Данное изобретение касается в общих чертах области лечения неопластических (опухолевых) заболеваний. Более конкретно, данное изобретение касается новых иммуноконъюгатов и их применения в лечении неопластического заболевания.
Описание затрагиваемой проблемы
Неопластическое заболевание является одной из основных причин смертности и распространенности болезней в Западном мире. Неопластические состояния, например, разные типы "рака", имеют по меньшей мере одну общую характеристику, а именно, нарушения в регуляторном процессе роста клеток.
Процесс, при помощи которого нормальные клетки превращаются в злокачественные клетки, был предметом интенсивного исследования в течение десятилетий. Более недавно исследование было сконцентрировано на роли онкогенов в процессе развития рака. Онкогены представляют собой гены, которые обладают способностью трансформировать эукариотические клетки таким образом, что они растут по типу роста опухолевых клеток.
Онкоген образуется при мутации, реаранжировке или амплификации нормального гена или прото-онкогена. Одним из таких онкогенов является прото-онкоген c-erbB-2 (HER -2/neu). В дальнейшем его называют c-erb B-2. Этот ген кодирует белок, похожий на рецептор эпидермального фактора роста. Амплификация этого прото-онкогена может приводить к каскаду клеточных событий, приводящих к неконтролируемому росту клеток.
Антитела представляют собой белки, продуцируемые иммунной системой животного, обычно в ответ на чужеродные антигены или антигенные детерминанты. Антитела связываются со специфическим антигеном, к которому они направлены. Получение специфических моноклональных антител обеспечило исследователей возможным инструментом для селективного направления терапевтических агентов к клеткам, экспрессирующим избыточное количество определенных антигенов.
Была предположена избыточная экспрессия прото-онкогена c-erb B-2 при неопластической (опухолевой) трансформации клеток. Некоторые типы рака человека, в том числе рак молочной железы и некоторые карциномы яичников, обнаруживают амплифицированный ген c-erb B-2. Кроме того, амплификация и последующая избыточная экспрессия гена c-erb B-2 коррелировали с плохим прогнозом заболевания. Поэтому в этой области исследований существует большая потребность и желание разработки способа селективного направления химиотерапевтического агента к клетке, обнаруживающей избыточную экспрессию онко-гена c-erb B-2 для модулирования роста клеток, усиленно экспрессирующих этот белок. Данное изобретение обеспечивает средства для достижения этой цели.
Краткое описание изобретения
Данное изобретение обеспечивает новую композицию, содержащую конъюгат направляющей к клеткам части молекулы, например, антигенсвязывающего района, имеющего специфичность связывания в отношении белка c-erb В-2, и модулятора роста клеток, например, токсина или подавляющего рост реагента. Такая композиция может действовать как иммунотоксин для специфического направления модулятора роста клеток к опухолевым клеткам, усиленно экспрессируйщих белок c-erb В-2.
Так, в одном из вариантов данного изобретения обеспечена новая композиция, содержащая конъюгат направляющей части молекулы со специфичностью связывания в отношении белка c-erb В-2, например, TAb 250 моноклональнах антител, и цитотоксической части молекулы. Цитотоксической частью может быть токсин, вызывающее разрушение клеток лекарственное средство, цитостатическое средство или модулятор биологической ответной реакции. В одном из отдельных вариантов цитотоксической частью является гелонин.
Другой вариант данного изобретения обеспечивает способ лечения неопластического состояния, например, заболевания, характеризующегося амплификацией или избыточной экспрессией онкогена c-erb B-2, предусматривающий введение эффективной для разрушения клеток дозы иммунотоксина данного изобретения индивидууму, нуждающемуся в таком лечении.
Другой вариант обеспечивает способ убивания опухолевых клеток in vitro с последующим обратным введением их хозяину. Например, при лечении неопластического состояния костного мозга его удаляют из индивидуума, имеющего неопластическое заболевание, и обрабатывают композицией данного изобретения.
В другом варианте данного изобретения обеспечен способ предотвращения рецидива неопластического заболевания. Рецидив предотвращают введением эффективного для разрушения клеток количества направленного токсина, например, такого иммунотоксина, как TAb 250 антитела-гелонин.
Еще в одном варианте данного изобретения обеспечены новые композиции, содержащие слитые конструкции направляющих частей молекул со связывающей аффинностью в отношении белка c-erb B-2 и цитотоксической части молекулы. Предпочтительно, направляющей частью является антитело, узнающее внеклеточный эпитоп c-erb В-2, например, TAb 250, и цитотоксическая часть относительно инертна при введении ее отдельно от направляющей части, например, гелонин. В других вариантах данного изобретения обеспечены способы продления времени жизни имеющего опухоль млекопитающего путем введения направленных токсинов данного изобретения этому млекопитающему, а также способ замедления скорости роста опухолей путем введения направленных токсинов данного изобретения. В типичном случае токсины могут быть направлены к опухолевым клеткам при помощи иммунологического района связывания, например, связывающего антитела сегмента. Кроме того, обеспечена фармацевтическая композиция, содержащая иммунотоксин, состоящий из цитотоксический части, конъюгированной с моноклональным антителом. Наиболее предпочтительно, антитело представляет собой TAb 250, а цитотоксическая часть представляет собой гелонин.
Фиг. 1 демонстрирует измеренные при помощи ELISA действия антител ZME, антител TAb 250 или иммуноконъюгатов TAb 250 и гелонина на клетки SKOV-3.
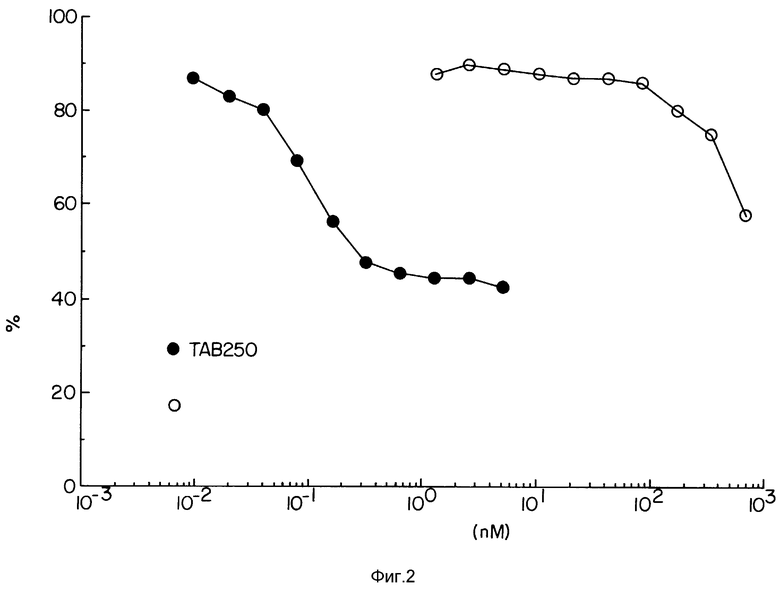
Фиг. 2 демонстрирует цитотоксичность конструкции TAb 250-гелонин на клетках SKOV-3.
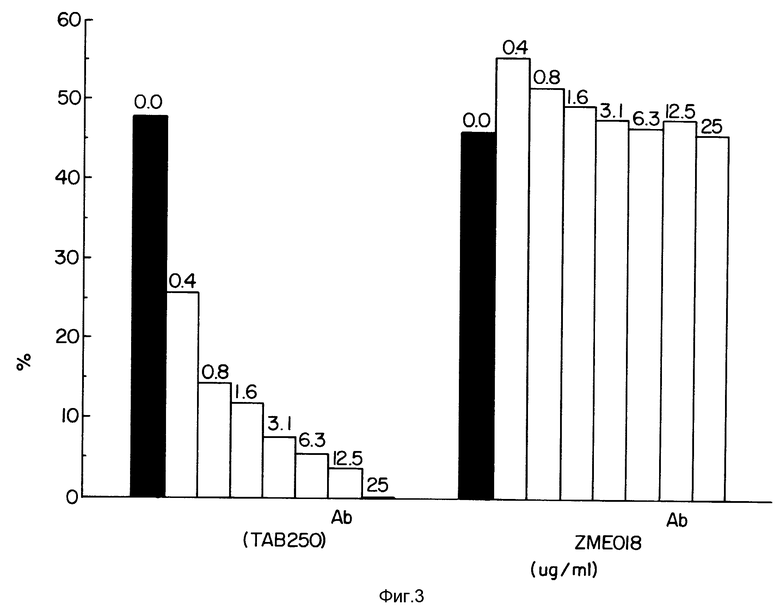
Фиг. 3 демонстрирует конкуренцию специфического (в противоположность неспецифическому) антитела с конъюгатом на клетках SKOV-3.
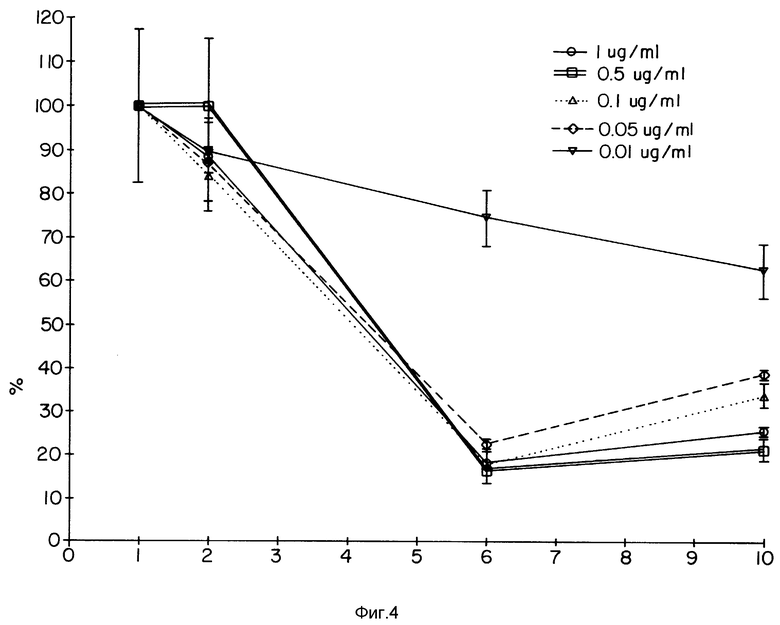
Фиг. 4 демонстрирует зависимость ответной реакции от дозы и эффекты конъюгата TAb 250-гелонин на клетках SKOV-3.
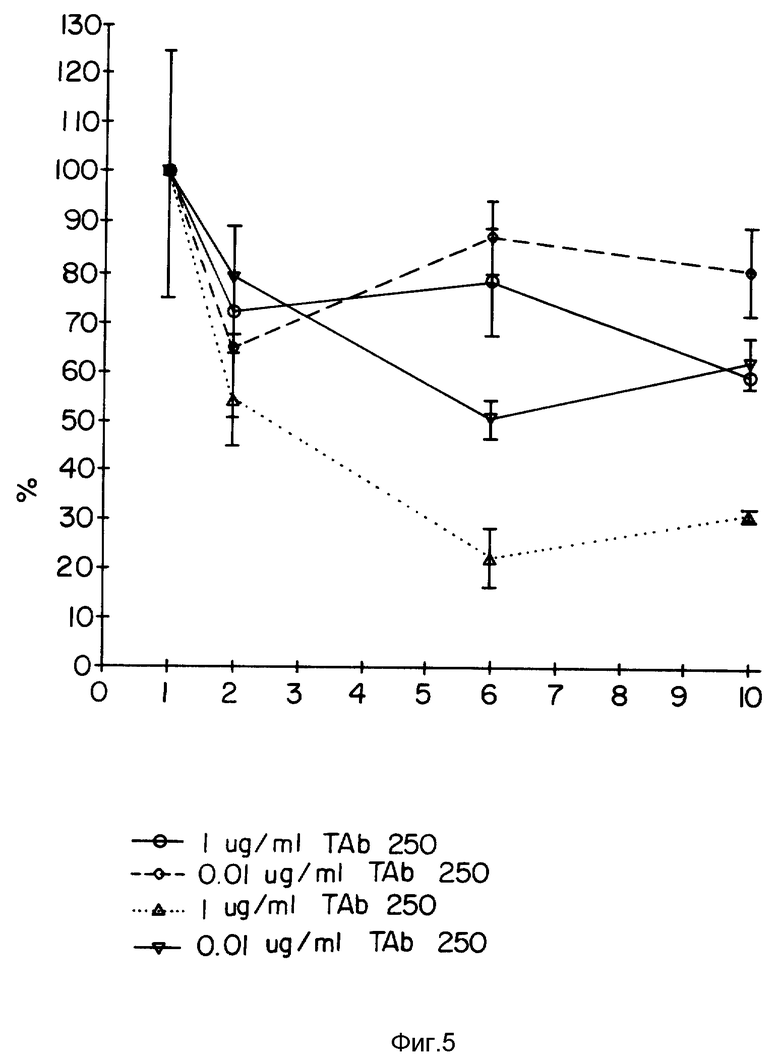
Фиг. 5 демонстрирует цитотоксичность TAb 250 и конъюгата TAb 250-гелонин на клетках SKOV-3.
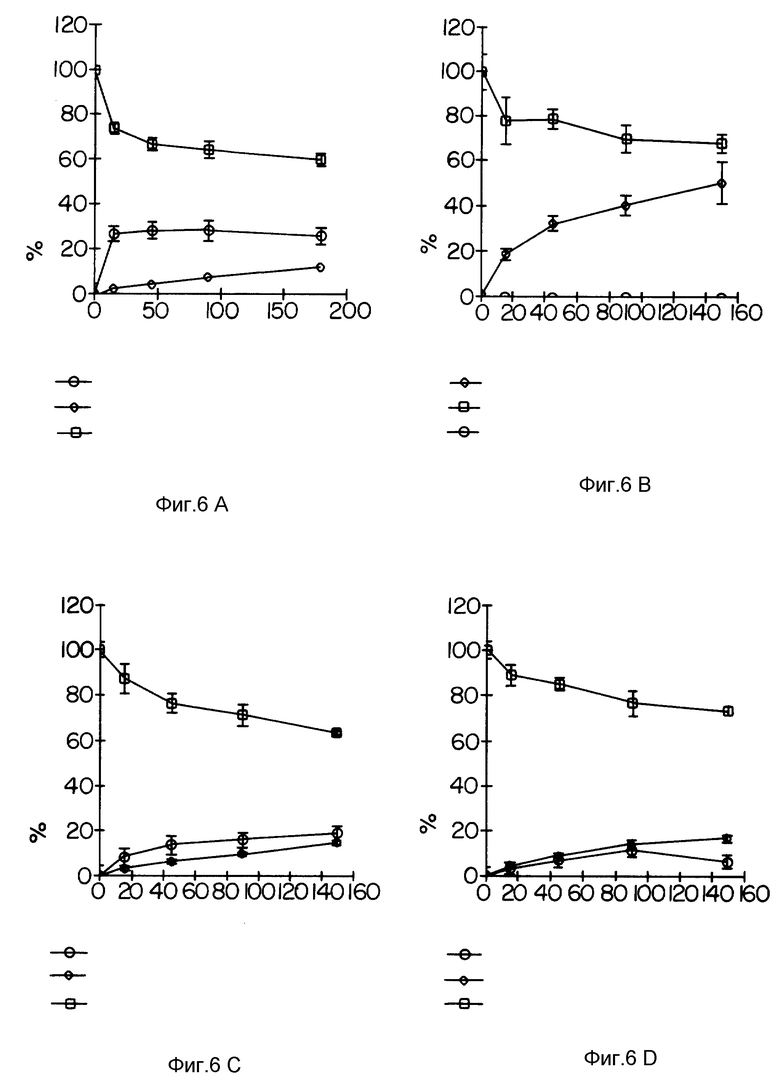
Фиг. 6 демонстрирует способность антител TAb 250 к интернализации в различные клеточные линии.
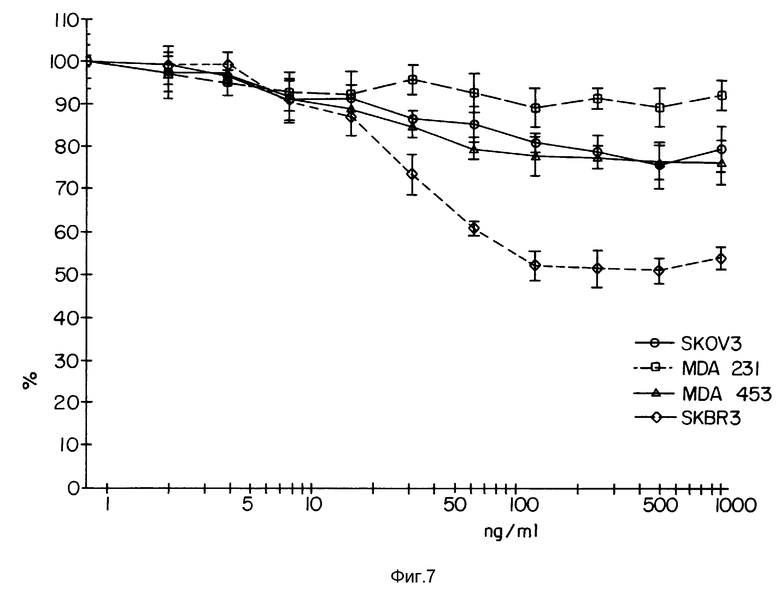
Фиг. 7 демонстрирует цитотоксичность иммуноконъюгата TAb 250-гелонин в МТТ-тесте.
Детальное описание изобретения
Как описано здесь, направляющая к клеткам часть молекулы способна селективно связываться с белком c-erb B-2, экспрессирующимся на клетке, обычно на ее поверхности. Направляющая часть содержит как лиганд, специфически связывающийся с белком c-erb B-2, так и антигенсвязывающий район, например, интактные антитела или их эпитопсвязывающие фрагменты. Это могут быть как классические молекулы антител, химерные разновидности, одиночная цепь, так и модифицированные фрагменты антител, сохраняющие эпитопсвязывающую специфичность и аффинность.
Термином "иммуноглобулин" или "пептид (пептиды) антител" называют целый иммуноглобулин или целое антитело или любой функциональный связывающий фрагмент молекулы иммуноглобулина. Примерами таких пептидов являются полные молекулы антител, фрагментны антител, такие как Fab, F(ab')2, CDR, VL, VH и любая другая часть антитела, в частности, часть антитела, обнаруживающая антигенсвязызающую специфичность и аффинность. Например, молекула антитела IgG состоит из двух легких цепей, каждая из которых соединена дисульфидными связями с двумя тяжелыми цепями. Тяжелые цепи, в свою очередь, соединены одна с другой дисульфидными связями в зоне, известной как шарнирная область антитела. Отдельная молекула IgG обычно имеет мол.массу приблизительно 150-160 кД и содержит два антигенсвязывающих сайта. Фрагменты этих молекул, например, тяжелая или легкая цепь отдельно, иногда могут связывать антиген. Антитела, фрагменты антител и индивидуальные цепи могут быть функционально эквивалентны иммуноглобулинам.
Тяжелая или легкая цепь нормального антитела имеет N-концевую (NH2) вариабельную (V) область и C-концевую (-COOH) константную (C) область. Вариабельную область тяжелой цепи называют VH (в том числе, например, Vγ), а вариабельную область легкой цепи называют VL (в том числе Vκ или Vλ). Вариабельная область является частью молекулы, которая связывается с родственным антителу антигеном, тогда как Fc район (второй или третий домены C-области) определяет эффекторную функцию антитела (например фиксацию комплемента, опсонизацию). "Легкие цепи" иммуноглобулина или антитела полной длины (в основном приблизительно 25 кД, приблизительно 214 аминокислот) кодируются геном вариабельной области на N-конце (приблизительно 110 аминокислот) и геном κ (каппа) или λ (лямбда) константной области на COOH-конце. "Тяжелые цепи" иммуноглобулина или антитела полной длины (в основном приблизительно 50 кД, приблизительно 446 аминокислот) подобным образом кодируются геном вариабельной области (кодирующим приблизительно 116 аминокислот) и одним из генов константной области, например, гамма (кодирующим 330 аминокислот). Обычно "VL" содержит часть легкой цепи, кодируемой сегментами гена VL и/или JL, (J - или соединительного района), а "VH" содержит часть тяжелой цепи, кодируемую сегментами гена VH и (или) DH (D - или diversity района) и JH. См. Roitt et al., Immunology, Chapter 6 (2d. ed. 1989) и Paul, Fundamental Immunology. Raven Press (2d ed. 1989), на которые здесь ссылаются.
Вариабельная область легкой или тяжелой цепи иммуноглобулина состоит из "каркасной" области, прерываемой тремя гипервариабельными участками, также называемыми участками, определяющими комплементарность, или CDR. Степень каркасной области и CDR была определена (см., "Seguences of Profeins of Immunological Interest", E. Kabat, et al., US Department of Health and Human Services (1987), включены здесь в виде ссылки). Последовательности каркасных областей различных легких и тяжелых цепей относительно консервативны внутри вида. Каркасные области антитела, представляющие собой объединенные каркасные области легких и тяжелых цепей, служат для расположения и построения CDR в трехмерном пространстве. CDR в первую очередь ответственны за связывание с эпитопом антигена. Их обычно называют, CDR1, CDR2 и CRD3 при последовательной нумерации от N-конца.
Два типа легких цепей, к и λ, называют изотипами. Изотипические детерминанты обычно находятся в константной области легкой цепи, называемой в общем CL, а в частности, CK или Cλ. Константная область молекулы тяжелой цепи, известная также как CH, определяет изотип антитела. Антитела классифицируются как IgM, IgD, IgG, IgA и IgE в зависимости от изотипа тяжелой цепи. Эти изотипы кодируются сегментами гена константной области тяжелой цепи мю (μ), дельта (Δ), гамма (γ), альфа (α) и эпсилон (ε), соответственно. Кроме того, имеется несколько субтипов γ.
Изотип тяжелой цепи определяет различные эффекторные функции антитела, такие как опсонизацию или фиксацию комплемента. Кроме того, изотип тяжелой цепи определяет секретированную форму антитела. Секретированные изотипы IgG, IgD и IgE обычно обнаруживают в состоящей из одной единицы или мономерной форме. Секретированный изотип IgM обнаружен в пентамерной форме, а секретированный IgA можно обнаружить как в мономерной, так и в димерной форме.
Фрагмент F(ab')2 не содержит C-концевой части константной области тяжелой цепи и обычно имеет мол.массу приблизительно 110 кД. Он сохраняет два антигенсвязывающих сайта и дисульфидные связи между цепями в шарнирной области, но не имеет эффекторных функций интактной молекулы IgG. Фрагмент F(ab')2 может быть получен из молекулы IgG протеолитическим перевариванием пепсином при pH 3,0-3,5 при помощи стандартных способов, например, описанных в Harlow and Zane, infra.
Фрагмент "Fab" содержит легкую цепь и N-концевую часть тяжелой цепи, которые соединены дисульфидными связями. Обычно он имеет мол.массу приблизительно 50 кД и содержит один антигенсвязывающий сайт. Фрагменты Fab могут быть получены из фрагментов F(ab')2 ограниченным восстановлением или из целого антитела перевариванием папаином в присутствии восстанавливающих агентов (См., Harlow and Lane infra). В некоторых случаях концентрация восстанавливающего агента, необходимая для поддержания активности папаина в присутствии атмосферного кислорода, достаточна для полного восстановления дисульфидных связей между цепями антитела. Это может привести к потере узнавания антигена. Для снятия этой проблемы папаин можно активировать и затем ввести в буфер с концентрацией восстанавливающего агента, совместимой с поддержанием антигенсвязывающей активности. Переваривание антител обычно проводят в атмосфере инертного газа для предотвращения дезактивации папаина.
Следующий ниже протокол является примером этого способа:
A) Активация папаина: папаин, полученный в виде 10 мг/мл (NH4)2SO4 суспензии, растворяют в 10 мМ ЭДТА, 20 мМ цистеине, pH 8,0, до конечной концентрации 2 мг/мл. Раствор дегазируют и инкубируют 2 часа при комнатной температуре под азотом.
B) Активированный папаин переносят в 20 мМ Na3PO4, pH 7,0, содержащий 150 мМ NaCl, 10 мМ ЭДТА, 30 мкМ ДТТ.
C) Переваривание антител: 1 мг активированного папаина добавляют к каждым 100 мг антител и раствор диализуют против большого избытка 20 мМ NaPO4, pH 7,0, содержащего 150 мМ NaCl, 10 мМ ЭДТА, 30 мкМ ДТТ при непрерывном барботировании гелием. Диализ применяют для поддержания молярного избытка восстанавливающего агента в процессе переваривания.
D) После 2-4 часов при комнатной температуре переваривание останавливают добавлением иодацетамида.
E) Фрагменты Fab отделяют от непереваренных или частично переваренных антител стандартными хроматографическими способами.
Термин "Fab" или любые другие фрагменты антител имеют классификации при применении в данном изобретении, сходные с классификациями, примененными для основных терминов "антитела" или "иммуноглобулины". Так, Fab белок "млекопитающих", "химерный Fab" и т.п. применяют аналогично соответствующим определениям общего использования, как изложено в последующих параграфах.
Терминами "химерные антитела" или "химерные пептиды" называют здесь такие антитела или пептиды антител, в которых одна часть пептида имеет аминокислотную последовательность, происходящую из соответствующей последовательности или гомологичную соответствующей последовательности в антителе или пептиде, происходящих из первого источника генов, тогда как оставшийся сегмент этой цепи (цепей) гомологичен соответствующим последовательностям другого источника генов. Например, химерный пептид тяжелой цепи антител мажет содержать мышиную вариабельную область и человеческую константную область. Два источника генов в типичном случае являются двумя разными видами, но иногда могут быть представителями одного вида.
Химерные антитела или пептиды обычно получают при помощи рекомбинантных молекулярных и(или) клеточных способов. Во многих случаях химерные антитела имеют вариабельные области как легких, так и тяжелых цепей, которые имитируют вариабельные области антител, происходящих из одного вида млекопитающего, тогда как константные и (или) каркасные части гомологичны последовательностям в антителах, происходящих из второго, отличающегося вида млекопитающих.
Однако применяемое здесь определение химерного антитела не ограничивается этим примером. Химерным является любое антитело, в котором одна или обе тяжелые или легкие цепи составлены из сочетаний последовательностей, имитирующих последовательности в антителах из различных источников, независимо от того, относятся ли эти источники к различным классам, отличающимся ответной реакцией на антиген, или к различным видам, а также от того, находится ли точка слияния в месте связи между вариабельной и константной областями или вне этого сайта. Например, химерные антитела могут быть антителами, в которых каркасная область и определяющие комплементарность области (CDR) происходят из разных источников. К примеру, нечеловеческие CDR могут быть интегрированы в человеческие каркасные области, соединенные с человеческой константной областью с образованием "очеловеченных" (humanised") антител. См., например, PCT Application Publication N WO 87/02671; US Patent N 4816567; EP Patent Application 0173494; Jones, et al., Nature, 321:522-525 (1986) и Verhoeyen, et al., Science, 239: 1534-1536 (1988), на все эти источники здесь ссылаются.
Применяемый здесь термин "подобная человеческой каркасная область" обозначает каркасную область каждой цепи антител, которая обычно содержит по меньшей мере 70 аминокислотных остатков, в типичном случае 75-85 или более остатков. Аминокислотные остатки подобной человеческой каркасной области по меньшей мере на 80%, предпочтительно на 80-85% и наиболее предпочтительно более чем на 85% гомологичны аминокислотным остаткам человеческого иммуноглобулина. Эта общая черта с другими эндогенными антителами применима для создания направляющей части молекулы, которая дает лишь небольшую иммунную реакцию, например, механизм, который сводит к минимуму ответную реакцию на аутоантигенные маркеры.
Применяемым здесь термином "очеловеченный" ("humanised") или "подобный человеческому иммуноглобулин" называют иммуноглобулин, содержащий подобную человеческой каркасную область и константную область, в основном гомологичную константной области человеческого иммуноглобулина, например, имеющую по меньшей мере 80% или более предпочтительно 85-90% или более и наиболее предпочтительно приблизительно 95% или более гомологии. Таким образом, большинство частей иммуноглобулина, подобного человеческому иммуноглобулину, за исключением, возможно, CDR, в основном гомологичны соответствующим частям одной или нескольких нативных последовательностей человеческого иммуноглобулина.
Применяемым здесь термином "гибридное антитело" называют антитело, в котором каждая цепь отдельно гомологична цепи антитела млекопитающего, но их сочетание представляет собой новый ансамбль, так что это антитело может узнавать два различных антигена. В гибридных антителах одна пара тяжелой и легкой цепей гомологична паре, обнаруженной в антителе, выработанном против одного признака узнавания антигена, тогда как другая пара тяжелой и легкой цепей гомологична паре, обнаруженной в антителе, выработанном против другого эпитопа. Это приводит к свойству мультифункциональной валентности, т.е. способности связывать по меньшей мере два различных эпитопа одновременно. Такие гибриды можно, конечно, получать также при помощи химерных цепей.
Термин "моноклональные антитела" обозначает здесь композицию антител, узнающую отдельную антигенную детерминанту. Термин применяют без ограничения в отношении источника антитела или способа, которым оно получено.
При помощи стандартных способов, хорошо известных в данной области исследований, вариабельные области и CDR могут быть получены из гибридомы, продуцирующей специфические для c-erb В-2 антитела. Последовательности нуклеиновых кислот данного изобретения, способные в конечном счете экспрессировать желаемые химерные антитела, могут быть образованы из множества различных нуклеотидных последовательностей (геномной или кДНК, РНК, синтетических олигонуклеотидов и т.д.) и компонентов (например, V-, J-, D- и C-районов), а также множеством различных способов. Соединение подходящих геномных последовательностей является в настоящее время одним из обычных способов, но можно применять также кДНК (см., European Patent Publication N 0239400 и Reichmann, Z., et al., Nature, 332:323-327 (1988), на которые дается ссылка).
Последовательности ДНК константной области человека предпочтительно выделяют из иммортализованных В-клеток, см., например, Heiter, et al., Cell, 22: 197-207 (1980), но могут быть выделены или синтезированы из множества других источников. Нуклеотидная последовательность гена CγI человеческого иммуноглобулина описана в Ellison et al., Nucl. Acids Res., 10:4071 (1982); Beidler, et al., J Immunol., 141:4053 (1988); Ziu et al., Proc. Natl. Acad. Sci. USA 84:3439 (1987) (все включены здесь ссылкой).
CDR для получения иммуноглобулинов данного изобретения предпочтительно получают из моноклональных антител, способных связываться с целевым антигеном, c-erb В-2, и продуцируют в любом удобном источнике (млекопитающем), в том числе, в клетках мышей, крыс, кроликов, хомяков или других позвоночных, способных продуцировать антитела, хорошо известными способами. Подходящие клетки для выделения ДНК и хозяйские клетки для экспрессии и секреции иммуноглобулина могут быть получены из разных источников, таких как American Type Culture Collection (АТСС) "Catalogue of Cell Lines and Hybridomas", Fifth Edition, (1985) Rpckville, Maryland, USA, включен в ссылки.
Кроме химерных пептидов антител, описанных здесь, можно легко спроектировать и приготовить иммуноглобулины, в основном гомологичные, при помощи различных способов рекомбинантных ДНК, известных специалистам в этой области. Модификации этих генов можно легко получить множеством хорошо известных способов, таких как сайт-направленный мутагенез (см. Jillman and Smifh, Jene, 8: 81-97 (1979) и Roberts, S., et al., Nature, 328:731-734 (1987), на которых здесь даны ссылки). Такими модификациями могут быть добавления аминокислот, делеции, замены, предпочтительно консервативные, и другие изменения в последовательности полипептида с сохранением однако необходимых свойств или биологической активности. Альтернативно, могут быть получены полипептидные фрагменты, содержащие только часть первичной структуры антител и обладающие связывающей и(или) эффекторной активностями. Поскольку относящиеся к иммуноглобулинам гены, подобно многим генам, содержат отдельные функциональные районы, каждый из которых имеет одну или несколько особых активностей, эти гены могут быть слиты с функциональными районами из других генов для получения слитых белков (например, иммунотоксинов), имеющих новые свойства или новые сочетания свойств.
Клонированные вариабельные и константные области можно выделить из плазмид и лигировать вместе в экспрессирующий вектор млекопитающих, например, pSV2-neo или pRSV-gpt для образования функциональной транскрипционной единицы. Предпочтительным хозяином являются мышиные миеломные клетки, такие как SP 2/0 или РЗХ, поскольку они не секретируют белок эндогенного иммуноглобулина и содержат все компоненты, необходимые для экспрессии иммуноглобулина. Миеломные клетки могут быть трансфицированы при помощи подходящих способов, как описано выше.
В этой области известны другие типы промоторов и усилителей (энхансеров), специфические для других клеток-хозяев. См., Kameyoma, K., et al., Supra.
Например, последовательность ДНК, кодирующая аминокислотную последовательность химерного антитела, может быть соединена с промоторами и энхансерами дрожжей и трансфицирована в дрожжи известными в этой области способами. См. Krigler, supra.
Тот же самый подход может быть применен для выделения c-erb В-2 специфических CDR из одного источника, такого как один вид млекопитающего, и каркасных областей другого источника, например, другого вида млекопитающего. Затем эти CDR могут быть лигированы с каркасными областями и константными областями с образованием химерного антитела. См., PCT N GB 88/00731 (1989) и U. S.S. N 07/808 462, filed Decemder 12, 1991, на которые даются ссылки. Эти CDR можно клонировать в экспрессирующем векторе, содержащем, например, человеческие каркасные и константные области.
Другим примером является рекомбинантная последовательность ДНК, кодирующая CDR1, CDR2 и CDR3 тяжелой и(или) легкой цепи одного вида, например, мыши, и каркасные области тяжелой цепи человека, для получения антитела, специфического для c-erb В-2. Другие возможности предусматривают применение CDR, специфических для c-erb В-2; применение вариабельной области, охватывающей CDR1 и CDR2 из одного вида млекопитающих, и затем лигирование этой последовательности с другой, кодирующей каркасные области млекопитающего второго вида, с CDR3 первого; или трансфекцию линии клеток хозяина рекомбинантной последовательностью ДНК, кодирующей специфические для c-erb В-2 CDR тяжелой цепи, происходящей из первого вида млекопитающего, перемешанной внутри каркасной области второго вида с последовательностью ДНК легкой цепи, содержащей ДНК вариабельной области, полученную из первого вида, и ДНК константной области, полученную из второго вида.
Экспрессирующие векторы рекомбинантной ДНК, содержащие последовательности антител, могут быть трансфицированы путем электропорации в клетки хозяина. Для выделения клонов, продуцирующих специфические для c-erb B-2 химерные антитела, применяют стандартные способы отбора.
Антитела могут быть экспрессированы с удобной степенью складчатости молекулы, в том числе в виде одноцепочечных антител, из бактерий, таких как E. coli. См. , Pluckthun, Biotechnology, 9:545 (1991), Huse, et al., Science 246: 1275 (1989) и Ward, et al., 341:544 (1989), на которых даются ссылки.
Последовательности ДЕК пептидных антител могут быть амплифицированы клонированием с применением полимеразной цепной реакции (PCR), способа, применяемого для амплификации целевой ДНК при помощи термостабильной ДНК-полимеразы, например Tag полимеразы, и олигонуклеотидных праймеров, как описано в PCR Protocols, ed. Innis, et al., Academic Press, Inc. (1990). См. также Orlandi, supra и Larrick, et al., Biotechnology, 7:934 (1989), включенные в ссылки.
Белок c-erb B-2 (называемый здесь просто c-erb В-2) представляет собой мембранный гликопротеин с мол. массой 185 кД, имеющий тирозинкиназную активность и родственный рецептору эпидермального фактора роста (EGFR), но отличающийся от него. Подобно белку FGFR, белок c-erb B-2 имеет внеклеточный домен, содержащий два богатых цистеином повторяющихся кластера, трансмембранный домен и внутриклеточный киназный домен. Кроме того, аминокислотная последовательность c-erb В-2 белка, а также соответствующая нуклеотидная последовательность описаны Coussens, et al., Science, 230:1132 (1985), включенным в ссылки.
Белок c-erb B-2 кодируется c-erb В-2 онкогеном, описанным в 1985 г. тремя разными группами исследователей: Semba, et al., Proc. Natl. Acad. Sci. USA, 82: 6497 (ген назван c-erb B-2); Coussens, et al., supra (ген назван HER-2); и King, et al., Science, 229:1132 (ген назван v-erb В). Таким образом, последовательность гена c-erb В-2 и соответствующая последовательность белка хорошо известны и описаны. Белок c-erb В-2 имеет определенную внутриклеточную часть, трансмембранный район и внеклеточный район. В типичном случае направляющие части молекул данного изобретения связываются с внеклеточным районом, который находится на наружной поверхности неопластической клетки. Направляющая часть молекулы, как правило, узнает обнаруживаемые здесь признак или признаки, в том числе лигандсвязывающий район или сайты узнавания антигена, например, эпитопы. Эпитопами могут быть чистые полипептидные эпитопы, детерминанты с линейной пептидной последовательностью или конформационные детерминанты, но также и эпитопы, имеющие углеводные компоненты. Эпитопы могут также представлять собой объединенные белок/углеводные компоненты или только углеводные компоненты. Другие модификации данного белка, нормальные или аномальные, могут представлять собой также важные эпитопные детерминанты.
Обнаружение белка c-erb В-2 может быть выполнено при помощи хорошо известных иммунотестов с применением антител, специфических для c-erb B-2 белка, например, описанных здесь. Такие антитела коммерчески доступны, например, из Chemical International, Inc., Temecula, CA, или могут быть получены стандартными иммунологическими способами. См., например, Harlow and Lane, Antibodies; A Laboratory Manual, Cold spring Harbor Publication, N.Y. (1988), включено в ссылки.
Определение c-erb В-2 белка будет здесь относиться также к белкам, происходящим из других систем хозяев, например, к белкам, которые иммунологически близки к человеческому c-erb В-2. Например, близкий крысиный ген (обозначенный neu) был сообщен Schecter, et al., Science, 229:976 (1985).
Удобными эпитопами, к которым можно легко получить антитела, являются внеклеточные эпитопы, обнаруженные на целевых клетках. Эти эпитопы в основном представляют собой белковые эпитопы, например, линейные или конформационные эпитопы белка, обнаруженные на неопластических (опухолевых) клетках. Другими применимыми эпитопами являются углеводные или иные модификации, обычно посттрансляционные, обнаруженные на c-erb В-2 белке. Антитела и другие связывающие участки, обладающие специфичностью в отношении избыточно экспрессируемого c-erb В-2, могут быть выработаны против фрагментов этого белка.
Были получены мышиные моноклональные антитела против внеклеточной части c-erb В-2. Одним из примеров таких антител является TAb 250, депонированный в АТСС Rockville, Maryland под N НВ10646.
Альтернативно, направляющая часть молекулы может быть получена любым другим способом, обеспечивающим аффинность и специфичность в отношении экспрессирующей c-erb В-2 клетки. Например, в качестве направляющей части молекулы мог бы применяться лиганд, узнаваемый и связываемый c-erb В-2 белком. См. , например, Ciccodicola, et al., (1989) Embo J. 3:1987-1991; Ciardiello, et al., (1991) Cancer Research 51:1051-1054; и Ciardiello, et al., (1991) P. N. A. S. USA 88:7792-7796, которые описывают CRIPTO, молекулу, которая, по-видимому, служит в качестве лиганда для рецептора EGF, а также связывается специфически с c-erb В-2 белком.
Что касается описываемых здесь последовательностей, следует понимать, что к ним относятся также такие их разновидности, как мутации в виде замены, добавления и(или) делеции, или любая другая последовательность, обладающая связывающей активностью, сходной с активностью последовательности, из которой она получена или которой она подобна.
Для этого изобретения антитело или другой пептид является специфическим для c-erb B-2 белка, если антитело или пептид связывают или способны связывать c-erb В-2, что измеряют или определяют стандартными тестами антиген-антитело или лиганд-рецептор, например, конкурентными тестами, тестами насыщения или стандартными иммуноанализами, такими как ELISA или RIA. Это определение специфичности применимо к одиночным тяжелым и(или) легким цепям, CDR, слитым белкам или фрагментам тяжелой и(или) легкой цепи, которые также специфичны в отношении белка c-erb В-2, если они связывают белок c-erb В-2 в отдельности или они способны специфически связывать белок c-erb В-2 после включения в конформацию иммуноглобулина с комплементарными вариабельными областями и константными областями.
В конкурентных тестах способность антитела или пептидного фрагмента связывать антиген может быть определена обнаружением способности пептида конкурировать со связыванием соединения, антигенсвязывающая способность которого известна. Известны многочисленные типы конкурентных тестов, которые здесь обсуждаются. Альтернативно, можно также применять тесты, в которых измеряют связывание тест-соединения в отсутствие ингибитора. Например, способность молекулы или другого соединения связывать белок c-erb В-2 можно детектировать мечением непосредственно целевой молекулы или целевая молекула может быть немеченой и детектироваться косвенно с применением "сэндвич"- тестов. Известны многочисленные типы тестов связывания, таких как тесты конкурентного связывания. (См. , например, U.S. Patent NN 3376110 и 4016043, Harlow and Lane, Antibodies: a Laboratory Manual, Cold Spring Harbor Publications, N. Y. (1988) и Coligan, et al., /eds/, Current Protocol in Immunolory, Wiley and Sons, N.Y., на которые здесь ссылаются). Пригодны также тесты измерения связывания тест-соединения с одним компонентом без конкуренции. Например, можно применять иммуноглобулины для идентификации присутствия белка c-erb В-2. Можно применять стандартные способы для тестирования моноклональных антител, такие как ELISA (см., Harlow and Lane, supra). Для обозрения различных продуцирующих сигнал систем, которые могут быть использованы, см. U.S. Patent N 4391904, включенный в ссылки.
Далее, специфичность связывающих частей молекул в отношении c-erb В-2 может быть определена по их аффинности. Такая специфичность существует, если константа диссоциации (KD=1/К, где К - константа аффинности) части молекулы менее 1 мкМ, предпочтительно менее 100 нМ и наиболее предпочтительно менее 1 нМ. Молекулы антител обычно имеют KD в более низких диапазонах. К = [R-L] /[R] [L] , где [R], [L] и [R-L] - концентрации при равновесии рецептора или c-erb В-2 (R), лиганда, антитела или пептида (L) и рецептор-лигандного комплекса (R-L), соответственно. В типичном случае связывающими взаимодействиями между лигандом или пептидом и рецептором или антигеном являются обратимые нековалентные связи, такие как электростатическое притяжение, ван-дер-вальсовы силы и водородные связи.
Другие тесты могут предусматривать детектирование присутствия или отсутствия различных физиологических или химических изменений, которые могут быть результатом взаимодействия, таких как отрицательная модуляция, интернализация или увеличение фосфорилирования, как описано в U.S. Patent Application N 07/644361 filed 1/18/91. См. также Receptor-Effector Coupling - a Practical approach, ed. Hulme, IRL Press, Oxford (1990).
Предпочтительный пептид, специфический для белка c-erb В-2, индуцирует увеличение фосфорилирования этого белка при помещении в контакт с опухолевыми клетками, экспрессирующими белок c-erb В-2. Молекула, индуцирующая увеличение фосфорилирования белка c-erb В-2, вызывает детектируемое увеличение во включении фосфата в белок по сравнению с включением в отсутствие этой молекулы. Обычно наблюдают двукратное или большее увеличение в фосфорилировании, предпочтительно более чем трехкратное увеличение над контролем. Фосфорилирование можно измерять способами, известными в этой области, применяемыми для детектирования фосфорилирования рецепторов. См., например, Cooper et al., Methods in Engymology, 99:387-402 (1983); Antoniades and Pantajis, Methods in Engymology, 147:36-40 (1987); и Lesniak, et al., Methods in Engymology, 150:717-723 (1987), на которые даны ссылки.
В типичном случае фосфорилирование можно измерять путем in vivo фосфорилирования интактных клеток (Lesniak, supra) или путем in vitro аутофосфорилирования (Antoniadis, supra). Для измерения in vivo фосфорилирования, например, тесты можно проводить помещением несущих белок c-erb В-2 клеток в контакт с радиоактивно меченым фосфатом. Для детектирования фосфорилирования рецептора белка c-erb В-2 в тесте ni vivo предпочтительно инкубировать тестируемые клетки в течение 12-18 часов с меченым фосфатом. Клетки делят на две или более экспериментальных серий и некоторые экспонируют с молекулами, которые предпочтительно должны увеличивать фосфорилирование, а другие являются контролями. Затем аликвоты иммунопреципитируют, рецептор детектируют, например, при помощи электрофореза в ПААГ с DDC Na или способом радиоавтографии, и статистически значимым увеличением в фосфорилировании считается двукратное или большее увеличение в пробе, экспонированной с тестируемой молекулой, по сравнению с контролями.
Для измерения in vitro аутофосфорилирования, например, клетки или клеточные экстракты можно инкубировать в присутствии или в отсутствие пептида, специфического для c-erb B-2. После иммунопреципитации антителами против c-erb B-2 иммунный комплекс можно инкубировать с γ32 Р-АТФ и анализировать при помощи радиоавтографии ПААГ с ДДС Na.
Другим предпочтительным пептидом, специфическим для белка c-erb В-2, является пептид, вызывающий отрицательное модулирование белка c-erb B-2. "Отрицательное модулирование белка c-erb В-2" определяется детектируемым уменьшением присутствия рецептора c-erb В-2 на опухолевых клетках. Такое отрицательное модулирование детектируют по снижению способности антител или других специфически связывающих частей молекул связывать или узнавать рецепторный белок c-erb В-2 на опухолевых клетках. Например, отрицательное модулирование может быть определено инкубированием опухолевых клеток, несущих рецептор белка c-erb В-2, с целевым пептидом, промыванием клеток с последующим контактированием клеток с мечеными (предпочтительно радиоактивно мечеными) антителами, специфическими для белка c-erb B-2. Степень связывания моченых антител против c-erb В-2 с клетками, экспонированными с пептидом, специфическим для белка c-erb В-2, сравнивают со степенью связывания этих антител с контрольными клетками (т.е. не экспонированными со специфическим для c-erb В-2 пептидом). Предпочтительно в этих тестах, чтобы клетки непосредственно подвергались контакту с мечеными антителами против c-erb В-2 после промывания.
Наблюдаемое отрицательное (направленное вниз) модулирование обычно зависит от дозы, т. е. степень его увеличивается с увеличением количества пептида, специфического для белка c-erb В-2, экспонированного с белком c-erb B-2. Предпочтителен пептид, вызывающий снижение на 90% или более связывания обработанными клетками по сравнению с контрольными клетками с антителами против c-erb В-2.
Другим предпочтительным пептидом, специфическим для белка c-erb B-2, является пептид, который связывает опухолевые клетки, экспрессирующие белок c-erb B-2, и интернализуется при помещении в контакт с опухолевыми клетками. "Интернализация" происходит, если пептид становится секвестрированным (разрушенным) в цитоплазме клеток. При интернализации рецептор и(или) пептид может деградироваться в лизосомах клетки или может быть вновь направлен к поверхности клетки. Способ определения интернализации лиганд-рецепторного комплекса также описан в Haigler, et al. J. Biol. Chem., 255:1239-1241 (1980). Модулятор роста клеток представляет собой молекулу, которая действует на рост клетки, к которой она направлена. В типичном случае модулятор может быть интернализован в клетку-мишень, но эта функция обычно обеспечивается интернализацией, вызываемой направляющей частью молекулы.
Модулирование обычно является снижением скорости метаболизма или роста, предпочтительно цитотоксического типа, но может быть полезным также и значительное увеличение скорости метаболизма или роста. При значительном увеличении скорости метаболизма иди роста для убивания только этих клеток можно применять яд с коротким периодом действия.
Для модуляторов, снижающих скорость метаболизма или роста, предпочтительно, чтобы модулятор был очень сильным, например, обладающим высокой активностью. Такой токсин может представлять собой неорганические или простые органические молекулы, но, обычно, более сильными являются биологические молекулы. Хотя существуют вирусные и грибные токсины, наиболее высокими удельными активностями обладают некоторые бактериальные и растительные токсины. Остановка роста может происходить в результате предотвращения любой из ряда основных клеточных функций, в том числе синтеза нуклеиновых кислот, белкового синтеза и клеточного метаболизма, основного или специфического. Например, эндотоксин Pseudomonas и дифтерийный токсин необратимо останавливают белковый синтез в эукариотических клетках. Оба ферментативно инактивируют фактор элонгации 2, являющийся основным компонентом белкового синтеза. Мишенями других токсинов могут быть другие факторы элонгации. В противоположность этому рицин, токсин растений, действует непосредственно на рибосому, действуя на 28 S pPHK.
Предпочтительно модуляторы роста имеют ферментативные активности с высоким числом оборота, так что интернализация очень малого числа молекул может убивать целевые клетки. См., Pastain, et al., Science, 254-1173-1177, данную в ссылке.
Гелонин представляет собой гликопротеин (мол. масса приблизительно 29-90000 кД), очищенный из семян Jelonium multiforum и относящийся к классу сильных инактивирующих рибосомы растительных токсинов, другими членами этого класса являются цепи абрина, рицина и модекцина. Гелонин, подобно абрину и рицину, ингибирует белковый синтез путем повреждения 60S субъединицы рибосом млекопитающих. По-видимому, гелонин является стабильным к химической и физической обработке. Кроме того, сам гелонин не связывается с клетками и обычно нетоксичен (за исключением высоких концентраций) при введении его отдельно и безопасен для манипулирования в лаборатории. Инактивация им рибосом является необратимой, не требует кофакторов и происходит с эффективностью, которая заставляет предполагать, что гелонин действует ферментативно.
Гелонин и рицин ингибируют белковый синтез и принадлежат к наиболее активным токсинам при расчете на вес белка. Гелонин в 10-1000 раз более активен в ингибировании белкового синтеза, чем A-цепь рицина. Пептиды, подобные рицину и абрину, состоят из двух цепей, A-цепи, являющейся токсической единицей, и B-цепи, которая связывается с клетками. В противоположность рицину и абрину, гелонин состоит из одной цепи и вследствие отсутствия в ней B-цепи для связывания с клетками он сам по себе относительно инертен или нетоксичен для интактных клеток. Это свойство, гораздо более низкое действие на клетки в отсутствие конъюгирования его со связывающими или направляющими частями молекул, является важным свойством для различных вариантов данного изобретения. Эта дифференциальная токсичность очень важна для высокой специфичности в отношении экспрессирующих c-erb B-2 клеток.
По-видимому, клетки млекопитающих не обладают способностью связывать и(или) интернализовать нативную молекулу гелонина. Конъюгаты гелонина с направляющим к опухоли реагентом, таким как моноклональные антитела TAb 250, направленные к опухолеспецифическому антигену, присутствующему на некоторых опухолевых клетках, обеспечивают как специфический способ связывания гелонина с клеткой, так и путь для интернализации комплекса гелонин-антитела.
Цитотоксической частью иммунотоксина может быть цитотоксическое лекарственное средство или ферментативно активный токсин бактериального или растительного происхождения или ферментативно активный фрагмент ("A-цепь") такого токсина. Предпочтительны ферментативно активные токсины и их фрагменты, представленные на примере гелонина, цепи A дифтерийного токсина, A-цепи экзотоксина (из Pseudomonas aeruginosa), A-цепи рицина. A-цепи абрина, A-цепи модекцина, α-сарцина, белков Aleurites fordii, белков диантина, белков Phytoiacca americana (PAPI, PAPII и PAP-S), ингибитора Momordica charantia, курцина, кротина, ингибитора Saponaria officinalis, митогеллина, рестриктоцина, феномицина и эномицина. Активные токсины могут действовать одним из ряда механизмов, каждый из которых влияет на физиологию и рост клеток. Токсины могут быть метаболическими ингибиторами или ядами, ингибиторами синтеза нуклеиновых кислот, ингибиторами белкового синтеза или любыми иными медиаторами аномальных или разрушительных функций. Наиболее предпочтительна конъюгация с гелонином.
Активными фрагментами и производными являются соединения, которые имеют ту же самую структуру кора, что и структура гелонина полной длины, но не содержат всей первичной последовательности. Эти фрагменты или производные имеют такую же или улучшенную биологическую или цитотоксическую активность, что и гелонин. Цитотоксичность фрагментов или производных гелонина может быть определена рутинными способами специалистами в этой области при помощи теста с применением лизата кроличьих ретикулоцитов.
Модификаторами биологической ответной реакции, которые могут быть соединены с антителами TAb 250 и применены в данном изобретении, могут быть (но не ограничиваются ими) лимфокины и цитокины, такие как IL-1, IL-2, интерфероны (α, β или γ), TNF, LT, TGF- β и IL-6. Эти модификаторы биологического ответа оказывают различное действие на опухолевые клетки. Среди этих эффектов - усиленное убивание опухолевых клеток путем прямого действия, а также усиленное убивание опухолевых клеток путем усиления процессов, медиированных защитной системой организма. Конъюгирование антител TAb 250 с модификаторами биологического ответа делает возможными селективную локализацию или направление к опухолям или клеткам, избыточно экспрессирующим - c-erb B-2, улучшенные антипролиферативные эффекты. Неспецифические эффекты, приводящие к токсичности для нецелевых клеток, сводятся к минимуму, поскольку выбранный медиатор роста клеток неэффективен в отсутствие направляющего компонента.
Цитотоксическими лекарственными средствами (и их производными), применимыми в данном изобретении, могут быть (но не ограничиваются ими) адриамицин, комплекс цис-платины, блеомицин и метотрексат. Эти цитотоксические лекарственные средства применимы для клинического лечения рецидивирующих опухолей, но их применение осложнено серьезными побочными эффектами и повреждениями, наносимыми нецелевым (здоровым) клеткам. Антитело TAb 250 может служить в качестве удобного носителя таких лекарственных средств, обеспечивая эффективное средство как для доставки к опухоли, так и для вхождения в сами опухолевые клетки. Кроме того, доставка цитотоксических лекарственных средств при помощи специфических антител к опухолям обеспечивает защиту от разрушительного действия химиотерапевтических агентов на чувствительные зоны, которые не экспрессируют избыточно c-erb B-2, такие как печень или костный мозг. Применение лекарственных средств, конъюгированных с антителом TAb 250 в качестве системы носителя, делает возможной более низкую дозировку самого лекарственного средства, так как все частицы лекарственного средства конъюгированы с антителами, концентрирующимися на неопластических клетках и обычно интернализующимися в них.
Направляющая часть молекулы и модулятор клеточного роста могут быть конъюгированы при помощи множества бифункциональных связывающих белок веществ. Примерами таких реагентов являются N-сукцинимидил-3- (2-пиридилтио)(пропионат) /SPDP/, 2-IT, 4-сукцинимидилоксикарбонил- -α- метил -α )(2 -пиридилдитио)толуол /SMPT/, бифункциональные производные имидоэфиров, такие как диметиладипимидат, HCl, активные сложные эфиры, такие как дисукцинимидилсуберат, альдегиды, такие как глутаровый альдегид, бис-азидосоединения, такие как бис-(р-азидобензоил)гександиамин, производные бис-диазония, такие как бис-(р-диазоний-бензоил)-этилендиамин, диизоцианаты, такие как толуол-2,6-диизоцианат, и бис-активные соединения фтора, такие как 1,5-дифтор-2,4-динитробензол.
Перед применением в этих исследованиях клетки Sp2/0-Ag14 растут исходно в присутствии 0,1 мкг/мл нативного гелонина. Через несколько месяцев концентрацию гелонина постепенно повышают до тех пор, пока клетки не будут поддерживаться при концентрации гелонина 10 мг/мл. Затем клетки клонируют лимитирующим разведением в присутствии 10 мг/мл гелонина и размножают образующиеся колонии, устойчивые к гелонину. Затем гелонин удаляют из культуральной среды при помощи двух пассажей и клетки вновь подвергают воздействию гелонина для подтверждения развития стабильно устойчивых клонов. После тестов для подтверждения продуцирования и активности химерных TAb 250 устойчивые к гелонину клетки SP 2/0, продуцирующие антитела, выращивают и кДНК для антител TAb 250 удаляют из тотальной ДНК инкубированием с рестриктазой. Параллельно кДНК из E. coli J M105, экспрессирующей оптимизированный гелонин, удаляют, очищают и кодирующую гелонин ДНК высвобождают после переваривания Hind 111 и Eco R1. Ген гелонина лигируют во фрагмент тяжелой цепи и эту инсерцию вводят в устойчивые к гелонину клетки SP2/0. Затем клетки субклонируют лимитирующим разведением и клоны подвергают скринингу как на продуцирование химерных антител, так и на содержание гелонина. Наконец, положительные клоны размножают и тестируют как в тестах на цитотоксичность in vitro, так и в испытаниях in vivo распределения в ткани, фармакокинетических, терапевтических испытаниях и испытаниях на токсичность. Для определения преимуществ и недостатков проводят сравнение свойств слитого белка TAb 250 гелонин с характеристиками описанных ранее конструкций TAb 250-гелонин. На основе этих исследований клиническое исследование химерного слитого белка TAb 250-гелонин можно проводить на больных с прогрессированным раком молочной железы.
Введение иммунотоксинов данного изобретения индивидууму с диагнозом наличия неопластических клеток, например, опухоли с нежелательным уровнем экспрессии онкогена c-erb B-2, позволит направлять и концентрировать цитотоксический агент в месте, где он необходим для убивания опухолевых клеток. В результате такого направления цитотоксических агентов неспецифическая токсичность для других органов, тканей и клеток будет исключена, сведена к минимуму или по меньшей мере снижена.
При применении для in vivo терапии иммунотоксины данного изобретения вводят больному или животному в терапевтически эффективных количествах, т.е. количествах, снимающих или уменьшающих предрасположение к опухолям. Обычно их вводят парентерально, предпочтительно внутривенно, но могут применяться и другие пути введения. Доза или схема приема будет зависеть от природы рака (первичная или метастатическая) и его популяции, свойств применяемого иммунотоксина, например, от его терапевтического индекса, от больного, истории болезни и других факторов. Обычно количество вводимого иммунотоксина лежит в диапазоне 0,1-10 мг/кг веса больного. Схему применения продолжают для оптимизации эффективности с одновременным соблюдением баланса против отрицательных эффектов лечения. См. Remingtons Pharmaceutical Science, 17 th Ed. /1990/ Mark Publishing Co. , Easton, Penn.; and Joodman and Jilman's: The Pharmacological Basis of Therapeutics 8 th Ed /1990/ Pergamon Press; включенные в виде ссылок.
Для парентерального введения иммунотоксины обычно готовят в пригодной для введения инъекцией форме стандартной дозировки (раствора, суспензии, эмульсии) в соединении с фармацевтически приемлемым парентеральным носителем. Такие носители предпочтительно не являются токсическими и терапевтическими. Примерами таких носителей являются вода, солевой раствор, раствор Рингера, раствор декстрозы и 5%-ный человеческий сывороточный альбумин. Можно применять также неводные носители, такие как фиксированные масла и этилолеат. В качестве носителей могут быть использованы липосомы. Носитель может содержать небольшие количества добавок, таких как вещества, усиливающие изотоничность и химическую стабильность, например, буферы и консерванты. Обычно иммунотоксин готовят в таких носителях в концентрациях приблизительно 0,1 мг/мл - 10 мг/мл.
Иммунотоксины данного изобретения можно также применять в in vitro способе. Например, способ можно применять для убивания опухолевых клеток из костного мозга. В этом способе костный мозг сначала удаляют из индивидуума, имеющего неопластическое заболевание. Затем его обрабатывают эффективной для разрушения клеток дозой иммунотоксина данного изобретения для устранения оставшихся опухолевых клеток. Обработанные клетки костного мозга можно вновь ввести больному для восстановления иммунной системы после получения интенсивной химиотерапии и(или) радиотерапии для устранения всех эндогенных ноопластических гемотоксических клеток.
Иммунотоксины данного изобретения можно также применять для продления времени жизни опухолевых млекопитающих и для замедления скорости роста опухолей, содержащих раковые клетки, у млекопитающих. Например, мышей, несущих ксенотрансплантаты опухолей человека, растущие подкожно или внутрибрюшинно, можно обработать дозами иммунотоксина, одного антитела, одного токсина или солевым раствором в дозе 25-100 мг/кг. Ингибирование роста опухоли может быть измерено по изменению в физическом размере подкожных опухолей или по удлинению времени жизни у мышей, несущих внутрибрюшинные опухоли, такие как SKOV-3-клетки. Подобные исследования могут быть полезны для разработки методологий и могут применяться для других млекопитающих, в том числе для приматов.
Нижеследующие примеры дают детальное описание получения, характеристики и применения иммунотоксинов данного изобретения. Эти примеры ни в какой мере не ограничивают данное изобретение.
ПРИМЕР 1
Очистка гелонина
Способ выделения гелонина описан в Stirpe, et al., J. Biol. Chem., 256: 6947-6953 (1980). Семена Jelonium multiflorum очищали от скорлупы и ядра растирали в гомогенизаторе с 8 объемами 0,14 M NaCl, содержащим 5 мМ фосфата натрия (pH 7,4). Гомогенат оставляли на ночь при 4oC при непрерывном перемешивании, охлаждали на льду и центрифугировали при 30000g в течение 20 минут при 0oC. Супернатант удаляли, диализовали против 5 мМ фосфата натрия (pH 6,5) и концентрировали при помощи фильтра pm10. Пробу наносили на ионообменную колонку CM-52 (20 • 1,5 см), уравновешенную 5 мМ фосфатом натрия (pH 6,5). Материал, связавшийся с ионообменной смолой, элюировали 400 мл линейного градиента NaCl от 0 до 0,3 М при скорости 25 мл/час при 4oC. Собирали фракции по 5 мл. Фракции наблюдали при 280 нм в спектрофотометре. Гелонин элюировался приблизительно во фракциях 55-70 и был последним большим пиком элюции. Фракции 55-70 объединяли, диализовали против бидистиллированной воды и концентрировали лиофилизацией. Чистоту и мол. массу каждого препарата проверяли при помощи жидкостной хроматографии высокого давления при помощи TSK 3000 гельпроникающей колонки с буфером из 50 мМ фосфата натрия с pH 7,4 и электрофореза в 15%-ном ПААГ с ДДС Na. Гелонин мигрировал в виде одной полосы с приблизительной мол. массой 29-30000 дальтон.
ПРИМЕР 2
Тест на активность гелонина
Активность гелонина определяли в тексте ингибирования бесклеточного белкового синтеза. Тест ингибирования бесклеточного белкового синтеза проводили путем последовательного добавления к 50 мкл лизата кроличьих ретикулоцитов с перемешиванием после каждого добавления следующих компонентов: 0,5 мл 0,2 М Трис-HCl (pH 7,8), 8,9 мл этиленгликоля и 0,25 мл 1 М HCl.
Затем добавляли 20 мкл смеси соль-аминокислоты-энергетический материал (SAEM), состоящей из 0,375 М KCl, 10 мМ Mg(CH3CO2)2, 15 мМ глюкозы, 0,25-10 мМ аминокислот (кроме лейцина), 5 мМ ATФ, 1 мМ ГТФ, 50 мМ Трис-HCl (pH 7,6), 10 мкл креатининфосфат-креатининфосфокиназы, 12 мкл 3H-лейцина (Amersham, 74 мКи) (ммоль) и 1,5 мкл растворов, содержащих варьирующие концентрации гелониновой смеси. Смесь инкубировали 60 минут при 30oC. Включение H-лейцина определяли в аликвоте смеси путем осаждения синтезированного белка на фильтрах из стеклянного волокна, промывания в 10%-ной ТХУ и ацетоне и мониторинга радиоактивности в Бета-счетчике с применением сцинтилляционной жидкости Aguasol. Гелонин с удельной активностью не менее 4 • 109 Е/мг был использован для конъюгирования с антителами. Единицей активности гелонина является количество белка гелонина, которое вызывает 50%-ное ингибирование включения 14C-лейцина в белок в бесклеточном тесте.
ПРИМЕР 3
Конъюгирование TAb 250 с гелонином.
Получение модифицированного 2-IT гелонина
Гелонин в солевом растворе с фосфатным буфером концентрировали приблизительно до 10 мг/мл в концентраторе Centriprep 10. Гидрохлорид триэтаноламина (TEA/HCl) с pH 8,0 и ЭДТА добавляли до конечных концентраций 60 мМ TEA/HCl и 1 мМ ЭДТА, pH 8,0. Исходный раствор 2-иминотиолана (2-IT) (500 мМ в 60 мМ буфере TEA/HCl, содержащем 1 мМ ЭДТА, pH 8,0) добавляли до конечной концентрации 1 мМ и пробу инкубировали 30 минут при 4oC под током газообразного азота при перемешивании. Избыток иминотиолана удаляли гель-фильтрацией на колонке Sephadex G-25 (1 • 24 см), предварительно уравновешенной фосфатным буфером с ЭДТА, pH 7,5, содержащим 0,01 М Na2HPO4, 0,0018 М KH2PO4, 0,0034 М KCl, 0,001 М ЭДТА и 0,17 М NaCl. Фракции анализировали на содержание белка в микротитровальных планшетах при помощи теста Bio-Rad. Гелонин элюировали в свободном объеме (приблизительно фракции 21-23). Эти фракции объединяли и хранили при 4oC.
ПРИМЕР 4
Получение моноклональных антител
BalB/c мышей иммунизировали интраперитонеально (i.p.) и подкожно (s.c.) 2 • 106 - 1 • 107 клетками NIH3T3T (клетки NIH373, трансфицированные c-erb B-2), эмульгированными 1: 1 в полном адъюванте Фрейнда. Животных реиммунизировали каждые 2-4 недели. После того как были детектированы положительные титры при помощи ELISA (описано ниже), проводили конечную i.p. или внутривенную (i.v.) иммунизацию за 4 дня до слияния. Клетки селезенки сливали с миеломными клетками РЗ-Х63Ag 8.653, поддерживаемыми в RPMI 1640, 10%-ном FBS и 2 мМ L-глутамине. Супернатанты гибридом текстировали на положительную реактивность в ELISA (см. ниже) и реактивность внеклеточного домена определяли при помощи непрямой иммунофлуоресценции с применением нефиксированных NIH3T3 и NIH3T3т клеток при 4oC с последующим анализом при помощи проточной цитометрии. Моноклональные антитела TAb 250 можно поручить из любого источника. Наиболее предпочтительными для применения в данном изобретения являются человеческие или мышиные антитела.
Гибридомные клетки, продуцирующие Tab 250, выращивали в 2-х литровом непрерывном перфузионном биореакторе. Клеточный супернатант из биореактора фильтровали и затем пропускали через систему с Protein-G-Trio с последующей ионообменной хроматографией. Затем материал концентрировали и стерильно фильтровали. Испытание конечного продукта предусматривает тесты на тотальную ДНК, на чистоту белка, pH (IEF), общий белок, эндотоксин, силу, идентичность и антиген белка - G.
Данное изобретение, Inter alia, использует химерное антитело, в том числе гибридное антитело или "очеловеченное" (humaniged) или подобное человеческому антитело. В предпочтительном варианте вариабельная последовательность происходит из последовательности мышиного TAb 250 антитела и в основном идентична ей. Такое химерное антитело названо BACh 250.
ПРИМЕР 5
Твердофазный иммуноферментный анализ (ELISA)
Стериальные планшеты на 96-ячеек предварительно обрабатывали в течение 2 часов при 37oC бычьим коллагеном в концентрации 1 мг/мл в стерильном PBS. NIH3T3T клетки (NIH353 клетки, трансформированные вектором) выращивали до 80% конфлюэнтности и собирали теплым версеном Пука (0,02% ЭДТА в PBS), промывали и обрабатывали 10%-ным нейтральным содержащим буфер формалином с последующей стадией блокирования 1%-ным бычьим сывороточным альбумином BSA/PBS. Супернатанты проб или разведения антитела добавляли затем на планшеты и инкубировали в течение 2 часов при 37oC с последующим инкубированием с конъюгированными со щелочной фосфатазой козьими вторичными антителами, специфическими для антител против Fc мышиного IgG в течение 1 часа при 37oC. Планшеты промывали PBS, добавляли р-нитро-фенилфосфат и диэтаноламин в качестве субстрата и инкубировали 15 минут при комнатной температуре, после чего измеряли A405. Супернатанты или антитела, которые реагировали с трансфицированными клетками при поглощении на 0,2-1,0 большем, чем поглощение для отрицательных контрольных антител, рассматривались как положительные.
ПРИМЕР 6
Получение и процессирование 125I-TAb 250
TAb 250 радиоактивно метили с применением Iodobeads/Piercet в соответствии с предписаниями изготовителя. Проводили реакцию не содержащего носителя Na1251 (400 мкКи IMS.30. Amersham) с 25 мкг TAb 250 в 100 мМ Na-фосфатном буфере (200 мкл, pH 7,4) в присутствии 3 Iodobeads (иодогранул). Это привозило к приблизительному соотношению 1 атом иода/молекулу IgG. Включение проводили при комнатной температуре 7,5 минут при прерываемом перемешивании. Реакционную смесь удаляли из гранул и после 5 минут объем доводили до 0,5 мл Na-фосфатным буфером. 2 мкл брали для оценки удельной активности (см. ниже). Оставшийся объем обессоливали гель-фильтрацией при помощи колонки АР-5 (Pharmacia), уравновешенной PBS, содержащим 0,1%-ный BSA и 0,02%-ный азид. Радиоактивно меченые антитела элюировали в 1 мл колоночного буфера и хранили при 4oC до 6 недель без видимой потери связывающей активности. Обессоленный материал был практически свободен от невключившегося иода, поскольку более 95% осаждалось ТХУ.
Удельную активность радиоактивно меченых антител оценивали осаждением при помощи ТХУ материала перед стадией обессоливания. Так, 2 мкл реакционной смеси разбавляли в 500 раз колоночным буфером и аликвоты в двух повторностях смешивали с равным объемом охлажденной льдом 20%-ной ТХУ. Через 25 минут стояния на льду осажденный материал собирали центрифугированием (10 мин, 3000g). Супернатанты и осадки просчитывались отдельно и включение выражали в процентах осаждаемых ТХУ импульсов. Включение, полученное в отдельных иодинированных материалах, составляло 27% - 45%, давая оценки удельной активности 3,9-7,2 мкКи/мкг. Перед каждым экспериментом по связыванию требуемое количество 125I-TAb 250 обессоливали гель-фильтрацией на NAP-5 колонке, уравновешенной буфером связывания. Эта процедура удаляла азид и давала материал, который обычно более чем на 98% осаждался ТХУ.
ПРИМЕР 7
Клеточная культура
Использовали клеточные линии аденокарциномы молочной железы человека SKBR-3, MDA-MB-453, MDA-MB-231 и аденокарциномы яичников SKOV-3, SKBR-3, MDA-MB-231 и MDA-MB-453 клетки поддерживали в минимальной основной среде с добавлением 10% FBS и 2 мМ L-глутамина. Среда для клеток MDA-MB-453 содержала также 1% незаменимой аминокислоты и 1% витаминов. Клетки SKOV-3 культивировали в модифицированной Iscove среде Dulbecco с 10% FBS и 2 мМ L-глутамина. Все культуры инкубировали при 37oC либо в 5%, либо в 10% CO2.
ПРИМЕР 8
Интернализация 125I-TAb 250
Интернализацию 125I-TAb 250 оценивали путем определения количества радиоактивности в чувствительных к кислоте и нечувствительных компартментах. Клетки собирали и ресуспендировали в охлажденном льдом буфере связывания с одним 125I-TAb 250 (6 нг/мл - 153 нг/мл) или с избытком немеченого TAb 250 для определения неспецифического связывания. После того как связывание клеточной поверхностью радиоактивно меченых антител достигали равновесия, клетки осаждали при 200g в течение 5 минут при 4oC и промывали трижды ледяным буфером связывания для удаления несвязавшихся антител. Клеточные осадки ресуспендировали в ледяной среде связывания и брали аликвоты для определения количества исходного поверхностного связывания 125I-TAb 250. Для инициации интернализации радиоактивно меченого антитела клетки подогревали до 37oC. В периоды от 15 до 150 минут удаляли аликвоты и клетки собирали центрифугированием (1400g, 5 минут, 4oC). Супернатанты, которые содержали диссоциированные или рециркулирующие антитела, собирали. Осадки ресуспендировали дважды в кислой промывке (100 мкл PBS на пробирку, 1% глюкозы, pH 1). Супорнатанты, содержащие связанные с поверхностями клеток антитела, объединяли и просчитывали на радиоактивность. Кончики пробирок, содержащие остаточную связанную с клетками радиоактивность, отрезали и просчитывали.
Фиг. 6 демонстрирует, что моноклональные антитела TAb 250 не интернализуются в клетки MDA-MB-231 (B). В противоположность этому клетки SKBR-3 интернализовали антитела TAb 250 наиболее эффективно (A), тогда как интернализация антител в клетки SKOV-3 и MDA-MB-453 была промежуточной (C и D, соответственно).
ПРИМЕР 9
Модификация моноклонального антитела TAb 250 при помощи SPDP
N-сукцинимидил-3-(2-пиридилдитио)(пропионат) (SPDP) в диметилформамиде готовили в виде исходного раствора 3 мг/мл в сухом диметилформамиде. Поскольку кристаллический SPDP мог подвергаться гидролизу, истинную концентрацию химически реактивного сшивающего средства определяли спектрометрически путем определения поглощения при 260 нм в двухлучевом спектрофотометре. Концентрацию исходного раствора SPDP рассчитывали из уравнения: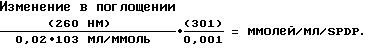
Один миллиграмм моноклональных антител TAb 250 в 1,0 мл солевого раствора с фосфатным буфером (PBS) добавляли в стеклянную пробирку. Исходный раствор SPDP медленно добавляли в пробирку при приблизительно 5-кратном молярном избытке при постоянном перемешивании. Смесь инкубировали 30 минут при комнатной температуре с перемешиванием через каждые 5 минут в течение инкубационного периода.
Избыток непрореагировавшего SPDP удаляли из пробы гель-фильтрационной хроматографией на Sephadex G-25 (1 • 24 см), предварительно уравновешенной 100 мМ Na-фосфатным буфером, рН 7,0, содержащим 0,5 мМ ЭДТА (буфер A). Фракции (0,5 мл) собирали и анализировали на содержание белка по методу Бредфорда (Bradford, Anal. Biochem. 72: 248-254 (1976)). Поглощение (600 нм) прослеживали в планшете из 96 ячеек при помощи автоматизированного считывающего устройства Bio-TEK Microplate. Антитела элюировались в свободном объеме (фракции 14-20) и их объединяли и хранили при 4oC. Белок концентрировали в микроконцентраторе Centricon-30. Удержанный материал промывали 100 мМ Na-фосфатным буфером с pH 7,0, содержащим 0,5 мМ ЭДТА. Антитела концентрировали до конечного объема приблизительно 0,5-0,75 мл.
ПРИМЕР 10
Конъюгирование модифицированных SPDP моноклональных антител TAb 250 с модифицированным иминотиоланом гелонином
Конъюгирование модифицированного 2-IT гелонина TAb 250
TAb 250-гелонин, соединенный с SMPT, готовят соединением модифицированного 2-IT гелонина с модифицированными SMPT моноклональными антителами TAb 250. Вкратце, для модификации TAb 250 SMPT 10 мг антител в 1,0 мл PBS разбавляют 1: 1 2-х боратным буфером (0,05 М борат натрия, 1,7% хлорид натрия, pH 9,0) и медленно добавляют к раствору антител 52 мкл 4 мМ SMPT в сухом DMF (диметилформамиде)). Реакцию инкубируют при комнатной температуре 2 часа под N2 при перемешивании. Избыток SMPT удаляют проведением реакционной смеси через Sephadex G-25 колонку, содержащую фосфатный буфер с ЭДТА, pH 7,5, и содержащие антитела фракции оценивают при помощи Bio-Rad - теста. Фракции объединяют и хранят при 4oC под N2. Сшивание с 2-IT проводят при 27oC под N2 с перемешиванием в течение 96 часов. Конечный продукт очищают, как описано для SPDP в примере 9. Один миллиграмм очищенного гелонина (2 мг/мл в PBS), полученного, как описано в примере 1, модифицировали иминотиоланом, как описано в примере 3. Моноклональные антитела, модифицированные, как описано в примере 9, смешивали с равным весом модифицированного гелонина. Эта пропорция соответствовала 5-кратному молярному избытку гелонина по сравнению с антителами. pH смеси доводили до 7,0 добавлением 0,05 М TEA/HCl буфера с pH 8,0 и смесь инкубировали 20 часов при 4oC под азотом. Иодацетамид (0,1 М) добавляли до конечной концентрации 2 мМ для блокирования оставшихся свободных сульфгидрильных групп и инкубирование продолжали еще в течение часа при приблизительно 25oC. Реакционную смесь хранили при 4oC до очистки при помощи гель-фильтрации.
ПРИМЕР 11
Очистка комплексов гелонин-моноклональные антитела TAb 250
Неконъюгированный гелонин и низкомолекулярные продукты удаляли из реакционных смесей примера 10 гель-фильтрацией на колонке Sephadex S-300 (1,6 • 31 см), предварительно уравновешенную PBS.
Реакционные смеси из примера 10 концентрировали приблизительно до 1 мл при помощи микроконцентратора Centricon 30 перед нанесением на колонку Sephadex. Колонку промывали PBS. Фракции по 1 мл собирали и аликвоты по 50 мкл анализировали на белок при помощи метода Бредфорда.
Неконъюгированные антитела удаляли из конъюгированных с гелонином антител аффинной хроматографией на колонке (1 • 24 см) Blue Sepharose CL-6B, предварительно уравновешенной 10 мМ фосфатным буфером, pH 7,2, содержащим 0,1 М NaCl. После нанесения пробы элюата S-300 колонку промывали 30 мл того же буфера для полной элюции неконъюгированных антител.
Конъюгированные с гелонином антитела, связанные с колонкой, элюировали линейным солевым градиентом 0,2-2 М NaCl в 10 мМ фосфатном буфере с pH 7,2. Комплекс антитела-гелонин элюировался приблизительно при 0,7 М NaCl. Содержание белка элюированных фракций определяли по Бредфорду. Содержащие белок фракции объединяли и распределение белков в элюированных фракциях подтверждали электрофорезом на градиентном 5-20% невосстанавливающем полиакриламидном геле. Проходящий через колонку пик (фракции 14-20) содержал только свободные антитела, тогда как фракции 50-80, элюированные высокой концентрацией соли, содержал конъюгат TAb 250-гелонин, свободный от неконъюгированных гелонина или антител. Конечный продукт содержал антитела TAb 250, сопряженные с 1, 2 и 3 молекулами гелонина. Среднее содержание гелонина было 1,5 молекул на молекулу антител. Для оценки активности гелонина практически чистого комплекса гелонин-TAg 250 применяли систему трансляции in vitro кроличьих ретикулоцитов. За одну единицу активности в таком тесте было взято количеством белка, необходимое для обеспечения 50%-ного ингибирования белкового синтеза по сравнению с необработанными контролями. При помощи этого теста были определены удельная активность нативного гелонина (2 • 108 Е/мг) и удельная активность TAb 250-гелонин-конъюгата (8,2 • 105 Е/мг). Практически чистый комплекс гелонин-TAb 250 активен в тесте с лизатом ретикулоцитов. Разведение 1:1000 исходной пробы вызывало приблизительно 50%-ное ингибирование белкового синтеза, т.е. 50%-ное снижение включения 14C-лейцина в белок. Таким образом, активность исходного препарата была 1000 Е/мл.
К композициям данного изобретения относятся также слитые конструкции моноклональных антител TAb 250 и цитотоксической части молекулы. Слитые конструкции иммунотоксина данного изобретения можно приготовить следующим способом. Нуклеотидную последовательность V областей H- и L-цепей Tab 250 можно легко определить. Например, из продуцирующих TAb 250 клеток выделяют общую РНК при помощи тиоциананта квандиния. Поли/A/+-РНК можно выделить при помощи хроматографии на олиго /dT/-олигоцеллюлозе. Требуемые гены можно выделить при помощи стандартных способов, в том числе обратной транскрипции и способа PCR (попимеразной цепной реакции). Цепь кДНК можно синтезировать из выделенной мРНК с применением олиго-dT-праймера и обратной транскриптазы. Праймер вблизи поли-A-"хвоста" мРНК может быть основан либо на последовательности поли-A, либо на общих смежных последовательностях, обнаруженных в мышиных иммуноглобулинах. См. Devereaux, Jenetic Computer Jroup, University of Wisconsin Biofechnology Centre and its associated seguence databases.
Праймер на другом конце гена может быть выбран из общих последовательностей, обнаруженных в мышиных иммуноглобулинах. См. Orlandi, et al., (1989) PNAS 86:3833-3837 и Larrick, et al., (1989) Bio / Thehnology 7:934-938. Ген тяжелой цепи TAb 250 имеет 5' предшествующую (upstream) последовательность АТАТАГ ЦАГГАЦ ЦАГГАЦ и начинает кодирование с АТГАА СТТГГ ГГЦТЦ. Ген легкой цепи имеет 5' - предшествующую (upstream) последовательность ТТТАЦ ТТЦЦТ ТАТТТ и начинает кодирование с АТГГГ, ЦАТЦА АГАТГ. Эти праймеры могут быть использованы для амплификации этих генов способом PCR и клонированы в плазмидные экспрессирующие векторы.
Трансфекция ДНК в мышиные клетки при помощи электропорации
Для этих генов можно применять стандартные способы трансфекции. Например, ДНК можно вводить в клетки мышиных гибридом Sp2/O-AG14 при помощи электропорации. 1-2 • 103 активно растущих клеток SP2/O-AG14 промывают и ресуспендируют в 1,0 мл стерильного РВS. 30 мкг каждой из химерных, IgK и IgG1, плазмид добавляют к клеточной суспензии. ДНК/клетки переносят в предварительно охлажденную электрошоковую кювету, инкубируют на льду не менее 5 минут и затем подают электроимпульс 0,5 кв/см в течение 10 мсек. (Transfector 300, ВТХ). После электрошока смесь ДНК/клетки возвращают на лед, выдерживают 10 минут, разбавляют в 10 мл DMEM, содержащей 5% NCTC-109 и 10% FCS, и инкубируют при комнатной температуре 10 минут. Наконец клетки переносят в инкубатор при 37oC с 7% CO2 и выдерживают в нем 48 часов перед посевом в селективную среду, содержащую 1 мкг/мл ксантина. Клетки можно высевать на планшеты с 96 ячейками при 3/104 клеток на 1 ячейку. Супернатанты культур анализируют при помощи ELISA на антитела, связанные с положительными (содержащими антиген TAb 250) целевыми клетками.
ПРИМЕР 12
Определение МТТ
Определения бромида 3-(4,5-диметил/тиазолил)-2,5-дифенилтетразолия (МТТ ) проводили путем удаления клеток из колб для культуры ткани версеном 1: 5000, центрифугирования при 500g в течение 5 минут и ресуспендированием клеток в среде при концентрации 1 • 105 клеток/мл. Клетки высевали по 100 мкл на ячейку в 96-ячейковые микротитровальные планшеты и инкубировали в увлажненном CO2-инкубаторе при 37oC в течение 24 часов.
На следующий день добавляли TAb 250 или TAb 250-гелонин. Сразу же после помещения наивысшей концентрации антител в первую колонку ячеек проводили разведения 1:2 непосредственно в микротитровальных планшетах при помощи многоканальной пипетки. Затем планшеты инкубировали в течение 3 дней с последующим добавлением 10 мкл МТТ на ячейку. МТТ готовили в виде раствора 5 мг/мл PBS, стерильно фильтровали и хранили при 4oC в темноте. Затем пластинки инкубировали дополнительно 4 часа в темноте при 37oC. Кристаллы МТТ растворяли смешиванием содержимого с 100 мкл изопропанола, содержащего 0,04 N HCl и 3% додецилсульфата натрия. Поглощение при 570 нм определяли с применением считывающего устройства твердофазного иммуноферментного анализа (ELISA).
ПРИМЕР 13
Тест пролиферации клеток (CPA)
Тест пролиферации клеток (CPA) измеряет рост клеток путем определения числа клеток и их жизнеспособности. В 0-ой день клетки при 80-90% конфлюэнтности высвобождали из колбы для культуры ткани, осаждали при 200g при 20oC в течение 6 минут и ресуспендировали в MEM ISCOVE, содержащей 2 мМ глутамина и 10% плодной бычьей сыворотки до концентрации 6000 клеток/мл. Суспензию клеток добавляли на планшеты с 24 ячейками по 1 мл на ячейку. Планшеты инкубировали при 37oC, 5% CO2, в течение 24 часов. На первый день клетки в трех ячейках промывали PBS и высвобождали из планшета при помощи 1 мл 0,05% трипсина в PBS и аликвоты 500 мкл считали при помощи Caulter Counter. К оставшимся ячейкам добавляли 20 мкл PBS, TAb 250 или TAb 250-гелонин при концентрациях, указанных на чертежах. Планшеты возвращали в инкубатор. Во временных точках, указанных на чертежах, клетки удаляли трипсином и считали, как и в первый день. Оставшиеся 500 мкл клеток окрашивали иодидом пропидия для определения жизнеспособности клеток при помощи проточной цитометрии. Для каждой точки на графике определяли среднее количество клеток (n = 3) и умножали на процент живых клеток для определения количества жизнеспособных клеток. Это количество делили на количество живых клеток, обработанных PBS, для выражения процента от контроля на оси Y.
ПРИМЕР 14
Цитотоксичность гелонина и комплекса гелонин-антитела TAb 250
Как можно видеть на фиг. 1, антитела ZME практически не влияли на клетки SKOV-З. В противоположность этому, иммуноконъюгат TAb 250-гелонин был высокоактивным.
Фиг. 2 иллюстрирует цитотоксичность иммуноконъюгата TAb 250-гелонин на клетках SKOV-З по сравнению с одним гелонином. При той же концентрации иммуноконъюгат TAb 250-гелонин был приблизительно в 10000 раз более цитотоксичен, чем один гелонии.
Фиг. 3 демонстрирует, что не относящиеся к исследуемой проблеме антитела, моноклональные антитела ZME 18, не обладали конкурентным действием на цитотоксичность иммуноконъюгата TAb 250-гелонин. В противоположность этому, увеличивающиеся концентрации моноклональных антител TAb 250 снижали цитотоксичность иммуноконъюгата зависимым от дозы образом.
Фиг. 4 иллюстрирует зависимое от дозы действие иммуноконъюгата TAb 250-гелонин на клетки SKOV-3. Как видно в CPA-тесте, доза более 0,1 мкг/мл вызывала 80%-ное ингибирование при 6 днях воздействия.
Фиг. 5 демонстрирует действие либо одного моноклонального антитела TAb 250, либо конъюгата TAb 250 с гелонином на клетки SKOV-З. Как можно видеть, существует зависимое от дозы ингибирование иммуноконъюгатом TAb 250-гелонин в CPA-тесте.
Фиг. 7 изображает действие конъюгата TAb 250/гелонин на четыре разные клеточные линии. Как и следовало ожидать, наибольшая токсичность наблюдается на линии клеток SKBR-3. Промежуточная токсичность была обнаружена на клетках SKOV-З и MDA-MB-453, тогда как на клетках MDA-MB-231 практически не наблюдалась цитотоксичность. Таким образом, цитотоксичность иммуноконъюгата TAb 250-гелонин коррелирует с количеством рецепторов клеточной поверхности в этих клетках.
Таким образом, очевидно, что данное изобретение вполне пригодно для достижения целей и получения результатов, изложенных вначале. В способе и устройстве могут быть сделаны определенные изменения без отхода от духа и сферы действия данного изобретения. Понятно, что возможны изменения, и предполагается, что каждый элемент или стадия, перечисленные в формуле изобретения, следует относить ко всем эквивалентным элементам или стадиям при достижении тех же результатов тем же или эквивалентным способом. Предполагается расширение сферы действия данного изобретения, в какой бы форме ни применялись его принципы. Таким образом, данное изобретение хорошо приспособлено для достижения целей и получения результатов и упомянутых преимуществ, а также и иных присущих ему преимуществ.
Изобретение относится к медицине и касается иммунотоксина, обладающего неопластическим действием, композиции, обладающей неопластической активностью, содержащей иммунотоксин, и способа обработки неопластической клетки. Задача изобретения заключается в разработке способа селективного направления химиотерапевтического агента в клетке. Сущность изобретения состоит в том, что иммунотоксин, обладающий неопластическим действием, представляет собой конъюгат гелонина и моноклонального антитела, проявляющего специфичность связывания в отношении антигенного домена с - erb В-2 и представляющего собой Tab 250. Композиция, обладающая неопластической активностью, представляет собой иммунотоксин и фармацевтически приемлемый носитель. Способ обработки неопластической клетки включает введение эффективной дозы иммунотоксина в эту клетку. Технический результат заключается в усилении селективности доставки химиотерапевтического агента к клетке. 3 с. и 7 з.п. ф-лы, 7 ил.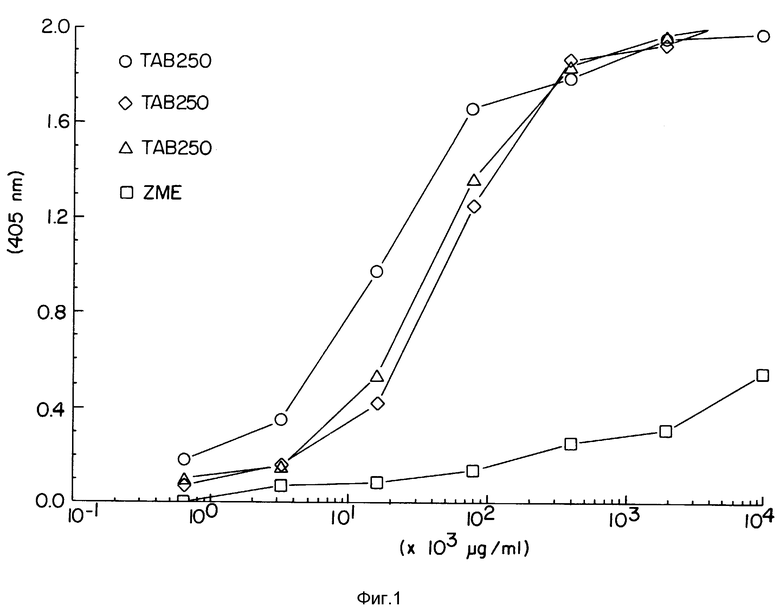
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Штамм гидридных культивируемых клеток животных MUS мUSсULUS,используемый для получения моноклональных антител к дифференцировочному антигену В-лимфоцитов человека | 1987 |
|
SU1420020A1 |
| УСТРОЙСТВО ДЛЯ АВТОМАТИЧЕСКОЙ ПОДАЧИ ЛЕНТОЧНОГО и ПОЛОСОВОГО МАТЕРИАЛА | 0 |
|
SU206533A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Tecce et al., Production and characterization immunotoxines to distinct epitops of the extracellular domain of the Hep-2 qp.185, Anticancer Research, 1990, v.10, N5, p.1454 | |||
| Martin D | |||
| el al., Role of Murine tumor models in cancer treatment Research, Cancer Research, 1986, v.46, pp.2189 - 2192. | |||

