Изобретение относится к медицинской технике, биотехнологии и фармацевтической промышленности.
Изобретение может быт использовано в области медицинской и клинической биохимии, а также молекулярной фармакологии при исследовании биологически активных веществ (БАВ) и препаратов, "мишенью", которых являются природные и синтетические соединения, образующие сшивки между двухцепочечными линейными молекулами ДНК.
Большинство существующих методов определения БАВ, в частности ферментов, основано на измерении их активности с использованием либо природных, либо синтетических субстратов. Количественная оценка продуктов расщепления, а, следовательно, и пропорциональной им концентрации фермента, производится, как правило, при помощи оптического метода (спектрофотометрия) [1]
Несмотря на свою простоту, такой подход к определению ферментов не позволяет достигнуть высокой чувствительности. При помощи оптического метода удается достоверно определить количество ферментов, не превышающее 10-6 г [2]
Другой способ определения ферментов это использование радиоактивных субстратов. Однако здесь возникает ряд трудностей, связанных с нестабильностью радиоактивной метки, радиационным расщеплением субстрата или фермента и их инактивацией, необходимостью работы высококвалифицированного персонала и отсутствием высокочувствительных счетчиков радиоактивности.
Чувствительность определения ферментов можно повысить до 10-8 г, используя иммунохимический метод [3]
Однако и этот способ имеет ряд ограничений, определяемых его дороговизной и трудоемкостью, а также необходимостью в высококвалифицированном персонале. Кроме того, данные иммунохимического определения затрудняют вывод о том, имеет ли автор дело с активной формой фермента или произошла его инактивация. Это означает, что при определении наличия в среде БАВ, в частности ферментов, актуальной является задача по созданию простых и высокочувствительных методов, основанных на других принципах.
Известно, что двухцепочечные молекулы ДНК низкой молекулярной массы (1 х 106) образуют дисперсные жидкокристаллические фазы, представляющие собой упорядоченный ансамбль молекул ДНК, который имеет микроскопический размер (0,3-1,0 мкм) [4]
Жидкие кристаллы ДНК имеют ряд отличительных особенностей. К ним относятся: во-первых, высокая плотность упаковки и упорядоченный характер укладки молекул ДНК в жидких кристаллах, практически совпадающие с аналогичными параметрами, характерными для ДНК в составе биологических объектов; во-вторых, высокая лабильность пространственной структуры жидких кристаллов в ответ на изменение внешних условий и, наконец, аномальная оптическая активность, наблюдаемая при образовании жидких кристалллов ДНК холестерического типа, которая проявляется, в частности, в наличии интенсивной полосы в спектре кругового дихроизма (КД), расположенной в области поглощения азотистых оснований ДНК ( λ ≈260 нм). Амплитуда этой полосы может в десятки и сотни раз превышать амплитуду полосы в спектре КД, обусловленную молекулярной оптической активностью, характерной для исходных линейных молекул ДНК.
Взаимодействие некоторых природных при синтетических соединений с молекулами ДНК, сопровождающееся образованием не только комплексов с ДНК, но и сшивок между этими молекулами, не нарушает способности молекул ДНК к образованию дисперсной фазы в условиях фазового исключения [5] В составе частиц образующейся фазы молекулы ДНК по-прежнему упорядочены, однако в этом случае свойства частиц могут отличаться от свойств частиц дисперсной холестерической фазы, формируемой в этих же условиях исходными молекулами ДНК. В частности, дисперсные фазы комплексов могут не иметь аномальной оптической активности. Это означает, что гидролиз соединений, образующих сшивки между молекулами ДНК, вызываемый в контролируемых условиях ферментации или действием некоторых физических факторов, может привести к восстановлению структуры холестерического жидкого кристалла, которая характерна для исходных двухцепочечных ДНК. При этом процесс восстановления холестерической структуры жидкого кристалла будет сопровождаться появлением аномальной оптической активности. Следовательно, аномальная оптическая активность жидких кристаллов ДНК может быть использована в качестве нового критерия при определении наличия ферментов, БАВ или физических факторов, вызывающих гидролиз молекул природных или синтетических веществ, образующих сшивки между ДНК.
Предлагается новый биодатчик, обладающий высокой чувствительностью и позволяющий быстро определять БАВ, вызывающие гидролиз молекул, образующих сшивки между ДНК.
Для этого осуществляют формирование жидких кристаллов микроскопического размера из молекул комплекса ДНК-природное соединение. Отличительная особенность предлагаемого датчика состоит в том, что жидкие кристаллы сформированы из молекул комплекса ДНК-природное соединение таким образом и в таких условиях, что их аномальная оптическая активность, регистрируемая в ответ на расщепление природного соединения под действием фермента, выступает в качестве индикатора перестроения структуры жидкого кристалла.
Предлагаемый биодатчик представляет собой частицы лиотропной дисперсно жидкокристаллической фазы, сформированной из комплекса ДНК-природное или синтетическое соединение в водно-солевом растворе полимера, и помешенные в оптическую кювету.
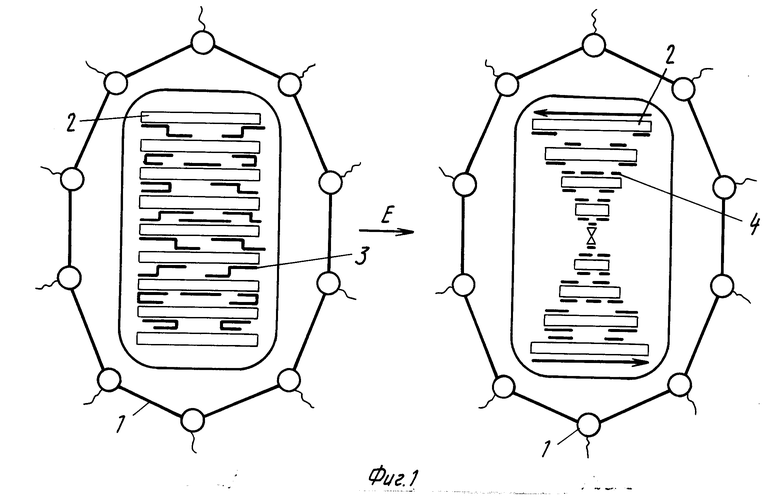
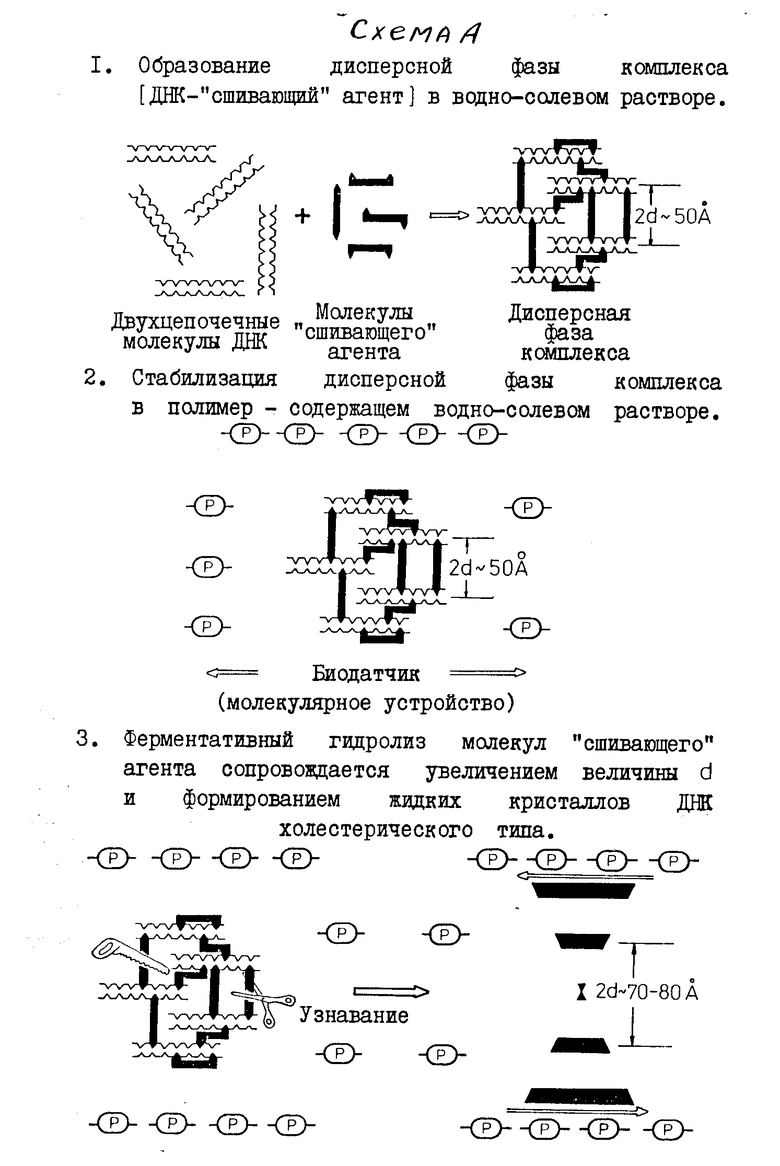
Схема предлагаемого биодатчика показана на фиг. 1, где 1 молекулы полимера, в водно-солевом растворе которого формируются жидкие кристаллы комплексов ДНК-стеллин В; 2 молекулы ДНК, образующие жидкие кристаллы (стрелки на правой части рисунка отражают наличие макроскопической спиральной закрутки слоев из молекул ДНК, образующих холестерический жидкий кристалл); 3 молекулы стеллина В, образующие сшивки между молекулами ДНК; 4 фрагменты молекул стеллина в после их ферментативного расщепления трипсином (Е).
Принцип действия датчика, состоит в том, что сначала из молекул ДНК и природного соединения формируют комплекс, затем из комплекса ДНК-природное соединение формируют дисперсную жидкокристаллическую фазу и расщепляют молекулы природного соединения в составе комплексов ДНК-природное соединение, образующих жидкие кристаллы, при помощи протеолотического фермента. В результате действия фермента сшивки между молекулами ДНК, образованные молекулами природного соединения, разрушаются и сами жидкие кристаллы переходят из оптически неактивного в оптически активное состояние. Принцип действия датчика показан на схеме А, где приводится структура молекулярного устройства (биодатчика), сконструированного на основе жидких кpисталлов двухцепочечных молекул ДНК, сшитых стеллином В. Приведенные конкретные параметры биодатчика установлены в результате рентгенографического анализа жидких кристаллов комплекса ДНК-стеллин В.
Изменение аномальной оптической активности, в частности спектров ДНК, наблюдаемое при перестроении структуры жидкого кристалла, позволяет не только детектировать наличие фермента в исследуемом растворе, но и открывает возможность для определения его типа, класса или группы по характеру наблюдаемых изменений.
П р и м е р 1. Приготовление исходных растворов.
1.1. Навеску NaCl (17,53 г) помещают в мерную колбу (N 1000 мл); в эту же колбу помещают навески NaH2PO4 ˙ H2O (1,315 г) и Na2HPO4 ˙ 12H2O (14,51 г), добавляют дистиллированную воду, растворяя вещества, и доводят объем до 1000 мл (Раствор 1.1).
1.2. Препарат ДНК молок осетровых рыб растворяют в растворе 1.1, дополнительно очищают от примесей, деполимеризуют (УЗ-дезинтегратор УЗДН-1 У4.2, частота 35 кГц, время обработки 1 мин, 4оС) и диализуют против раствора 1.1 для удаления низкомолекулярных примесей (Раствор 1.2).
Оптическая плотность раствора 1.2 при λ= 260 нм составляет 0,8 оптич.ед.
1.3 Препарат протамина (стеллин В) из гонад (50 мг) растворяют в 25 мл раствора 1.1 (Раствор 1.3).
1.4. 1 мг препарата трипсина растворяют в 10 мл раствора 1.1 (Раствор 1.4).
1.5. 0,1 мл раствора 1.4 помещают в мерную колбу (V 100 мл) и доводят объем до метки раствором 1.1 (раствор 1.5).
1.6. Навеску синтетического инертного, нейтрального и не поглощающего в УФ-области полимера (34 г), помещают в мерную колбу на 100 мл и добавляя раствор 1.1, доводят объем до метки (раствор 1.6).
П р и м е р 2. Получение комплекса ДНК-стеллин В.
2.1. К 10 мл раствора 1.2 при постоянном перемешивании добавляют 1 мл раствора 1.3. Таким способом получают раствор комплекса ДНК-стеллин В, в котором степень нейтрализации отрицательно заряженных групп ДНК положительно заряженными боковыми радикалами стеллина В составляет ≈ 0,6. (Раствор 2.1).
2.2. Контролируют свойства полученного комплекса при помощи оптических методов (УФ-спектроскопия, круговой дихроизм, оптическое вращение).
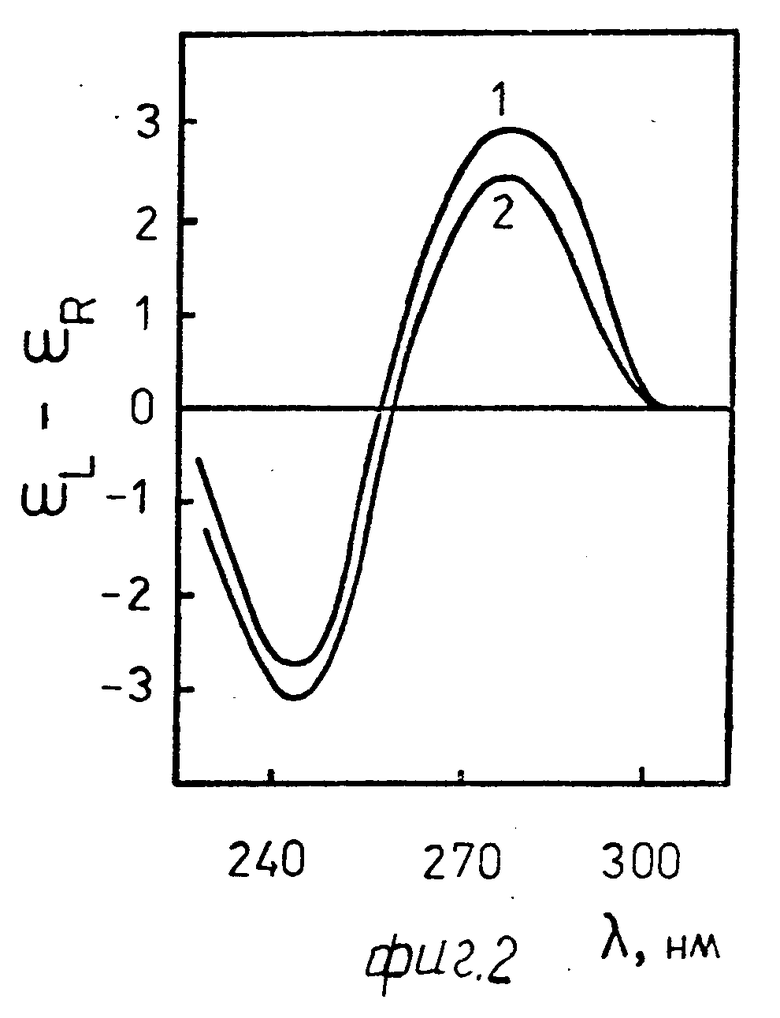
На фиг. 2 показаны спектры кругового дихроизма (КД).
Сопоставление кривых 1 и 2 на фиг. 2 показывает, что спектр КД водно-солевого раствора комплекса ДНК-стеллин В (кривая 2, r 0,6) практически не отличается от спектра КД, характерного для водно-солевого раствора исходной, линейной В-формы ДНК (кривая 1).
П р и м е р 3. Получение жидких кристаллов комплекса ДНК-стеллин В в водно-солевом растворе полимера.
3.1. Смешивают равные объемы растворов 1.6 и 2.1, интенсивно перемешивают полученную смесь и оставляют ее на 2 часа при комнатной температуре.
3.2. Контролируют свойства полученного жидкого кристалла при помощи оптических методов (УФ-спектроскопия, круговой дихроизм или оптическое вращение).
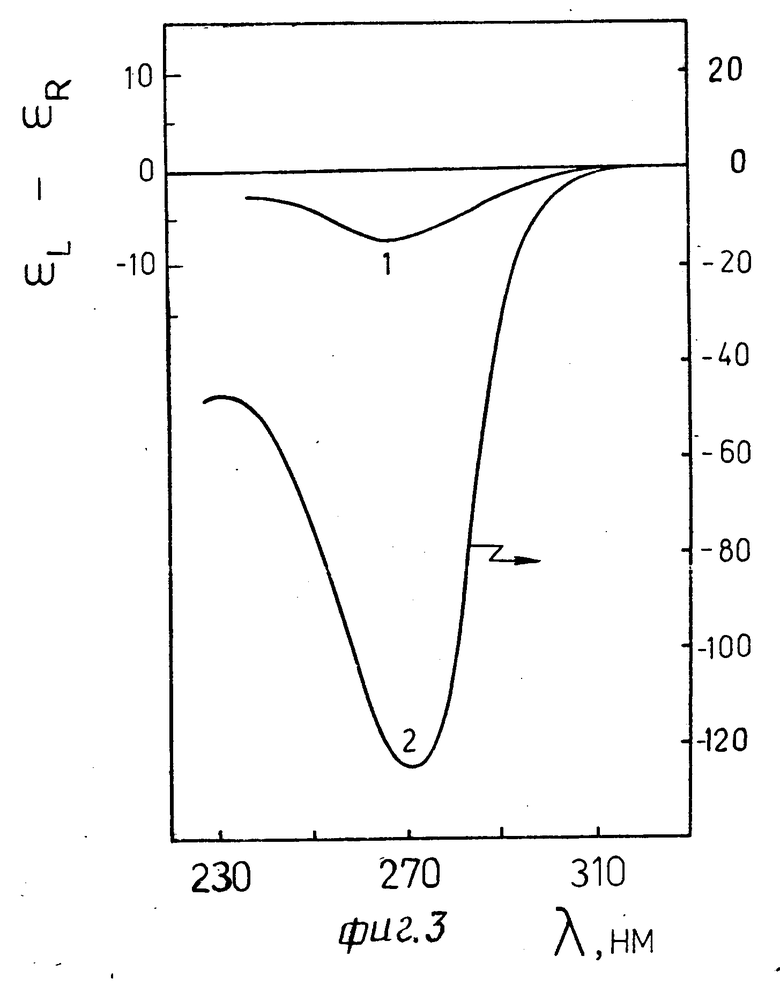
На фиг. 3 изображены спектры, КД жидкокристаллической дисперсии, сформированной в водно-солевом растворе полимера из молекул комплексам ДНК-стеллин В (r ≈0,6) до (кривая 1) и после (кривая 2) обработки ее трипсином (концентрация полимера 170 мг/мл; ионная сила раствора 0,15. Отсутствие аномальной оптической активности в полосе поглощения азотистых оснований ДНК свидетельствует об образовании в результате наличия сшивок между молекулами ДНК оптически неактивной жидкокристаллической фазы.
П р и м е р 4. Гидролиз трипсином молекул стеллина В в составе комплексов ДНК-стеллин В, образующих частицы дисперсной жидкокристаллической фазы в водно-солевом растворе полимера (10-6 г трипсина).
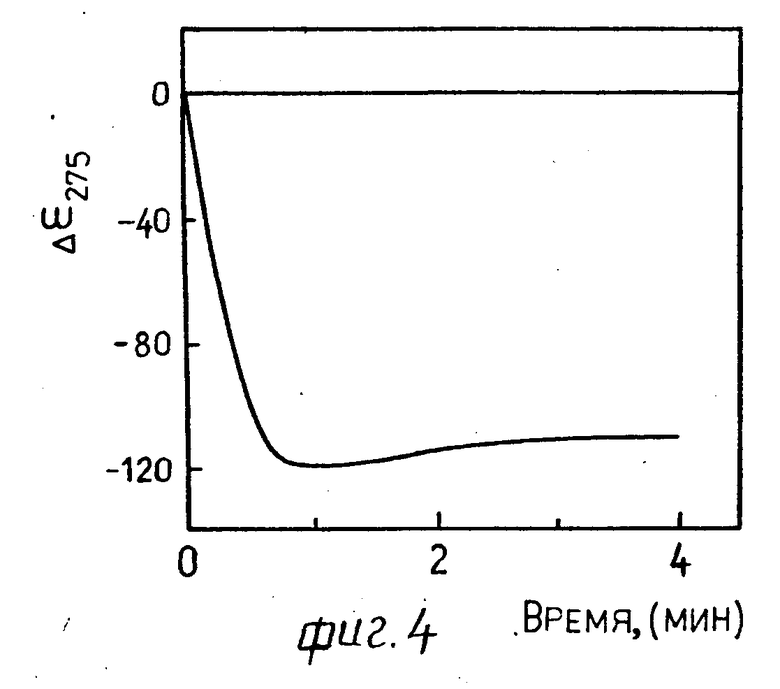
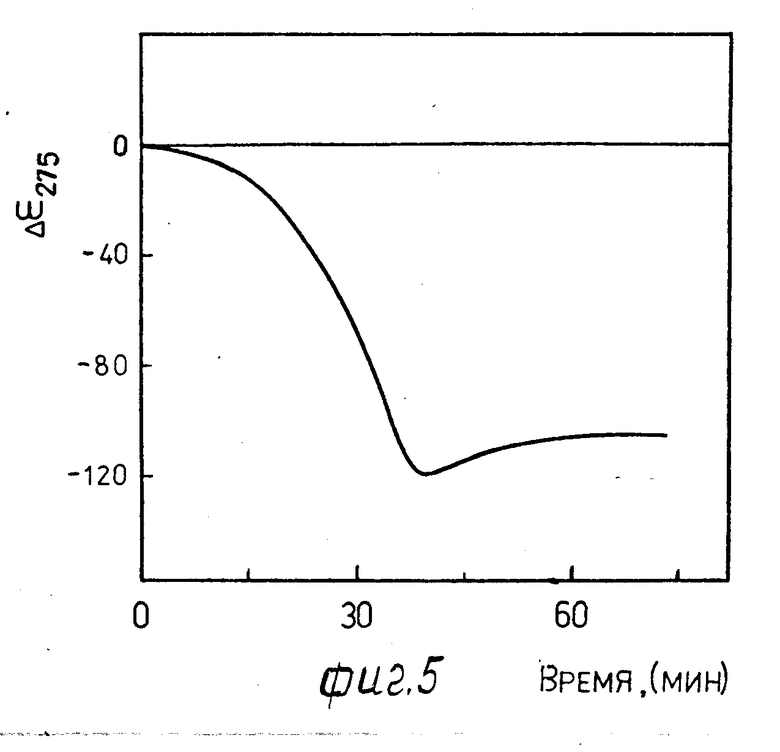
4.1. Раствор 3.1 инкубируют 10 мин при 37оС и добавляют 10 мкм раствора 1.4. Гидролиз проводят при 37оС.
На фиг. 4, 5 в качестве примера приведена кинематическая кривая нарастания аномальной оптической активности при λ275 нм в спектре КД жидкокристаллической дисперсии комплекса ДНК-стеллин В (r 0,6) после добавления трипсина (10-6 г фиг. 4; 10-9 г фиг. 5). Появление аномальной оптической активности в спектре КД жидкокристаллической дисперсии, сформированной по п. 3.1, свидетельствует о гидролизе молекул стеллина В, образующих сшивки между молекулами ДНК, трипсином, сопровождающимся перестройкой ее структуры, и приводящей к образованию жидкокристаллической фазы холестерического типа (ср. кривые 1 и 2 на фиг. 3).
П р и м е р 5. Гидролиз трипсином молекул стеллина В в составе комплексов ДНК-стеллин В, образующих частицы дисперсной жидкокристаллической фазы в водно-солевом растворе полимера (10-9 г трипсина).
5.1. Раствор 3.1 инкубируют 10 мин при 37оС и добавляют 10 мкл раствора 1.5. Гидролиз проводят при 37оС.
В полном соответствии с данными, приведенными на фиг. 3 и 4, в спектре КД также появляется аномальная оптическая активность; однако в отличие от случая п. 4.1 максимальный оптический эффект достигается уже не через 2, а приблизительно через 30-40 мин (см. данные фиг. 4 и 5).
Следует отметить, что при добавлении ингибитора трипсина диизопропилфторфосфата к растворам, приготовленным по п.п. 4 и 5, интенсивная отрицательная полоса в спектре КД не появляется. Этот факт показывает, что полоса в спектре КД появляется только в тех случаях, когда молекулы стеллина В в составе комплексов ДНК-стеллин В гидролизуются трипсином.
П р и м е р 6. Оптические текстуры тонких слоев жидких кристаллов, формируемых в водно-солевом растворе полимера из исходных молекул ДНК и их комплексов со стеллином В.
6.1. Препараты тонких слоев жидких кристаллов готовили по стандартной методике [4]
Текстура тонкого слоя ( ≈20 мкм) жидкого кристалла, сформированного из молекул комплекса ДНК-стеллин В, наблюдаемая в поляризованном свете, по своему внешнему виду напоминает закристаллизованную поликристаллическую массу; такого рода текстуры жидких кристаллов полимеров искусственного и биологического происхождения относятся к "неспецифическому" типу.
Гидролиз трипсином молекул стеллина В, образующих сшивки между молекулами ДНК, сопровождается перестроением структуры жидкого кристалла. Текстура характеризуется периодическим чередованием темных и светлых полос. Эта текстура известна по названием текстуры "отпечатков пальцев"; она характерна для холестерических жидких кристаллов, образованных низкомолекулярными соединениями, а также полимерами синтетического и биологического происхождения [7] Следовательно, результаты оптической микроскопии полностью подтверждают вывод о перестройке структуры жидкого кристалла комплекса ДНК-стеллин В, сделанный на основании измерений аномальной оптической активности.
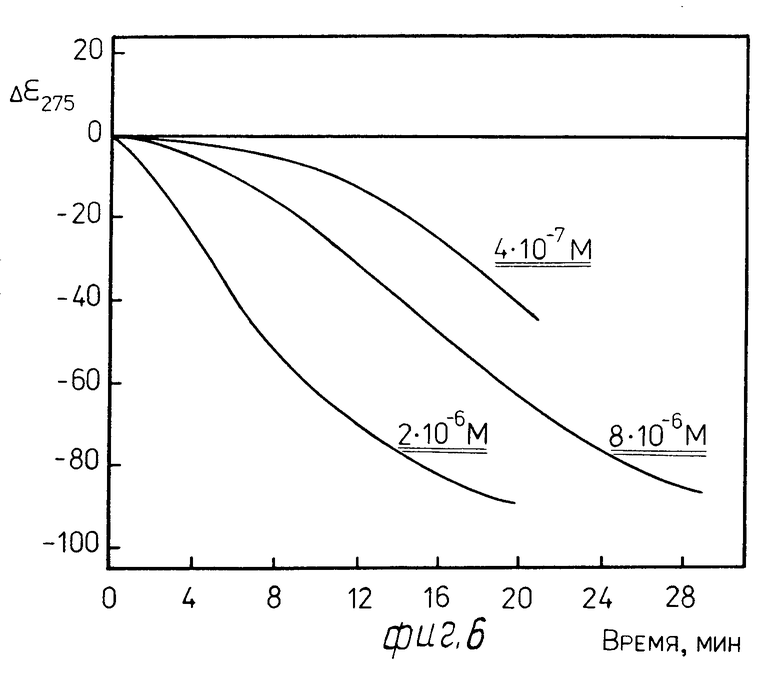
П р и м е р 7. Биодатчик, приготовленный по примеру 3 в растворе 1.1, содержащем 0,04 М трис НСl (рН 8,0) и 0,001 М СаCl2, был обработан по примеру 4 протеолитическим ферментом папаином.
Концентрация используемого фермента показана на фиг. 6.
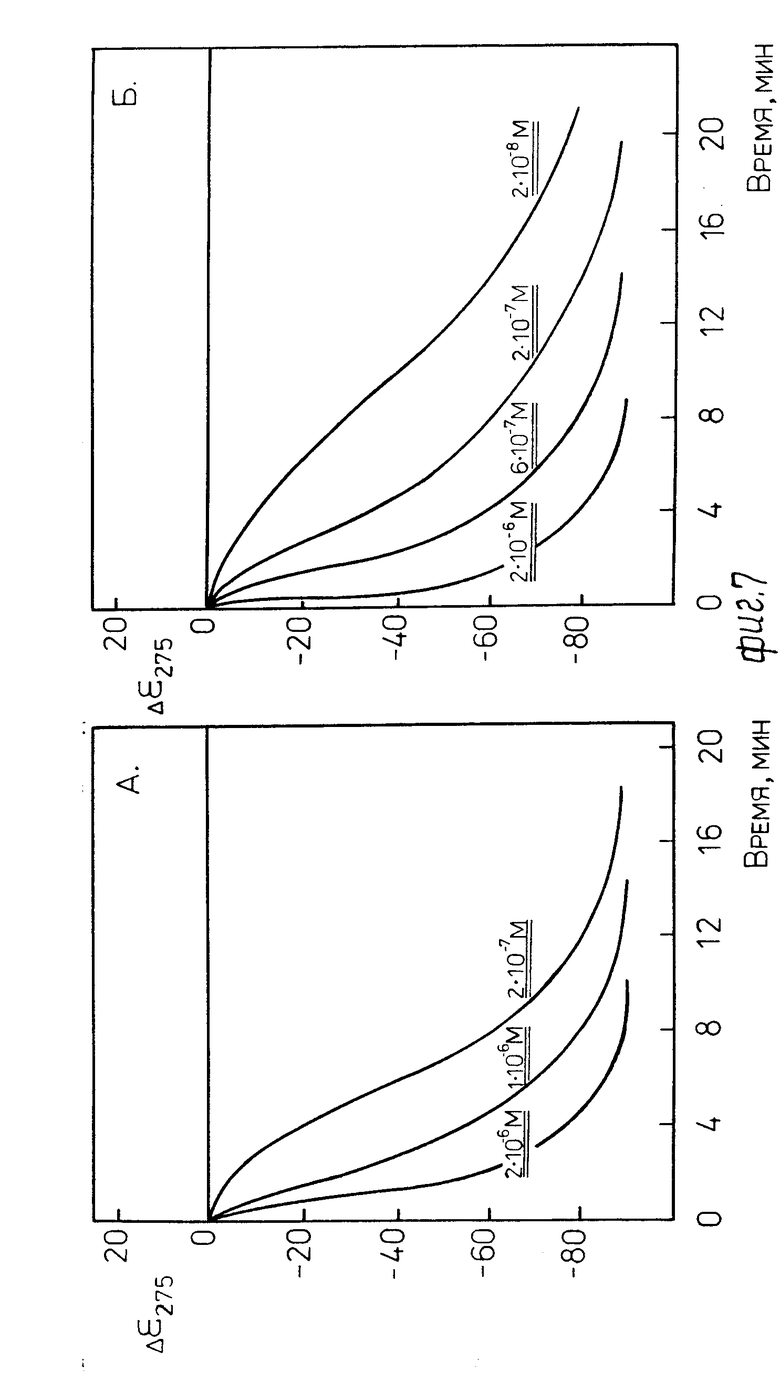
На фиг. 7 показало изменение во времени амплитуды отрицательной полосы при λ 275 нм в спектре КД жидкокристаллической дисперсии комплекса ДНК-стеллин В (r 0,6) после добавления проназы Р (А) и α -химотрипсина (Б). Кривые нарастания аномальной полосы в спектре КД свидетельствуют о том, что по мере расщепления молекул стеллина В ферментом происходит изменение характера упаковки молекул ДНК, составляющих основу биодатчика. Этот результат означает, что биодатчик "срабатывает" при наличии папаина в среде.
П р и м е р 8. Биодатчик, приготовленный по примеру 3 в растворе 1.1, содержащем 0,04 М трис НСl (рН 8,0) и 0,001 М CaCl2, был обработан по примеру 4 протеолитическим ферментом проназой Р.
Концентрация используемого фермента указана на фиг. 7А. Объяснение наблюдаемых кривых аналогично примеру 7.
Биодатчик, приготовленный по примеру 3 в растворе 1.1, содержащем 0,04 М трис НСl (рН 8,0) и 0,001 М СаСl2, был обработан по примеру 4 протеолитическим ферментом α -химотрипсином.
Концентрация используемого фермента указана на фиг. 7Б. Объяснение наблюдаемых кривых аналогично примеру 7.
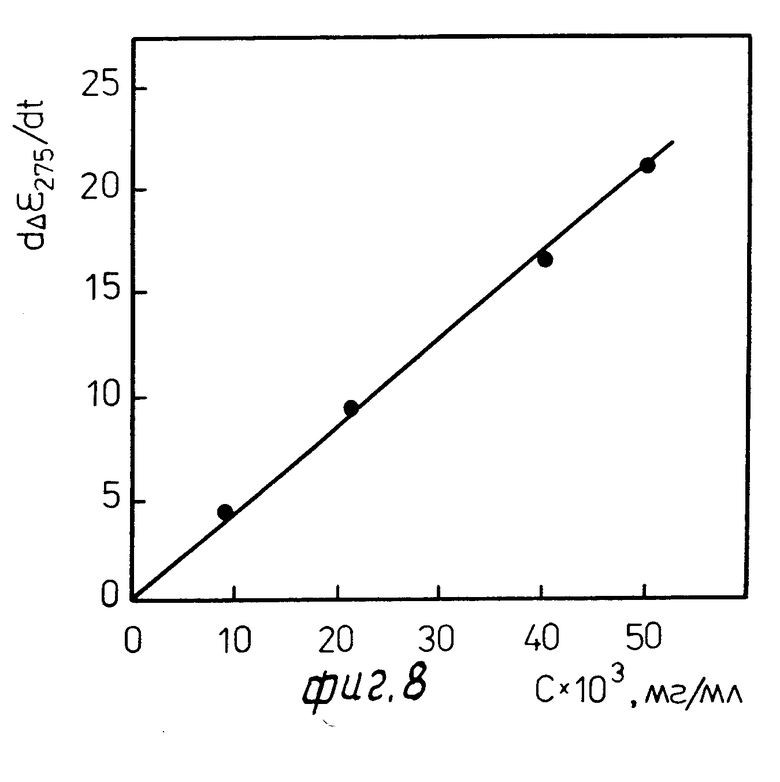
Фиг. 6 и 7 показывают, что присутствие протеолитических ферментов в среде быстро и просто определяется по "срабатыванию" биодатчика, т.е. по факту появления аномальной полосы в спектре КД. Более того, как показывает фиг. 8, приведенная в качестве примера для проназы Р, имеются простые координаты, пользуясь которыми можно определять не только факт наличия протеолитических ферментов, но и их концентрацию в анализируемой среде.
Изобретение относится к средствам для определения биологически активных веществ и может найти применение в области медицинской техники, а также молекулярной фармакологии. Предложен датчик, представляющий собой частицы дисперсной лиотропной жидкокристаллической фазы, сформированной из молекул комплекса ДНК - субстрат определяемого БАВ, распределенные в водно-солевом растворе полимера. 8 ил.
ЖИДКОКРИСТАЛЛИЧЕСКИЙ БИОДАТЧИК ДЛЯ ОПРЕДЕЛЕНИЯ БИОЛОГИЧЕСКИ АКТИВНЫХ ВЕЩЕСТВ (БАВ), предоставляющий собой дисперсную фазу из линейных двухцепочных молекул ДНК, сшитых молекулами субстрата для определяемого БАВ, распределенную в водно-полимерном матриксе.
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| livolant F | |||
| Eur | |||
| j.Cell Biol., 1984, vol.33, no.2, p.300-311. | |||
