Изобретение относится к вектору секреции для получения чужеродного белка, микроорганизмам, трансформированным с помощью данного вектора, и способу получения гирудина HV1 или HV3 при использовании указанных трансформированных микроорганизмов.
Вектор секреции чужеродного белка, такого как аналог гирудина, описанный в данном изобретении, позволяет промышленным путем получать данный белок, поскольку обеспечивает высокий внеклеточный выход гирудина при использовании микроорганизмов E.coli, трансформированных указанным вектором.
Гирудин - антикоагуляционный фактор, секретируемый слюнными железами медицинской пиявки (Hirudo medicinalis), который представлен смесью пептидов, состоящих из 65-66 аминокислот. Структура гирудина I (HV1) определена Dodt и др. (FEBS Lett, 165, 180, 1984). Другой вариант гирудина (HV2) (Harvey и др. , Proc. Natl. Acad Sei, США, 83, 1084, 1986) имеет девять различных аминокислотных последовательностей по сравнению с гирудином НV1, и еще один вариант гирудина (HV3) (Dodt и др., Biol. Chem. Hoppe-Seyler, 161, 803, 1986) имеет десять различных аминокислотных последовательностей по сравнению с HV1, чья аминокислотная последовательность совпадает с последовательностью HV2 вплоть до Ser32, и дополнительную аминокислоту (Ala63) на C-концевом участке. Структура аналогов гирудина приведена на фиг. 1. Известна патентная заявка (выложенная для ознакомления заявка на японский патент 3-164184, 1991), касающаяся получения синтетического гена HV1 и HV3 и экспрессии их в E.coli.
Кроме того, были предложены различные системы для получения гирудина методами генной инженерии, однако их нельзя считать удовлетворительными. В частности, с промышленной точки зрения желательно, чтобы белок секретировался из клетки, поскольку в этом случае упрощается разделение и очистка целевого продукта, и, кроме того, обеспечивается его защита от переваривания бактериальными протеазами внутри клетки.
Для получения секретируемого гирудина были предложены способы с использованием в качестве хозяев Bacillus subtilis, дрожжей или E.coli.
Способы, основанные на использовании в качестве хозяев Bacillus subtilis, имеют недостатки. Так например, плазмиды обычно нестойки в бактериях, что приводит к их частому удалению из клеток. Это затрудняет стабильную продукцию белка. Кроме того, содержащиеся в питательной среде продукты расщепляются эндогенными бактериальными протеазами. Способы, предложенные для получения гирудина (например согласно открытому для ознакомления японскому патенту 2-35084, 1990 г.), пока еще не разрешили этих проблем, и выход продукта составляет всего лишь около 100 мг/л.
Известно, что при использовании дрожжей в качестве хозяев C-терминальные аминокислоты целевых продуктов, как правило, гидролизуются карбоксипептидазой.
В опубликованном ранее сообщении (N.Riehi-Bellon и др., Biochemistry 1989, 28, 2941-2949) описывается получение побочных продуктов, содержащих 1 или 2 аминокислотных остатка отщепленных от C-терминального участка HV2.
Для решения данной проблемы был предложен способ, в котором в качестве хозяина использовался дрожжевой штамм с дефицитом карбоксипептидазы; однако этот способ не обеспечил достаточной продуктивности (выложенная для ознакомления заявка на японский патент 2-104279, 1990).
Известен способ с применением E.coli в качестве хозяина. При этом используется сигнальная последовательность щелочной фосфатазы (J.Dodt и др., FEBS Lett, 202, 373-377, 1986). Хотя это секреционная система, продукт высвобождается главным образом в периплазматическое пространство, что неудовлетворительно, поскольку необходим разрыв бактериальной клетки за счет дополнительного процесса, такого как осмотический шок, и выход целевого продукта составляет лишь 4 мг/л.
Изобретение относится к способу внеклеточного продуцирования чужеродных белков с высоким выходом при использовании Е. coli в качестве хозяина. Авторы изобретения установили, что в том случае, когда в качестве оргужина репликации вектора секреции, содержащего последовательности ДНК сигнального пептида, и промотора, используется оргужин (ori) плазмиды pUC, то чужеродный белок секретируется клетками E.coli в больших количествах.
Настоящее изобретение относится к трансформированным микроорганизмам, полученным путем трансформации E. coli векторами экспрессии, содержащими последовательность ДНК, аналога гирудина с высокой актитромбиновой активностью, и способу получения аналогов гирудина при использовании указанных выше трансформированных микроорганизмов. Для этого, как указано в последнем примере, были сконструированы секретирующие гирудин HV1 плазмиды pMTSHV1 и pMKSHV1, которыми трансформировали E.coli с последующим получением секретируемого HV1. Плазмида pMTSHV1 содержит промотор (Ptrp), последовательность ДНК, кодирующую сигнальный пептид (PhoA сигнал), последовательность ДНК, кодирующую аминокислотную последовательность гирудина HV1, оргужин репликации (ori) и последовательность ДНК, включающую терминационный сигнал трансляции (rrnBT1T2), как показано на фиг. 3. Согласно данному изобретению для замещения аминокислотной последовательности гирудина HV1 после 53-й аминокислотной группы, аминокислотной последовательностью гирудина HV3, начинающейся после 53-й аминокислотной группы, секретирующую HV1 плазмиду pMTSHV1 расщепляют ферментом рестрикции для удаления последовательности ДНК, кодирующей аминокислотную последовательность, начинающуюся после 53-й аминокислотной группы гирудина HV1. С другой стороны, с использованием фермента рестрикции из экспрессионной плазмиды pUCHV3 вырезают последовательность ДНК, кодирующую аминокислотную последовательность, начинающуюся после 53-й аминокислотной группы гирудина HV3. Согласно данному изобретению ДНК с удаленным участком, кодирующим последовательность, начинающуюся после 53-й аминокислотной группы гирудина HV1, полученную при расщеплении плазмиды pMTSHV1, и ДНК, кодирующую аминокислотную последовательность, начинающуюся после 53-й аминокислотной группы гирудина HV3, полученную при расщеплении плазмиды pUCHV3, сшивают ДНК лигазой с получением плазмиды pMTSHV1C3, содержащей ДНК, кодирующую аминокислотную последовательность аналога гирудина, отвечающего данному изобретению.
Плазмида pMTSHV1C3 содержит последовательность ДНК, кодирующую промотор, сигнальный пептид и оргужин репликации (ori), полученные из плазмиды pMTSHV1, последовательность ДНК, кодирующую с 1-й по 52-ю аминокислотные группы гирудина HV1, и терминационный сигнал трансляции, а также последовательность ДНК, кодирующую аминокислотную последовательность, начиная с 53-й аминокислотной группы гирудина HV3, полученную из плазмиды pUCHV3, которая вставляется между указанной последовательностью ДНК, кодирующей с 1-й по 53-ю аминокислоты гирудина HV1 и сигналом терминации трансляции.
Аналог гирудина образуется в клетке и поступает в культуральную среду при трансформации E.coli плазмидой pMTSHV1C3 с последующим культивированием трансформированного микроорганизма.
Согласно способу данного изобретения аналог гирудина выделяют общеизвестным способом и очищают такими методами, как обратно-фазовая жидкостная хроматография высокого разрешения или хроматография на колонке.
Полученный аналог гирудина проявляет более высокую антитромбиновую активность и эффективнее снижает кровоточивость, чем гирудин HV1, при испытании на животных.
Кроме того, как уже отмечалось ранее, настоящее изобретение относится к вектору секреции для получения чужеродного белка, в частности к вектору секреции для внеклеточного получения гирудина или его аналогов в больших количествах.
Настоящее изобретение относится также к вектору секреции, содержащему последовательность ДНК, кодирующую гирудин или его аналоги, микроорганизму (E. coli), трансформированному данным вектором секреции, и способу получения гирудина или его аналогов путем культивирования указанного трансформированного микроорганизма и выделения целевого продукта из культуральной среды.
Вектор секреции чужеродного белка, отвечающий данному изобретению, содержит последовательность ДНК, оргужина репликации (ori), полученного из плазмиды pUC, последовательность ДНК промотора tac или промотора trp, последовательность ДНК, кодирующую сигнальный пептид, и последовательность ДНК, кодирующую чужеродный белок.
Фрагмент ДНК, содержащий оргужин репликации (ori) плазмиды pUC, может быть получен путем расщепления коммерчески доступных плазмид семейства pUC, например рUС9, рUC18, или pUC19, ферментами рестрикции в соответствующей комбинации. Так например, фрагмент ДНК размером около 1440 пар оснований (вр), включая оргужин репликации (ori), полученный путем расщепления плазмиды pUC18 рестриктазами PvuI, или PvuI и PvuII, совместно может быть использован как оргужин репликации плазмиды pUC.
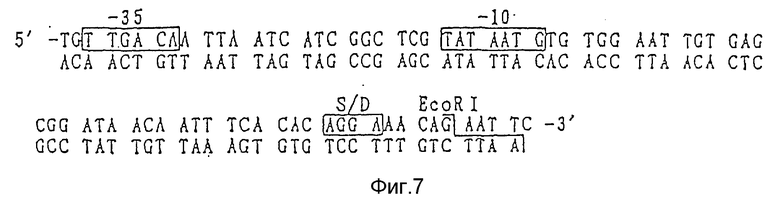
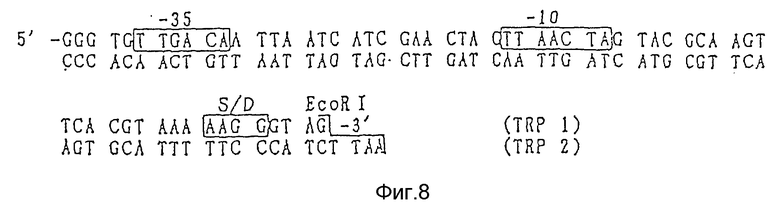
Фрагмент промоторов tac или trp имеет нуклеотидные последовательности, показанные на фиг. 7 и 8 соответственно, и они могут быть легко синтезированы с помощью ДНК синтезатора и другими методами. Таким образом, указанные промоторы могут быть синтезированы и соединены с фрагментами, содержащими оргужин репликации (ori) плазмиды pUC18. С другой стороны, они могут быть относительно легко получены путем легирования фрагментов ДНК, образованных при расщеплении коммерчески доступных плазмид ферментами рестрикции. Так например, плазмида рМК2 может быть получена с использованием Т4 ДНК лигазы путем соединения фрагментов ДНК, включающих ДНК последовательность промотора, полученных при расщеплении коммерчески доступной плазмиды рКК223-3 (Pharmacia) рестриктазами PvuI и NruI, и фрагмента ДНК, включающего оргужин репликации (ori), полученного путем расщепления коммерчески доступной плазмиды pUC18 рестриктазами PvuI и PvuII.
С другой стороны, плазмида pMT1 может быть получена при удалении фрагмента ДНК, соответствующего промотору tac, путем расщепления указанной выше плазмиды рМК2 ферментами рестрикции EcoRI и Есо47III, и последующей вставке фрагмента, соответствующего промотору trp, синтезированного с помощью ДНК-синтезатора.
В качестве последовательности ДНК сигнального пептида может быть использована последовательность ДНК, кодирующая сигнальный пептид белков, локализирующихся в периплазме E.coli, например ферментов, таких как щелочная фосфатаза (phoА) и B-лактамаза (bla) или белков наружной мембраны. Например, OmpA, OmpB и OmpF. Указанные фрагменты ДНК могут быть легко получены с помощью ДНК-синтезатора.
Хотя в настоящем изобретении любой белок может считаться чужеродным, наиболее подходящими являются гирудин и его варианты. Аминокислотная последовательность этих белков приведена на фиг. 1 (гирудины HV1, HV2, HV3 и HV1C3).
Краткое описание чертежей.
Фиг. 1 иллюстрирует аминокислотную последовательность гирудинов HV1, HV2, HV3, HV1C3.
Фиг. 2 иллюстрирует ДНК последовательность сигнального пептида phoA.
Фиг. 3 дает общую схему построения плазмиды pMTSHV1, секретирующей гирудин HV1.
Фиг. 4 дает общую схему построения плазмиды pMKSHV1.
Фиг. 5 дает общую схему построения плазмиды pMTSHV1CS, секретирующей гирудин HV1C3.
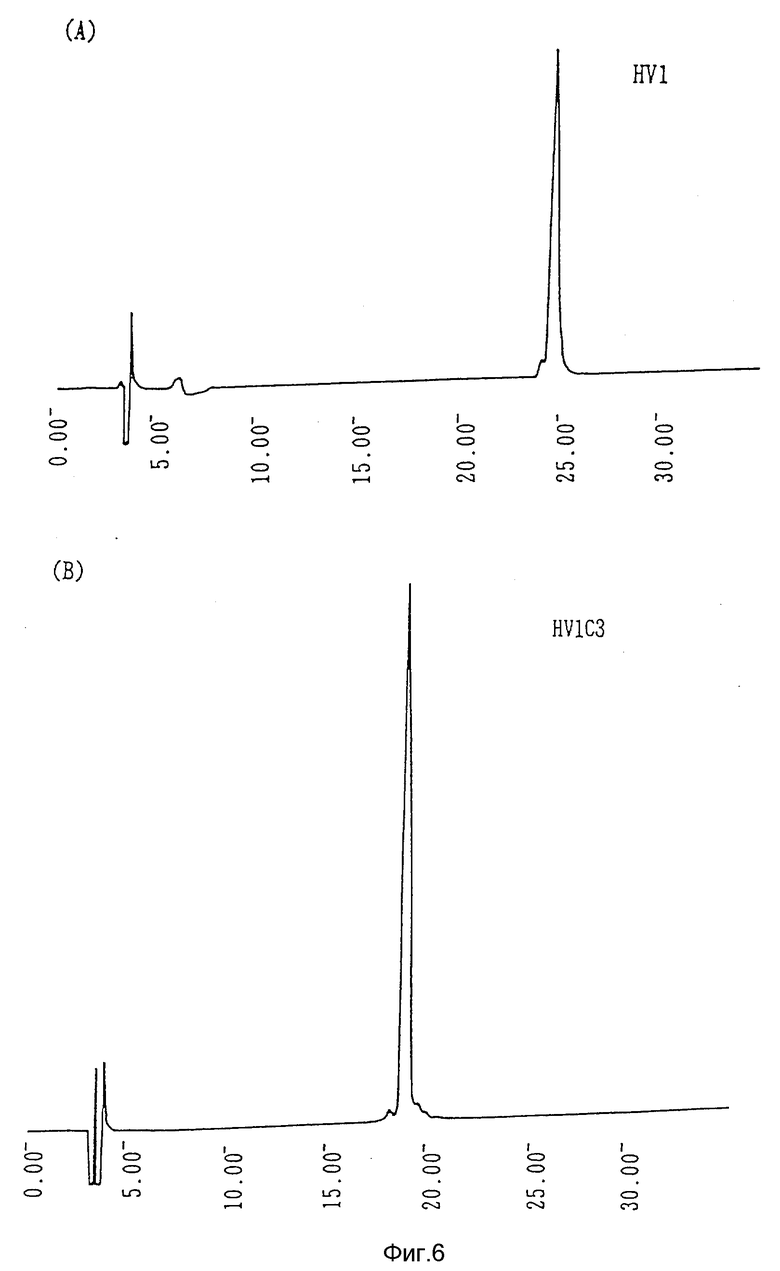
Фиг. 6 - (А) кривая С4 обратно-фазовой жидкостной хроматографии высокого разрешения, показывающая профиль гирудина HV1; (В) кривая С4 обратно-фазовой жидкостной хроматографии высокого разрешения, показывающая профиль гирудина HV1C3.
Фиг.7 - последовательность ДНК промотора tac, используемого согласно данному изобретению.
Фиг.8 - последовательность ДНК промотора trp, используемого согласно данному изобретению
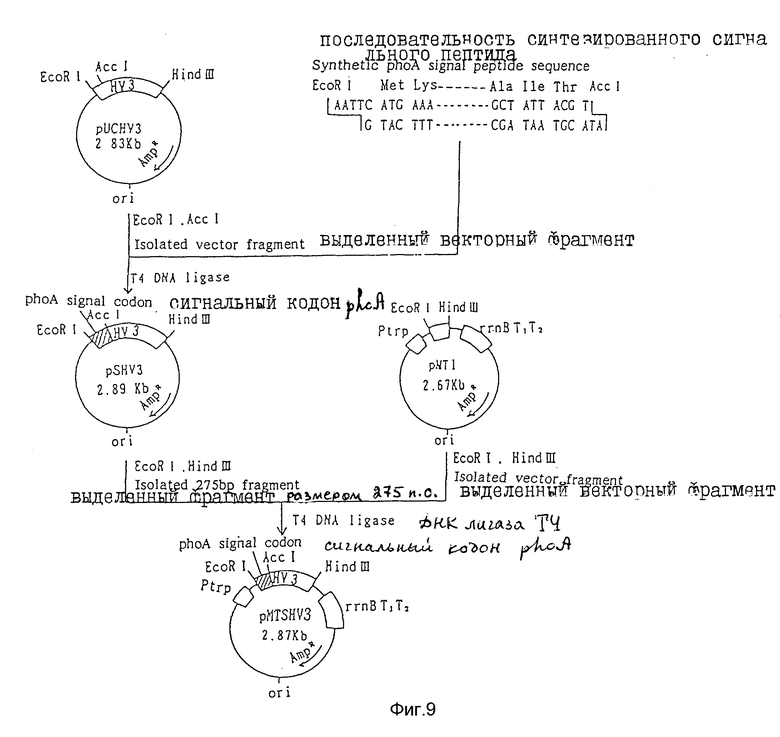
Фиг. 9 дает общую схему построения плазмиды pMTSHVS, секретирующей гирудин HV3.
Фиг. 10 иллюстрирует ДНК последовательность сигнального пептида phoA для секреции гирудина HV3, используемую в примере 5.
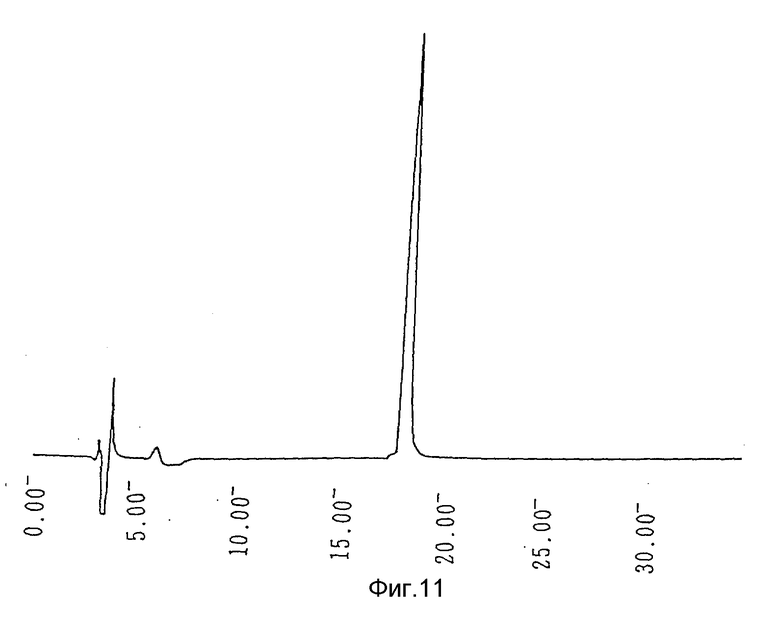
Фиг. 11 - кривая С4 обратно-фазовой жидкостной хроматографии высокого разрешения, показывающая профиль очищенного гирудина HV3.
Наилучший вариант осуществления данного изобретения.
В нижеследующих примерах описывается конструирование плазмид pUCHV1, pMT1 и рМК2 для получения плазмиды pHTSHV1 или pMKSHV1 секретирующей HV1, которая используется в данном изобретении, а также конструирование плазмиды pUCHV3 для получения плазмиды pMTSHV1C3.
Пример 1. Получение плазмид pUCHV1 и pUCHV3.
10 мкг коммерчески доступной плазмиды pUC18 обрабатывают 30 ед. EcoRI и 30 ед. Hind III при температуре 37oC в течение 2 ч. Затем векторную часть отделяют и экстрагируют путем электрофореза в агарозном геле. Затем удаляют белки с помощью фенольной экстракции, осаждают их холодным этанолом и растворяют в 50 мкл буферного раствора ТE (10 мМ Трис-HCl, pH=8,0, 1 мМ этилендиаминтетрауксусной кислоты). К количеству данного раствора, содержащего 50 нг ДНК, добавляют 10 мкл раствора (66 мМ Трис-HCl, pH=7,5, 5 мМ MgCl2, 5 мМ DTT,1 мМ АТП и 300 ед. Т4 ДНК лигазы), содержащего двухнитевую ДНК гирудина НV1 или HV3. Далее проводят реакцию в течение ночи при температуре 16oC и получают плазмиду pUCHV1 или pUCHV3, в которые вставлен соответственно ген HV1 или ген HV3 между EcoRI и Hind III сайтом плазмиды pUC18.
Пример 2. Получение плазмид рМК2 и PMT1.
a) Получение плазмиды рМК2.
Фрагмент, содержащий tac-промотор, полученный путем расщепления промышленно доступной плазмиды рКК223-3 (Pharmacia) ферментами рестрикции PvuI и NruI, фрагмент, содержащий оргужин репликации (ori), полученный путем расщепления промышленно доступной плазмиды pUC18 ферментами рестрикции PvuI и PvuII и фрагмент, содержащий ген устойчивости к ампициллину, сшивают с помощью ДНК лигазы Т4. Полученный таким образом фрагмент вводят в штамм E.coli JM109 с последующим культивированием и селекцией по устойчивости к ампициллину, и получают вектор, содержащий как промотор, так и оргужин репликации (ori) плазмиды pUC18. Данный вектор обозначают рМК2.
(б) Получение плазмиды pMT1.
10 мкг плазмиды рМК2 обрабатывают 30 ед.ЕсоRI и Eсо47 III, удаляют фрагмент, содержащий tac-промотор, и затем выделяют фрагмент, содержащий оргужин репликации (ori) путем электрофореза в агарозном геле. Одновременно с этим синтезируют фрагмент ДНК, содержащий trp-промотор, с помощью синтезатора ДНК. Указанный фрагмент сшивают с полученным выше (расщепление плазмиды рМК2 EcoR1 и Есо47 III) при использовании ДНК лигазы Т4 при температуре 16oС в течение ночи. Полученным конструктом трансформируют E.coli JM109 для получения вектора, содержащего как trp-промотор, так и ориджин репликации (ori) плазмиды рМК2. Последовательность ДНК данной плазмиды определяют методом Сэнгера и др., и указанную плазмиду обозначают pMT1.
Настоящее изобретение более подробно иллюстрируется следующими примерами.
Пример 1.
(1) Получение плазмиды секреции pMTSHV1 гирудина HV1.
Плазмиду pMTSHV1 получают как показано на фиг. 3. Синтезируют четыре олигонуклеотида, приведенных на фиг. 2, для построения фрагмента ДНК, соответствующего сигнальному пептиду щелочной фосфатазы (phoA) E.coli, и фрагмента ДНК, кодирующего Val1-Val2 N-терминальной аминокислотной последовательности гирудина HV1. После снятия защиты каждый нуклеотид очищают путем электрофореза в 10%-ном полиакриламидном геле.
После фосфорилирования 500 пкМ каждого из двух олигонуклеотидов (S2, S4) 20 пкМ каждого из четырех олигонуклеотидов смешивают, отжигают и затем обрабатывают при 16oC в течение ночи 20 мкл раствора, содержащего ДНК лигазу Т4. После удаления белка фенолом и хлороформом и осаждения холодным этанолом, получают нужный фрагмент двухнитевой ДНК. 1/10 количества полученного таким образом фрагмента и 100 нг плазмиды pUCHV1 (выложенная для ознакомления заявка на японский патент 3-164184, 1991 г.), расщепленной ферментами рестрикции EcoRI и АссI, обрабатывают ДНК лигазой Т4 при 16oC в течение ночи. Гибридную плазмиду pSHV1, содержащую ген слияния, в котором последовательность ДНК, кодирующая гирудин HV1, расположена непосредственно после последовательности, кодирующей сигнальный пептид phoА, получают путем трансформации штамма E.coli JM109. Последовательность ДНК плазмиды pSHV1 определяют методом Сэнгера и др.
10 мкг плазмиды pSHV1 обрабатывают ферментами рестрикции EcoRI (30 ед.), и Hind III (30 ед.) и фрагмент слитого гена размером 276 пар оснований (вр) очищают путем электрофореза в агарозном геле.
100 нг полученного таким образом фрагмента ДНК и 100 нг фрагмента ДНК, полученного при расщеплении вектора экспрессии E.coli pMT1 (выложенная для ознакомления заявка на японский патент 3-76579, 1991 г.) ферментами рестрикции EcoRI и Hind III с последующей очисткой фрагментов ДНК электрофорезом в агарозном геле, соединяют с помощью ДНК лигазы Т4 при температуре 16oC в течение ночи. Указанной реакционной смесью трансформируют E.coli JM109 для получения плазмиды pMTSHV1, секретирующей HV1, в которой ген слияния, кодирующий сигнальный пептид phoA и гирудин HV1, расположен за trp-промотором. Последовательность ДНК плазмиды pMTSHV1 определяют методом Сэнгера и др.
(2) Получение плазмиды pMKSHV1, секретирующей гирудин HV1.
Плазмиду pMKSHV1 получают, как показано на фиг. 4.
Указанную выше ДНК слияния, кодирующую сигнальный пептид phoA, и гирудин HV1 и фрагмент ДНК, полученный при расщеплении плазмид экспрессии E.coli рМК2 (выложенная для ознакомления заявка на японский патент 3-76579, 1991 г. ) ферментами рестрикции EcoRI и Hind III сшивают с помощью ДНК лигазы Т4, трансформируют E.coli JM109, и получают плазмиду pMKSHV1, секретирующую гирудин HV1, в которой ген слияния расположен ниже tac-промотора. Последовательность ДНК плазмиды pMKSHV1 определяют методом Сэнгера и др.
(3) Секреторная продукция гирудина HV1 с помощью вектора DMTSHV1.
Клетки E. coli JM109, трансформированные плазмидами pMTSHV1 и pMKSHV1, полученными как указано выше в (1) и (2), культивируют в среде 2 х УТ (16 г/л бактотриптона, 10 г/л бактодрожжевого экстракта и 5 г/л NaCl), содержащей 100 мкг/мл ампициллина. После культивирования при 37oC в течение 24 ч собирают 1 мл культуральной среды.
Осажденные клетки каждого из образцов суспендируют в 1 мл 50 мМ трис-HCl (pH=7,5), содержащего 25% сахарозы и 1 мМ ЕДТА, затем выдерживают при комнатной температуре в течение 10 мин. После сбора клеток путем центрифугирования при 6000 х g в течение 10 мин, их суспендируют в 1 мл холодной воды для высвобождения вещества в периплазматическое пространство путем осмотического шока. Затем клетки удаляют из периплазматической фракции центрифугированном при 6000 х g в течение 10 мин. Количество секретированного и накопившегося в периплазме гирудина определяют путем измерения его антитромбиновой активности в супернатанте. Антитромбиновую активность определяют по количественному измерению интенсивности окраски создаваемой гидролитическим действием тромбина на синтетический хромогенный субстрат хромозим ТН (тоцилглицил - пролиларгинин - 4-нитроанилидацетат, Boeringer-Mannhaim) и ингибирующему действию гирудина на тромбин, которое подавляет проявление окраски.
В результате выявляется, что штамм, содержащий плазмиду pMTSHV1, обнаруживает 450 ед. антитромбиновой активности (ATU) на 1 мл культуральной среды.
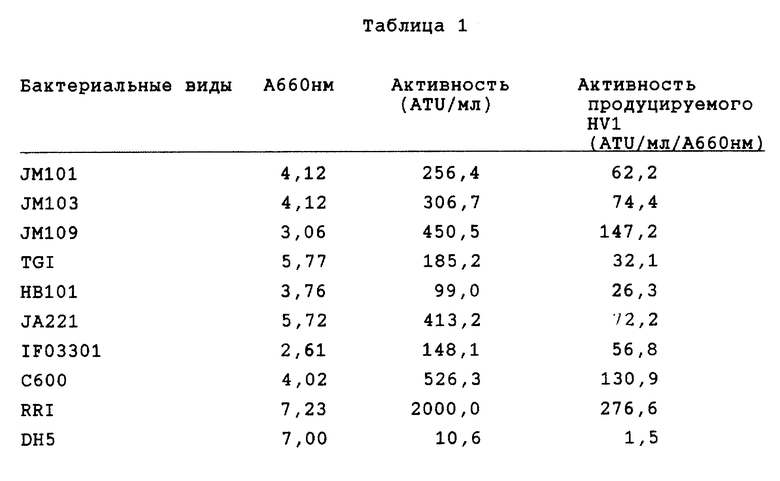
Кроме того, штамм, содержащий плазмиду pMKSHV1, обнаруживает 360 ATU на 1 мл среды разведения. В табл. 1 представлены результаты других исследований, касающихся получения гирудина при использовании различных штаммов E. coli, таких как JM101, JM103, JM109, TG1, HB101, JA221, IF03301, C600, RR1 и DH5, в которые методом трансформации по Hanahan et al. введена плазмида pMTSHV1.
При использовании RRI в качестве хозяина внеклеточная продукция HV1 составляет примерно 2000 ATU/мл.
(4) Секреция гирудина HV1 в культуральную среду при использовании трансформированных клеток E.coli JM109/pMTSHV1.
При культивировании штамма E.coli JM109, трансформированного плазмидой pMTSHV1 (E.coli JM109/pMTSHV1) (FERM BP-3266), в 2 л питательной среды 2хУТ, содержащей 2% глюкозы, в 5-литровом ферментере с перемешиванием и аэрацией при 37oC в течение 24 ч внеклеточная продукция гирудина составляет около 5300 ATU на 1 мл среды
(5) Выделение гирудина HV1 из питательной среды.
После ферментации собирают 1,5 л питательной среды и центрифугируют при 6000 х g в течение 10 мин для отделения супернатанта от клеточного дебриса. Поскольку концентрация соли в супернатанте составляет 1,3%, то супернатант четырехкратно разбавляют 10 мМ буферного раствора фосфата калия (pH=7,0) и фильтруют через фильтр с диаметром пор 3,2 мкм (Pole Со.Ltd). Полученный фильтрат наносят на колонку с QAE-toyopearl (4,4 х 7 см), уравновешенную 10 мМ буферным раствором фосфата калия (pH=7,0). После загрузки образца колонку уравновешивают буферным раствором с последующей ступенчатой элюцией гирудина HV1 0,2 М NaCl. Элюированный раствор концентрируют на мембране Diaflow (YM5) фирмы Amicon, и затем осуществляют гель-фильтрацию с помощью Сефакрила Sephacryl S-100HR, предварительно уравновешенного 10 мМ буферным раствором фосфата калия (pH=7,0) для обессоливания.
Активные фракции наносят на колонку с DEAE-toyopearl (4,4 х 40 см), уравновешенную 10 мМ буферным раствором фосфата калия (pH=7,0), тщательно промывают и затем элюируют линейным градиентом, создаваемым между 3 л уравновешивающего буфера и 3 л 0,3 М NaCl в уравновешивающем буфере. Окончательную очистку осуществляют посредством обратно-фазовой жидкостной хроматографии высокого разрешения при использовании колонки С4 с Delta-prep 3000, полученным от фирмы Waters. Очищенный гирудин HV1 получают элюированием его из хроматографической колонки с помощью линейного градиента 15-30%-ного (по объему) ацетонитрила, содержащего 0,065% (по объему) трифторацетата.
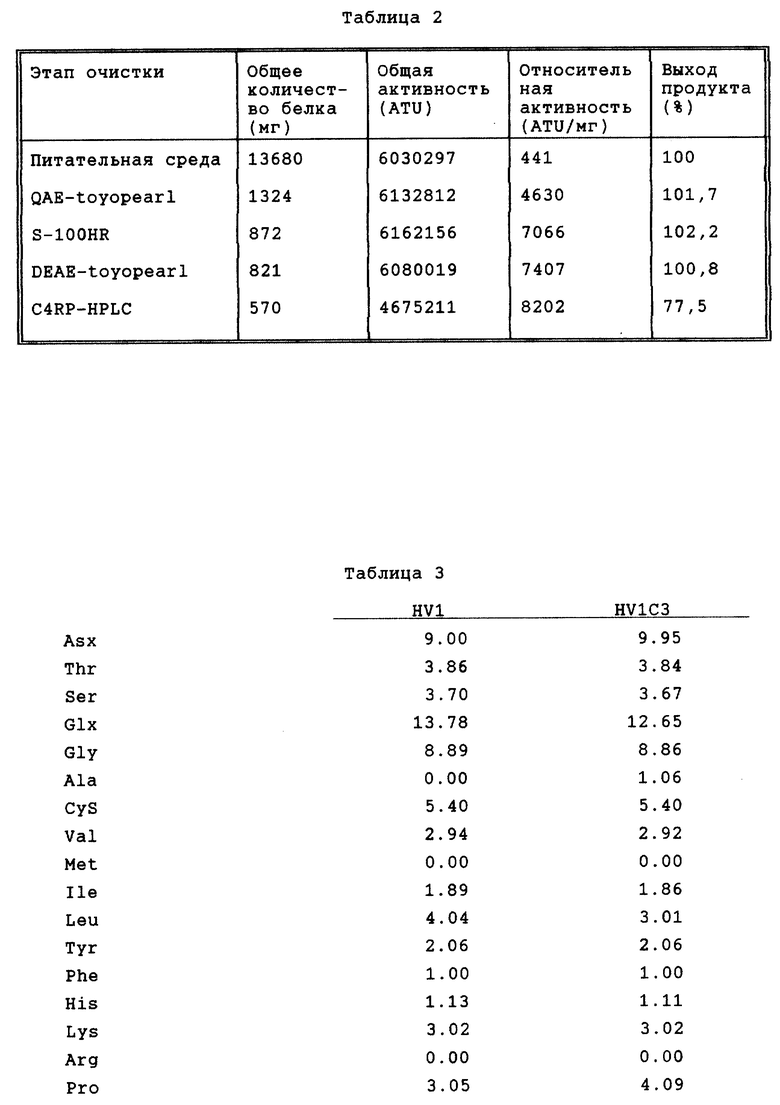
Степень очистки, достигаемая на каждом этапе, приведена в табл. 2.
Аминокислотный состав полученного таким образом гирудина HV1 является постоянным по сравнению с природным гирудином HV1, как показано в табл. 3. Антитромбиновая активность составляет 8202 ATU/мг.
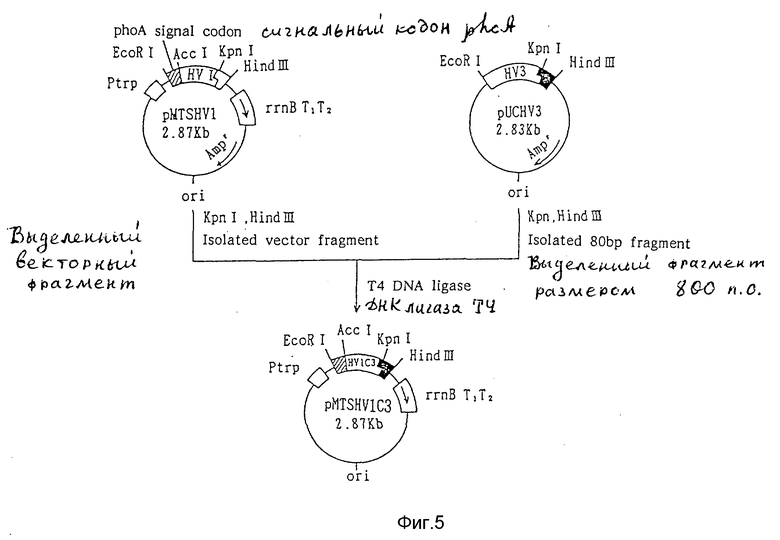
6) Получение плазмиды pMTSHV1C3, секретирующей химерный гирудин.
Плазмиду pMTSHV1C3 получают согласно способу, показанному на фиг. 5. Для замены аминокислотных групп гирудина HV1, начиная с 53-й аминокислоты последовательностью гирудина HV3, плазмиду pMTSHV1 (10 мкг), секретирующую HV1 обрабатывают ферментами рестрикции Крn 1 (30 ед.) и Hind III (30 ед.) с последующим электрофорезом в агарозном геле для очистки фрагмента ДНК размером 2,8 т.п.о.
Аналогичным образом 10 мкг плазмиды pUCHV3 (выложенная для ознакомления заявка на японский патент 3-164184, 1991) также обрабатывают Kpn 1 и Hind III и получают фрагмент ДНК размером 80 п.о., кодирующий С-терминальную аминокислотную последовательность гирудина HV3.
100 нг обоих фрагментов ДНК обрабатывают ДНК лигазой Т4 при температуре 16oC в течение ночи, и полученную реакционную смесь используют для трансформации E. coli JM109 с выделением плазмиды pMTSHV1C3, секретирующей химерный гирудин HV1C3. Последовательность ДНК плазмиды pMTSHV1C3 определяют методом Сэнгера и др.
(7) Получение химерного гирудина HV1C3 с помощью плазмиды секреции.
Клетки E.coli RRI (FERM ВР-3130), трансформированные плазмидой pMTSHV1C3, полученной как описано выше (6), культивируют в питательной среде 2хУТ, содержащей 100 мкг/мл ампициллина. После культивирования в течение 24 ч при температуре 37oC, собирают 1 мл культуральной среды и с помощью осмотического шока измеряют антитромбиновую активность переплазматической фракции.
Результирующая антитромбиновая активность составляет 3060 ATU на 1 мл культуральной среды.
(8) Секреция химерного гирудина HV1C3 в ферментационную или культуральную среду трансформированным штаммом E.coli JM109/pMTSH1C3
При культивировании штамма E.coli JM109 (FERM ВР-3104), трансформированного плазмидой pMTSHV1C3, в 2 л питательной среды 2хУТ, содержащей 2% глюкозы, в 5-литровом ферментере с перемешиванием и аэрацией при температуре 37oC в течение 24 ч, обнаруживают общую антитромбиновую активность равную 6050 ATU/мл. Активность в периплазме и культуральной среде составляет 350 ATU/мл и 5700 ATU/мл соответственно.
(9) Очистка химерного гирудина HV1C3.
Гирудин HV1C3 выделяют из культуральной среды, полученной как указано выше (8), согласно способу, описанному в (5). При определении аминокислотной последовательности очищенного химерного гирудина HV1C3, было выявлено, что его С-терминальный участок отличен от такового HV1, как показано на фиг. 1. Специфическая активность составляет 11 250 ATU/мг. Профили HPLC для очищенных HV1 и HV1C3 приведены на фиг. 6. В эксперименте используют хроматографическую колонку с VYDAC С4 (0,46 х 25 см) с линейным градиентом ацетонитрила от 15 до 30% при скорости потока 1 мл/мин в течение 30 мин.
Пример 2.
Ингибирующее действие гирудина на смертельный исход, вызванный тромбином.
В организм мышей самцов (весом 20-25 г) без анестезии вводят внутривенно тромбин (10 NIH ед/10 г) и определяют антитромбиновую активность испытываемого соединения при наблюдении за исчезновением уплотнения при рефлексии и смертельном исходе, что служит показателями этой активности.
При индуцированной тромбином смертельной реакции в условиях "ин виво", химерный гирудин (HV1C3), полученный согласно данному изобретению, обнаруживает примерно в 3-4 раза более сильную ингибирующую активность по сравнению с гирудином HV1.
Получение гирудина HV3.
(1) Конструирование плазмиды, секретирующей гирудин HV3.
(i) Как показано на фиг. 9, ориджин репликации (ori), полученный при расщеплении плазмиды pUCHV3, сконструированной как описано в примере 1, ферментами рестрикции EcoRI и AccI, векторный фрагмент, содержащий ген устойчивости к ампициллину, и синтетический ген, кодирующий сигнальную последовательность щелочной фосфатазы (phoA), строение которой приведено на фиг. 10, сшивают с помощью ДНК лигазы Т4. В результате получают плазмиду pSHV3, в которой последовательность ДНК, кодирующая сигнальный пептид щелочной фосфатазы (pho А), присоединена к последовательности ДНК, кодирующей HV3. Данную плазмиду вводят в клетки E.coli JM109, культивируют их и отбирают клоны, устойчивые к ампициллину.
(ii) Далее плазмиду pSHV3 расщепляют ферментами рестрикции EcoRI и Hind III и получают фрагмент размером 275 п.о., соответствующий последовательности ДНК сигнального пептида щелочной фосфатазы (phoА), расположенный перед последовательностью ДНК, кодирующей гирудин HV3.
(iii) Одновременно получают плазмиду pMT1, как описано в примере 2, Данная плазмида содержит последовательность ДНК, включающую ориджин репликации (ori) плазмиды pUC18 и trp-промотор, как описано в примере 2. Векторный фрагмент получают при расщеплении плазмиды pMT1 ферментами рестрикции EcoRI и Hind III.
(iv) Указанный выше фрагмент размером 275 п.о. и векторный фрагмент сшивают ДНК лигазой Т4, вводят в клетки Е.coli JM109, культивируют и отбирают клоны, устойчивые к ампициллину. В результате получают плазмиду pMTSHV3, вектор экспрессии HV3 размером 2,87 п.о., который содержит последовательность ДНК ориджина репликации (ori) плазмиды pUC, последовательность ДНК trp-промотора, последовательность ДНК, кодирующую сигнальный пептид щелочной фосфатазы (phoА) и последовательность ДНК, кодирующую HV3.
(2) Получение гирудина HV3 с помощью секреции.
Клетки E. coli RRI (E.coli RRI /pMTSHV3) (PERM BP-3267), трансформированные плазмидой pMTSHV3, которую получают указанным выше способом, культивируют в 2 л питательной среды 2 х УТ, содержащей 100 мкг/мл ампициллина и 2% глюкозы. Культивирование осуществляют в 2 л питательной среды в 5-литровом сосуде при 37oC в течение 24 ч. Обнаруживаемая антитромбиновая активность составляет 6073 ATU на 1 мл питательной среды.
(3) Выделение гирудина HV3 из культуральной среды.
Гирудин HV3 получают из культуральной среды способом, описанным в примере 1 (5). После центрифугирования культуральной среды для отделения супернатанта от клеток, его четырехкратно разбавляют 10 мМ буферным раствором фосфата калия (pH= 7,0) и фильтруют. Полученный фильтрат наносят на колонку (4,4 х 13 см) с QAE-toypearl, как описано в примере 5, и осуществляют гель-фильтрацию при использовании колонки с Sephacryl C-100HR.
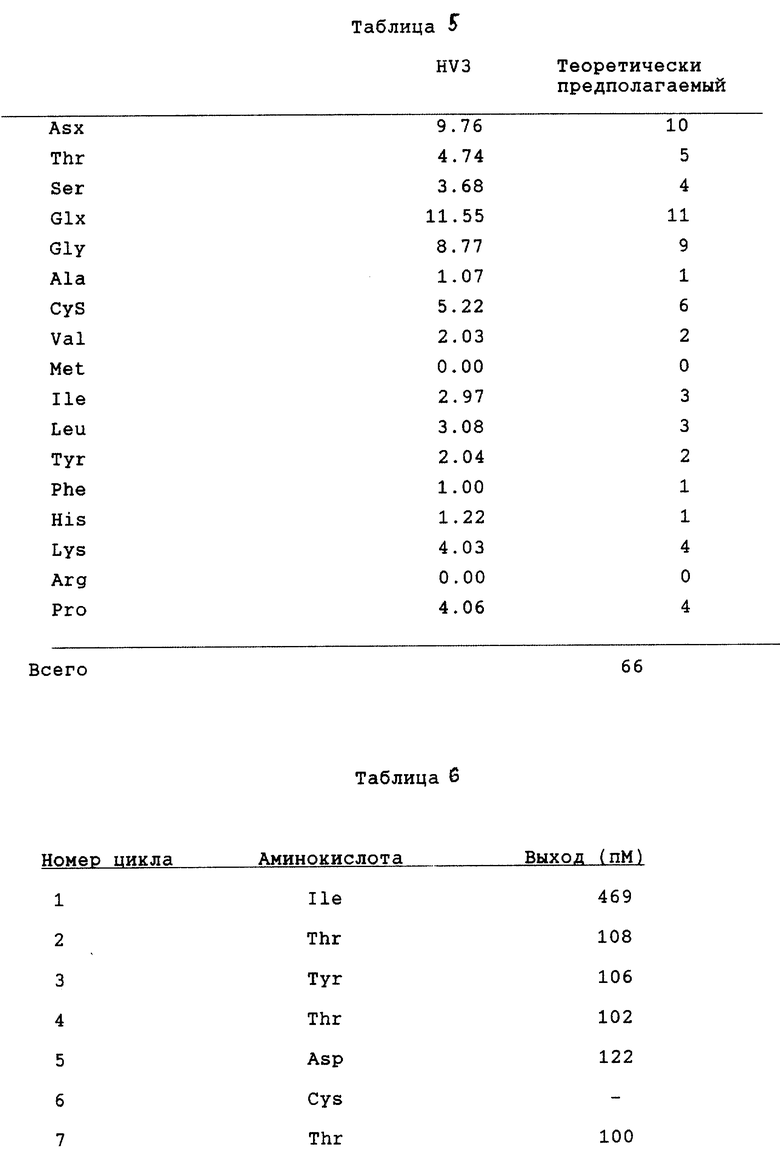
Активную фракцию наносят на колонку с DEAE toyopearl (4,4 х 40 см), уравновешенную буферным раствором, и затем элюируют с помощью линейного градиента от 0 до 0,3 М NaCl в уравновешивающем буферном растворе. И, наконец, гирудин HV3 очищают посредством обратно-фазовой жидкостной хроматографии высокого разрешения с использованием колонки С4 Vydac С4 (4,7 х 30 см) путем элюирования линейным градиентом (1%/мин, 30 мин или более) ацетонитрила от 15 до 30% (по объему). Профиль элюции показан на фиг. 11. Степень очистки, достигаемая на каждом этапе, приведена в табл. 4.
Аминокислотный анализ продукта показывает, что аминокислотный состав гирудина HV3 постоянен и сопоставим с теоретическим, как указано в табл. 5. Как приведено в табл. 6, вплоть до 15-й N-терминальной аминокислоты, аминокислотная последовательность полученного продукта идентична таковой гирудина HV3. Эти результаты показывают, что осуществлен правильный процессинг сигнального пептида и полученным продуктом является гирудин HV3.
Таким образом, при использовании векторов секреции и трансформированных микроорганизмов согласно изобретению предложенный способ позволяет получать внеклеточно чужеродные белки с высоким выходом.






Изобретение относится к биотехнологии, генной инженерии. Способ получения гирудина HV1 осуществляют трансформацией рекомбинантного штамма Escherichia coli JM109 (BP-32266) вектором секреции pMTSHV1 или pMKSHV1 с последующим его культивированием и выделением целевого продукта из среды или периплазмы клеток. Вектор секреции содержит 1440 п.о. PvuI - PvuII-фрагмент, содержащий область репликации pUC18 плазмиды, trp или tac промотор, ДНК фрагмент, кодирующий сигнальный пептид щелочной фосфатазы, ДНК фрагмент, кодирующий гирудин HV1, соединенные в сайте AccI и ген резистентности к ампициллину в качестве генетического маркера. Гирудин HV1 проявляет более высокую антитромбоцитарную активность, эффективно снижает кровоточивость. 4 с. и 1 з.п. ф-лы, 6 табл., 11 ил.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| "FEBS Lett", v.202, 1986, с.373 - 377. | |||

