00
о со
00
Изобретение относится к сельскому хозяйству, в частности к растениеводству, и мбжет быть использовано для диагностики завершения стадии яровизации как в практике фитотронии, так и при производственном выращивании озимой пшеницы в случае необходимости установления факта прохождения стадии яровизации после неблагоприятных осенне-зимних условий для роста и развития озимых растений при решении вопроса о целесообразности пересева после перезимовки ослабленных вследствие осенней засухи посевов. Ранняя диагностика окончания либо незавершения стадии яровизации при производственных посевах позволяет ускорить пересев и испольт зовать при этом лучше весеннюю влагу.
В фитотронии тестирование завершения срока яровизации позволяет значительно (на 40%) уменьшить время эксплуатации климатических камер в научно-исследовательских учреждениях, что обеспечивает существенную экономию электроэнергии и расходы по техническому обслуживанию фитотрона.
Целью изобретения является повышение эффективности способа за счет возможности ранней и быстрой диагностики завершения срока яровизации и сокращение сроков ее определения.
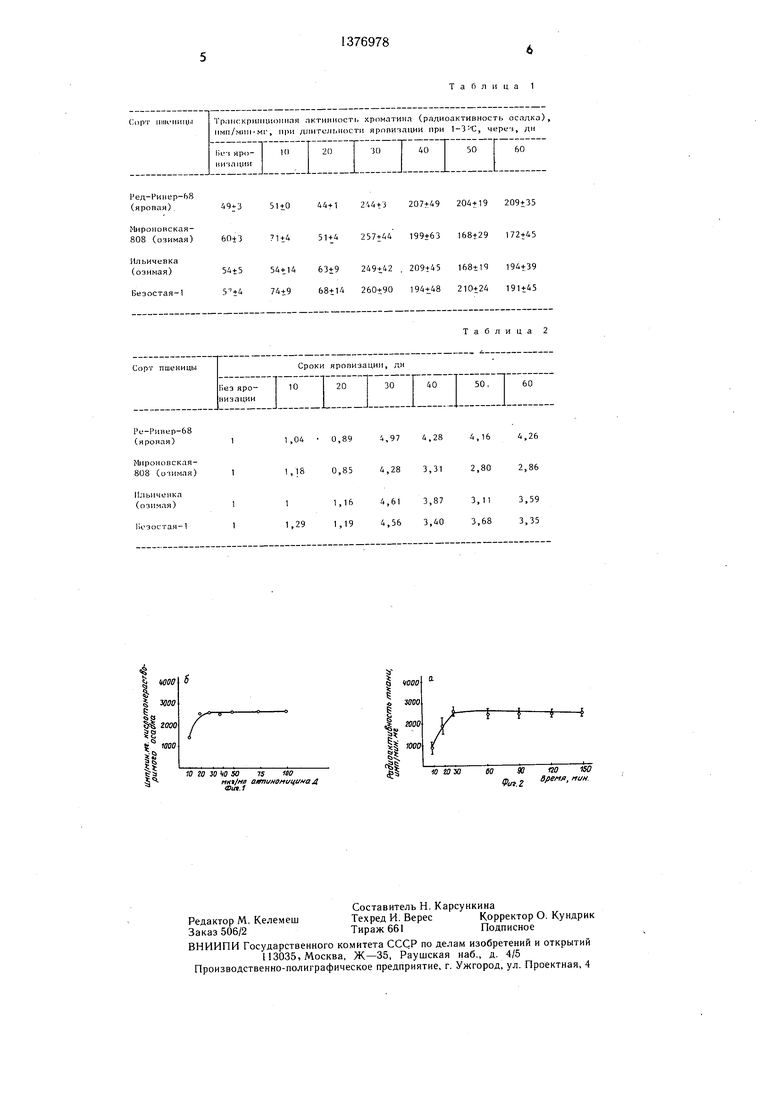
На фиг. 1 изображена кинетика поглощения НАМД тканями меристем главного стебля озимой пшеницы Мироновская 808; на фиг. 2 - зависимость поглощения НАМД тканями меристем главного стебля от его концентрации.
В основу изобретения взято свойство хроматина изменить свою конформацию и характер белково-нуклеинового взаимодействия, связанного с изменением активности определенных генов при переключении программы онтогенеза растений. Транскрипционную активность хроматина оценивали по способности ДНК образовывать комплексы с антибиотиком актиномицином Д. Актиноми- цин Д обладает высокой избирательностью действия, образуя комплексы с гуаминовы- ми основаниями ДНК, более доступными благодаря разной степени конденсации надмолекулярной структуры ДНК, в которой отражена транскрипционная активность генетического аппарата клеток. Поэтому регистрация уровня связывания актиномицина Д с ДНК хроматина позволяет количественно тестировать транскрипционную активность выделяемого из тканей растения дезокси- рибонуклеопротеида.
Способ осуществляется следующим образом.
Из 7-10 опытных растений на разных этапах яровизации изолируют конус нарастания в основном побеге вместе с отрезком прилегающего листа длиной около 1 мм (изоляция конуса нарастания занимает больше времени, чем отрезание ко
0
5
нуса вместе с кусочком листа, а на конечный результат это не влияет). Изолированные части растений помещают в водный раствор с меченым по тритию Н-ак- тиномицином Д (НАМД) при концентрации раствора 30 мкг/мл и удельной радиоактивности 1 мкКи в миллилитре на 0,5-1 ч при комнатной температуре. После экспозиции образцы промывают проточной холодной водопроводной водой, гомогенизируют, осаждают двойным объемом 20%-ной три- хлоруксусной кислоты (ТХУ) и определяют радиоактивность нерастворимого осадка в кислоте осадка на сцинтилляционном счетчике. Поскольку НАМД специфически взаимодействует в основном с гуаниновыми основаниями ДНК, а при осаждении ТХУ
в осадке содержится ДНК, РНК и белки, то основное количество включенной радиоактивности приходится на ДНК. Измеренная радиоактивность отражает транскрипционную (относительную) хроматина яровизированных растений и сравниваем ее с активностью неяровизированных растений для установления факта завершения процесса яровизации. По соотношению транскрипционных активностей яровизированных и неяровизированных растений судят о моменте завершения яровизации, считая завершившими яровизацию те растения, у которых наблюдается резкое увеличние этого соотношения.
Установлено, что после завершения процесса яровизации активность хроматина, тестируемая по уровню комплексооб- разования с актиномицином Д, резко повышается по сравнению с транскрипционной активностью хроматина неяровизированных растений.
Пример. Из сортов озимых пшениц Мироновская 808, Ильичевка, Безостая - 1 на разных этапах яровизации (через каждые 10 сут), и контрольных растений (неяро- 0 визированных), а также у ярового сорта Ред-Ривер-68 произвольно отбирали 7-10 растений (метод позволяет использовать и меньшее количество растений, что. важно при селекционной работе), изолировали конус нарастания с отрезком листа около 1 мм. Образцы помещали на 0,5-1 ч в водный раствор с меченым по тритию актиномицином Д. Концентрация антибиотика - 30 мкг/мл; удельная радиоактивность - 1 мкКи/мл. Оптимум концентрации и длительность экспозиции установлены экспериментально (см. фиг. 1 и 2).
После экспозиции образцы промывали водопроводной водой, гомогенизировали с водой (1:1 - навеска:вода), добавляли раствор 20%-ной ТХУ до конечной концентрации 10% ТХУ. Полученные осадки переносили на нитроцеллюлозные фильтры, промывали сначала раствором 5%, затем водой, высушивали на воздухе и определяли
0
5
0
5
радиоактивность на сцинтилляционном счетчике. Вся процедура заняла 2-3 ч.
Яровизацию растений указанных сортов озимых пшениц проводили путем выдерживания растений при 1-3°С в течение определенного времени, а именно 60 дн, и через каждые 10 сут отбирали пробы для определения транскрипционной активности указанным способом.
В табл. 1 приведены сравнительные данные определения момента завершения стадии яровизации согласно предлагаемому способу у растений, яровизированных с помощью низких температур, и неяровизированных растений.
Из приведенных в табл. 1 данных видно, что . максимальная транскрипционная активность наблюдалась, начиная с 30-го дня яровизации растений. Этот период оказался достаточным для того, чтобы растения выколосились.
Если сравнить транскрипционную активность хроматина у яровизированных и неяровизированных растений, то оказывается, что момент завершения стадии яровизации наступает, когда резко увеличивается соотношение транскрипционных активностей у яровизированных к неяровизированным растениям.
В табл. 2 представлено соотношение транскрипционных активностей яровизированных к неяровизированным растениям (отношение радиоактивности кислотонерастворимого осадка у яровизированных растений к неяровизированным).
Так, у неяровизированных растений сорта Мироновская 808 транскрипционная активность, тестируемая по интенсивности связывания актиномицина Д, составляет бОмп/мин мг . При яровизации растений до 20 дн включительно резкого повышения транскрипционной активности не обнаружено (соотношение транскрипционной активности яровизированных растений к неяровизированным сортам Мироновская 808, прошедших яровизацию в течение 10 дн, составляет 71:60 1,18; 20 дн - 51:60 0,85). При более длительном выдерживании растений при низких температурах происходит скачкообразное повышение этого показателя (30 дн - 257:60 4,25; 40 дн - 168:60 2, 80; 60 дн - 171:60 2,86).
Для сорта Безостая-1 соотношение транскрипционных активностей яровизированных растений при низких температурах к неяровизированным составляло: после 10 дн.- 1,28; 20 дн - 1,19; 30 дн - 4,5; 40 дн - 3,40; 50 дн - 3,68; 60 дн - 3,35;. Аналогичные данные получены и для других
сортов.
Таким образом, резкое повышение транскрипционной активности хроматина у завершивших яровизацию растений можно.расценивать, как переключение программы онтогенеза.
Преимущества предлагаемого способа заключается в следующем.
Значительное сокращение времени тестирования - 2-3 ч (быстрая оценка селекционного материала на продолжитель5 ность яровизации дает возможность на ранних этапах выявлять высокозимостойкие, длинностадийные формы).
Раннее определение срока завершения процесса яровизации в климатических камерах приводит к освобождению площади,
0 эксплуатация которой составляет 300 руб на 1 м за 30 дн (возможность ранней оценки производственных посевов позволяет своевременно решить вопрос о необходимости пересева при неблагоприятных условиях осен5 ней вегетации после перезимовки и т.е. поднять урожайность зерновых культур на 4-5 ц/га).
Минимальное количество аналитического материала для определения окончания момента завершения стадии яровизации (точки
0 роста из 7-10 растений), а при необходимости анализа уникальных селекционных форм можно ограничиться одним растением или одной точкой роста.
Формула изобретения
Способ диагностики завершения стадии яровизации озимой пшеницы, включающий отбор и анализ растений на биохимический показатель, значение которого связано с завершением стадии яровизации, отличающийся тем, что, с целью повышения эффективности способа и сокращения сроков определения, в качестве биохимического показателя используют транскрипционные активности хроматина, которые определяют в айикальных отрезках основных побегов растений, вычисляют соотнощение данного показателя у яровизированных и неяровизированных растений, считая завершившими стадию яровизации те растения, у которых это соотношение составляет 2,8-4,9.
Таблица 1

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ ОКОНЧАНИЯ ЯРОВИЗАЦИИ МНОГОЛЕТНИХ КОРМОВЫХ ТРАВ | 1999 |
|
RU2176444C2 |
| СПОСОБ В.И.БАБЕНКО ОПРЕДЕЛЕНИЯ ФОТОПЕРИОДИЧЕСКОЙ ЧУВСТВИТЕЛЬНОСТИ ФОРМ ОЗИМОЙ ПШЕНИЦЫ | 1987 |
|
SU1559455A1 |
| Способ выращивания озимой пшеницы | 1989 |
|
SU1727628A1 |
| Способ изменения типа развития у злаковых культур | 1990 |
|
SU1752283A1 |
| СПОСОБ СЕЛЕКЦИИ РАСТЕНИЙ ЗЛАКОВЫХ КУЛЬТУР | 1997 |
|
RU2125365C1 |
| Способ подготовки семян озимых культур к посеву | 1976 |
|
SU588940A2 |
| Способ экологической защиты озимых культур от неблагоприятных условий перезимовки | 1987 |
|
SU1435195A1 |
| Способ оценки засухоустойчивости растений пшеницы | 1990 |
|
SU1687109A1 |
| СПОСОБ ОЦЕНКИ ВЛИЯНИЯ ИНГИБИТОРА ДЕАЦЕТИЛИРОВАНИЯ БЕЛКОВ НА ИНДУКЦИЮ РОСТОВОГО МОРФОГЕНЕЗА РАСТЕНИЙ | 2009 |
|
RU2404585C1 |
| Способ определения морозоустойчивости пшеницы | 1987 |
|
SU1528395A1 |
Изобретение относится к сельскому хозяйству. Цель изобретения - повышение эффективности способа и сокращение сроков определения. Из анализируемых растений вычленяют отрезки с конусом нарастания в основном побеге, определяют в них транскрипционную активность хроматина. По соотношению транскрипционных активностей яровизированных и неяровизированных растений судят о моменте завершения яровизации, считая заверц1ившими яровизацию те растения, у которых это соотношение составляет 2,8-4,9. Резкое повышение транскрипционной активности хроматина у завершивших яровизацию растений можно расценивать как переключение .программы онтогенеза. 2 ил., 2 табл. (в
49 -351+044+1244 + 3207+49204+19209 + 35
60+371+451+4257+44199+63168+29172+45
54±554t1463+9249+42. 209+45168+19194+39
5- +474+968+14260+90194+48210+24191 + 45
1,04 0,894,974,284,164,26
1,18
0,854,28 3,312,802,86
11,164,613,873,113,59
1,291,194,56 3,403,683,35
Таблица 2
| Бассарская М | |||
| А | |||
| О биологической диагностике стадий развития растений.- Яровизация, 1936, 6, с | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
