Область техники
Настоящее изобретение относится к экзосомам, нагруженным FAF1, и их применению в качестве противоопухолевого средства.
Уровень техники
Опухоль является результатом беспорядочной гиперпролиферации аномальных клеток, и когда такая опухоль обладает деструктивными пролиферативными, инвазивными и метастатическими свойствами, то ее классифицируют как злокачественную опухоль, то есть рак. Смертность от рака занимает первое место среди причин смертности в Корее и является одной из важнейших причин смертности в развитых странах. К настоящему времени для лечения рака разрабатывалось и применялось хирургическое лечение до 1950-х годов, лучевая терапия - в 1960-х годах, и противоопухолевая лекарственная терапия - в 1970-х годах. В 1980-х годах благодаря радикальному развитию фундаментальной науки, особенно иммунологии и молекулярной биологии, начали появляться иммунотерапия и генная терапия, и многие исследования в настоящее время продолжаются, но их все еще недостаточно для достижения прорыва в существующих методах лечения.
Fas (также известный как Fas-рецептор, CD95, Apo1 или TNFRSF6) представляет собой рецептор, обладающий наиболее высокой способностью к киллингу клеток среди рецепторов клеточной гибели, и Fas и лиганд Fas (Fas L) играют важную роль в апоптозе. Когда три лиганда Fas связываются с тремя молекулами Fas, то адаптерный белок, называемый Fas-ассоциированным белком с доменом смерти (FADD), связывается с доменом смерти (DD) Fas. Затем, когда эффекторный домен смерти (DED) FADD объединяется с эффекторным доменом смерти (DED) неактивной каспазы-8 (также известной как FLICE или MACH), то каспаза-8 активируется, и эти Fas, FADD и каспаза-8 образуют сигнальный комплекс, индуцирующий клеточную гибель Fas (Fas-DISC), и в конечном итоге активируют эффекторную каспазу, которая вызывает клеточную гибель, тем самым индуцируя гибель клеток.
Fas-ассоциированный фактор 1 (FAF1) представляет собой белок, связанный с путем Fas, и он был обнаружен у мышей (Chu et al., 1995; Becker et al., 1997), перепелов (Frohlich et al., 1998) и людей (Ryu и др., 1999). Белки имеют высокую гомологию аминокислотных остатков друг с другом: FAF1 у мышей состоит из 649 аминокислот, FAF1 у перепелов состоит из 648 аминокислот, и FAF1 у человека состоит из 650 аминокислот.
В предыдущих исследованиях FAF1 было обнаружено, что FAF1 обладает способностью индуцировать апоптоз, и минимальной областью индукции является область из аминокислот 181-381 FAF1. Установлено, что FAF1 является членом Fas-DISC, и DISC образуется в результате взаимодействия DED каспазы-8 и FADD, и области из аминокислот 181-381 FAF1, структурно подобной DED (Ryu and Kim, 2003).
Сообщалось, что уровень экспрессии FAF1 снижается при различных видах рака, включая рак легкого, рак толстого кишечника, рак печени, рак предстательной железы, рак мозга и рак молочной железы (Feng et al., 2017), и белок FAF1 привлекает внимание в качестве супрессора опухолей, и известно, что FAF1 подавляет рост опухолей посредством ингибирования NF-κB и подавляет метастазы опухолей через сигналы TGF-β (Park et al., 2004; Park et al., 2007). Кроме того, известно, что FAF1 ингибирует пролиферацию опухолей, останавливая клеточный цикл в опухолях в фазе G2/M посредством ингибирования Aurora-A (Jang et al., 2008). В выложенной публикации корейского патента №1020040101707 раскрывается, что фрагмент специфической области FAF1 обладает активностью ингибирования ангиогенеза и образования трубочек, а также пролиферации клеток, и, таким образом, данный фрагмент можно использовать в качестве супрессора опухолей.
Внеклеточные везикулы были впервые открыты 50 лет назад, и с тех пор было обнаружено, что везикулы находятся во всех тестированных биологических жидкостях, и известно, что клеточные линии in vitro выделяют везикулы на различных уровнях. Среди внеклеточных везикул экзосома представляет собой везикулу, состоящую из фосфолипидного бислоя и имеющую размер 50-200 нм, которая продуцируется в клетке и секретируется наружу и, как было обнаружено, играет важную роль в межклеточной коммуникации посредством межклеточной доставки специфического репертуара нуклеиновых кислот (ДНК и РНК), белков и липидов, важных для гомеостаза. Например, экзосомы участвуют в основных физиологических процессах, таких как нейротрансмиссия, презентация антигенов, иммунный ответ, развитие органов и репродуктивная функция, и также вовлечены в развитие некоторых патологических расстройств, включая прогрессирование рака, сердечно-сосудистые заболевания, воспаления и передачу прионов.
Экзосомы были впервые открыты в процессе высвобождения и удаления внутриклеточных белков на конечной стадии созревания эритроцита, в результате чего в эритроците остается только гемоглобин, и при исследовании с помощью электронного микроскопа было установлено, что эти экзосомы высвобождаются и секретируются во внеклеточное пространство, происходя из специфических компартментов клетки, называемых мультивезикулярными тельцами (MVB), а не непосредственно при отделении от плазматической мембраны. То есть, когда мультивезикулярные тельца сливаются с плазматической мембраной, то такие везикулы высвобождаются во внеклеточную среду, и называются экзосомами.
Проводится множество исследований по изучению молекулярного механизма образования экзосом, и известно, что различные типы иммунных клеток, включая B-лимфоциты, Т-лимфоциты, дендритные клетки, мегакариоциты, макрофаги и т.п., а также стволовые клетки и опухолевые клетки, продуцируют и секретируют экзосомы. Хотя сообщалось, что продукция экзосом зависит или не зависит от эндосомального сортировочного комплекса, необходимого для транспорта (ESCRT), трудно определить точный механизм.
В выложенной публикации корейского патента №20040015508 в качестве примера близкого уровня техники раскрывается способ, в котором ген специфического антигена вводят в клеточную линию, и белок введенного гена стабильно экспрессируется в клеточной линии и высвобождается во внеклеточное пространство через экзосомы, и раскрывается способ применения экзосом в качестве вакцины. Кроме того, корейский патент №102053065 относится к рН-чувствительной экзосомальной композиции с использованием гиалуроновой кислоты и доксорубицина, и в нем раскрывается способ получения рН-чувствительной экзосомы с использованием доксорубицина и высокомолекулярного вещества, химически связывающего гиалуроновую кислоту и 3-диэтиламинопропиламин, и киллинговый эффект экзосомы для опухолевых клеток. Кроме того, выложенная публикация корейского патента №1020180078173 относится к новому противоопухолевому агенту на основе экзосом, и в ней раскрывается рекомбинантная экзосома, в которой на ее поверхности находится рецепторная тирозинкиназа и SIRP, которые представляют собой белки, способствующие фагоцитозу, и рекомбинантная экзосома, включающая аспарагиназу, белковый токсин, антитело, специфичное к опухолевому антигену, или фрагмент антитела, ген-супрессор опухоли или антиангиогенный фактор и т.п., которые являются противоопухолевыми белками. Кроме того, в заявке на патент Кореи №1020190059724 раскрывается способ скрининга потенциальных лекарственных препаратов для лечения нейродегенеративных заболеваний с использованием подтверждения того, что FAF1 нагружается в экзосомы и секретируется во внеклеточное пространство, но не сообщалось о результатах исследований, связанных с опухолевыми клетками.
При использовании экзосом, вещества-грузы защищены от внеклеточной среды, что приводит к более длительному периоду полураспада и, возможно, проникновению в клетку-мишень. Однако не все белки загружаются в экзосомы в качестве веществ-грузов, и даже при загрузке в экзосомы нагруженные вещества-груза не проникают во все клетки. Кроме того, технически сложно загружать высокомолекулярные белки большого размера внутрь экзосомы, и эффективность нагруженного белка может существенно снизиться, если белок сконструирован для загрузки. Сообщалось, что FAF1 спонтанно загружается в экзосому нервной клетки, но вопрос о том, насколько FAF1, загруженный в экзосому, проникает в клетку и правильно функционирует, зависит от типа клетки, и существует ограничение в том, что это должно быть доказано экспериментально. Следовательно, если будет подтверждено, что доставка FAF1 с использованием экзосом функционирует правильно в опухолевых клетках и станет возможным массовое производство экзосом, нагруженных FAF1, то экзосомы, нагруженные FAF1, можно использовать в качестве супрессора опухолей при различных видах рака.
Следовательно, для определения того, насколько FAF1, известный как обладающий функцией подавления опухолей, загружается в экзосомы, для его доставки к различным типам опухолей, и насколько FAF1, загруженный в экзосомы, подавляет образование опухолей, авторы настоящего изобретения выделили экзосомы из клеток, в которых FAF1 сверхэкспрессируется, проверили тот факт, что FAF1 спонтанно загружается в выделенные экзосомы, подтвердили, что при обработке опухолевых клеток, обработанных выделенными экзосомами, нагруженными FAF1, образование колоний подавляется, и подтвердили эффект достоверного ингибирования роста опухолей по сравнению с другими контрольными группами, включая группы, обработанные рекомбинантными белками FAF1, когда экзосомы, нагруженные белком FAF1, вводили в различные опухолевые модели на мышах, которым трансплантировали опухолевые клетки, и тем самым завершили настоящее изобретение.
Техническая проблема
Целью настоящего изобретения является обеспечение фармацевтической композиции для лечения рака, содержащей экзосомы, нагруженные белком FAF1, в качестве активного ингредиента.
Другой целью настоящего изобретения является обеспечение способа получения экзосом, нагруженных белком FAF1.
Еще одной целью настоящего изобретения является обеспечение способа скрининга противоопухолевых терапевтических средств, для опухолей, имеющих пониженную экспрессию FAF1.
Решение технической проблемы
Для достижения вышеуказанных целей, настоящее изобретение обеспечивает фармацевтическую композицию для лечения рака, содержащую экзосомы, нагруженные белком FAF1, в качестве активного ингредиента.
Кроме того, настоящее изобретение относится к способу получения экзосом, нагруженных белком FAF1, где способ включает стадии: 1) введение полинуклеотида, кодирующего белок FAF1, в клетки для получения трансформированных клеток; 2) культивирование трансформированных клеток; и 3) выделение экзосом из культивированных клеток.
Кроме того, настоящее изобретение относится к способу скрининга противоопухолевых терапевтических агентов, где способ включает стадии: 1) обработку тестируемым веществом опухолевых клеток, имеющих пониженную экспрессию FAF1; 2) измерение уровня белка FAF1 в экзосомах, выделенных из клеток на стадии 1); и 3) выбор тестируемого вещества, при котором уровень белка FAF1 на стадии 2) повышается по сравнению с контрольной группой, которую не обрабатывали тестируемым веществом.
Преимущественные эффекты изобретения
Настоящее изобретение относится к экзосомам, нагруженным белком Fas-ассоциированного фактора 1 (FAF1), и их применению для лечения рака. Авторы настоящего изобретения выделили экзосомы, нагруженные белком FAF1, из клеток HEK 293, в которых, как известно, FAF1, обладающий функцией подавления опухолей, сверхэкспрессируется, и ввели экзосомы, нагруженные белком FAF1, в опухолевую модель, для получения которой клетки рака поджелудочной железы (MIA PaCa-2), клетки рака легкого (A549), клетки колоректального рака (HCT 116), клетки рака печени (Hep3B), клетки рака молочной железы (MDA-MB-231), клетки рака почки (Caki-1) и клетки рака шейки матки (HeLa клетки) трансплантировали голым мышам посредством интратуморальной инъекции и подтвердили, что наблюдались достоверные эффекты ингибирования роста опухолей по сравнению с контрольной группой, не обработанной FAF1, и, таким образом, экзосомы, нагруженные FAF1, по настоящему изобретению можно использовать в качестве терапевтического средства при различных видах рака.
Краткое описание фигур
На фиг. 1 приведены результаты вестерн-блоттинга, показывающие количество FAF1, нагруженного в экзосомы, выделенные с использованием ультрацентрифуги из клеток HEK 293, в которых FAF1 сверхэкспрессируется.
На фиг. 2 приведены результаты вестерн-блоттинга, показывающие количество FAF1, нагруженного в экзосомы, выделенные с использованием ультрацентрифуги из клеток HeLa, в которых FAF1 сверхэкспрессируется.
На фиг. 3 приведены результаты вестерн-блоттинга, показывающие количество FAF1, нагруженного в экзосомы, выделенные с использованием набора Exo-quick-TC™ из клеток HEK 293, в которых FAF1 сверхэкспрессируется.
На фиг. 4 приведены результаты вестерн-блоттинга, показывающие количество FAF1, нагруженного в экзосомы, выделенные с использованием набора Exo-quick-TC™ из клеток HeLa, в которых FAF1 сверхэкспрессируется.
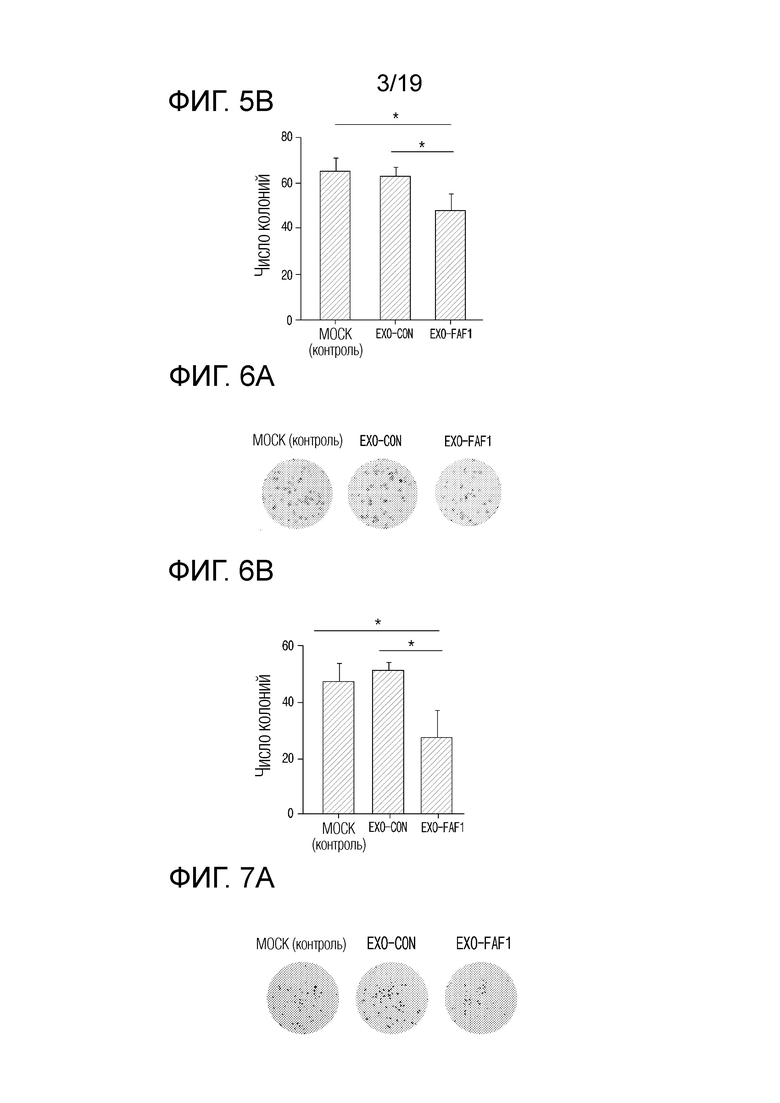
На фиг. 5а приведены фотографии, показывающие эффект ингибирования образования колоний при обработке клеток MIA PaCa-2 экзосомами, выделенными из клеток HEK 293, в которых FAF1 сверхэкспрессируется.
На фиг. 5b представлен график, показывающий число колоний при обработке клеток MIA PaCa-2 экзосомами, выделенными из клеток HEK 293, в которых FAF1 сверхэкспрессируется.
На фиг. 6а приведены фотографии, показывающие эффект ингибирования образования колоний при обработке клеток А549 экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 6b представлен график, показывающий число колоний при обработке клеток A549 экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
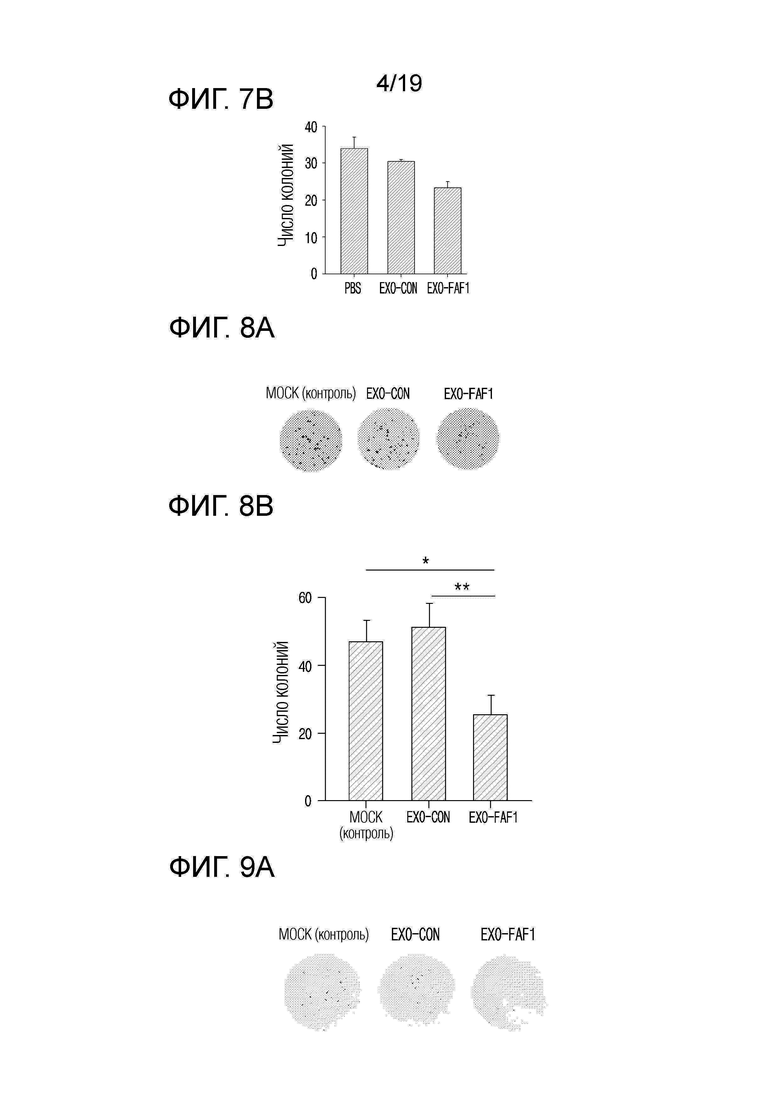
На фиг. 7а приведены фотографии, показывающие эффект ингибирования образования колоний при обработке клеток HCT 116 экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 7b представлен график, показывающий число колоний при обработке клеток HCT 116 экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 8а приведены фотографии, показывающие эффект ингибирования образования колоний при обработке клеток Hep3B экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 8b представлен график, показывающий число колоний при обработке клеток Hep3B экзосомами, выделенными из клеток HEK 293, в которых FAF1 сверхэкспрессируется.
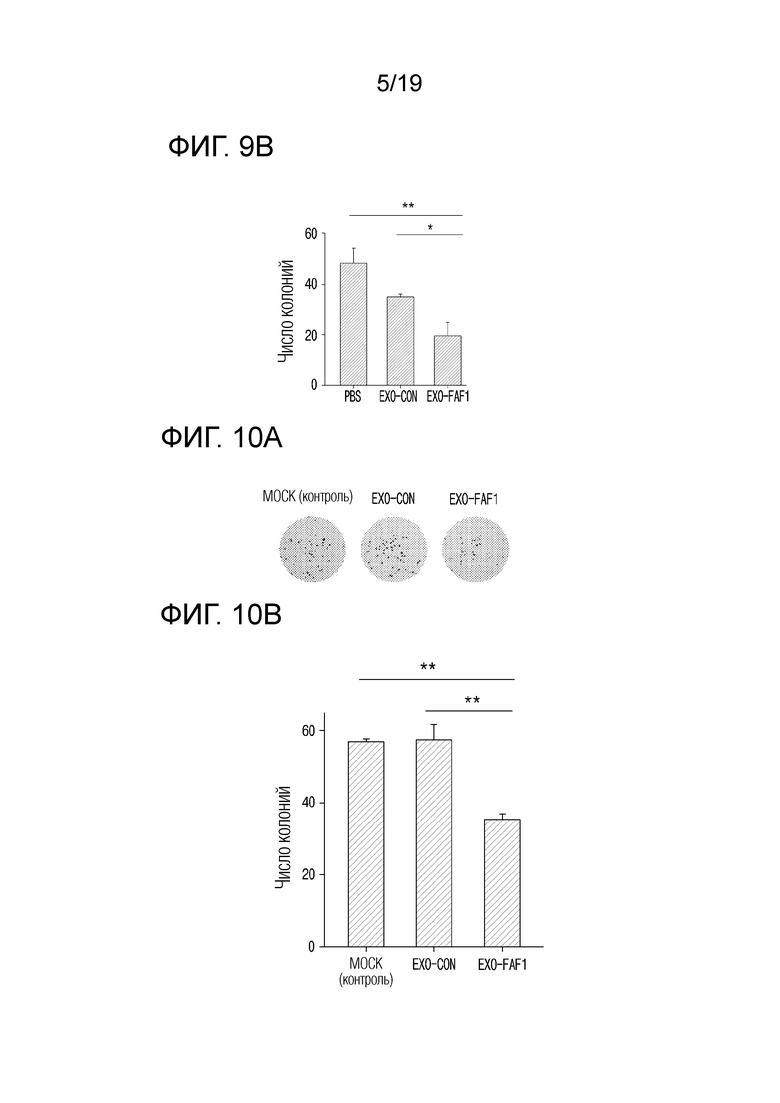
На фиг. 9а приведены фотографии, показывающие эффект ингибирования образования колоний при обработке клеток MDA-MB-231 экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 9b представлен график, показывающий число колоний при обработке клеток MDA-MB-231 экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 10а приведены фотографии, показывающие эффект ингибирования образования колоний при обработке клеток HeLa экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 10b представлен график, показывающий число колоний при обработке клеток HeLa экзосомами, выделенными из клеток HEK 293, в которых сверхэкспрессируется FAF1.
На фиг. 11a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MIA PaCa-2.
На фиг. 11b представлен график, показывающий объемы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MIA PaCa-2.
На фиг. 11c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MIA PaCa-2.
На фиг. 11d представлен график, показывающий массы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MIA PaCa-2.
На фиг. 11e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MIA PaCa-2.
На фиг. 12a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток A549.
На фиг. 12b представлен график, показывающий объемы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток A549.
На фиг. 12c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток A549.
На фиг. 12d представлен график, показывающий массы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток A549.
На фиг. 12e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток A549.
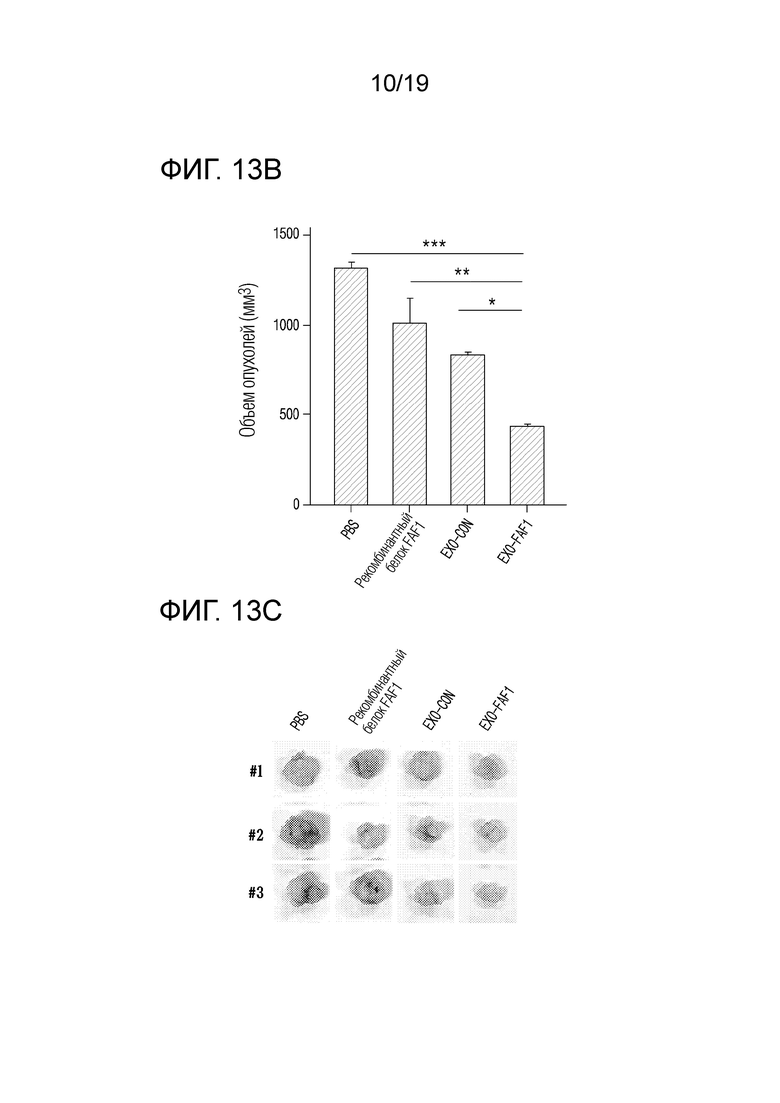
На фиг. 13a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HCT 116.
На фиг. 13b представлен график, показывающий объемы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HCT 116.
На фиг. 13c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HCT 116.
На фиг. 13d представлен график, показывающий массы опухолей для каждой группы MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HCT 116.
На фиг. 13e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HCT 116.
На фиг. 14a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Hep3B.
На фиг. 14b представлен график, показывающий объемы опухолей для каждой группы MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Hep3B.
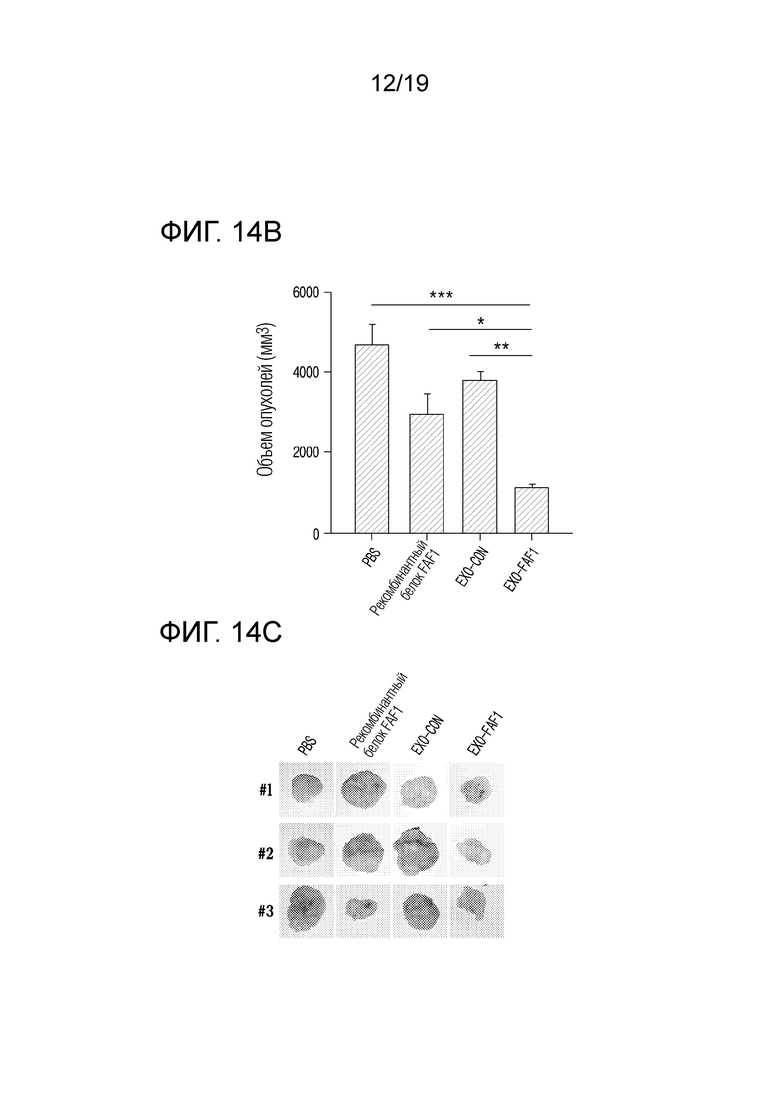
На фиг. 14c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Hep3B.
На фиг. 14d представлен график, показывающий массы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Hep3B.
На фиг. 14e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Hep3B.
На фиг. 15a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MDA-MB-231.
На фиг. 15b представлен график, показывающий объемы опухолей для каждой группы MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MDA-MB-231.
На фиг. 15c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MDA-MB-231.
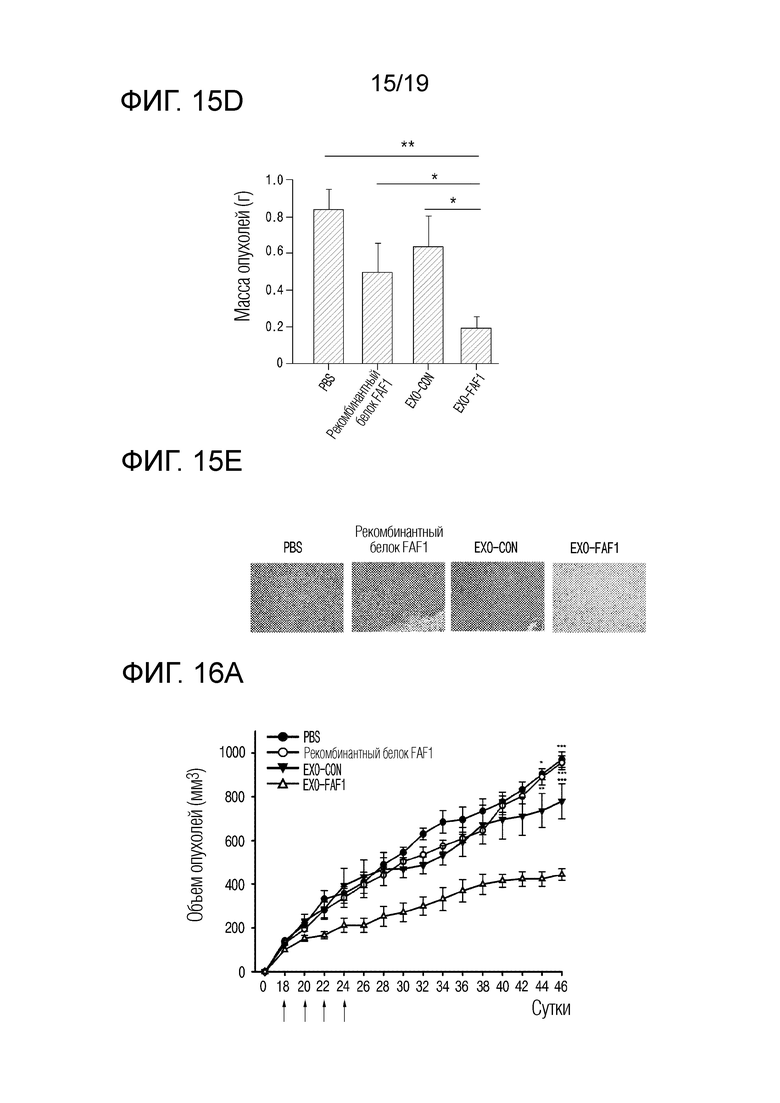
На фиг. 15d представлен график, показывающий массы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MDA-MB-231.
На фиг. 15e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для каждой группы MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток MDA-MB-231.
На фиг. 16a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Caki-1.
На фиг. 16b представлен график, показывающий объемы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Caki-1.
На фиг. 16c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Caki-1.
На фиг. 16d представлен график, показывающий массы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Caki-1.
На фиг. 16e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток Caki-1.
На фиг. 17a представлен график, показывающий изменения объема опухолей во времени для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HeLa.
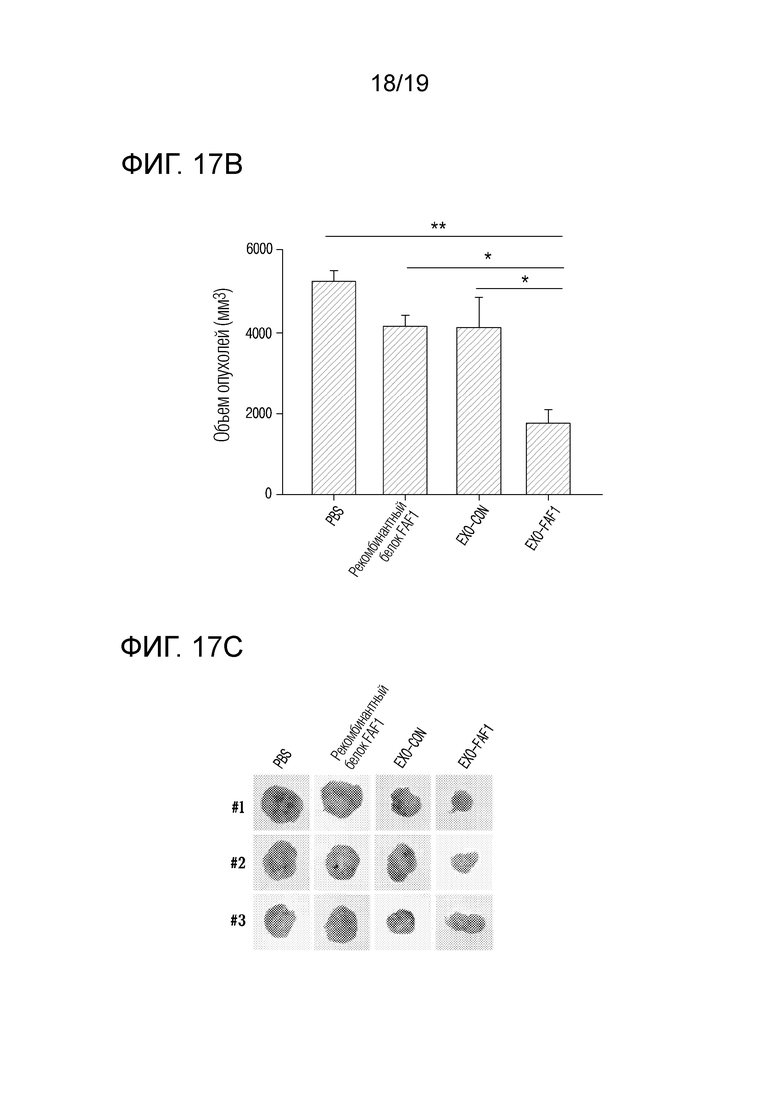
На фиг. 17b представлен график, показывающий объемы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1 непосредственно перед энуклеацией опухолей, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HeLa.
На фиг. 17c приведены фотографии опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HeLa.
На фиг. 17d представлен график, показывающий массы опухолей для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HeLa.
На фиг. 17e приведены фотографии, показывающие изменения в морфологии клеток опухолевой ткани для групп MOCK (контроля), рекомбинантного белка FAF1, EXO-CON и EXO-FAF1, когда экзосомы, выделенные из клеток HEK 293, в которых FAF1 сверхэкспрессируется, вводили голым мышам после ксенотрансплантации клеток HeLa.
Преимущественный способ осуществления изобретения
Далее настоящее изобретение будет описано подробно.
Настоящее изобретение относится к фармацевтической композиции для лечения рака, содержащей экзосомы, нагруженные белком FAF1, в качестве активного ингредиента.
Белок FAF1 по настоящему изобретению включает аминокислотную последовательность, показанную в SEQ ID NO: 1.
FAF1 по настоящему изобретению представляет собой сигнальный комплекс, индуцирующий клеточную гибель Fas (Fas-DISC), и индуцирует апоптоз. FAF1 известен как белок, который активирует несколько путей и индуцирует апоптоз.
FAF1 опосредует некроз клеток через JNK-зависимую митохондриальную дисфункцию, участвует в пролиферации клеток посредством подавления фазы G2/M в клеточном цикле путем негативной регуляции Aurora-A, участвует в убиквитин-протеасомном пути посредством связывания с убиквитинированным белком и валозин-содержащим белком (VCP) для регуляции деградации белка, и «лишний» FAF1 подвергается убиквитинированию с участием Parkin и разрушается на протеасомном пути.
Кроме того, FAF1 участвует в различных биохимических процессах, включая апоптоз, воспаление, пролиферацию клеток и гомеостаз белков. FAF1 является супрессором опухолей, который играет роль в подавлении роста опухолей за счет ингибирования NF-κB, а также подавляет метастазы опухолей посредством сигналов TGF-β.
Экзосомы по настоящему изобретению представляют собой небольшие везикулы, состоящие из двух слоев фосфолипидной мембраны, секретируемые клетками, и, как известно, они осуществляют передачу сигналов между клетками, образуют специфичные для заболевания нуклеиновые кислоты и белки и высвобождают их в жидкости организма, а также играют важную роль в межклеточной коммуникации посредством межклеточной трансдукции специфических репертуаров нуклеиновых кислот, белков и липидов, которые важны для гомеостаза. Например, экзосомы принимают участие в основных физиологических процессах, таких как нейротрансмиссия, презентация антигена, иммунный ответ, развитие органов, и репродуктивная функция, а также вовлечены в развитие некоторых патологических расстройств, включая прогрессирование рака, сердечно-сосудистые заболевания, воспаления и передачу прионов.
Экзосомы секретируются во внеклеточную среду после слияния поздней эндосомы, известной как мультивезикулярное тельце (MVB), содержащей внутрипросветные везикулы (ILV), с клеточной мембраной. Мультивезикулярное тельце может сливаться с клеточной мембраной, высвобождая ILV, тогда как мультивезикулярное тельце может сливаться с лизосомой, разрушая содержимое. Вещества, находящиеся в экзосомах, не аналогичны по составу цитоплазматическим веществам, и экзосомы могут необязательно содержать РНК и белки.
Хотя сообщалось, что продукция экзосом зависит или не зависит от эндосомального сортировочного комплекса, необходимого для транспорта (ESCRT), трудно определить точный механизм. Клетки секретируют белки, среди которых находятся сигнальные пептиды, через эндоплазматическую сеть-аппарат Гольджи. Везикулы, содержащие белки, среди которых присутствуют сигнальные пептиды, движутся к клеточной мембране, сливаются с клеточной мембраной и высвобождают белки во внеклеточное пространство. Однако белки без сигнальных пептидов могут секретироваться альтернативным неклассическим секреторным путем. Когда секретируются белки без сигнальных пептидов, то белки секретируются либо в отсутствии, либо в присутствии везикул, и хотя точный механизм невезикулярного секреторного пути неизвестен, некоторые белки секретируются посредством мембранных пор и АТФ-связывающих кассетных транспортеров. Везикулярная секреция осуществляется посредством внеклеточных везикул, в том числе экзосом, и осуществляется посредством везикул различного размера.
Экзосомы могут иметь средний диаметр от 50 до 300 нм, но настоящее изобретение не ограничивается указанными размерами.
Экзосомы представляют собой внеклеточные везикулы наноразмера, которые выделяются большинством типов клеток и содержат небольшие РНК, липиды и белки и имеют ряд преимуществ перед доступными в настоящее время синтетическими средствами доставки лекарственных средств. Такие преимущества включают способность преодолевать естественные барьеры, присущие свойства нацеливания на клетки, улучшенную проницаемость, поддерживающий эффект и биосовместимость. С учетом основной функции, заключающейся в доставке биологической информации, применение экзосом в качестве терапевтического агента привлекает внимание.
Рак представляет собой рак, при котором экспрессия FAF1 снижена.
Рак включает, помимо прочего, рак шейки матки, рак поджелудочной железы, рак печени, рак легкого (включая мелкоклеточный рак легкого и немелкоклеточный рак), рак молочной железы, рак почки, колоректальный рак, плоскоклеточный рак головы и шеи, рак мочевого пузыря, рак предстательной железы, рак желудка, рак эндометрия, рак головного мозга, рак яичника, рак яичка, рак головы, рак шеи, рак кожи (включая меланому и базальноклеточную карциному), рак мезотелиальной выстилки, рак пищевода, рак надпочечников, рак щитовидной железы, рак кости, глиобластому, мезотелиому, саркому, хориокарциному, базальноклеточный рак кожи, семиному яичка, лейкоз и злокачественную лимфому. В предпочтительном аспекте рак представляет собой рак поджелудочной железы, немелкоклеточный рак легкого, колоректальный рак, рак печени, тройной негативный рак молочной железы, рак почки или рак шейки матки.
Фармацевтическая композиция по настоящему изобретению может включать носитель, разбавитель, эксципиент или их смесь, которые обычно используются для фармацевтических составов. Можно использовать любой фармацевтически приемлемый носитель, если он подходит для доставки композиции в живой организм. В частности, носителем могут быть соединения, описанные в Merck Index, 13th ed. Merck & Co. Inc., физиологический раствор, стерилизованная вода, раствор Рингера, раствор декстрозы, раствор мальтодекстрина, глицерин, этанол или их смесь. Кроме того, при необходимости могут быть добавлены типичные добавки, такие как антиоксиданты, буферы и бактериостаты.
При формуляции фармацевтической композиции могут быть добавлены разбавители или эксципиенты, такие как наполнители, разбавители, связующие вещества, смачивающие агенты, разрыхлители и поверхностно-активные вещества, которые обычно используются.
Фармацевтическая композиция по настоящему изобретению может быть формулирована в виде препаратов для перорального или парентерального введения. Препараты для перорального введения могут включать твердые препараты и жидкие препараты. Твердые препараты могут представлять собой таблетки, пилюли, порошки, гранулы, капсулы или пастилки, и твердые препараты можно приготовить добавлением к композиции, по меньшей мере, одного эксципиента. Эксципиентом может быть крахмал, карбонат кальция, сахароза, лактоза, желатин или их смесь. Кроме того, твердые препараты могут включать смазывающее вещество, например стеарат магния и тальк. При этом жидкие препараты могут представлять собой суспензии, растворы, эмульсии или сиропы. В этом случае жидкие препараты могут содержать эксципиенты, такие как смачивающие агенты, подсластители, ароматизаторы и консерванты.
Препараты для парентерального введения могут включать инъекционные растворы, суппозитории, порошки для респираторных ингаляций, аэрозольные спреи, порошки, кремы и тому подобное. Инъекционные препараты могут содержать стерилизованные водные растворы, неводные растворители, суспендирующие агенты, эмульгаторы и т.п. В этом случае в качестве неводных растворителей или суспендирующих агентов можно использовать пропиленгликоль, полиэтиленгликоль и растительное масло, такое как оливковое масло, или сложный эфир для инъекций, такой как этилолеат, или тому подобное.
Фармацевтическую композицию по настоящему изобретению можно вводить перорально или парентерально любым желаемым способом. Парентеральное введение может включать, помимо прочего, метод интрабронхиальной ингаляции, внутрибрюшинную инъекцию, ректальное введение, подкожную, внутривенную, внутримышечную, интраторакальную или интратуморальную инъекцию. В предпочтительном аспекте фармацевтическую композицию по настоящему изобретению можно вводить методом интратуморальной инъекции.
Фармацевтическую композицию можно вводить в фармацевтически эффективном количестве. Оно может варьироваться в зависимости от типа заболевания, тяжести заболевания, активности лекарственного препарата, чувствительности пациента к лекарственному препарату, времени введения, пути введения, продолжительности лечения, одновременно используемых лекарственных препаратов и т.п. Однако для желаемого эффекта количество активного ингредиента, содержащегося в фармацевтической композиции по настоящему изобретению, может составлять 0,0001-1000 мг/кг, в частности, 0,001-500 мг/кг. Введение может осуществляться от одного до нескольких раз в сутки.
Фармацевтическую композицию по настоящему изобретению можно вводить самостоятельно или в комбинации с другими терапевтическими средствами. При комбинированном введении введение можно осуществлять последовательно или одновременно.
Кроме того, настоящее изобретение относится к способу получения экзосом, нагруженных белком FAF1, где способ включает стадии:
1) введение в клетки полинуклеотида, кодирующего белок FAF1, для получения трансформированных клеток;
2) культивирование трансформированных клеток; и
3) выделение экзосом из клеток.
Клетки на стадии 1) представляют собой любую одну или более клеток, выбранных из группы, состоящей из В-лимфоцитов, Т-лимфоцитов, дендритных клеток, мегакариоцитов, макрофагов, стволовых клеток, опухолевых клеток и клеток HEK293, не ограничиваясь этим. В предпочтительном аспекте клетки представляют собой клетки HEK 293 или клетки HeLa.
Трансдукцию можно осуществлять обработкой от 5×106 до 7×106 клеток 5-10 мкг плазмиды 3X Flag-FAF1 и культивированием клеток, предпочтительно обработкой 6,5×106 клеток 8 мкг плазмиды 3X Flag-FAF1 и культивированием клеток.
Экзосомы можно выделить методом, обычно используемым в данной области техники для выделения экзосом. Для выделения экзосом можно использовать эксклюзионную хроматографию, ионообменную хроматографию, центрифугирование в градиенте плотности, дифференциальное центрифугирование, ультрафильтрацию, тангенциальную проточную фильтрацию, осаждение экзосом, набор для полного выделения экзосом, иммуноабсорбентный захват, аффинный метод, например, аффинный захват, аффинную очистку, иммуноанализ, микрофлюидное разделение или их комбинацию.
Согласно конкретному варианту осуществления настоящего изобретения культуральную среду собирают и центрифугируют при 300 g в течение 10 мин, при 2000 g в течение 10 мин и при 10000 g в течение 30 мин для отделения супернатанта, супернатант фильтруют, используя 0,2 мкм фильтр и центрифугируют с использованием ультрацентрифуги при 150000 g в течение 70 мин. Отцентрифугированный супернатант удаляют, осадок промывают добавлением PBS, и затем снова центрифугируют при 150000 g в течение 70 мин для удаления супернатанта, тем самым выделив экзосомы, оставшиеся в нижнем слое.
Выделенные экзосомы можно хранить при температуре -20°С и -80°С.
В настоящем документе термин «трансформация» представляет собой молекулярно-биологический метод, при котором фрагмент цепи ДНК или плазмида и т. д., содержащие экзогенный ген, отличный от гена исходной клетки, проникает в клетку и связывается с ДНК, находящейся в исходной клетке, тем самым изменяя генетическую природу клетки.
Экзосомы, нагруженные FAF1, содержат белки FAF1 и служат переносчиками для транспорта белков FAF1 к опухолевым клеткам или тканям.
Белки FAF1 секретируются во внеклеточное пространство через экзосомы.
Белки FAF1, секретированные внеклеточно, могут индуцировать апоптоз в других клетках.
Апоптоз представляет собой вид запрограммированной гибели клеток, которая может иметь место в многоклеточных организмах. Апоптоз приводит к гибели клеток в результате изменений в морфологии клеток и биохимических изменений внутри клетки. Данный процесс заканчивается набуханием и расщеплением клеток, изменением клеточных мембран, конденсацией хроматина и расщеплением хромосом, а также поглощением клетки фагоцитом.
В отличие от некроза, который представляет собой гибель клеток, вызванную острым клеточным повреждением, апоптоз не повреждает организмы, а в ряде случаев дает преимущества в их жизненном цикле. Формирование пальцев рук и ног в процессе дифференцировки эмбриона человека является характерным примером апоптоза. Кроме того, апоптоз является важным механизмом замещения клеток, ремоделирования тканей и удаления поврежденных клеток.
Апоптоз происходит в двух случаях: в первом, для устранения ненужных частей в процессе развития и дифференцировки, и в другом, для защиты других клеток, когда клетка имеет серьезные повреждения и может переродиться опухолевую клетку. Однако клетки, у которых возникают проблемы в процессе апоптоза, становятся опухолевыми клетками, и опухолевые клетки не погибают, и постепенно размножаются и растут посредством повторного деления.
В рамках настоящего изобретения, термин «культивирование» относится к способу размножения клеток или микроорганизмов в искусственно контролируемых условиях окружающей среды. В настоящем изобретении способ культивирования трансформанта можно осуществлять с использованием способа, широко известного в данной области техники.
Среда относится к известной среде, используемой для культивирования клеток животных, и может быть выбрана из группы коммерчески доступных бессыворочных сред, сред, не содержащих белков, и сред с определенным химическим составом.
Кроме того, настоящее изобретение относится к способу скрининга противоопухолевых терапевтических агентов, включающему стадии:
1) обработку тестируемым веществом опухолевых клеток, имеющих пониженную экспрессию FAF1;
2) измерение уровня белка FAF1 в экзосомах, выделенных из клеток на стадии 1); и
3) выбор тестируемого вещества, при котором уровень белка FAF1 на стадии 2) повышается по сравнению с контрольной группой, которую не обрабатывали тестируемым веществом.
В способе клетки на стадии 1) не ограничиваются, но в предпочтительном аспекте представляют собой клетки MIA PaCa-2, A549, HCT 116, Hep3B, MDA-MB-231, Caki-1 или HeLa.
В данном способе тестируемое вещество на стадии 1) представляет собой вещество, которое повышает экспрессию FAF1, и может представлять любое вещество, выбранное из группы, состоящей из пептида, белка, непептидного соединения, синтетического соединения, продукта ферментации, клеточного экстракта, растительного экстракта и экстракт животной ткани, не ограничиваясь этим.
В качестве маркера для выбора вещества-кандидата, используемого в настоящем изобретении, уровень белка FAF1 может быть измерен с использованием метода, известного в данной области. Например, антитела, которые специфически детектируют белок и являются коммерчески доступными, можно использовать для детектирования белка с использованием вестерн-блоттинга, соиммунопреципитации, иммунофлуоресценции, иммуноферментного анализа и т.п. Используя такие антитела, можно специфически подтвердить уровень экспрессии конкретного белка, но настоящее изобретение не ограничивается этим.
Повышение уровня экспрессии специфического белка можно использовать для основы для выбора вещества-кандидата в качестве противоопухолевого терапевтического средства по сравнению с контрольной группой, которую не обрабатывают тестируемым веществом.
Первое, было подтверждено с учетом повышения количества FAF1 в экзосомах из клеток HEK 293 и клеток HeLa, в которых FAF1 сверхэкспрессируется, что сверхэкспрессия FAF1 в клетках HEK 293 и клетках HeLa увеличивает количество FAF1 в клетках HEK 293 и клетках HeLa, находящегося в экзосомах и делает возможной секрецию FAF1 через экзосомы (см. фиг. 1-4). FAF1, фактор, способствующий апоптозу, представляет собой белок, который может ингибировать образование опухолей. Следовательно, для подтверждения того, насколько экзосомы, выделенные из клеток HEK 293, сверхэкспрессирующих FAF1, обладают эффектом подавления образования опухолей, были проведены тесты на подавление образования опухолей на клеточных линиях MIA PaCa-2, A549, HCT 116, Hep3B, MDA-MB-231 или HeLa, и было подтверждено, что число колоний, образовавшихся при обработке экзосомами, выделенными из клеток HEK 293, сверхэкспрессирующих FAF1, было ниже, чем экзосомами, выделенными из клеток HEK 293, не сверхэкспрессирующих FAF1 (см. фиг. 5a, 5b, 6a, 6b, 7a, 7b, 8а, 8b, 9а, 9b, 10а и 10b). Кроме того, было подтверждено, что объем и масса опухолей в случае экзосом, нагруженных FAF1, уменьшались при интратуморальной инъекции экзосом, выделенных из клеток HEK 293, сверхэкспрессирующих FAF1, голым мышам после ксенотрансплантации клеток MIA PaCa-2, A549, HCT 116, Hep3B, MDA-MB-231, Caki-1 или HeLa, и было подтверждено, что некроз клеток развивается в результате обработки FAF1 (см. фиг. 11a-11e, фиг. 12a-12e, фиг. 13a-13е, 14а-14е, 15а-15е, 16а-16е и 17а-17е). Вышеприведенные результаты демонстрируют, что нагруженный FAF1 может секретироваться во внеклеточное пространство через экзосомы, экзосомы, нагруженные FAF1, должным образом доставляются к опухолевым клеткам-мишеням, и FAF1 может эффективно подавлять рост опухолей, означая, что экзосомы, нагруженные FAF1, можно использовать в качестве средства для лечения рака.
Далее настоящее изобретение будет описано подробно со ссылкой на следующие примеры.
Однако следующие примеры предназначены только для иллюстрации настоящего изобретения, и настоящее изобретение не ограничивается ими.
Варианты осуществления изобретения
Пример 1. Выделение экзосом из клеток HEK 293, сверхэкспрессирующих FAF1
1-1. Культивирование клеток
Клетки HEK 293, представляющие собой клеточные линии, полученные из эмбриональной почки человека, поддерживали при субкультивировании один раз в 2 суток в условиях с 5% CO2 и при 37°C в среде Игла, модифицированной по способу Дульбекко (DMEM, WelGENE, Корея), содержащей антибиотики, 10% фетальной бычьей сыворотки (FBS, Atlas Biologicals, США) и антибиотик-антимикотик (пенициллин/стрептомицин, GIBCOBRL, США).
1-2. Внутриклеточная трансдукция
Для сверхэкспрессии FAF1 в клетках плазмиду 3x Flag tag-FAF1 (8 мкг) трансдуцировали в 6,5×106 клеток.
В частности, клетки высевали на культуральную чашку диаметром 150 мм, и затем клетки культивировали в среде DMEM, содержащей 10% FBS, в течение 24 ч в условиях с 5% CO2 и при 37°C. 3x Flag tag-FAF1 готовили известным ранее методом (Yu et al., 2016). В целом, приготовили 180 мкл смеси в соотношении ДНК:BioT (Bioland Scientific, США)=1:1,5 с использованием DMEM без FBS и антибиотиков, и смесь подвергали взаимодействию при комнатной температуре в течение 5 мин. Смесь добавляли к клеткам, культивированным в течение 24 ч, и затем клетки реагировали в течение 24 ч в условиях с 5% CO2 и при 37°C.
1-3. Выделение экзосом
1-3-1. Выделение экзосом с использованием Exo-quick-TC™
Экзосомы выделяли из клеток, трансдуцированных плазмидой 3x Flag tag-FAF1, с использованием набора Exo-quick-TC™ (System Biosciences, США).
В частности, через 24 ч после трансдукции клетки обрабатывали DMEM (System Biosciences, США), обработанной 10% обедненной экзосомами FBS, и культивировали в течение 48 ч. Среду для культивированных клеток собирали и центрифугировали при 3000 об/мин в течение 15 мин для сбора супернатанта, из которого удаляли клеточный дебрис. Собранную среду (10 мл) обрабатывали 2 мл буфера Exo-quick, встряхивали и хорошо перемешивали так, чтобы соотношение среда:буфер Exo-quick составляло 5:1. После культивирования в течение ночи при 4°С процесс центрифугирования среды при 1500 g в течение 30 мин повторяли дважды для получения экзосом.
1-3-2. Выделение экзосом с использованием ультрацентрифуги
Экзосомы выделяли из клеток, трансдуцированных плазмидой 3x Flag tag-FAF1, с использованием ультрацентрифуги (optima XE-100, BECKMAN Coulter, США).
В частности, через 24 ч после трансдукции клетки обрабатывали DMEM без FBS и антибиотиков и снова культивировали в течение 24 ч. Среду для культивированных клеток собирали и центрифугировали при 300 g в течение 10 мин, при 2000 g в течение 10 мин и при 10000 g в течение 30 мин для отделения супернатанта, и супернатант фильтровали с использованием шприцевого фильтра 0,2 мкм (BioFACT, Корея).
Для выделения экзосом для использования в экспериментах in vitro и in vivo супернатант центрифугировали при 150000 g в течение 70 мин с ротором 45Ti на ультрацентрифуге BECKMAN Coulter optima XE-100, супернатант отбрасывали, и осадок промывали PBS. Промытый PBS осадок снова центрифугировали при 150000 g в течение 70 мин, супернатант затем отбрасывали, и остаток собирали в пробирку EP с использованием PBS.
Выделенные экзосомы хранили при -20°С и -80°С, и перед использованием размораживали при температуре 4°С.
Пример 2. Выделение экзосом из клеток HeLa, сверхэкспрессирующих FAF1
2-1. Культивирование клеток
Клетки HeLa, полученные от пациенток с раком шейки матки, поддерживали при субкультивировании один раз в 2 суток в условиях с 5% CO2 и при 37°C в среде DMEM (WelGENE, Корея), содержащей антибиотики, 10% FBS (Atlas Biologicals, США) и антибиотик-антимикотик (пенициллин/стрептомицин, GIBCOBRL, США).
2-2. Внутриклеточная трансдукция
Для сверхэкспрессии FAF1 в клетках HeLa плазмиду 3x Flag tag-FAF1 (8 мкг) трансдуцировали в клетки в количестве 6,5×106.
Конкретный экспериментальный метод был таким же, как описан в примере 1-2.
2-3. Выделение экзосом
Экзосомы выделяли из клеток HeLa, трансдуцированных плазмидой 3x Flag tag-FAF1, с использованием набора Exo-quick-TC™ или ультрацентрифуги.
Конкретный экспериментальный метод был таким же, как описан в примере 1-3.
Пример 3. Культивирование опухолевых клеточных линий
MIA PaCa-2 (клетки рака поджелудочной железы), Hep3B (клетки рака печени), MDA-MB-231 (клетки тройного негативного рака молочной железы), Caki-1 (клетки рака почки) и HeLa (клетки рака шейки матки) культивировали в инкубаторе в следующих условиях: DMEM, 10% FBS, 1% антибиотика-антимикотика (пенициллин/стрептомицин, GIBCO BRL, США), 5% CO2 и 37°C, субкультивировали один раз в 3 суток. A549 (клетки немелкоклеточного рака легкого) и HCT 116 (клетки колоректального рака) культивировали в инкубаторе в среде RPMI 1640 в условиях: 10% FBS, 1% антибиотика-антимикотика, 5% CO2, 37°C и субкультивировали один раз в 3 суток.
Экспериментальный пример 1. Подтверждение повышенного уровня FAF1 в экзосомах при сверхэкспрессии FAF1 в клетках HEK 293 и клетках HeLa
Для выяснения того, насколько больше FAF1 проникает в экзосомы, когда FAF1 сверхэкспрессируется в клетках HEK 293 и клетках HeLa, уровень экспрессии FAF1 в экзосомах, выделенных с использованием ультрацентрифуги и набора Exo-quick-TC™, из клеток HEK 293 и клеток HeLa, трансдуцированных плазмидой 3x Flag tag-FAF10 (8 мкг) из примера 1, определяли вестерн-блоттингом.
В частности, для проведения вестерн-блоттинга к осадку после центрифугирования добавляли 50 мкл буфера для лизиса клеток млекопитающих, от которого отделяли супернатант и растворяли на льду в течение 30 мин. Для приготовления 8-10% акриламидного геля использовали набор для постановки вестерн-блоттинга (Amersham Biosciences, Великобритания), растворенный осадок смешивали с 5 мкл буфера для образцов SDS, кипятили в течение 3 мин, и затем наносили на гель в заранее определенном количестве и применяли напряжение 80-100 В для разделения белков, находящихся в осадке, по размеру. Акриламидный гель, содержащий разделенные белки, располагали таким образом, чтобы он перекрывал нитроцеллюлозную мембрану, и в течение 2 ч прикладывали ток силой 200 мА для переноса белков на нитроцеллюлозную мембрану. Нитроцеллюлозную мембрану, на которую были перенесены белки, блокировали обезжиренным сухим молоком, на нее наносили антитела к FAF1, и затем присоединяли вторичные антитела, количество белка FAF1 определяли с использованием набора для вестерн-блоттинга (AbFrontier, Корея) и визуализирующей системы ChemiDoc-It (UVP, США).
Когда экзосомы выделяли с использованием ультрацентрифуги, то экспрессия FAF1 была несколько выше в экзосомах, выделенных из клеток HEK 293, в которых FAF1 сверхэкспрессируется, по сравнению с экзосомами, выделенными из клеток HEK 293, в которых FAF1 не сверхэкспрессируется (фиг. 1). В случае клеток HeLa редко наблюдалась экспрессия FAF1 в экзосомах, выделенных из клеток, в которых FAF1 не сверхэкспрессируется, и клеток, в которых FAF1 сверхэкспрессируется (фиг. 2).
Когда экзосомы выделяли с помощью набора Exo-quick-TC™, то уровень экспрессии FAF1 в экзосомах, выделенных из клеток, сверхэкспрессирующих FAF1, был достоверно выше в клетках HEK 293 (фиг. 3), и FAF1 не детектировали в экзосомах, выделенных из клеток, сверхэкспрессирующих FAF1, в клетках HeLa, но с помощью Flag, который является белком-меткой, было подтверждено, что количество FAF1 было повышенным (фиг. 4).
Способ выделения экзосом с использованием набора Exo-quick-TC™ является более простым, чем способ с использованием ультрацентрифуги, и имеет преимущество, заключающееся в том, что сокращается время, но имеет недостаток: при нем имеет место загрязнение во время процесса выделения. Кроме того, поскольку способ с использованием ультрацентрифуги позволяет выделить экзосомы, имеющие более высокую чистоту, то авторы настоящего изобретения выделили экзосомы с использованием ультрацентрифуги в следующих экспериментальных примерах, чтобы подтвердить эффект чистых экзосом.
Экспериментальный пример 2. Подтверждение противоопухолевого эффекта in vitro экзосом, выделенных из клеток HEK 293, сверхэкспрессирующих FAF1
С использованием экзосом, выделенных из клеток HEK 293, сверхэкспрессирующих FAF1, оценивали способность ингибировать образование колоний в клеточных линиях рака поджелудочной железы, немелкоклеточного рака легких, рака печени и рака шейки матки.
В частности, клетки HEK 293, трансдуцированные 3x Flag tag-FAF1 (0 или 8 мкг), культивировали в течение 24 ч, культивировали в среде, не содержащей FBS и антибиотиков, в течение 24 ч, для стимуляции секреции экзосом, и затем среду собирали и экзосомы выделяли с использованием ультрацентрифуги (optima XE-100, BECKMAN Coulter). Количественное определение выделенных экзосом проводили на спектрофотометре (MECASYS, Корея). Для выявления in vitro эффекта ингибирования опухолевых колоний под действием экзосом, нагруженных FAF1, в клеточных линиях MIA PaCa-2, A549, HCT 116, Hep3B, MDA-MB-231 или HeLa, клетки MIA PaCa-2, Hep3B или HeLa высевали в 6-луночный планшет по 100 клеток, и клетки A549, HCT116 и MDA-MB-231 высевали по 200 клеток. Через 24 ч клетки обрабатывали экзосомами без загрузки FAF1 (экзосомный контроль: EXO-CON) и экзосомами, нагруженными FAF1 (EXO-FAF1). В случае MIA PaCa-2 экзосомы анализировали на спектрофотометре (MECASYS, Корея), клетки обрабатывали 6 мкг экзосом, и затем культивировали в течение 12 суток; в случае А549 экзосомы анализировали с использованием NTA, клетки обрабатывали 3×108 экзосом, и затем культивировали в течение 10 суток; в случае HCT 116 экзосомы анализировали с использованием NTA, клетки обрабатывали 3,16×109 экзосом, и затем культивировали в течение 10 суток; в случае Hep3B экзосомы определяли на спектрофотометре (MECASYS, Корея), клетки обрабатывали 12 мкг экзосом, и затем культивировали в течение 7 суток; в случае MDA-MB_231 экзосомы анализировали с использованием NTA, клетки обрабатывали 8,4×109 экзосом, и затем культивировали в течение 12 суток; и в случае HeLa экзосомы определяли с использованием NTA, клетки обрабатывали 1,65×1010 экзосом, и затем культивировали в течение 10 суток. Среду заменяли свежей средой один раз в 3 суток, и при замене среды клетки обрабатывали экзосомами в соответствующих концентрациях. После завершения экспериментов число окрашенных колоний определяли промывкой PBS, обработкой 1 мл метанола в течение 20 мин и обработкой 1 мл раствора кристаллического фиолетового (SIGMA-ALDRICH, США) в течение 5 мин.
2-1. Подтверждение возможности ингибировать образование колоний при раке поджелудочной железы
Для подтверждения возможности ингибировать образование колоний за счет противоопухолевых эффектов экзосом, нагруженных FAF1, при раке поджелудочной железы, клетки MIA PaCa-2, которые представляют собой клеточные линии рака поджелудочной железы, обрабатывали экзосомами, нагруженными FAF1, и затем проводили анализ.
В результате, как показано на фиг. 5а и 5b, было подтверждено, что наименьшее число колоний образовалось в группе, обработанной EXO-FAF1. Определив числовые значения, было подтверждено, что EXO-FAF1 приводил к снижению образования колоний на 24,2±7,27% по сравнению с EXO-CON и на 26,2±7,08% по сравнению с MOCK (контролем).
Вышеприведенные результаты позволяют предположить, что экзосомы, нагруженные FAF1, подавляют образование опухолей при раке поджелудочной железы.
2-2. Подтверждение возможности ингибировать образование колоний при немелкоклеточном раке легкого
Для подтверждения противоопухолевых эффектов экзосом, нагруженных FAF1, при немелкоклеточном раке легкого, клетки A549, которые представляют собой клеточные линии немелкоклеточного рака легких, обрабатывали экзосомами, нагруженными FAF1, и затем подтверждали возможность ингибировать образований колоний.
В результате, как показано на фиг. 6а и 6b, было подтверждено, что наименьшее число колоний образовалось в группе, обработанной EXO-FAF1. Было установлено, что EXO-FAF1 приводил к снижению числа колоний на 46,9±12,88% по сравнению с EXO-CON и на 42,7±13,91% по сравнению с MOCK (контролем).
Вышеприведенные результаты позволяют предположить, что экзосомы, нагруженные FAF1, подавляют образование опухолей при немелкоклеточном раке легких.
2-3. Подтверждение возможности ингибировать образование колоний при колоректальном раке
Для подтверждения противоопухолевых эффектов экзосом, нагруженных FAF1, при колоректальном раке, клетки HCT 116, которые представляют собой клеточные линии колоректального рака, обрабатывали экзосомами, нагруженными FAF1, и затем подтверждали наличие способности ингибирования образования колоний.
В результате, как показано на фиг. 7а и 7b, было подтверждено, что наименьшее количество колоний образовалось в группе, обработанной EXO-FAF1. Было установлено, что EXO-FAF1 приводил к снижению числа колоний на 23,0±9,07% по сравнению с EXO-CON и на 30,9±8,14% по сравнению с MOCK (контролем).
Вышеприведенные результаты позволяют предположить, что экзосомы, нагруженные FAF1, подавляют образование опухолей при колоректальном раке.
2-4. Подтверждение возможности ингибировать образование колоний при раке печени
Для подтверждения противоопухолевых эффектов экзосом, нагруженных FAF1, при раке печени, клетки Hep3B, которые представляют собой клеточные линии рака печени, были обработаны экзосомами, нагруженными FAF1, и затем подтверждали возможность ингибировать образований колоний.
В результате, как показано на фиг. 8a и 8b, было подтверждено, что наименьшее число колоний образовалось в группе, обработанной EXO-FAF1. Было установлено, что EXO-FAF1 приводил к снижению числа колоний на 50,6±10,21% по сравнению с EXO-CON и на 46,1±11,15% по сравнению с MOCK (контролем).
Вышеприведенные результаты позволяют предположить, что экзосомы, нагруженные FAF1, подавляют образование опухолей при раке печени.
2-5. Подтверждение возможности ингибировать образование колоний при тройном негативном раке молочной железы
Для подтверждения возможности ингибировать образование колоний за счет противоопухолевых эффектов экзосом, нагруженных FAF1, при тройном негативном раке молочной железы, клетки MDB-MB-231, которые представляют собой клеточные линии тройного негативного рака молочной железы, обрабатывали экзосомами, нагруженными FAF1, и затем подтверждали наличие возможности ингибировать образований колоний.
В результате, как показано на фиг. 9а и 9b было подтверждено, что наименьшее число колоний образовалось в группе, обработанной EXO-FAF1. Определив числовые значения, было подтверждено, что EXO-FAF1 приводил к снижению образования колоний на 43,8±10,40% по сравнению с EXO-CON и на 59,3±7,59% по сравнению с MOCK (контролем).
Вышеприведенные результаты позволяют предположить, что экзосомы, нагруженные FAF1, подавляют образование опухолей при тройном негативном раке молочной железы.
2-6. Подтверждение возможности ингибировать образование колоний при раке шейки матки
Для подтверждения возможности ингибировать образование колоний в результате противоопухолевого эффекта экзосом, загруженных FAF1, при раке шейки матки, клетки HeLa, которые представляют собой клеточные линии рака шейки матки, обрабатывали экзосомами, нагруженными FAF1, и затем подтверждали возможность ингибировать образований колоний.
В результате, как показано на фиг. 10а и 10b, было подтверждено, что наименьшее число колоний образовалось в группе, обработанной EXO-FAF1. Определив числовые значения, было подтверждено, что EXO-FAF1 приводил к снижению образования колоний на 38,4±6,45% по сравнению с EXO-CON и на 38,0±6,49% по сравнению с MOCK (контролем).
Вышеприведенные выше результаты позволяют предположить, что экзосомы, нагруженные FAF1, подавляют образование опухолей при раке шейки матки.
Экспериментальный пример 3. Подтверждение противоопухолевого эффекта in vivo экзосом, выделенных из клеток HEK 293, сверхэкспрессирующих FAF1, на животных моделях ксенотрансплантированных опухолей
Для демонстрации наличия in vivo эффекта подавления опухолей под действием экзосом, выделенных из клеток HEK293, сверхэкспрессирующих FAF1, 2-7×106 каждой из клеточных линий MIA PaCa-2, A549, HCT 116, Hep3B, MDA-MB-231, Caki-1 и HeLa смешивали со 100 мкл PBS и 100 мкл матригеля (Corning Life Science, США), и затем ксенотрансплантаты вводили в дорсальную область 8-недельных самцов мышей BALB/cSLC nu/nu (Central Lab. Animal Inc., Корея) через шприц 26G. Объем опухолей рассчитывали по формуле: (длинная ось) × (короткая ось) 2 × 0,5. Когда объем опухолей достигал 80-140 мм3, то PBS, рекомбинантный FAF1 (AngioLab, Корея), EXO-CON и EXO-FAF1 вводили мышам в общей сложности четыре раза, один раз в 2 суток интратуморальной инъекцией соответственно. Объем опухолей клеточной линии MIA PaCa-2 измеряли до 43 суток, объем опухолей клеточной линии A549 измеряли до 48 суток, объем опухолей клеточной линии HCT 116 измеряли до 30 суток, объем опухолей клеточной линии Hep3B измеряли до 35 суток, объем опухолей клеточной линии MDA-MB-231 измеряли до 40 суток, объем опухолей клеточной линии Caki-1 измеряли до 46 суток, и объем опухолей клеточной линии HeLa измеряли до 41 суток. В последний день, после того как мышей подвергли эвтаназии, опухоли извлекали и взвешивали, и внутреннюю часть опухоли анализировали после окрашивания H&E.
Извлеченную ткань для окрашивания H&E хранили в растворе формалина. Через 24 ч проводили дегидратацию в течение 20 мин в 70% этаноле, дегидратацию проводили дважды по 20 мин в 95% этаноле и, наконец, дегидратацию проводили дважды в 100% этаноле в течение 20 мин. Затем проводили обработку ксилолом и парафином соответственно в течение 3 ч с использованием тканевого процессора (Leica, Германия). Для приготовления тканевых срезов ткань подвергали затвердеванию в парафине, резали на микротоме до толщины 5 мкм и помещали на предметное стекло. Перед началом окрашивания проводили обработку при 60°C в течение 20 мин с использованием подогревателя предметных стекол. Затем проводили обработку ксилололом три раза по 3 мин 30 с. После этого проводили дважды обработку 100% этанолом и 95%, 80% и 70% этанолом по порядку один раз, по 3 мин 30 с каждый раствором. Затем проводили промывку под проточной водой в течение 3 мин. Далее проводили окрашивание гематоксилином в течение 5 мин с последующей промывкой в течение 10 мин HCl в течение 4 с, 1% аммиаком в течение 10 с, промывкой в течение 5 мин, и окрашивание эозином в течение 1 мин 10 с по порядку. Затем предметное стекло помещали в 70% этанол и 85% этанол по порядку на 1 мин 30 с, и затем проводили дважды обработку 100% этанолом в течение 1 мин 30 с. После обработки этанолом проводили обработку раствором, в котором были смешаны 100% этанол и ксилол в соотношении 1:1, в течение 1 мин 30 с. Наконец, проводили обработку ксилолом в течение 3 мин, и затем снова со свежим ксилолом в течение 4 мин. Предметное стекло закрывали покровным стеклом с использованием синтетической среды для заключения гистологических препаратов Shandon™ (Thermo Scientific, США). После завершения окрашивания препараты исследовали под микроскопом.
3-1. Оценка эффективности на животных моделях ксенотрансплантатов рака поджелудочной железы
Клетки MIA PaCa-2, которые представляют собой клеточные линии рака поджелудочной железы, ксенотрансплантировали иммунодефицитным голым мышам, для определения того, насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 11a и 11b, было подтверждено, что объем опухолей уменьшился на 60,46±13,33% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 55,40±15,04% по сравнению с группой, получавшей EXO-CON, и на 59,71±13,58% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 11c и 11d, было установлено, что масса опухолей уменьшалась на 52,55±12,13% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 46,74±13,61% по сравнению с группой, получавшей EXO-CON, и на 45,28±13,98% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея).
Кроме того, в результате исследования морфологии клеток опухолевой ткани после окрашивания H&E, как показано на фиг. 11e, размер цитоплазмы опухолевой ткани группы HEK Exo-FAF1 был меньше, чем у животных в группе MOCK (контроле), группе с рекомбинантным FAF1 и группе с HEK Exo-CON, и, таким образом, было подтверждено, что некроз клеток имел место за счет обработки HEK Exo-FAF1.
Вышеприведенные результаты показывают, что FAF1 через экзосомы также подавляет образование опухолей in vivo.
3-2. Оценка эффективности на животных моделях ксенотрансплантатов немелкоклеточного рака легких
Клетки A549, которые представляют собой клеточные линии немелкоклеточного рака легких, ксенотрансплантировали иммунодефицитным голым мышам, для определения того насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 12a и 12b, было подтверждено, что объем опухолей уменьшился на 75,45±9,89% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 72,77±10,97% по сравнению с группой, получавшей EXO-CON, и на 75,14±10,01% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 12c и 12d, было установлено, что масса опухолей уменьшилась на 59,07±13,88% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 61,31±13,12% по сравнению с группой, получавшей EXO-CON, и на 63,19±12,48% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 12e, было подтверждено, что некроз имел место внутри опухолей в группе, обработанной EXO-FAF1.
3-3. Оценка эффективности на животных моделях ксенотрансплантатов колоректального рака
Клеточные линии HCT 116, которые представляют собой клеточные линии колоректального рака, ксенотрансплантировали иммунодефицитным голым мышам, для определения того, насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 13a и 13b, было подтверждено, что объем опухолей уменьшался на 66,87±7,62% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 47,87±12,00% по сравнению с группой, получавшей EXO-CON, и на 56,93±9,91% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 13c и 13d, было установлено, что масса опухолей уменьшалась на 71,88±5,66% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 40,12±12,05% по сравнению с группой, получавшей EXO-CON, и на 60,09±8,03% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 13e, было подтверждено, что некроз имел место в опухолях в группе, обработанной EXO-FAF1.
3-4. Оценка эффективности на животных моделях ксетрансплантатов рака печени
Клеточные линии Hep3B, которые представляют собой линии клеток рака печени, ксенотрансплантировали иммунодефицитным голым мышам, для определения того, насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 14a и 14b, было подтверждено, что объем опухолей уменьшился на 76,16±11,61% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 70,74±14,25% по сравнению с группой, получавшей EXO-CON, и на 62,44±18,29% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 14c и 14d, было установлено, что масса опухолей уменьшилась на 73,48±33,22% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 66,62±42,16% по сравнению с группой, получавшей EXO-CON, и на 44,44±70,87% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 14e, было подтверждено, что некроз имел место в опухолях в группе, обработанной EXO-FAF1.
3-5. Оценка эффективности на животных моделях ксенотрансплантатов тройного негативного рака молочной железы
Клеточные линии MDA-MB-231, которые представляют собой клеточные линии тройного негативного рака молочной железы, ксенотрансплантировали иммунодефицитным голым мышам, для определения того насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 15a и 15b, было подтверждено, что объем опухолей уменьшился на 80,38±8,34% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 68,32±13,46% по сравнению с группой, получавшей EXO-CON, и на 67,22±13,93% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 15c и 15d, было установлено, что масса опухолей уменьшилась на 77,03±11,83% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 69,63±15,66% по сравнению с группой, получавшей EXO-CON, и на 62,50±19,33% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 15e, было подтверждено, что некроз имел место в опухолях в группе, обработанной EXO-FAF1.
3-6. Оценка эффективности на животных моделях ксенотрансплантатов рака почки
Клеточные линии Caki-1, которые представляют собой клеточные линии рака почки, ксентрансплантировали иммунодефицитным голым мышам, для определения того насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 16a и 16b, было подтверждено, что объем опухолей уменьшился на 54,23±12,40% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 43,05±15,21% по сравнению с группой, получавшей EXO-CON, и на 53,56±12,60% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 16c и 16d, было установлено, что масса опухолей уменьшилась на 42,11±16,22% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 31,25±19,26% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 16e, было подтверждено, что некроз имел место в опухолях в группе, обработанной EXO-FAF1.
3-7. Оценка эффективности на животных моделях ксенотрансплантатов рака шейки матки
Клетки HeLa, которые представляют собой клеточные линии рака шейки матки, ксенотрансплантировали иммунодефицитным голым мышам, для определения того насколько экзосомы, нагруженные FAF1, обладают способностью подавлять образование опухолей.
В результате, как показано на фиг. 17a и 17b, было подтверждено, что объем опухолей уменьшился на 66±11,33% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 56,4±14,52% по сравнению с группой, получавшей EXO-CON, и на 57,1±14,30% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 17c и 17d, было установлено, что масса опухолей уменьшилась на 60±17,59% в группе, получавшей экзосомы, нагруженные FAF1, по сравнению с группой, получавшей PBS, на 48,6±21,39% по сравнению с группой, получавшей EXO-CON, и на 70,7±12,17% по сравнению с группой, получавшей рекомбинантный FAF1 (AngioLab, Корея). Кроме того, как показано на фиг. 17e, было подтверждено, что некроз имел место в опухолях в группе, обработанной EXO-FAF1.
--->
Список последовательностей
<110> Kainos medicine, Inc.
<120> ЭКЗОСОМЫ, НАГРУЖЕННЫЕ FAS-АССОЦИИРОВАННЫМ ФАКТОРОМ 1 (FAF1), И ИХ
ПРИМЕНЕНИЕ В КАЧЕСТВЕ ПРОТИВООПУХОЛЕВОГО СРЕДСТВА
<130> 2022FPO-02-005PCT
<150> KR 10/20210040990
<151> 2021-03-30
<160> 1
<170> KoPatentIn 3.0
<210> 1
<211> 650
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Белок FAS-ассоциированного фактора 1
<400> 1
Met Ala Ser Asn Met Asp Arg Glu Met Ile Leu Ala Asp Phe Gln Ala
1 5 10 15
Cys Thr Gly Ile Glu Asn Ile Asp Glu Ala Ile Thr Leu Leu Glu Gln
20 25 30
Asn Asn Trp Asp Leu Val Ala Ala Ile Asn Gly Val Ile Pro Gln Glu
35 40 45
Asn Gly Ile Leu Gln Ser Glu Tyr Gly Gly Glu Thr Ile Pro Gly Pro
50 55 60
Ala Phe Asn Pro Ala Ser His Pro Ala Ser Ala Pro Thr Ser Ser Ser
65 70 75 80
Ser Ser Ala Phe Arg Pro Val Met Pro Ser Arg Gln Ile Val Glu Arg
85 90 95
Gln Pro Arg Met Leu Asp Phe Arg Val Glu Tyr Arg Asp Arg Asn Val
100 105 110
Asp Val Val Leu Glu Asp Thr Cys Thr Val Gly Glu Ile Lys Gln Ile
115 120 125
Leu Glu Asn Glu Leu Gln Ile Pro Val Ser Lys Met Leu Leu Lys Gly
130 135 140
Trp Lys Thr Gly Asp Val Glu Asp Ser Thr Val Leu Lys Ser Leu His
145 150 155 160
Leu Pro Lys Asn Asn Ser Leu Tyr Val Leu Thr Pro Asp Leu Pro Pro
165 170 175
Pro Ser Ser Ser Ser His Ala Gly Ala Leu Gln Glu Ser Leu Asn Gln
180 185 190
Asn Phe Met Leu Ile Ile Thr His Arg Glu Val Gln Arg Glu Tyr Asn
195 200 205
Leu Asn Phe Ser Gly Ser Ser Thr Ile Gln Glu Val Lys Arg Asn Val
210 215 220
Tyr Asp Leu Thr Ser Ile Pro Val Arg His Gln Leu Trp Glu Gly Trp
225 230 235 240
Pro Thr Ser Ala Thr Asp Asp Ser Met Cys Leu Ala Glu Ser Gly Leu
245 250 255
Ser Tyr Pro Cys His Arg Leu Thr Val Gly Arg Arg Ser Ser Pro Ala
260 265 270
Gln Thr Arg Glu Gln Ser Glu Glu Gln Ile Thr Asp Val His Met Val
275 280 285
Ser Asp Ser Asp Gly Asp Asp Phe Glu Asp Ala Thr Glu Phe Gly Val
290 295 300
Asp Asp Gly Glu Val Phe Gly Met Ala Ser Ser Ala Leu Arg Lys Ser
305 310 315 320
Pro Met Met Pro Glu Asn Ala Glu Asn Glu Gly Asp Ala Leu Leu Gln
325 330 335
Phe Thr Ala Glu Phe Ser Ser Arg Tyr Gly Asp Cys His Pro Val Phe
340 345 350
Phe Ile Gly Ser Leu Glu Ala Ala Phe Gln Glu Ala Phe Tyr Val Lys
355 360 365
Ala Arg Asp Arg Lys Leu Leu Ala Ile Tyr Leu His His Asp Glu Ser
370 375 380
Val Leu Thr Asn Val Phe Cys Ser Gln Met Leu Cys Ala Glu Ser Ile
385 390 395 400
Val Ser Tyr Leu Ser Gln Asn Phe Ile Thr Trp Ala Trp Asp Leu Thr
405 410 415
Lys Asp Ser Asn Arg Ala Arg Phe Leu Thr Met Cys Asn Arg His Phe
420 425 430
Gly Ser Val Val Ala Gln Thr Ile Arg Thr Gln Lys Thr Asp Gln Phe
435 440 445
Pro Leu Phe Leu Ile Ile Met Gly Lys Arg Ser Ser Asn Glu Val Leu
450 455 460
Asn Val Ile Gln Gly Asn Thr Thr Val Asp Glu Leu Met Met Arg Leu
465 470 475 480
Met Ala Ala Met Glu Ile Phe Thr Ala Gln Gln Gln Glu Asp Ile Lys
485 490 495
Asp Glu Asp Glu Arg Glu Ala Arg Glu Asn Val Lys Arg Glu Gln Asp
500 505 510
Glu Ala Tyr Arg Leu Ser Leu Glu Ala Asp Arg Ala Lys Arg Glu Ala
515 520 525
His Glu Arg Glu Met Ala Glu Gln Phe Arg Leu Glu Gln Ile Arg Lys
530 535 540
Glu Gln Glu Glu Glu Arg Glu Ala Ile Arg Leu Ser Leu Glu Gln Ala
545 550 555 560
Leu Pro Pro Glu Pro Lys Glu Glu Asn Ala Glu Pro Val Ser Lys Leu
565 570 575
Arg Ile Arg Thr Pro Ser Gly Glu Phe Leu Glu Arg Arg Phe Leu Ala
580 585 590
Ser Asn Lys Leu Gln Ile Val Phe Asp Phe Val Ala Ser Lys Gly Phe
595 600 605
Pro Trp Asp Glu Tyr Lys Leu Leu Ser Thr Phe Pro Arg Arg Asp Val
610 615 620
Thr Gln Leu Asp Pro Asn Lys Ser Leu Leu Glu Val Lys Leu Phe Pro
625 630 635 640
Gln Glu Thr Leu Phe Leu Glu Ala Lys Glu
645 650
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭКЗОСОМАЛЬНЫЕ БИОМАРКЕРЫ | 2015 |
|
RU2712223C2 |
| НОВЫЕ МЕДИЦИНСКИЕ АГЕНТЫ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2739073C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СТИМУЛЯЦИИ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ | 2020 |
|
RU2839664C2 |
| ПЕПТИД, ПОЛУЧЕННЫЙ ИЗ DEPDC1, И СОДЕРЖАЩАЯ ЕГО ВАКЦИНА | 2016 |
|
RU2765574C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К BCMA И BCMA-CAR-NK-КЛЕТКИ | 2020 |
|
RU2795443C2 |
| ПЕПТИД, ПОЛУЧЕННЫЙ ИЗ FOXM1, И ВКЛЮЧАЮЩАЯ ЕГО ВАКЦИНА | 2016 |
|
RU2738418C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К BCMA И BCMA-CAR-T-КЛЕТКИ | 2020 |
|
RU2762942C1 |
| ПЕПТИД, ПОЛУЧЕННЫЙ ИЗ MPHOSPH1, И ВКЛЮЧАЮЩАЯ ЕГО ВАКЦИНА | 2016 |
|
RU2731099C2 |
| ВЕЗИКУЛЫ, СОДЕРЖАЩИЕ ИНГИБИТОР PTEN, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2800729C2 |
| НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, ЭКСПРЕССИРУЮЩАЯ ЯКОРНЫЙ БЕЛОК ЭКЗОСОМ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ВАКЦИНЫ | 2017 |
|
RU2761642C2 |












Группа изобретений относится к биотехнологии. Представлены применения экзосом, нагруженных белком fas-ассоциированного фактора 1 (FAF1), и способ лечения рака. Суть изобретения заключается в выделении экзосом, нагруженных белком FAF1, из клеток HEK 293, в которых сверхэкспрессируется FAF1, обладающий известной опухоль-супрессирующей функцией, затем вводили экзосомы, нагруженные белком FAF1, в опухолевую модель, где клетки рака поджелудочной железы (MIA PaCa-2), клетки рака легкого (A549), клетки рака толстого кишечника (HCT 116), клетки рака печени (Hep3B), клетки рака молочной железы (MDA-MB-231), клетки рака почки (Caki-1) и клетки рака шейки матки (клетки HeLa) трансплантировали голым мышам интратуморальной инъекцией и выявили, что наблюдались эффекты ингибирования роста опухолей, более выраженные, чем в контрольной группе, не обработанной FAF1. Таким образом, экзосомы, нагруженные FAF1, по настоящему изобретению можно использовать в качестве терапевтического агента для лечения различных видов рака. 4 н. и 8 з.п. ф-лы, 19 ил., 3 пр.
1. Применение экзосом, нагруженных белком Fas-ассоциированного фактора 1 (FAF1), для лечения рака, где рак представляет собой рак, при котором экспрессия FAF1 снижена.
2. Применение по п.1, где белок FAF1 содержит аминокислотную последовательность, показанную в SEQ ID NO: 1.
3. Применение по п.1, где экзосомы имеют диаметр 50-200 нм.
4. Применение по п.1, где белок FAF1, нагруженный в экзосомы, подавляет образование опухолей из других клеток.
5. Применение по п.1, где рак представляет собой рак шейки матки, рак поджелудочной железы, рак печени, рак легкого, рак молочной железы, рак почки, колоректальный рак, плоскоклеточный рак головы и шеи, рак мочевого пузыря, рак предстательной железы, рак желудка, рак эндометрия, рак мозга, рак яичника, рак яичка, рак головы, рак шеи, рак кожи, рак мезотелиальной выстилки, рак пищевода, рак надпочечников, рак щитовидной железы, рак кости, глиобластому, мезотелиому, саркому, хориокарциному, базальноклеточную карциному кожи, семиному яичка, лейкоз или злокачественную лимфому.
6. Применение по п.1, где экзосомы доставляют FAF1 к опухолевым клеткам-мишеням.
7. Применение по п.1, где экзосомы вводят посредством перорального введения, интрабронхиальной ингаляции или внутрибрюшинной инъекции, ректально, подкожной, внутривенной, внутримышечной, интраторакальной или интратуморальной инъекции.
8. Применение по п.1, где экзосомы вводят самостоятельно или одновременно и последовательно с другими терапевтическими агентами.
9. Применение по п.2, где FAF1 SEQ ID NO: 1 загружен в экзосомы.
10. Применение композиции для лечения рака, где рак представляет собой рак, при котором экспрессия FAF1 снижена, где композиция содержит экзосомы, нагруженные белком Fas-ассоциированного фактора 1 (FAF1).
11. Способ лечения рака, где рак представляет собой рак, при котором экспрессия FAF1 снижена, включающий введение экзосом, нагруженных белком FAF1, субъекту, который имеет сниженную экспрессию FAF1.
12. Применение экзосом, нагруженных белком FAF1, для производства лекарственного средства для лечения рака, где рак представляет собой рак, при котором экспрессия FAF1 снижена.
| GYEONGRIN PARK et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ использования делительного аппарата ровничных (чесальных) машин, предназначенных для мериносовой шерсти, с целью переработки на них грубых шерстей | 1921 |
|
SU18A1 |
| WO 9640864 A1, 19.12.1996 | |||
| CN 101899443 A, 01.12.2010 | |||
| RANJAN UPADHYAY et al | |||
| A Critical Role for Fas-Mediated Off-Target Tumor Killing in T-cell Immunotherapy, Cancer Discov., | |||

