ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к способу получения характеристик молекул нуклеиновой кислоты, который включает создание удлиняемых направленных вверх фрагментов, являющихся следствием расщепления нуклеиновой кислоты в лишенном основания звене.
УРОВЕНЬ ТЕХНИКИ
Получение характеристик целевых нуклеиновых кислот (нуклеиновых кислот - мишеней) является чрезвычайно важной задачей по нескольким причинам, связанным с подтверждением наличия или отсутствия гена в образце, подтверждением части или всей нуклеотидной последовательности и проверкой присутствия известных и неизвестных вызывающих заболевания мутаций, приводящих к наследуемой болезни, а также наличия в ДНК природных вариаций. Несмотря на то, что имеется много известных способов получения характеристик нуклеиновых кислот и обнаружения неизвестных изменений нуклеотидных последовательностей, возрастающее количество появляющейся новой генетической информации обусловливает необходимость разработки новых, лучших и более быстрых способов получения характеристик нуклеиновых кислот.
Заявка WO 97/03210 раскрывает применение фермента ДНК-гликозилазы, различающей модифицированное основание, для прямого обнаружения известных и неизвестных мутаций в образце целевой нуклеиновой кислоты. Как правило, способ включает амплификацию образца целевой нуклеиновой кислоты с использованием комбинации нормальных нуклеотидов-предшественников ДНК и одного или более модифицированных нуклеотидов-предшественников, причем модифицированный нуклеотид-предшественник замещает один из нормальных нуклеотидов-предшественников, что является субстратом для ДНК-гликозилазы. После удаления гликозилазой модифицированного основания производят расщепление в полученном лишенном основания звене и анализируют продукты расщепления. Этот способ позволяет обнаруживать мутации в предполагаемых локусах. Однако способ заявки WO 97/03210 имеет определенные ограничения. Например, с помощью этого способа невозможно обнаружить различия молекул нуклеиновой кислоты в нуклеотидных последовательностях без регистрации подобия последовательностей, и поэтому невозможно провести одновременный анализ нескольких образцов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение предлагает способ получения характеристик молекул нуклеиновой кислоты, включающий следующие этапы:
i) введение в молекулу ДНК модифицированного основания, которое является субстратом для ДНК-гликозилазы;
ii) удаление модифицированного основания с помощью указанной ДНК-гликозилазы для создания лишенного основания звена;
iii) расщепление ДНК в лишенном основания звене таким образом, чтобы создать направленный вверх (в направлении от 5'-конца к 3'-концу) фрагмент ДНК, который можно удлинить; и
iv) инкубация способного к удлинению (удлиняемого) направленного вверх фрагмента в присутствии фермента, обеспечивающего его удлинение, и матричной нуклеиновой кислоты и анализ полученного фрагмента (полученных фрагментов).
Изобретение предоставляет новый, гибкий и простой способ, использующий указанные выше удлиняемые направленные вверх фрагменты ДНК, который позволяет характеризовать нуклеиновые кислоты и который имеет преимущества перед существующими способами, как указано в нижеследующем описании.
Одним из наиболее важных применений (но не единственным применением) способа в соответствии с изобретением является сканирование или проверка фрагмента ДНК (целевой нуклеиновой кислоты) на наличие или отсутствие мутации. Неотъемлемыми составными частями способа являются: 1) создание удлиняемых (способных к удлинению) направленных вверх фрагментов ДНК и 2) последующее использование этих фрагментов в анализе образца ДНК (т.е. в обнаружении мутации).
Предпочтительно используемое модифицированное основание представляет собой урацил, гипоксантин или (8-ОН)-гуанин.
Предпочтительно модифицированные основания получены из модифицированных нуклеотидов-предшественников, которые при включении в ДНК генерируют указанные модифицированные основания,
Так, предпочтительными модифицированными нуклеотидами-предшественниками являются дезоксиуридин-трифосфат (dUTP), дезоксинозин-трифосфат (dITP) и (6-ОН)-дезоксигуанозин-трифосфат ((8-OH)-dGTP), после включения которых в ДНК создаются основания - субстраты для гликозилазы: соответственно урацил, гипоксантин и (8-ОН)-гуанин. Каждый из модифицированных нуклеотидов-предшественников представляет собой сахарофосфат основания, содержащий указанное основание и остаток фосфата сахара. Урацил в ДНК специфически распознается урацил-ДНК-гликозилазой (УДГ) и выщепляется из ДНК. УДГ распознает также и другие родственные урацилу основания, если они присутствуют в ДНК. Гипоксантин специфически распознается алкилпурин-ДНК-гликозилазами (АДГ) и выщепляется из ДНК. Этот фермент также распознает и выщепляет N3-метиладенин, N3-метилгуанин, О2-метилцитозин и О2-метилтимин, когда они присутствуют в ДНК. (8-ОН)-гуанин специфически распознается и выщепляется из ДНК формамидопиримидин-ДНК-гликозилазой (ФПГ). Этот фермент также распознает и выщепляет пурины с разомкнутым кольцом, если они присутствуют в ДНК. Тимин-ДНК-гликозилаза распознает и выщепляет урацил и тимин, расположенные в ДНК напротив гуаниновых оснований.
Использованные здесь термины "модифицированный нуклеотид-предшественник" и "модифицированные нуклеотиды-предшественники" обозначают модифицированный нуклеотид или модифицированные нуклеотиды, которые могут быть включены в нуклеиновую кислоту таким образом, что создаются модифицированное основание или модифицированные основания, которые распознаются и могут быть выщеплены из ДНК ферментом ДНК-гликозилазой.
После введения модифицированного основания ДНК-продукт обрабатывают подходящим типом фермента ДНК-гликозилазы, которая распознает и выщепляет основание - субстрат для гликозилазы, присутствующее в образце-мишени, вследствие чего формируется апуриновое (лишенное пурина) или апиримидиновое (лишенное пиримидина) АП-звено (или АП-сайт), в зависимости от природы модифицированного основания (модифицированных оснований). "АП-звено" (или "АП-сайт") является термином, присвоенным звену в ДНК, где остаток основания в нуклеотиде утрачен или удален, с сохранением в фосфодиэфирном скелете ДНК неповрежденного дезоксирибофосфата. АП является аббревиатурой термина "апуриновый" или "апиримидиновый", в зависимости от того, какое основание было присоединено к кольцу рибозы - пурин или пиримидин. "АП-звено" обозначает также лишенное основания звено и является общим термином для апуринового и апиримидинового звена.
Выщепление из образца нуклеиновой кислоты основания - субстрата для гликозилазы приводит, например, в случае урацила к появлению апиримидинового звена, а в случае гипоксантина или (8-ОН)-гуанина - апуринового звена. В общем виде такие звенья называются лишенными основания звеньями.
Необходимо, чтобы полученные расщеплением фрагменты имели на 3'-конце гидроксильные группы, а ДНК в непосредственном соседстве вниз от указанного 3'-конца не была блокирована таким способом, который препятствовал бы удлинению фрагментов от указанных 3'-концов. После расщепления создаются фрагменты, имеющие на 3'-конце гидроксил, и удаляются расположенные ниже блокирующие группы, препятствующие удлинению фрагмента на указанном 3'-конце при использовании в качестве матрицы той ДНК, из которой был получен удлиняемый (способный к удлинению) фрагмент.
ДНК в звене, лишенном основания, можно расщепить разными способами, чтобы создать указанный направленный вверх фрагмент ДНК, как подробнее описано далее.
Например, фосфатные связи в лишенном основания звене можно расщепить обработкой, выбранной среди способов обработки щелочным раствором или другой химической обработки, обработки теплом и/или обработки ферментом.
Согласно одному из вариантов осуществления изобретения направленный вверх фрагмент создается расщеплением ДНК с 5'-стороны лишенного основания звена, так что 3'-конец направленного вверх фрагмента имеет гидроксильную группу.
Термины "удлиняемый фрагмент" ("способный к удлинению фрагмент") и "направленный вверх фрагмент ДНК" здесь используются как взаимозаменяемые.
После того как таким образом создан 3'-ОН конец, далее вплоть до стадии (iv) нет необходимости в дальнейшей обработке (процессинге) направленных вверх фрагментов.
Обработка основными растворами (щелочами) при высокой температуре или химическим реагентом, таким как пиперидин, или ферментом, специфически разрезающим нуклеиновую кислоту в лишенных основания звеньях, таким как эндонуклеаза IV Е.coli, приводит к расщеплению в лишенном основания звене с 5'-стороны. В данном осуществлении подходящее расщепление осуществляется 5'-АП эндонуклеазой.
В соответствии с альтернативными вариантами осуществления изобретения направленный вверх фрагмент создается расщеплением на 5'-стороне лишенного основания звена таким образом, чтобы сохранить фосфатную группу на 3'-конце направленного вверх фрагмента, и удалением фосфатной группы таким образом, что направленный вверх фрагмент имеет на 3'-конце гидроксильную группу.
В качестве альтернативы можно применить расщепление щелочью или использовать специфическую для лишенных основания звеньев эндонуклеазную активность ФПГ, после чего удаляют 3'-фосфат. Например, можно удалить 3'-фосфатные группы ферментативными методами, используя ферменты с активностью по отношению к 3'-фосфатам - как, например, Т4 полинуклеотидкиназу.
В соответствии с другим вариантом осуществления направленный вверх фрагмент создается расщеплением на 3'-стороне лишенного основания звена таким образом, чтобы на 3'-конце направленного вверх фрагмента осталась группа дезоксирибозофосфата, с последующим удалением дезоксирибозной группы, чтобы на 3'-конце осталась гидроксильная группа.
Это можно осуществить с помощью фермента с 3'-дезоксирибофосфодиэстеразной активностью или с помощью ФПГ с последующей обработкой 3'-фосфатазой.
В альтернативном варианте осуществления удаляют расположенные вниз от 3'-конца направленного вверх фрагмента остатки 5'-дезоксирибозы таким образом, чтобы направленный вверх фрагмент можно было удлинить на матрице.
Предпочтительно остатки 5'-дезоксирибозы удаляют 5'-дезоксирибофосфодиэстеразой.
Обработка только высокой температурой или 3'-АП-эндонуклеазой приводит к полному расщеплению на 3'-стороне лишенного основания звена.
Расщепление с участием гликозилазы после отщепления модифицрованного основания ДНК-гликозилазой приводит к разрезанию удлиненного праймера в лишенном основания звене с получением 3'-концов с 3'-ОН или 3'-фосфатной группами или с дезоксирибофосфатными группами. За исключением случая, где образуются концы 3'-ОН, при образовании всех других концов перед удлинением направленного вверх фрагмента необходима дополнительная обработка.
Расщеплением с участием гликозилазы по способу в соответствии с данным изобретением называется расщепление как в 5'-, так и в 3'-положении, включая любые последующие обработки, необходимые для образования на 3'-конце направленного вверх фрагмента 3'-ОН группы.
Предпочтительно модифицированное основание вводится путем ферментативной амплификации ДНК.
Предпочтительно ДНК (носящая здесь также название "целевая нуклеиновая кислота") амплифицируется с использованием нормальных нуклеотидов-предшественников ДНК и по крайней мере одного модифицированного нуклеотида-предшественника.
Нуклеотидами-предшественниками в случае процесса амлификации ДНК называют дезоксирибонуклеотиды dATP (дезоксиаденозин-трифосфат), dCTP (дезоксицитидин-трифосфат), dGTP (дезоксигуанозин-трифосфат) и dTTP (дезокситимидин-трифосфат), которые здесь носят название "нормальные" нуклеотиды-предшественники ДНК.
Термин "амплификация" используется здесь для обозначения любого процесса in vitro для увеличения числа копий нуклеотидной последовательности или нуклеотидных последовательностей. Амплификация молекулы целевой нуклеиновой кислоты приводит к включению нуклеотидов-предшественников в амплифицируемую ДНК. Обычно амплификацию целевого образца проводят в полимеразной цепной реакции (ПЦР), используя подходящие праймеры. В ходе амплификации праймеры комплементарно присоединяются к целевой нуклеиновой кислоте и с помощью ДНК-полимеразы удлиняются в направлении 5'→3' на целевой нуклеиновой кислоте, которая служит матрицей для синтеза новой ДНК. Использование фланкирующих праймеров, которые здесь названы первоначальными праймерами, присоединяющихся к верхней и нижней нитям молекулы ДНК, обеспечивает экспоненциальную амплификацию сегмента ДНК, ограниченного верхним и нижним праймерами.
Амплификация обычно включает амплификацию целевой нуклеиновой кислоты с использованием комбинации нормальных нуклеотидов-предшественников ДНК и одного или больше модифицированных нуклеотидов-предшественников, причем модифицированный нуклеотид-предшественник замещает полностью или частично один из нормальных нуклеотидов-предшественников. Амплификация нуклеиновой кислоты с использованием нормальных нуклеотидов-предшественников ДНК приводит к включению в ДНК четырех нормальных оснований: гуанина (G), аденина (А), тимина (Т) или цитозина (С). Амплификация нуклеиновой кислоты с использованием модифицированного нуклеотида-предшественника вместо одного из нормальных нуклеотидов-предшественников приводит к включению в ДНК основания - субстрата для гликозилазы вместо одного из четырех нормальных оснований G, А, Т или С.
Обычно образец целевой нуклеиновой кислоты представляет собой ДНК. Однако можно использовать также РНК после ее превращения в ДНК с помощью обратной транскрипции.
Если модифицированный нуклеотид-предшественник частично замещает один из нормальных нуклеотидов-предшественников, отношение модифицированного нуклеотида-предшественника к замещаемому нормальному нуклеотиду-предшественнику таково, что в каждую нить амплифицированной ДНК оптимально включается один модифицированный нуклеотид-предшественник. Это позволяет затем после осуществления, как описано здесь, контакта с ДНК-гликозилазой и агентом, способным вести расщепление в лишенном основания звене, расщепить одну нить амплифицированной ДНК на два фрагмента. Этот способ расщепления назван здесь "расщеплением с участием гликозилазы". Для получения более одного сайта расщепления на одну нить амплифицированной ДНК используют более высокое отношение модифицированного нуклеотида-предшественника к нормальному нуклеотиду-предшественнику. Включение модифицированного нуклеотида-предшественника в продукт амплификации приводит к появлению в продукте амплификации одного или более модифицированных оснований в одном или более положений, различаемых ферментом ДНК-гликозилазой.
Если в этапе (i) используется полное замещение нормального нуклеотида-предшественника модифицированным нуклеотидом-предшественником, это позволяет с помощью расщепления с участием гликозилазы расщепить праймер, удлиненный в реакции амплификации, в первом 3'-положении удлиненного праймера, где нормальное основание замещено модифицированным основанием.
Так, если CTAG - последовательность матрицы, следующая в 3'-сторону сразу вслед за положением, где гибридизируется праймер, и модифицированный нуклеотид-предшественник представляет собой dUTP (дезоксиуридин-трифосфат), замещающий dTTP, тогда модифицированное основание - урацил (U) будет включаться напротив А в матричной нити. Таким образом, в этой ситуации праймер будет после амплификации и расщепления с участием гликозилазы удлинен на 3'-конце двумя нуклеотидами (праймер-GA 3'). Эти удлиненные праймеры, полученные после первоначального удлинения и расщепления с участием гликозилазы, названы здесь среди прочего удлиняемыми (способными к удлинению) фрагментами, как указано выше, тогда как праймеры до удлинения названы здесь первоначальными праймерами.
Удлиненная 3'-конечная последовательность удлиняемого фрагмента синтезирована ферментативным способом и непосредственно соответствует нуклеиновой кислоте, характеризуемой как нуклеиновая кислота, действующая как матрица для ее синтеза. Таким образом, 3'-конец удлиняемого фрагмента комплементарен нуклеиновой кислоте. Поэтому определение любым способом природы 3'-конца удлиняемого фрагмента позволяет охарактеризовать нуклеиновую кислоту, по которой он получен. Если первоначальный праймер поместить вплотную к локусу, в котором имеется вариация нуклеотидной последовательности ДНК - такая, как полиморфизм или мутация, чтобы первое модифицированное основание, включенное в удлиненный праймер, было в локусе с мутацией, тогда первоначальный праймер после расщепления с участием гликозидазы будет удлиняться до различной длины, в зависимости от того, присутствует или нет мутация в данном локусе. Потом способные к удлинению фрагменты обрабатывают, если необходимо, таким образом, что они могут быть использованы как праймеры для последующей реакции удлинения. Поскольку нуклеотидная последовательность на З'-концах удлиняемых фрагментов различна в зависимости от того, присутствует или отсутствует мутация в анализируемом на мутацию локусе, анализ возможности удлиняемого фрагмента функционировать в последующей реакции удлинения с использованием матричной нуклеиновой кислоты позволяет определить, присутствует ли или отсутствует мутация в анализируемом на мутацию локусе. В качестве матричной нуклеиновой кислоты, обеспечивающей подлежащую проверке способность удлиняемого фрагмента работать в качестве праймера, может быть сконструирована и/или отобрана любая природная или синтезированная ферментативным или химическим способом матрица, которая полностью или частично гибридизируется со способным к удлинению фрагментом.
Если в реакции амплификации часть нормального нуклеотида-предшественника замещается модифицированным нуклеотидом-предшественником, расщепление с участием гликозилазы праймера, удлиненного в реакции амплификации, даст набор (популяцию) способных к удлинению фрагментов различной длины, так как различные молекулы расщепляются в разных точках в зависимости от того, где включился модифицированный нуклеотид-предшественник. Длина каждого фрагмента определяется местом включения модифицированного нуклеотида-предшественника в ходе удлинения с 3'-конца первоначального праймера.
Амплификация целевой нуклеиновой кислоты с использованием нуклеотидов-предшественников dATP, dCTP, dGTP, dTTP и небольшого количества модифицированного нуклеотида-предшественника dUTP приводит к получению аплифицированной ДНК, в которой тимин заменен случайным образом (статистически) на урацил. Урацил в ходе процесса амплификации включается во вновь синтезируемую нить ДНК в положениях, где он комплементарен остаткам аденина в матричной нити ДНК. Амплификация целевой нуклеиновой кислоты с использованием нуклеотидов-предшественников dATP, dCTP, dGTP, dTTP и небольшого количества модифицированного нуклеотида-предшественника dITP приводит к получению амплифицированной ДНК, в которой гуанин предпочтительно заменен случайным образом на гипоксантин. Гипоксантин в ходе процесса амплификации включается во вновь синтезируемую нить ДНК в положениях, где он комплементарен остаткам цитозина в матричной нити ДНК, если количество других нуклеотидов-предшественников не является лимитирующим. Амплификация целевой нуклеиновой кислоты с использованием нуклеотидов-предшественников dATP, dCTP, dGTP, dTTP и небольшого количества модифицированного нуклеотида-предшественника (8-OH)-dGTP приводит к получению амплифицированной ДНК, в которой гуанин предпочтительно заменен случайным образом на (8-ОН)-гуанин. (8-ОН)-гуанин в ходе процесса амплификации включается во вновь синтезируемую нить ДНК в положениях, где он комплементарен остаткам цитозина в матричной нити ДНК, если количество других нуклеотидов-предшественников не является лимитирующим.
Нити амплифицированной ДНК можно разделить для анализа по отдельности соответствующих нитей. Разделенные нити дополнительно могут быть иммобилизованы, что можно осуществить различными способами. Общепринятый способ иммобилизации и/или разделения нитей ДНК состоит в использовании взаимодействия биотина со стрептавидином, причем обычно ДНК содержит биотиновую метку, а стрептавидин прикреплен к нерастворимому носителю. Однако способ в соответствии с данным изобретением пригоден для любых процедур иммобилизации, обеспечивающих иммобилизацию направленного вверх фрагмента, нити, несущей направленный вверх фрагмент, нити комплементарной к нити, несущей направленный вверх фрагмент, матричной нуклеиновой кислоты, целевой нуклеиновой кислоты и продуктов, полученных при расщеплении с участием гликозилазы амплифицированных или удлиненных нуклеиновых кислот, содержащих модифицированные основания.
Вместо применения техники амплификации, такой как полимеразная цепная реакция (ПЦР), модифицированное основание может быть введено путем химической модификации нуклеиновой кислоты.
Существует несколько методов, в которых обработка ДНК специфическими реагентами модифицирует существующие основания таким образом, что они распознаются специфическими разновидностями фермента ДНК-гликозилазы. Например, обработка ДНК алкилирующими агентами, такими как метилнитрозомочевина, дает несколько типов алкилированных оснований, включая N3-метиладенин и N3-метилгуанин, которые распознаются и выщепляются алкилпурин-ДНК-гликозилазой. Обработка ДНК бисульфитом натрия приводит к дезаминированию в ДНК остатков цитозина с образованием остатков урацила, которые распознаются и выщепляются урацил-ДНК-гликозилазой. Обработка ДНК сульфатом железа и EDTA (этилендиаминтетраацетатом) вызывает окисление в ДНК остатков гуанина с образованием в ДНК остатков (8-ОН)-гуанина, которые распознаются и выщепляются формамидопиримидин-ДНК-гликозилазой.
Таким образом, имеющиеся в нуклеиновой кислоте или, в особенности, в способном к удлинению фрагменте, получаемом в этапе (iii), основания могут быть превращены химическими методами в модифицированные основания. Часть остатков цитозина или все они могут быть легко превращены при использовании бисульфита натрия в урацил, что делает амплифицированный образец восприимчивым к действию урацил-ДНК-гликозилазы в местах превращения цитозина. Если синтезировать верхний или нижний праймер таким образом, чтобы он содержал вместо цитозина 5-метилцитозин, то в таком случае праймер будет устойчив к расщеплению с участием урацил-ДНК-гликозилазы, поскольку дезаминирование 5-метил цитозина происходит с меньшей скоростью по сравнению с цитозином и приводит к образованию тимина, а не урацила.
Перед обработкой подходящей ДНК-гликозилазой двунитевую ДНК можно обработать экзонуклеазой I. Эта обработка позволяет разрушить неиспользованные праймеры и любые неспецифические однонитевые продукты амплификации ДНК, что повышает отношение сигнала к шуму.
В том случае, когда модифицированным нуклеотидом-предшественником является dUTP, в амплифицированном образце целевой нуклеиновой кислоты в соответствующих тимину положениях будет возникать модифицированное основание - урацил. Добавление к образцу урацил-ДНК-гликозилазы приводит к выщеплению урацила из образца. В том случае, когда модифицированным нуклеотидом-предшественником является dITP, в амплифицированном образце целевой нуклеиновой кислоты в соответствующих гуанину положениях будет возникать модифицированное основание - гипоксантин. Добавление к образцу алкилпурин-ДНК-гликозилазы приводит к выщеплению гипоксантина из образца. В том случае, когда модифицированным нуклеотидом-предшественником является (8-OH)-dGTP, в амплифицированном образце целевой нуклеиновой кислоты в соответствующих гуанину положениях будет возникать модифицированное основание - (8-ОН)-гуанин. Добавление к образцу формамидопиримидин-ДНК-гликозилазы приводит к выщеплению (8-ОН)-гуанина из образца.
Праймер или один или более из участвующих в ферментативной амплификации нуклеотидов могут быть помечены.
В частности, может быть помечен используемый первоначальный праймер. Введение метки в праймеры может быть осуществлено многими способами, включая добавление к праймеру радиоактивного, флуоресцирующего или различаемого каким-либо способом лиганда в ходе или после синтеза праймера.
В одном из вариантов осуществления данного изобретения используемый в этапе (iv) фермент является полимеразой, которую можно инкубировать со способным к удлинению направленным вверх фрагментом в присутствии одного или более нуклеотидов.
В этом же варианте желательно, чтобы один или более из нуклеотидов на этапе (iv) был дидезокси-нуклеотидом.
Кроме того, один или более из нуклеотидов этапа (iv) может быть помечен.
Для удлинения 3'-конца способного к удлинению фрагмента на матричной нуклеиновой кислоте могут быть использованы различные полимеразы нуклеиновой кислоты. В литературе описаны многие полимеразы, которые удлиняют 3'-концы праймеров на матричной ДНК. Например, могут быть использованы ДНК-полимеразы, выделенные из фагов и мезофильных и термофильных бактерий.
Несколько ДНК-полимераз, включая Т7 ДНК-полимеразу, при удлинении праймеров включают как концевые (обрывающие цепь) дидезокси-нуклеотиды, так и нормальные нуклеотиды-предшественники. Обычно для амплификации нуклеиновых кислот путем удлинения праймеров в повторяющихся циклах используют термофильные ДНК-полимеразы. Направленные вверх фрагменты ДНК, получаемые на этапе (iii), способны служить праймерами для всех полимераз нуклеиновых кислот, способных удлинять стандартные праймеры нуклеиновой кислоты.
Применение в любой из реакций удлинения меченого нуклеотида-предшественника или концевого дидезокси-нуклеотида облегчает обнаружение удлиненного способного к удлинению фрагмента. Прямые методы окрашивания ДНК, такие как окрашивание серебром или бромистым этидием, облегчают обнаружение всех продуктов удлинения после их разделения по размерам на основе электрофоретической подвижности.
Благодаря тому что способные к удлинению фрагменты могут функционировать в последующей реакции удлинения с использованием матричной нуклеиновой кислоты и нормальных нуклеотидов или концевого дидезокси-нуклеотида (нуклеотид, который после внедрения в цепь препятствует дальнейшему удлинению праймера по матрице), создается набор длин ("лестница") фрагментов, что позволяет определить распределение и полное число положений модифицированного нуклеотида-предшественника в одной или в обеих нитях амплифицированной целевой нуклеиновой кислоты. Наличие вариации нуклеотидной последовательности или мутации приводит к появлению или исчезновению фрагмента - продукта расщепления, о чем судят по сравнению с известной нуклеотидной последовательностью ДНК для амплифицированной молекулы. Анализ размеров фрагментов позволяет определить точную локализацию и нуклеотидную последовательность подлежащей определению мутации в образце целевой нуклеиновой кислоты.
Таким образом, если появляется такая вариация нуклеотидной последовательности, что создается дополнительное место включения модифицированного нуклеотида-предшественника, при анализе лестницы продуктов расщепления будет наблюдаться дополнительный фрагмент - продукт расщепления. Если встречается такая вариация нуклеотидной последовательности, что место включения модифицированного нуклеотида-предшественника утрачивается, при анализе лестницы продуктов расщепления соответствующий фрагмент - продукт расщепления не будет наблюдаться.
В этом случае выбранной матрицей будет служить исходно интактная или расщепленная при участии гликозидазы и амплифицированная нуклеиновая кислота. В тех случаях, когда используется расщепленная с участием гликозидазы амплифицированная нуклеиновая кислота, ее можно обработать для удаления остаточных групп, расположенных вниз от удлиняемого фрагмента, которые препятствуют удлинению способного к удлинению фрагмента полимеразой нуклеиновой кислоты. Конкретно, матричную ДНК можно обработать таким образом, чтобы удалить остаточную 5'-дезоксирибозную группировку. Это достигается инкубацией матричной ДНК с 5'-дезоксирибозо-фосфодиэстеразой, такой как эндонуклеаза RecJ Е.coli или формамидопиримидин-ДНК-гликозилаза. В качестве матричной нуклеиновой кислоты для проверки способности удлиняемого фрагмента работать в качестве праймера может быть сконструирована и/или отобрана любая природная или синтезированная ферментативным или химическим способом матрица, которая полностью или частично гибридизируется с удлиняемым фрагментом.
Благодаря тому что способные к удлинению фрагменты могут функционировать в последующей реакции удлинения с использованием матричной нуклеиновой кислоты и комбинации немеченых и меченых нормальных нуклеотидов или концевого дидезокси-нуклеотида, обеспечивается возможность обнаружения вариаций нуклеотидной последовательности и мутаций. Удлинение способных к удлинению фрагментов на матричной нуклеиновой кислоте, из которой они были получены и которая гетерозиготна в отношении вариации нуклеотидной последовательности, с использованием меченого концевого дидезокси-нуклеотида, имеющего тип комплементарного спаривания, отличный от типа спаривания модифицированного нуклеотида-предшественника, и немеченого концевого дидезоксинуклеотида, имеющего такой же тип спаривания, как и модифицированный нуклеотид-предшественник, обеспечивает обнаружение только вариантных или мутантных локусов, тогда как неизмененные локусы не проявляются. Этот аспект изобретения особенно полезен, поскольку детектирование только вариаций нуклеотидной последовательности обеспечивает высокую эффективность сканирования и детектирования мутаций и позволяет осуществить каталогизацию ("дактилоскопию") нуклеиновых кислот, основанную на вариациях их нуклеотидной последовательности.
Необходимо отметить, что амплификация любой целевой ДНК, которая гетерозиготна по мутации или полиморфизму, приводит к появлению четырех различных дуплексных ДНК, так как (если для примера взять мутацию G→A в положении Х в последовательности ДНК) одна четверть будет гомодуплексом с парой оснований GC в положении X, одна четверть - гомодуплексом с парой оснований AT в положении X, одна четверть - гетеродуплексом с парой оснований GT в положении Х и одна четверть - гетеродуплексом с парой оснований АС в положении X. Аналогично, гетеродуплексную ДНК можно легко получить, денатурируя и перегибридизируя две гомодуплексных ДНК, имеющих различие (различия) в нуклеотидной последовательности.
Таким образом, природу нуклеотидной последовательности 3'-конца удлиняемых фрагментов можно определить по их способности функционировать в качестве праймеров в последующей реакции удлинения с использованием матричной нуклеиновой кислоты. Необходимо, чтобы такое определение было основано на возможности 3'-конца удлиняемого фрагмента гибридизироваться с выбранной матрицей при выбранных условиях. После частичной или полной гибридизации способный к удлинению фрагмент можно удлинить, используя полимеразу нуклеиновой кислоты и предшественники нуклеиновой кислоты или их выбранные комбинации, как здесь описано. Следует подчеркнуть, что для выбора матричных молекул существует много возможностей. Тем не менее удлинение способного к удлинению фрагмента является мерой его гибридизации или отсутствия гибридизации с выбранной матричной молекулой, и поэтому определение природы нуклеотидной последовательности 3'-конца способных к удлинению фрагментов произведено на этой основе, так как эта 3'-конечная последовательность является индикатором нуклеотидной последовательности исходной целевой нуклеиновой кислоты.
Обычно матричную молекулу выбирают таким образом, что она имеет частичную или полную комплементарность нуклеотидной последовательности по отношению к направленному вверх фрагменту. Направленный вверх фрагмент может быть удлинен на один или более нуклеотидов на матричной молекуле с использованием полимеразы нуклеиновой кислоты и предшественников нуклеиновой кислоты, или их комбинации, или концевых дидезокси-нуклеотидов, или комбинации нормальных предшественников нуклеиновой кислоты с концевыми дидезокси-нуклеотидами.
Удлинение в этапе (iv) может быть осуществлено с помощью реакции амплификации с использованием указанного способного к удлинению фрагмента ДНК.
В качестве альтернативы удлинение в этапе (iv) может быть достигнуто с помощью реакции амплификации с включением праймера дополнительно к использованию указанного способного к удлинению фрагмента ДНК.
Повторяющееся удлинение направленного вверх фрагмента на матричной нуклеиновой кислоте в сочетании со вторым фланкирующим праймером, который может удлиняться на копии матрицы, обеспечивает амплификацию матричной нуклеиновой кислоты. Такие амплифицированные продукты можно легко обнаруживать стандартными методами окрашивания нуклеиновых кислот (например, бромистым этидием) после разделения электрофорезом.
В качестве альтернативы можно выбрать матричную молекулу таким образом, чтобы ее можно было удлинять с использованием в качестве матрицы направленного вверх фрагмента, и так, чтобы удлинение было основано на гибридизации с 3'-концом направленного вверх фрагмента.
Кроме того, направленный вверх фрагмент можно анализировать, используя его способность функционировать в пробах с 5'-нуклеазой. В ходе удлинения направленного вверх фрагмента полимеразой с 5'→3'-нуклеазной активностью 5'→3'-нуклеаза разрушает направленную вниз репортерную молекулу, гибридизированную (отожженную) с той же самой матричной нитью, что и указанный направленный вверх фрагмент.
Дополнительная возможность состоит в удлинении направленного вверх фрагмента на синтетической матрице, содержащей репортерную и ослабляющую метки, после чего расщеплением полученной двунитевой ДНК отделяют репортер от ослабителя и определяют сигнал. Обычно такое расщепление производят ферментом, распознающим двунитевую молекулу ДНК. Обычно таким ферментом является рестриктаза.
Кроме того, 3'-концевую нуклеотидную последовательность направленного вверх фрагмента можно определить с помощью его гибридизации с другими молекулами нуклеиновой кислоты.
Дополнительная возможность состоит в том, что продукты, удлиненные или амплифицированные с использованием направленных вверх фрагментов, могут быть обнаружены на основе их свойств фильтрации и/или осаждения.
При анализе удлинения и включения нуклеотидов в этапе (iv), если в реакции удлинения используются направленные вверх фрагменты, важно проверять, что каждое наблюдаемое удлинение является специфическим результатом удлинения направленных вверх фрагментов, а не результатом удлинения исходных праймеров, не использованных при первоначальной амплификации, если она применяется на этапе (i), или результатом удлинения возможно возникающих неспецифически направленных вверх фрагментов, которые могут образовываться при неспецифическом разрыве или разрушении ДНК на предыдущих этапах процедуры. Чтобы избежать появления этого "шума", пред обработкой с участием гликозилазы можно обрабатывать ДНК нуклеазой, специфической к однонитевой ДНК, - например, экзонуклеазой I, которая разрушит неиспользованные праймеры и неспецифическую однонитевую ДНК и которую можно затем инактивировать прогревом. ДНК можно обработать смесью 3'-АП-эндонуклеаза/лиаза, которая разрежет ДНК и праймеры у всех предсуществующих АП-звеньев, созданных специфически или появившихся в ходе разрушения ДНК. Затем 3'-АП-эндонуклеазу/лиазу удаляют из реакции. Поскольку эндонуклеаза/лиаза разрезает цепь на 3'-стороне звена, получаемые примесные направленные вверх фрагменты не способны к удлинению и не будут смешиваться с создаваемыми после этого направленными вверх фрагментами. Кроме этих обработок, можно для проверки неспецифических удлинений провести контрольную реакцию. Так, на этапе (iii) данного способа можно сделать разрезы в АП-звеньях 3'-АП-эндонуклеазой/лиазой, получая таким образом не способные к удлинению (неудлиняемые) направленные вверх фрагменты. Однако если в следующем этапе (iv) наблюдается удлинение и включение нуклеотидов, тогда можно измерить или определить уровень получаемого в ходе этой процедуры неспецифического удлинения.
Важно также гарантировать, что включение нуклеотидов, меченых или немеченых, на этапе (iv) есть включение именно нуклеотидов, внесенных на этапе (iv), а не нуклеотидов, попавших из предыдущих этапов. Это особенно важно, если в реакции происходит включение дидезокси-нуклеотидов. Перед или вслед за расщеплением ДНК в лишенных основания звеньях и создания направленных вверх фрагментов реакционную смесь можно обработать фосфатазой, которая разлагает все присутствующие в реакции невключенные нуклеотиды. Можно использовать, например, щелочную фосфатазу креветок, которую затем можно инактивировать прогревом.
В следующем варианте осуществления изобретения используемый на этапе (iv) фермент является лигазой, которую можно инкубировать вместе с удлиняемым направленным вверх фрагментом в присутствии репортерного олигонуклеотида.
Репортерный олигонуклеотид может быть частично дегенерированным.
Способ по настоящему изобретению может быть применен, среди прочего, для обнаружения известной или неизвестной мутации и для обнаружения различий и подобий в геномах. Эти аспекты изобретения дополнительно иллюстрированы далее.
Способ в соответствии с данным изобретением предоставляет в одном из аспектов инструмент для создания случайных праймеров простым легким приемом. По сути, введение в амплифицируемый ДНК-продукт модифицированного основания, затем расщепление с участием гликозилазы и последующая обработка продуктов расщепления таким образом, что они могут быть удлинены полимеразой нуклеиновой кислоты, дает метод быстрого создания случайных праймеров. Последующее использование таких праймеров - например, способных к удлинению направленных вверх фрагментов в случайной, статистической амплификации целевых нуклеиновых кислот, обеспечивает амплификацию дискретных молекул ДНК среди целевых нуклеиновых кислот, позволяя таким образом охарактеризовать нуклеиновую кислоту на основе подобий и различий амплифицированных продуктов. Поскольку эти праймеры по существу ведут свое происхождение от указанных целевых нуклеиновых кислот, конкретно от их 3'-концов, они являются более хорошими праймерами для последующего анализа путем статистической амплификации указанной нуклеиновой кислоты, и амплификация более специфична. В ходе статистической амплификации нуклеиновых кислот создается множество дискретных ДНК-продуктов. Отдельный ДНК-продукт можно отделить от других продуктов фракционированием по размерам. Способ в соответствии с данным изобретением позволяет получать праймеры из всего отделенного продукта или из его части. Использование исходного праймера в случайной амплификации нуклеиновой кислоты, которая позволяет иммобилизовать отделенный продукт, обеспечивает выделение верхних и нижних праймеров, удлиненных до первой точки расщепления с участием гликозилазы. 3'-Концы таких направленных вверх фрагментов получены из целевой нуклеиновой кислоты и поэтому обеспечивают более специфическую амплификацию целевой нуклеиновой кислоты или родственных нуклеиновых кислот. 3'-Концы таких направленных вверх фрагментов могут быть короткими или длинными. Здесь под коротким 3'-концом подразумевается конец, содержащий от одного до трех нуклеотидов, а под длинным 3'-концом подразумевается конец из более чем трех нуклеотидов. Предпочтительны более длинные 3'-концы таких направленных вверх фрагментов, так как они обеспечивают высокую специфичность амплификации нуклеотидной последовательности целевой нуклеиновой кислоты. Направленные вверх фрагменты, образующиеся с более длинными 3'-концами, можно отобрать с помощью методов фракционирования по размерам. В качестве альтернативы можно так сконструировать первоначальные праймеры амплификации, что они способствуют связыванию белка, который защищает участок 3'-области исходного удлиненного праймера. Поэтому такая область устойчива к расщеплению с участием гликозилазы благодаря защите белком, и включение такого конструирования праймера и белка позволяет получать направленные вверх фрагменты с более длинными 3'-концами.
Этот вариант осуществления изобретения предоставляет быстрый и простой способ получения случайных и специфических праймеров для амплификации нуклеиновой кислоты без необходимости знать заранее нуклеотидную последовательность нуклеиновой кислоты.
Необходимо учитывать, что случайная амплификация нуклеиновой кислоты с использованием произвольно выбранных праймеров является уже известным способом обнаружения подобий и различий в геномах. Такая случайная амплификация основана на гибридизации произвольно выбранных праймеров с целевыми образцами с последующими множественными циклами ферментативной амплификации, причем праймеры удлиняются с использованием в качестве матрицы геномной ДНК или комплементарной ДНК (кДНК). Использование таких праймеров в таком способе приводит к амплификации дискретных молекул ДНК. Анализ таких молекул позволяет исследовать подобия и различия между различными образцами. Обычно для исследования генома или кДНК выбирают большое число различных случайных праймеров, и такие праймеры синтезируют химически и конструируют случайным образом, предполагая, что они будут в процессе амплификации гибридизироваться с целевой нуклеиновой кислотой. Способ в соответствии с настоящим изобретением позволяет легко и быстро получать праймеры из целевой нуклеиновой кислоты, которые затем могут быть использованы для случайной амплификации той же самой или другой целевой нуклеиновой кислоты.
В дополнение к удлинению с помощью полимеразной реакции, как указано выше, направленные вверх фрагменты в этапе (iv) могут быть также удлинены ферментативной пришивкой (лигированием) другой однонитевой молекулы ДНК, в результате чего получаются удлиненные направленные вверх фрагменты большего размера, чем первоначальные направленные вверх фрагменты, получаемые расщеплением с участием гликозилазы. Молекула ДНК, к которой пришивают лигазой направленный вверх фрагмент, здесь названа репортерным олигонуклеотидом, и она может быть различной длины. Пришивка направленных вверх фрагментов к репортерному олигонуклеотиду зависит от обеих молекул ДНК (направленный вверх фрагмент и репортерный олигонуклеотид), гибридизирующихся с матричной молекулой в смежных участках таким образом, что концы направленного вверх фрагмента и репортерного олигонуклеотида примыкают друг к другу. Это означает, что 3'-концевое основание направленного вверх фрагмента примыкает к 5'-концевому основанию репортерного олигонуклеотида. Обычно репортерный олигонуклеотид является синтетическим олигонуклеотидом, однако он может быть любым иным типом молекулы ДНК или молекулы РНК.
Матричную молекулу выбирают таким образом, что она имеет частичную или полную комплементарность нуклеотидной последовательности с направленным вверх фрагментом и репортерным олигонуклеотидом. Предпочтительно матрицей может быть исходно интактная или расщепленная с участием гликозилазы амплифицированная нуклеиновая кислота. Кроме того, матрица может быть однонитевой молекулой ДНК, т.е. синтетическим олигонуклеотидом, который может иметь различную длину и с которым могут комплементарно гибридизироваться направленный вверх фрагмент (фрагменты) и репортерный олигонуклеотид (олигонуклеотиды). Матричная ДНК может быть по своей природе однонитевой или двунитевой. Двунитевая ДНК, служащая матрицей, состоит из матричной нити и комплементарной нити. Двунитевую ДНК можно вначале денатурировать и затем перегибридизировать ее в присутствии направленного вверх фрагмента и репортерного олигонуклеотида. При этом направленный вверх фрагмент и репортерный олигонуклеотид конкурируют с комплементарной нитью в гибридизации с матричной нитью двунитевой матричной молекулы.
Для удлинения 3'-конца удлиняемого фрагмента сшиванием его с репортерным олигонуклеотидом на матричной нуклеиновой кислоте можно использовать различные ДНК-лигазы и РНК-лигазы. Для пришивки репортерного олигонуклеотида к удлиняемому фрагменту могут быть использованы ДНК-лигазы из многих источников, включая ДНК-лигазы, выделенные из фагов - например, Т4 ДНК-лигаза, а также из мезофильных и термофильных бактерий. Термофильные ДНК-лигазы могут быть использованы в повторных циклах пришивок удлиняемых фрагментов к репортерным олигонуклеотидам.
Как обсуждалось выше, способ по данному изобретению включает создание направленного вверх фрагмента, имеющего 3'-гидроксильную группу. Это является необходимым требованием для удлинения молекулы путем добавления нуклеотидов полимеразой, а также для удлинения путем пришивки лигазой репортерного олигонуклеотида. Кроме наличия 3'-гидроксильной группы у направленного вверх фрагмента, для осуществления пришивки необходимо, чтобы репортерный олигонуклеотид имел 5'-фосфатную группу. Кроме этого, пришивка лигазой позволяет при необходимости обнаруживать направленный вниз фрагмент. Направленный вниз фрагмент - это остаток нити ДНК, из которой был выщеплен направленный вверх фрагмент. В этом случае необходимо, чтобы репортерный олигонуклеотид имел концевой 3'-гидроксил, а направленный вниз фрагмент имел на 5'-конце фосфатную группу.
Поскольку расщепление с участием гликозилазы может приводить в одной реакции к образованию нескольких различных направленных вверх фрагментов из-за присутствия в целевой нуклеиновой кислоте нормальных и мутантных аллелей или из-за случайного (статистического) включения модифицированных оснований, удлинение с помощью реакции пришивки может иметь дело с несколькими различными направленными вверх фрагментами в дополнение к нескольким различным репортерным олигонуклеотидам и матричным нуклеиновым кислотам. Кроме того, можно одновременно охарактеризовать несколько различных нуклеиновых кислот, так как для получения характеристик каждой индивидуальной исследуемой нуклеиновой кислоты можно использовать индивидуальный репортерный олигонуклеотид и/или индивидуальную матричную нуклеиновую кислоту. Удлиненный направленный вверх фрагмент можно обнаружить любым из нескольких способов, включая анализ по размерам, гибридизацию и амплификацию. Кроме того, молекулу ДНК, полученную сшивкой лигазой, можно далее амплифицировать в полимеразной цепной реакции.
Репортерный олигонуклеотид, направленный вверх фрагмент или матричную нуклеиновую кислоту можно пометить. Например, кроме метки биотином или дигоксигенином, можно применить флуоресцентную или радиоактивную метку. Удобной меткой для репортерного олигонуклеотида является 5'-концевой радиоактивный фосфат, т.е. 32Р или 33Р. Радиоактивный фосфат служит как меткой для обнаружения ДНК, так и необходимым 5'-фосфатом репортерной молекулы. Метка биотином на репортерном олигонуклеотиде, направленном вверх фрагменте или матричной нуклеиновой кислоте служит для иммобилизации направленного вверх фрагмента на нерастворимом носителе - или прямо, или путем гибридизации. Иммобилизация в сочетании с множественными различными реакциями удлинения лигазой позволяет получить очень эффективную и высокопроизводительную систему для получения характеристик молекул ДНК.
Настоящее изобретение с использованием удлинения путем пришивки лигазой благодаря способности идентифицировать мутантный направленный вверх фрагмент в смеси с нормальными направленными вверх фрагментами можно также использовать для идентификации в нуклеиновой кислоте неизвестных изменений нуклеотидной последовательности.
В реакции пришивки лигазой можно использовать частично вырожденный репортерный олигонуклеотид. Можно также получить селективную пришивку направленного вверх фрагмента, полученного в результате расщепления молекулы ДНК в мутантном звене, путем использования вырожденного репортерного олигонуклеотида, у которого 5'-конец комплементарен нормальной аллели. В дополнение к этому изобретение может быть применено для исследования всех динуклеотидных участков CpG (p - фосфатная группа) во фрагменте ДНК с использованием полностью комплементарного репортерного олигонуклеотида, сконструированного таким образом, чтобы он гибридизировался с каждым участком CpG. Длина любого получаемого продукта пришивки лигазой укажет участок CpG, который подвергся мутации.
Полезно, чтобы все удлиненные фрагменты, получаемые на этапе (iv), обнаруживались с помощью гибридизации.
Способ в соответствии с настоящим изобретением обладает следующими существенными преимуществами перед известными способами:
а) Указанный способ позволяет определить подобия или различия или же подобия и различия между образцами нуклеиновой кислоты. В частности, он позволяет определить это в большом числе множественных различных локусов в нуклеиновой кислоте. При том что для достижения этой цели можно использовать другие способы, данный способ обладает преимуществами, так как он предлагает одиночный процесс, который может быть легко масштабирован и позволяет быстро и легко охарактеризовать молекулы нуклеиновой кислоты.
б) Следует учитывать, что амплификация нуклеиновых кислот является распространенным методом получения характеристик и обнаружения нуклеиновых кислот. Амплификация зависит от праймеров, которые удлиняются в процессе амплификации. Способ по данному изобретению обеспечивает создание праймеров высокой специфичности для амплификации нуклеиновых кислот без необходимости знать предварительно какую-либо нуклеотидную последовательность нуклеиновой кислоты. Таким образом, данный способ в высокой степени пригоден для получения характеристик нуклеиновых кислот путем анализа полученных из них продуктов амплификации. Описанные для получения характеристик нуклеиновых кислот подходы к амплификации в соответствии с настоящим изобретением позволяют получить характеристики нуклеиновых кислот таким путем, который не был возможен до настоящего изобретения.
в) В данной области имеется потребность в упрощенных способах обнаружения специфических мутаций в предполагаемых локусах. Способ в соответствии с настоящим изобретением предоставляет такой упрощенный способ для обнаружения таких мутаций. В частности, направленный вверх фрагмент, создаваемый на локусах нуклеиновых кислот с вариациями нуклеотидной последовательности, позволяет анализировать такие локусы с использованием большого числа различных аналитических методов и обеспечивает точное и упрощенное обнаружение вариаций нуклеотидной последовательности в таких локусах.
г) Способ в соответствии с данным изобретением обеспечивает возможность получения характеристик нуклеиновых кислот с лучшей воспроизводимостью и меньшей вероятностью ошибок.
Способ в соответствии с настоящим изобретением может быть также применен для анализа содержания CpG в ДНК на основе обнаружения в ДНК переходов С (цитозина) в Т (тимин).
При анализе данных по мутациям и спектров мутаций, полученных в течение многих лет мутационных исследований, была идентифицирована горячая точка для мутаций во всех организмах (в том числе и у людей), содержащих в своей геномной ДНК 5-метилцитозин. Такими мутациями являются мутации в динуклеотидах CpG. Динуклеотиды CpG являются в человеческих клетках точками метилирования цитозина и вовлечены в выполнение многих структурных и регуляторных функций соответственно в организации генома и в экспрессии генов. Цитозин в ДНК в норме подвержен дезаминированию (с низкой, но доступной измерению эффективностью) с образованием урацила. Если не происходит репарация, это событие является мутагенным. Однако после метилирования в 5-м положении чувствительность кольца цитозина к спонтанному дезаминированию увеличивается. В итоге остаток 5-метилцитозина дезаминируется и становится тимином. После дезаминирования динуклеотид CpG превращается в динуклеотид TpG. Таким образом, поскольку 5-метилцитозин имеется только в участках CpG и главная причина мутации в этом сайте является следствием дезаминирования 5-метилцитозина, в ДНК появляются мутированные последовательности CpG в виде динуклеотидных последовательностей TpG, что является классической мутацией перехода ("транзицией") С→Т. Поскольку в исследованиях в области генетики человека было показано, что CpG является такой горячей точкой мутаций, крайне полезен быстрый поиск для улучшения обнаружения мутаций в динуклеотидах CpG в испытуемом фрагменте ДНК.
Для улучшения обнаружения мутаций в последовательностях CpG можно после амплификации целевой ДНК, следующей за расщеплением с созданием направленных вверх фрагментов, выполнить одну из следующих процедур.
Этап (iv) проводится в присутствии полимеразы, дидезокси-ТТР (ddTTP), меченого дидезокси-СТР (ddCTP) и ДНК дикого типа в качестве матрицы, направленные вверх фрагменты удлиняются включением дидезоксинуклеотидов, но только места, где CpG претерпели мутацию в TpG, станут мечеными после включения меченого ddCTP. Поскольку ddCTP является нуклеотидом, обрывающим цепь, то ДНК не будет удлиняться за этой точкой. Таким образом, ДНК будет удлинена в мутированных последовательностях CpG и будет также меченой и доступной для обнаружения.
Этап (iv) проводится в присутствии полимеразы, ДНК дикого типа в качестве матрицы, dTTP и меченого дидезокси-GTP (ddGTP), направленные вверх фрагменты удлиняются включением dTMP (дезокситимидинмонофосфата, в который превращается dTTP после включения в нить ДНК), но мечеными после включения меченого ddGTP будут только места, где после Т следует G, то есть последовательности TpG. Так как ddGTP является обрывающим цепь нуклеотидом, ДНК не будет удлиняться дальше этой точки. Таким образом, ДНК будет теперь удлинена по всем последовательностям TpG и будет также меченой и доступной для обнаружения. Поскольку динуклеотиды TpG естественным образом содержатся в ДНК, а также возникают вследствие дезаминирования CpG, ДНК дикого типа будет давать характерное распределение зон (обычно видно только несколько зон). Метками могут быть, например, радиоактивный 33P, флуоресцентная метка или биотин, которые вводятся способами, которые известны специалистам.
Этап (iv) проводится в присутствии полимеразы, dUTP, dGTP, dATP, меченого dCTP и ДНК дикого типа в качестве матрицы, направленные вверх фрагменты удлиняются включением дезоксинуклеотидов и становятся мечеными. Затем удлиненные направленные вверх фрагменты расщепляются урацил-ДНК-гликозилазой и агентом, расщепляющим цепь в лишенном основания звене. Остается удлиненным и меченым только такой направленный вверх фрагмент, который возникает при расщеплении в мутантном звене. Эта процедура обнаруживает все мутации, вызывающие появление точек включения Т, в том числе мутации С→Т в участках CpG. Эта процедура имеет то преимущество, что в этапе (iv) используют включение дезоксинуклеотидов, которые включаются всеми полимеразами более эффективно, чем дидезоксинуклеотиды.
В дополнение к этому в гетерозиготных образцах мутированные CpG порождают нарушения спаривания T/G (или U/G после включения в качестве модифицированного нуклеотида dUTP). Для специфического расщепления у этих точек отсутствия спаривания T/G или U/G в мутированных последовательностях CpG можно использовать тимин-ДНК-гликозилазу.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1 представляет схему лишенного основания звена (АП-звена), получаемого на одном этапе способа по данному изобретению, и нескольких путей, которыми ДНК может быть расщеплена у лишенного основания звена таким образом, чтобы создать направленный вверх фрагмент ДНК, который может быть удлинен.
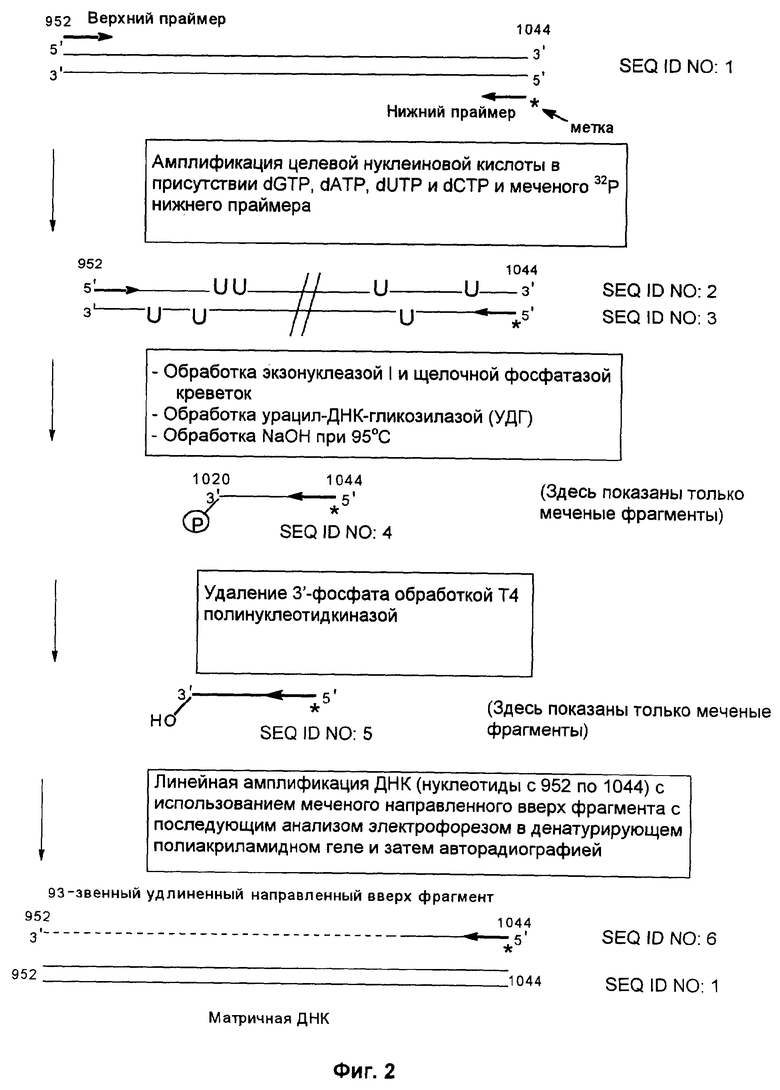
Фиг.2 схематически представляет способ по данному изобретению, как он описан в примере 1.
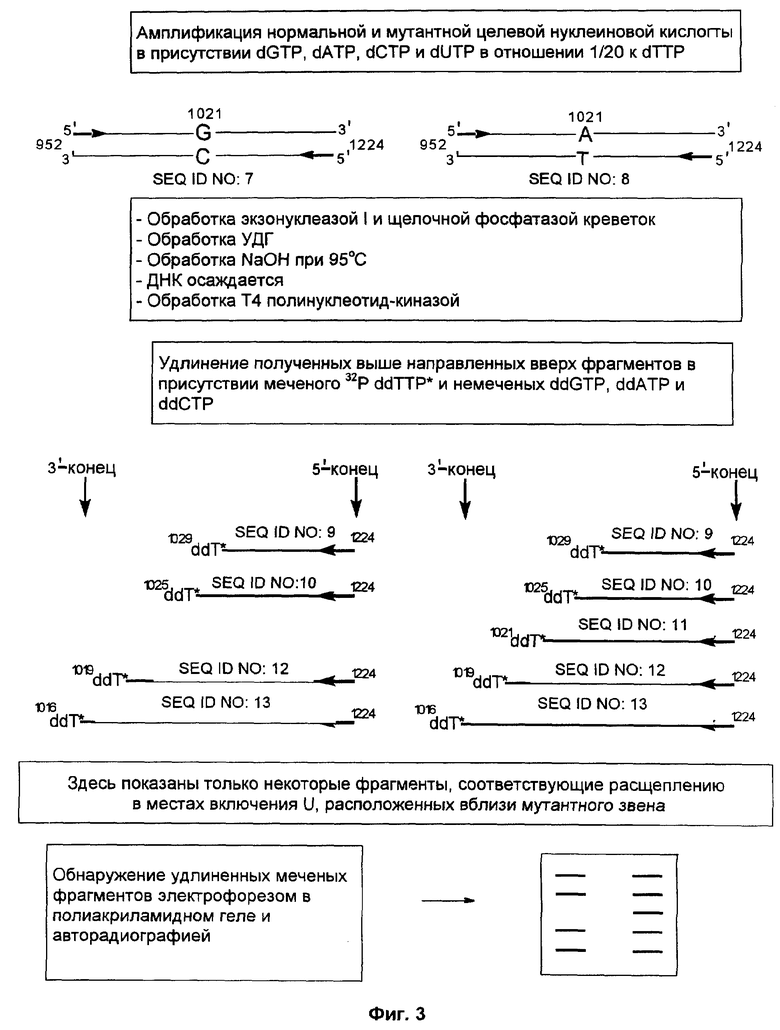
Фиг.3 представляет схематическое изображение способа по настоящему изобретению, как он описан в примере 2, где в реакции линейной амплификации был использован меченый ddTTP, после чего следовало создание направленного вверх способного к удлинению фрагмента.
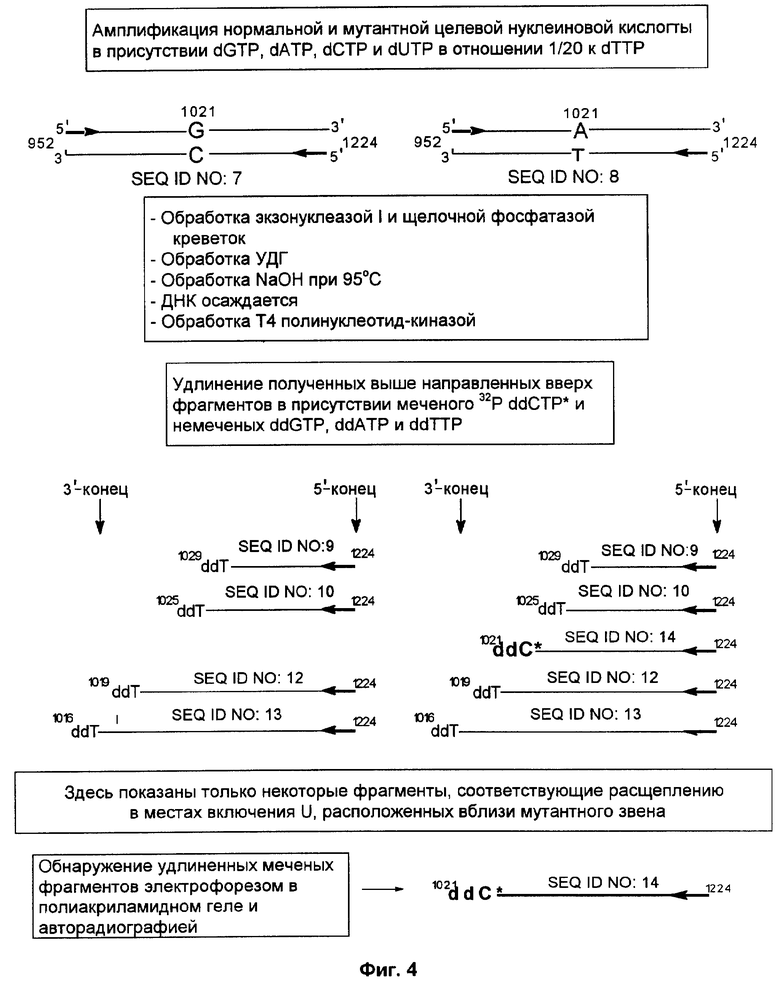
Фиг.4 представляет схематическое изображение способа по настоящему изобретению, как он описан в примере 2, где в реакции линейной амплификации был использован меченый ddCTP, после чего следовало создание направленного вверх способного к удлинению фрагмента.
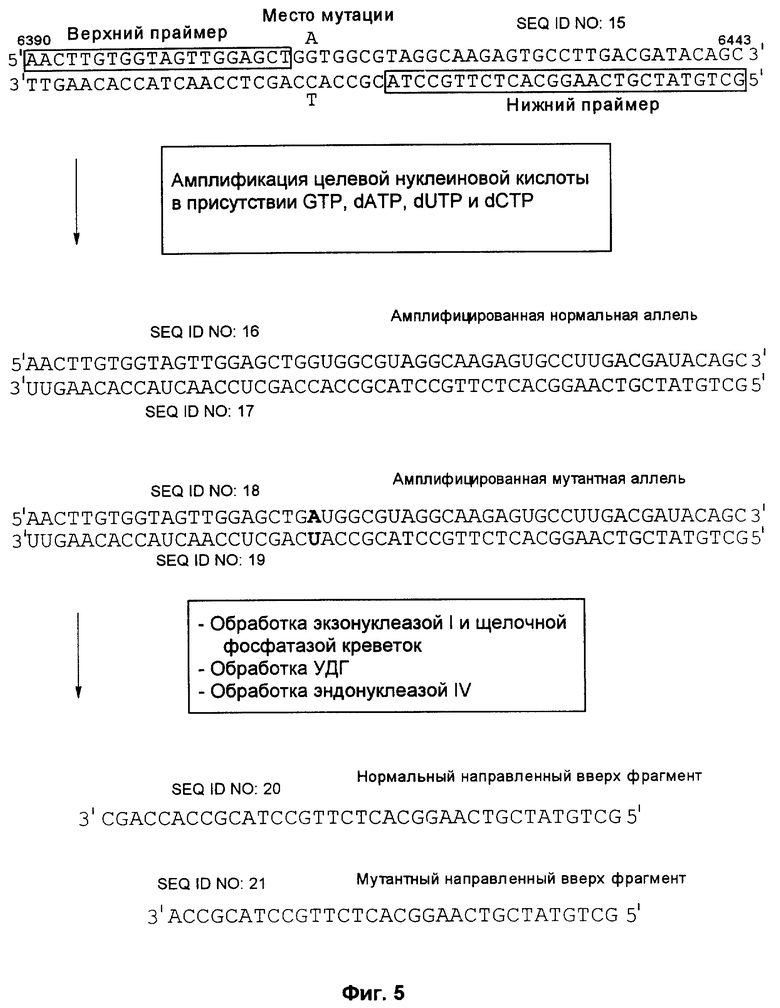
Фиг.5 представляет схематическое изображение способа по настоящему изобретению, как он описан в примере 3.
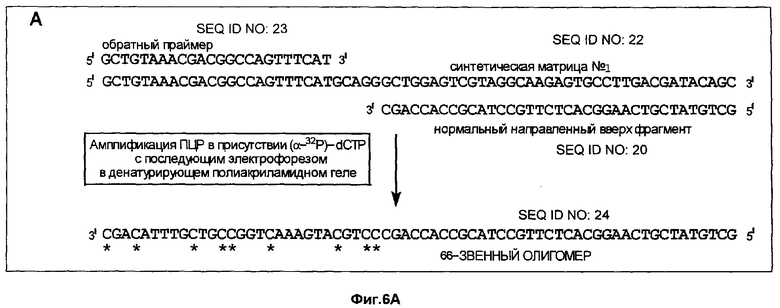
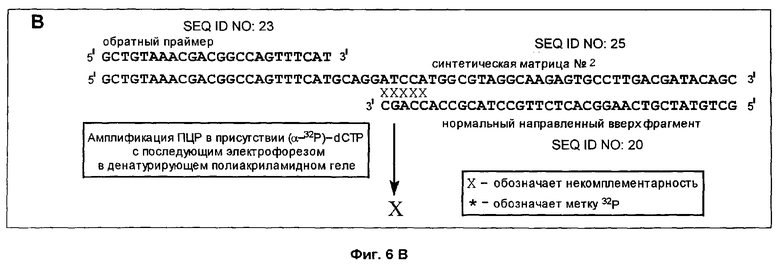
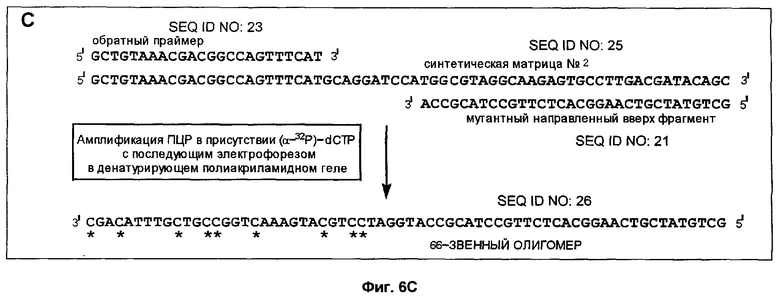
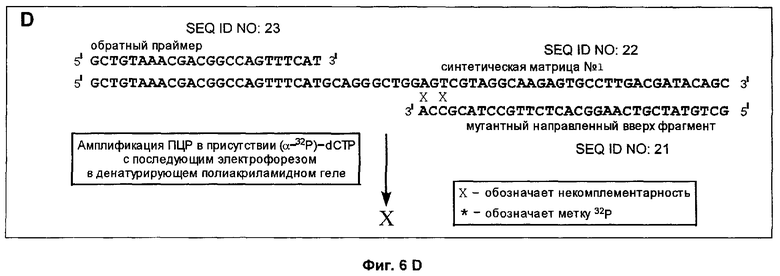
Фиг.6A-6D дает схематическое изображение продуктов удлинения, полученных в примере 3 после электрофореза и авторадиографии и анализа авторадиограмм; и
Фиг.7A-7D дает схематическое изображение реакций пришивки лигазой, осуществленных в примере 4 с направленными вверх фрагментами, и полученных таким путем продуктов.
На фиг.1 изображена одиночная нить ДНК в ориентации 5'→3'. Показано расположение двух оснований, т.е. основания 1 и основания 3. Основание 2 было удалено, и в этом положении находится АП-звено. Вертикальные линии означают кольцо рибозы, которое связано с основанием. Диагональные линии со значком "Р" в кружке обозначают фосфодиэфирные связи, связывающие между собой остатки рибозы. Как показано стрелками, ДНК может быть разрезана на 5'-стороне или на 3'-стороне АП-звена.
Если ДНК разрезана на 3'-стороне АП-звена, т.е. в месте Z, тогда направленный вверх фрагмент (содержащий основание 1) имеет на своем 3'-конце остаток дезоксирибозо-фосфата, как показано в рамке С. Расщепление ДНК на 3'-стороне АП-звена может быть достигнуто обработкой смесью 3'-АП-эндонуклеаза/лиаза или тепловой обработкой, как описано выше.
АП-звено может также быть отрезано двумя различными способами расщеплением на 5'-стороне АП-звена, т.е. в местах Х и Y. Расщепление в месте Х приводит к образованию на 3'-конце направленного вверх фрагмента ОН-группы, как показано в рамке А. Расщепление в месте Х может быть осуществлено обработкой 5'-АП-эндонуклеазой, как описано выше. Расщепление в месте Y приводит к образованию на 3'-конце направленного вверх фрагмента фосфатной группы, как показано в рамке В. Расщепление в месте Y может быть осуществлено обработкой теплом и щелочью, как описано выше.
Далее изобретение будет иллюстрировано сопутствующими примерами.
ПРИМЕРЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Пример 1
Способ в соответствии с изобретением был применен для демонстрации получения удлиненного фрагмента ДНК удлинением от 3'-ОН группы направленного вверх фрагмента, который был создан расщеплением содержащей урацил ДНК в месте включения модифицированного нуклеотида. Целевой нуклеиновой кислотой была область гена RYR1 (нуклеотиды с 952 по 1044), амплифицированная с кДНК человека с использованием верхнего (нуклеотиды с 952 по 972) и нижнего (нуклеотиды с 1024 по 1044) праймеров для получения двунитевой ДНК длиной 93 пары оснований. (Номера нуклеотидов соответствуют нуклеотидной последовательности гена RYR1).
Фиг.2 представляет собой схему целевой нуклеиновой кислоты и верхнего и нижнего праймеров (праймеры содержат стандартные основания G, А, Т и С). В нижний праймер (6 пикомолей) вводили концевую метку инкубацией в течение 30 минут при 37°С в реакционной смеси состава: 1 ед. Т4 полинуклеотид-киназы (коммерческий продукт фирмы New England Biolabs), 70 мМ трис-HCl (рН 7,6), 10 мМ MgCl2, 5 мМ дитиотреитол и 1 микрокюри (γ-32Р)-АТР (3000 кюри/ммоль). Образец целевой нуклеиновой кислоты амплифицировали с помощью ПЦР в реакционной смеси, содержащей в общем объеме 19 мкл: 0,2 мМ dATP, dCTP, dGTP и dUTP, 6 пикомолей меченого 32P нижнего праймера и немеченого верхнего праймера. Затем на реакционную смесь наслаивали равный объем минерального масла и осуществляли горячий запуск ПЦР посредством нагревания реакционной смеси до 94°С в течение 5 мин, перед добавлением 1 единицы Taq полимеразы (полимераза из Thermus aquaticus, продукт фирмы Promega), общий объем доводили до 20 мкл. В термоциклере проводили 30 циклов денатурации, гибридизации и удлинения. Реакционную смесь, содержащую амплифицированную целевую нуклеиновую кислоту, обрабатывали затем экзонуклеазой I (продукт фирмы Amersham Life Sciences), чтобы разрушить праймеры, которые не были удлинены в стадии амплификации, и щелочной фосфатазой креветок (SAP) (фирмы Boehringer Mannheim), чтобы разрушить дезоксинуклеозид-трифосфаты (dNTPs), не включившиеся на стадии амплификации. Это осуществляли инкубацией 10 мкл реакционной смеси ПЦР с 0,5 ед. экзонуклеазы I и 1 ед. SAP при 37°С в течение 30 мин. Затем экзонуклеазу I и SAP инактивировали теплом, инкубируя реакционную смесь при 80°С в течение 15 мин.
После этого добавляли 0,5 ед. урацил-ДНК-гликозилазы (продукт фирмы New England Biolabs) и инкубацию продолжали при 37°С в течение 30 мин. После обработки урацил-ДНК-гликозилазой проводили исчерпывающее расщепление в лишенных основания звеньях, созданных в амплифицированном продукте. Для этого добавляли NaOH до конечной концентрации 0,05 М и прогревали смесь в течение 15 мин при 95°С. Затем расщепленную ДНК осаждали добавлением 3 М ацетата натрия (10% от объема смеси) и 2 объемов этанола. Осадок ресуспендировали в 5 мкл воды. После этого расщепленную ДНК обрабатывали 0,5 ед. Т4 полинуклеотид-киназы (PNK), которая удаляет с 3'-конца фосфатную группу.
Затем проводили реакцию линейной амплификации, используя продукты предшествующей реакции расщепления, причем интересующий фрагмент - это меченый направленный вверх способный к удлинению фрагмент. В ходе этой реакции направленный вверх способный к удлинению фрагмент удлиняется термостабильной ДНК-полимеразой в ходе циклической реакции в общем объеме 10 мкл. Матрицей для этой реакции служит амплифицированная целевая нуклеиновая кислота (последовательность с 952 по 1044 нуклеотид гена RYR1), которая освобождена от праймеров предварительной обработкой экзонуклеазой I.
К образцу добавляли равный объем формамидного покрывающего красителя (90% формамид, 0,025% бромфенол голубой, 0,025% ксиленцианол), затем образец прогревали при 85°С в течение 5 мин. После этого образец наносили на 20% денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез в течение 3-4 ч при 60 В для анализа продуктов удлинения по размерам. После электрофореза проводили авторадиографию, экспонируя гель в контакте с рентгеновской фотопленкой в течение 12 ч при -70°С.
При анализе авторадиограммы, полученной для случая, когда меченым был нижний праймер, был обнаружен продукт длиной 93 нуклеотида. Этот продукт не наблюдался, если в вышеуказанную процедуру не была включена обработка Т4 полинуклеотидкиназой или реакция линейной амплификации.
Пример 2
Способ согласно настоящему изобретению был применен для обнаружения присутствия мутации G→A в положении 1021 гена RYR1 человека. Амплифицировали кДНК нормального индивидуума и индивидуума со злокачественной гипертермией, используя верхний (нуклеотиды с 952 по 972) и нижний (нуклеотиды с 1204 по 1224) праймеры для создания фрагмента двунитевой ДНК (нуклеотиды с 952 по 1224) (как и в примере 1, номера нуклеотидов относятся к нуклеотидной последовательности гена RYR1).
Фиг.3 и фиг.4 представляют собой схемы целевой нуклеиновой кислоты и верхнего и нижнего праймеров (праймеры содержат стандартные основания G, А, Т и С). Целевую нуклеиновую кислоту амплифицировали ПЦР в реакционной смеси, содержащей в общем объеме 19 мкл целевую нуклеиновую кислоту, по 0,2 мМ dATP, dCTP, dGTP, 0,19 мМ dTTP и 0,01 мМ dUTP, а также 6 пикомолей верхнего и нижнего праймеров. На реакционную смесь наливали сверху равный объем минерального масла и осуществляли горячий запуск ПЦР посредством прогревания реакционной смеси при 94°С в течение 5 мин перед добавлением 1 единицы Taq полимеразы (общий объем смеси становился равным 20 мкл). В термоциклере проводили 30 циклов денатурации, гибридизации и удлинения. Реакционную смесь, содержащую амплифицированную целевую нуклеиновую кислоту, обрабатывали затем экзонуклеазой I, чтобы разрушить праймеры, которые не были удлинены в стадии амплификации, и щелочной фосфатазой креветок (SAP), чтобы разрушить dNTPs, не включившиеся на стадии амплификации. Это осуществляли инкубацией 10 мкл реакционной смеси ПЦР с 0,5 ед. экзонуклеазы I и 1 ед. SAP при 37°С в течение 30 мин. Затем экзонуклеазу I и SAP инактивировали теплом, инкубируя реакционную смесь при 80°С в течение 15 мин.
После этого добавляли 0,5 ед. урацил-ДНК-гликозилазы и инкубацию продолжали при 37°С в течение 30 мин. После обработки урацил-ДНК-гликозилазой проводили исчерпывающее расщепление в лишенных основания звеньях, созданных в амплифицированном продукте. Для этого добавляли NaOH до конечной концентрации 0,05 М и прогревали смесь в течение 15 мин при 95°С. Затем расщепленную ДНК осаждали добавлением 3 М ацетата натрия (10% от объема смеси) и 2 объемов этанола. Осадок ресуспендировали в 5 мкл воды. После этого расщепленную ДНК обрабатывали 0,5 ед. Т4 полинуклеотид-киназы, которая удаляет с 3'-концов фосфатную группу.
Затем проводили реакцию линейной амплификации, используя продукты предшествующей реакции расщепления, т.е. различные способные к удлинению направленные вверх фрагменты. В ходе амплификации направленный вверх фрагмент удлиняется термостабильной ДНК-полимеразой ("Термосеквеназа" - продукт фирмы Amersham Life Sciences) в циклической реакции в общем объеме 10 мкл. Матрицей в этой реакции служит амплифицированный фрагмент нормальной кДНК (нуклеотиды с 952 по 1224 гена RYR1), очищенный от немеченых праймеров предобработкой экзонуклеазой I. Реакцию удлинения проводили в присутствии 1 мМ каждого из трех концевых дидезоксинуклеотидов (ddNTP) и 0,02 мМ меченого 33Р ddNTP.
К образцу добавляли равный объем формамидного покрывающего красителя (90% формамид, 0,025% бромфенол голубой, 0,025% ксиленцианол), затем образец прогревали при 85°С в течение 5 мин. После этого образец наносили на денатурирующий (7 М мочевина) гель 6% полиакриламида и проводили электрофорез в течение 3-4 ч при 60 В для анализа продуктов удлинения по размерам. После электрофореза проводили авторадиографию, экспонируя гель в контакте с рентгеновской фотопленкой в течение 12 ч при -70°С.
При анализе авторадиограммы для случая, когда меченым ddNTP был ddTTP (фиг.3), обнаружена "лестница" меченых фрагментов, соответствующая расстоянию от 5'-конца праймеров до места включения dUMP (показано удлинение только нижней нити). Эти места включения соответствуют дикому типу "Т-распределения" для ДНК. Если для создания удлиненных праймеров использовали мутантную целевую нуклеиновую кислоту, в Т-распределении линий наблюдали появление дополнительной линии (203 нуклеотида). Анализ авторадиограммы, полученной в случае, когда меченым был ddCTP (фиг.4), не обнаружил меченых линий, если источником направленных вверх фрагментов была амплифицированная нормальная целевая нуклеиновая кислота. Однако если использовали амплифицированную мутантную целевую нуклеиновую кислоту, анализ показывал как раз одну зону. Размер меченого фрагмента соответствует расстоянию между 5'-концом нижнего праймера и мутантным звеном (т.е. 203 нуклеотида), что демонстрирует наличие у данного индивидуума мутации G→A в положении 1021 гена RYR1.
Пример 3
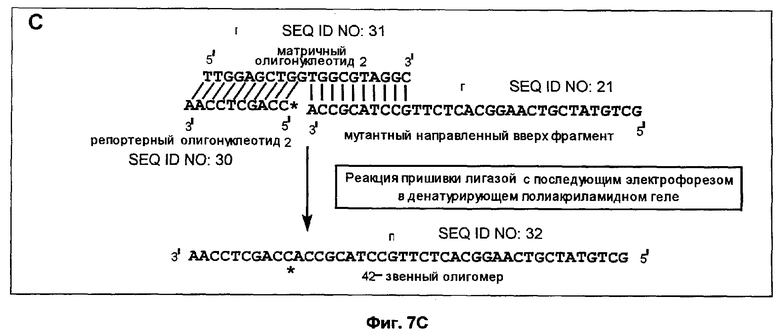
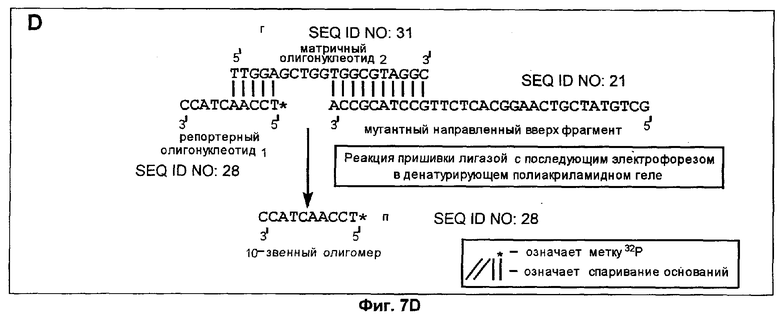
Способ согласно настоящему изобретению был применен для обнаружения присутствия мутации G→A в положении 6411 (кодон 12) гена Ki-ras человека. Амплифицировали геномную ДНК из нормальной ткани и опухолевой ткани индивидуума с раком толстой кишки, используя верхний (нуклеотиды с 6390 по 6409) и нижний (нуклеотиды с 6417 по 6443) праймеры для создания двунитевого фрагмента ДНК размером 54 пары оснований (нуклеотиды с 6390 по 6443). Номера нуклеотидов соответствуют геномной нуклеотидной последовательности гена Ki-ras, включая интроны.
На фиг.5 схематически изображены целевая нуклеиновая кислота и верхний и нижний праймеры (праймеры содержат стандартные основания G, А, Т и С). Целевую нуклеиновую кислоту амплифицировали ПЦР в реакционной смеси, содержащей в общем объеме 19 мкл целевую нуклеиновую кислоту, по 0,2 мМ dATP, dCTP, dGTP и dUTP, а также 6 пикомолей верхнего и нижнего праймеров. На реакционную смесь наливали сверху равный объем минерального масла и осуществляли горячий запуск ПЦР посредством прогревания реакционной смеси при 94°С в течение 5 мин перед добавлением 1 единицы Taq полимеразы (общий объем смеси становился равным 20 мкл). В термоциклере проводили 30 циклов денатурации, гибридизации и удлинения. Реакционную смесь, содержащую амплифицированную целевую нуклеиновую кислоту, обрабатывали затем экзонуклеазой I, чтобы разрушить праймеры, которые не были удлинены в стадии амплификации, и щелочной фосфатазой креветок (SAP), чтобы разрушить dNTPs, не включившиеся на стадии амплификации. Это осуществляли инкубацией 20 мкл реакционной смеси ПЦР с 0,5 ед. экзонуклеазы I и 1 ед. SAP при 37°С в течение 30 мин. После этого экзонуклеазу 1 и SAP инактивировали теплом, инкубируя реакционную смесь при 80°С в течение 15 мин. Затем добавляли 0,5 ед. урацил-ДНК-гликозилазы и 1 ед. эндонуклеазы IV и продолжали инкубацию при 37°С в течение 30 мин, чтобы обеспечить полное выщепление всех имеющихся в амплифицированной ДНК урацилов и полное расщепление во всех полученных лишенных основания звеньях. В результате этого расщепления получали направленные вверх фрагменты, содержащие 3'-гидроксильную группу на своем 3'-конце, который можно удлинить с помощью ДНК-полимеразы.
Затем проводили реакции удлинения в реакционной смеси (10 мкл), содержащей продукты вышеуказанной реакции расщепления, т.е. способные к удлинению фрагменты (2 мкл реакционной смеси после расщепления, приблизительно 1 пикомоль способных к удлинению фрагментов) и 100 фемтомолей синтетического матричного олигонуклеотида, 0,2 мМ каждого из dATP, dTTP и dGTP, 0,02 мМ dCTP, 1 мккюри (α-32P)-dCTP, 6 пикомолей обратного праймера и 1 ед. Taq ДНК-полимеразы. Реакцию продолжали до получения 40 циклов денатурации, гибридизации и удлинения.
К образцу добавляли равный объем формамидного покрывающего красителя (90% формамид, 0,025% бромфенол голубой, 0,025% ксиленцианол), затем образец прогревали при 85°С в течение 5 мин. После этого образец наносили на денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез в течение 3-4 ч при 60 В для анализа продуктов удлинения по размерам. После электрофореза проводили авторадиографию, экспонируя гель в контакте с рентгеновской фотопленкой в течение 12 час. при -20°С.
Анализ ДНК из нормальной ткани приводит к появлению после расщепления с участием гликозилазы и использования указанных выше верхнего и нижнего праймеров способного к удлинению фрагмента длиной 37 нуклеотидов (фиг.5). Подобный анализ ДНК из опухолевой ткани приводит к появлению способного к удлинению фрагмента длиной 32 нуклеотида (фиг.5). Анализ авторадиографа показывает после анализа ДНК из нормальной ткани с использованием матричного олигонуклеотида 1 появление линии, соответствующей длине в 66 нуклеотидов (фиг.6А). Эта линия не наблюдалась, если в указанном выше анализе использовали матричный олигонуклеотид 2 (фиг.6В). Анализ авторадиографа показал наличие линии 66 нуклеотидов после проведения анализа ДНК из опухолевой ткани и использования матричного олигонуклеотида 2 (фиг.6С). Эта линия не наблюдалась, если в указанном выше анализе использовали матричный олигонуклеотид 1 (фиг.6D). Таким образом, наличие мутации в кодоне 12 гена Ki-ras определено по способности направленного вверх фрагмента к удлинению на мутантном матричном олигонуклеотиде, тогда как направленный вверх фрагмент не удлинялся на нормальном матричном олигонуклеотиде. Обратная ситуация была в отсутствие мутации.
Пример 4
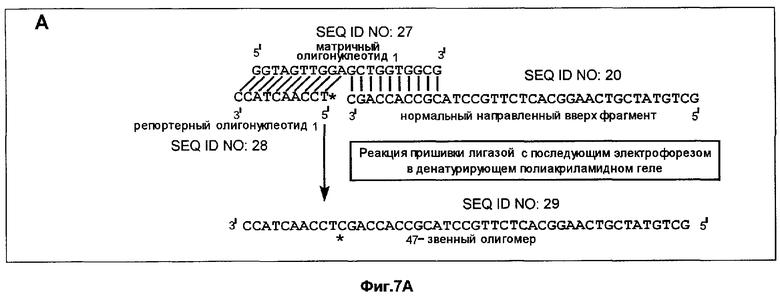
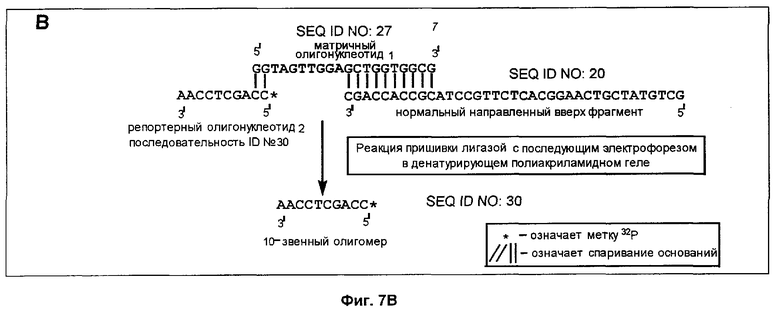
Пример 3 был повторен до стадии, когда было проведено полное расщепление в лишенных основания звеньях. Это расщепление приводило к появлению направленных вверх фрагментов, имеющих, как указано выше, 3'-гидроксильную группу на своем 3'-конце.
Репортерные олигонуклеотиды 1 и 2 (фиг.7) (комплементарные к участкам соответственно с нуклеотидами с 6397 по 6406 (5' ТССААСТАСС 3', №1 (SEQ ID NO: 28)) и с нуклеотидами с 6402 по 6411 (5' CCAGCTCCAA 3', №2 (SEQ ID NO: 30)) гена Ki-ras) были помечены на 5'-концах с помощью (γ-32Р)-АТР и Т4 полинуклеотидкиназы. Это позволяло пометить полученный пришитый лигазой фрагмент и обеспечивало также наличие у репортерного олигонуклеотида 5'-концевого фосфата, необходимого для любой лигазной пришивки.
Реакции пришивки лигазой проводили (при 16°С в течение 16 мин) с использованием продуктов указанной выше реакции расщепления, т.е. способных к удлинению направленных вверх фрагментов (5 мкл смеси из реакции расщепления, 2 пикомоля способных к удлинению фрагментов), 4 пикомолей меченого репортерного олигонуклеотида 1 или 2 (соответственно 5'GGTAGTTGGAGCTGGTGGCG3' (SEQ ID NO: 27) (нуклеотиды с 6397 по 6416) или 5TTGGAGCTGGTGGCGTAGGC3' (SEQ ID NO: 31) (нуклеотиды с 6402 по 6421) (фиг.7) и 1 ед. Т4 ДНК-лигазы. Общий объем реакционной смеси составлял 20 мкл. В ходе реакции направленный вверх фрагмент удлиняется пришивкой репортерного олигонуклеотида.
К образцу добавляли равный объем формамидного покрывающего красителя (90% формамид, 0,025% бромфенол голубой, 0,025% ксиленцианол), затем образец прогревали при 85°С в течение 5 мин. После этого образец наносили на денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез в течение 3-4 ч при 60 В для анализа продуктов удлинения по размерам. После электрофореза проводили авторадиографию, экспонируя гель в контакте с рентгеновской фотопленкой в течение 3 ч при -20°С.
Анализ ДНК из нормальной ткани дает способный к удлинению фрагмент размером 37 нуклеотидов после расщепления с участием гликозилазы и использования указанных выше верхнего и нижнего праймеров (фиг.5). Подобный же анализ ДНК из опухолевой ткани дает способный у удлинению фрагмент длиной 32 нуклеотида (фиг.5). Просмотр авторадиографа после анализа ДНК из нормальной ткани и использования репортерного олигонуклеотида 1 и матричного олигонуклеотида 1 выявляет линию, соответствующую длине 47 нуклеотидов (фиг.7А). Эта линия не обнаруживается, если в указанном выше анализе использован репортерный олигонуклеотид 2 (фиг.7В). Просмотр авторадиографа после анализа ДНК из опухолевой ткани и использования репортерного олигонуклеотида 2 и матричного олигонуклеотида 2 выявляет линию, соответствующую длине 42 нуклеотида (фиг.7С). Эта линия не обнаруживается, если в указанном выше анализе использован репортерный олигонуклеотид 1 (фиг.7D). Таким образом, наличие мутации в кодоне 12 гена Ki-ras было определено по присутствию линии 42 нуклеотида, в то время как наличие нормальной аллели было определено по присутствию линии 47 нуклеотидов. Образцы, содержавшие нормальную и опухолевую ДНК, давали обе линии - и 42, и 47 нуклеотидов.
Описанный выше анализ был проведен также с введением 5'-концевой метки 32P в нижний праймер. Для этого было необходимо, чтобы репортерные олигонуклеотиды были фосфорилированы с использованием немеченого АТР в качестве донора фосфата. Результаты были подобны описанным выше в том, что линия 42 нуклеотида свидетельствовала о наличии мутантного гена Ki-ras (кодон 12), тогда как линия 47 нуклеотидов свидетельствовала об отсутствии мутации в кодоне 12 гена Ki-ras. Кроме того, описанный выше анализ был также выполнен с использованием в качестве матрицы в ходе реакции пришивки лигазой первоначально амплифицированного фрагмента гена Ki-ras из нормального или мутантного образца. Нормальный амплифицированный продукт был использован вместо матричного олигонуклеотида 1, а мутантный амплифицированный продукт был использован вместо матричного олигонуклеотида 2. Вновь были получены такие же результаты, как и описанные выше.
Как указывалось выше, способ по настоящему изобретению имеет много преимуществ перед известными способами, особенно перед способом по заявке WO 97/03210. В заявке WO 97/03210 после расщепления с участием гликозилазы (которое может быть выполнено большим числом способов и которое дает большое число различных 3'-концов) полученные фрагменты ДНК далее не подвергаются обработке и анализируются непосредственно. В настоящем изобретении после этапа расщепления с участием гликозилазы следует этап, позволяющий удлинить 3'-концы, полученные при расщеплении с участием гликозилазы.
Основным преимуществом настоящего изобретения является то, что настоящее изобретение обеспечивает обнаружение различий в нуклеотидной последовательности между молекулами нуклеиновой кислоты, таких как мутации и полиморфизмы, без определения подобия последовательностей, что невозможно осуществить в случае применения известных способов. Как указано выше, способом по заявке WO 97/03210 невозможно обнаружить различия в нуклеотидной последовательности молекул нуклеиновой кислоты без определения совпадений нуклеотидной последовательности. Это является недостатком способа по заявке WO 97/03210, так как нельзя объединить несколько образцов для их одновременного анализа. Настоящее изобретение делает также возможным одновременный анализ многих генов или сегментов генов. Более того, настоящее изобретение позволяет создавать специфические праймеры для амплификации нуклеиновых кислот без необходимости иметь предварительную информацию о нуклеотидной последовательности нуклеиновой кислоты. Известные способы не дают такой возможности. Кроме того, настоящее изобретение позволяет получать из нуклеиновых кислот специфические праймеры единственным путем, и такие праймеры можно затем анализировать путем полимеразного удлинения, чтобы определить природу нуклеотидной последовательности у 3'-концов указанных праймеров.
Изобретение относится к области молекулярной биологии и биохимии и может быть использовано в диагностических системах, предназначенных для выявления в биологических образцах определенных генов, а также известных или неизвестных мутаций этих генов. Предложен способ характеристики последовательности нуклеиновой кислоты путем установления наличия (или отсутствия) в этой последовательности вариаций и определения характера обнаруженных вариаций. Способ предусматривает ферментативное копирование анализируемой последовательности в присутствии, по крайней мере, одного вида дНТФ с модифицированным основанием, являющимся субстратом ДНК-гликозилазы; обработку полученной копии ДНК-гликозилазой и затем агентом, расщепляющим последовательность с образованием фрагментов, которые содержат ОН-группу на 3'-конце, инкубацию этих фрагментов в присутствии матричной нуклеиновой кислоты и фермента, способного обеспечить их наращивание, и анализ продуктов удлинения. Применение предлагаемого способа в диагностических целях повышает эффективность процедуры тестирования. 19 з.п. ф-лы, 7 ил.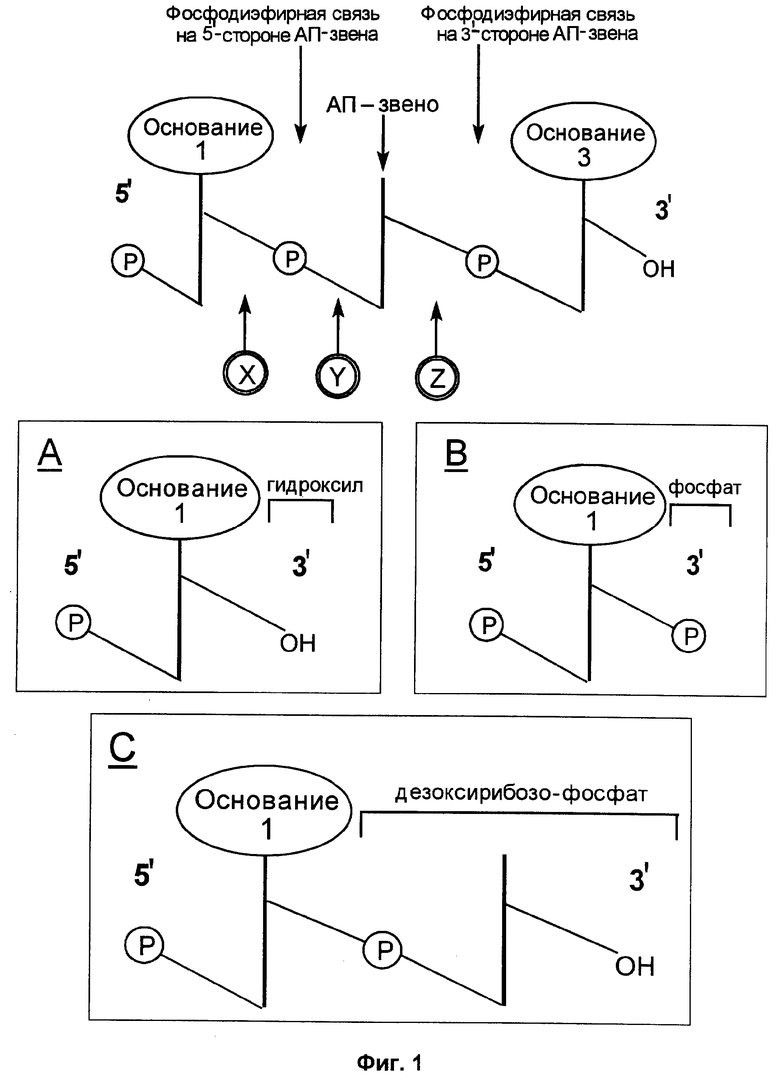
i) ферментативное копирование анализируемой последовательности в присутствии всех необходимых видов дезоксирибонуклеозидтрифосфатов (дНТФ) и, по меньшей мере, одного вида дНТФ, включающего модифицированное основание, которое является субстратом для ДНК-гликозилазы;
ii) обработка копированной последовательности, полученной на стадии (i), ДНК-гликозилазой, обеспечивающей удаление включившихся модифицированных оснований с образованием лишенных основания звеньев;
iii) обработка последовательности, полученной на стадии (ii), обеспечивающая расщепление фосфодиэфирной связи в лишенном основания звене с образованием фрагментов с ОН-группой на 3'-конце или с последующим переводом в указанную форму фрагментов с блокированными 3'-концами, которые способны к последующему удлинению; и iv) инкубация полученных на стадии (iii) фрагментов ДНК в присутствии фермента, обеспечивающего их удлинение, и матричной нуклеиновой кислоты, частично или полностью комплементарной последовательности удлиняемых фрагментов, и анализ полученных после удлинения фрагментов.
| WO 9712061, 03.04.1997 | |||
| WO 9703210, 30.01.1997 | |||
| P.VAUGHAN, V.Mc CARTHY | |||
| Nucl | |||
| Acids | |||
| Res | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| З.А.ШАБАРОВА и др | |||
| Химические основы генетической инженерии | |||
| - М.: Изд-во Московского университета, 1994. | |||

