Изобретение относится к способу определения наличия или отсутствия одной или нескольких последовательностей вариантных нуклеотидов посредством экстракопирования или его отсутствия и к наборам для определения.
Изобретение имеет особое значение при диагностическом скрининге образцов ДНК для наследованных состояний, предрасположений или соматических мутаций и обеспечивает общий способ легкого определения точечных мутаций. Кроме того, оно пригодно для определения и типизации инфекционных патогенов посредством анализа их ДНК или РНК.
Известно, что у людей существуют несколько сотен генетических заболеваний, которые являются результатом определенных мутаций на уровне ДНК. Молекулярное основание для некоторых из этих болезней уже известно, причем исследование быстро выявляет молекулярное основание для тех генетических заболеваний, для которых в настоящее время неизвестна природа мутации. Когда точное молекулярное основание для наследованного состояния не известно, может быть предоставлен диагноз нарушения или расположения носителей в информативных родословных посредством RFLR (ограничивающий фрагмент протяжения полиморфизма) с использованием зондов ДНК в генетической связи с областью заболевания. Таким образом, в настоящее время мышечная дистрофия Дюшенна, цистный фиброз и хорея Хантингтона могут диагностироваться в качестве примера использования технологии RFLR. Однако такое тестирование необходимо осуществлять отдельно в отношении каждого состояния и требуется значительное количество работы, причем для каждого случая требуется очистка ДНК, расщепление ограничивающего фермента, электрофорез агарозного геля, окрашивание по Соутерну, гибридизация, определение пробы гибридизованного гена и анализ родословной. Некоторые другие наследованные состояния связаны с одноточечными мутациями в генах, однако каждое из этих состояний должно быть проанализировано отдельно, причем возникают дополнительные специфические затруднения, когда точечные мутации являются гетерогенными. Так, например, более чем 40 различных точечных мутаций могут вызвать бета-талласаэмию и по меньшей мере 5, а вероятно больше чем 12 точечных мутаций могут вызвать гемофилию А. В отношении этих гетерогенных состояний каждая потенциальная точка мутации в настоящее время, возможно, нуждается в отдельном анализе. Это может включать сложный RFLR гаплотипный анализ с множеством ограничивающих ферментов.
Ряд точечных мутаций в соматических клетках был вовлечен в развитие различных видов рака, например точечные мутации в ras онкогена (Дж. Л. Боос и сотр. Nature, т.327, с.293,1987).
В заявке на Европатент N 87302196.8 (публикация N 237362) фирмы Цетус Корп. описан способ определения наличия или отсутствия по меньшей мере одной нуклеофильной вариации в последовательности из одной или нескольких нуклеиновых кислот, которые содержатся в образце, который включает:
а) обработку образца вместе или последовательно с четырьмя различными нуклеотидными трифосфатами, реагентом для полимеризации нуклеотидных трифосфатов и одного олигонуклеотидного праймера для каждого витка каждой нуклеиновой кислоты, в которой предполагается содержание указанной вариации при условиях гибридизации, такой, что для каждого витка нуклеиновой кислоты, содержащего различные вариации, подлежащие определению, синтезируется продукт распространения каждого праймера, который является комплементарным к каждому витку нуклеиновой кислоты, в котором указанный праймер или праймеры выбираются таким образом, чтобы быть существенно комплементарными к каждому витку нуклеиновой кислоты, содержащему любую вариацию, так что продукт распространения, синтезированный из одного праймера, когда он отделятся от своего комплемента (хромосомного набора), может служить в качестве матрицы для синтеза продукта распространения другого праймера;
б) обработку образца в условиях денатурации, чтобы отделить продукты распространения праймера от их матрицы, если присутствуют вариации, подлежащие определению;
в) обработку образца вместе или последовательно указанными четырьмя трифосфатами нуклеотида, реагентом для полимеризации трифосфатов нуклеотида и праймерами олигонуклеотидов такими, чтобы синтезировался продукт распространения праймера с использованием каждого отдельного витка, полученного на стадии б) в качестве матрицы, при которой стадии б) и в) повторяются достаточное число раз, чтобы привести к обнаруживаемому экстракопированию нуклеиновой кислоты, содержащей последовательность вариаций, если они имеются;
г) фиксирование продукта стадии в) на мембране;
д) обработку мембраны в условиях гибридизации меченой последовательностью специфическим олигонуклеотидным зондом, способным к гибридизации только с экстракопированной последовательностью, является комплементарной по отношению к области экстракопированной последовательности; и
е) определение, если зонд гибридизовался на экстракопированной последовательности в образце нуклеиновой кислоты.
Определение наличия или отсутствия по меньшей мере одной нуклеотидной вариации в определенных специальных ситуациях может быть достигнуто с помощью различных методик. Так, в необычных случаях, если точечная мутация создает или разрушает ограничивающий сайт (например серповидная клеточная анемия), усвоение ограничивающего фермента может применять или до, или после экстракопирования (Ф. Ф. Чехаб и сотр. Nature, т.329, с.293, (1987)). Более того, в отношении больших исключенных последовательностей нуклеиновой кислоты праймеры для экстракопирования могут быть приготовлены для областей внутри предполагаемого исключения, такого как 23 кIо исключение, вызывающее альфа-талассаэмию; в таких случаях неуспех с экстракопированием исключенной последовательности подтверждает исключение и таким образом, например, является диагнозом альфа-талассаэмии (Ф.Ф.Чехаб и сотр. Nature, т.329, с.293, (1987)).
Способ экстракопирования по публикации Европатента N 237362 предоставляет некоторые преимущества перед методиками RFLР (ограничивающий фрагмент протяжения полиморфизма) и аллельного специфического олигонуклеотида, которая описана, например, Каном и Дози, Труды Национальной Академии наук США (Proc.Nat.Acad.Sci.) т. 75, с.5631, 1978 г. Рубином и Каном, Lancet, 1985-1, c. 75, 1985 г. Коннером и сотр. Труды национальной Академии наук США, т. 80, с.78, 1983 г. Киддом и сотр. Nature, т.304, с.230, 1983 г. и Пиратсу и сотр. New England J.Medicine (Журнал медицины Новой Англии), т.309, с.284, 1983 г.
Тем не менее в публикации Европатента N 237362 описан способ, который включает недискриминативное экстракопирование представляющей интерес определенной последовательности, что неизбежно приводит к необходимости ряда дополнительных стадий определения, на которые затрачивается время и которые включают или дальнейшее использование меченой последовательности - специфического олигонуклеотидного зонда, который, возможно, должен быть способным различать последовательности, отличающиеся столь незначительно, как на единственный нуклеотид и/или использование специфической ограничивающей эндонуклеазы в тех ограниченных случаях, когда представляющая интерес точечная мутация создает или разрушает опознающую последовательность фермента, и/или использование методов непосредственного последования на экстракопированной ДНК (смотри работу С.Wong и сотр. Nature, т.330, с.384, 1987 г.
Существует потребность в простом способе непосредственного определения по меньшей мере одного основного различия в нуклеиновых кислотах, таких как геномная ДНК, в котором сведены к минимуму стадии определения, что приводит к способу, который может быть осуществлен быстро, точно и легко при минимальной квалификации оператора.
Изобретение основано на открытии того, что путем соответствующего выбора нуклеотидной последовательности, олигонуклеотидного праймера возможно селективное достижение распространения праймера либо последовательности, содержащей предполагаемый вариантный нуклеотид, либо соответствующей последовательности, содержащей нормальный нуклеотид, или предотвращение распространения такого праймера, таким образом существенно упрощаются необходимые методики определения.
В соответствии с одной характерной чертой изобретения, разработан способ определения наличия или отсутствия по меньшей мере одного варианта нуклеотида в одной или нескольких нуклеиновых кислотах, которые содержались в образце.
Этот способ включает обработку образца, вместе или последовательно с соответствующими нуклеозид-трифосфатами, реагентом для полимеризации нуклеозид-трифосфатов и диагностическим праймером для диагностической части основной последовательности мишени в условиях гибридизации, причем нуклеотидная последовательность указанного диагностического праймера является таковой, что она существенно комплементарна к этой диагностической части, терминальный нуклеотид диагностического праймера является либо комплементарным к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду, посредством этого синтезируется продукт распространения диагностического праймера, тогда указанный терминальный нуклеотид диагностического праймера комплементарен по отношению к соответствующему нуклеотиду в основной последовательности мишени, причем никакой продукт распространения не синтезируется, если указанный терминальный нуклеотид диагностического праймера не комплементарен к соответствующему нуклеотиду в основной последовательности мишени, а также детектирование наличия или отсутствия продукта распространения.
Следует признать, что хотя этот способ представляет особый интерес при определении наличия или отсутствия точечных мутаций, способ в равной степени применим к определению наличия или отсутствия исключений, включая исключения более чем одного нуклеотида, а также к определению наличия или отсутствия замещений более чем одного нуклеотида. В этом отношении просто необходимо знать соответствующие нуклеотиды, особенно соответствующий терминальный нуклеотид, с тем чтобы могли быть спланированы надлежащим образом необходимые диагностические праймеры (праймер).
Можно признать, что любой образовавшийся продукт распространения может быть определен в любой удобной форме, например в одно- или двукратно спиральной форме.
Кроме того, можно признать, что любой полученный продукт распространения по желанию может быть экстраполирован с помощью реакции полимеразной цепи (PCR), которая описана в патентах США N 4683195 и 4683202 с использованием Ку-бета-репликазы, которая описана в публикации патента "Пи-Си-Ти" N W087/06270 и 6-м томе Биотехнологии, октябрь 1988, посредством использования экстракопирования нуклеиновой кислоты на основе транскрипции фирмы "Сиска Корпорейшн" как описано в публикации патента "Аи-Си-Ти" N W088/10315, или посредством использования линейного экстракопирования. В этой связи выражение "линейное экстракопирование" (или амплификация) используется здесь для ссылки на амплификацию с использованием единственного праймера для каждой диагностической части в присутствии реагента для полимеризации и соответствующего нуклеотид-трифосфата, посредством чего амплификация осуществляется распространением праймера на основе использования одной спирали образца нуклеиновой кислоты в качестве матрицы.
В первом, и особенно предпочтительном, варианте воплощения настоящего изобретения способ включает:
1) обработку образца вместе или последовательно соответствующими нуклеозид-трифосфатами, реагентом для полимеризации нуклеозид-трифосфатов, диагностическим праймером для диагностической части основной последовательности мишени и соответствующим праймером в условиях гибридизации, причем эта нуклеотидная последовательность указанного диагностического праймера является такой, что она существенно комплементарна к указанной диагностической части, терминальный нуклеотид диагностического праймера является либо комплементарным к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду, в результате чего синтезируется продукт распространения диагностического праймера, когда упомянутый терминальный нуклеотид диагностического праймера комплементарен к соответствующему нуклеотиду в основной последовательности мишени, причем никакой продукт распространения не синтезируется, если терминальный нуклеотид диагностического праймера не является комплементарным к соответствующему нуклеотиду в основной последовательности мишени; причем любой образовавшийся продукт распространения диагностического праймера способен служить в качестве матрицы для синтеза продукта распространения упомянутого амплификационного праймера после отделения от его комплемента;
2) обработку образца в условиях денатурирования, для того чтобы отделить продукт распространения праймера от его матрицы, на которой образовался такой продукт распространения;
3) контактирование полученных на стадии 2 отдельных спиралей либо вместе, либо последовательно с соответствующими нуклеозид- трифосфатами, реагентом для полимеризации нуклеозид-трифосфатов, диагностическим праймером и амплификационным праймером, который определен здесь, посредством чего там, где возможно, синтезируются дополнительные продукты распространения с использованием в качестве матриц отдельных спиралей, полученных на стадии 2.
4) повторение стадий 2 и 3 достаточное число раз, чтобы получить заметную амплификацию соответствующей нуклеотидной последовательности;
5) определение наличия или отсутствия предполагаемого вариантного нуклеотида по наличию или отсутствию продукта амплификации, полученного на стадии 4.
Во втором варианте воплощения изобретения указанный образец обрабатывают вместе или последовательно, либо:
а) первым диагностическим праймером, имеющим последовательность, которая существенно комплементарна к диагностической части первой последовательности нуклеиновой кислоты, причем первый диагностический праймер имеет терминальный нуклеотид, который комплементарен к указанному предполагаемому вариантному нуклеотиду, и вторым диагностическим праймером, имеющим последовательность, существенно комплементарную к диагностической части второй последовательности нуклеиновой кислоты, причем второй диагностический праймер имеет терминальный нуклеотид, который комплементарен к комплементарному предполагаемому варинатному нуклеотиду;
б) первым диагностическим праймером, имеющим последовательность существенно комплементарную к диагностической части первой последовательности нуклеиновой кислоты, причем первый диагностический праймер имеет терминальный нуклеотид, который комплементарен к нормальному нуклеотиду, который соответствует упомянутому предполагаемому вариантному нуклеотиду, и вторым диагностическим праймером, имеющим последовательность существенно комплементарную к диагностической части второй последовательности нуклеиновой кислоты, причем второй диагностический праймер имеет терминальный нуклеотид, комплементарный к нормальному нуклеотиду, который соответствует указанному предполагаемому вариантному нуклеотиду; причем упомянутый терминальный нуклеотид первого диагностического праймера и терминальный нуклеотид второго диагностического праймера оба находятся при конце 5' или оба при конце 3' соответствующих праймеров, и первая последовательность нуклеиновой кислоты находится в обратном отношении ко второй последовательности нуклеиновой кислоты.
Следовательно, в этом варианте воплощения второй диагностический праймер может считаться амплификационным праймером, о чем упоминалось выше и в последующем.
Этот второй вариант воплощения может обеспечивать дискриминацию и специфичность, которые возрастают, так как для любого искусственного продукта требуется, чтобы прайминг (зарождение) происходил на соответствующем терминальном конце (обычно терминальный конец 3'-) двух непарных олигонуклеотидов, а не на единственном конце, как в случае, когда используется только единственный диагностический праймер.
Определение наличия или отсутствия предполагаемого вариантного нуклеотида может быть осуществлено, например, так, как описано ниже.
В третьем варианте воплощения изобретения образец, включающий ДНК, содержащую предполагаемый вариантный нуклеотид, подвергается амплификации, например, посредством линейного экстракопирования, которое определено здесь или, например, как описано в патентах США N 4683195 и 4683202, в публикации патента Пи- Си-Ти W087/06270, в кн. Биотехнология, т. 6, октябрь 1988 или в публикации патента Пи-Си-Ти W088/10315, и продукт амплификации обрабатывается диагностическим праймером для диагностической части последовательности оснований мишени в условиях гибридизации, в присутствии соответствующих нуклеозид-трифосфатов, в присутствии соответствующих нуклеозид-трифосфатов, и агента для полимеризации нуклеозид-трифосфатов, причем нуклеотидная последовательность указанного диагностического праймера такова, что она существенно комплементарна к указанной диагностической части, терминальный нуклеотид диагностического праймера является либо комплементарным к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду.
Таким образом, в третьем воплощении изобретения традиционная амплификация может быть осуществлена в желаемом числе циклов и гибридизация с диагностическим праймером, предпринятая на следующей стадии, до стадии определения. Отпадает необходимость использования амплификационного праймера.
Это третье воплощение представляет интерес, так как количество реагентов, требующихся для полимеризации, может быть существенно снижено (например, по меньшей мере на половину), разумеется это может быть количество используемых нуклеозид-трифосфатов и число применяемых нагревательных устройств для реакции полимеразной цепи (PCR). Таким образом, это третье воплощение обеспечивает существенное уменьшение затрат, которое может быть достигнуто. Кроме того, поскольку стадия амплификации может осуществляться в интервале удобных температур без ухудшения результатов. В этом третьем воплощении требуется только, чтобы более чувствительная к нагреву предпринятая гибридизация с диагностическим зондом осуществлялась в свое время, таким образом еще снижается дополнительный риск неправильного зарождения терминального непарного (обычно 3'- непарного) диагностического праймера. Таким образом, это третье воплощение обеспечивает потенциально более гибкий и ясный способ для использования обычным неэкспертным персоналом, который менее чувствителен к ошибке оператора. Кроме того, в этом третьем варианте исключена потребность в дополнительной контрольной стадии реакции полимеразной цепи, так как начальная амплификация предоставляет собственный внутренний контроль.
По желанию диагностический праймер может нести сигнал или метку, которую не следует подвергать риску деструкции, например, в методике высокотемпературного цикла, такой как реакция полимеразной цепи. Например, помечивание может быть осуществлено с использованием соответствующей меченой или сигнальной функции, такой как щелочная фосфатаза или пероксидаза хрена.
В этой связи может представлять интерес применение термостабильных ферментов для помечивания, таких как фосфатаза, произведенная из Thermus aquaticus.
В четвертом и предпочтительном воплощении изобретения модифицирован его третий вариант путем введения признака использования двух диагностических праймеров, как описано во втором воплощении изобретения, причем второй диагностический праймер потенциально служит как амплификационный праймер.
Таким образом, в четвертом воплощении настоящего изобретения образец, включающий ДНК, содержащую предполагаемый вариантный нуклеотид, подвергается амплификации, и экстракопированный продукт обрабатывается одновременно или последовательно, либо:
а) первым диагностическим праймером, имеющим последовательность существенно комплементарную к диагностической части первой последовательности нуклеиновой кислоты, причем первый диагностический праймер имеет терминальный нуклеотид, комплементарный к указанному предполагаемому вариантному нуклеотиду, и вторым диагностическим таймером, имеющим последовательность существенно комплементарную к диагностической части второй последовательности нуклеиновой кислоты, причем второй диагностический праймер имеет терминальный нуклеотид, комплементарный к нуклеотиду, который является комплементарным к упомянутому вариантному нуклеотиду;
б) первым диагностическим праймером, имеющим последовательность, существенно комплементарную к диагностической части первой последовательности нуклеиновой кислоты, причем первый диагностический праймер имеет терминальный нуклеотид, комплементарный к нормальному нуклеотиду, который соответствует указанному предполагаемому вариантному нуклеотиду, и вторым диагностическим праймером, имеющим последовательность, существенно комплементарную к диагностической части второй последовательности нуклеиновой кислоты, причем этот второй диагностический праймер имеет терминальный нуклеотид, комплементарный к нуклеотиду, который комплементарен к указанному нормальному нуклеотиду, который соответствует предполагаемому вариантному нуклеотиду;
причем указанные терминальный нуклеотид первого диагностического праймера и терминальный нуклеотид второго диагностического праймера оба находятся либо при 5'-конце, либо при 3'-конце соответствующих праймеров, и первая последовательность нуклеиновой кислоты комплементарна к второй последовательности нуклеиновой кислоты.
В общем, указанный терминальный нуклеотид первого диагностического праймера и терминальный нуклеотид второго диагностического праймера каждый находится при 3'-конце их соответствующих праймеров.
Таким образом, в четвертом варианте воплощения изобретения объединены потенциальные преимущества указанных выше второго и третьего воплощений изобретения, которые обладают потенциально повышенной специфичностью, пониженными затратами и более ясной и располагающей потребителя методикой.
Определение наличия или отсутствия предполагаемого вариантного нуклеотида может быть осуществлено, например, так, как указано в последующем.
Можно признать, что амплифицированный продукт, однажды обработанный как указано выше (либо а, либо б), может быть вовлечен в один или несколько финальных циклов по желанию. Когда осуществляются многочисленные циклы, тогда можно получить дополнительное количество продукта, который представляет собой гибрид продуктов распространения диагностических праймеров. Эти различные продукты будут образовываться в соотношениях, зависящих от относительных соотношений исходных PCR (реакция полимеразной цепи) праймерных олигонуклеотидов и добавленных диагностических праймерных олигонуклеотидов.
Когда амплификация осуществляется либо путем применения диагностических и амплификационных праймеров, либо посредством использования двух диагностических праймеров, например, как описано в первом и втором вариантах воплощения изобретения, либо как часть методики амплификации, описанной в публикации Европатента N 237362, то стадии а) денатурирования для отделения продуктов распространения праймера от их матриц и б) контактирования, полученных таким образом отдельных спиралей либо одновременно, либо раздельно с соответствующими нуклеозид-трифосфатами и соответствующими праймерами, чтобы синтезировать дополнительные продукты распространения, предпочтительно повторяются по меньшей мере пять раз (циклов) и до неопределенно большого числа, особенно если праймер является устойчивым к амплификации без ущерба для изобретения. Более предпочтительно, что применяются 15-60, например 15-30, раз (циклов), если образец содержит человеческую геномную ДНК. Если образец включает клетки, то предпочтительно их нагревают до стадии а), чтобы подвергнуть имеющиеся в них нуклеиновые кислоты воздействию реагентов. Эта стадия исключает очистку нуклеиновых кислот до добавления реагентов. В связи с этим можно признать, что изобретение представляет существенное усовершенствование по сравнению с предыдущими способами, даже если очистка ДНК из образца осуществлялась до предпринятой амплификации.
Следует признать, что на стадии б контакт между полученными на стадии а отдельными спиралями и подходящими нуклеозид-трифосфатами, реагентом для полимеризации нуклеозид-трифосфатов, праймером (праймерами), например диагностическим праймером и/или амплификационным праймером, может быть осуществлен или посредством добавления этих материалов в реакционную смесь с последующим отделением продукта распространения праймера от его матрицы (стадия а), либо можно надеяться на материалы, которые уже присутствуют в реакционной смеси. Разумеется, любой один или несколько различных нуклеозид-трифосфатов и/или реагент для полимеризации, и/или праймер (праймеры), например диагностический праймер и/или ампликационный праймер, могут быть добавлены на любой стадии предложенного способа.
В соответствии с пятым и предпочтительным воплощением изобретения предоставляется способ, определенный выше, в котором амплификация осуществляется посредством распространения праймера, основанного на использовании отдельной спирали образца нуклеиновой кислоты в качестве матрицы.
Таким образом, в этом воплощении распространение праймера осуществляется на основе использования той же самой спирали образца нуклеиновой кислоты в качестве матрицы, причем ампликационный праймер отсутствует. Такая амплификация скорее является арифметической, а не экспоненциальной, причем экспоненциальная амплификация может быть достигнута по крайнем мере теоретически с реакцией полимеразной цепи (PCR). Преимущество этого пятого варианта (также называемого здесь линейной амплификацией) состоит в том, что артефактные продукты, если они получаются, сами по себе не могут подвергаться экспоненциальной амплификации.
Линейная амплификация может осуществляться с помощью любого удобного средства и таким образом может осуществляться путем использования комплементарных нуклеозид-трифосфатов в присутствии реагента для полимеризации нуклеозид-трифосфатов, для того чтобы получить продукты распространения праймера неопределенной протяженности, если имеется достаточная степень комплементарности между диагностическим праймером и образцом нуклеиновой кислоты. Предпочтительно, когда все комплементарные нуклеозид-трифосфаты должны использоваться, образец нуклеиновой кислоты подвергается эндонуклеазному расщеплению, причем ограничительная эндонуклеаза выбирается таким образом, чтобы гарантировать то, что расщепление образца нуклеиновой кислоты осуществится при сайте, соответствующем возможности образования продуктов распространения праймера фиксированной длины. Однако преимущественно линейная амплификация может осуществляться в присутствии только одного, преимущественно только двух или предпочтительно только трех нуклеозид-трифосфатов, так что диагностический праймер в своем связанном состоянии (т.е. гибридизован к нуклеиновой кислоте образца) может распространиться только на столько, на сколько позволят только 1,2 или 3 нуклеозид- трифосфат. Когда нуклеозид-трифосфат присутствует в нуклеиновой кислоте образца, для которой отсутствует комплементарный нуклеозид-трифосфат, тогда распространение праймера будет прекращено.
По желанию линейная амплификация может осуществляться при температуре плавления (Тпл) последовательности. При этой температуре диагностический праймер, гибридизованный к комплементарной последовательности в нуклеиновой кислоте образца, находится в равновесии с диагностическим праймером, свободно находящимся в растворе, и таким образом диагностический праймер (необязательно в продленной форме) быстро гибридизуется и денатурируется от нуклеиновой кислоты образца. По желанию линейная амплификация также может осуществляться путем термической осцилляции. Обычно такая термическая осцилляция может включать быструю температурную флуктуацию вблизи температуры плавления последовательности.
Если присутствуют только 1,2 или 3 нуклеозид-трифосфата, тогда диагностический праймер может распространиться только на столько, на сколько позволят эти присутствующие нуклеозид-трифосфаты. Как указано выше, если имеется несоответствие между, например 3'-терминальным концом диагностического праймера и соответствующего нуклеозид-трифосфата в нуклеиновой кислоте образца, то не будет осуществляться распространение праймера. Однако, если 3'- терминальный нуклеозид-трифосфат комплементарен с соответствующим нуклеозид-трифосфатом в нуклеиновой кислоте образца, то распространение праймера будет осуществляться.
Когда используются только 1,2 или 3 нуклеозид-трифосфат и при этом терминальный нуклеозид-трифосфат протяженного диагностического праймера используется только однажды, тогда может быть выгодным использование дидеоксинуклеозид-трифосфата в качестве нуклеозид-трифосфата, который при использовании будет составлять терминальный нуклеозид-трифосфат продукта распространения диагностического праймера. Это будет способствовать при получении четко ограниченного распространения диагностического праймера.
По желанию один или несколько нуклеозид-трифосфатов, присутствующих в реакционной смеси для целей включения в распространненый праймер (праймеры), могут быть помечены или маркированы любым удобным образом. Так, например, один или несколько нуклеозид-трифосфатов могут быть помечены флуоресцентно. Это помечивание нуклеозид-трифосфатов представляет особый интерес в связи с пятым вариантом воплощения изобретения, когда производство продукта распространения диагностического праймера может быть детектировано путем определения меченого или маркированного нуклеозид-трифосфата, включенного в продукт распространения. Если продукт распространения не образуется, то включение не имеет места, и меченый или маркированный нуклеозид-трифосфат может быть, например, удален промывкой. Более конкретно, в пятом варианте изобретения устранена проблема амплификации артефактных продуктов, и таким образом обеспечивается хорошая дискриминация, которая достигается в присутствии меченого или маркированного нуклеозид-трифосфата. Когда амплификация осуществляется, например, путем использования PCR, то любое производство артефактного продукта может привести к амплификации этого продукта и таким образом к включению меченого или маркированного нуклеозид-трифосфата, тем самым снижается дискриминация.
В дополнение к изложенному выше может быть желательным, чтобы диагностический праймер имел одного представителя иммунологической связывающей пары, например антиген или антитело, или одного представителя пары, образующей комплекс, например биотин, для привязывания к другому представителю указанной связывающей пары, или для образования пары с целью фиксации на твердой фазе.
В соответствии с дополнительной характеристикой изобретения в нем представлен набор для определения наличия или отсутствия по меньшей мере одного вариантного нуклеотида в одной или нескольких нуклеиновых кислотах, содержащихся в образце, причем этот набор включает:
1) диагностический праймер для каждой диагностической части основной последовательности мишени, причем нуклеотидная последовательность каждого диагностического праймера является таковой, что она существенно комплементарна к указанной диагностической части, терминальный нуклеотид диагностического праймера является либо комплементарным к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду, так что при использовании синтезируется продукт распространения диагностического праймера, когда указанный терминальный нуклеотид диагностического праймера является комплементарным к соответствующему нуклеотиду в основной последовательности мишени, причем продукт распространения не синтезируется, когда указанный терминальный нуклеотид диагностического праймера не является комплементарным к соответствующему нуклеотиду в основной последовательности мишени;
2) каждый из четырех различных нуклеозид-трифосфатов;
3) реагент для полимеризации нуклеозид-трифосфатов на стадии 2.
Преимущественно набор настоящего изобретения, кроме того, включает амплификационный праймер, соответствующий каждому диагностическому праймеру, причем нуклеотидная последовательность амплификационного праймера является таковой, что любой продукт распространения соответствующего диагностического праймера после отделения от его комплемента, может служить в качестве матрицы для синтеза продукта распространения амплификационного праймера. Например, набор настоящего изобретения включает каждый или оба ряда диагностических праймеров, описанных выше в связи с вторым вариантом воплощения изобретения.
Набор настоящего изобретения может также включать праймеры внутреннего контроля, где это целесообразно.
Однако особенно предпочтительно, чтобы набор настоящего изобретения включал PCR (реакция полимеразной цепи) праймеры и диагностический праймер (который определен в дальнейшем) в отношении к каждому предполагаемому вариантному нуклеотиду. По желанию этот набор может дополнительно содержать каждый или оба набора диагностических праймеров, которые подробно указаны выше в связи со вторым вариантом воплощения изобретения.
Каждый из материалов, указанных в 1, 2 и 3, и/или амплифицированный праймер, могут быть для удобства упакованы в отдельные контейнеры, но предпочтительно все они могут быть объединены в единственном контейнере, в который добавляется материал, подлежащий анализу. Преимущественно этот единственный контейнер может дополнительно содержать буфер.
Следует признать, что когда набор настоящего изобретения содержит оба набора диагностических праймеров а и б, указанных выше в связи с вторым вариантом воплощения изобретения, то оба набора диагностических праймеров могут не присутствовать вместе в одном контейнере, хотя каждый набор праймеров может присутствовать вместе с каждым из материалов, указанных подробно в 2 и 3, и/или с амплификационными праймерами в раздельных контейнерах. Когда подлежащий испытанию образец необходимо первоначально экстраполировать согласно публикации Европатента N 237362, может быть выгодным включить PCR-праймер, а также диагностические праймеры в один контейнер вместе с указанными выше в 2 и 3 материалами. Однако диагностические праймеры могут присутствовать в отдельных контейнерах для последующего использования, после завершения амплификации.
Термин "нуклеозид-трифосфат" используется здесь для обозначения нуклеозидов, присутствующих либо в ДНК, либо в РНК, и таким образом, включает нуклеозиды, которые содержат в качестве основания аденин, цитозин, гуанин, тимин и урацил, причем функциональной группой сахара является деоксирибоза или рибоза. Обычно деоксирибонуклеозиды могут применяться в сочетании с полимеразой ДНК. Однако можно признать, что могут применяться другие модифицированные основания, способные к спариванию оснований с одним из традиционных оснований аденина, цитозина, гуанина, тимина и урацила. Такие модифицированные основания включают, например 8-азагуанин и гипоксантин.
Термин "нуклеотид", используемый здесь, может относиться к нуклеотидам, присутствующим либо в ДНК, либо в РНК, и таким образом включает нуклеотиды, которые содержат в качестве основания аденин, цитозин, гуанин, тимин и урацил, причем функциональной группой сахара является деоксирибоза или рибоза. Однако следует признать, что другие модифицированные основания, способные к спариванию оснований с одним из традиционных оснований, аденином, цитозином, гуанином, тимином и урацилом, могут использоваться в диагностическом праймере и амплификационном праймере, применяемом в настоящем изобретении. Такие модифицированные основания включают, например, 8-азагуанин и гипоксантин.
Следует признать, что когда способ предполагают использовать для определения наличия или отсутствия предполагаемого варианта нуклеотида, который находится по соседству с частью последовательности оснований мишени, которая не содержит всех четырех различных нуклеотидов, тогда продукт распространения диагностического праймера и по желанию продукт распространения амплификационного праймера могут образоваться только в присутствии надлежащих соответствующих нуклеозид-трифосфатов, и все четыре различных нуклеозид-трифосфата не будут необходимыми.
Реагентом для полимеризации нуклеозид-трифосфатов может быть любое соединение, которое будет способствовать синтезу продуктов для распространения праймера, включая ферменты. Подходящие для этой цели ферменты включают например, полимеразу 1 ДНК E.coli, фрагмент Кленова полимеразы 1 ДНК E.coli, Т4 ДНК полимеразу, другие доступные ДНК-полимеразы, реверсивные транскриптазы и другие ферменты, включая термостабильные ферменты. Термин "термостабильный фермент", используемый здесь, относится к ферменту, который является стабильным к нагреванию и обладает термической стабильностью, и катализирует (ускоряет) сочетание нуклеотидов надлежащим образом, чтобы образовались продукты распространения праймера, которые комплементарны к каждой спирали нуклеиновой кислоты. Синтез может инициироваться при 3'-конце каждого праймера и может протекать в направлении 5' вдоль спирали матрицы, пока синтез не прекратится, образуя молекулы различной длины. Например, там могут быть ферменты, например термостабильные ферменты, которые, однако, инициируют синтез при конце 5', причем он протекает в другом направлении с использованием того же самого процесса, что описан выше. Предпочтительным термостабильным ферментом, который может применяться в способе по изобретению, является тот, который может быть экстрагирован и очищен от Thermus aquaticus. Такой фермент имеет молекулярный вес примерно 86000-90000 единиц Дальтона, как описано в публикации Европатента N 237362 (см. также публикацию Европатента N 258017). Штамм VTI Thermus aquaticus является доступным без ограничений из Коллекции Культур Американского Типа, 12301 Парклоун Драйв, г. Роквилл, шт. Мэриленд, США, как АТСС 25 104.
Выражение "диагностическая часть", которое используется здесь, обозначает ту часть основной последовательности мишени (которая определена в дальнейшем), которая содержит в качестве терминального нуклеотида потенциальный вариантный нуклеотид, присутствие или отсутствие которого необходимо определить. Обычно этот потенциальный вариантный нуклеотид будет находиться при 5'-терминальном конце диагностической части, поскольку в общем синтезе продуктов распространения праймера будет инициироваться при 3'-конце каждого праймера, как описано выше. Однако, когда необходимо использовать реагент для полимеризации, который инициирует синтез при 5'-конце диагностического праймера и направляет его в направлении 3' вдоль матричной спирали, до тех пор пока синтез не прекратится, "диагностическая часть" будет содержать потенциальный вариантный нуклеотид при своем 3'-конце. Диагностические праймеры также будут соответственно спроектированы в этом отношении, как изложено ниже.
Выражение "основная последовательность мишени", используемое здесь, означает нуклеотидную последовательность, включающую по меньшей мере одну диагностическую часть (как определено выше). Так, например, в отдельном испытании на бета-талассаэмию последовательность мишени может содержать до 60, например 50, диагностических частей, причем каждая диагностическая часть содержит отдельный потенциальный вариантный нуклеотид.
Термин "олигонуклеотид", используемый здесь, определяется как молекула, включающая два или более деоксирибонуклеотидов или рибонуклеотидов, предпочтительно больше трех. Точный его размер будет зависеть от многих факторов, таких как температура реакции, концентрация соли, наличие формамида и присутствие других закрытых мутаций, таких как заболевание срощенных клеток HbC, которые, в свою очередь, зависят от конечной функции или использования олигонуклеотида. Точная последовательность олигонуклеотида также может зависеть от ряда факторов, которые описаны в последующем. Эти олигонуклеотиды могут быть получены синтетически или путем клонирования.
Термин "праймер", используемый здесь, относится к олигонуклеотиду независимо от того, имеют ли они природное происхождение, как в очищенном ограничивающем дайджесте, или получаются синтетически, который способен действовать как место инициирования синтеза, когда его помещают в условия, в которых вызывается синтез продукта распространения праймера, который комплементарен к спирали нуклеиновой кислоты, т.е. в присутствии соответствующих нуклеозид-трифосфатов и реагента для полимеризации, такого как ДНК-полимераза в подходящем буфере (термин "буфер" включает pH, ионную силу раствора, сопутствующие факторы и др.) и при подходящей температуре.
Предпочтительно праймер является односпиральным для максимальной эффективности при амплификации, но может быть альтернативно двухспиральным. Если он двухспирален, то праймер сначала обрабатывается, чтобы выделить его спирали до использования, чтобы приготовить продукты распространения. Предпочтительно праймер является олигодеоксирибонуклеотидом. Праймер должен быть достаточно длинным, чтобы зарождать синтез продуктов распространения в присутствии реагента для полимеризации. Точные значения длины праймеров будут зависеть от многих факторов, включая температуру и источник праймера и использование способа. Например, в зависимости от сложности последовательности мишени диагностические и амплификационные праймеры содержат 12-35, например 15-35, нуклеотидов, хотя они могут содержать больше или меньше нуклеотидов. Обычно для коротких молекул праймера требуются пониженные температуры, чтобы образовался достаточно стабильный гибридный комплекс с матрицей.
Термин "комплементарный к" используется здесь и связан с нуклеотидами и означает нуклеотид, который может спаривать основание с другим специфическим нуклеотидом. Так оденозин-трифосфат комплементарен к уридин-трифосфату или тимидин-трифосфату, а гуанозин-трифосфат комплементарен к цитидин-трифосфату. Признано, что хотя тимидин-трифосфат и гуанозин-трифосфат могут в определенных условиях спаривать основания, они не считаются комплементарными для целей этого описания. Кроме того, следует признать, что хотя цитозин-трифосфат и аденозинтрифосфат могут спаривать основания в определенных условиях, они не считаются комплементарными для целей этого описания. То же самое приложимо к цитозин-трифосфату и урацил-трифосфату.
Праймеры в этом изобретении выбираются таким образом, чтобы они были "существенно" комплементарны к различным спиралям каждой специфической последовательности, которая подлежит амплификации. Это означает, что праймеры должны быть достаточно комплементарны, чтобы гибридизоваться их соответствующими спиралями. Поэтому последовательность праймера не должна отражать точную последовательность матрицы. Например, когда диагностический праймер включает последовательность нуклеотида, в которой 3'-терминальный нуклеотид комплементарен либо к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду, не комплементарный нуклеотидный фрагмент может присоединяться к 5'-концу праймера, причем остальная часть последовательности праймера является комплексной к диагностической части основной последовательности мишени. Однако обычно праймеры обладают строгой комплементарностью за исключением того, насколько некомплементарные нуклеотиды могут присутствовать при заданном конце праймера, как было описано ранее.
В некоторых обстоятельствах синтез продукта распространения диагностического праймера может инициироваться даже в присутствии некомплементарного 3'-терминального остатка. Этот артефактный результат может возникать вследствие использования слишком низкой температуры, в этом случае температура может быть повышена; вследствие слишком длительного периода инкубации/отпуска, в этом случае время может быть уменьшено; вследствие слишком высокой концентрации соли, в этом случае концентрация соли может быть понижена; вследствие слишком высокой концентрации фермента, слишком высокой концентрации нуклеозид-трифосфата, неправильного значения pH или неправильной длины олигонуклеотидного праймера. Все эти факторы обсуждаются в публикации Европатента N 237362. Вероятно, что основным источником артефактных продуктов является возможность слишком большого снижения температуры реакции, что обуславливает слишком низкую точность, например, путем удаления реакционной смеси из устройства для циклического нагрева даже на короткий период, например, для добавления реагента для полимеризации (например, Таg-полимеразы), особенно в первом реакционном цикле. Кроме того, обнаружено, что такие артефактные результаты также могут возникать от использования диагностического праймера, который особенно обогащен остатками G (гуанозин) и С (цитидин). Диагностический праймер может вызывать затруднения в связи с этим, если в целом он обогащен G/C или особенно если он обогащен G/C при соответствующем, обычно 3', конце. Более того, точная природа спаривания оснований в области соответствия, обычно 3'-конца, диагностического праймера при использовании может быть причиной артефактного результата. Так присутствие As (аденозина) при спаривании оснований в области соответствия, обычно 3'-, когда наличие диагностического праймера приводит к улучшению специфичности, хотя наличие Gs (гуанозина) не приводит к этому. Более того, точная природа непарности при соответствующем, обычно 3'-, конце диагностического праймера может быть важным фактором того, получится артефактный результат или нет. Так например, АА или СТ непарность обычно не приводит к гибридизации, но GT или AC непарность могут привести к значительной степени гибридизации, в результате чего образуется артефактный продукт (продукты). Артефактных продуктов можно избежать путем умышленного введения одного или нескольких дополнительных непарных остатков, или исключений или включений внутри диагностического праймера, чтобы дестабилизировать праймер путем дальнейшего уменьшения связывания в ходе гибридизации.
Например, любой или несколько (из 10), например шесть, нуклеотидов, соседних с терминальной непарностью, могут быть изменены, чтобы ввести дополнительную непарность. Вообще только одна непарность, кроме терминальной, может быть необходимой, которая расположена, например, на 1, 2 или 3 основания от терминальной непарности. Например, в связи с определением наличия нормальной гомозиготы, гетерозиготы или поражения гомозиготы, в отношении Z-аллеля альфа 1-антитрипсинового гена мы обнаружили, что могут быть получены хорошие результаты, если третий нуклеотид от 3'-терминального нуклеотида изменен, чтобы порождать непарность при использовании. Так например, мы обнаружили, что наличие С вместо А в третьем нуклеотиде от 3'-терминуса диагностического праймера обеспечивает нормальные гомозиготы, гетерозиготы и пораженные гомозиготы в отношении Z-аллеля, который подлежит быстрому различению.
Таким образом, наилучший дизайн диагностического праймера может быть определен путем непосредственного эксперимента на основе указанных выше критериев, причем такой эксперимент может хорошо осуществить специалист по молекулярной биологии.
Термин "диагностический праймер", используемый здесь, относится к праймеру, который имеет такую нуклеотидную последовательность, что его терминальный нуклеотид выбирается таким образом, чтобы быть комплементарным к предполагаемому вариантному нуклеотиду или соответствующему нормальному нуклеотиду, так что продукт распространения диагностического праймера синтезируется, когда терминальный нуклеотид диагностического праймера комплементарен к соответствующему терминальному нуклеотиду соответствующей диагностической части последовательности оснований мишени, когда терминальный нуклеотид диагностического праймера не является гомологом соответствующего терминального нуклеотида соответствующей диагностической части последовательности оснований мишени.
Термин "амплификационный праймер", используемый здесь, относится к праймеру, который способен гибридизоваться к спирали нуклеиновой кислоты, который комплементарен к спирали нуклеиновой кислоты, с которой диагностический праймер способен гибридизоваться, причем "амплификационный праймер" имеет такую нуклеотидную последовательность, что он способен гибридизоваться с продуктом распространения диагностического праймера после отделения от его комплемента, посредством чего продукт распространения диагностического праймера служит в качестве матрицы для синтеза продукта распространения амплификационного праймера, тем самым облегчается амплификация.
Таким образом, изобретение способствует, по крайней мере отчасти, усовершенствованию процессов амплификации, таких как способ, описанный в публикации Европатента N 237362, в котором усовершенствование дает возможность селективно амплифицировать либо последовательность, содержащую предполагаемый вариантный нуклеотид, если он имеется, либо последовательность, содержащую соответствующий нормальный нуклеотид, если он присутствует, таким образом упрощается определение и пока избегают определения последовательности аллельных специфических олигонуклеотидов и ограничительного расщепления. Так для данной нуклеотидной вариации, например точечной мутации, ее наличие или отсутствие может быть определено либо
1) проектированием диагностического праймера таким образом, чтобы он имел соответствующий терминальный нуклеотид, который комплементарен к предполагаемой нуклеотидной вариации, так что синтез амплифицированного продукта будет указанием наличия предполагаемой нуклеотидной вариации, а отсутствие амплифицированного продукта будет указанием отсутствия предполагаемой нуклеотидной вариации, либо
2) посредством проектирования диагностического праймера, чтобы он имел соответствующий терминальный нуклеотид, который комплементарен к соответствующему нормальному нуклеотиду, так что синтез амплифицированного продукта будет указанием отсутствия предполагаемой нуклеотидной вариации, а отсутствие амплифицированного продукта будет указанием присутствия предполагаемой нуклеотидной вариации. В этом отношении ссылки в описании на "соответствующий терминальный нуклеотид" означают терминальный нуклеотид праймера, из которого при использовании может быть инициирован синтез, если это возможно. Так как в общем реагент для полимеризации будет инициировать синтез при 3'-конце праймера, то соответствующий терминальный нуклеотид в общем будет 3'-терминальным нуклеотидом.
Подтверждение наличия или отсутствия, например, заданной точечной мутации может быть получено посредством принятой как альтернативной методики 1, так и альтернативной методики 2, которые представлены выше. Сочетание этих двух подходов обеспечивает способ определения гетерозигот, которые могут представлять интерес для анализа доминирующих наследованных состояний, и при определении носителей рецессивных наследованных состояний.
В предпочтительном варианте изобретение направлено на определение наличия или отсутствия более чем одного предполагаемого вариантного нуклеотида в одном и том же образце. Способность изобретения к селективному амплифицированию последовательностей, зависящая от предварительно заданной нуклеотидной последовательности диагностических праймеров, обеспечивает простое различие сложных продуктов амплификации с высокой точностью при минимальной квалификации оператора, таким образом возможно предоставление ясной методики для скрининга единственного образца для сложных нуклеотидных вариаций. Таким образом, изобретение представляет особый интерес при скрининге отдельного образца ДНК или РНК для ряда наследованных состояний, таких как генетические нарушения, предрасположения и соматические мутации, приводящие к различным заболеваниям. Такие ДНК или РНК, например, могут экстрагироваться из крови или тканевого материала, такого как хорионные ворсинки или амниотичные клетки, с помощью множества методик, таких как те, которые описаны Маниатисом и сотр. в журнале Molecular Cloning (1982), с.280-281. Кроме того, когда молекулярные основания для других наследованных состояний станут известными, эти дополнительные состояния могут быть просто включены в методику скрининга настоящего изобретения.
Сложные продукты амплификации могут быть распознаны с помощью множества методик. Например, могут применяться зонды для каждого предполагаемого амплифицированного продукта, причем каждый зонд содержит различный и способный к распознаванию сигнал или остаток, способный производить сигнал.
Такие сигналы и остатки, способные производить сигнал, обсуждаются подробно в публикации Европатента N 246864, но могут, например, включать твердофазную систему амплификации, описанную Вангом Си.Джи в журнале World Biotech Report, 1986, т. 2, ч.2, с.33-37 (Труды конференции по диагностике в здравоохранении, проведенной в ноябре 1986 в г. Сан-Франциско), в которой к зонду присоединены микрослои, полученные с многими выбранными следовыми элементами. Наличие специфических зондов может быть определено методом рентгено-флуоресцентного анализа. Такие методики обычно могут быть простыми и могут использоваться непосредственно, так как необходимо будет только определить существование продукта амплификации, а не распознать последовательности, различающиеся столь незначительно, как на единственный нуклеотид.
Гораздо более простой и предпочтительный способ распознавания продуктов амплификации включает выбор такой последовательности нуклеотидов амплифицированных праймеров, чтобы длина каждого амплифицированного продукта, образовавшегося в ходе процесса настоящего изобретения, была различной. В связи с этим число пар оснований, присутствующих в амплифицированном продукте, диктуется расстоянием между диагностическими и амплификационными праймерами. Амплификационные праймеры могут быть спроектированы таким образом, чтобы каждый потенциальный вариантный нуклеотид был связан с потенциальным амплификационным продуктом различной длины.
Наличие или отсутствие данного потенциального вариантного нуклеотида может быть таким образом удобно определено по методике электрофореза, при которой полученные различные амплифицированные продукты могут быть распределены в соответствии с их молекулярным весом и тем самым идентифицированы, например, методами ауторадиографии или флуоресценции. Длины различных продуктов могут отличаться только единственным нуклеотидом, однако предпочтительно эти длины могут различаться по меньшей мере на 3 нуклеотида. Способ предпочтительно осуществлять путем использования интеркаляционного красителя, такого как этидиумбромид, который может наблюдаться как оранжевая флуоресценция при ультрафиолетовом облучении. Таким образом наличие или отсутствие множества потенциальных вариантных нуклеотидов в отдельном образце может быть определено быстро, легко и точно. По желанию диагностические и/или амплификационные праймеры могут маркироваться или метиться, например, путем использования флуорофора. Например, каждый отличающийся диагностический праймер или амплификационный праймер может содержать различный флуорофор, так что результаты ряда испытаний могут быть считаны с геля электрофореза, например методом лазерного сканирования, таким образом обеспечивается автоматизация способа по изобретению. Альтернативно наличие или отсутствие амплифицированного продукта может быть просто определено посредством использования растворителя, способного селективно растворять нуклеозид-трифосфаты, но не способного растворять нуклеотидную последовательность (например ДНК). Примером такого растворителя является трихлоруксусная кислота (ТХУК). Например, наличие или отсутствие амплифицированного продукта может быть определено путем осаждения ТХУК амплифицированных реакционных смесей. Если внедрение соответствующих нуклеозид-трифосфатов происходит в экспоненциальной реакционной последовательности, тогда будут присутствовать значительно увеличенные количества нерастворимого в трихлоуксусной кислоте материала, чем когда не происходит никакого распространения диагностического праймера. Количественное определение нерастворимого материала может быть выполнено известными методами. Например, нуклеозид-трифосфаты могут быть помечены (например радиоактивной или флуоресцентной меткой), реакционная смесь может быть подвергнута, например, центрифугированию, присутствующая жидкость декантирована, и либо жидкость, либо нерастворимый продукт подвергают соответствующему анализу определения, такому как радиоактивный счетчик или определение флуоресценции.
В соответствии с дополнительной характеристикой изобретения мы предоставляем нуклеотидную последовательность примерно от 5 до 50 bp для использования в способе по изобретению, причем терминальный нуклеотид указанной последовательности является комплементарным либо к предполагаемому вариантному нуклеотиду, связанному с известными генетическими нарушениями, либо к соответствующему нормальному нуклеотиду, причем остальная часть указанной последовательности существенно комплементарна к соответствующей последовательности оснований мишени, которая содействует с предлагаемым вариантным нуклеотидом или соответствующим нормальным нуклеотидом; указанная нуклеотидная последовательность такова, что при использовании в качестве диагностического праймера продукт распространения диагностического праймера синтезируется, если этот терминальный нуклеотид диагностического праймера комплементарен к соответствующему нуклеотиду в последовательности оснований мишени, причем продукт распространения не синтезируется, если указанный терминальный нуклеотид диагностического праймера не является комплементарным к соответствующему нуклеотиду в последовательности оснований мишени.
Удобно, чтобы терминальный нуклеотид, комплементарный либо к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду, находился при 3'-конце нуклеотидной последовательности. Предпочтительно предполагаемый вариантный нуклеотид возникает из точечной мутации соответствующей нормальной последовательности.
Указанная нуклеотидная последовательность может иметь, например, от 10 до 50 bp, например от 10 до 36 bp.
Согласно четырем дополнительным характеристикам изобретения мы предоставили нуклеотидные последовательности, которые определены в предыдущем абзаце и в которых терминальный нуклеотид нуклеотидной последовательности является комплементарным к вариантному нуклеотиду, полученному от изменения соответствующего нормального нуклеотида в (I)A, (II)G, (III)C, (IV)T или V соответственно.
В соответствии с дополнительной характеристикой изобретения мы предоставили набор двух нуклеотидных последовательностей, которые определены выше, причем терминальный нуклеотид одной последовательности является комплементарным к предполагаемому вариантному нуклеотиду, связанному с известными генетическими нарушениями, а терминальный нуклеотид другой последовательности комплементарен к соответствующему нормальному нуклеотиду.
Согласно еще одной характеристике изобретения в нем разработаны зонды, включающие нуклеотидную последовательность, которая определена выше, содержащую меченый или маркированный компонент.
Мутации, вызывающие генетические заболевания, приведены в таблице.
Часто для анализа доступны лишь небольшие количества геномной ДНК. Было установлено, что специфичность диагностических праймеров для релевантных нормальных или вариантных нуклеотидных последовательностей можно легко оценить путем увеличения числа копий нуклеотидных последовательностей при гибридизационном анализе. Это достигается, например, конструированием двунитевой "кассеты", включающей в себя нормальную последовательность, соединенную с вариантной последовательностью, причем непарность двух нуклеотидов в соединенных цепочках предпочтительно должна быть направлена к середине кассеты. Копии кассеты затем удобно получать, вставляя ее в плазмиду-хозяина с последующей репликацией плазмиды и выделением нужных последовательностей, что делается по хорошо известным методикам. Этот метод проиллюстрирован на фиг. 10.
Для более полного понимания данного изобретения оно описывается ниже с помощью примеров и фигур, где: фиг.1а и 1в иллюстрируют первый вариант данного изобретения, фиг.1с иллюстрирует типичный результат такого испытания, как это можно показать электрофоретически; фиг.2а и 2в иллюстрируют второй вариант данного изобретения; фиг.3 иллюстрирует предпочтительный способ различения продуктов множественной амплификации; фиг.4а и 4в иллюстрируют третий вариант данного изобретения; на фиг.5а, в и с проиллюстрирован четвертый вариант данного изобретения; на фиг.6 (не в масштабе/ показан ген человеческого α1-антитрипсина; на фиг.7 представлен результат визуализации геля, полученного в примерах 1 и 2; на фиг.8 представлен результат визуализации геля, где треки 1-4 являются результатом примера 3, треки 5-8 это результаты примера 4 и трек 9 соответствует маркеру размеров; на фиг. 9 представлены результаты визуализации геля, полученного по примеру 4; фиг.10 иллюстрирует использование кассетной плазмидной системы для амплификации нужного ДНК-дуплекса; фиг. 11 иллюстрирует пятый вариант (линейную амплификацию) данного изобретения; на фиг. 12 показаны результаты визуализации геля, полученного по примеру 5; на фиг. 13 показаны результаты визуализации геля, полученного по примеру 6.
На фиг. 1 проиллюстрирован способ по первому варианту данного изобретения. На фиг.1а показаны нити денатурированной геномной ДНК, содержащие нормальный нуклеотид /N/ в том положении, которое соответствует, например, тому положению, в котором может находиться вариантный нуклеотид, являющийся результатом генетического нарушения. Как показано на фиг. 1в, контакт цепочки нуклеиновых кислот в условиях гибридизации с диагностической пробой, имеющей 3'-концевой нуклеотид, комплементарный нормальному нуклеотиду /-N/ в присутствии соответствующих нуклеозидтрифосфатов и агента полимеризации нуклеозидтрифосфатов приводит к удлинению цепи диагностического праймера в направлении 3'. Удлинение цепи не наблюдается, если используется диагностический праймер, в котором 3'-концевой нуклеотид комплементарен к ожидаемому вариантному нуклеотиду /-М/. Любой продукт удлинения цепи диагностического праймера может быть затем денатурирован и амплифицированный праймер /А/, гибридизован до полученного денатирурованного продукта с получением дальнейшего удлинения цепи продукта. Продукт удлинения цепи можно затем денатурировать и повторить процесс с достижением амплификации. Участок геномной ДНК, подвергаемый амплификации диагностическим праймером, обозначен DG на фиг.1в. Диагностические продукты этого участника соответствуют полосам, обозначенным DG на фиг. 1с. В качестве контроля PCR /полимеразная цепочечная реакция/ праймеры /Р/, обозначенные Р на фиг. 1в, можно использовать в отдельном сайте цепочки нуклеиновых кислоты или в другой цепочке нуклеиновых кислот, например в другой хромосоме человеческой ДНК. Этот отдельный сайт обозначен PCR на фиг. 1с и PCR-контрольные продукты этого сайта соответствуют полосам, обозначенным PCR на фиг. 1с. Понятно, что полосы PCR могут соответствовать большим или меньшим продуктам, чем продукт, соответствующий полосе DG.
На фиг. 1с представлен результат, полученный способом, описанным на фиг. 1а и в, в форме полос, разделенных электрофорезом на агарозном геле для генетической мутации, вызванной точечной мутацией и проанализированной соответственно диагностическим праймером -М или -N. При этом тест-объектом является носитель /С/ полосы генетического нарушения, что видно как для нормальной /N/ нуклеотидной последовательности, так и для вариантной /М/ нуклеотидной последовательности. Нормальные гомозиготы /N/ будут давать полосы, соответствующие нормальной нуклеотидной последовательности, но не полосы соответствующие вариантному нуклеотиду, а гомозиготы вызванной заболеванием мутации /подвергнутые генетическому нарушению D / не будут давать полос, присутствующих в случае нормальной нуклеотидной последовательности, и полос, относящихся к вариантной нуклеотидной последовательности.
На фиг. 2 показан второй вариант данного изобретения. Диагностический праймер используют для каждой цепочки двунитевой нуклеиновой кислоты. В /а/ диагностические праймеры обозначены таким образом, что 3'-концевой нуклеотид комплементарен нормальному нуклеотиду. Способ осуществляется так, как описано на фиг.1, но присутствуют два различных диагностических праймера, инициирующих амплификацию. Таким образом в данном варианте второй диагностический праймер эквивалентен амплификационному праймеру /А/ на фиг.1. Как видно из фиг. 2а, амплификация инициируется праймерами /-N и N-/, но не диагностическими праймерами, имеющими 3'-концевые нуклеотиды, комплементарные вариантной нуклеотидной последовательности /-М и М- /. Обратное справедливо для фиг. 2в, так как вариантная последовательность присутствует в цепочке нуклеиновой кислоты образца. Результаты осуществления способа, описанного на фиг. 2, могут быть представлены в виде, аналогичном изображенному на фиг. 1с, но нормальные и вариантные /N и М/ последовательности будут соответствовать парам таких последовательностей. На фиг. 2 буквой Р обозначены PCR-праймеры, используемые в качестве контроля.
На фиг. 3 показан один из путей различения продуктов множественной амплификации, что дает возможность испытать один образец РНК или ДНК для набора условий. Две нити /денатурированные/ ДНК или РНК образца обозначены имеющими три отдельные зоны 1,2 и 3. Дополнительный выделенный из этих зон участок используют для контрольной PCR-амплификации. Для зоны /1/ 20-мерный диагностический праймер /5'-N 3'/ используют вместе с дополнительным 20-мерным /3'N-5'/ диагностическим праймером. Если амплификация протекает по зоне /1/, то получают 39-мерный продукт амплификации. На чертеже 3'-концевой нуклеотид является нормальным как релевантный нуклеотид в испытуемом образце, и поэтому амплификация будет протекать. Аналогичным образом амплификация будет происходить, если релевантный нуклеотид в испытуемом образце является вариантным нуклеотидом и используемые диагностические праймеры несут также 3'-концевой вариантный нуклеотид. Однако амплификация не будет протекать если имеется непарность между релевантным нуклеотидом образца и 3'-концевым нуклеотидом диагностического праймера. Для зоны /2/ диагностические праймеры таковы, что дают 49-мерный амплификационный продукт при протекании амплификации, а праймеры для зоны /3/ дают 59-мерный амплификационный продукт в ходе амплификации. Из вышеизложенного ясно, что размер полосы амплификационного продукта является суммой размеров двух диагностических праймеров минус один, и можно проводить множественные испытания на одном образце в одном реакционном сосуде, соответствующим образом конструируя диагностические праймеры для характеризации каждой интересующей зоны. Так как размер суммы этих размеров диагностических праймеров увеличивается, то продукты амплификации можно разделить, например, на агарозном геле. Улучшенное разделение возможно, если диагностические праймеры помечены любым удобным способом и затем разделены, например, на акриламидном геле. На фиг.3 символом Р обозначены PCR-праймеры, используемые в качестве контроля.
На фиг.4 показан третий вариант данного изобретения, в котором проводят обычную амплификацию последовательности нуклеиновой кислоты, содержащей нормальный нуклеотид в том положении, где ожидается возможный вариантный нуклеотид, и здесь можно использовать, например, обычные PCR-праймеры. Аналогичным образом обычная амплификация будет протекать, если ожидаемый вариантный нуклеотид присутствует вместо нормального нуклеотида. Продукт амплификации контактирует с диагностическим праймером, как описано выше для третьего варианта изобретения. Если продукт амплификации содержит последовательность нуклеиновой кислоты, имеющую нормальный нуклеотид, как описано выше, а используемый диагностический праймер имеет 3'-концевой нормальный нуклеотид, то продукт удлинения цепи диагностического праймера будет образовываться с использованием в качестве матрицы нуклеотидной последовательности, получаемой обычной PCR-амплификацией, обозначаемой здесь как PCR-контрольный продукт, если присутствуют соответствующие нуклеозидтрифосфаты и агент полимеризации нуклеозидтрифосфатов. Нет необходимости в присутствии праймера амплификации, поскольку матрица для получения продукта удлинения цепи диагностического праймера всегда будет амплифицирована. Присутствие или отсутствие продукта удлинения цепи диагностического праймера, обозначенного как 1G на фиг.4в, а также присутствие PCR-контрольного продукта, обозначенного как PCR на фиг.4в, можно зарегистрировать, например, с помощью электрофореза.
Понятно, что продукт удлинения цепи диагностического праймера образуется также, если PCR-контрольный продукт содержит вариантный нуклеотид и используемый диагностический праймер имеет 3'-концевой комплементарный вариантный нуклеотид.
Полосы результирующего продукта могут быть визуализированы, например, так, как показано на фиг.4в. Символы N-, M-, C-, N, D, DG и PCR пояснены на фиг.1с. Однако полоса продукта удлинения цепи диагностического праймера имеет меньший молекулярный вес, чем PCR-контрольная полоса /как это показано на фиг. 4в/. В этом случае контрольная полоса является реальным внутренним контролем, поскольку один из PCR-праймеров служит амплификационным праймером для амплификации продукта удлинения цепи диагностического праймера. Если PCR-контрольный продукт содержит вариантный нуклеотид, а диагностический праймер имеет 3'-концевой нормальный нуклеотид, то продукт удлинения цепи не образуется; это верно и для обратного варианта.
При необходимости третий вариант данного изобретения можно осуществлять, вводя испытуемый образец в начале испытания в контакт не только с PCR-праймером, но и с диагностическим праймером. Поэтому нет необходимости откладывать использование диагностического праймера до достижения нужной степени амплификации PCR-контрольного продукта. Действительно, диагностический праймер можно использовать как одновременно, так и в любой момент после использования PCR-праймеров. Следовательно, испытание можно проводить, например, простым добавлением испытуемого образца в один контейнер, содержащий PCR-праймеры, диагностический праймер, соответствующие нуклеозидтрифосфаты и агент полимеризации нуклеозидтрифосфатов. Получаемые продукты и соответственно полосы будут такими, как описано выше и обозначено на фиг.4.
На фиг.5 описан четвертый вариант данного изобретения, где обычную амплификацию проводят так, как описано на фиг.4. Полученный таким образом продукт амплификации вводят затем в контакт в условиях гибридизации либо с /а/ двумя вышеописанными отдельными диагностическими праймерами, каждый из которых имеет 3'-концевой нормальный нуклеотид, или /в/ двумя вышеописанными диагностическими праймерами, каждый из которых имеет 3'-концевой вариантный нуклеотид /см. фиг. 5а/. Образование продуктов удлинения цепи в результате одного цикла покажет, содержит ли образец нуклеиновой кислоты нормальный или ожидаемый вариантный нуклеотид, и электрофорезом будут получены полосы, сходные с полосами, изображенными на фиг.5в. Однако следует понимать, что при использовании двух диагностических праймеров один из диагностических праймеров служит амплификационным праймером для другого. Поэтому при необходимости любой продукт /продукты/ удлинения цепи сам может быть подвергнут амплификации. Таким образом после многократных дополнительных циклов продукты более низкого молекулярного веса, такие как продукты, соответствующие дополнительной полосе на рис.5с, будут получены в количестве, соответствующем относительному содержанию первоначальных PCR-праймерных олигонуклеотидов и добавленных диагностических праймеров. PCR-амплификационные продукты можно легко отличить от амплификационных продуктов удлинения цепи диагностического праймера, например, по увеличению или уменьшению расстояния от связывающих сайтов PCR-праймеров в геноме.
При желании можно осуществить четвертый вариант данного изобретения введением испытуемого образца в начале испытания в контакт не только с PCR-праймерами, но и с диагностическими праймерами. Нет необходимости откладывать использование диагностического праймера до достижения нужной степени амплификации PCR-контрольного продукта. Действительно, диагностические праймеры можно использовать одновременно или в любой момент после использования PCR-праймеров. Поэтому испытание можно проводить, например, простым добавлением испытуемого образца в один контейнер, содержащий PCR-праймеры, диагностические праймеры, соответствующие нуклеозидтрифосфаты и агент полимеризации нуклеозидтрифосфатов. Если испытание проводить таким образом, то диагностические продукты более низкого молекулярного веса, указанные выше, образуются и дадут дополнительные полосы, указанные на фиг.5.
На фиг.6 (без соблюдения масштаба) изображен ген человеческого a1-антитрипсина, хорошо известный в данной области /Long G.L et al. Biochemistry, vol. 23, p.4828-4837, 1984/, причем показано также взаимное расположение эксонов III и IV, которые потенциально несут S и Z мутации a1-антитрипсинового белка. В частности, на чертеже изображен эксон III гена a1-антитрипсина и показано положение S-варианта, где кодон GAA мутирован в GТA, что приводит к получению валинового остатка в аминокислотном положении 264 вместо остатка глутаминовой кислоты. Праймер 1 имеет следующую последовательность:
5' CCCACCTTCCCCTCTCTCTCCAGGAAATGGG 3' I
которая гибридизируется в положения 7440-7469 гена a1- антитрипсина и либо праймер II имеет следующую последовательность:
5' GGGCCTCAGTCCCAACATGGCTAAGAGGTG 3' II
либо праймер IIа /СТ-3'-непарность в II/ имеет следующую последовательность:
5' GGGCCTCAGTCCCAACATGGCTAAGAGGTT 3' IIа
где каждый из них соответствует положению 7770-7799 гена a1- антитрипсина и может быть легко использован.
На фигуре изображен также эксон V гена a1-антитрипсина и положение Z-варианта, в котором кодон GAG мутирован до AAG, что приводит к получению лизинового остатка в аминокислотном положении 342 вместо остатка глутаминовой кислоты. Праймер III имеет следующую последовательность:
5' TGTCCAGGTGAGCCTTCGAGGCCTGGG 3'III
и гибридизуется в положениях 9890-9919 гена a1-антитрипсина, а праймер IV имеет следующую последовательность:
5' GAGACTTGGTATTTTGTTCAATCATTAAG 3' IV
и гибридизуется в положениях 10081-10109 гена a1-антитрипсина, и эти праймеры можно легко использовать. Вышеуказанные номера оснований приведены в соответствии с Biochem. 23, 4828-4837, 1984.
На фиг.7 показан результат визуализации геля, полученного по примерам 1 и 2. Трек 1 соответствует плазмиде, содержащей эксон V и расщепленной с помощью AluI; трек 2 соответствует маркеру размера, представляющему собой ДНУ бактериофага Х174, расщепленного Нае III; трек 3 соответствует амплификации эксона III праймерами I и II, как это описано выше, и амплификации эксона V праймерами III и IV, как это описывалось выше, в одной реакционной смеси с получением продуктов длиной соответственно 360 и 220 пар оснований. Из трека 4 видно, что не происходит амплификации эксона III при использовании праймеров I и IIa, как описывалось выше, однако происходит амплификация эксона V праймерами III и IV, как это указывалось выше. Трек 5 соответствует отрицательному контролю, где праймеры I, II, III и IV присутствуют в условиях, эффективных для промотирования амплификации, но не присутствует геномная ДНК.
На фиг.8 треки 1-3 соответствуют результатам примера 3, треки 5-8 - результатам использования нормального и мутантного диагностических праймеров, где не происходит дестабилизация в соответствии с примером 4, а трек 9 это маркер размера из бактериофага Х174. На треке 1 показан результат использования следующей нуклеотидной последовательности:
5'TGGTGATGATATCGTGGGTGTTCATTTT V
в качестве диагностического праймера и олигонуклеотида 1, описанного выше, в качестве амплификационного праймера. Используют человеческую геномную ДНК из нормальной гомозиготы, имеющую нормальный нуклеотид /А/ в потенциальном положении S мутации гена человеческого a1-антитрипсина. Ясно виден ожидаемый амплификационный продукт длиной 267 пар оснований. Трек 2 показывает результат использования следующей нуклеотидной последовательности:
5'TGGTGATGATATCGTGGGTGAGTTCATTTA VI
в качестве диагностического праймера и олигонуклеотида 1, описанного выше, в качестве амплификационного праймера. Используют человеческую геномную ДНК из нормальной гомозиготы, имеющую нормальный нуклеотид /А/ в потенциальном положении S мутации гена человеческого a1-антитрипсина. В присутствии А-А непарности продукт амплификации длиной 267 пар оснований не образуется. На треке 3 показана контрольная полоса на основе гена человеческого аполипопротеина В, где использованы следующие последовательности:
AATGAATTTATGAGCCAAAACTTTTACAGG XIII
и
CTCTGGGAGCACAGTACGAAAAACCACTT XIV
а трек 4 оставлен чистым.
На треке 5 показан результат использования нуклеотидной последовательности
5'CCGTGCATAAGGCTGTGCTGACCATCGAGG 3' VII
в качестве диагностического праймера по примеру 4. Диагностический праймер несет 3'-концевой нуклеотид /G/, способный образовывать пары оснований с нормальным нуклеотидом /C/ в потенциальной точке мутации Z и, с другой стороны, полностью комплементарный образцу ДНК. Образец ДНК получен нормальной гомозиготы, имеющей нормальный нуклеотид /C/, присутствующий в точке Z потенциальной мутации гена человеческого a1-антитрипсина. Трек 6 является результатом применения использования следующей нуклеотидной последовательности:
5'CCGTGCATAAGGCTGTGCTGACCATCGACA 3' VIII
1 качестве диагностического праймера в примере 4. Диагностический праймер содержит 3'-концевой нуклеотид /A/, способный к образованию пары с мутантным нуклеотидом /T/ в точке Z мутации, а в остальной части полностью комплементарный образцу ДНК. Используют образец ДНК из нормальной гомозиготы, имеющий нормальный нуклеотид /C/ в точке Z потенциальной мутации гена человеческого a1-антитрипсина. Трек 7 показывает результат использования нуклеотидной последовательности формулы VII в качестве диагностического праймера в примере 4. Используют образец ДНК из гомозиготы с генетическим нарушением человеческого a1-антитрипсина, имеющий мутантный нуклеотид /Т/ в точке Z мутации. Трек 8 является результатом использования нуклеотидной последовательности формулы VIII в качестве диагностического праймера по примеру 4. Используемый образец ДНК получен из гетерозиготы с генетическим нарушением человеческого a1-антитрипсина и имеет мутантный нуклеотид /Т/ в точке Z мутации. Нуклеотидная последовательность амплификационного праймера, использованная в каждом из испытаний, показанных треками 5-8, имеет вышеописанную формулу IV. Можно видеть, что выработка амплификационного продукта длиной 150 пар оснований не полностью подавляется присутствием А-С непарности /трек 6/ и еще меньше подавляется присутствием непарности GT /трек 7/.
На фиг. 9 показан результат визуализации геля, полученного в примере 4. Треки 1 и 14 соответствуют маркеру размеров, являющемуся продуктом расщепления ДНК бактериофага Х174 с помощью Нае III, трек 2 показывает результат использования следующей нуклеотидной последовательности:
5'CCGTGCATAAGGCTGTGCTGACCATAGACG 3' IX
в качестве диагностического праймера в присутствии образца ДНК, имеющего нормальный нуклеотид /С/ в точке Z потенциальной мутации гена человеческого a1-антитрипсина /образец ДНК из нормальной гомозиготы/. Диагностический праймер содержит изменение /подчеркнутое в последовательности IX A вместо С/ в пятом нуклеотиде от 3'-окончания, но 3'-концевой нуклеотид /G/, способный образовывать пару оснований с нормальным нуклеотидом /C/, присутствует в точке потенциальной мутации Z. Трек 3 показывает результат использования следующей нуклеотидной последовательности:
5'CCGTGCATAAGGCTGTGCTGACCATAGACA 3' X
в качестве диагностического праймера в присутствии образца ДНК, имеющего нормальный нуклеотид /С/ в точке возможной мутации /образец ДНК из нормальной гомозиготы/. Диагностический праймер несет изменение /подчеркнутое в последовательности Х-А вместо С/ в пятом нуклеотиде от 3'-окончания и 3'-концевой нуклеотид /А/, способный образовывать пару оснований с мутантным нуклеотидом /Т/, если он имеется в точке возможной мутации Z; трек 4 показывает результат использования нуклеотидной последовательности IX /указанной выше/ в качестве диагностического праймера в присутствии образца ДНК из гетерозиготы для мутации Z и, таким образом, носитель генетического нарушения дефицита человеческого a1-антитрипсина; трек 5 показывает результат использования нуклеотидной последовательности Х /как описано выше/ в качестве диагностического праймера в присутствии образца ДНК из гетерозиготы для мутации Z и, таким образом, носителя генетического нарушения дефицита человеческого a1-антитрипсина. Трек 6 показывает результат использования нуклеотидной последовательности IX /описанной выше/ в качестве диагностического праймера в присутствии образца ДНК из гомозиготы, имеющей генетическое нарушение человеческого a1-антитрипсина, а трек 7 является результатом использования нуклеотидной последовательности X /описанной выше/ в качестве диагностического праймера в присутствии образца ДНК из гомозиготы, подвергнутой генетическому нарушению человеческого a1-антитрипсина.
Трек 8 является результатом использования следующей нуклеотидной последовательности:
5'CCGTGCATAAGGCTGCTGACCATCGCCG3' XI
в качестве диагностического праймера в присутствии образца ДНК, имеющего нормальный нуклеотид /С/, присутствующий в точке потенциальной мутации Z гена человеческого a1-антитрипсина /образец ДНК из нормальной гомозиготы/. Диагностический праймер несет изменение /подчеркнутое в последовательности XI C вместо А/ в третьем нуклеотиде от 3'-конца, но 3'-окончание /G/, способное образовывать пару оснований с нормальным нуклеотидом /C/, присутствующим в потенциальной точке мутации Z; трек 9 показывает результат использования следующей нуклеотидной последовательности:
5' CCGTGCATAAGGCTGTGCTGACCATGCCA3' XII
в качестве диагностического праймера в присутствии образца ДНК, имеющего нормальный нуклеотид /C/, присутствующий в точке Z потенциальной мутации /образец ДНК из нормальной гомозиготы/. Диагностический праймер содержит изменение /подчеркнутое в последовательности XII -C вместо А/ в третьем нуклеотиде от 3'-окончания и 3'-концевой нуклеотид /A/, способный образовывать пару оснований с мутантный нуклеотидом /T/, если тот присутствует в потенциальной точке мутации Z; трек 10 показывает результат использования нуклеотидной последовательности XI, описанной выше, в качестве диагностического праймера в присутствии образца ДНК из гетерозиготы для мутации Z и таким образом носителя генетического нарушения дефицита человеческого a1-антитрипсина; трек 11 является результатом использования вышеописанной нуклеотидной последовательности XII в качестве диагностического праймера в присутствии образца ДНК из гетерозиготы для мутации Z и таким образом носителя генетического нарушения человеческого a1-антитрипсина. На треке 12 показан результат использования нуклеотидной последовательности XI в качестве диагностического праймера в присутствии образца ДНК из гомозиготы, подвергшейся генетическому нарушению дефицита человеческого a1-антитрипсина.
На треке 13 представлен результат использования нуклеотидной последовательности XII в качестве диагностического праймера в присутствии ДНК из гомозиготы, подвергшейся генетическому нарушению человеческого a1-антитрипсина.
В каждом из треков 2-13 нуклеотидную последовательность формулы IV используют в качестве амплификационного праймера, а в качестве контроля - нуклеотидные последовательности формулы
AATGAATTTATCAGCCAAAACTTTTACAGG XIII
и формулы
CTCTGGGAGCACAGTACGAAAAACCACTT XIV
Полосы, соответствующие контролю, обозначены /С/ на фиг.9 и соответствуют амплификационному продукту длиной 510 пар оснований из эксона 26 гена человеческого аполипопротеина В.
На фиг.10 проиллюстрировано использование кассетной плазмидной системы. Плазмиду рАТ 153, имеющую участки, ответственные за устойчивость к антибиотику ампициллину /Apn/ и тетрациклину /Tcn/, а также рестрикционные сайты EcoRI, BamHI и SalI, расщепляют с помощью Eco RI и BamHI. Как показано, синтетическую кассету, содержащую ДНК-дуплекс с непарным нуклеотидом в центре, обусловленным присутствием как нормальной, так и вариантной последовательностей, связывают с использованием Т4-ДНК-лигазы с плазмидой-хозяином. Затем, как показано, плазмиду реплицируют и отбирают.
Система более подробно иллюстрируется следующим образом: получают синтетические ДНК-кассеты, каждая из которых содержит два денатурированных 65-мера, имеющиx рестрикционные концы EcoRI и BamHI и непарную пару оснований в середине, обусловленную вариантным нуклеотидом. Как описывалось выше, эти кассеты вводят в плазмиду-хозяина рАТ153 и после трансформирования выделяют чувствительные к тетрациклину клоны и экстрагируют плазмиду. Затем плазмидные экстракты используют для вторичной трансформации с получением клонов, имеющих нормальную или вариантную вставки. Для подтверждения того, что клоны содержат полностью нормальную или мутантную последовательности, приблизительно 2 мг каждого образца секвентируют после денатурации с NaOH с использованием Т4-ДНК-полимеразы /Sequenase US Biochemicals/ и 32Р-меченного праймера. Если находят по крайней мере один нормальный и один вариантный тип, то соответствующие клоны каждого выращивают в массе, плазмиды экстрагируют и очищают в градиенте CsCl. Затем кассеты расщепляют с SalI, экстрагируют смесью фенол/хлороформ, осаждают и промывают 70%-ным этанолом. Затем кассеты легко использовать, например, в полимеразных цепных реакциях для определения чувствительности и специфичности различных олигонуклеотидов.
На фиг. 11 проиллюстрирован пятый вариант /линейная амплификация/ данного изобретения.
а/ Нормальная геномная ДНК- последовательность показана вместе с двумя диагностическими праймерами, один из которых комплементарен в своем 3'-окончании нормальной геномной ДНК-последовательности, а второй праймер имеет нуклеотид, комплементарный к ожидаемому вариантному нуклеотиду в своем 3'-окончании.
b/ Вариантная геномная ДНК-последовательность представлена вместе с этими же двумя диагностическими праймерами. Если каждую из геномных ДКН-последовательностей ввести в контакт с праймерами в условиях, позволяющих протекать комплементарной гибридизации, то в обоих случаях протекает гибридизация. Прибавление всех четырех нуклеозидтрифосфатов в условиях, позволяющих удлинять цепь праймера, будет приводить лишь к образованию удлиненного продукта в случае а/ праймера с нормальной последовательностью и b/ праймера с вариантной последовательностью, а удлинение другого праймера будет предотвращаться непарностью.
Кроме того, как показано в с/, если вместо четырех нуклеозидтрифосфатов добавить лишь три или меньше, то удлинение соответствующего праймера предотвращается в заданной точке из-за отсутствия соответствующего нуклеозидтрифосфата /нуклеозидтрифосфатов/.
На фиг.12 показаны результаты визуализации геля, полученного по примеру 5.
Треки 1 и 2 соответствуют ДНК нормальной гомозиготы /МM/, треки 3 и 4 соответствуют ДНК гетерозиготы /MS/, а треки 5 и 6 соответствуют ДНК вариантной гомозиготы /SS/. Для треков 1,3 и 5 был использован олигонуклеотид XXII /нормальный/, а для треков 2, 4 и 6 был использован олигонуклеотид XXI /вариантный/. Ожидаемое удлинение олигонуклеотида не наблюдается для треков 2 и 5. Обнаруженные полосы продукта обозначены стрелками.
На фиг.13 показаны результаты визуализации геля, полученного по примеру 6.
На крайнем левом треке представлены маркеры размеров, использованные для подтверждения размеров полученных продуктов. Треки 1 и 2 соответствуют ДНК из нормальной гомозиготы /ММ/, треки 3 и 4 представляют собой ДНК из гетерозиготы /MZ/, а треки 5 и 6 соответствуют ДНК из вариантной гомозиготы /ZZ/. Для треков 1,3 и 5 использовали праймеры XIX и XI; для треков 2, 4 и 6 использовали праймеры XII и XX. Как и ожидалось, удлинение праймеров не наблюдалось для треков 2 или 5. Обнаруженные полосы продукта отмечены стрелками.
Последовательность локуса S a1-антитрипсина /эксон III/.
Представленные на фиг.14-16 основания отражают нормальную последовательность каждого локуса. Подчеркнутые основания в праймерах показывают непарности, введенные для дестабилизации праймеров. Координаты указаны в соответствии с Long et al. /1984/, Biochemistry, 23; 4828-4837.
Последовательность локуса Z a1-антитрипсина /эксон V/.
Показанные на фиг.17 основания соответствуют нормальной последовательности каждого локуса. Подчеркнутые основания в праймерах соответствуют непарностям, введенным для дестабилизации праймеров.
Изобретение иллюстрируется, но не ограничивается, представленными ниже примерами. За исключением специально указанных случаев в примерах указаны следующие количества и концентрации соединений:
Cубстратная ДНК: 1 мкг человеческой геномной ДНК.
Олигонуклеотиды: 100 пмоль каждого соответствующего нуклеотида.
Дезоксинуклеозидтрифосфаты: результирующая концентрация каждого 1,5 мМ.
Буфер /результирующие концентрации в реакционной смеси/:
67 мМ Трис /pH доводят до 8,8 с помощью HCl/.
16,6 мМ Cульфата аммония.
6,7 мМ Хлорида магния.
10 мМ a-Меркаптоэтанола.
6,7 мкМ ЭДТУК.
Загрузочный буфер:
35% Ficoll 70 /Ficoll 70 представляет собой синтетический полимер, полученный сополимеризацией сахарозы и эпихлоргидрина, имеющий средний молекулярный вес около 70000, производства Pharmacia/ 200 мМ Трис-ацетата.
100 мМ Ацетата натрия.
5 мМ ЭДТУК.
0,2% Бромфенолового голубого.
Маркеры размеров: бактериофаг Х174, ДНК которого расщеплена Нае III; полосы соответствуют следующим количествам пар оснований 1353, 1078, 872, 603, 310, 281, 271, 234, 194, 118 и 72.
Пример 1. 1 мкг Человеческой геномной ДНК, 100 пикомоль каждого из вышеописанных олигонуклеотидов I, II, III и IV, по 1,5 мМ /результирующая концентрация/ каждого из четырех дезоксинуклеозидтрифосфатов и буфер в указанных выше результирующих концентрациях смешивают в 1,5 мл микроцентрифужной завинчивающейся пробирке и объем доводят до 100 мкл стерильной дистиллированной водой. Пробирку закрывают и помещают в баню с кипящем водой на 5 мин. Реакцию инициируют, добавляя 1 мкл раствора Tag-полимеразы, содержащей 1 ед. фермента /партия 3 Anglian Biotech, разбавленная до 1 ед/мкл вышеуказанным буфером/. Пробирку инкубируют при 58oC 4 мин и затем 2 мин при 91oC. Режим нагрева/охлаждения 58oC/91oC повторяют затем еще в 6 циклах и затем прибавляют еще 1 ед. фермента /как описано выше/, затем еще 5 циклов с прибавлением дополнительной 1 ед. фермента /как описано выше/ и затем еще 2 цикла с последующим прибавлением еще 1 ед. фермента /как описано выше/. После этого инкубируют при 58oC 20 мин.
Определение продуктов амплификации проводят, соединяя 15 мкл реакционной смеси с 5 мкл загружающего гелевого буфера, и проводят электрофорез на агарозном геле /3% "Nu Sieve"/, содержащем этидий бромид /2 мкл/мл/. Электрофорез проводят вместе с маркерами размеров для подтверждения правильного размера продуктов амплификации. Гель визуализируют в проходящем свете /длина волны 300 нм/ и делают поляроидную фотографию. Трек 3 на фиг.7 является результатом этой визуализации. Из трека 3 видна амплификация эксона III праймерами I и II, как указано выше, с получением продукта длиной 360 пар оснований и амплификация эксона V праймерами III и IV, как описано выше, с получением продукта длиной 220 пар оснований.
Пример 2. Воспроизводят пример 1 за исключением того, что праймер II заменяют на праймер IIa. Трек 4 на фиг. 7 является результатом визуализации геля, полученного по данному примеру. Как можно видеть, амплификация эксона III не протекает при использовании вышеописанных праймеров I и IIa, но протекает амплификация эксона V с праймерами III и IV, что дает продукт длиной 220 пар оснований, как в примере I.
Таким образом пример 2 показывает, что непарность в 3'-окончании олигонуклеотидного праймера по крайней мере в основном подавляет инициирование полимеразной активности.
Пример 3. S-Аллель человеческого a1-антитрипсина.
1 мкг Человеческой геномной ДНК, по 100 пикомоль каждого из описываемых ниже олигонуклеотидов, по 1,5 мМ /результирующая концентрация/ каждого из четырех дезоксинуклеозидтрифосфатов и буфер в описанной выше концентрации смешивают в 1,5 мл завинчивающей микроцентрифуражной пробирке и объем доводят до 100 мкл стерильной дистиллированной водой. Пробирку завинчивают и помещают в баню с кипящей водой на 5 мин. Реакцию инициируют, добавляя 1 мкл раствора Tag-полимеразы, содержащего 1 ед. фермента /партия 3 Agnlian Biotech, разбавленная до 1 ед/мкл вышеуказанным буфером/. Пробирку инкубируют 4 мин при 58oC и затем 2 мин при 91oC. Цикл нагрев-охлаждение 58oC/91oC повторяют 5 раз и затем прибавляют еще 1 ед. фермента /как описано выше/. Вышеуказанный цикл нагрев/охлаждение повторяют еще 6 раз и прибавляют еще 1 ед. фермента, как описано выше. Затем этот цикл повторяют еще 6 раз и после этого прибавляют 1 дополнительную единицу фермента, затем проводят еще 5 циклов и прибавляют еще 1 ед. фермента, после этого еще 2 цикла и затем прибавляют еще 1 ед. фермента так, как описано выше. После этого проводят инкубирование в течение 20 мин при 58oC.
Проводят следующие испытания.
а/ Используют олигонуклеотид
5'TGGTGATGATATCGTGGGTGAGTTCAATTTT V
и описанный выше олигонуклеотид 1. Используют человеческую геномную ДНК из нормальной гомозиготы, не подвергшейся генетическому нарушению человеческого a1-антитрипсина;
b/ используют следующий олигонуклеотид:
TGGTGATGATATCGTGGGTGAGTTCATTTA VI
и вышеописанный олигонуклеотид I. Используют человеческую геномную ДНК из нормальной гомозиготы, не имеющей генетического нарушения человеческого b1-антитрипсина.
Для определения амплифицированных продуктов соединяют 15 мкл реакционной смеси с 5 мкл загрузочного гелевого буфера и проводят электрофорез на агарозном геле /3% "Nu Sieve"/, содержащем этидий бромид /2 мкг/мл/. Электрофорез проводят с маркерами размеров /трек 9 на фиг.8/ для подтверждения правильного размера продуктов амплификации. Проводят визуализацию геля в проходящем свете /длина волны 300 нм/ и делают поляроидную фотографию. Треки 1 и 2 на фиг.8 являются результатом этой визуализации. Как видно по треку 1, амплификация протекает при использовании нуклеотидной последовательности формулы V с 3'-концевым нуклеотидом, комплементарным соответствующему нуклеотиду в возможной точке мутации S в образце ДНК, при этом образуется продукт амплификации длиной 267 пар оснований. Трек 2 показывает, однако, что амплификация не протекает, если непарность имеется в 3'-окончании диагностического праймера, где используемый диагностический праймер имеет нуклеотид /А/ в 3'-окончании, непарный по отношению к нуклеотиду /А/ в потенциальной точке мутации S образца ДНК из нормальной гомозиготы.
Пример 4. Z-Аллель.
1 мкг Человеческой геномной ДНК, по 100 пикомоль каждого из указываемых ниже олигонуклеотидов, 1,5 мМ /результирующая концентрация/ каждого из четырех дезоксинуклеозидтрифосфатов и буфер в указанной выше результирующей концентрации смешивают в 1,5 мл завинчивающейся микроцентрифужной пробирке и объем доводят до 100 мкл стерильной дистиллированной водой. Пробирку закрывают и помещают в баню с кипящей водой на 5 мин. Реакцию инициируют, прибавляя 1 мкл Tag-полимеразы, содержащей 0,5 ед фермента /партия 9 Anglian Biotech, разбавленная до 0,5 ед/мкл вышеуказанным буфером/. Пробирку инкубируют 4 мин при 60oC и затем 2 мин при 91oC. Цикл нагревание/охлаждение 60oC/91oC проводят еще 5 раз, и затем прибавляют еще 0,5 ед. фермента /как описано выше/. Затем цикл нагревание/охлаждение 60oC/91oC проводят еще 6 раз, прибавляя затем еще 0,5 ед. фермента /как описано выше/. Вышеуказанный цикл нагрев/охлаждение повторяют еще 6 раз и затем прибавляют еще 0,5 ед. фермента /как описано выше/, затем еще 5 раз с последующим прибавлением 0,5 ед. фермента /как описано выше/ и еще 3 раза с последующим дополнительным прибавлением 0,5 ед. фермента /как описано выше/. Затем проводят инкубирование в течение 20 мин при 60oC.
На основе вышеописанной методики проводят следующие испытания.
а/ Используют вышеописанные олигонуклеотиды IX и IV и человеческую геномную ДНК из нормальной гомозиготы, не имеющей генетического нарушения человеческого a1-антитрипсина.
b/ Используют вышеописанные олигонуклеотиды X и IV и человеческую геномную ДНК из нормальной гомозиготы, не имеющей генетического нарушения человеческого a1-антитрипсина.
с/ Используют вышеуказанные олигонуклеотиды IX и IV и человеческую геномную ДНК из гетерозиготы Z-аллели человеческого a1-антитрипсина.
d/ Используют вышеуказанные олигонуклеотиды X и IV и человеческую геномную ДНК из гетерозиготы Z-аллели человеческого a1-антитрипсина
е/ Используют вышеописанные олигонуклеотиды IX и IV и человеческую геномную ДНК из гомозиготы /ZZ/, имеющей нарушение a1-антитрипсина.
f/ Используют вышеописанные олигонуклеотиды X и IV и человеческую геномную ДНК из гомозиготы /ZZ/, имеющей нарушение a1-антитрипсина.
g/ Используют вышеописанные олигонуклеотиды IX и IV и человеческую геномную ДНК из нормальной гомозиготы, не имеющей генетического нарушения человеческого a1-антитрипсина.
h/ Используют вышеописанные олигонуклеотиды XII и IV и человеческую геномную ДНК из нормальной гомозиготы, не имеющей генетического нарушения человеческого a1-антитрипсина.
i/ Используют вышеописанные олигонуклеотиды IX и IV и человеческую геномную ДНК из гетерозиготы Z-аллели человеческого a1-антитрипсина.
j/ Используют вышеописанные олигонуклеотиды XII и IV и человеческую геномную ДНК из гетерозиготы для Z-аллели человеческого a1-антитрипсина.
k/ Используют вышеописанные олигонуклеотиды IX и IV и человеческую геномную ДНК из гомозиготы /ZZ/ с нарушением a1-антитрипсина.
l/ Используют вышеописанные олигонуклеотиды XII и IV и человеческую геномную ДНК из гомозиготы /ZZ/, с нарушением a1- антитрипсина.
В каждом испытании нуклеотидные последовательности XIII и XIV используют для контроля амплификации.
Определение продуктов амплификации проводят, соединяя 15 мкл реакционной смеси с 5 мкл загрузочного гелевого буфера и проводя электрофорез на 1,4%-ном агарозной геле, содержащем этидий бромид /0,5 мкг/мл/. Электрофорез проводят с маркерами размеров, описанными выше, для подтверждения правильного размера продуктов амплификации. Проводят визуализацию геля в проходящем свете /длина волны 300 нм/ и делают поляроидную фотографию. Треки 2-13 на фиг. 9 показывают результат этой визуализации, а треки 1 и 14 являются полосами маркеров размера. Как видно из треков 2-7, дестабилизация диагностического праймера изменением пятого от 3'-окончания нуклеотида не совсем достаточна для того, чтобы праймер различал образцы нормальной и мутантной ДНК в этих реакционных условиях. Тем не менее треки 8-13 показывают, что изменение третьего от 3'-окончания нуклеотида в диагностическом праймере позволяет праймеру эффективно различать образцы нормальной и мутантной ДНК и быть диагностическим праймером для нормальных гомозигот, гетерозигот /носителей/ или ZZ-гомозигот, имеющих нарушение человеческого a1- антитрипсина.
Вышеописанные эксперименты по данному примеру проводят также с использованием диагностического праймера, в котором не проводят дополнительных дестабилизирующих изменений нуклеотидов и в котором седьмой нуклеотид от 3'-окончания изменен /А на С/ для достижения дестабилизации. В каждом случае диагностический праймер недостаточно эффективен для того, чтобы различать нормальные и мутантные образцы ДНК и этот факт проиллюстрирован в треках 5-8 на фиг. 8 для диагностического праймера, не имеющего дополнительных дестабилизирующих нарушений нуклеотидов.
Пример 5. В качестве праймеров используют олигонуклеотиды XXII /нормальный/ и XXI /вариантный/. В этом примере смесь дезоксинуклеозидтрифосфатов A, G и Т дает удлинение праймера на 7 оснований на матрице без необходимости введения нуклеотидтрифосфата С. Tm для этих олигонуклеотидов, рассчитанная по формуле Tm 4/G + C/ и 2/А+Т/, равна 94oC, но это считается верным только для олигонуклеотидов до 23-меров, поэтому выбирают предпочтительную температуру 75oC.
Олигонуклеотиды /по 8 пикомоль каждого/ метят по 5'-концевой окси-группе с помощью d32P ATP /Amersham 2 мкК/ и прибавляют Т4-полинуклеотидкиназу /4 ед./ в 80 мкл реакционного объема, содержащего 5 мМ Трис-HCl с pH 7,6, 10 мМ MgCl2, 5 мМ DTT, 100 мкМ спермидин и 100 мкМ ЭДТУК. Киназную реакцию проводят 20 мин при 37oC и затем меченые олигонуклеотиды проверяют электрофорезом на 15% -ном полиакриламидном денатурирующем геле с 8М мочевиной /предварительный режим 1 ч при 500 В, основной режим 4 ч при 800 В/.
Используют две плазмиды, одна из которых содержит нормальную последовательность /ММ/ S-локуса эксона III гена человеческого a1- антитрипсина, а другая содержит вариантную последовательность, соответствующую вариантной гомозиготе /SS/. Подготавливают 6 пробирок с 2 возможными типами ДНК. Для гомозигот используют 1 фмоль соответствующей плазмиды, а гетерозиготы стимулируют использованием 0,5 фмоль каждой из плазмид. 200-Кратный избыток меченого олигонуклеотида создают в каждой пробирке, прибавляя 200 фмоль /2 мкл меченого раствора/ нормального /XXII/ или вариантного /XXI/ олигонуклеотида к каждому типу ДНК. Каждая пробирка содержит A, G и T /Pharmacia/ в результирующей концентрации 5 мМ каждый в реакционном объеме 20 мкл, содержащем 6,7 мМ ЭДТУК, 6,7 мМ MgCl2, 67 мМ Трис-HCl c pH 8,8, 10 мМ меркаптоэтанола и 16,6 мМ сульфата аммония. Реакционный объем утраивают, т.е. в каждом случае используют 60 мкл для уменьшения эффекта упаривания. Пробирки кипятят в течение 5 мин и затем хранят на льду, после чего в каждую пробирку прибавляют по 3 ед. Tag-полимеразы /Cetus I, разбавление в 5 раз с помощью 1,5 мМ MgCl2, 50 mM KCl, 10 mM Трис с pH 8,3 и 0,01% желатины с получением 1 ед. /мкл/. Пробирки центрифугируют при 13000 об/мин в течение 2 мин и аликвоты по 2 мкл прибавляют к 5 мкл смеси формамида с красителем бромфеноловым голубым. Затем пробирки оставляют в водяной бане при 75oC. Через 4 ч пробирки вынимают, центрифугируют 1 мин при 13000 об/мин и отбирают еще аликвоту объемом 2 мкл и прибавляют ее к 5 мкл красителя. Затем пробирки возвращают в водяную баню с температурой 75oC. Через 6 ч пробирки снова вынимают, в каждую пробирку прибавляют еще по 3 ед. Tag-полимеразы, центрифугируют 1 мин при 13000 об/мин и прибавляют 1 каплю легкого минерального масла /Sigma/ для уменьшения испарения, но не отбирают аликвоту. Затем пробирки выдерживают в течение ночи при 75oC. Через 24 ч от конца первого центрифугирования пробирки вынимают из водяной бани, центрифугируют 1 мин и последние аликвоты объемом по 2 мкл прибавляют к 5 мкл красителя. Все аликвоты подвергают электрофорезу на предобработанном /1 ч при 500 В/ 15%-ном денатурирующем полиакриламидном геле с 8М мочевиной в 1 х ТВЕ буфере /0,089М Трис-борат, 0,089 борная кислота, 0,002М ЭДТУК/ в течение 5,5 ч при 880 В. Затем гели оценивают авторадиографией в течение ночи при комнатной температуре с помощью фотографической малочувствительной пленки без диафрагмы. Результаты представлены на фиг. 12.
Треки 1 и 2 соответствуют ДНК для нормальной гомозиготы /ММ/, треки 3 и 4 ДНК для гетерозиготы /MS/, а треки 5 и 6 ДНК для вариантной гомозиготы /SS/. Для треков 1, 3 и 5 был использован олигонуклеотид XXII /нормальный/, а для треков 2,4 и 6 использован олигонуклеотид XXI /вариантный/. Как и ожидалось, удлинение олигонуклеотидов не наблюдается в треках 2 или 5. Обнаруживаемые полосы продукта обозначены стрелками.
Пример 6. 1 нг Кассетной плазмидной ДНК, 100 пикомоль каждого из следующих олигонуклеотидных праймеров:
TTACTTTCACCAGCGTTTCTGGGTGAGCAA и
TATGCGACTCCCTGCATTAGGAGCCCA,
1,5 мМ /результирующая концентрация/ каждого из четырех дезоксинуклеозидтрифосфатов и буфер с указанной выше результирующей концентрацией смешивают в 1,5 мл завинчивающейся микроцентрифужной пробирке и объем доводят до 100 мкл стерильной деионизированной водой /Milli-Q/. Пробирки закрывают и помещают в баню с кипящей водой на 5 мин. Реакцию инициируют прибавлением 2 ед. Tag-полимеразы /Cetus/ и смесь закрывают легким минеральным маслом /Sigma/ для предотвращения испарения. Пробирку инкубируют при 60oC в течение 4 мин и затем 2 мин при 90oC. Эту операцию циклически повторяют 30 раз /общее число циклов/.
Затем отбирают по 60 мкл реакционной смеси и прибавляют 60 пикомоль либо праймеров XXI и XI /нормальные/, либо праймеров XII и XX /вариантные/. Затем для инициирования дальнейшей реакции прибавляют 1 ед. Tag-полимеразы /Cetus/. Смесь инкубируют 2 мин при 92oC и 4 мин при 60oC. Эту операцию повторяют в целом 4 раза. Определение продуктов амплификации проводят, соединяя 10 мкл аликвоты реакционной смеси с гелеобразующим буфером и проводя электрофорез на 3% -ном агарозном геле /"Nu Siewe", FMC Bioproduct/, содержащем 0,5 мкг/мл этидий бромида. Гель визуализируют в проходящем свете /длина волны 300 нм/ и делают фотографию на поляроиде. Результаты представлены на фиг.13. Крайний левый трек соответствует маркерам размеров, используемым для подтверждения размеров получаемых продуктов. Треки 1 и 2 соответствуют ДНК из нормальной гомозиготы /ММ/, треки 3 и 4 ДНК из гетерозиготы /MZ/, а треки 5 и 6 ДНК из вариантной гомозиготы /ZZ/. Для треков 1,3 и 5 использовали праймеры XIX и XI; для треков 2,4 и 6 использовали праймеры XII и XX. Как и ожидалось, удлинение праймеров не наблюдается в треках 2 или 5. Обнаруженные полосы продукта помечены стрелками.
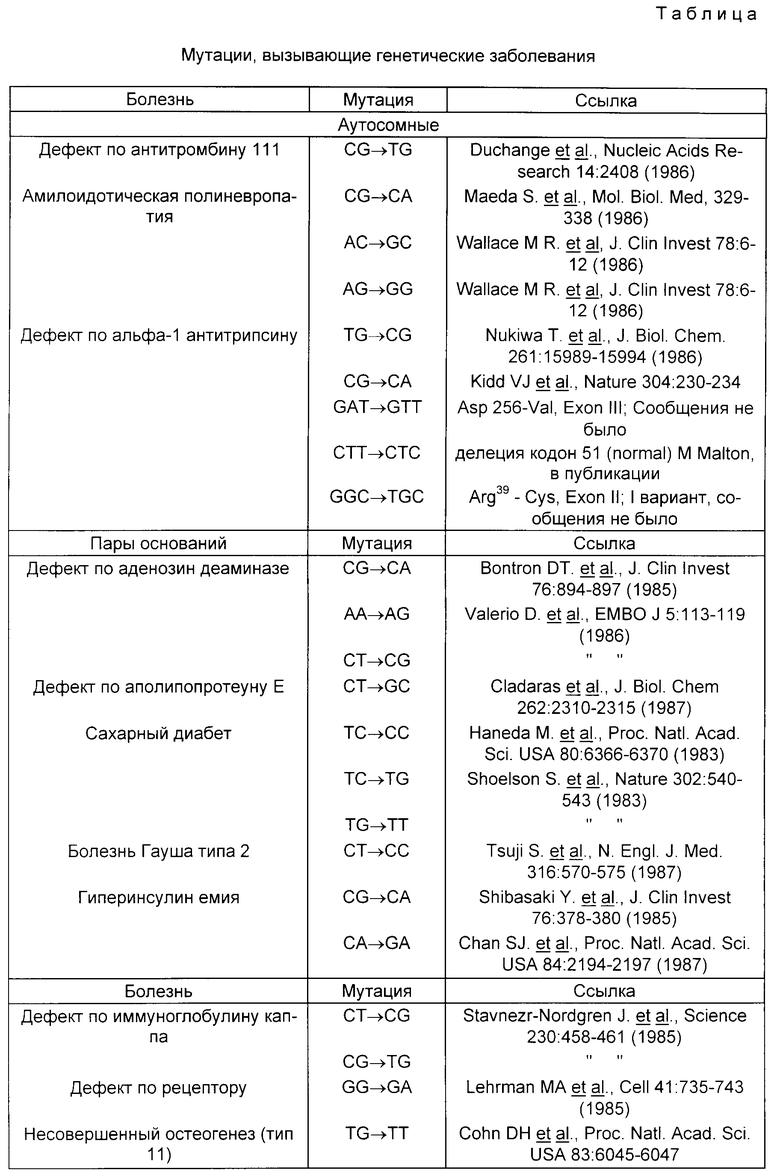
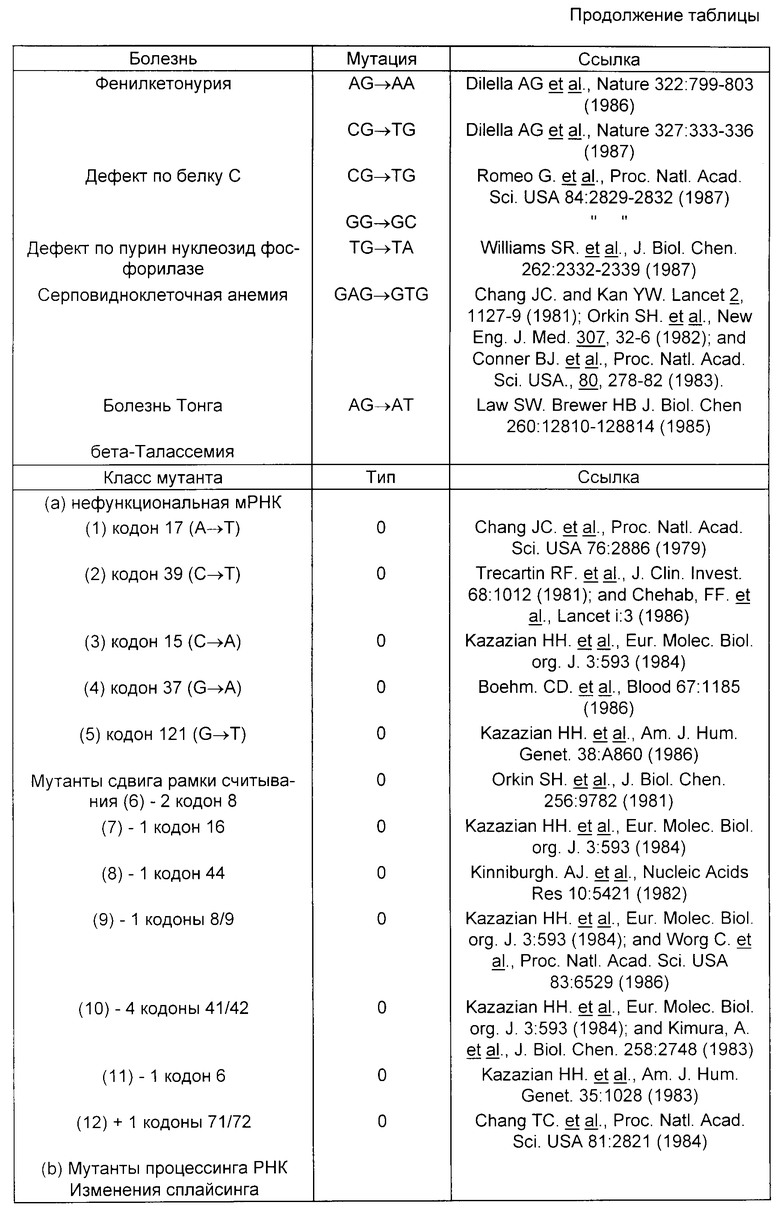

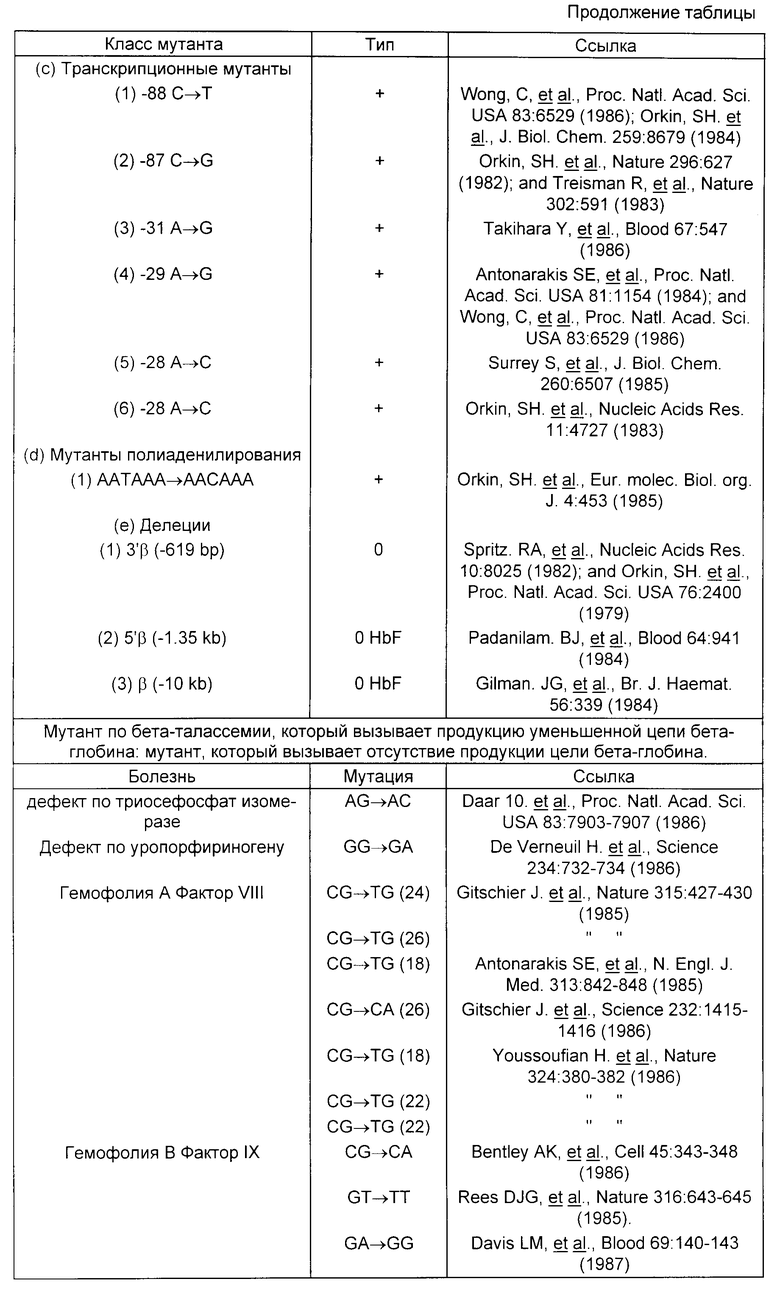
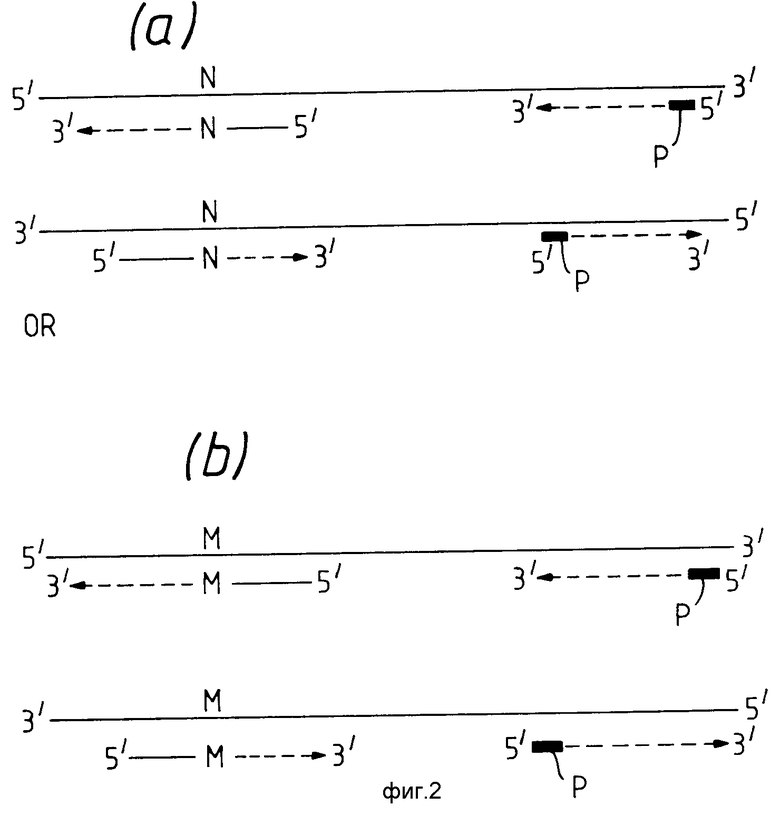
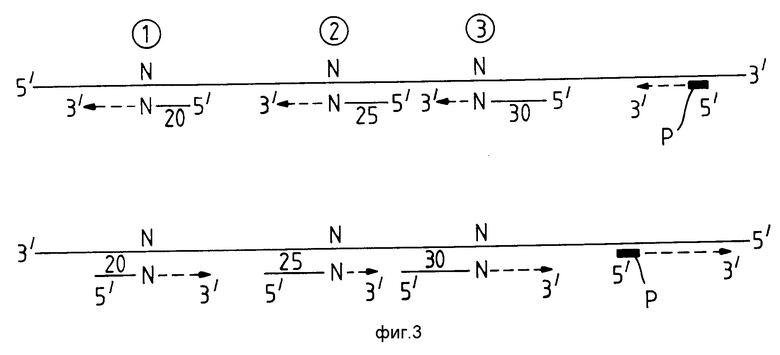
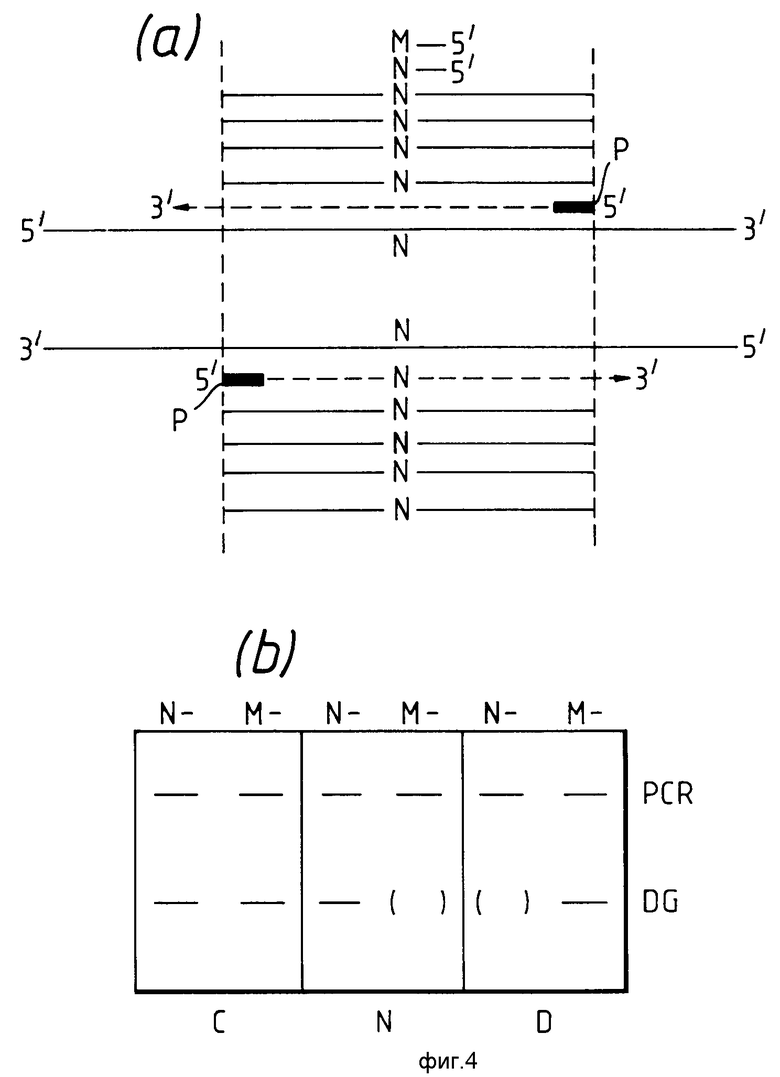
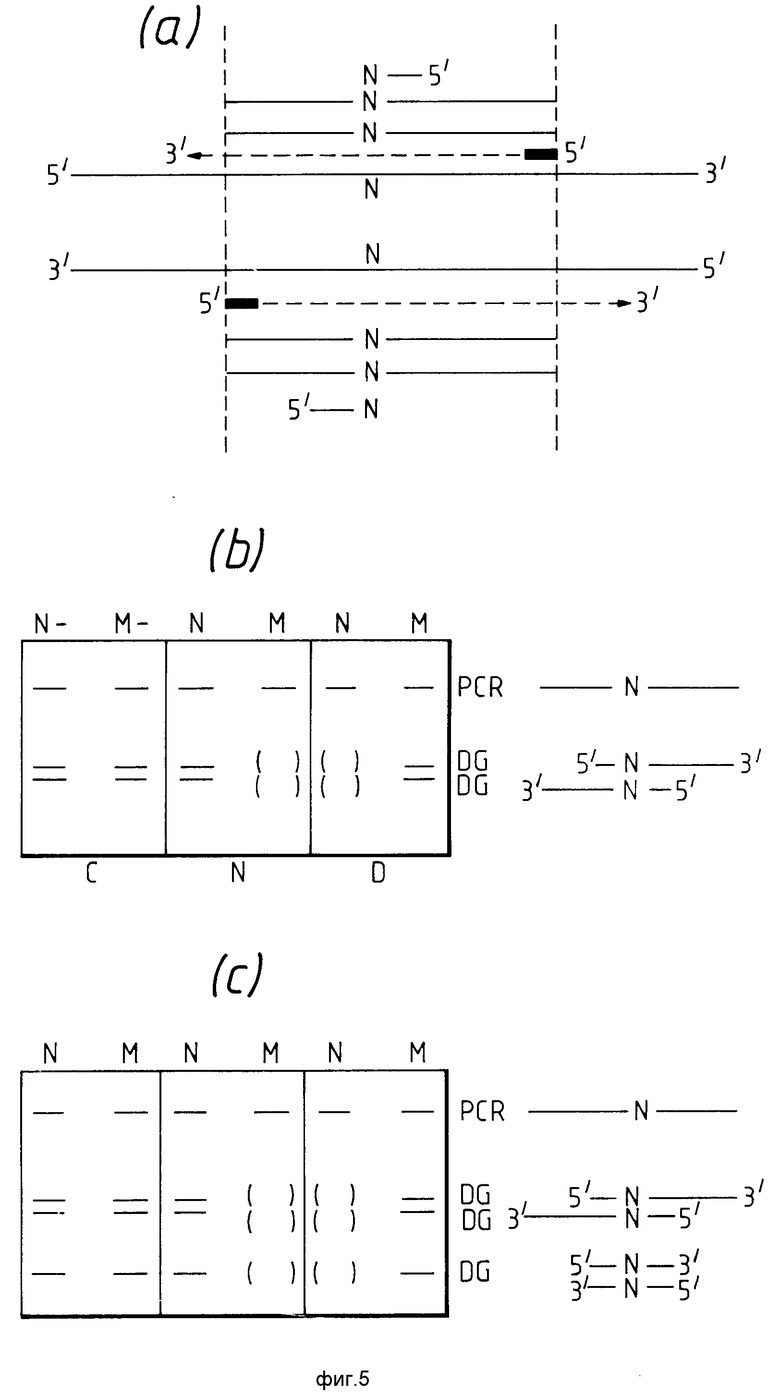
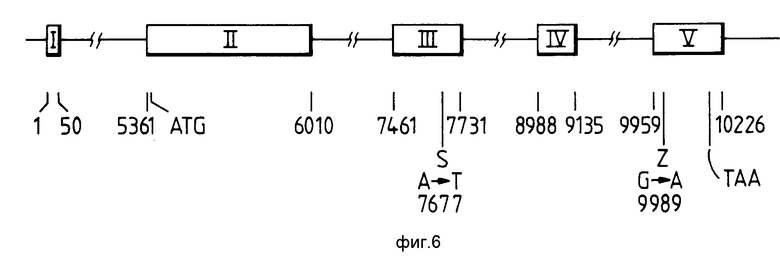
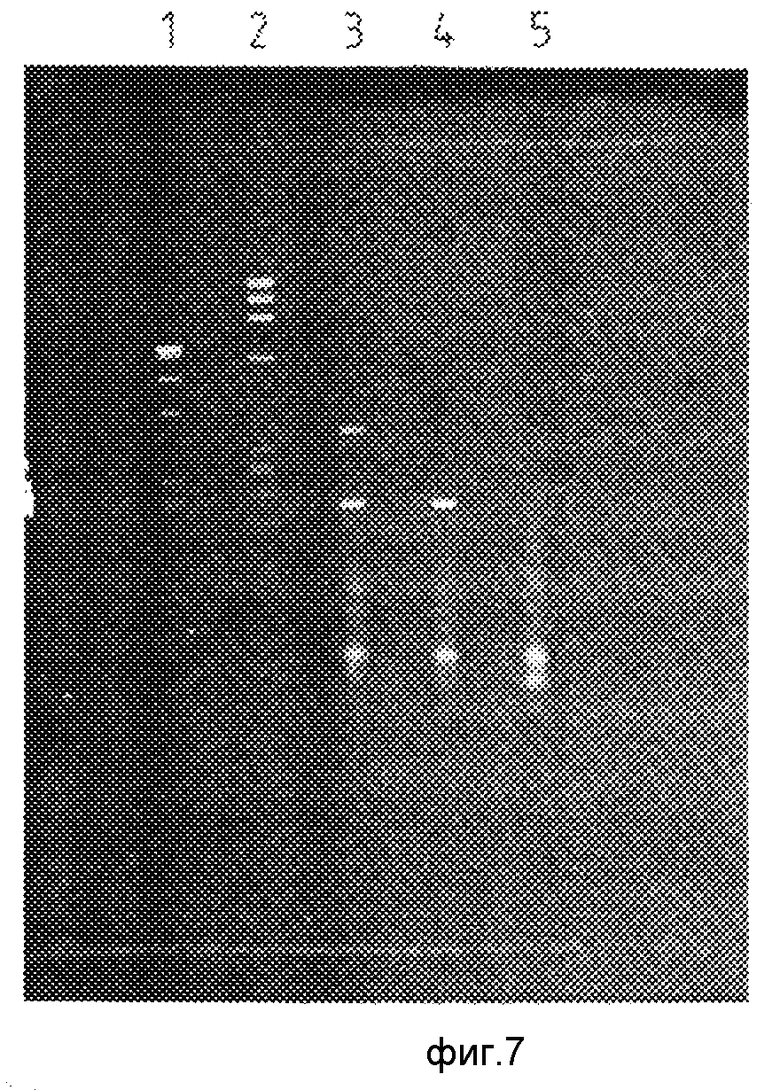

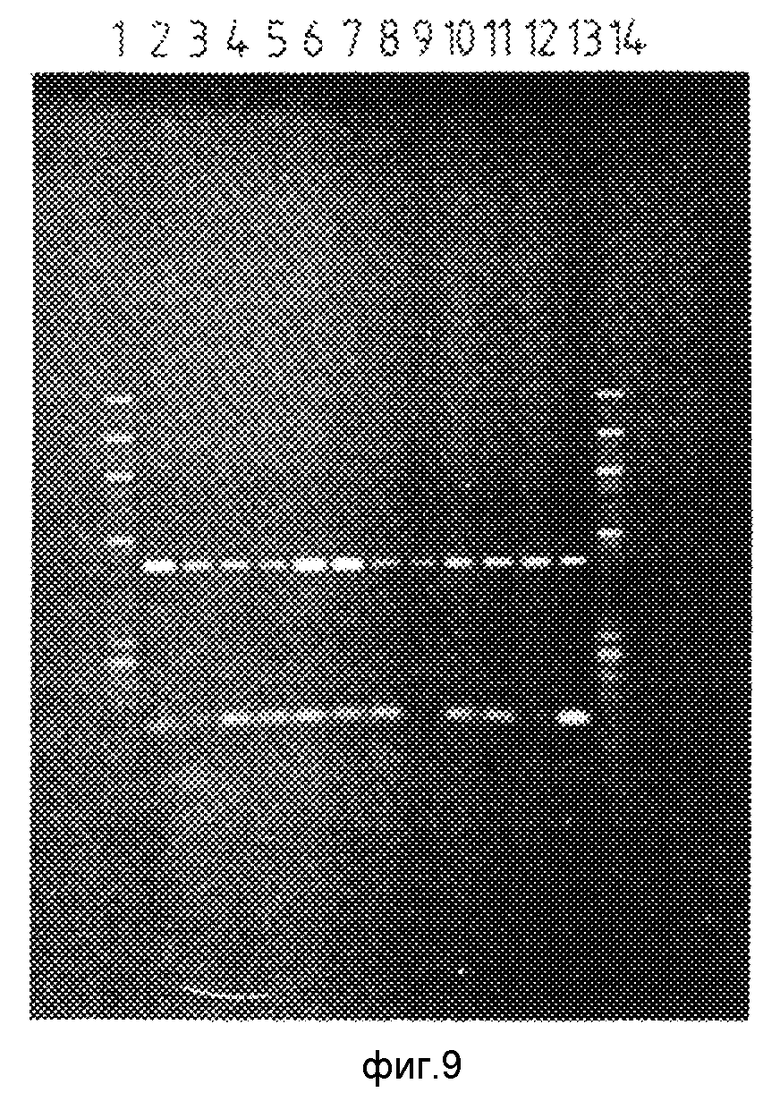
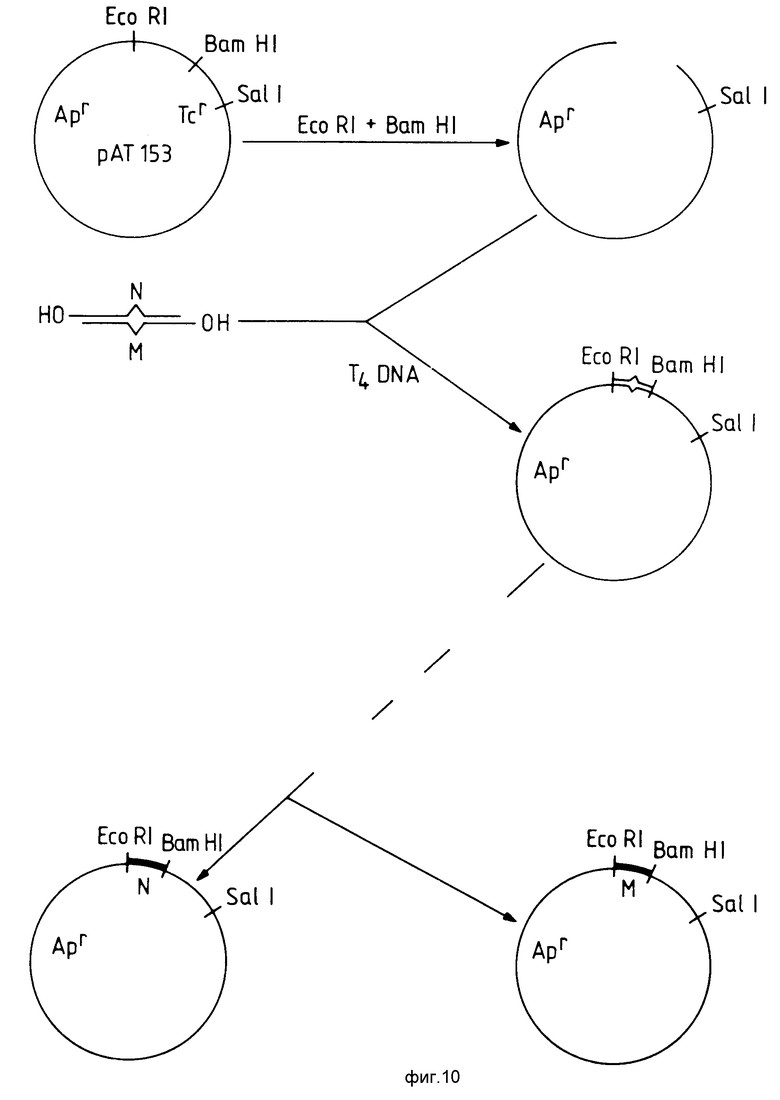
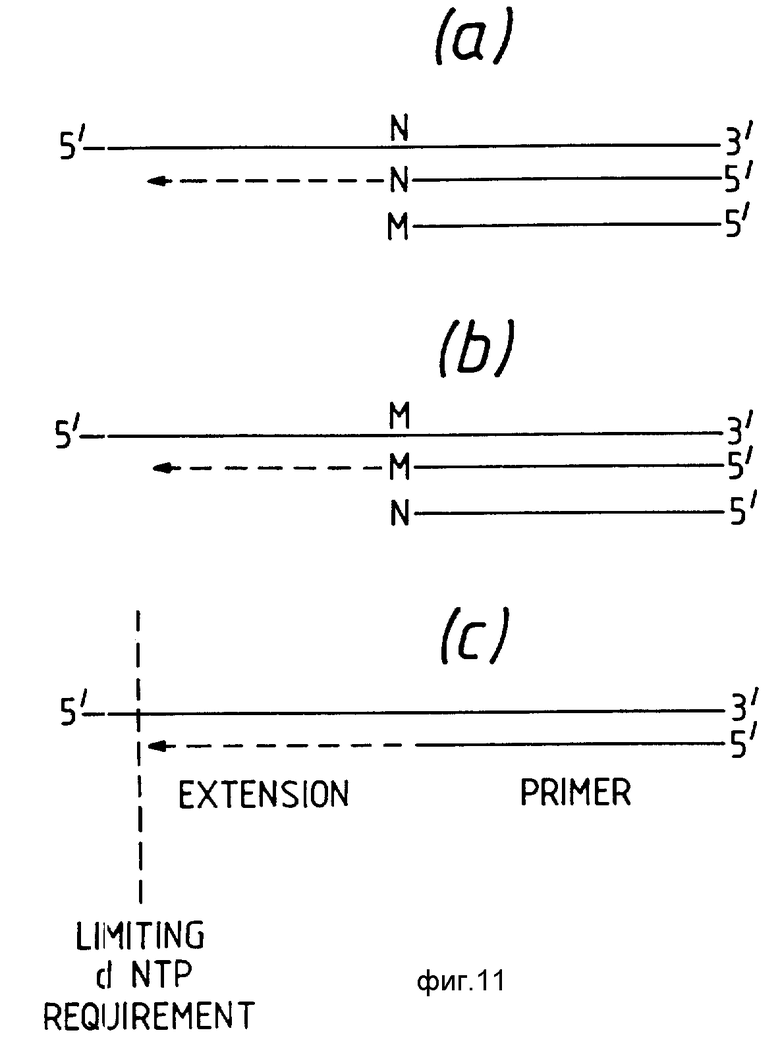
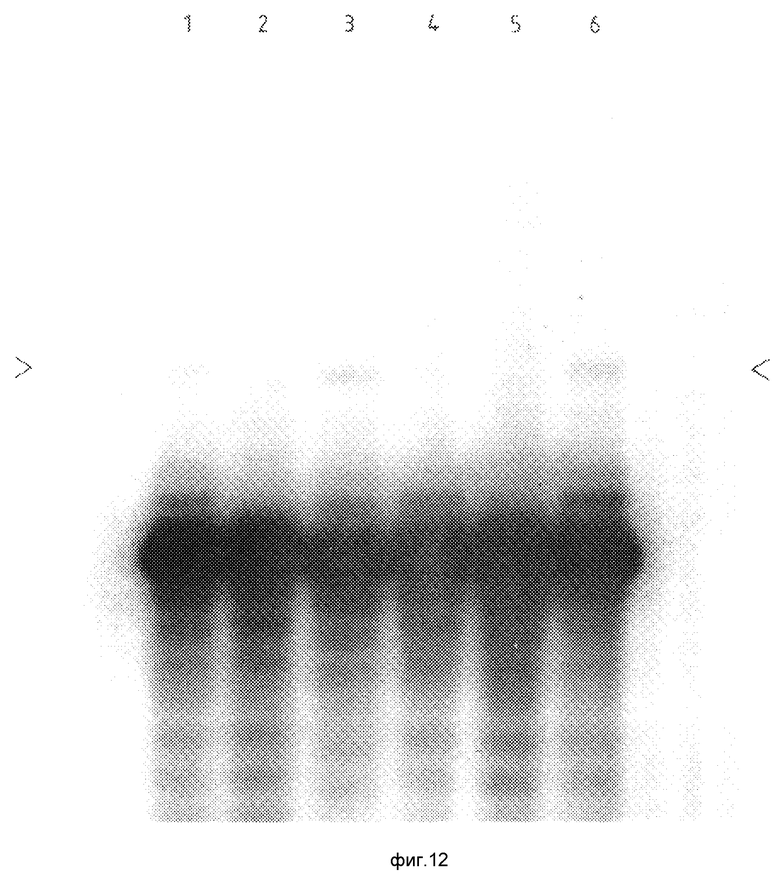
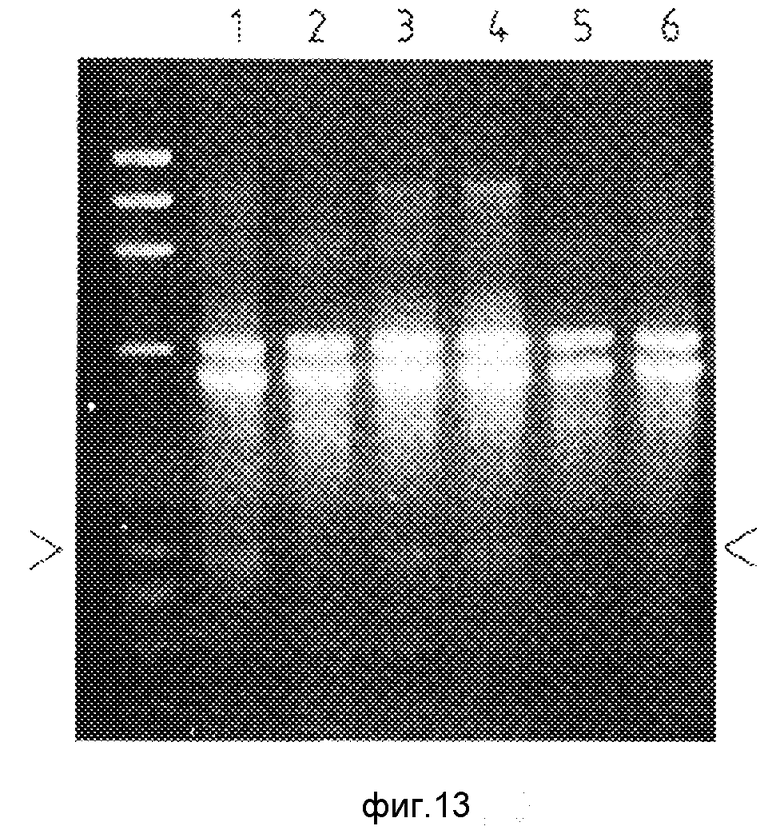




Способ для определения наличия или отсутствия одной или нескольких последовательностей нуклеотидов. Способ включает контактирование образца нуклеиновой кислоты с диагностическим праймером, который является существенно комплементарным в отношении диагностической части последовательности основания мишени, посредством чего достигается только распространение диагностического праймера на матрицу мишени при соответствующих условиях, если терминальный нуклеотид диагностического праймера комплементарен либо в отношении к предполагаемому вариантному нуклеотиду, либо к соответствующему нормальному нуклеотиду последовательности основания мишени. 1 табл., 17 ил.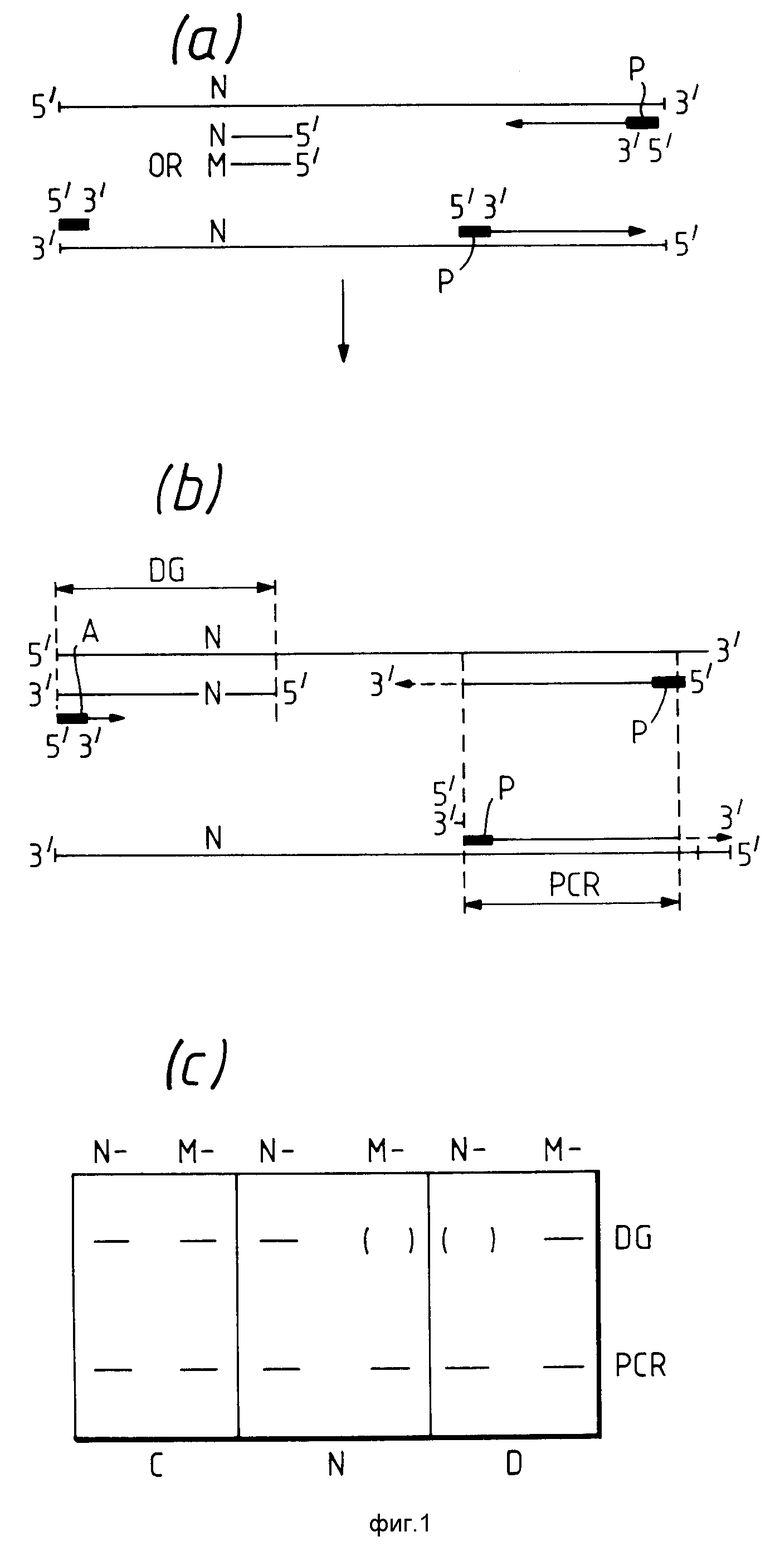
Способ обнаружения единичных нуклеотидных изменений в нуклеиновых кислотах образца, предусматривающий амплификацию нуклиновых кислот образца путем проведения реакции удлинения цепи с использованием праймеров, нуклеозидтрифосфатов и фермента полимеризации с последующим выявлением изменений, отличающийся тем, что по крайней мере один из праймеров является диагностическим, имеющим на 5' или 3' конце терминальный нуклеотид, комплементарный измененному нуклеотиду или соответствующему нормальному нуклеотиду, а изменение выявляют по наличию продуктов амплификации, если терминальный нуклеотид праймера комплементарен к соответствующему нуклеотиду в образце нуклеиновой кислоты, или по отсутствию их, если терминальный нуклеотид праймера не комплементарен соответствующему нуклеотиду в образце нуклеиновой кислоты.
| EP, патент N 0237362, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
