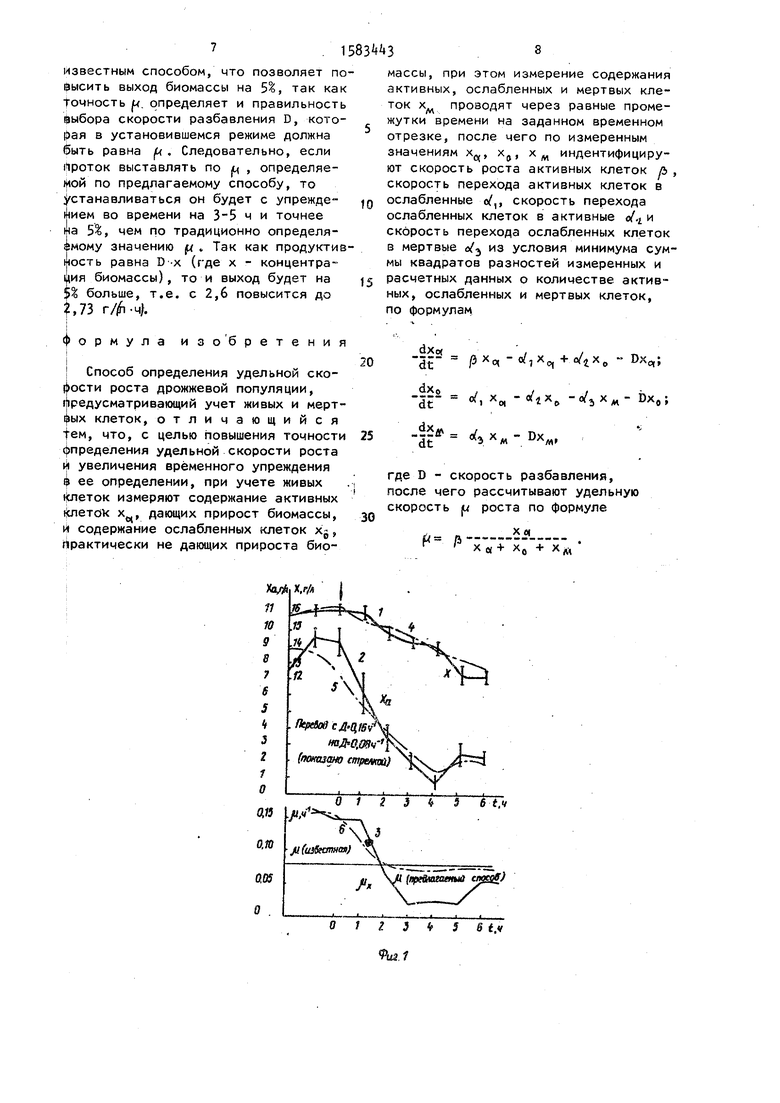
культивирования Candida maltosa ВСБ-5&9 нэ среде с парафином снача- ла при D 0,15 ч( , затем при смене D на 0,25 ч 1, где показано стрелкой 1 «; 2.5 - х; 3,6 - |U (кривые 1-3 - экспериментальные данные; кривые k-6 - расчетные данные).
Способ заключается в том, что группу живых клеток разбивают на активные, дающие прирост биомассы, и ослабленные, практически не дающие прироста биомассы, клетки, определяет относительное содержание активных клеток в популяции, идентифицируют коэффициент пропорциональности из условия минимума суммы квадратов разностей экспериментальных и расчетных цанных о количестве активных, ослаб- пенных и мертвых клеток на заданном этрезке времени, умножают коэффици- энт пропорциональности на относительное содержание активных клеток, полу™ чая тем самым искомый параметр. | На сканирующем микроскопе, соединенном с ЭВМ, анализируют через равные промежутки времени многослойные препараты пробы, взятой из ферментера, и выдают следующие морфологофизиологи- ческие параметры популяции: распределе ие клеток по размерам, форме, содер- 1кание липидных включений и вакуолей9 тепени гетерогенности протоплазмы (моменты распределения ее оптической Плотности), а также процентное содер- Кание мертвых и почкующихся клеток. Всю биомассу представляют в виде суммы клеток (х) активных (х), ос- Ьабленных (хл) и мертвых (х,,):
vгЛ
V
х « х D +
г
(1)
Активные, ослабленные и мертвые клетки идентифицируют по значениям оптической плотности цитоплазмы, которые для активных клеток заключены в пределах 5,5-6,5, ослабленных ,0- 5,, мертвых - ниже ,0 и более 7,0. Соответственно скорость роста
dxa
dt
+
dx(
dt
dxM dt
(2)
Клетки группы ха образуют биомассу со скоростью д за счет роста (увеличения объема) и почкования. Часть клеток этой группы переходит в труп- пу ослабленных хв со скоростью а/, . В свою очередь клетки группы х а воз1583 йЗ4
зращаются в группу активных х
0(. Обозначим эту скорость е/г. Также вероятна гибель другой части клеток группы ослабленных хв со скоростью с/э .
Составляют систему балансных уравнений по каждой из трех групп клеток при непрерывном культивировании:
dxc
dt
( -
+ оЛ, X,. - DX
at
-j - dt
5 Х °Хй
- ./ v - DV - «Э ллч
(3)
Подставляют (3) в (2), получают
3d
dx
----- - /э хс
(Dx н- Dx0 + DxM)
/ЗХа - D(xa + х0
Известно, что dx
-зг Х (0
(5)
Сравнивают уравнения () и (5) , получают расчетную формулу для удельной скорости роста микроорганизмов:
и- л
XJ
(5)
или с учетом уравнения (1) оконча- 3§ тельно получают
/5
Хч
xw +
хо + хм
(6)
0
5
В процессе промышленного культивирования микроорганизмов через определенные интервалы времени (например, через 2 ч) определяют количест- во активных х«5 ослабленных х„ и мертвых клеток xw. Используя эти экспериментальные данные, по значениям хд, х0, хм определяют коэффициенты , с/, ,е(г , о/э из системы балансных уравнений (3) подбором таких значений указанных коэффициентов, которые обеспечили бы в заданном интервале времени максимальное сближение расчетных и экспериментальных
, т.е. из условия
значении ха, х 9
минимума суммы квадратов разностей экспериментальных и расчетных значений активных, ослабленных и мертвых клеток на заданном отрезке времени. Затем умножают коэффициент пропорциональности р на относительное содер(--3) и полужание активных клеток
чают искомый параметр - значение удельной скорости роста |и „
Пример 1. В начале эксперимента, когда популяция дрожжей Candida maltosa ВСБ-569 находится в стационарном состоянии при D 0,16 , концентрация биомассы составляет 16,5 г/л, из которых 9,5 г/л приходится на активные, 7 г/л - на ослабленные клетки, мертвых клеток не обнаружено. Определение содержания (в %)
ч«
м
в популяции проводят в
результате сопоставления данных по морфологии, скорости роста клеток и их оптической плотности при использовании сканирующего комплекса микроскопа - ЭВМ Протва. К активным клеткам относят клетки с условной оптической плотность1; 5,5-6,5. Обозначена эта активная группа клеток как xq. Мертвые клетки характеризуются низкими значениями (ниже А) и высокими (более 7) значениями оптической плотности. Эти клетки не изменяют своего объема. Эта группа клеток обозначена xw. Остальная часть популяции отнесена к ослабленной группе х. Она характеризуется значением оптической плотности 5,-,0. После резкого уменьшения скорости разбавления до 0,08 происходит быстрое снижение количества активных клеток почти до нуля (до 0,5 г/л через А ч после смены протока) , а затем число их вновь увеличивается до 2 г/л и стабилизируется на этом уровне (фиг.1, кривая 2). Среднеквадратическая ошибка при оп- , ределении активных клеток экспериментальным путем составляет 10%. Расчетные данные коэффициентов по активным клеткам составляют: р 0,4; /, 0,3; /t 0,02. Кривая изменения скорости роста, полученная с помощью уравнения (6), показывает более плавное изменение (и (фиг.1, кривая 6), соответствующее более плавному теоретическому изменению количества активных клеток (фиг.1, кривая 5) по сравнению с экспериментальными данными.
Расчетные данные позволяют выявить на 5 ч ранее, чем экспериментальные, стабилизацию величины удельной скорости роста после смены протока (фигч кривые 3 и 6) . Следовательно,
подобранные значения кинетических
коэффициентов /э; ; к формулам (3) и (6) корректны, а сама формула (6) может быть использована для оцен- ки и в переходном и в установившемся, но с определенной лимитацией.
Через 3 ч Хо, 2,5; х 13, следовательно
10
М °.°77.
Точность Ј (фиг.1) определяется как Ј -.-- .100%,так как
Iм юе (4 , го
0
Миае V го Ј 3,75%.
Известным способом (Л можно определить через 6 ч с точностью Ј
( use- у
иэв
- 10%. Точность опре5
деления р составляет 3,75% через 3 ч, в го время как при определении известным способом 10% и только через 6 ч.
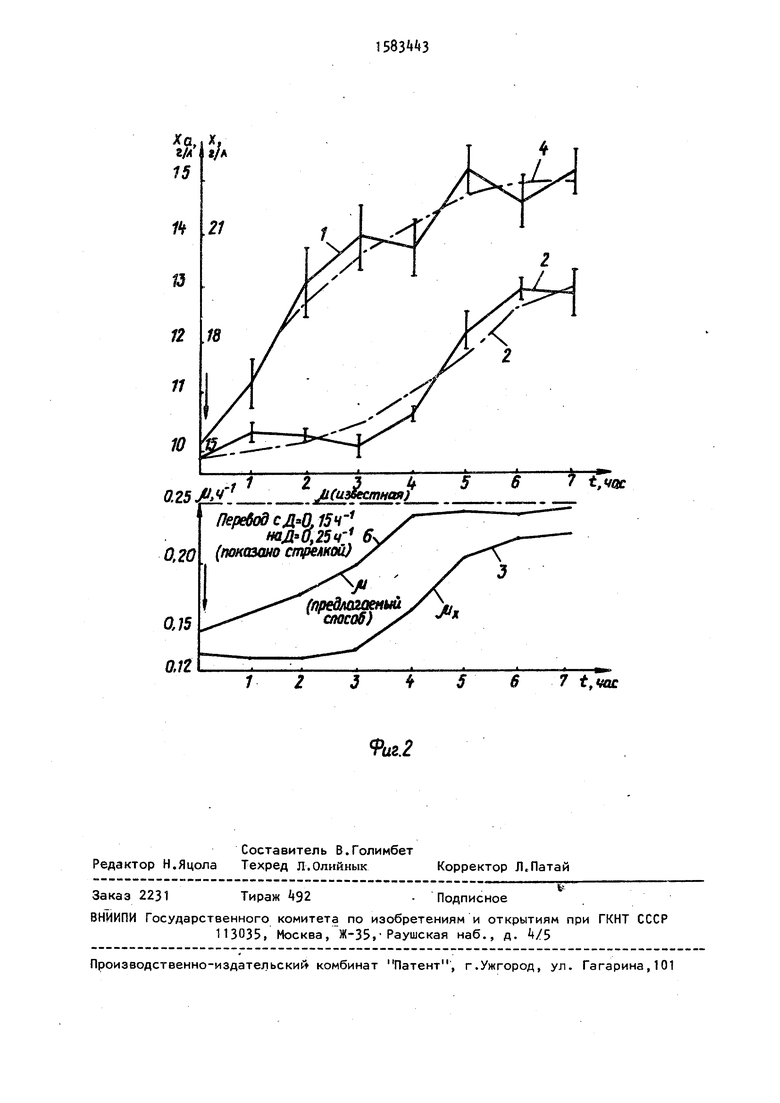
П р и м е р 2. Популяция Candida maltosa ВСБ-569 находится сначала в стационарном состоянии при D
0,15 ч
-i
Т,5 г/л,
0
9,8 г/л, х ,7 г/л, хм практио - м чески нет. После увеличения скорости
разбавления до 0,25 происходит быстрое возрастание количества активных клеток до ха 13,3 г/л и более плавное (инерционное изменение
5 общего количества биомассы х, х0и х определяют как и в предыдущем примере. Через 5 ч после смены протока х 18 г/л, хм 15,7 г/л. Ис- . i пользуя полученные экспериментальные
0 данные.рассчитывают кинетические коэффициенты р ; о,, {г и изменение во врегени теоретических значений х, хй, fX по уравнениям (3), (5) и (6). Через 4 ч хс( 12,5 г/л, х 23 г/л,
5 р 0,f. Точность определения и составляет Ј k%-t pMJ6 0,23.
Величина р, определенная через 7 ч известным способом, составляет и 0,21, соответственно точность
0 определения будет . В данном примере точность определения t в 2 раза выше, а и определено на нее.
ч ра
Таким образом, определение удельной скорости роста дрожжевой популяции по предлагаемому способу позволяет определить удельную скорость роста на 3-5 ч ранее и на 5-6% точнее, чем
известным способом, что позволяет повысить выход биомассы на 5%, так как точность и определяет и правильность Эыбора скорости разбавления D, кото- рая в установившемся режиме должна быть равна и . Следовательно, если Кроток выставлять по (j , определяемой по предлагаемому способу, то устанавливаться он будет с упрежде- нием во времени на 3-5 ч и точнее на 5%, чем по традиционно определя- 4мому значению . Так как продуктивность равна D -х (где х - концентрация биомассы), то и выход будет на $% больше, т.е. с 2,6 повысится до 2,73 r//h4
формула изобретения
Способ определения удельной скорости роста дрожжевой популяции, предусматривающий учет живых и мерт- фых клеток, отличающийся |ем, что, с целью повышения точности фпределения удельной скорости роста и увеличения временного упреждения $ ее определении, при учете живых клеток измеряют содержание активных клеток х, дающих прирост биомассы, и содержание ослабленных клеток xg, практически не дающих прироста био
массы, при этом измерение содержания активных, ослабленных и мертвых клеток хм проводят через равные промежутки времени на заданном временном отрезке, после чего по измеренным значениям х, х0, хм индентифициру- ют скорость роста активных клеток jb скорость перехода активных клеток в ослабленные ,, скорость перехода ослабленных клеток в активные /.г и скорость перехода ослабленных клеток в мертвые э из условия минимума суммы квадратов разностей измеренных и расчетных данных о количестве активных, ослабленных и мертвых клеток, по формулам
20
, , + Dx«;
dx0 dt
dt
о/, хи - , -0/3 хм- Dx
о
d
13 x«
- Dx
tA
где D - скорость разбавления, после чего рассчитывают удельную скорость р роста по формуле
х«
г Р
X м + X. + X
м


| название | год | авторы | номер документа |
|---|---|---|---|
| Система управления процессом культивирования микроорганизмов | 1988 |
|
SU1555698A2 |
| СПОСОБ ПОЛУЧЕНИЯ МИКРОСОМАЛЬНОЙ ФРАКЦИИ АЛКАНОКИСЛЯЮЩИХ ДРОЖЖЕЙ CANDIDA MALTOSA | 1991 |
|
RU2032735C1 |
| СПОСОБ ПОЛУЧЕНИЯ БИОМАССЫ МИКРООРГАНИЗМОВ | 1992 |
|
RU2093570C1 |
| Питательная среда для выращивания дрожжей | 1987 |
|
SU1578188A1 |
| ШТАММ ДРОЖЖЕЙ CANDIDA TROPICALIS - ПРОДУЦЕНТ БИОМАССЫ, ОБОГАЩЕННОЙ БЕЛКОМ | 1992 |
|
RU2031115C1 |
| СПОСОБ ПОЛУЧЕНИЯ БИОМАССЫ | 1994 |
|
RU2092560C1 |
| СПОСОБ ПОЛУЧЕНИЯ АЛЛЕРГЕНА ДЛЯ ВЫЯВЛЕНИЯ СЕНСИБИЛИЗАЦИИ ОРГАНИЗМА ГРИБАМИ РОДА CANDIDA | 1991 |
|
RU2031657C1 |
| СПОСОБ АКТИВАЦИИ МИКРОБИОЛОГИЧЕСКИХ ПРОЦЕССОВ, РОСТА РАСТЕНИЙ И КЛЕТОК РАСТЕНИЙ | 1995 |
|
RU2092547C1 |
| СПОСОБ ПОЛУЧЕНИЯ БИОМАССЫ ДРОЖЖЕЙ | 1987 |
|
SU1526223A1 |
| СПОСОБ ПОЛУЧЕНИЯ БИОМАССЫ ДРОЖЖЕЙ | 1988 |
|
RU1586177C |
Изобретение касается микробиологической промышленности и может быть использовано в системах автоматизированного контроля и управления процессом культивирования микроорганизмов при производстве белково-витаминных концентратов. Цель изобретения - повышение точности определения удельной скорости роста микробной популяции и увеличения временного упреждения в ее определении. Сущность способа заключается в том, что группу живых клеток разбивают на активные, дающие прирост биомассы, и ослабленные, практически не дающие прироста биомассы клетки, определяют относительное содержание активных клеток в популяции, идентифицируют коэффициент пропорциональности из условия минимума суммы квадратов разностей экспериментальных и расчетных данных о количестве активных , ослабленных и мертвых клеток на заданном отрезке времени, затем умножают коэффициент пропорциональности на относительное содержание активных клеток. 2 ил.
&
п
Лфйов c&Qf$v на&0,08ч (тшаано стрелкой)
/.«
О 1 2 S « 5
XV
jufia&aima }
(яргвлагагный
.
а 1 г з if 5 в t.v
Физ1
(яргвлагагный
.
1
«/А
.21
18
0.25 Л ч 1 1 Д(иЈктная$ ЧК
0,20
ПеребодсД 0.15ч-1
,25ч & (показано стрелкой)
0,15
O.J2
$567 ±,час
Фиг.2
| Кафаров В.В., Винаров А.Ю., Гордеев Л.С | |||
| Моделирование биохимических реакторов | |||
| - М.: Лесная промышленность, 1979.1 Печуркин Н.С | |||
| Динамика возвращения непрерывной культуры в активное состояние после снятия неблагоприятного воздействия | |||
| Инженерные проблемы микробиологического синтеза | |||
| - М.,ВИИИА, J969, с.249-251 | |||
| ( СПОСОБ ОПРЕДЕЛЕНИЯ УДЕЛЬНОЙ СКОРОСТИ РОСТА ДРОЖЖЕВОЙ ПОПУЛЯЦИИ |
