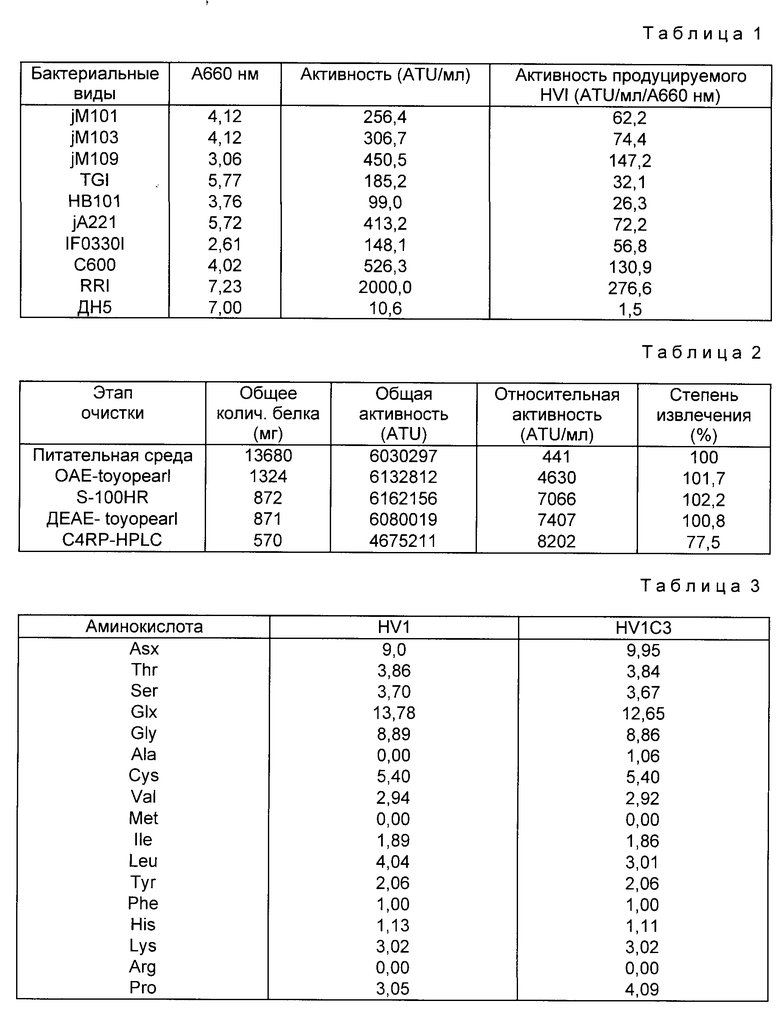
Настоящее изобретение касается нового гирудинового аналога, последовательности ДНК, кодирующей данный гирудиновый аналог, способа получения данного гирудинового аналога с использованием последовательности ДНК и антикоагулянта.
Настоящее изобретение касается также вектора секреции для продуцирования инородного белка, микроорганизмов, трансформированных данным вектором, и способа получения гирудина или его аналогов с использованием указанного трансформированного микроорганизма.
Новый гирудиновый аналог, описанный в данном изобретении, используется как антикоагулянт благодаря его более высокой антитромбиновой активности, а также более низкой тенденции к кровоизлиянию по сравнению с известным гирудином HVI.
Кроме того, вектор секреции инородного белка, такого как гирудиновый аналог, описанный в данном изобретении, дает возможность промышленным путем продуцировать инородный белок, такой как гирудиновый аналог, поскольку он обеспечивает высокий выход продуцируемого внеклеточным путем инородного белка при использовании E.coli, трансформированной указанным вектором.
Предпосылки для создания изобретения
Гирудин является антикоагуляционным фактором, секретируется из слюнных желез медицинской пиявки (Herudo medicinalis) и представляет собой смесь пептидов, состоящих из 65 - 66 аминокислот. Структура гирудина определена Dodt и др. (FEBS Lett, 165, 180, 1984) как разновидность гирудина 1 (HVI). Другая разновидность гирудина HV2 (Hervey и др. Proc Natl Acad Sci, США, 83, 1084, 1986) имеет девять различий в аминокислотной последовательности по сравнению с гирудином HVI, а еще одна разновидность гирудина HV3 (Dodt и др. Bid.Chem. Heppe Sey ler 367, 803, 1986) имеет десять различий в аминокислотной последовательности по сравнению с HVI и одинаковую последовательность с HV2 вплоть до Ser32 и дополнительную аминокислоту (Ala63) в C-концевой зоне. Структура этих трех гирудиновых разновидностей показана на фиг.1.
Эти варианты естественного типа включают 65 или 66 аминокислот и имеют два домена, которые являются сферической структурированной N-терминальной зоной с тремя дисульфидными связями и кислотной C-терминальной зоной, проявляющей гомологию с зоной расщепления тромбина протромбиновой молекулы или с зоной расщепления фибриногена.
При сравнении C-терминальной аминокислотной последовательности разновидностей HV1 и HV3 естественного гирудина авторы данного изобретения обнаружили, что Leu64 - Gln65 гирудина HV1 заменены Asp65 - Glu66 в HV3. Подана также патентная заявка (открытый для ознакомления японский патент 3-164184, 1991), касающаяся синтеза синтетического гена HV1 и HV3 и экспрессии их в E. coli.
Названные разновидности гирудина HV1, HV2 и HV3, имеющие антитромботическую активность, не могут быть лекарственными препаратами, пригодными для клинического использования, ввиду серьезных побочных эффектов, таких как увеличение продолжительности кровоизлияния.
С другой стороны, были предложены различные системы для продуцирования гирудина методом генной инженерии, однако не был разработан отвечающий требованиям метод. В частности, с промышленной точки зрения, желательна система внеклеточного продуцирования, поскольку, если продуцируемый белок может быть секретирован внеклеточно, то достигается не только упрощение разделения и очистки данного продукта ввиду его присутствия в активной форме, но и защита продукта от переваривания бактериальной протеазой внутри клетки.
Для продуцирования гирудина путем секреции были предложены способы, в которых как хозяева использовались Bacillus subtiles, дрожжи или E.coli.
Способы с использованием Bacillus subtilis в качестве хозяев имеют недостатки. Так, например, то, что плазмиды обычно нестойки в бактериях, в результате приводит к частому удалению их из клеток и затрудняет стабильное продуцирование; или то, что продукты в питательной среде иногда перевариваются бактериальными эндогенными секретируемыми протеазами. Способы, предложенные для продуцирования гирудина (например согласно выложенному для ознакомления японскому патенту 2-35084, 1990), пока еще не разрешили этих проблем, и выход продукта составляет лишь примерно 100 мг/л.
Известно, что в способах, осуществляемых с использованием дрожжей в качестве хозяев, C-терминальные аминокислоты данных продуктов обычно гидролизуются карбоксипептидазой.
В опубликованном ранее отчете (N.Riehi-Bellon и др. Biochemistry 1989, т. 28, 2941 - 2949) описывается получение побочных продуктов, в которых 1 или 2 аминокислотные группы отщеплены от C-конца HV2.
Для разрешения данной проблемы был предложен способ, в котором в качестве хозяина использовался дрожжевой штамм с дефицитом карбоксипептидазы, однако этот способ не обеспечил достаточной продуктивности.
В качестве способа с использованием E.coli как хозяина был предложен способ, использующий сигнальную последовательность щелочной фосфатазы (J.Dodt и др. FEBS Lett 202, 373/377, 1986). Хотя это секреционная система, продукт секретируется главным образом в периплазматическое пространство, что не разрешает задачу, поскольку требуется разрыв бактериальной клетки за счет ведения дополнительного процесса, такого как осмотический шок, и выход продуцируемого продукта составляет лишь 4 мг/л.
Описание изобретения
Авторы данного изобретения получали различные гирудиновые аналоги, в основе которых лежит исходная структура гирудина HV1, HV2 и HV3, и установили, что гирудиновый аналог (химерный гирудин), имеющий аминокислотную последовательность гирудина HV1, у которого аминокислотная последовательность после 53-й аминокислотной группы была заменена аминокислотной последовательностью HV3 после 53-й аминокислотной группы, проявляет не только высокую антитромбиновую активность, но и пониженный эффект продления времени кровотечения. На основании этого открытия было сделано изобретение.
Кроме того, авторы данного изобретения основательно изучили способ внеклеточного продуцирования инородных белков с высоким выходом с использованием E.coli в качестве хозяина и установили, что в случае, когда в качестве источника репликации вектора секреции с последовательностью ДНК, сигнального пептида и промотора используется источник репликации (ori) плазмиды pUC, то инородный белок секретируется из E.coli в больших количествах.
Таким образом, данное изобретение касается нового гирудинового аналога, имеющего высокую антитромбиновую активность.
Далее, настоящее изобретение касается последовательности ДНК, кодирующей аминокислотную последовательность гирудинового аналога, имеющего указанную высокую антитромбиновую активность и низкую тенденцию к кровоистечению, трансформированных микроорганизмов, получаемых путем трансформации E. coli экспрессионными векторами, включающими указанную последовательность ДНК, и способа получения гирудиновых аналогов путем экспрессии гирудиновых аналогов с использованием указанных выше трансформированных микроорганизмов и их потомков.
Далее, настоящее изобретение касается антикоагуляционного лекарственного препарата, заключающего в себе указанный гирудиновый аналог в качестве активного ингредиента.
Этот новый гирудиновый аналог HVIC3, отвечающий данному изобретению, имеет аминокислотную последовательность, показанную ниже.

Последовательность ДНК, кодирующая аминокислотную последовательность данного аналога, может быть представлена следующей формулой
Гирудиновый аналог, отвечающий данному изобретению, представленный формулой (I), может быть синтезирован способом химического синтеза или может быть продуцирован способом генной инженерии.
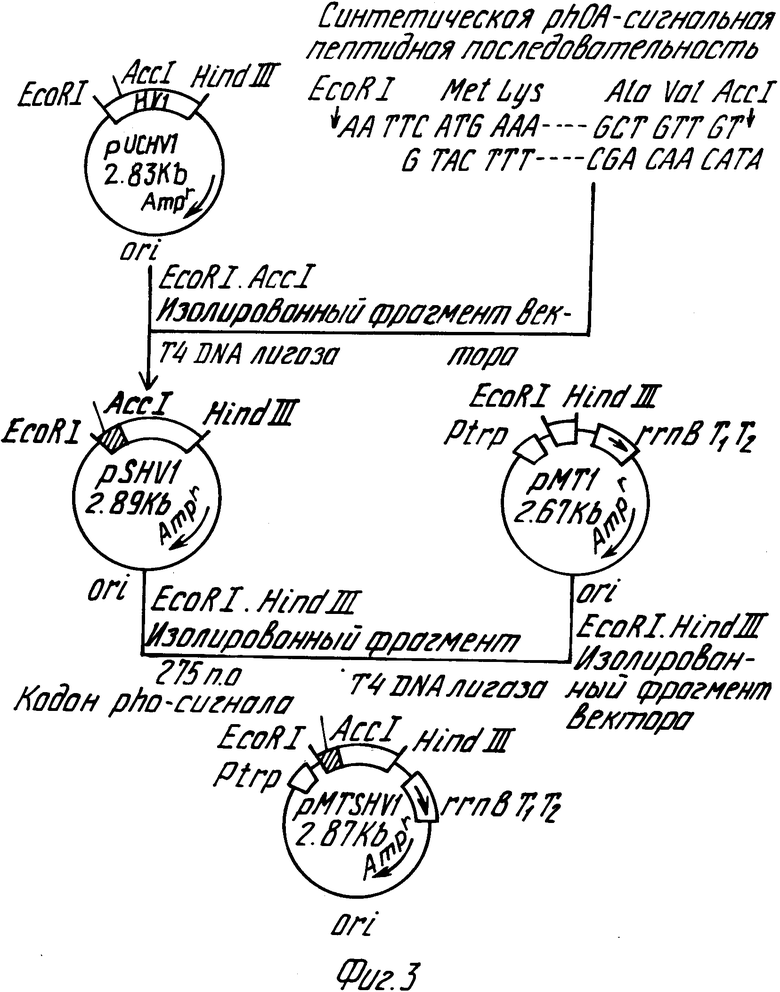
Для получения способом генной инженерии прежде всего, как указано в последнем примере, были построены секретирующие гирудин HV1 плазмиды pMTs HV1 и pMKSHV1, ими был трансформирован микроорганизм E.coli и был продуцирован HV1 путем секреции с использованием данного трансформированного микроорганизма. Плазмида pMTS HV1 включает промотор (Ptrp), последовательность ДНК, кодирующую сигнальный пептид (PhoA сигнал), последовательность ДНК, кодирующую аминокислотную последовательность гирудина HV1, источник репликации (ori) и последовательность ДНК, включающую трансляционный терминирующий сигнал (rm BT1T2), как показано на фиг.3. Согласно данному изобретению для замещения аминокислотной последовательности гирудина HV1, начиная с 53-й аминокислотной группы, аминокислотной последовательностью гирудина HV3, начинающийся с его 53-й аминокислотной группы, секретирующая HV1 плазмида pMTSHV1 была расщеплена для удаления последовательности ДНК, кодирующей аминокислотную последовательность, начинающуюся с 53-й аминокислотной группы гирудина HV1. С другой стороны, с использованием фермента рестрикции была изолирована последовательность ДНК, кодирующая аминокислотную последовательность, начинающуюся с 53-й аминокислотной группы экспрессионной плазмиды pUCHV3 гирудина HV3. Согласно данному изобретению ДНК с удаленной последовательностью, кодирующей полипептид, начинающийся с 53-й аминокислотной группы гирудина HV1, полученная из плазмиды pMTSHV1, и ДНК, кодирующая аминокислотную последовательность, начинающуюся с 53-й аминокислотной группы гирудина HV3, полученная из pUCHV3, сшивались ДНК-лигазой, в результате чего была построена плазмида pMTSHVIC3, заключающая в себе ДНК, кодирующую аминокислотную последовательность гирудинового аналога, отвечающего данному изобретению.
Плазмида pMTSHVIC3 включает происходящую из плазмиды pMTSHVI последовательность ДНК, кодирующую промотор, сигнальный пептид и источник репликации (ori), последовательность ДНК, кодирующую аминокислотную последовательность, начиная с 1-й по 52-ю аминокислотную группу гирудина HVI, и трансляционный терминирующий сигнал, а также последовательность ДНК, кодирующую аминокислотную последовательность, начиная с 53-й аминокислотной группы гирудина HV3, происходящую из плазмиды pUCHV3, которая вставляется между указанной последовательностью ДНК, кодирующей аминокислотную последовательность от 1-й до 53-й аминокислотной группы гирудина HVI, и трансляционным терминирующим сигналом.
Гирудиновый аналог, отвечающий данному изобретению, продуцируется в клетке и в культуральной среде после введения плазмиды pMTSHVIC3 в E.coli для трансформации и последующего культивирования данного трансформированного микроорганизма.
Согласно способу данного изобретения гирудиновый аналог отделяется общеизвестным способом и очищается такими способами как обращенно-фазовая жидкостная хроматография высокого разрешения или хроматография в колонке.
Полученный гирудиновый аналог проявляет более высокую антитромбиновую активность и тенденцию к более низкому кровотечению по сравнению с гирудином HVI при испытании на животных. Он может быть использован как прекрасное антикоагуляционное лекарство путем первоначального приготовления рецептуры общеизвестными способами. Иными словами, гирудиновый аналог, отвечающий данному изобретению, может быть далее приготовлен в форме рецептуры любыми общеизвестными способами с использованием носителей, которые обычно применяются для фармацевтических препаратов, или присадок для фармацевтических препаратов. Ввод таких препаратов в организм может осуществляться внутривенно, внутрикожно, подкожно или внутримышечно, или локально, но не орально. Хотя точное количество препарата для каждого случая определяется такими факторами, как симптомы заболевания, возраст и пол пациента, обычно это количество составляет от 0,1 до 100 мг в день для взрослого пациента, и это общее количество препарата должно вводиться в организм 1 - 7 раз.
Кроме того, как уже отмечалось ранее, настоящее изобретение охватывает вектор секреции для продуцирования инородного белка, в частности вектор секреции для внеклеточного продуцирования с высоким выходом гирудина или его аналогов.
Настоящее изобретение касается также вектора секреции, заключающего в себе последовательность ДНК, кодирующую гирудин или его аналоги, микроорганизма (E. coli), трансформированного данным секреторным вектором, и способа получения гирудина или его аналогов путем культивирования указанного трансформированного микроорганизма и извлечения продукта из культуральной среды.
Вектор секреции инородного белка, отвечающий данному изобретению, включает ДНК последовательность источника репликации (ori), происходящего из плазмиды pUC, ДНК последовательность промотора tac или промотора trp, ДНК последовательность, кодирующую сигнальный пептид, и ДНК последовательность, кодирующую инородный белок.
ДНК фрагмент, заключающий в себе источник репликации (ori) плазмиды pUC, отвечающей данному изобретению, может быть получен путем расщепления коммерчески доступных плазмид семейства pUC, например pUC9, pUC18 или pUC19, ферментами рестрикции в соответствующей комбинации. Так, например, фрагмент ДНК примерно 1440 пар оснований (bp), включающий источник репликации (ori) и полученный путем расщепления pUC18 ферментами рестрикции PVUI или PVUI и PVU II, может быть использован как источник репликации плазмиды pUC.
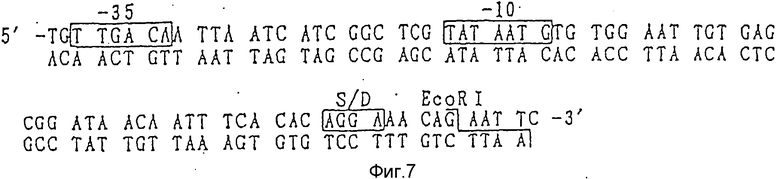
Фрагмент промотора tac или просмотра trp согласно данному изобретению имеет нуклеотидные последовательности, показанные на фиг.7 и 8 соответственно, и они могут быть легко синтезированы посредством ДНК синтезатора и др. Далее, данные синтезированные промоторы могут быть сшиты с фрагментами, содержащими источник репликации (ori) плазмиды pUC18. Как другой возможный вариант, они могут быть относительно легко получены путем сшивки фрагментов ДНК, получаемых путем расщепления коммерчески доступных плазмид ферментами рестрикции. Так, например, плазмида pMK2 может быть получена путем сшивки с использованием T4 ДНК лигазы фрагментов ДНК, включающих ДНК последовательность промотора, полученного путем расщепления коммерчески доступной плазмиды pKK223-3 (Pharmacia) ферментом рестрикции pVUI и NruI, и фрагмента ДНК, включающего источник репликации (ori), полученного расщеплением коммерчески доступной плазмиды pUC18 ферментами рестрикции PVUI и PVUII.
С другой стороны, плазмида PMTI может быть получена после удаления фрагмента, включающего последовательность ДНК промотора tac, путем расщепления указанной выше плазмиды pMK2 ферментами рестрикции EcoRI и Eco47III и последующей вставки фрагмента, включающего последовательность промотора trp, которая синтезирована посредством ДНК синтезатора.
Для последовательности ДНК сигнального пептида согласно данному изобретению может быть использована последовательность ДНК, кодирующая сигнальный пептид белков, локализирующихся в периплазме E.coli, например ферментов, таких как щелочная фосфатаза (phoA) и β - лактамаза (bla), или белков наружной мембраны, таких как OmpA, OmpB и OmpF. Эти ДНК фрагменты могут быть легко получены с использованием ДНК синтезатора.
Инородным белком может быть любой белок, хотя, как описано в данном изобретении, наиболее подходящими являются гирудин и его варианты. Аминокислотная последовательность этих белков показана на фиг. 1 как гирудины HVI, HV2, HV3 и HVIC3.
Фиг.1 иллюстрирует аминокислотную последовательность гирудинов HVI, HV2, HV3, HVIC3.
Фиг.2 иллюстрирует ДНК последовательность сигнального пептида phoA.
Фиг.3 дает общую схему, иллюстрирующую построение плазмиды pMTSHVI, секретирующей гирудин HVI.
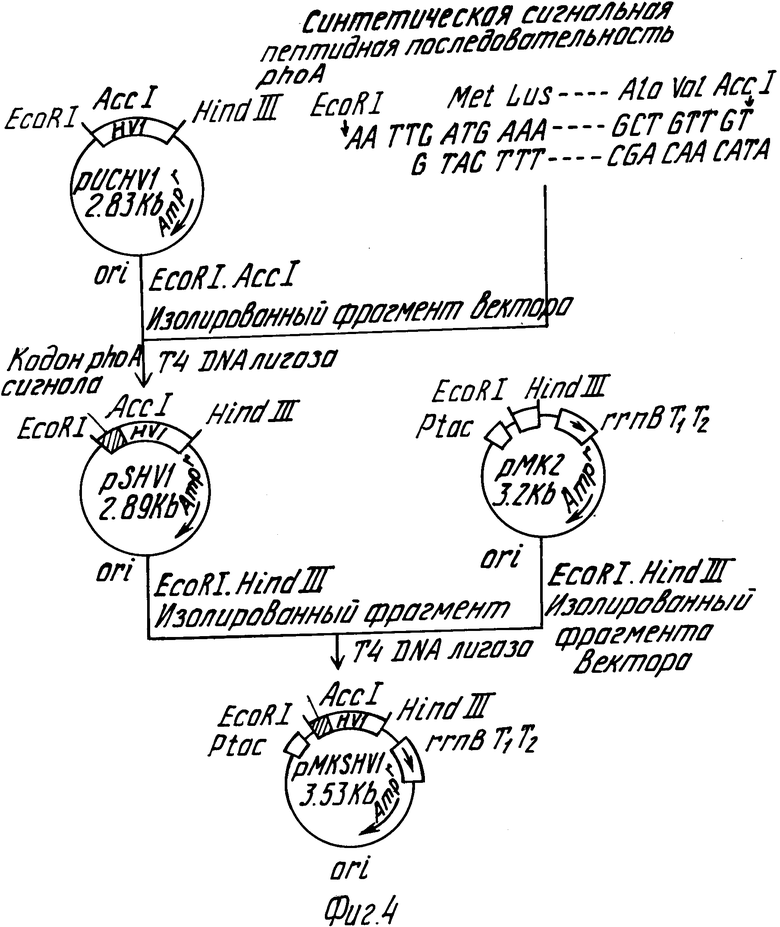
Фиг.4 дает общую схему, иллюстрирующую построение плазмиды pMKSHVI.
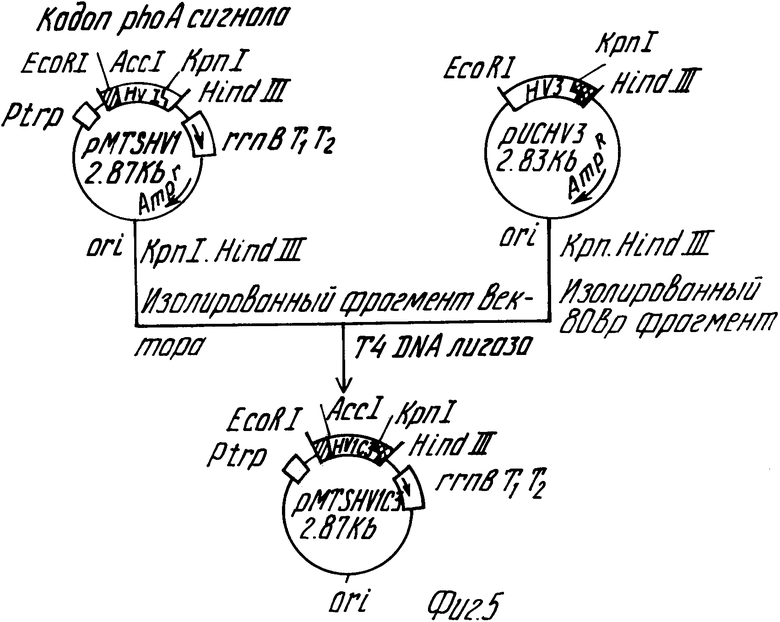
Фиг. 5 дает общую схему, иллюстрирующую построение плазмиды pMTSHVIC3, секретирующей гирудин HVIC3.
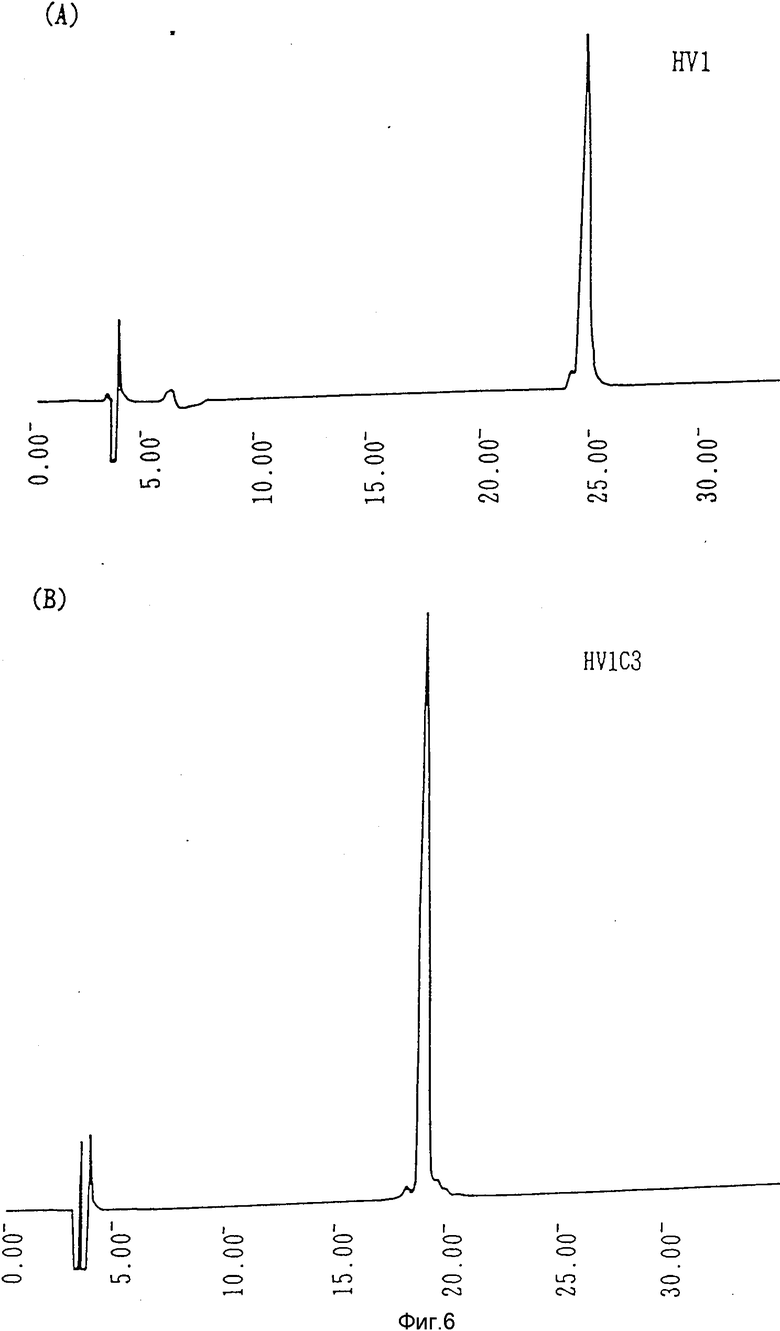
Фиг.6 (A) - кривые C4-обращенно-фазовой жидкостной хроматографии высокого разрешения гирудина HVI; (B) - кривые C4-обращенно-фазовой жидкостной хроматографии высокого разрешения соответственно гирудина HVIC3.
Фиг. 7 - ДНК последовательность промотора tac, используемого согласно данному изобретению.
Фиг. 8 - последовательность ДНК промотора trp, используемого согласно данному изобретению.
Фиг. 9 дает общую схему, иллюстрирующую построение плазмиды pMTS HV3, секретирующей гирудин HV3.
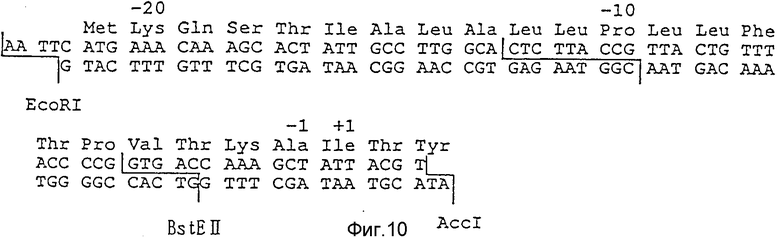
Фиг. 10 иллюстрирует ДНК последовательность сигнального пептида phoA для секреции гирудина HV3, используемую в примере 5.
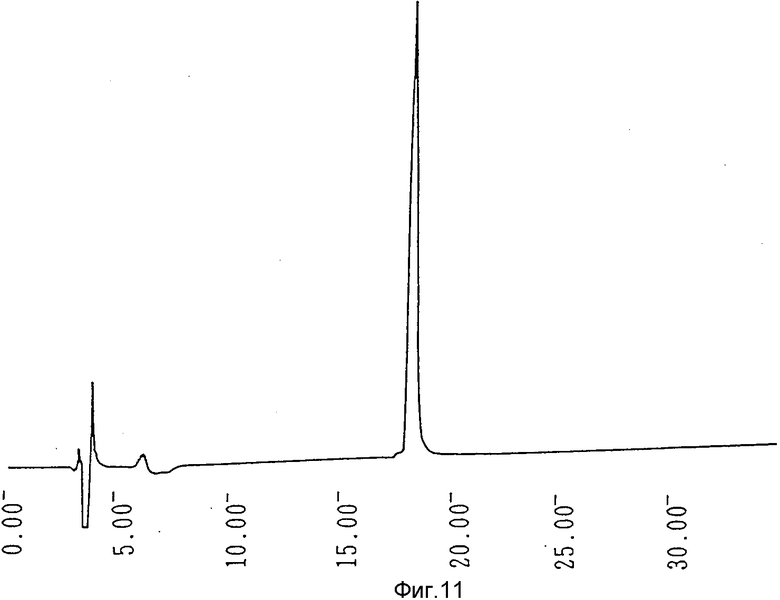
Фиг. 11 иллюстрирует кривую обращенно-фазовой (С4)жидкостной хроматографии очищенного гирудина HV3.
Наилучший вариант способа осуществления данного изобретения
В нижеследующих сравнительных примерах описывается процедура построения плазмиды pUCHVI, pMTI и pMK2 для построения pMTIS HVI как плазмиды секреции HVI, используемой в данном изобретении, и плазмиды pUCHV3 для построения плазмиды pMTSH VIC3.
Сравнительный пример 1
получение плазмиды pUCHVI и плазмиды pUCHV3
10 мг коммерчески доступной плазмиды pUC18 переваривалось 30 ед. Eco RI и 30 ед. HindIII при температуре 37oC в течение 2 ч. Затем векторная часть отделялась и экстрагировалась путем электрофореза на агарозном геле, белки удалялись путем экстракции фенолом, осаждались холодным этанолом и растворялись в 50 мкл буферного раствора TE (10 мM трис-HCI, pH = 8,0, 1 мМ этилендиаминтетрауксусной кислоты). К указанному количеству раствора, содержащего 50 нг ДНК, добавляли 10 мкМ раствора (66 мМ трис-HCl, pH = 7,5, 5 мМ MgCl2, 5 мМ ДТТ, 1 мМ АТР и 300 ед. Т4 ДНК лигазы), содержащего двухнитевую ДНК для гирудина HVI или HV3, с последующей реакцией в течение ночи при температуре 16oC, в результате чего получалась плазмида pUCHVI или pUCHV3, в которые был вставлен соответственно ген HVI или ген HV3 между Eco RI и Hind III сайтом плазмиды pUC18.
Сравнительный пример 2
Построение плазмиды pМК2 и плазмиды pMTI
(а) Получение плазмиды pMK2
Осуществлялась сшивка фрагмента, содержащего промотор tac, который был получен путем расщепления промышленно доступной плазмиды pKK223-3 (Pharmacia) ферментами рестрикции PVUI и NruI, фрагмента, содержащего источник репликации (ori), полученного путем расщепления промышленно доступного pUC18 ферментами рестрикции PVUI и PVUII, и фрагмента, содержащего ген устойчивости к ампициллину с использованием Т4 ДНК лигазы. Полученный таким путем фрагмент вставлялся в штамм E.coli jM109 с последующим культивированием, отбором по устойчивости к ампициллину, и выделялся вектор, заключающий в себе как промотор, так и источник репликации (ori) плазмиды pUC18; и этот вектор был назван pMK2.
(б) Получение плазмиды pMTI
10 мкг плазмиды pMK2 переваривались 30 ед. EcoRI и Eco47III, затем удаляли фрагмент, содержащий промотор tac, и выделяли фрагмент, содержащий источник репликации (ori) путем электрофореза на агарозном геле. Одновременно с этим был синтезирован фрагмент ДНК, содержащий промотор trp, с использованием синтезатора ДНК. Этот фрагмент был сшит с указанным выше фрагментом, полученным путем переваривания pMK2 EcoRI и Eco47III, с использованием Т4 ДНК лигазы при температуре 16oC в течение ночи. Он был использован для трансформации E. coli jM109 получения вектора, включающего как промотор trp, так и источник репликации (ori) плазмиды pMK2. Последовательность ДНК данной плазмиды была подтверждена методом Sanger и др., и эта плазмида была названа pMTI.
Настоящее изобретение более подробно иллюстрируется нижеследующими примерами.
Пример 1
(1) Построение плазмиды pMT SHVI, секретирующей гирудин HVI
Была построена плазмида pMTS HVI способом, проиллюстрированным на фиг. 3. Были синтезированы четыре олигонуклеотида, показанных на фиг. 2, для построения фрагмента ДНК, соответствующего сигнальному пептиду щелочной фосфатазы (phoA) E.coli, и фрагмента ДНК, кодирующего Val1 Val2 группы N-терминальной аминокислоты гирудина HVI. После снятия защиты каждый нуклеотид был очищен путем электрофореза на 10% полиакриламидном геле.
После того, как было фосфорилировано 500 пмолей каждого из двух олигонуклеотидов (S2, S4), 20 пмолей каждого из 4 олигонуклеотидов смешивались, гибридизировались и затем обрабатывались при 16oC в течение ночи в 20 мкл раствора, содержащего Т4 ДНК лигазу. После удаления белка фенолом и хлороформом и осаждения холодным этанолом был получен желаемый фрагмент двухнитевой ДНК. 1/10 количества полученного таким путем фрагмента и 100 нг плазмиды pUCHVI (выложенное для общего ознакомления описание японского патента 3-164184, 1991), расщепленной ферментами рестрикции EcoRI и AccI, сшивали Т4 ДНК лигазой при 16oC в течение ночи. Была получена гибридная плазмида PSHVI, содержащая ген слияния, в котором последовательность ДНК, кодирующая гирудин HVI, следовала сразу же после последовательности, кодирующей сигнальный пептид phoA, после трансформации E.coli jM109. Данная последовательность ДНК этой плазмиды PSHVI была подтверждена путем осуществления способа Sanger и др.
10 мкг pSHVI переваривалось ферментами рестрикции EcoRI (30 ед) и HindIII (30 ед), и фрагмент слитого гена размером 276 пар оснований (bp) был очищен путем электрофореза на агарозном геле.
100 нг полученного таким путем фрагмента ДНК и 100 нг фрагмента ДНК, полученного путем переваривания вектора экспрессии E.coli pMTI (выложенная для ознакомления публикация японского патента 3-76579, 1991) ферментами рестрикции EcoRI и Hind III с последующей очисткой путем электрофореза на агарозном геле, сшивались с использованием Т4 ДНК лигазы при температуре 16oC в течение ночи. Полученная реакционная смесь использовалась для трансформации E. coli jM109 для получения плазмиды pMTS HVI, секретирующей HVI, в которой ген слияния, кодирующий сигнальный пептид phoA и гирудин HVI, следует за участком промотора trp. Эта последовательность ДНК плазмиды pMTS HVI была подтверждена путем осуществления способа Sanger и др.
(2) Построение плазмиды, pMKSHVI, секретирующей гирудин HVI
Плазмиды pMKSHVI была построена согласно способу, иллюстрированному на фиг. 4.
Указанная выше ДНК слияния, кодирующая сигнальный пептид phoA, и гирудин HVI, и фрагмент ДНК, полученный путем переваривания плазмиды экспрессии pMK2 (выложенная для ознакомления публикация японского патента 3-76579, 1991) ферментами рестрикции EcoRI и Hind III, сшивались с использованием Т4 ДНК лигазы, использовались для трансформации E.coli jM109, и была получена плазмида pMKSHVI, секретирующая гирудин HVI через ген слияния, вставленный в сайт за промотором tac. Данная последовательность ДНК плазмиды pMKSHVI была подтверждена посредством способа, описанного Sanger и др.
(3) Продуцирование путем секреции гирудина HVI с использованием pMTSHVI
E.coli jM109, трансформированная плазмидой pMTISHVI или pMKSHVI, построенными как указано выше в (1) и (2), была культивирована в среде 2 х УТ (16 г/л бактотриптона, 10 г/л бактодрожжевого экстракта и 5 г/л NaCl), содержащей 100 мкг/мл ампициллина. После культивирования при 37oC в течение 24 ч было собрано 1 мл среды культивирования.
Осажденные клетки каждого из образцов суспендировались в 1 мл 50 мМ трис-HCl (pH= 7,5) содержащего 25% сахарозы и 1 мМ ЭДТА, после чего осуществлялась обработка при комнатной температуре в течение 10 мин. После сбора клеток путем центрифугирования при 6000 х g в течение 10 мин клетки суспендировались в 1 мл холодной воды, в результате чего происходило выделение вещества в периплазматическое пространство клетки посредством осмотического шока. Клетки отделялись от периплазматической фракции путем центрифугирования при 6000 х g в течение 10 мин. Количество секретированного и накапливаемого в периполазме гирудина определяли путем измерения антитромбиновой активности в супернатанте от центрифугирования. Антитромбиновая активность определяется по количественному измерению интенсивности цвета, создаваемого гидролитической активностью тромбина на синтетическом хромогенном субстрате на хромозиме ТН (Тоцилглицил-пролиларгинин-4-нитроанилидацетат, Boeringer-Mannhaim), и ингибирующему действию гирудина на тромбин, которое подавляет проявление цвета.
Данная реакция осуществляется следующим образом. В 1 мл реакционной пробы, содержащий 0,36 ед. NIH человеческого тромбина (Сигма) и буферный раствор (100 мМ трис-HCl (pH = 8,5), 150 мМ NaCl и 0,1% полиэтиленгликоля-6000), вводят стандартный гирудин или неизвестный образец, и этот раствор предварительно инкубируется при 37oC в течение 3 мин.
Как субстрат, так и хромозим ТН добавляют до конечной концентрации 200 мкМ с замером выделения пара-нитроанилида по увеличению поглощения (при 450 нм) раствора в 1 мин и рассчитывают антитромбиновую активность (ATU).
В результате штамм, использующий плазмиду pMTIS HVI, проявляет антитромбиновую активность, равную 450 ATU на 1 мл среды разведения. А штамм, использующий плазмиду pMKS HVI, проявляет антитромбиновую активность 360 ATU на 1 мл среды разведения. В табл. 1 представлены результаты других исследований по продуцированию гирудина с использованием различных штаммов E.coli, таких как jM101, jM103, jM109, TGI, HB101, jA221, IF03301, C600, RRI и ДН5, в которые введена плазмида pMTS HVI способом трансформации согласно Hanahan и др.
В случае использования RRI в качестве хозяина происходит внеклеточное продуцирование примерно 2000 TU/мл.
(4) Секреция гирудина HVI в среду разведения трансформированный E.coli jM109/pMTSHVI
При трансформации штамма E. coli jM109 плазмидой pMTSHVI (E. coli jM109/pMTS HVI) (FERM ВР-3266) и культивировании в 2 л питательной среды 2 х УТ, содержащей 2% глюкозы, в 5-литровом ферментере с перемешиванием и аэрацией при 37oC в течение 24 ч продуцируется внеклеточно примерно 5300 ATU на 1 мл гирудина HVI.
(5) Очистка гирудина HVI от питательной среды
После ферментации собирается 1,5 л питательной среды и центрифугируется при 600 х g в течение 10 мин с отделением супернатанта от клеточных осколков. Поскольку концентрация соли в поверхностном слое от центрифугирования составляет 1,3% при измерении солевым измерителем, то этот супернатант четырехкратно разбавляется 100 мМ буферным раствором фосфата калия (pH = 7,0) и фильтруется через 3,2 мкм фильтр (Pole Co.LTd). Полученный фильтрат вводится в колонку с QAE-toyopeare (4,4 см х 7 см), уравновешенную 10 мМ буферным раствором фосфата калия (pH = 7,0). После загрузки образца колонка уравновешивается буферным раствором с последующим ступенчатым элюированием гирудина HVI 0,2 М NaCl. Элюированный раствор концентрируется посредством мембраны Diaflow (УМ5) Amicon, после чего осуществляется гель-фильтрация на Sephacryls-100R предварительно уравновешенным 10 мМ буферным раствором фосфата калия (pH = 7,0) для обессоливания.
Активные фракции вводятся в колонку с DEAE-toyopearl (4,4 см х 40 см), уравновешенную 10 мМ буферным раствором фосфата калия (pH = 7,0), тщательно промываются и затем элюируются линейным градиентом, создаваемым в пределах между 3 л уравновешивающего буфера и 3 л 0,3 М NaCl в уравновешивающем буфере. Окончательная очистка осуществляется посредством обращенно-фазной С4 жидкостной хроматографии высокого разрешения в колонке с Дельта-преп 3000, представляющим собой продукт Waters. Очищенный гирудин HVI получается путем элюирования его с хроматографической колонки линейным градиентом 15 - 30%-ного (об/об) ацетонитрила, содержащего 0,065% (об/об) трифторацетата.
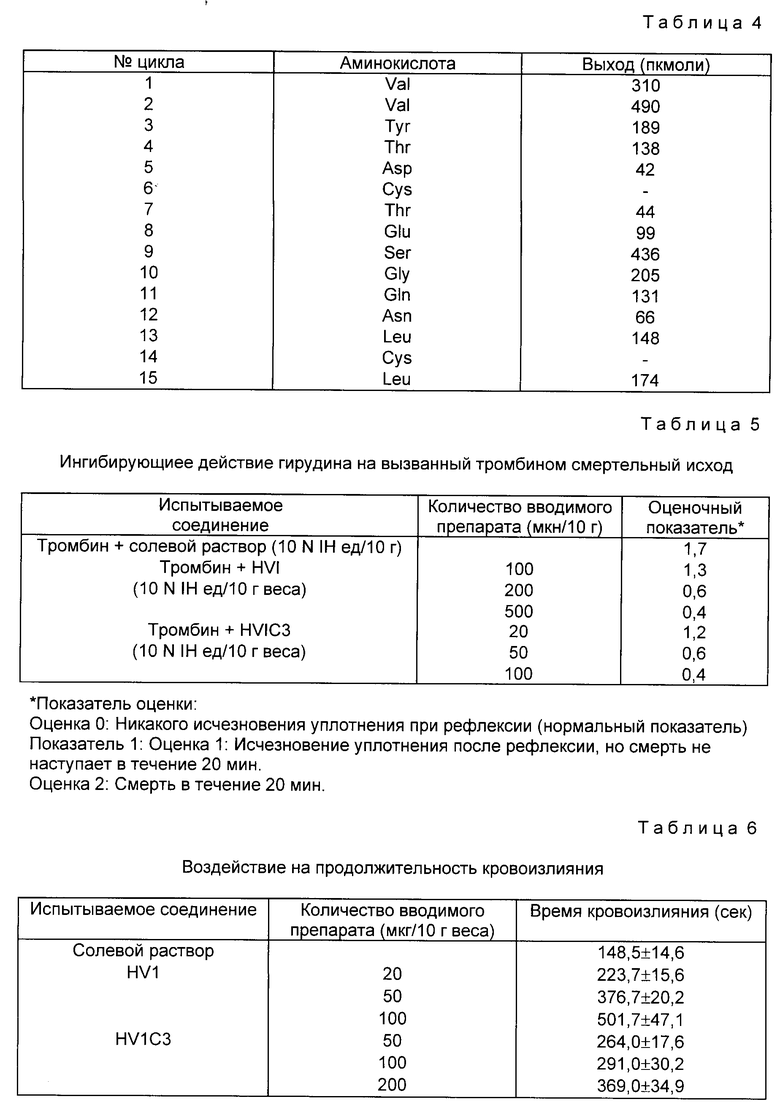
Степень очистки в каждом этапе приводится в табл. 2
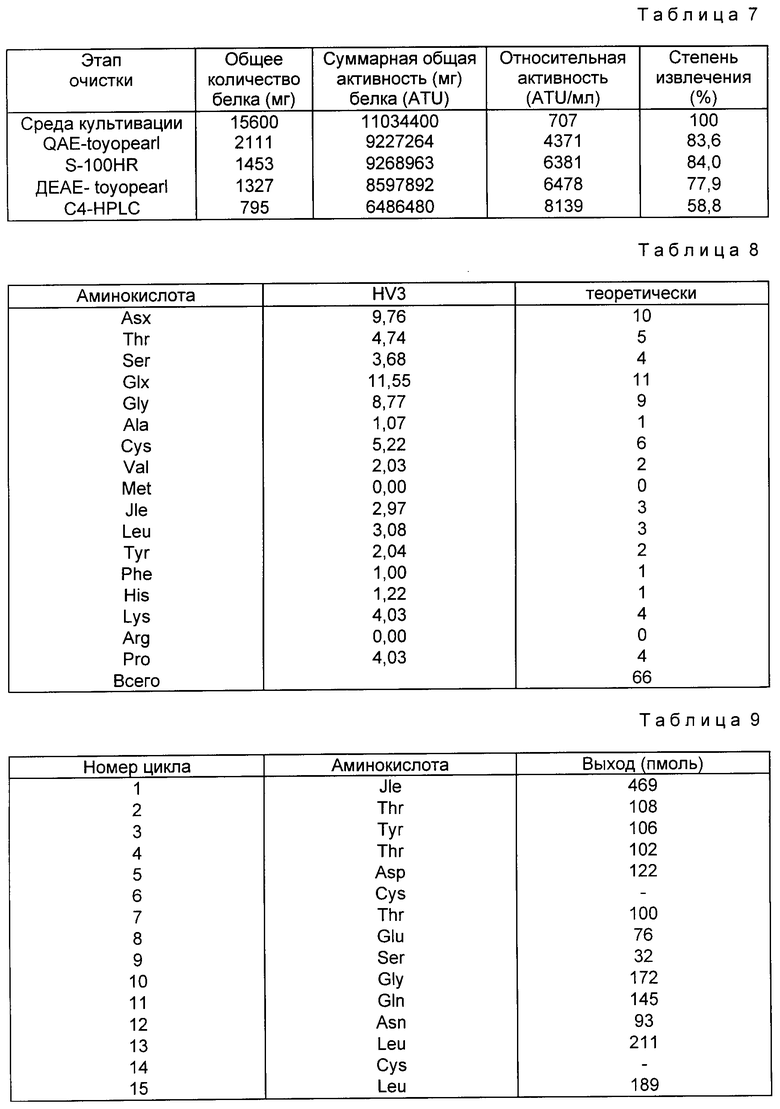
Аминокислотный состав полученного таким путем гирудина HVI является сходным с естественным гирудином HVI, как показано в табл. 3. N-терминальная аминокислотная последовательность очищенного гирудина HVI начинается с Val-Val-Tyr, как показано в табл. 4, что говорит о том, что происходит правильное отщепление сигнального пептида phoA. Антитромбиновая активность составляет 8202 ATU/мг.
(6) Получение плазмиды pMTSHVIC3, секретирующей химерный гирудин HVIC3
Плазмида pMTSHVIC3 была построена согласно способу, проиллюстрированому на фиг. 5. Для замены аминокислотных групп гирудина HVI, начиная с 53-й группы, последовательностью гирудина HV3 плазмида pMTSHVI, секретирующая HVI (10 мкг), была переварена ферментами рестрикции Kpn I (30 ед) и Hind III (30 ед) с последующим электрофорезом на агарозном геле для очистки фрагмента ДНК на 2,8 Квр.
Аналогичным образом, 10 мкг плазмиды pUCHV3 (выложенная для ознакомления публикация японского патента 3-164184, 1991) также переваривалось Kpn I и Hind III, и был очищен 80 bp-фрагмент ДНК, кодирующий С-терминальную аминокислотную последовательность гирудина HV3.
100 нг обоих фрагментов ДНК взаимодействовали в присутствии Т4 ДНК лигазы при температуре 16oC в течение ночи, и полученная реакционная смесь использовалась для трансформации E.coli jM109, в результате чего была получена плазмида pMTSHVIC3, секретирующая химерный гирудин HVIC3. Данная ДНК последовательность плазмиды pMTSHVIC3 была подтверждена с помощью способа Sanger и др.
(7) Продуцирование путем секреции химерного гирудина HVIC3 с использованием плазмиды, секретирующей гирудин
E. coli RRI (FERM ВР-3130), трансформированная плазмидой pMTSHVIC3, которая была построена, как описано выше (в разд. 6), культивировалась в питательной среде 2 х УТ, содержащей 100 мкг/мл ампициллина. После культивирования в течение 24 ч при температуре 37oC собирали 1 мл среды культивирования и осуществляли осмотический шок для измерения антитромбиновой активности периплазматической фракции.
В результате была выявлена активность антитромбина, равная 3060 ATU на 1 мл среды культивирования.
(8) Секреция химерного гирудина HV1C3 в ферментационную среду или в среду культивирования трансформированным штаммом E. coli jM109/pMTSH1C3
В случае культивации E. coli jM109 (FERM BP-3104), трансформированной плазмидой pMTSHVIC3, в 2 л среды культивирования 2 х УТ, содержащей 2% глюкозу, в 5-литровом ферментере с перемешиванием и эрацией при температуре 37oC в течение 24 ч, обнаруживалась общая антитромбиновая активность 60450 ATU/мл; 350 ATU/ мл в периплазме и 5700 ATU/мл в среде культивирования.
(9) Очистка химерного гуридина HV1C3
Гирудин HV1C3 был очищен из культуральной среды, полученной, как указано выше (в разд. 8), согласно способу, также описанному выше, в разд. (5). При определении аминокислотной последовательности очищенного химерного гирудина HV1C3 было подтверждено, что C-терминальная аминокислотная последовательность отлична от HV1, как иллюстрируется на фиг. 1. Удельная активность составляла 11 250 ATU/мг. Хроматограммы HPLC для очищенных HV1 и HV1C3 иллюстрируются на фиг. 6. Данный эксперимент осуществлялся в хроматографической колонне с VVDAC C4 (0,46 х 25 см) с линейным градиентом ацетонитрила от 15 до 30% при скорости потока 1 мл/мин в течение 30 мин.
Пример 2
Ингибирующее действие гирудина на смертный исход, вызванный тромбином
В организм мышей самцов (весом 20 - 25 г) без анестезии вводили внутривенно тромбин (10N 1H ед/10 г) и определяли антромбиновое действие испытываемого соединения по наблюдению за исчезновением уплотнения при рефлексии и смертельном исходе, что служило в качестве показателей этого действия. Результаты представлены в табл. 5.
Как ясно следует из табл. 5, в индуцированной тромбином смертельной реакции в условиях "ин виво" химерный гирудин (HV1C3), отвечающий данному изобретению, показывает примерно в 3 - 4 раза более сильную ингибирующую реакцию по сравнению с гирудином HV1.
Пример 3
Влияние на удлинение времени кровоизлияния
Образцы вводили путем инъекции в организм мышей самцов (весом 20 - 25 г) через хвостовую вену при анестезии пентобарбиталем натрия (40 мг/кг, внутрибрюшинно). Делали колотую рану путем вставки иглы 21G (наружным диаметром 0,85 мм) в другую сторону хвостовой вены через 5 мин после ввода испытываемого соединения и определяли продолжительность кровоизлияния из раны. На рану накладывали фильтровальную бумагу, которую меняли через каждые 15 с. Время кровоизлияния определялось как время, требуемое до тех пор, пока на фильтровальной бумаге не появлялось красное пятно. Результаты представлены в табл. 6.
Обычно продолжительность кровоистечения является одним из побочных эффектов антикоагулянтов. Точно подтверждено, что химерный гирудин (HV1C3), отвечающий данному изобретению, дает более низкую тенденцию к кровоистечению, чем гирудин HV1.
Пример 4.
Препарат, включающий в качестве ингредиента химерный гирудин HV1C3
Очищенный химерный гирудин HV1C3, отвечающий примеру 1-(9), обессоливался посредством Сафадекса G25 (фармацевтический продукт), после чего его фильтровали через фильтр (0,22 мкм) в стерильных условиях. Этот раствор разливался в пробирки и лиофилизировался. Полученный лиофилизированный порошок химерного гирудина HV1C3 мог быть растворен в солевом растворе и использован как инъекционно вводимое в организм лекарство.
Пример 5
Продуцирование гирудина HV3
(1) Построение плазмиды, секретирующей гирудин HV3
(I) Как показано на фиг. 9, источник репликации (ori), полученный путем переваривания плазмиды pUCHV3, полученной, как описано в сравнительном примере 1, ферментом рестрикции EcoRI и AccI, векторный фрагмент, включающий ген устойчивости к ампициллину, и синтезированный ген, кодирующий сигнальную последовательность щелочной фосфатазы (phoA), последовательность которой показана на фиг. 10, сшивались с использованием Т4 ДНК лигазы. В результате была получена плазмида pSHV3, к которой последовательность ДНК, кодирующая сигнальный пептид гена щелочной фосфатазы (phoA), была присоединена перед последовательностью ДНК, кодирующей HV3. Данная плазмида была введена в E. coli jM109, культивирована и отобрана по устойчивости к ампициллину.
(II) Далее, плазмида pHV3 была расщеплена ферментами рестрикции EcoRI и Hind III, в результате чего был получен фрагмент размером 275 bp, имеющий последовательность ДНК, кодирующую сигнальный пептид гена щелочной фосфатазы (phoA), присоединенный перед последовательностью ДНК, кодирующей гирудин HV3.
(III) Одновременно была получена плазмида pMTI способом, описанным в сравнительном примере 2. Данная плазмида заключает в себе последовательность ДНК, включающую источник репликации (ori) плазмиды pUC18 и промотор trp, как описано в сравнительном примере 2. Данный векторный фрагмент был получен путем расщепления плазмиды pMTI ферментами рестрикции EcoRi и Hind III.
(IV) Указанный выше фрагмент из 275 bp и векторный фрагмент сшивались Т4 ДНК лигазой, вводились в E. coli jM109, культивировались и отбирались на стойкость к ампициллину, и в результате получалась плазмида pMTSHV3 - вектор экспрессии HV3 из 2,87 kb, который включал последовательность ДНК с источником репликации (ori) плазмиды pUC, последовательность ДНК протомора trp, последовательность ДНК, кодирующую сигнальный пептид гена щелочной фосфотазы (phoA) и последовательность ДНК, кодирующую HV3.
(2) Продуцирование гирудина HV3 путем секреции с помощью плазмиды, секретирующей HV3.
E. coli RRI (E. coli RRI/pMTS HV3) (FERM BP-3267), трансформированная плазмидой pMTS HV3, которая была построена указанным выше способом, культивировалась в 2 л среды культивирования 2 х УТ, содержащей 100 мкг/мл ампициллина и 2% глюкозы. Культивирование осуществлялось в 2 л питательной среды в 5-литровом сосуде при 37oC в течение 24 ч. В результате обнаруживалась антитромбиновая активность 6073 ATU на 1 мл питательной среды.
(3) Очистка гирудина HV3 из среды культивирования
гирудин HV3 был получен из питательной среды способом, описанным в примере 1-(5). После центрифугирования среды культивирования для отделения поверхностного слоя от клеток супернатант разбавлялся четырехкратно 10 мМ буферным раствором фосфата калия (pH = 7,0) и фильтровался. Полученный фильтрат вводился в колонку (4,4 х 13 см) с QAE - toyopearl, как описано в примере 5, и затем осуществлялась гель-фильтрация на колонке с Sephacryls - 100HR.
Активность фракции вводилась в колонку с DEAE toyopear (4,4 х 40 см), уравновешенную буферным раствором, и затем элюировалась линейным градиентом от 0 до 0,3 М в уравновешивающем буферном растворе. И, наконец, гирудин HV3 очищался посредством обращенно-фазовой C4 жидкостной хроматографии высокого разрешения с использованием колонки Vydae C4 (4,7 х 30 см) с элюированием линейным градиентом (1% миним., 30 мин или более) ацетонитрила от 15 до 30% (об/об). Хроматограмма показана на фиг. 11. Степень очистки в каждом этапе указана в табл. 7.
При анализе аминокислотного состава и N-терминальной аминокислотной последовательности очищенного продукта аминокислотный состав был сопоставим с теоретическим составом гирудина HV3, как показано в табл. 8.
Как показано в табл. 9, этот состав вплоть до 15-й N-терминальной аминокислотной последовательности был идентичен гирудину HV3. Эти результаты показывают, что осуществлен правильный процессинг сигнального пептида, и полученным продуктом является гирудин HV3.
Промышленная применимость
Настоящее изобретение предлагает новый гирудиновый аналог. Гирудиновый аналог, отвечающий данному изобретению, используется как антикогаулянт, имеющий сильную антитромбиновую активность и более низкую тенденцию к кровоиспусканию.
Кроме того, настоящее изобретение предусматривает векторы секреции для секреции инородного белка, включая указанный выше гирудиновый аналог, трансформированные микроорганизмы и способ получения инородного белка с использованием указанных трансформированных микроорганизмов. При использовании способа, описанного в данном изобретении, инородные белки продуцируются внеклеточно с высоким выходом, что позволяет успешно получать инородные белки в промышленном масштабе.
Рекомендация микроорганизма:
(1) E. coli jM109/pMTS HV1C3
(Ссылка на организм E. coli jM109/pMTРНОН V1C3 была исправлена)
Дата депонирования:
18 сентября 1990 года
Регистрационный номер: FERM BP-3104
(2) E. coli RR1/pMTS HV1C3
Дата депонирования:
11 октября 1990 года
Регистрационный номер: FERM BP-3130
(3) E. coli jM109/pMTS HV1
Дата депонирования:
6 февраля 1991 года
Регистрационный номер: FERM BP-3266
(4) E. coli jRR1/pMTS HV3
Дата депонирования:
6 февраля 1991 года
Регистрационный номер: FERM BP-3267м



Изобретение может быть использовано в биотехнолоогии при получении аналога гирудина. Аналог гирудина, обладающий антикоагуляционной активностью, превышающей активность HVI, получают путем трасформации штамма E.coli рекомбинантным вектором секреции, содержащим фрагмент ДНК, кодирующий целевой полипептид, а также последовательность точки начала репликации, полученной из плазмиды pUC. Проводят культивирование упомянутого штамма и аналог гирудина извлекают из клеток или культуральной среды. 4 с.п. ф-лы, 11 ил., 9 табл.

2. Последовательность ДНК, кодирующая аналог гирудина по п.1.
PvuI - PvuII фрагмент pUC 18 размером 1140 bp;
tгр промотор;
фрагмент ДНК, кодирующий сигнальный пептид щелочной фосфатазы;
фрагмент ДНК, кодирующий аналог гирудина HV1C3 и имеющий последовательность
ген устойчивости к ампициллину.
| FEBS Lett, v | |||
| Приспособление к тростильной машине для прекращения намотки шпули | 1923 |
|
SU202A1 |

