Данное изобретение касается в общих чертах области лечения неопластических /опухолевых/ заболеваний. В частности, данное изобретение касается новых иммуноконъюгатов и их применения в лечении неопластического заболевания. Более конкретно, изобретение касается новых иммуноконъюгатов, цитотоксических в отношении лейкозных клеток, характеризующихся экспрессией антигена CD 33.
Неопластическое заболевание является одной из основных причин смертности и распространенности болезней в Западном мире. Все неопластические заболевания или типы "рака" имеют по меньшей мере одну общую характеристику, а именно, нарушения в регуляторном процессе роста клеток.
Антигены, расположенные на поверхности раковых клеток, были применены в дифференцировке лимфолейкоза и не лимфатического лейкоза, субтипировании острого миелопоэтического лейкоза, предсказании терапевтического результата, а также в терапии in vivo или ex vivo при помощи очистки костного мозга. Антигены, определяющие клетки не лимфатического лейкоза, применимы также в идентификации нормальных гематопоэтических /кроветворных/ клеток во время ранних стадий их развития.
Антиген CD 33 представляет собой гликопротеин с мол. массой 67 кД, обнаруженный на нормальных гранулоцитах-моноцитах колониеобразующей единицы /CFU-GM/, на фракции эритроидов бурст-образующей единицы (области дифференцировки ранних клеток-предшественников эритроидного ряда: BFU-E и CFU -гранулоцит, эритроид, моноцит, мегакариоцит) CFU-GEMM и отсутствующий в нормальных полипотентных эмбриональных клетках.
Антитела представляют собой белки, обычно продуцируемые иммунной системой животного в ответ на антигенные детерминанты. Антитела связываются со специфическим антигеном, на который они нацелены. Выработка специфических моноклональных антител обеспечила исследователей возможным инструментом для избирательного направления химиотерапевтических агентов к клеткам, которые экспрессируют избыточное количество опухолеспецифических антигенов.
Иммунотоксины представляют собой гибридные молекулы, состоящие из моноклональных антител, ковалентно связанных с токсином. Иммунотоксины имеют несколько возможных преимуществ по сравнению с обычными противоопухолевыми средствами, в том числе избирательность в отношении опухолевых клеток и способность потенциальной доставки исключительно сильных токсинов.
Присутствие антигена CD 33 на клетках острого не лимфатического лейкоза и клетках острого миелопоэтического лейкоза предоставляет возможность усовершенствования селективных иммунотоксинов. В настоящее время не существует таких эффективных иммунотоксинов. Поэтому по-прежнему существует в этой области исследований огромная необходимость и желание получения соединений и разработки способов селективного уничтожения лейкозных клеток.
Данное изобретение обеспечивает новую композицию, содержащую конъюгат антигенсвязывающего района, проявляющего специфичность связывания относительно белка CD 33, и модулятора роста клеток. Такая композиция действует как иммунотоксин, специфически убивая опухолевые клетки, отличающиеся экспрессией белка CD 33.
Так, в одном из вариантов данного изобретения обеспечена новая композиция, содержащая конъюгат антигенсвязывающего района, проявляющего специфичность связывания относительно белка CD 33, и модулятора роста клеток. Модулятором роста клеток может быть токсин, разрушающее клетки лекарственное средство, цитостатическое средство или модулятор биологической ответной реакции. Предпочтительным модулятором роста клеток является гелонин.
В другом варианте данного изобретения обеспечен способ лечения неопластического заболевания, предусматривающий введение эффективной для разрушения клеток дозы иммунотоксина данного изобретения индивидууму, нуждающемуся в таком лечении.
Еще в одном варианте данного изобретения обеспечен способ убивания опухолевых клеток в костном мозге, предусматривающий удаление костного мозга индивидуума, имеющего неопластическое заболевание, обработку костного мозга композицией данного изобретения и инфузию обработанного костного мозга обратно этому индивидууму.
В другом варианте данного изобретения обеспечен способ предотвращения рецидива неопластического заболевания, характеризующегося экспрессией белка CD 33. Рецидив предотвращают введением эффективного для разрушения клеток количества иммунотоксинов данного изобретения.
Еще в одном варианте изобретения обеспечена новая композиция, содержащая слитый белок, образованный слиянием антиген CD 33-связывающего района и модулятора роста клеток. Предпочтительным модулятором роста клеток является гелонин.
В последующих вариантах данного изобретения обеспечены способы продления времени жизни млекопитающего, имеющего опухоль, путем введения иммунотоксина данного изобретения этому млекопитающему. В дальнейших вариантах обеспечен способ замедления скорости роста опухолей введением иммунотоксина данного изобретения.
Фиг. 1 изображает ингибирование синтеза белка гелонином и иммунотоксином M195 на клетках HL 60.
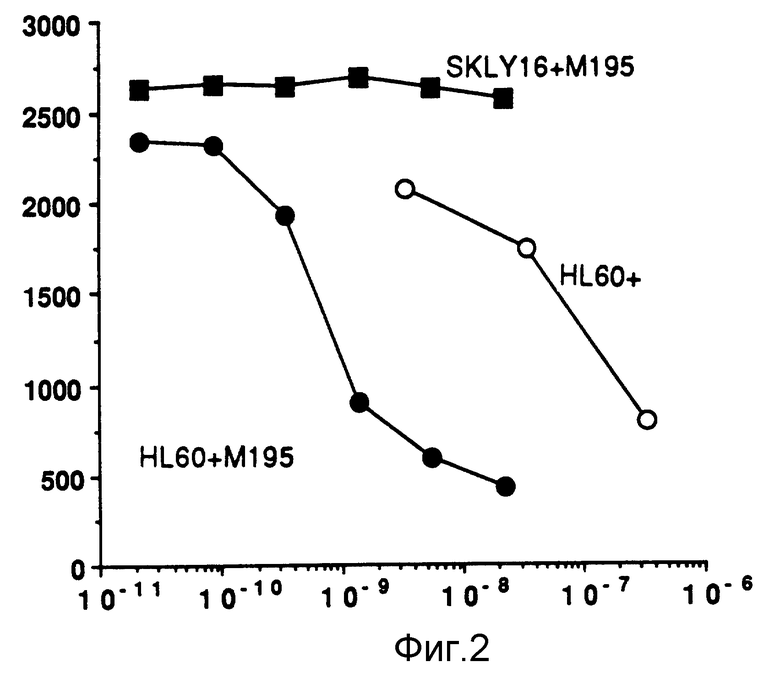
Фиг. 2 демонстрирует ингибирование синтеза белка гелонином и МI95-гелонином на клеточных линиях SKL YI6 и HL 60.
Фиг. 3 иллюстрирует ингибирование синтеза белка путем трехдневного инкубирования иммунотоксина M195 с HL 60.
Фиг. 4 изображает ингибирование синтеза белка путем пятидневного инкубирования иммунотоксина /IT/ МI95 с клетками HL 60.
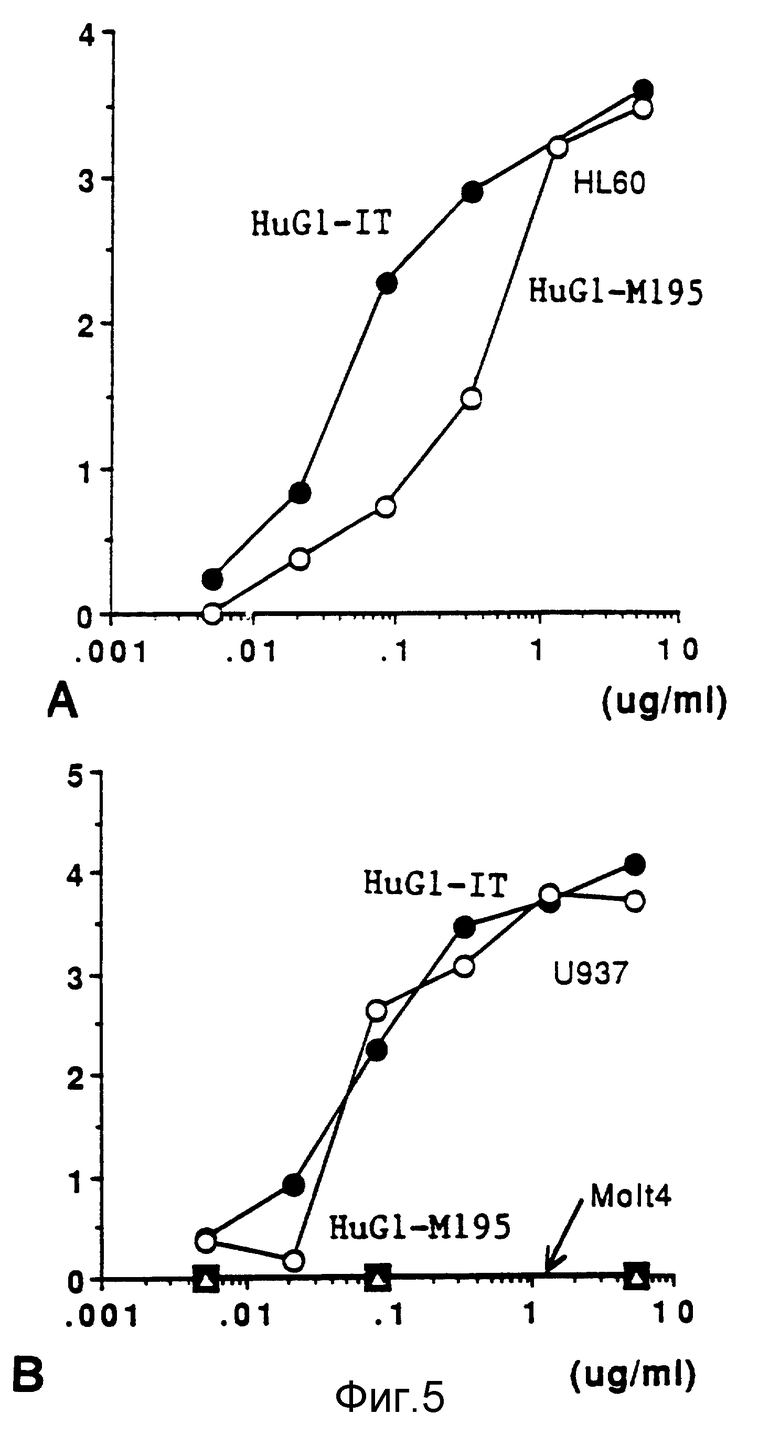
Фиг. 5 демонстрирует связывание Hug1 и Hug1-IT с клетками HL 60, U 937, MOL Т4.
Фиг. 6 иллюстрирует ингибирование синтеза ДНК, вызываемого Hug1-IT в клетках HL 60.
Фиг. 7 изображает конкуренцию между Hug1-IT и Hug1 или FD 79.
Фиг. 8 иллюстрирует электрофоретическое распределение одного M195, реакционной смеси, одного гелонина или очищенного комплекса M195-гелонин.
В описании терминами "химерные антитела" или "химерные пептиды" называются такие антитела или пептиды антител, в которых одна часть пептида имеет аминокислотную последовательность, происходящую из соответствующей последовательности или гомологичную соответствующей последовательности в антителе или пептиде, происходящих из первого источника генов, тогда как оставшийся сегмент этой цепи /цепей/ гомологичен соответствующим последовательностям другого источника генов. Например, химерный пептид тяжелой цепи антител может содержать мышиную вариабельную область и человеческую константную область. Два источника генов в типичном случае являются двумя разными видами, но иногда могут быть представителями одного вида.
Химерные антитела или пептиды обычно получают при помощи рекомбинатных молекулярных и/или/ клеточных способов. Во многих случаях химерные антитела имеют вариабельные области как легких, так и тяжелых цепей, которые имитируют вариабельные области антител, происходящих из одного вида млекопитающих, тогда как константные и/или/ каркасные части гомогологичны последовательностям в антителах, происходящих из второго, отличающегося вида млекопитающих.
Однако применяемое здесь определение химерного антитела не ограничивается этим примером. Химерное антитело представляет собой любое антитело, в котором одна или обе из тяжелых и легких цепей составлены из сочетаний последовательностей, имитирующих последовательности в антителах из различных источников, независимо от того, относятся ли эти источники к различным классам, отличающимся ответной реакцией на антиген, или к различным видам, а также от того, находятся ли точка слияния в месте связи между вариабельной и константной областями или вне этого места. Например, химерные антитела могут быть антителами, в которых каркасные области и определяющие комплементарность области /CDR/ происходят из разных источников. К примеру, нечеловеческие CDR могут быть интегрированы в человеческие каркасные области, соединенные с человеческими константной областью с образованием "гуманизованных /"humanized"/ антител. См. , например, РСТ Application Publication N WO 87/02671; US Patent N 4816567; ЕР Раtent Application 0173494; Jones, et al., Nature, 321: 522-525 /1986/; и Verhoeyen, et al., Science 239:1534-1536 /1988/, на все эти патенты здесь даются ссылки.
Применяемый здесь термин "подобная человеческой каркасная область" обозначает каркасную область каждой цепи антител, которая обычно содержит по меньшей мере 70 аминокислотных остатков, в типичном случае 75 - 85 или более остатков. Аминокислотные остатки подобной человеческой каркасной области по меньшей мере на 80%, предпочтительно на 80 - 85% и наиболее предпочтительно более, чем на 85% гомологичны аминокислотным остаткам человеческого иммуноглобулина. Эта общая черта с другими эндогенными антителами применима для создания направляющей части молекулы, которая вызывает лишь небольшую иммунную реакцию, например, механизм, сводящий к минимуму ответную реакцию на аутоантигенные маркеры.
Применяемым здесь термином "гуманизованный" или "подобный человеческому иммуноглобулин" называют иммуноглобулин, содержащий подобную человеческой каркасную область и константную область, гомологичную в основном константной области человеческого иммуноглобулина, например, имеющую не менее 80% или более, предпочтительно приблизительно 85 - 90% или более и наиболее предпочтительно 95% или более гомологии. Таким образом, большинство частей иммуноглобулина, подобного человеческому иммуноглобулину, за исключением, возможно, CDR, в основном гомологичны соответствующим частям одной или нескольких нативных последовательностей человеческого иммуноглобулина.
Применяемым здесь термином "гибридное антитело" называют антитело, в котором каждая цепь отдельно гомологична цепи антитела млекопитающего, но их сочетание представляет собой новый ансамбль, так что это антитело может узнавать два различных антигена. В гибридных антителах одна пара тяжелой и легкой цепей гомологична паре, обнаруженной в антителе, выработанном против одного признака узнавания антигена, например, эпитопа, тогда как другая пара тяжелой и легкой цепей гомологична паре, обнаруженной в антителе, выработанном против другого эпитопа. Это приводит к свойству мультифункциональной валентности, т.е. способности связывать по меньшей мере два различных эпитопа одновременно. Такие гибриды можно, конечно, получать также при помощи химерных цепей.
Термин "моноклональные антитела" обозначает здесь композицию антител, узнающую отдельную антигенную детерминанту. Термин применяют здесь без ограничения в отношении источника антитела или способа, которым оно получено.
Для данного изобретения антитело или другой пептид является специфическим в отношении CD 33, если это антитело или пептид связывают или способны связывать CD 33, что определяют, например, при помощи стандартных способов определения комплексов антитело-антиген или лиганд-рецептор, например, конкурентными тестами, тестами насыщения или стандартными способами иммуноанализа, такими как ELISA /твердофазный иммуноферментный анализ/, или RIA /радиоиммуноанализ/. Это определение специфичности применимо к отдельным тяжелым и/или/ легким цепям, CDR, слитым белкам или фрагментам тяжелых и/или/ легких цепей, которые также специфичны в отношении CD 33, если они связывают только CD 33 или, если после соответствующего включения в молекулу иммуноглобулина с комплементарными вариабельными и константными областями они способны специфически связывать CD 33.
В конкурентных анализах способность антитела или пептидного фрагмента связывать антиген может быть определена детектированием способности этого пептида конкурировать со связыванием соединения, способность которого связывать этот антиген известна. Многочисленные типы конкурентных анализов известны и обсуждаются здесь. Альтернативно, можно также применять анализы, в которых измеряют связывание тест-соединения в отсутствие ингибитора. Например, способность молекулы или другого соединения связывать белок c-erbB-2 можно обнаружить либо путем мечения целевой молекулы, либо путем непосредственного детектирования немеченной целевой молекулы при помощи различных "сэндвич"-тестов. Известны многочисленные типы тестов связывания, таких как тесты конкурентного связывания (см., например, US Patent NN 3376110, 4016043 и Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Publications, N. Y. /1988/), которые даются здесь в виде ссылок). Имеются также способы для измерения связывания тест-соединения только с одним компонентом без использования конкурентного анализа. Например, для идентификации присутствия CD 33 можно применять иммуноглобулины. Можно использовать стандартные способы определения моноклональных антител, такие как ELISA /см. Harlow and Lane, выше/. Для просмотра различных продуцирующих сигнал систем, которые можно использовать, см. US Patent 4391904, на который здесь дается ссылка.
Далее, специфичность связывающих частей молекул в отношении CD 33 можно определить по их афинности. Такая специфичность существует, если константа диссоциации /KD = 1/K, где K - константа афинности/ этой части молекулы менее 1 мкМ, предпочтительно менее 100 нМ и наиболее предпочтительно менее 1 нМ. Молекулы антител в типичном случае имеют К в более низких диапазонах. KD = [R - L] [R] [L], где [R], [L] и [R - L] - концентрации при равновесии рецептора или CD 33 /R/, лиганда, антитела или пептида /L/ и комплекса рецептора с лигандом /R - L/, соответственно. Обычно между лигандом или пептидом и рецептором или антигеном могут быть обратимые нековалентные связи, такие как электростатическое притяжение, ван-дервальсовы силы и водородные связи.
Другие тест-способы могут предусматривать детектирование присутствия или отсутствия различных физиологических или химических изменений, вызываемых взаимодействием, таких как модулирование, интернализация или увеличение фосфорилирования, как описано в US Patent Application 07/644 361 filed 1/18/91, на который здесь дается ссылка. См. также Receptor-Effector Coupling - A Practical Approach, ed. Hulme, IRL Press, Oxford /1990/.
Гелонин представляет собой гликопротеин /мол. масса приблизительно 29 - 30000 Кд/, очищенный из семян Gelonium multiforum. Гелонин относится к классу сильных инактивирующих рибосомы токсинов растений. Другими членами этого класса инактивирующих рибосомы токсинов растений являются цепи абрина, рицина и модекцина. Гелонин, подобно арбину и рицину, ингибирует белковый синтез путем повреждения субъединицы 60 S рибосом млекопитающих. По-видимому, гелонин стабилен к химической и физической обработке. Кроме того, сам гелонин не связывается с клетками и, следовательно, нетоксичен /за исключением его высоких концентраций/ и безопасен при манипулировании в лаборатории. Инактивация им рибосом является необратимой, по-видимому, не требует кофакторов и происходит с такой эффективностью, которая заставляет предполагать, что гелонин действует ферментативно. Гелонин и рицин относятся к наиболее активным токсинам, которые ингибируют белковый синтез в расчете на массу белка. Гелонин в 10 - 1000 раз более активен в ингибировании белкового синтеза, чем А-цепь рицина. Пептиды, подобные рицину и абрину, состоят из двух цепей, из которых А-цепь представляет собой токсическую единицу, а В-цепь действует путем связывания с клетками. В отличие от рицина и абрина гелонин состоит из одной цени и вследствие отсутствия у него В-цепи, необходимой для связывания с клетками, он сам относительно нетоксичен для интактных клеток.
Клетки млекопитающих, по-видимому, не способны связывать и /или/ интернализовать нативную молекулу гелонина. Конъюгаты гелонина с моноклональными антителами, такими как M195, направляемыми на опухолеспецифический антиген, присутствующий на некоторых опухолевых клетках, обеспечивают как специфический способ связывания гелонина с клеткой, так и путь интернализации комплекса гелонина с антителом.
Цитотоксической частью иммунотоксина может быть цитотоксическое лекарственное средство или ферментативно активный токсин бактериального или растительного происхождения или ферментативно активный фрагмент /"А-цепь"/ такого токсина. Ферментативно активные токсины и их фрагменты являются предпочтительными и иллюстрируются на примере гелонина, А-цепи дифтерийного токсина, несвязывающихся активных фрагментов дифтерийного токсина, А-цепи экзотоксина /из Pseudomanas aeruginosa/, А-цепи, А-цепи абрина, А-цепи модекцина, α-сарцина, белков Aleurites fordii, белков диантина, белков Phytoiacca americana /PAPI, PAPII, и PAP-S/, ингибитора Momordica charantia, курцина, кротина ингибитора, Saponaria officinalis, митогеллина, рестриктоцина, феномицина и эномицина. Наиболее предпочтительна конъюгация с гелонином.
К активным фрагментам и производным относятся любые соединения, которые имеют ту же самую структуру кора /центральной части/, что и гелонин со структурой полной длины, но не имеют всей первичной последовательности. Эти фрагменты или производные обладают такой же или улучшенной биологической или цитотоксической активностью, что и гелонин. Цитотоксичность фрагментов или производных гелонина могут определять квалифицированные в этой области специалисты при помощи рутинного теста с применением лизатов ретикулоцитов кролика.
Модификаторами биологической ответной реакции, которые могут быть соединены с антителами M195 и применены в данном изобретении, являются лимфокины и цитокины, такие как IL-1, IL-2, интерфероны / α, β или γ / TNF, LT, TGF- β и IL-6. Эти модификаторы биологического ответа оказывают различные воздействия на опухолевые клетки. Среди этих воздействий - усиленное убивание опухолевых клеток путем прямого действия, а также усиленное убивание опухолевых клеток путем усиления процессов, обусловленных защитными силами организма. Конъюгирование антител M195 с модификаторами биологического ответа делают возможной их селективную локализацию внутри опухолей и, следовательно, улучшенное антипролиферативное действие с одновременным подавлением неспецифических эффектов, ведущих к токсичности для нормальных клеток.
Цитотоксическими лекарственными средствами /и их производными/, которые применимы в данном изобретении, являются адриамицин, комплекс цис-платины, блеомицин и метотрексат /перечень не является ограничивающим/. Они применимы для клинического лечения рецидивирующих опухолей, однако их применение осложняется вредными побочными эффектами и повреждениями, вызываемыми ими в нормальных клетках. Антитела M195 могут служить в качестве подходящего носителя таких лекарственных средств, обеспечивая эффективное средство как доставки к опухоли, так и усиленного вхождения в сами опухолевые клетки. Кроме того, доставка цитотоксических лекарственных средств к опухолям при помощи специфических антител обеспечивает защиту чувствительных мест, таких как печень, которая не экспрессирует CD 33, и стволовые клетки костного мозга, от вредного действия химиотерапевтических агентов. Применение лекарственных средств, конъюгированных с антителами M195, в качестве системы доставки делает возможной более низкую дозировку самого лекарственного средства, поскольку все молекулы лекарственного средства конъюгированы с антителами, которые концентрируются внутри опухоли или лейкоза.
Конъюгаты моноклональных антител могут быть приготовлены с использованием различных бифункциональных связывающих белки веществ. Примерами таких реагентов являются SPDP, иминотиолан (IT), бифункциональные производные имидоэфиров, например, диметиладипимидат, HCl, активные сложные эфиры, например, дисукцинимидилсуберат, альдегиды, например, глутаровый альдегид, бис-азидосоединения, например, бис(р-азидобензоил)гександиамин, производные бис-диазония, например, бис(р-диазонийбензоил)-этилендиамин, диизоцианаты, например, толуол-2,6-диизоцианат, и бис-активные соединения фтора, например, 1,5-дифтор-2,4-динитробензол.
Введение иммунотоксинов данного изобретения индивидууму с диагнозом лейкоза, характеризующегося экспрессией белка CD 33, делает возможным направление цитотоксического агента к клеткам-мишени и концентрирование его в месте, где он необходим для убивания опухолевых клеток. В результате такого направления цитотоксических агентов к клеткам-мишени неспецифическая токсичность по отношению к другим органам, тканям и клеткам будет исключена или уменьшена.
При применении in vivo для терапии иммунотоксины вводят больному человеку или животному в терапевтически эффективных количествах, т.е. количествах, устраняющих или снижающих предрасположение к опухоли, или в количествах, устраняющих остаточное заболевание после лечения с применением химиотерапии или лучевой терапии. Обычно их вводят парентерально, предпочтительно внутривенно. Доза и схема приема зависит от природы лейкоза и его популяции, свойств применяемого иммунотоксина, например, его терапевтического индекса, от больного и истории болезни данного больного. Обычно количество вводимого иммунотоксина находится в диапазоне приблизительно 0,01 - 1,0 мг/кг веса больного.
Для парентерального введения иммунотоксины готовят в виде стандартной дозы, пригодной для инъекций /раствора, суспензии, эмульсии/ в сочетании с фармацевтически приемлемым парентеральным носителем. Такие носители должны быть нетоксичными и нетерапевтическими. Примерами таких носителей являются вода, солевой раствор, раствор Рингера, раствор декстрозы и 5%-ный человеческий сывороточный альбумин. Можно применять также неводные носители, такие как фиксированные масла и этиолеат. В качестве носителей можно применять липосомы. Наполнитель может содержать минорные количества- добавок, таких как вещества, увеличивающие изотоничность и химическую стабильность, например буферы и консерванты. Иммунотоксин обычно готовят в таких наполнителях при концентрациях приблизительно 0,1 мг/мл - 10 мг/мл.
Иммунотоксины данного изобретения можно применять также в способе убивания опухолевых клеток в костном мозгу. В этом способе костный мозг сначала удаляют из индивидуума с таким неопластическим заболеванием, как лейкоз. Затем костный мозг обрабатывают эффективной для разрушения клеток дозой иммунотоксина данного изобретения.
Представленные далее примеры дают детальное описание получения, характеристики и применения иммунотоксинов данного изобретения. Эти примеры не ограничивают рамки данного изобретения.
Пример 1. Очистка гелонина.
С семян Gelonium multiflorum снимали скорлупу и измельчали их в гомогенизаторе с 8 объемами 0,14 М NaCl, содержащим 5 мМ фосфат натрия /pH 7,4/. Гомогенат оставляли на ночь при 4oC при непрерывном перемешивании, охлаждали на льду и центрифугировали при 35000 g в течение 20 минут при 0oC. Супернатант удаляли, диализовали против 5 мМ фосфата натрия /pH 6,5/ и концентрировали при помощи фильтра pm 10. Пробу наносили на ионообменную колонку СМ-52 /20 • 1,5 см/, уравновешенную 5 мМ фосфатом натрия /pH 6,5/. Материал, связавшийся с ионообменной смолой, элюировали 400 мл линейного градиента 0 - 0,3 М при скорости 25 мл/час при 4oC. Собирали фракции по 5 мл. Фракции наблюдали при 280 нм в спектрофотометре. Гелонин элюировался приблизительно во фракциях 55 - 70 и был последним большим пиком элюиции. Фракции 55 - 70 сливали вместе, диализовали против бидистиллированной воды и концентрировали лиофилизацией. Чистоту и мол. массу каждого препарата проверяли при помощи жидкостной хроматографии высокого давления с использованием гель-фильтрационной колонки ТК 3000 с 50 мМ фосфатом натрия с pH 7,4 в качестве буфера и при помощи электрофореза в 15%-ном ПААГ с ДДС Na. Гелонин перемещался в виде одной полосы с приблизительно мол. массой 29 - 30000 дальтон.
Пример 2. Определение активности гелонина.
Активность гелонина контролировали в тесте ингибирования бесклеточного белкового синтеза. Тест проводили путем последовательного добавления к 50 мкл лизата ретикулоцитов кролика с перемешиванием после каждого добавления следующих компонентов: 0,5 мл 0,2 М Трис-HCl /pH 7.8/, 8,9 мл этиленгликоля и 0,25 мл 1 М HCl.
К 20 мкл смеси соли-аминокислоты-энергетические соединения /SAEM/, состоящей из 0,375 М KCl, 10 мМ Mg (CH3CO2)2, 15 мМ глюкозы, 0,25 - 10 мМ аминокислот /кроме лейцина/, 5 мМ АТФ, 1 мМ ГТФ, 50 мМ Трис-HCl /pH 7,6/. 10 мкл креатининфосфатакреатининфосфокиназы, 8 мкл 14С-лейцина /Amersham, 348 мКи/моль/ добавляли 1,5 мкл растворов, содержащих вариирующие концентрации гелониновой смеси. Смесь инкубировали 60 минут при 30oC. Включение 14С-лейцина определяли в аликвоте смеси путем осаждения синтезированного белка на фильтрах из стеклянного волокна, промывания в 10%-ной ТХУ и ацетоне и определения радиоактивности в Бета-счетчике с применением сцинтилляционной жидкости Aquasol. Гелонин с удельной активностью не менее 4 • 109 Е/мг применяли для конъюгирования с антителами. Единицей активности гелонина является количество белка гелонина, вызывающее 50%-ное ингибирование включения 14С-лейцина в белок в бесклеточном тесте.
Пример 3. Модификация гелонина иминотиоланом.
Получение 2-1Т-модифицированного гелонина.
Гелонин в забуференном фосфатом солевом растворе концентрировали приблизительно до 10 мг/мл в концентраторе Centriprep 10. Гидрохлорид триэтаноламина /TEA/HCl/, pH 8,0, и ЭДТА добавляли до конечной концентрации 60 мМ TEA/HCl и 1 мМ ЭДТА, pH 8,0. Исходный раствор 2-иминотиолана /500 мМ в 60 мМ буфере TEA/HCl, содержащем 1 мМ ЭДТА, pH 8,0/ добавляли до конечной концентрации 1 мМ и пробу инкубировали в течение 90 минут при 4oC под током азота при перемешивании. Избыток иминотиолана удаляли гель-фильтрацией на колонке Sephadex G-25 /1 x 24 см/, предварительно уравновешенной буфером фосфат-ЭДТА, pH 7,5, содержащим 0,01 М Na2HPO4, 0,0018 М KHPO4, 0,0034 М KCl , 0,001 М ЭДТА и 0,17 М NaCl. Фракции анализировали на содержание белка в микротитровальных планшетах при помощи Bio-Rad-теста. Гелонин элюировали в свободном объеме /приблизительно фракции 21-23/. Эти фракции соединяли и хранили при 4oC.
M195, соединенный с 4-сукцинимидилоксикарбонил-α-метил-α(2-пиридилтио)толуолом /SMPT/ получали соединением модифицированного 2-1Т гелонина с модифицированными SMPT MAB M195. Вкратце, для модификации M195 при помощи SMPT 10 мг антител в 1,0 мл PBS разбавляли 1:1 2 х боратным буфером /0,05 М борат натрия, 1,7% хлорид натрия, pH 9,0/ и к раствору антител медленно добавляли 52 мкл 4 мМ SMPT в сухом DMF. Реакцию проводили при комнатной температуре в течение 2 часов при перемешивании под N2. Избыток SMPT удаляли пропусканием реакционной смеси через колонку Sephadex G-25, содержащую буфер фосфат-ЭДТА, pH 7,5, содержащие антитела фракции оценивали при помощи Bio-Rad анализа. Фракции соединяли и хранили при 4oC под N2. Перекрестное сшивание с 2-1Т проводили при 27oC под N2 при перемешивании в течение 96 часов. Конечный продукт очищали, как описано ранее для SPDP.
Пример 4. Получение мышиных моноклональных антител M195.
Мышиные моноклональные антитела M195 получали из гибридов, полученных в результате слияния мышиных миеломных клеток NS-1 и клеток селезенки 5-недельных BAL B/c мышей, иммунизированных лейкозными клетками из больного с острым нелимфатическим лейкозом /FAB-M2/. Супернатанты из клонированных гибридомных культур подвергали скринингу против панели лейкозных клеточных линий и исходных ANLL лейкозных клеток при помощи розеткообразования эритроцитов с использованием белка A /PA/ Staphylococcus aureus. Повторно субклонированную гибридому M195 размножали в дважды пристан-примированных (C57 BL/6 раз BAL B/c) F1 мышах.
M195 очищали на PA-Sepharose афинной хроматографией с использованием последовательных РН ступенчатых разведений. Чистоту определяли на гелях ПААГ с ДДС Na, окрашенных Кумасси /brilliant blue/. "Гуманизованные" /"очеловеченные"/ моноклональные антитела M195 готовили, как описано Co et al. , "Chimeric and Humanized Antibodies with Specificity for the CD 33 Antigen", J. Immunol., 148:1149-1154 /1992/, и получали из гибридом и очищали как описано выше.
Вкратце, гипервариабельный домен N-район для H /тяжелой/ и L /легкой/ цепей мышиных антител M195 клонировали при помощи якорной PCR /полимеризной цепной реакции/. Сначала клетки гибридомы M195 лизировали и экстрагировали из них РНК при помощи горячего фенола. Из РНК синтезировали кДНК путем инкубирования РНК с вирусной обратной транскриптазой. Затем конструировали праймеры РСР и в праймеры, расположенные по ходу транскрипции и против хода транскрипции, включали сайты Eco RI и Hind III для удобства субклонирования. PCR-реакции проводили в программируемом нагревающем блоке с применением 30 циклов температурой циклизации /92oC в течение 1 минуты, 50oC в течение 2 минут и 72oC в течение 3 минут/. Продукты полимеразной цепной реакции разделяли при помощи электрофореза на агарозном геле с низкой точкой плавления. Полосы вырезали, переваривали рестриктазами и клонировали в вектор PUC 18 для секвенирования. Затем мышиные H- и L-цепи расщепляли в плазмиды, содержащие комплементарные H- и L-цепи человеческих антител, при помощи различных векторов. Затем векторы, кодирующие химерные "мышь-человек" L- и H-цепи, трансфицировали в клетки SP 2/0 при помощи электропорации. Выжившие колонии высевали на чашки и тестировали на способность продуцировать (очеловеченные) "гуманизованные" антитела α CD 33.
Пример 5. Конъюгирование модифицированного SPDP моноклонального антитела M195 с модифицированным иминотиоланом гелонином.
1 мг очищенного гелонина /2 мг/мл в PBS/, полученного, как описано в примере 1, модифицировали иминотиоланом, как описано в примере 3. Моноклональные антитела M195, модифицированные, как описано в примере 4, смешивали с равным по весу количеством модифицированного гелонина. Это соотношение соответствовало 5-кратному избытку молярному гелонина по отношению к антителу. pH смеси доводили до 7,0 путем добавления 0,05 М буфера TEA/HCl с pH 8,0 и смесь инкубировали в течение 20 часов при 4oC под азотом. Для блокирования оставшихся свободных сульфгидрильных групп добавляли иодацетамид /0,1 М/ до конечной концентрации 2 мМ и инкубирование продолжали еще в течение 1 часа при приблизительно 25oC Реакционную смесь хранили при 4oC до очистки при помощи гель-фильтрации.
Пример 6. Очистка комплексов гелонин-моноклональное антитело M195.
Неконъюгированный гелонин и низкомолекулярные продукты удаляли из реакционных смесей примера 6 гель-фильтрацией на колонке Sephadex S-300 /1,6 x 31 см/, предварительно уравновешенной PBS.
Реакционные смеси из примера 5 концентрировали приблизительно до 1 мл при помощи микроконцентратора Centricon 30 перед нанесением на колонку Sephadex. Колонку промывали PBS. Собирали фракции по 1 мл и аликвоты 50 мкл анализировали на белок по методу Бредфорда.
Для удаления неконъюгированного M195 высокомолекулярный пик /фракции 28 - 40/ из колонки S-300 наносили на колонку для афинной хроматографии Blue Sepharose CL-6B /1 x 24см/, предварительно уравновешенную 10 мМ фосфатным буфером /pH 7,2/, содержащим 0,1 М NaCl. После нанесения пробы колонку промывали 30 мл буфера для полной элюции неконъюгированных антител. Колонку элюировали линейным солевым градиентом 0,1 - 2 М NaCl в 10 мМ фосфатном буфере с pH 7,2. Содержание белка в элюированных фракциях определяли по методу Бредфорда.
Неконъюгированные антитела удаляли из конъюгированных с гелонином антител при помощи афинной хроматографии на колонке /1 x 24 см/ Blue Sepharose CL-6B, предварительно уравновешенной 10 мМ фосфатным буфером pH 7,2, содержащим 0,1 М NaCl. После нанесения пробы S-300 элюата колонку промывали 30 мл того же буфера для полной элюции неконъюгированных антител.
Конъюгированные с гелонином антитела, связанные с колонкой, элюировали линейным солевым градиентом 0,2 - 2,0 М NaCl в 10 мМ фосфатном буфере с pH 7,2. Комплекс антитело-гелонин элюировался приблизительно при 0,7 М NaCl. Содержание белка в элюированных фракциях определяли по методу Бредфорда. Содержащие белок фракции объединяли и анализировали при помощи электрофореза на 5-20%-ном градиентном невосстанавливающем ПААГ. Фиг. 8 иллюстрирует электрофоретическое распределение одного M195, неочищенной реакционной смеси M195, генолина и комплекса M195-гелонин, одного гелонина или очищенного комплекса M195-гелонин. Проходящий через колонку пик /фракции 14-20/ содержит только свободные антитела, тогда как фракции 50 - 80, элюированные высокой концентрацией соли, содержат конъюгат M195-гелонин, не содержащий неконъюгированного гелонина или антител. Конечный продукт содержал антитело M195, сопряженное с 1, 2 или 3 молекулами гелонина. Среднее содержание гелонина было 1,5 молекул на 1 молекулу антитела. Кроличьи ретикулоциты в системе трансляции in vitro применяли для оценки активности гелонина в практически чистом комплексе гелонин-антитело M195. Одной единицей активности в этом тесте считали количество белка, требуемое для 50%-ного ингибирования белкового синтеза по сравнению с необработанными контролями. При помощи этого теста определили, что удельная активность нативного гелонина и конъюгата M195-гелонин была равно 2 • 108 Е/мг и 8,2 • 105 Е/мг, соответственно. Очищенный комплекс гелонин-антитело M195 активен в тесте с лизатом ретикулоцитов. Разведение 1: 1000 исходной пробы вызывало приблизительно 50%-ное снижение включения 14С-лейцина в белок. Таким образом, активность исходного препарата была 1000 Е/мл.
Пример 7.
Композиции данного изобретения могут включать слитые конструкции моноклонального антитела M195 и цитотоксической части молекулы. Такие слитые конструкции иммунотоксина данного изобретения можно приготовить по способу Co, et al., на который дается ссылка.
Перед использованием в этих исследованиях клетки Sp2/0-AgI4 выращивают исходно в присутствии 0,1 мкг/мл нативного гелонина. После несколько месяцев концентрацию гелонина постепенно увеличивают, пока клетки не будут поддерживаться при концентрации до 10 мг/мл. Затем клетки клонируют путем лимитирующего разведения в присутствии 10 мг/мл гелонина и образующиеся колонии, устойчивые к гелонину, размножают. Затем гелонин удаляют из культуралъной среды в течение двух пассажей и клетки вновь подвергают действию гелонина для подтверждения образования стабильно резистентных клонов. После тестов, подтверждающих образование и активность "гуманизованных" антител M195 выращивают резистентные к гелонину клетки SP2/0, продуцирующие антитела, и кДНК для антител M195 удаляют из тотальной ДНК инкубированием с рестриктазой. Параллельно выделяют кДНК из E.coli JMI05, экспрессирующей оптимизированный гелонин, очищают и высвобождают ДНК, кодирующую гелонин, после переваривания рестриктазами Hind III и Eco RI. Ген гелонина лигируют во фрагмент тяжелой цепи и инсерцию вводят в устойчивые к гелонину клетки SP2/0. Затем клетки субклонируют путем лимитирующего разведения и клоны подвергают скринингу как на содержание гелонина, так и на образование "гуманизованных" антител. Наконец, положительные в этом отношении клоны размножают и рекомбинантный слитый белок очищают и тестируют как в тестах на цитотоксичность, так и в испытаниях распределения в ткани in vitro, фармакокинетики, терапевтических тестах и тестах на токсичность. Проводят сравнение свойств слитого белка M195-гелонина с характеристиками описанных ранее конструкций M195-гелонин для определения преимуществ и недостатков каждого из комплексов. На основе этих исследований клиническое исследование фазы I химерного слитого белка M195-гелонина можно проводить на больных с прогрессированным раком молочной железы.
Пример 8.
Как показано на фиг. 1, иммунотоксин M195 тестировали на его способность убивать клетки HL 60 по сравнению со свободным гелонином. Ингибирование белкового синтеза (при помощи теста включения тритированных смешанных аминокислот) 0,5 мкКи/мл; New England Nuclear Corp.) в осаждаемый трихлоруксусной кислотой /ТХУ/ белок) использовали как меру активности примененного агента. Конечные концентрации иммунотоксина M195-гелонина были 4 мкг/мл - 5 нг. Конечные концентрации гелонина простирались от 44 мкг/мл до 0,6 мкг/мл.
Иммунотоксин M195-гелонин был приблизительно в 600 раз более сильным, чем один свободный гелонин. ID50 для иммунотоксина M195-гелонина был приблизительно 0,4 нМ. Ингибирование белкового синтеза иммунотоксином M195-гелонином приводит затем либо к прекращению деления клеток, либо к смерти клеток. Смерть клеток подтверждали экспериментами с применением trupan blue-исключения для определения общего числа живых клеток и процента живых клеток. На (фиг. 1 показано, что клетки HL 60 с конечной концентрацией 1 • 106 клеток на мл инкубировали в течение трех дней при 37oC в присутствии иммунотоксина M195-гелонина или одного гелонина.
На фиг. 2 показано, что клетки HL 60 и SKL YI6 с конечной концентрацией 5 • 105 клеток/мл инкубировали в течение трех дней при 37oC в присутствии либо одного гелонина, либо иммунотоксина M195-гелонина. Конечные концентрации иммунотоксина простирались от 4 мкг/мл до 15,2 nг/мл. Конечные концентрации гелонина простирались от 10 мкг/мл до 0,1 мкг/мл. Уровни белкового синтеза определяли по пятичасовому включению тритированных аминокислот в осаждаемый трихлоруксусной кислотой белок.
Как можно видеть на фиг. 2, концентрации иммунотоксина M195-гелонина ингибировали белковый синтез HL 60 клеток более, чем на 80%; сравнимые концентрации иммунотоксина не влияли на клетки SKL YI6. Таким образом, очевидной является селективность иммунотоксина M195-гелонина.
На фиг. 3 и 4 изображены результаты исследований, в которых клетки HL 60 в конечной концентрации 1 • 106 инкубировали в течение 3 /фиг. 3/ или 5 /фиг. 4/ дней при 37oC в присутствии иммунотоксина M195-гелонина. Конечная концентрация иммунотоксина были в диапазоне 4 мкг/мл - 0,9 нг/мл. Уровни белкового синтеза определяли путем пятичасового включения тритированных аминокислот в осаждаемый трихлоруксусной кислотой белок.
Как видно при сравнении фиг. 3 и 4, продолжительность времени экспозиции с иммунотоксином влияет на его активность. Действительно после 5 дней инкубирования можно наблюдать 10-кратное увеличение активности иммунотоксина по сравнению с трехдневным инкубированием.
На фиг. 5 показаны результаты испытаний связывания "гуманизованного "очеловеченного" M195 (Hug1)-иммунотоксина с клетками HL 60, U 937, MOL T4. Hug1 и Hug1-иммунотоксин добавляли к клеткам HL 60 /фиг. 5, А/ или по клеточным линиям U 937, MOL T4 /фиг. 5, В/ в диапазоне концентраций 5 мкг/мл - 5 нг/мл/. Клетки инкубировали на льду в течение 1 часа перед вымыванием избытка антител. Затем к клеткам добавляли козьи антитела FITC против человека и клетки вновь инкубировали в течение 1 часа на льду. После вымывания избытка антител клетки фиксировали в присутствии 0,5%-ного параформальдегида и снимали показания на протонном цитометре /EPICS Profile flow cytometer/.
Фиг. 5 показывает, что иммунотоксин, содержащий конъюгат "гуманизованных" "очеловеченных" антител M195 с гелонином, способен специфически связываться с клетками-мишенью. С применением непрямой проточной цитометрии этот иммунотоксин обнаружил более специфическое связывание с экспрессирующимии CD 33 клеточными линиями /HL 60/ и с U 937 по сравнению со свободными "гуманизованными" антителами M195. Он не связывался с отрицательными по CD 33 клеточными линиями /MOL T4; фиг. 5, В/.
На фиг. 6 представлены результаты экспериментов, в которых клетки HL 60 в конечной концентрации 3 • 104 и 5 • 104 инкубировали в течение 5 дней при 37oC в присутствии иммунотоксина, содержащего "гуманизованные" очеловеченные" антитела M195 и гелонин, при конечной концентрации в диапазоне 4 мкг/мл - 0,2 нг/мл. Конечные концентрации гелонина были в диапазоне 50 мкг/мл - 0,5 мкг/мл. Синтез ДНК определяли путем пятичасовой инкубации с тритированным тимидином. Как показывает фиг. 6, ингибированный иммунотоксином белковый синтез приблизительно в 4500 раз выше белкового синтеза в присутствии одного гелонина.
Фиг. 7 иллюстрирует способность иммунотоксина, содержащего "гуманизованные" "очеловеченные" антитела M195 и гелонин, убивать клетки HL 60. Как видно из фиг. 7, ID50 для этого иммунотоксина был менее 10 микомолей. ID50 для этого иммунотоксина более чем в 4000 раз ниже, чем ID50 одного гелонина.
Как видно из фиг. 8, HL 60 клетки инкубировали в течение 1 часа на льду в присутствии "гуманизованных" "очеловеченных" антител M195 или Fd 79, изотипически совместимых контрольных антител Hug1. Иммунотоксин, содержащий "гуманизованные" "очеловеченные" антитела M195 и гелонин, добавляли в концентрации 0,15 мкг/мл. Клетки инкубировали 90 часов при 37oC. Количество живых клеток определяли способом исключения с применением trupan blue. Процент ингибирования обозначает снижение числа клеток, убитых в присутствии конкурирующего антитела, по сравнению с клетками, убитыми в присутствии одного иммунотоксина.
Как видно из фиг. 8, "гуманизованные" "очеловеченные" антитела M195 были способны блокировать цитотоксичность иммунотоксина, содержащего эти антитела вместе с гелонином, и блокирование зависело от дозы. В противоположность этому, Fd 79, неспецифические "гуманизованные "очеловеченные" Ig GI, не оказывали влияние при тех же самых уровнях.
Таким образом, очевидно, что данное изобретение и предусмотренные им варианты хорошо применимы для достижения целей и получения результатов, изложенных вначале. В способе и устройствах могут быть сделаны определенные изменения без отхода от характера и сферы действия данного изобретения. Понятно, что возможны изменения, и предполагается, что каждый элемент или каждая ступень, описанные в каждом из следующих далее пунктов формулы изобретения, следует относить ко всем эквивалентным элементам или ступеням для достижения тех же результатов теми же или равноценными способами. Предлагается расширение сферы действия данного изобретения в любой форме, в которой могут быть использованы его принципы. Таким образом, данное изобретение хорошо приспособлено для достижения целей и получения преимуществ, упомянутых выше, а также других присущих изобретению преимуществ.





Данное изобретение обеспечивает новые иммунотоксины и способы лечения неопластических (опухолевых) заболеваний. Эти иммунотоксины образованы конъюгированием связывающего антиген района, обладающего специфичностью связывания относительно белка СD 33, и модулятора роста клеток гелонина и являются фармацевтической композицией. Иммунотоксины данного изобретения специфически и селективно убивают опухолевые клетки, отличающиеся экспрессией антигена СD 33. Поэтому новые иммунотоксины могут быть применимы в способе лечения лейкозов как острых, так и хронических, и других миелодиспластических синдромов. Способ и композиция обеспечивают замедление роста опухолей введением фармацевтической композиции данного изобретения в опухолевую клетку. 2 с и 9 з.п.ф-лы, 8 ил.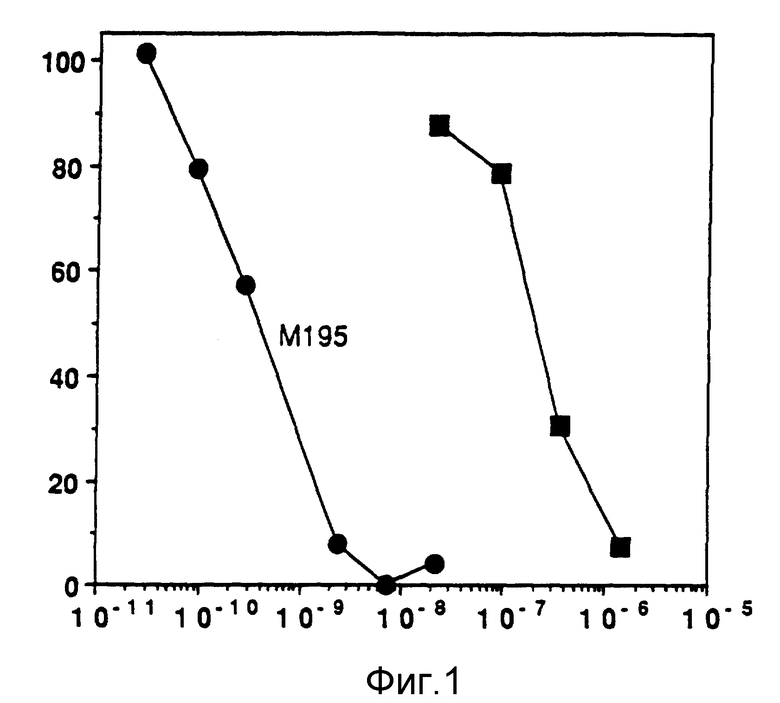
| US 4766106, 23.08.88 | |||
| Алахов В.Ю | |||
| и др | |||
| Новый принцип создания иммунотерапевтических соединений направленного действия .. | |||
| Докл | |||
| АН СССР | |||
| Механическая топочная решетка с наклонными частью подвижными, частью неподвижными колосниковыми элементами | 1917 |
|
SU1988A1 |

