Изобретение относится к биотехнологии, в частности к способам биосинтеза рибонуклеотидов из циклических фрагментов с помощью микроорганизмов.
Известны способы биосинтеза рибонуклеотидов из экзогенных пуринов и пиримидинов, при которых азотистые основания, по отдельности или в комбинации, вносят в позднюю 48- или 72-часовую периодическую культуру микроорганизма, растущую на ферментационной среде, и инкубируют ее для накопления в среде рибонуклеотидов в течение 48-72 ч при температуре 30oC [1]
При осуществлении известных способов биосинтеза в нативной микробной культуре концентрации накапливающихся в среде рибонуклеодитов достигают всего 5,0-6,0 мМ в силу того, что метаболические процессы, протекающие в нормально функционирующей микробной клетке, конкурентны за общие промежуточные субстраты и кофакторы тем реакциям, которые участвуют в конденсации экзогенных пуринов и пиримидинов. По этой причине не удается в полной мере раскрыть пурин- и пиримидинконденсирующий потенциал используемых продуцентов.
Известен способ микробиологического синтеза никотинамидадениннуклеотида, НАД+, состоящий в конденсации с помощью пермеабилизованных ксилолом клеток Corynebacterium ammoniagenes ЛИА 0950 экзогенного аденина и никотинамида на структуре эндогенно образующегося из глюкозы рибоза-5'-фосфата, включающий проведение процедур по выращиваню клеточной массы и проведение катализируемых ею реакций раздельно [2] [3]
Известен штамм Corynebacterium species ВСТИ-301 продуцент нуклеотидов (АТФ, АДФ, АМФ, ГТФ) [4] ближайший аналог.
Для выращивания биомассы Corynebacterim species ВСТИ-301 применяют известную ферментационную среду, содержащую, в мас.
глюкоза 10, сульфат магния селиеводный 1, хлорид кальция 0,01, одно- и двузамещенный фосфат калия по 1, пептон 0,5, биотин 30 мкг/л.
Для выращивания инокулята применяют известную посевную среду, содержащую, в мас.
пептон 1,5, глюкоза 2, однозамещенный фосфат калия 0,1, сульфат магния семиводный 0,03, хлорид натрия 0,3, сульфат железа семиводный - 0,01, биотин 30 мкг/л.
Продолжительность процедуры выращивания инокулята при температуре 30oC составляет 24 ч, выращивания биомассы 72 ч.
Ферментацию проводят на круговой качалке в течение трех суток и вносят в колбы по 30 мг аденина и 0,15 сл ксилола. Затем ферментацию продолжают еще сутки.
Выход АТФ, АДФ, АМФ составляет соответственно 1,5, 055 и 1,5 мг/мл, ГТФ
0,42 мг/мл.
Технический результат, достигаемый предложенным способом, заключается в увеличении выхода рибонуклеотидов.
Для этого в способе биосинтеза рибонуклеотидов, включающем конденсацию с помощью Corynebacterium ammoniagenes ЛИА 0950 азотистых оснований на структуре рибозо-5'-фосфата, образуемого клетками из гексоз, и катализ этих процессов с помощью пермеабилизованных клеток, конденсацию азотистых оснований и других фрагментов рибонуклеозидмоно-, ди- и трифосфатов и их производных проводят на структуре экзогенного рибозо-5'-фосфата, осуществляя процесс инкубированием суспензией пермеабилизованных ксилолом или другими апротонными липидоэкстрагентами клеток Corynebacterium ammoniagenes ЛИА 0950 с плотностью (по сухой биомассе) 100-105 мг/мл в реакционной смеси, содержащей, в рибозо-5'-фосфат, в динатриевой форме 1,9 2,0, глюкозу 3,0 3,2, азотистые основания, отдельно или в комбинации, по 0,80 0,84, однозамещенный фосфат калия 0,1 0,2, двузамещенный фосфат калия 1,4 1,5, карбонат магния 0,6 0,7 (pH 7,8), остальное дистиллированная вода, в течение 24 30 ч при температуре 34oC и постоянном перемешивании, до момента полного потребления клетками глюкозы. При этом получение задаваемого в реакционную смесь рибозо-5'-фосфата осуществляют инкубированием суспензии пермеабилизованных ксилолом или другими апротонными экстрагентами липидов клеток Corynebacterium ammoniagenes ЛИА 0950 с плотностью (по сухой биомассе) 130 136 мг/мл в глюкозо-солевой среде, содержащей, в глюкозу 20,0 21,0, однозамещенный фосфат калия 0,4 0,5, двузамещенный фосфат калия 2,44 2,6, карбонат магния 0,8 0,9 (pH 7,8), остальное дистиллированная вода, в течение 24 30 ч при температуре 34oC и постоянном перемешивании, до накопления в среде рибозо-5'-фосфата в количестве 25,0 мг/мл, с последующим выделением его в виде препарата, содержащего основное вещество в динатриевой форме в количестве 87,0 93,0% При этом суспендируемую в реакционной смеси и в глюкозо-солевой среде пермеабилизованную клеточную массу Corynebacterium ammoniagenes ЛИА 0950 получают выращиванием инокулята клеток в количестве 10,0 об. на стерильной ростовой среде, содержащей, в глюкозу, патоку или другой глюкозосодержащий питательный ингредиент, эквивалентный по свободной глюкозе 5,6 5,9, дрожжевой экстракт 0,5 0,6, мочевину 0,4 0,5, соли одно- и двузамещенного фосфата калия, по 0,3 0,4, карбонат магния 0,20 0,3, остальное дистиллированная вода, в течение 24 30 ч при температуре 30oC и аэрации, например в ферментере, до достижения плотности клеточной популяции (по сухой биомассе) 12,0 13,0 мг/мл, с последующей пермеабилизацией клеточной массы ксилолом или другими апротонными липидоэкстрагентами и получением ее в виде концентрата с массовой долей влаги 87,0 90,0% путем механического отделения от среды осаждением или фильтрацией.
По результатам специально проведенного исследования процессов, катализируемых пермеабилизованными ксилолом клетками Corynebacterium ammoniagenes ЛИА 0950, суспендированными в глюкозо-солевой среде, неожиданно обнаружено, что протекающие в клетках (в одной и той же клеточной биомассе) процессы эндогенный биогенез рибозо-5'-фосфата и экзогенный биосинтез рибонуклеотидов имеют разную физиологическую направленность (катаболизм глюкозы, помимо биогенеза рибозо-5'-фосфата, служит одновременно процессом распада гексоз до конечных продуктов) и разномасштабность скоростей протекания реакций, и по этой причине несовместимы для проведения в одной и той же клеточной биомассы. Скорость катаболизации глюкозы до конечных продуктов, CO2 и лактата, достигающая у клеток 0,28 мМ•мг-1•ч-1, превышает скорость конденсации фрагментов, например НАД+ 0,02 мМ•мг-1•ч-1 почти на целый порядок. Установлено, что экогенный задаваемый во внеклеточную среду - рибозо-5'-фосфат в динатриевой форме может проникать во внутрь пермеабилизованной ксилолом и другими апротонными липидоэкстрагентами клетки и вовлекаться тем самым в реакции конденсации рибонуклеотидных фрагментов при подпитке клетки небольшим количеством глюкозы, фруктозы и других усвояемых гексоз в качестве источника энергии; а также то, что такие клетки, суспендированные в глюкозо-солевой среде концентрированного состава, но не содержащей азотистых оснований, способны превращать глюкозу со значительной эффективностью в рибозо-5'-фосфат и накапливать его во внеклеточной среде.
Эти факты послужили основой для разделения катализируемых клетками реакций превращения гексоз в рибозо-5'-фосфат и конденсации на его структуре рибонуклеотидных фрагментов на две осуществляемые раздельно технологические процедуры, составляющие, вместе с тем, одну общую технологическую схему биосинтеза рибонуклеотидов из экзогенных фрагментов фиг. 1.
Технологические процессы, объединенные в блок 2 реакции трансформации гексоз в рибозо-5'-фосфат, и в блок 3 конденсации экзогенных фрагментов рибонуклеотидов (фиг. 1) катализируются раздельно с помощью одинаковой (полученной одним и тем же способом) пермеабилизованной клеточной биомассы (ПКБ); гибкая технология производства рибонуклеотидов (блок 3) основана на применении экзогекнного рибозо-5'-фосфата (Р5Ф) однотипного сахарофосфатного фрагмента, на структуре которого по выбору в зависимости от потребностей, осуществляется конденсация азотистых и других фрагментов (АФ); для получения клеточной биомассы в любом масштабе используется унифицированная по составу ростовая среда.
Пример 1. Получение каталитического биоагента (биомассы пермеабилизованных клеток Corynebacterium ammoniagenes ЛИА 0950).
Свежевыращенный на ростовой среде инокулят в количестве 300 мл, содержащий 12,0 мг клеток/мл (по сухой биомассе), вносят асептически в ферментер с рабочим объемом 5 л, заполненный 3 л стерильной ростовой среды, приготовленной на дистиллированной воде и содержащей, в патоку, в расчете на свободную глюкозу 5,6; дрожжевой экстракт 0,5, мочевину (вносят 30%-ный стерильный раствор дробно) 0,4, соли одно- и двузамещенного фосфата калия, по 0,3, карбонат магния 0,2 (pH 7,4), устанавливают аэрацию с помощью мешалки с числом оборотов 900 об/мин, температуру среды 30oC и выращивают 24 30 ч, контролируя плотность клеточной популяции нефелометрически при λ = 630 нм.
По достижении плотности популяции 12,0 мг/мл по сухой биомассе (по калибровочной кривой), в культуру вносят 30 мл ксилола (смесь о-, м- и п-изомеров) и инкубируют ее для экстракции липидов из клеточной мембраны (пермеабилизации клеток) в течение 0,5 ч при перемешивании с помощью мешалки с числом оборотов 200 об/мин. Пермеабилизованную таким образом клеточную биомассу отделяют от среды центрифугированием при числе оборотов ротора 5000 об/мин. Получают биомассу, содержащую 31 г сухих клеток.
Пример 2. Получение рибозо-5'-фосфата.
Пермеабилизованную ксилолом клеточную биомассу в количестве 26,0 г ( по сухому весу) суспендируют в 200 мл глюкозо-солевой среды в колбе 500 мл, содержащей, в дистиллированной воде, в глюкозу 20,0; однозамещенный фосфат калия 0,4; двузамещенный фосфат калия 2,4; карбонат магния 0,8 (pH 7,8). Полученную суспензию инкубируют на круговой качалке при температуре 34oC в течение 24 30 ч, контролируя накопление в среде рибозосодержащих веществ (колориметрически, с помощью орцинового реактива) в количестве 25,0 мг/мл.
Выделение рибозо-5'-фосфата из среды проводят с помощью ионообменной хроматографии на смоле AB-17. Полученный в количестве 2,0 г препарат рибозо-5'-фосфата содержит основное вещество в динатриевой форме в количестве 87,0%
Пример 3. Биосинтез аденозинмоно-, ди- и трифосфатов (пуриновых рибонуклеотидов).
Пермеабилизованную ксилолом клеточную биомассу в количестве 1,0 г (по сухому весу) суспендируют в 10 мл реакционной смеси в колбе на 100 мл, содержащей, в дистиллированной воде, в рибозо-5'-фосфат (87,0%-ный препарат, в динатриевой форме, по основному веществу) 1,9; глюкозу 3,0; аденин 0,8; однозамещенный фосфат калия 0,1; двузамещенный фосфат калия - 1,4; карбонат магния 0,6 (pH 7,8). Полученную суспензию инкубируют на круговой качалке при температуре 34oC в течение 24 30 ч, контролируя на приборе "Эксан-Г" момент полного потребления клетками глюкозы.
Содержание накопившихся в среде адениннуклеотидов составляет: АТФ 14,5 мг/мл, АДФ 5,6 мг/мл, АМФ 2,1 мг/мл.
Пример 4. Биосинтез уридинмонофосфата (пиримидиновых рибонуклеотидов).
Процесс биосинтеза и способ его проведения осуществляют таким же образом, как в примере 3, только реакционная смесь вместо аденина содержала эквимолярное количество урацила.
Содержание накопившегося в среде УМФ составляет 20,2 мг/мл.
Пример 5. Биосинтез 6-тиогуанинрибонуклеотида (аналогов пуриннуклеотидов).
Процесс биосинтеза и способ его проведения осуществляют таким же образом, как в примере 3, только реакционная смесь вместо аденина содержала эквимолярное количество 6-тиогуанина.
Содержание накопившегося в среде 6-тио ГМФ составляет 12,3 мг/мл.
Пример 6. Биосинтез 5-фторурацилрибонуклеотида (аналогов пиримидиннуклеотидов).
Процесс биосинтеза и способ его проведения осуществляют таким же образом, как в примере 3, только реакционная смесь вместо аденина содержала эквимолярное количество 5-фторурацила.
Содержание накопившегося в среде 5-фтор УМФ составляет 14,7 мг/мл.
Пример 7. Биосинтез никотинамидадениндинуклеотида (адениннуклеотидных коферментов).
Процесс биосинтеза и способ его проведения осуществляют таким же образом, как в примере 3, только реакционная смесь в комбинации с аденином содержала эквимолярное количество никотинамида.
Содержание накопившегося в среде кофермента НАД+ составляет 19,6 мг/мл.
Пример 8. Пермеабилизация клеточной биомассы.
Биосинтез НАД+ и способ его проведения осуществляют таким же образом, как в примере 7, только суспензия клеток вместо пермеабилизованной ксилолом и другими апротонными экстрагентами липидов биомассы содержала пермеабилизованную протонными экстрагентами липидов, например диметилсульфоксидом, биомассу.
Содержание накопившегося в среде НАД+ составляет следовые количества.
Установленные в ходе практических испытаний значения экономического коэффициента роста клеточной биомассы Y г/г (количество биомассы, в граммах, отнесенное к количеству потребленной глюкозы, в граммах) и НАД+ -синтетической активности пермеабилизованной ксилолом биомассы (определенной так, как в примере 7) у популяций клеток Corynebacterium ammoniagenes ЛИА 0950, выращенных на ферментационной и заявляемой ростовой средах в течение 24, 48, 72 и 96 ч, сведены в прилагаемую таблицу.
Как видно из приведенных результатов, в примерах конкретного исполнения использована оптимальная для выращивания клеточной биомассы и катализируемых ею реакций питательная среда и оптимальный возраст популяций выросших клеток.
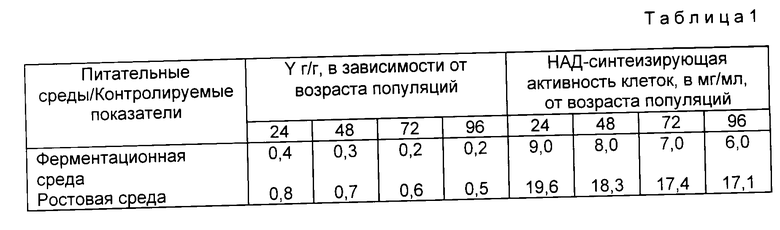
На фиг. 2 приведена в ростовой среде ингредиентов (концентрация ингредиентов в ростовой среде, использованной в примере 1, принята за 100%).
Из него видно, что плотность популяций (в мг сухой биомассы в мл среды), выросших за 24 ч на средах с низкой концентрацией ингредиентов ниже, чем в контроле; на средах с более высокой концентрацией ингредиентов кривая ее зависимости переходит на плато. Во избежание лишнего перерасхода ингредиентов для выращивания биомассы выбрана ростовая среда, содержащая 100 105% концентрации ингредиентов.
Для пермеабилизации биомассы испытано более 15 протонных (с нулевым дипольным моментом) веществ липидоэкстрагентов. Наиболее эффективными для экскреции из клеток рибозо-5'-фосфата и рибонуклеотидов оказались апротонные липидоэкстрагенты, такие как бензол, толуол, о-, м- и п-изомеры ксилола, из поверхностно-активных веществ цетилипиридиний хлорид, обеспечивающие минимальное рассеивание протонного 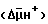 потенциала клеточной мембраны, необходимого для энергообеспечения акта экскреции (выбрасывания из клеток наружу) данных фосфороорганических веществ.
потенциала клеточной мембраны, необходимого для энергообеспечения акта экскреции (выбрасывания из клеток наружу) данных фосфороорганических веществ.
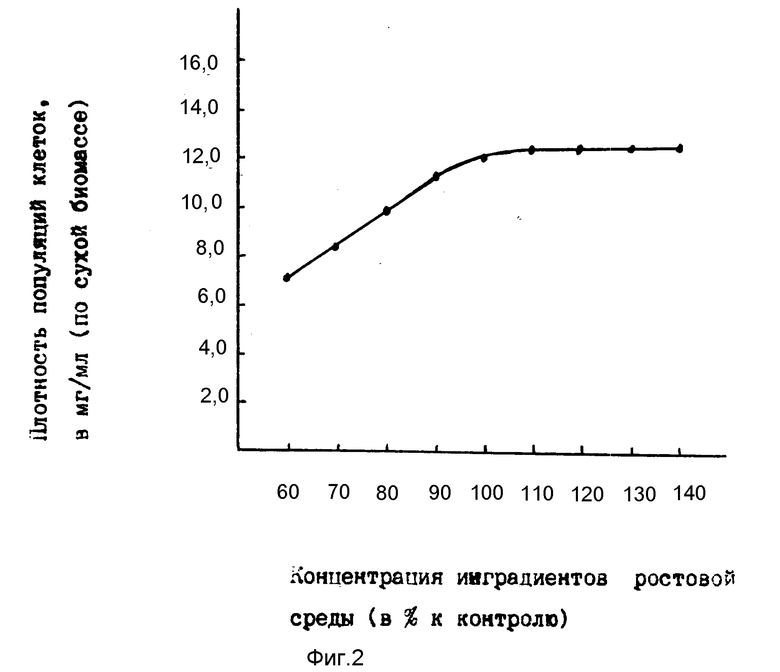
На фиг. 3 приведена зависимость накопления в среде рибозо-5'-фосфата от концентрации компонентов суспензий в глюкозо-солевой среде (концентрация компонентов суспензии, использованной в примере 2, принята за 100%).
Как видно из данных результатов, за равный промежуток времени (24 ч) инкубации у суспензий, имеющих меньшую, чем в контроле концентрацию компонентов (плотность биомассы, концентрацию глюкозы и других ингредиентов), содержание накапливающегося в среде рибозо-5'-фосфата ниже чем в контроле; у суспензий, имеющих более высокие, чем в контроле, значения концентрации компонентов, кривая накопления рибозо-5'-фосфата выходит на плато.
Во избежании перерасхода компонентов суспензии выбраны такие значения концентраций, которые составляют 100 105% к контролю.
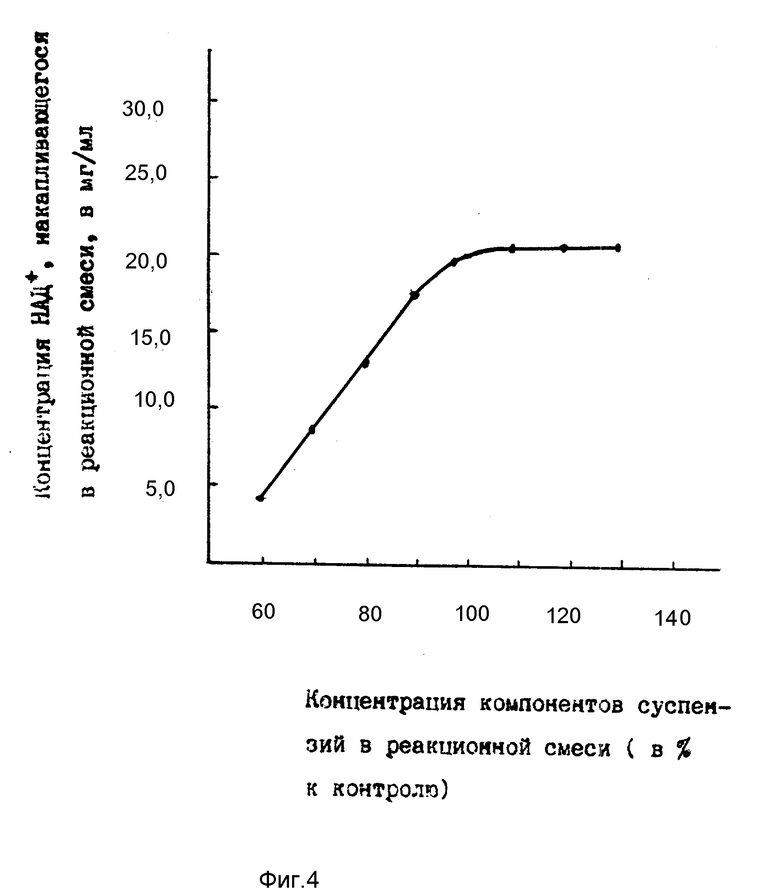
На фиг. 4 показана зависимость накопления в среде НАД+ от концентрации компонентов суспензий в реакционной смеси (компоненты суспензий, использованных в примерах 3 8, приняты за 100%).
Как видно на фиг. 4 у суспензий, имеющих меньшую, чем в контроле, концентрацию компонентов, за 24 ч содержание накопившегося в среде НАД+ ниже, чем в контроле; у суспензий, имеющих более высокие, чем в контроле, концентрации компонентов, кривая накопления НАД+ в среде также переходит на плато.
Из соображений эффективного использования компонентов суспензий выбраны такие пределы их концентраций, которые составляют 100 105% от контроля.
Таким образом, заявляемое изобретение в совокупности позволяет раскрыть в полной мере гексозотрансформирующий и на его основе нуклеотидсинтетический потенциалы Corynebacterium ammoniaqenes ЛИА 0950 (ВСТИ 404), получить в препаративном количестве рибозо-5'-фосфат и провести на его основе препаративный синтез практически всей номенклатуры пурин- и пиримидинрибонуклеозид-моно-, ди и трифосфатов и их производных природного и синтетического происхождения. Изобретение позволяет накопить в реакционной смеси рибонуклеотиды в количестве 28 30 мМ.
Источники информации:
1. Цыренов В.Ж. Микробиологический синтез нуклеозидфосфатов. М. Наука, 1990, с. 200.
2. Гончиков Г. Г. Ленхобоев В.Д. Абросимова Е.В. Бархутова Д.Д. Характеристика НАД синтезирующей активности продуцента Corynebacterium ammoniagenes ВСТИ 404. Тезисы докладов конференций "Проблемы микробного синтеза витаминов и их производных" Ташкент, 1990, с. 12.
3. US, патент, N 3705080, кл. C 12 D 13 /06, 1968.
4. SU, авторское свидетельство, N 726161, кл. C 12 P 19/30, 1980.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ КСАНТОЗИН-5'-МОНОФОСФАТА, ШТАММ CORYNEBACTERIUM AMMONIAGENES - ПРОДУЦЕНТ КСАНТОЗИН-5'-МОНОФОСФАТА (ВАРИАНТЫ) | 2000 |
|
RU2209249C2 |
| ШТАММЫ БАКТЕРИЙ Bacillus subtilis И Bacillus amyloliquefaciens-ПРОДУЦЕНТЫ ИНОЗИНА И СПОСОБ ПРОДУКЦИИ ИНОЗИНА С ИХ ИСПОЛЬЗОВАНИЕМ | 2005 |
|
RU2333949C2 |
| ШТАММ БАКТЕРИЙ CORYNEBACTERIUM AMMONIAGENES - ПРОДУЦЕНТ УРИДИН-5'-МОНОФОСФАТА (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ УРИДИН-5'- МОНОФОСФАТА | 1999 |
|
RU2203948C2 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ БАКТЕРИЙ SERRATIA MARCESCENS, ПРОДУЦЕНТА ХИТИНАЗЫ | 2003 |
|
RU2241744C2 |
| Универсальная питательная среда плотная для выращивания биомассы бруцелл | 2020 |
|
RU2748808C1 |
| Способ получения @ -инозиновой кислоты | 1984 |
|
SU1264579A1 |
| ШТАММ CORYNEBACTERIUM AMMONIAGENES - ПРОДУЦЕНТ 5'-КСАНТИЛОВОЙ КИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ 5'-КСАНТИЛОВОЙ КИСЛОТЫ | 2003 |
|
RU2312137C2 |
| СПОСОБ ПОЛУЧЕНИЯ СУММЫ ЭКСТРАКТИВНЫХ ВЕЩЕСТВ, ОБЛАДАЮЩИХ АДАПТОГЕННОЙ АКТИВНОСТЬЮ | 1993 |
|
RU2076727C1 |
| Штамм аRтнRовастеR SpecIeS ВСТИ-5-продуцент нуклеотидов и рибозо-5-фосфата | 1981 |
|
SU960258A1 |
| ШТАММ БАКТЕРИЙ AGROBACTERIUM TUMEFACIENS, ОСУЩЕСТВЛЯЮЩИЙ ДЕГРАДАЦИЮ ФЕНОЛА | 1993 |
|
RU2077575C1 |
Инокулят штамма Corynebacterium ammoniagenes ЛИА 0950 выращивают в стерильной ростовой среде, далее биомассу обрабатывают апротонным липидоэкстрагентом и отделяют пермеабилизованную биомассу, часть пермеабилизованной биомассы суспендируют и выращивают в глюкозо-солевой среде при постоянном перемешивании до накопления в среде рибозо-5'-фосфата в динатриевой форме, далее полученный продукт вносят в содержащую азотистое основание питательную среду для выращивания другой части пермеабилизованной биомассы, выделяют целевой продукт. 1 з.п. ф-лы, 4 ил., 1 табл.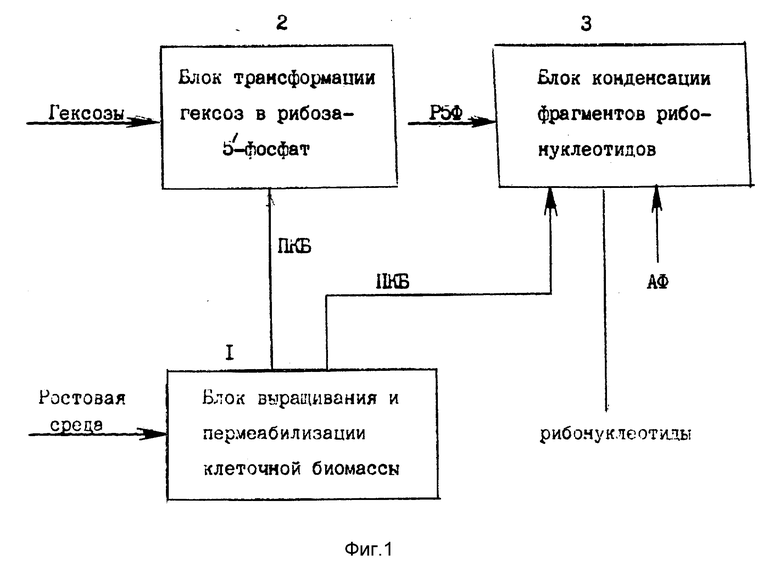
Глюкоза или патока в количестве, эквивалентном по свободной глюкозе 5,6 5,9
Дрожжевой экстракт 0,5 0,6
Мочевина 0,4 0,5
Соли одно- и двузамещенного фосфата калия По 0,3 0,4
Карбонат магния 0,2 0,3
Дистиллированная вода Остальное
и культивирование ведут до достижения плотности клеточной популяции по сухой биомассе 12,0 13,0 мг/мл, далее биомассу обрабатывают апротонным липидоэкстрагентом и отделяют пермеабилизованную биомассу с массовой долей влаги 87,0 90,0% затем часть пермеабилизованной биомассы с плотностью по сухой биомассе 130 136 мг/мл суспендируют и выращивают в глюкозосолевой среде, содержащей следующие компоненты, мас.
Глюкоза 20,0 21,0
Однозамещенный фосфат калия 0,4 0,5
Двузамещенный фосфат калия 2,4 2,6
Карбонат магния 0,8 0,9 (pH 7,8)
Дистиллированная вода Остальное
при температуре 34oС и постоянном перемешивании до накопления в среде рибозо-5'-фосфата в динатриевой форме в количестве 25,0 мг/мл среды, далее полученный продукт вносят в питательную среду для выращивания другой части пермеабилизованной биомассы, осуществляемого в тех же условиях на питательной среде следующего состава, мас.
Рибозо-5'-фосфат в динатриевой форме 1,9 2,0
Глюкоза 3,0 3,2
Азотистое основание, отдельно или в комбинации По 0,80 0,84
Однозамещенный фосфат калия 0,1 0,2
Двузамещенный фосфат калия 1,4 1,5
Карбонат магния 0,6 0,7
до полного потребления глюкозы, затем выделяют целевой продукт.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Гончиков Г.Г., Ленхобоев В.Д., Абросимова Е.В., Бархутова Д.Д | |||
| Катодный усилитель с промежуточными контурами и батарейным коммутатором для цепей сетки | 1923 |
|
SU404A1 |
| Тезисы докладов конференций "Проблемы микробного синтеза витаминов и их производных" | |||
| - Ташкент, 1990, с.12 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| SU, авторское свидетельство, N 726161, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
