ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к инактивированным токсинам ApxIA, ApxllA и ApxlllA, вакцинам, содержащим указанные инактивированные токсины, и их применению для иммунизации и защиты млекопитающих.
УРОВЕНЬ ТЕХНИКИ
Actinobacillus pleuropneumoniae (APP) представляет собой грам-отрицательные бактерии и является членом семейства Pasteurellaceae. APP представляет собой этиологический агент плевропневмонии свиней, тяжелого легочного заболевания свиней, которое вызывает сильные экономические потери в свиноводстве по всему миру. Заболевание характеризуется геморрагическим, фибринозным и некротическим поражением легких. Свиньи, которые выжили после заболевания, часто становятся бессимптомными носителями APP и являются основной причиной бактериальной диссеминации.
На данный момент идентифицировано 18 серотипов на основе антигенных свойств капсульных полисахаридов (КПС, CPS) и в результате генотипического анализа. Основные факторы вирулентности APP представляют собой экзотоксины, КПС, липополисахариды (ЛПС, LPS) и мембранные белки. Наиболее важные факторы вирулентности представляют собой экзотоксины Apx, которые принадлежат к порообразующим повторам в семействе токсинов (RTX). Известно, что токсины являются высокоиммуногенными и крайне необходимыми для получения защитного иммунитета против APP, связанного с плевропневмонией. По меньшей мере четыре различных токсина Apx продуцируются APP, обозначаемые как Apxl, Apxll, Apxlll и ApxIV. Apxl демонстрирует сильную, а Apxll демонстрирует низкую гемолитическую активность. Оба являются цитотоксическими и активными против широкого спектра клеток различных организмов-хозяев. Apxlll не является гемолитическим, но является крайне цитотоксическим, основными мишенями которого являются макрофаги и нейтрофилы альвеол свиней. ApxIV не демонстрирует цитотоксической и только очень слабую гемолитическую активность. Никакой из серотипов APP не продуцирует все четыре токсина Apx. Большинство серотипов продуцируют три токсина Apx. Схема продуцирования токсина Apx связана с вирулентностью, при этом серотипы 1, 5, 9 и 11 продуцируют Apxl и Apxll, являющиеся наиболее вирулентными.
По меньшей мере четыре гена отвечают за продуцирование и секрецию активных токсинов Apx. Ген A кодирует структурный токсин, а ген C кодирует ацилтрансферазу, необходимую для пост-трансляционной активации токсина. Гены B и D кодируют два мембранных белка, которые необходимы для секреции зрелого токсина. Гены Apx организованы как опероны. Опероны токсинов Apxl и Apxlll состоят из генов CABD, тогда как оперон Apxll содержит только гены CA. Следовательно, секреция ApxllA зависит от активных генов B и D оперона Apxl.
В настоящий момент плевропневмонию, возникающую вследствие инфекции APP свиней, обычно лечат антибиотиками. Однако, было установлено, что APP часто демонстрирует антибиотикоустойчивость к по меньшей мере одному из антибиотиков, обычно используемых для лечения инфекции APP.
Вакцинация против APP представляет собой перспективную профилактическую стратегию для профилактики плевропневмонии. На рынке имеется несколько вакцин. Коммерчески доступные вакцины являются или химически инактивированными цельноклеточными вакцинами, или субъединичными вакцинами, или их комбинацией. Иммунологическая реакция животных, вакцинированных цельноклеточными вакцинами, в основном направлена на поверхностные структуры, такие как КПС и ЛПС. Отсутствие секретируемых белков, таких как токсины Apx, которые, как известно, являются высокоиммуногенными и необходимыми для защиты, объясняет ограниченную защиту, наблюдаемую у бактериальных вакцин. Кроме того, цельноклеточные вакцины APP обеспечивают только гомологическую защиту против серотипа, используемого для получения вакцины.
Коммерчески доступная субъединичная вакцина (патент EP0453024 B1) содержит химически инактивированные токсины ApxA, а также наружный белок мембран. Инактивация токсинов ApxA денатурирующими веществами, такими как формальдегид, потенциально приводит к снижению иммуногенности токсоидов. Недостатками такой вакцины являются недостаточная защита вследствие денатурирующих токсинов, а также сопутствующие серьезные побочные реакции, вероятно, вследствие остаточной токсичности, из-за неполной инактивации токсинов ApxA. Следовательно, вследствие сниженной иммуногенности после инактивации необходимо использовать большие количества токсинов, что приводит к увеличению количеств загрязняющего ЛПС в вакцинах. Высокое содержание ЛПС может вызвать побочные эффекты, как это наблюдается для коммерческих субъединичных вакцин APP.
Можно сделать вывод, что имеющиеся на данный момент коммерческие вакцины не обладают удовлетворительными профилями безопасности и/или эффективности против инфекции APP.
Более того, существуют некоторые другие экспериментальные подходы к вакцинам APP.
В патентной заявке W02004045639 (Al) раскрыта живая ослабленная вакцина против плевропневмонии свиней, содержащая штамм APP, который модифицирован в трансмембранных доменах генов, кодирующих токсины ApxIA и ApxllA. Для определения степени ослабления свиней в возрасте три месяца прививали модифицированными живыми штаммами APP. Через семь дней после привития животных умерщвляли и регистрировали макроскопические поражения органов дыхания. Все животные продемонстрировали изменение поведения и поражения легких при вскрытии. Эффективность такой живой вакцины не изучали.
В патентной заявке EP0810283 (A2) и патенте EP0861319 (B1) описаны живые ослабленные штаммы APP, содержащие делеции в генах активатора ApxA apxC. Модифицированный штамм APP не продуцирует белок-активатор ApxC в функциональной форме, а следовательно токсины ApxIA и ApxllA не активировались ацилированием. Мышей вакцинировали штаммом ΔapxC и заражали вирулентными полевыми штаммами APP. Вакцинированные мыши были защищены от гомологического заражения и частично защищены от гетерологического заражения. Было выполнено одно исследование на свиньях. У 1 из 6 вакцинированных свиней наблюдались поражения легких после гетерологического заражения при вскрытии. Эти ослабленные живые вакцины, вероятно, являются эффективными, но такие вакцины несут риск безопасности. Вакцинные штаммы продуцируют токсины Apx, которые не активируются ацилированием вследствие отсутствия apxC. Однако эти токсины обладают исходной аминокислотной последовательностью токсического ApxA. Очень вероятно, что гетерологическая ацилтрансфераза может ацилировать ApxA, превращая его в его активную токсическую форму. На большинстве свиноферм по всему миру есть бессимптомные носители вирулентных штаммов APP, а также свиньи, инфицированные штаммами APP с низкой вирулентностью. Если таких свиней вакцинировали с помощью вакцинного штамма ΔapxC, неактивированные токсины ApxA вакцинного штамма могут быть активированы функциональными белками ApxC полевых штаммов. Кроме того, существуют возможность комплементации делеции apxC путем захвата функциональных генов apxC. Существует возможность того, что ослабленный штамм вновь станет патогенным, что приведет к заболеванию привитых животных.
На сегодняшний момент на рынке нет ослабленных живых вакцин.
Следовательно, существует необходимость в усовершенствованных вакцинах против APP, которые являются безопасными и способными индуцировать перекрестный иммунитет против всех существенных серотипов у свиней и/или поросят.
Следовательно, объектом данного изобретения является получение инактивированных токсинов ApxIA, ApxllA и ApxlllA и применение этих токсоидов в безопасных и эффективных субъединичной и живой вакцинах против плевропневмонии свиней.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В данном изобретении раскрыты модификации сайтов ацилирования токсинов ApxIA, ApxllA и ApxlllA для создания неактивных, но полностью иммуногенных токсоидных форм этих белков. Замена обеих аминокислот двух сайтов ацилирования на другие предотвращает их ацилирование. Следовательно, эти модифицированные токсины Apx не могут инициировать связывание с мембраной клетки-мишени и не обладают цитотоксической или гемолитической активностью. Нет необходимости химически инактивировать токсины, что дает высокоиммуногенные белки. Требуемое количество этих белков для получения эффективной субъединичной вакцины является относительно низким. Следовательно, содержание загрязняющего ЛПС также является относительно низким, что приводит к незначительным побочным эффектам, таким как лихорадка, рвота и апатия. Вследствие лучшей иммуногенности по сравнению с химически инактивированными субъединицами Apx, субъединичные вакцины по данному изобретению обеспечивают лучший профиль безопасности.
Кроме того, изобретение можно применять в живых вакцинах, поскольку замена двух аминокислот основана на генетической модификации генов apxA. Таким образом, они могут быть перенесены в штаммы APP, которые затем продуцируют инактивированные и высокоиммуногенные белки Apx. Эти штаммы можно использовать в качестве живых вакцин, которые продуцируют токсоиды ApxA, а также широкий спектр других белков APP. Эти вакцины индуцируют комплексный иммунный ответ и перекрестный иммунитет против широкого спектра серотипов APP. Вследствие генетической модификации генов apxA полученные токсоиды Apx не могут быть активированы гетерологичными ацилтрансферазами. Существует еще несколько преимуществ при использовании живых вакцинных штаммов APP. Ожидается, что влияние исходных антител на вакцину будет ниже при использовании живых вакцин по сравнению с субъединичными вакцинами, состоящими из высокоиммуногенных антигенов. Следовательно, с высокой вероятностью вакцину можно использовать для поросят возрастом до 6 недель, и существующий иммунологический пробел среди имеющихся вакцин будет устранен. Дополнительные нокауты генов можно использовать для получения еще более ослабленного вакцинного штамма и для использования нокаутов генов как маркеров, чтобы различить инфицированных и вакцинированных животных (DIVA), например, по делеции гена apxIVA. Следовательно, в этом изобретении предложен лучший профиль безопасности и эффективности, чем для всех известных подходов к получению вакцин. Поскольку APP сильно приспособлен к свиньям, ослабленный вакцинный штамм APP можно также использовать в качестве вектора для антигенов других патогенов, которые имеют отношение к свиньям.
Изобретение определено прилагаемой формулой изобретения.
Раскрыты полипептиды ApxIA, ApxllA или ApxlllA Actinobacillus pleuropneumoniae, в которых
a. аминокислотная последовательность
i. ApxIA дикого типа, как представлено в SEQ ID №:1, содержит по меньшей мере одну модифицированную аминокислоту, выбранную из группы, состоящей из K560 и K686,
ii. ApxllA дикого типа, как представлено в SEQ ID №:2, содержит по меньшей мере одну модифицированную аминокислоту, выбранную из группы, состоящей из K557 и N687, и
iii. ApxlllA дикого типа, как представлено в SEQ ID №: 3, содержит по меньшей мере одну модифицированную аминокислоту, выбранную из группы, состоящей из K571 и K702,
и по меньшей мере одна аминокислота заменена на аминокислоту, не подверженную ацилированию; или
b. аналог или фрагмент модифицированной аминокислотной последовательности, определенной в a., по меньшей мере на 90% гомологичен модифицированной аминокислотной последовательности, определенной в a., и фрагмент содержит по меньшей мере 50% аминокислот подряд модифицированной аминокислотной последовательности, определенной в a., и причем указанный аналог или фрагмент содержит аминокислоту, не подверженную ацилированию, как определено в a; или
c. аминокислотная последовательность
i. ApxIA дикого типа, как представлено в SEQ ID №:1, содержит делеции, содержащие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из K560 и K686,
ii. ApxllA дикого типа, как представлено в SEQ ID №:2, содержит делеции, содержащие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из K557 и N687, и
iii. ApxlllA дикого типа, как представлено в SEQ ID №:3, содержит делеции, содержащие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из K571 и K702,
при этом указанные делеции удаляют не более 50% аминокислотной последовательности дикого типа;
d. аналог аминокислотной последовательности, содержащий делеции, как определено в c., по меньшей мере на 90% гомологичен аминокислотной последовательности, содержащей делеции, как определено в c.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Следующие заголовки служат только для организации раскрытия, и их не следует рассматривать как отделяющие один раздел описания от другого раздела описания. Признаки каждого раздела можно объединять с признаками любого другого раздела.
ОБЩИЕ ОПРЕДЕЛЕНИЯ
Термин «фрагмент или аналог», при использовании в данном документе, имеют следующие определения:
«Аналог» можно рассматривать как последовательность аминокислот, демонстрирующую уровень гомологичности, составляющий по меньшей мере 60%, 70%, 80%, 85%, 90%, 95% или 97% по сравнению с исходной аминокислотной последовательностью. Гомология, при использовании в данном документе, означает идентичность. Таким образом последовательности могут отличаться друг от друга на основании консервативного замещения, делеции или вставки.
Степень идентичности можно определить с использованием программы для белков Blast с помощью алгоритма Blastp с параметрами по умолчанию, которые представляют собой, например, ожидаемый порог: 10, длина сегмента: 3, матрица: BLOMSUM62, цена гэпа: наличие: 11, расширение: 1, и композиционная корректировка: условная композиционная корректировка матрицы замен (BLAST представляет собой зарегистрированную торговую марку Национальной медицинской библиотеки). Программу можно использовать для поиска в белковой базе данных используя запрос по белку. Идентичность свидетельствует о точных совпадениях между выровненным запрашиваемым белком и последовательностями из базы данных.
Консервативные аминокислотные «замены» предпочтительно представляют собой результат замещения одной аминокислоты на другую аминокислоту с подобными структурными и/или химическими свойствами, т.е. консервативные замены аминокислот. Замены аминокислот могут быть выполнены на основе схожести в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфифильной природе вовлеченных остатков. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; и отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту.
«Вставки» или «делеции», как правило, находятся в диапазоне около 1, 2 или 3 аминокислот. Разрешенный вариант может быть определен экспериментально путем систематического введения вставок или делеций аминокислот в белок с использованием рекомбинантных технологий ДНК и анализа активности полученных рекомбинантных вариантов. Это не требует более, чем обычных экспериментов для специалиста в данной области техники.
«Фрагмент» полипептида содержит по меньшей мере 50%, 60%, 70%, 80%, 90%, 95% или 97% от исходного полипептида. Эти фрагменты можно использовать как исключительный активный ингредиент в вакцине в соответствии с данным изобретением.
«Токсин» APP представляет собой полипептид, который состоит из аминокислотной последовательности ApxIA, ApxllA или ApxlllA (например, как представлено в SEQ ID №: 1-3) и демонстрирует цитотоксическую и/или гемолитическую активность. «Токсоид» в этом описании представляет собой полипептид, который представляет собой модифицированную форму «токсина», причем модификацию получают путем замещения или делеции аминокислоты, которая подвержена ацилированию, in vivo в APP, и не демонстрирует какой-либо цитотоксической или гемолитической активности.
Термин «содержащий», при использовании в данном документе в контексте композиции вакцины, означает, что могут присутствовать дополнительные активные или иммуногенные компоненты. «Состоящий из» означает, что не присутствует никаких дополнительных компонентов, а «состоящий по существу из» означает, что могут присутствовать конкретные дополнительные компоненты, а именно те, которые существенно не влияют на ключевые характеристики вакцины (т.е. неактивные или неиммуногенные ингредиенты).
Род Actinobacillus включает грам-отрицательные, неспорообразующие и в основном инкапсулированные бактериальные виды, которые колонизируются на поверхностях слизистых респираторной и мочеполовой систем. Релевантные для ветеринарии виды представляют собой, например, A. pleuropneumoniae, A. suis, A. equuli и A. lignieresii, которые являются предпочтительными видами Actinobacillus в этом описании. Род Actinobacillus обычно демонстрирует сильную специфичность к хозяину. Предпочтительные серотипы APP представляют собой серотипы 1, 5, 7, 8, 9 и 11.
ПОЛИПЕПТИДЫ
Описание относится к полипептидам ApxIA, ApxllA или ApxlllA (далее взаимозаменяемо обозначены как полипептиды ApxA для краткости), причем полипептиды ApxIA, ApxllA или ApxlllA дикого типа модифицированы путем замены или делеции аминокислот в положениях аминокислот, подверженных ацилированию в полипептиде дикого типа. При замещении аминокислоты вводится аминокислота, которая не подвержена ацилированию. Неожиданно было установлено, что полипептиды ApxIA, ApxllA или ApxlllA, модифицированные вышеописанным способом, более не демонстрируют гемолитическую и/или цитотоксическую активность. Таким образом, они могут проявлять меньше побочных эффектов при использовании для вакцинирования млекопитающего, при этом обеспечивая иммунологическую защиту от APP. Следовательно, их можно рассматривать как инактивированные токсины, т.е. токсоиды. Следовательно, полипептиды по изобретению можно применять безопасно в вакцинах для иммунизации субъектов, в частности млекопитающих, против инфекции APP.
Полипептиды, содержащие замены в положениях аминокислот, подверженных ацилированию, в полипептиде дикого типа
В частности описание относится к полипептиду ApxIA, ApxllA или ApxlllA Actinobacillus pleuropneumoniae, причем
a. аминокислотная последовательность
i. ApxlA дикого типа, как представлено в SEQ ID №: 1, содержит по меньшей мере одну модифицированную аминокислоту, выбранную из группы, состоящей из K560 и K686,
ii. ApxllA дикого типа, как представлено в SEQ ID №: 2, содержит по меньшей мере одну модифицированную аминокислоту, выбранную из группы, состоящей из K557 и N687, и
iii. ApxlllA дикого типа, как представлено в SEQ ID №: 3, содержит по меньшей мере одну модифицированную аминокислоту, выбранную из группы, состоящей из K571 и K702,
и по меньшей мере одна аминокислота заменена на аминокислоту, не подверженную ацилированию.
Полипептид может вызывать гуморальный и/или клеточный иммунологический ответ против одного или более серотипов APP у млекопитающего, в частности свиньи, при введении указанному млекопитающему. Полипептид может вызывать гуморальный и/или клеточный иммунологический ответ против одного или более штаммов APP, например 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 штаммов APP, у млекопитающего, в частности свиньи, при введении указанному млекопитающему. Предпочтительно, полипептиды вызывают стерильный иммунитет против APP, штаммов APP или серотипов APP.
Каждый из ApxIA, ApxllA или ApxllIA содержит две аминокислоты, которые подвержены ацилированию. Модификации могут затрагивать одну или обе указанные аминокислоты.
Описание также относится к аналогу и/или фрагменту модифицированной аминокислотной последовательности, определенной выше, который по меньшей мере на 90% гомологичен модифицированной аминокислотной последовательности, определенной выше, и фрагмент содержит по меньшей мере 50% аминокислот подряд модифицированной аминокислотной последовательности, определенной выше, и аналог или фрагмент содержит аминокислоту, не подверженную ацилированию, как определено выше.
Как определено выше, аналог может отличаться от дикого типа за счет делеций или консервативных замен. Однако аналог всегда содержит те аминокислоты, которые заменяют аминокислоты, подверженные ацилированию.
Поскольку модифицированный полипептид не содержит аминокислоты, подверженные ацилированию, модифицированный полипептид не может быть ацилирован. Полипептиды, модифицированные таким образом, не могут быть ацилированы с помощью любой эндогенной или экзогенной ацилтрансферазы (например, ApxC Actinobacillus pleuropneumoniae). Как экспериментально показано ниже, такие модифицированные полипептиды больше не проявляют значительных патологических эффектов и, следовательно, могут безопасно использоваться в композициях вакцин. Эти модифицированные полипептиды являются более безопасными для применения в композициях вакцин, чем вакцины, в которых удалена ацилтрансфераза ApxC, поскольку они остаются патологическими, даже если ацилтрансфераза поступает извне, или за счет встречающегося в природе штамма APP, или за счет любого другого источника ацилтрансферазы in vivo. Те же рассуждения применяются к следующим полипептидам, в которых одна или две аминокислоты APP, которые подвержены ацилированию, удалены.
Полипептиды, содержащие делеции в положениях аминокислот, подверженных ацилированию, в полипептиде дикого типа
Вместо замены аминокислот, подверженных ацилированию, в полипептиде дикого типа, аминокислоты также можно удалить, чтобы ликвидировать патологические эффекты полипептида дикого типа.
Следовательно, описание также относится к полипептиду ApxIA, ApxllA или ApxlllA Actinobacillus pleuropneumoniae, при этом аминокислотная последовательность
i. ApxIA дикого типа, как представлено в SEQ ID №:1, содержит делеции, содержащие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из K560 и K686,
ii. ApxllA дикого типа, как представлено в SEQ ID №:2, содержит делеции, содержащие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из K557 и N687, и
iii. ApxlllA дикого типа, как представлено в SEQ ID №:3, содержит делеции, содержащие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из K571 и K702.
Полипептид может вызывать гуморальный и/или клеточный иммунологический ответ против одного или более серотипов APP у млекопитающего, в частности свиньи, при введении указанному млекопитающему. Полипептид может вызывать гуморальный и/или клеточный иммунологический ответ против одного или более штаммов APP, например 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 штаммов APP, у млекопитающего, в частности свиньи, при введении указанному млекопитающему. Предпочтительно, полипептиды вызывают стерильный иммунитет против APP, серотипов APP, штаммов APP или серотипов APP.
Как указано выше, каждый из ApxIA, ApxIIA или ApxIIIA содержит две аминокислоты, которые подвержены ацилированию. Делеция указанных аминокислот может включать одну или обе из указанных аминокислот.
Следовательно, делеции могут включать точечные делеции, при этом удаляют только любую одну или две аминокислоты, подверженные ацилированию, в каждой последовательности дикого типа. Также предусмотрено, что делециями также удаляют аминокислоты в области, смежной с одной или двумя аминокислотами, подверженными ацилированию. Таким образом, соответствующие делеции могут включать 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 200, 300, 350, 400 аминокислот при условии, что делеция содержит одну из двух аминокислот, подверженных ацилированию. Размер делеции для каждой аминокислоты, подверженной ацилированию, может не зависеть от других или иметь такой же размер. Также рассматриваются делеции, которые охватывают непрерывный участок аминокислот между двумя аминокислотами, подверженными ацилированию.
Делеции удаляют не более 50% аминокислотной последовательности дикого типа.
Описание также относится к аналогу модифицированной аминокислотной последовательности, определенной выше, который по меньшей мере на 90% гомологичен модифицированной аминокислотной последовательности, определенной выше, и аналог не содержит аминокислоту, не подверженную ацилированию, как определено выше. Такой аналог может содержать консервативные замены и делеции, как определено в разделе, который относится к общим определениям. Таким образом, полипептид может содержать дополнительные делеции за пределами удаленного участка, содержащего одну или две аминокислоты, подверженные ацилированию, но эти делеции будут удалять только 1, 2, 3 аминокислоты.
АМИНОКИСЛОТЫ, НЕ ПОДВЕРЖЕННЫЕ АЦИЛИРОВАНИЮ
Аминокислоты, подверженные ацилированию, представляют собой встречающиеся в природе аминокислоты, например лизин и/или аспарагин. Аминокислоты, не подверженные ацилированию, известны специалисту в данной области техники и могут использоваться для замены одной или обеих аминокислот, подверженных ацилированию. Например, аминокислота, не подверженная ацилированию, может быть независимо выбрана из группы, состоящей из аланина, глицина, изолейцина, лейцина, метионина, валина, серина, треонина, аспарагина, глутамина, аспарагиновой кислоты, гистидина, аспарагиновой кислоты, цистеина, пролина, фенилаланина, тирозина, триптофана и глутаминовой кислоты. В частности, аминокислота, не подверженная ацилированию, может быть независимо выбрана из группы, состоящей из аланина, глицина, серина, треонина, валина, изолейцина и лейцина.
Кроме того, аминокислота, не подверженная ацилированию, может быть независимо выбрана из группы, состоящей из аланина, глицина и серина. Наиболее предпочтительная аминокислота, не подверженная ацилированию, представляет собой аланин.
ПОЛИПЕПТИД APXIA, APXIIA ИЛИ APXIIIA С ДВУМЯ МОДИФИЦИРОВАННЫМИ АМИНОКИСЛОТАМИ
Предпочтительно, в каждом из ApxIA, ApxllA или ApxlllA модифицированы две аминокислоты, подверженные ацилированию.
Следовательно, последовательность ApxIA дикого типа, как представлено в SEQ ID №:1, модифицирована в K560 и K686. Следовательно, последовательность ApxllA дикого типа, как представлено в SEQ ID №:2, модифицирована в K557 и N687. Следовательно, последовательность ApxlllA дикого типа, как представлено в SEQ ID №: 3, модифицирована в K571 и K702.
Раскрыто также то, что указанные две аминокислоты, подверженные ацилированию, в каждом из ApxIA, ApxllA или ApxlllA изменены на аланин.
Таким образом, раскрытый модифицированный полипептид может иметь последовательность, представленную в SEQ ID №: 4, 5 или 6, или представляет собой ее аналог или фрагмент, как определено выше.
НУКЛЕИНОВЫЕ КИСЛОТЫ
Также раскрыты нуклеиновые кислоты, содержащие последовательность нуклеиновой кислоты, способной кодировать описанные выше полипептиды. Раскрытая нуклеиновая кислота может представлять собой кДНК, ДНК, РНК, кРНК или ПНК (пептидо-нуклеиновая кислота). Термин «последовательность нуклеиновой кислоты» относится к гетерополимеру нуклеотидов последовательности этих нуклеотидов. Нуклеиновая кислота может содержать нуклеиновую кислоту, представленную в SEQ ID №: 10, 11 или 12.
Нуклеиновая кислота может входить в состав вектора, пригодного для клонирования или экспрессирования нуклеиновых кислот по изобретению. Иллюстративные векторы представляют собой pEX-A258 (SEQ ID №: 19), pQE-80L (SEQ ID №: 20) или pQE-60 (SEQ ID №: 21). Нуклеиновые кислоты или векторы могут содержать дополнительные регулирующие некодирующие элементы, такие как индуцируемые или не-индуцируемые промоторы, операторы (например, lac-оператор), или нуклеиновые кислоты, кодирующие другие белки AP. Дополнительно, предпочтительно, чтобы композиция вакцины в соответствии с данным изобретением дополнительно характеризовалась полинуклеотидом, который интегрирован в вектор, при этом полинуклеотид функционально связан областью контроля экспрессии в векторе. Таким образом, также описан экспрессионный вектор, при этом экспрессионный вектор предпочтительно содержит одну или более регуляторных последовательностей в дополнение к последовательностям нуклеиновой кислоты, кодирующим модифицированные формы ApxA. Термин «экспрессионный вектор» в общем относится к плазмиде, или фагу, или вирусу, или вектору для экспрессии полипептида из последовательности ДНК (РНК). Экспрессионный вектор может содержать единицу транскрипции, содержащую сборку из (1) генетического элемента или элементов с регуляторной ролью при экспрессии гена, например промоторы или энхансеры, (2) структурной или кодирующей последовательности, которая транскрибируется в мРНК и транслируется в белок, и (3) соответствующих последовательностей начала или конца транскрипции. Структурные единицы, предназначенные для применения в дрожжевых или эукариотических экспрессионных системах, предпочтительно включают лидерную последовательность, которая делает возможной внеклеточную секрецию транслируемого белка клеткой-хозяином. Альтернативно, когда рекомбинантный белок экспрессируется без лидерной или транспортной последовательности, он может включать N-концевой метиониновый остаток. Этот остаток затем может отщепляться или не отщепляться от экспрессионного рекомбинантного белка, чтобы получить конечный продукт.
МИКРООРГАНИЗМЫ
Также раскрыты микроорганизмы, содержащие раскрытые нуклеиновые кислоты и/или вектора, описанные выше. Микроорганизмы могут представлять собой Escherichia coli (E. coli), штамм E. coli, например штамм ToplOF' E. Coli,, или Actinobacillus pleuropneumoniae, штамм Actinobacillus pleuropneumoniae, например ST2 (например, APP23), ST2 (например, 07/07), ST5 (например, DZY47), ST7 (например, DZY33) или ST8 (например, DZY49). Микроорганизмы также включают нуклеиновые кислоты и/или векторы, кодирующие другие гены.
КОМПОЗИЦИЯ ВАКЦИНЫ
Также раскрыта композиция вакцины, содержащая по меньшей мере один модифицированный полипептид ApxIA, ApxllA или ApxlllA, как определено выше, молекулу нуклеиновой кислоты, как определено выше, или вектор, как определено выше, или микроорганизм, как определено выше. Композиция вакцины может содержать по меньшей мере фармацевтический носитель, разбавитель и/или адъювант.
Получение композиции вакцины в соответствии с изобретением известно в данной области техники и описано в справочных руководствах, известных специалисту в данной области техники. Для получения композиции вакцины в соответствии с данным изобретением используют фармацевтически приемлемые носители, разбавители или адъюванты, которые включают, но не ограничиваются следующим: адъюванты на основе минеральных солей (например, на основе алюминия, кальция, железа, циркония), поверхностно-активные адъюванты (например, Quil A, QS-21, другие сапонины), адъюванты, полученные из бактерий (например, N-ацетилмурамил-L-аланил-D-изоглутамин (MDP), липополисахариды (ЛПС), монофосфориллипид A, димиколат трегалозы (TDM), ДНК, CpGs, бактериальные токсины), эмульгирующие адъюванты (например, FIA, Montanide, адъювант 65, Lipovant), липосомные адъюванты, полимерные адъюванты и носители, цитокины (например, гранулоцитарно-моноцитарный колониестимулирующий фактор), углеводные адъюванты, живые системы доставки антигенов (например, бактерии, особенно модифицированные APP). Более того, носители также могут включать сухие составы, такие как покрытые пластыри, выполненные из титана или полимера. Способы получения составов и введения вакцин по данному изобретению можно также найти в "Remington, The Science and Practice of Pharmacy", 22-е издание.
Композиция вакцины в виде единичной композиции может содержать 0,01-2,0 мг белка, 0,01-2,0 мг нуклеиновой кислоты или 0,5-200 мг (или 1x104 КОЕ-1x1010КОЕ, колониеобразующих единиц) микроорганизмов. Необходимое активное количество белка, нуклеиновой кислоты или микроорганизма может определяться рутинными способами тестирования специалистом в данной области техники, например у свиней или поросят.
Композиция вакцины, содержащая по меньшей мере один полипептид ApxIA, ApxllA или ApxlllA, как описано выше, может представлять собой субъединичную вакцину. Субъединичная вакцина может содержать дополнительные полипептиды APP.
Также подразумевается, что вакцина содержит по существу только нуклеиновую кислоту или вектор, например, представляет собой ДНК вакцину. ДНК вакцины представляют собой третье поколение вакцин. Вакцины на основе нуклеиновых кислот или ДНК содержат ДНК/нуклеиновые кислоты, которые кодируют конкретные белки (ApxAs) от APP. ДНК/нуклеиновую кислоту вводят в организм млекопитающего, и она поглощается клетками, нормальные метаболические процессы которого синтезируют белки на основе генетического кода в нуклеиновой кислоте, которую они поглотили. Поскольку эти белки содержат области аминокислотных последовательностей, которые являются характерными для APP, они распознаются как чужеродные агенты, и когда они процессируются клетками-хозяевами и отображаются на их поверхности, иммунная система активизируется, что затем запускает иммунные ответы.
Альтернативно, нуклеиновая кислота может быть заключена в белок для облегчения вхождения в клетку. Если этот капсидный белок включен в нуклеиновую кислоту, полученная вакцина может объединять активность живой вакцины без рисков «обратного переключения».
Микроорганизмы, как описано выше, в композиции вакцины, могут представлять собой род Actinobacillus, например Actinobacillus pleuropneumoniae, Actinobacillus suis, штамм бактерий рода Actinobacillus, например штамм Actinobacillus pleuropneumoniae, Actinobacillus suis или конкретный серотип штамма бактерий рода Actinobacillus. Композиция вакцины может содержать по меньшей мере 1, например 1, 2, 3, 4, 5, 6 или 7 штаммов бактерий рода Actinobacillus или серотипов штаммов бактерий рода Actinobacillus. Каждый из серотипов, штаммов или бактерий рода Actinobacillus может кодировать по меньшей мере один полипептид, как определено выше.
Микроорганизм, как описано выше, в композиции вакцины, может не содержать нуклеиновые кислоты APP, кодирующие белки APP, которые отличаются от описанных модифицированных полипептидов ApxIA, ApxllA или ApxlllA. Например, микроорганизм может содержать делеции гена apxlV или/и гена sxy.
Кроме того, по меньшей мере один из Actinobacillus pleuropneumoniae в композиции вакцины может содержать одну или по меньшей мере две из следующих модификаций (например, единичная или множественные делеции): ΔtbpA, ΔtonB2, ΔsodC, ΔdsbA, Δfur, ΔmlcA, ΔmglA, ΔexbB, ΔureC, вариант с двумя мутациями ΔexbB ΔureC, ΔfhuA, ΔhlyX, вариант с двумя мутациями ΔapxIC и ΔapxllC, вариант с тремя мутациями ΔapxIC ΔapxllC Δorf1, вариант с шестью мутациями ΔapxllA ΔureC ΔdmsA ΔhybB ΔaspA Δfur, вариант с двумя мутациями ΔapxlllB ΔapxlllD, вариант с двумя мутациями ΔcIpP и ΔapxllC, ΔznuA, ΔapfA, вариант с двумя мутациями ΔapxllA и ΔureC, вариант с пятью мутациями ΔapxIC ΔapxllC Δorfl ΔcpxAR ΔarcA, вариант с двумя мутациями ΔapxIC ΔompP2, вариант с двумя мутациями ΔapxllC ΔapxIVA, инактивированный apxllC, инактивированный apxIC, Δlip40, ΔcpxA/cpxR, ΔpotD2, ΔtolC2, ΔsapA или ΔPdxS/PdxT.
Токсоидная форма ApxIA, а также композиция вакцины, содержащая по меньшей мере один полипептид ApxIA, ApxllA или ApxlllA, как раскрыто выше, демонстрирует значительно возросшую иммуногенность и профилактическую эффективность. Токсоидная форма ApxIA , а также композиция вакцины, содержащей по меньшей мере один полипептид ApxIA, ApxllA или ApxlllA, как описано выше, не демонстрирует цитотоксического эффекта и гемолитического эффекта и не вызывает значительного повышения температуры тела после иммунизации, которое может наблюдаться после традиционной иммунизации.
Вышеуказанные модификации описаны в:
Baltes, Nina; Hennig-Pauka, Isabel; Gerlach, Gerald-F (2002): Both transferrin binding proteins are virulence factors in Actinobacillus pleuropneumoniae serotype 7 infection. В FEMS microbiology letters 209 (2), pp. 283-287,
Sheehan, B. J.; Bosse, J. T.; Beddek, A. J.; Rycroft, A. N.; Kroll, J. S.; Langford, P. R. (2003): Identification of Actinobacillus pleuropneumoniae Genes Important for Survival during Infection in Its Natural Host. В Infection and immunity 71 (7), pp. 3960-3970. DOI: 10.1128/IAI.71.7.3960- 3970.2003,
Baltes, N.; Tonpitak, W.; Gerlach, G. F.; Hennig-Pauka, I.; Hoffmann-Moujahid, A.; Ganter, M.; Rothkötter, H. J. (2001): Actinobacillus pleuropneumoniae iron transport and urease activity. Effects on bacterial virulence and host immune response. В Infection and immunity 69 (1), pp. 472-478. DOI: 10.1128/IAI.69.1.472-478.2001,
Jacques, Mario (2004): Surface polysaccharides and iron-uptake systems of Actinobacillus pleuropneumoniae. В Canadian journal of veterinary research = Revue canadienne de recherche veterinaire 68 (2), pp. 81-85,
Baltes, Nina; N'diaye, Mohamed; Jacobsen, Ilse D.; Maas, Alexander; Buettner, Falk F. R.; Gerlach, Gerald-F (2005): Deletion of the anaerobic regulator HlyX causes reduced colonization and persistence of Actinobacillus pleuropneumoniae in the porcine respiratory tract. В Infection and immunity 73 (8), pp. 4614-4619. DOI: 10.1128/IAI.73.8.4614-4619.2005,
Lin, Liwen; Bei, Weicheng; Sha, Yonggang; Liu, Jinlin; Guo, Yi; Liu, Weihong et al. (2007): Construction and immunogencity of a DeltaapxIC/DeltaapxllC double mutant of Actinobacillus pleuropneumoniae serovar 1. В FEMS microbiology letters 274 (1), pp. 55-62. DOI: 10.1111/j.1574-6968.2007.00813.x,
Yuan, Fangyan; Liu, Jinlin; Guo, Yi; Tan, Chen; Fu, Shulin; Zhao, Jin et al. (2011): Influences of 0RF1 on the virulence and immunogenicity of Actinobacillus pleuropneumoniae. В Current microbiology 63 (6), pp. 574-580. DOI: 10.1007/s00284-011-0016-0,
Maas, Alexander; Jacobsen, Ilse D.; Meens, Jochen; Gerlach, Gerald-F (2006): Use of an Actinobacillus pleuropneumoniae multiple mutant as a vaccine that allows differentiation of vaccinated and infected animals. В Infection and immunity 74 (7), pp. 4124-4132. DOI: 10.1128/IAI.00133-06,
Park, Changbo; Ha, Yooncheol; Kim, Soohee; Chae, Chanhee; Ryu, Doug-Young (2009): Construction and characterization of an Actinobacillus pleuropneumoniae serotype 2 mutant lacking the Apx toxin secretion protein genes apxlllB and apxlllD. В The Journal of veterinary medical science 71 (10), pp. 1317-1323,
Xie, Fang; Li, Gang; Zhou, Long; Zhang, Yanhe; Cui, Ning; Liu, Siguo; Wang, Chunlai (2017): Attenuated Actinobacillus pleuropneumoniae double-deletion mutant S-8AclpP/apxllC confers protection against homologous or heterologous strain challenge. В BMC veterinary research 13 (1), p. 14. DOI: 10.1186/S12917-016-0928-9,
Yuan, Fangyan; Liao, Yonghong; You, Wujin; Liu, Zewen; Tan, Yongqiang; Zheng, Chengkun et al. (2014): Deletion of the znuA virulence factor attenuates Actinobacillus pleuropneumoniae and confers protection against homologous or heterologous strain challenge. В Veterinary microbiology 174 (3-4), pp. 531-539. DOI: 10.1016/j.vetmic.2014.10.016,
Zhou, Yang; Li, Lu; Chen, Zhaohui; Yuan, Hong; Chen, Huanchun; Zhou, Rui (2013): Adhesion protein ApfA of Actinobacillus pleuropneumoniae is required for pathogenesis and is a potential target for vaccine development. В Clinical and vaccine immunology: CVI 20 (2), pp. 287-294. DOI: 10.1128/CVI.00616-12,
Tonpitak, Walaiporn; Baltes, Nina; Hennig-Pauka, Isabel; Gerlach, Gerald F. (2002): Construction of an Actinobacillus pleuropneumoniae serotype 2 prototype live negative-marker vaccine. В Infection and immunity 70 (12), pp. 7120-7125,
Yuan, Fangyan; Liu, Jinlin; You, Wujin; Bei, Weicheng; Wang, Chunlai; Zhao, Jin et al. (2018): Generation, safety and immunogenicity of an Actinobacillus pleuropneumoniae quintuple deletion mutant SLW07 (ΔapxICΔapxllCΔorf1ΔcpxARΔarcA). В Vaccine 36 (14), pp. 1830-1836. DOI: 10.1016/j.vaccine.2018.02.083,
Liu, Qiong; Gong, Yuheng; Cao, Yuqin; Wen, Xintian; Huang, Xiaobo; Yan, Qigui et al. (2013): Construction and immunogenicity of a ΔapxlC/ompP2 mutant of Actinobacillus pleuropneumoniae and Haemophilus parasuis. В The Onderstepoort journal of veterinary research 80 (1), p. 519. DOI: 10.4102/ojvr.v80i1.519,
Liu, Jinlin; Chen, Xia; Lin, Liwen; Tan, Chen; Chen, Yan; Guo, Yi et al. (2007): Potential use an Actinobacillus pleuropneumoniae double mutant strain DeltaapxllCDeltaapxIVA as live vaccine that allows serological differentiation between vaccinated and infected animals. В Vaccine 25 (44), pp. 7696-7705. DOI: 10.1016/j.vaccine.2007.07.053,
Bei, Weicheng; He, Qigai; Yan, Lin; Fang, Liurong; Tan, Yadi; Xiao, Shaobo et al. (2005): Construction and characterization of a live, attenuated apxllCA inactivation mutant of Actinobacillus pleuropneumoniae lacking a drug resistance marker. В FEMS microbiology letters 243 (1), pp. 21-27. DOI: 10.1016/j.femsle.2004.11.033,
Xu, Fu-Zhou; Shi, Ai-Hua; Chen, Xiao-Ling; Yang, Bing; Wang, Jin-Luo (2007): Construction and immunogenicity of an attenuated mutant of Actinobacillus pleuropneumoniae by insertional inactivation of apxIC. В Wei sheng wu xue bao = Acta microbiologica Sinica 47 (5), pp. 923-927,
Prideaux, C. T.; Lenghaus, C.; Krywult, J.; Hodgson, A. L. (1999): Vaccination and protection of pigs against pleuropneumonia with a vaccine strain of Actinobacillus pleuropneumoniae produced by sitespecific mutagenesis of the Apxll operon. В Infection and immunity 67 (4), pp. 1962-1966,
Liu, 2018 Outer Membrane Lipoprotein Lip40 Modulates Adherence, Colonization, and Virulence of Actinobacillus pleuropneumoniae. Front Microbiol. 2018 Jul 3;9:1472. doi: 10.3389/fmicb. 2018.01472,
Li, 2018 The CpxA/CpxR Two-Component System Affects Biofilm Formation and Virulence in Actinobacillus pleuropneumoniae. Front Cell Infect Microbiol. 2018 Mar 20;8:72. doi: 10.3389/fcimb. 2018.00072,
Zhu 2017 Polyamine-binding protein PotD2 is required for stress tolerance and virulence in Actinobacillus pleuropneumoniae. Antonie Van Leeuwenhoek. 2017 Dec;110(12):1647-1657. doi: 10.1007/S10482-017-0914-7,
Li 2017 TolC2 is required for the resistance, colonization and virulence of Actinobacillus pleuropneumoniae. J Med Microbiol. 2017 Jul 31. doi: 10.1099/jmm.0.000544,
Xie 2017 The SapA Protein Is Involved in Resistance to Antimicrobial Peptide PR-39 and Virulence of Actinobacillus pleuropneumoniae. Front Microbiol. 2017 May 10;8:811. doi: 10.3389/fmicb.2017.00811, и
Xie 2017 Pyridoxal phosphate synthases PdxS/PdxT are required for Actinobacillus pleuropneumoniae viability, stress tolerance and virulence. PLoS One. 2017 Apr 27;12(4):e0176374. doi: 10.1371/journal.pone.0176374.
Ожидается, что комбинация модифицированных полипептидов, как описано в данном документе, с дополнительными известными модификациями, перечисленными выше, будет приводить к синергетическому ослаблению Actinobacillus pleuropneumoniae.
МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ ИЛИ СПОСОБ
Описанную композицию вакцины можно использовать при профилактике, метафилактике или лечении пневмонии, плеврита или плевропневмонии, в частности пневмонии, плеврита или плевропневмонии, вызванных Actinobacillus pleuropneumoniae у субъекта. Субъект, подлежащий лечению, представляет собой млекопитающее, в частности свинью. Композицию вакцины можно вводить внутримышечно, внутрикожно, внутривенно, подкожно или нанесением на слизистую (например, интраназально).
Композицию вакцины можно вводить путем по меньшей мере одного, например, одного или двух введений с использованием единичной композиции, как описано выше. В частности, композицию можно вводить в первый раз в день рождения, через 3 дня, через 1 неделю, 2 недели, 4 недели, 6 недель, 8 недель, 10 недель или 12 недель после рождения субъекта. Следовательно, вакцину предпочтительно можно вводить в ранний момент жизни млекопитающего. Однако, также предполагается, что вакцину можно вводить в любой момент жизни млекопитающего.
Композицию вакцины можно вводить во второй раз, при этом период времени между двумя введениями может составлять от 1 до 4 недель, от 1 до 3 недель или от 1 до 2 недель. Композицию вакцины, содержащей микроорганизм, предпочтительно вводить только один раз.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1 иллюстрирует осуществление анализа цитотоксичности с использованием клеток BL-3 и токсинов ApxA, выделенных из супернатантов APP.
Фигура 2 иллюстрирует цитотоксический анализ с клетками BL-3 и генетически модифицированным токсином ApxIA и токсином ApxIA дикого типа (A) и генетически модифицированным токсином ApxlllA и токсином ApxlllA дикого типа (B).
Фигура 3 иллюстрирует осуществление анализа на гемолиз с использованием крови овцы и токсинов ApxA, выделенных из супернатантов APP.
Фигура 4 иллюстрирует анализ на гемолиз с использованием крови овцы и генетически модифицированного токсина ApxIA и токсина ApxIA дикого типа.
Фигура 5: ректальная температура тела через 6 часов после первой вакцинации (A) и второй вакцинации (B). Каждая точка представляет ректальную температуру тела отдельного животного. Столбцы представляют медианы с 95%-ым доверительным интервалом. Критерий множественных сравнений Данна; * = p < 0,05; ** = p < 0,01.
Фигура 6: Количественное определение IgG, специфичных к ApxIA, ApxllA и ApxIIIA, в сыворотке свиней. IgG, связывающийся с различными антигенами, количественно определяли с использованием ELISA. Анализировали сыворотку свиней из различных групп до вакцинации (д0), через 2 недели после первой вакцинации (д14) и через 2 недели после второй вакцинации (д28). Рассчитанные средние значения IgG для групп для каждого момента времени сравнивали с соответствующим моментом времени для группы 1. Критерий множественных сравнений Данна; * = p < 0,05; ** = p < 0,01. Столбцы представляют медианы. Отн. ед.: относительные единицы.
Фигура 7: Анализ сывороточной нейтрализации с использованием 6 пг активного токсина ApxIA и сыворотки от свиней через две недели после второй вакцинации. Жизнеспособность клеток оценивали с использованием WST-1, который метаболизировался метаболически активными клетками. Легенда отображает количества животных, а в скобках данные ELISA относительно IgG, специфичного к ApxlA, (A). X-величины (концентрации сыворотки) ymax/2 рассчитывали с помощью криволинейных регрессионных кривых (B). Критерий множественных сравнений Данна; ** = p < 0,01; n.s. = не значимо. Столбцы представляют медианы. Отн. ед.: относительные единицы.
Фигура 8: Суммарные клинические показатели после заражения сероваром 9 APP в трех группах, получающих «вакцину 1» (группа 1), Porcilis® APP (группа 2) или плацебо (группа 3). Суммарные клинические показатели сравнивали с соответствующим клиническим показателем группы 3. Критерий множественных сравнений Данна; * = p < 0,05; ** = p < 0,01. Столбцы представляют медианы.
Фигура 9: Выживаемость свиней после заражения сероваром 9 APP в трех группах, получающих «вакцину 1» (группа 1), Porcilis® APP (группа 2) или плацебо (группа 3). Выживаемость в процентах представляет собой 100% для группы 1 и группы 2, тогда как 37,5% животных в группе 3 умерли или были умерщвлены в соответствии с требованиями содержания животных из-за тяжелых симптомов, вызванных APP.
Фигура 10: Показатели поражения легких после заражения сероваром 9 APP в трех группах, получающих «вакцину 1» (группа 1), Porcilis® APP (группа 2) или плацебо (группа 3). Показатели поражения легких группы 1 и группы 2 сравнивали с соответствующим показателем поражения легких контрольной группы (группа 3). Критерий множественных сравнений Данна; * = p < 0,05; ** = p < 0,01. Столбцы представляют медианы.
ПРИМЕРЫ
В следующих примерах представлены экспериментальные данные, которые объясняют данное изобретение, и их необходимо понимать только в качестве иллюстрации изобретения. В частности, подразумевается, что каждый конкретный признак, раскрытый в примерах, можно объединить с признаками предшествующей части описания.
ПРИМЕР 1: КЛОНИРОВАНИЕ APXIA, APXIIA ИЛИ APXIIIA ДИКОГО ТИПА ИЛИ МОДИФИЦИРОВАННЫХ В PEX-A258
Последовательности белков ApxlA-ApxlllA дикого типа получали из базы данных Uniprot, т.е. ST5b ApxIA APP (Uniprot: A3N292), ST3 ApxllA APP (Uniprot BOBPPO, с модификацией S148G) и ST8 ApxlllA (Uniprot: P55131).
Кроме того, были получены последовательности белков, содержащие следующие модификации:
Таблица 1
Нуклеиновые кислоты были получены из белковых последовательностей, при этом последовательности нуклеиновых кислот были кодон-оптимизированы и синтетически получены (Eurofins Genomics) без инициирующего кодона. Каждая последовательность нуклеиновой кислоты была перенесена в плазмиду pEX-A258 (SEQ ID №: 19), фланкированную сайтами рестрикции BamHI на 5'-конце и Sacl на 3'-конце.
ПРИМЕР 2: КЛОНИРОВАНИЕ И ЭКСПРЕССИЯ APXIA-APXIIIA ДИКОГО ТИПА И МОДИФИЦИРОВАННЫХ В PQE-80L (SEQ ID №: 20)
1 мкг pEX-A258, содержащего соответствующие ApxlA-ApxlllA, рестриктировали BamHI FD (ThermoFisher) и Sacl FD (ThermoFisher) в соответствии с инструкциями производителя и разделяли на 1% агарозном геле. Фрагменты нуклеиновой кислоты соответствующего размера экстрагировали из агарозного геля (QUIAEX-II). pQE-80L рестриктировали BamHI FD (ThermoFisher) и Sacl FD (ThermoFisher) в соответствии с инструкциями производителя. Лигирование pQE-80L и соответствующего фрагмента нуклеиновой кислоты в соотношении 1:5 выполняли в течение ночи при комнатной температуре с использованием T4-ДНК-лигазы (ThermoFisher). Лигированную ДНК электропорировали в Topf10F’ E. Coli и подтверждали успешное клонирование форм дикого типа и модифицированных форм ApxlA-ApxlllA с помощью рестрикционного анализа и секвенирования. pQE-80L включает устойчивость к ампициллину.
ПРИМЕР 3: КЛОНИРОВАНИЕ ГЕНОВ APXIC, APXIIC И APXIIIC В PQE-60
Гены apxIC-IIIC амплифицировали из APP-ДНК посредством полимеразной цепной реакции (ПЦР) с использованием праймеров, указанных в таблице 2, а затем очищали.
Фрагменты ПЦР, содержащие соответствующие нуклеиновые кислоты apxIC-IIIC и pQE-60 рестриктировали с помощью эндонуклеаз рестрикции, указанных в таблице 2 ниже, в соответствии с инструкциями соответствующего производителя и разделяли на 1% агарозном геле.
Таблица 2
Фрагменты нуклеиновой кислоты подходящего размера экстрагировали из агарозного геля (QUIAEX-II). Лигирование pQE-60 и соответствующего фрагмента нуклеиновой кислоты в соотношении 1:5 выполняли в течение ночи при комнатной температуре с использованием T4-ДНК-лигазы (ThermoFisher).
Лигированную ДНК электропорировали в Topf10F’ E. coli и подтверждали успешное клонирование с помощью рестрикционного анализа и секвенирования.
ПРИМЕР 4: КЛОНИРОВАНИЕ И ЭКСПРЕССИЯ ГЕНОВ APXIC, APXIIC И APXIIIC В PACYC184
Соответствующие гены apxIC, apxllC и apxlllC амплифицировали из соответствующих векторов pQE-60 примера 2 с использованием праймеров, указанных в таблице 3 ниже.
Таблица 3
Clal/Bscl и Nrul (apxllC)
Таким путем промотор/lac-оператор и сайты связывания рибосомы также содержались в ампликонах (SEQ ID №: 16, 17 и 18).
Соответствующие продукты ПЦР были ограничены с использованием эндонуклеаз рестрикции, указанных в таблице 3 и клонированных в pACYC184 посредством сайтов рестрикции Xbal и Nrul (apxIC и apxlllC) или Clal/Bscl и Nrul (apxllC) с использованием того же способа и условий, как указано выше. pACYC184 включает устойчивость к хлорамфениколу.
Лигированную ДНК электропорировали в Topf10F’ E. coli и подтверждали успешное клонирование с помощью рестрикционного анализа и секвенирования.
ПРИМЕР 5: КОТРАНСФОРМАЦИЯ PACYC184, СОДЕРЖАЩЕГО ГЕНЫ APXIC, APXIIC ИЛИ APXIIIC, И PQE-80L, СОДЕРЖАЩЕГО APXIA, APXIIA ИЛИ APXIIIA
ApxC необходим для активации токсинов ApxA. Следовательно, pACYC184, содержащий соответствующие гены apxIC, apxllC или apxlllC, полученный в примере 4, котрансформировали с pQE-80L, содержащим соответствующий ген apxIA, apxllA или apxlllA, в TopflOF’ E. Coli, и успешную котрансформацию выбирали на агаровых пластинах с LB, содержащих 50 мкг/мл ампициллина и 15 мкг/мл хлорамфеникола. Успешную трансформацию подтверждали рестрикционным анализом.
ПРИМЕР 6: ЭКСПРЕССИЯ И ВЫДЕЛЕНИЕ ТОКСИНОВ И ТОКСОИДОВ APXA, В ЧАСТНОСТИ APXIA
Колонию E. Coli,, полученную Примере 5 и содержащую pACYC184 и pQE-80L, кодирующие соответствующие гены ApxA и ApxC, использовали для инокуляции 40 мл среды LB (100 мкг/мл ампициллина и 30 мкг/мл хлорамфеникола). Культуру инкубировали при встряхивании при 30°C в течение ночи. На следующий день ночную культуру инокулировали в 400 мл среды LB (100 мкг/мл ампициллина и 30 мкг/мл хлорамфеникола), так что была достигнута оптическая плотность 0,1 (оптическая плотность при 600 нм = OD600). Полученную культуру инкубировали при встряхивании до получения OD600, составляющей 0,5. Для индуцирования экспрессии был добавлен рекомбинантный белок 1 мМ IPTG (изопропил-β-D-1-тиогалактопиранозид). Культуру инкубировали при встряхивании при 30°C в течение еще 4 часов. Затем культуру центрифугировали при 7000g в течение 10 минут. После удаления супернатанта осадок бактерий ресуспендировали в буфере LEW (50 мМ NaH2PO4, 300 мМ NaCI в дистиллированной воде; pH 8,0), 5 мл буфера LEW на 1 г осадка бактерий, и хранили при -20°C.
К 10 мл ресуспендированного осадка бактерий добавляли 1 мл 10x Bug Buster (Novagen), 5 мкл бензоазы (25 ед./мкл, Novagen) и 10 мг лизоцима. Полученную суспензию медленно вращали при комнатной температуре в течение 20 минут. После центрифугирования при 4°C и 15000 g в течение 60 минут осадок промывали буфером LEW (10 мл буфера LEW на 1 мг осадка) и центрифугировали (4 °C, 15000 g, 60 мин). Промытый осадок ресуспендировали в 2 мл денатурирующего солюбилизирующего буфера (8 M мочевина в буфере LEW; pH=8,0) на 1 г осадка, а затем в 1 мл Bug Buster (Novagen) на 10 мл ресуспендированного осадка. Суспензию инкубировали при 4°C в течение 1 часа, а затем центрифугировали (4 °C, 18000 g, 15 мин). Супернатант, содержащий белок, использовали на следующих стадиях для колоночной очистки белка.
В соответствии с инструкциями производителя колонки (колонки Protino Ni-IDA 2000, Machery-Nagel) уравновешивали 4 мл равновесного буфера (8 M мочевины в буфере LEW; pH=8,0). Затем в колонку загружали половину содержащего белок супернатанта и дважды промывали 4 мл равновесного буфера (всего 8 мл буфера). Белки дважды элюировали 3 мл элюирующего буфера (равновесный буфер с 250 мМ имидазола, всего 6 мл буфера). Процедуру повторяли со второй половиной содержащего белок супернатанта. Элюаты, содержащие белок, объединяли и диализировали в ФСБ, pH 8,0, содержащем 1 M мочевины при 4°C в течение ночи. Диализный буфер заменяли на 0,5 М мочевины в ФСБ (pH 8,0), и диализ продолжали еще 4 часа. Затем буфер заменяли на ФСБ (pH 8,0) без мочевины и диализировали еще 4 часа. Концентрацию белка определяли с помощью набора для анализа белков Pierce BCA (ThermoFisher) в соответствии с инструкциями производителя. Белок хранили при -80°C до использования.
ПРИМЕР 7: ВЫДЕЛЕНИЕ ТОКСИНОВ APXA ИЗ APP
Раствор APPVX содержит 1 г D(+)-глюкозы, моногидрата, 0,1 г L-глутамина, 0,362 г L-цистеина гидрохлорида моногидрата и 10 мл безнуклеазной H2O. Изоляты APP высевали на шоколадный агар, содержащий IsovitaleX, и инкубировали при 37°C в течение ночи. Три колонии APP объединяли и использовали, чтобы инокулировать 3 мл колумбийской среды (10 мМ Ca2+, 0,2 мг/мл NAD, 1 % APPVX), и полученную культуру инкубировали при 37°C при встряхивании в течение 5 часов. Затем культуру центрифугировали при 3200g и 4°C в течение 20 минут. Прозрачный супернатант переносили в новую пробирку и постепенно добавляли 44,05 г сульфата аммония и перемешивали до полного растворения сульфата аммония. Затем раствор инкубировали при 4°C в течение ночи. Осажденные белки центрифугировали при 3200 g и 4°C в течение 3 часов, и супернатант удаляли. Белковый осадок сушили на воздухе, а затем суспендировали в 1 мл ФСБ, содержащего 6 М мочевину. Для удаления остаточного буфера полученный раствор белка очищали с помощью гель-фильтрации на колонках PDMidiTrap G25 (GE Healthcare). После уравновешивания ФСБ, содержащим 6 М мочевину, 1 мл раствора белка загружали в гель-фильтрационную колонку, а затем элюировали 1,5 мл ФСБ, содержащего 6 М мочевины. Затем элюированные белки хранили при -20°C. Идентификацию элюированных белков проводили с помощью SDS-PAGE.
ПРИМЕР 8: ПРОВЕДЕНИЕ АНАЛИЗА НА ЦИТОТОКСИЧНОСТЬ
Для определения цитотоксической активности рекомбинантных белков ApxA (дикого типа и токсоидных форм) проводили анализ на цитотоксичность с использованием клеток BL3 (BL3.1 CRL-2306) в качестве клеток-мишеней. Клетки BL3 получали из бычьих B-клеток, и известно, что они могут быть лизированы токсинами ApxA (Tu, A. H.; Hausler, C.; Young, R.; Struck, D. K. (1994): Differential expression of the cytotoxic and hemolytic activities of the ApxllA toxin from Actinobacillus pleuropneumoniae. В Infection and immunity 62 (5), pp. 2119-2121). Токсины ApxA, полученные в Примере 7, использовали в качестве положительного контроля, а ФСБ, содержащий 6 М мочевины, служил в качестве отрицательного контроля.
Клетки BL3 культивировали в среде (DMEM, содержащей 100 ед./мл пенициллина, 100 мкг/мл стрептомицина, 10 % FCS и заменимые аминокислоты, 1x) при 37 °C и 5 % CO2. Для культивирования использовали колбы T75 с 25 мл среды. Клетки BL3 пересевали раз в неделю, при этом 50000 клеток/см2 использовали для засевания в новую колбу.
Для применения в цитотоксическом анализе 50 мкл клеток BL3 в среде переносили в лунку 96-луночного планшета. Рекомбинантные белки и контроли разбавляли в DMEM. 50 мкл разбавленных рекомбинантных белков и контролей добавляли в каждую лунку, содержащую клетки. Затем планшет инкубировали при 37 °C и 5% CO2 в течение 24 часов. После оценки морфологии клеток под микроскопом в каждую лунку добавляли 10 мкл WST-1 (Sigma Aldrich), и планшет инкубировали при 37°C и 5% CO2 в течение еще 24 часов.
Затем определяют поглощение с помощью ридера ELISA при 450 нм. WST-1 расщепляется посредством сложного клеточного механизма на клеточной поверхности жизнеспособных клеток до растворимого формазана. Следовательно, количество образовавшегося красителя формазана и величина экстинкции, полученная на ридере для ELISA, напрямую коррелируют с количеством метаболически активных клеток в культуре.
ПРИМЕР 9: ПРОВЕДЕНИЕ АНАЛИЗА НА ГЕМОЛИЗ
Для определения гемолитической активности рекомбинантных белков ApxA (дикого типа и токсоидных форм) осуществляли анализ на гемолиз. Использовали планшеты для культивирования клеток, содержащие колумбийский агар с 5% овечьей крови (Bection Dickinson). В планшетах для культивирования клеток вырезали отверстия диаметром 3 мм. 10 мкл соответствующих образцов рекомбинантного белка и контроля наливали в отверстие, и планшеты инкубировали при 37°C в течение ночи. Токсины ApxA, полученные в Примере 7, использовали в качестве положительного контроля, а ФСБ, содержащий 6 М мочевины, служил в качестве отрицательного контроля.
Считают, что гемолитическая активность присутствует, когда агар с овечьей кровью обесцвечивался и таким образом становился беловатым.
ПРИМЕР 10: ВЫДЕЛЕНИЕ ТОКСИНОВ APXA ИЗ APP
Штаммы APP ST2, ST5, ST7 и ST8 использовали для выделения супернатантов, содержащих токсины ApxA, с использованием методов экспрессии и выделения, описанных в примере 7. С помощью SDS-PAGE было установлено, что супернатанты содержали полосы при ожидаемых величинах молекулярной массы, соответствующих ApxIA, ApxllA, ApxlllA и ApxIVA (данные не показаны). Таким образом, каждый из полученных супернатантов соответствующих штаммов ST2, ST5, ST7 и ST8 содержит смесь ApxIA, ApxllA, ApxlllA и ApxIVA.
ПРИМЕР 11: ЦИТОТОКСИЧНОСТЬ APXA из APP В КЛЕТКАХ BL-3
Используя способ из примера 8, 50 мкл токсинов ApxA примера 7, полученных из APP (штаммы APP ST2, ST5, ST7 и ST8), тестировали при возрастающих разбавлениях в DMEM (1:8, 1:16, 1:32; 1:64, 1:128; 1:256, 1:512; 1:1024, 1:2048, 1:4096).
Наблюдали цитотоксический эффект супернатантов APP, который уменьшался с увеличением разбавления (см. фигуру 1). Выделенные токсины ST8 продемонстрировали наиболее высокую цитотоксичность, тогда как выделенные токсины ST7 показали наиболее низкую токсичность. Не наблюдалось никакого влияния ЛПС в препарате ApxA на цитотоксичность (данные не показаны).
ПРИМЕР 12: ЦИТОТОКСИЧНОСТЬ РЕКОМБИНАНТНЫХ ФОРМ APXIA И APXIIIA В КЛЕТКАХ BL-3
С использованием способа из примера 8 были протестированы следующие образцы.
Таблица 4
50 мкл каждого образца тестировали при возрастающих разбавлениях в DMEM (1:2, 1:4, 1:8, 1:16, 1:32; 1:64, 1:128; 1:256, 1:512).
В контроле с клетками использовали разбавления ФСБ, чтобы оценить влияние ФСБ. Не наблюдали никакого эффекта (см. фигуру 2). Результаты показаны на фигуре 2.
Как проиллюстрировано на фигуре 2A, анализ цитотоксичности продемонстрировал, что рекомбинантный ApxIA дикого типа лизировал клетки BL3 и таким образом являлся активным. Начиная с разбавления 1:64 (13,1 мкг/мл токсина ApxIA дикого типа), величины жизнеспособности клеток были сопоставимы со значениями для контроля ФСБ. Жизнеспособность клеток BL3, которые инкубировали с токсоидной формой ApxIA, была сопоставима со значениями для контроля ФСБ. Это также применимо к морфологии клеток (данные не показаны).
Поскольку токсоидная форма ApxIA имеет точно такое же влияние на жизнеспособность и морфологию клеток, что и контроль ФСБ, тогда как форма ApxIA дикого типа обладает сильным цитотоксическим эффектом на клетки BL3, можно сделать вывод, что токсоидная форма ApxIA не обладает цитотоксическим эффектом. Подобные результаты были получены для токсоидных форм ApxllA (данные не показаны) и ApxlllA (фигура 2 B).
ПРИМЕР 13: ГЕМОЛИТИЧЕСКАЯ АКТИВНОСТЬ APXA ИЗ APP
С использованием способа примера 9, были протестированы 10 мкл токсинов ApxA из примера 7, полученных из APP (штаммы ST2, ST5 и ST8 APP).
Результаты показаны на фигуре 3. Штамм ST8 продемонстрировал сильный гемолиз, ST5 - гемолиз средней степени, и ST2 - не продемонстрировал гемолиза, как можно заключить из снижения интенсивности белого цвета слева направо на фигуре 3. Известно, что не каждый из ApxIA, ApxllA и ApxllA способен лизировать эритроциты. Поскольку различные штаммы секретируют различные количества соответствующих форм ApxA, супернатанты из соответствующих штаммов, которые содержат смеси различных форм ApxA могут демонстрировать различную гемолитическую активность.
В целом, можно продемонстрировать, что гемолитическая активность токсинов ApxA может быть выявлена в способе, включающем применение колумбийского агара, содержащего 5% овечьей крови, как описано в примере 9.
ПРИМЕР 14: ГЕМОЛИТИЧЕСКАЯ АКТИВНОСТЬ РЕКОМБИНАНТНЫХ ФОРМ APXIA
С использованием способа из примера 9 были протестированы следующие образцы.
Таблица 5
Результаты показаны на фигуре 4. Тогда как форма ApxIA дикого типа лизирует эритроциты, токсоидная форма ApxIA не лизирует эритроциты. Форма ApxlllA дикого типа не демонстрирует гемолитической активности, что также описано в литературе.
Поскольку токсоидная форма ApxIA имела точно такой же, т.е. отсутствие, гемолитический эффект, что и контроль ФСБ, тогда как форма ApxIA дикого типа обладала сильным гемолитическим эффектом на эритроциты овцы, можно сделать вывод, что токсоидная форма ApxIA не обладает гемолитическим эффектом. Подобные результаты были получены для токсоидных форм ApxllA (данные не показаны).
ПРИМЕР 15: ИММУНОГЕННОСТЬ ПОЛИПЕПТИДОВ APXIA, APXIIA И APXIIIA ПО ИЗОБРЕТЕНИЮ У СВИНЕЙ
(1) Получение антигенов и составов вакцин
Нуклеиновые кислоты, кодирующие модифицированные ApxIA (SEQ ID №: 4), ApxllA (SEQ ID №: 5) и ApxlllA (SEQ ID №: 6), кодон-оптимизированные для экспрессии в E. coli, были получены синтетически (Eurofins Genomics), и последовательность введена в плазмиду pEX-A258 (SEQ ID №: 19), фланкированную сайтами рестрикции BamHI на 5'-конце и Sacl на 3'-конце. 1 мкг pEX-A258, содержащего соответствующие ApxlA и ApxlllA, рестриктировали BamHI FD (ThermoFisher) и Sacl FD (ThermoFisher) в соответствии с инструкциями. Нуклеотиды, кодирующие AA 439-801 ApxllA, амплифицировали с помощью ПЦР, используя pEX-A258 (SEQ ID №: 19), содержащий ген для ApxllA и праймеры (прямой: GCAGGATCCCAGGGGTATGATTCACGC и обратный GGTGAGCTCTTAGGTGATGGAGTCGTTGCC), вводящие сайт рестрикции BamHI на 5'- и Sacl на 3'-конце. Продукт ПЦР очищали с использованием набора для очистки ПЦР QIAquick 250 (Qiagen) и рестриктировали с помощью BamHI FD (ThermoFisher) и Sacl FD (ThermoFisher) в соответствии с инструкциями производителя. Рестриктированные ApxIA, ApxllA (усеченный) и ApxlllA разделяли в 1% агарозном геле. Фрагменты нуклеиновой кислоты соответствующего размера экстрагировали из агарозного геля (QUIAEX-II). pQE-80L рестриктировали с помощью BamHI FD (ThermoFisher) и Sacl FD 10 (ThermoFisher) в соответствии с инструкциями производителя. Лигирование pQE-80L и соответствующего фрагмента нуклеиновой кислоты в соотношении 1:5 выполняли в течение ночи при комнатной температуре с использованием T4-ДНК-лигазы (ThermoFisher). Лигированную ДНК электропорировали в TopflOF’ E. Coli и подтверждали успешное клонирование модифицированных форм ApxIA, ApxllA (усеченный) или ApxlllA с помощью рестрикционного анализа и секвенирования. pQE-80L включает устойчивость к ампициллину. Соответствующий pQE-80L, кодирующий модифицированный ApxIA, ApxllA (усеченный) или ApxlllA, котрансформировали в TopflOF’ E. Coli с плазмидой pACYC184, кодирующей соответствующий apxIC, apxllC и apxlllC, полученной в Примере 4, в соответствии со способом, описанным в Примере 5. После котрансформации TopflOF’ E. Coli селектировали на агаровых пластинках с LB, содержащих 50 мкг/мл ампициллина и 15 мкг/мл хлорамфеникола. Полученные колонии E. Coli, содержащие:
- PQE-80L, кодирующий модифицированный ApxIA, и pACYC184, кодирующий ApxIC
- PQE-80L, кодирующий усеченный ApxllA (усеченный), и pACYC184, кодирующий ApxllC
- PQE-80L, кодирующий модифицированный ApxlllA, и pACYC184, кодирующий ApxlllA
использовали для инокуляции 40 мл среды LB (дополненной 100 мкг/мл ампициллина, 30 мкг/мл хлорамфеникола). Культуру инкубировали при встряхивании при 30°C в течение ночи. Ночную культуру инокулировали в 400 мл среды LB (дополненная 100 мкг/мл ампициллина, 30 мкг/мл хлорамфеникола), так что была достигнута оптическая плотность 0,1 (оптическая плотность при 600 нм = OD600). Полученную культуру дополнительно инкубировали при встряхивании до получения OD600, составляющей 0,5. Для индуцирования экспрессии был добавлен рекомбинантный белок 1 мМ IPTG (изопропил-β-D-1-тиогалактопиранозид). Культуру инкубировали при встряхивании при 30°C в течение еще 4 часов. Затем культуру центрифугировали при 7000g в течение 10 минут. После удаления супернатанта осадок бактерий ресуспендировали в буфере LEW (50 мМ NaH2PO4, 300 мМ NaCI в дистиллированной воде; pH 8,0), 5 мл буфера LEW на 1 г осадка бактерий, и хранили при -20°C. К 10 мл ресуспендированного осадка бактерий добавляли 1 мл 10x Bug Buster (Novagen), 5 мкл бензоазы (25 ед./мкл, Novagen) и 10 мг лизоцима. Полученную суспензию медленно вращали при комнатной температуре в течение 20 минут. После центрифугирования при 4°C и 15000 g в течение 60 минут осадок промывали буфером LEW (10 мл буфера LEW на 1 мг осадка) и центрифугировали (4 °C, 15000 g, 60 мин). Промытый осадок ресуспендировали в 2 мл денатурирующего солюбилизирующего буфера (8 M мочевина в буфере LEW; pH=8,0) на 1 г осадка, а затем в 1 мл Bug Buster (Novagen) на 10 мл ресуспендированного осадка. Суспензию инкубировали при 4°C в течение 1 часа, а затем центрифугировали (4 °C, 18000 g, 15 мин). Супернатант, содержащий белок, использовали на следующих стадиях для колоночной очистки белка. В соответствии с инструкциями производителя колонки (колонки Protino Ni-IDA 2000, Machery-Nagel) уравновешивали 4 мл равновесного буфера (8 M мочевины в буфере LEW; pH=8,0). Затем в колонку загружали половину содержащего белок супернатанта и дважды промывали 4 мл равновесного буфера (всего 8 мл буфера). Белки дважды элюировали 3 мл элюирующего буфера (равновесный буфер с 250 мМ имидазола, всего 6 мл буфера). Процедуру повторяли со второй половиной содержащего белок супернатанта. Фракции элюатов, содержащих белок, объединяли и диализировали в ФСБ, pH 8,0, содержащем 1 M мочевины при 4°C в течение ночи. Диализный буфер заменяли на 0,5 М мочевины в ФСБ (pH 8,0), и диализ продолжали еще 4 часа. Затем буфер заменяли на ФСБ (pH 8,0) без мочевины и диализировали еще 4 часа. Концентрацию белка определяли с помощью набора для анализа белков Pierce BCA (ThermoFisher) в соответствии с инструкциями производителя. Соответствующие антигены ApxA фильтровали с помощью стерилизующего фильтрования (0,2 мкм) и хранили при 4°C до включения в состав.
Montanide ISA 201 VG (серия 36075M, Seppic) использовали в соответствии с инструкцией производителя для получения тестовых партий с соответствующими модифицированными ApxIA, ApxllA (усеченный) и ApxlllA (Таблица 6). Вкратце, адъювант и антиген предварительно нагревали до 31°C на водяной бане. Объем антигена(-ов) делали равным объему адъюванта (50:50, мас./об.) путем добавления стерильного ФСБ и дополнительно выдерживали при 31°C. Montanide ISA 201 VG медленно перемешивали в стакане. Затем антиген добавляли к адъюванту и дополнительно перемешивали при 31°C в течение 5 мин. Состав хранили в течение 1 ч при 20°C в стеклянных флаконах и далее хранили при 4°C до использования.
Таблица 6: Состав различных тестовых партий для изучения иммуногенности
(2) Исследование на животных (иммуногенность)
Иммуногенность составов вакцин определяли путем количественного определения антител IgG, специфичных к ApxIA, ApxllA и ApxlIIA, в ELISA и оценки ApxlA-нейтрализующих свойств в сыворотке крови после вакцинации свиней возрастом 8 недель.
Животные
Свиней обоих полов (самцы [кастрированные] и самки) приобретали у животновода, разводящего свиней (BHZP, 21337 Люнебург, Zuchtbetrieb Garlitz (DE-LWL 1051)). Каждую свинью идентифицировали с помощью уникального идентификационного номера на ухе и подвергали ветеринарному осмотру относительно соответствия критериям включения/исключения, указанным в протоколе исследования. Все животные не демонстрировали клинических признаков инфекции APP, и у животных не было клинических признаков (одышка, кашель, диарея, хромота) в начале исследования. Было подтверждено, что животные являлись сероотрицательными, что установлено с помощью коммерчески доступного анализа ELISA APP-ApxlV (IDEXX) до включения в исследование. Свиней содержали на огороженной территории животноводческого хозяйства IDT Biologika GmbH, Am Pharmapark, D-06861 Дессау-Рослау (Германия). В каждом блоке было две отдельных комнаты/площадки одинакового размера на сплошном настиле. В качестве подстилки использовали солому, которую меняли раз в день. Свиней кормили (пеллетами, продажная смесь) в необходимом количестве с использованием вручную заполненных кормушек для сухих кормов. Воду качества водопроводной подавали с помощью крановой и сосковой поилок. Все помещения убирали раз в день.
Дизайн исследования
В общем 25 свиней в возрасте 8 недель случайным образом распределяли в 5 групп, содержащих 5 животных на группу. Трем группам внутримышечно вводили 2 раза с интервалом 14 дней 1,5 мл «вакцины 1», «вакцины 2» или «вакцины 3». Одна группа из пяти животных получала 2 мл коммерчески доступной вакцины против Actinobacillus Pleuropneumoniae (Porcilis® APP, MSD Animal Health, партия: A784A01) дважды с интервалом 14 дней и служила положительным контролем. Другая группа получала вакцину-плацебо и служила отрицательным контролем. Обзор групп и типов воздействия представлен в Таблице 7.
Таблица 7: Группа исследования и воздействие
* свинья под номером 5670 (группа 4) была исключена из исследования в 14 день исследования вследствие инфекции, не связанной с тестируемыми вакцинами.
Образцы сыворотки собирали в 0 день исследования (день первой вакцинации), 14 (день второй вакцинации) и 28 (конец исследования) от каждого из животных.
Местные и системные реакции, связанные с вакцинацией, оценивали один раз на день, начиная с 1 дня до и заканчивая 3 днями после каждой из двух вакцинаций. Для оценки местных реакций оценивали покраснение и отек мест инъекции.
Ректальную температуру тела измеряли один раз на день, начиная с 1 дня до и заканчивая 3 днями после каждой из двух вакцинаций. В день вакцинации температуру измеряли до и через 6 ч после вакцинации. Результаты относительно температуры тела показаны в таблице 8 и на фигуре 5.
Результаты:
Таблица 8: Средняя ректальная температура тела на группу (возрастание температуры обозначено)
(3) Серологические данные (ELISA)
Анализы ELISA осуществляли для количественного определения антител IgG, специфичных к ApxIA, ApxllA или ApxllIA, в сыворотке свиней. Вкратце, ApxIA, ApxllA или ApxlllA экспрессировали, очищали и количественно определяли, как описано в Примере 5. Соответствующий ApxA разбавляли в покрывающем буфере lxELISA (120125; Candor Bioscience) до конечной концентрации 5 мкг/мл. 100 мкл соответствующего ApxA в покрывающем буфере ELISA помещают в каждую лунку 96-луночного планшета (планшет для ELISA Nunc MaxiSorb (44-2404-21, ThermoFisher)) и инкубируют при 4°C в течение ночи. Затем покрывающий буфер удаляют и планшеты 3 раза промывают промывающим буфером (ФСБ, 0,05% твина 20 (об./об.)). Планшеты блокируют с использованием блокирующего раствора, содержащего ФСБ с 10% (об./об.) FCS, в течение 1 ч при комнатной температуре (к.т.) на шейкере. Затем блокирующий раствор удаляют, а все лунки промывают 3 раза промывающим буфером. Сыворотку свиней предварительно разбавляют 1:100 блокирующим раствором, и готовят ряд разбавлений 1:5 в блокирующем растворе (включая разбавления 1:100-1:312500). 50 мкл каждого разбавления пипеткой переносят в одну из лунок планшета для ELISA. Гипериммунизированная сыворотка, полученная от животного, инфицированного A. Pleuropneumoniae несколько раз, служит в качестве положительного контроля и разбавлена, как описано ранее. Через 1 ч инкубации при к.т. на шейкере разбавленную сыворотку удаляют и каждую лунку промывают 3 раза промывающим буфером. IgG козы против IgG свиньи (H+L)- HRPO, без MinX (Dianova) служит в качестве вторичного антитела, и его разбавляют 1:4000 в блокирующем буфере, и 50 мкл добавляют в каждую лунку 96-луночного планшета. Через 1 ч инкубации при к.т. на шейкере разбавленные вторичные антитела удаляют и каждую лунку промывают 3 раза промывающим буфером. Затем 50 мкл субстрата TMB (555214, BD) помещают в каждую лунку планшета для ELISA и инкубируют в течение 6 минут при к.т. в темноте. Реакции останавливают путем добавления 20 мкл 2 н. H2SO4 (X873.1, Roth) на лунку. Поглощение в каждой лунке при длине волны 450 нм измеряют на ридере для ELISA. Данные поглощения обрабатывают с использованием GraphPad Prism 8.0 с логарифмическим превращением величин поглощения и нелинейной регрессией с сигмовидной зависимостью доза-ответ. Для статистического анализа результаты ELISA анализировали с использованием критерия Крускала-Уоллиса и критерия множественного сравнения Данна. Данные каждой группы сравнивали с соответствующими данными группы, получавшей плацебо (группа 1). p-величину меньше или равную 0,05 считали значимой. Результаты показаны на фигуре 6.
(4) Серологические данные (нейтрализационный анализ)
Нейтрализационный анализ позволяет количественно оценить нейтрализующую емкость рекомбинантного ApxlA дикого типа в соответствующей сыворотки свиней.
Клетки BL3 (BL3.1 CRL-2306) применяют в качестве клеток-мишеней. Клетки BL3 получали из бычьих B-клеток, а лизис этой клеточной линии токсинами ApxIA был описан (Tu, A. H.; Hausler, C.; Young, R.; Struck, D. K. (1994)). Клетки BL3 культивировали в среде (DMEM, содержащей 100 ед./мл пенициллина, 100 мкг/мл стрептомицина, 10 % FCS и заменимые аминокислоты, 1x) при 37 °C и 5 % CO2. Для культивирования использовали колбы T75 с 25 мл среды. Клетки BL3 пересевали раз в неделю, при этом 50000 клеток/см2 использовали для засевания в новую колбу. Для применения в нейтрализационном анализе 50 мкл клеток BL3 в среде переносили в каждую лунку 96-луночного планшета.
Сыворотку свиней инактивируют нагреванием при 56°C в течение 30 мин. Затем готовят ряд 2-кратных разбавлений сыворотки в DMEM, включая разбавления. Рекомбинантный ApxIA дикого типа (активный токсин) готовят в соответствии с Примером 7 и разбавляют до конечной концентрации 240 мкг/мл. 70 мкл каждого разбавления сыворотки смешивают с 30 мкл разбавлений ApxIA дикого типа (конечное количество ApxIA на лунку: 6 мкг) в 96-луночном планшете (что дает разбавления сыворотки в диапазоне 1:2 - 1:256) и инкубируют в течение 1 ч при 37°C. 50 мкл содержимого каждой лунки переносят в круглодонный 96-луночный планшет, содержащий 50000 клеток BL3/см2 в 50 мкл DMEM), что приводит к конечным разбавлениям сыворотки в диапазоне 1:4 - 1:512. Планшеты инкубируют в течение 24 ч при 37°C в атмосфере, содержащей 5% CO2 и при 100% влажности. Через 24 ч в каждую лунку планшета добавляют субстрат WST-1 (Sigma) в соответствии с инструкцией производителя. Планшеты инкубируют в течение еще 24 ч при 37°C в атмосфере, содержащей 5% CO2 и при 100% влажности. Затем определяли поглощение с помощью ридера для ELISA при 450 нм. WST-1 расщепляется посредством сложного клеточного механизма на клеточной поверхности жизнеспособных клеток до растворимого формазана. Следовательно, количество образовавшегося красителя формазана и величина экстинкции, полученная на ридере для ELISA, напрямую коррелирует с количеством метаболически активных клеток в культуре. Нейтрализационный анализ применяли для сыворотки, полученной в 28 день серологического анализа животных в группах 1, 2 и 3. Результаты показаны на фигуре 7.
ПРИМЕР 16: ЭФФЕКТИВНОСТЬ ПОЛИПЕПТИДОВ APXIA, APXIIA И APXIIIA ПО ИЗОБРЕТЕНИЮ У СВИНЕЙ
(1) Получение антигенов и составов вакцин
Получение антигена и состава осуществляли, как описано в Примере 15. Таблица 9 обобщает состав различных тестовых партий вакцины.
Таблица 9: Состав различных тестовых партий для изучения эффективности
(2) Исследование на животных (эффективность)
Целью этого исследования было оценить эффективность защиты, полученной при вакцинации с использованием состава вакцин на основе рекомбинатно экспрессируемых, модифицированных ApxIA, ApxllA и ApxlllA против серовара 9 Actinobacillus pleuropneumoniae (APP) у свиней.
Животные
Свиней обоих полов (самцы [кастрированные] и самки) приобретали у животновода, разводящего свиней (Dalmandi Mezögazdasági Zrt., 7211 Далманд, Felszabadulás u. 42). Каждую свинью идентифицировали с помощью уникального идентификационного номера на ухе и подвергали ветеринарному осмотру относительно соответствия критериям включения/исключения, указанным в протоколе исследования.
Источник (стадо) не имел APP и Mycoplasma. Поросята получены от самок, которые не были вакцинированы против APP, чтобы предотвратить взаимодействие антител, полученных от матери, с вакцинацией. Животные не имели клинических признаков заболевания (одышка, кашель, диарея, хромота). Кроме того, животные являлись сероотрицательными, что установлено с помощью коммерчески доступного анализа ELISA APP-ApxlV (IDEXX) до включения в исследование. Поросята были хорошо развиты, здоровы без каких-либо нарушений и не имели врожденных пороков развития.
Животных содержали на огражденной территории животноводческого хозяйства PROPHYL Ltd, Top. № 045/7, Бар H-7711, Венгрия). Свиней кормили (нашинкованный корм, продажная смесь) в необходимом количестве с использованием вручную заполненных кормушек для сухих кормов. Воду качества водопроводной подавали с помощью крановой и сосковой поилок. Все помещения убирали раз в день.
Дизайн исследования
Эффективность, вызванную вакциной, содержащей 25 пг каждого модифицированного ApxIA, ApxllA (усеченный) и ApxlllA, которые введены в состав с Montanide ISA 201 VG (партия 36075M, Seppic), по отношению к интраназальному заражению сероваром 9 APP сравнивали с эффективностью, вызванной зарегистрированной вакциной против APP (Porcilis® APP, MSD Animal Health, партия: A739A01) и группой плацебо, получающей ФСБ. Обзор групп и типов воздействия представлен в Таблице 10.
Таблица 10: Обзор групп и воздействие
В возрасте 42-45 дней все свиньи были внутримышечно вакцинированы 2,0 мл соответствующей вакцины в соответствии с группой лечения (0 день исследования). Через 28 дней свиньи получили 2-ю вакцинацию соответствующей вакциной (28 день исследования) с использованием внутримышечного пути. Через две недели после 2-й вакцинации всем животным было произведено внутриназальное заражение сероваром 9 APP (5x107 КОЕ/свинья) (42 день исследования).
Клиническое обследование производили каждый день в течение 7 дней, начиная с дня заражения. Каждый из осмотров приводил к показателю, зависящему от клинических наблюдений и в соответствии с таблицей 11. Клинические показатели накапливают для каждого животного с течением времени, и рассчитывали средний показатель на группу за весь период наблюдения. Для статистического анализа значения клинических показателей анализировали с использованием критерия Крускала-Уоллиса и критерия множественного сравнения Данна. Данные каждой группы сравнивали с соответствующими данными группы, получавшей плацебо (группа 3). p-величину меньше или равную 0,05 считали значимой. Результаты показаны на фигуре 8. Результаты относительно выживаемости животных после заражения показаны на фигуре 9.
Таблица 11: Оценка клинического осмотра
Через 7 дней после заражения все оставшиеся животные были гуманным образом умерщвлены и подвергнуты посмертному осмотру. Легкие всех животных были подвернуты вскрытию для регистрации поражений, вызванных APP. Оценку легких производили в соответствии с системой оценивания, представленной в таблице 12. Поражения легких оценивали от 0 до 5 на долю в соответствии с процентным содержанием уплотненных масс в легких. Результаты оценивания легких выражали как кумулятивные показатели легких (диапазон от 0 до 35) для каждой свиньи. Кумулятивные показатели легких анализировали с использованием критерия Крускала-Уоллиса и критерия множественного сравнения Данна. Данные каждой группы сравнивали с соответствующими данными группы, получавшей плацебо (группа 3). p-величину меньше или равную 0,05 считали значимой. Результаты показаны на фигуре 10.
Таблица 12: Оценка поражений легких
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> IDT Biologika GmbH
<120> Инактивированные токсины ApxIA, ApxIIA и ApxIIIA
<130> P46559-WO
<150> 19154466.7
<151> 2019-01-30
<160> 34
<170> BiSSAP 1.3.6
<210> 1
<211> 1022
<212> Белок
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIA дикого типа
<400> 1
Met Ala Asn Ser Gln Leu Asp Arg Val Lys Gly Leu Ile Asp Ser Leu
1 5 10 15
Asn Gln His Thr Lys Ser Ala Ala Lys Ser Gly Ala Gly Ala Leu Lys
20 25 30
Asn Gly Leu Gly Gln Val Lys Gln Ala Gly Gln Lys Leu Ile Leu Tyr
35 40 45
Ile Pro Lys Asp Tyr Gln Ala Ser Thr Gly Ser Ser Leu Asn Asp Leu
50 55 60
Val Lys Ala Ala Glu Ala Leu Gly Ile Glu Val His Arg Ser Glu Lys
65 70 75 80
Asn Gly Thr Ala Leu Ala Lys Glu Leu Phe Gly Thr Thr Glu Lys Leu
85 90 95
Leu Gly Phe Ser Glu Arg Gly Ile Ala Leu Phe Ala Pro Gln Phe Asp
100 105 110
Lys Leu Leu Asn Lys Asn Gln Lys Leu Ser Lys Ser Leu Gly Gly Ser
115 120 125
Ser Glu Ala Leu Gly Gln Arg Leu Asn Lys Thr Gln Thr Ala Leu Ser
130 135 140
Ala Leu Gln Ser Phe Leu Gly Thr Ala Ile Ala Gly Met Asp Leu Asp
145 150 155 160
Ser Leu Leu Arg Arg Arg Arg Asn Gly Glu Asp Val Ser Gly Ser Glu
165 170 175
Leu Ala Lys Ala Gly Val Asp Leu Ala Ala Gln Leu Val Asp Asn Ile
180 185 190
Ala Ser Ala Thr Gly Thr Val Asp Ala Phe Ala Glu Gln Leu Gly Lys
195 200 205
Leu Gly Asn Ala Leu Ser Asn Thr Arg Leu Ser Gly Leu Ala Ser Lys
210 215 220
Leu Asn Asn Leu Pro Asp Leu Ser Leu Ala Gly Pro Gly Phe Asp Ala
225 230 235 240
Val Ser Gly Ile Leu Ser Val Val Ser Ala Ser Phe Ile Leu Ser Asn
245 250 255
Lys Asp Ala Asp Ala Gly Thr Lys Ala Ala Ala Gly Ile Glu Ile Ser
260 265 270
Thr Lys Ile Leu Gly Asn Ile Gly Lys Ala Val Ser Gln Tyr Ile Ile
275 280 285
Ala Gln Arg Val Ala Ala Gly Leu Ser Thr Thr Ala Ala Thr Gly Gly
290 295 300
Leu Ile Gly Ser Val Val Ala Leu Ala Ile Ser Pro Leu Ser Phe Leu
305 310 315 320
Asn Val Ala Asp Lys Phe Glu Arg Ala Lys Gln Leu Glu Gln Tyr Ser
325 330 335
Glu Arg Phe Lys Lys Phe Gly Tyr Glu Gly Asp Ser Leu Leu Ala Ser
340 345 350
Phe Tyr Arg Glu Thr Gly Ala Ile Glu Ala Ala Leu Thr Thr Ile Asn
355 360 365
Ser Val Leu Ser Ala Ala Ser Ala Gly Val Gly Ala Ala Ala Thr Gly
370 375 380
Ser Leu Val Gly Ala Pro Val Ala Ala Leu Val Ser Ala Ile Thr Gly
385 390 395 400
Ile Ile Ser Gly Ile Leu Asp Ala Ser Lys Gln Ala Ile Phe Glu Arg
405 410 415
Val Ala Thr Lys Leu Ala Asn Lys Ile Asp Glu Trp Glu Lys Lys His
420 425 430
Gly Lys Asn Tyr Phe Glu Asn Gly Tyr Asp Ala Arg His Ser Ala Phe
435 440 445
Leu Glu Asp Thr Phe Glu Leu Leu Ser Gln Tyr Asn Lys Glu Tyr Ser
450 455 460
Val Glu Arg Val Val Ala Ile Thr Gln Gln Arg Trp Asp Val Asn Ile
465 470 475 480
Gly Glu Leu Ala Gly Ile Thr Arg Lys Gly Ala Asp Ala Lys Ser Gly
485 490 495
Lys Ala Tyr Val Asp Phe Phe Glu Glu Gly Lys Leu Leu Glu Lys Asp
500 505 510
Pro Asp Arg Phe Asp Lys Lys Val Phe Asp Pro Leu Glu Gly Lys Ile
515 520 525
Asp Leu Ser Ser Ile Asn Lys Thr Thr Leu Leu Lys Phe Ile Thr Pro
530 535 540
Val Phe Thr Ala Gly Glu Glu Ile Arg Glu Arg Lys Gln Thr Gly Lys
545 550 555 560
Tyr Glu Tyr Met Thr Glu Leu Phe Val Lys Gly Lys Glu Lys Trp Val
565 570 575
Val Thr Gly Val Gln Ser His Asn Ala Ile Tyr Asp Tyr Thr Asn Leu
580 585 590
Ile Gln Leu Ala Ile Asp Lys Lys Gly Glu Lys Arg Gln Val Thr Ile
595 600 605
Glu Ser His Leu Gly Glu Lys Asn Asp Arg Ile Tyr Leu Ser Ser Gly
610 615 620
Ser Ser Ile Val Tyr Ala Gly Asn Gly His Asp Val Ala Tyr Tyr Asp
625 630 635 640
Lys Thr Asp Thr Gly Tyr Leu Thr Phe Asp Gly Gln Ser Ala Gln Lys
645 650 655
Ala Gly Glu Tyr Ile Val Thr Lys Glu Leu Lys Ala Asp Val Lys Val
660 665 670
Leu Lys Glu Val Val Lys Thr Gln Asp Ile Ser Val Gly Lys Arg Ser
675 680 685
Glu Lys Leu Glu Tyr Arg Asp Tyr Glu Leu Ser Pro Phe Glu Leu Gly
690 695 700
Asn Gly Ile Arg Ala Lys Asp Glu Leu His Ser Val Glu Glu Ile Ile
705 710 715 720
Gly Ser Asn Arg Lys Asp Lys Phe Phe Gly Ser Arg Phe Thr Asp Ile
725 730 735
Phe His Gly Ala Lys Gly Asp Asp Glu Ile Tyr Gly Asn Asp Gly His
740 745 750
Asp Ile Leu Tyr Gly Asp Asp Gly Asn Asp Val Ile His Gly Gly Asp
755 760 765
Gly Asn Asp His Leu Val Gly Gly Asn Gly Asn Asp Arg Leu Ile Gly
770 775 780
Gly Lys Gly Asn Asn Phe Leu Asn Gly Gly Asp Gly Asp Asp Glu Leu
785 790 795 800
Gln Val Phe Glu Gly Gln Tyr Asn Val Leu Leu Gly Gly Ala Gly Asn
805 810 815
Asp Ile Leu Tyr Gly Ser Asp Gly Thr Asn Leu Phe Asp Gly Gly Val
820 825 830
Gly Asn Asp Lys Ile Tyr Gly Gly Leu Gly Lys Asp Ile Tyr Arg Tyr
835 840 845
Ser Lys Glu Tyr Gly Arg His Ile Ile Ile Glu Lys Gly Gly Asp Asp
850 855 860
Asp Thr Leu Leu Leu Ser Asp Leu Ser Phe Lys Asp Val Gly Phe Ile
865 870 875 880
Arg Ile Gly Asp Asp Leu Leu Val Asn Lys Arg Ile Gly Gly Thr Leu
885 890 895
Tyr Tyr His Glu Asp Tyr Asn Gly Asn Ala Leu Thr Ile Lys Asp Trp
900 905 910
Phe Lys Glu Gly Lys Glu Gly Gln Asn Asn Lys Ile Glu Lys Ile Val
915 920 925
Asp Lys Asp Gly Ala Tyr Val Leu Ser Gln Tyr Leu Thr Glu Leu Thr
930 935 940
Ala Pro Gly Arg Gly Ile Asn Tyr Phe Asn Gly Leu Glu Glu Lys Leu
945 950 955 960
Tyr Tyr Gly Glu Gly Tyr Asn Ala Leu Pro Gln Leu Arg Lys Asp Ile
965 970 975
Glu Gln Ile Ile Ser Ser Thr Gly Ala Leu Thr Gly Glu His Gly Gln
980 985 990
Val Leu Val Gly Ala Gly Gly Pro Leu Ala Tyr Ser Asn Ser Pro Asn
995 1000 1005
Ser Ile Pro Asn Ala Phe Ser Asn Tyr Leu Thr Gln Ser Ala
1010 1015 1020
<210> 2
<211> 956
<212> Белок
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIA дикого типа
<400> 2
Met Ser Lys Ile Thr Leu Ser Ser Leu Lys Ser Ser Leu Gln Gln Gly
1 5 10 15
Leu Lys Asn Gly Lys Asn Lys Leu Asn Gln Ala Gly Thr Thr Leu Lys
20 25 30
Asn Gly Leu Thr Gln Thr Gly His Ser Leu Gln Asn Gly Ala Lys Lys
35 40 45
Leu Ile Leu Tyr Ile Pro Gln Gly Tyr Asp Ser Gly Gln Gly Asn Gly
50 55 60
Val Gln Asp Leu Val Lys Ala Ala Asn Asp Leu Gly Ile Glu Val Trp
65 70 75 80
Arg Glu Glu Arg Ser Asn Leu Asp Ile Ala Lys Thr Ser Phe Asp Thr
85 90 95
Thr Gln Lys Ile Leu Gly Phe Thr Asp Arg Gly Ile Val Leu Phe Ala
100 105 110
Pro Gln Leu Asp Asn Leu Leu Lys Lys Asn Pro Lys Ile Gly Asn Thr
115 120 125
Leu Gly Ser Ala Ser Ser Ile Ser Gln Asn Ile Gly Lys Ala Asn Thr
130 135 140
Val Leu Gly Gly Ile Gln Ser Ile Leu Gly Ser Val Leu Ser Gly Val
145 150 155 160
Asn Leu Asn Glu Leu Leu Gln Asn Lys Asp Pro Asn Gln Leu Glu Leu
165 170 175
Ala Lys Ala Gly Leu Glu Leu Thr Asn Glu Leu Val Gly Asn Ile Ala
180 185 190
Ser Ser Val Gln Thr Val Asp Ala Phe Ala Glu Gln Ile Ser Lys Leu
195 200 205
Gly Ser His Leu Gln Asn Val Lys Gly Leu Gly Gly Leu Ser Asn Lys
210 215 220
Leu Gln Asn Leu Pro Asp Leu Gly Lys Ala Ser Leu Gly Leu Asp Ile
225 230 235 240
Ile Ser Gly Leu Leu Ser Gly Ala Ser Ala Gly Leu Ile Leu Ala Asp
245 250 255
Lys Glu Ala Ser Thr Glu Lys Lys Ala Ala Ala Gly Val Glu Phe Ala
260 265 270
Asn Gln Ile Ile Gly Asn Val Thr Lys Ala Val Ser Ser Tyr Ile Leu
275 280 285
Ala Gln Arg Val Ala Ser Gly Leu Ser Ser Thr Gly Pro Val Ala Ala
290 295 300
Leu Ile Ala Ser Thr Val Ala Leu Ala Val Ser Pro Leu Ser Phe Leu
305 310 315 320
Asn Val Ala Asp Lys Phe Lys Gln Ala Asp Leu Ile Lys Ser Tyr Ser
325 330 335
Glu Arg Phe Gln Lys Leu Gly Tyr Asp Gly Asp Arg Leu Leu Ala Asp
340 345 350
Phe His Arg Glu Thr Gly Thr Ile Asp Ala Ser Val Thr Thr Ile Asn
355 360 365
Thr Ala Leu Ala Ala Ile Ser Gly Gly Val Gly Ala Ala Ser Ala Gly
370 375 380
Ser Leu Val Gly Ala Pro Val Ala Leu Leu Val Ala Gly Val Thr Gly
385 390 395 400
Leu Ile Thr Thr Ile Leu Glu Tyr Ser Lys Gln Ala Met Phe Glu His
405 410 415
Val Ala Asn Lys Val His Asp Arg Ile Val Glu Trp Glu Lys Lys His
420 425 430
Asn Lys Asn Tyr Phe Glu Gln Gly Tyr Asp Ser Arg His Leu Ala Asp
435 440 445
Leu Gln Asp Asn Met Lys Phe Leu Ile Asn Leu Asn Lys Glu Leu Gln
450 455 460
Ala Glu Arg Val Val Ala Ile Thr Gln Gln Arg Trp Asp Asn Gln Ile
465 470 475 480
Gly Asp Leu Ala Ala Ile Ser Arg Arg Thr Asp Lys Ile Ser Ser Gly
485 490 495
Lys Ala Tyr Val Asp Ala Phe Glu Glu Gly Gln His Gln Ser Tyr Asp
500 505 510
Ser Ser Val Gln Leu Asp Asn Lys Asn Gly Ile Ile Asn Ile Ser Asn
515 520 525
Thr Asn Arg Lys Thr Gln Ser Val Leu Phe Arg Thr Pro Leu Leu Thr
530 535 540
Pro Gly Glu Glu Asn Arg Glu Arg Ile Gln Glu Gly Lys Asn Ser Tyr
545 550 555 560
Ile Thr Lys Leu His Ile Gln Arg Val Asp Ser Trp Thr Val Thr Asp
565 570 575
Gly Asp Ala Ser Ser Ser Val Asp Phe Thr Asn Val Val Gln Arg Ile
580 585 590
Ala Val Lys Phe Asp Asp Ala Gly Asn Ile Ile Glu Ser Lys Asp Thr
595 600 605
Lys Ile Ile Ala Asn Leu Gly Ala Gly Asn Asp Asn Val Phe Val Gly
610 615 620
Ser Ser Thr Thr Val Ile Asp Gly Gly Asp Gly His Asp Arg Val His
625 630 635 640
Tyr Ser Arg Gly Glu Tyr Gly Ala Leu Val Ile Asp Ala Thr Ala Glu
645 650 655
Thr Glu Lys Gly Ser Tyr Ser Val Lys Arg Tyr Val Gly Asp Ser Lys
660 665 670
Ala Leu His Glu Thr Ile Ala Thr His Gln Thr Asn Val Gly Asn Arg
675 680 685
Glu Glu Lys Ile Glu Tyr Arg Arg Glu Asp Asp Arg Phe His Thr Gly
690 695 700
Tyr Thr Val Thr Asp Ser Leu Lys Ser Val Glu Glu Ile Ile Gly Ser
705 710 715 720
Gln Phe Asn Asp Ile Phe Lys Gly Ser Gln Phe Asp Asp Val Phe His
725 730 735
Gly Gly Asn Gly Val Asp Thr Ile Asp Gly Asn Asp Gly Asp Asp His
740 745 750
Leu Phe Gly Gly Ala Gly Asp Asp Val Ile Asp Gly Gly Asn Gly Asn
755 760 765
Asn Phe Leu Val Gly Gly Thr Gly Asn Asp Ile Ile Ser Gly Gly Lys
770 775 780
Asp Asn Asp Ile Tyr Val His Lys Thr Gly Asp Gly Asn Asp Ser Ile
785 790 795 800
Thr Asp Ser Gly Gly Gln Asp Lys Leu Ala Phe Ser Asp Val Asn Leu
805 810 815
Lys Asp Leu Thr Phe Lys Lys Val Asp Ser Ser Leu Glu Ile Ile Asn
820 825 830
Gln Lys Gly Glu Lys Val Arg Ile Gly Asn Trp Phe Leu Glu Asp Asp
835 840 845
Leu Ala Ser Thr Val Ala Asn Tyr Lys Ala Thr Asn Asp Arg Lys Ile
850 855 860
Glu Glu Ile Ile Gly Lys Gly Gly Glu Arg Ile Thr Ser Glu Gln Val
865 870 875 880
Asp Lys Leu Ile Lys Glu Gly Asn Asn Gln Ile Ser Ala Glu Ala Leu
885 890 895
Ser Lys Val Val Asn Asp Tyr Asn Thr Ser Lys Asp Arg Gln Asn Val
900 905 910
Ser Asn Ser Leu Ala Lys Leu Ile Ser Ser Val Gly Ser Phe Thr Ser
915 920 925
Ser Ser Asp Phe Arg Asn Asn Leu Gly Thr Tyr Val Pro Ser Ser Ile
930 935 940
Asp Val Ser Asn Asn Ile Gln Leu Ala Arg Ala Ala
945 950 955
<210> 3
<211> 1052
<212> Белок
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIIA дикого типа
<400> 3
Met Ser Thr Trp Ser Ser Met Leu Ala Asp Leu Lys Lys Arg Ala Glu
1 5 10 15
Glu Ala Lys Arg Gln Ala Lys Lys Gly Tyr Asp Val Thr Lys Asn Gly
20 25 30
Leu Gln Tyr Gly Val Ser Gln Ala Lys Leu Gln Ala Leu Ala Ala Gly
35 40 45
Lys Ala Val Gln Lys Tyr Gly Asn Lys Leu Val Leu Val Ile Pro Lys
50 55 60
Glu Tyr Asp Gly Ser Val Gly Asn Gly Phe Phe Asp Leu Val Lys Ala
65 70 75 80
Ala Glu Glu Leu Gly Ile Gln Val Lys Tyr Val Asn Arg Asn Glu Leu
85 90 95
Glu Val Ala His Lys Ser Leu Gly Thr Ala Asp Gln Phe Leu Gly Leu
100 105 110
Thr Glu Arg Gly Leu Thr Leu Phe Ala Pro Gln Leu Asp Gln Phe Leu
115 120 125
Gln Lys His Ser Lys Ile Ser Asn Val Val Gly Ser Ser Thr Gly Asp
130 135 140
Ala Val Ser Lys Leu Ala Lys Ser Gln Thr Ile Ile Ser Gly Ile Gln
145 150 155 160
Ser Val Leu Gly Thr Val Leu Ala Gly Ile Asn Leu Asn Glu Ala Ile
165 170 175
Ile Ser Gly Gly Ser Glu Leu Glu Leu Ala Glu Ala Gly Val Ser Leu
180 185 190
Ala Ser Glu Leu Val Ser Asn Ile Ala Lys Gly Thr Thr Thr Ile Asp
195 200 205
Ala Phe Thr Thr Gln Ile Gln Asn Phe Gly Lys Leu Val Glu Asn Ala
210 215 220
Lys Gly Leu Gly Gly Val Gly Arg Gln Leu Gln Asn Ile Ser Gly Ser
225 230 235 240
Ala Leu Ser Lys Thr Gly Leu Gly Leu Asp Ile Ile Ser Ser Leu Leu
245 250 255
Ser Gly Val Thr Ala Ser Phe Ala Leu Ala Asn Lys Asn Ala Ser Thr
260 265 270
Ser Thr Lys Val Ala Ala Gly Phe Glu Leu Ser Asn Gln Val Ile Gly
275 280 285
Gly Ile Thr Lys Ala Val Ser Ser Tyr Ile Leu Ala Gln Arg Leu Ala
290 295 300
Ala Gly Leu Ser Thr Thr Gly Pro Ala Ala Ala Leu Ile Ala Ser Ser
305 310 315 320
Ile Ser Leu Ala Ile Ser Pro Leu Ala Phe Leu Arg Val Ala Asp Asn
325 330 335
Phe Asn Arg Ser Lys Glu Ile Gly Glu Phe Ala Glu Arg Phe Lys Lys
340 345 350
Leu Gly Tyr Asp Gly Asp Lys Leu Leu Ser Glu Phe Tyr His Glu Ala
355 360 365
Gly Thr Ile Asp Ala Ser Ile Thr Thr Ile Ser Thr Ala Leu Ser Ala
370 375 380
Ile Ala Ala Gly Thr Ala Ala Ala Ser Ala Gly Ala Leu Val Gly Ala
385 390 395 400
Pro Ile Thr Leu Leu Val Thr Gly Ile Thr Gly Leu Ile Ser Gly Ile
405 410 415
Leu Glu Phe Ser Lys Gln Pro Met Leu Asp His Val Ala Ser Lys Ile
420 425 430
Gly Asn Lys Ile Asp Glu Trp Glu Lys Lys Tyr Gly Lys Asn Tyr Phe
435 440 445
Glu Asn Gly Tyr Asp Ala Arg His Lys Ala Phe Leu Glu Asp Ser Phe
450 455 460
Ser Leu Leu Ser Ser Phe Asn Lys Gln Tyr Glu Thr Glu Arg Ala Val
465 470 475 480
Leu Ile Thr Gln Gln Arg Trp Asp Glu Tyr Ile Gly Glu Leu Ala Gly
485 490 495
Ile Thr Gly Lys Gly Asp Lys Leu Ser Ser Gly Lys Ala Tyr Val Asp
500 505 510
Tyr Phe Gln Glu Gly Lys Leu Leu Glu Lys Lys Pro Asp Asp Phe Ser
515 520 525
Lys Val Val Phe Asp Pro Thr Lys Gly Glu Ile Asp Ile Ser Asn Ser
530 535 540
Gln Thr Ser Thr Leu Leu Lys Phe Val Thr Pro Leu Leu Thr Pro Gly
545 550 555 560
Thr Glu Ser Arg Glu Arg Thr Gln Thr Gly Lys Tyr Glu Tyr Ile Thr
565 570 575
Lys Leu Val Val Lys Gly Lys Asp Lys Trp Val Val Asn Gly Val Lys
580 585 590
Asp Lys Gly Ala Val Tyr Asp Tyr Thr Asn Leu Ile Gln His Ala His
595 600 605
Ile Ser Ser Ser Val Ala Arg Gly Glu Glu Tyr Arg Glu Val Arg Leu
610 615 620
Val Ser His Leu Gly Asn Gly Asn Asp Lys Val Phe Leu Ala Ala Gly
625 630 635 640
Ser Ala Glu Ile His Ala Gly Glu Gly His Asp Val Val Tyr Tyr Asp
645 650 655
Lys Thr Asp Thr Gly Leu Leu Val Ile Asp Gly Thr Lys Ala Thr Glu
660 665 670
Gln Gly Arg Tyr Ser Val Thr Arg Glu Leu Ser Gly Ala Thr Lys Ile
675 680 685
Leu Arg Glu Val Ile Lys Asn Gln Lys Ser Ala Val Gly Lys Arg Glu
690 695 700
Glu Thr Leu Glu Tyr Arg Asp Tyr Glu Leu Thr Gln Ser Gly Asn Ser
705 710 715 720
Asn Leu Lys Ala His Asp Glu Leu His Ser Val Glu Glu Ile Ile Gly
725 730 735
Ser Asn Gln Arg Asp Glu Phe Lys Gly Ser Lys Phe Arg Asp Ile Phe
740 745 750
His Gly Ala Asp Gly Asp Asp Leu Leu Asn Gly Asn Asp Gly Asp Asp
755 760 765
Ile Leu Tyr Gly Asp Lys Gly Asn Asp Glu Leu Arg Gly Asp Asn Gly
770 775 780
Asn Asp Gln Leu Tyr Gly Gly Glu Gly Asn Asp Lys Leu Leu Gly Gly
785 790 795 800
Asn Gly Asn Asn Tyr Leu Ser Gly Gly Asp Gly Asn Asp Glu Leu Gln
805 810 815
Val Leu Gly Asn Gly Phe Asn Val Leu Arg Gly Gly Lys Gly Asp Asp
820 825 830
Lys Leu Tyr Gly Ser Ser Gly Ser Asp Leu Leu Asp Gly Gly Glu Gly
835 840 845
Asn Asp Tyr Leu Glu Gly Gly Asp Gly Ser Asp Phe Tyr Val Tyr Arg
850 855 860
Ser Thr Ser Gly Asn His Thr Ile Tyr Asp Gln Gly Lys Ser Ser Asp
865 870 875 880
Leu Asp Lys Leu Tyr Leu Ser Asp Phe Ser Phe Asp Arg Leu Leu Val
885 890 895
Glu Lys Val Asp Asp Asn Leu Val Leu Arg Ser Asn Glu Ser Ser His
900 905 910
Asn Asn Gly Val Leu Thr Ile Lys Asp Trp Phe Lys Glu Gly Asn Lys
915 920 925
Tyr Asn His Lys Ile Glu Gln Ile Val Asp Lys Asn Gly Arg Lys Leu
930 935 940
Thr Ala Glu Asn Leu Gly Thr Tyr Phe Lys Asn Ala Pro Lys Ala Asp
945 950 955 960
Asn Leu Leu Asn Tyr Ala Thr Lys Glu Asp Gln Asn Glu Ser Asn Leu
965 970 975
Ser Ser Leu Lys Thr Glu Leu Ser Lys Ile Ile Thr Asn Ala Gly Asn
980 985 990
Phe Gly Val Ala Lys Gln Gly Asn Thr Gly Ile Asn Thr Ala Ala Leu
995 1000 1005
Asn Asn Glu Val Asn Lys Ile Ile Ser Ser Ala Asn Thr Phe Ala Thr
1010 1015 1020
Ser Gln Leu Gly Gly Ser Gly Met Gly Thr Leu Pro Ser Thr Asn Val
1025 1030 1035 1040
Asn Ser Met Met Leu Gly Asn Leu Ala Arg Ala Ala
1045 1050
<210> 4
<211> 1022
<212> Белок
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIA K560A K686A
<400> 4
Met Ala Asn Ser Gln Leu Asp Arg Val Lys Gly Leu Ile Asp Ser Leu
1 5 10 15
Asn Gln His Thr Lys Ser Ala Ala Lys Ser Gly Ala Gly Ala Leu Lys
20 25 30
Asn Gly Leu Gly Gln Val Lys Gln Ala Gly Gln Lys Leu Ile Leu Tyr
35 40 45
Ile Pro Lys Asp Tyr Gln Ala Ser Thr Gly Ser Ser Leu Asn Asp Leu
50 55 60
Val Lys Ala Ala Glu Ala Leu Gly Ile Glu Val His Arg Ser Glu Lys
65 70 75 80
Asn Gly Thr Ala Leu Ala Lys Glu Leu Phe Gly Thr Thr Glu Lys Leu
85 90 95
Leu Gly Phe Ser Glu Arg Gly Ile Ala Leu Phe Ala Pro Gln Phe Asp
100 105 110
Lys Leu Leu Asn Lys Asn Gln Lys Leu Ser Lys Ser Leu Gly Gly Ser
115 120 125
Ser Glu Ala Leu Gly Gln Arg Leu Asn Lys Thr Gln Thr Ala Leu Ser
130 135 140
Ala Leu Gln Ser Phe Leu Gly Thr Ala Ile Ala Gly Met Asp Leu Asp
145 150 155 160
Ser Leu Leu Arg Arg Arg Arg Asn Gly Glu Asp Val Ser Gly Ser Glu
165 170 175
Leu Ala Lys Ala Gly Val Asp Leu Ala Ala Gln Leu Val Asp Asn Ile
180 185 190
Ala Ser Ala Thr Gly Thr Val Asp Ala Phe Ala Glu Gln Leu Gly Lys
195 200 205
Leu Gly Asn Ala Leu Ser Asn Thr Arg Leu Ser Gly Leu Ala Ser Lys
210 215 220
Leu Asn Asn Leu Pro Asp Leu Ser Leu Ala Gly Pro Gly Phe Asp Ala
225 230 235 240
Val Ser Gly Ile Leu Ser Val Val Ser Ala Ser Phe Ile Leu Ser Asn
245 250 255
Lys Asp Ala Asp Ala Gly Thr Lys Ala Ala Ala Gly Ile Glu Ile Ser
260 265 270
Thr Lys Ile Leu Gly Asn Ile Gly Lys Ala Val Ser Gln Tyr Ile Ile
275 280 285
Ala Gln Arg Val Ala Ala Gly Leu Ser Thr Thr Ala Ala Thr Gly Gly
290 295 300
Leu Ile Gly Ser Val Val Ala Leu Ala Ile Ser Pro Leu Ser Phe Leu
305 310 315 320
Asn Val Ala Asp Lys Phe Glu Arg Ala Lys Gln Leu Glu Gln Tyr Ser
325 330 335
Glu Arg Phe Lys Lys Phe Gly Tyr Glu Gly Asp Ser Leu Leu Ala Ser
340 345 350
Phe Tyr Arg Glu Thr Gly Ala Ile Glu Ala Ala Leu Thr Thr Ile Asn
355 360 365
Ser Val Leu Ser Ala Ala Ser Ala Gly Val Gly Ala Ala Ala Thr Gly
370 375 380
Ser Leu Val Gly Ala Pro Val Ala Ala Leu Val Ser Ala Ile Thr Gly
385 390 395 400
Ile Ile Ser Gly Ile Leu Asp Ala Ser Lys Gln Ala Ile Phe Glu Arg
405 410 415
Val Ala Thr Lys Leu Ala Asn Lys Ile Asp Glu Trp Glu Lys Lys His
420 425 430
Gly Lys Asn Tyr Phe Glu Asn Gly Tyr Asp Ala Arg His Ser Ala Phe
435 440 445
Leu Glu Asp Thr Phe Glu Leu Leu Ser Gln Tyr Asn Lys Glu Tyr Ser
450 455 460
Val Glu Arg Val Val Ala Ile Thr Gln Gln Arg Trp Asp Val Asn Ile
465 470 475 480
Gly Glu Leu Ala Gly Ile Thr Arg Lys Gly Ala Asp Ala Lys Ser Gly
485 490 495
Lys Ala Tyr Val Asp Phe Phe Glu Glu Gly Lys Leu Leu Glu Lys Asp
500 505 510
Pro Asp Arg Phe Asp Lys Lys Val Phe Asp Pro Leu Glu Gly Lys Ile
515 520 525
Asp Leu Ser Ser Ile Asn Lys Thr Thr Leu Leu Lys Phe Ile Thr Pro
530 535 540
Val Phe Thr Ala Gly Glu Glu Ile Arg Glu Arg Lys Gln Thr Gly Ala
545 550 555 560
Tyr Glu Tyr Met Thr Glu Leu Phe Val Lys Gly Lys Glu Lys Trp Val
565 570 575
Val Thr Gly Val Gln Ser His Asn Ala Ile Tyr Asp Tyr Thr Asn Leu
580 585 590
Ile Gln Leu Ala Ile Asp Lys Lys Gly Glu Lys Arg Gln Val Thr Ile
595 600 605
Glu Ser His Leu Gly Glu Lys Asn Asp Arg Ile Tyr Leu Ser Ser Gly
610 615 620
Ser Ser Ile Val Tyr Ala Gly Asn Gly His Asp Val Ala Tyr Tyr Asp
625 630 635 640
Lys Thr Asp Thr Gly Tyr Leu Thr Phe Asp Gly Gln Ser Ala Gln Lys
645 650 655
Ala Gly Glu Tyr Ile Val Thr Lys Glu Leu Lys Ala Asp Val Lys Val
660 665 670
Leu Lys Glu Val Val Lys Thr Gln Asp Ile Ser Val Gly Ala Arg Ser
675 680 685
Glu Lys Leu Glu Tyr Arg Asp Tyr Glu Leu Ser Pro Phe Glu Leu Gly
690 695 700
Asn Gly Ile Arg Ala Lys Asp Glu Leu His Ser Val Glu Glu Ile Ile
705 710 715 720
Gly Ser Asn Arg Lys Asp Lys Phe Phe Gly Ser Arg Phe Thr Asp Ile
725 730 735
Phe His Gly Ala Lys Gly Asp Asp Glu Ile Tyr Gly Asn Asp Gly His
740 745 750
Asp Ile Leu Tyr Gly Asp Asp Gly Asn Asp Val Ile His Gly Gly Asp
755 760 765
Gly Asn Asp His Leu Val Gly Gly Asn Gly Asn Asp Arg Leu Ile Gly
770 775 780
Gly Lys Gly Asn Asn Phe Leu Asn Gly Gly Asp Gly Asp Asp Glu Leu
785 790 795 800
Gln Val Phe Glu Gly Gln Tyr Asn Val Leu Leu Gly Gly Ala Gly Asn
805 810 815
Asp Ile Leu Tyr Gly Ser Asp Gly Thr Asn Leu Phe Asp Gly Gly Val
820 825 830
Gly Asn Asp Lys Ile Tyr Gly Gly Leu Gly Lys Asp Ile Tyr Arg Tyr
835 840 845
Ser Lys Glu Tyr Gly Arg His Ile Ile Ile Glu Lys Gly Gly Asp Asp
850 855 860
Asp Thr Leu Leu Leu Ser Asp Leu Ser Phe Lys Asp Val Gly Phe Ile
865 870 875 880
Arg Ile Gly Asp Asp Leu Leu Val Asn Lys Arg Ile Gly Gly Thr Leu
885 890 895
Tyr Tyr His Glu Asp Tyr Asn Gly Asn Ala Leu Thr Ile Lys Asp Trp
900 905 910
Phe Lys Glu Gly Lys Glu Gly Gln Asn Asn Lys Ile Glu Lys Ile Val
915 920 925
Asp Lys Asp Gly Ala Tyr Val Leu Ser Gln Tyr Leu Thr Glu Leu Thr
930 935 940
Ala Pro Gly Arg Gly Ile Asn Tyr Phe Asn Gly Leu Glu Glu Lys Leu
945 950 955 960
Tyr Tyr Gly Glu Gly Tyr Asn Ala Leu Pro Gln Leu Arg Lys Asp Ile
965 970 975
Glu Gln Ile Ile Ser Ser Thr Gly Ala Leu Thr Gly Glu His Gly Gln
980 985 990
Val Leu Val Gly Ala Gly Gly Pro Leu Ala Tyr Ser Asn Ser Pro Asn
995 1000 1005
Ser Ile Pro Asn Ala Phe Ser Asn Tyr Leu Thr Gln Ser Ala
1010 1015 1020
<210> 5
<211> 956
<212> Белок
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIA S148G K557A N687A
<400> 5
Met Ser Lys Ile Thr Leu Ser Ser Leu Lys Ser Ser Leu Gln Gln Gly
1 5 10 15
Leu Lys Asn Gly Lys Asn Lys Leu Asn Gln Ala Gly Thr Thr Leu Lys
20 25 30
Asn Gly Leu Thr Gln Thr Gly His Ser Leu Gln Asn Gly Ala Lys Lys
35 40 45
Leu Ile Leu Tyr Ile Pro Gln Gly Tyr Asp Ser Gly Gln Gly Asn Gly
50 55 60
Val Gln Asp Leu Val Lys Ala Ala Asn Asp Leu Gly Ile Glu Val Trp
65 70 75 80
Arg Glu Glu Arg Ser Asn Leu Asp Ile Ala Lys Thr Ser Phe Asp Thr
85 90 95
Thr Gln Lys Ile Leu Gly Phe Thr Asp Arg Gly Ile Val Leu Phe Ala
100 105 110
Pro Gln Leu Asp Asn Leu Leu Lys Lys Asn Pro Lys Ile Gly Asn Thr
115 120 125
Leu Gly Ser Ala Ser Ser Ile Ser Gln Asn Ile Gly Lys Ala Asn Thr
130 135 140
Val Leu Gly Gly Ile Gln Ser Ile Leu Gly Ser Val Leu Ser Gly Val
145 150 155 160
Asn Leu Asn Glu Leu Leu Gln Asn Lys Asp Pro Asn Gln Leu Glu Leu
165 170 175
Ala Lys Ala Gly Leu Glu Leu Thr Asn Glu Leu Val Gly Asn Ile Ala
180 185 190
Ser Ser Val Gln Thr Val Asp Ala Phe Ala Glu Gln Ile Ser Lys Leu
195 200 205
Gly Ser His Leu Gln Asn Val Lys Gly Leu Gly Gly Leu Ser Asn Lys
210 215 220
Leu Gln Asn Leu Pro Asp Leu Gly Lys Ala Ser Leu Gly Leu Asp Ile
225 230 235 240
Ile Ser Gly Leu Leu Ser Gly Ala Ser Ala Gly Leu Ile Leu Ala Asp
245 250 255
Lys Glu Ala Ser Thr Glu Lys Lys Ala Ala Ala Gly Val Glu Phe Ala
260 265 270
Asn Gln Ile Ile Gly Asn Val Thr Lys Ala Val Ser Ser Tyr Ile Leu
275 280 285
Ala Gln Arg Val Ala Ser Gly Leu Ser Ser Thr Gly Pro Val Ala Ala
290 295 300
Leu Ile Ala Ser Thr Val Ala Leu Ala Val Ser Pro Leu Ser Phe Leu
305 310 315 320
Asn Val Ala Asp Lys Phe Lys Gln Ala Asp Leu Ile Lys Ser Tyr Ser
325 330 335
Glu Arg Phe Gln Lys Leu Gly Tyr Asp Gly Asp Arg Leu Leu Ala Asp
340 345 350
Phe His Arg Glu Thr Gly Thr Ile Asp Ala Ser Val Thr Thr Ile Asn
355 360 365
Thr Ala Leu Ala Ala Ile Ser Gly Gly Val Gly Ala Ala Ser Ala Gly
370 375 380
Ser Leu Val Gly Ala Pro Val Ala Leu Leu Val Ala Gly Val Thr Gly
385 390 395 400
Leu Ile Thr Thr Ile Leu Glu Tyr Ser Lys Gln Ala Met Phe Glu His
405 410 415
Val Ala Asn Lys Val His Asp Arg Ile Val Glu Trp Glu Lys Lys His
420 425 430
Asn Lys Asn Tyr Phe Glu Gln Gly Tyr Asp Ser Arg His Leu Ala Asp
435 440 445
Leu Gln Asp Asn Met Lys Phe Leu Ile Asn Leu Asn Lys Glu Leu Gln
450 455 460
Ala Glu Arg Val Val Ala Ile Thr Gln Gln Arg Trp Asp Asn Gln Ile
465 470 475 480
Gly Asp Leu Ala Ala Ile Ser Arg Arg Thr Asp Lys Ile Ser Ser Gly
485 490 495
Lys Ala Tyr Val Asp Ala Phe Glu Glu Gly Gln His Gln Ser Tyr Asp
500 505 510
Ser Ser Val Gln Leu Asp Asn Lys Asn Gly Ile Ile Asn Ile Ser Asn
515 520 525
Thr Asn Arg Lys Thr Gln Ser Val Leu Phe Arg Thr Pro Leu Leu Thr
530 535 540
Pro Gly Glu Glu Asn Arg Glu Arg Ile Gln Glu Gly Ala Asn Ser Tyr
545 550 555 560
Ile Thr Lys Leu His Ile Gln Arg Val Asp Ser Trp Thr Val Thr Asp
565 570 575
Gly Asp Ala Ser Ser Ser Val Asp Phe Thr Asn Val Val Gln Arg Ile
580 585 590
Ala Val Lys Phe Asp Asp Ala Gly Asn Ile Ile Glu Ser Lys Asp Thr
595 600 605
Lys Ile Ile Ala Asn Leu Gly Ala Gly Asn Asp Asn Val Phe Val Gly
610 615 620
Ser Ser Thr Thr Val Ile Asp Gly Gly Asp Gly His Asp Arg Val His
625 630 635 640
Tyr Ser Arg Gly Glu Tyr Gly Ala Leu Val Ile Asp Ala Thr Ala Glu
645 650 655
Thr Glu Lys Gly Ser Tyr Ser Val Lys Arg Tyr Val Gly Asp Ser Lys
660 665 670
Ala Leu His Glu Thr Ile Ala Thr His Gln Thr Asn Val Gly Ala Arg
675 680 685
Glu Glu Lys Ile Glu Tyr Arg Arg Glu Asp Asp Arg Phe His Thr Gly
690 695 700
Tyr Thr Val Thr Asp Ser Leu Lys Ser Val Glu Glu Ile Ile Gly Ser
705 710 715 720
Gln Phe Asn Asp Ile Phe Lys Gly Ser Gln Phe Asp Asp Val Phe His
725 730 735
Gly Gly Asn Gly Val Asp Thr Ile Asp Gly Asn Asp Gly Asp Asp His
740 745 750
Leu Phe Gly Gly Ala Gly Asp Asp Val Ile Asp Gly Gly Asn Gly Asn
755 760 765
Asn Phe Leu Val Gly Gly Thr Gly Asn Asp Ile Ile Ser Gly Gly Lys
770 775 780
Asp Asn Asp Ile Tyr Val His Lys Thr Gly Asp Gly Asn Asp Ser Ile
785 790 795 800
Thr Asp Ser Gly Gly Gln Asp Lys Leu Ala Phe Ser Asp Val Asn Leu
805 810 815
Lys Asp Leu Thr Phe Lys Lys Val Asp Ser Ser Leu Glu Ile Ile Asn
820 825 830
Gln Lys Gly Glu Lys Val Arg Ile Gly Asn Trp Phe Leu Glu Asp Asp
835 840 845
Leu Ala Ser Thr Val Ala Asn Tyr Lys Ala Thr Asn Asp Arg Lys Ile
850 855 860
Glu Glu Ile Ile Gly Lys Gly Gly Glu Arg Ile Thr Ser Glu Gln Val
865 870 875 880
Asp Lys Leu Ile Lys Glu Gly Asn Asn Gln Ile Ser Ala Glu Ala Leu
885 890 895
Ser Lys Val Val Asn Asp Tyr Asn Thr Ser Lys Asp Arg Gln Asn Val
900 905 910
Ser Asn Ser Leu Ala Lys Leu Ile Ser Ser Val Gly Ser Phe Thr Ser
915 920 925
Ser Ser Asp Phe Arg Asn Asn Leu Gly Thr Tyr Val Pro Ser Ser Ile
930 935 940
Asp Val Ser Asn Asn Ile Gln Leu Ala Arg Ala Ala
945 950 955
<210> 6
<211> 1052
<212> Белок
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIIA K571A K702A
<400> 6
Met Ser Thr Trp Ser Ser Met Leu Ala Asp Leu Lys Lys Arg Ala Glu
1 5 10 15
Glu Ala Lys Arg Gln Ala Lys Lys Gly Tyr Asp Val Thr Lys Asn Gly
20 25 30
Leu Gln Tyr Gly Val Ser Gln Ala Lys Leu Gln Ala Leu Ala Ala Gly
35 40 45
Lys Ala Val Gln Lys Tyr Gly Asn Lys Leu Val Leu Val Ile Pro Lys
50 55 60
Glu Tyr Asp Gly Ser Val Gly Asn Gly Phe Phe Asp Leu Val Lys Ala
65 70 75 80
Ala Glu Glu Leu Gly Ile Gln Val Lys Tyr Val Asn Arg Asn Glu Leu
85 90 95
Glu Val Ala His Lys Ser Leu Gly Thr Ala Asp Gln Phe Leu Gly Leu
100 105 110
Thr Glu Arg Gly Leu Thr Leu Phe Ala Pro Gln Leu Asp Gln Phe Leu
115 120 125
Gln Lys His Ser Lys Ile Ser Asn Val Val Gly Ser Ser Thr Gly Asp
130 135 140
Ala Val Ser Lys Leu Ala Lys Ser Gln Thr Ile Ile Ser Gly Ile Gln
145 150 155 160
Ser Val Leu Gly Thr Val Leu Ala Gly Ile Asn Leu Asn Glu Ala Ile
165 170 175
Ile Ser Gly Gly Ser Glu Leu Glu Leu Ala Glu Ala Gly Val Ser Leu
180 185 190
Ala Ser Glu Leu Val Ser Asn Ile Ala Lys Gly Thr Thr Thr Ile Asp
195 200 205
Ala Phe Thr Thr Gln Ile Gln Asn Phe Gly Lys Leu Val Glu Asn Ala
210 215 220
Lys Gly Leu Gly Gly Val Gly Arg Gln Leu Gln Asn Ile Ser Gly Ser
225 230 235 240
Ala Leu Ser Lys Thr Gly Leu Gly Leu Asp Ile Ile Ser Ser Leu Leu
245 250 255
Ser Gly Val Thr Ala Ser Phe Ala Leu Ala Asn Lys Asn Ala Ser Thr
260 265 270
Ser Thr Lys Val Ala Ala Gly Phe Glu Leu Ser Asn Gln Val Ile Gly
275 280 285
Gly Ile Thr Lys Ala Val Ser Ser Tyr Ile Leu Ala Gln Arg Leu Ala
290 295 300
Ala Gly Leu Ser Thr Thr Gly Pro Ala Ala Ala Leu Ile Ala Ser Ser
305 310 315 320
Ile Ser Leu Ala Ile Ser Pro Leu Ala Phe Leu Arg Val Ala Asp Asn
325 330 335
Phe Asn Arg Ser Lys Glu Ile Gly Glu Phe Ala Glu Arg Phe Lys Lys
340 345 350
Leu Gly Tyr Asp Gly Asp Lys Leu Leu Ser Glu Phe Tyr His Glu Ala
355 360 365
Gly Thr Ile Asp Ala Ser Ile Thr Thr Ile Ser Thr Ala Leu Ser Ala
370 375 380
Ile Ala Ala Gly Thr Ala Ala Ala Ser Ala Gly Ala Leu Val Gly Ala
385 390 395 400
Pro Ile Thr Leu Leu Val Thr Gly Ile Thr Gly Leu Ile Ser Gly Ile
405 410 415
Leu Glu Phe Ser Lys Gln Pro Met Leu Asp His Val Ala Ser Lys Ile
420 425 430
Gly Asn Lys Ile Asp Glu Trp Glu Lys Lys Tyr Gly Lys Asn Tyr Phe
435 440 445
Glu Asn Gly Tyr Asp Ala Arg His Lys Ala Phe Leu Glu Asp Ser Phe
450 455 460
Ser Leu Leu Ser Ser Phe Asn Lys Gln Tyr Glu Thr Glu Arg Ala Val
465 470 475 480
Leu Ile Thr Gln Gln Arg Trp Asp Glu Tyr Ile Gly Glu Leu Ala Gly
485 490 495
Ile Thr Gly Lys Gly Asp Lys Leu Ser Ser Gly Lys Ala Tyr Val Asp
500 505 510
Tyr Phe Gln Glu Gly Lys Leu Leu Glu Lys Lys Pro Asp Asp Phe Ser
515 520 525
Lys Val Val Phe Asp Pro Thr Lys Gly Glu Ile Asp Ile Ser Asn Ser
530 535 540
Gln Thr Ser Thr Leu Leu Lys Phe Val Thr Pro Leu Leu Thr Pro Gly
545 550 555 560
Thr Glu Ser Arg Glu Arg Thr Gln Thr Gly Ala Tyr Glu Tyr Ile Thr
565 570 575
Lys Leu Val Val Lys Gly Lys Asp Lys Trp Val Val Asn Gly Val Lys
580 585 590
Asp Lys Gly Ala Val Tyr Asp Tyr Thr Asn Leu Ile Gln His Ala His
595 600 605
Ile Ser Ser Ser Val Ala Arg Gly Glu Glu Tyr Arg Glu Val Arg Leu
610 615 620
Val Ser His Leu Gly Asn Gly Asn Asp Lys Val Phe Leu Ala Ala Gly
625 630 635 640
Ser Ala Glu Ile His Ala Gly Glu Gly His Asp Val Val Tyr Tyr Asp
645 650 655
Lys Thr Asp Thr Gly Leu Leu Val Ile Asp Gly Thr Lys Ala Thr Glu
660 665 670
Gln Gly Arg Tyr Ser Val Thr Arg Glu Leu Ser Gly Ala Thr Lys Ile
675 680 685
Leu Arg Glu Val Ile Lys Asn Gln Lys Ser Ala Val Gly Ala Arg Glu
690 695 700
Glu Thr Leu Glu Tyr Arg Asp Tyr Glu Leu Thr Gln Ser Gly Asn Ser
705 710 715 720
Asn Leu Lys Ala His Asp Glu Leu His Ser Val Glu Glu Ile Ile Gly
725 730 735
Ser Asn Gln Arg Asp Glu Phe Lys Gly Ser Lys Phe Arg Asp Ile Phe
740 745 750
His Gly Ala Asp Gly Asp Asp Leu Leu Asn Gly Asn Asp Gly Asp Asp
755 760 765
Ile Leu Tyr Gly Asp Lys Gly Asn Asp Glu Leu Arg Gly Asp Asn Gly
770 775 780
Asn Asp Gln Leu Tyr Gly Gly Glu Gly Asn Asp Lys Leu Leu Gly Gly
785 790 795 800
Asn Gly Asn Asn Tyr Leu Ser Gly Gly Asp Gly Asn Asp Glu Leu Gln
805 810 815
Val Leu Gly Asn Gly Phe Asn Val Leu Arg Gly Gly Lys Gly Asp Asp
820 825 830
Lys Leu Tyr Gly Ser Ser Gly Ser Asp Leu Leu Asp Gly Gly Glu Gly
835 840 845
Asn Asp Tyr Leu Glu Gly Gly Asp Gly Ser Asp Phe Tyr Val Tyr Arg
850 855 860
Ser Thr Ser Gly Asn His Thr Ile Tyr Asp Gln Gly Lys Ser Ser Asp
865 870 875 880
Leu Asp Lys Leu Tyr Leu Ser Asp Phe Ser Phe Asp Arg Leu Leu Val
885 890 895
Glu Lys Val Asp Asp Asn Leu Val Leu Arg Ser Asn Glu Ser Ser His
900 905 910
Asn Asn Gly Val Leu Thr Ile Lys Asp Trp Phe Lys Glu Gly Asn Lys
915 920 925
Tyr Asn His Lys Ile Glu Gln Ile Val Asp Lys Asn Gly Arg Lys Leu
930 935 940
Thr Ala Glu Asn Leu Gly Thr Tyr Phe Lys Asn Ala Pro Lys Ala Asp
945 950 955 960
Asn Leu Leu Asn Tyr Ala Thr Lys Glu Asp Gln Asn Glu Ser Asn Leu
965 970 975
Ser Ser Leu Lys Thr Glu Leu Ser Lys Ile Ile Thr Asn Ala Gly Asn
980 985 990
Phe Gly Val Ala Lys Gln Gly Asn Thr Gly Ile Asn Thr Ala Ala Leu
995 1000 1005
Asn Asn Glu Val Asn Lys Ile Ile Ser Ser Ala Asn Thr Phe Ala Thr
1010 1015 1020
Ser Gln Leu Gly Gly Ser Gly Met Gly Thr Leu Pro Ser Thr Asn Val
1025 1030 1035 1040
Asn Ser Met Met Leu Gly Asn Leu Ala Arg Ala Ala
1045 1050
<210> 7
<211> 3078
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIA дикого типа
<400> 7
ggatccgcaa attctcaact ggaccgcgta aagggcctca ttgacagcct caaccaacac 60
accaaatcgg ctgcgaaatc cggtgcgggt gcgctgaaaa atggtctggg tcaggtgaaa 120
caggcgggtc agaaactgat cttatacatc ccgaaagatt atcaggcttc aaccggctcc 180
agtcttaatg atttggtaaa agccgccgaa gcactgggta ttgaagtaca tcgcagcgaa 240
aaaaatggga ctgcgttagc gaaagagctg tttggcacaa cggaaaagct gctgggcttt 300
agtgaacgtg gcattgcgct gtttgcacca cagttcgata aattgttaaa taagaatcag 360
aaactgagca agtccttagg cggctcatca gaagccctgg gtcagcgtct gaacaaaact 420
cagacagccc tgagtgcatt acagtcattt ctcgggacgg caattgcagg catggatctg 480
gatagcctgc ttcgccgtcg gcgtaacggt gaggatgtct cgggcagcga gctggcaaaa 540
gccggcgttg acctggcagc ccagttggtt gacaacatcg catctgccac cggcactgtt 600
gacgcttttg ccgaacaact gggcaaatta gggaacgcgc tgtctaacac gcgcttatct 660
gggctcgcat ccaaactcaa caacttgccg gacttgtcgc tggcaggacc tggctttgac 720
gcggtttcag gcattctgtc cgtggttagt gcttcgttca ttcttagcaa caaagacgcc 780
gatgcgggta ccaaggcggc cgctgggatt gaaatcagca cgaaaatctt gggtaatatt 840
ggcaaggcag tgtcgcagta tatcatcgct caacgtgtgg ccgcaggcct cagcaccacc 900
gcggccacag gtggcctgat tggtagtgtc gtcgctctgg ccatttctcc cctgtcgttc 960
ctgaacgttg cggacaaatt tgaacgcgct aaacagctgg aacagtattc tgaacgcttt 1020
aaaaaatttg gatatgaagg cgatagcctg ctggcatcct tttaccgcga aaccggtgcg 1080
attgaagccg cattgaccac cattaactcg gtactcagtg ctgccagtgc aggggtggga 1140
gcggcagcca caggttctct ggtcggagcc ccggttgctg ccttagtgtc tgccattacc 1200
gggattatct cgggcatcct tgatgcgtct aaacaggcga ttttcgaacg cgtggcgacg 1260
aaactggcga acaaaattga tgaatgggaa aagaaacacg gtaagaacta ctttgagaac 1320
ggctatgatg cccgtcatag tgcgtttctc gaagatacct ttgaactttt gtcgcaatac 1380
aacaaagagt atagcgtgga gcgcgtagtt gctattacac agcagcgttg ggatgtgaat 1440
attggggaat tggcggggat cacccgtaaa ggcgcggatg caaaatccgg taaagcttac 1500
gtcgatttct tcgaggaagg gaaactgctg gaaaaggacc cggatcgctt tgataagaaa 1560
gtgttcgacc cgcttgaggg taaaattgac ttgtcatcca ttaacaaaac gacgttgctg 1620
aaattcatta cgcccgtgtt tacggcggga gaagaaattc gcgaacgcaa acagactggg 1680
aaatacgagt acatgactga actgtttgtt aaagggaagg aaaagtgggt agtcaccggt 1740
gttcagtcgc acaatgcgat ttacgattat actaacctga ttcagctggc aatcgacaag 1800
aaaggtgaaa aacgccaagt gaccatcgag agtcaccttg gcgaaaagaa cgatcgtatc 1860
tatctgagca gtggcagctc aatcgtctat gccggcaacg gtcatgacgt tgcctattac 1920
gataaaacgg ataccggata tctgacgttt gacggtcaaa gcgcgcagaa agcgggtgaa 1980
tatatcgtca ctaaggaatt gaaagccgac gttaaagtcc tgaaagaggt ggtcaaaacc 2040
caagatatct cagtaggcaa acgcagcgag aaactggagt accgcgatta tgagctgagc 2100
ccgtttgaac ttggtaacgg gattcgtgcg aaagatgaac tgcatagcgt ggaagagatt 2160
attggatcga atcggaaaga caaattcttt ggtagccgct tcaccgatat ctttcatggc 2220
gccaaaggcg acgacgagat ttatgggaat gatgggcatg atatcctgta tggcgacgat 2280
ggcaacgatg tgattcatgg aggcgacggt aatgatcact tagtgggcgg caatgggaat 2340
gaccgtctga tcggcgggaa aggcaataat ttcctgaatg gcggtgatgg cgatgatgaa 2400
cttcaggtgt tcgaggggca atataatgtg ctgttaggcg gcgccggcaa cgatatttta 2460
tacggtagtg atggtaccaa cctcttcgat ggtggcgtgg gtaatgataa aatctatggt 2520
ggcctgggca aagatatcta tcgctattct aaagaatacg gtcggcatat tatcattgaa 2580
aaaggtggtg acgacgacac cctgctgttg agcgatcttt ccttcaagga cgtcggtttc 2640
attcgtattg gagatgatct gctggtgaac aaacggatcg gcggcaccct gtattaccac 2700
gaggattaca acggtaatgc gcttacaatc aaagattggt ttaaagaggg taaggaaggt 2760
cagaataaca aaattgaaaa gatcgttgac aaagatggag cttatgtcct gtcccagtat 2820
ttgaccgaac tcacggcgcc tggtcgtggc attaattact tcaacggatt agaagagaag 2880
ttatactacg gtgaaggcta caacgcgctc ccgcaactgc gcaaagatat tgaacaaatc 2940
atctccagca ccggagcttt aacgggcgaa catggccaag ttctggtagg agcgggtggc 3000
ccactggcgt attcaaattc tccgaacagc attccaaacg cgttttcgaa ttatctgact 3060
cagagcgctt aagagctc 3078
<210> 8
<211> 2880
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIA дикого типа
<400> 8
ggatcctcga aaattacctt gtccagcctg aaatccagct tacagcaagg tctgaagaat 60
gggaagaaca aactgaacca agccggtacc acgctgaaga atggattgac gcaaacaggt 120
catagtctgc agaatggtgc gaagaaatta atcctctaca ttccgcaggg ctatgactca 180
ggccaaggca acggggtcca ggacctcgta aaagccgcga atgatctggg cattgaggtg 240
tggcgcgaag aacgtagtaa cctggatatt gcgaaaactt cgttcgatac cacccagaaa 300
attctgggat tcaccgaccg gggcatcgtg ttgttcgcac cccagctgga taatctcctg 360
aagaagaacc cgaaaatcgg caacacgctg ggcagcgcat caagcatctc gcagaacatc 420
ggcaaagcca ataccgtctt aggtggaatc cagagcattc tgggtagcgt gctttctggc 480
gtgaacttaa acgaattact tcagaacaaa gacccgaatc agctggaact ggcgaaagct 540
ggactggaac ttacgaacga actggtaggt aacattgcca gctctgtaca aaccgttgat 600
gcgtttgcgg agcagatcag taaactgggt tctcatttgc aaaatgttaa agggctgggt 660
ggtctgagta acaaattgca gaatctgccg gacctgggca aggcttcttt ggggctggat 720
attatctcgg ggctgttgtc tggcgcaagc gctggcctga ttttagcgga taaggaggct 780
tcgaccgaaa agaaggccgc tgccggcgta gaatttgcga accagattat tgggaacgtg 840
accaaagcgg tgagtagcta tattttggcg caacgtgttg cctccggttt gagtagtact 900
ggtccagtcg cagcactgat tgccagtact gtggcactgg cggttagtcc actgtccttt 960
ctgaatgtcg cggataaatt taaacaagcc gatctcatca aatcttatag tgaacgtttc 1020
cagaaattag gttatgatgg cgatcgctta ctggcggact ttcaccggga aacaggcacc 1080
attgatgcaa gcgtgaccac gattaatacc gcgcttgcag caatcagcgg tggtgttggg 1140
gcagcgagtg ctggaagcct ggttggcgcg cctgttgctc tgcttgtagc gggtgtaacg 1200
ggcttaatca caacgatcct cgaatattcc aaacaagcga tgtttgagca tgtcgctaac 1260
aaagtccacg accgcatcgt ggaatgggaa aaaaaacaca ataaaaacta ctttgagcag 1320
ggctacgaca gccgtcattt ggcagatctg caggacaata tgaaattcct catcaactta 1380
aacaaagaac ttcaggctga acgtgtcgtg gcgattacgc aacaacgttg ggataaccag 1440
attggcgacc tggcggccat ttcccgtcgc actgacaaaa tttcctcggg taaagcatat 1500
gtggacgcgt ttgaggaagg tcagcaccag agctatgata gttcagtgca gttagacaat 1560
aaaaatggta ttattaacat ttcaaacaca aaccgcaaga cccaatcagt actgtttcgc 1620
acgccgttac tgaccccggg tgaagaaaac cgcgagcgca ttcaggaggg taagaactcc 1680
tacattacca aacttcacat tcagcgcgta gattcatgga cggtgacgga tggcgatgcg 1740
tcatcgagcg tagatttcac caatgtggtt cagcgtattg cggtgaaatt cgatgatgca 1800
ggaaacatta tcgagtcaaa agatactaaa attatcgcca atctcggtgc cggtaatgac 1860
aatgtctttg taggcagcag cacgacggtg attgacggcg gtgacggcca tgatcgcgtt 1920
cattattcgc gtggcgaata cggcgctctg gtcattgacg caactgccga gacagagaaa 1980
ggtagctact ccgttaaacg ttatgttggt gactcaaaag cgctgcacga aacgattgcc 2040
actcatcaga ccaatgtggg gaatcgcgaa gagaaaatcg agtatcgccg cgaagatgat 2100
cggttccata ccgggtatac tgtgaccgat tcgctgaaaa gcgtcgaaga aattatcgga 2160
tctcaattca atgacatctt caaaggttcc cagtttgatg acgtgtttca tggcggtaat 2220
ggcgttgata ccattgacgg caacgatggc gacgaccatc tgtttggcgg ggctggtgat 2280
gacgtgatcg atggcggcaa cggcaacaac tttctggtgg gtggtaccgg gaacgatatc 2340
atttcaggcg gcaaagataa cgacatctac gttcacaaga caggggatgg gaatgatagc 2400
attactgatt cgggtggcca agataagctg gccttttctg acgttaatct gaaagatctg 2460
actttcaaaa aggtggattc tagcctggaa atcattaatc agaaaggaga gaaggtccgc 2520
attggaaatt ggtttctcga agatgatctt gcctcgaccg tggcaaacta caaagcgacc 2580
aatgaccgca aaatcgaaga aatcattggg aaaggcggtg aacgcatcac ctctgaacag 2640
gtggataaac tgatcaaaga aggcaataac cagatttccg ctgaagcgtt gtctaaagtc 2700
gtcaacgatt acaacacgtc caaagatcgc caaaacgtta gcaattcact tgccaaactg 2760
atctcgagcg ttggatcttt tacatcctcg agcgatttcc gtaacaacct cggcacctat 2820
gttccttctt cgattgatgt cagtaataat atccaactgg cccgtgccgc ctaagagctc 2880
<210> 9
<211> 3168
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIIA дикого типа
<400> 9
ggatcctcta cctggtcctc catgctggct gatctgaaga agcgtgcgga agaagccaaa
60
cgtcaggcga aaaaaggtta tgacgtcacg aagaacggcc tgcagtacgg tgttagtcaa
120
gccaaactgc aggctctggc agccggtaaa gctgttcaga aatacggtaa caaacttgtg
180
ttagtaattc cgaaagaata tgatgggtcg gtgggtaatg gcttcttcga cctggtcaaa
240
gccgcggaag aactgggcat tcaggtgaaa tatgtcaacc gcaacgaact cgaggttgcg
300
cataaatctc tgggaacggc ggatcagttt ctgggtctga ctgagcgtgg cctgaccctg
360
tttgcaccgc agcttgacca gttcttacag aaacacagca aaatctcaaa tgtggttggt
420
tcttcaactg gtgacgcggt gagcaaactg gcaaagtctc agacaatcat ttctggaatc
480
cagtcggtgt tgggcactgt actcgctggg atcaacttaa atgaagccat catttcaggc
540
ggctctgaat tggagctggc ggaagctggc gtaagcctgg cgtcggaact ggtgtcgaat
600
atcgccaaag gcaccacgac aattgatgcg tttaccaccc aaattcagaa ttttggaaaa
660
ctggtcgaaa acgcgaaggg cctgggtggc gtgggtcgtc agctgcaaaa catttccggt
720
agcgccttga gcaaaactgg tttgggctta gacatcatca gcagcttgct gagtggagtc
780
acagcgagct ttgcgctggc aaacaagaat gcctctacct ccaccaaagt tgctgccggt
840
tttgaactgt cgaatcaagt tattggaggc attaccaaag cggtcagctc atatatcctg
900
gcccaacgtc tggcggcggg tttgagcacg actggacccg ctgcagcgct gattgcatct
960
agtattagtc tggccatcag cccgctggcg tttctgcgcg tagctgataa tttcaaccgc
1020
agcaaagaaa tcggtgagtt cgcggaacgc tttaagaaat taggctatga tggggataag
1080
ctgctgtctg aattttacca tgaggctggc accattgacg catctatcac gacaatttcg
1140
acagccctgt cagcgattgc cgcagggact gcagccgctt cagccggcgc cctggttggt
1200
gcaccgatta cgcttcttgt cacgggtatt accggcttga ttagcggcat cctggaattt
1260
tccaaacagc cgatgctgga ccatgtagca agtaaaattg gcaacaaaat cgacgagtgg
1320
gaaaagaaat atggtaagaa ctacttcgag aatggttatg atgctcgcca taaagccttc
1380
ttggaagatt cgttcagcct gctcagttcc ttcaataagc aatatgaaac cgaacgcgcc
1440
gtgctgatca cccaacagcg ctgggatgag tacattgggg aactggcggg tattacgggt
1500
aagggtgaca aattatcaag cgggaaggca tacgttgatt attttcagga gggtaaactt
1560
ctggagaaga aacccgacga tttcagcaaa gtagtgtttg atccgacaaa aggcgagatc
1620
gatatcagca attcccagac ctctaccctg ctgaaatttg tgacccctct cctgacccca
1680
ggaactgaaa gccgtgaacg cactcaaacg ggaaagtacg aatacattac taaactcgtg
1740
gtaaaaggga aagataaatg ggtcgtgaac ggcgtgaaag ataaaggcgc ggtatacgat
1800
tacaccaact taattcagca tgctcacatt tcctcttccg tggcgcgtgg cgaagagtat
1860
cgggaagtcc gcctggttag ccatttgggt aatggcaatg ataaagtctt tctggctgcg
1920
ggctcggcag aaattcacgc cggcgaaggc catgatgtcg tatattacga taaaacggac
1980
accggactcc tggtgatcga tggcacaaaa gcaaccgagc aagggcgcta tagtgttacg
2040
cgtgagctga gtggcgcgac caaaattttg cgtgaagtga tcaaaaatca gaaatcggca
2100
gttggcaagc gcgaagaaac gttagagtat cgcgattacg aactcacgca aagcggtaac
2160
agtaatctga aagcacacga cgaactgcat tctgttgaag aaattattgg cagcaatcag
2220
cgcgatgaat tcaaaggctc gaagtttcgt gatatctttc atggtgcaga cggggatgac
2280
ctcctgaacg gcaatgatgg ggatgacatt ttatatggag acaaagggaa cgatgaactg
2340
cgcggcgata acgggaacga ccaactgtac ggtggtgagg ggaatgataa actgttgggc
2400
gggaacggca ataactattt atcgggcgga gatggtaacg atgaacttca ggttctgggg
2460
aacggtttca acgtccttcg tggaggcaaa ggtgacgata aactttatgg tagcagcggc
2520
tccgacctcc ttgatggtgg agaaggcaac gattacttag aaggtggtga tgggagtgat
2580
ttctatgtgt atcggagcac cagtggcaat cacaccattt atgatcaggg taaatcctcc
2640
gatctggaca aattatacct gtcagatttc tcgtttgacc gcctcttggt ggagaaagtg
2700
gatgacaacc tcgttttgcg ttccaatgag agtagccaca ataatggcgt tttaaccatc
2760
aaagactggt ttaaagaagg gaacaagtac aaccacaaaa tcgagcagat tgtcgataag
2820
aatggccgca aactgacggc cgaaaatctg ggtacctatt ttaaaaacgc cccaaaagcg
2880
gacaaccttc tgaactatgc gaccaaagag gaccagaacg aatcaaattt atcttcattg
2940
aaaacggaac tgagtaaaat tatcactaat gcgggtaatt ttggcgttgc caaacaaggt
3000
aacaccggca ttaataccgc agcgcttaac aacgaagtga acaaaatcat ttcgtcagcg
3060
aacaccttcg ctacgagtca actcggtggc tcggggatgg gcacactgcc ttccacgaac
3120
gtgaatagca tgatgctggg caatcttgcc cgggcggcgt aagagctc
3168
<210> 10
<211> 3078
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIA K560A K686A
<400> 10
ggatccgcta attctcagtt agatcgtgtg aaaggattaa tcgatagcct gaaccagcac 60
accaaatccg cagcaaaatc aggagctggc gctctgaaga atggactggg ccaggtgaaa 120
caggcagggc agaaactgat tctctatatt ccgaaagact atcaagcttc caccggttcc 180
agcttgaatg atttggtgaa agcagcggaa gctctgggaa ttgaagtcca tcgtagcgag 240
aagaacggaa ccgcattggc gaaagagtta tttgggacca cggaaaaact gttaggcttt 300
agtgaacggg gtattgcact gtttgcacct cagtttgaca aactgcttaa caaaaaccag 360
aagttaagca aatcactggg tggcagctcc gaagccttgg gccagcgcct gaacaaaacc 420
cagacggcat tgagtgcact gcaaagtttt ctgggcacgg ctatcgccgg aatggatctg 480
gattctctcc tgcgtcggcg tcgcaacggt gaagatgtaa gtggatcgga gctggcgaaa 540
gctggtgtcg atctcgcggc acaattggtt gacaatattg cgtcggccac aggaacggtg 600
gatgcgtttg cggaacaact cggtaaactg ggcaacgcgt tatctaacac acgcttatcc 660
ggcttagcct ctaagctgaa taaccttccg gatctgtcct tggccggccc cggctttgat 720
gcggttagtg gcattcttag cgttgtgtcg gcgagtttca ttctgtccaa caaagatgca 780
gatgcgggca ccaaagccgc cgcgggcatt gaaatttcga cgaaaattct tggtaacatc 840
ggcaaggctg tgtctcagta tatcattgct caacgtgtgg ccgccgggtt aagcactaca 900
gcagcgacag gtggcctgat tggctctgtg gtagcactcg caattagtcc gctgtcattt 960
ctgaatgtcg cggataaatt tgaacgtgcc aaacaactgg aacagtatag cgaacgcttc 1020
aagaaatttg gttatgaagg ggatagtttg cttgcgagct tctaccgcga aaccggtgcc 1080
attgaagcgg ccctgaccac gatcaactcg gtgttgtcag ccgccagcgc cggggtcggt 1140
gcggcagcga cgggttcctt ggtaggtgca ccagttgcgg cgcttgtatc agccattact 1200
ggcatcatct ccggcattct ggatgccagc aagcaagcta tcttcgaacg tgtggcgacg 1260
aaactggcga acaaaattga tgaatgggag aaaaagcacg gcaaaaatta cttcgaaaac 1320
ggttatgatg cgcgccattc ggcgtttctg gaggatacat ttgaacttct gagccagtac 1380
aacaaagaat actctgtgga gcgtgtcgta gcgatcaccc agcaacggtg ggatgttaac 1440
attggtgaac tggccggcat tacccgcaaa ggcgctgacg cgaaatcagg caaagcatac 1500
gtcgacttct ttgaagaggg gaaactgttg gaaaaggacc cggaccgttt tgataagaaa 1560
gtctttgatc cgctggaagg gaaaattgat ctgtcctcta tcaacaaaac tacgttgctg 1620
aaattcatca cgcccgtttt cacagcgggt gaggaaattc gcgagcgtaa gcagactggg 1680
gcgtatgaat acatgaccga actgttcgtc aaaggtaaag agaaatgggt ggtgaccggc 1740
gttcagtcgc ataacgccat ctacgattat accaacttaa tccagctggc gattgacaaa 1800
aaaggtgaaa agcgtcaggt gaccattgag agccatctgg gcgagaagaa tgaccgcatc 1860
tatttaagca gtggtagttc gattgtctat gctggcaatg ggcatgatgt cgcctattat 1920
gacaaaaccg atacgggcta tttgaccttt gacggtcaga gcgcccagaa agcaggcgaa 1980
tatatcgtaa ccaaagaact gaaagctgat gtgaaagtcc tgaaagaggt tgtgaaaacg 2040
caagatatta gcgttggcgc ccgctcggag aaattagaat atcgcgatta tgagctgagc 2100
ccattcgaac tcggtaacgg cattcgcgcg aaagatgaac tccactctgt ggaagaaatt 2160
attggttcta atcgtaaaga taaattcttc gggtcacgct ttaccgatat ctttcatggc 2220
gcgaaagggg acgatgaaat ctatggcaac gacggccacg acattctgta tggtgatgat 2280
gggaacgacg ttatccatgg aggggatggc aatgatcatc tggttggcgg caatggaaat 2340
gaccgtctta tcggcggcaa aggtaataac tttcttaacg gtggtgatgg ggacgacgaa 2400
cttcaggtct ttgaaggtca gtataacgta ctgctgggcg gtgctgggaa cgacatcctt 2460
tatggtagtg atggtactaa tctgttcgac ggcggcgtgg gtaacgacaa gatctacgga 2520
gggttgggta aggacattta ccgctactcg aaagaatacg gccggcacat cattattgag 2580
aaaggcggag atgacgatac cctcctgctg tcggatttaa gctttaaaga cgtgggcttc 2640
attcgcatcg gggatgatct gttagtgaac aaacgcattg gaggcacgct gtactaccac 2700
gaagattaca atggcaatgc tctgaccatt aaggactggt tcaaagaagg caaggaaggt 2760
caaaacaata agatcgagaa aatcgtagat aaagacggcg cctatgttct cagccagtat 2820
ctcactgaac tgactgcacc tggccgcggt attaattact tcaatggtct ggaagagaaa 2880
ctgtactatg gcgagggtta taacgccctg ccacaactgc gcaaagacat tgaacagatt 2940
atcagctcta ccggggcgct gactggtgag catggtcaag ttctcgttgg tgcgggtggt 3000
ccgctggcgt actcaaattc gccgaatagc atcccgaatg ccttttccaa ttacctgacc 3060
cagtcagcat aagagctc 3078
<210> 11
<211> 2880
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIA S148A K557A K686A
<400> 11
ggatccagca aaattaccct ctcatccctc aaatcaagcc tgcagcaggg gctgaagaac 60
ggcaagaaca agctgaatca ggcgggtacg actctgaaga atggcttaac gcaaacaggg 120
cactcattac agaacggtgc gaaaaaactg atcctgtaca tcccgcaagg ttacgactcg 180
gggcagggca atggggtcca agatcttgtt aaagcggcca atgacctggg aatcgaagtg 240
tggcgtgaag aacggagcaa cctggacatt gcgaaaacca gctttgatac gacccagaaa 300
atcctgggtt tcaccgaccg cggaattgta ctgtttgccc cgcaacttga taaccttctg 360
aaaaaaaacc cgaaaattgg aaatacattg ggttcggcaa gctctatcag tcagaacatt 420
ggcaaagcta acactgtgtt aggtggtatt cagtctattc tggggtcggt cttgtccggc 480
gttaacctga atgaactgct tcaaaacaaa gatccaaatc aactggaact ggccaaagcg 540
ggcctggagt taacgaacga gttggtagga aacattgcct cctctgtgca gacggtggat 600
gcatttgctg aacagattag caaattaggt tctcatctgc aaaacgtcaa aggactgggc 660
ggcctcagca acaaactgca gaatttaccg gatctgggca aagccagcct tgggttagat 720
attatctccg ggctcttgtc aggcgcgtca gccggtctta tcctggcgga caaagaagca 780
agcaccgaaa agaaagctgc cgcaggcgtc gagtttgcga accagattat tggaaatgtc 840
accaaagcgg tcagttccta catcctggcg cagcgtgtag cgtcgggcct gtcatctact 900
ggtccagttg ccgcgttgat tgcgtctacg gtcgccttag cggtttcgcc gctgtccttt 960
ctgaacgtgg cggataagtt caaacaagcg gatttgatca aaagctatag tgaacgcttt 1020
cagaaattag gttatgatgg tgaccgtctg ctggcggatt tccaccgcga aaccgggacc 1080
attgatgcct ctgtcactac catcaatacg gctttggcag cgatctccgg tggggtgggc 1140
gcggcgagcg ctggctcact ggtaggagcc cctgttgctc ttttggttgc aggcgttaca 1200
ggcctgatta cgaccatttt agagtatagc aaacaagcga tgtttgagca cgtcgcaaat 1260
aaggtacacg atcgtattgt agagtgggaa aagaaacata acaagaatta cttcgaacag 1320
gggtatgatt cacgccatct ggcagatctt caggacaaca tgaaatttct gatcaatctg 1380
aacaaagagc tgcaagcaga acgcgttgtg gcgattacac aacagcgttg ggataaccag 1440
attggcgatc tggcagcaat ctcgcgccgt acggacaaaa tttcttcggg taaagcctac 1500
gtggacgcct ttgaagaagg acagcatcag tcttatgatt cctcggtaca gctggacaac 1560
aaaaatggaa tcatcaacat tagtaatacc aaccgcaaga cccaaagtgt cctgtttcgc 1620
actcccctgc tcaccccggg cgaagagaac cgcgaacgca ttcaagaggg tgcaaactca 1680
tacattacca aattgcacat tcagcgtgtg gatagttgga ctgtcactga cggtgatgcg 1740
agcagcagtg ttgactttac taacgtggtt caacgcattg ccgtcaaatt cgacgacgct 1800
gggaatatta ttgaaagcaa agatacgaaa attatcgcta atttgggcgc tggtaatgat 1860
aatgtgttcg tgggttccag caccacggtt attgatggtg gcgatgggca tgaccgcgtg 1920
cattactcgc gtggcgaata tggcgccctt gtaatcgacg ccaccgcgga aacagagaaa 1980
ggtagctatt cggtgaaacg ctatgtgggt gattctaaag cgttacacga aaccattgcc 2040
acacatcaga ccaacgtggg cgctcgtgaa gagaaaatcg aatatcggcg tgaagatgat 2100
cgttttcata caggctacac cgtaacggac tctctcaaaa gcgttgaaga aattatcggc 2160
agccagttca atgatatctt caaaggctca caattcgacg atgtgttcca tggaggtaat 2220
ggggttgaca ccattgatgg caatgacggc gatgatcatc tgtttggcgg cgctggtgac 2280
gatgtgatcg atggtggtaa cggtaataat tttctcgtcg gcggtaccgg gaacgatatt 2340
atcagcggcg gtaaggacaa tgatatttat gtgcacaaga ctggtgatgg caacgactcc 2400
atcaccgata gcggtggcca ggataaactg gcatttagcg atgtgaacct caaagatctg 2460
acctttaaaa aagtggactc tagtctggaa atcattaacc agaagggcga gaaagtccgc 2520
attggtaact ggtttctgga agatgactta gctagcaccg tggcgaacta taaagccacg 2580
aatgatcgca aaattgaaga aatcatcggc aagggcggcg agcgtattac ttcggaacag 2640
gtggataaac tgatcaaaga gggtaacaat caaattagcg ccgaagcgct gtcgaaagta 2700
gtgaacgact acaatacgag taaagatcgc cagaatgttt ccaactcgct ggccaagctg 2760
atctccagtg ttggctcatt caccagttcg agtgatttcc ggaacaatct cggtacgtat 2820
gttccttctt ccatcgacgt tagtaacaat attcagttgg cacgcgcagc ataagagctc 2880
<210> 12
<211> 3168
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIIA K571A K702A
<400> 12
ggatccagca catggagcag tatgcttgcg gacttgaaga aacgggcgga agaagcgaaa
60
cgccaagcga agaaaggata tgatgtgact aagaacggcc tgcagtacgg tgtatcgcaa
120
gctaaactgc aggccctggc ggccgggaaa gcggttcaga agtatggtaa caaactcgtc
180
cttgtgattc ccaaagaata cgatggcagc gtgggtaatg gtttctttga tctggttaaa
240
gcggccgagg aactcggcat tcaggttaaa tatgtcaatc gtaacgagct tgaggttgcg
300
cacaaatccc tgggtactgc ggatcaattc ctcggcctca cggaacgtgg tctgactctg
360
ttcgccccac agctcgatca gttcttacag aagcacagca agatttccaa tgtagtcggg
420
agtagcactg gtgatgctgt cagcaaactt gctaaatcgc agactatcat ctcgggtatt
480
cagtcagtgc tgggcacggt tctggcgggc atcaacctga acgaagctat tatcagtggt
540
ggttccgaat tagaactggc cgaagccggg gtgagtcttg cctcagaact ggtgagcaat
600
attgcgaaag gcacgaccac gattgatgct ttcaccaccc aaatccagaa ttttgggaaa
660
ttggtggaaa atgcgaaagg cctgggaggc gtaggccgcc agctgcaaaa catttcgggg
720
agcgccctct ctaaaaccgg cctggggtta gacattatct cttcgctgct gtcaggagtt
780
accgcgagct ttgccctggc aaacaagaat gccagcacgt ctactaaagt ggcagcgggt
840
tttgaactct caaatcaggt cattggtggc attactaaag ccgtatcgag ttacattctg
900
gcgcaacgtc tggctgcagg cctgtctacg accggccctg cggcagcgct gattgcgtct
960
tctatctcgt tggcgatttc gcctctggcg tttctgcgtg ttgctgataa tttcaatcgg
1020
tccaaagaaa tcggggagtt tgcagagcgt tttaagaagc tgggatatga cggtgataaa
1080
ttgctgagcg aattttacca tgaggcgggc actatcgacg cctccattac caccatttca
1140
accgcactga gcgcgatcgc agcaggaacg gcggcagctt cggcgggtgc attagtggga
1200
gcaccgatta ccctgttggt taccgggatt accgggttaa ttagcggcat tctggaattc
1260
agtaaacaac cgatgctgga tcatgtcgca agcaaaatcg gtaacaaaat tgacgaatgg
1320
gaaaaaaaat acggcaaaaa ctacttcgaa aatggctatg acgcgcgcca caaggccttt
1380
ctcgaagatt cattttcact gttgagcagc ttcaataagc agtatgaaac ggaacgtgcc
1440
gttttgatca cgcagcagcg ttgggatgaa tacattggcg aattagccgg aattacgggc
1500
aaaggtgaca aactgtcctc tggcaaagcc tatgtggact acttccaaga aggtaaactg
1560
ctcgaaaaaa aaccagatga tttctcgaaa gtggtctttg atcccacgaa gggtgagatc
1620
gacatcagca attctcagac atctaccctg ctgaagtttg ttacgccgtt gttgaccccg
1680
ggtacggaat cacgcgaacg cacacagaca ggcgcctatg agtacattac caaattagtg
1740
gtaaaaggca aagataaatg ggtagttaac ggcgtgaaag acaaaggagc agtgtatgac
1800
tacacgaact taattcagca tgcacacatt agctcctccg tcgcacgcgg tgaagagtac
1860
cgtgaagttc gccttgtaag tcatttgggt aacggaaatg ataaagtgtt cctcgccgca
1920
ggaagtgcgg aaattcatgc tggcgaaggt catgacgtcg tgtactatga taaaaccgat
1980
acaggccttc tggttattga cggcactaaa gctaccgaac aggggcgtta cagtgtgacc
2040
cgcgaacttt ccggtgctac caaaatctta cgcgaagtca tcaagaatca gaaaagtgcc
2100
gtaggcgcgc gtgaagagac actggaatat cgggactatg agttaacgca aagcgggaac
2160
agtaatttaa aggctcacga tgaactgcat tccgtagagg agattatcgg cagcaaccag
2220
cgtgatgaat ttaaaggcag caaattccgc gatatcttcc atggtgccga cggtgacgac
2280
cttcttaacg ggaatgatgg ggatgacatc ctgtatggtg acaagggcaa tgatgaactg
2340
cgtggcgata acggcaatga ccagctgtac ggtggtgagg ggaatgacaa attgttaggt
2400
ggcaacggaa acaactatct gtccggcggc gacggtaatg atgagttgca agttcttggc
2460
aatggcttta acgtgctgcg cggcggcaaa ggtgatgata aactgtatgg tagttcaggc
2520
tcagatttgc tggatggcgg ggaaggtaac gattatctgg aaggcggtga tggctctgat
2580
ttctatgtgt atcgctcgac ctctggcaac cataccattt atgaccaggg aaaatcctcg
2640
gatctggata aactctatct gtccgatttt agttttgatc gcctgctggt cgagaaagtt
2700
gatgacaatc tggttctgcg ctcgaacgaa tctagccaca acaatggtgt gcttaccatc
2760
aaagattggt ttaaagaggg taacaaatac aatcacaaaa tcgagcaaat tgtggataag
2820
aatggtcgca agctgaccgc cgagaattta gggacttact ttaaaaatgc cccgaaagcg
2880
gataatctcc tgaactatgc aaccaaggag gaccagaacg aatcgaacct gtcctctctg
2940
aaaacggaac tgagcaaaat cattaccaac gcaggcaact ttggcgtggc taaacaaggg
3000
aacacaggga tcaataccgc ggcgctgaac aacgaagtca acaaaattat cagtagcgcc
3060
aacacatttg cgacctctca actgggtggt agcggtatgg gaaccttgcc gtcaacgaat
3120
gtcaacagca tgatgctggg caacttagcc cgcgctgcat aagagctc
3168
<210> 13
<211> 527
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIC
<400> 13
tcatgagtaa aaaaattaat ggatttgagg ttttaggaga ggtggcatgg ttatgggcaa 60
gttctccttt acatcgaaag tggccgcttt ctttgttagc aattaatgtg ctacctgcga 120
ttgagagtaa tcaatatgtt ttgttaaagc gtgacggttt tcctattgca ttttgtagct 180
gggcaaattt gaatttggaa aatgaaatta aataccttga tgatgttgcc tcgctagttg 240
cggatgattg gacttccggc gatcgtcgat ggtttataga ttggatagca ccgttcggag 300
acagtgccgc attatacaaa catatgcgag ataacttccc gaatgagctg tttagggcta 360
ttcgagttga tccggactct cgagtaggga aaatttcaga atttcatgga ggaaaaattg 420
ataagaaact ggcaagtaaa atttttcaac aatatcactt tgaattaatg agtgagctaa 480
aaaataaaca aaattttaaa ttttcattag taaatagcta aagatct 527
<210> 14
<211> 488
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIC
<400> 14
acatgttaaa aaatgatttt aacgtattgg gacaaattgc ttggttatgg gcaaattctc 60
caatgcaccg aaattggtca gtttcactgt taatgaagaa tgttattcct gcaattgaaa 120
atgaccaata tttgttacta gttgatgatg gttttcctat tgcatattgc agttgggcca 180
aattaactct agagagtgag gctcgctatg taaaggacac caattcatta aaaatagatg 240
attggaatgc aggagatcgt atatggatca ttgattggat tgccccattc ggggattcat 300
ctctattgta taaacatatg agacaacgtt ttccatacga tattggaagg gcaattagaa 360
tctatcctag caaaaaagat actggaaaaa tcatatattt aaaaggagga aaaataacaa 420
aaaaagtagc tgaaaagaca tttcttcagt atgagcaaga gttaataaca gctctacaat 480
aaagatct 488
<210> 15
<211> 530
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIIC
<400> 15
tcatgagtta taaaaatgtt aaaaatttaa cagatgattt tacaacttta gggcatatcg 60
cttggttgtg ggctaattct ccgttacata aggagtggtc tatctctttg tttactaaga 120
atattttgcc agccattcaa catgatcaat atattttact tatgcgagat gagttccctg 180
tagcgttttg tagttgggca aatttaacgt taactaatga agtgaagtat gtacgtgatg 240
tgacgtcatt gacttttgaa gattggaatt caggagaacg aaaatggttg atcgactgga 300
ttgcgccatt tggggataac aatacgcttt atagatatat gcgtaaaaaa tttcctaatg 360
aagtattccg ggccattcga gtatatcctg gttctacaga agcgaaaatc attcatgttc 420
aaggaggaca aattaataaa tttacagcta aaaaattaat acaacaatat caggaagaac 480
ttattcaagt tcttaacaat cacaaaaaaa ttgtaagagg ataaagatct 530
<210> 16
<211> 639
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIC с промотором/lac-оператором и RBS
<400> 16
tctagaaaat cataaaaaat ttatttgctt tgtgagcgga taacaattat aatagattca 60
attgtgagcg gataacaatt tcacacagaa ttcattaaag aggagaaatt aaccatgagt 120
aaaaaaatta atggatttga ggttttagga gaggtggcat ggttatgggc aagttctcct 180
ttacatcgaa agtggccgct ttctttgtta gcaattaatg tgctacctgc gattgagagt 240
aatcaatatg ttttgttaaa gcgtgacggt tttcctattg cattttgtag ctgggcaaat 300
ttgaatttgg aaaatgaaat taaatacctt gatgatgttg cctcgctagt tgcggatgat 360
tggacttccg gcgatcgtcg atggtttata gattggatag caccgttcgg agacagtgcc 420
gcattataca aacatatgcg agataacttc ccgaatgagc tgtttagggc tattcgagtt 480
gatccggact ctcgagtagg gaaaatttca gaatttcatg gaggaaaaat tgataagaaa 540
ctggcaagta aaatttttca acaatatcac tttgaattaa tgagtgagct aaaaaataaa 600
caaaatttta aattttcatt agtaaatagc taatcgcga 639
<210> 17
<211> 600
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIC с промотором/lac-оператором и RBS
<400> 17
atcgataaat cataaaaaat ttatttgctt tgtgagcgga taacaattat aatagattca 60
attgtgagcg gataacaatt tcacacagaa ttcattaaag aggagaaatt aaccatgtta 120
aaaaatgatt ttaacgtatt gggacaaatt gcttggttat gggcaaattc tccaatgcac 180
cgaaattggt cagtttcact gttaatgaag aatgttattc ctgcaattga aaatgaccaa 240
tatttgttac tagttgatga tggttttcct attgcatatt gcagttgggc caaattaact 300
ctagagagtg aggctcgcta tgtaaaggac accaattcat taaaaataga tgattggaat 360
gcaggagatc gtatatggat cattgattgg attgccccat tcggggattc atctctattg 420
tataaacata tgagacaacg ttttccatac gatattggaa gggcaattag aatctatcct 480
agcaaaaaag atactggaaa aatcatatat ttaaaaggag gaaaaataac aaaaaaagta 540
gctgaaaaga catttcttca gtatgagcaa gagttaataa cagctctaca ataatcgcga 600
<210> 18
<211> 642
<212> ДНК
<213> Actonbacillus pleuropneumoniae
<220>
<223> ApxIIIC с промотором/lac-оператором и RBS
<400> 18
tctagaaaat cataaaaaat ttatttgctt tgtgagcgga taacaattat aatagattca 60
attgtgagcg gataacaatt tcacacagaa ttcattaaag aggagaaatt aaccatgagt 120
tataaaaatg ttaaaaattt aacagatgat tttacaactt tagggcatat cgcttggttg 180
tgggctaatt ctccgttaca taaggagtgg tctatctctt tgtttactaa gaatattttg 240
ccagccattc aacatgatca atatatttta cttatgcgag atgagttccc tgtagcgttt 300
tgtagttggg caaatttaac gttaactaat gaagtgaagt atgtacgtga tgtgacgtca 360
ttgacttttg aagattggaa ttcaggagaa cgaaaatggt tgatcgattg gattgcgcca 420
tttggggata acaatacgct ttatagatat atgcgtaaaa aatttcctaa tgaagtattc 480
cgggccattc gagtatatcc tggttctaca gaagcgaaaa tcattcatgt tcaaggagga 540
caaattaata aatttacagc taaaaaatta atacaacaat atcaggaaga acttattcaa 600
gttcttaaca atcacaaaaa aattgtaaga ggataatcgc ga 642
<210> 19
<211> 2446
<212> ДНК
<213> Плазмида pEX-A258
<220>
<223> pEX-A258
<400> 19
gtggcagctc tagagctagc gaattctttg gtgaaattgt tatccgctca caattccaca 60
caacatacga gccggaagca taaagtgtaa agcctggggt gcctaatgag tgagctaact 120
cacattaatt gcgttgcgct cactgcccgc tttccagtcg ggaaacctgt cgtgccagct 180
gcattaatga atcggccaac gcgcggggag aggcggtttg cgtattgggc gctcttccgc 240
ttcctcgctc actgactcgc tgcgctcggt cgttcggctg cggcgagcgg tatcagctca 300
ctcaaaggcg gtaatacggt tatccacaga atcaggggat aacgcaggaa agaacatgtg 360
agcaaaaggc cagcaaaagg ccaggaaccg taaaaaggcc gcgttgctgg cgtttttcca 420
taggctccgc ccccctgacg agcatcacaa aaatcgacgc tcaagtcaga ggtggcgaaa 480
cccgacagga ctataaagat accaggcgtt tccccctgga agctccctcg tgcgctctcc 540
tgttccgacc ctgccgctta ccggatacct gtccgccttt ctcccttcgg gaagcgtggc 600
gctttctcat agctcacgct gtaggtatct cagttcggtg taggtcgttc gctccaagct 660
gggctgtgtg cacgaacccc ccgttcagcc cgaccgctgc gccttatccg gtaactatcg 720
tcttgagtcc aacccggtaa gacacgactt atcgccactg gcagcagcca ctggtaacag 780
gattagcaga gcgaggtatg taggcggtgc tacagagttc ttgaagtggt ggcctaacta 840
cggctacact agaagaacag tatttggtat ctgcgctctg ctgaagccag ttaccttcgg 900
aaaaagagtt ggtagctctt gatccggcaa acaaaccacc gctggtagcg gtggtttttt 960
tgtttgcaag cagcagatta cgcgcagaaa aaaaggatct caagaagatc ctttgatctt 1020
ttctacgggg tctgacgctc agtggaacga aaactcacgt taagggattt tggtcatgag 1080
attatcaaaa aggatcttca cctagatcct tttaaattaa aaatgaagtt ttaaatcaat 1140
ctaaagtata tatgagtaaa cttggtctga cagttaccaa tgcttaatca gtgaggcacc 1200
tatctcagcg atctgtctat ttcgttcatc catagttgcc tgactccccg tcgtgtagat 1260
aactacgata cgggagggct taccatctgg ccccagtgct gcaatgatac cgcgcgaacc 1320
acgctcaccg gctccagatt tatcagcaat aaaccagcca gccggaaggg ccgagcgcag 1380
aagtggtcct gcaactttat ccgcctccat ccagtctatt aattgttgcc gggaagctag 1440
agtaagtagt tcgccagtta atagtttgcg caacgttgtt gccattgcta caggcatcgt 1500
ggtgtcacgc tcgtcgtttg gtatggcttc attcagctcc ggttcccaac gatcaaggcg 1560
agttacatga tcccccatgt tgtgcaaaaa agcggttagc tccttcggtc ctccgatcgt 1620
tgtcagaagt aagttggccg cagtgttatc actcatggtt atggcagcac tgcataattc 1680
tcttactgtc atgccatccg taagatgctt ttctgtgact ggtgagtact caaccaagtc 1740
attctgagaa tagtgtatgc ggcgaccgag ttgctcttgc ccggcgtcaa tacgggataa 1800
taccgcgcca catagcagaa ctttaaaagt gctcatcatt ggaaaacgtt cttcggggcg 1860
aaaactctca aggatcttac cgctgttgag atccagttcg atgtaaccca ctcgtgcacc 1920
caactgatct tcagcatctt ttactttcac cagcgtttct gggtgagcaa aaacaggaag 1980
gcaaaatgcc gcaaaaaagg gaataagggc gacacggaaa tgttgaatac tcatactctt 2040
cctttttcaa tattattgaa gcatttatca gggttattgt ctcatgagcg gatacatatt 2100
tgaatgtatt tagaaaaata aacaaatagg ggttccgcgc acatttcccc gaaaagtgcc 2160
acctgacgtc taagaaacca ttattatcat gacattaacc tataaaaata ggcgtatcac 2220
gaggcccttt cgtctcgcgc gtttcggtga tgacggtgaa aacctctgac acatgcagct 2280
cccggagacg gtcacagctt gtctgtaagc ggatgccggg agcagacaag cccgtcaggg 2340
cgcgtcagcg ggtgttggcg ggtgtcgggg ctggcttaac tatgcggcat cagagcagat 2400
tgtactgaga gtttggcaat tggtcgacct cgagggcgcg cccgta 2446
<210> 20
<211> 4751
<212> ДНК
<213> Плазмида pQE-80L
<220>
<223> pQE-80L
<400> 20
ctcgagaaat cataaaaaat ttatttgctt tgtgagcgga taacaattat aatagattca
60
attgtgagcg gataacaatt tcacacagaa ttcattaaag aggagaaatt aactatgaga
120
ggatcgcatc accatcacca tcacggatcc gcatgcgagc tcggtacccc gggtcgacct
180
gcagccaagc ttaattagct gagcttggac tcctgttgat agatccagta atgacctcag
240
aactccatct ggatttgttc agaacgctcg gttgccgccg ggcgtttttt attggtgaga
300
atccaagcta gcttggcgag attttcagga gctaaggaag ctaaaatgga gaaaaaaatc
360
actggatata ccaccgttga tatatcccaa tggcatcgta aagaacattt tgaggcattt
420
cagtcagttg ctcaatgtac ctataaccag accgttcagc tggatattac ggccttttta
480
aagaccgtaa agaaaaataa gcacaagttt tatccggcct ttattcacat tcttgcccgc
540
ctgatgaatg ctcatccgga atttcgtatg gcaatgaaag acggtgagct ggtgatatgg
600
gatagtgttc acccttgtta caccgttttc catgagcaaa ctgaaacgtt ttcatcgctc
660
tggagtgaat accacgacga tttccggcag tttctacaca tatattcgca agatgtggcg
720
tgttacggtg aaaacctggc ctatttccct aaagggttta ttgagaatat gtttttcgtc
780
tcagccaatc cctgggtgag tttcaccagt tttgatttaa acgtggccaa tatggacaac
840
ttcttcgccc ccgttttcac catgggcaaa tattatacgc aaggcgacaa ggtgctgatg
900
ccgctggcga ttcaggttca tcatgccgtt tgtgatggct tccatgtcgg cagaatgctt
960
aatgaattac aacagtactg cgatgagtgg cagggcgggg cgtaattttt ttaaggcagt
1020
tattggtgcc cttaaacgcc tggggtaatg actctctagc ttgaggcatc aaataaaacg
1080
aaaggctcag tcgaaagact gggcctttcg ttttatctgt tgtttgtcgg tgaacgctct
1140
cctgagtagg acaaatccgc cctctagatt acgtgcagtc gatgataagc tgtcaaacat
1200
gagaattgtg cctaatgagt gagctaactt acattaattg cgttgcgctc actgcccgct
1260
ttccagtcgg gaaacctgtc gtgccagctg cattaatgaa tcggccaacg cgcggggaga
1320
ggcggtttgc gtattgggcg ccagggtggt ttttcttttc accagtgaga cgggcaacag
1380
ctgattgccc ttcaccgcct ggccctgaga gagttgcagc aagcggtcca cgctggtttg
1440
ccccagcagg cgaaaatcct gtttgatggt ggttaacggc gggatataac atgagctgtc
1500
ttcggtatcg tcgtatccca ctaccgagat atccgcacca acgcgcagcc cggactcggt
1560
aatggcgcgc attgcgccca gcgccatctg atcgttggca accagcatcg cagtgggaac
1620
gatgccctca ttcagcattt gcatggtttg ttgaaaaccg gacatggcac tccagtcgcc
1680
ttcccgttcc gctatcggct gaatttgatt gcgagtgaga tatttatgcc agccagccag
1740
acgcagacgc gccgagacag aacttaatgg gcccgctaac agcgcgattt gctggtgacc
1800
caatgcgacc agatgctcca cgcccagtcg cgtaccgtct tcatgggaga aaataatact
1860
gttgatgggt gtctggtcag agacatcaag aaataacgcc ggaacattag tgcaggcagc
1920
ttccacagca atggcatcct ggtcatccag cggatagtta atgatcagcc cactgacgcg
1980
ttgcgcgaga agattgtgca ccgccgcttt acaggcttcg acgccgcttc gttctaccat
2040
cgacaccacc acgctggcac ccagttgatc ggcgcgagat ttaatcgccg cgacaatttg
2100
cgacggcgcg tgcagggcca gactggaggt ggcaacgcca atcagcaacg actgtttgcc
2160
cgccagttgt tgtgccacgc ggttgggaat gtaattcagc tccgccatcg ccgcttccac
2220
tttttcccgc gttttcgcag aaacgtggct ggcctggttc accacgcggg aaacggtctg
2280
ataagagaca ccggcatact ctgcgacatc gtataacgtt actggtttca cattcaccac
2340
cctgaattga ctctcttccg ggcgctatca tgccataccg cgaaaggttt tgcaccattc
2400
gatggtgtcg gaatttcggg cagcgttggg tcctggccac gggtgcgcat gatctagagc
2460
tgcctcgcgc gtttcggtga tgacggtgaa aacctctgac acatgcagct cccggagacg
2520
gtcacagctt gtctgtaagc ggatgccggg agcagacaag cccgtcaggg cgcgtcagcg
2580
ggtgttggcg ggtgtcgggg cgcagccatg acccagtcac gtagcgatag cggagtgtat
2640
actggcttaa ctatgcggca tcagagcaga ttgtactgag agtgcaccat atgcggtgtg
2700
aaataccgca cagatgcgta aggagaaaat accgcatcag gcgctcttcc gcttcctcgc
2760
tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct cactcaaagg
2820
cggtaatacg gttatccaca gaatcagggg ataacgcagg aaagaacatg tgagcaaaag
2880
gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc cataggctcc
2940
gcccccctga cgagcatcac aaaaatcgac gctcaagtca gaggtggcga aacccgacag
3000
gactataaag ataccaggcg tttccccctg gaagctccct cgtgcgctct cctgttccga
3060
ccctgccgct taccggatac ctgtccgcct ttctcccttc gggaagcgtg gcgctttctc
3120
atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag ctgggctgtg
3180
tgcacgaacc ccccgttcag cccgaccgct gcgccttatc cggtaactat cgtcttgagt
3240
ccaacccggt aagacacgac ttatcgccac tggcagcagc cactggtaac aggattagca
3300
gagcgaggta tgtaggcggt gctacagagt tcttgaagtg gtggcctaac tacggctaca
3360
ctagaaggac agtatttggt atctgcgctc tgctgaagcc agttaccttc ggaaaaagag
3420
ttggtagctc ttgatccggc aaacaaacca ccgctggtag cggtggtttt tttgtttgca
3480
agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc ttttctacgg
3540
ggtctgacgc tcagtggaac gaaaactcac gttaagggat tttggtcatg agattatcaa
3600
aaaggatctt cacctagatc cttttaaatt aaaaatgaag ttttaaatca atctaaagta
3660
tatatgagta aacttggtct gacagttacc aatgcttaat cagtgaggca cctatctcag
3720
cgatctgtct atttcgttca tccatagttg cctgactccc cgtcgtgtag ataactacga
3780
tacgggaggg cttaccatct ggccccagtg ctgcaatgat accgcgagac ccacgctcac
3840
cggctccaga tttatcagca ataaaccagc cagccggaag ggccgagcgc agaagtggtc
3900
ctgcaacttt atccgcctcc atccagtcta ttaattgttg ccgggaagct agagtaagta
3960
gttcgccagt taatagtttg cgcaacgttg ttgccattgc tacaggcatc gtggtgtcac
4020
gctcgtcgtt tggtatggct tcattcagct ccggttccca acgatcaagg cgagttacat
4080
gatcccccat gttgtgcaaa aaagcggtta gctccttcgg tcctccgatc gttgtcagaa
4140
gtaagttggc cgcagtgtta tcactcatgg ttatggcagc actgcataat tctcttactg
4200
tcatgccatc cgtaagatgc ttttctgtga ctggtgagta ctcaaccaag tcattctgag
4260
aatagtgtat gcggcgaccg agttgctctt gcccggcgtc aatacgggat aataccgcgc
4320
cacatagcag aactttaaaa gtgctcatca ttggaaaacg ttcttcgggg cgaaaactct
4380
caaggatctt accgctgttg agatccagtt cgatgtaacc cactcgtgca cccaactgat
4440
cttcagcatc ttttactttc accagcgttt ctgggtgagc aaaaacagga aggcaaaatg
4500
ccgcaaaaaa gggaataagg gcgacacgga aatgttgaat actcatactc ttcctttttc
4560
aatattattg aagcatttat cagggttatt gtctcatgag cggatacata tttgaatgta
4620
tttagaaaaa taaacaaata ggggttccgc gcacatttcc ccgaaaagtg ccacctgacg
4680
tctaagaaac cattattatc atgacattaa cctataaaaa taggcgtatc acgaggccct
4740
ttcgtcttca c
4751
<210> 21
<211> 3431
<212> ДНК
<213> Плазмида pQE-60
<220>
<223> pQE-60
<400> 21
ctcgagaaat cataaaaaat ttatttgctt tgtgagcgga taacaattat aatagattca
60
attgtgagcg gataacaatt tcacacagaa ttcattaaag aggagaaatt aaccatggga
120
ggatccagat ctcatcacca tcaccatcac taagcttaat tagctgagct tggactcctg
180
ttgatagatc cagtaatgac ctcagaactc catctggatt tgttcagaac gctcggttgc
240
cgccgggcgt tttttattgg tgagaatcca agctagcttg gcgagatttt caggagctaa
300
ggaagctaaa atggagaaaa aaatcactgg atataccacc gttgatatat cccaatggca
360
tcgtaaagaa cattttgagg catttcagtc agttgctcaa tgtacctata accagaccgt
420
tcagctggat attacggcct ttttaaagac cgtaaagaaa aataagcaca agttttatcc
480
ggcctttatt cacattcttg cccgcctgat gaatgctcat ccggaatttc gtatggcaat
540
gaaagacggt gagctggtga tatgggatag tgttcaccct tgttacaccg ttttccatga
600
gcaaactgaa acgttttcat cgctctggag tgaataccac gacgatttcc ggcagtttct
660
acacatatat tcgcaagatg tggcgtgtta cggtgaaaac ctggcctatt tccctaaagg
720
gtttattgag aatatgtttt tcgtctcagc caatccctgg gtgagtttca ccagttttga
780
tttaaacgtg gccaatatgg acaacttctt cgcccccgtt ttcaccatgc atgggcaaat
840
attatacgca aggcgacaag gtgctgatgc cgctggcgat tcaggttcat catgccgtct
900
gtgatggctt ccatgtcggc agaatgctta atgaattaca acagtactgc gatgagtggc
960
agggcggggc gtaatttttt taaggcagtt attggtgccc ttaaacgcct ggggtaatga
1020
ctctctagct tgaggcatca aataaaacga aaggctcagt cgaaagactg ggcctttcgt
1080
tttatctgtt gtttgtcggt gaacgctctc ctgagtagga caaatccgcc gctctagagc
1140
tgcctcgcgc gtttcggtga tgacggtgaa aacctctgac acatgcagct cccggagacg
1200
gtcacagctt gtctgtaagc ggatgccggg agcagacaag cccgtcaggg cgcgtcagcg
1260
ggtgttggcg ggtgtcgggg cgcagccatg acccagtcac gtagcgatag cggagtgtat
1320
actggcttaa ctatgcggca tcagagcaga ttgtactgag agtgcaccat atgcggtgtg
1380
aaataccgca cagatgcgta aggagaaaat accgcatcag gcgctcttcc gcttcctcgc
1440
tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct cactcaaagg
1500
cggtaatacg gttatccaca gaatcagggg ataacgcagg aaagaacatg tgagcaaaag
1560
gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc cataggctcc
1620
gcccccctga cgagcatcac aaaaatcgac gctcaagtca gaggtggcga aacccgacag
1680
gactataaag ataccaggcg tttccccctg gaagctccct cgtgcgctct cctgttccga
1740
ccctgccgct taccggatac ctgtccgcct ttctcccttc gggaagcgtg gcgctttctc
1800
atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag ctgggctgtg
1860
tgcacgaacc ccccgttcag cccgaccgct gcgccttatc cggtaactat cgtcttgagt
1920
ccaacccggt aagacacgac ttatcgccac tggcagcagc cactggtaac aggattagca
1980
gagcgaggta tgtaggcggt gctacagagt tcttgaagtg gtggcctaac tacggctaca
2040
ctagaaggac agtatttggt atctgcgctc tgctgaagcc agttaccttc ggaaaaagag
2100
ttggtagctc ttgatccggc aaacaaacca ccgctggtag cggtggtttt tttgtttgca
2160
agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc ttttctacgg
2220
ggtctgacgc tcagtggaac gaaaactcac gttaagggat tttggtcatg agattatcaa
2280
aaaggatctt cacctagatc cttttaaatt aaaaatgaag ttttaaatca atctaaagta
2340
tatatgagta aacttggtct gacagttacc aatgcttaat cagtgaggca cctatctcag
2400
cgatctgtct atttcgttca tccatagttg cctgactccc cgtcgtgtag ataactacga
2460
tacgggaggg cttaccatct ggccccagtg ctgcaatgat accgcgagac ccacgctcac
2520
cggctccaga tttatcagca ataaaccagc cagccggaag ggccgagcgc agaagtggtc
2580
ctgcaacttt atccgcctcc atccagtcta ttaattgttg ccgggaagct agagtaagta
2640
gttcgccagt taatagtttg cgcaacgttg ttgccattgc tacaggcatc gtggtgtcac
2700
gctcgtcgtt tggtatggct tcattcagct ccggttccca acgatcaagg cgagttacat
2760
gatcccccat gttgtgcaaa aaagcggtta gctccttcgg tcctccgatc gttgtcagaa
2820
gtaagttggc cgcagtgtta tcactcatgg ttatggcagc actgcataat tctcttactg
2880
tcatgccatc cgtaagatgc ttttctgtga ctggtgagta ctcaaccaag tcattctgag
2940
aatagtgtat gcggcgaccg agttgctctt gcccggcgtc aatacgggat aataccgcgc
3000
cacatagcag aactttaaaa gtgctcatca ttggaaaacg ttcttcgggg cgaaaactct
3060
caaggatctt accgctgttg agatccagtt cgatgtaacc cactcgtgca cccaactgat
3120
cttcagcatc ttttactttc accagcgttt ctgggtgagc aaaaacagga aggcaaaatg
3180
ccgcaaaaaa gggaataagg gcgacacgga aatgttgaat actcatactc ttcctttttc
3240
aatattattg aagcatttat cagggttatt gtctcatgag cggatacata tttgaatgta
3300
tttagaaaaa taaacaaata ggggttccgc gcacatttcc ccgaaaagtg ccacctgacg
3360
tctaagaaac cattattatc atgacattaa cctataaaaa taggcgtatc acgaggccct
3420
ttcgtcttca c
3431
<210> 22
<211> 4245
<212> ДНК
<213> Плазмида pACYC184
<220>
<223> pACYC184
<400> 22
gaattccgga tgagcattca tcaggcgggc aagaatgtga ataaaggccg gataaaactt
60
gtgcttattt ttctttacgg tctttaaaaa ggccgtaata tccagctgaa cggtctggtt
120
ataggtacat tgagcaactg actgaaatgc ctcaaaatgt tctttacgat gccattggga
180
tatatcaacg gtggtatatc cagtgatttt tttctccatt ttagcttcct tagctcctga
240
aaatctcgat aactcaaaaa atacgcccgg tagtgatctt atttcattat ggtgaaagtt
300
ggaacctctt acgtgccgat caacgtctca ttttcgccaa aagttggccc agggcttccc
360
ggtatcaaca gggacaccag gatttattta ttctgcgaag tgatcttccg tcacaggtat
420
ttattcggcg caaagtgcgt cgggtgatgc tgccaactta ctgatttagt gtatgatggt
480
gtttttgagg tgctccagtg gcttctgttt ctatcagctg tccctcctgt tcagctactg
540
acggggtggt gcgtaacggc aaaagcaccg ccggacatca gcgctagcgg agtgtatact
600
ggcttactat gttggcactg atgagggtgt cagtgaagtg cttcatgtgg caggagaaaa
660
aaggctgcac cggtgcgtca gcagaatatg tgatacagga tatattccgc ttcctcgctc
720
actgactcgc tacgctcggt cgttcgactg cggcgagcgg aaatggctta cgaacggggc
780
ggagatttcc tggaagatgc caggaagata cttaacaggg aagtgagagg gccgcggcaa
840
agccgttttt ccataggctc cgcccccctg acaagcatca cgaaatctga cgctcaaatc
900
agtggtggcg aaacccgaca ggactataaa gataccaggc gtttccccct ggcggctccc
960
tcgtgcgctc tcctgttcct gcctttcggt ttaccggtgt cattccgctg ttatggccgc
1020
gtttgtctca ttccacgcct gacactcagt tccgggtagg cagttcgctc caagctggac
1080
tgtatgcacg aaccccccgt tcagtccgac cgctgcgcct tatccggtaa ctatcgtctt
1140
gagtccaacc cggaaagaca tgcaaaagca ccactggcag cagccactgg taattgattt
1200
agaggagtta gtcttgaagt catgcgccgg ttaaggctaa actgaaagga caagttttgg
1260
tgactgcgct cctccaagcc agttacctcg gttcaaagag ttggtagctc agagaacctt
1320
cgaaaaaccg ccctgcaagg cggttttttc gttttcagag caagagatta cgcgcagacc
1380
aaaacgatct caagaagatc atcttattaa tcagataaaa tatttctaga tttcagtgca
1440
atttatctct tcaaatgtag cacctgaagt cagccccata cgatataagt tgtaattctc
1500
atgtttgaca gcttatcatc gataagcttt aatgcggtag tttatcacag ttaaattgct
1560
aacgcagtca ggcaccgtgt atgaaatcta acaatgcgct catcgtcatc ctcggcaccg
1620
tcaccctgga tgctgtaggc ataggcttgg ttatgccggt actgccgggc ctcttgcggg
1680
atatcgtcca ttccgacagc atcgccagtc actatggcgt gctgctagcg ctatatgcgt
1740
tgatgcaatt tctatgcgca cccgttctcg gagcactgtc cgaccgcttt ggccgccgcc
1800
cagtcctgct cgcttcgcta cttggagcca ctatcgacta cgcgatcatg gcgaccacac
1860
ccgtcctgtg gatcctctac gccggacgca tcgtggccgg catcaccggc gccacaggtg
1920
cggttgctgg cgcctatatc gccgacatca ccgatgggga agatcgggct cgccacttcg
1980
ggctcatgag cgcttgtttc ggcgtgggta tggtggcagg ccccgtggcc gggggactgt
2040
tgggcgccat ctccttgcat gcaccattcc ttgcggcggc ggtgctcaac ggcctcaacc
2100
tactactggg ctgcttccta atgcaggagt cgcataaggg agagcgtcga ccgatgccct
2160
tgagagcctt caacccagtc agctccttcc ggtgggcgcg gggcatgact atcgtcgccg
2220
cacttatgac tgtcttcttt atcatgcaac tcgtaggaca ggtgccggca gcgctctggg
2280
tcattttcgg cgaggaccgc tttcgctgga gcgcgacgat gatcggcctg tcgcttgcgg
2340
tattcggaat cttgcacgcc ctcgctcaag ccttcgtcac tggtcccgcc accaaacgtt
2400
tcggcgagaa gcaggccatt atcgccggca tggcggccga cgcgctgggc tacgtcttgc
2460
tggcgttcgc gacgcgaggc tggatggcct tccccattat gattcttctc gcttccggcg
2520
gcatcgggat gcccgcgttg caggccatgc tgtccaggca ggtagatgac gaccatcagg
2580
gacagcttca aggatcgctc gcggctctta ccagcctaac ttcgatcact ggaccgctga
2640
tcgtcacggc gatttatgcc gcctcggcga gcacatggaa cgggttggca tggattgtag
2700
gcgccgccct ataccttgtc tgcctccccg cgttgcgtcg cggtgcatgg agccgggcca
2760
cctcgacctg aatggaagcc ggcggcacct cgctaacgga ttcaccactc caagaattgg
2820
agccaatcaa ttcttgcgga gaactgtgaa tgcgcaaacc aacccttggc agaacatatc
2880
catcgcgtcc gccatctcca gcagccgcac gcggcgcatc tcgggcagcg ttgggtcctg
2940
gccacgggtg cgcatgatcg tgctcctgtc gttgaggacc cggctaggct ggcggggttg
3000
ccttactggt tagcagaatg aatcaccgat acgcgagcga acgtgaagcg actgctgctg
3060
caaaacgtct gcgacctgag caacaacatg aatggtcttc ggtttccgtg tttcgtaaag
3120
tctggaaacg cggaagtccc ctacgtgctg ctgaagttgc ccgcaacaga gagtggaacc
3180
aaccggtgat accacgatac tatgactgag agtcaacgcc atgagcggcc tcatttctta
3240
ttctgagtta caacagtccg caccgctgtc cggtagctcc ttccggtggg cgcggggcat
3300
gactatcgtc gccgcactta tgactgtctt ctttatcatg caactcgtag gacaggtgcc
3360
ggcagcgccc aacagtcccc cggccacggg gcctgccacc atacccacgc cgaaacaagc
3420
gccctgcacc attatgttcc ggatctgcat cgcaggatgc tgctggctac cctgtggaac
3480
acctacatct gtattaacga agcgctaacc gtttttatca ggctctggga ggcagaataa
3540
atgatcatat cgtcaattat tacctccacg gggagagcct gagcaaactg gcctcaggca
3600
tttgagaagc acacggtcac actgcttccg gtagtcaata aaccggtaaa ccagcaatag
3660
acataagcgg ctatttaacg accctgccct gaaccgacga ccgggtcgaa tttgctttcg
3720
aatttctgcc attcatccgc ttattatcac ttattcaggc gtagcaccag gcgtttaagg
3780
gcaccaataa ctgccttaaa aaaattacgc cccgccctgc cactcatcgc agtactgttg
3840
taattcatta agcattctgc cgacatggaa gccatcacag acggcatgat gaacctgaat
3900
cgccagcggc atcagcacct tgtcgccttg cgtataatat ttgcccatgg tgaaaacggg
3960
ggcgaagaag ttgtccatat tggccacgtt taaatcaaaa ctggtgaaac tcacccaggg
4020
attggctgag acgaaaaaca tattctcaat aaacccttta gggaaatagg ccaggttttc
4080
accgtaacac gccacatctt gcgaatatat gtgtagaaac tgccggaaat cgtcgtggta
4140
ttcactccag agcgatgaaa acgtttcagt ttgctcatgg aaaacggtgt aacaagggtg
4200
aacactatcc catatcacca gctcaccgtc tttcattgcc atacg
4245
<210> 23
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 23
gcatcatgag taaaaaaatt aatggatttg agg 33
<210> 24
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 24
ggtagatctt tagctattta ctaatgaaaa tttaa 35
<210> 25
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 25
gcaacatgtt aaaaaatgat tttaacgtat tgg 33
<210> 26
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 26
ggtagatctt tattgtagag ctgttattaa ct 32
<210> 27
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 27
gcatcatgag ttataaaaat gttaaaaatt taaca 35
<210> 28
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 28
ggtagatctt tatcctctta caattttttt gtg 33
<210> 29
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 29
gcatctagaa aatcataaaa aatttatttg cttt 34
<210> 30
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 30
agctcgcgat tagctattta ctaatgaaaa ttta 34
<210> 31
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 31
gcaatcgata aatcataaaa aatttatttg cttt 34
<210> 32
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 32
agctcgcgat tattgtagag ctgttattaa ct 32
<210> 33
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 33
gcatctagaa aatcataaaa aatttatttg cttt 34
<210> 34
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 34
agctcgcgat tatcctctta caattttttt gt 32
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ БЕЛОК, СВЯЗЫВАЮЩИЙ ФАКТОР Н | 2017 |
|
RU2773464C1 |
| КОМПОЗИЦИИ NEISSERIA MENINGITIDIS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2739504C2 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2017 |
|
RU2773880C1 |
| СТАБИЛИЗИРОВАННЫЕ БЕЛКИ F RSV И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2795459C2 |
| РЕКОМБИНАНТНЫЕ ВАКЦИНЫ ОТ FMDV И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2745373C2 |
| АНТИГЕНЫ И АНТИГЕННЫЕ КОМПОЗИЦИИ | 2013 |
|
RU2727476C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ИНФЕКЦИИ, ВЫЗВАННОЙ STREPTOCOCCUS SUIS | 2015 |
|
RU2735101C2 |
| ВАКЦИНА ПРОТИВ БЕШЕНСТВА | 2014 |
|
RU2712743C2 |
| Слитые молекулы, происходящие от Cholix-токсина, для пероральной доставки биологически активных нагрузок | 2015 |
|
RU2723178C2 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2012 |
|
RU2754446C2 |
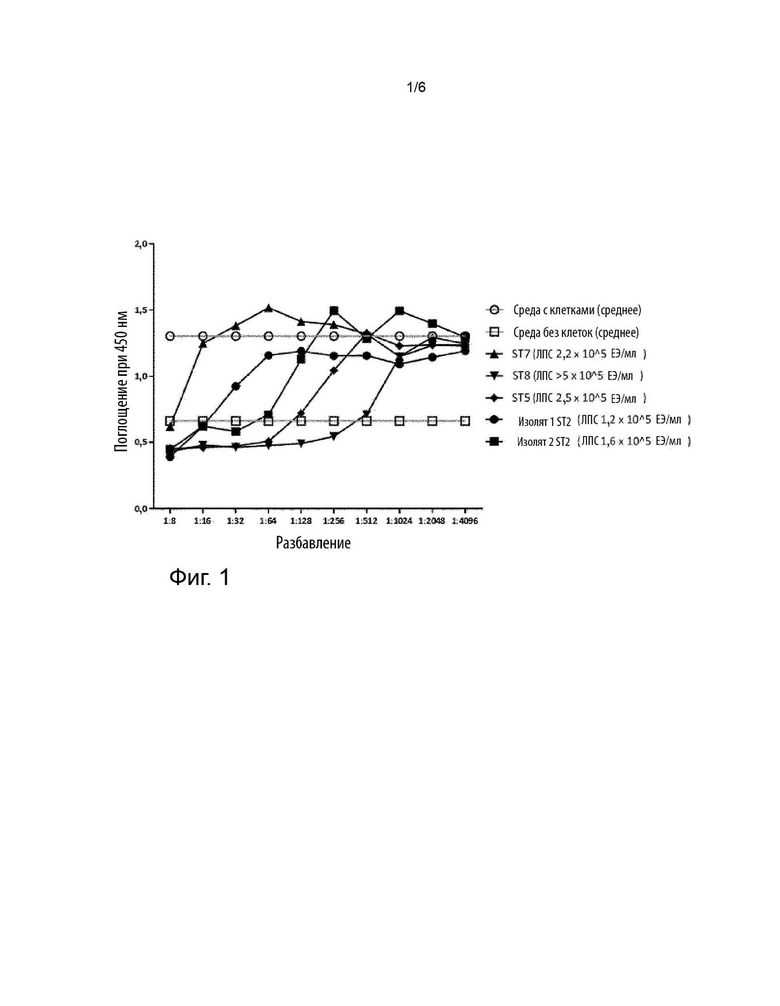
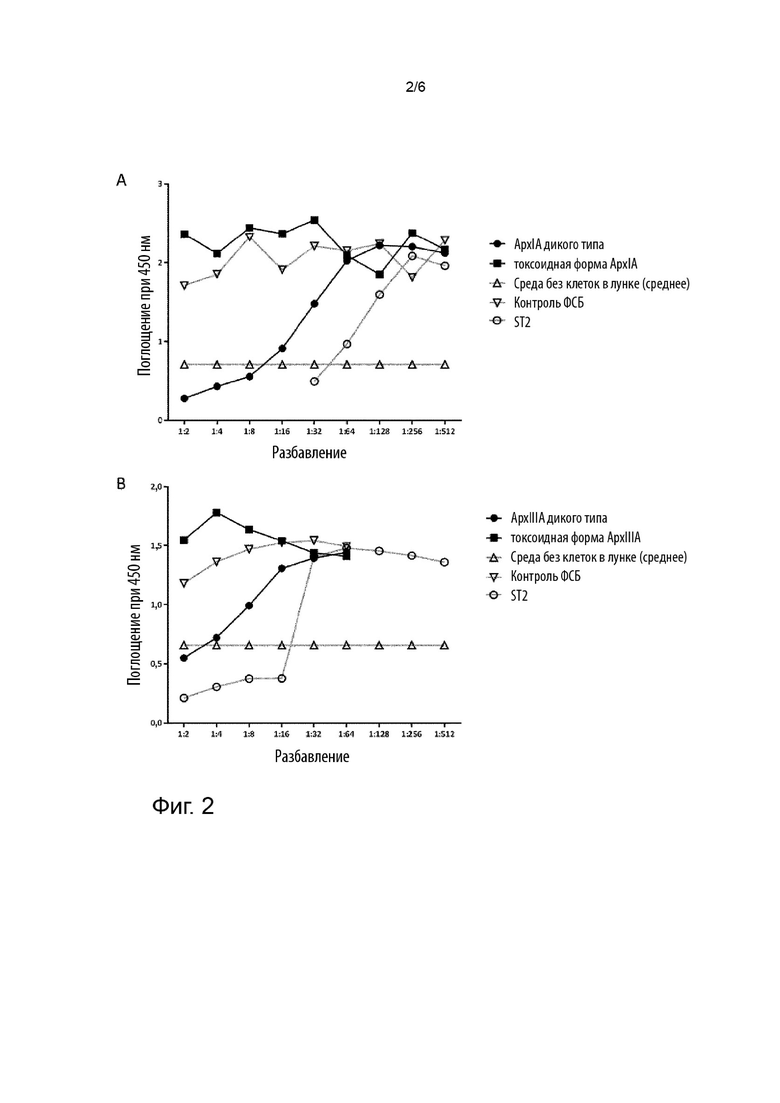
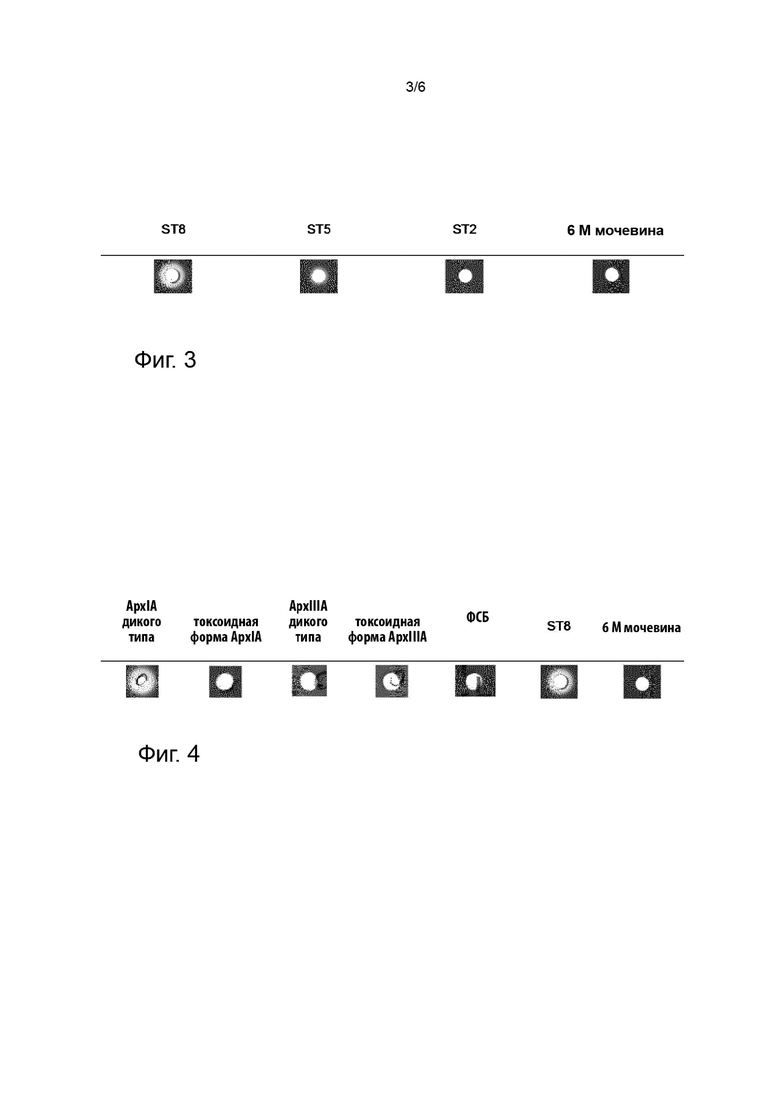
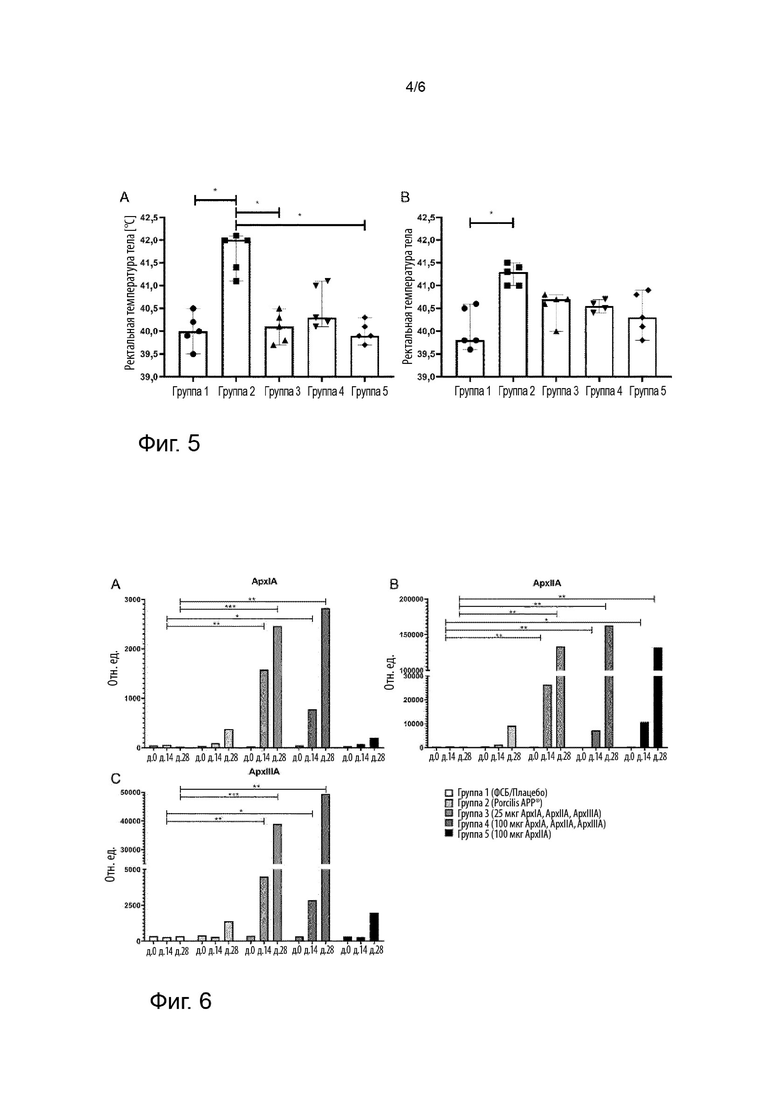
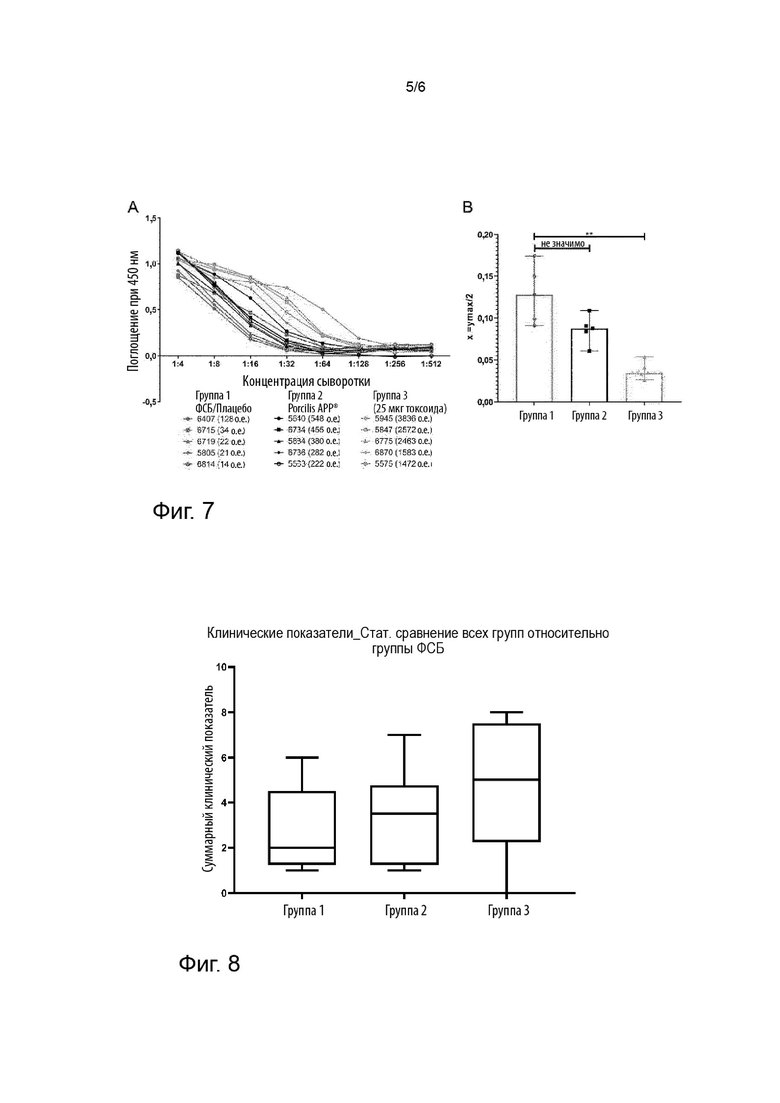
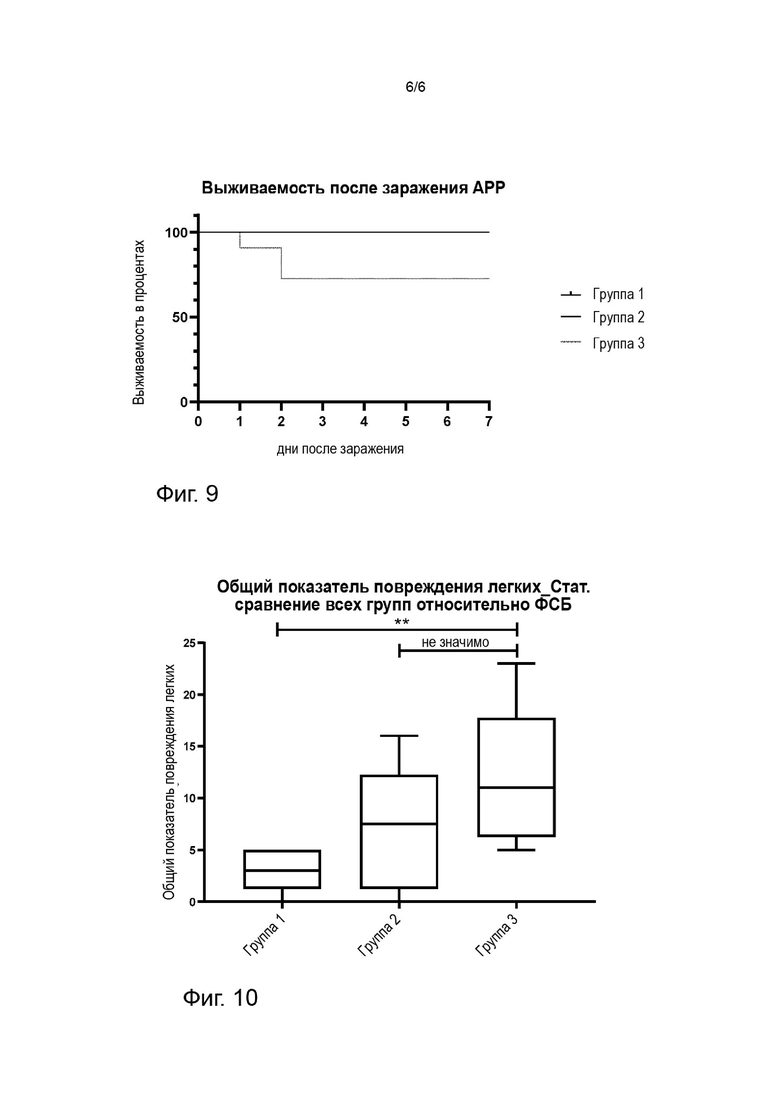
Изобретение относится к композиции вакцины для иммунизации и защиты путем профилактики или метафилактики свиней от плевропневмонии, вызванной Аctinobacillus pleuropneumoniаe, содержащей модифицированные полипептиды ApxIA, ApxIIA и ApxIIIA от Actinobacillus pleuropneumoniae. Также предложено применение указанной композиции вакцины для получения лекарственного средства для иммунизации и защиты путем профилактики или метафилактики свиней от плевропневмонии, вызванной Actinobacillus pleuropneumoniae, для получения гуморального и/или клеточного иммунологического ответа, причем композиция вакцины выполнена с возможностью введения внутримышечно, внутрикожно, внутривенно, подкожно или путем нанесения на слизистую. Изобретение обеспечивает эффективную иммунизацию и защиту свиней от плевропневмонии, вызванной Аctinobacillus pleuropneumoniаe. 2 н. и 12 з.п. ф-лы, 10 ил., 12 табл., 16 пр.
1. Композиция вакцины для иммунизации и защиты путем профилактики или метафилактики свиней от плевропневмонии, вызванной Аctinobacillus pleuropneumoniae, содержащая полипептид ApxIA, ApxIIA и ApxIIIA от Actinobacillus pleuropneumoniae, для получения гуморального и/или клеточного иммунологического ответа, в которой
a) аминокислотная последовательность дикого типа
i) ApxIA, как представлено в SEQ ID NO: 1, имеет замены K560 и K686,
ii) ApxIIA, как представлено в SEQ ID NO: 2, имеет замены K557 и N687, и
iii) ApxIIIA, как представлено в SEQ ID NO: 3, имеет замены K571 и K702,
и, по меньшей мере, аминокислота заменена аминокислотой, не подверженной ацилированию; или
b) аналог или фрагмент модифицированной аминокислотной последовательности, определенной в a), вызывающий гуморальный и/или клеточный иммунологический ответ, который по меньшей мере на 90% гомологичен модифицированной аминокислотной последовательности, определенной в a), и указанный аналог или фрагмент содержит, по меньшей мере, аминокислоту, замененную аминокислотой, не подверженной ацилированию, как определено в a);
и, по меньшей мере, фармацевтический носитель, разбавитель и/или адъювант.
2. Композиция вакцины по п. 1, отличающаяся тем, что аминокислота, не подверженная ацилированию, независимо выбрана из группы, состоящей из аланина, глицина, изолейцина, лейцина, метионина, валина, серина, треонина, глутамина, гистидина, цистеина, пролина, фенилаланина, тирозина, триптофана и глутаминовой кислоты; предпочтительно выбрана из группы, состоящей из аланина, глицина, серина, треонина, валина, изолейцина и лейцина; наиболее предпочтительно выбрана из группы, состоящей из аланина, глицина, серина.
3. Композиция вакцины по любому предшествующему пункту, отличающаяся тем, что аминокислотная последовательность дикого типа
a) ApxIA, как представлено в SEQ ID NO: 1, имеет замены K560 и K686,
b) ApxIIA, как представлено в SEQ ID NO: 2, имеет замены K557 и N687, и
c) ApxIIIA, как представлено в SEQ ID NO: 3, имеет замены K571 и K702,
и, по меньшей мере, аминокислота заменена аминокислотой, не подверженной ацилированию.
4. Композиция вакцины по любому из пп. 1-3, отличающаяся тем, что
a) модифицированная аминокислотная последовательность дикого типа ApxIA представляет собой SEQ ID NO: 4,
b) модифицированная аминокислотная последовательность дикого типа ApxIIA представляет собой SEQ ID NO: 5, и
c) модифицированная аминокислотная последовательность дикого типа ApxIIIA представляет собой SEQ ID NO: 6.
5. Композиция вакцины по любому из пп. 1-4, отличающаяся тем, что полипептид ApxIA, ApxIIA или ApxIIIA кодируется молекулой нуклеиновой кислоты, входящей в состав конструкта нуклеиновой кислоты, содержащего последовательность нуклеотидов, которая кодирует полипептид ApxIA, ApxIIA или ApxIIIA.
6. Композиция вакцины по п. 5, отличающаяся тем, что молекула нуклеиновой кислоты входит в состав вектора, пригодного для экспрессирования нуклеиновой кислоты по изобретению.
7. Композиция вакцины по любому одному из пп. 5 или 6, отличающаяся тем, что молекула нуклеиновой кислоты входит в состав микроорганизма по изобретению.
8. Композиция вакцины по п. 7, отличающаяся тем, что микроорганизм, содержащий нуклеиновую кислоту, представляет собой штамм Escherichia coli или штамм Actinobacillus, предпочтительно штамм Actinobacillus pleuropneumoniae.
9. Композиция вакцины по любому из пп. 1-8, отличающаяся тем, что представляет собой субъединичную вакцину.
10. Композиции вакцины по п. 9, отличающаяся тем, что субъединичная вакцина содержит по меньшей мере один дополнительный полипептид Actinobacillus pleuropneumoniae.
11. Композиция вакцины по любому из пп. 1-8, отличающаяся тем, что представляет собой живую вакцину, предпочтительно содержащую по меньшей мере один штамм Actinobacillus pleuropneumoniae.
12. Композиция вакцины по п. 11, отличающаяся тем, что содержит по меньшей мере два различных штамма Actinobacillus pleuropneumoniae.
13. Композиция вакцины по любому одному из пп. 11 или 12, отличающаяся тем, что штамм Actinobacillus pleuropneumoniae содержит делецию гена apxIV или/и гена sxy.
14. Применение композиции вакцины по любому одному из пп. 1-13 для получения лекарственного средства для иммунизации и защиты путем профилактики или метафилактики свиней от плевропневмонии, вызванной Actinobacillus pleuropneumoniae, для получения гуморального и/или клеточного иммунологического ответа, причем композиция вакцины выполнена с возможностью введения внутримышечно, внутрикожно, внутривенно, подкожно или путем нанесения на слизистую.
| US 6500435 B1, 31.12.2002 | |||
| US 6770275 B1, 03.08.2004 | |||
| KR 20150086672 A, 29.07.2015 | |||
| ВАКЦИНА, НАПРАВЛЕННАЯ ПРОТИВ АКТИНОБАЦИЛЛЕЗНОЙ ПЛЕВРОПНЕВМОНИИ, И СПОСОБ ПОЛУЧЕНИЯ ТАКОЙ ВАКЦИНЫ | 2010 |
|
RU2554812C2 |

