Настоящее изобретение относится к экспериментальной биологии в области сельского хозяйства, в частности к оптическим методам детектирования патогенов растений, касается способа досимптомной детекции вирусных инфекций в растениях с помощью РАМ-флуометрии, который может быть применен для отбора устойчивых к вирусным инфекциям сортов растений при селекции, а также при оценке эффективности разрабатываемых средств защиты растений.
Патогены являются серьезной проблемой при производстве сельскохозяйственной продукции, вызывая значительные экономические потери. Увеличение доли сельскохозяйственной продукции, выращиваемой в закрытом грунте, усиливает проблему, поскольку создаются благоприятные для размножения патогенов условия (Nazarov et al., 2020; Lu, et al., 2021; Singh, et al., 2023). Успешная стратегия борьбы с патогенами подразумевает их раннее обнаружение, поскольку любая задержка, как и неправильная идентификация заболевания, ведут к массовому поражению растений. В число методов детектирования входят традиционная визуальная оценка болезней, серологические методы анализа, молекулярно-генетические и дистанционные оптические методы (Scala et al., 2018; Singh et al., 2020; Buja et al., 2022). Последние базируются на оценке оптических свойств растения в различных областях спектра электромагнитного излучения и использовании датчиков, которые измеряют отражательную способность, температуру или флуоресценцию (Terentev et al., 2022, Tanner et al., 2022). Оптические методы отличаются неинвазивностью, быстротой получения и обработки информации. К их преимуществам также относится относительно низкая стоимость анализа в силу отсутствия расходов на реактивы. Одним из самых чувствительных оптических методов является метод РАМ-флуориметрии, который позволяет измерять непосредственно активность фотосинтеза (Корнеев, 2002; Гольцев и др., 2016).
Принцип метода РАМ-флуориметрии (pulse amplitude modulation) базируется на следующем. Существует три пути реализации энергии поглощенных хлорофиллом квантов света: фотохимические реакции, флуоресценция и тепловая диссипация энергии. Эти три процесса являются конкурентными, вследствие чего изменение активности одного из них неизбежно вызывает изменение активности других. Это позволяет на основании регистрации флуоресценции хлорофилла определять интенсивность фотохимических реакций и тепловой диссипации энергии, важнейших характеристик активности фотосинтеза. Перед процедурой измерения растение должно быть адаптировано к темноте в течение 10-15 минут; за такое время происходит полное окисление акцепторов электронов фотосистемы П. Измерение начинается подачей слабого импульсного света, который вызывает флуоресценцию хлорофилла, но не вызывает фотохимических реакций, ML (Measuring Light). Производится регистрация начального (нулевого) уровня флуоресценции Fo. Затем включается насыщающий импульс SP (Saturating Pulse) - короткий и мощный импульс света, который вызывает восстановление всех акцепторов электронов в фотосистеме II. Вследствие восстановления акцепторов временно блокируются фотохимические реакции, в результате чего резко повышается выход флуоресценции. Интенсивность флуоресценции хлорофилла в предварительно адаптированном к темноте объекте достигает максимального значения (Fm). После снижения уровня флуоресценции до исходного темнового значения F0, включается непрерывно действующий актиничный свет, AL (Actinic Light), с заданной плотностью потока фотосинтетически активных фотонов. Через некоторое время после включения AL подаются следующие измерительный ML и насыщающий SP импульсы. Производится регистрация соответственно текущей флуоресценции F и максимальной флуоресценции Fm'. Такие серии измерительных и насыщающих импульсов подаются с заданной частотой. Измерения флуоресценции хлорофилла с использованием РАМ-флуориметра обеспечивает быструю оценку эффективности преобразования энергии фотонов в энергию химических связей органических соединений в процессе фотосинтеза у исследуемых объектов. Важнейшими измеряемыми параметрами являются отношение переменной флуоресценции (Fv) и максимальной флуоресценции (Fv/Fm=(Fm-F0)/Fm)), квантовый выход фотохимических реакций фотосистемы II (ФPSII=(Fm' - F)/Fm') и нефотохимическое тушение флуоресценции (NPQ=(Fm-Fm')/Fm'). Изменение данных показателей флуоресценции хлорофилла имеет место под действием различных факторов (Корнеев 2002 Гольцев и др., 2016).
Известны работы, в которых продемонстрирована возможность детектирования патогенов в растениях с помощью метода РАМ-флуориметрии (Kyselakova et al., 2011; Granum et al., 2015; Pineda et al., 2018 и др.). В работах на различных растениях продемонстрировано, что такие показатели флуоресценции хлорофилла как Fv/Fm, а также ФPSII и NPQ, зарегистрированные в стационарном (адаптированном к свету состоянии) изменяются при инфицировании. Однако, регистрация параметров осуществлялась на симптомных стадиях заболевания, при наличии визуальных признаков болезни. Другим ограничением является проведение измерений на инокулированном листе, который непосредственно подвергался искусственному инфицированию с нанесением механического повреждения. Также отсутствовал надежный контроль пространственного распределения патогенов в исследуемом объекте.
Известен способ дистанционного определения функционального состояния фотосинтетического аппарата растений (RU 2453829 С2, кл. G01N 21/64, опубл. 20.06.2012), включающий дистанционное определение содержания хлорофилла и фотосинтетической активности растения. Определяют, посылая лазерные импульсы излучения в темное время суток, возбуждающие флуоресценцию хлорофилла, принимают часть излучения флуоресценции, подвергают ее спектральной селекции, регистрируют полученную информацию и подвергают ее компьютерной обработке и записи. Обнаружение нарушения фотосинтеза позволяет на ранней стадии выявить заболевания растений, ухудшение состояния окружающей среды и своевременно помочь растениям. Для данного способа не приведены доказательства возможности детектирования заболевания растений на досимптомной стадии инфицирования.
Известен способ оценки зимостойкости плодовых растений (RU 2514400 С2, кл. A01G 7/00, А01Н 1/04, опубл. 27.04.2014), в котором оценку поврежденных побегов производят по величине максимального квантового выхода фотохимических реакций, которые определяют РАМ-флуориметром, затем регистрируют минимальный уровень флуоресценции и изменения этого показателя под действием актиничного света плотностью 190 μmol/(m2s) и после воздействия на объект импульса света высокой интенсивности (10000 μmol/(m2s), 450 нм). Способ позволяет ускорить оценку повреждений плодовых растений морозом. Описанный способ базируется на применении метода РАМ-флуориметрии для оценки действия на растения абиотических, а не биотических стрессоров. Не продемонстрирована возможность детекции патогенов.
Известен дистанционный способ обнаружения стрессовых состояний растений (RU 2646937 С1, кл. G01N 21/6402, A01G 7/00, опубл. 12.03.2018), включающий дистанционное обнаружение участков растительности в стрессовом состоянии путем лазерного возбуждения флуоресценции хлорофилла растения и регистрации интенсивности флуоресценции. Способ обеспечивает повышение надежности обнаружения стрессового состояния растений. Для данного способа не продемонстрирована возможность детектирования патогенов.
Известна технология количественного определения флуоресценции в реальном времени (CN 104450868, кл. C12Q 1/68, C12Q 1/06, G01N 21/64, опубл. 25.03.2015), которая может достоверно отражать количество патогена в семенах и обеспечивает основу для раннего прогнозирования и раннего предупреждения заболеваний. Используется метод ПЦР для обнаружения возбудителя угловых пятен на листьях огурца и ее применение. Описанный метод не базируется на регистрации флуоресценции хлорофилла. Описанный метод детекции патогенов является инвазивным, затратным по времени, необходимо использовать реактивы для реализации метода.
Наиболее близким по технической сущности и достигаемому результату к предлагаемому изобретению является способ детекции патогенов растений описанный в работе «Предсимптомное выявление вирусной инфекции в листьях табака с помощью флуорометрии РАМ» Гришина А., Шерстнева А., Гринберг М., Здобнова Т.И др., 2021, 10, 2782, https://doi.org/10.3390/plants10122782, принятый за ближайший аналог (прототип). В данной работе продемонстрировано, что показатели флуоресценции хлорофилла PSII и NPQ в переходном состоянии (через короткое время после включения актиничного света) обладают большей чувствительностью к инфицированию в сравнении с таковыми в стационарном состоянии.
Недостатком подхода является отсутствие анализа пространственной неоднородности показателей флуоресценции хлорофилла в здоровых листьях, что может привести к ложноположительным заключениям; отсутствие анализа влияния интенсивности AL на различия в значениях PSII и NPQ в переходном состоянии между инфицированными и здоровыми областями листа, что важно для работы с конкретными моделями РАМ-флуориметров, отсутствие анализа возможности работы на отделенных от растения листьях. Последнее может иметь большое значение при работе с растениями, имеющими плотно перекрывающееся расположение листьев, а также при работе на стационарном РАМ-флуориметре.
В задачу заявляемого изобретения положена разработка нового способа детектирования вирусной инфекции на основании показателей флуоресценции хлорофилла листьев растений, который наряду с выявлением инфицирования на ранней стадии при производстве сельскохозяйственной продукции может быть использован в селекции для отбора устойчивых к патогенам сортов растений и при разработке агрохимикатов для оценки эффективности новых средств защиты растений.
Технический результат от использования заявляемого изобретения заключается в повышении точности детектирования за счет исключении ложноположительных заключений. Также предлагаемый способ обеспечивает возможность использования стационарных флуориметров при работе с листьями крупных растений, которые невозможно поместить в прибор, что обеспечивает большую скорость анализа при одновременном наблюдении нескольких листьев, отделенных от различных растений.
Это достигается тем, что способ досимптомного детектирования вирусной инфекции включает помещение исследуемого растения, или отделенного листа исследуемого растения, в систему для регистрации флуоресценции хлорофилла методом РАМ, адаптацию растений к темноте в течение 10-15 минут, регистрацию начальной флуоресценции хлорофилла F0, регистрацию максимальной флуоресценции хлорофилла Fm в адаптированном к темноте состоянии, включение актиничного света, регистрацию текущего уровня флуоресценции хлорофилла F, а также максимальный выход флуоресценции Fm' через короткое время после включения актиничного света в течение 30-80 секунд, на основе показателей флуоресценции хлорофилла с помощью импульсно-модулированного флуориметра с функцией имиджинга получают изображения, представляющие пространственное распределение величин ФPSII и NPQ в переходном состоянии, после чего осуществляют расчет показателя ФPSII по формуле ФPSII=(Fm'–F)/Fm', где ФPSII - квантовый выход фотохимических реакций фотосистемы II, Fm' -максимальный выход флуоресценции, F - текущий выход флуоресценции, и расчет показателя NPQ по формуле NPQ=(Fm - Fm')/Fm', где NPQ - нефотохимическое тушение флуоресценции, Fm максимальный выход флуоресценции в адаптированном к темноте состоянии, Fm' - максимальный выход флуоресценции, затем осуществляют выбор в пределах исследуемого растения или отделенного листа исследуемого растения на изображениях ФPSII и NPQ областей, различающихся по величине анализируемого показателя, нахождение отношения показателя, выявление инфицированных участков листа осуществляют на основании величины отношения, превышающей 1,5-2 для NPQ и превышающий 1,5-2,5 для ФPSII, а также на основании пространственного распределения неоднородности; показатели флуоресценции хлорофилла регистрируют с помощью системы визуализации импульсно-модулированной флуоресценции хлорофилла IMAGING-РАМ MINI; актиничный свет, представляющий собой синий свет с интенсивностью 102 мкмоль м-2 с-1, включают через 10 секунд, при этом его интенсивность обеспечивает величину показателя эффективного квантового выхода фотосистемы II ФPSII близкую к 0,5; в качестве насыщающего света используют вспышки синего света 3 000 мкмоль м-2 с-1, длительностью 800 мс; используют сельскохозяйственные растения, произрастающие в условиях закрытого или открытого грунта, или листья таких растений, отделенные от них; значение квантового выхода фотохимических реакций фотосистемы II ФPSII и нефотохимического тушения флуоресценции NPQ рассчитывают автоматически с помощью программы для регистрации флуоресценции.
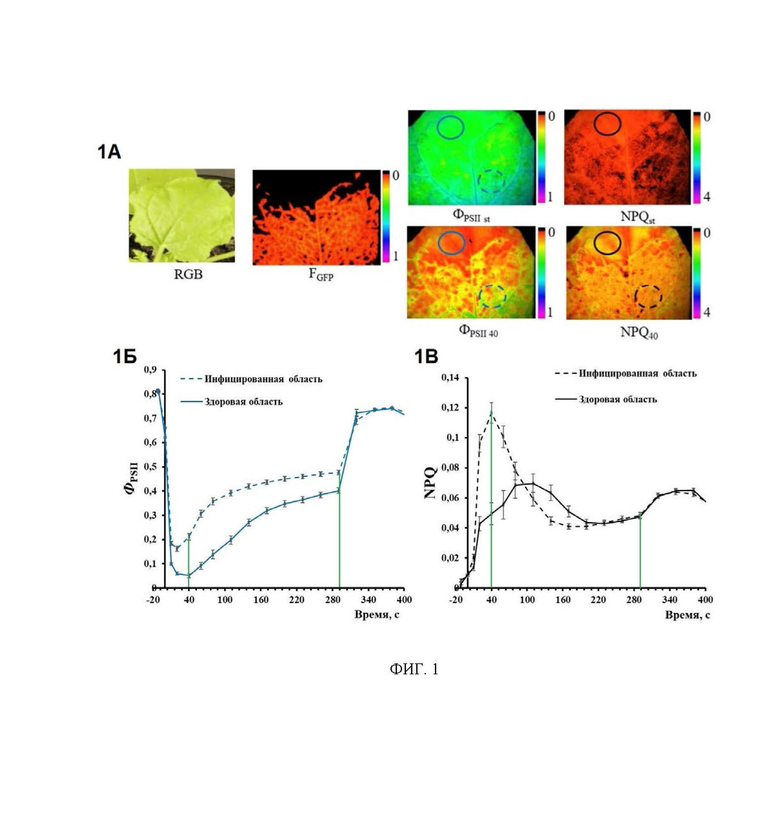
На фиг. 1 показаны изображения десятого листа табака, системно инфицированного вирусом PVX на четвертый день распространения вируса в лист, полученные различными методами (панель 1А), где: RGB - стандартное RGB-изображение, полученное с помощью цифровой камеры, демонстрирует внешний вид листа; Fgfp - флуоресцентное изображение вирусных частиц PVX с GFP в капсиде, полученное при λex=460 нм и λem=500-540 нм, отражает локализацию вируса, меченного флуоресцентным белком GFP, в изучаемом листе; ФPSII st - квантовый выход фотосистемы II в стационарном состоянии; ФPSII 40 квантовый выход фотосистемы II в переходном состоянии; NPQst - нефотохимическое тушение флуоресценции в стационарном состоянии; NPQ 40 - не фотохимическое тушение флуоресценции в переходном состоянии. Изображения ФPSII st и NPQst получены через 290 секунд после включения актиничного света. Изображения ФPSII40 и NPQ40 получены через 40 секунд после включения актиничного света. Сплошные и пунктирные круги показывают пример расставления областей интереса (ROI), от которых получали данные для построения динамики ФPSII и NPQ. 1Б и 1В - индуцированная включением актиничного света динамика ФPSII и NPQ инфицированной и здоровой областей десятого листа табака системно инфицированного PVX. На графиках для оси абсцисс за начало отсчета принят момент включения актиничного света. Вертикальные зеленые линии показывают время после включения актиничного света для максимальных (переходное состояние, 40 с) и минимальных (стационарное состояние, 290 с) различий определяемых параметров между здоровой и инфицированной зонами листа.
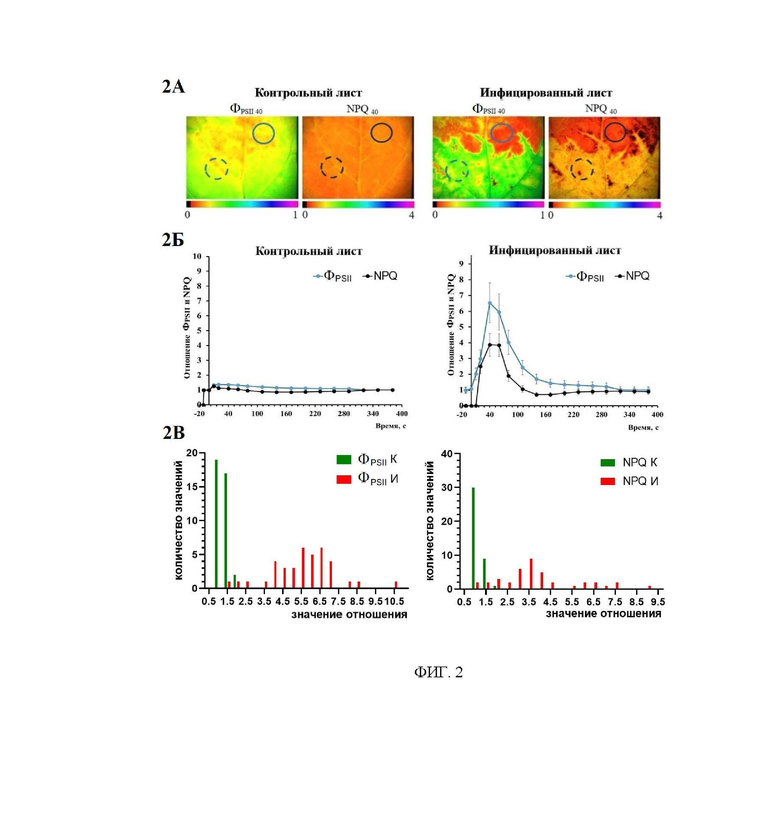
На фиг. 2 представлены изображения ФPSII40 и NPQ40 для контрольных и инфицированных 10-ых листьев табака (панель 2А). На данных изображениях были выбраны наиболее контрастные по величинам ФPSII и NPQ области (показаны сплошными и пунктирными кругами), значения показателей для которых были использованы для расчета отношений ФPSII и NPQ. На графиках панели 2Б приведены зависимости отношений показателей ФPSII и NPQ в переходном состоянии в здоровой и инфицированной частях инфицированного десятого листа и соответствующих областях десятого листа контрольного растения. На панели 2 В приведены диаграммы распределения величин отношения показателей ФPSII и NPQ для контрастных участков здорового листа (зеленые столбики) и величин отношения ФPSII40 и NPQ40 для инфицированных и здоровых участков инфицированного листа (красные столбики).
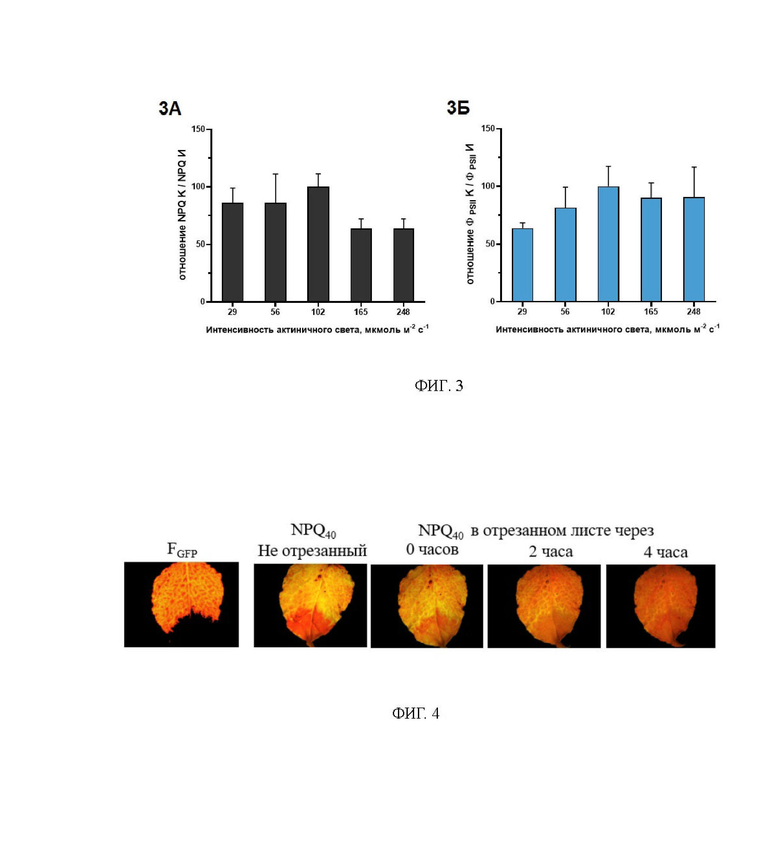
На фиг. 3 представлены отношения величин NPQ40 (панель 3А) и ФPSII40 (панель 3Б) для инфицированных и здоровых участков инфицированного листа, полученные при использовании AL различной интенсивности. Увеличение интенсивности AL в диапазоне от примерно 30 до 100 мкмоль м-2 с-1 ведет к увеличению отношения ФPSII40 инфицированной части листа к здоровой. При дальнейшем увеличении интенсивности AL до значений около 250 мкмоль м-2 с-1 имеет место тенденция к снижению отношения. Отношение NPQ=инфицированной части листа к здоровой имеет высокие значения в диапазоне интенсивности AL от 30 до 100 мкмоль м-2 с-1. При дальнейшем увеличение интенсивности AL происходит снижение отношения.
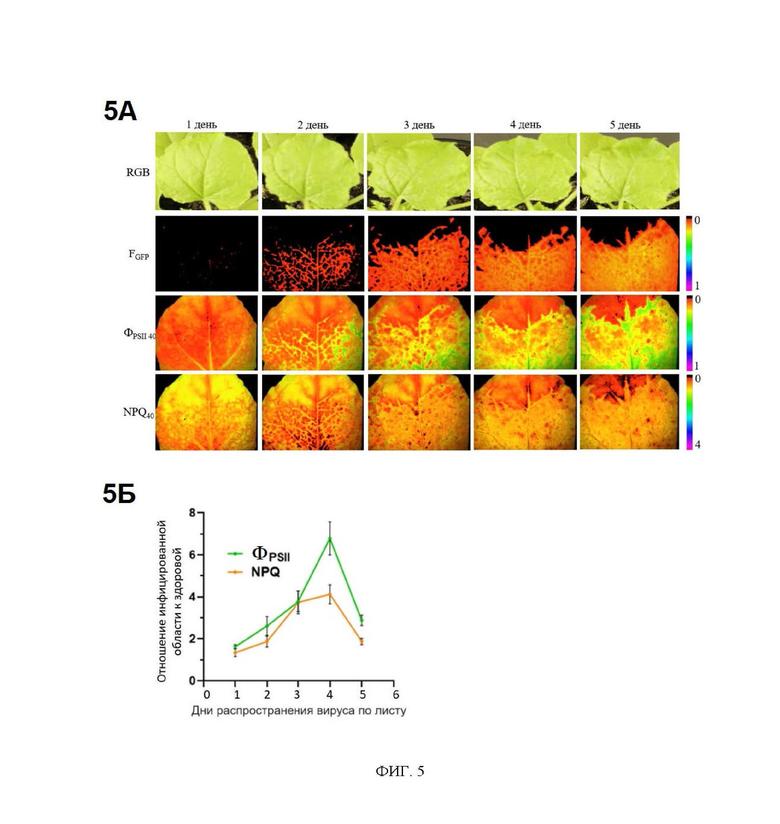
На фиг. 4 представлены изображения показателя NPQ40 для инфицированного листа до и через различные интервалы времени после его отделения от растения. Fgfp флуоресцентное изображение вирусных частиц PVX с GFP в капсиде, полученное при λех=460 нм и λem=500-540 нм, отражает локализацию вируса, меченного флуоресцентным белком GFP, в изучаемом листе; NPQ40 - нефотохимическое тушение флуоресценции в переходном состоянии. В отделенном листе сохраняются типичные различия в NPQ между инфицированной и здоровой частями листа.
На фиг. 5 приведены изображения десятого листа, системно инфицированного PVX, в разные дни после появления вируса в листе (панель 5А): Fgfp флуоресцентные изображения распространения вирусных частиц с GFP в капсиде по листу (λex - 460 нм, λem - 500-540 нм); ФPSII40 - изображения ФPSII, полученные через 40 с после включения актиничного света; NPQ40 изображения NPQ, полученные через 40 с после включения актиничного света. На панели 5Б представлена динамика отношений ФPSII40 и NPQ40 в разные дни после распространения вируса в исследуемый лист.
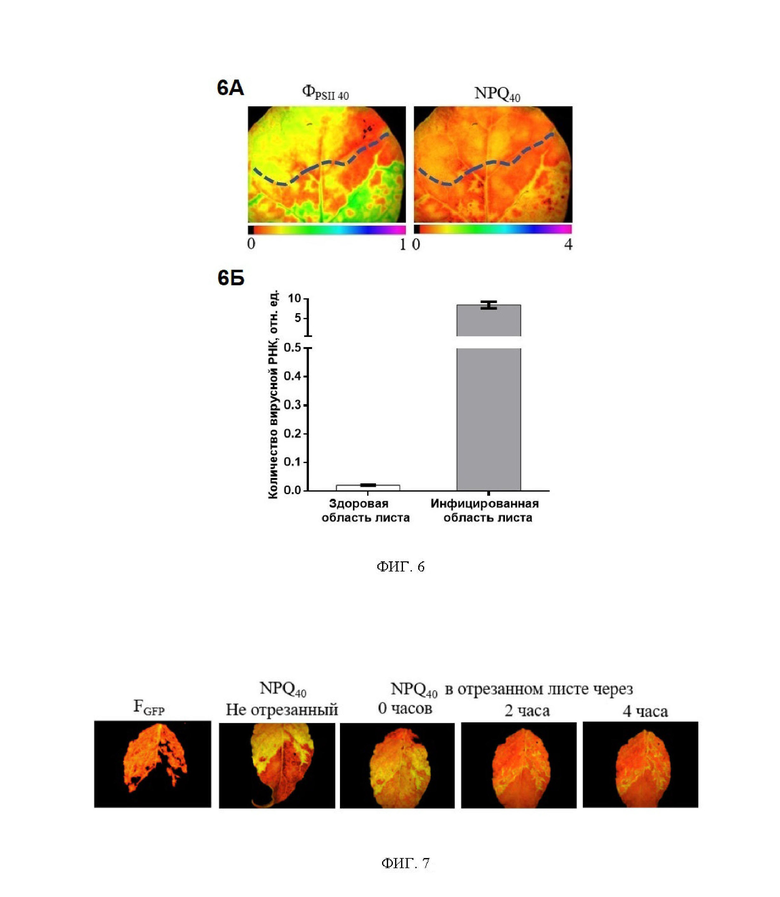
На фиг. 6 показаны изображения листа табака, инфицированного вирусом PVX, полученные методом РАМ-имиджинга (панель 6А). ФPSII40 квантовый выход фотосистемы II в переходном состоянии; NPQ40 - нефотохимическое тушение флуоресценции в переходном состоянии. Пунктиром показана граница, разделяющая инфицированную и здоровую часть листа, проведенная на основании разницы в показателях ФPSII40 и NPQ40. На панели 6Б представлена диаграмма, показывающая в относительных единицах определенное методом полимеразной цепной реакции (ПЦР) в реальном времени количество вирусной РНК в областях листа, идентифицированных на основании показателей флуоресценции хлорофилла как здоровая и инфицированная.
На фиг. 7 показана возможность оценки наличия вирусной инфекции в отделенном от растения листе на основании анализа изображения показателя NPQ40. Приведены изображения показателя NPQ40 для инфицированного листа до и через различные интервалы времени после его отделения от растения. Fgfp - флуоресцентное изображение вирусных частиц PVX с GFP в капсиде, полученное при λех=460 нм и λem=500-540 нм, отражает локализацию вируса, меченного флуоресцентным белком GFP, в изучаемом листе; NPQ40 - нефотохимическое тушение флуоресценции в переходном состоянии.
Предлагаемый способ детектирования вирусной инфекции в растениях методом РАМ-флуориметрии осуществляют следующим образом.
Растения помещают в систему для регистрации импульсно-модулированной флуоресценции хлорофилла и адаптируют к темноте не менее 10 минут. После темновой адаптации производят регистрацию начальных параметров F0 и Fm, затем включают актиничный свет, и через короткое время после включения актиничного света (в течение 30-80 секунд) осуществляют регистрацию текущего уровня флуоресценции F, а также Fm' с использованием вспышки насыщающего света. Измеряемые параметры флуоресценции хлорофилла используются для расчета показателей ФPSII и NPQ, осуществляют с помощью программного обеспечения оборудования.
На основе показателей флуоресценции хлорофилла получают изображения с помощью импульсно-модулированного флуориметра с функцией имиджинга, представляющие пространственное распределение величин ФPSII и NPQ в переходном состоянии (полученные через короткое время после включения актиничного света), используют в дальнейшем анализе. Выбор времени получения изображений обусловлен тем, что именно в таком диапазоне времени после включения актиничного света имеет место максимальная разница между инфицированной и неинфицированной областями листа (фиг. 1, фиг. 2).
Затем осуществляют расчет показателя Opsii по формуле:
ФPSII=(Fm'-F)/Fm', где ФPSII - квантовый выход фотохимических реакций фотосистемы II, Fm' - максимальный выход флуоресценции, F текущий выход флуоресценции; и расчет показателя NPQ по формуле:
NPQ=(Fm - Fm')/Fm', где NPQ - нефотохимическое тушение флуоресценции,
Fm максимальный выход флуоресценции в адаптированном к темноте состоянии, Fm' - максимальный выход флуоресценции.
Производят сопоставление величин показателей ФPSII и NPQ в максимально контрастных областях анализируемого изображения. Если отношение показателей в различных областях листа больше порогового значения, то эта область идентифицируется, как инфицированная. Для неинфицированных листьев не характерно превышение отношения в неоднородных областях листа больше 1,5-2 для показателя NPQ и больше 1,5-2,5 для показателя ФPSII (фиг. 2В). Соотношение указанных показателей может варьироваться в зависимости от вида растения и вида патогена. Дополнительным критерием отнесения участка листа к инфицированной области является пространственное распределение неоднородностей показателей. В растениях системное распространение вирусов происходит преимущественно по флоэме, что обуславливает поступление вируса в лист через черешок с последующим распространением по центральной и боковым жилкам. Такой пространственный паттерн служит дополнительным критерием отнесения области листа к инфицированной вирусом.
Так по данным, полученным на инфицированных вирусом PVX растениях табака на досимптомной стадии, при отсутствии визуальных симптомов инфицирования, имеет место более быстрое изменение показателей ФPSII и NPQ при включении актиничного света в инфицированной области листа в сравнении с неинфицированной. Это ведет к тому, что величины показателей ФPSII и NPQ в переходном состоянии в инфицированной области листа превышают таковые в неинфицированной. Физиологическую основу обнаруженных различий объясняет рассмотрение фотосинтетических процессов. Скорость светоиндуцированных изменений ФPSII и NPQ и время выхода их на стационарный уровень определяются скоростью достижения равновесия в продукции и потреблении АТФ и НАДФН (Гольцев и др., 2016; Leucker и др, 2017). Ускорение выхода на стационарный уровень показателей фотосинтетической активности может свидетельствовать о более высокой скорости потребления АТФ в инфицированных клетках, что может быть связано с потребностью в дополнительной энергии для защитных процессов (Wang и др., 2019).
Следует отметить, что различия в показателе ФPSII между инфицированными и здоровыми областями листа зависят не только от времени после включения актиничного света, но также и от его интенсивности. На фиг. 3 приведены отношения показателей ФPSII и NPQ, зарегистрированных при различных значениях интенсивности AL (в диапазоне от 30 до 250 мкмоль м-2 с-1. Максимальные различия между инфицированной и неинфицированными областями листа имеют место при интенсивности AL около 100 мкмоль м-2 с-1 как в отношении ФPSII, так и в отношении NPQ (фиг. 3). Проведение измерений нужно выполнять при интенсивности AL, обеспечивающий максимальный контраст. В связи с тем, что в различных моделях флуориметров источники AL могут иметь различные спектральные характеристики, а фотосинтетически активными являются кванты синего и красного света, целесообразно предварительно выбирать интенсивность AL исходя из стационарного уровня ФPSII.Стационарный уровень ФPSII обратно пропорционален интенсивности AL (Гольцев и др., 2016). Интенсивность AL, обеспечивающие максимальные различия в Opsii между инфицированной и здоровой областями, обеспечивала стационарный уровень ФPSII в диапазоне 0,45-0,55. Значения AL, обеспечивающие такую величину ФPSII целесообразно выбирать при исходных настройках оборудования.
Предложенный способ обеспечивает выявление инфицированных участков листа растения на досимптомной стадии болезни на основании данных о работе фотосинтетического аппарата, которая зависит от наличия вируса в клетках. Это позволяет выявлять наличие заболевания на ранней стадии при выращивании сельскохозяйственных культур и тем самым снизить возможные потери урожая и повысить качество сельхозпродукции за счет внесения меньшего количества химикатов, оценивать устойчивость сортов растений к патогенам при селекции, оценивать эффективность в отношении патогенов разрабатываемых средств защиты растений.
Анализ контраста неоднородностей на здоровом листе позволяет предложить количественный критерии (1,5-2 - для показателя NPQ и 1,5-2,5 - для показателя ФPSII), по которым можно отделить неоднородности, вызванные заболеванием от присущих здоровым, что позволяет избежать ложноположительных заключений.
Наряду с работой с целыми растениями предлагаемый способ детектирования вирусной инфекции может быть реализован и в работе с отделенным листом растения. В этом случае в РАМ флуориметр помещается не целое растение, а исследуемый отделенный лист растения. Процедура измерения полностью повторяет таковую, описанную выше. Фиг. 4 демонстрирует пространственное распределение NPQ в инфицированном листе до его срезания, а также через различные интервалы времени после срезания. Различия между инфицированной и неинфицированными областями листа сохраняются в течение как минимум нескольких часов после срезания. Проведение измерений на отделенных листьях, а не на целых растениях, обеспечивает возможность использования стационарных флуориметров при работе с крупными растениями, которые невозможно поместить в прибор. Также такой подход также обеспечит большую скорость анализа при одновременном наблюдении нескольких листьев, отделенных от различных растений. Ниже приведены примеры осуществления предлагаемого изобретения.
Пример 1
Эксперимент осуществляли на растениях Nicotiana benthamiana. Инфицирование четырехнедельных растений N. benthamiana, выращенных в контролируемых условиях вегетационной камеры (24°С, 16-часовой световой период), проводили с помощью агоробактериальной инфильтрации. Для этого использовали два варианта трансформантов агробактерий A. tumefaciens штамма С58С1, несущие два различных вида агробактериальных векторов: (i) вектор pBin-PVX-GFP, обеспечивающий экспрессию вируса PVX с GFP-белком, (ii) вектор pLH-P19, обеспечивающий экспрессию белка Р19 супрессора РНК-интерференции растений. Инокуляция осуществляли в 4-ый настоящий лист растения. Регистрацию показателей флуоресценции хлорофилла в инфицированном и неинфицированном участках неинокулированных 10-ых листьев табака оценивали с помощью метода РАМ-флуориметрии с использованием IMAGING-PAM MINI Version (Heinz Walz GmbH, Efreltrich, Германия). Исследуемые листья, не отделяя от растения, закрепляли в держателе РАМ-флуориметра, после чего осуществляли темновую адаптацию растения в течение 15 минут. Затем определяли темновой (F0), а также максимальный (Fm) выход флуоресценции с использованием насыщающего импульса (460 нм, интенсивность 6 ммоль м-2 с-1, длительность 800 мс). После этого измеряли текущий (F), а также максимальный выход флуоресценции Fm' с использованием насыщающих вспышек, генерируемых каждые 10 с. Актиничный свет (синий свет, 460 нм, интенсивность 102 мкмоль м-2 с-1,) включали через 25 с; после включения актиничного света запись параметров флуоресценции хлорофилла осуществляли в течение 7 минут.Значение квантового выхода фотохимических реакций ФСП и нефотохимического тушения NPQ рассчитывали автоматически с помощью программы, предоставленной производителем системы РАМ-имиджинга. Наряду с регистрацией флуоресценции хлорофилла IMAGING-PAM MINI позволяет регистрировать флуоресценцию GFP при выборе соответствующего фильтра эмиссии. Флуоресценцию GFP возбуждали при λem 460, регистрировали в диапазоне 500-540 нм. Флуоресцентный сигнал GFP позволил наблюдать распространение вируса по листу (фиг. 5). Измерения производили один раз в день в течение 5 дней после распространения вируса в 10-ый неинокулированный лист.
С 5-го дня после инокуляции в 4-й лист вирус уверенно детектировался по флуоресцентному сигналу в 10-м листе. В следующие три дня происходил быстрый рост инфицированной площади. Суммарная площадь заражения к концу эксперимента составляла около 80% от общей площади листа (фиг. 5А). Вместе с ростом площади инфицирования происходило увеличение площади листа, в пределах которой имело место изменение показателей ФPSII40 и NPQ40. Величины ФPSII40 и NPQ40 в областях листа, где наблюдался флуоресцентный сигнал GFP вируса, были выше таковых в областях листа, где сигнал вируса отсутствовал. Отношение величин ФPSII40 и NPQ40 в областях, где наблюдался сигнал вируса к таковым, где он отсутствовал представлено на диаграмме фиг. 5Б. С увеличением времени после распространения вируса в лист происходило увеличение отношения. Величина отношения превышала 2 начиная со второго дня после регистрации флуоресцентного сигнала от сцепленного с вирусом GFP. Максимум отношения имел место спустя 4 дня после распространения вируса в лист.
Необходимо отметить, что в течение периода времени, когда происходили описанные выше изменения в показателях ФPSII40 и NPQ40, на исследуемых листьях отсутствовали визуальные симптомы заболевания. Такие симптомы, представляющие собой формирование сетчатой структуры и частичное изменение окраски в инфицированной области, возникали на 4-5-й дни после обнаружения по флуоресцентному сигналу вируса в исследуемом листе.
Полученный результат говорит о том, что сопоставление показателей ФPSII40 и NPQ40 позволяет обнаружить вирусную инфекцию в листе исследуемого растения на досимптомной стадии заболевания. Периодические измерения позволяют определить скорость изменения площади, подверженной инфицированию.
Пример 2
Инокуляцию растений N. benthamiana осуществляли так же, как описано в примере 1. Регистрацию показателей флуоресценции хлорофилла в инфицированном и неинфицированном участках целых неинокулированных 10-ых листьев табака оценивали с помощью метода РАМ-флуориметрии с использованием IMAGING-PAM MINI Version (Heinz Walz GmbH, Effeltrich, Германия) как описано в примере 1.
Из фиг. 6 представлены изображения ФPSII 40 и NPQ40 десятого листа растения, подвергнутого инокуляции. Изображения были получены через 5 дней после инокуляции 4-го листа. На изображениях присутствуют характерные для вирусной инфекции изменения показателей ФPSII40 и NPQ40. Изменения ФPSII40 и NPQ40 имеют место в нижней (базальной) части листа, преимущественно в областях, примыкающих к жилкам. Величина отношения ФPSII40 и NPQ40 для указанных областей превышает 2. Область листа с более высокими значениями ФPSII40 и NPQ40 была идентифицирована нами как инфицированная.
Для проверки сделанного заключения выполняли анализ содержания вируса в определенных как зараженная и незараженная части листа методом ПЦР. Листовую пластину разделяли на две части как обозначено линией на фиг. 6. После чего фрагменты листа были исследованы методом ПЦР. Для этого замороженные в жидком азоте части листа измельчали до мелкого порошка. Тотальную РНК экстрагировали с помощью ExtractRNA («Евроген», Россия). Качество образцов проверяли на 1,2% агарозном геле, концентрацию РНК оценивали с помощью спектрофотометра NanoVue Plus (GE Healthcare, Чикаго, Иллинойс, США). Полученную РНК подвергали реакции обратной транскрипции с использованием набора MMLV RT с Oligo(dT)17-праймером («Евроген», Москва, Россия). Количественное определение PVX-GFP проводили с использованием системы ПЦР в реальном времени 7500 и Power SYBR Green PCR Master Mix (Applied Biosystems, Фостер-Сити, США). Последовательности праймеров (прямой - 5'-AAGCCTGAGCACAAATTCGC-3', обратный - 5'-GCTTCAGACGGTGGCCG-3'), позволяющих амплифицировать фрагмент гена белка оболочки (CP) длиной 101 п. о. PVX, были взяты из статьи (Mortimer-Jones и др., 2009). Размер ампликона проверяли разделением на 1,5% агарозном геле. Программа ПЦР была следующей: (1) инициация при 50,0°С, 2 мин, (2) «горячий старт» при 95,0°С, 10 мин, (3) денатурация при 95,0°С, 15 с, (4) праймер. отжиг и синтез ДНК при 60,0°С в течение 1 мин. Этапы 3 4 повторялись 40 раз. Ct (пороговый цикл) определяли с помощью программного обеспечения 7500 версии 2.0.4 (Applied Biosystems, Фостер-Сити, Калифорния, США). Данные ПЦР были представлены как 2-Ct (как описано для ненормализованных отдельных точек данных в (Schmittgen and Livak, 2008). Содержание вирусной РНК выражали в условных единицах: N=2-Ct × 104. Содержание вируса в различных областях листа представлено на диаграмме фиг. 6Б.
Согласно полученным результатам, концентрация вирусной РНК в той области листа, которая была идентифицирована как инфицированная на основании показателей флуоресценции хлорофилла ФPSII40 и NPQ40 более чем в 4000 раз превышает содержание вирусной РНК в той области листа, которая была идентифицирована как не инфицированная. Полученные результаты подтверждают возможность детектирования вирусной инфекции на досимптомной стадии с помощью заявляемого способа.
Пример 3
Инокуляцию растений N. benthamiana осуществляли так же, как описано в примере 1. Исходно проводили регистрацию флуоресценции GFP вируса и показателей флуоресценции хлорофилла в неотделенном от растения 10-м листе (неинокулированный лист). Через 3 дня после обнаружения флуоресценции GFP вируса в исследуемом листе, лист срезался и в нем осуществлялась регистрация показателей флуоресценции хлорофилла. Процедура регистрации была аналогична таковой для неотделенного листа и описана в примере 1.
Фиг. 7 демонстрирует показатель NPQ40 в областях листа, где имела место и отсутствовала флуоресценция GFP, сцепленного с вирусом. В инфицированной области показатель NPQ40 превышает таковой для области, где отсутствовала флуоресценция GFP вируса. Как демонстрирует фиг. 7 непосредственно после срезания листа, а также через 2 и 4 часа сохраняется контраст между инфицированной и неинфицированной областями листа.
Полученный результат указывает на возможность использования срезанных листьев растений в качестве объекта наблюдения в течение нескольких часов после отделения.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ прогнозирования продуктивности пшеницы на основании показателей флуоресценции хлорофилла | 2022 |
|
RU2792444C1 |
| СПОСОБ ПОВЫШЕНИЯ ПРОДУКТИВНОСТИ РАСТЕНИЙ КАРТОФЕЛЯ В ОПТИМАЛЬНЫХ И СТРЕССОВЫХ УСЛОВИЯХ ВЫРАЩИВАНИЯ | 2019 |
|
RU2711577C1 |
| СПОСОБ ОЦЕНКИ ЗИМОСТОЙКОСТИ ПЛОДОВЫХ РАСТЕНИЙ | 2012 |
|
RU2514400C2 |
| УСТРОЙСТВО ДЛЯ УПРАВЛЕНИЯ РОСТОМ ИЛИ СВОЙСТВАМИ РАСТЕНИЙ | 2008 |
|
RU2462025C2 |
| Способ определения морозостойкости винограда | 2016 |
|
RU2653016C2 |
| СИСТЕМА ДЕТЕКТИРОВАНИЯ ФЛУОРЕСЦЕНЦИИ ДЛЯ ОПРЕДЕЛЕНИЯ ЗНАЧИМЫХ ПАРАМЕТРОВ РАСТИТЕЛЬНОСТИ | 1998 |
|
RU2199730C2 |
| СПОСОБ ПОВЫШЕНИЯ УРОЖАЙНОСТИ ПШЕНИЦЫ | 2022 |
|
RU2785518C1 |
| СПОСОБ ФЛУОРОМЕТРИЧЕСКОГО ОПРЕДЕЛЕНИЯ ПАРАМЕТРОВ ФОТОСИНТЕЗА ФОТОАВТОТРОФНЫХ ОРГАНИЗМОВ, УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ И ИЗМЕРИТЕЛЬНАЯ КАМЕРА | 2006 |
|
RU2354958C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СОСТОЯНИЯ ДРЕВОСТОЯ ПО ПОБЕГАМ КРОН | 2023 |
|
RU2824521C1 |
| СПОСОБ ОЦЕНКИ ГАЗОУСТОЙЧИВОСТИ ДРЕВЕСНЫХ РАСТЕНИЙ | 2021 |
|
RU2766187C1 |
Настоящее изобретение относится к биохимии, к оптическим методам детектирования патогенов растений. Заявлен способ досимптомной детекции вирусных инфекций в растениях с помощью РАМ-флуометрии, который может быть применен для отбора устойчивых к вирусным инфекциям сортов растений при селекции, а также при оценке эффективности разрабатываемых средств защиты растений. Способ основан на анализе показателей флуоресценции хлорофилла с помощью импульсно-модулированного флуориметра с функцией имиджинга. Выявление инфицированных участков листа осуществляют на основании величины отношения, превышающей 1,5-2 для NPQ и превышающий 1,5-2,5 для ΦPSII, а также на основании пространственного распределения неоднородности. Технический результат от использования заявляемого изобретения заключается в повышении точности детектирования за счет исключения ложноположительных заключений. Также предлагаемый способ обеспечивает возможность использования стационарных флуориметров при работе с листьями крупных растений, которые невозможно поместить в прибор, что обеспечивает большую скорость анализа при одновременном наблюдении нескольких листьев, отделенных от различных растений. 5 з.п. ф-лы, 7 ил., 3 пр.
1. Способ досимптомного детектирования вирусной инфекции в растениях, включающий помещение исследуемого растения или отделенного листа исследуемого растения в систему для регистрации флуоресценции хлорофилла методом PAM, адаптацию растений к темноте в течение 10-15 минут, регистрацию начальной флуоресценции хлорофилла F0, регистрацию максимальной флуоресценции хлорофилла Fm в адаптированном к темноте состоянии, включение актиничного света, регистрацию текущего уровня флуоресценции хлорофилла F, а также максимальный выход флуоресценции Fm' через короткое время после включения актиничного света в течение 30-80 секунд, на основе показателей флуоресценции хлорофилла с помощью импульсно-модулированного флуориметра с функцией имиджинга получают изображения, представляющие пространственное распределение величин ФPSII и NPQ в переходном состоянии, после чего осуществляют расчет показателя ΦPSII по формуле
ΦPSII = (Fm' − F)/Fm', где
ΦPSII – квантовый выход фотохимических реакций фотосистемы II,
Fm' – максимальный выход флуоресценции,
F – текущий выход флуоресценции,
и расчет показателя NPQ по формуле
NPQ = (Fm − Fm')/Fm', где
NPQ – нефотохимическое тушение флуоресценции,
Fm – максимальный выход флуоресценции в адаптированном к темноте состоянии,
Fm' – максимальный выход флуоресценции,
затем осуществляют выбор в пределах исследуемого растения или отделенного листа исследуемого растения на изображениях ΦPSII и NPQ областей, различающихся по величине анализируемого показателя, нахождение отношения показателя, выявление инфицированных участков листа осуществляют на основании величины отношения, превышающей 1,5-2 для NPQ и превышающей 1,5-2,5 для ΦPSII, а также на основании пространственного распределения неоднородности.
2. Способ по п. 1, отличающийся тем, что показатели флуоресценции хлорофилла регистрируют с помощью системы визуализации импульсно-модулированной флуоресценции хлорофилла IMAGING-PAM MINI.
3. Способ по п. 1, отличающийся тем, что актиничный свет, представляющий собой синий свет с интенсивностью 102 мкмоль м-2 с-1, включают через 10 секунд, при этом его интенсивность обеспечивает величину показателя эффективного квантового выхода фотосистемы II FPSII, близкую к 0,5.
4. Способ по п. 1, отличающийся тем, что в качестве насыщающего света используют вспышки синего света 3000 мкмоль м-2 с-1 длительностью 800 мс.
5. Способ по п. 1, отличающийся тем, что используют сельскохозяйственные растения, произрастающие в условиях закрытого или открытого грунта, или листья таких растений, отделенные от них.
6. Способ по п. 1, отличающийся тем, что значение квантового выхода фотохимических реакций фотосистемы II ΦPSII и нефотохимического тушения флуоресценции NPQ рассчитывают автоматически с помощью программы для регистрации флуоресценции.
| СПОСОБ ОЦЕНКИ ЗИМОСТОЙКОСТИ ПЛОДОВЫХ РАСТЕНИЙ | 2012 |
|
RU2514400C2 |
| Дистанционный способ обнаружения стрессовых состояний растений | 2016 |
|
RU2646937C1 |
| СПОСОБ ФЛУОРЕСЦЕНТНОГО МЕЧЕНИЯ кДНК ВИРУСА ГРИППА ТИПА А | 2011 |
|
RU2480525C2 |
| СПОСОБ ДИАГНОСТИКИ ГЕРПЕСВИРУСНОЙ ИНФЕКЦИИ | 2001 |
|
RU2192473C1 |
| GRISHINA A | |||
| et al., Pre-Symptomatic Detection of Viral Infection in Tobacco Leaves Using PAM Fluorometry, Plants, 2021, 10, 2782 | |||
| S | |||
| HUPP et al., Noninvasive Phenotyping of Plant-Pathogen Interaction: Consecutive In Situ Imaging of | |||

