Изобретение относится к физиологии и биохимии растений и может быть применено к анализу механизмов ростового морфогенеза растений, а именно при исследовании процессов, происходящих на уровне супраструктур интерфазного клеточного ядра, которые иногда называют «молекулярными морфогенезами», физико-химическая природа которых мало исследована, что необходимо для получения дополнительной информации в разработках и построении компьютерных моделей организации генных и эпигенных сетей управления.
Известен способ оценки влияния гиббереловой кислоты и кинетина на белки клеточных ядер [1], в котором был описан способ оценки влияния физиологически активных веществ на белки клеточных ядер проростков растений. Недостаток этого метода заключается в том, что с его помощью невозможно оценить влияние физиологически активных веществ на индукцию ростового морфогенеза растений.
Вышеуказанный способ оценки влияния гиббереловой кислоты и кинетина на белки клеточных ядер [1] был принят за основу, в котором первоначально осуществляют обработку семян в процессе набухания физиологически активными веществами, инкубацией в среде с 2-14С-ацетатом и Na2H32PO4, с последующей изоляцией клеточных ядер по методам Ро, Чипчейза и Кюля [см. 1], получением ядерных экстрактов растворами солей: 0,14 М, 2 М хлористого натрия, определением турбидиметрически содержание белка в пробах, фракционный состав белков определяли с помощью электрофореза. Недостаток этого метода заключается в том, что в анализ берется только один временной период роста растения, а именно 2 суток, что не позволяет оценить морфогенетические изменения в растении в процессе индукции ростовых процессов, происходящие под влиянием физиологически активных веществ, клеточные ядра выделяются методами Ро, Чипчейза и Кюля [см. 1], что ведет к низкому выходу нативных ядер, фракционирование клеточных ядер ограничивается получением только 2-х фракций, содержание белка в которых определяется турбидиметрически, т.е. методом с низким порогом чувствительности (до 3-5 мкг белка в 0,1 мл раствора).
Цель изобретения - предлагается использование Арг-Х протеолиза, как одного из молекулярно-генетических механизмов крупномасштабной пространственной организации хроматина в условиях гиперацетилированного состояния протеома клеточных ядер при индукции ростового морфогенеза растений.
Указанная цель достигается тем, что в способе оценки влияния ингибитора деацетилирования белков на индукцию ростового морфогенеза растений первоначально проращивают семена пшеницы в 0,004 мМ растворе бутирата натрия, в определенные интервалы времени от начала замачивания 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводят отделение зародышей от эндосперма с последующей консервацией зародышей в забуференном 80-90% глицерине при минус 25°С, выделяют клеточные ядра, проводят экстракцию ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом и определяют в вышеперечисленных ядерных фракциях протеолитическую и ингибиторную активности.
Изобретение иллюстрируется следующим примером.
Пример. Опыты проводили на элитных семенах пшеницы (Thticum aestivum L.) сортов Мироновская 808 (озимая) и Мироновская яровая (любезно присланные нам из коллекции Мироновского научно-исследовательского института селекции и семеноводства пшеницы им. В.Н. Ремесло). Проращивание зародышей осуществляли в темноте при 22±1°С. Соответственно, в одном варианте опыта (контроль) использовали дистиллированную воду, в другом варианте - 0,004 мМ NaВ(бутират натрия), растворенный в дистиллированной воде. Бутират натрия использовали для ингибирования in vivo процессов деацетилирования гистонов (возможно и других негистоновых белков). То есть для поддержания удлиненной (пролонгированной) стадии повышенного уровня ацетилирования гистонов (возможно и других ядерных белков), необходимого для стимуляции транскрипции, NaB был синтезирован на базе Института органической химии УНЦ РАН. Элементный анализ бутирата натрия находился в пределах: С - 43,5; Н - 6,3; Na - 21. Оптимальную концентрацию NaB подбирали по увеличению всхожести семян пшеницы Мироновской яровой через 21 ч после индукции ростовых процессов (таблица). В определённые интервалы времени - 0 ч (воздушно-сухое семя) и от начала замачивания семян: 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводили отделение зародышей от эндосперма. Клеточные ядра выделяли по методу [2]. Надмолекулярные структуры: нуклеоплазму (Нп), хроматин непрочно- (Хр-I) и прочно- (Хр-II) связанный с ядерным матриксом (ЯМ), а также ЯМ выделяли из очищенных клеточных ядер соответственно при повышении ионной силы раствора: 0,14М NaCl, 0,35М NaCl, 2М NaCl в 0,01М трис-HCl буфере, рН 6.8. ЯМ извлекали 6М гуанидингидрохлоридом (Gu·HCI) с 0,004%-ным β-меркаптоэтанолом в том же буфере. Количество белка в ядрах и ядерных фракциях определяли методом Бредфорда в нашей модификации [3]. Арг-Х (триптазную) активность оценивали по расщеплению Арг-Х связей в аргининбогатом белке - протамине- Salmine-A-I («Merk») (молекула которого состоит из 33 аминокислот: 22-х молекул Apг; 4-х молекул Сер; 3-х молекул Про; по 2 молекулы Глу и Вал) во всех вышеперечисленных фракциях ядер [3]. Удельную активность триптаз выражали в нмоль аргинина·с-1·мг·белка.
Анализ таблицы показал, что максимальная всхожесть семян на 21 ч прорастания соответствует концентрации бутирата натрия 0,004 мМ.
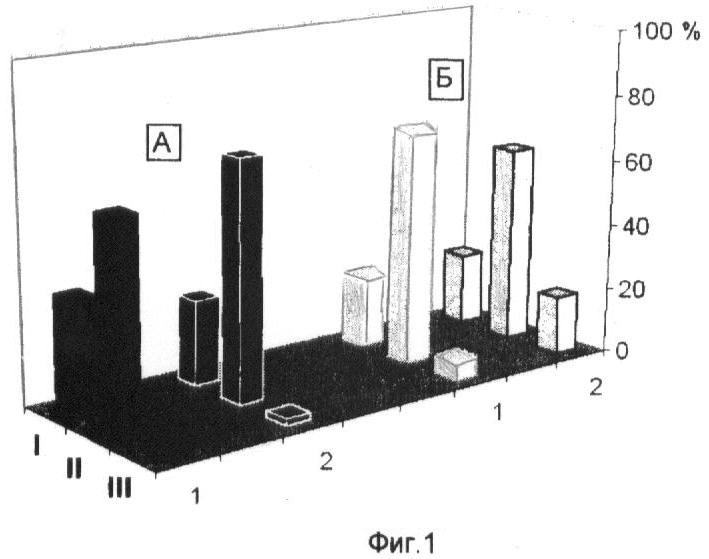
На фиг.1 представлена всхожесть семян пшениц Мироновская яровая (А) и Мироновская озимая (Б), контроли (1), обработанные бутиратом натрия (2). I - не проклюнулись, II - проклюнулись: общий размер проростка 0,3-0,5 см, III - активный рост главного корня (более 0,1 см). На оси ординат показан процент всхожести семян.
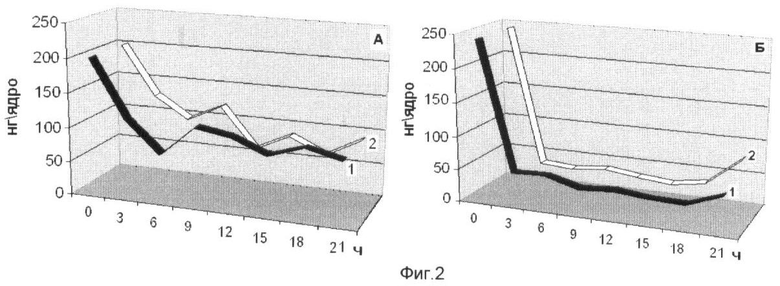
На фиг.2 представлена динамика внутриядерного протеома G1 фазы клеточных ядер зрелых зародышей Мироновской яровой (А) и Мироновской 808 (озимой) (Б) пшениц в нормальных условиях (1) и в присутствии ингибитора деацетилирования белков (2). На оси абсцисс показано время прорастания зародышей пшеницы. На оси ординат - масса 1 зародыша, мг.
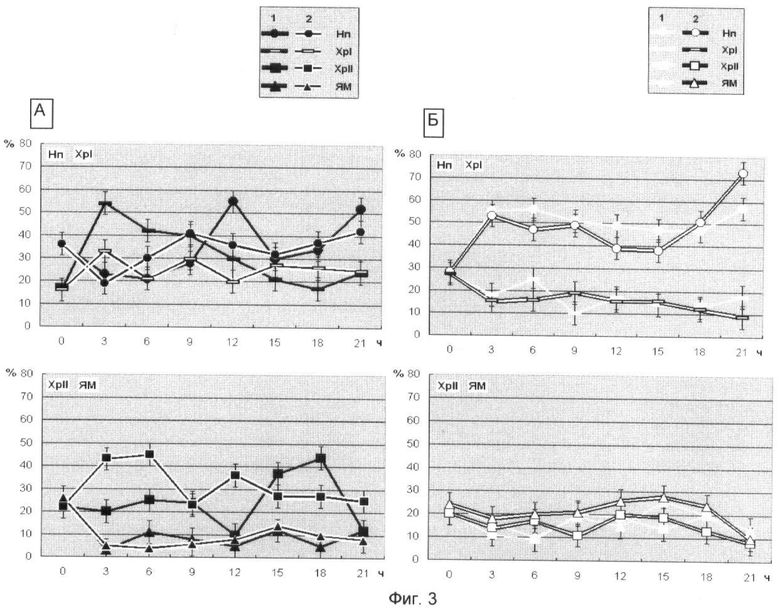
На фиг.3 представлен выход белковых компонентов во фракциях клеточных ядер зрелых зародышей Мироновской яровой (А) и Мироновской 808 (озимой) (Б) пшениц в нормальных условиях (1) и в присутствии ингибитора деацетилирования белков (2). Использованы следующие обозначения: Нп-нуклеоплазма, Хр-I - хроматин непрочносвязанный с ядерным матриксом, Хр-II - прочносвязанный с ядерным матриксом, ЯМ - ядерный матрикс. На оси абсцисс показан возраст зародышей пшеницы, по оси ординат - процент выхода белковых компонентов.
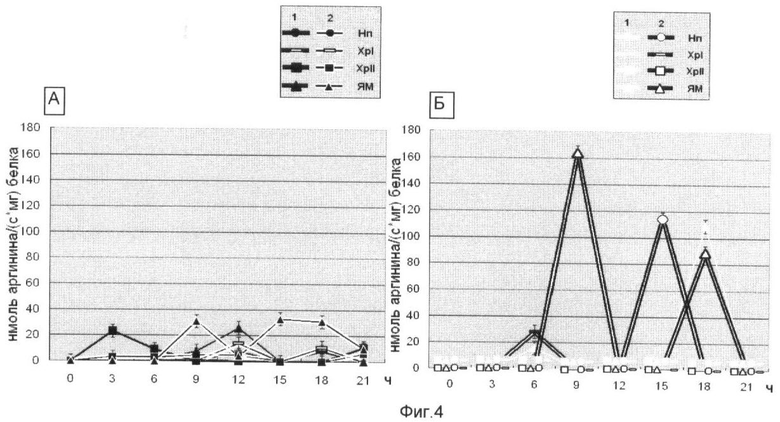
На фиг.4 представлена Арг-Х протеолитическая активность во фракциях клеточных ядер зрелых зародышей Мироновской яровой (А) и Мироновской 808 (озимой) (Б) пшениц в нормальных условиях (1) и в присутствии ингибитора деацетилирования белков (2). Использованы следующие обозначения: Нп - нуклеоплазма, Хр-I - хроматин непрочносвязанный с ядерным матриксом, Хр-II - прочносвязанный с ядерным матриксом, ЯМ - ядерный матрикс. На оси абсцисс указан возраст зародышей пшеницы, по оси ординат - единицы протеолитической активности нмоль аргинина·с-1·мг·белка.
Анализ фиг.4 показал, что протеазочувствительность супраструктур клеточного ядра в присутствии ингибитора деацетилирования ядерных белков резко влияет на экспонированность определенных участков ядерного матрикса (9 ч), нуклеоплазмы (15 ч) при сохраненности Арг-Х протеолиза на уровне ядерного матрикса в контрольном варианте опыта (18 ч). 18-ч фаза клеточного цикла у пшеницы интересна тем, что в этот временной период наблюдается репликация и переход к синтезу ДНК, т.е. клетка осуществляет процессы, необходимые для их сохранения в митозе.
Данное изобретение рекомендуется для молекулярно-генетического анализа механизмов ростового морфогенеза растений, а именно при исследовании процессов, происходящих на уровне супраструктур интерфазного клеточного ядра.
Изобретение относится к физиологии и биохимии растений. Первоначально проращивают семена пшеницы в 0,004 мМ растворе бутирата натрия, в определенные интервалы времени от начала замачивания 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводят отделение зародышей от эндосперма с последующей консервацией зародышей в забуференном 80-90% глицерине при минус 25°С, выделяют клеточные ядра, проводят экстракцию ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом и определяют в вышеперечисленных ядерных фракциях протеолитическую и ингибиторную активности. Изобретение позволяет реализовать указанное назначение. 1 табл., 4 ил.
Способ оценки влияния ингибитора деацетилирования белков на индукцию ростового морфогенеза растений, включающий проращивание семян пшеницы в 0,004 мМ растворе бутирата натрия, отделение зародышей от эндосперма в определенные интервалы времени от начала замачивания: 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч, консервацию зародышей в забуференном 80-90% глицерине при минус 25°С, с последующим выделением клеточных ядер, экстракцией ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом, определением в вышеперечисленных ядерных фракциях протеолитической и ингибиторной активности.
| Иванова Э.А., Ахметов P.P | |||
| Модификация негистоновых белков в проростках растений// Физиология растений, 1987, Т.34, № 3, С.507-512 | |||
| Способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью | 1990 |
|
SU1733471A1 |
| Способ выделения растительных клеточных ядер | 1990 |
|
SU1701747A1 |

