ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к биотехнологии и может быть использовано в сельском хозяйстве для увеличения выхода оздоровленных (безвирусных) миниклубней в гидропонных условиях или на торфе; при химической защите растений от действия абиотических стрессоров при последующем выращивании на открытом грунте или в сооружениях защищенного грунта.
УРОВЕНЬ ТЕХНИКИ
Из научной литературы известно, что в настоящее время производство 90% всей мировой агрономической продукции достигается за счет выращивания тридцати видов сельскохозяйственных культур, подавляющее большинство которых являются гликофитами. Картофель является четвертой по значимости основной продовольственной культурой в мире после риса, пшеницы и кукурузы. Растения картофеля диких видов относительно устойчивы к засолению, тогда как современные сорта, являющиеся продуктом долговременной селекции, значительно менее солеустойчивы. Картофель относится к орошаемым культурам, поэтому он часто сталкивается с неблагоприятным воздействием засоления.
Существующие в настоящее время агрономические и инженерные технологии минимизации засоления почв не способны решить эту проблему на глобальном уровне из-за их низкой эффективности, высоких трудозатрат и негативного влияния на окружающую среду.
Повышение стресс-устойчивости растений во многом определяется факторами гормональной природы. Среди фитогормонов как эффективных и экологически безопасных стресс-протекторных регуляторов наибольший интерес представляют стероидные гормоны растений брассиностероиды.
Известен способ повышения устойчивости растений рапса к повреждающему действию интенсивного хлоридного засоления стероидным гормоном растений – 24-эпибрассинолидом (ЭБЛ). Растения рапса культивируют на жидкой питательной среде в течение трех недель, последующие две недели растения подвергают хлоридному засолению 125 мМ с однократным внесением в раствор одновременно с началом засоления ЭБЛ в концентрации 0.01 мкМ. Показано, что протекторное действие гормона выражается в снижении отрицательного воздействия засоления на фотосинтетический аппарат растений за счет стабилизации транскрипции хлоропластных генов (Пат. RU 2514641).
Основной недостаток способа заключается в необходимости использования высокой концентрации действующего вещества – 0.01 мкМ, приводящий к повышенному расходу фитогормонов.
Известен способ повышения урожайности картофеля и томатов путем трехкратной в течение вегетации внекорневой подкормки растений раствором фитогормона 6-бензиламинопурина с концентрацией 100 мкМ при расходе 300–400 л/га (Пат. RU 2243658).
Основной недостаток способа – многократная обработка препаратом надземной части растений с использованием высокой концентрации гормона, что сопровождается увеличением временных и финансовых затрат.
Известен способ регуляции клубнеобразования и продуктивности растений картофеля при обработке растений картофеля брассинолидом, 24-эпибрассинолидом и 28-гомобрассинолидом. Способ повышает накопление фотосинтетических пигментов, ускоряет клубнеобразование и увеличивает продуктивность растений (Пат. RU 2660918, приятно за прототип).
Основной недостаток способа-прототипа заключается в его относительной низкой эффективности, так, например, гомобрассинолид и эпибрассинолид увеличивали количество миниклубней только на 6 или 23% соответственно. Другим недостатком способа-прототипа можно считать косвенную оценку влияния гормонов. Отсутствуют опытные данные о функционировании ассимиляционного аппарата, что немаловажно для конечной продуктивности растений. Кроме того, в известном способе эффективность гормонального воздействия рассмотрена только в оптимальных условиях выращивания, способность фитогормонов повышать продуктивность и урожай растений картофеля при действии техногенных стрессовых факторов не доказана.
ЦЕЛЬ ИЗОБРЕТЕНИЯ
Целью изобретения является разработка экономичного способа увеличения продуктивности растений картофеля, как в обычных условиях выращивания, так и при действии хлоридного засоления с использованием экологически чистых фитогормонов.
Задача решается краткосрочной обработкой корневой системы растений картофеля стероидными фитогормонами: 24-эпикастастероном и 24-эпибрассинолидом.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Для решения поставленной задачи в способе повышения продуктивности растений картофеля, включающем кратковременную обработку корневой системы растений картофеля раствором биологически активного вещества, в процессе адаптации растений к жидкой питательной среде увеличивают площадь ассимилирующей поверхности, содержание в них фотосинтетических пигментов и эффективность столонобразования, однократно обрабатывая корневую систему растений раствором 24-эпикастастерона в концентрации 0.01-1 нМ в течение 4-5 часов, а также повышают активность антиоксидантных ферментов, увеличивают эффективность фотосинтеза и снижают избыточное поступление токсичных неорганических ионов, обрабатывая корневую систему растений раствором 24-эпибрассинолида в концентрации 0.01-1 нМ в течение 4-5 часов до солевого воздействия.
Известно, что продуктивность растения определяется не только его массой, но и его фотосинтетической активностью листьев, которая напрямую определяется площадью ассимилирующей поверхности и содержанием фотосинтетических пигментов [Zivcak et al., 2017]. В условиях действия хлоридного засоления важно поддерживать не только оптимальную фотосинтетическую активность, но и ограничивать поступление токсичных неорганических ионов или нейтрализовать их избыток за счёт стимулирования ферментативной и неферментативной защитных систем. В процессе экспериментального выбора наиболее эффективных ростовых гормонов, их концентрации от 0.01 нМ до 1 нМ и длительности воздействия этим характеристикам было уделено особое внимание. Обработку корневой системы гормоном варьировали от 1 до 24 часов, оптимальный период – не менее 4х часов, увеличение времени замачивания свыше 5 часов вплоть до одних суток не дает заметного эффекта. Среди исследованных фитогормонов наиболее значимые результаты показали кетонсодержащий (24-эпикастастерон) и лактонсодержащий (24-эпибрассинолид) стероидные гормоны.
Значительное повышение продуктивности растений в отсутствии действия стрессоров и протекторное действие стероидных гормонов различной химической структуры при хлоридном засолении в отношении растений картофеля иллюстрируется таблицами и рисунками. Стимулирующий и стресс-протекторный эффекты брассиностероидов доказан при концентрации гормонов 0,01 – 1 нМ (10-9 – 10-11 М), преимущественно 0,1 нМ, при продолжительности воздействия 4-5 часов для ЭПК и от 4 до 5 часов для ЭБЛ (в качестве примеров в таблицах и на рисунках приведены данные только для этих вариантов обработки растений).
ПОЯСНЯЮЩИЕ ТАБЛИЦЫ И ГРАФИКИ
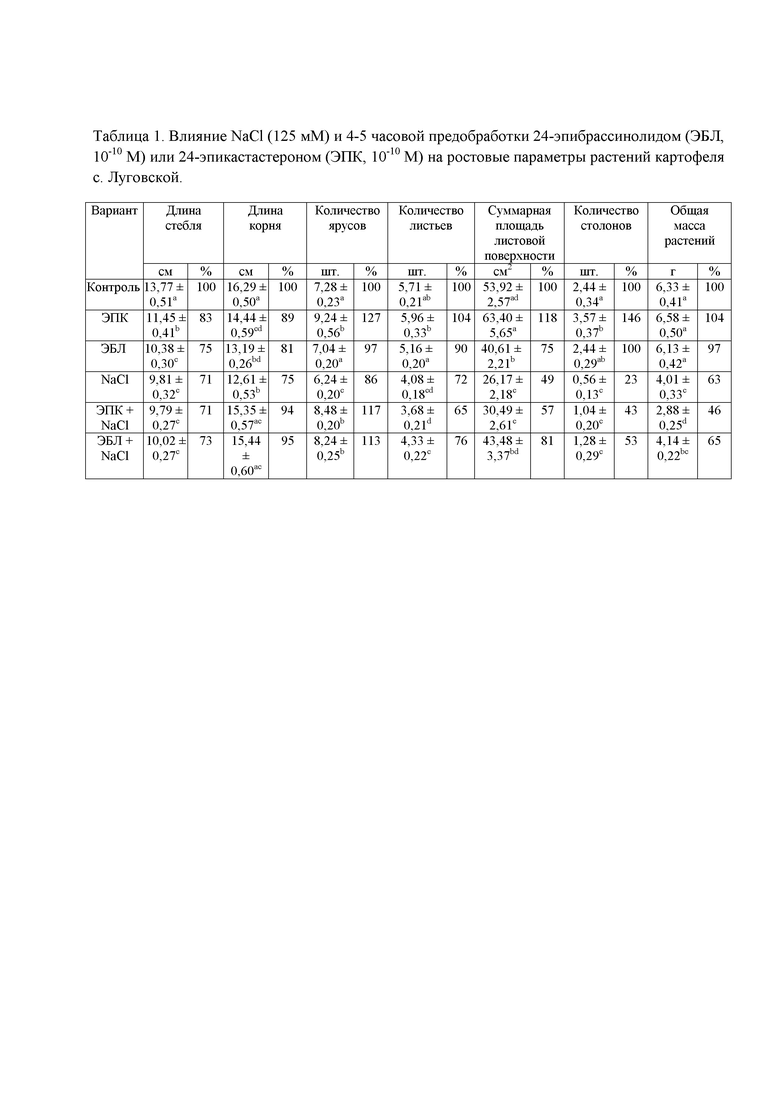
В таблице 1 представлены ростовые показатели растений картофеля. Буквы указывают на наличие достоверных различий (p˂0.05) между вариантами.
В таблице 2 отражена фотохимическая активность фотосинтетического аппарата растений. Буквы указывают на наличие достоверных различий (p˂0.05) между вариантами
Y(II) – эффективный квантовый выход ФС II;
ETR – скорость переноса электронов;
qP и qL – коэффициенты фотохимического тушения флуоресценции, которые оценивают долю открытых реакционных центров ФС II;
qN – коэффициент нефотохимического тушения флуоресценции (отражает степень диссипации поглощенной энергии в тепло);
NPQ – нефотохимическое тушение флуоресценции: количественная оценка нефотохимического тушения, альтернативная расчетам qN;
Y(NO) – квантовые выходы нерегулируемой нефотохимической диссипации световой энергии в ФС II;
Y(NPQ) – квантовые выходы регулируемой нефотохимической диссипации световой энергии в ФС II;
Fo' – минимальный уровень флуоресценции, возбуждаемый измерительным светом низкой интенсивности, так что при его действии все реакционные центры ФС II остаются открытыми;
Fm' – максимальный выход флуоресценции, в момент подачи сильного светового импульса при закрытых реакционных центрах ФС II.
FV/FM – максимальный квантовый выход фотосистемы II (ФС II).
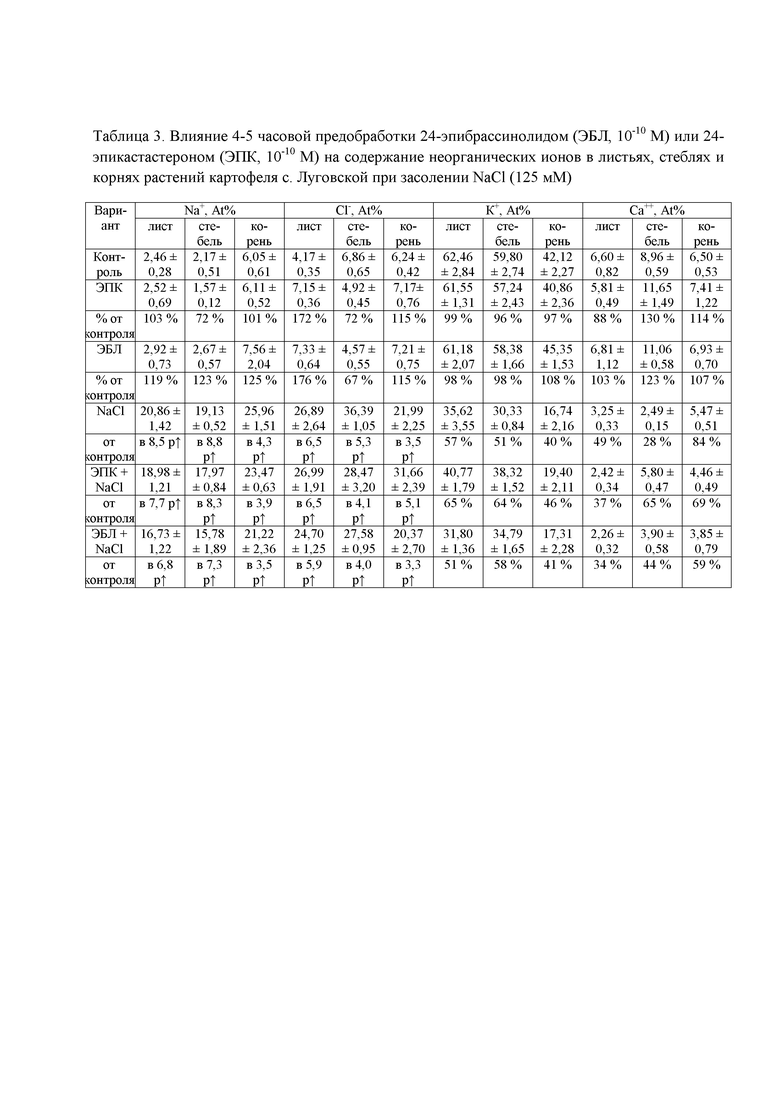
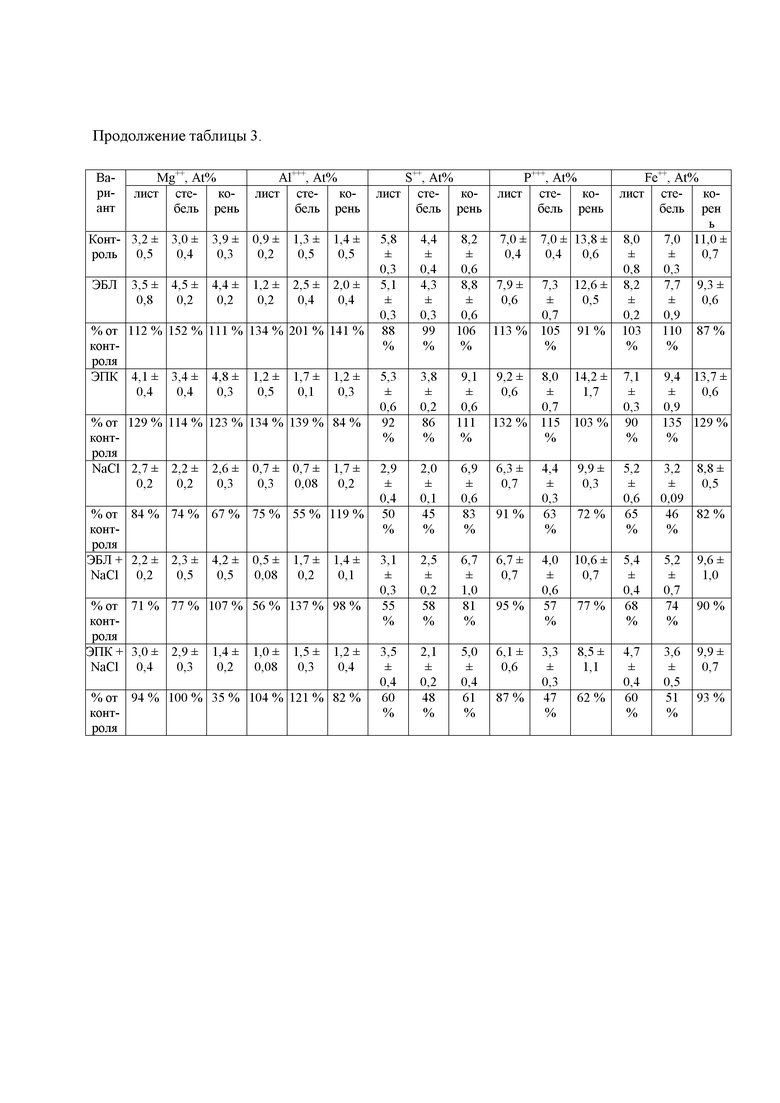
В таблице 3 показана динамика неорганических ионов в разных частях растения.
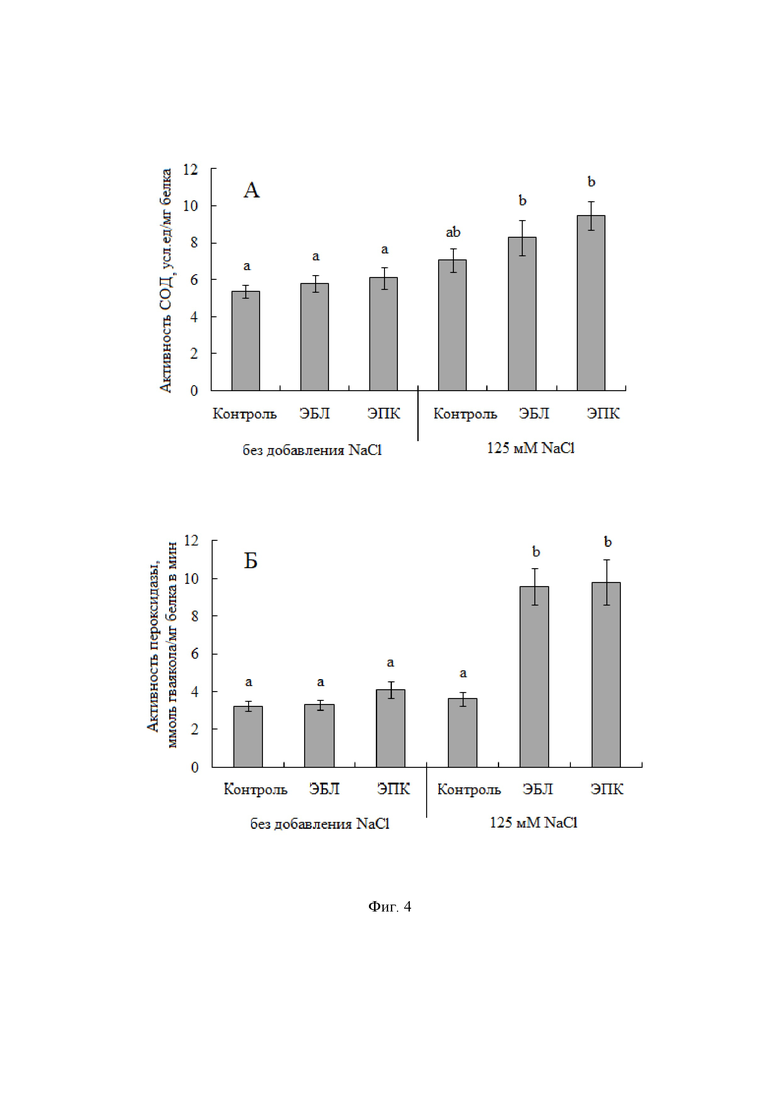
На фиг. 1, 2, 3, 4 показаны гистограммы, отражающие физиологическое состояние растений картофеля на примере следующих показателей – фотосинтетических пигментов, осмотического потенциала клеточного содержимого, пролина, малонового диальдегида, антиоксидантных ферментов.
Проведены испытания влияния гормонов как в оптимальных условиях, так и при последующем солевом стрессовом факторе (NaCl в концентрации 125 мМ).
ПРИМЕРЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Примеры иллюстрируют действие на растение стероидных гормонов при разной химической структуре молекулы (кетонсодержащим 24-эпикастастероном или лактонсодержащим 24-эпибрассинолидом). Опыты проведены на растениях Solanum tuberosum L. среднеспелого сорта Луговской (идентификатор 8301891). В оптимальных условиях картофель данного сорта дает стабильно высокий урожай, его клубни характеризуются высокой лёжкостью и устойчивостью к ряду заболеваний, в том числе, к фитофторозу. Оздоровленные растения-регенеранты картофеля in vitro получали из апикальной меристемы и на протяжении 30 суток культивировали на агаризованной питательной среде Мурасиге и Скуга с половинным составом среды (½ МС). Корни растений отмывали от агаризованной среды и проводили недельную адаптацию микроклонов к жидкой половинной среде Мурасиге и Скуга (½ МС) и условиям воздушной среды под люминесцентными лампами L36W/77 Fluora («Osram», Германия) при плотности потока квантов ФАР 100–150 мкмоль∙м-2∙с-1 в фитотроне с 16-часовым фотопериодом и температурой 20 ± 3°C.
После двухнедельного роста на гидропонной установке в среде МС растения переносили на непродолжительное время на ту же самую среду в отсутствие (контрольный вариант) или с добавлением 24-эпикастастерона (ЭПК) до различной концентрации в пределах 0.01-1 нМ. Также исследовали влияние солевого стрессового фактора на контрольные и обработанные гормонами экземпляры: растения помещали на питательную среду МС в отсутствии (для оценки эффективности гормонального воздействия в оптимальных условиях роста), а также моделировали стрессовые условия (присутствие NaCl в концентрации125 мМ).
Полученную свежую биомассу растительного материала оценивали гравиметрическим методом. Для оценки уровня фотосинтетических пигментов листья картофеля растирали в 96%-ном этаноле, гомогенат центрифугировали (10 мин при 8 тыс. об./мин, центрифуга MiniSpin "Eppendorf", Германия). Оптическую плотность пробы (супернатанта) оценивали на спектрофотометре Genesys 10 ThermoElectron (США). Концентрацию пигментов в спиртовой вытяжке рассчитывали согласно методу, предложенному H.K. Lichtenthaler, 1987.
Измерение флуоресцентных показателей, отражающих фотохимическую активность фотосинтетического аппарата растений, проводили на РАМ-флуориметре (Junior-PAM, “Heinz-Walz”, Germany). Параметры флуоресценции регистрировали после 20-минутной темновой адаптации.
Экстракцию и определение свободного пролина проводили по методу, предложенному L.S. Bates, 1973.
Осмотический потенциал клеточного экссудата определяли на криоскопическом осмометре Osmomat 030 (Германия) в соответствии с инструкцией производителя. Клеточный сок отжимали из размороженных образцов листьев растений.
Определение активности антиоксидантных ферментов (супероксиддисмутазы (ЕС 1.15.1.1) и пероксидазы (ЕС 1.11.1.7) в листьях растений оценивали по методам, описанным Ch. Beauchamp, 2002. Концентрацию белка в полученных ферментных препаратах оценивали по методу, предложенному A.A. Esen, 1978.
Содержание неорганических ионов (натрия, калия, хлора, алюминия, фосфора, магния, кальция, серы и железа) определяли в листьях, стеблях и корнях методом энергодисперсионного анализа на электронном ионно-растровом сканирующем микроскопе «Quanta 200 3D» EDAX (Нидерланды). Данные представлены в атомарных процентах (At %) от общего количества элементов в данной точке образца (100 At %) за вычетом доли углерода и кислорода. Сканирование каждого варианта проводили не менее шести раз.
В качестве ключевых показателей ответа растений на гормональное и солевое воздействие использовали интегральные показатели: размеры осевых органов, суммарная площадь ассимилирующей поверхности, количество ярусов и столонов.
Результаты экспериментов показали повышение продуктивности растений в оптимальных условиях произрастания и снижение эффекта негативного воздействия хлоридного засоления на ростовые и физиологические показатели картофеля.
В оптимальных условиях выращивания растений экзогенный 24-эпибрассинолид практически не оказывает никакого воздействия на ростовые показатели растений картофеля, в то время как его предшественник – 24-эпикастастрон, способствует увеличению количества столонов, ярусов и суммарной площади листовой поверхности на 46, 27 и 18% соответственно (табл. 1). Содержание зелёных пигментов (хлорофиллов a и b) повышалось при 4х и более часовом воздействии ЭПК в бóльшей степени для хлорофилла b (в среднем на 33%).
Известно, что NaCl в высоких концентрациях оказывает не только прямое токсическое действие на клеточный метаболизм и вызывает осмотический стресс, но и стимулирует генерацию АФК и развитие окислительного стресса. Основная причина окислительного стресса в этом случае связана с закрыванием устьиц, снижением доступности СО2 и повышением энергии возбуждения электронов, что сопровождается интенсивной генерацией АФК.
Для выявления протекторного эффекта стероидных гормонов в условиях действия абиотического стрессора, нами была проведена оценка их эффективности на фоне хлоридного засоления (см. табл. 1). Использованная в эксперименте концентрация NaCl 125 мМ подавляет столонообразование в 4.4 раза и снижает суммарную площадь листовой поверхности в 2.0 раза. Длина осевых органов уменьшалась на 25%, общая масса растений в ответ на действие стрессора снижалась в 1.5 раза. Обработка растений гормонами снижает ингибирующее действие засоления 125 мМ NaCl на длину корня, площадь ассимилирующего аппарата, количество ярусов, листьев и столонов. Негативный эффект NaCl проявляется не только на ростовых характеристиках, но и на содержании фотосинтетических пигментов; уровень зелёных пигментов в контроле уменьшался на 15-20% (фиг. 1).
Эффективность защитного действия в отношении площади листовой поверхности и столонообразования зависит от химической структуры вещества. Так, протекторный эффект ЭПК отмечен на уровне столонообразования, в то время как специфика действия ЭБЛ проявляется в стимуляции развития ассимилирующего аппарата (табл. 1). Негативное действие хлоридного засоления на накопление фотосинтетических пигментов полностью снимается предварительной обработкой ЭПК; вместе с тем положительный эффект экзогенного ЭБЛ был наиболее выражен – содержание хлорофиллов и каротиноидов превышало контрольные значения на 20% (фиг. 1). Проведенные дополнительные опыты с 28-гомобрассинолидом до начала действия хлоридного засоления (NaCl 100 мМ) также подтвердили, что лактонсодержащие стероидные гормоны более эффективны для борьбы со стрессором за счёт аккумуляции пролина и каротиноидов, обладающих выраженными антиоксидантными и стресс-протекторными свойствами.
Учитывая ключевую роль фотосинтетического аппарата в продуктивности растений и в их выживании при неблагоприятных условиях произрастания, большое значение имеет оценка показателей, отражающих функционирование ассимиляционного аппарата растений.
Максимальная квантовая эффективность фотосистемы II (Fv/Fm) контрольных растений составляла 0.83, хлоридное засоление снижало данный показатель до 0.78, что свидетельствует о нарушении функционирования фотосинтетического аппарата (табл. 2). В соответствии с этим эффективный квантовый выход ФС II (Y(II)) понижался на 20%. Коэффициенты фотохимического тушения флуоресценции qP и qL в ответ на действие стрессора также снизились. Кроме того, о нарушении эффективности работы ФС II свидетельствует снижение скорости переноса электронов ETR и увеличение квантового выхода регулируемой утилизации Y(NPQ).
Кратковременная обработка растений гормонами снижает последующее ингибирующее действие соли в отношении максимального и эффективного квантовых выходов фотосистемы II, а также коэффициенты фотохимического тушения флуоресценции qP до контрольных значений. В ответ на гормональное воздействие частично восстанавливались скорость передачи электронов и коэффициент фотохимического тушения qL. В отношении квантового выхода регулируемой утилизации выявлена специфика действия для кетон- и лактонсодержащих стероидных соединений – ЭПК полностью восстанавливал данный показатель, в то время как ЭБЛ удерживал данный показатель на уровне солевого контроля (табл. 2).
Величина осмотического потенциала клеточного содержимого листьев в течение эксперимента в контрольных растениях составляла (– 0,75 МПа). Хлоридное засоление вызывало снижение химического потенциала клеточного экссудата до (– 1,4 МПа), что было 1,8 раза ниже аналогичных значений контрольного варианта (фиг. 2). Гормональный прайминг с последующим солевым стрессом не оказывал достоверного влияния на поступление воды в растения по сравнению с действием стрессора (фиг. 2); поддержание осмотического потенциала на низком уровне позволяет растению восстанавливать направление градиента водного потенциала между питательной средой и клетками растения и поглощать воду из среды с низким водным потенциалом.
Решающую роль в формировании осмотического потенциала клеточного содержимого играют неорганические ионы. Нами проанализировано содержание ионов натрия, хлора, калия, кальция, магния, алюминия, фосфора, серы и железа в листьях, стеблях и корнях растений картофеля. Как следует из полученных данных (табл. 3), в отсутствие стрессора атомарная концентрация (At%) натрия в подземной части растений примерно в три раза превышала его содержание в листьях и стеблях, а At% ионов калия, наоборот, была выше в надземной части. Среди преобладающих ионов можно отметить ионы калия. В ответ на воздействие NaCl на корневую систему At% неорганических ионов принципиально изменялось. Основную массу среди девяти анализируемых ионов составляли ионы натрия, хлора и калия. Максимальное увеличение процентной доли ионов натрия и хлора показано для надземной части – примерно в 9 и 6 раз соответственно, минимальное наблюдалось в корнях – в 4 раза (табл. 3). At% К+ уменьшилась во всех частях растения в ответ на действие стрессора примерно в два раза. Атомарная концентрация кальция, серы и железа снизилась примерно в два раза в надземной части и на 15% в корнях растений; доля ионов магния во всех частях растения уменьшилась на 16-33%.
Обработка растений картофеля лактонсодержащим 24-эпибрассинолидом с последующим хлоридным засолением приводила к снижению атомарной доли ионов натрия хлора во всех частях растения и незначительному увеличению At% ионов калия, что свидетельствует о снижении негативного эффекта стрессора (табл. 3).
Авторы полагают, что снижение концентрации неорганических ионов при воздействии фитогормонами происходит за счет повышения эффективности барьерных механизмов, которые задерживают поступление токсичных ионов в растения. Защитное действие кратковременной обработки кетонсодержащим 24-эпикастастероном было менее выражено и проявлялось только в снижении атомарной доли ионов натрия; несмотря на снижение At% ионов хлора в стеблях растений, в корнях наблюдалось его увеличение, что не может гарантировать положительное влияние данного гормона на накопление ионов хлора в растении.
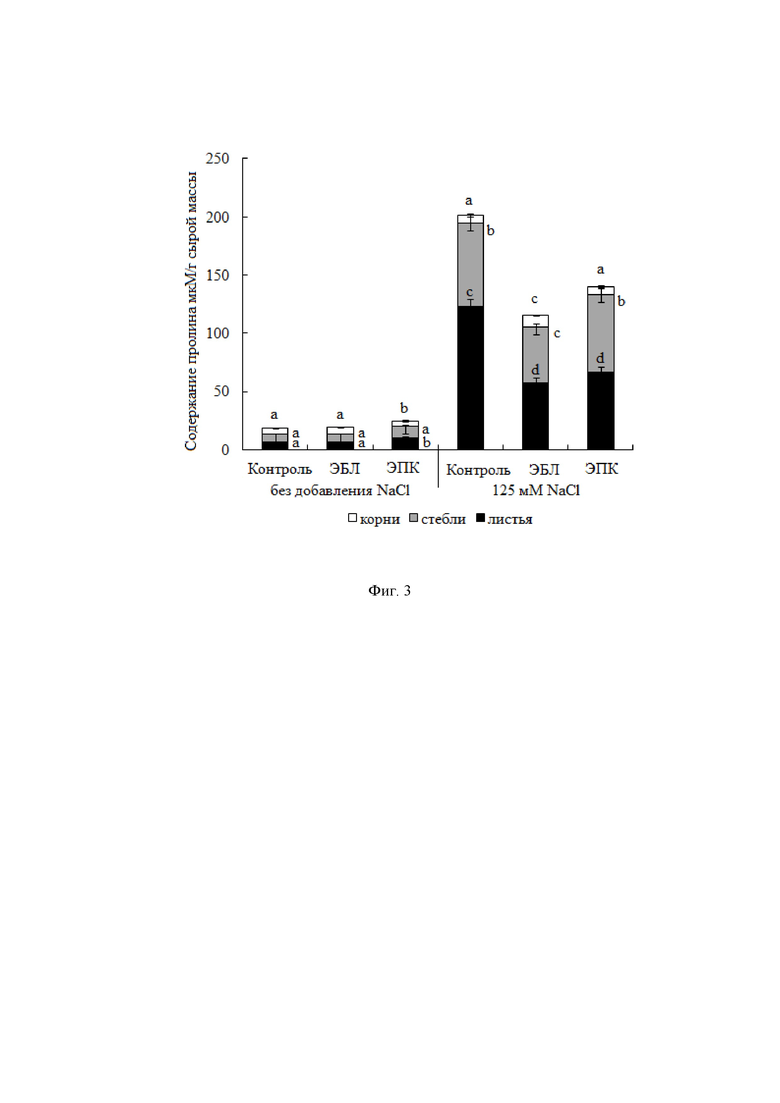
При адаптации растений к нарушению водного статуса и к токсичному действию избытка неорганических ионов, прежде всего натрия, важная роль принадлежит совместимым осмолитам, обладающим свойствами химических шаперонов. Нами была выявлена органоспецифичность в накоплении универсального совместимого осмолита пролина в растениях картофеля в ответ на действующие факторы. Содержание пролина во всех частях растений в отсутствие стрессора составляло 5.4-6.5 мкМ/г сырого веса (фиг. 3). Хлоридное засоление активировало накопление пролина в листьях и стебле растений в 19 и 10 раз соответственно (фиг. 3). Накопление пролина при стрессе способствует сохранению клеточного гомеостаза. Об этом свидетельствует тот факт, что пролин при стрессе играет роль не только осморегулятора, хотя при засолении это крайне важно, но и реализует целый ряд других стресс-протекторных функций, таких как функции химического шаперона, антиоксиданта, регулятора экспрессии стресс-регулируемых генов, источника углерода, азота и восстановительных эквивалентов, регулятора внутриклеточного рН-стата и т.д.) (Mansour M.M.F., Ali E.F., 2017). Содержание пролина при кратковременном гормональном воздействии с последующим хлоридным засолением зависит от химической структуры гормона. Так, корневая обработка растения лактонсодержащим ЭБЛ до солевого стресса (NaCl 125 мМ) способствовала более эффективному снижению уровня пролина по сравнению с его содержанием в растениях, подвергнутых действию ЭПК, особенно для стеблей и листьев (фиг. 3).
При развитии окислительного стресса в ответ на действие хлоридного засоления растение отвечает формированием клеточной антиоксидантной системы, которая включает в себя кроме низкомолекулярных органических соединений, антиоксидантные ферменты – супероксиддисмутазу (СОД), пероксидазу и др. Хлоридное засоление способствовало незначительной активации ферментативных систем в листьях растений картофеля; активность СОД и гваякол-зависимой пероксидазы повышалась на 32 и 12% соответственно (фиг. 4). Кратковременная обработка растений ЭПК и ЭБЛ до стрессового воздействия вызывала увеличение активности супероксиддисмутазы на 54-76% (фиг. 4А), пероксидазы в 3 раза и практически не зависит от химической структуры использованных гормонов (фиг. 4Б).
Таким образом, экспериментально показано, что кратковременная корневая предобработка растений картофеля 24-эпикастастероном способствует увеличению количества столонов, ярусов, суммарной площади листовой поверхности и содержанию хлорофиллов в отсутствие действия стрессора. Корневая предобработка растений картофеля 24-эпибрассинолидом снижает отрицательное воздействие последующего хлоридного засоления на ростовые показатели, содержание фотосинтетических пигментов, максимальный и эффективный квантовые выходы второй фотосистемы, коэффициенты фотохимического тушения флуоресценции qP. Протекторный эффект гормонов достигается за счёт снижения атомарной доли ионов хлора и натрия и увеличения активности антиоксидантных ферментов, содержание пролина при этом уменьшается. Более выраженный защитный эффект при действии хлоридного засоления показал лактонсодержащий ЭБЛ.
На основании этих результатов предложен способ повышения продуктивности растений картофеля в оптимальных условиях произрастания и его агрономической устойчивости в условиях хлоридного засоления согласно заявленной формуле изобретения.
Использованные источники
1. Пат. RU 2514641. Опубл.: 27.04.2014.
2. Пат. RU 2243658. Опубл: 10.01.2005.
3. Пат. RU 2660918. Опубл: 11.07.2018.
4. Mansour M.M.F., Ali E.F. Evaluation of proline functions in saline conditions // Phytochemistry. 2017. Vol. 140. P. 52–68.
5 Zivcak M., Olsovska K., Brestic M. Photosynthetic Responses Under Harmful and Changing Environment: Practical Aspects in Crop Research // Photosynthesis: Structures, Mechanisms, and Applications. 2017. P. 203-248.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ повышения продуктивности растений картофеля эпибрассинолидом в оптимальных условиях и при хлоридном засолении | 2023 |
|
RU2821570C1 |
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ РАСТЕНИЙ РАПСА К ИНТЕНСИВНОМУ ХЛОРИДНОМУ ЗАСОЛЕНИЮ | 2015 |
|
RU2603091C2 |
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ РАСТЕНИЙ РАПСА К ИНТЕНСИВНОМУ ХЛОРИДНОМУ ЗАСОЛЕНИЮ | 2013 |
|
RU2522519C1 |
| СПОСОБ СТАБИЛИЗАЦИИ ТРАНСКРИПЦИИ ХЛОРОПЛАСТНЫХ ГЕНОВ РАПСА В УСЛОВИЯХ ХЛОРИДНОГО ЗАСОЛЕНИЯ | 2012 |
|
RU2514641C1 |
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ РАСТЕНИЙ РАПСА К ХЛОРИДНОМУ ЗАСОЛЕНИЮ | 2012 |
|
RU2515726C1 |
| СПОСОБ РЕГУЛИРОВАНИЯ КЛУБНЕОБРАЗОВАНИЯ И ПРОДУКТИВНОСТИ РАСТЕНИЙ КАРТОФЕЛЯ В УСЛОВИЯХ ГИДРОПОНИКИ | 2017 |
|
RU2660918C1 |
| СПОСОБ ПОВЫШЕНИЯ СТРЕССОУСТОЙЧИВОСТИ ЗЕРНОВЫХ КУЛЬТУР | 2018 |
|
RU2703022C1 |
| СПОСОБ ОЦЕНКИ АДАПТИВНОГО ПОТЕНЦИАЛА ЗЕРНОВЫХ КУЛЬТУР ПРИ СТРЕССОВОМ ВОЗДЕЙСТВИИ | 2017 |
|
RU2651284C1 |
| СПОСОБ РЕГУЛЯЦИИ ТРАНСКРИПЦИИ ХЛОРОПЛАСТНЫХ ГЕНОВ ЯЧМЕНЯ (HORDEUM VULGARE) | 2011 |
|
RU2460279C1 |
| ОРГАНОМИНЕРАЛЬНОЕ УДОБРЕНИЕ ДЛЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР | 2016 |
|
RU2633796C1 |



Изобретение относится к биотехнологии и может быть использовано в сельском хозяйстве для увеличения выхода оздоровленных миниклубней картофеля в гидропонных условиях или на торфе. Способ включает обработку растений раствором биологически активных веществ. При этом в процессе адаптации к жидкой питательной среде увеличивают площадь ассимилирующей поверхности растений, содержание в них фотосинтетических пигментов и эффективность столонобразования путем однократной обработки корневой системы растений раствором 24-эпикастастерона в концентрации 0.01-1 нМ в течение 4-5 ч. Также повышают активность антиоксидантных ферментов и снижают поступление токсичных неорганических ионов путем обработки корневой системы растений раствором 24-эпибрассинолида в концентрации 0.01-1 нМ в течение 4-5 ч до солевого воздействия. Техническим результатом является увеличения продуктивности растений картофеля как в обычных условиях выращивания, так и при действии хлоридного засоления. 4 ил.,3 табл.
Способ повышения продуктивности растений картофеля в оптимальных и стрессовых условиях выращивания, включающий кратковременную обработку растений раствором биологически активного вещества, отличающийся тем, что в процессе адаптации к жидкой питательной среде увеличивают площадь ассимилирующей поверхности растений, содержание в них фотосинтетических пигментов и эффективность столонобразования, однократно обрабатывая корневую систему растений раствором 24-эпикастастерона в концентрации 0.01-1 нМ в течение 4-5 ч, а также повышают активность антиоксидантных ферментов и снижают избыточное поступление токсичных неорганических ионов, обрабатывая корневую систему растений раствором 24-эпибрассинолида в концентрации 0.01-1 нМ в течение 4-5 ч до солевого воздействия.
| СПОСОБ РЕГУЛИРОВАНИЯ КЛУБНЕОБРАЗОВАНИЯ И ПРОДУКТИВНОСТИ РАСТЕНИЙ КАРТОФЕЛЯ В УСЛОВИЯХ ГИДРОПОНИКИ | 2017 |
|
RU2660918C1 |
| ПРИСПОСОБЛЕНИЕ ДЛЯ ПОДАЧИ МАТЕРИАЛА К ПРОКАТНОМУ СТАНУ С ВОЗВРАТНО-ПОСТУПАТЕЛЬНЫМ ДВИЖЕНИЕМ | 1926 |
|
SU5698A1 |
| Способ выращивания картофеля | 1981 |
|
SU1015868A1 |
| JP S63170306 A, 14.07.1988 | |||
| CN 108094104 A, 01.06.2018 | |||
| US 8119855 B2, 21.02.2012 | |||
| CN 105940901 B, 21.05.2019. | |||

