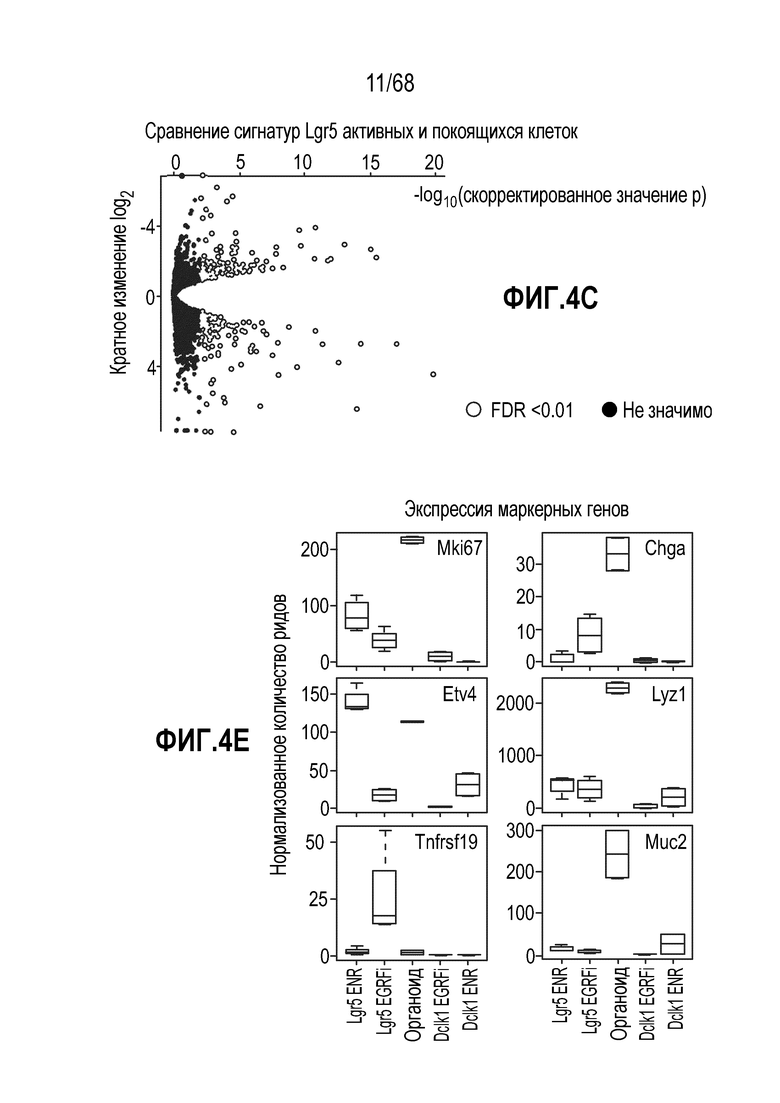
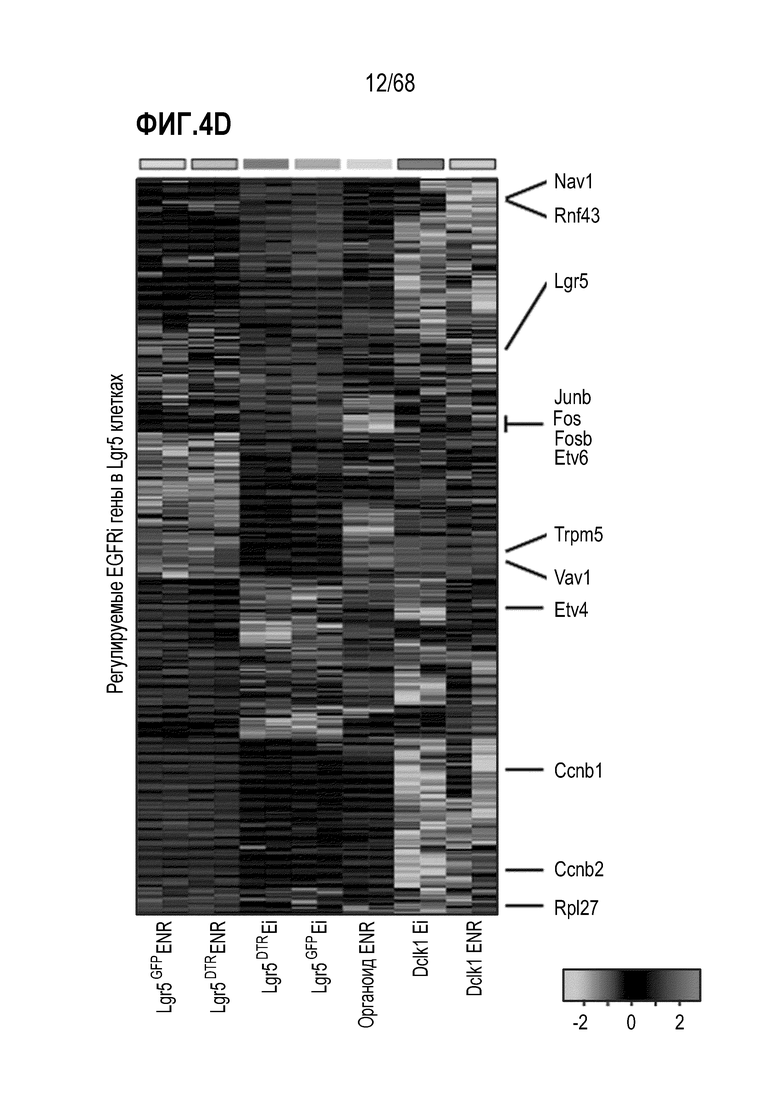
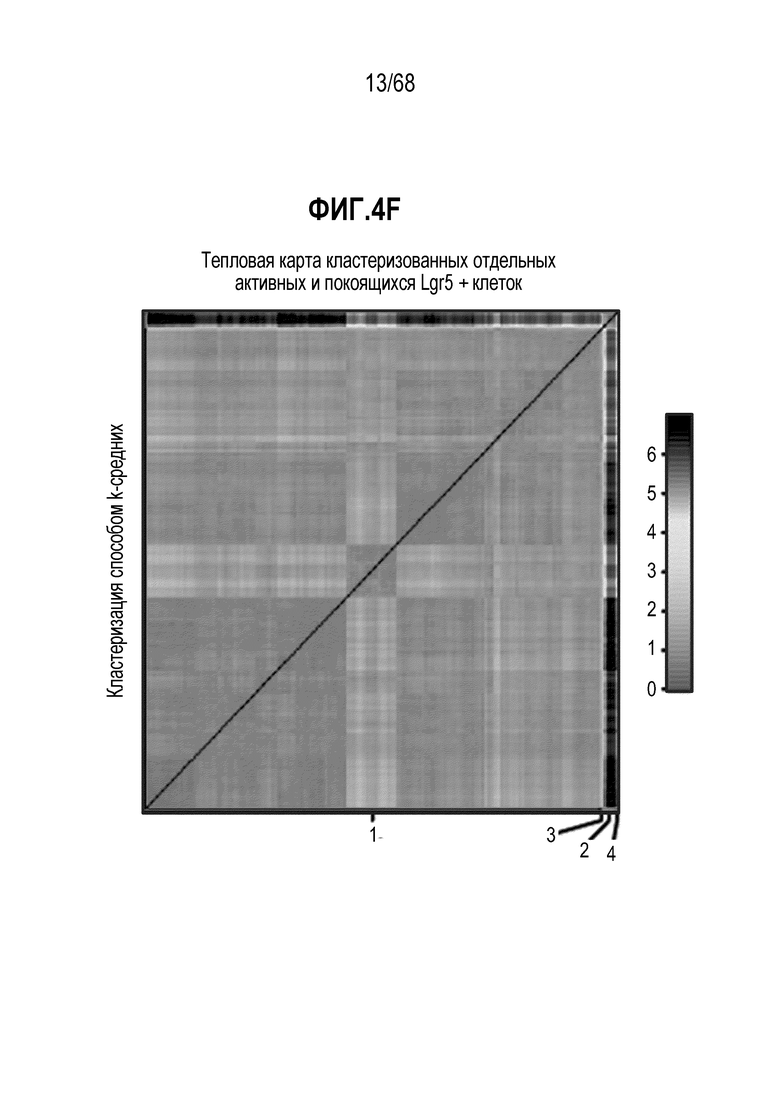
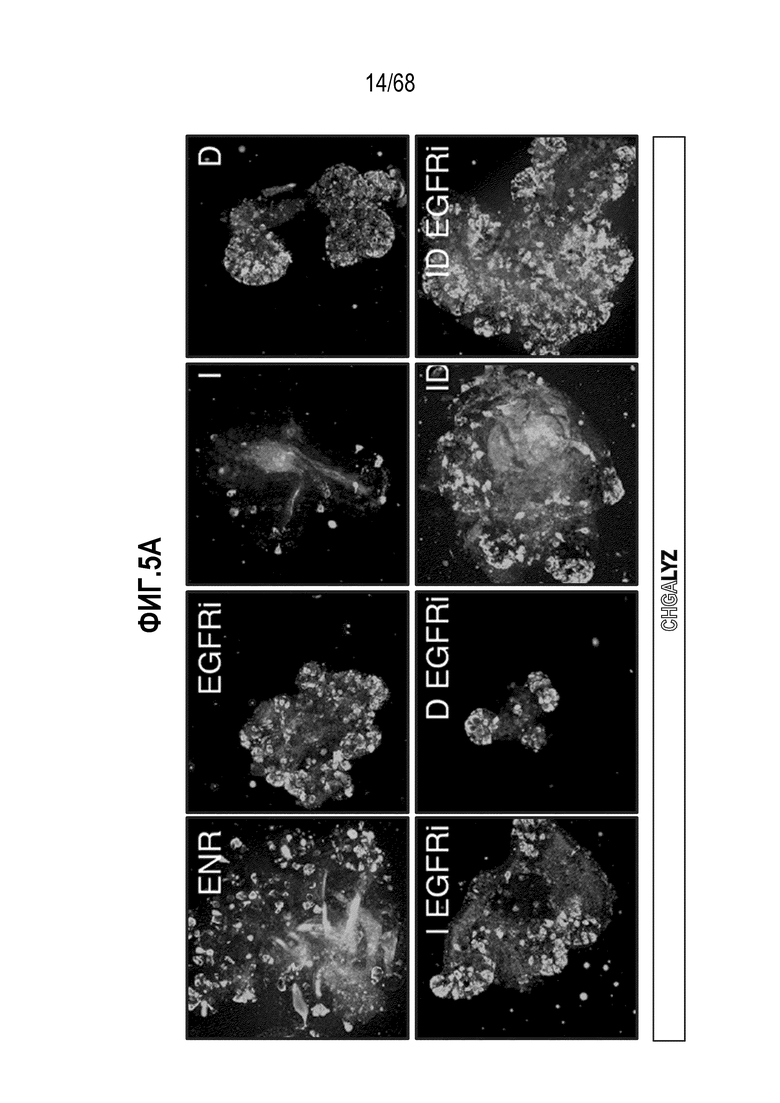
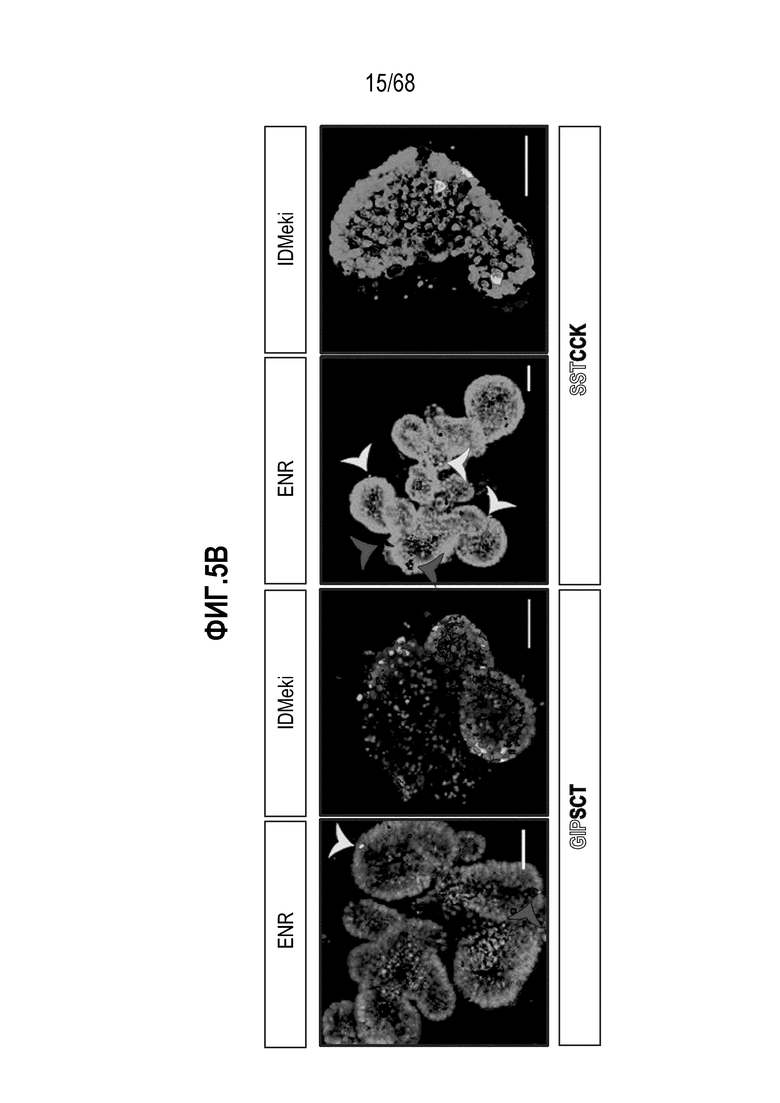
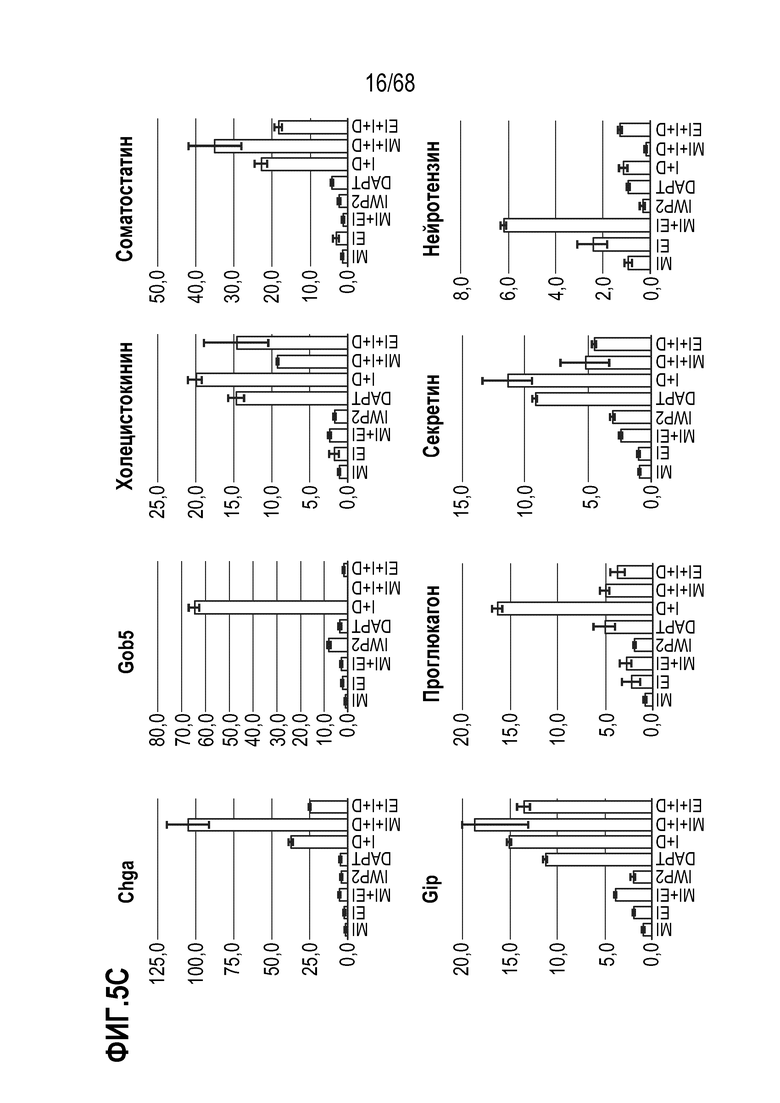
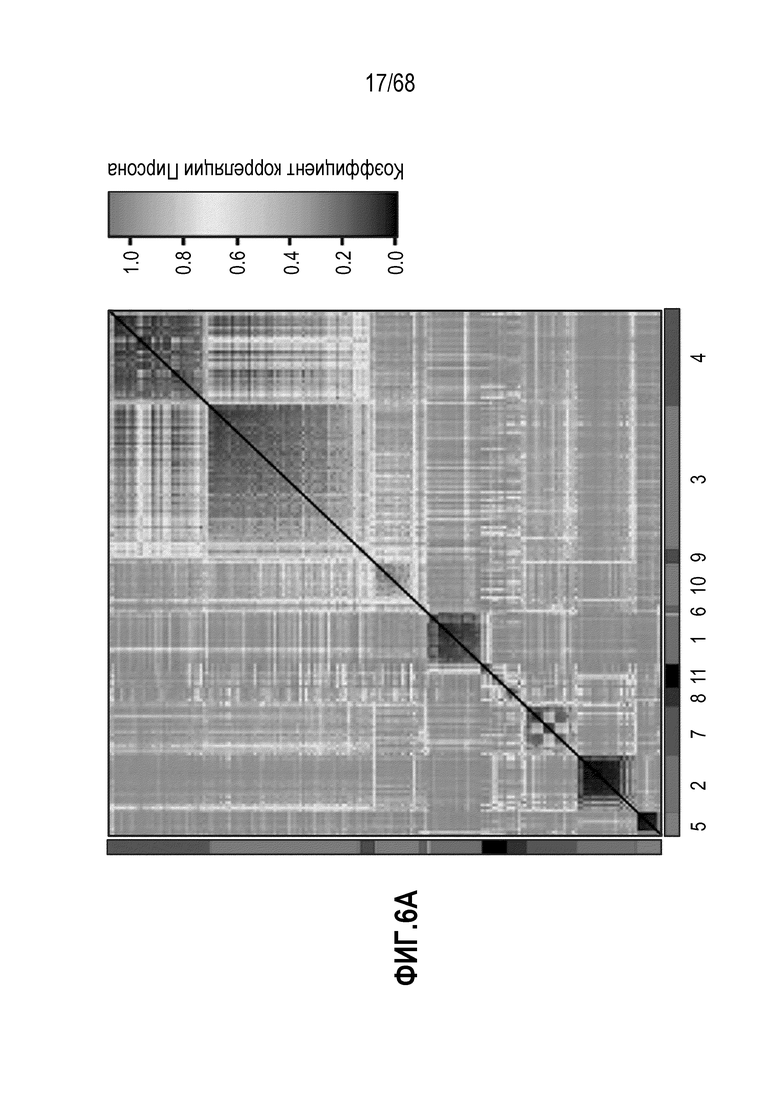
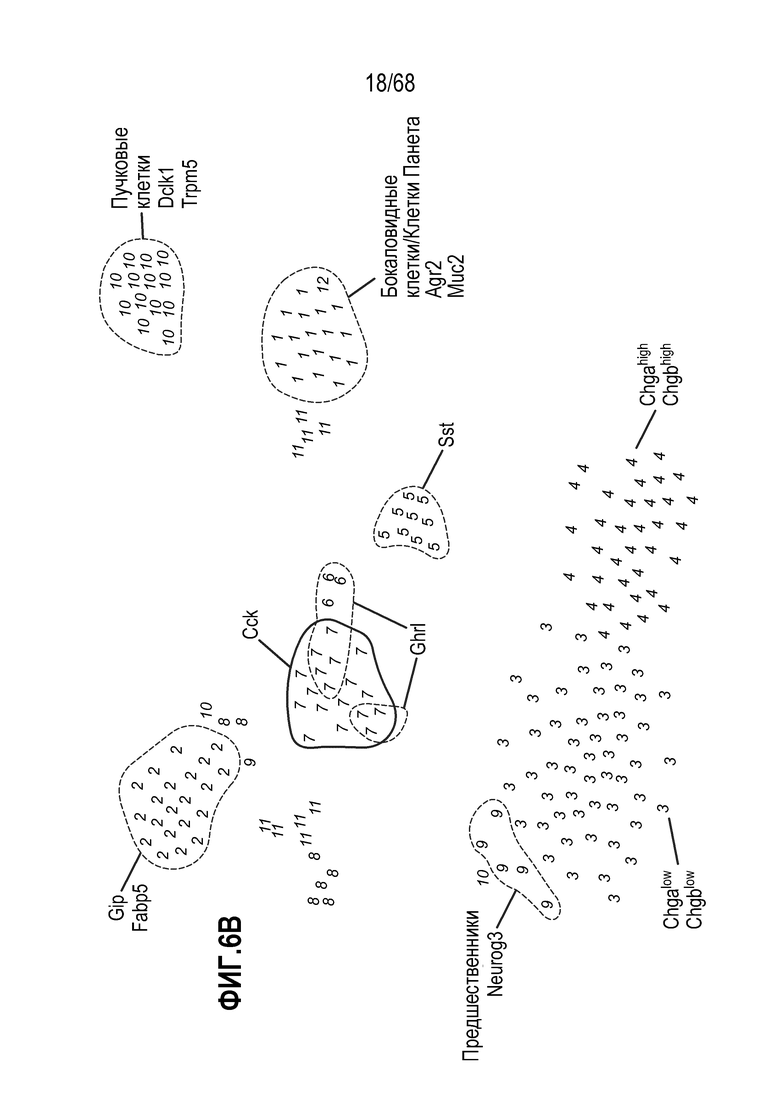




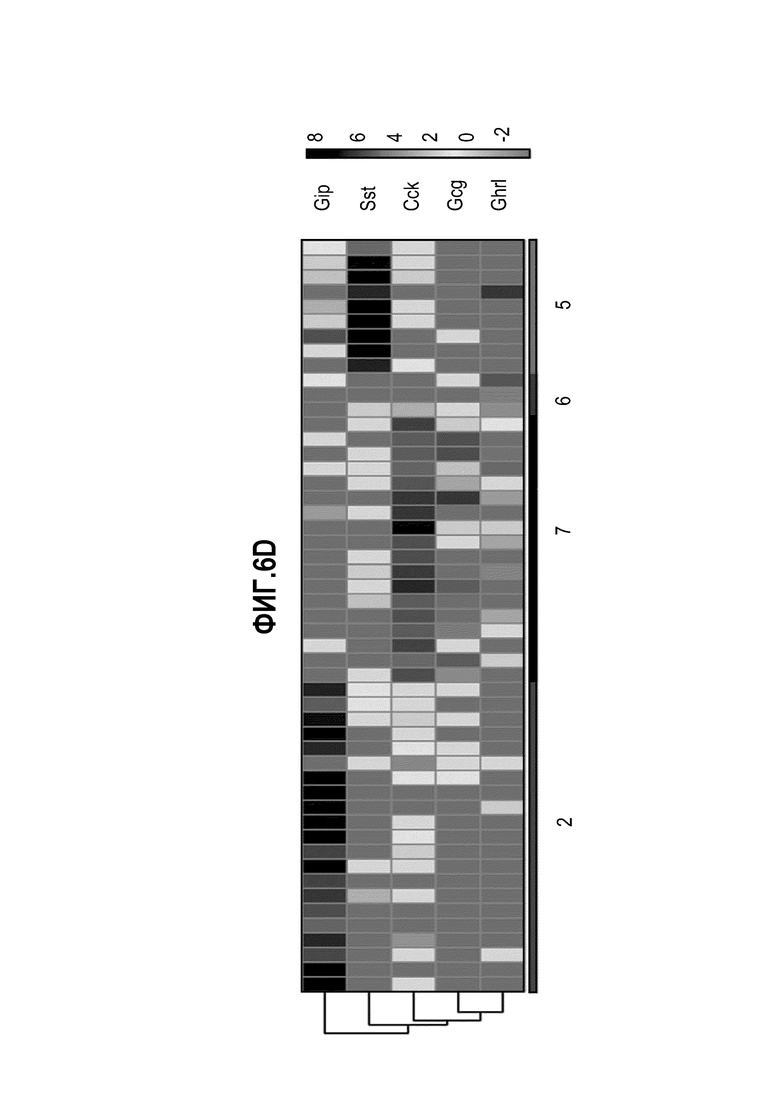

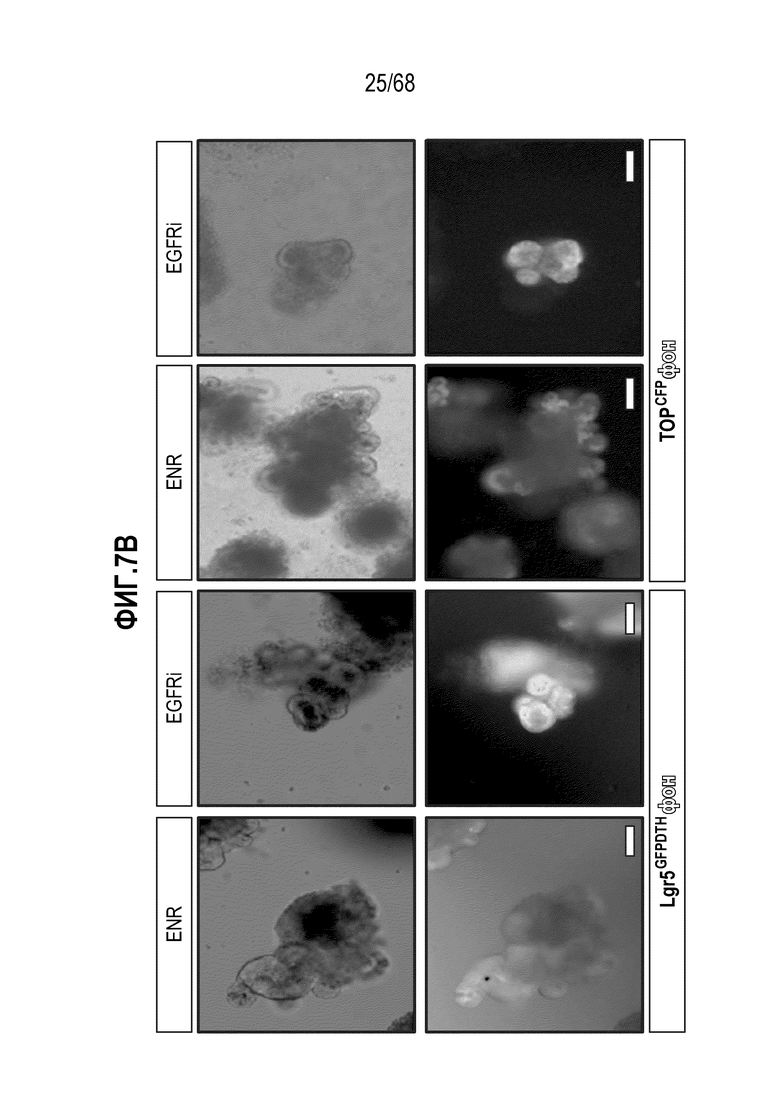
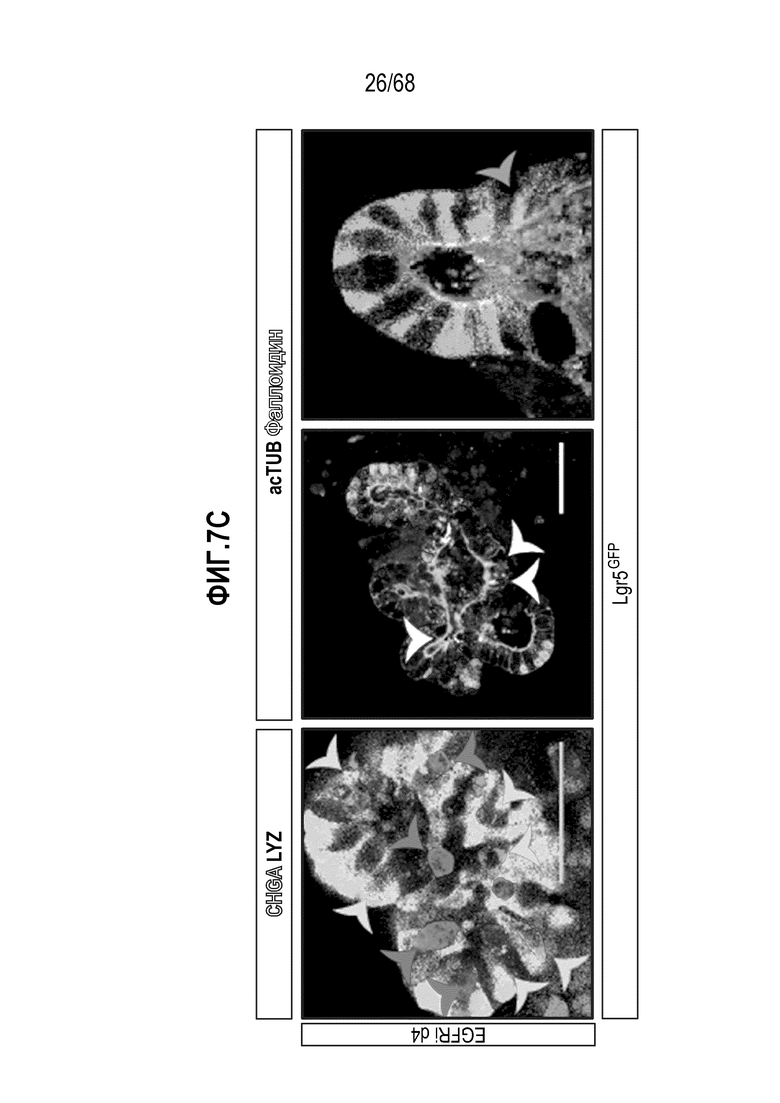
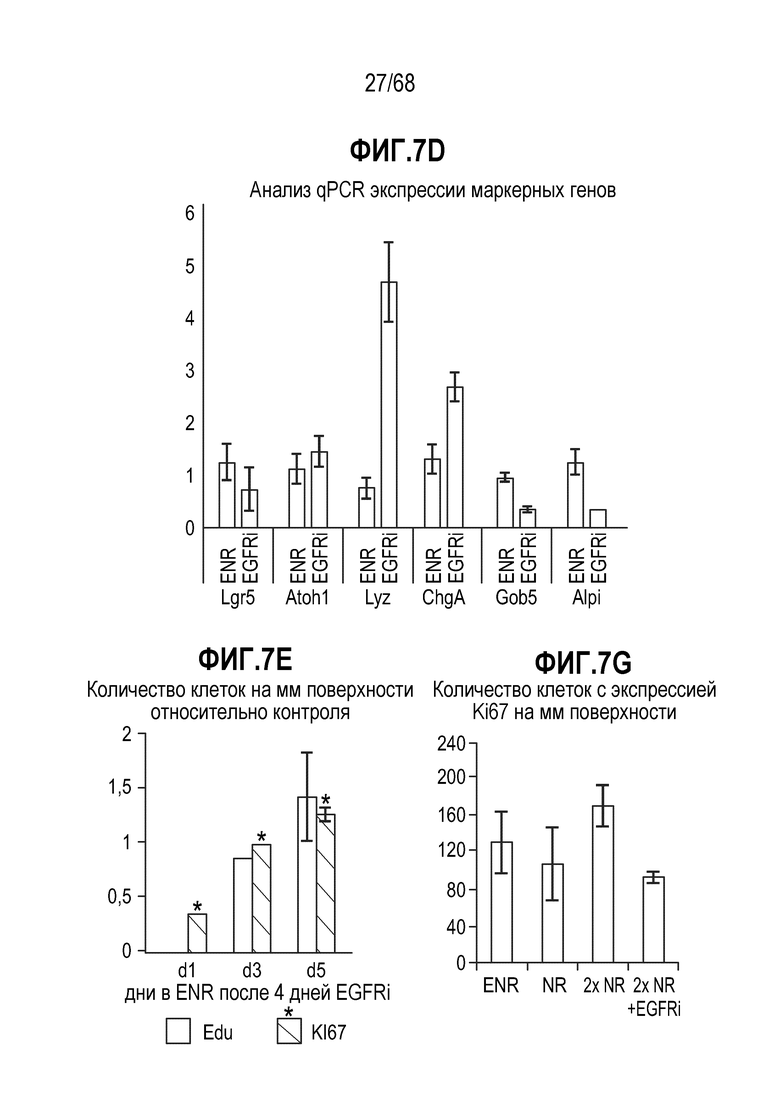
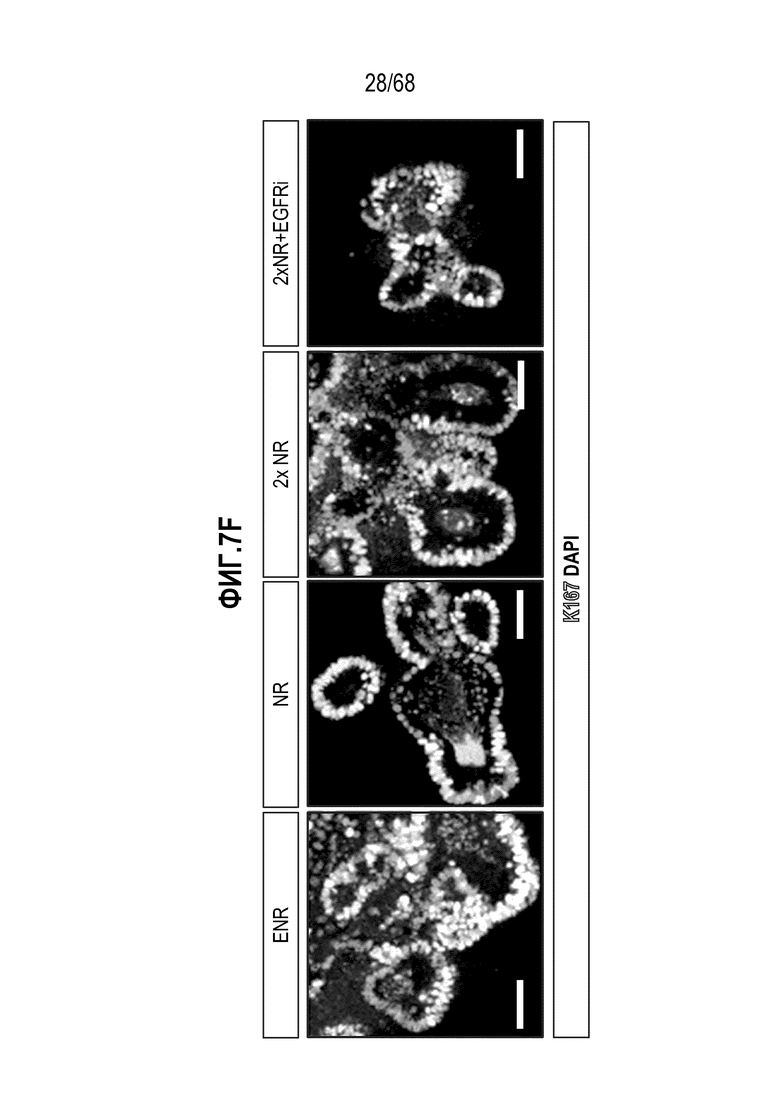
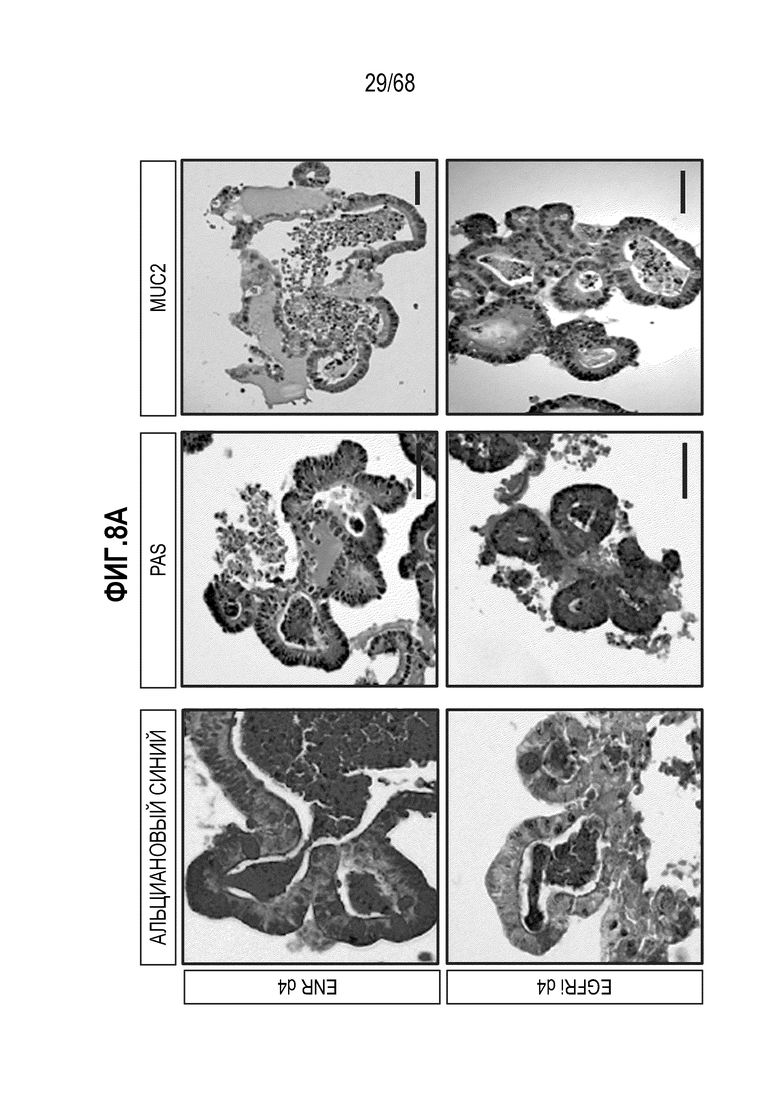
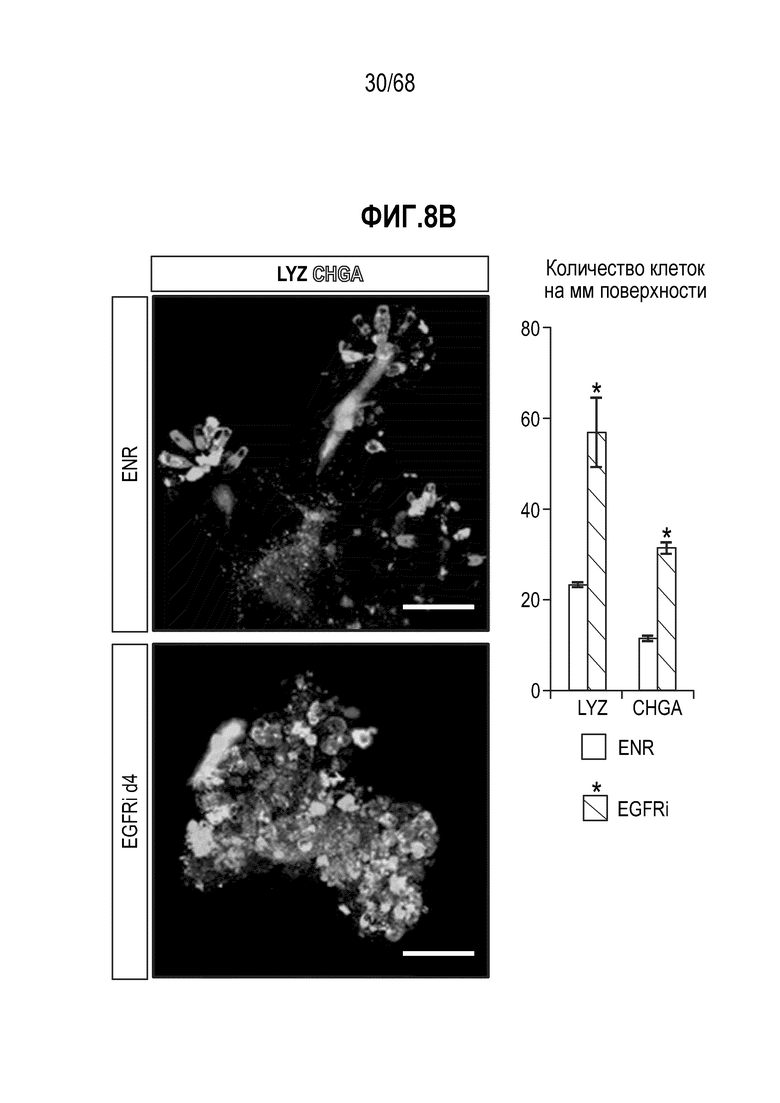

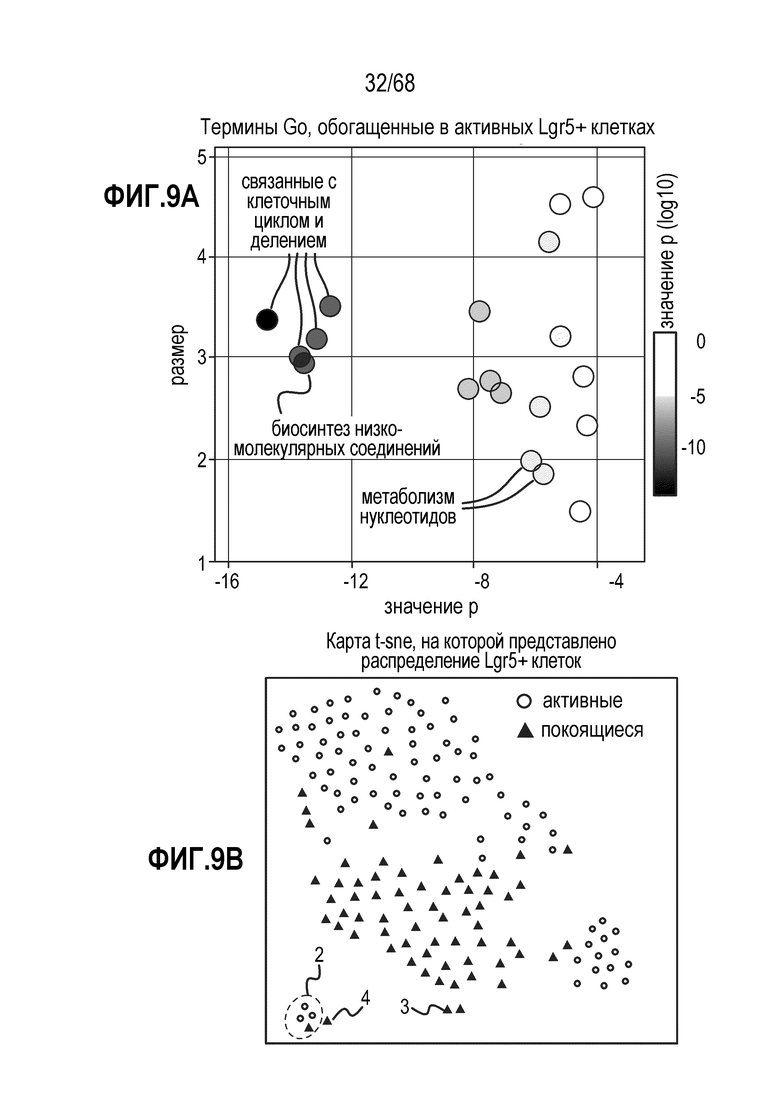

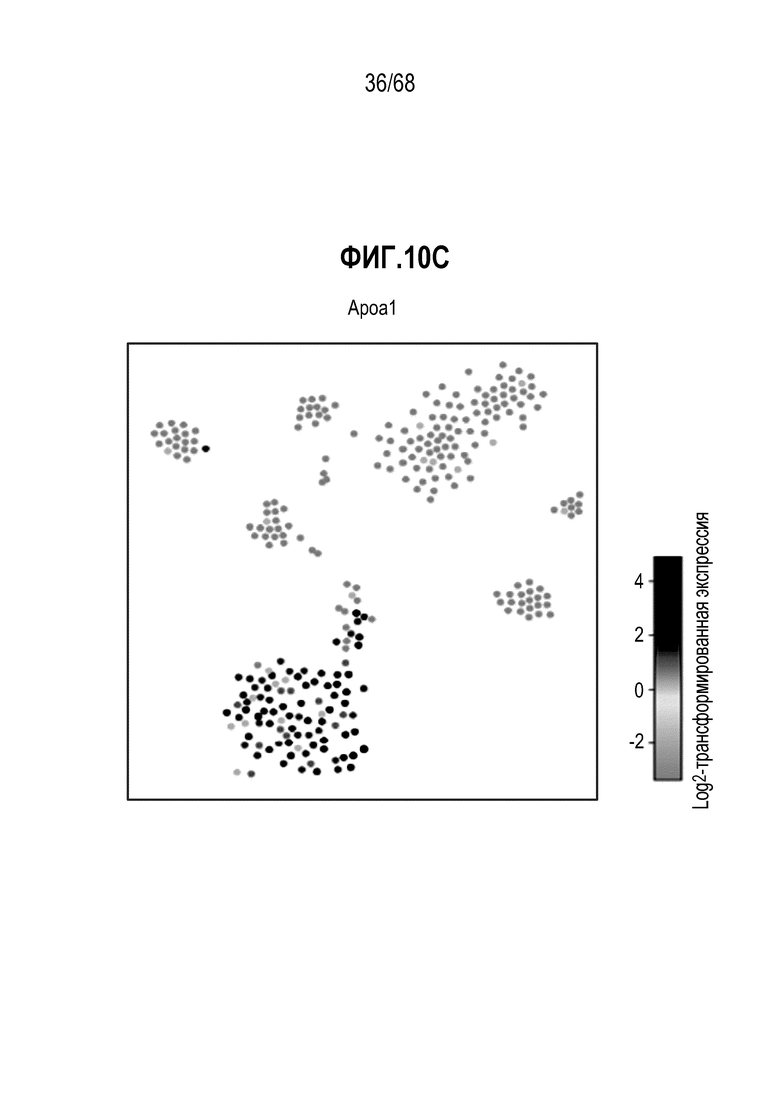
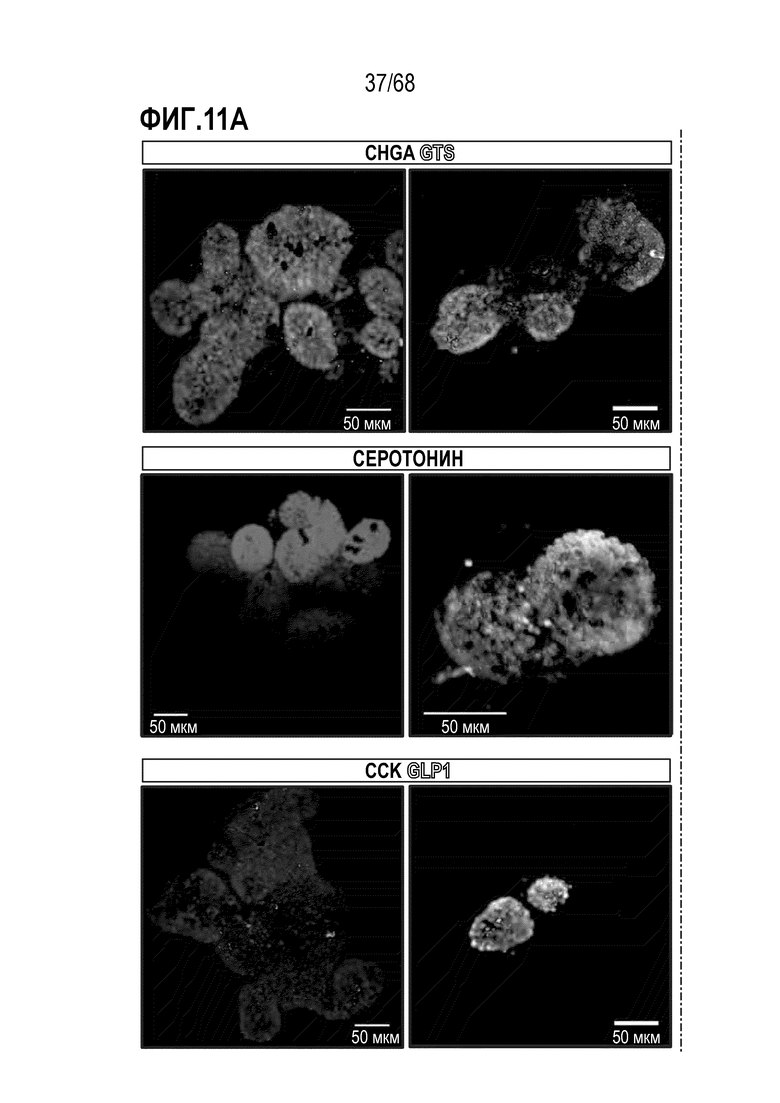
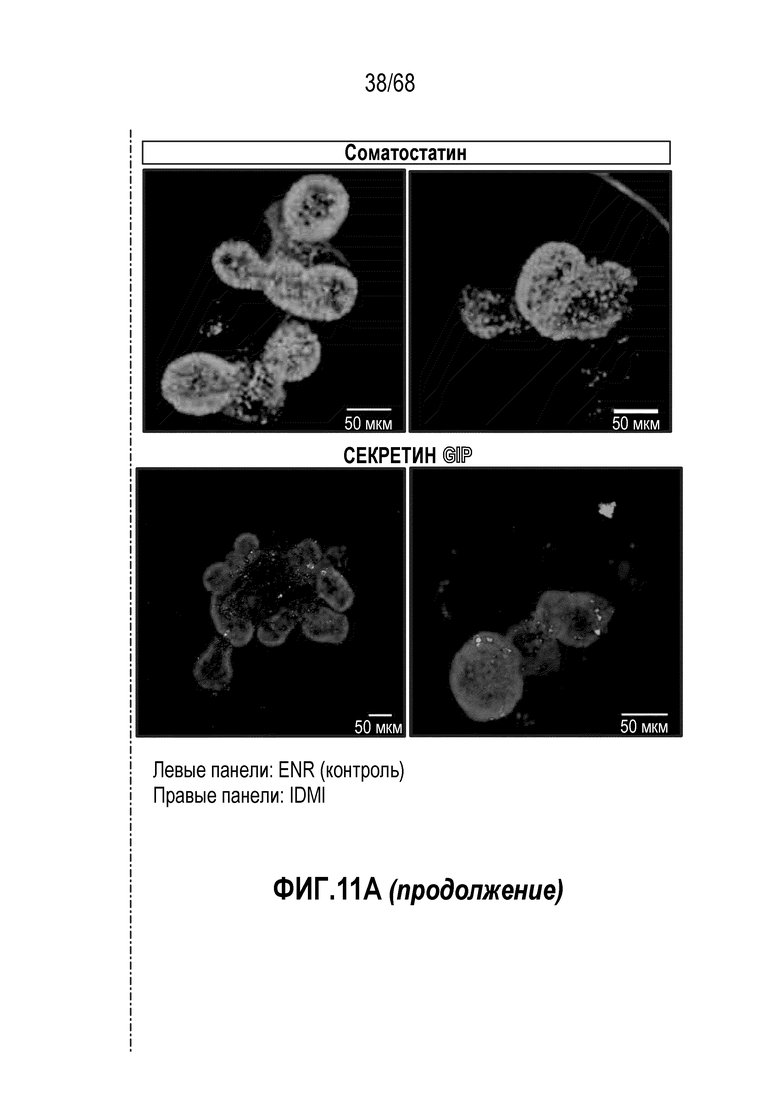
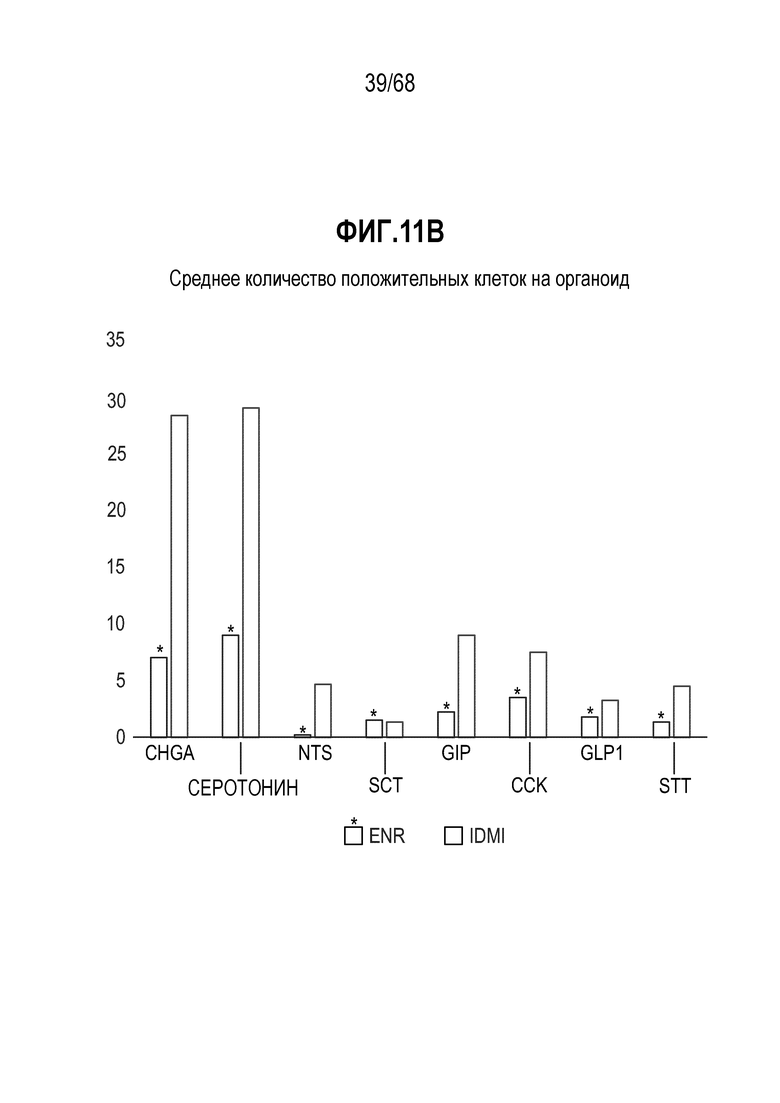
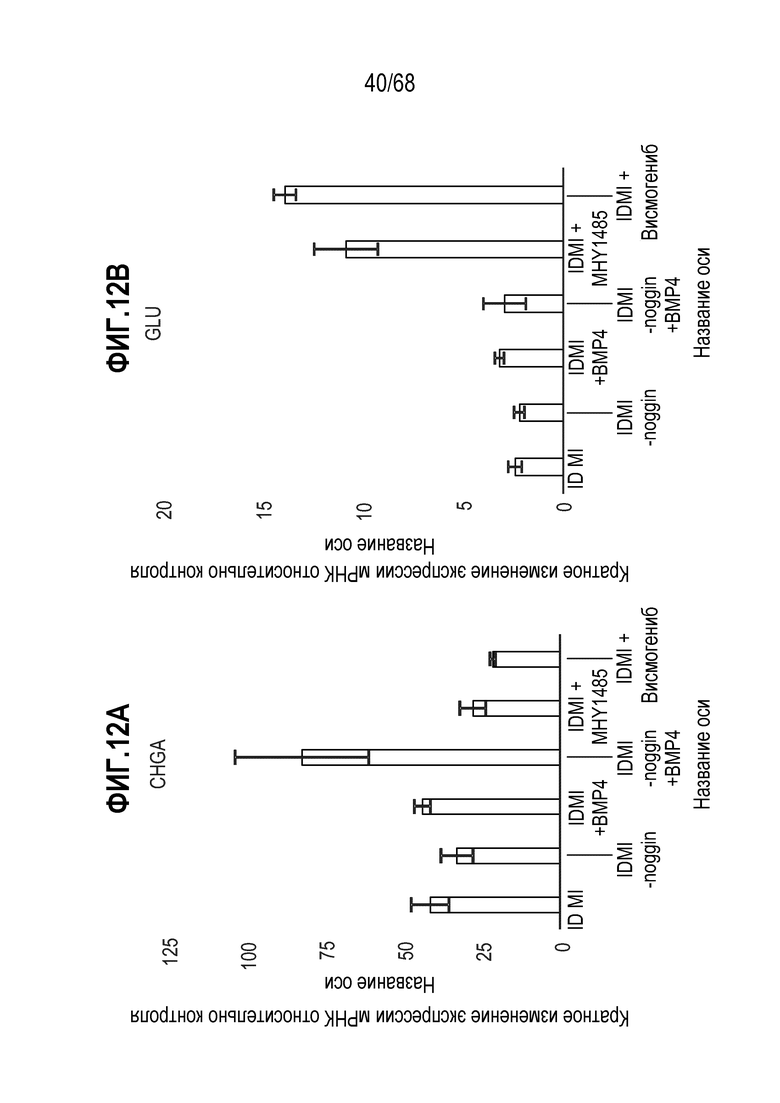
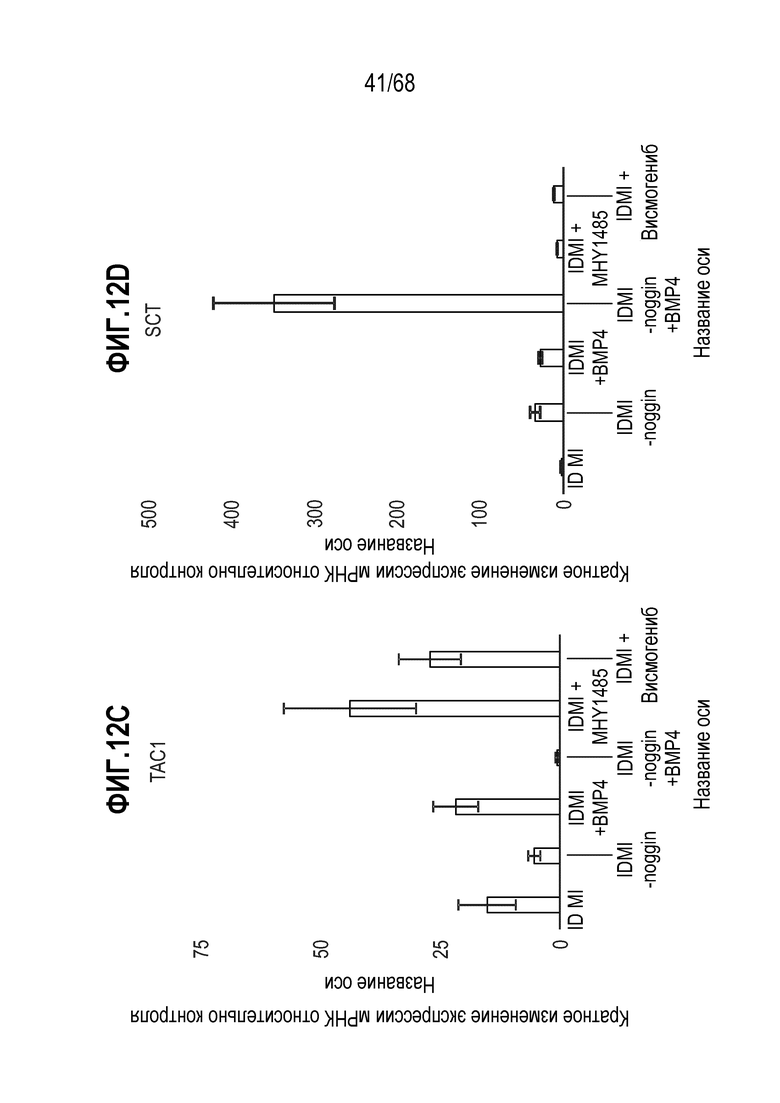
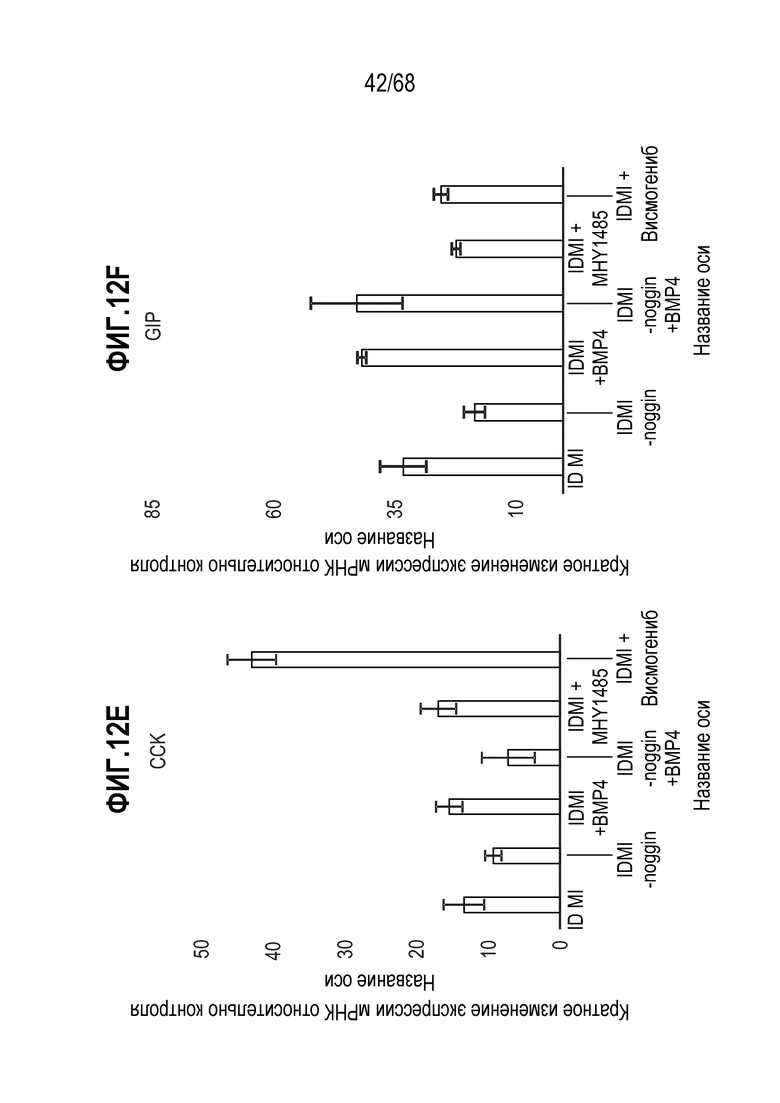
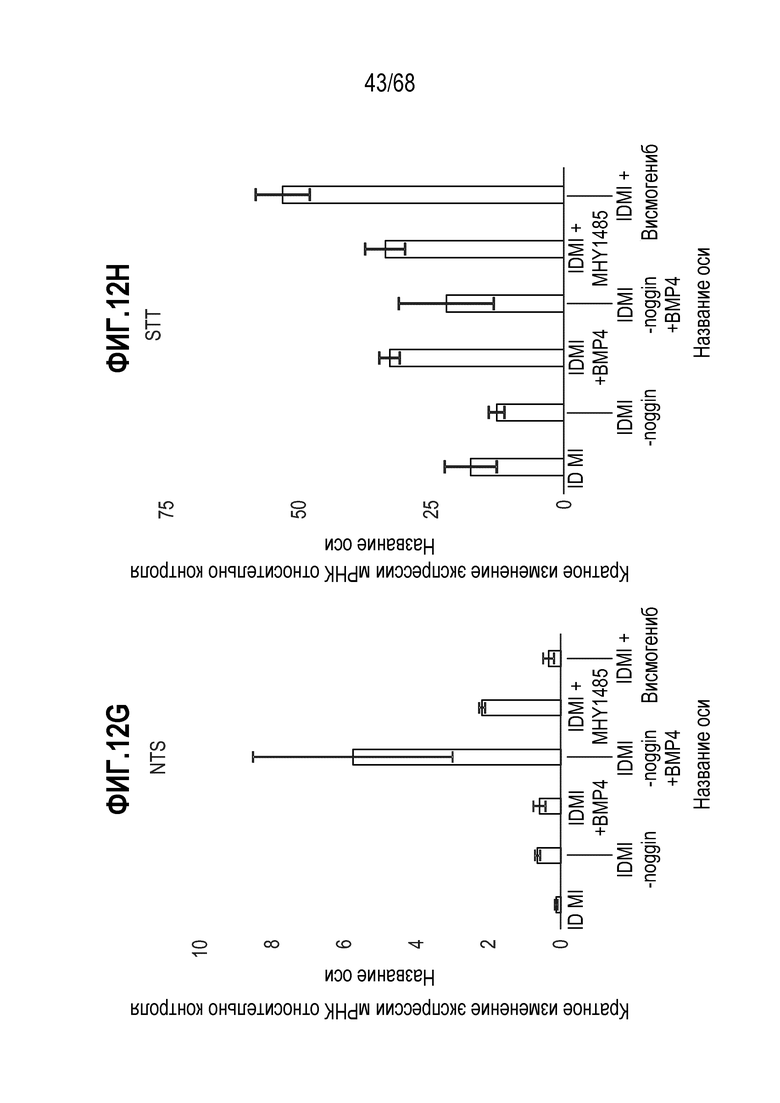
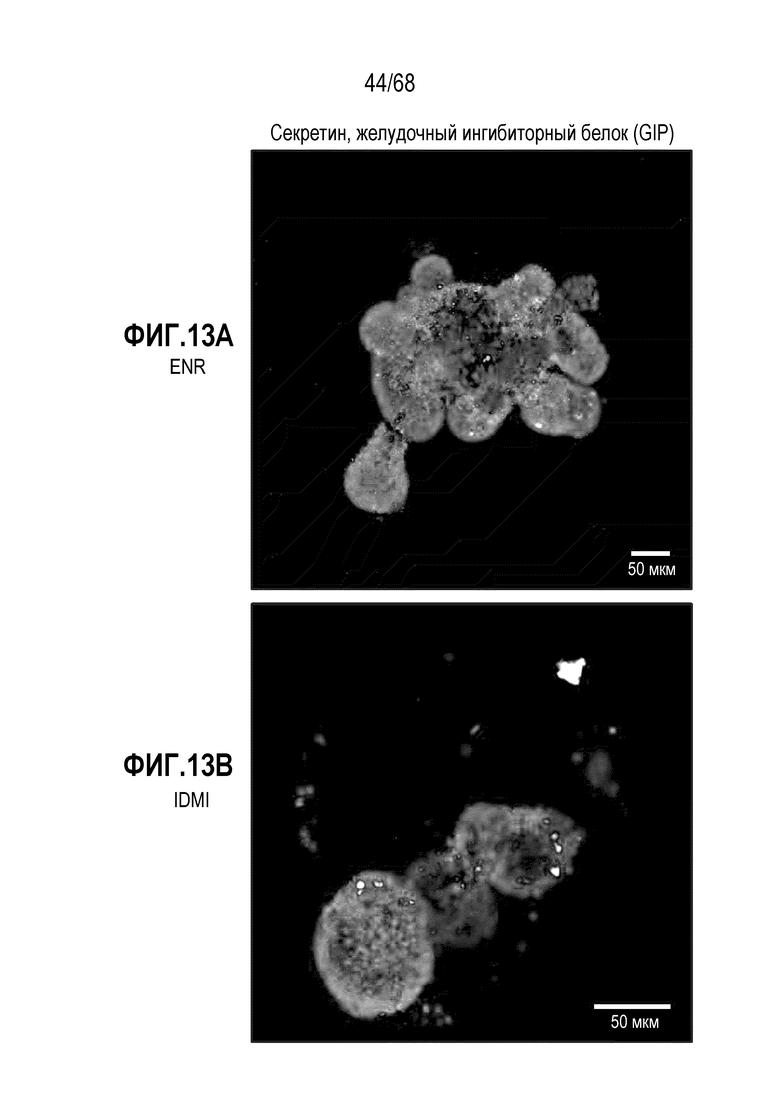
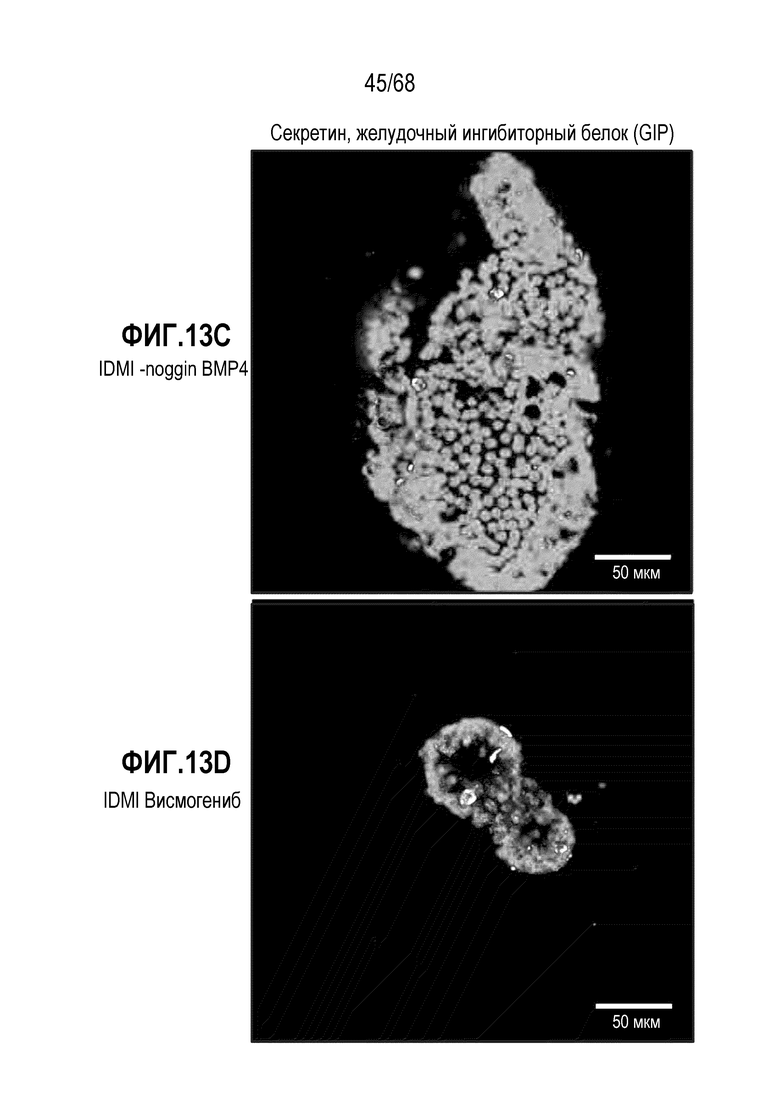
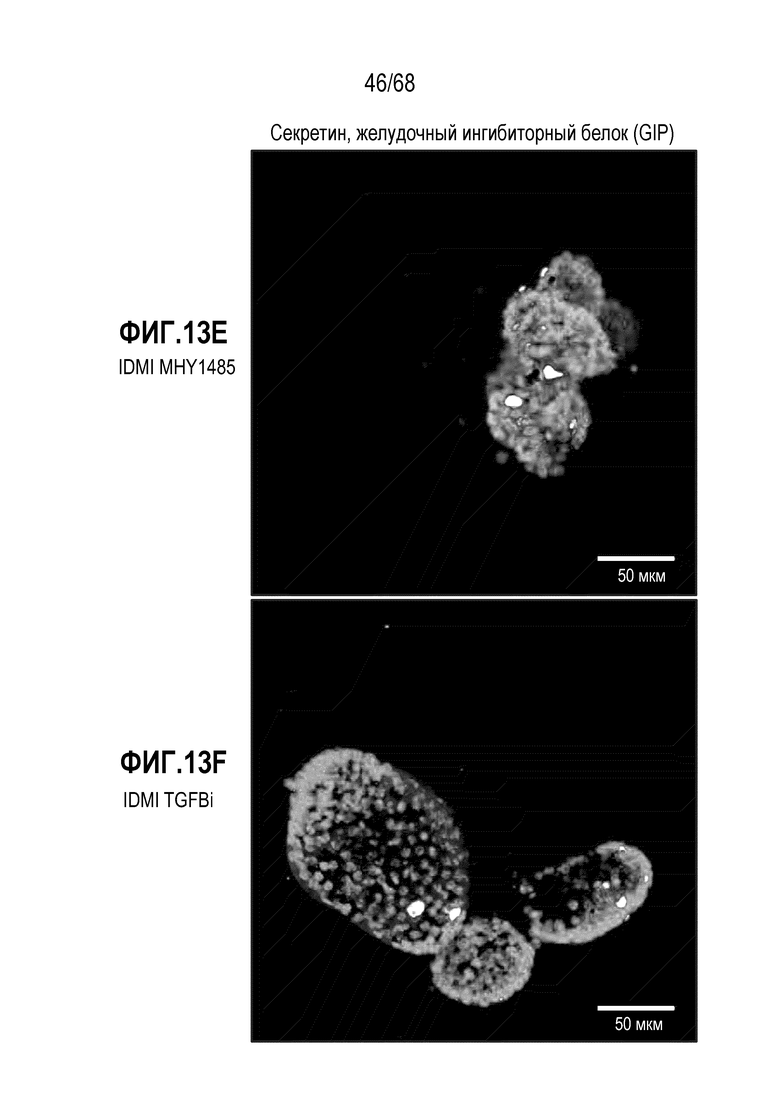
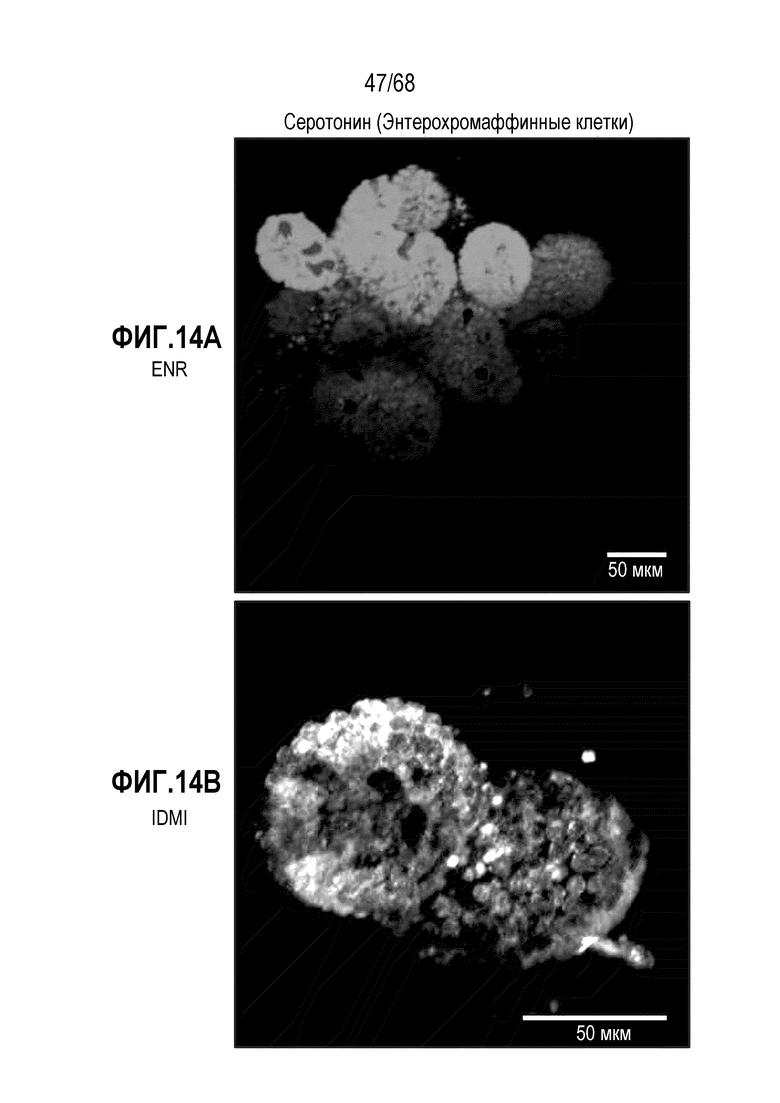
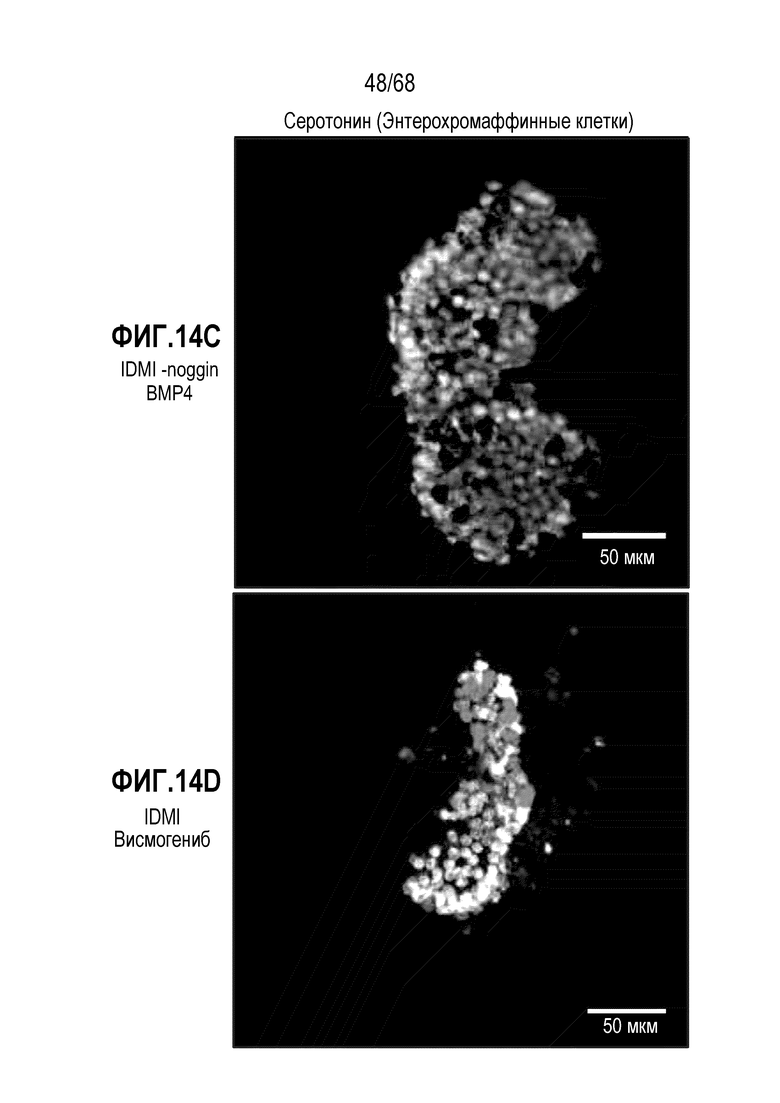
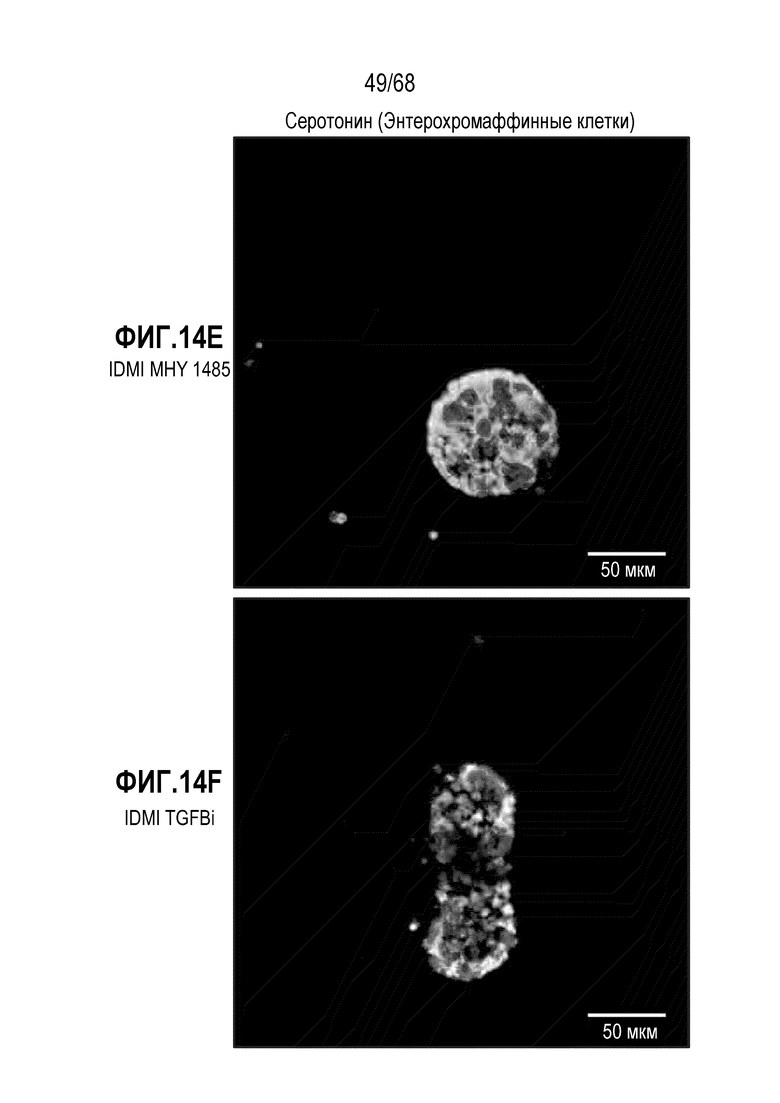
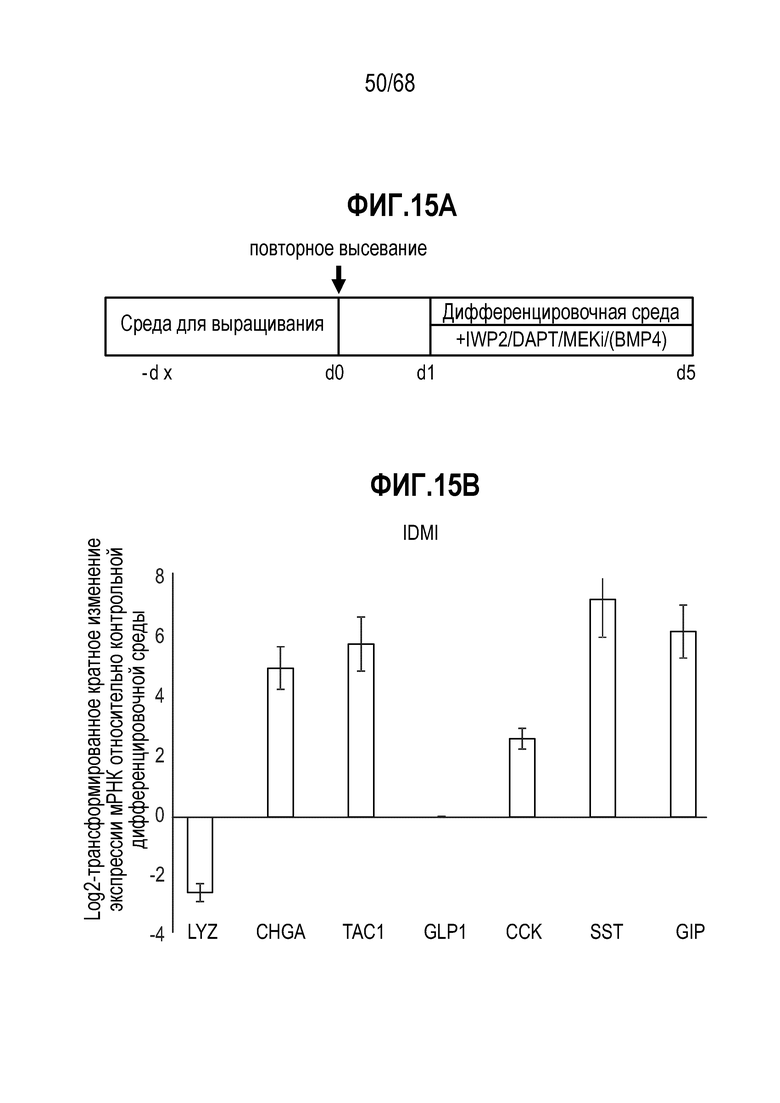
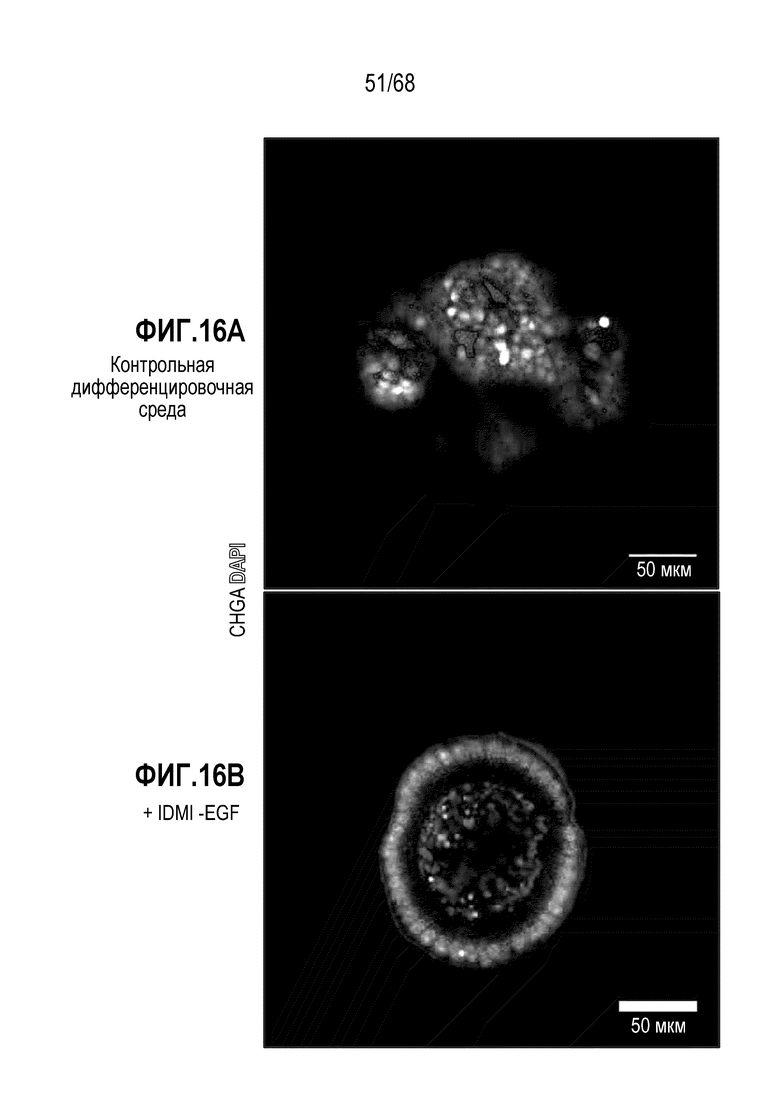
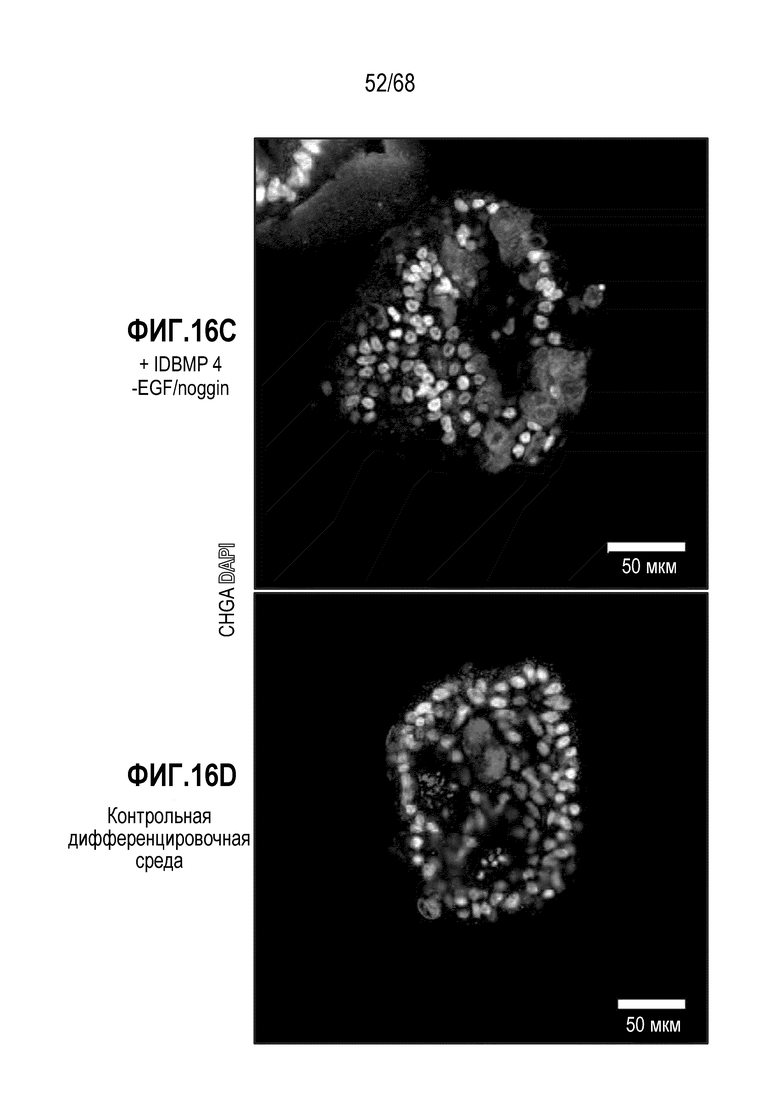
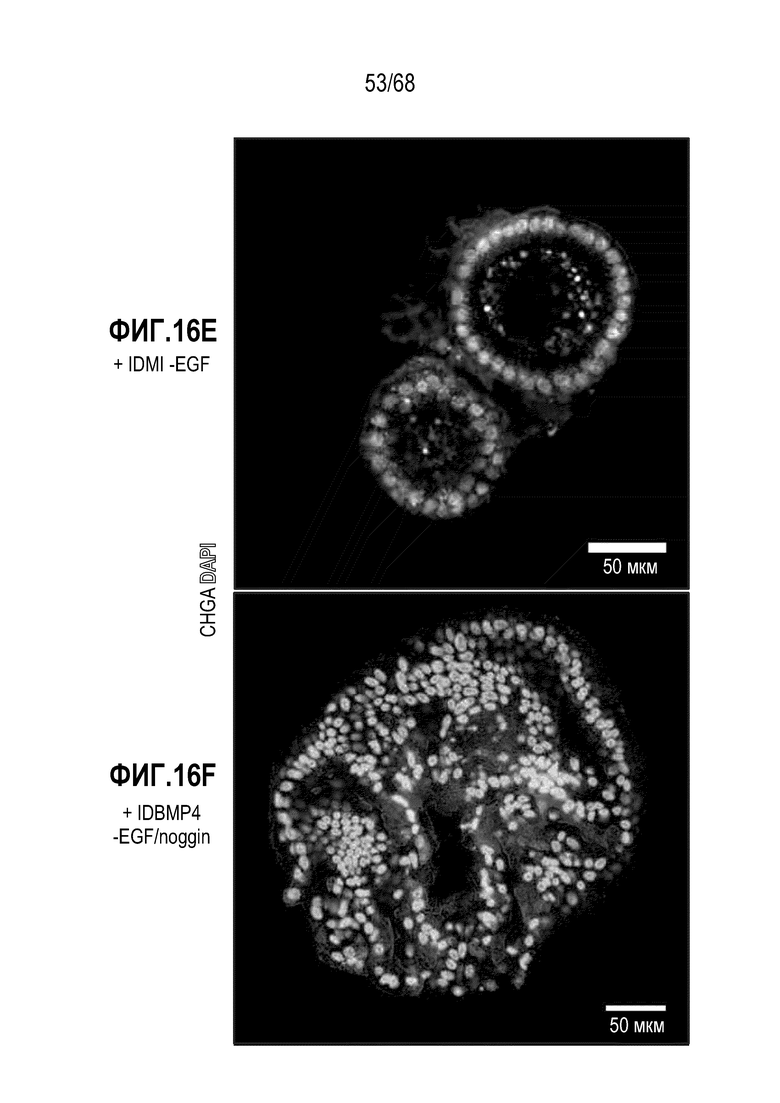
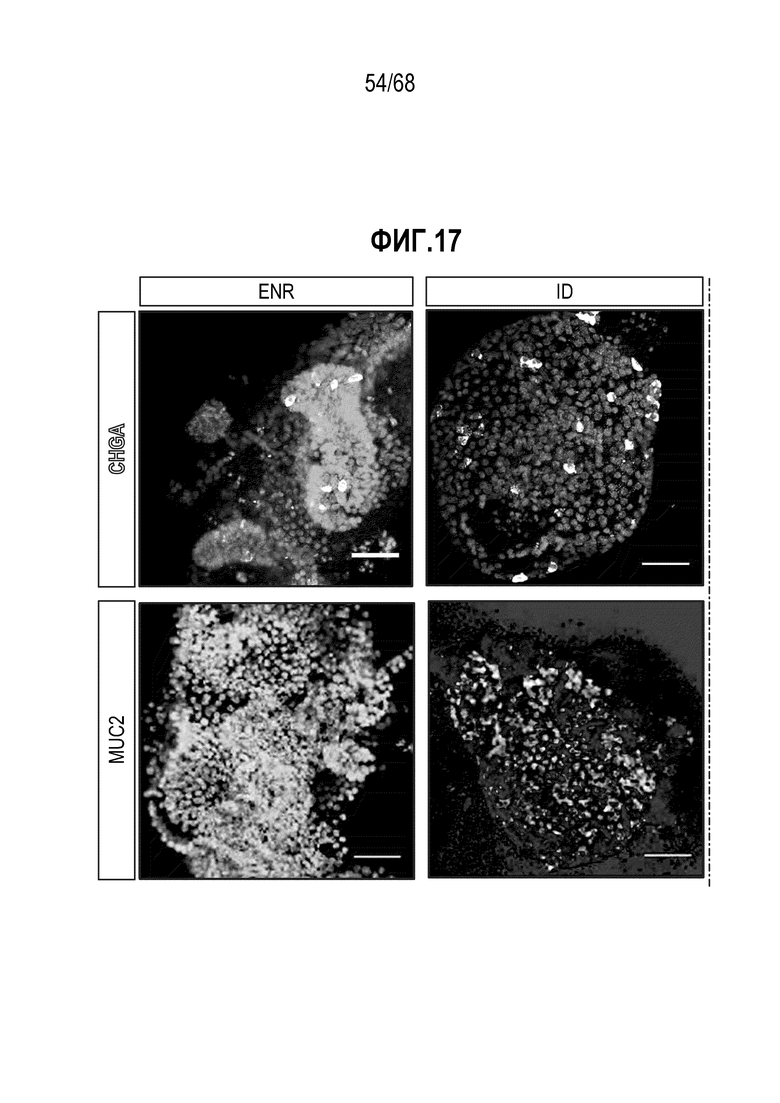
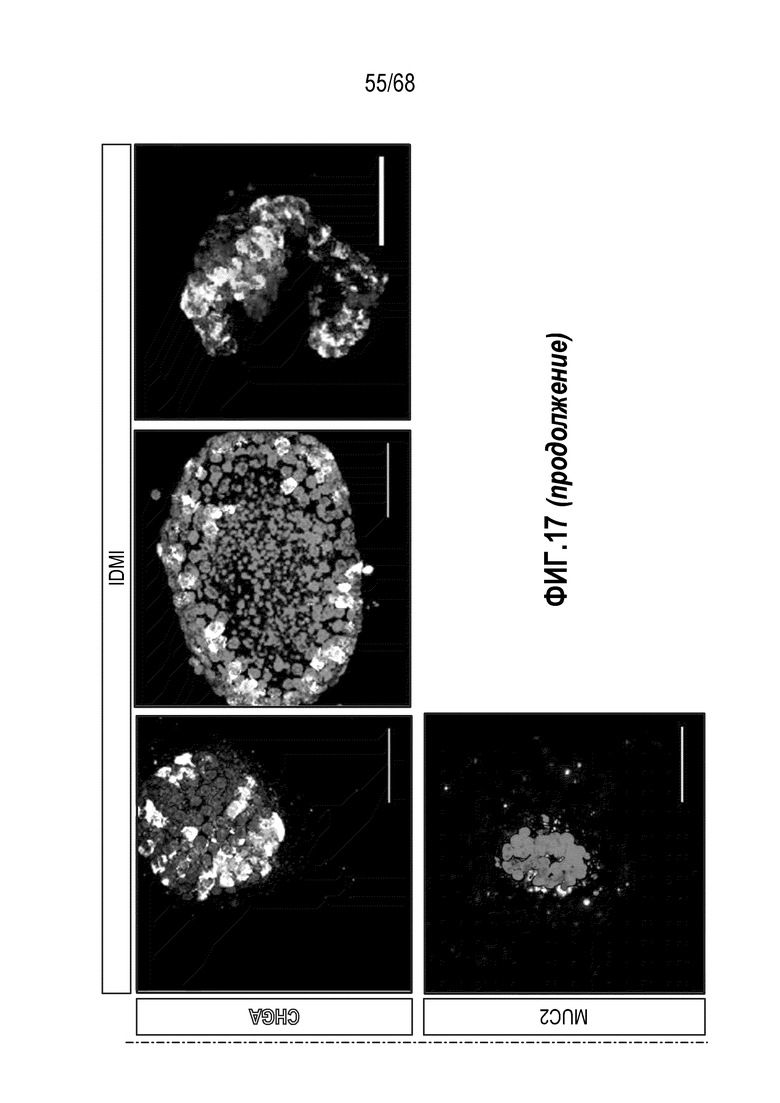
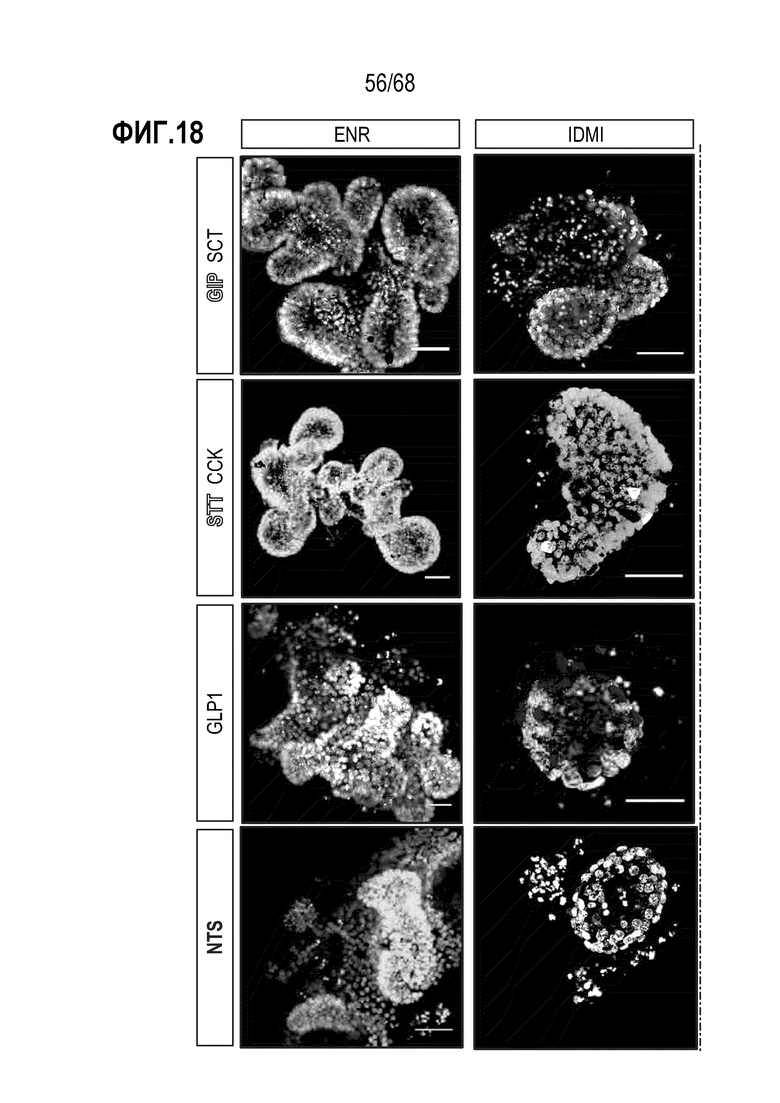
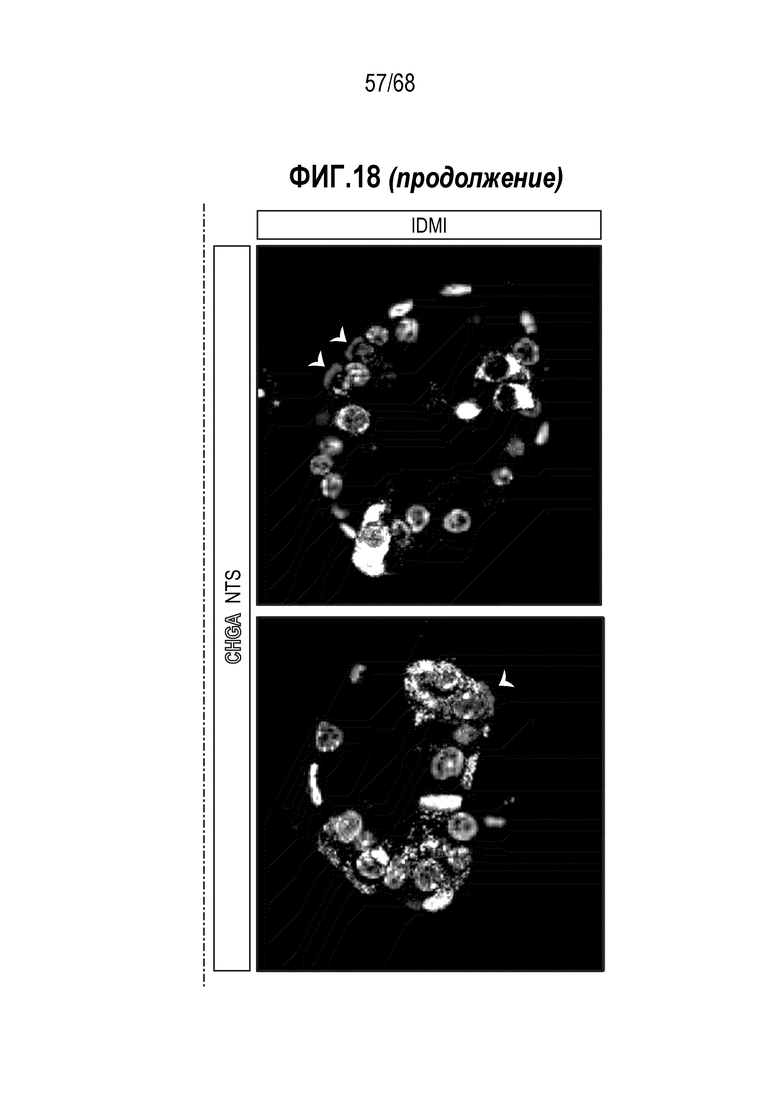
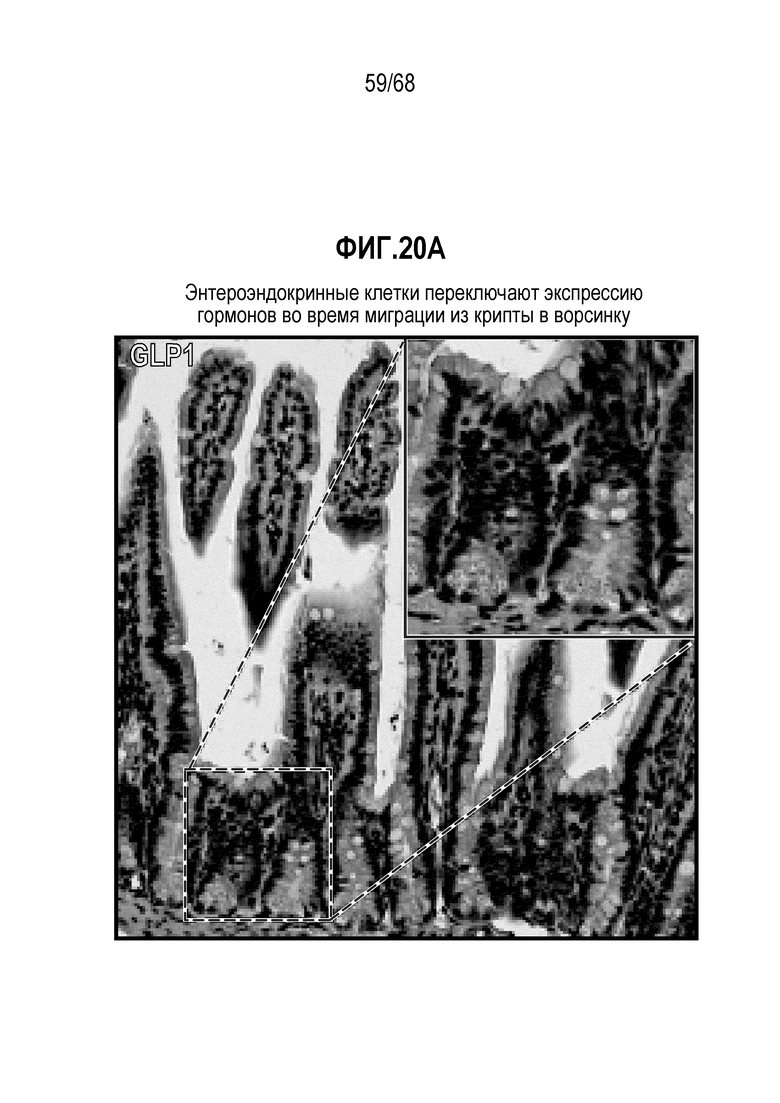

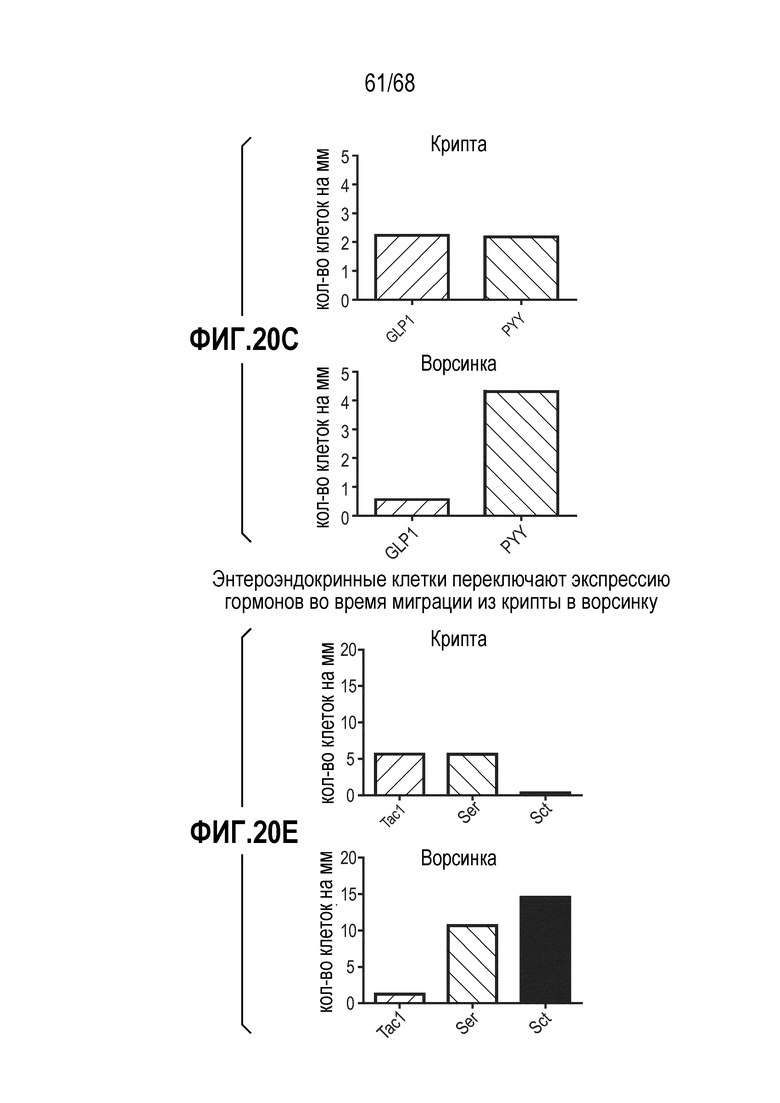
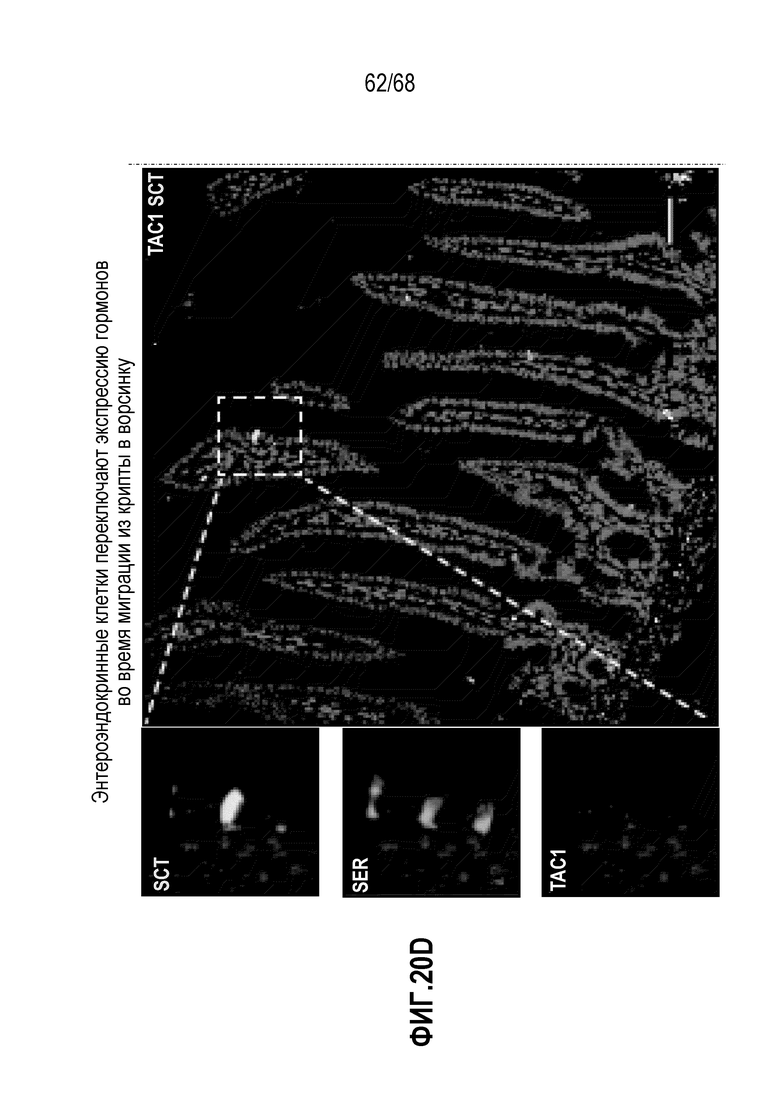
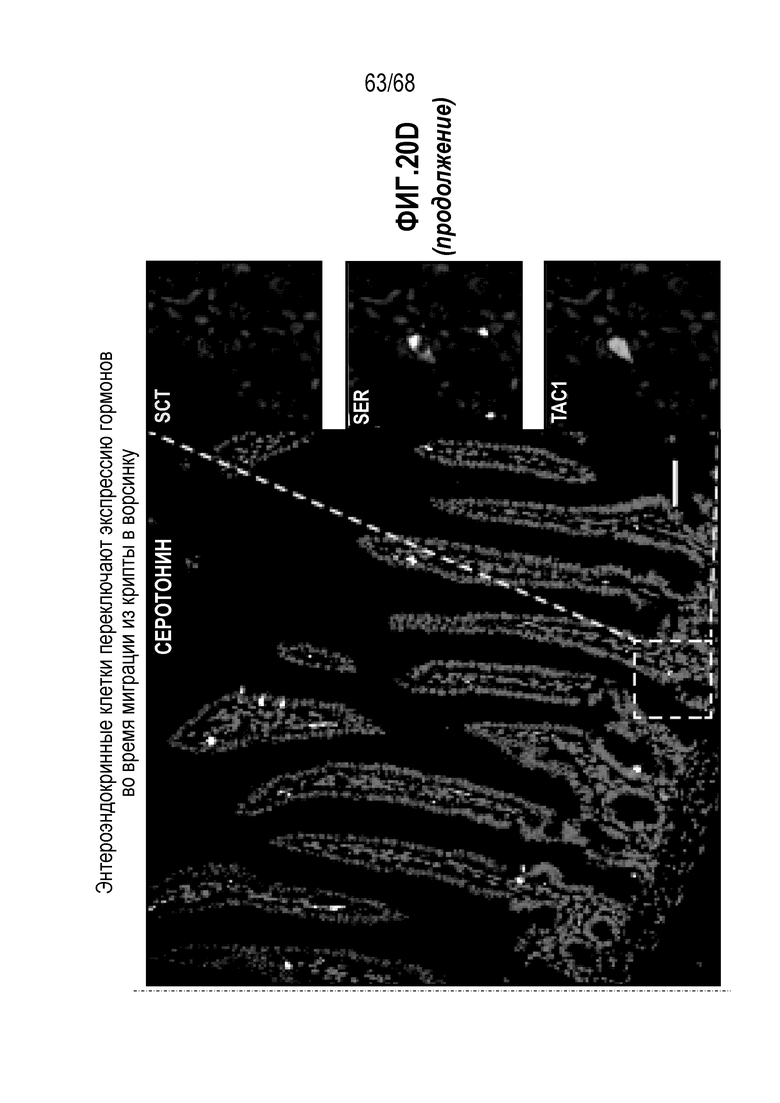
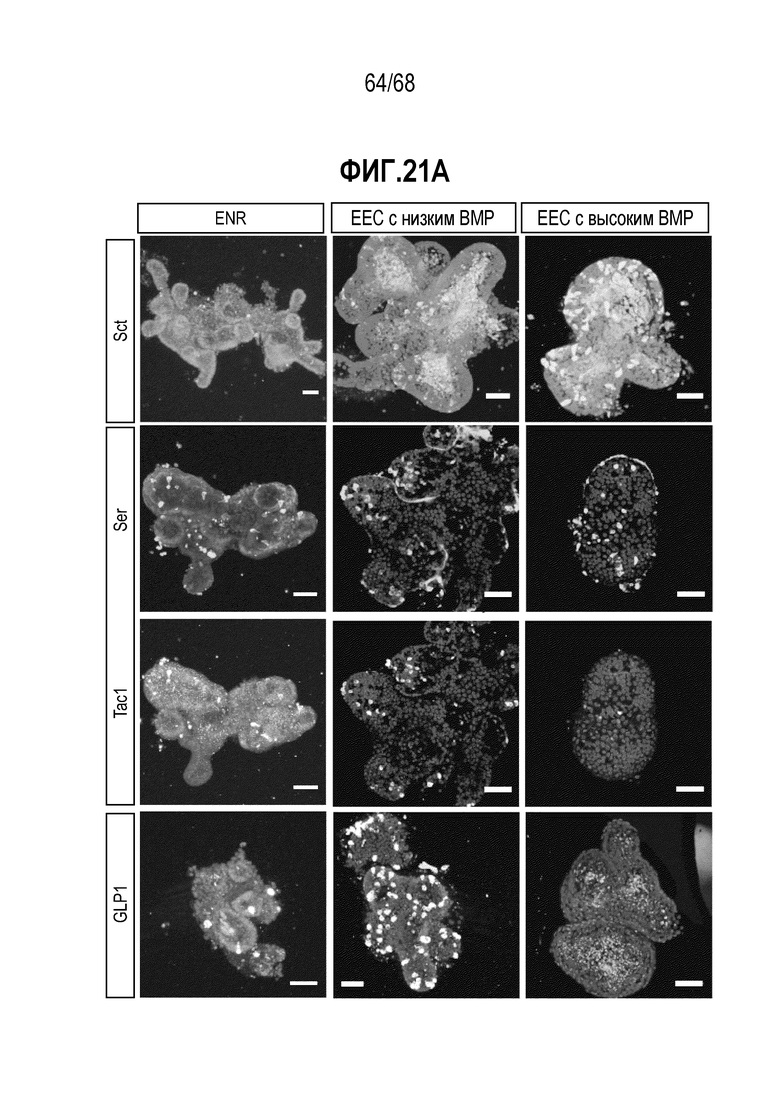

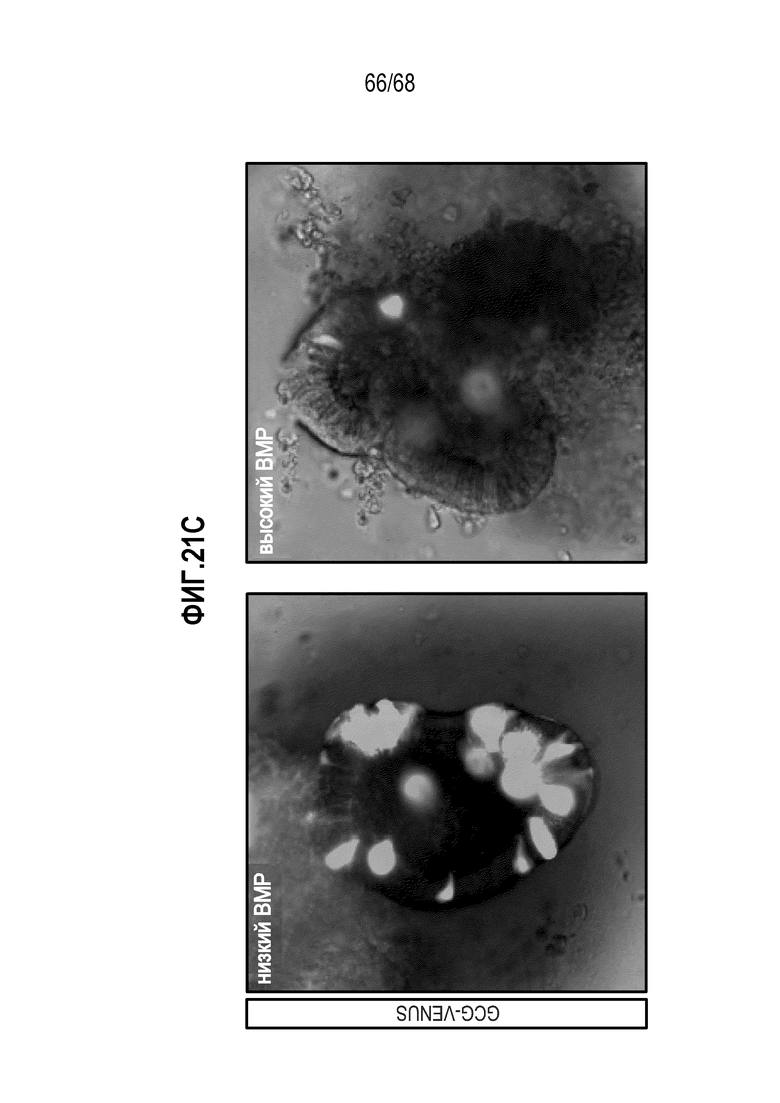
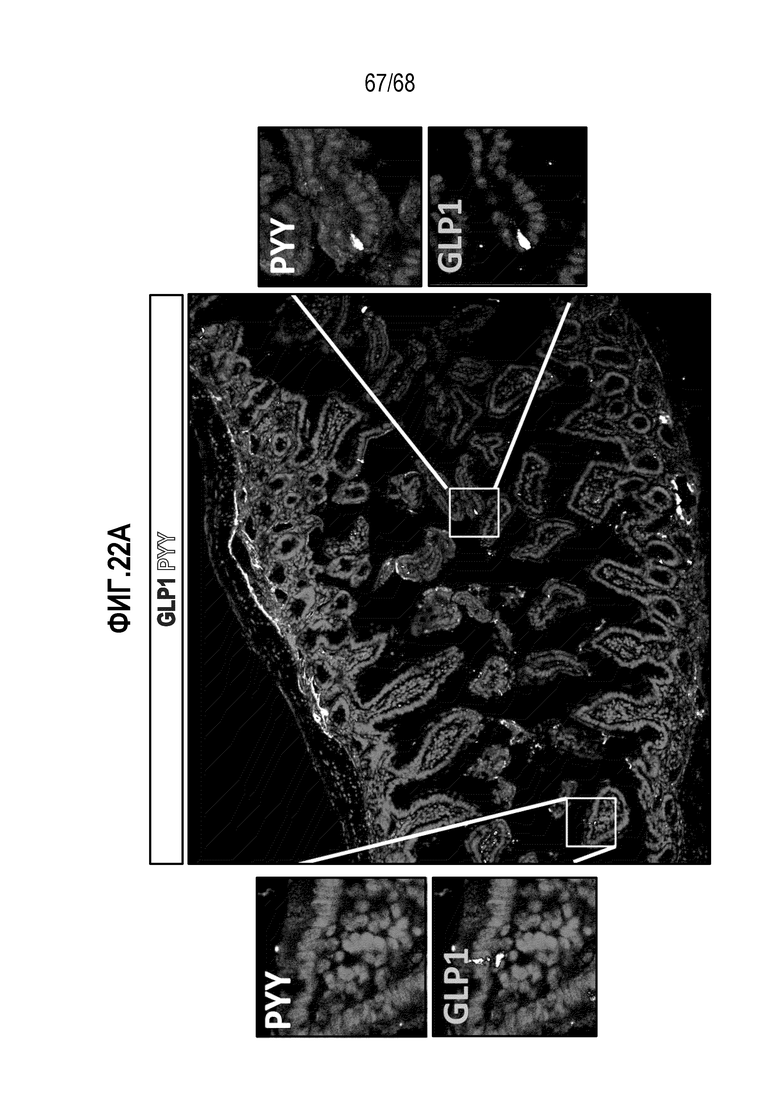
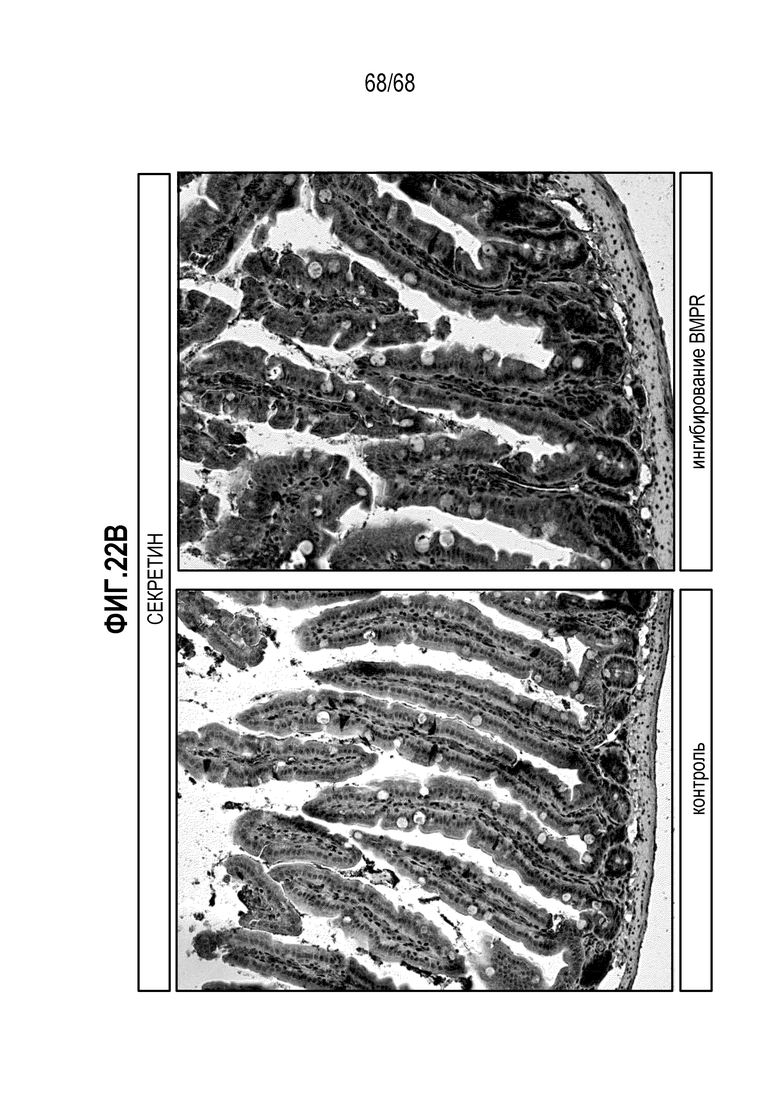
Изобретение относится к области клеточной биологии и медицине, в частности к способам дифференцировки клеток-предшественников с целью получения аргентофильных клеток. Способы осуществляют с применением особой дифференцировочной среды, содержащей минимальную среду и дополнительно содержащей один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt. Способы используют для получения трёхмерных органоидов, которые могут быть использованы в различных областях медицины, в частности в скрининге для обнаружения лекарственных средств и других, а также в различных исследованиях, в частности в исследованиях эмбриологии ткани, линий клеток и путей дифференцировки и других. Изобретение также раскрывает способы скрининга лекарственных или косметических средств, а также способы получения популяции клеток, обогащенной аргентофильными клетками, в частности GLP1-секретирующими или секретин-секретирующими клетками. Изобретение позволяет повысить эффективность дифференцировки клеток-предшественников в направлении аргентофильных клеток. 22 н. и 29 з.п. ф-лы, 87 ил., 7 табл., 6 пр.
1. Способ дифференцировки эпителиальных клеток-предшественников, включающий:
культивирование клеток в дифференцировочной среде, содержащей минимальную среду и дополнительно содержащей один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt,
где эпителиальные клетки-предшественники не происходят из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
2. Дифференцировочная среда для дифференцировки эпителиальных клеток-предшественников, где дифференцировочная среда содержит минимальную среду и дополнительно содержит один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt.
3. Способ по п. 1 или дифференцировочная среда по п. 2, где один или несколько ингибиторов пути EGFR выбраны из: (1) ингибитора EGFR, (2) ингибитора EGFR и ErbB2, (3) ингибитора пути RAS-RAF-MAPK, (4) ингибитора пути PI3K/AKT и (5) ингибитора пути JAK/STAT.
4. Способ или дифференцировочная среда по п. 3, где один или несколько ингибиторов пути EGFR включают ингибитор EGFR.
5. Способ или дифференцировочная среда по п. 4, где ингибитор EGFR представляет собой гефитиниб.
6. Способ или дифференцировочная среда по любому из пп. 3-5, где один или несколько ингибиторов пути EGFR включают ингибитор EGFR и ErbB-2.
7. Способ или дифференцировочная среда по п. 6, где ингибитор EGFR и ErbB-2 представляет собой афатиниб.
8. Способ или дифференцировочная среда по п. 3, где один или несколько ингибиторов пути EGFR включают ингибитор пути RAS-RAF-MAPK.
9. Способ или дифференцировочная среда по п. 8, где один или несколько ингибиторов пути EGFR включают ингибитор MEK и/или ингибитор ERK.
10. Способ или дифференцировочная среда по п. 9, где ингибитор MEK представляет собой PD0325901 и/или ингибитор ERK представляет собой SCH772984.
11. Способ или дифференцировочная среда по любому из пп. 1-10, где ингибитор Notch является ингибитором гамма-секретазы.
12. Способ или дифференцировочная среда по п. 11, где ингибитором гамма-секретазы является DAPT или дибензодиазепин (DBZ), или бензодиазепин (BZ), или LY-411575.
13. Способ или дифференцировочная среда по любому из пп. 1-12, где один или несколько ингибиторов Wnt выбраны из: (1) ингибитора секреции Wnt, (2) конкурентного или неконкурентного ингибитора взаимодействия между Wnt или R-спондином и рецепторным комплексом Wnt, (3) ингибитора, стимулирующего деградацию компонентов рецепторного комплекса Wnt, (4) ингибитора белков семейства Dishevelled, (5) активатора, стимулирующего активность деструктивного комплекса, (6) ингибитора деолигомеризации деструктивного комплекса и/или (7) ингибитора экспрессии гена-мишени β-катенина.
14. Способ или дифференцировочная среда по п. 13, где один или несколько ингибиторов Wnt включают ингибитор секреции Wnt.
15. Способ или дифференцировочная среда по п. 14, где ингибитор секреции Wnt представляет собой ингибитор Porc, выбранный из IWP 2, LGK974 и IWP 1.
16. Способ или дифференцировочная среда по любому из пп. 1-15, где дифференцировочная среда содержит менее 1 мМ EGF.
17. Способ или дифференцировочная среда по любому из пп. 1-16, где дифференцировочная среда дополнительно содержит
один или несколько компонентов, выбранных из группы, состоящей из: ингибитора p38, ингибитора TGF-бета, гастрина, глюкокортикоида, лиганда рецепторных тирозинкиназ, активатора пути BMP, активатора пути цАМФ, активатора Hedgehog, ингибитора Hedgehog, модулятора передачи сигнала mTOR, B27 и N2; или
один или несколько компонентов, выбранных из группы, состоящей из: ингибитора p38, ингибитора TGF-бета, гастрина, глюкокортикоида, лиганда рецепторных тирозинкиназ, ингибитора BMP, активатора пути цАМФ, активатора Hedgehog, ингибитора Hedgehog, модулятора передачи сигнала mTOR, B27 и N2; или
активатор BMP, выбранный из BMP7, BMP4 или BMP2; или
ингибитор BMP, выбранный из Noggin, склеростина, хордина, CTGF, фоллистатина, гремлина, tsg, sog, LDN193189 или дорсоморфина.
18. Способ дифференцировки клеток-предшественников кишечника для получения популяции клеток кишечника, обогащенной аргентофильными клетками, включающий:
культивирование клеток-предшественников кишечника в дифференцировочной среде по любому из пп. 2-17,
где эпителиальные клетки-предшественники кишечника не получают из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
19. Способ культивирования эпителиальных стволовых клеток, включающий:
культивирование эпителиальных стволовых клеток в присутствие среды для выращивания эпителиальных стволовых клеток для получения выращенных эпителиальных стволовых клеток; а затем
культивирование одной или нескольких выращенных клеток в дифференцировочной среде по любому из пп. 2-17,
где эпителиальные стволовые клетки не происходят из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
20. Способ по любому из пп. 1-19, где клетки культивируют в контакте с внеклеточным матриксом.
21. Способ по любому из пп. 1-20, где клетки получают из кишечника, желудка, поджелудочной железы, печени, предстательной железы, легких, молочной железы, яичника, слюнной железы, волосяного фолликула, кожи, пищевода, мочевого пузыря, уха или щитовидной железы.
22. Способ по п. 21, где клетки получают из кишечника, желудка, поджелудочной железы или легких.
23. Способ по любому из пп. 1-22, где клетки являются эпителиальными клетками-предшественниками или стволовыми клетками млекопитающих.
24. Способ по п. 23, отличающийся тем, что клетки-предшественники или стволовые клетки млекопитающих представляют собой клетки-предшественники или стволовые клетки человека.
25. Способ по любому из пп. 1-24, который дополнительно включает получение и/или выделение популяции дифференцированных клеток или дифференцированного органоида.
26. Способ культивирования эпителиальных стволовых клеток кишечника, включающий:
культивирование одной или нескольких эпителиальных стволовых клеток кишечника, приведенных в контакт с внеклеточным матриксом, в присутствие среды для выращивания; а затем
культивирование одной или нескольких выращенных эпителиальных стволовых клеток кишечника, приведенных в контакт с внеклеточным матриксом, в присутствие дифференцировочной среды по любому из пп. 2-17,
где эпителиальные клетки-предшественники кишечника не получают из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
27. Способ по п. 26, отличающийся тем, что среда для выращивания включает минимальную среду и дополнительно содержит: лиганд рецепторной тирозинкиназы, ингибитор BMP и агонист Wnt.
28. Способ по п. 27, в котором среда для выращивания дополнительно содержит вальпроевую кислоту и ингибитор GSK-3.
29. Способ по п. 28, где ингибитор GSK-3 представляет собой CHIR99021.
30. Способ по любому из пп. 26-29, отличающийся тем, что способ дополнительно включает получение дифференцированного органоида кишечника.
31. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в скрининге для обнаружения лекарственных средств.
32. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в анализе токсичности.
33. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в диагностике.
34. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях эмбриологии ткани, линий клеток и путей дифференцировки.
35. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях для идентификации химических и/или нейронных сигналов, приводящих к высвобождению соответствующих гормонов.
36. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях экспрессии генов, включая экспрессию рекомбинантных генов.
37. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях механизмов, участвующих в повреждении и репарации ткани.
38. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях воспалительных и инфекционных заболеваний.
39. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях патогенетических механизмов.
40. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в исследованиях механизмов трансформации клеток и этиологии злокачественных новообразований.
41. Трехмерный органоид, полученный способом по п. 25 или 30, где органоид получен из кишечника, имеет просвет, и в котором по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 99% клеток экспрессируют маркеры аргентофильных клеток, для использования в регенеративной медицине, при которой органоид трансплантируется пациенту.
42. Органоид по п. 30, отливающийся тем, что органоид получен от человека или мыши.
43. Органоид по любому из пп. 30-41, где маркеры аргентофильных клеток выбраны из Chga, Chgb, Tac1, Tph1, Gip, Fabp5, Ghrl, Pyy, Nts, Neurod1, Sst, Sct, холецистокинина, глюкагона и/или проглюкагона.
44. Фармацевтический состав, содержащий один или несколько ингибиторов пути EGFR, ингибитор Notch и один или несколько ингибиторов Wnt для стимуляции дифференцировки стволовых клеток in vivo.
45. Способ скрининга терапевтического или профилактического фармацевтического лекарственного средства, включающий:
приведение трехмерного органоида по любому из пп. 31-43 в контакт с молекулой-кандидатом или библиотекой молекул-кандидатов,
оценку указанного органоида на любые эффекты или изменение в органоиде; и
идентификацию молекулы-кандидата, вызывающей указанные эффекты, в качестве потенциального лекарственного средства.
46. Способ скрининга косметического средства, включающий:
приведение трехмерного органоида по любому из пп. 31-43 в контакт с молекулой-кандидатом или библиотекой молекул-кандидатов,
оценку указанного органоида на любые эффекты или изменение в органоиде; и
идентификацию молекулы-кандидата, вызывающей указанные эффекты, в качестве потенциального косметического средства
47. Способ по п. 45, дополнительно включающий получение указанной молекулы-кандидата в качестве фармацевтического или средства.
48. Способ по п. 46, дополнительно включающий получение указанной молекулы-кандидата в качестве косметического средства.
49. Способ получения популяции клеток, обогащенной аргентофильными клетками (EEC), включающий культивирование популяции эпителиальных клеток-предшественников в дифференцировочной среде по любому из пп. 2-17, и, где эпителиальные клетки-предшественники не происходят из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
50. Способ получения популяции клеток, обогащенной GLP1-секретирующими EEC, включающий культивирование популяции эпителиальных клеток-предшественников в дифференцировочной среде по любому из пп. 2-17, где дифференцировочная среда содержит ингибитор BMP и, где эпителиальные клетки-предшественники не происходят из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
51. Способ получения популяции клеток, обогащенной секретин-секретирующими EEC, включающий культивирование популяции эпителиальных клеток-предшественников в дифференцировочной среде по любому из пп. 2-17, где дифференцировочная среда содержит активатор пути BMP и, где эпителиальные клетки-предшественники не происходят из эмбриональных стволовых клеток или линий эмбриональных стволовых клеток.
| WO 2014159356 A1, 0.10.2014 | |||
| WO 2010090513 A2, 12.08.2010 | |||
| XIAOLEI YIN et al., Niche-independent high-purity cultures of Lgr5+ intestinal stem cells and their progeny, Nature methods, vol.11, No.1, 01.12.2013, pp.106-112 | |||
| ПРОСКУРЯКОВ С.Я., и др., Стволовые клетки кишечного эпителия механизмы выживаемости и роль микробиоты, Биомедицинская химия, |

