Изобретение относится к области биотехнологии и, в частности к генетической инженерии и может быть использовано в производстве препаратов против колорадского жука и других насекомых отряда Coleoptera.
Известен штамм бактерий Bacillus thuringiensis - продуцент дельтаэндотоксина Cry IIIA - типа против колорадского жука (SU патент N 1814520 кл. A 01 N 63/00).
Однако данный штамм имеет те же недостатки, что и другие природные штаммы Bacillus thuringiensis.
Биоинсектициды, создаваемые на их основе содержат жизнеспособные споры и кристаллические включения токсина, освобождающиеся при лизисе спорулирующих клеток. Токсин в таких препаратах быстро разрушается под воздействием природных факторов, а внесение в окружающую среду значительных количеств спор может в конечном счете негативно влиять на экологическую обстановку.
Известны штаммы Bacillus thuringiensis, синтезирующие белки, токсичные против насекомых отряда Coleoptera и известны последовательности этих токсинов и кодирующих их генов (ЕР N 0498537 A 2, C 12 N 15/32, 1992 и заявка PCT WO 91/14778, c 12 N 15/32, 1991). В цитируемых документах предлагаются возможные способы использования данных штаммов, генов дельта-эндотоксинов и кодируемых ими продуктов, включая конструирование новых рекомбинантных микроорганизмов.
Однако в этих источниках не содержится подробной информации о способах конструирования новых штаммов и их характеристиках. Кроме того, данные гены кодируют токсины, отличные от токсина Cry IIIA - типа, продуцируемого заявляемым штаммом P. putida.
Mycogen Corporation осуществляет разработку и испытание инкапсулированных биопестицидов на основе убитых клеток Pseudomonas fluoresoens, в том числе клеток с Cry IIIA - белком. Однако конкретные данные относительно системы регуляции и уровня экспрессии клонированных Cry-генов в рекомбинантных штаммах псевдомонад не сообщаются.
(Feitelson J. , Payne J., Kim L.// Bio/ Technol., 1992, v.10, p. 271 - 275).
Однако конкретные данные относительно системы регуляции и уровня экспрессии клонированного cry-гена в рекомбинантных штаммах псевдомонад, не сообщаются.
Задача предлагаемого изобретения - конструирование рекомбинантной плазмидной ДНК, кодирующей синтез инсектицидного белка и создание штамма Pseudomonas putida - продуцента кристаллического дельта-эндотоксина, активного против колорадского жука и других насекомых отряда Coleoptera.
Создана плазмида pBTN11, кодирующая дельта - эндотоксин Cry IIIA -типа и способная поддерживаться в клетках различных грамотрицательных бактерий, состоящая из следующих элементов:
полной последовательности сконструированной векторной плазмиды pBTN4 (размер 14,8 т.п.н.) с широким кругом бактериальных хозяев;
полной последовательности HindIII-фрагмента ДНК B.thuringiensis subsp. tenebrionis (размер 3,0 т.п.н.) и содержит:
интактный ген дельта-эндотоксина B.thuringiensis subsp. tenebrionis;
ген устойчивости к канамицину (Kmr);
гены, ответственные за мобилизацию плазмиды при конъюгативном переносе (oriT, mob);
промотор гена cryIA(b) B. thuringiensis subsp. berliner 1715;
промотор Pm оперона мета-пути деградации ароматических углеводородов плазмиды pWWO;
ген xylS, участвующий в позитивной регуляции этого оперона;
сайты расщепления эндонуклеазой PstI с координатами 0; 5,3; 5,7, 7,1 и 17,8 т.п.н.;
сайты расщепления эндонуклеазой HindIII с координатами 2,4, 6,5 и 9,5 т. п.н.;
сайты расщепления эндонуклеазой BamHI с координатами 3,2; 4,8 и 6,0 т.п. н.;
сайты расщепления эндонуклеазой BglII с координатами 3,6 и 9,4 т.п.н.;
сайты расщепления эндонуклеазой EcoRI с координатами 6,0; 8,1; 8,8 и 9,8 т.п.н.;
Размер плазмиды pBTN11 равен 17,8 т.п.н.
Способ конструирования плазмиды pBTN11, кодирующей синтез Coleoptera-специфического эндотоксина, заключающейся том, что ДНК плазмиды pBTO22, несущей ген cryIA(b) B. thuringiensis subsp. berliner 1715, расщепляют эндонуклеазой Clal, смешивают с ДНК плазмиды pC194, подвергнутой частичному гидролизу Taql, фрагменты ДНК соединяют с помощью ДНК-лигазы и полученной смесью трансформируют клетки Escherichia coli и трансформанты высевают на агаризованную среду, содержащую ампициллин (Ap) и хлорамфеникол (Cm), и из клеток устойчивых к Am и Cm клонов выделяют ДНК плазмиды pBTO3, в структуре которой меньший Clal-фрагмент плазмиды pBTO22 заменен на фрагмент pC194, состоящий из трех Taql (A, D и E) фрагментов, ДНК плазмиды pBTO3 и экспрессирующего вектора с широким кругом бактериальных хозяев - плазмиды pNM185 - гидролизуют EcoRI, продукты гидролиза соединяют с помощью ДНК-лигазы и после трансформации клеток E.coli клоны, содержащие гибридную плазмиду, отбирают по устойчивости к канамицину и хлорамфениколу, отобранные клоны проверяют на отсутствие индуцируемой 3-метилбеназоатом (0,5 - 1 мМ) устойчивости к стрептомицину и из полученных клонов выделяют ДНК плазмиды pBTN3, в структуре которой ориентации промоторов Pm и cryIA(b) - гена совпадают, ДНК плазмиды pBTN3 подвергают частичному гидролизу эндонуклеазой ClaI и обрабатывают ДНК-лигазой фага T4, полученным препаратом трансформируют клетки E.coli и отбирают клоны, устойчивые к канамицину, но неспособные расти на среде с хлорамфениколом, из этих клонов выделяют ДНК плазмиды pBTN4, затем ДНК плазмиды pUC19 и B. thuringiensis subsp. tenebriones расщепляют HindIII, образовавшиеся фрагменты лигируют, трансформируют лигазной смесью клетки E.coli JM103 и отбирают бесцветные колонии на среде Мак-Конки с ампициллином, отобранные клоны проверяют на наличие последовательностей ДНК, гомологичных соответствующим последовательностям cryIIIA-гена B. thuringiensis subsp. tenebrionis, из клонов, содержащих эти последовательности, выделяют ДНК плазмиды PBTT51 с cryIIIA-геном, смесью ДНК pBTT51 и pBTN4, последовательно обработанной эндонуклеазой HindIII и ДНК-лигазой, трансформируют клетки E.coli, отбирают устойчивые к канамицину клоны, проверяют их на индуцируемую 3-метилбеназоатом устойчивость к стрептомицину, и из клеток чувствительных к стрептомицину клонов выделяют плазмиду pBTN11, содержащую HindIII-фрагмент c cryIIIA-геном в прямой ориентации к промотору Pm.
Создан штамм:
Pseudomonas putida IPM-36, содержащий рекомбинантную плазмиду ДНК pBTNII - продуцент кристаллического инсектицидного дельта-эндотоксина Cry IIIA - типа, активного против насекомых отряда Coleoptera. Штамм депонирован в Центральной коллекции микроорганизмов Российского акционерного общества "Биопрепарат", регистрационный номер - ЦКМ В - 63 И
Штамм P.putida IPM-36 получен в результате трансформации плазмиды pBTN11 в клетки штамма E.coli S17 - 1 и последующего конъюгационного переноса этой плазмиды из E.coli S17-1 (pBTN11) в клетки штамма P. putida BS1356.
Полученный штамм характеризуется следующими признаками.
Культурно-морфологические признаки.
Клетки палочковидные, размером (2,0 - 3,2) х (0,7 - 0,9) мкм, подвижные, перетрихи, грамотрицательные, спор не образуют. (Размеры клеток даны для односуточной культуры, выращенной на МПА при 30oC.)
Штамм хорошо растет на следующих средах: МПБ (мясопептонный бульон), LB (триптон - 10 г/л, дрожжевой экстракт - 5 г/л, хлористый натрий - 10 г/л, pH 7,2), Кинга B (пептон - 20 г/л, глицерин - 10 мл/л, MgSO4 x 7H2O - 1,5 г/л), питательном агаре на основе гидролизата кильки (ДИПС, г. Махачкала) - 35 г/л, на минимальной синтетической среде M9 следующего состава: K2HPO4 - 7 г/л; KH2PO4 - 3 г/л; NH4Cl - 1 г/л; глюкоза - 1 г/л.
На мясо-пептонном агаре колонии гладкие, блестящие, с ровными краями. В мясо-пептонном бульоне наблюдается помутнение, осадок и желто-зеленый флуоресцирующий пигмент.
На стандартных средах Кинга продуцирует желто-зеленый флуоресцирующий пигмент, феназиновые пигменты не образует.
Физиолого-биохимические признаки.
Облигатный аэроб, температурный оптимум роста 28-30oC, не растет при 42oC, оптимум pH 6,8 - 7,6.
Нитраты редуцирует, молоко не пептонизирует, не использует в качестве источников углерода трегалозу, и д-коилозу, утилизирует этиловый спирт.
Ассимилирует аммонийный и нитратный азот. В качестве источника углерода использует д-глюкозу, д-фруктозу, сахарозу, мальтозу, лактозу, ацетат, пропинат, бутират, сукцинат, пируват, фумарат, бензоат, лактат, цитрат, катехол, протокатехоат.
Оксидазная реакция положительная.
Аргининдегидролаза присутствует.
Устойчивость к антибиотикам.
Проявляет устойчивость к канамицину (50-100 мкг/мл) и стрептомицину (200 мкг/мл), обусловленную присутствием плазмиды pBTN11, а также устойчивость к ампициллину (500 мкг/мл).
Условия хранения штамма.
Штамм может храниться в лиофильно-высушенном состоянии или в 0,6%-агаризованной среде (Nutrient Broth (Difco) - 4 г/л, NaCl - 5 г/л, агар - 6 г/л, pH 6,8) под вазелиновым маслом.
Штамм не патогенен для теплокровных животных.
Полученный штамм Pseudomonas putida IPM-36- продуцент дельта-эндотоксина обладает следующими преимуществами перед B. thuringiensis:
возможность защиты энтомотоксина от действия факторов внешней среды клеточной стенкой продуцента в связи с его внутриклеточной локализацией;
уменьшение возможных отрицательных последствий для окружающей среды при использовании препаратов на основе инактивированных вегетативных клеток по сравнению с препаратами, содержащими кристаллы и споры;
повышение продуктивности и технологичности при производстве.
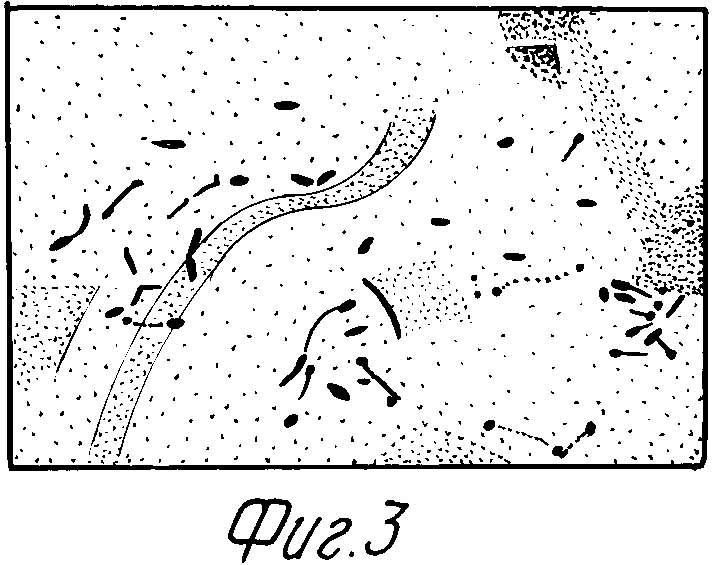
На фиг. 1 представлена схема получения векторной плазмиды pBTN4; на фиг. 2 - карта плазмиды pBTN11; на фиг. 3 - клетки P. putida IPM-36 с внутриклеточными включениями энтомоцидного белка.
Пример 1. Конструирование рекомбинантной плазмиды pBTN11.
Выделяют плазмидные ДНК из клеток E.coli и Bacillus cereus с последующей очисткой в градиенте плотности хлористого цезия в присутствии бромистого этидия.
ДНК плазмиды pBTO22 гидролизуют рестрикционой эндонуклеазой ClaI при 37oC в буфере A (10 мм трис-HCl, pH 7,5, 10 мМ MgCl2, 1 мМ дитиотрейтол). ДНК плазмиды pC194 подвергают частичному расщеплению рестриктазой Taql при 65oC в том же буфере. В инкубационную смесь вносят 5 М раствор NaCl до конечной концентрации 0,1 М, ДНК осаждают добавлением двух объемов этанола и растворяют в буфере ТЕ (10 мМ трис-HCl, pH 7,4, 1 мМ ЭДТА, pH 8,0).
Гидролизаты ДНК pBTO22 и pC194 смешивают в соотношении 1:5, добавляют буфер для лигирования (конечная концентрация - 66 мМ трис-HCl, pH 7,5, 5 мМ MgCl2, 5 мМ дитиотрейтол, 1 мМ АТФ) и ДНК-лигазу фага Т4 (1 ед/мкг ДНК). Концентрация ДНК составляет 50-100 мкг/мл, объем инкубационной смеси - 50 мкл. Реакцию проводят при 8oC в течение ночи. Полноту гидролиза и лигирования ДНК контролируют с помощью электрофореза в геле 0,7% агарозы.
Для получения компетентных клеток, культуру клеток Escherichia coli DH5 α выращивают в среде LB при 37oC до середины логарифмической фазы роста. Клетки осаждают центрифугированием. (Эту и дальнейшие операции проводят при 2-4oC). Осадок ресуспендируют в 1/2 первоначального объема 10 мМ раствора CaCl2, выдерживают 20 мин и после центрифугирования ресуспендируют в 1/50 первоначального объема 50 мМ CaCl2. Через 12-24 ч хранения при 2-4oC 0,2 мл суспензии смешивают с 10 мкл раствора лигированной ДНК (10-50 мкг/мл) и инкубируют 30-60 мин на льду. Трансформационную смесь переносят на 2 мин в водяную баню при 42oC, добавляют к суспензии 1 мл LB, инкубируют 1 ч при 37oC и высевают на агаризованную среду LB с хлорамфениколом (5 мкг/мл).
Чашки инкубируют при 37oC в течение 20-24 ч, выросшие колонии отбирают, из полученных клонов выделяют плазмидные ДНК и проводят рестрикционный анализ выделенных плазмид. Рекомбинантную плазмиду, в которой ClaI-фрагмент cryIA(b) гена в плазмиде pBTO22 заменен на фрагмент ДНК pC194, состоящий из трех TaqI (A, D и E) фрагментов, с прямой ориентацией гена хлорамфениколрезистентности (CAT) по отношению к промотору cryIA(b) детерминанта, обозначают как pBTO3.
ДНК плазмид pBTO3 и pNM185 (по 2 мкг) смешивают и обрабатывают эндонуклеазой EcoRI в 50 мкл буфера B (10 мМ трис-HCl, pH 7,5, 50 мМ NaCl, 10 мМ MgCl2, 1 мМ дитиотрейтол). Лигирование продуктов гидролиза, получение компетентных клеток E. coli DH5lgЛК50 = lgCм-σ(ΣL-0,5), и трансформацию этих клеток рекомбинантными молекулами ДНК проводят как описано выше. Суспензию трансформированных клеток высевают на агаризованную среду LB с канамицином (50 мкг/мл) и хлорамфениколом (5 мкг/мл).
Выросшие колонии перекалывают, дублируя чашки с этой средой и средой LB, содержащей канамицин (50 мкг/мл), хлорамфеникол (5 мкг/мл), стрептомицин (100 мкг/мл) и 3-метилбензоат (68 мкг/мл), и после выращивания в течение 20-24 ч при 30oC отбирают колонии клонов, способных к росту только на среде без стрептомицина. Из клеток этих клонов выделяют плазмидные ДНК и проводят рестрикционный анализ. Плазмиду, в которой ориентация промотора Pm и cryIA(b) гена совпадают, обозначают как pBTN3.
ДНК плазмиды pBTN3 подвергают частичному гидролизу рестриктазой ClaI, обрабатывают ДНК-лигазой и трансформируют компетентные клетки E. coli DH5σ как описано выше. Клетки трансформантов высевают на агаризованную среду LB с канамицином (50 мкг/мл), и выросшие колонии перекалывают, дублируя чашки с этой средой и средой, содержащей канамицин (50 мкг/мл) и хлорамфеникол (5 мкг/мл).
Из колоний клонов, не способных к росту на среде с хлорамфениколом, выделяют плазмидные ДНК и анализируют их структуру.
Плазмиду, представляющую собой укороченный вариант плазмиды pBTN3 с делецией ClaI-фрагмента, содержащего ген CAT, обозначают как pBTN4.
Из клеток штамма Bacillus thuringiensis subsp. tenebrionis, синтезирующего энтомоцидный белок против личинок колорадского жука, выделяют ДНК. Бактерии выращивают в 1 л среды LB до поздней логарифмической фазы роста при 30oC, клетки собирают центрифугированием, суспендируют в 50 мл TES-буфера (0,01 М Трис-HCl, pH 8,0, 0,001 М ЭДТА, 0,1 М NaCl) и добавляют 5 мл раствора лизоцима (20 мг/мл). Суспензию клеток инкубируют при 37oC в течение 30 мин, затем добавляют 3 мл 20% раствора додецилсульфата натрия. Лизат экстрагируют дважды равным объемом смеси хлороформа и изоамилового спирта (24: 1). Препарат ДНК обрабатывают РНК-азой (конечная концентрация 50 мкг/мл) в течение 1 ч при 37oC и осаждают этанолом. Чистоту и концентрацию препаратов ДНК определяют спектрофотометрически. Затем смешивают 0,1 мкг ДНК pUC19, выделенной из клеток Escherichia coli, и 2 мкг ДНК B. thuringiensis subsp. tenebrionis и гидролизуют рестриктазой HindIII (10 ед) в 50 мкл буфера Б, как указано выше для фермента EcoRI, при 37oC в течение 2 ч. Полученные фрагменты ДНК осаждают спиртом, перерастворяют, лигируют и трансформируют компетентные клетки E. coli JM103, как описано выше. Клетки высевают на среду МакКонки (Difco) с ампициллином и отбирают бесцветные колонии.
Из клеток отобранных колоний выделяют плазмидные ДНК и, используя эти ДНК в качестве матриц, проводят полимеразную цепную реакцию (ПЦР). Праймерами в ПЦР служат олигодезоксинуклеотиды, имеющие следующую структуру: 5' - GGTTCCAACCAGGATATT - 3' и 5' - CAGACCGCAAGATTTGAT - 3'. Первый соответствуют фрагменту cryIIIA - гена B. thuringiensis subsp. tenebrionis с 1034 по 1051 нуклеотид и второй комплементарен последовательности этого детерминанта с 1215 по 1232 нуклеотид. Последовательности ДНК, исследуемые на наличие детерминанта Coleoptera-специфического инсектицидного белка, амплифицируют in vitro с помощью полимеразной цепной реакции в 50 мкл реакционной смеси, содержащей ДНК (около 10 нг), праймеры (концентрация каждого праймера - 0,3 мкм), 60 мМ трис-HCl, 16 мМ (NH4SO4, 1,5 мМ MgCl2, 1 мМ дитиотрейтол, 0,01% тритона X-100, 0,01% Твин-20, бычий сывороточный альбумин (0,1 мг/мл), смесь дезоксинуклеозидтрифосфатов (дАТФ, дГТФ, дЦТФ и дТТФ - по 0,2 мМ) и 2 ед. Tth-полимеразы; pH смеси - 8,8 при 25oC. Реакцию амплификации проводят под вазелиновым маслом в течение 30 циклов: 94oC - 1 мин, 45oC - 2 мин и 67oC - 2 мин.
Продукты ПЦР разделяют с помощью электрофореза в геле 2,5% агарозы и отбирают клоны, содержащие плазмиды, являющиеся матрицей для синтеза фрагментов ДНК размером около 0,2 т.п.н. при использовании в ПЦР, представленных выше праймеров. Плазмиду, в структуре которой ген дельта-эндотоксина входит в состав HindIII-фрагмента размером около 3 т.п.н. и находится в прямой ориентации по отношению к lac-промотору, обозначают как pBTT51.
ДНК плазмид pBTT51 и pBTN4 смешивают в соотношении 10:1 (общее количество - 2 мкг) гидролизуют эндонуклеазой HindIII, полученные фрагменты объединяют с помощью ДНК-лигазы фага T4 и трансформируют рекомбинантными молекулами ДНК CaCl2-обработанные клетки E. coli DH5ΣL как описано выше. Трансформированные клетки высевают на агаризованную среду LB с канамицином (50 мкг/мл) и выросшие через 20-24 ч инкубации при 37oC колонии перекалывают, дублируя, на чашки с этой средой и средой, содержащей дополнительно стрептомицин (100 мкг/мл) и 3-метилбензоат (68 мкг/мл). Чашки инкубируют при 30oC в течение 20-24 ч. Из клеток трансформантов, не способных к росту на среде с стрептомицином и 3-метилбензоатом, выделяют плазмидные ДНК и проводят рестрикционный анализ выделенных ДНК. Плазмиду, содержащую HindIII-фрагмент с геном энтомотоксина в прямой ориентации к промотору Pm обозначают как pBTN11.
Схема конструирования векторной плазмиды pBTN4 и карта плазмиды pBTN11 представлены на фиг. 1 и 2.
Пример 2. Получение штамма-продуцента.
Для получения на основе штамма Pseudomonas putida BS1356 продуцента, способного к индуцированному синтезу энтомоцидного белка в виде "телец включения" конструируют донорный штамм E. coli S17-1 (pBTN11), из которого плазмида pBTN11 может быть передана путем конъюгации в клетки различных грамотрицательных бактерий. В состав генома E. coli S17-1 входят гены tra-системы плазмиды RP4 с этой целью ДНК плазмиды pBTN11 трансформируют компетентные клетки E. coli S17-1, полученные как описано в примере 1. Отбор трансформантов проводят на агаризованной среде LB, содержащей канамицин (50 мкг/мл).
Скрещиваемые штаммы E. coli S17-1 (pBTN11) и P. putida BS1356 выращивают в течение ночи при 30oC. Культуры донора и реципиента смешивают в соотношении 1:10 (общий объем 1 мл). Суспензию клеток фильтруют через нитроцеллюлозный фильтр (средний диаметр пор - 0,45 мкм, диаметр фильтра - 25 мм) и фильтр помещают на поверхность агаризованной среды LB. Инкубацию проводят 16 - 20 ч при 30oC, затем клетки смывают с фильтра 1 мл 0,9% раствора NaCl и для отбора трансконъюгантов высевают на чашки со средой, содержащей канамицин (50 мкг/мл) и ампициллин (200 мкг/мл).
В клетках трансконъюгантов определяют продукцию дельта-эндотоксина с помощью твердофазного иммуноферментного анализа (ИФА). Для построения калибровочной кривой в ИФА используют очищенный препарат кристаллического белка B. thuringiensis subsp. tenebrionis. Результаты выражают в процентах от общего содержания клеточного белка, растворимого в 0,1 М NaOH, определяемого по Лоури. В качестве стандартов используют серию разведений кристаллического белка B. thuringiensis subsp. tenebrionis и бычьего сывороточного альбумина.
Пример 3. Для получения дельта-эндотоксина свежую 16 ч ночную культуру штамма P. putida IPM-36, полученную инкубированием в среде LB при 30oC разводят в 100 раз в среде LB с канамицином (25 мкг/мл), растят в течение 4 ч при 30oC, добавляют 0,5 мМ индуктора 3-метилбенарата и продолжают инкубировать в тех же условиях еще 16 ч. Количество дельта-эндотоксина, синтезируемого клетками штамма P. putida IPM-36 в указанных выше условиях составляет 500-700 мг/л или 62% от общего растворимого клеточного белка. Дельта-эндотоксин локализуется в клетках в виде "телец-включения" (фиг. 3).
Пример 4. Оценку инсектицидной активности штамма P. putida IPM-36 проводят на личинках колорадского жука (Leptinotarsa decemlineata) первого или второго возраста, отродившихся из одной природной популяции.
Бактериальную культуру разводят в 5 раз 0,9% NaCl и готовят серию из пяти разведений культуры с шагом 5. В качестве положительного контроля используют серию разведений споро-кристаллической смеси B. thuringiensis subsp. tenebrionis, которую получают инкубированием этой бациллы при 30oC сначала в среде LB в течении 16 ч, а затем, после разведения в 40 раз, в среде NB или другой среде для споруляции еще в течении 48 ч. В приготовленные суспензии опускают листья картофеля примерно одного размера, вынимают, дают им высохнуть при 20oC и помещают в чашки Петри. В качестве отрицательного контроля используют раствор 0,9% NaCl. На каждое разведение используют по 10 личинок жука и по 2-5 листьев картофеля. Учет гибели личинок проводят через 3 сут инкубирования в термостате при температуре 21oC и фотопериоде 18 ч.
Биологическую активность штаммов, выраженную в ЛК50, вычисляют по формуле Кербера в процентах концентрации культуральной жидкости (КЖ) в суспензии:
α,
где Cм - максимальная из испытанных концентраций;
ΣL - логарифм отношения каждой предыдущей концентрации к последующей (логарифм кратности разведения);
σ - сумма значений L (доли погибших личинок от числа испытуемых), найденных для всех концентраций с учетом поправки на гибель в контроле по методу Аббота:
L = (pо - pk) : (1-pk),
где pо - доля погибших личинок при испытании культуры;
pk - доля погибших личинок в контрольном опыте.
Штамм P. putida IPM-36 по своей инсектицидной активности (ЛК50 равна 0,13% КЖ) не уступает штамму Bacillus thuringiensis subsp. tenebrionis (ЛК50=0,48% КЖ) и даже превосходит его.
Предлагаемое изобретение позволяет получить новые энтомопатогены на основе грамотрицательных бактерий, в том числе микроорганизмов, колонизирующих растения, и, в частности псевдомонад, широко представленных в эпифитной микрофлоре. Использование таких штаммов, благодаря их высокой колонизирующей активности и внутриклеточной локализации инсектицидного белка, предохраняющей его от действия повреждающих факторов внешней среды, позволяет снизить количество вносимого энтомоцидного препарата и уменьшить число обработок.
Положительный эффект предлагаемого изобретения достигается за счет свойств сконструированной новым способом плазмиды pBTN11, наличию в ее структуре детерминанта инсектицидного белка CryIIIA-типа и генетических элементов, определяющих ее способность к передаче путем мобилизации широкому кругу грамотрицательных бактерий, автономной репликации и поддержанию в клетках этих микроорганизмов.
Возможность индуцированного синтеза целевого продукта, благодаря включению его детерминанта под контроль промотора Pm, позволяет также использовать штаммы Pseudomonas spp. , в том числе предлагаемый штамм P. putida IPM-36, для создания энтомоцидных препаратов против колорадского жука и ряда других насекомых отряда Coleoptera на основе инактивированных клеток бактерий, содержащих большое количество энтомоцидного белка в виде внутриклеточных включений. Уровень продукции дельта-эндотоксина в клетках P. putida IPM-36 достигает 62% от общего клеточного белка при сохранении его биологической активности (ЛК50 препаратов дельта-эндотоксина, синтезированных в клетках P. putida IPM-36 и B. thuringiensis subsp. tenebrionis равны соответственно 0,13% КЖ и 0,48% КЖ). Существующие способы фиксации бактериальных клеток могут сохранять интактной клеточную оболочку, защищающую энтомоцидный белок от действия внешних факторов.
Другими положительными свойствами предлагаемого штамма P. putida IPM-36 по сравнению с известными продуцентами энтомотоксинов - штаммами Bacillus thuringiensis - являются возможность получения целевого продукта за более короткий период времени (16 - 20 и 40 - 48 ч соответственно), существование для псевдомонад более дешевых питательных сред и технологий их выращивания, с выходом биомассы, значительно превышающим продуктивность культур Bacillus thuriensis.

Использование: биотехнология, получение средств защиты растений в отношении жесткокрылых насекомых на основе рекомбинантных грамотрицательных бактерий. Сущность изобретения: Сконструирована рекомбинантная плазмидная ДНК pBT N 11, определяющая синтез кристаллического инсектицидного дельта-эндотоксина Cry IIIA - типа и на ее основе получен штамм бактерий Psendomonas putida, содержащий вышеуказанную плазмиду, продуцирующий дельта-эндотоксин Cry IIIA - типа, активный в отношении насекомых отряда Coleoptera. 3 с.п. ф-лы, 3 ил.
| EP, патент, 0498537, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
