Изобретение касается способа пастеризации водного раствора иммуноглобулина и относится к нагреванию растворов иммуноглобулина с целью снижения или устранения опасности передачи вирусов при клиническом использовании этих материалов и для снижения содержания ферментных загрязнителей, которые могут вызывать клинические побочные эффекты, при одновременном отсутствии вредного влияния на структуру или функцию иммуноглобулина.
Коммерческие иммуноглобулиновые препараты содержат в основном молекулы иммуноглобулина G(IgG) и широко используются для заместительной терапии в случае агаммаглобулинемии и гипогаммаглобулинемии при проведении пассивной иммунизации против патогенных организмов, при профилактике Rh-сенсибилизации и при лечении аутоиммунных состояний, таких как идиопатической тромбоцитопенической пурпуры (JTP). Основной способ введения - путем внутривенной или внутримышечной инъекции. Однако сообщалось, что высокие дозировки иммуноглобулинов при внутривенном введении передают гепатит Non A Non B и возможно другие вирусные инфекции.
Иммуноглобулины обычно выделяют из плазмы или плацентарной крови с помощью холодно-спиртовых методов по Кону, ионообменной хроматографии и множество других опубликованных методов, которые зависят от физических, химических или иммунологических характеристик участвующих белков.
Применяется несколько методик для инактивации вирусов, непреднамеренно загрязняющих концентраты фактора VIII, другой белковый препарат, получаемый из тех же источников человеческой плазмы, как и раствору иммуноглобулина. Предприняты попытки использовать "паровую обработку", сухо-тепловую обработку" или химическую обработку, например с применением бета-пропионлактона (BPL) и ультрафиолетовое облучение или воздействие холата тринитробутилфосфата (ТНВФ). Концентрат фактора VIII, после нагревания в течение 72 ч при 60оС в лиофилизированном состоянии (или после обработки сухим теплом, передавал NANB-гепатит II из 13 больных гемофилией (см. Colombo "M et" al, Lancet, 1985, 2: 1-4). Случаи NANB-гепатита не зарегистрированы, но трое пациентов, получавших концентрат, нагревавшийся во влажном состоянии под парциальным давлением пара (см. Schimpf K. et al. , New England J of Me dicine 316, 918 (1987)). Однако эти методики не дают в итоге продукт, не имеющий концентрации вирусов гепатита. Использование BPL (УФ-облучения), хотя и устраняет вирусные загрязнители, может приводить к химической модификации белковых молекулы и к образованию нео-антигенов, которые могут вызывать клинические реакции.
Лабораторные исследования показали, что холат (ТNBP) весьма эффективен с точки зрения удаления покрытых липидами вирусов из продуктов коагуляции. Ограниченное клиническое изучение на семи пациентах, ранее не подвергавшихся воздействию препаратов крови, показало, что передачи гепатита не происходит (см. Horavit MS. et al. Thrombosis and Hemostasis, 1987, 58 (1) р. 371. Однако этот метод применим лишь к разрушению окруженных липидами вирусов. Последующее удаление использованных при осуществлении метода химических веществ оказывается, к сожалению, довольно затруднительным.
Пастеризация, т. е. тепловое воздействие на растворы, весьма эффективна для инактивации вирусов и других организмов. Пастеризационная обработка молока обычно осуществляется при 71,7оС в течение 15 с (см. Pelezar M. J. et al. , Microbiology 4-е издание. Изд-во Me Graiv Hill, 1977, р. 833-835). Показано, что эта обработка эффективно инактивирует Mycobacterium tuberculosis наиболее стойкий из ныне известных патогенов. Случаи передачи сывороточного гепатита или какого-либо иного вирусного агента пастерированным нормальным альбумином сыворотки не известны на протяжении более, чем 35 лет (см. Edsall J. T. , vox Sang 46 : 338-340. 1984). Пастеризация концентратора фактора VIII в присутствии стабилизаторов, таких как сахара и аминокислоты, сильно сократила передачу гепатита (см. Schimps K et al. , op. cit). Пастеризация антитромбина III, еще одного препарата плазмы, в присутствии 0,5 М цитрата была необходимой процедурой в течение ряда лет и после ее применения о передаче гепатита не соблюдалось. Тем самым, пастеризация иммуноглобулина представляется весьма привлекательной возможностью инактивации вируса. Однако известно, что молекулы JgG агрегируются при нагревании растворов (см. Rosen gvist E et al, Molecular lmmunolody 24, N 5, р. 495-501. 1987. Действительно, нагревание раствора IgG при 63оС в течение 15 мин является широко распространенным методом получения растворимых агрегатов IgG. Такие агрегаты обладают свойствами, аналогичными свойствам комплексов антиген-антитела, а именно они фиксируют комплемент, связываются с макрофагами и вызывают реакции Артуса (см. Christian C. L. lmmunogy 84, 112-121). , Активация системы комплимента FC-областями агрегированных молекул IgG обусловила многочисленные попытки получения формы IgG, пригодной для внутривенного использования, либо путем модифицикации процедуры выделения IgG из плазмы, либо модификацией или удалением FC-области (см. Мс Сеlland D. B. L. Yap P. L. Clinics in Haematology, 13. 1, февраль 1984). Следовательно, общая идея о том, что иммуноглобулины не могут быть нагреты в растворенном состоянии без образования агрегатов, является неотъемлемой частью концепций химии иммуноглобулинов на протяжении многих лет. Fernandes P. M. et al (см. патент США N 4440679) и Hirao Y et al. (см. опубликованная заявка на Европейский патент N 0196761) заявляют, что иммуноглобулин может быть нагрет в растворе и сохраняет свои биохимические характеристики, но эта процедура требует присутствия значительных количеств сахаров или сахароспиртов в качестве первичного стабилизатора (до 50% мас. /об. ) и выше). Hirao et al. заявляют, что дополнительный стабилизатор, которым может быть соль органической карбоновой кислоты или ПАВ, обеспечивает дополнительную стабильность. Однако присутствие больших количеств сахаров в качестве стабилизаторов также может стабилизировать вирусы. Сахароза используется в стабилизирующем препарате для консервации риккетский и вирусов после лииофилизации (см. Bovarnich et al, J. Bac teriology 59, 509-522, 1950), Ng P. K et al. (см. Thrombosis Resеarch 39, 439-446. 1985). Известно, что добавление сахарозы обеспечивает защиту от инактивации парвовирусу свиней при пастеризации раствора фактора VIII. Кроме того весьма затруднительно удаление значительных количеств сахаров или сахароспиртов.
Предлагаемый способ позволяет осуществить пастеризацию растворов иммуноглобулина в отсутствии какого-либо стабилизатора для инактивации вирусов без существенного изменения молекул IgG или их физиологической активности. Было показано, что когда растворы иммуноглобулина нагревают при низком уровне ионной силы и мягкокислотном рН, биологическая активность сохраняется и происходит минимальное агрегирование (менее 5% ). Это целиком укладывается в критерии, рекомендованные ВОЗ в отношении иммуноглобулиновых препаратов, пригодных для внутривенного употребления (см. WHO/BS (83.1396. Приложение 2).
В соответствии с предлагаемым способом пастеризацию разбавленных водных растворов IgG обычно в виде водного раствора концентрацией менее 5% (мас. /об. ), предпочтительно 2% (мас. /об. ), проводят при умеренно повышенной температуре, обычно в диапазоне 45 - 75оС, предпочтительно 50 - 60оС, в мягко-кислотных условиях, обычно при рН 3,5 - 6,5, предпочтительно рН 4 - 6, при низкой ионной силе, обычно эквивалентной водному раствору. Ионная сила не превышает ионной силы водного 0,05 мМ раствора NaCl.
Раствор иммуноглобулина, обработанный в соответствии с изобретением, обычно имеет низкую концентрацию белка, обычно менее 5 мас. % , предпочтительно менее 2 мас. /об. % .
Время нагревания раствора белка для обеспечения инактивации присутствующих вирусов зависит от типа и концентрации присутствующих вирусов и температуры нагрева. В типовом случае необходимо время нагревания не менее 10 ч.
В экспериментах с использованием в качестве модельных вирусов Синдбис человеческого иммунодефицита-1 коровьей оспы было показано, что нагревание разбавленного раствора IgG при рН 4 - 6 в течение 10 ч при 56 - 60оС было как минимум столь же эффективным с точки зрения разрушения вирусов, как и пастеризация 25% -ного нормального раствора альбумина сыворотки при этой температуре, и более эффективным, чем пастеризация раствора фактора VIII в присутствии сахаров и глицина в качестве стабилизаторов. Пастеризация иммуноглобулинов также ведет к снижению содержания или устранению некоторых ферментных загрязнителей, которые могут вызывать клинические реакции (см. Aloing B. M. et al. lmmunohemotherapy под ред. Nydegger. Изд-во Academic press. 1981. Такие загрязнители могут вызывать сокращение времени превращения неактивированного частичного тромбопластина и образование кининогенинов при анализе in vitro.
Предлагаемый способ не ограничивается пастеризацией иммуноглобулинов, которые получают для клинического употребления существующими коммерческими методами, такими как фракционирование по Кону, но метод получения иммуноглобулинов не имеет отношения к полезности предлагаемого процесса и иммунноглобулины, полученные из плазмы другими способами или полученные методами генной инженерии и выделенные из ферментационных бульонов, а также растворы, полученные от траногенных животных, в равной степени могут быть подвергнуты пастеризации этим методом.
Коммерческими являются иммуноглобулины типа, который отличается мультивалентностью, т. е. они являются продуктом, являющимся смесью антител против большого числа эпитопов. Эти иммуноглобулины известны также под названием поликлональных. Предлагаемый способ пастеризации применим также к так называемым моноклональным антителам, которые получают из гибридомных клеток и которые не являются мультивалентными, а специфичны для одного конкретного эпитопа.
Поэтому предлагаемый способ применим для пастеризации широкого множества растворов нативных и модифицированных иммуноглобулинов, в том числе растворов гаммаглобулина (JgG), смеси иммуноглобулинов, полученных непосредственно из крови и представляющих собой главным образом гаммаглобулин вместе с менее, чем 5% других классов иммуноглобулинов, в том числе JgA, IgM, IgE, внутривенного иммуноглобулина (NIG) гипериммунных глобулинов, таких как иммуноглобулин Rho(D) и иммуноглобулин гепатита В, моноклонального антитела класса IgG, предпочтительно подкласса IgG1, и белка человеческой миеломы, относимого к классу IgG, предпочтительно к подклассу IgG1.
П р и м е р 1. Этот пример иллюстрирует пастеризацию раствора гаммаглобулина.
Гаммаглобулин получают из фракции II + III по Кону, основываясь на методе осаждения спиртом по Кону (см. J. A. C. S т. 68, р. 459-475, 1946). Фракцию II получают по методике Кистлера и соавторов (см. vok Sanguinis 7,414. 1962). Фракцию II растворяют и осветляют фильтрованием, рН доводят до 4,0±0,3. Этанол и соли удаляют диафильтрацией на ультрафильтрационном аппарате Millipore Cassitle System M. W. CO 100.000). Для обеспечения удаления соли раствор диализируют дополнительно относительно дистиллированной воды в течение 60 ч при 4оС. Концентрацию белка доводят до 1 мас. /об. % дистиллированной водой и рН доводят до 5,0 0,1 н. хлористоводородной кислотой. Раствор нагревают 10 ч при 60оС.
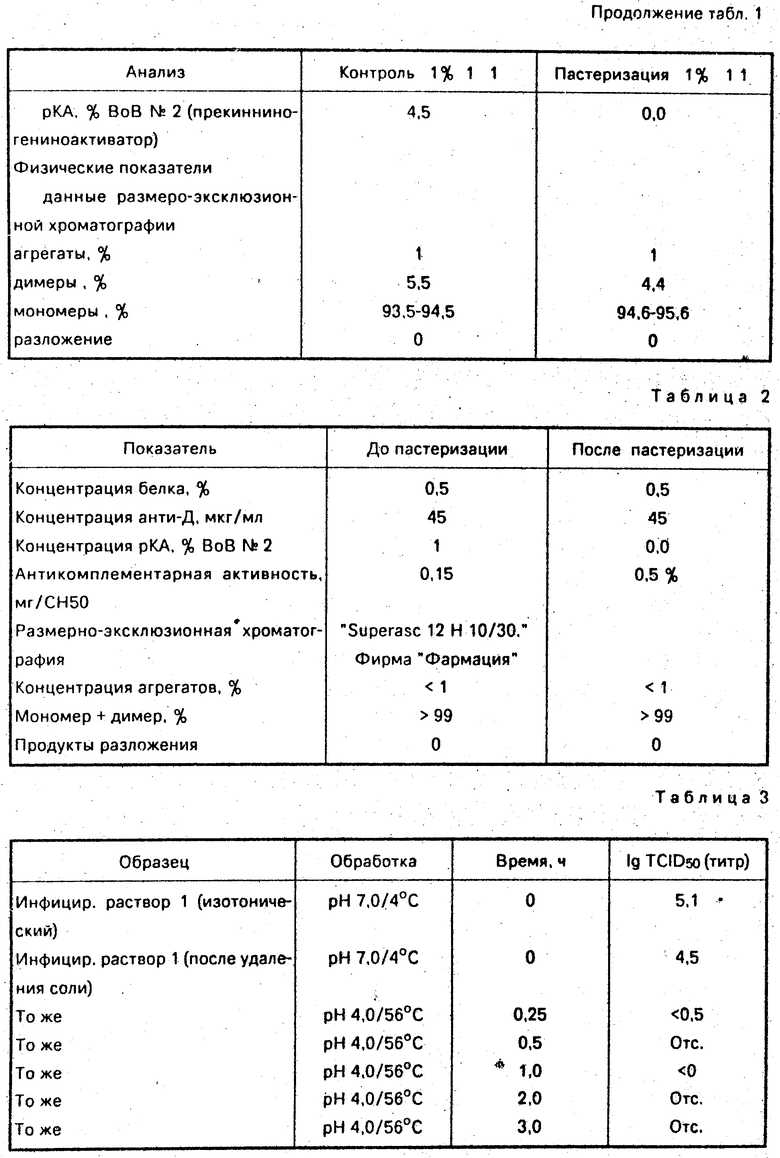
Пробы до и после пастеризации анализируют на концентрацию различных вирусов и бактериальных антител. Какого-либо снижения концентрации какого-нибудь из антител обнаружено не было. Не было отмечено также какого-либо вредного эффекта в результате пастеризации при испытании проб на антикомплементарную активность, прекининогенино-активаторную активность (рКА), распределение размеров молекул (с помощью размеро-эксклюзионной хроматографии), и СДН ПАГЭ (для определения состава белков). Нежелательная рКА-активность в продукте фактически устранялась в результате пастеризации.
Полученные результаты сведены в табл. 1.
СДН-ПАГЭ После пастеризации никаких дополнительных полос не наблюдалось.
П р и м е р 2. Этот пример иллюстрирует пастеризацию внутривенного иммуноглобулина (1V1G).
Лиофилизированный препарат внутривенного иммуноглобулина, состоящий из нативного 1gG, не подвергнутого предварительно модификации химическими или ферментными средствами, разбавляют дистиллированной водой. Раствор глубоко диализируют относительно дистиллированной воды при 4оС для удаления глюкозы и солей. Концентрация белка снижают до 1% (мас. /об. ) добавлением дистиллированной воды, а рН доводят до 5,0 0,1 н. хлористоводородной кислотой. Проводимость раствора составляет 110 микроS (25оС). Пастеризацию проводят при 60оС в течение 10 ч.
Чтобы убедиться в отсутствии повреждений в структуре 1gG в результате пастеризации, проводят следующие проверки:
1. Определяют спектры кругового дихроизма в полосе 210-320 нм для растворов иммуноглобулина до и после пастеризации. Эти спектры получают на спектрополяриметре ORD/CD-15 при 25оС. КД-спектра были идентичными.
2. УФ-спектры двух растворов, до и после пастеризации также были идентичными в полосе 240-350 нм при использовании спектрофотометра CaRY 219 VV/Vi1.
Поэтому никаких значительных конформационных изменений не происходит в трехмерной структуре белков в результате пастеризации раствора в течение 10 ч при 60оС и рН 5,0.
Концентрация агрегатов, определенная методом размероэксклюзионной хроматографии, увеличилась с менее, чем 1% , до 2% после пастеризации, что существенно ниже 5% -предела, рекомендованного Всемирной организацией здравоохранения (ВОЗ) для раствора иммуноглобулина, пригодного для внутривенного употребления. Продукты разложения не выявлены.
Однако, растворы иммуноглобулина до и после пастеризации сохраняют свою способность агрегироваться после восстановления изотоничности и нейтрального рН и после погружения в водяную баню при 62,5оС на 14 мин оба образца обеспечили получение около 20% агрегатов. После пастеризации не выявлено никакого изменения антикомплементарной активности и концентраций различных антител. Концентрация рКА снизилась после пастеризации до "недетектируемого" уровня.
Биологическую активность in vitro растворов иммуноглобулина до и после пастеризации оценивали с помощью анализов, включавших связывание иммуноглобулинов с первым компонентом комплемента (С1) и изменение конкурентного связывания с несущими Fe-рецептор промоноцитными клетками линии V937. Характеристики Fe-рецепторов в линии клеток V937 оказались аналогичны по сродству и структуре полипептидной цепи показателям для нормальных человеческих моноцитов и макрофагов (см. Anderson C. L. J. Exp. Med 156, 1794-1806, 1982). С1-связывание в обоих образцах демонстрирует нормальную активность с 50% -ным ингибированием на 6,5 мкМ и 5 мкМ соответственно. Те же образцы после теплового агрегирования при 62,5оС в течение 14 мин в изотонических условиях демонстрируют 50% -ное ингибирование С1-связывания на 0,0013 мкМ и 0,0035 мкМ, что соответствует более, чем 100-кратному увеличению связывания с С1.
Концентрации белка, необходимые для достижения 50% -ного ингибирования при конкурентном связывании с FC-рецепторами U937 для непастеризованных и пастеризованных образцов составили 15 нМ и 16 нМ. Это согласуется со значением 20 нМ (см. Law et al Molecular lmmunology, ) 23, 331-338. 1986) для препаратов иммуноглобулина с интактной FC-функцией. Сообщается, что для достижения того же эффекта требуется в два раза большее количество гамиммуна (товарный знак фирмы "Каттер лэбс"), восстановленного и алкилированного препарата иммуноглобулина.
Поэтому пастеризация не ведет к существенному увеличению связывающей способности IgG относительно С1 и не влияет на его взаимодействие с моноцитами и макрофагами. Хотя довольно трудно коррелировать результаты испытаний in vitro с характеристиками in vitro, эти наблюдения свидетельствуют об отсутствии значительных различий опсонической функции (благодаря взаимодействию между FC и нормальными человеческими моноцитами и макрофагами) и способности восполнять систему комплемента после связывания антигена, между иммуноглобулиновыми препаратами до и после пастеризации.
П р и м е р 3. Иллюстрирует пастеризацию иммуноглобулина Rho(D).
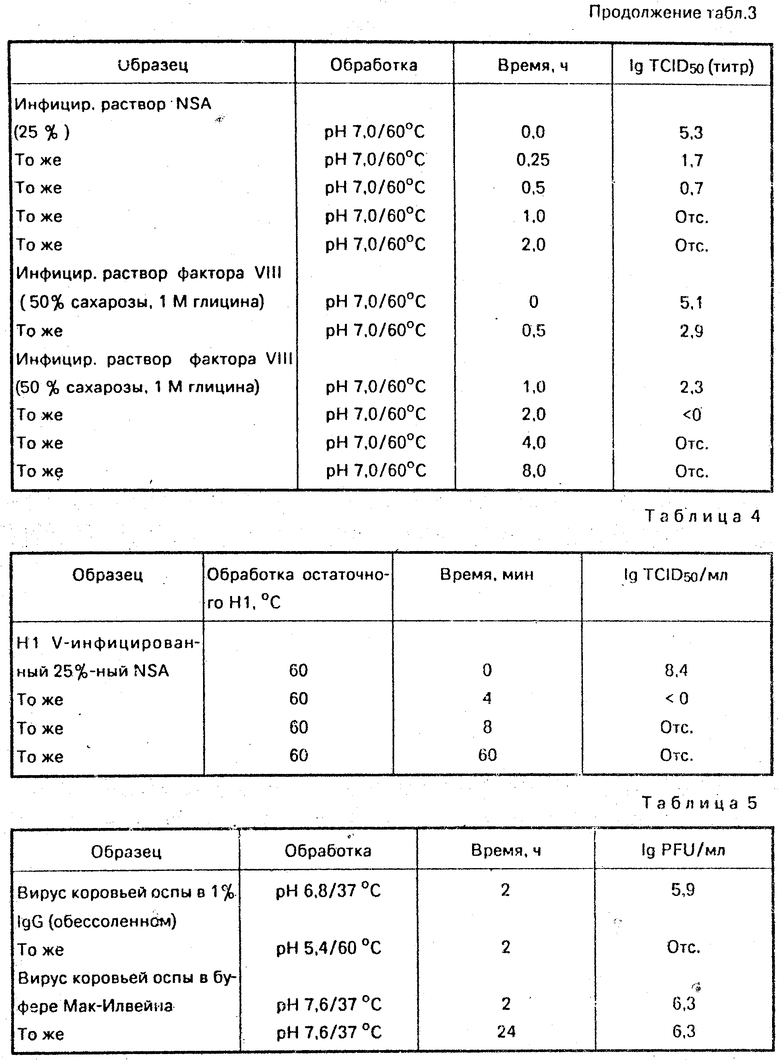
Порошок иммуноглобулина PhO(D) (112 г), полученный спиртовым фракционированием по Кону и обработанный с помощью ДЭАЭ-сефадекса А50 (фирма "Фармация", Швеция), растворяют в 706 мл дистиллированной воды. Раствор осветляют фильтрованием через мембранный фильтр и соль удаляют пропусканием через колонку сефадекс-G 25 (100 см х 10 см). Собирают объем выделений, содержащих белок, и разбавляют до концентрации приблизительно 0,5% (мас. /об. ).
рН доводят до 4,9 добавлением 4 мл 1,0 н. хлористоводородной кислоты. Разбавленный раствор заливают во флаконы по 500 мл, их герметизируют и пастеризуют в течение 11 ч на водяной бане при 60оС. Не выявлено снижения анти-D-способности. Концентрация агрегатов в образцах до и после пастеризации ниже 1% . После пастеризации не выявлено никакой рКА-активности и антикомплементарная активность не изменилась. Полученные результаты сведены в табл. 2.
П р и м е р 4. Иллюстрирует инактивирование вируса Синдбис и человеческого вируса-1 иммунодефицита (Н1V-1) в растворе иммуноглобулина в результате пастеризации.
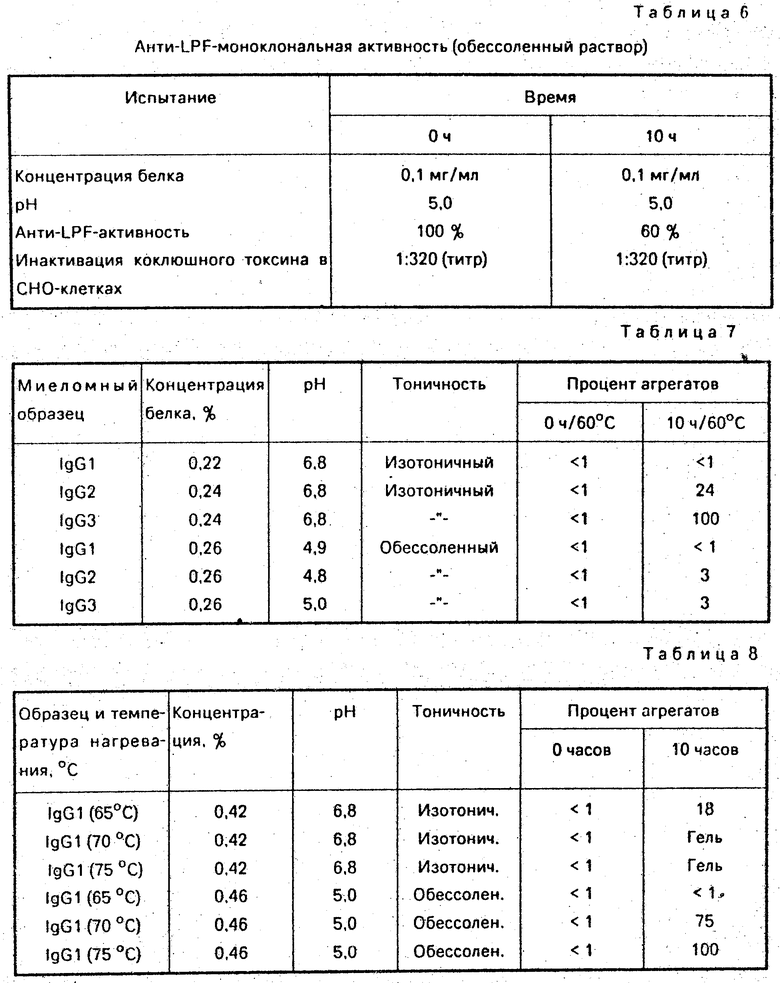
Сравнение инактивации вируса в растворах иммуноглобулина (рН 4,0 и бессолевой) и в растворе фактора VIII, содержащем 50% сахарозы, 1 М глицина (рН 7,0), проводили с использованием вируса Синдбис. К 1% -ному раствору иммуноглобулина (после удаления соли и при рН 4,0) добавляют затравтку вируса Синдбис. Раствор нагревают при 56оС на водяной бане. Пробы отбирают на определение инфективности вируса по прошествии 0,25; 0,5; 1,0; 2,0 и 3,0 ч и сразу же нейтрализуют добавлением 4,5% раствора карбоната натрия (рН 8,0).
Аналогично, 1% раствора фактора VIII, содержащий 50% сахарозы и 1,0 М глицина при рН 7,0, использовали для точечного введения идентичной затравки вируса Синдбис и нагревали при 60оС на водяной бане. Пробы отбирали и анализировали спустя 0,5; 1,0; 2,0; 4,0 и 8,0 ч.
Результаты определения остаточной вирусной инфективности, выраженные в виде десятичного логарифма инфективной дозы тканевой культуры (ТС1D50), сведены в таблицу 3.
Минимальное снижение, равное 4,5 lg ТС1D50, достигалось спустя всего 0,25 ч нагревания при 56оС в случае бессолевого раствора иммуноглобулина при рН 4,0. Никакой вирусной активности не выявлено спустя 2 и 3 ч. Аналогичная скорость инактивации вируса Синдбис в 25% -ном растворе нормального альбумина сыворотки (NSA) достигается при нагревании при 60оС. В случае стабилизированного раствора фактора VIII, даже при проведении нагревания при 60оС, снижение lg ТС1D50 составило всего 2,3 по прошествии 1 ч, и присутствие вируса не определяется спустя 4 ч нагревания при 60оС.
2. Аналогичный эксперимент повторяют на суспензии человеческого вируса-1 иммунодефицита (H1V-1) в 25% -ном NSA растворе. Оказалось, что скорость инактивации H1V в 25% -ном NSA-растворе гораздо выше, чем в случае вируса Синдбис (см. табл. 4). Все 8,5 lg TC1D50 исчезали спустя 8 мин при 60оС. Из-за более медленной скорости инактивирования вируса Синдбис в растворе иммуноглобулина при 56оС и в 25% -ном растворе при 60оС, вполне разумно предположить, что инфективности вируса Н1V, если он присутствует в растворе иммуноглобулина, будет значительно снижена в результате пастеризации при 60оС в течение 10 ч. Полученные результаты представлены в табл. 4.
П р и м е р 5. Этот пример иллюстрирует инактивацию вируса коровьей оспы в результате пастеризации.
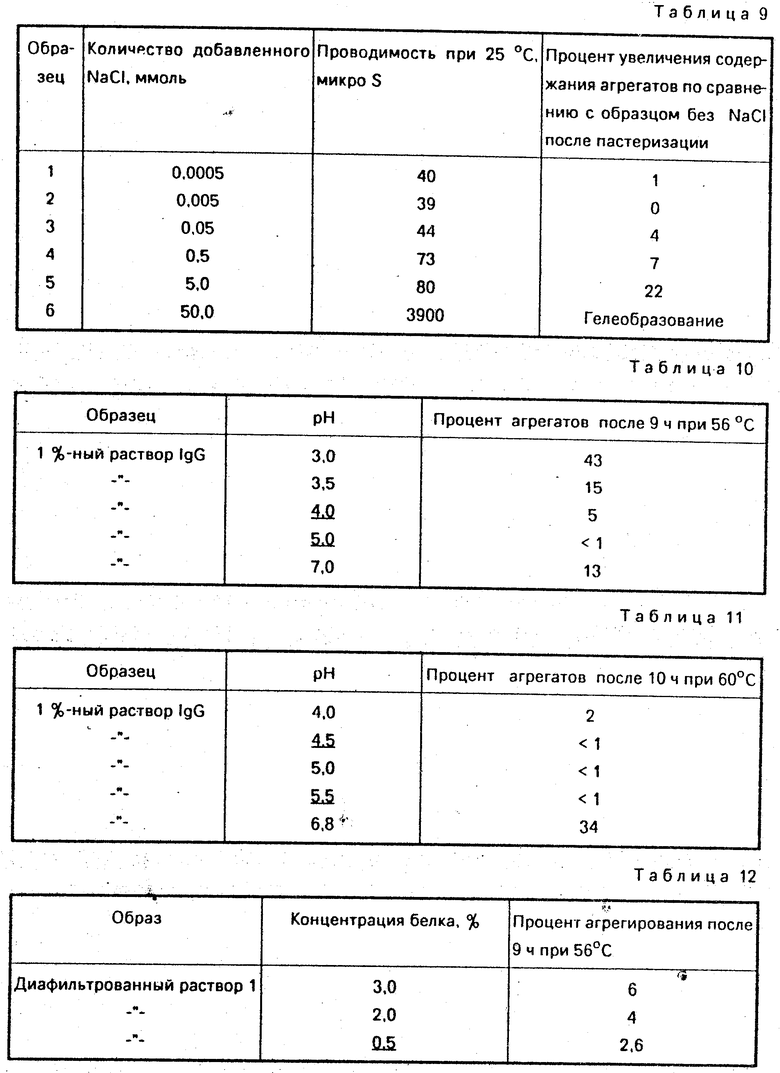
Затравку вируса коровьей оспы добавляют к обессоленному раствору иммуноглобулина. рН раствора составил 5,4. Пастеризацию проводят при 60оС в течение 2 ч. Позитивный контроль проб обеспечивался точечным заражением 1% -ного обессоленного раствора иммуноглобулина (рН 6,8) идентичной вирусной затравкой и инкубированием при 37оС в течение 2 ч. Остаточную вирусную инфективность определяют титрованием в оплодотворенных куриных яйцах и выражают величиной lg PFU (пустуло-образующих единиц) на миллилитр.
Полученные результаты сведены в таблицу 5.
Вся вирусная инфективность (lg PFU 6,3) разрушалась за 2 ч при 60оС. Инактивация происходила вовсе не из-за присутствия какой-либо возможной противооспененной активности, которая могла иметься в исходном растворе иммуноглобулина. Вирусная инфективность аналогичным образом зараженного раствора IgG оказалась неподверженной влиянию при инкубировании в течение 2 ч при 37оС. Индентичная оспенная затравка при разбавлении буфером Мак-Илвейна оказалась стабильной на протяжении 24 ч при 37оС. Hilfenhaus et al b vox Songuina 50, 208 (1986) сообщают о снижении инфективности вируса коровьей оспы, составляющей 6,2 lg ID50/мл, спустя 10 ч пастеризации в растворе фактора VIII, содержащем 54-ной сахарозы и 1,8 М глицина. Однако полная инактивация всего вируса коровьей оспы (6,7 lg ID50/мл), присутствовавшего до тепловой обработки, не достигалась к моменту окончания периода обработки.
Поэтому пастеризация обессоленных растворов иммуноглобулина при 60оС является более эффективной с точки зрения инактивации вируса коровьей оспы, чем пастеризация стабилизированного раствора фактора VIII, содержащего 50% -ной сахарозы и 1,0 - 2,0 М глицина.
П р и м е р 6. Этот пример иллюстрирует пастеризацию моноклональных антител.
Мононоклональные тела мышей на лимфоцито-промовирующий фактор (LPF, коклюшный токсин) выделяют из асцитической жидкости мышей. Антитела относятся к классу IgG1. После обессоливания раствора антител (0,5 мг/мл) на колонке G-25, собирали фракции выделительных объемов. рН доводят до 5,0 добавлением 0,06 М НСl. Пробу позитивного контроля готовят добавлением 0,02 М, трис-гидроглорида/0,12 М хлорида натрия, рН 6,8. Анти-LPF-активность определяют методом ELISA до и после пастеризации. Неспособность инактивировать коклюшный токсин в клетках яичника китайского хомяка (СНО) после пастеризации не нарушалось.
Полученные результаты сведены в нижеследующую табл. 6.
П р и м е р 7. Этот пример иллюстрирует пастеризацию человеческой миеломы IgG1, IgG2 и IgG3.
Сыворотку получают от трех пациентов с подтвержденными диагнозами множественной миеломы. Три миеломных белка относились к подклассам IgG1, IgG2 и IgG3. IgG выделяют из всех проб с помощью осаждения сульфатом аммония и ДЭАЭ-целлюлозной хроматографии (см. Stevenson G. T. u Dorrington K. J. Biochemikal Journal 118, 703. 1970).
Очищенные IgG (1,5 мл) глубоко диализируют относительно дистиллированной воды при 4оС. Диализированные растворы разбавляют до концентрации белка 0,26% (мас. /об. ) и доводят до рН 4,8±0,2 добавлением 0,06 М хлористоводородной кислоты. Образцы нагревают при 60оС в течение 10 ч. Образование агрегатов не выявлено в растворе IgG1, тогда как в растворах IgG2 и IgG3 после пастеризации определена концентрация агрегатов 3,3% , и 2,7% соответственно. Пробы позитивного контроля для трех белков были получены добавлением 0,02 М трис-гидрохлорида и 0,1,2 М хлорида натрия, рН 6,8. Эти растворы нагревают при 60оС в течение 10 ч. Изотонический раствор IgG1 был менее чувствительным, чем IgG2 или IgG3к агрегированию при нагревании при 60оС. Действительно, обнаружено менее 1% агрегатов.
Изотонический раствор IgG2 содержал 24% агрегатов после нагревания при 60оС, тогда как 100% изотонического раствора IgG3 было преобразовано в нерастворимые агрегаты.
Полученные результаты сведены в табл. 7.
Как можно видеть из табл. 7, при разбавленной концентрации белка IgG1, наблюдается слабое агрегирование или его совсем нет, независимо от того, является ли раствор изотоничным или обессоленным в условиях нагревания. Этот результат можно сравнивать с результатом, полученным при более высоких концентрациях и температурах (см. табл. 8).
П р и м е р 8. Этот пример иллюстрирует пастеризацию человеческого миеломного IgG1.
Очищенный IgG1 пациента с подтвержденным диагнозом множественной миеломы выделяют описанными в предыдущем примере путем. Очищенный раствор IgG1 при концентрации белка 18,3 мг/мл (1 мл) подвергают удалению соли на колонке G-25 (6 см х 1 см). Фракции выделительных объемов, содержащие белок, собирают воедино и разбавляют дистиллированной водой до концентрации белка 0,46% (мас. /об. ) рН раствор доводят до 5,0 добавлением 0,06 М хлористоводородной кислоты. Пробу позитивного контроля готовят добавлением 0,02 М трис-гидрохлорида и 0,12 М хлорида натрия, рН 6,8. Образцы нагревают в течение 10 ч при 65, 70 и 75оС.
Результаты определения распределения размеров молекул сведены в табл. 8.
Этот конкретный обессоленный миеломный IgG не дает агрегатов после нагревания при 65оС в течение 10 ч. Агрегирование происходит только когда материал нагревают при 70оС в течение 10 ч. Изотоничный IgG1, который не дает агрегатов при нагревании при 60оС в предыдущем примере, содержит 18% агрегатов после нагревания при 65оС. Гелеобразование происходит, когда нагревание проводят при 70оС или более высокой температуре.
П р и м е р 9. Этот пример иллюстрирует эффект ионной силы на агрегирование иммуноглобулинов.
К 0,5% -ному раствору иммуноглобулина, подвергнутому глубокому диализу, добавляют разные количества хлорида натрия. рН доводят до 5,0 добавлением 2 М хлористоводородной кислоты. Пастеризацию проводят при 60оС в течение 10 ч для каждого образца. Процент содержания агрегатов после пастеризации определяют методом размеро-эксклюзионной хроматографии (колонка "Фармация FPLC, супероза-12"). Полученные результаты приведены в табл. 9.
Как можно видеть из полученных результатов, добавление всего лишь 0,05 мМоль хлористого натрия ведет к значительному усилению образования агрегатов после пастеризации. Добавление 50 мМоль хлористого натрия приводит к гелеобразованию раствора, что свидетельствует о денатурации белка.
Из этих результатов совершенно ясно, что раствор иммуноглобулина должен иметь перед пастеризацией минимальную ионную силу, чтобы избежать агрегации иммуноглобулина. Это может быть достигнуто многими физико-химическими методами, такими как диализ, электродиализ, гель-фильтрация или ионообмен.
П р и м е р 10. Этот пример иллюстрирует влияние рН на агрегирование иммуноглобулином.
Образцы 1% -ного раствора иммуноглобулина доводят до рН 3,0; 3,5; 4,0; 5,0 и 7,0 добавлением либо 0,1 н. хлористоводородной кислоты, либо 0,5 н. гидроксида натрия. После этого образцы глубоко диализируют относительно воды, предварительно доведенной до требуемого рН. После диализа каждый раствор иммуноглобулина дополнительно доводят до исходного рН добавлением 0,1 н. хлористоводородной кислоты. рН образцов, первоначально имевших рН 5,0 и 7,0, доводки не требует. Пастеризацию проводят при 56оС в течение 9 ч для каждого образца. Результаты образования агрегатов в образцах сведены в табл. 10, .
Из приведенных в табл. 10 результатов видно, что лишь образцы при рН 4,0 и 5,0 дают минимальные концентрации агрегатов (5% ).
Аналогичный эксперимент проведен на 1% -ных растворах IgG при рН 4,0; 4,5; 5,0; 5,5; и 6,8. Растворы нагревают при 60оС в течение 10 ч. Приемлемые концентрации агрегатов (< 5% ) достигаются при рН в диапазоне 4,0 - 5,5. Полученные результаты представлены в табл. 11.
П р и м е р 11. Этот пример иллюстрирует влияние концентрации белка на агрегирование иммуноглобулинов
Раствор иммуноглобулина при рН 4,0 подвергают глубокой диафильтрации относительно дистиллированной воды, используя кассетную систему миллипорной ультрафильтрации (100, 000 MWCO) для удаления соли и этанола. Получают концентрированный раствор иммуноглобулина 10% (мас. /об. ) Разбавления этого раствора доводили до 3% ; 2% и 0,5% (мас. /об. ) дистиллированной водой. рН растворов доводили до рН 4,0 и растворы пастеризовали при 56оС в течение 9 ч. Полученные результаты сведены в табл. 12.
Концентрация агрегатов была наивысшей для 3% (мас. /об. ) раствора иммуноглобулина. Наинизшая из полученных концентраций агрегатов была для 0,5% -ного раствора. Очевидно, что для пастеризации с минимальным образованием агрегатов предпочтительна меньшая концентрация белка.
П р и м е р 12. Этот пример иллюстрирует влияние этанола на агрегирование иммуноглобулинов.
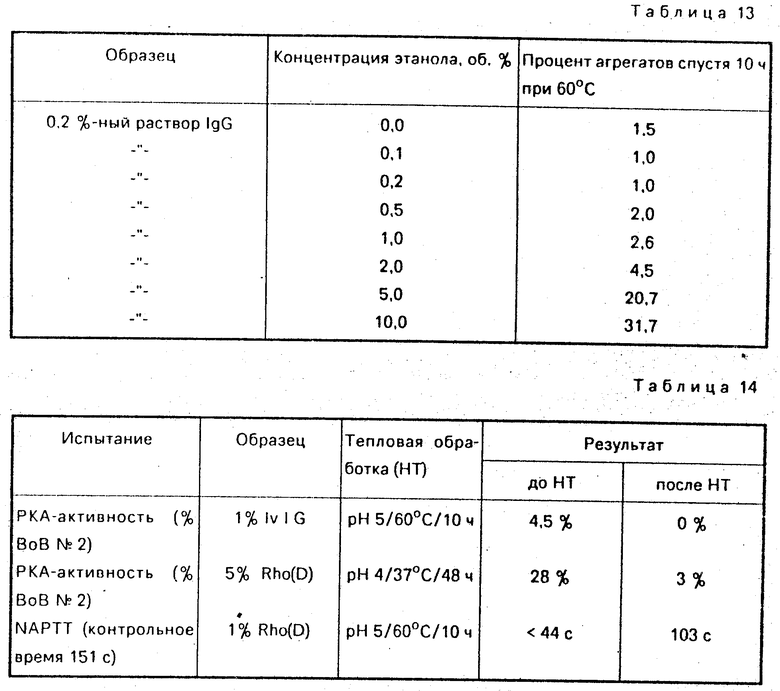
Раствор иммуноглобулина подвергают глубокому диализу при рН 5,0. Концентрацию белка доводят до 0,2% (мас. /об. ) добавлением дистиллированной воды. Добавляют разные количества этанола для доведения его концентрации в образцах до 0,1; 0,2; 0,5; 1,0; 2,0; 5,0 и 10% (мас. /об. ) Растворы подвергают пастеризации при 60оС в течение 10 ч. Полученные результаты представлены в сводном виде в табл. 13.
Данные в отношении образования агрегатов, представленные в табл. 13, показывают, что концентрация этанола свыше 0,5 мас. /об. вызывают значительную агрегацию молекул IgG.
П р и м е р 13. Этот пример иллюстрирует снижение содержания ферментных загрязнений в загрязненных растворах иммуноглобулина, обусловленных пастеризацией.
Ферментные загрязнители, которые могут присутствовать в иммуноглобулиновых препаратах, значительно снижают свое содержание в результате пастеризации. Влияние на прекининогениновый активатор (РКА) и время преобразования неактивированного частичного тромбопластина (NAPTT) представлено в табл. 14.
Пастеризация ликвидирует РКА-активность в 1% -ном растворе иммуноглобулина для внутривенного введения. Каждая обработка при рН4 и 37оС в течение 48 ч существенно снижает РКА-концентрацию с 28 до 3% . Величина NAPTT для 1% Rho(D) после пастеризации существенно удлиняется с менее чем 44 с до 103 с. Это указывает на ослабление прокоагулянтной активности, имеющей место в препарате.
Следовательно согласно изобретению предлагается новый способ пастеризации растворов иммуноглобулинов. Такая пастеризация инактивирует вирусы и снижает ферментную активность, не вызывая агрегирования или утраты биологической активности. В рамках предлагаемого способа возможны модификации. (56) Патент ЕР N 0196761, кл. A 61 L 2/04.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РАСТВОРА ГАММА-ГЛОБУЛИНА, ПРЕДНАЗНАЧЕННОГО ДЛЯ ВНУТРИВЕННОГО ВВЕДЕНИЯ, И ПРОДУКТ, ПОЛУЧАЕМЫЙ ЭТИМ СПОСОБОМ | 1998 |
|
RU2198668C2 |
| Способ получения иммуноглобулинов | 2016 |
|
RU2708394C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЛОБУЛИНОВ ДЛЯ ВНУТРИВЕННОГО ВВЕДЕНИЯ И ДРУГИЕ ИММУНОГЛОБУЛИНОВЫЕ ПРОДУКТЫ | 1999 |
|
RU2197500C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОМПОЗИЦИИ IgG ПОСРЕДСТВОМ ТЕПЛОВОЙ ОБРАБОТКИ | 2013 |
|
RU2636049C2 |
| ПРЕПАРАТ ИММУНОГЛОБУЛИНА ЧЕЛОВЕКА ПРОТИВ ЦИТОМЕГАЛОВИРУСА ДЛЯ ВНУТРИВЕННОГО ВВЕДЕНИЯ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2009 |
|
RU2404250C1 |
| СОСТАВ ДЛЯ СТАБИЛИЗАЦИИ ПЛАЗМЫ КРОВИ В ХОДЕ ПАСТЕРИЗАЦИИ И СПОСОБ ПАСТЕРИЗАЦИИ ПЛАЗМЫ КРОВИ | 1991 |
|
RU2045902C1 |
| СПОСОБ ПРОДУЦИРОВАНИЯ В ВЫСОКОЙ СТЕПЕНИ ВИРУСОБЕЗОПАСНЫХ КОМПОНЕНТОВ ДЛЯ ПОЛУЧЕНИЯ ФИБРИНОВОГО КЛЕЯ ИЗ ПУЛА ПЛАЗМЫ ЧЕЛОВЕКА | 1998 |
|
RU2236237C2 |
| ПРЕПАРАТ ИММУНОГЛОБУЛИНА IgG И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2005 |
|
RU2337109C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВЫСОКОЭФФЕКТИВНОГО ЧЕЛОВЕЧЕСКОГО АЛЬБУМИНА ДЛЯ ПРИМЕНЕНИЯ В ДЕТОКСИКАЦИОННОЙ ТЕРАПИИ | 2008 |
|
RU2424822C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЛОБУЛИНА ЧЕЛОВЕКА | 2013 |
|
RU2614119C9 |

Использование : изобретение относится к медицине и касается пастеризации водного раствора иммуноглобулина. Целью изобретения является повышение качества целевого продукта. Сущность изобретения заключается в тепловой обработке водного раствора иммуноглобулина в течение 0,25 - 10 ч при температуре 56 - 60С рН 4,0 - 5,5. Положительный эффект заключается в инактивации вирусов и снижении ферментативной активности . 7 з. п. ф-лы, 14 табл.
СПОСОБ ПАСТЕРИЗАЦИИ ВОДНОГО РАСТВОРА ИММУНОГЛОБУЛИНА путем его тепловой обработки, отличающийся тем, что, с целью повышения качества целевого продукта, длительность тепловой обработки составляет 0,25 - 10,0 ч при 56 - 60oС и рН 4,0 - 5,5.

