Область науки, к которой относится изобретение.
Настоящее изобретение относится к области иммуноконъюгат и более конкретно к использованию иммуноконъюгат при лечении рака. Изобретение относится также к лечению меланомы конъюгатами моноклональных антител (монАт) и цитотоксичных составляющих, таких как гелонин, рибосомноингибирующий белок, иные цитотоксичные составляющие растительного происхождения или же цитотоксичные или цитостатические модификаторы биологических реакций.
Предпосылки к созданию изобретения.
Необходимость точной направленности терапии рака имеет критическое значение по той причине, что адекватная реакция опухоли зависит от введения и поддержания в опухоли должной лечебной концентрации лекарства. Достижение нацеленной точно на опухоль терапии стало целью нескольких исследователей, использующих моноклональные антитела в качестве специализированных носителей терапевтических средств.
Рак является одной из основных причин смертности и заболеваемости в западном мире. Существует много разновидностей рака, каждая из которых имеет свои особенности. Однако, по меньшей мере одна особенность является общей для всех разновидностей рака. Она связана с дефектами процесса регулирования роста клеток.
Меланома, наиболее опасная разновидность рака кожи, является болезнью с активным развитием метастаз, поражающей оба пола и почти обязательно приводящей к смертельному исходу в течение пяти лет после установления диагноза. Доказано, что хирургическое удаление выявленных злокачественных образований может быть эффективным только тогда, когда появились первичные признаки заболевания. Если болезнь получила развитие, хирургическое вмешательство должно быть дополнено иными, более общими процедурами уничтожения больных или злокачественных клеток. Большинство обычно применяемых дополнительных процедур, таких как облучение или химиотерапия, не ограничиваются опухолевыми клетками, и хотя они и оказывают пропорционально более разрушающее действие на злокачественные клетки, но в определенной степени часто поражают нормальные клетки.
Многие опухоли экспрессируют антигены или антигенные детерминанты, которые в очень слабой степени экспрессируются или не экспрессируются вовсе нормальными клетками. Некоторые опухолевые клетки экспрессируют антигены, которые экспрессируются клетками эмбрионов, но не экспрессируются нормальными клетками взрослого животного. Эти аномально экспрессируемые антигены известны как антигены, ассоциированные с опухолями. Особенностью этих антигенов является то, что в наше время как определенный антиген может быть экспрессирован более чем одной опухолью, этот антиген обычно экспрессируется всеми или большинством клеток определенных опухолей, экспрессирующих его. Клетка опухоли может экспрессировать один или несколько ассоциированных с опухолью антигенов. Эти ассоциированные с опухолью антигены могут быть экспрессированы на поверхности клетки (антиген поверхности клетки), могут быть секретированы опухолевой клеткой (секретированные антигены) или могут оставаться внутри клетки (внутриклеточный антиген).
Присутствие этих ассоциированных с опухолью антигенов используется для обнаружения, диагностики и локализации опухоли. В некоторых случаях присутствие ассоциированных с опухолью антигенов на клетках опухоли позволяет нацеливать определенные лекарственные средства или другие лечебные средства точно на клетки опухоли.
Антитела являются белками, которые иммунная система животного образует в ответ на чуждые антигены или антигенные детерминанты. Антитела связываются с конкретными антигенами, против которых они направлены. Так, например, антитела к другим антигенам меланомы используются для того, чтобы после систематического интраперитонального введения показать расположение определенной опухоли.
Один из методов нацеливания терапевтических средств на опухолевые клетки и недопущения их воздействия на нормальные клетки стал возможен после создания монАт, нацеленных против антигенов на опухолевых клетках, не встречающихся на нормальных клетках. Моноклональные антитела, направленные к определенным антигенам или антигенным детерминантам, могут быть получены в больших количествах.
Антитела могут быть помечены, чтобы их можно было использовать для локализации и лечения злокачественных образований. Такие меченые радиоактивными изотопами моноклональные антитела к антигенам поверхности опухолевых клеток с успехом использовались для представления опухоли у пациента посредством наружной сцинтографии. (Deland, Lemin. Nucl. Med. 19(3) : 158-65 (Обзор) (1989) Tuhl. Heptagastroenterology 36(1) : 27-32 (Обзор), (1989)). Антитела в комплексе с лекарственными средствами могут быть использованы как система доставки, с помощью которой лекарственные средства нацеливаются на опухолевые клетки определенного типа, против которых действует антитело, что связано с уникальной способностью антител локализовать место опухоли после систематического введения. Антитела могут также объединяться с токсинами и действовать таким образом как система доставки, предназначенная для нацеливания токсинов непосредственно на определенные опухолевые клетки.
Цитотоксичные средства, часто применяемые в конъюгатах антител, подразделяются в целом на три основных класса: токсины, радионуклиды и химиотерапевтические средства. Конъюгаты антител с каждым из этих типов средств предлагают многообещающие возможности, но при этом возникают и определенные проблемы (Frankel и др. Aun. Rev. Med. 37 : 125 - 142 (1986), Reimann и др. T. Clin. Invest 82(1) : 129-138 (1988)). Иммуноконъюгаты, включающие растительные токсины, обладают уникальными преимуществами по сравнению с другими типами конъюгатов антител, поскольку:
1. Дозы иммунотоксинов, необходимых для противоопухолевого воздействия в целом, значительно ниже, чем требуется в конъюгатах антител с лекарственными средствами.
2. Сцепление токсинов с антителами, видно, не оказывает отрицательного воздействия на связуемость антител.
Гелонин является гликопротеином (мол. м. приблизительно 29-30000 Кд), очищенным из семян Gelonium multiflorum. Гелонин относится к классу действенных рибосомо-инактивирующих растительных токсинов. Другими членами этого класса рибосомо-инактивирующих растительных токсинов являются цепи абрина, рицина и модецина. Гелонин так же, как абрин и рицин, ингибирует синтез белка, повреждая подгруппу 60 S рибосом млекопитающих. Хотя цепь A рицина (РТА) нашла широкое применение для использования в иммунотоксинах, гелонин оказывается более устойчивым к химическому и физическому воздействию, чем РТА (Basbieri и др. Cancer Surv. 1 : 439-520 (1982)). Кроме того, сам гелонин не связывается с клетками и поэтому нетоксичен (за исключением случаев высокой концентрации) и безопасен при лабораторных исследованиях. Инактивация рибосом необратима, похоже, тем, что не влечет за собой побочных воздействий и происходит с эффективностью, позволяющей предполагать, что гелонин действует ферментативно.
Многие исследователи в прошлом уже сообщили или предлагали соединение цитотоксичных средств с антителами с целью получения "иммунотоксинов". Особый интерес представляет иммунотоксины из моноклональных антител, сцепленных с ферментативно активными частями (цепями A) токсинов бактериального или растительного происхождения, таких как рицин или абрин (Nevetta и др., Immunol. Rev. 62 : 75 - 92), Ross и др., European J. Biochem 104 (1980), Vitteta и др. , Immunol. Rev. 62 : 158 - 183 (1982), Ross и др., Cancer, Rev. 42 (1982) 457-464, Trowriolge и Domingo, Nature (Cond.) 294 : 181 - 173 (1982).
Гелонин и рицин относятся к числу наиболее активных токсинов, ингибирующих синтез белка на основе массы белка. Гелонин в 10 - 1000 раз более активен в ингибировании синтеза белка по сравнению с цепью A рицина. Пептиды типа рицина и абрина состоят из двух цепей, из которых цепь A является токсичной составляющей, а цепь B осуществляет связь с клетками. В отличие от рицина и абрина гелонин состоит из одной цепи и, поскольку у него отсутствует цепь B для связи с клетками, он сам по себе относительно нетоксичен для здоровых клеток (Stirpe и др. J. Biol. Chem. 255 : 6947 - 6953 (1980)). Клеткам млекопитающих явно недостает способности к связи и/или внедрению природной молекулы гелонина. Конъюгаты гелонина с нацеленным на опухоль моноклональным антителом, таким как моноклональное тело ZME, направленное к антигену, присутствующему на некоторых опухолевых клетках, таких как клетки меланомы, обеспечивают как определенный способ связывания гелонина с клеткой, так и путь внедрения комплекса гелонин-антитело. Одним из преимуществ использования токсина гелонина по сравнению с использованием таких токсинов, как цепь A рицина, является снижение токсичности к нормальным клеткам в сравнении с цепью A рицина. Гелонин в комплексе с моноклональным, направленный к ассоциированному с опухолью антигену, является активным иммунотоксичным средством избирательного действия для терапии опухолей.
Предыдущие исследования описывают ряд конъюгат антитело токсин, содержащих гелонин (Lambert и др., J.Biol. Chem. 260 : 12035-12038 (1985) Jhorpe и др. , Eur. J. Biochem. 116 : 447-454 (1981) Lingh и др., J.Biol. Chem. 264 : 3089-95 (1989) Lcott и др., J. Natl. Cancer Lnst. 79 : 1163-72 (1987) Tedoler и др., J. Immunol. 137(4) : 1387-91 (1986)). Недавно Озавари и др. Int. J. Cancer. 43 : 152 - 157 создал гелониновый иммунотоксин, включающий антитело B 467, которое связывается с клеточным рецептором эпидермального фактора роста (ЕФР). Этот конъюгат B 467 и гелонина обладает высокой токсичностью по отношению к клеткам, экспрессирующим рецептор ЕФР, но не токсичен для клеток, не имеющих рецептора. Сивэм и др. (Cancer Research 47 : 3169-3173 (1987)) создал конъюгат антимеланомного антитела 9.2.2.7 с гелонином и сравнивал in vivo и in vitro цитотоксичное действие с конъюгатом 9.2.2.7 с цепью A абрина и рицина. Эти исследования показали, что конъюгаты с гелонином демонстрируют in vitro значительный избирательный цитотоксичный эффект по отношению к клеткам, положительным по антигену. Эксперименты in vivo показали, что гелониновые конъюгаты не токсичны при дозе всех антител, достигающей 2 мг/мышь и что множественное введение I.V. гелонинового иммунотоксина значительно замедляет рост подкожного ксенотрансплантата человеческой опухоли у голой мыши. По сравнению с конъюгатами с арбином и рицином конъюгаты гелонина обладают сходной действенностью, лучшей избирательностью и локализацией опухоли с более значительными терапевтическими свойствами in vivo.
Поскольку антитело, с которым объединяется лекарственное средство, токсин или радиоизотопная метка, связывается только с опухолевыми клетками, экспрессирующими определенный антиген, убиваются только опухолевые клетки. В отличие от этого при лучевой терапии радиация от соединений, меченых радиоактивными изотопами, не ограничивается только опухолевыми клетками, на которые направлено облучение. Так, например, метаболическое или ферментативное разложение антител может привести к высвобождению радиоактивной метки и позволить ей распространиться на другие ткани, такие как почки или костный мозг, ведя к неприемлемому радиационному повреждению этих органов. Антитела с радиоактивными метками сталкиваются с проблемами, которые ограничивают или осложняют их использование в качестве терапевтических средств.
Краткое изложение существа изобретения.
Настоящее изобретение предлагает иммуноконъюгаты с антителом (обозначенным здесь как ZME - 018), которое распознает антиген GP 240 на раковых клетках меланомы. Одно из антител (225. 28S), рассмотренных Вилсоном и др. (Wilson и др. Int. J. Cancer 28 : 293 (1981)) распознает этот антиген мембраны меланомы. Этот антиген обозначается здесь символом GP 240. Антитело 225.28S , которое связывается с антигеном GP 240, обозначено и упоминается далее как ZME-018. В одном варианте осуществления антитело объединяется с токсином, выбранным из группы, состоящей из гелонина, цепи A рицина и цепи A абрина. В другом варианте осуществления антитело ZME может быть объединено с цитокидальным средством, таким как адриамицин, или с модификатором биологической реакции, таким как лимфокин или цитокин. Еще в другом варианте осуществления антитело может быть помечено различной меткой, такой как метка радиоактивным изотопом, хемолюминисцером, флюоресцером или ферментной меткой. Цитокидальные иммуноконъюгаты полезны для лечения и предупреждения рецидива опухолей, несущих ассоциированный с опухолью GP 240 путем введения этих цитокинальных иммуноконъюгатов в соответствии с индивидуальными особенностями в каждом отдельном случае. Различимо помеченные иммуноконъюгаты ZME полезны для диагностики и локализации опухолей средствами, известными специалистам в данной области. Эти меченые иммуноконъюгаты полезны также при анализе на присутствие антигена GP 240 в биологических образцах и при локализации места опухоли in vivo средствами, известными специалистам.
Одной из целей настоящего изобретения является предложение специального состава, который избирательно связывается с опухолевыми клетками и убивает их. В особенности целью настоящего изобретения является предложение цитотоксичного состава, который избирательно будет связываться и убивать опухолевые клетки, экспрессирующие антиген GP 240 так, как описано выше. Антитело ZME-018 было приготовлено "Хайбритекс" с применением солевого фракционирования и хроматографии на ДЭАЭ, а гомогенность которого была определена с помощью SDS PAGE (Wilson и др., Int. J. Cancel. 28 : 293-300 (1981)). Другой аспект настоящего изобретения касается способа уничтожения клеток меланомы или любых других опухолевых клеток, экспрессирующих антиген ZME (GP 240), путем осуществления контакта клеток с цитокидально эффективным количеством иммунотоксина.
Еще одной целью настоящего изобретения является предложение такого состава, который был бы токсичным для опухолевых клеток, но наносил бы минимальный вред здоровым тканям.
Описание чертежей.
На фиг. 1 показана схема соединения и очистки комплекса ZME-гелонин.
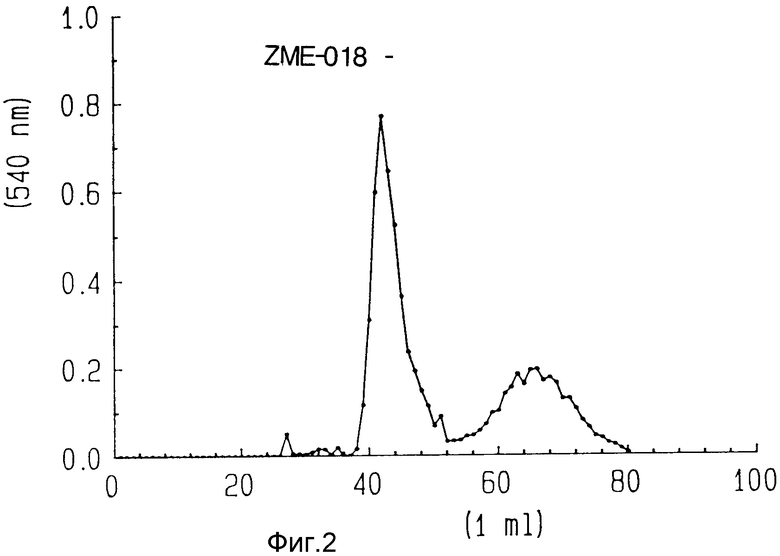
На фиг. 2 показана очистка ZME-гелонина методом эксклюзионной хроматографии на геле S-300.
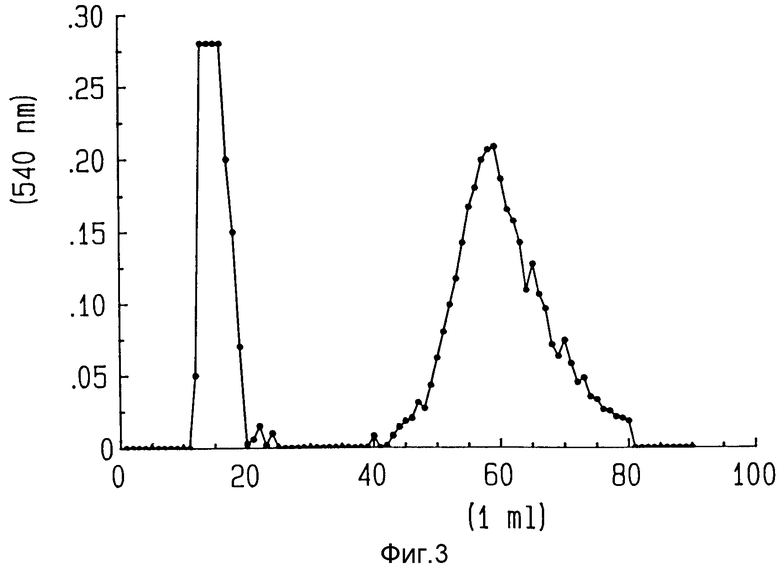
На фиг. 3 показан профиль элюирования в колонке Сибахрон-Блу на сефарозе после поступления материала с высокой молекулярной массой после хроматографии на S-300 и его элюирования с линейным солевым градиентом (0-300 мМ NaCl). Наблюдаются два белковых пика: пик протекания (фракция 14-20) и граничный пик элюции при высоком солевом содержании (фракции 44-75).
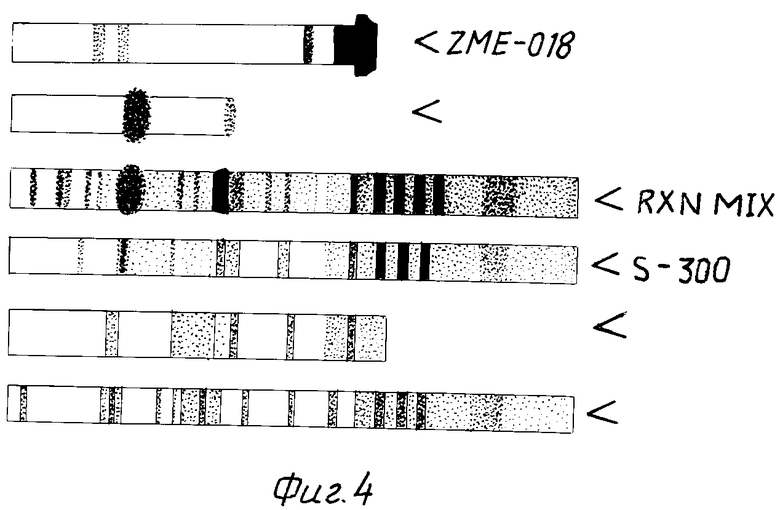
На фиг. 4 показана электрофорезная картина гелонина и конъюгата ZME-гелонин.
На фиг. 5 показаны сравнительные данные по анализу ELISA по ZME (белые кружки) и ZME-гелонина (черные кружки).
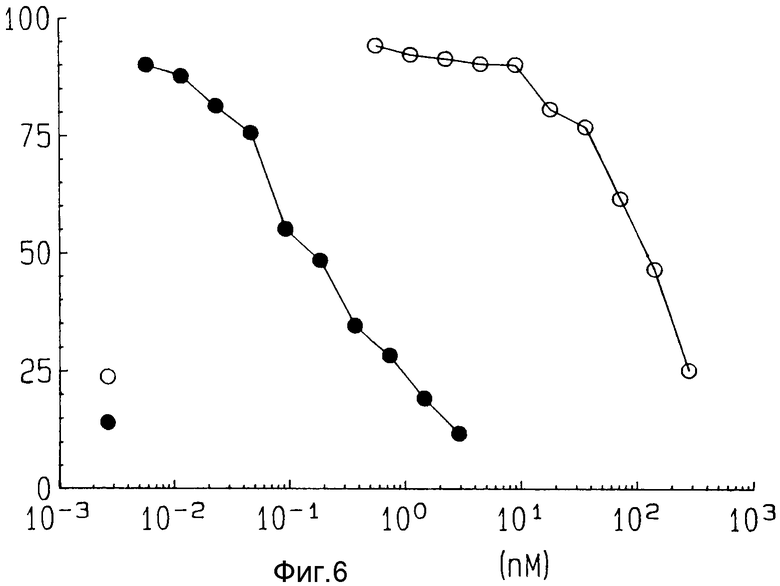
На фиг. 6 показана цитотоксичность ZME-гелонина и свободного гелонина на лог-фазе клеток AAB-527 после 72 часов экспозиции.
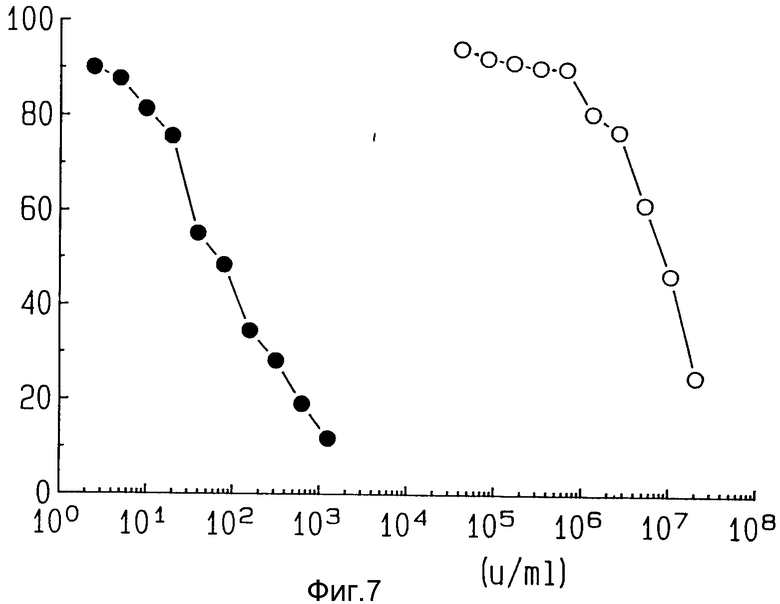
На фиг. 7 показана цитотоксичность ZME-гелонина и свободного гелонина на лог-фазе клеток AAB-527.
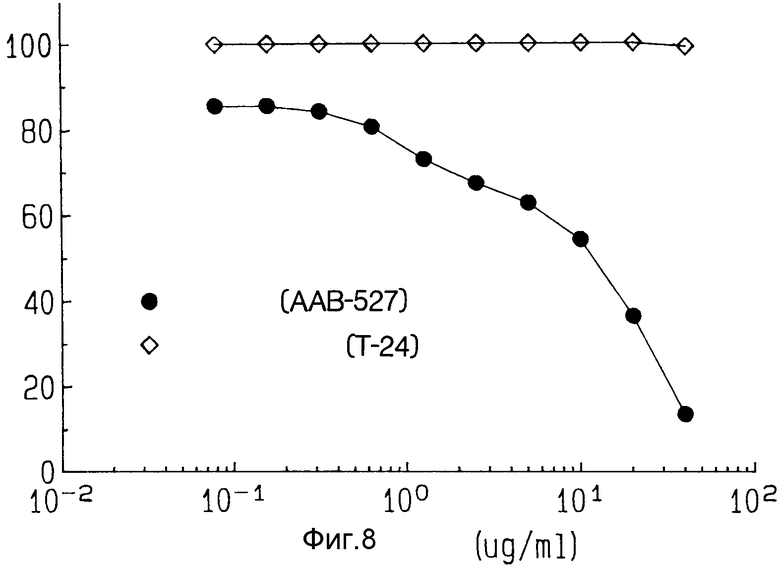
На фиг. 8 показана цитотоксичность ZME-гелонина к положительным по антигену клеткам-мишеням меланомы (AAB-527) и к отрицательным по антигену клеткам T-24 в культуре.
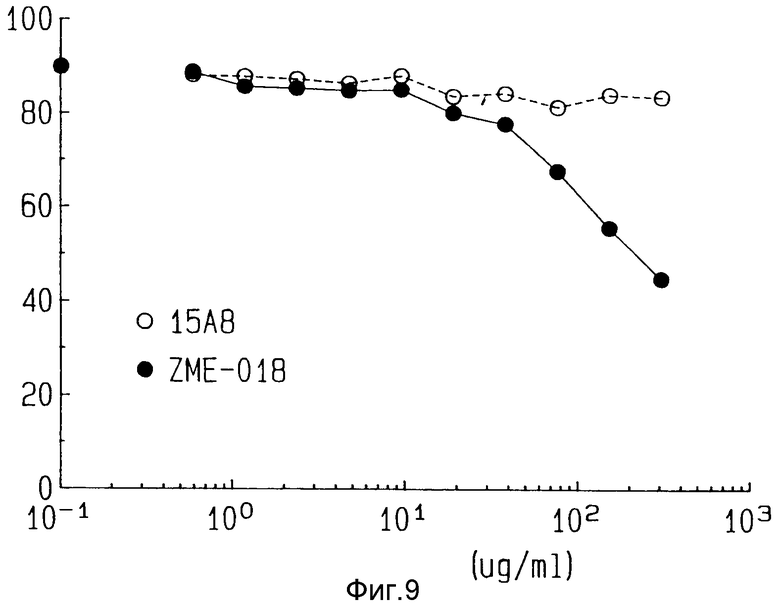
На фиг. 9 показано влияние свободных антител на цитотоксичность ZME-гелонина.
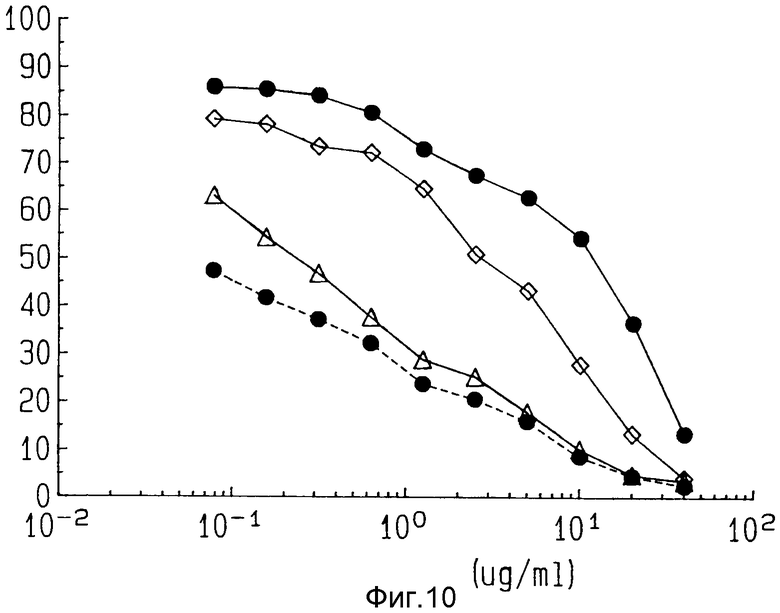
На фиг. 10 показано влияние, которое оказывает IFN-α , IFN-γ и TNF на цитотоксичность ZME-гелонина. Черные кружки показывают реакцию на дозу только ZME-гелонина. Белые ромбы показывают реакцию на дозу ZME-гелонина и IFN - γ . Белые треугольники показывают реакцию на дозу ZME-гелонина в присутствии заданного количества TNF - α . Черные кружки с пунктирной линией представляют кривую реакцию на дозе ZME-гелонина в присутствии фиксированного количества IFN - α .
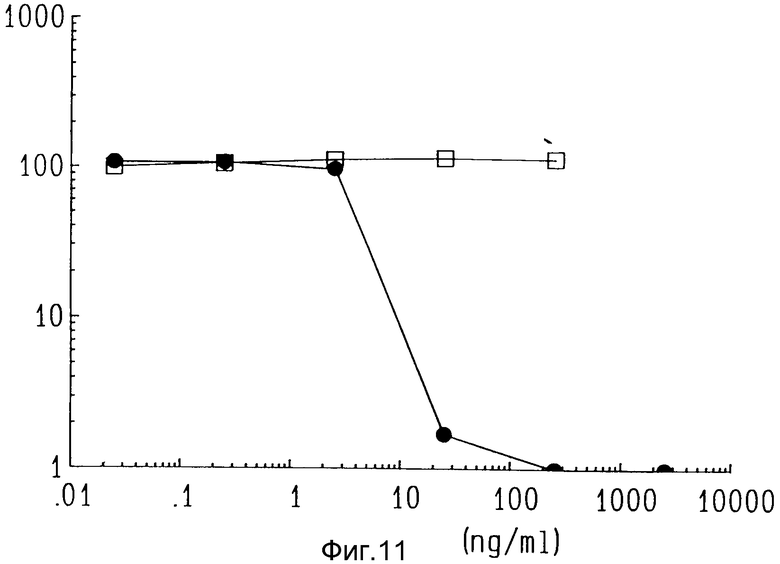
На фиг. 11 показано воздействие ZME-гелонина на положительные (A-375, черные кружки) и отрицательные (CEM, белые квадраты) по антигену клетки в пробе стволовой клетки человеческой опухоли.
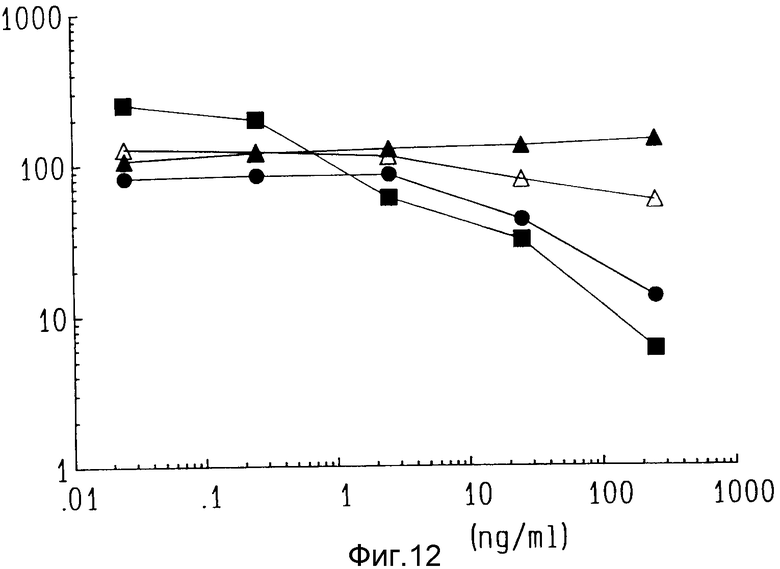
На фиг. 12 показан цитотоксичный эффект ZME-гелонина на выживаемость различных линий стволовых клеток, полученных из свежих образцов для биопсии от 4 различных пациентов.
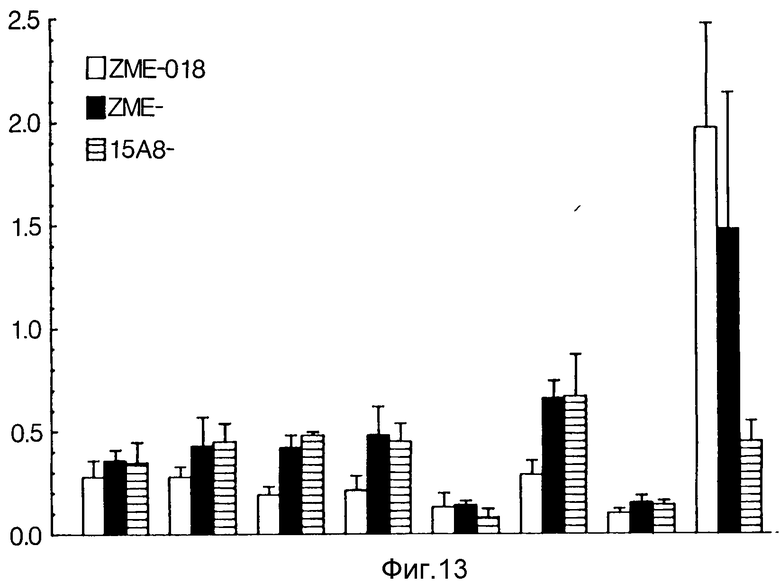
На фиг. 13 показано распределение ткани антител ZME и конъюгата ZME-гелонина у голых мышей, несущих ксенотрансплантат человеческой меланомы.
Подробное описание изобретения.
Используемый здесь термин "моноклональное антитело" означает состав, обладающий однородной популяцией антител. Не предполагается наличия ограничений как по источнику антител, так и по способу их получения.
Клетки меланомы экспрессируют на поверхности клеток антиген 240 кД (GP 249). Производят антитела к этому антигену. Антитело ZME-018 (от "Хайбритекс") является мышиным моноклональным антителом Ig G2, распознающим гликопротеин 240 кД, присутствующий на большинстве клеток человеческой меланомы. Могут быть получены моноклональные антитела изотопов Ig G1, Ig G2a и Ig G2b, которые различают эпитопы этого антигена 240 кД. Этот эпитоп 240 кД антигена ZME будет обозначен для целей настоящего изобретения как эпитоп ZME. Таким образом, все антитела, распознающие эпитоп ZME, функционально эквивалентны.
Эти представительные культуры гибридом, чьи клетки секретируют антитела одного одиотипа, например, все распознают эпитоп ZME, были заложены в Американской коллекции типовых культур на 12301 Парклаун Драйв, Роквилл, Шт. Мэриленд 20852 ("ATCC").
Эти моноклональные антитела могут быть получены способами, известными специалистам в этой области. Характеристики и процедура получения культур этих клеток гибридом, создающих эти антитела, подробно описанных. (Wilson и др. Int. J. Cancer 28 : 293 (1981), Jmai и др., Transplant broc. 12: 380-383 (1980)). Вкратце можно сказать, что гибридомы были получены из клеток миеломы мышей линии Sp2/0-Ag-14 и спленоцитов мышей, иммунизированных с клетками миеломы линии М21, как это описано Имаи и др. ((1980) Transplant broc. 12: 380-383). Гибридомы, секретирующие моноклональные антитела (монАн) 225. 28 и 465. 12 субклонированы и репродуцированы in vitro и in vivo. Оба моноклональных антитела относятся к подклассу Ig и G2a и перед использованием были очищены от асцитической жидкости мышей путем абсорбции и элюирования из сефарозы 4B белка A ("Фармациа", Пискэтуэй, шт. Нью-Джерси, США).
Гибридомы, образующие антитела, реагирующие с клетками меланомы человека, но не с нормальными клетками человека, характеризовались далее. Антитела, образованные линией клеток ZME и гибридомой, образующей функционально эквивалентные антитела, реагировали с антигеном ZME на клетках меланомы человека. Они реагировали также с 70-80% произвольным образом выбранных испытываемых меланом и не продемонстрировали реакции с различными тканями, как показано в таблице.
Термин "Функционально эквивалентный", применяемый здесь к взятым в качестве примера мышиным моноклональным антителам, действующим против меланомы человека, означает моноклональное антитело, которое: (1) перекрестно блокируется с приведенным в качестве примера моноклональным антителом, (2) избирательно связывается с клетками, экспрессирующими антиген ZME, такими как клетки меланомы человека, (3) имеет изотипы G или M, (4) связывается с антигеном ZME, выявленным иммунопреципитацией или сэндвич-иммуноанализом, и (5) при сцеплении с гелонином его доза ингибирования культуры тканей составляет по меньшей мере 50% против по меньшей мере одной из линий клеток ААВ-527 или А 375 при использовании дозы 80-100 ед/мл.
Антитело ZME сцепляется с гелонином с использованием N-суццинил-3-/2-пиридилтио/пропионата (СПДП) или 2-иминотиолана в качестве соединительного реагента. Конъюгаты испытывали на клетках ААВ-527 и А 375 при экспозиции на тканевой культуре в течение 72 ч. Конъюгаты антител продемонстрировали приемлемое антипролиферативное действие (доза ингибирования культуры тканей 50% от менее чем 10 ед/мл) против обеих линий этих клеток.
Дополнительные детали, характеризующие антитела, представлены ниже в примерах.
Иммунохимикаты
Иммунохимическими производными настоящего изобретения, обладающими первостепенной важностью, являются иммунотоксины (конъюгаты антител ZME и цитотоксичной составляющей или модификатора биологической реакции) и меченые (например, радиоактивными изотопами, ферментами или люминофорами) производные, в которых метка является средством идентификации иммунных комплексов, включающих меченое антитело.
Цитотоксичная составляющая иммунотоксина может быть представлена цитотоксичным лекарственным средством или ферментативно активным токсином бактериального или растительного происхождения (гелонин), или же ферментативно активным фрагментом (цепь А) такого токсина. Энзиматически активные токсины и их фрагменты предпочтительны и их примерами могут служить гелонин, цепь А дифтерии, несвязывающиеся активные фрагменты токсина дифтерии, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модеццина, альфасерцин, белки Aleuretes fosdii, белки дианатина, белки Phytoiacca americana (РАРI, РАРII и РАР-S) ингибитор момдика харантиа, курцин, кротин; ингибитор спонария оффициналис, митогаллин, рестриктоцин, феномицин и эномицин. Наиболее предпочтительной является соединение с гелонином.
Модификаторы биологической реакции, которые могут быть соединены с антителом ZME и использованы в настоящем изобретении, включают лимфокины и цитокины, такие как IL - 1, IL - 2, интерфероны (α,β или γ/ TNF, LT, TGF-β и IL - 6, но не ограничиваются ими. Эти модификаторы биологической реакции оказывают на опухолевые клетки разнообразное воздействие. Среди вариантов воздействия - увеличение убитых опухолевых клеток под прямым воздействием, а также увеличение числа убитых клеток за счет процессов, способствующих иммунной защите. Соединение антитела ZME с этими модификаторами биологической реакции способствует избирательной локализации внутри клеток и поэтому улучшению антиполиферационного действия при одновременном подавлении нежелательных эффектов, ведущих к токсичности по отношению к клеткам, не являющимся целями.
Цитотоксичные лекарственные средства, полезные для настоящего изобретения, включают адриамицин (и его производные), цис-платиновый комплекс (и его производные), блеомицин и метотрексан (и его производные), но не ограничиваются ими. Эти цитотоксичные лекарства иногда полезны для клинического лечения рецидивных опухолей и в особенности меланомы, но их использование осложняется серьезными побочными эффектами и повреждениями клеток, не являющихся целями. Антитело ZME может служить полезным носителем таких лекарств, обеспечивая как эффективные пути доставки к опухоли, так и улучшения внедрения в сами опухолевые клетки. Кроме того, целенаправленная доставка антителами цитотоксичных лекарств к опухолям обеспечит защиту чувствительных органов, таких как печень, почки и костный мозг, от отрицательного воздействия химиотерапевтических средств. Использование лекарств, соединенных с антителами ZME как системой доставки, позволяет снизить дозу самих лекарств, поскольку все лекарственные составляющие соединяются с антителами, концентрирующимися внутри опухоли.
Конъюгаты моноклонального антитела могут быть получены с помощью различных биофункциональных белковых соединительных средств. Примерами таких реагентов могут служить SPD p. IT, биофункциональные производные имидоэстеров, таких как диметиладипимидат, HCl, активные сложные эфиры, такие как дисуццинимидил суберат, альдегиды, такие как глутаральдегид, соединения бис-азидо, такие как бис /n-азидобензоил /гексанэдиамин, производные бис-диазония, такие как бис-/n-диазонийбензоил/-этилендиамин, диизоцианаты, такие как толуол 2,6-ди-изоцианат и бис-активные соединения фтора, такие как 1,5-ди-фторо-2,4-динитробензол.
При использовании для уничтожения клеток меланомы человека in vitro в диагностических и терапевтических целях конъюгаты обычно будут добавляться к среде культуры с концентрацией по меньшей мере около 10 нМ. Технология приготовления и способ введения при использовании in vivo не имеют решающего значения. Обычно будут применяться водные составы, совместимые с культурой или средой перфузии.
Цитотоксичные радиофармацевтические препараты для диагностики и лечения опухолей, несущих антиген ZME, таких как меланома, могут быть получены путем соединения изотопов с высокой линейной передачей энергии с антителами. Термин "цитотоксичная составляющая", применяемый здесь, должен включать такие изотопы.
Метки, которые применяются при приготовлении меченых вариантов антител, включают составляющие, которые могут быть выявлены непосредственно, такие как флуоресцины и радиоактивные метки, а также составляющие, такие как ферменты, которые должны включаться в реакцию или быть введены для того, чтобы быть отмеченными. Примерами таких меток, являются 32P, 125I, 3H, 14C, флуоресцин и его производные, родамин и его производные, дансил, умбеллиферон, люцифериа, 2,3-дигидрофталзаинедионы, пероксидаза хрена, щелочная фосфатаза, лисозима и дигидрогеназа глюкозы-6-фосфата. Антитела могут быть связаны с такими метками известными способами. Так, возможно использование для соединения антител с перечисленными выше флуоресцентными, хемиолюминесцентными или ферментными метками соединительных агентов, таких как альдегиды, карбодиимиды, дималеимиды, имидаты, сукцинимиды, бис-диазотированный бензадин и тому подобное.
Антитела и меченые антитела могут быть использованы в различных иммуновизуализационных и иммуноаналитических операциях с целью выявления присутствия у пациента опухолей, экспрессирующих антиген ZME, таких как меланому, или для определения состояния такого ракового заболевания у пациента, диагноз которому уже поставлен. При использовании для определения состояния рака может быть применен количественный иммуноанализ. Такие обследования проводят периодически, а результаты сопоставляют, чтобы определить, растет или уменьшается опухоль пациента. Обычные методы анализа, которые могут быть применены, включают прямой и косвенный анализ. Прямой анализ предусматривает выращивание образца ткани или клеток пациента с меченым антителом. Если входящие в пробу несущие антиген ZME клетки включают клетки меланомы, меченое антитело свяжется с этими клетками. После промывания ткани или клеток с целью удаления несвязанного меченого антитела образец ткани обследуется на предмет присутствия меченых иммунных комплексов.
Для использования в диагностических целях антитела будут предлагаться в форме наборов. Эти наборы обычно будут включать: антитела в меченой форме в подходящей упаковке, реагенты для выращивания и промывки и питательные среды или отделяющие реагенты в зависимости от характера метки. Кроме того, могут также быть включены средства контроля антигена ZME и инструкции.
Введение иммунотоксинов, являющихся предметом настоящего изобретения, лицу, которому был поставлен диагноз наличия опухоли с антигенной детерминантой ZME, позволит нацелить и сосредоточить цитотоксичный агент в том месте, которое необходимо для уничтожения клеток опухоли. Такое нацеливание цитотоксичных реагентов позволяет избежать или уменьшить нежелательное токсичное воздействие на другие органы, ткани и клетки.
При использовании в целях терапии in vivo иммунотоксины вводятся пациенту в терапевтически эффективных количествах (т.е. количествах, которые позволяют ликвидировать или уменьшить опухоль пациента). Обычно они будут вводиться парентерально, предпочтительно внутривенно. Доза и схема приема лекарственного средства будут обычно зависеть от характера рака (первичный или от метастаз) и его популяции, характеристик конкретного иммунотоксина, например его терапевтического индекса, пациента, истории болезни. Количество иммунотоксина, вводимого пациенту, будет обычно находиться в пределах от 0,1 до приблизительно 10 мг/кг веса пациента.
Для парентерального введения иммунотоксины будут приготовлены в форме, удобной для дозирования и инъекций (раствор, суспензия, эмульсия), в сочетании с фармацевтически приемлемым парентеральным наполнителем. Такие наполнители по сути своей нетоксичны и не дают лечебного эффекта. Примерами таких наполнителей являются вода, физиологический раствор, раствор Рингера, раствор декстрозы и 5% сывороточный альбумин человека. Безводные наполнители, такие как нелетучие масла и этиловый олеат, могут применяться равным образом. В качестве носителей могут применяться липосомы. Наполнитель может содержать небольшие количества добавок, таких как вещества, улучшающие стабильность химическую и изотоничность, например буферы и консерванты. Иммунотоксин обычно будет приготавливаться с такими наполнителями при концентрации около 0,1 - 10 мг/мл.
Токсин гелонин очищался из семян gelonium multiflorum способом Стирпа и др. , Вкратце, гелонин экстрагировали из посева путем гомогенизации в буферном солевом растворе (pH 7,4). Супернатант подвергли концентрации после диализа против 5 мМ фосфата натрия (pH 6,5), после чего гелонин подвергли дальнейшей очистке ионнообменной хроматографией; как показано в примере 1. Чистоту токсина гелонина определяли методом жидкостной хроматографии высокого давления (ЖХВД) и электрофорезом в геле додецилсульфат-полиациламина натрия (СДС-пейдж). Токсин гелонин перемещается как единая полоса с приблизительной молекулярной массой 29-30000 дальтон.
Действенность токсина гелонина измеряли как описано в примере 2 на ингибировании синтеза белка в бесклеточной системе.
Антитело МЕ-018, модифицированное, как описано в примере 5, SPDP, было соединено с гелонином, модифицированным иминотиоланом, как описано в примерах 3 и 6. Антитело, связанное с гелонином, подвергли очистке, как описано в примере 7 методом колоночной хроматографии в колонке Сефадекс G - 75.
Токсичность антитела, соединенного с гелонином определяли по ингибированию синтеза белка, и с помощью опытов in vitro и in vivo определяли его антипролиферативное действие.
Приведенные ниже примеры дают подробное описание приготовления, определения характеристик и использования иммунотоксинных моноклональных антител, являющихся предметом настоящего изобретения. Эти примеры никоим образом не ограничивают сферу действия настоящего изобретения.
Пример 1.
Очистка гелонина
Семена Gelonium multiflorum были освобождены от оболочки, а сердцевину измельчили в гомогенизаторе с восемью объемами 0,14 М NaCl, содержащими 5 мМ фосфата натрия (pH 7,4). Гомогенат оставили на ночь при 4oC при непрерывном перемешивании, охладили на льду и центрифугировали при 35 тыс. g в течение 20 мин при температуре 0oC. Супернатант сняли, подвергли диализу против 5 мМ фосфата натрия (pH 6,5) и концентрации с помощью фильтра 10 млн-1. Образец поместили в ионообменную колонку СМ-52 (20 • 1,5 см), уравновесив 5 мМ фосфата натрия (pH 6,5). Материал, приставший к ионообменной смоле, элюировали 400 мл 0-0,3 М линейного градиента при расходе 25 мл/ч при температуре 4oC. Было получено пять фракций по 1 мл. Фракции замерялись на спектрофотометре при длине волны 280 нм. Гелонин элюировался приблизительно во фракциях 55-70 и на них приходится последний пик гелонина. Фракции 55-70 объединили, подвергли диализу против дистиллированной воды и концентрации путем лиофилизации. Чистоту и молекулярную массу каждого препарата проверяли с помощью жидкостной хроматографии высокого давления, используя метод эксклюзионной хроматографии в колонке TSK 3000 с буфером 50 мМ фосфата натрия, pH 7,4 и с помощью электрофореза в 15% геле доцецилсульфата-полиакриламида натрия (SDS - page). Гелонин мигрировал в форме отдельной полосы с приблизительной молекулярной массой 29-30 тыс. дальтон.
Пример 2
Определение активности гелонина
Активность гелонина определяли в ходе опытов по ингибированию синтеза бесклеточного белка. Опыты по ингибированию синтеза бесклеточного белка выполняли, последовательно добавляя к 50 мкс лизата ретикулоцита кролика, подвергнутого оттаиванию непосредственно перед применением, следующие компоненты: 0,5 мл 0,2 М Трис HCl (pH 7,8) 7 8,9 мл этиленгликоля и 0,25 мл 1 M HCl, перемешивая после каждого добавления.
Двадцать микролитов смеси соль-аминокислота-энергия (САЕМ), состоящей из 0,375 M KCl; 10 мМ Mg (CH3CO2)2, 15 мМ глюкозы, 0,25-10 мМ аминокислот (исключая лейцин), 15 мМ ATP, 1 мМ Трис-HCl (pH 7,6), 10 ул креатенин-фосфат-креатинин фосфокиназы, 8 ул 14C лейцина (Amersham 348 mCi мМ), с добавлением 1,5 ул растворов, содержащих различные концентрации смеси гелонина. Смесь инкубировали в течение 60 мин при 30oC. Внедрение 14C - лейцина прослеживали по аликвоте смеси путем осаждения синтезированного белка на стекловолоконных фильтрах, промывания трихлоруксусной кислотой и ацетоном и измерением радиоактивности счетчиком бета-частиц с использованием сцинциллирующей жидкости Аквасол. Для сцепления с антителами использовали гелонин с удельной активностью не менее 4 • 109 ел/мг. Единицей активности гелонина является количество белка гелонина, вызывающего 50% ингибирование внедрения [14C] лейцина в белок в свободной пробе клеток.
Пример 3.
Модификация гелонина иминотиоланом
Концентрация гелонина в физиологическом растворе с фосфатным буфером была доведена на микроконцентраторе Центрилон 10 приблизительно до 2 мг/мл. К нему добавили гидрохлорид триэтаноламина (TEA/HCl) pH 8,8, этилендиаминтетрауксусную кислоту (ЭДТА) при окончательной концентрации 60 мМ TEA/HCl и мМ ЭДТА pH 8,0. Добавили основной раствор 2 - иминтиолана (20 мМ) до окончательной концентрации 1 мМ, после чего образец инкубировали в течение 90 мин при 4oC в струе газообразного азота.
Избыток иминотиолана удалили путем гель-фильтрации в колонке Сефадекс G - 25/1 • 24 см/, предварительно уравновешенной 5 мМ бис-трис-ацетатного буфера, pH 5,8, содержащего 5 мМ NaCl и 1 мМ ЭДТА. Фракции анализировали на белок на микроплатах с использованием метода Брадфорда на связывание красителя. Для этого в каждую ячейку добавили 40 мкл образца, 100 ул физиологического раствора с фосфатным буфером и 40 ул концентрата красителя. Оптическую плотность при 600 нм определяли на автоматическом считывающем устройстве "Дюнатекс микроэлиса". Гелонин элюирован в пустом объеме (около фракций 14-20). Эти фракции слили вместе и концентрировали на микроконцентраторе Центрикон 10.
Пример 4.
Приготовление и идентификация моноклонального антитела к антигену ZME меланомы
Гибридомы, секретирующие антитела
Самке мыши BAL B/c (Питомник SCRF Ла Джолла, шт. Калифорния) 8-недельного возраста интраперитонеально ввели 107 клеток М21 меланомы человека, дополнив их двумя неделями позже 5 • 10 клеток М21. Самцу мыши NZB/B (Питомник SCRF) 50-недельного возраста ввели сначала 5 • 106 клеток BW5 меланомы, дополнив это пятью инъекциями по 5 • 106 клеток BW5, М51, Коло 38, BW5 и М21 меланомы с месячными интервалами. Через три дня после дополнительной инъекции мышь забивали, извлекали селезенку, скальпелем разделяли спленоциты с целью получения клеточной суспензии. Суспензию клеток селезенки в течение 10 мин обрабатывали 0,17 J H4Cl в 0,01 M Трис, pH 7,2 с целью лизирования красных кровяных телец. Затем эти спленоциты сливали с клетками SP2/OAg 14, как описано Гефтером и др./ (1977) Somat. Cell. Genet 3:231 /со следующими небольшими модификациями : 5 • 107 клеток селезенки и 107 клеток SP2/OAg 14 гибридизировали с 0,3 мл 30% (об/об) полиэтиленгликоля 1000 (ПЭГ) в МЕМ. После инкубирования с ПЭГ клетки промывали и выращивали в течение ночи при концентрации 2 • 106 клеток/мл в D-MEM. На следующий день клетки развели в виде суспензии в 40 мл среды НАТ и пипеткой внесли в 400 ячеек/0,1 мл/ячейку/микроплатах (Костар N 3596, Кембридж, шт. Массачусетс). Через недельные интервалы добавляли по капле прибл. 25 мл среды НАТ. Через 2-3 недели в D-MEM с 10% ФБС были выращены гибридомы, выбранные для дальнейших исследований. Гибридомы развели в культуре ткани и выращивали в полости брюшины мыши BALB/c, которой сначала ввели 0,5 мл Пристана/Фальц энд Бауэр", Стемфорд, шт. Коннектикут/, отработанная питательная среда и акцитическая жидкость были использованы в качестве источника антител.
Клоны гибридомы выращивали in vitro в соответствии с известными методами выращивания тканевых культур, как описано Коттеном и др. Eur. J. Immunol. 3: 136 (1973).
Затем идентифицировались гибридомы, образующие антитела, которые вступают в реакцию с клетками миеломы человека, но не со здоровыми клетками человека.
Как показано в таблице, они не вступают в реакцию с большинством здоровых тканей, прошедших испытания. Антитела, образованные клетками линии ME, и функционально эквивалентные антитела, образованные гибридомами, вступали в реакцию с антигенами ME на клетках меланомы человека, таких как М21.
Пример 5
Модификация моноклонального антитела ME-018 посредством СПДП
N - сукцинимид 3-/2-пиридилтио/пропионат/ (СПДП) в диметилформамиде был приготовлен как основной раствор 3 мг/мл в сухом диметилформамиде. Поскольку кристаллический СПДП подвержен гидролизу, фактическую концентрацию химически активного элемента, образующего поперечные связи, определяли спектрофотометрическим методом анализа оптической плотности при 260 нм в двухлучевом электрофотометре. Концентрацию основы СПДП рассчитывали по следующей формуле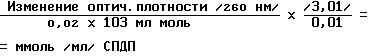
Один миллиграмм моноклонального антитела ME в 1,0 мл физиологического раствора с фосфатным буфером (ФБС) ввели в стеклянную трубку. Основной раствор СПДП медленно добавляли в трубку при приблизительно пятикратном молярном избытке, непрерывно перемешивая. Смесь инкубировали в течение 30 мин при комнатной температуре, перемешивая в течение инкубационного периода каждые 5 мин.
Избыток непрореагировавшего СПДП удалили из образца с помощью эксклюзионной хроматографии на колонке Сефадекс С-25 (1 х 24 см), предварительно уравновешенной 100 мМ буфером из фосфата натрия pH 7,0, содержащим 0,5 мМ ЭДТА (буфер A). Фракции (0,5 мл) собирали и анализировали на содержание белка с помощью метода Брэдфорда на связывание красителя (Брэдфорд, Anal. Biochem. 72: 248-254 (1976)). Оптическую плотность (600 нм) определяли на 96-ячеечной микроплате с использованием автоматического считывающего устройства "Био-текс микроплейт". Антитела элюировали в пустом объеме (фракции 14-20), эти фракции слили вместе и выдерживали при 4oC. Белок концентрировали на микроконцентраторе "Центрикон-30". Осадок в "Центриконе" промыли 100 мМ буфером из фосфата натрия pH 7,0, содержащем ЭДТА (0,5 мМ). Антитела сконцентрировали до конечного объема приблизительно 0,5 - 0,75 мл.
Пример 6
Соединение модифицированных СПДП моноклональных антител ZME-018 с модифицированным имидотиоланом галонином
Один миллиграмм очищенного гелонина (2 мг/мл в ФБС), приготовленный так, как описано в примере 1, модифицировали имидотиоланом так, как описано в примере 3. Моноклональные антитела ME, модифицированные так, как описано в примере 4, перемешали с равным по весу количеством гелонина, модифицированного так, как показано в примере 3. Такая пропорция соответствует пятикратному молярному избытку гелонина в сравнении с антителами. Значение pH смеси было доведено до 7,0 путем добавления 0,05 М буфера ТЕА/HCl pH 8,0, после чего смесь инкубировали в течение 20 ч при 4oC в атмосфере азота. Иодоацетомид (0,1 М) добавили до конечной концентрации 2 мМ с целью блокировать все оставшиеся свободные сульфидрильные группы, после чего еще час продолжали инкубацию при температуре около 25oC. Реакционную смесь хранили при 4oC до очистки путем гель-фильтрации.
Пример 7
Очистка комплексов гелонин-моноклональные антитела 15А8
Гелонин, не вошедший в конъюгат, и продукты с низкой молекулярной массой удаляли из реакционных смесей пример 6 путем гель-фильтрации на колонке Сефадекс S-300 (1,6 х 31 см), предварительно уравновешенной ФБС.
Перед загрузкой в колонку Сефадекс реакционную смесь из примера 6 концентрировали на концентраторе "Центрикон 30" приблизительно до 1 мл. Колонку промыли ФБС. Были отобраны фракции по 1 мл и аликвоты по 50 ул подвергли анализу на белок методом Брэдфорда на связывание красителя (Брэдфорд, Anal Biochem, 72: 248 (1976)).
Как показано на фиг. 2, свободные и не связанные с гелонином антитела элюированы в пустом объеме (около фракций 28-40), в то время как несвязанный гелонин элюирован около фракции 45 - 65.
Для удаления несвязанного ZME-018 высокомолекулярный пик (фракция 28-40) колонки S-300 применили к колонке аффинной хроматографии Сефароза Блу CL-6B (1 x 24 см), предварительно уравновешенной 10 мМ фосфатным буфером (pH 7,2), содержащим 0,1 М NaCl. После загрузки образцов колонку промывали 30 мл буфера до полной элюции несвязанных антител. Колонку элюировали с линейным солевым градиентом от 0,1 до 2 М NaCl в 10 М фосфатном буфере pH 7,2. Содержание белка в элюированных фракциях определяли методом Брэдфорда на связывание красителя.
На фиг. 2 показан профиль элюции в колонке S-300 и показано, что гелонин можно отделить от конъюгата гелонин-антитело и несвязанных антител, которые совместно элюируются в первом пике (около фракций 28-40). Эта картина элюции подтверждается электрофорезом 50 ул аликвот на 5-20% градиентных невосстановительных SDS полиакриламидных гелях, как показано на фиг. 4. Соединительная смесь была помещена на дорожку 3. Показаны полосы, соответствующие свободному гелонину (дорожка 2), свободным антителам (дорожка 1) и одной молекуле гелонина, связанной с молекулой антитела и двумя молекулами, соединенными в молекулой антитела. Пик в пустом объеме колонки S-300, соединяющей свободное антитело и конъюгат антитело-гелонин был помещен на дорожку 4.
Несвязанные антитела отделяли от антител, связанных с гелонином, посредством аффинной хроматографии на колонке Сефароза Блу CL - 6B (1 х 24 см), предварительно уравновешенной 10 мМ фосфатным буфером, pH 7,2, содержащим 0,1 М NaCl. После загрузки образца элюата S-300 колонку промыли 30 мл того же буфера, чтобы полностью элюировать несвязанные антитела.
Антитела, связанные с гелонином и задержанные в колонке, элюировали с линейным солевым градиентом 0,1-2 NaCl в 10 мМ фосфатном буфере, pH 7,2. Комплекс антитело-гелонин элюировали приблизительно 0,7 М NaCl как показано на фиг. 3, где изображен профиль элюции в колонке Сефароза Блу. Содержание белка в элюированных фракциях определяли методом Брадфорда по связыванию красителя. Содержащие белок фракции слили вместе, и картина элюция была подтверждена электрофорезом на 5-20% градиентном невосстановительном полиакриламидном геле. Картина электрофореза комплекса ZME-гелонин показана на фиг. 4. Пик протекания (фракции 14-20) содержит только свободные антитела (фиг. 4, дорожка 5), в то время как фракции 50-80, элюированные с высоким содержанием соли, содержат конъюгат ZME-гелонин, свободный от несвязанного гелонина или антител (фиг. 4, дорожка 6). Конечный продукт содержит антитела ZME, связанные с 1, 2 и 3 молекулами гелонина. Среднее содержание гелонина составляет 1,5 молекулы на молекулу антител.
Система трансляции ретикулоцита кролика in vitro, описанная в примере 2, была использована для оценки активности галонина в исключительно чистом комплексе гелонин-антитело ZME. Одна единица активности в этих опытах была определена как количество белка, необходимого для 50% ингибирования синтеза белка по сравнению с необработанным контрольным образцом. Проведение этих опытов гелонина и конъюгата ME-гелогин равна соответственно 2 • 108 ед/мг и 8,2 • 105 ед/мг. Исключительно чистый гелонин-антитело ME активен в пробе лизата ретикулоцита. Разведение в отношении 1:1000 первоначального образца привело к приблизительно 50% ингибированию синтеза белка, т.е. 50% сокращению внедрения 14C-лейцина в белок. Таким образом, активность исходного препарата составила 1000 ед/мл.
Пример 8
Методы получения клеточной культуры
Отрицательные к антигену ZME клетки карциномы мочевого пузыря человека (Т-24), карциномы шеи человека или положительные к антигену клетки опухоли метастической меланомы человека А375М или ААВ-527 культивировали с использованием минимального дополнения основной среды (МЕМ) с 10% дезактивированной термообработкой зародышевой бычьей сывороткой плюс 100 мМ неэфирных аминокислот, 2 мМ L-глютамина, 1 мм натриевой соли пировиноградной кислоты, витамины и антибиотики. Выращенные клетки подвергли обычному скринингу для удаления примесей микоплазмы.
A. Опыт по пролиферации клеток
Клеточные линии сохраняли в культуре в полной среде при 37oC в инкубаторе с 5% CO2 - увлажненном воздухе. Для опытов с комбинациями TNF, иммунотоксинов IFNα и IFNγ культуры промывали, разделяли версеном и повторно суспендировали в полной среде при плотности 25 • 103 клеток/мл. Аликвоты по двести мл распределялись по 96-ячеечным микропластам, после чего допускалось прилипание клеток. Это привело к получению редкой популяции клеток. Через 24 ч среду заменили средой, содержащей различные концентрации иммунотоксинов, токсинов, TNF, IFNg или IFNa. Клетки инкубировали в течение 72 час после чего выполнили анализ на относительную пролиферацию клеток путем окрашивания кристаллическим фиолетовым.
B. Окрашивание кристаллическим фиолетовым.
Клетки трижды промыли ФБС, содержащим связанные кальций и магний, и окрасили 20% (об/об) метанолом, содержащим 0,5% (об/об) кристаллической фиолетовой. Связанный краситель элюировали 150 ул буфера цитрата Соренсона /0,1 М цитрат натрия, pH 4,2 - 50% /об/об этанола/ в течение 2 ч при комнатной температуре. Оптическую плотность измеряли при 600 нм с использованием считывателя микроплат "Био-Текс". Относительную пролиферацию клеток (ОПК) рассчитывали следующим образом:
C. Проба колонии клеток человеческой опухоли
Образцы опухоли для биопсии были получены у пациентов, страдающих меланомой в ходе клинически предписанных процедур биопсии. Асептически приготовили суспензии опухолевых клеток (Лейбовиц и др., Int. J. Cell Cloning 1 : 478 - 485 (1983)). Наряду с этим испытывали линии клеток меланомы А375Р и лейкемии CEM из Американской коллекции типовых культур (Роквилл, шт. Мэриленд). Определение воздействия МЕ-гелонина на суспензии свежих клеток меланомы и линии клеток определяли в пробах колонии клеток человеческой опухоли с использованием стандартных процедур пластинчатой разводки клеток опухоли в полутвердой среде (агарозе) в присутствии полной среды, содержащей 10% зародышевой сыворотки теленка, каждая из 0,5 мл пластинок с культурой содержала 100000 клеток свежей опухоли и 10000 клеток из линий клеток (Гамбургер и др. , Science 197 : 461 - 463 (1977), Салмон и др., N. Engl. J. Med. 298 : 1321 - 1327 (1987), Салмон и др. J. clin Oncol. : 1346 - 1350 (1989)). Конъюгат ZME-гелонин, приготовленный так, как описано выше, испытывали путем добавления на пластины с культурой вскоре после нанесения опухолевых клеток. ZME-гелонин добавляли на трипликатные пластины при каждой из трех концентраций от 0,025 до 250 нг/мл. В дополнение к необработанным контрольным пластинам параллельно испытывали несвязанные моноклональные антитела ZME-18 и свободный гелонин. Культуры линий клеток и опухолевых клеток инкубировали в среднем в течение 10 суток при температуре 37oC и 50% CO2 на воздухе в инкубаторе с увлажнением, с определением формирования колонии по окраске жизнеспособности (Шумейкер и др., Cancer Res. 45 : 2145 - 2153 (1985)) и с помощью автоматизированного прибора анализа изображения, оптимизированного для подсчета колоний (Салмон и др., Int. J. Cell. Cloning 2 : 142 - 160 (1984)). Процент выживания культур, обработанных ZME-018 в сравнении с одновременно необработанными контрольными образцами определяли в ходе тех же экспериментов. После этого графически были построены кривые реакции на дозу.
Пример 9
Сопоставление силы связи антитела ZME, связанного и не связанного с гелонином, с клетками-целями.
Определяли способность связанных и не связанных с гелонином антител ZME к соединению с клетками-целями. Связь ZME-гелонинового иммунотоксина с клетками положительными (клетки AAB-527) и отрицательными (клетки Т-24) по антигену определяли методом проб ELISA.
В каждую ячейку на микроплате добавили 50 тыс. клеток-целей (клетки AAB-527) или клеток, не являющихся целями (Т-24). Клетки высушивали на пластинах в течение ночи при температуре 37oC. После этого клетки трижды промывали тремя сменами фосфатного буфера и в течение ночи сушили на воздухе. После этой обработки антигенные детерминанты поверхности клеток оставались антигенно активными.
После нанесения клеток микроплаты промыли промывочным буфером (9,68 г Трис, 64,8 г хлорида натрия, 16 мл Твин 20, 800 мг тимеразола в 8 л двойной дистилляции). Образцы антител развели в промывочном буфере, содержащем 1% зародышевой бычьей сыворотки (вес/об) (разводящий буфер). В ячейки добавляли по 50 мкл различной концентрации от 0,02 до 50 уг/мл или со связанными, или не связанными антителами ME. После инкубации в течение часа при 4oC супернатант сняли, а ячейки дважды промывали промывочным буфером.
В каждую ячейку добавили 50 мкл пероксидазы хрена обыкновенного, связанной с козьей анти-мышиной Ig G, полученной из Био-Рад и разведенной в отношении 1 : 1000 (об/об) (HPGAM) в разводящем буфере. Пластины инкубировали в течение 1 ч при 4oC, а ячейки дважды промыли промывочным буфером. После инкубации пластин с 50 мл раствора субстрата (80 мМ цитрата фосфата), pH 5,0 (1 мм 2,2 азино-бис/3-этил бенц-тиазолин-6-сульфокислоты) (АВТС) диаммониевая соль ("сигма кемикл") и 4 ул 30% перекиси водорода в темноте в течение 30 мин при комнатной температуре в каждую ячейку добавили 25 ул четырехнормальной серной кислоты. Оптическую плотность при 492 нм определяли на счетчике "Элиса".
Как показано на фиг. 5, как природный ZME, так и конъюгат ZME-гелонин хорошо связываются с клетками-цепями после 60 минут выдержки. Как ни удивительно, но конъюгат ZME-гелонин связывается с клетками-целями лучше, чем природное антитело. Это улучшение не связано с модификацией антител посредством СПДП, поскольку модифицированное СПДП ZME ведет себя идентично природному ZME. Улучшение не связано также со связью клеток-целей с гелониновой частью молекулы, поскольку предварительная обработка клеток-целей природным гелонином не оказывает влияния на связь антитела, так и иммунотоксина.
В ходе опытов "ЭЛИСА" не определялась связь с отрицательными по антигену клетками Т-24 ни ZME, ни ZME-гелонина.
Пример 10
Цитотоксичность гелонина и комплекса гелонин-антитело ZME
Исследования цитотоксичности конъюгата ZME-гелонин проводили на положительных по антигену клетках после продолжительной (72 ч) экспозиции иммунотоксину или природному гелонину. Как показано на фиг. 6, когда положительные по антигену клетки AAB-527 подвергались воздействию 0,1/нм ZME-гелонина, наблюдалась гибель 50%-клеток. Когда клетки подвергли воздействию природного гелонина, потребовалась концентрация в 100 нм гелонина, чтобы уменьшить количество клеток до 50% от их числа в необработанной контрольной пробе.
Клетки-цепи обрабатывали после этого различными концентрациями ZME-гелонина или только гелонина на единичной основе, как определено в примере 2. Как показано на фиг. 7, 50% цитотоксичности достигали, используя 50 ед/мл конъюгата ME-гелонина, в то время как для достижения такого же эффекта требовалось 1 • 107 ед/мл свободного гелонина.
Эффект ZME-гелонина определяли на отрицательных по антигену клетках T-24 в культуре лог-фазы. Как показано на фиг. 8 только гелонин обеспечивает 50% цитотоксичности клеток AAB-527 при концентрации 100 ед/мл, что близко с тем, что выяснено для клеток AAB-527. *ZME-гелонии обеспечивает 50% цитотоксичность клеток-целей T-24 при концентрации 10 мкг/мл. Однако иммунотоксин ZME-гелонин не обладает цитотоксичностью против не являющихся целями клеток T-24, даже при наивысших испробованных концентрациях.
Для того, чтобы продемонстрировать, что цитотоксичность ZME-гелонина опосредуется через поверхностный антиген ZME, фиксированная доза а ZME-гелонина, достигающая 80% цитотоксичности, была добавлена к лог-фазе клеток-целей меланомы в культуре в присутствии свободных антител ZME или не относящихся к данной ситуации антител (15AB, антитело, которое не связывается с клетками меланомы). Как показано на фиг. 9, присутствие растущих количеств антитела ZME подавляет цитотоксичность конъюгата ZME-гелонин, в то время как антитела 15AB влияния не оказывают. Таким образом, цитотоксичность конъюгата ZME-гелонин непосредственно связана со сцеплением антитела ZME с антигеном ZME на клетке-цели.
Пример 11
Модулирование цитотоксичности ME-геллонина посредством IFNα, IFNγ и TNF
Для того, чтобы продемонстрировать влияние, которое оказывает на цитотоксичность иммунотоксина обработка различными модификаторами биологической реакции, лог-фазу клеток меланомы обрабатывали в течение 24 ч фиксированными дозами IFNα/ (200 ед/ мл), IFN/(20000 ед/мл) или rTNFα/(20000 ед/ мл). Предварительно было определено, что эти дозы обладают минимальным (приблизительно 20%) цитотоксичным действием на эти клетки. После этого клетки в течение 72 ч обрабатывали различными дозами ZME-гелонина. Как показано на фиг. 10, обработка rIFNγ ведет к двукратному повышению чувствительности к иммунотоксину ZME-гелонин. Однако, предварительная обработка и rIFNα и TNF в обоих случаях ведет к двукратному повышению чувствительности к иммунотоксину. Добавление фиксированных доз rIFNα, rIFNγ или rTNF к положительным по антигену клеткам ведет к усилению цитотоксичности токсина ZME-гелонин. Обработка rTNFα обеспечивает наибольшее увеличение цитотоксичности иммунотоксина, вслед за которым следует обработка rIFNα и rIFNγ.
Значительное усиление цитотоксичного воздействия ZME-гелонина наблюдалось при предварительной обработке rIFNα и rTNF, но не rIFNγ . В то время как наблюдалось, что IFNα и IFNγ могут оказывать регулирующее воздействие в сторону повышения на некоторые поверхностные антигены меланомы, такие как P-97, они оказывают небольшое воздействие на антиген с большой молекулярной массой (GP 240), распознаваемый ZME (Мюррей и др. Proc, Ain. Assoc. Cancer. Res. 27:313 (1986), Грейнер и др., Cancer Res. 44:3208-3214 (1984), Грейнер и др. Cancer. Res. 46:4984-4990 (1986), Гроакомини и др., J. Jmmunol 133:1649-1655 (1984), Имаи и др., J. Jmmunol 127:505-509, Мюррей и др., J. Biol. Res. Mod. 7: 152-161 (1988)). Поэтому связанный с использованием TNFα и IFNα механизм усиления активности ZME-гелонина неясен, но может включать изменения в скорости внедрения антител, изменения в клеточном действии иммунотоксина или модуляцию любого из нескольких ферментов, опосредованных интефероном.
Поскольку иммунотоксин гелонин-ZME убивает только клетки, которые содержат на своей поверхности антиген ZME, этот иммунотоксин является эффективным способом выявления и уничтожения клеток, содержащих антиген ZME, связанный с опухолью, одновременно сводя к минимуму или предупреждая повреждение здоровых клеток, не связанных с опухолью.
Пример 12
Воздействие иммунотоксина ZME-гелонин на пробу колонии клеток человеческой опухоли
Активность ZME-гелонина оценивали также, используя пробу колонии клеток человеческой опухоли в соединении с клетками, полученными биопсией у четырех пациентов с меланомой.
In vitro цитотоксичность против клеток человека в культуре оценивали также по колонии клеток, описанной в примере 8C. Различные дозы иммунотоксина ZME-гелонин добавляли к линиям клеток, положительным (A-375 меланомы) и отрицательным (CEM) по антигену. Выживаемость колоний определяли через 72 ч после добавления. Как показано на фиг. 11, дозы иммунотоксина в пределах от 0,25 до 2500 нг/мл способствовали подавлению колоний линий клеток, положительных к антигену (черные кружки). ZME-018 и свободный гелонин по отдельности или вместе не были цитотоксичны. Не оказано воздействия на линию, отрицательную по антигену (CEM) даже при наивысшей концентрации испытываемого иммунотоксина (светлые квадраты).
Воздействие ZME-гелонина на 4 различных свежих образца биопсии показаны на фиг. 12. При наивысшей испытанной дозе иммунотоксина (250 нг/мл) на двух образцах обнаружено сокращение выживания клеток меланомы, образующих колонию на 80 - 90%. Один пациент продемонстрировал при этой дозе подавление роста клеток на 50%, в то время как один пациент не показал цитотоксичности иммуноконъюгата. На третьем образце было отмечено только умеренное (25%) уменьшение колонии. В четвертом образце при наивысшей дозе иммунотоксина было отмечено усиление роста. Кроме того, усиление роста наблюдалось на одном образце при низких дозах, в то время как более высокие дозы вызывали значительную токсичность. Как и в экспериментах с клеточными линиями, добавление несвязанных ZME-018 и свободного гелонина при испробованных дозах не было цитотоксичным.
В то время как пробы на колониях клеток не безошибочны, приблизительно 75% клинически активных противоопухолевых средств положительны в этой системе испытаний. Средства, неактивные на колониях клеток, до сих пор оказались неактивными и в клинических условиях. Следовательно, активность конъюгата ZME-гелонин имеет 75% вероятности продемонстрировать положительную клиническую ценность.
Пример 13
Распределение антител ZME в ткани
Распределение в ткани меченого 125I антитела ME сравнивали с иммунотоксином, относящимся к данному случаю (ZME-гелонин) и иммунотоксином, не относящимся к делу (15A8-гелонин). Каждое антитело или конъюгат антитела вводили внутривенно в хвостовую вену пяти голых мышей, несущих ксенотрансплантат меланомы человека. Каждое животное получило 10 мг общего белка, меченного 0,5 мкС 125I в общем объеме 100 мкл физиологического раствора с фосфатным буфером.
Как показано на фиг. 13, не относящийся к данному случаю конъюгат 15A8-гелонин не локализовался специально на опухолевых тканях (отношение T/B равно 0,5). В отличие от этого как ZME, так и конъюгат ZME-гелонин, относящиеся к данному случаю, продемонстрировали целенаправленную локализацию (отношения T/B равны соответственно 2,0 и 1,5). Статистически значительных различий в поглощении 125I опухолью после введения ZME или ZME-гелонина не наблюдалось. Специалист в данной области может легко прийти к выводу, что настоящее изобретение хорошо подходит для достижения целей и получения упомянутых результатов и преимуществ, также как и присущих ему. Соединения, методы, процедуры и приемы, описанные здесь, являются представительными для предпочтительных вариантов осуществления и приведены в качестве примеров, но не должны обозначать пределы сферы настоящего изобретения. Специалисты в данной области могут представить себе изменения, которые находятся в пределах существа, а также объема данного изобретения, определенного помещенной ниже формулой изобретения.

Коньюгат антимеланомного антитела и цитотоксического агента представляет собой коньюгат моноклонального антитела ZME-018 к антигену GP 240 раковых клеток меланомы, а в качестве цитотоксического агента он содержит токсины, или цитокидальные средства, или цитостатики, способ ингибирования меланомы заключается во введении млекопитающему с меланомой коньюгата парентерально в дозе от 0,1 до 10 мг на 1 кг веса тела. 2 с. и 2 з.п.ф-лы, 13 ил, 1 табл.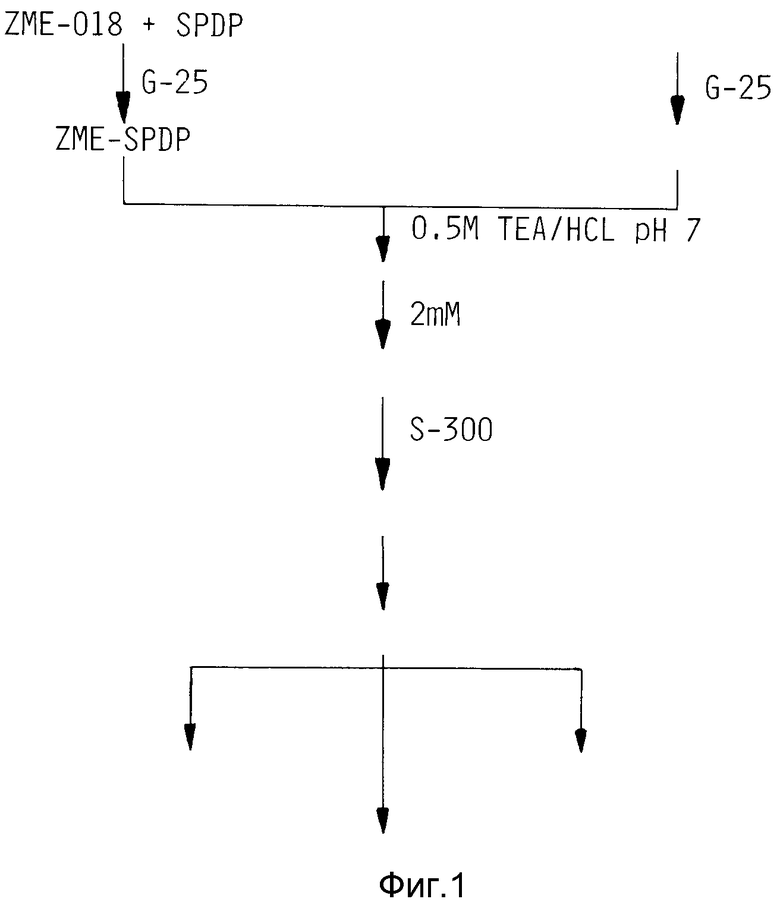
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Selvam et al., Cancer Reslarch., 1987, v.47, pp.3169 - 3173 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Gallego et al., Jnt | |||
| J.Cancer, 1984, v.33, pp.737 - 479 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Zambert et al., J | |||
| Biol | |||
| Chem., 1985, v.260, pp.12035 - 12038. | |||

