Изобретение относится к области иммунологии, в частности к получению и использованию моноклональных антител. Более конкретно, оно относится к моноклональным анти-IgM-антителам, гибридомам, которые продуцируют эти антитела, иммунохимическим агентам, получаемым из этих антител, и их использованию.
Со времени доклада Кохлера и Милстейна, в котором было описано получение моноклональных антител, развитие технологии получения лимфоцитов, способных продуцировать антитела определенной заранее специфичности, было направлено на развитие как прикладных, так и фундаментальных научных исследований.
Антитела являются эндогенными протеинами, продуцируемыми иммунной системой в ответ на антигенное стимулирование. Эти протеины специфически связываются с молекулами антигена в определенных участках (эпитопах). Поликлональные антитела получают при помощи иммунизации животных антигенами, такие антитела связываются с молекулой антигена в нескольких участках (эпитопах). Моноклональные же антитела представляют собой вполне определенное множество антител, которые получают из одного клона (моноклона) клеток, продуцирующих специфическое антитело. В отличие от поликлональных антител, моноклональные антитела связываются только с одним специфическим эпитопом на молекуле антигена.
Хотя технология продуцирования моноклональных антител достаточно хорошо разработана, современные методы все же остаются весьма длительными по времени, трудоемкими и часто приводят к продуцированию антител, которые, хотя и являются специфическими относительно целевого антигена, обладают относительно низким сродством к нему, и, таким образом, имеют ограничения при использовании.
Среди трудностей, возникающих при получении эффективных и клинически полезных моноклональных антител, важнейшая заключается в обилии антител подтипа IgM, получаемых из гибридом, продуцированных с использованием стандартных процедур иммунизации in vivo или in vitro. Антитела подтипа IgM обладают в общем случае низким сродством, трудно поддаются очистке и часто содержат множество антител, продуцируемых гибридомами. Кроме того, в смешанных культурах клеток гибридом, секретирующих IgM и IgG, клетки, секретирующие IgM, часто перерастают гибридные клетки, секретирующие IgG.
Частью трудоемкой процедуры получения гибридом является удаление клеток гибридомы, продуцирующих IgM, полученных после слияния клеток. Это в общем случае делают путем клонирования клеток при помощи ограниченного разбавления, выращивания отдельных клеток в колониях и испытания каждой колонии отдельно с целью определения, какие колонии продуцируют антитела подтипа IgG. Обычно клетки гибридомы, продуцирующие IgG, затем подвергают последующему анализу с тем, чтобы определить антигенную специфичность продуцируемых антител.
Есть сообщения об антителах, которые направлены против эпитопов на IgM-антителе, но все эти антитела, испытанные до настоящего времени, оказались также химически активными и с антителами подтипа IgG.
Заявителем, а также другими авторами предложены различные цитотоксичные агенты, связывающиеся с антителами с образованием так называемых иммунотоксинов. В последнее время интерес исследователей был направлен в основном на иммунотоксины, в которых моноклональные антитела конъюгированы с ферментативно активными участками (A-цепями) токсинов бактериальной или растительной природы через гетеро-бифункциональные агенты ( Nevelle, D.M. и Youle, R.J., Immunol Rev (1982) т. 62, стр. 75-91, Ross, W.C.J. и др. European J. Biochem. (1980), стр. 104, Vitteta, E.S. и др. Immunol. Rev. (1982), т. 62, стр. 158-183, Raso, V. и др. Cancer. Res. (1982), т. 42, стр. 457-464, Trowbridge, I.W. и Domingo D.L., Nature (Лондон) (1981), т. 294, стр. 171-173.
Настоящее изобретение относится к получению и использованию моноклональных антител крысы, которые:
(a) селективно связываются с антителами подтипа IgM;
(b) являются IgG;
(c) не связываются с подтипами IgG1 или IgG2.
(Предпочтительный вариант осуществления этих антител 2G10 и его функциональные эквиваленты).
Более конкретно, настоящее изобретение относится к иммунотоксинам, получаемым при помощи конъюгации
(a) описанных выше моноклональных антител и
(b) цитотоксичной составляющей или магнитных шариков.
В соответствии с еще одним аспектом, настоящее изобретение относится к способу подавления секреции лимфоцитами IgM-антител при помощи контактирования этих клеток с цитоцидально эффективным количеством одного или нескольких описанных выше иммунотоксинов.
К дополнительным аспектам настоящего изобретения относятся прямые или косвенные иммуноанализы, направленные на определение клеток, продуцирующих IgM-антитела, или на выявление антител IgM-изотипа. Эти анализы включают инкубирование клеток с моноклональными антителами или его мечеными производными. Если используют меченые производные, присутствие меченых бинарных иммунных комплексов на клетках считывается непосредственно. Если используют немеченое антитело, клетки далее инкубируют с меченым антителом против моноклонального антитела и выявляют присутствие меченых третичных комплексов на клетках.
Фиг. 1 и 2 демонстрирует тестирование надосадочных слоев гибридомы на IgM-специфическое антитело.
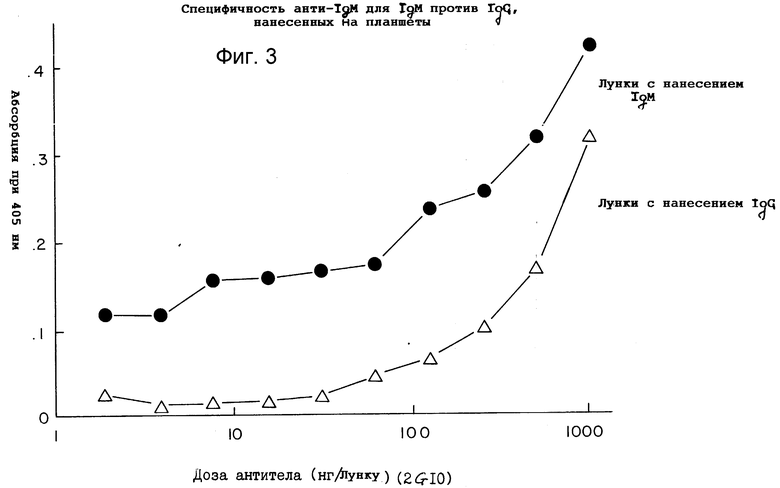
Фиг. 3 демонстрирует специфическое связывание антитела 2G10 в зависимости от дозы.
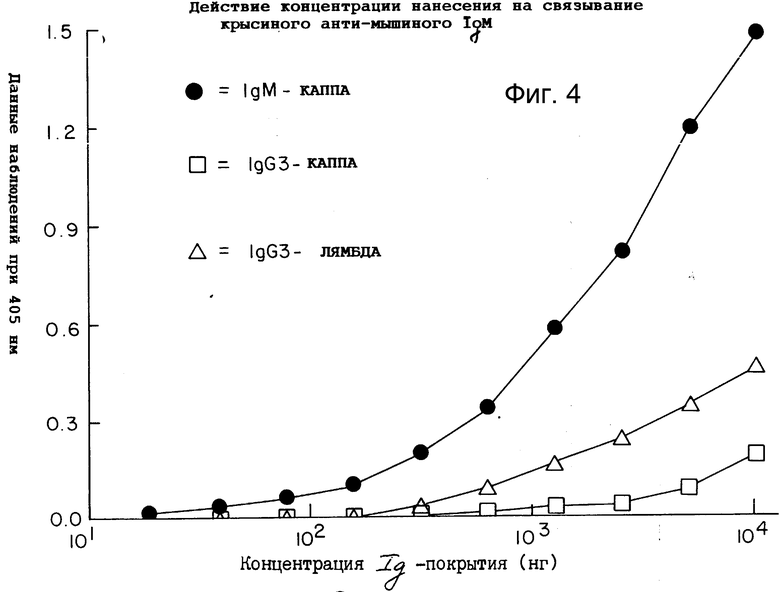
Фиг. 4 демонстрирует действие повышения содержания абсорбированного иммуноглобулина мыши при связывании 2G10.
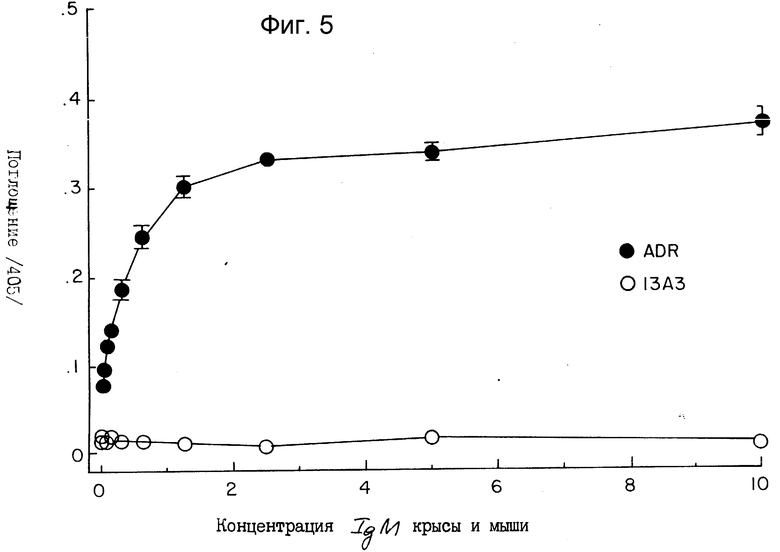
На фиг. 5 показано селективное распознавание IgM мыши прямым твердофазным иммуноферментным анализом (ELISA) с помощью 2G10.
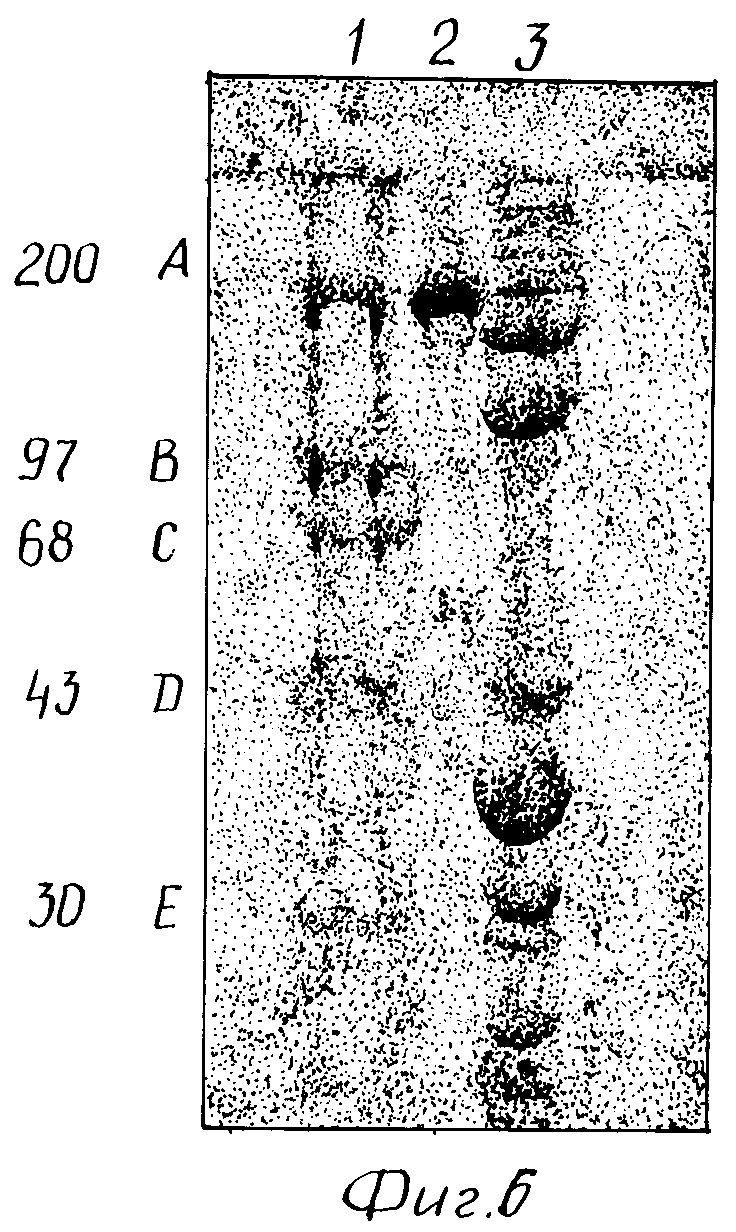
На фиг. 6 показана чистота антитела 2G10 с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PACE).
Фиг. 7 характеризует подкласс крысиного антитела 2G10 иммунодиффузией Ouchterlony.
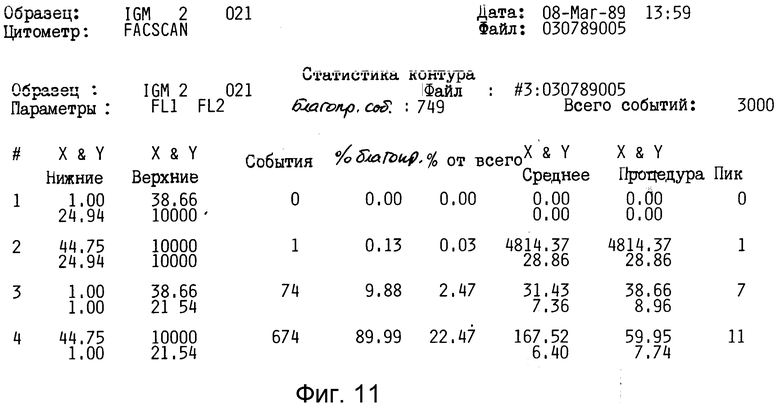
Фиг. 8-11 демонстрируют использование 2G10 для связывания с клетками, экспрессирующими антитела подкласса IgM.
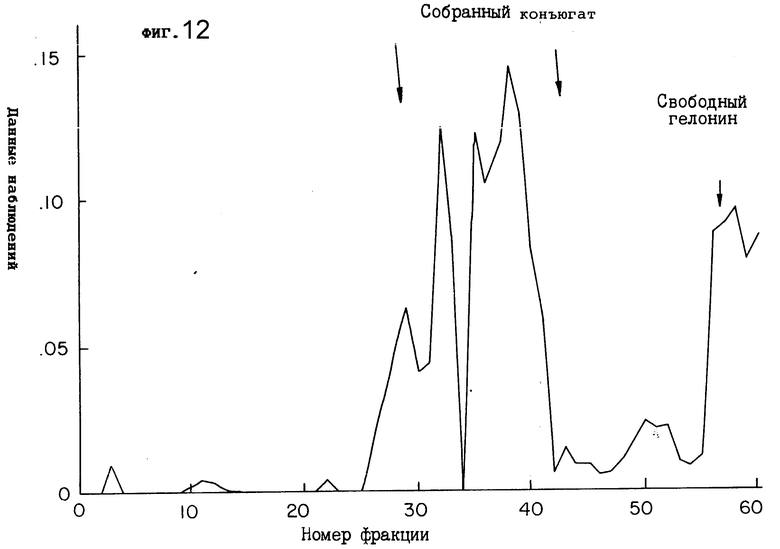
Фиг. 12 иллюстрирует профиль элюирования иммунотоксина (полученного от связывания 2G10 с гелонином) на матрице для гель-фильтрации.
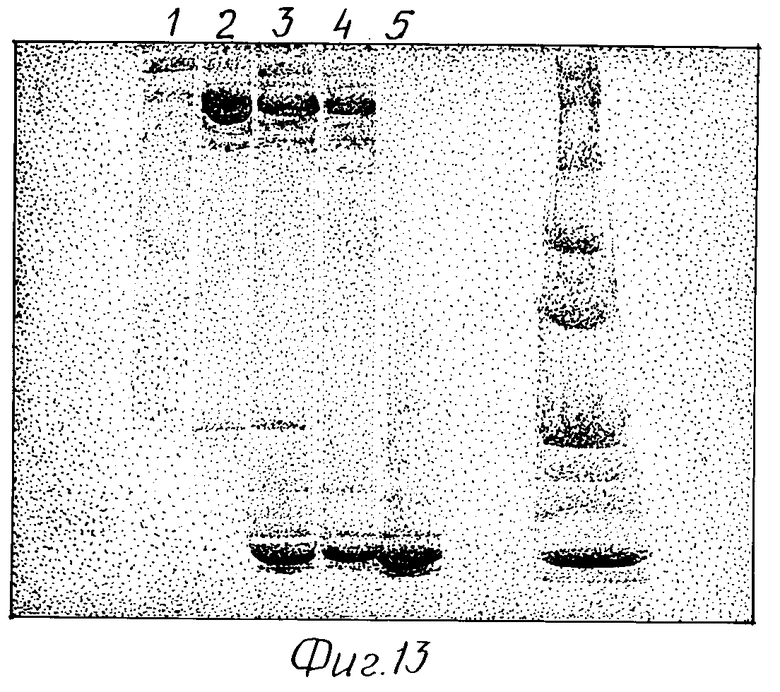
Фиг. 13 демонстрирует чистоту иммунотоксина 2G10-гелонина с помощью SDS-PAGE.
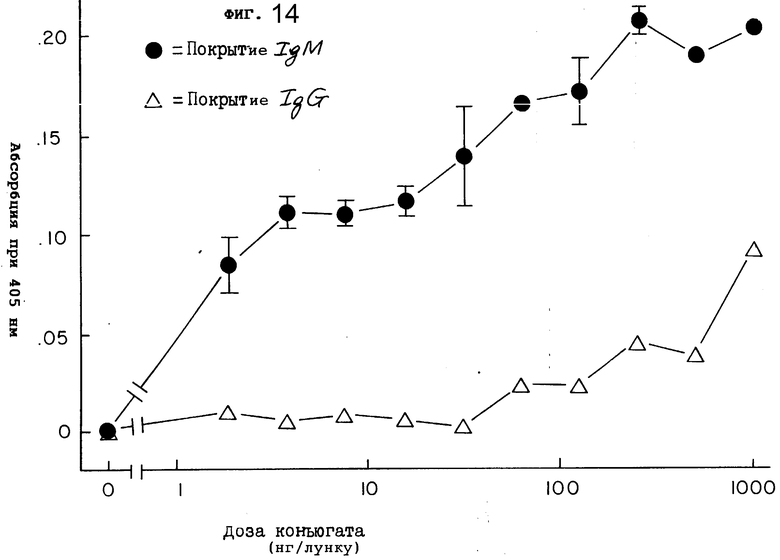
На фиг. 14 показано специфическое связывание иммунотоксина (2G10-гелонина) с IgM мыши.
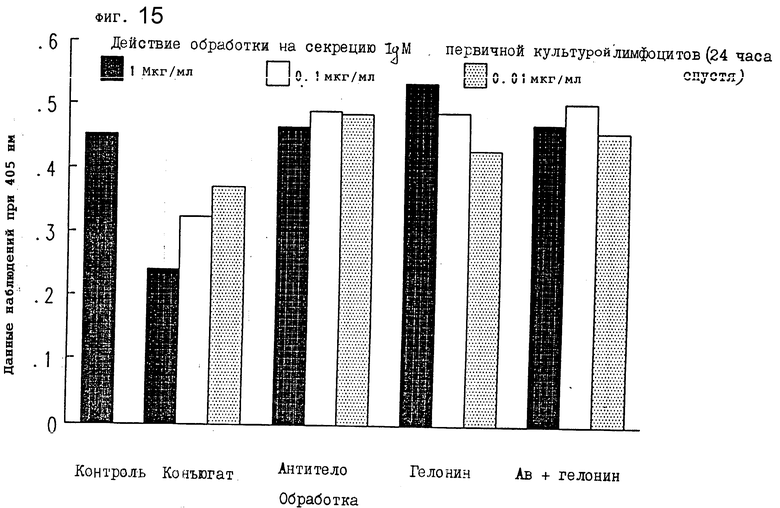
На фиг. 15 показано действие обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов после 24 ч I.
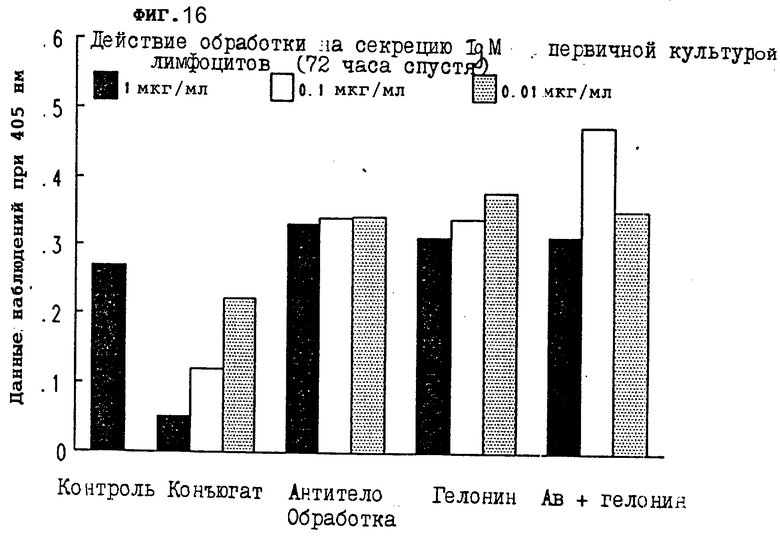
На фиг. 16 показано действие обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов через 72 ч I.
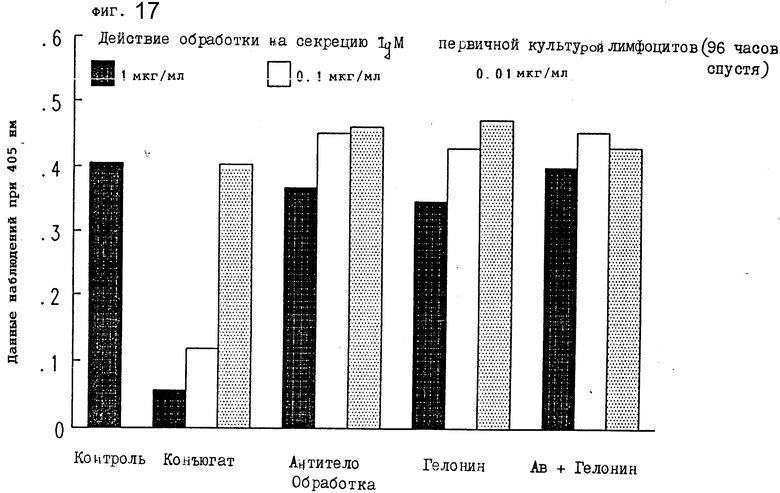
На фиг. 17 показано действие обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов после 96 ч I.
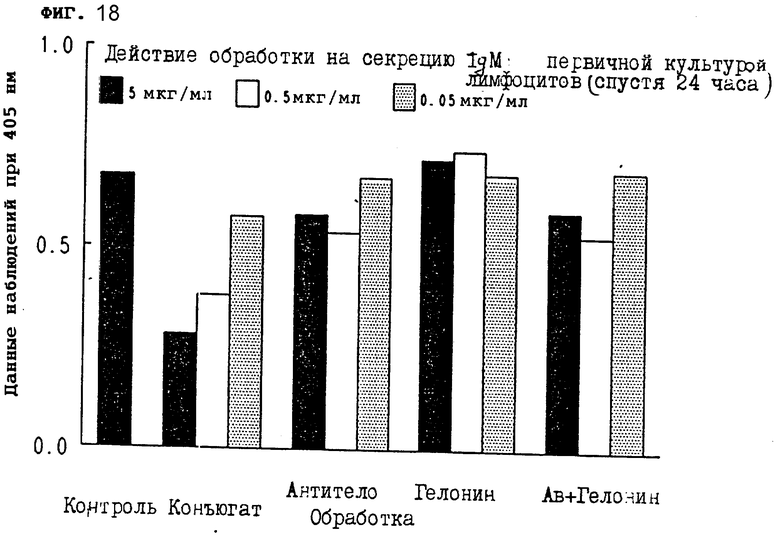
На фиг. 18 показано действие обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов через 24 ч II.
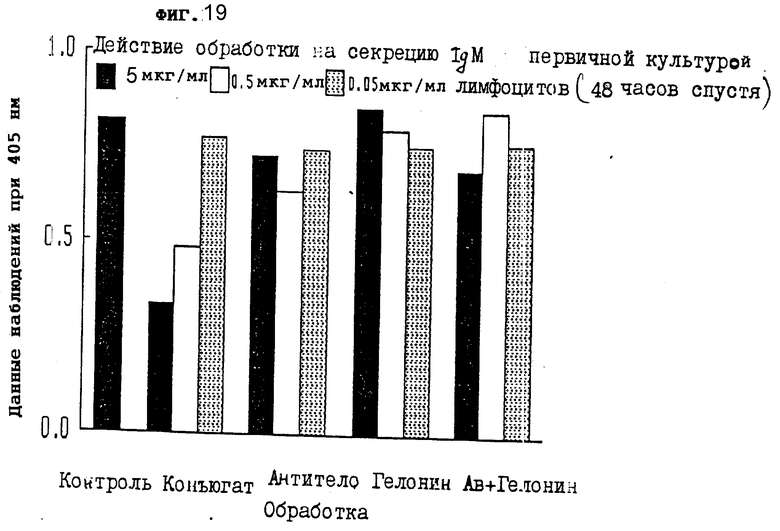
На фиг. 19 показано действие обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов через 48 ч II.
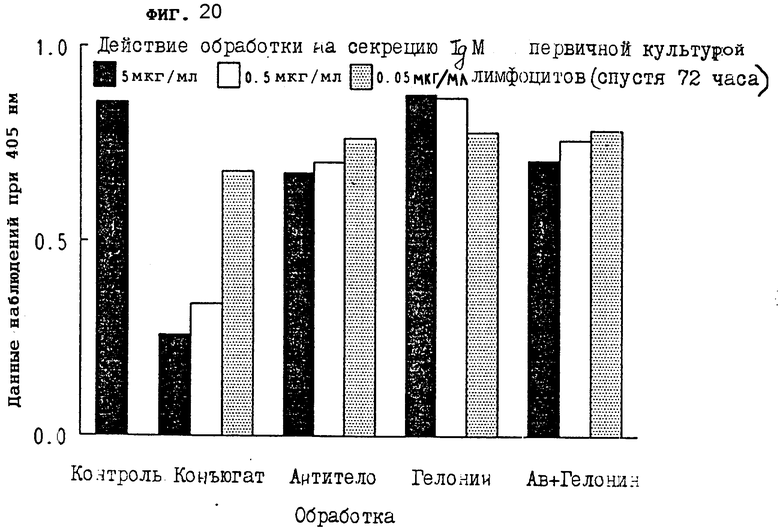
На фиг. 20 показано действие обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов через 72 ч II.
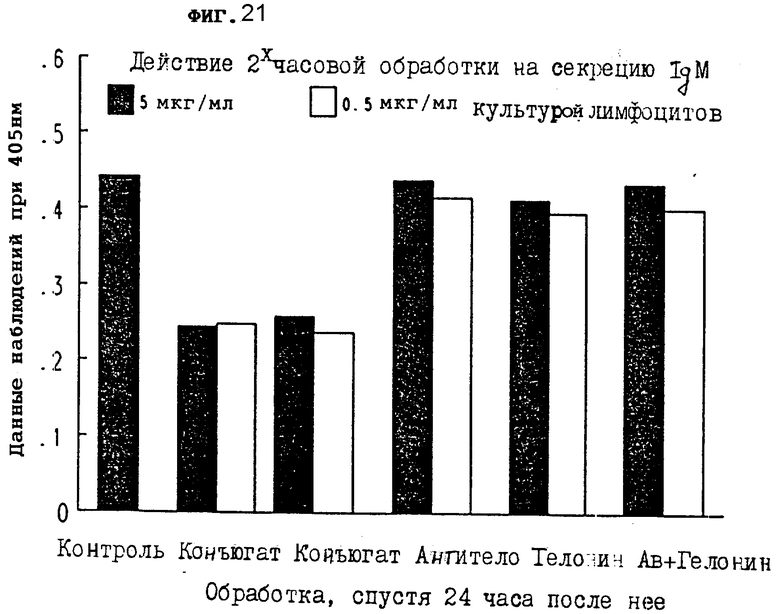
На фиг. 21 показано действие 2-часовой обработки конъюгатом на секрецию IgM первичной культурой лимфоцитов через 24 ч III.
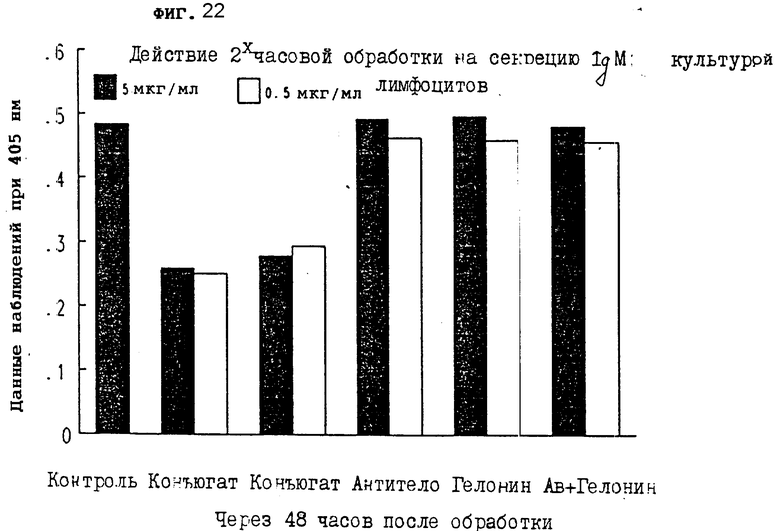
На фиг. 22 показано действие 2-часовой обработки на секрецию IgM первичной культурой лимфоцитов через 48 ч III.
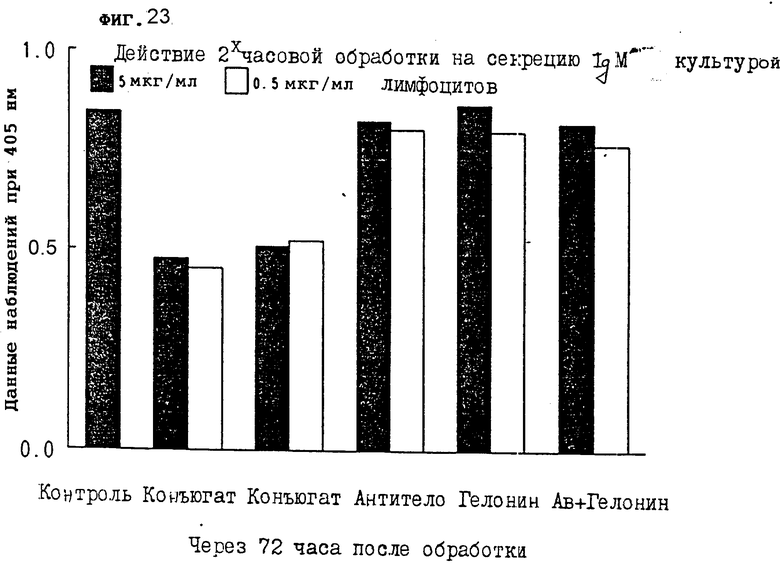
На фиг. 23 показано действие 2-часовой обработки на секрецию IgM первичной культурой лимфоцитов через 72 ч III.
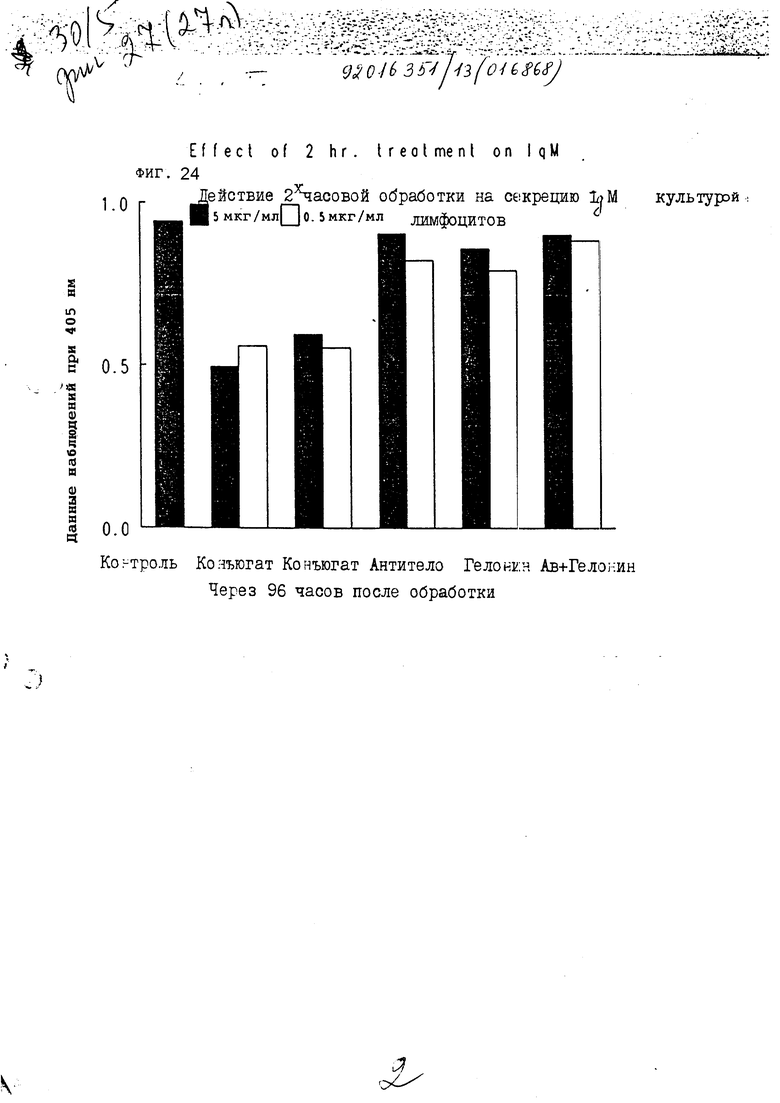
На фиг. 24 показано действие 2-часовой обработки на секрецию IgM первичной культурой лимфоцитов через 96 ч III.
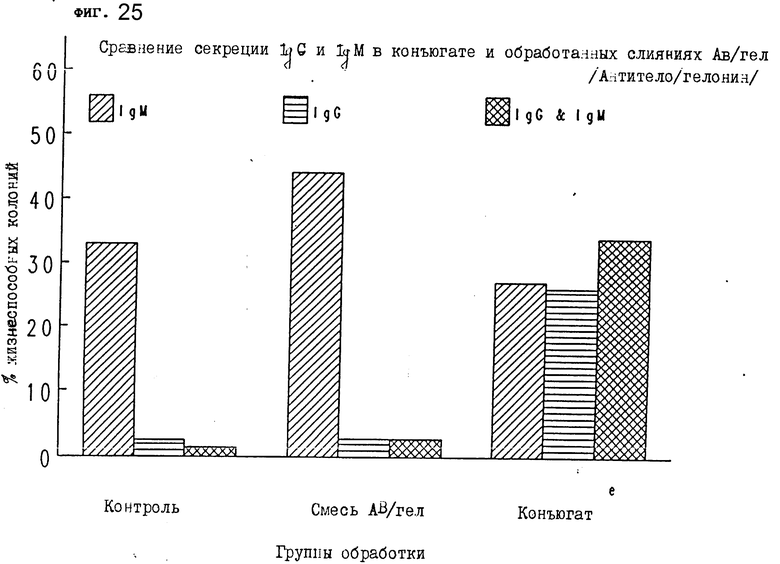
На фиг. 25 показано сравнение секреции IgM и IgG в конъюгате и обработанных слияниях AB/гел/антитело/гелонин/.
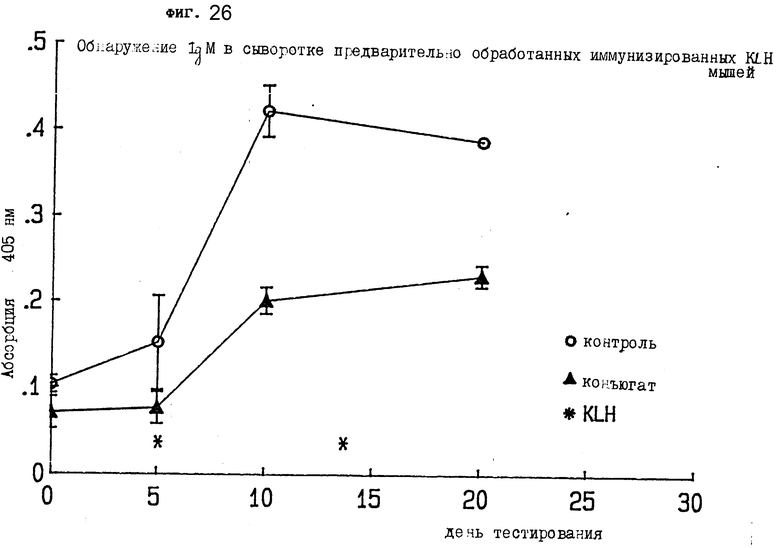
На фиг. 26 показано обнаружение IgM в сыворотке мыши I, предварительно иммунизированной KLH.
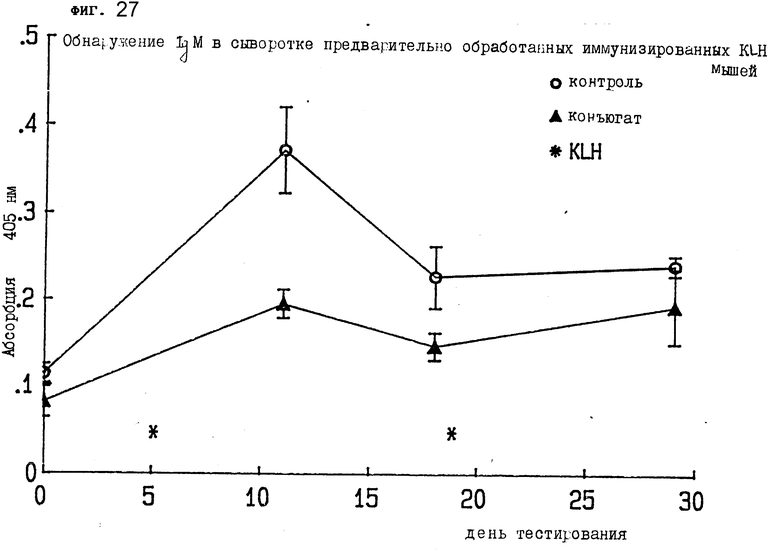
На фиг. 27 показано обнаружение IgM в сыворотке мыши II, предварительно иммунизированной KLH.
Используемый в настоящем тексте термин "моноклональное антитело" обозначает композицию антител, представленную однородной их популяцией.
Термин "функциональный эквивалент" относительно, например, крысиных моноклональных антимышиных IgM-антител, обозначает моноклональное антитело, которое: (a) кроссблокирует крысиное моноклональное антитело, (b) селективно связывается с мышиным IgM-антителом, (c) имеет q-изотип, и (d) не связывается ни с изотипом IgG1, ни с изотипом IgG2.
Термин "потомство", как он здесь используется относительно гибридом, продуцирующих моноклональные антитела, являющиеся предметом настоящего изобретения, предназначен для того, чтобы "охватить" все производные и продукты родительской гибридомы.
Настоящее изобретение может быть использовано для продуцирования антител, которые будут связываться с IgM-антителами любых видов. Описание настоящего изобретения раскрывает получение линии клеток гибридомы, которая стабильна и продолжает продуцировать анти-IgM-антитело, направленное на иммунизирующий вид. В предпочтительном варианте анти-IgM моноклональное антитело, являющееся предметом настоящего изобретения, направлено на мышиный или человеческий IgM.
Продуцирование моноклонального антитела
Партнеры для слияние, обеспечивающего получение гибридомы настоящего изобретения, получают при помощи иммунизации крыс мышиным IgM-антителом. Крысам прививали подкожным и внутрибрюшинным способом иммуногенное количество мышиного IgM-антитела в адъюванте Фрейнда, которое затем увеличивали путем введения аналогичного количества иммуногена в адъюванте. Через несколько дней после последней активации иммунизации селезенки иммунизированных крыс извлекали и из них получали суспензию клеток для использования при слиянии.
Гибридомы получали из спленоцитов и клеток опухоли крысы, используя технику гибридизации соматических клеток Колера Б. и Милстейна К. (Nature (1975), т. 256, стр. 495-497) в модификации Бака и др. (In vitro, (1982), т. 18, стр. 377-381). Для гибридизации можно использовать доступные линии миеломы крысы, такие, как УВ2/0 и УЗ-Ag 1.2.3.
В основном, эти приемы включают слияние клеток опухоли и спленоцитов, используя вещество, способствующее слиянию клеток, такое, как полиэтиленгликоль. После слияния клетки отделяли от среды и выращивали в селективной среде, такой, например, как НАТ-среда, чтобы удалить негибридизированные родительские клетки. Гибридомы размножали, если это необходимо, а супернатанты анализировали на анти-мышиную IgM-активность при помощи известных приемов иммуноанализа (например, при помощи радиоиммуноанализа, ферментного иммуноанализа или флуоресцентного иммуноанализа), используя иммунизирующий агент IgG1, IgG2 и IgM/мышиное IgM-антитело/ в качестве антигена. Далее характеризовали положительные клоны с тем, чтобы определить, удовлетворяют ли они критерию антител, являющихся предметом настоящего изобретения.
Гибридомы, которые продуцируют такие антитела, могут выращиваться in vitro или in vivo с использованием известных приемов. Моноклональные антитела могут быть выделены из культурной среды (или жидкостей организма в зависимости от случая) при помощи известных приемов очистки иммуноглобулина, таких как осаждение сульфатом аммония, гелевый электрофорез, диализ, хроматография и ультрафильтрация, если это необходимо.
Селекция/характеристика моноклонального антитела
Важными характеристиками моноклональных антител являются (1) класс иммуноглобулина, (2) селективность относительно мышиного IgM-антитела и (3) применимость при идентификации и связывании с клетками гибридомы, продуцирующей мышиный IgM.
Селективность и спектр данного антитела определяли при помощи его испытания с (1) IgG1, IgG2 и IgM-продуцирующими клетками гибридомы и (2) IgG1, IgG2 и IgM-антителами. При селекции предлагаемых антител первоначально скринировали приблизительно 162 растущих культур гибридомы. Девять клонов взаимодействовало с мышиным IgM-антителом, но не с IgG. Один из этих клонов выбирали для дальнейшей характеристики.
Антитела, проявляющие приемлемые селективность и спектры, конъюгировали с гелонином, используя N-сукцинимидил-3-(2-пиридилтио)пропионат)СПДП) или иминотиолан (ИТ) в качестве связывающего агента. Конъюгаты испытывали против покрытых IgM и IgG пластинок (фиг. 2), чтобы определить, сохраняется ли специфичность антитела после химического спаривания с токсином.
Дальнейшие подробности характеристики этого антитела будут даны в промерах, приводимых ниже.
Иммунохимические агенты
Иммунохимические производные моноклональных антител, являющихся предметом настоящего изобретения, которые представляют особую важность, являются иммунотоксинами/конъюгатами антитела и цитотоксичной составляющей и меченых, например, радиомеченых, меченых ферментами, магнитномеченых или меченых люминофором, производных, в которых метка представляет собой средство для идентификации и/или сортировки иммунных комплексов, включающих меченое антитело/.
Цитотоксичной составляющей иммунотоксина может быть или цитотоксичный препарат, или ферментативно активный токсин бактериальной или растительной природы, или ферментативно активный фрагмент (A-цепь) такого токсина. Ферментативно активные токсины и их фрагменты являются предпочтительными, их примерами являются гелонин, A-цепь токсина дифтерии, несвязывающие активные фрагменты токсина дифтерии, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь арбина, A-цепь модекцина, альфа-сарцин, протеины Aleurites fordii, диантиновые протеины, протеины Phytoiacca americana (PAPI, PAPII и PAP-S), ингибитор Momordica Charantia, курцин, кротин, ингибитор Saponaria officinalis, митоцеллин, рестриктоцин, феномицин и эномицин. Гелонин является наиболее предпочтительным. Конъюгаты моноклонального антитела и такие цитотоксичные составляющие могут быть получены с использованием самых разнообразных соединяющих агентов, предназначенных для получения бифункциональных протеинов. Примерами таких реагентов являются СПДП, ИТ, бифункциональные производные сложных имидоэфиров, такие, как диметиладипимидат-НС1, активные сложные эфиры, такие, как дисукцинимидилсуберат, альдегиды, такие, как глютаральдегид, бис-азидосоединения, такие, как бис/пара-азидопензоил/-гександиамин, бис-диазониевые производные, такие, как бис-/пара-диамоний-бензоил/-этилендиамин, диизоцианаты, такие, как толилен 2,6-диизоцианат, и бис-активные соединения фтора, такие, как 1,5-дифтор-2,4-динитробензол.
При использовании с целью подавления продукции IgM-антител гибридомами in vitro, конъюгаты в общем случае добавляют в культурную среду клеток в концентрации не менее примерно 10нМ. Форма и способ применения в экспериментах in vitro не играют решающей роли. В общем случае используют водные формы, которые совместимы с культуральной или перфузионной средой. Цитотоксичность может быть "считана" при помощи известных приемов для определения присутствия или концентрации клеток гибридомы, продуцирующих IgM.
При использовании in vivo с целью подавления секреции клетками IgM, иммунотоксины вводят иммунизированным животным в терапевтически эффективных количествах (т.е. количествах, которые исключают или снижают продуцирование IgM спленоцитами). Они обычно применяются парентерально или, в предпочтительном варианте, внутривенным способом. Доза и режим дозировки будут зависеть от природы продуцирующей IgM клетки, подлежащей подавлению, характеристик конкретного иммунотоксина, например, его терапевтического показателя, и активности действия.
Количество применяемого иммунотоксина может изменяться в области от примерно 0,1 до примерно 10 мг/кг массы организма.
При парентеральном применении иммунотоксины используют в виде единичной дозы инъектируемой формы (раствора, суспензии, эмульсии) вместе с приемлемым с фармацевтической точки зрения парентеральным носителем. Примерами таких носителей являются вода, физиологический раствор, раствор Рингера, раствор декстрозы и 5%-ный человечий сывороточный альбумин. Можно также использовать неводные носители, такие как нелетучие масла и этилолеат. В качестве носителей можно использовать липосомы. Носитель может содержать небольшие количества добавок, таких, как материалы, которые увеличивают изотоничность и химическую стабильность, например, буферы и консервирующие агенты. Иммунотоксин в общем случае включают в такие носители в концентрациях от примерно 1 мг/мл до 1 мг/мл.
Цитоксичные радиофармацевтические агенты для подавления клеток, продуцирующих IgM, могут быть получены путем комбинирования изотопов (например, V, Pt), испускающих переносчики высокой линейной энергии (ПЛЭ), с антителами. Термин "цитотоксичная составляющая", как он здесь используется, включает такие изотопы.
Метки, которые используются при получении меченых вариантов антител, включают составляющие, которые могут быть обнаружены непосредственно, такие, например, как люминофоры и радио-метки, а также такие, как ферменты, которые в результате реакции могут образовывать производные, которые затем могут быть обнаружены. Примерами таких меток являются 32P, 125I, 3H, 14C, флюоресцентные агенты и их производные, родамин и его производные, дансил, умбеллиферон, 2,3-дигидрофталазин-дионы, пероксидаза хрена обыкновенного, щелочная фосфатаза, лизоцим и глюкоза 6-фосфат дегидрогеназа. Антитела могут быть снабжены такими метками при помощи известных приемов. Например, можно использовать соединяющие агенты, такие как альдегиды, карбодиимиды, дималеимид, имидаты, сукцинимиды, бис-диазотированный бензидин и т.п., для того чтобы снабдить антитела указанными выше флуоресцентными, хемилюминесцентными и ферментными метками. Антитела могут быть также мечены с использованием магнитных шариков, применяемых в режиме "магнитной селекции".
Антитела и меченые антитела могут быть использованы в самых разнообразных процедурах сортировки клеток, чтобы отделить гибридомные клетки, продуцирующие IgM, от гибридомных клеток, продуцирующих IgG, или чтобы удалить клетки гибридомы, продуцирующей IgM, из культур, содержащих такие клетки.
Приемы, которые могут быть использованы для анализа, известны и включают прямые и непрямые методики. Прямые анализы включают инкубирование гибридомы или антител неизвестного изотипа с меченым антителом, являющимся предметом настоящего изобретения. Если образец включает продуцирующие IgM клетки, меченое антитело будет связываться с такими клетками. После промывки клеток с тем, чтобы удалить несвязанные меченые антитела, образец анализируют на присутствие меченых иммунных комплексов. При непрямых анализах образец клеток инкубируют с немеченным моноклональным антителом. Образец затем обрабатывают меченым антителом против моноклонального антитела (например, меченого антикрысиного антитела), промывают и анализируют на присутствие меченых третичных комплексов.
Приводимые ниже примеры дают подробное описание получения, характеристики и использования одного из представителей моноклонального антитела по изобретению. Эти примеры не следует рассматривать в качестве ограничения настоящего изобретения в какой-либо степени.
Пример 1. Источник и характеристика анти-мышиных IgM моноклональных антител крысы
Гибридому крысы, обозначаемую через 58,6, получали от доктора Иоанна Триала (Joanne Trial), Факультет иммунологии Андерсоновского Онкологического Центра. Сначала линия клеток 58,6 секретировала крысиное антитело. Линия клеток не была стабильной и, спустя примерно четыре субкультуры, клетки 58,6 прекращали продуцировать какие-либо антитела. Первоначальный источник клеток затем клонировали при помощи ограничивающих разбавлений с тем, чтобы получить линию клеток, которая была бы стабильной и продолжала продуцировать анти-IgM антитела.
A) Клонирование при помощи ограничивающего разбавления
Клетки 58,6 культивировали в среде Айскоува в течение 3 дней при температуре 37oC во влажной атмосфере с 5% CO2 в воздухе.
Когда размножающаяся культура клеток достигала 50%-ного слияния, их собирали центрифугированием и осуществляли подсчет с использованием гемоцитометра. Клетки разбавляли 50% свежей среды и 50% обработанной среды (среды, в которой клетки 58,6 выращивали в течение 7 дней) и помещали на пластинки с 96 углублениями в расчете приблизительно одна клетка на каждое углубление пластины. Когда в углублениях, содержащих по одной клетке, вырастали небольшие колонии (приблизительно 12 дней), среду удаляли и анализировали на анти-IgM антитело, как описано в примере 2. Положительные клетки размножали до значительных объемов антител, клеточное сырье замораживали, а антитела затем подвергали анализу и характеристике. Когда линии клеток были полностью проанализированы на тип и специфичность продуцируемых антител, соответствующие линии клеток снова клонировали и размножали для последующего замораживания клеточного сырья и введения мышам Nude, обработанным пристаном, с целью получения асцитных жидкостей.
B) Замораживание гибридных клеток
Когда гибридные клетки, помещенные в колбы Т75, достигали 70% слияния, клетки собирали центрифугированием, и осадок снова суспендировали в 0,9 мл фетальной сыворотке теленка. Непосредственно перед замораживанием клетки переносили в виалы для замораживания и в каждую виалу добавляли 0,1 мл диметилсульфооксида (ДМСО). Виалы хранили в жидком азоте.
C) Получение асцитных жидкостей
Приблизительно 107 клеток гибридомы промывали в среде, не содержащей сыворотки, и инъецировали внутрибрюшинным способом мыши Nude, которая предварительно получала внутрибрюшинным способом 0,5 мл пристана за 7-14 дней до этого. Асцитные жидкости в основном образовывались в течение 1-3 недель, и их собирали из брюшной полости, используя большую калибровочную иглу. Жидкость собирали в пробирки, содержащие 5 мл ПБС (физраствор, забуференный фосфатом) с 20 мМ ЭДТА (этилендиаминтетрауксусная кислота). После центрифугирования со скоростью 2000 • g в течение 10 мин супернатант собирали, обрабатывали 0,1%-ным азидом натрия и хранили при температуре 4oC или замораживали при -20oC малыми порциями. Эта жидкость обеспечивала богатый источник моноклонального антитела (приблизительно 5-10 мг-мл).
Пример 2. Селекция и анализ клеток гибридомы, продуцирующих крысиные анти-мышиные антитела
Колонии гибридомы, которые выращивали до плотности приблизительно 500-1000 клеток в течение 2 недель, отбирали для последующего анализа с тем, чтобы определить, какие из клеток гибридомы продуцировали антитела, которые связывают мышиные IgM-антитела. Культуральную среду этих колоний анализировали на присутствие анти-мышиного IgM крысы при помощи иммуноферментного анализа, осуществляемого в соответствии с процедурой Боллера и др. 100 мг 1 мг/мл очищенного мышиного IgM или IgG (фирма Сигма Кемикал Компани, Сент-Луис, Mo) разбавляли в покрывающем буфере (50 мМ NaHCO3, pH 9,8) и абсорбировали в течение ночи на микротитровальных пластинах с 96 углублениями путем инкубирования при температуре 4oC во влажной камере. Затем углубления промывали три раза забуференным фосфатом физраствором, содержащим 0,2% Твина-20 (ПБС-Твин). После промывки и удаления всех следов жидкости в углублениях промоканием 100 μг супернатанта гибридомы добавляли в покрытые IgM углубления и инкубировали при комнатной температуре в течение 2 ч. Пластины снова промывали при помощи ПБС-Твина и инкубировали в течение 1 ч при комнатной температуре с 100 μл разбавленного в соотношении 1:1000 ПБС-Твином конъюгированного с пероксидазой анти-крысиного IgG козы. После этого углубления снова промывали, как это описано выше, а затем осуществляли реакцию с 100 μл 1 мМ АВТС (2,2-азино-ди)3-этилбензтиазолин сульфоновая кислота в 0,1 М цитратном буфере, pH 4,2, содержащем 0,03% перекиси водорода в течение 20-60 мин при температуре 37oC. Оптическую плотность измеряли в диапазоне 405 нм на MikroElisa Reader.
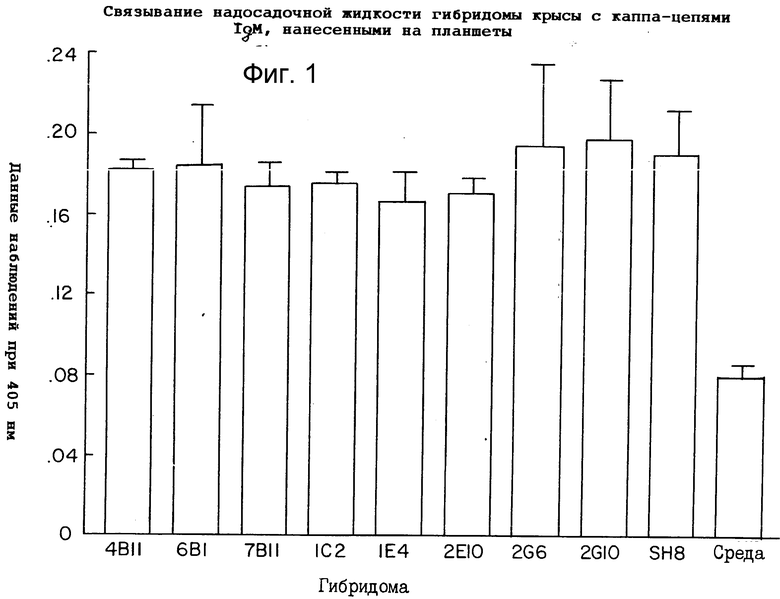
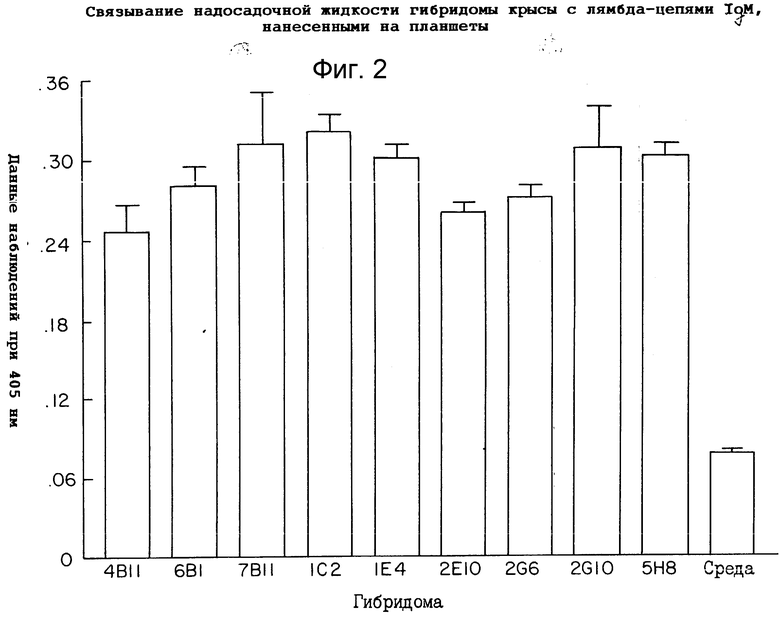
Для того чтобы оценить, был ли эпитоп, узнанный анти-IgM антителами, расположен на тяжелой цепи или легкой цепи антитела, испытывали связывание 9 супернатантов гибридомы с покрытыми IgM-ламбда и IgM-каппа пластинками. Гибридомы 1C2 и 2G10 в равной мере хорошо связывались с покрытыми IgM-каппа и IgM-ламбда углублениями, что указывает на то, что связывающая специфичность анти-IgM-антител находится на тяжелой (mu) цепи IgM-антитела. На то, что другие 7 антител, подвергнутые испытаниям, также связываются с мышиным IgM, указывают результаты, приведенные на фиг.1. Как можно видеть на фиг. 1 и 2, испытывали девять различных гибридом против пластинок, покрытых либо IgM-каппа, либо IgM-ламбда. Стандартные ELISA-анализы осуществляли для того, чтобы оценить иммунизацию крысы, связанную с каждой пластинкой. Было установлено, что все антитела связываются в углублениях, покрытых как IgM-каппа, так и IgM-ламбда. Среди антител с самым высоким связыванием для дальнейшего изучения выбрали 1C2 и 2G10. С учетом лучших характеристик роста и продуцирования антител, в конечном итоге было выбрано антитело 2G10. Оно давало оптическую плотность 0,3±0,03 (стандартное отклонение). Точкой отсчета было 0,06 единиц оптической плотности (О.П.) при использовании среды без крысиного моноклонального антитела. Углубления, которые давали реакцию на анти-мышиное IgM-антитело, превышающую или равную 0,3 О.П., отбирали.
Специфичность моноклонального 2G10-антитела крысы испытывали при помощи ELISA при самых различных условиях. Чтобы определить селективное узнавание IgM в сравнении с IgG при помощи 2G10, 100 нг мышиного IgM-ламбда или IgG3-ламбда абсорбировали на микротитровальных пластинках (100 μл) и инкубировали с возрастающими концентрациями антитела 2G10. Как показано на фиг. 2, в области изменения доз антитела 2-1000 нг, 2G10 связывался с пластинками, покрытыми IgM, в более значительной степени, чем с пластинками, покрытыми IgG. При дозах антитела 2G10 50 нг и ниже на пластинках, покрытых IgG, никакого связывания не было отмечено, в то время как углубления, покрытые IgM, узнавались при помощи 2G10.
Как показано на фиг. 3, антитело 2G10 (добавленное в дозе 100 нг в 100 μл), инкубированное в углублениях, покрытых IgG3 и IgM, указывает на селективное связывание с покрытыми IgM пластинками при всех концентрациях покрытия. В десять раз более значительную химическую активность измеряли при помощи ELISA на пластинках, покрытых IgM, по сравнению с покрытыми IgG, при дозе покрытия 10000 нг IgM или IgG (сравните IgG3-каппа к IgM-каппа). Этим снова было подтверждено селективное IgM-связывание.
Антитело 2G10 также испытывали на его способность к селективному узнаванию IgM в непрямом ELISA-анализе. Антигены наносили на микротитровальные пластинки и инкубировали с культуральной средой гибридомы, которая содержала антитело, узнающее эти антигены. В этом анализе использовали антитела подклассов IgM и IgG1. После инкубирования пластинок, покрытых антигеном, с взаимодействующими с ним антителами, в углублениях добавляли возрастающие концентрации крысиного антитела 2G10, и при помощи ELISA определяли связывание 2G10. Как показано на фиг. 5, 2G10 чувствительно и селективно узнает IgM, связанный с соответствующим ему антигеном (темные кружочки), но не в состоянии обнаружить IgG (светлые кружочки) при тех же условиях, хотя присутствие IgG1 можно было легко обнаружить антителами, активными относительно всех подтипов мышиного иммуноглобулина. Эти результаты подтверждают селективное узнавание иммуноглобулинов мышиных IgM-подклассов при помощи крысиного моноклонального антитела 2G10. Моноклональное антитело крысы могло также узнать IgM-антитело в пикограммовых дозах, что подтверждает его высокое сродство с мышиным IgM-антителом.
Пример 3. Характеристика анти-мышиных IgM-антител крысы
Мышиным иммуноглобулином различных подтипов (IgM-каппа и ламбда, IgG1, IgG2 и IgG3) покрывали микротитровальные пластинки, как в примере 2, а затем более подробно изучали связывание анти-мышиных IgM-антител крысы, продуцированных клетками гибридомы, которые были положительны в примере 2. Все пластинки "считывали" в области с длиной волны 405 нм. Поглощение (в сравнении с контролем) рассматривали как указание на присутствие антитела против мышиного IgM.
Пример 4. Очистка и характеризация моноклонального антитела крысы 2G10
Как было показано в примере 3, антитело 2G10 давало положительную реакцию с IgM-каппа и ламбда. Это антитело подвергали очистке при помощи центрифугирования и фракционирования с использованием сульфата аммония.
Линию клеток гибридомы, обозначенной через 2G10, сдали на хранение в Американское Собрание Типов Культур (АТСС), Роквилл, Мд, США 23 апреля 1990 г., где она имеет шифр хранения N HB 10436. Эти культуры доступны в соответствии с патентным законодательством и регулированием в этой области в Соединенных Штатах Америки и тех иностранных государствах, в которых поданы аналоги настоящей патентной заявки. Доступность сданной на хранение культуры не включает в себя разрешение на осуществление изобретения, являющегося предметом настоящей патентной заявки, а также патента, выданного по настоящей патентной заявке или любой раздельной заявке, или ее продолжение.
Супернатант культуры 2G10 (или асцитную жидкость, полученную от мышей Nude) доводили до 45%-ного насыщения сульфатом аммония (высаливание) при помощи медленного добавления равного объема раствора сульфата аммония с 90%-ным насыщением. Образец перемешивали в течение 30 мин при температуре 4oC, а затем центрифугировали со скоростью 20000 x g в течение 30 мин. Осадок снова суспендировали в растворе сульфата аммония с 40%-ным насыщением, перемешивали 30 мин и снова получали осадок центрифугированием, как это описано выше. Этот осадок повторно суспендировали в воде и подвергали диализу против 100 объемов ПБС. Порции раствора использовали для определения содержания протеина (по оптической плотности при 280 нм), чистоты (при помощи SDC-PAGE) и специфичности связывания (при помощи ELISA). Оставшийся раствор антитела замораживали при -20oC и хранили до употребления.
IgM-антитела подвергали очистке при помощи осаждения сульфатом аммония и гель-фильтрации на колонке размером 2,6x40 см, заполненной агарозой, декстраном и/или акриламидом, элюируя смесью ПБС/ 0,01% раствор азида натрия при комнатной температуре с объемной скоростью потока 1 мл/мин.
Подкласс моноклонального антитела крысы 2G10 определяли при помощи процедуры Оухтерлони (Ouchterlony и Nilsson (1958) в книге Handbook of Exp. Immun. , ред. Weir, изд. Blackwell Scientific, London, p. 19.1-19.44), используя комплект реактивов для иммунодиффузии, производимый фирмой ICN Immunobiologicals (Lisle, IL).
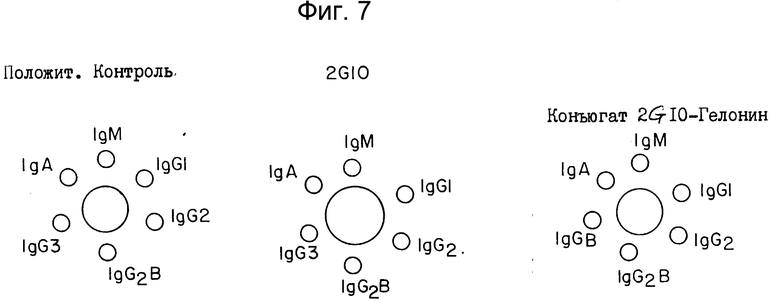
Подкласс антитела 2G10 очень важен при определении способа очистки. Чтобы осуществить анализ подкласса, использовали набор реактивов для иммунодиффузии по Оухтерлони. При этом антисыворотки против различных подтипов иммуноглобулина крысы помещали в соответствующие углубления. В центральное углубление добавляли известный стандарт или неизвестный образец и им давали возможность диффундировать в полутвердую среду. Полоса осаждения в сайте специфической антисыворотки указывает на подтип. Как показано на фиг. 7, положительные контрольные образцы, содержащие все антитела подтипов крысы, демонстрируют линии реакции во всех углублениях (для всех подтипов). С другой стороны, антитело 2G10 взаимодействовало только с антисывороткой подтипа IgG2A, что означает, что антитело 2G10 крысы является антителом IgG2a.
Для того чтобы оценить связывание производимого промышленностью антитела крысы с мышиным IgM, оценивали активность в ELISA-тесте моноклонального антитела крысы L-O-MM-9 (фирма Serotec, cat # MCA 199) против мышиного IgM. Сто нанограмм mu-каппа, гамма-каппа или гамма-ламбда помещали в каждое из углублений пластины с 96 углублениями. Затем добавляли различные количества антитела крысы LO-MM-9 и осуществляли ELISA-анализ на антитело крысы, как это было описано ранее. Как показано в табл. 1, какого-либо связывания этого антитела крысы с мышиным IgM-покрытием углублений отмечено не было.
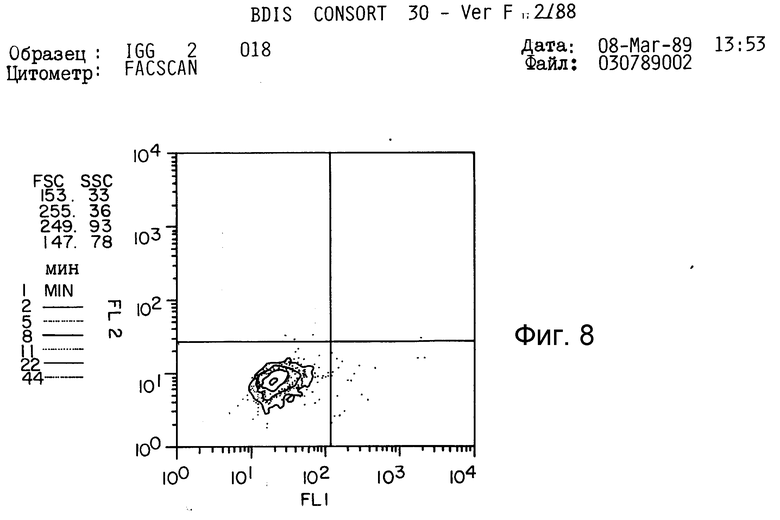
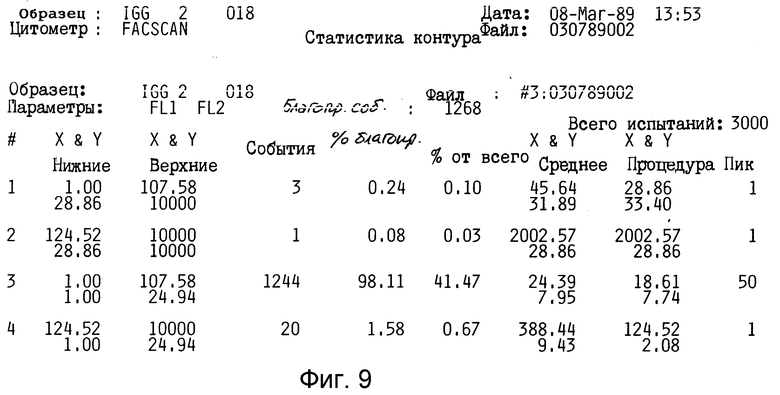
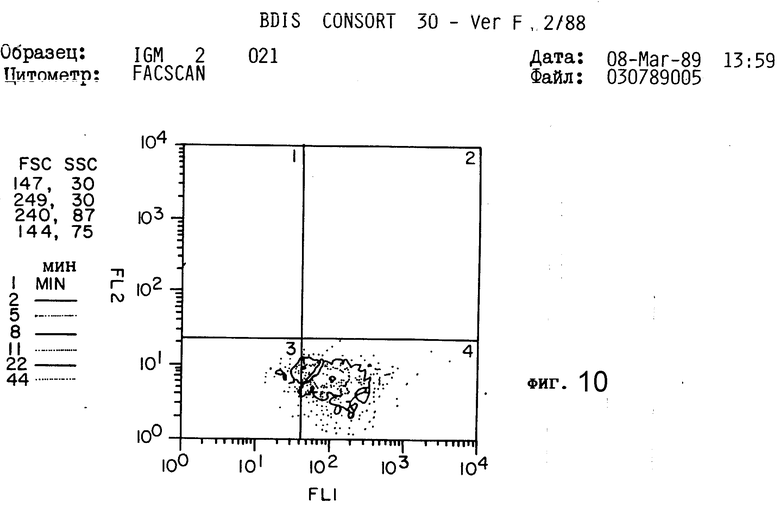
Таким образом, это антитело не представляется полезным для дальнейшего изучения. Кроме того, как показано на фиг. 8-11 (полоса 3), разделение этого антитела в SDS-PAGE давало по крайней мере три основные протеиновые полосы и по крайней мере пять слабых протеиновых полос.
Пример 5. Связывание антимышиного IgM крысы с клетками, продуцирующими IgM
Для того чтобы подтвердить, что антимышиное IgM-антитело крысы 2G10 связывается не только с пластиной с 96 углублениями, покрытыми очищенным IgM-антителом, но также и с клетками, продуцирующими IgM-антитела, осуществляли FACS-анализ на клетках 10C1 и клетках гибридомы 238-57ADR мыши, которые секретируют IgG и IgM, соответственно. Для этого 1•106 клеток центрифугировали со скоростью 500 • g в течение 3 мин, затем промывали три раза ПБС, а затем снова суспендировали в 3 мл ПБС.
Очищенный на основе сродства к флуоресцеину F(ab)2-фрагмент анти-мышиного иммуноглобулина IgM козла (фирма Каппел) разбавляли в пропорции 1:100 в ПБС (1х) и 20-40 μл разведения добавляли к 20 μл клеточной суспензии. После инкубирования в течение 15-20 мин в темноте при комнатной температуре клетки дважды промывали ПБС, центрифугируя со скоростью 500 об/мин в течение 3 мин. Чтобы зафиксировать клетки, добавляли порцию (300 μл) параформальдегида (1% в ПБС). Клетки инкубировали при 4oC, а затем сортировали при помощи проточной цитометрии.
Для непрямого окрашивания клетки гибридомы сначала инкубировали с антимышиным IgM-антителом 2G10 крысы, промывали, затем окрашивали конъюгированным с флуоресцеином F(ab)2-фрагментом антимышиного иммуноглобулина IgM козла и сортировали при помощи проточной цитометрии.
Как показано в таблице 2 и на фиг. 8-11, антитело 2G10 связано специфически с IgM, содержащимся на поверхности секретирующих IgM клеток гибридомы мышей, и не связано с секретирующим IgM клетками.
Как показано в табл. 2, "фонового" флуоресцеина у необработанных клеток отмечено не было. Произвольный IgG крысы также не связывался ни с IgG-гибридами, ни с IgM-гибридами. Не было отмечено также связывания антитела 2G10 с продуцирующими IgG клетками гибридомы (1,58% клеток являются положительными, фиг. 8,9 и табл. 2). Однако, как показано на фиг. 10 и 11 и в табл. 2, 90% клеток, продуцирующих IgM-антитела мыши, прочно связываются с антителом 2G10. Таким образом, связывание антитела 2G10 с клетками имеет место, благодаря узнаванию мышиного IgM на поверхности клетки.
Анализ цитотоксичности
Предлагаемые антитела подвергали конъюгации с A-цепью токсина рицина (RTA), обработанной СПДП, как это описано Карлссоном Я. и др. Biochem. Journ. (1978), т.173, стр. 723-727, или иминотиоланом (ИТ).
Пример 6. Соединение 2G10 с гелонином
Получали сырьевой раствор СПДП-реагента (N-сукцинидил-3-(2-пиридилдитио)проприоната)) 6 мг/мл) в сухом ДМФ. В 1 мл раствора ПБС, содержащего 1 мг антитела 2G10, в стеклянной пробирке размером 12х75 мм, медленно добавляли СПДП до 5-кратного молярного избытка (приблизительно 10 μл сырьевого раствора). Смесь перемешивали через каждые 5 мин в течение 30 мин при комнатной температуре.
Избыточный непрореагировавший СПДП удаляли из образца методом гель-фильтрации на колонке типа Сефадекс G-25 (Sephadex G-25) (1 x 24 см), которую предварительно уравновешивали 100 мМ Na-фосфатным буфером, pH 7,0, содержащим 0,5 мМ ЭДТА (Буфер А). Фракции (0,5 мл) собирали и анализировали на содержание протеина, используя анализ на связывание красителя Брэдфорда (Брэдфорд (1976) Anal. Biochem. т. 72, стр. 248-254). Анализировали поглощение (600 нм) на пластинке с 96 углублениями, используя автосчитыватель типа Bio-Tek Microplate. Антитело элюировалось в свободном объеме (фракции 14-20), эти фракции собирали и хранили при 4oC.
Токсин гелонина экстрагировали из семян Gelonium multiflorum и подвергали очистке до однородного состояния, используя процедуру Стирпа и др. (Stirpe и др. J. Biol. Chem., т. 255, стр. 6947-6953 (1980)). Один миллиграмм очищенного гелонина (2 мг/мл в ПБС) добавляли в НС1-триэтаноламиновый буфер (ТЭА (НС1) до конечной концентрации 60 мМ ТЭА/НС1, а pH устанавливали на уровне 8,0. Раствор делали содержащим 1 мМ ЭДТК. Добавляли сырьевой раствор 2-иминотиолана -/0,5 М в 0,5 М ТЭА/HCl, pH 8,0/ до финальной концентрации 1 мМ, и образец инкубировали 90 мин при 4oC.
Избыток реагента 2-иминотиолана удаляли при помощи гелевой фильтрации на колонке с Сефадексом G-25 (1 x 24 см), которую предварительно уравновешивали 5 мМ трис-ацетатным буфером, pH 5,8, содержащим 50 мМ NaCl и 1 мМ ЭДТА. Фракции (0,5 мл) собирали и анализировали на содержание протеина на микротитровальных пластинках с 96 углублениями, используя анализ на связывание красителя Брэдфорда. Гелонин элюировали во фракциях 14-20; эти фракции собирали и хранили при температуре 4oC. Антитело 2G10, модифицированное при помощи СПДП, смешивали с 5-кратным молярным избытком модифицированного 2-иммунотиоланом гелонина, pH смеси доводили до 7,0 при помощи добавления 0,05 М буфера ТЭА/HCl/pH 8,0/ и смесь инкубировали в течение 20 ч при температуре 4oC в атмосфере азота. Иодацетамид (0,1 М в ПБС) добавляли до финальной концентрации 2 мМ, чтобы блокировать любые оставшиеся свободные сульфгидрильные группы, а инкубирование продолжали еще в течение часа при температуре 25oC.
Очистка 2G10-гелониновых комплексов
Чтобы удалить продукты с низкой молекулярной массой и неконъюгированный гелонин, реакционную смесь наносили на колонку с Сефадекс G-300 (1,6 x 31 см), предварительно сбалансированную при помощи ПБС. Фракции (1,0 мл) собирали и порции по 50 μл анализировали на содержание протеина, используя анализ на связывание красителя Био-Рад. Чтобы удалить неконъюгированное 2G10, высокомолекулярный пик/фракции 17-23/ с колонки G-300 наносили на колонку для аффинной хроматографии типа Blue Sepharose GL-6B, которую предварительно уравновешивали при помощи 10 мМ фосфатного буфера (pH 7,2), содержащего 0,1 М NaCl. После загрузки образца колонку промывали 30 мл буфера до тех пор, пока не будет полностью элюировано неконъюгированное антитело. Колонку элюировали линейным солевым градиентом от 0,1 до 2 М NaCl в 10 мМ фосфатном буфере, pH 7,2. Содержание протеина в элюированных фракциях определяли при помощи анализа на связывание красителя, который был описан ранее.
"Суммарную" смесь, содержащую свободное антитело 2G10, 2G10-гелонин и свободный гелонин, сначала подвергали очистке при помощи гель-фильтрации на колонке G-300. Как показано на фиг. 12, обнаруживался пик с высокой молекулярной массой (фракции 25-42), а также пик с более низкой молекулярной массой (фракции 55-67). Фракции 26-42 собирали с целью анализа на чистоту и химическую активность конъюгата.
Осуществляли PAGE-анализ очищенного конъюгата 2G10-гелонина. Как можно видеть на фиг. 13, объединенная смесь 2G10-гелонин (полоса 3) содержала свободный 2G10, свободный гелонин (стрелки), а также 2G10, соединенное с 1 молекулой гелонина (единичная стрелка), и 2G10, соединенное с двумя молекулами гелонина (двойная стрелка). Как можно сидеть на полосе 1, окончательно очищенный конъюгат 2G10-гелонин содержит, главным образом, 2G10, соединенное с 1 молекулой гелонина, и меньшие количества 2G10, соединенного с 2 молекулами гелонина. Этот препарат не был загрязнен свободным гелонином или свободным антителом.
Чтобы определить, модифицировали ли химическая реакция и связывание антитела 2G10 с гелонином узнавание антитела 2G10 мышиным IgM, пластины с 96 углублениями покрывали либо мышиным IgG-антителом, либо IgM-антителом, как на фиг. 3. Вместо антитела 2G10 в углубления добавляли в различных концентрациях конъюгат 2G10-гелонин. Чтобы обнаружить антитело крысы, затем осуществляли стандартный ELISA-анализ. Как показано на фиг. 14, конъюгат 2G10-гелонин легко связывается с IgM и только в небольшой степени с пластинами, покрытыми IgG. Только в самых высоких испытываемых концентрациях (1000 нг) углубление/конъюгат 2G10-гелонин в существенно связывался с углублениями, покрытыми IgG. Напротив, при аналогичной концентрации конъюгат 2G10-гелонин в существенной степени связывался с углублениями, покрытыми IgM. Таким образом, иммунная активность конъюгата 2G10-гелонин относительно IgM сохраняется. Кроме того, конъюгат 2G10-гелонин не давал перекрестной реакции в какой-либо заметной степени с мышиным IgG, и, таким образом, селективность конъюгата 2G10-гелонин также не менялась. Эти данные указывают на то, что конъюгат 2G10-гелонин должен связываться с IgM, покрывающим мышиные клетки, по той же схеме, что и само антитело 2G10.
Пример 7. Продуцирование ОДС-моноклональных антител
A. In vitro иммунизация и продуцирование моноклонального антитела
Описания, используемые в настоящее время при получении мышиных моноклональных антител, предусматривают формирование гибридом, которые секретируют антитела подкласса IgM. Для того чтобы подтвердить наиболее общий результат, получаемый в соответствии с этими описаниями, использовали два приема для формирования моноклональных антител против декарбоксилазы орнитина (ОДС) крысы. Первый прием включает иммунизацию клеток селезенки мыши in vitro, т. е. в чашках для культур, ОДС-протеином.
Эту процедуру осуществляли в соответствии с описанием Лубена и др. (Luben и Mohler (1980), Molec. Immunol, т. 17, стр. 635-639), используя 25 мг очищенного ОДС печени крысы.
ОДС-протеин инкубировали с мышиными клетками в течение 72 ч при температуре 37oC в присутствии тимоцит-кондиционированной среды (источник специальных факторов роста для клеток, секретирующих иммуноглобулин). Клетки селезенки затем сливали с клетками МРС-миеломы, используя полиэтиленгликоль в качестве агента, стимулирующего слияние. Полученные в результате гибридные клетки испытывали на секретирование антитела, химически активного относительно ОДС-протеина. Полученные результаты приведены в табл. 3.
После первоначального скрининга ОДС-протеином гибридомы снова клонировали и снова испытывали. Получали семьдесят гибридом, которые взаимодействовали с ОДС-протеином в соответствии с несколькими критериями. Однако, как показывает табл. 3, все клоны секретировали антитела изотипа IgM. Ни один из них не секретировал IgG-антитела. Как было установлено, эти антитела имеют ограниченное использование в самых различных приложениях моноклональных антител.
B. In vivo иммунизация и продуцирование моноклональных антител.
Второй прием, используемый для получения моноклональных антител, состоял в иммунизации мышей при помощи введения путем инъекции очищенного ОДС-протеина. Клетки мышиной селезенки изолировали после иммунизации, сливали с клетками миеломы P3 • 63 Ag 8.653 (используя PEG, как это описано выше). Клоны изолировали, испытывали на химическую активность относительно ОДС-протеина, а затем характеризовали на использование в качестве специфического ОДС - узнающего реагента. Результаты этих испытаний приведены в табл. 4.
Размножали двадцать семь линий моноклональных клеток, 100% которых секретировали антитело IgM-подкласса. Повторно было показано, что эти антитела являются неадекватными для использования в качестве ОДС-специфических реагентов.
Пример 8. Сортировка клеток, активируемая потоком (FACS-процедура).
Секретирующие IgM клетки гибридомы 238-57ADR подвергали центрифугированию со скоростью 500 • g в течение 3 мин, промывали три раза при помощи ПБС и снова суспендировали в 3 мл ПБС.
Конъюгированный с флуоресцеином, очищенный по сродству F(ab)2-фрагмент антимышиного иммуноглобулина IgM (Каппел) разбавляли в отношении 1:100 ПБС, и 20-40 μл добавляли в 20 μл суспензии клеток.
После инкубирования в течение 15-20 мин в темноте при комнатной температуре клетки дважды промывали ПБС, центрифугируя со скоростью 500 об/мин в течение 3 мин.
Порцию в 300 μл параформальдегида (1% в ПБС) добавляли, чтобы зафиксировать клетки. Клетки инкубировали при температуре 40oC до сортировки при помощи проточной цитометрии.
Для непрямого окрашивания клетки гибридомы сначала инкулировали с антимышиным IgM-антителом крысы 2G10, промывали, затем окрашивали флуоресцеиновым F(ab)2-фрагментом антимышиного иммуноглобулина IgM козла и сортировали при помощи проточной цитометрии.
Пример 9. Магнитное разделение клеток.
Клетки гибридомы мыши 238-57 ADR промывали три раза в среде Искова, содержащей 1%-ную фетальную телячью сыворотку и 0,1%-ный гентамицин. Клетки разделяли в центробежном сепараторе при 500 • g 3 мин при комнатной температуре и затем просчитывали. Осадок повторно суспендировали в среде (0,5 мл) и инкубировали 60 мин на льду с очищенным крысиным антимышиным IgM-антителом (2G10) в концентрации приблизительно 0,425 мг/мл (около 0,1 мл использовалось на 106 клеток/мл). После промывания дважды в холодной среде Искова клетки повторно суспендировали в 0,2 мл среды и просчитывали. Магнитные гранулы промывались три раза в среде, свободной от сыворотки, с использованием магнитной пластины. Клеточный осадок смешивался с осадком из гранул в соотношении 20 гранул/клетку. Общий объем не должен превышать 0,4 мл. Смесь клетки/гранулы инкубировалась 1/2 ч на льду с перемешиванием через каждые 10 мин.
Смесь клетки/гранулы повторно суспендировалась по меньшей мере в 2 мл среды и разделялась магнитно в направлении, перпендикулярном к силе тяжести. После завершения разделения супернатант удалялся, не затрагивая магнитный осадок. Гранулы повторно суспендировались в 1-2 мл среды и исследовались микроскопически.
Пример 10. Исследование лимфоцитов конъюгатом 2G10-гелонин
Для демонстрации цитотоксичности конъюгата гелонина 2G10 использовались две самки мыши Balb (c) (Harlan Sprague Dawley), у которых были иссечены селезенки. В ламинарном проточном боксе кожа над селезенкой промывалась 70%-ным изопропиловым спиртом и вскрывалась асептическими или простерилизованными ножницами. С использованием пары стерильных пинцетов и ножниц вскрывалась брюшина и обнажалась селезенка. Селезенка помещалась в 100-миллилитровую стерильную чашку Петри с 10-15 мл питательной среды (Iscoves; Gibco) с 10%-ной фетальной телячьей сывороткой (Hyclone) и 50 г/мл гентамицина (Tri-Bio Laboratories). Селезенка промывалась в среде и затем переносилась в чистую чашку Петри со свежей питательной средой и закрывалась.
Подготовка лимфоцитов
В ламинарном проточном боксе с использованием стерильных инструментов от селезенок отсекались избыточные жировые и соединительные ткани (белый материал). Пинцетами селезенки переносили в новую чашку Петри с 20 мл питательной среды. Процедура промывания повторялась дважды. Селезенки диспергировали с использованием двух 10-миллилитровых шприцев с калиброванными иглами до тех пор, пока не осталось больших фрагментов ткани. Клетки селезенки (спленоциты) имели вид непрозрачных смесей в питательной среде и содержали как красные, так и белые клетки. Клетки и среда вносились пипеткой в 50-миллилитровую центрифужную пробирку (Corning), и оставшиеся крупные недиспергированные кусочки ткани осаждались. Клетки помещались на лед в закрытой пробирке на 10 мин.
Верхняя порция отбиралась пипеткой (оставляя более крупные секции недиспергированной ткани), и центрифугировалась в течение 10 мин со скоростью 1800 оборотов в минуту (об/мин) в центрифуге с верхней платформой Dupon Sorvall 2LC-2B; надосадочная жидкость сливалась. Осадок повторно суспендировали в 10 мл питательной среды и клетки просчитывали с использованием 5 μл клеточной суспензии и 95 μг раствора трипана голубого в разбавлении 1:2 с применением гемоцитометра и фазового контрастного микроскопа Nicon со 100-кратным увеличением. Общий просчет клеток составил 6,24 • 108 клеток. Раствор доводился до 20 мл с получением 3,12 • 107 клеток/мл питательной среды. По одному мл этой клеточной суспензии вносили в несколько 15-миллилитровых центрифужных пробирок.
Группа образцов 1: к клеткам добавлялась только контрольная среда; группа 2: к клеткам добавлялась только контрольная среда; группа 3: к клеткам добавлялась только контроль-среда; группа 4: 2G10 антитело (AB) с концентрацией 1 мг/мл; группа 5: 2G10 антитело с концентрацией 0,1 μг /мл; группа 6: 2G10 антитело с концентрацией 0,01 μг /мл; группа 7: гелонин с концентрацией 1 μг /мл, эквивалентной = 0,2 мг/мл; группа 8: гелонин с концентрацией 0,1 μг /мл, эквивалентной = 0,002 μг /мл; группа 9: гелонин с концентрацией 0,01 μг /мл, эквивалентной = 0,02 μг /мл; группа 10: Ab + гелонин с 5 μг /мл Ab + 0,1 г/мл гелонина; группа 11: Ab + гелонин с 0,05 μг/мл Ab + 0,02 μг /мл гелонина; группа 12: Ab + гелонин с 0,005 μг /мл Ab + 0,002 г/мл гелонина; группа 13: конъюгат 2G10 + гелонин с 1 г/мл; группа 14: конъюгат 2G10 + гелонин с концентрацией 0,1 μг /мл; и группа 15: конъюгат 2G10 + гелонин с концентрацией 0,01 μл /мл.
В пронумерованные соответствующим образом пробирки был добавлен белок и общий объем в каждой был доведен до 5 мл питательной средой Искова. Крышки приоткрывали и пробирку инкубировали 24 ч при 37oC в инкубаторе с CO2 (Queue) (5% CO2 - 95% воздуха). Затем проводили центрифугирование со скоростью 1500 об/мин 10 мин в центрифуге IEC Centra-7R при 4oC. Надосадочная жидкость удалялась и сохранялась. Клеточный осадок повторно суспендировался в 10 мл питательной среды и центрифугировался. Надосадочный слой выливался, и клетки промывались дважды 4-5 мл питательной среды центрифугированием. Обработанные клетки переносили в 5 мл питательной среды на планшете с 6 лунками (Falcon) и инкубировали при 37oC в CO2 инкубаторе. Впоследствии по определенным дням 300 μл питательной среды собиралось в нумерованную микроцентрифужную пробирку. В каждую такую пробирку добавляли 1 μл 10% азида натрия и пробирки оставляли при 4oC. Образцы питательной среды брались через 24 ч, 72 ч, 96 ч. Твердо-фазный иммуносорбентный анализ использовался для определения действия обработки на секрецию IgM из первичной культуры лимфоцитов.
50 μл 2 мг/мл козьего антимышиного IgG (раствор 2 мг/мл) (Sigma) помещали в 12 мл бидистиллированной воды (dd H2O). С использованием многоканальной пипетки (Titer-Tek) 50 μг этого раствора помещали в каждую лунку микротитровального планшета с 96 лунками (Falkon), при этом в каждую лунку помещалось 500 μл козьего антитела против иммуноглобулина G мыши; далее проводили высушивание инкубированием в течение ночи в открытом виде в печи при 37oC. 100 μл 5%-ного бычьего сывороточного альбумина (BSA) в физиологическом растворе с фосфатным буфером (PBS) добавляли к каждой лунке 96-луночного планшета и инкубировали 1,5 ч при комнатной температуре. Жидкость удалялась опрокидыванием планшета на бумажные полотенца. Каждая лунка 96-луночного планшета промывалась PBS (1x), содержащим Tween-20 (0,05%) (Biorad), и жидкость удалялась опрокидыванием планшета на сухие бумажные полотенца.
По 100 μл каждой надосадочной жидкости добавляли к отдельным лункам 96-луночного планшета и инкубировали 1,5 ч при комнатной температуре. Лунки трижды промывали PBS/Tween-20, добавляли 100 μл козьего антитела против mu-цепи иммуноглобулина мыши, химически конъюгированного с пероксидазой хрена (Sigma) в разбавлении 1:1000 (в 1 мг/мл BSA в PBS), и инкубировали 1,5 ч при комнатной температуре. Добавляли 100 μл ABTS/ /2,21-азино-бис/3-этилбензтиазолин-6-сульфокислота// (Sigma) в 0,1 М цитратном буфере (pH 4,2) с 0,03%-ной перекисью водорода (11 μг 30% H2O2 в 11 мл ABTS), и после 20 мин инкубации при комнатной температуре планшет помещался в аутосчитывающее устройство Bio-Tek Laboratories для считывания величины абсорбции. Конъюгат настоящего изобретения значительно тормозил секрецию IgM из первичных лимфоцитов через 24 ч (фиг. 11), 48 ч (фиг. 16) и 72 ч (фиг. 17). Результаты, показанные на фиг. 15-17, иллюстрируют, что обработка конъюгатами в зависимости от дозы снижала уровень IgM в этих первичных культурах лимфоцитов. Другие обработки не смогли изменить уровни IgM ни в одной из дозировок, которые тестировались. Следовательно, уровень IgM, продуцируемого лимфоцитами, подавлялся ковалентной ассоциацией антимышиного IgM моноклонального антитела 2G10 с гелонином.
Пример 11. Исследование лимфоцитов # 2 конъюгатом 2G10-гелонин
Процедура была той же самой, что и в примере 10, за исключением того, что окончательные концентрации белка были 5 μг, 0,5 μг и 0,05 μг, или составляли эквивалентную молярную концентрацию для гелонина.
На фиг. 18-20 показано, что конъюгат настоящего изобретения сильно тормозил секрецию IgM из первичных лимфоцитов через 24, 48 и 72 ч, соответственно.
Пример 12. Исследование лимфоцитов # 3 конъюгатом 2G10-гелонин
Клетки подготавливали, как в примерах 10 и 11, за исключением того, что они были открыты для обработки (контролем, антителом, гелонином, Ab + гелонином и конъюгатом) путем инкубации 2 ч на льду. Затем клетки центрифугировали и промывали дважды средой, переносили на 6-луночные планшеты и инкубировали. Образцы надосадочного слоя собирались через интервалы в 24, 48, 72 и 96 ч. Окончательная концентрация белка была 5 μг /мл (или эквивалентная для гелонина). Был получен новый конъюгат 2G10-гелонин, и он тестировался вместе с первым конъюгатом.
На фиг. 21-24 показано, что конъюгаты настоящего изобретения явно тормозят секрецию IgM антитела из лимфоцитов через 24, 48, 72 и 96 ч, соответственно. Очевидно, что короткой инкубации иммуноконъюгатом достаточно, чтобы получить изменения в секреции IgM из клеток.
Пример 13. Слияние клеток мышиной селезенки (Balb/c) с клетками мышиной миеломы (P3 x AG63 x 8)
Питательной средой для клеток миеломы P3 x AG63 x 8 была среда Искова (Gibco) с 500 мл, содержащими 10%-ную фетальную телячью сыворотку (Hyclone) и 0,5% мл гентамицина (Trio-Bio Laboratories).
Раствор 8-азо-гуанина (20 μл /мл) был заменен на обычную питательную среду для (P3 x AG63 x 8) миеломы приблизительно за неделю до времени слияния. HAT-раствор состоял из 0,5 мл гентамицина, 5 мл 100 • HT, 5 мл 100 • A и 20% B (фетальной телячьей сыворотки). Раствор 100 • HT состоял из 136 мг гипоксантина (10 мМ) и 39 мг тимидина (1,6 мМ) в 100 мл dd H2O (подогретой до 70-80oC). 100 • A состоял из 1,8 мг аминоптерина (100 мл H2O, pH 7,8).
Слияние проводилось после предварительного промывания клеток селезенки в свободной от сыворотки и гентамицина среде центрифугированием 5 мин при 200 • g (1000 об/мин в центрифуге с верхней платформой Sorvall CLC-2B). После повторного суспендирования в 10 мл среды, свободной от сыворотки, клетки разбавлялись 1:20 (5 μл в 100 μл среды) и просчитывались в гемоцитометре.
Использовали колбы со сливающимся слоем, на 50-60% заполненные P3 клетками миеломы; клетки собирались центрифугированием и промывались 4 раза в среде, свободной от сыворотки (5 мин при 500 • g), и просчитывались. Клетки селезенки и P3 соединяли в соотношении 4:1 (8 • 107 клеток селезенки и 2 • 107 клеток миеломы) и осаждали вместе центрифугированием в течение 5 мин при 1200 об/мин. Добавлялся 1 мл холодного раствора полиэтиленгликоля-4000-диметилсульфоксида /PEG-400-DMSO/, и клетки инкубировали в этой смеси 15 с, затем медленно разбавляли PHC-DMSO, добавляя по каплям в течение 3-минутного периода 20 мл среды, свободной от сыворотки. Смесь клеток центрифугировалась при 400 • g (166 об/мин) 6 мин с целью отделения разбавителя PEG. Клетки вновь суспендировали в 50 мл 1 • HAT и распределяли многоканальной пипеткой в пять 96-луночных планшетов (Falcon) по 100 μл/лунку и инкубировали. После высевания клеток они оставались в инкубаторе на 3-4 дня до добавления 100 μл свежей HAT-среды, затем оставлялись до формирования колоний.
Пример 14. Тестирование IgM и IgG
Когда надосадочные слои клеток слияния пожелтели, определяли секрецию IgM и IgG с использованием анализа ELISA. При этом жестких 96-луночных микротитровальных планшетов (Falcon) покрывали (500 нг/лунку) козьим антимышиным IgG-антителом, используя многоканальную пипетку, и высушивали в течение ночи в открытом виде при 37oC. Раствор блокировался 5% BSA в PBS (добавлением 100 μл к каждой лунке и инкубированием 1,5 ч при комнатной температуре). После промывания 3-х PBS, содержащим 0,05% Tween-20 (Bio-Rad), по 50 μл вносили пипеткой в каждую лунку на два отдельных планшета (один для тестирования IgG и другой - для тестирования IgM). К планшету со слиянием была добавлена заменяющая питательная среда, и он инкубировался 1,5 ч при комнатной температуре до промывания 3- x PBS-Tween-20. Вторичным антителом, тестируемым на наличие IgM, было козье x мышиное IgM-антитело (специфическая пероксидаза (Sigma)), разбавленное 1:1000 в 1 мг/мл BSA в 1 x PBS. Вторичное антитело было добавлено к соответствующим планшетам и инкубировалось 1,5 ч при комнатной температуре, и затем промывалось 3 раза PBS-Tween-20. Обнаружение проводили добавлением 100 # ABTS/2,21-азино-бис/3-этилбензтиазолин-6-сульфокислота/, содержащей 0,03% перекиси водорода; планшеты считывали на аутосчитывающем устройстве микропланшетов Bio-Tek Instruments на 405 нм.
Пример 15. Слияние клеток мышиной миеломы (P3 x AG63 x 8) с клетками селезенок обработанных самок мышей Balb/c
Группы обработаны следующим образом. Группа 1: контрольные мыши - PBS; группа 2: антителом 2G10 и гелонином; и группа 3: конъюгатом. Конъюгат вводился внутрибрюшинно в концентрации 500 μг 500 μл PBS в 1-ый и 4-ый дни. Смесь антитела 2G10 и гелонина вводилась мышам в концентрации 250 μг антитела + 50 г галонина в 500 μл PBS в 1-й и 4-й дни. Контрольная инъекция делалась внутрибрюшинно в объеме 500 μл в 1-ый и 4-ый дни. На 9-ый день селезенки иссекались и клетки готовились, как описано выше. Затем каждый образец селезенки сливался с равным количеством клеток миеломы с использованием для каждого слияния 8 • 107 клеток селезенки и 2 • 107 клеток миеломы. Надосадочные жидкости тестировались неделю спустя на секрецию IgG и IgM антител в каждой группе и оценивались. Процент жизнеспособных колоний, секретирующих IgM и IgG, и те лунки, которые содержали и IgM-, и IgG-антитела, показаны на фиг. 21. Очевидно, что IgM является основной формой иммуноглобулина, секретируемого из жизнеспособных клеток, полученных от животных, которым вводили физраствор или смесь 2G10 и гелонина. Обработка иммуноконъюгатом значительно повысила (от 3 до 30%) число колоний, секретирующих IgG, и колоний, секретирующих IgG и IgM (это подтверждается специфическим обнаружением IgG и IgM в лунках с надосадочной жидкостью использованной культуры, содержащих колонии). Таким образом, иммуноконъюгат увеличивает число жизнеспособных колоний и секрецию IgG жизнеспособными колониями после слияния клеток. Эти результаты показывают действие in vivo иммуноконъюгата на экспрессию иммуноглобулина в здоровом животном после иммортализации их лимфоцитов клеточным слиянием. Обработка животных иммуноконъюгатом была бы полезна в индуцировании получения IgG in vitro после обычных процессов иммортализации клеточным слиянием в схемах продуцирования моноклонального антитела.
Пример 16. Обнаружение IgM в сыворотке предварительно обработанных KLH-иммунизированных мышей 1
Самки мышей Balb/c обрабатывались и иммунизировались по следующей схеме. В день 0 из хвостовой вены каждой мыши отбирали 150 μл крови в 0,5 микроцентрифужную пробирку, центрифугировали 10 мин и отогнанную сыворотку аккуратно вводили пипеткой в чистую центрифужную пробирку с пометкой. Эти пробирки хранили при температуре -20oC до анализа. После забора крови делалась первая обработка. Мыши делились на 4 группы (2 мыши на группу). Первая группа представляла собой контрольную и получала 500 μл физраствора с фосфатным буфером внутрибрюшинно; вторая группа также внутрибрюшинно получала инъекцию 500 μг 2G10-антитела в PBS (500 μл). Третья группа получала 100 μг гелонина (молярная эквивалентная доза в 500 μл) в 500 μл PBS и четвертая группа получала внутрибрюшинную инъекцию конъюгата 2G10-гелонин (доза 500 μг в 500 μл PBS вводилась внутрибрюшинно).
На второй день каждую мышь подвергали второй обработке, которая повторяла первую обработку, но с половинной дозой. Все инъекции делали внутрибрюшинно. На 4-й день третья обработка была той же самой, что и вторая. На 5-й день от каждой мыши была взята кровь на анализ и сделана первая иммунизация иммуногеном [хромопротеид, полученный из моллюска Megathure Crenulata, - KLH (Keyhole-limpet-hemocianin)/DNP (динитрофенол)]. Каждой мыши внутрибрюшинно вводилась доза в 250 μг DNP-KLH в 200 μл PBS. На 6-ой день внутрибрюшинно делалась четвертая обработка: контрольным мышам - 125 μл PBS; антитело - 125 μл в 125 μл PBS; гелонин - 50 μл в 125 μл PBS; конъюгат - 125 μл в 125 μл PBS.
На 8-й день делалась пятая обработка. Доза была той же самой, что и в четвертой обработке. На 10-й день забиралась кровь на анализ. Затем проводилась шестая обработка, которая была такой же, что и пятая. На 12-й день проводилась седьмая обработка. Доза составила 250 μг 250 μл, а контрольные мыши получали 250 μл PBS. На 13-й день был введен KLH бустер. Бустер был тем же самым, что и в первой иммунизации. На 20-й день забирались анализы крови.
На 20-й день анализировали образцы сыворотки. Для тестирования на IgM в сыворотке предварительно обработанных - иммунизированных KLH мышей четыре 96-луночных планшета покрывались 150 нг DNP-KLH на лунку в объеме 50 μл. С использованием многоканальной пипетки по 50 μл добавлялось к каждой лунке в четырех планшетах; их оставляли открытыми и высушивали в течение ночи при 37oC. Планшеты блокировали 100 μл/лунку 5% BSA в PBS и инкубировали 1,5 ч при комнатной температуре. После промывания (3 x PBS Tween-20) каждая проба сыворотки разбавлялась 1:500 (2 μл/1мл). 1 мг/мл BSA в PBS. 100 μл вводили в каждую лунку и инкубировали 1,5 ч при комнатной температуре. После промывания (3 x PBS Tween-20) добавляли 100 μл вторичного антитела - козья • мышиная IgM-пероксидаза (Sigma). Проба разбавлялась 1:2000 1 мг/мл BSA-PBS и инкубировалась 1,5 ч при комнатной температуре. После промывания (3 x PBS Tween-20) добавляли 100 μл ABTS с 0,03% H2O2 и проводили считывание при 405 нм в аутосчитывающем устройстве микропланшетов. На фиг. 26 показано, что конъюгат настоящего изобретения подавляет IgM-реакцию на KLH in vivo, демонстрируя эффективность иммуноконъюгата у животных.
Пример 17. Обнаружение IgM в сыворотке предварительно обработанных KLH - иммунизированных мышей II
Самки мышей Balb/c обрабатывались и иммунизировались по следующей схеме. В день 0 от каждой мыши забирали и оставляли кровь на анализ. В 1-й день мыши делились на 4 группы: группа 1 представляла собой группу контрольных мышей, которые получили 500 μл физраствора в фосфатном буфере внутрибрюшинно; группа 2 получала 100 μг (эквивалентная доза в 500 μл) гелонина в 500 μл PBS внутрибрюшинной инъекцией; группа 3 получала смесь антитела-гелонина, состоящую из 250 μг 2G10-антитела и 50 μг гелонина в 250 μл и PBS внутрибрюшинно; группа 4 получала 500 μг конъюгата 2G10 - гелонин в 500 μл PBS также внутрибрюшинно. На 4-й день делалась вторая обработка. На 5-й день была проведена KLH-иммунизация, состоящая из 250 μг KLH-DNP в 200 μл PBS. На 12-й и 18-й дни забиралась кровь. На 19-й день делалась третья обработка (такая же, как первая и вторая). На 20-й день вводился KLH-бустер (тот же, что и при первой иммунизации). На 29-й день забиралась кровь. На 36-й день сыворотка тестировалась на наличие IgM у предварительно обработанных иммунизированных KLH мышей. На фиг. 27 показаны результаты, которые схожи с результатами фиг. 26.
Представленные примеры вместе с методами, процедурами, обозначениями и специфическими соединениями, описанными здесь, предназначены для иллюстрации предпочтительных вариантов и их не следует рассматривать как ограничивающие объем изобретения. Специалистам понятно, что возможны изменения и другие использования, не выходящие за рамки изобретения, определенные формулой изобретения.
Использование: биотехнология, иммунология. Сущность изобретения: способ снижения секреции IgM-антитела из лимфоцитов заключается в контактировании лимфоцитов с цитоцидально эффективным количеством конъюгата на основе анти-IgM-антитела. Конъюгат включает цитотоксический агент гелонин, связанный с моноклональным антителом, которое селективно взаимодействует с IgM-антителом или с клетками гибридомы, продуцирующими IgM-антитело, не взаимодействует с IgG1 и IgG2 антителом и относится к G-изотипу. 4 с. и 4 з.п. ф-лы, 4 табл., 27 ил.