Изобретение относится к способу усиления иммунного ответа против целевого антигена путем введения антигена в липосому вместе с дополнительным компонентом, содержащим, по крайней мере один сайт узнавания Т-лимфоцита-хелпера.
Известно, что антиген, используемый для изготовления вакцины, сообщает иммунитет к инфекции путем индуцирования в организме иммунного ответа. Отсюда очевидно, что антигены обладают различной степенью пригодности к их использованию для вакцины в зависимости от силы иммунного ответа, который они индуцируют. По своей природе, антигены могут быть сильными (высокоиммуногенными) и слабыми (низкоиммуногенными). Кроме того, антигены могут быть классифицированы как тимус-(Т-) зависимые и тимус -(Т-)-независимые антигены в зависимости от их способности продуцировать хелперную активность Т-клеток. Эта Т-клеточная активность ассоциируется с продуцированием антител класса IgG и IgA. Т-независимые антигены индуцируют образование антител IgM, но при этом, они не индуцируют B-клетки для переключения на синтез антител класса IgG или IgA. Продуцирование иммуноглобулинов класса IgM представляет собой временный процесс, который у лабораторных животных и у человека продолжается всего несколько месяцев; тогда как продуцирование антител класса IgG и IgA обычно продолжатся в течение нескольких лет. Поэтому, более предпочтительно индуцировать Т-зависимый ответ к какому-либо конкретному антигену. Антигены на основе углеводов или полисахаридов являются Т-независимыми антигенами и имеют ограниченное применение, например, они используются в качестве вакцин для детей. Аналогично, многие полипептиды, которые представляют собой сайты распознавания антитела, являются Т-независимыми антигенами, и поэтому неспособны генерировать IgG и IgA-антительные ответы, желательные для вакцины.
Известно, что более сильная иммунная реакция в ответ на "слабый" антиген может быть вызвана посредством конъюгирования (ковалентного связывания) антигена с хелперным белком, усиливающим иммунный ответ. Например, в пат. США 4761283 показано, что слабый иммуногенный ответ к некоторым бактериальным капсулярным полимерам может быть усилен путем конъюгирования антигена с бактериальным хелперным белком, который сам по себе способен индуцировать антигенный ответ. В качестве хелперного белка обычно используют токсин, например, такой, как дифтерийный токсоид (Diphtheria Toxoid). Однако, использование указанного токсичного хелперного белка имеет свой недостаток, который заключается в том, что природная токсичность хелперного белка может лимитировать дозу конъюгата "антиген-хелперный белок", а значит лимитировать его эффективность.
Известно также, что иммунный ответ на целевой антиген подавляется действием первичной иммунизации организма к хелперному белку, вызывающему эпитопную суппрессию. Если хозяйский организм примировать (т.е. подвергнуть первичной иммунизации) для реакции с хелперным белком, то этот организм будет очищать тело от конъюгата "целевой антиген-хелперный белок" перед тем, как этот организм сможет инициировать иммунологический ответ на целевой антиген.
Указанные ранее используемые конюъгаты формируют путем ковалентного связывания антигена с хелперным белком-носителем. Усиление иммунного ответа является особенно важным для Т-независимых антигенов, которые могут быть конъюгированы с пептидом, содержащим, по крайней мере, один сайт узнаваний Т-хелперной клетки, поскольку это позволяет получить Т-зависимый ответ на Т-независимый антиген. Используемые в настоящее время ковалентно связанные конъюгаты имеют, однако, ограниченные эффективности, обусловленные структурными ограничениями, вносимыми процессом ковалентного связывания, то есть, конформационными изменениями, потенциальной недоступностью сайтов связывания, и невозможностью варьировать соотношения компонентов. Кроме того, указанные конъюгаты могут обуславливать ограничения доз и вызывать эпитопную супрессию.
Липосомы представляют собой мембранные везикулы, образованные дисперсией липидов в водных средах. Способы получения липосом хорошо известны специалистам, и проиллюстрированы, например, в пат. США 4565696 и 4235871, вводимых в настоящее описание посредством ссылки (однако, эти способы не ограничиваются лишь указанными примерами). Липосомы обладают некоторыми свойствами, которые являются необходимыми для in vivo-носителя, а именно, низкой токсичностью, низкой иммуногенностью и низкой способностью к биодеградации. Было также показано, что липосомы могут усиливать иммунный ответ к антигенам у лабораторных животных. Антигенны либо захватываются вовнутрь водных участков липосомы, либо ассоциируются с бислоем. Например, в пат. США описан способ ковалентного связывания иммуногенов с поверхностным бислоем липосомы, и тем самым потенциирования иммунного ответа.
Изобретение относится к продуцированию усиленного антигенного ответа к целевому антигену путем введения этого антигена в липосомный препарат вместе, с по крайней мере, одним хелперным пептидом, содержащим, по крайней мере, один сайт узнавания Т-клетки. Изобретение может быть использовано для усиления иммунного ответа к любому антигену, например, такому, как Пфейффера палочка, менингококк, пневмококк, и стрептококк, а также пептиды, такие, как поверхностные белки гепатита B, поверхностные ВИЧ-белки, белки вируса группа, пептиды вируса парагриппа или гемагглютинин, поверхностные гликопротеины респираторно-синцитиального вируса, и поверхностные гликопротеины холеры.
Целевой антиген (по отношению к которому необходимо усилить иммуногенный ответ) может быть введен в липосомную везикулу посредством связывания антигена с одним из широкого ряда липидных материалов, содержащих активные функциональные группы. Такими липидными материалами являются фосфатидиловые простые эфиры или фосфатидиловые сложные эфиры (например, фосфатидилэтаноламин и фосфатидилхолин), глицериды, цереброзиды, ганглиозиды, сфингомиелины и стероиды (например, холестерин), и т.п. В пат. США 4565696 и 4235871 раскрываются дополнительные липидные материалы для использования при получении липосом.
Альтернативно целевой антиген может быть введен в липосомную везикулу с помощью гидрофобных сил, в случае, если антиген содержит липофильную группу (см., пат. США 4448765) или гидрофобная группа может быть связана с антигеном.
Введение хелперного пептида может быть также осуществлено с использованием либо ковалентных, либо гидрофобных взаимодействий. Например, гемаглютининовый белок (НА) вируса гриппа состоит из двух полипептидных цепей (НА1 и НА2). Полипептидная цепь НА2 содержит последовательность гидрофобных аминокислот, расположенных близ карбокси-конца полипептида, и имеет, по крайней мере, один сайт узнавания Т-хелперной клетки. Так, например, НА2 -цепь полипептида может быть введена в липосомы через трансмембранную гидрофобную область. В пат. США 4448765 описывается введение субъединиц вируса гриппа в липосомы (этот патент вводится в настоящее описание посредством ссылки). Альтернативно, гидрофобный компонент может быть добавлен к хелперному пептиду в целях облегчения ассоциирования с липосомной мембраной, либо халперный пептид может быть ковалентно связан непосредственно с липидными материалами, содержащими активные функциональные группы.
По сравнению с ранее используемыми конъюгатами "антиген-носитель", например, такими, как описаны в пат. США 4761283, изобретение имеет ряд преимуществ. В ковалентно связанных конъюгатах, благодаря ковалентной связи являются возможными изменения в конформации белка-носителя, а также изменения в конформации антигена. Указанные недостатки ранее используемых конъюгатов могут быть устранены с помощью изобретения, где целевой антиген и хелперный пептид ассоциируются с липосомой. Изобретение также позволяет избежать токсичности и эффектов эпитопной супрессии, имеющих место в известных способах. В изобретении применяется нетоксичные липиды, которые предпочтительно используются с выделенными нетоксичными пептидами, содержащими Т-хелперный сайт.
По сравнению с ранее используемыми конюъгатами, изобретение имеет также то преимущество, что оно допускает модификацию антигенной плотности, отношения целевого антигена к хелперному пептиду, и ведение более одного хелперного пептида, несущего сайт узнавания Т-хелперной клетки. Эти факторы могут оказывать влияние на количество специфических антител, продуцируемых против целевого антигена. В случае синтезирования конъюгатов, указанные факторы трудно контролировать, поскольку два компонента должны быть ковалентно связаны, что создает физические ограничения в отношении числа и типа факторов, которые могут быть ассоциированы вместе, затрудняя тем самым доступ к сайтам узнавания. Кроме того, изобретение имеет преимущества по сравнению с предшествующими конъюгатами, которое заключается в том, что введение антигена в липосому способствует усилению продуцирования антитела. Таким образом, изобретение использует указанное свойство липосом в целях дополнительного усиления продуцирования антител к данному антигену и особенно к "слабым" антигенам, которые не способны продуцировать достаточное количество антител.
Липосомные везикуля изобретения позволяют получить новое средство для изготовления синтетических вакцин. Множество копий целевого антигена и хелперного пептида или пептидов могут быть объединены быстрым и простым способом. Помимо обеспечения эффективной и усиленной иммунизации к целевому антигену, способ изобретения позволяет легко проводить тестирования антигенов и хелперных пептидов. Например, изобретение дает простое средство для сравнения способности различных Т-хелперных пептидов усиливать иммуногенный ответ к данному антигену.
Изобретение также дает ценное средство для исследования механизма иммунного ответа.
Другие варианты, особенности и преимущества изобретения будут проиллюстрированы по ходу описания предпочтительного варианта осуществления изобретения, представленного ниже.
Вакцинная композиция изобретения может быть использована для усиления иммунного ответа к любому целевому антигену, а наиболее предпочтительно к Т-независимым антигенам. Настоящее изобретение проиллюстрировано с использованием DNP-CapPE, который был хорошо исследован как Т-независимый антиген. Однако, каждому специалисту ясно, что изобретение может быть применено к любому антигену. В предпочтительном варианте осуществления изобретения, синтезируют синтетическую вакцину, которая индуцирует Т-зависимый иммунологический ответ на Т-независимый антиген. Целевой антиген вводят в липосомный препарат вместе с пептидом или пептидами, которые содержат сайт узнавания Т-клетки. В качестве хелперного пептида может быть использован любой пептид, который содержит сайт узнавания Т-клетки, например, нативный пептид или дезинтоксицированные пептиды, происходящие от токсоидов, например, группа, столбнячного токсина, дифтерийной палочки, стафилококка, стрептококка, коклюшной палочки и E.coli. Для того чтобы определить, содержит ли данный пептид сайт узнавания Т-клетки, может быть осуществлен простой тест. Для этого этот пептид вводят в липосомный препарат вместе с антигеном, таким, как DNP-CapPE, который содержит только B-клеточный эпитом. Если хозяйский организм дает IgG-ответ на полученный липосомный препарат, то испытуемый пептид содержит сайт узнавания Т-клетки и может быть использован в качестве хелперного пептида настоящего изобретения. Субъединица НА2 гемаглютининового белка вируса гриппа является предпочтительной для использования в качестве хелперного пептида, поскольку она хорошо изучена, и поскольку достоверно известно, что она содержит, по крайней мере, один сайт узнавания Т-хелперной клетки, и не содержит ни одного сайта узнавания В-клетки. Таким образом, этот пептид индуцирует Т-зависимый иммунологический ответ на конъюгированный антиген, но не индуцирует антительный ответ против самого себя. Указанное ограничение не является обязательным для осуществления изобретения; и в нем может быть также использован любой хелперный пептид, который содержит сайт узнавания В-клеток, если помимо ответа на нужный антиген необходимо или допустимо получить ответ на хелперный пептид. Однако, предпочтительно, чтобы хелперный пептид выступал в роли эпитопа Т-клетки-хелпера. Аналогичным образом, а изобретении может быть также использован любой хелперный пептид, который имеет сайт узнавания Т-цитотоксичного лимфоцита, если помимо ответа на целевой антиген необходимо или допустимо получить цитотоксичный ответ.
Кроме того, HA2-субъединица является подходящей для использования в качестве хелперного пептида, поскольку она содержит гидрофобную последовательность аминокислот близ карбокси-конца, который обычно проходит через липидную оболочку вируса. Эта трансмембранная область способствует облегчению ассоциирования с липидными бислоями липосом, а HA2 количественно ассоциируется с липоосмными везикулами. Как обсуждалось ранее, могут быть также использованы и альтернативные методы ведения халперного пептида, например, такие, как ковалентное связывание пептида с липидом, или связывание пептида с липидом, или связывание гидрофобной последовательности аминокислот с концом пепетида. Таким образом, каждому специалисту ясно, что для введения халперного пептида в липосому могут быть использованы различные способы.
В качестве целевого антигена был выбран DNP-CapPE, поскольку он является хорошо изученным Т-независимым антигеном. Таким образом, в представленных ниже экспериментах, Т-зависимый ответ на DNP-CapPE является результатом влияния хелперного пептида. Липосомный препарат изобретения может быть использован для усиления иммунного ответа в организме животного. Таким животным может быть любое животное, предпочтительно млекопитающее, а наиболее предпочтительно, собака, кошка, корова, лошадь или человек.
Пример 1. Получение липосом. Для осуществления изобретения может быть использован широкий ряд липидных материалов, включая фосфатидиловые простые эфиры или фосфатидиловые сложные эфиры (например, фосфатидилэтаноламин и фосфатидилхолин), глицериды, цероброзиды, ганглиозиды, сфингомиелины, и стероиды (например, холестерин). Ниже приводятся пример получения липосом с использованием фосфатидилхолина, хотя каждому специалисту должно быть понятно, что для получения липосом может быть использована любая техника, предусматривающая введение целевого антигена и хелперного пептида или белков. Перед получением липосомы может оказаться необходимым ковалентное связывание целевого антигена и/или хелперного пепетида с одним из липидных компонентов (в соответствии с одним из известных способов) для облегчения введения в липосому. Для получения липосом использовали фосфатидилхолин (PC) (Avanti polar lipids, Pelham, AI), очищенный их яичного желтка (EYPC). EYPC добавляли в колбу с круглым дном в нужном количестве, а хлороформ удаляли в роторном испарителе (Buchi 461). Осуществленную липидную пленку ресуспендировали в стерильной воде или фосфатно-солевом буфере (PBS), в количествах, достаточных для получения 10 мМ раствора EYPC. В случае введения в липосому антигена DNP-CapPE, в EYPC-раствор хлороформа добавляли N-2(2,4-динитрофенил) E-аминокапроилфосфатидилэтаноламин (DNP-CapPE). К липидной пленке, для включения в липосомы также добавляли очищенную HA2-субъединицу (поставляемую Doris Bucher, Mt. Sinie, NY), происходящую от гемогглютинина (HA) вируса A/USS R-90-77 H1N1. Затем были образованы липосомы либо путем сусупендирования смеси в стерильном PBS, либо путем растворения липидной смеси в н-октилглюкозиде (5% масс/об. в PBS) с последующим диализом против PBS в течение 16 ч. Количества DNP-CapPE и HA2, включая в отдельные липосомные препараты, варьировались в соответствии с требованиями эксперимента.
Пример 2. Характеризация липосом. Для оценки связывания HA2 с липосомными структурами использовали I125HA2. Для анализа включения DNP-CapPE в липосомы аликвоты исходной суспензии (10 мкл) растворяли в 990 мкл метанола и определяли оптическую плотность (при длине волны 360 мМ) с помощью спектрофотометра. Процент ассоциирования HA2 в препаратах с последующим восстановлением с использованием PBS и диализом не показал значительного изменения с изменением количеств HA2и составлял более 90%. Процент ассоциирования DNP-CapPE в препаратах составлял более 95% во всех препаратах. Высвобождения DNP между инъекциями не наблюдалось. DNP-CapPE оставался ассоциированным с липосомными структурами.
Пример 3. Схема иммунизации. Для эксперимента использовали самок мышей альбиносов аутбредной линии в возрасте 6-8 недель (Timco, Houston, Tx). У животных брали кровь из хвостовой вены, а затем вводили 0,1 мл плацебо (PBS), контрольные липосомы (без HA2 или гаптена) или липосомы, содержащие указанное количество гаптена и/или HA2-пептида. Через три-четыре недели у мышей снова брали кровь из хвостовой вены и проводили повторную иммунизацию с использованием тех же препаратов и в том же количестве, что и при первичной иммунизации. Через 9-14 дней после этого, у животных снова брали кровь из хвостовой вены. Затем, через 8 недель после последней иммунизации проводили третью иммунизацию (но это необязательно) и через две недели спустя также брали кровь. Все указанные инокуляции осуществляли внутримышечно (IM) в задние лапы. Мышей содержали в пластиковых клетках (по 4 животных на клетку) с пороговыми фильтрами, при этом животные получали воду и пищу ad Libitum. Сыворотку отделяли путем центрифугирования при 2000 x г в течение 5 мин при 4oC и хранили при -20oC до исследования. Все сывороточные препараты от каждого эксперимента анализировали на продуцирование антител в простом тесте.
Пример 4. Тесты на анти-динтрофенил (DNP)-сывороточные антитела. Для тестирования антигена использовали динитрофенилированный бычий сывороточный альбумин (DNP-BSA). В качестве контрольного антигена использовали недериватизированный BSA, DNP-BSA получали путем добавления диинитрофторбензола к раствору BSA в молярном соотношении 10:1, а триэтаноламин добавляли до тех пор, пока значение pH не достигнет 9 - 9,5. После 16 ч инкулирования при 20oC несвязанный DNP удаляли путем диализа против PBS. В предварительных экспериментах было установлено, что BSA, дериватизированный в указанном молярном отношении, обнаруживал сывороточное антитело более эффективно, чем белок, содержащий более высокие или более низкие молярные отношения DNP. Уровни сывороточного антитела определяли с помощью процедур ELISA и SPIRA.
ELISA. Планшеты из Иммулона 1 (Dynatech, Deyroit MI) покрывали DNP-BSA (10 мкг на лунку) в 0,05 М карбонатном буфере (pH 9,6), а контрольные лунки покрывали 10 мкг (на лунку) BSA. После 16 ч инкубирования при 4oC планшеты покрывали 5%-ным (об./об.) FCS в PBS. После этой и каждой последующей инкубации планшеты промывали фосфатно-солевым буфером (PBS), содержащим 1% околоплодной телячьей сыворотки (FCS) и 0,2% Твина-20. Серийные двухкратные разведения сыворотки (начиная с 1:100) получали в BPS, содержащем 1% BSA (в дупликации), и 0,1 мл образцов каждого разведения добавляли к антигену и в контрольные лунки. После 5 ч инкубации при 20oC, добавляли козьи антитела, специфичные к конъюгированной со щелочной фосфатной тяжелой цепи мышиного IgG (Fc Specific, Cappel lab, Wegt Chegter, PA), или козьи антимышиные иммуноглобулины подкласса IgG в концентрации, определенной в предварительных экспериментах и превышающей оптимальное количество, необходимое для обнаружения антитела. В случае использования подклассов IgG, затем добавляли конъюгированные со щелочной фосфатазой свиные антитела, специфичные к козьим IgG. После этого добавляли P-нитрофенилфосфат (1 мг/мл) в 10%-ном диэтаноламиновом буфере (pH 9,8) (0,1 мл на лунку). Оптическую плотность определяли спустя полчаса с помощью спектрофотометра Dynatech MP 600. Титры антител экспрессировали как сыворотку самого высокого разведения, что давало оптическую плотность, которая на 0,2 превышала плотность в контрольных лунках.
SPIRA (Твердофазный радиоиммуноанализ). С помощью SPIRA проводили оценку уровней DNP-антител. Эту процедуру осуществляли в соответствии с процедурой ELISA-анализа, но с двумя модификациями. Для анализа использовали гибкие поливинилхлоридные планшеты для микротитрования и для обнаружения связанного мышиного антитела, связанного с DNP-антигеном, использовали 125I-меченые аффинно-очищенные (специфичные к тяжелой цепи) кроличьи антимышиные IgG. Титры антител рассчитывали как описано выше, а специфичность меченых кроличьих антимышиных антител устанавливали путем стандартных перекрестно-штриховых тестов с использованием мышиных моноклональных антител, специфичных к HA2 вируса гриппа, описанных выше. Сравнение титров антител проводили в соответствии с тестом Mann-Withey в случае, если в одном эксперименте участвовали две группы мышей. Если в сравнении участвовали более двух групп, то использовали метод Krusdall-Wallis.
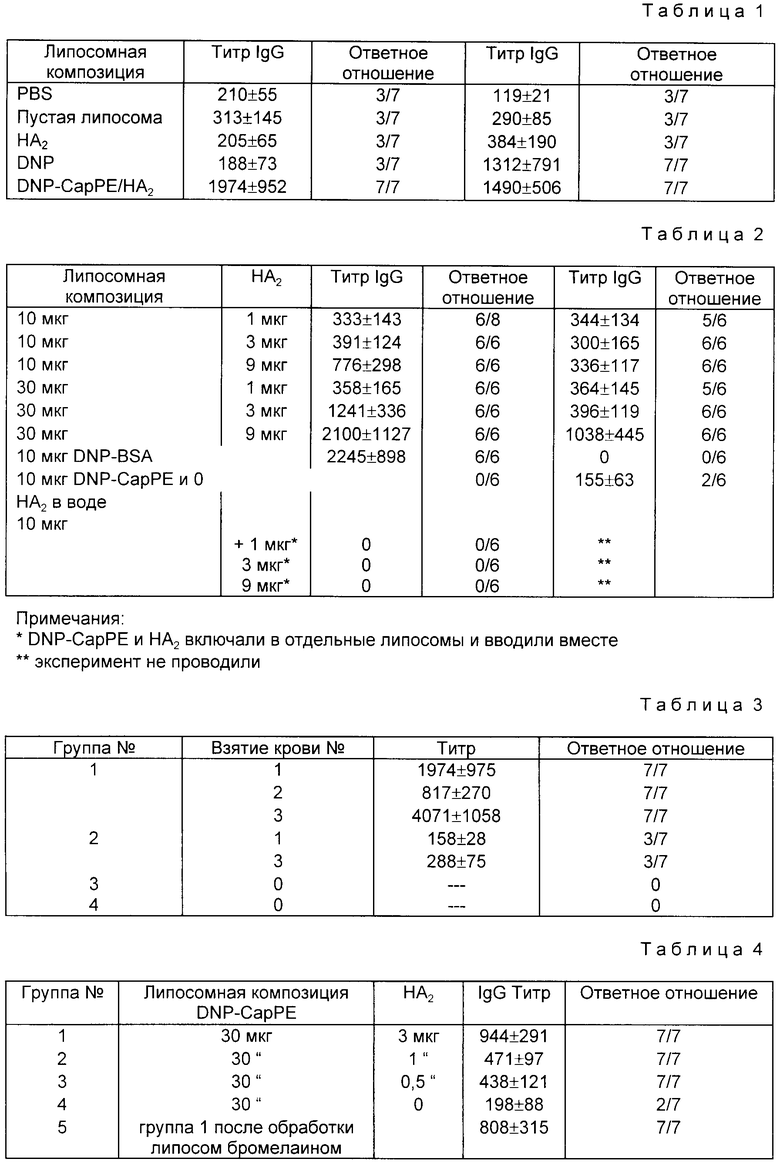
Пример 5. иммуногенность липосом, содержащих DNP-CapPE и/или субъединицу HA2. Группам аутбредных мышей инъецировали два раза (через 4 недели) препараты EYPC-липосом, содержащих HA2 (3,7 мкг), DNP-CapPE (30 мкг) или то и другое (табл. 1). IgG и IgM - ответы анализировали с помощью непрямого ELISA и экспрессировали как среднее из титров антител. Молярное отношение липида к DNP и HA2 составляло 6000:200:1. TLISA - отсчеты показали, что оба препарата, содержащих DNP-CapPE (с HA2 или без HA2), генерировали IgM-ответ у 7 из 7 мышей. Однако, при титровании анти-DNP-IgG-сывороточными антителами результаты показали резкое ухудшение для DNP-липосомной группы, то есть данные титры были аналогичны контрольным группам, тогда как группы, иммунизированные DNP-HA2-липосомами, обнаруживали значительно более высокие уровни сывороточных специфичных анти-DNP-IgG.
Это изменение в классах иммуноглобулинов можно приписать лишь связыванию HA2 и DNP в тех же самых липосомных структурах, поскольку анти-DNP-IgG антитела не генерировались при иммунизации мышей либо смесью HA2 и DNP-CapPE в водном растворе, либо DNP-CapPE-липосомами и HA2-липосомами, инъецированными вместе (табл. 2). Для стимулирования продуцирования анти DNP-IgG антител необходимо, чтобы оба компонента вместе находились в одной и той же липосоме.
Пример 6. Дозозависимые ответы на липосомные DNP и HA2. В этом эксперименте были исследованы три группы иммунизованных животных. Для данного количества DNP-CapPE (5, 10, 30 мкг на инъекцию) в липосомные препараты, состоящие из EYPC (750 мкг на инъекцию), были также включены различные количества HA2 (1, 3 или 9 мкг). У животных брали кровь за один день перед и через девять дней после введения идентичной бустерной инъекции. Сыворотки анализировали на анти-DNP-IgG и IgM с помощью ELISA-теста. Кроме того, две группы мышей были иммунизированы либо 10 мкг DNP-BSA, либо DNP-CapPE и HA2, смешанных вместе в водном растворе. При статическом анализе ELISA-величин IgM (табл.2 две группы, представляющие 5 и 10 мкг DNP-CapPE, не обнаруживали значительных различий с увеличением количества HA2, а поэтому результаты для группы с 5 мкг DNP-CapPE в табл. 2 не представлены. Однако значительное различие наблюдалось при иммунизации мышей липосомами, содержащими 30 мкг DNP-CapPE и 1,3 или 9 мкг HA2, причем, чем выше было их отношение, тем выше был титр IgM. При иммунизации мышей дозой 10 мкг DNP-CapPE никакого IgM-ответа не наблюдалось, а в группе, иммунизированной DNP-CapPE и HA2 в водном растворе, только 2 из 6 мышей давали IgM-ответ. При статическом анализе ELISA-величин IgG (табл. 2), ответы для данного количества DNP-CapPE (5, 10 или 30 мкг) показали значительное расхождение, что свидетельствует о наличии зависимости ответа от дозы как для DNP, так и для HA2. И снова это указывает на то, что продуцирование IgG повышается благодаря присутствию как целевого антигена, так и хелперного пептида в той же липосоме. Мыши, иммунизированные DNP-BSA, дают IgG-ответ, сравнимый с ответом группы, иммунизированной самыми высокими дозами HA2 и DNP в тестируемой липосоме. Для мышей, иммунизированных DNP/HA2 в водном растворе, IgG-ответа не наблюдалось.
Пример 7. Воздействие третьей инъекции DNP-CapPE-липосом на антительный ответ у мышей, предварительно иммунизированных DNP/HA2-липосомами. Анализ на появление вторичного иммунного ответа на целевой DNP-антиген у мышей, предварительно иммунизированных липосомами с DNP-CapPE/HA2, проводили с использованием группы из 7 мышей, которым в третий раз были инъецированы липосомы, содержащие только DNP-CapPE (то есть, сообщая при этом только B-клеточные сайты). Этот эксперимент показал, что вторичный иммунный ответ продуцируется только в том случае, если в исходном липосомном препарате присутствуют вместе DNP-CapPE на HA2 (табл.3). Таким образом, можно с уверенностью утверждать, что изобретение позволяет продуцировать тимус-зависимый иммунологический ответ и вторичный иммунологический ответ на целевой антиген, который может продуцировать DNP -антитела в ответ на целевой антиген без дополнительного требования присутствия сайта узнавания T-клеток. Как показано выше, мыши, иммунизированные липосомами, содержащими DNP-CapPE/HA2, генерируют образование IgG против DNP. Затем тем же самым мышам в третий раз инъецировали липосомы, содержащие только DNP-CapPE (группа I), в результате чего SPRITA-отсчеты резко возрастали по сравнению с первым или вторым взятием проб крови, что свидетельствует о рестимуляции специфических B-клеток при более высоком уровне, чем уровень, продуцированный при первой иммунизации. Мыши, иммунизированные HA2-липосомами (группа 3), не продуцировали какого-либо заметного титра анти-DNP-IgG-антител. Мыши, которым инъецировали DNP-CapPE/HA2-липосомы один раз (группа 2), давали уровень анти-DNP-IgG-антител для первичной иммунизации. Контрольная группа (группа 4), которой инъецировали пустые EYPC-липосомы, не обнаруживали продуцирования анти-DNP-IgG-антител.
Пример 8. HA2 наружной мембраны по сравнению с внутренней HA2 в иммунном ответе. Четыре группы аутбредных мышей иммунизировали EYPC-липосомами, содержащими 30 мкг DNP-CapPE и различные количества HA2 (3, 1, 0,5 и 0 мкг). Кроме того, после предварительной обработки липосом броммелаином (100 мкг/мл), группу мышей иммунизировали этими липосомами, содержащими DNP-CapPE/HA2 (30 и 3 мкг, соответственно). С помощью обработки броммелаином гидролизовали поверхностные белки, оставляя лишь входящий в мембрану конец и целую интернализованную HA2. При статическом анализе ELISA-титра, снова наблюдали ответ в зависимости от дозы с увеличением количества HA2 (табл. 4). Как видно из эксперимента, лишь 0,5 см мкг HA2 в DNP-CapPE-липосомах (на одну инъекцию) достаточно для индуцирования IgG ответа, который статически отличается от ответа на DNP-CapPE-липосомы. Однако, при иммунизации мышей бромелаин-обработанными липосомами, не наблюдалось статического отличия по отношению к ответу, полученному для тех же, но необработанных липосом. Полученные результаты показали, что липосомы с наружной HA2 не требуются и что липосомы должны быть процессированы в целях реализации комбинированного представления B- и T-клеточных эпитопов для иммунокомпетентных клеток.
Пример 9. Распределение подклассов иммуноглобулина IgG по время иммунного ответа. Анализ ELISA-отсчетов показал, что преобладающим подклассом антител был IgG1. IgG1-ответ аутбредных мышей к DNP был аналогичен ответу для низких доз DNP-CapPE (табл.1) при различных концентрациях HA2. Для более высокой концентрации DNP-CapPE (30 мкг) наблюдали эффект дозы для увеличивающихся количеств HA2. Липосомный препарат EYPC, DNP-CapPE и HA2 с 750, 30 и 9 мкг, соответственно, давал IgGI-ответ, сравнимый с ответом группы мышей, иммунизированных 10 мкг DNP-BSA, типичный системы, несущий гаптен. В отношении антител подклассов IgG2a и IgG2b значительных различий не наблюдалось (табл.5). Группы, для которых IgG2a- и IgG2b-антитела не могут быть обнаружены, является лишь группа мышей, иммунизированных EYPC-липосомами, содержащими 1 мкг HA2 и 10 мкг DNP-CapPE (IgG2a и IgG2b) и 3 мкг и 10 мкг DNP-CapPE (IgG2a). Для более высокой дозы DNP-CapPE или HA2 значительных различий в ответах не наблюдалось. Эти ответы были сравнимы с ответами, продуцированными мышами, которых иммунизировали DNP-BSA. В отношении антител подкласса IgG3 значительного различия не наблюдали. Для каждой группы, титр антител обнаруживали по крайней мере 4 из 6 иммунизированных мышей (табл.5). Однако только 2 мыши, иммунизированные 10 мкг DNP-BSA, давали нарастающий титр значительно ниже, чем группа мышей, иммунизированных липосомными препаратами.
Выше было представлено полное описание изобретения, однако каждому специалисту очевидно, что при этом возможным некоторые изменения и модификации, не вытекающие за рамки существа и объема изобретения.
Использование: изобретение относятся к медицине, а именно к иммунологии и касается способа усиления иммунного ответа на "слабый" антиген посредством его введения в липосому вместе с дополнительным компонентом, имеющим сайт узнавания Т-лимфоцита хелпера. Сущность изобретения заключается в создании липосомы, включающей в себя липоды широкого ряда с инкапсулированным, ДНР-Сар РЕ антигеном, который является независимым антигеном и хелперным пептидом, представляющим собой легкую цепь гемагглютипина вируса гриппа. Технический результат заключается в усилении иммунного ответа на "слабый" антиген. 1 с. и 5 з.п. ф-лы, 5 табл.
| Uichida T et al., Cell.Biol., v.80, pp.10-20, 1979. |

