Изобретение относится к получению производного гирудина с N-концевой последовательностью аминокислот Ала-Тре-Тир-Тре-Асп в секреторных мутантах E.coli.
Гирудин - это полипептид из 65 аминокислот, выделяемый из кровяного геля Hirudo medicinalis. Он действует как высокоспецифический ингибитор тромбина путем образования стабильных комплексов с ним и обладает благодаря этому широкими терапевтическими возможностями, в особенности для антикоагуляционной терапии (F. Markguardt, Biomed. Biochim. Acta. 44 (1985), 1007 - 1013).
После опубликования полной последовательности аминокислот гирудина (J. Dodt и др., FEBS LETTERS 165 (2), (1984), 180 - 184) были созданы предпосылки для получения гирудина с помощью техники рекомбинантных ДНК и экспрессии в микроорганизмах.
В EP-A-О 158564 (Transgene) представлены векторы клонирования для экспрессии гирудина или его аналога в клетке-хозяине, в частности в бактериальной клетке. Ген, кодирующий гирудин, получают при этом синтезом ДНК на матрице mРНК кровяного геля Hirudo medicinalis. Описано также производное гирудина с N-концевой последовательностью Иле-Тре-Тир-Тре-Асп и способ его получения.
В EP-A-O 168342 (Ciba Geigy) представлены ДНК-последовательности, кодирующие аминокислотную последовательность природного гирудина, который имеет N-концевую последовательность аминокислот Вал-Вал-Тир-Тре-Асп. Экспрессию гирудина осуществляют в микроорганизмах E.coli и Saccharomyces cerevisiae внутриклеточно.
В EP-A-O 171024 (Hoechst AG) представлен способ генно-инженерного получения пептидов с активностью гирудина, преимущественно в клетках E.coli, при этом клетки разрушают и из клеточного экстракта получают полипептид с активностью гирудина. Образующийся слитый белок можно разделить путем протеолитического или химического расщепления и очистить освободившиеся молекулы гирудина.
Предметом изобретения WO 86/03517 и DE-OS 3445571 (GEN-BJO-TEC) является последовательность ДНК, которая кодирует белок с биологической активностью гирудина, а также способ получения такого белка из клеток E. coli, которые трансформируют пригодным рекомбинантным вектором, путем лизиса клеток.
В работе Бергмана и др. (Biol. Chem. Hoppe Seyler 367, (1986), 731 - 740) описан синтез гирудина в E. coli. Выделение гирудина из клеток осуществляют путем обработки толуолом, причем достигается лишь незначительный выход около 500 нг/л культуры с плотностью A578 единиц.
В EP-A-O 200655 (Transgene), EP-A-O 252854 (Transgene) и EP-A-O 225633 (Ciba Geigy) представлено получение белка с активностью гирудина путем секреции из эукариотического хозяина, в особенностей дрожжей, трансформированных вектором, который содержит ДНК-последовательность сигнального пептида перед структурным геном. Описана секреция производных гирудина с N-концевой последовательностью Вал-Вал-Тир-Тре-Асп, а также с N-концевой последовательностью Иле-Тре-Тир-Тре-Асп в дрожжах. При этом указан выход до 100 мг/л.
В DE-OS 39 00 626 (Hoechst AG) представлено производное гирудина с N-концевой последовательностью Лей-Тре-Тир-Тре-Асп. Экспрессия преимущественно имеет место в дрожжах, причем применяют промотор и сигнальную последовательность гена феромона дрожжей MFα для экспрессии и секреции производного гирудина.
Все описанные выше способы получения производных гирудина имеют недостатки. Так, при применении дрожжей в качестве хозяина для продукции и секреции гирудина в культуральной среде получают относительно высокий выход, однако культивирование дрожжевых клеток является более длительным процессом, требующим соблюдения определенных условий, чем в случае с бактериями E. coli. В клетках E. coli, наоборот, имеет место относительно низкий выход и/или при увеличении числа клеток усложняется способ выделения.
Задачей изобретения в связи с эти является разработка простого способа получения производных гирудина, при котором эти производные можно получить с высоким выходом из бактериальных клеток, не прибегая к необходимости использования их избытка.
Предметом изобретения является способ получения производных гирудина с использованием секретирующих мутантов E. coli, по которому 1) конструируют рекомбинантный вектор, в котором ген, кодирующий производные гирудина, следует за ДНК-фрагментом, кодирующим в свою очередь бактериальный сигнальный пептид, 2) трансформируют секретирующие мутанты E. coli рекомбинантным вектором, сконструированным по п. 1), 3) трансформированные клетки культивируют в среде и 4) выделяют из среды производные гирудина.
Под понятием производного гирудина согласно изобретению понимают происходящие от гирудина белки, которые действуют как тромбинингибиторы и обладают специфической активностью, равной по меньшей мере 10000 AT-U/мг, где AT-U антитромбиновые единицы (Dodt и др., Biol. Chem. Hoppe Seyler 366 (1985), 379 - 385).
Способом согласно изобретению получают производные гирудина со следующей последовательностью аминокислот:
(X)m-Z-Тре-Tир-Тре-Acп,
где
m = 0 - 50;
X - одинаковые или различные генетически кодированные аминокислоты;
Z - генетически кодированная аминокислота из группы Лей, Иле, Ала, Вал, Гли, Сер, Асп, Глу, Асн, Глн, Гис, Мет, Фен и Тир.
Поскольку m>0, то последовательность X содержит протеолитически или химически расщепляемое место, предпочтительно у своего конца. Если, например, последняя аминокислота X - является Арг- остатком, то можно слитую последовательность X отщепить путем триптического переваривания (расщепление после Арг) и очистить активное производное гирудина. Отщепление слитой части, однако, можно проводить с использованием и других известных протеолитических ферментов или химических расщепляющих реагентов. Если, например, аминокислотная последовательность X оканчивается Мет- остатком, то можно слитый белок расщепить с помощью галогенциана (E. Gross и B. Wittkop, J. Am. Chem. Soc., 82 (1961), 1510 - 1517). Если, например, C - конец последовательности аминокислот X включает аминокислотную последовательность Иле-Глу-Глу-Арг, то расщепление можно проводить фактором Xa (EP-A 0025190 и EP-A 0161973).
Если в способе согласно изобретению m = 0, то Z - преимущественно ГЛН, Гис, Фен, Тир, Гли, Сер, Асп или Асн, в особенности Ала, Гли, Сер, Асп или Асн. Наиболее предпочтительно производное гирудина, если m = 0, а Z - Ала.
Таким образом, предметом изобретения являются также производные гирудина с N-концевой последовательностью А-Тре-Тир-Тре-Асп, где А-Ала-Глн, Гис, Фен, Тир, Гли, Сер, Асп или Асн, предпочтительно Ала, Гли, Сер, и Асп или Асн. Наибольшее предпочтение отдают производному с N-концевой последовательностью Ала-Тре-Тир-Тре-Асп. Это производное гирудина можно получить в культуре секреторного мутанта E. coli с выходом активной формы до 2 г/л среды.
Преимущество способа согласно изобретению также состоит в том, что благодаря секреции производного гирудина в среду дисульфидные мостики гирудина легко образуются при создании окислительных условий вне клетки.
Под определением "секреторные мутанты E. coli", согласно изобретению принимают штаммы E. coli, которые дают массовую секрецию белков в культуральную среду. Способ получения этих мутантов представлен в EP-A-O 338410. При получении пригодных мутантов E. coli можно исходить из E. coli DS 410/DSM 4513/ или E/coli BW 7261/DSM 5231/. Штамм E. coli трансформируют сначала плазмидой, которая включает ДНК-последовательность, кодирующую выделяемый белок. Затем трансформированный штамм E. coli подвергают мутагенезу, например, путем обработки N-метил-N'-нитро-N- нитрозогуанидином и проводят селекцию пригодных штаммов-мутантов. Если выбирают в качестве выделяемого белка, например, α -циклодекстрингликозилтрансферазу, то можно обнаружить мутанты по резистентности в D-циклосерину, веществу, проявляющему активность относительно стенок клетки. Далее секреция α -циклодекстрингликозилтрансферазы (CGTase) способствует гидролизу крахмала в окружающей среде, что при использовании амилопектин-азур-среды приводит к дополнительной возможности селекции мутантов.
В качестве рекомбинантного вектора для данного изобретения пригодны векторы, которые могут либо интегрировать в геном E. coli (например, бактериофаги λ), либо существовать в трансформированной клетке E. coli вне хромосомы (например, плазмиды). Преимущественно применяют плазмиды.
В рекомбинантный вектор встраивают ген, который кодирует белок, состоящий из сигнального пептида и производного гирудина, и находится преимущественно под контролем индуцируемого промотора, особенно trp-lac - промотора, который может быть индуцирован добавкой лактозы или ИРТГ (изопропил- β -D-тиогалактозид). Далее в векторе должен быть ген-маркер для селекции, в данном случае ген lac-репрессора.
В качестве бактериальной сигнальной последовательности, способствующей секреции производного гирудина, в принципе пригодны все известные сигнальные пептиды, отвечающие за прохождение через мембрану клеток E. coli. В основном применяют также сигнальные пептиды из грамм-отрицательных бактерий, например сигнальные пептиды следующих белков E. coli: наружный мембранный белок OmpA (DiRienzo и др., 1978 Ann. Rev. Biochem. 47: 481 - 532), щелочная фосфатаза PhoA (Inouye и др. , 1982 J. Bacteriol 149: 434 - 439), LamB - белок (Hedgpeth и др., 1980 Proc. Nat. Acad. Sci. USA 77: 2621 - 2625), мальтоза - связующий белок MalE (Bedouelle и др., 1980 Nature 285: 78 - 81). Преимущественно применяют сигнальный α -CGTase-пептид.
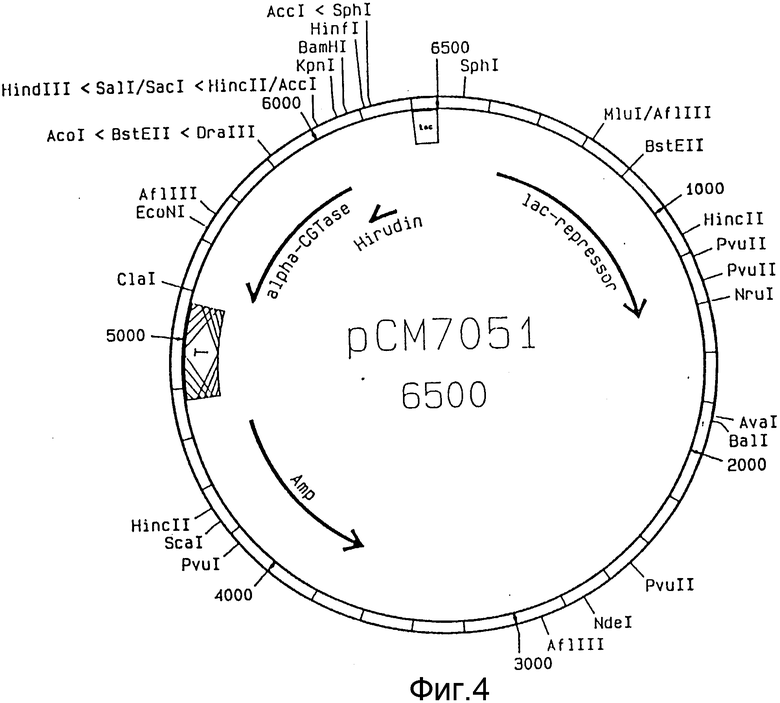
Вектором, пригодным для способа согласно изобретению, являются, например, плазмида pCM705 (фиг. 1), которую получают из плазмиды pCM703, описанный в EP-A-O 383410, путем выделения Nru I - фрагмента длиной около 1 kb. Этот вектор содержит ген резистентности к ампициллину, ген lac - репрессора и CGTase - ген с участком для кодирования сигнального пептида на 51-конце. Последовательность, кодирующую производные гирудина, интегрируют в вектор pCM705 с тем, чтобы внутри клетки образовались промежуточные молекулы с сигнальным пептидом α -CGTase на его N-конце. Сконструированный ген находится под контролем tac-промотора. Полученной таким образом плазмидой можно трансформировать мутантный штамм E. coli.
Положительно трансформированные клоны культивируют во встряхивающихся колбах или ферментерах. При достижении оптимальной плотности (OD600) около 1 проводят индукцию с помощью ИРТГ или лактозы.
С помощью теста тромбин-инактивация (Griesbach и др., Thrombosis Research 37, (1985), 347 - 350) определяют процесс образования производного пирудина. Накопление слитых белков анализируют с помощью жидкостной хроматографии высокого давления (обратимая фаза). Вспомогательную часть слитых белков потом можно отщепить и очистить образовавшееся активное производное гирудина.
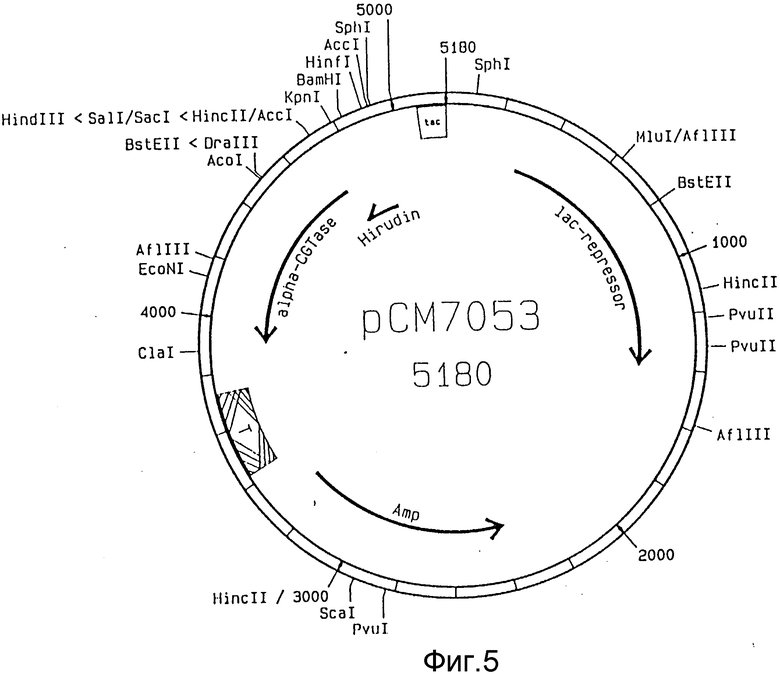
На фиг. 1 показана плазмида pCM705; на фиг. 2 - ДНК-последовательность синтетического гена гирудина из pK152; на фиг. 3 - последовательности олигонуклеотидов HJP1, HJP2 и HJP3: на фиг. 4 - плазмида pCM7051; на фиг. 5 - плазмида pCM7053.
Пример 1. Конструирование вектора секреции.
Плазмида pK152 несет синтетический ген гирудина, последовательность которого представлена в EP-A 0171024. Из этой плазмиды выделяют большой ДНК-фрагмент Hinf I - Hind III размером около 200 bp путем гель-электрофореза в агарозе, содержащий большую часть ДНК-последовательности, кодирующей гирудин (фиг. 2). Отсутствующая 51-концевая последовательность регенерируется вновь синтезированным олигонуклеотидом (HJP1). Последовательность олигонуклеотида представлена на фиг. 3. Благодаря слиянию Hinf1- концов получают производное гирудина с N-концевой последовательностью Ала-Тре-Тир-Тре-Асп.
Плазмиду pCM705 (фиг. 1) расщепляют с помощью Pst1 и Hind III. Оба места разрыва находятся в кодирующей области гена CGTase, благодаря чему выделяется ДНК-отрезок размером 1 kb. Pst I расщепляет точно в области, которая кодирует место разрыва для сигнальной пептидазы.
PstI-Hind III фрагмент pCM705, 6,3 kb, Hind I-Hind III фрагмент pK152 0,2 kb и олигонуклеотид HJP1 лигируют друг с другом, благодаря чему образуется плазмида pCM7051 (фиг. 4). Штамм HB101 (DSM 1607) E. coli трансформируют лигирующей смесью. Колонии, которые на селективной среде с аминопектином и азуром (окрашенный амилопектин) не показывают снижения крахмала и, тем самым, α -CGTase-экспрессии выделяют и очищают до однородности. ДНК-плазмиду выделяют из нескольких очищенных клонов и идентифицируют с помощью рестрикционного анализа.
ДНК-плазмиду, которая имеет корректную генную структуру для гирудина, расщепляют с помощью Nru I и Nde I и выделяют фрагмент размером 5,18 kb путем гель-электрофореза в агарозе.
После достраивания Nde I- сайта ферментом Кленова фрагмент циклизуют. Образованная плазмида обозначена как pCM7053 (фиг. 5).
Мутанты E. coli W CM100, которые получают описанным в EP-A-О 338410 методом, трансформируют полученной плазмидой pCM7053.
Пример 2. Тест на секрецию гирудина во встряхиваемых колбах.
В 10 мл αB -среды с 100 мкм/мл ампиллина и оптической плотностью OD420 = 0,1 засеивают свежую суточную культуру WCM100/pCM7053. Культуру взбалтывают при 30oC. По достижении оптической плотности OD420 = 1,0 добавляют индуктор лактозу в конечной концентрации 1%. После 48 ч берут пробу культуры, центрифугированием отделяют клетки и в фильтрате определяют концентрацию гирудина. Определение проводят с помощью теста тромбин-инактивации. Выход составляет до 4000 AT-U/мл или ≅ 250 мг/л.
Пример 3. Получение гирудина в 10-литровом ферментере
Минимально в 7 л среды с 100 мкм/мл ампиллина с оптической плотностью OD600 = 0,1 засевают свежую суточную культуру WCM100/pCM7053.
Условия ферментации: температура 30oC; скорость перемешивания 450 - 500 rpm; разрежение 0,5 - 1,5 Vvm; pH 7,0±0,1.
По достижении оптической плотности OD600 = 1,0 добавляют 0,5 ммоль ИПТГ / изопропил- β -D-тиогалактозид/.
Через 40 ч после добавки ИПТГ в фильтрате определяли 36000 AT-U/мл, или ≅ 2,25 г/л.
Пример 4. Секреция гирудина с N-концевой последовательностью Ала-Тре-Арг-Лей-Тре-Тир-Тре-Асп.
Опыт проводят аналогично примеру 1, но вместо олигонуклеотида HJP1 применяют олигонуклеотид HJP2 (фиг. 3). Получают слитый белок гирудин с N - концевой последовательностью Ала-Тре-Арг-Лей-Тре-Тир-Тре-Асп. Количество слитого белка в фильтрате определяют с помощью ЖХ-хроматографии высокого давления при использовании "обратимой фазы" (C18 -хроматографическая колонна). Ферментация штамма WCM100 с данной генной конструкцией дает выход слитого белка 25 мг/л. Благодаря расщеплению трипсином можно получить активный гирудин с N-концевой последовательностью Лей-Тре-Тир-Тре-Асп.
Пример 5. Секреция гирудина мутантом WCM88.
Мутант WCM88, полученный описанным в EP-A-О 338410 способом, трансформируют плазмидой pCM7053 (пример 1). Получение гирудина благодаря секреции в среду тестируют с использованием способа со встряхиваемыми колбами и ферментацией.
а) Опыт во встряхиваемых колбах.
Штамм WCM88/pCM7053 культивируют аналогично примеру 1. Через 48 ч определяют в фильтрате культуры концентрацию гирудина. Выход 1800 AT-U/мл или ≅110 мг/л.
б) Получение в 10-литровом ферментере.
Свежий штамм обрабатывают, как описано в примере 3. Через 45 ч после добавки ИПТГ в фильтрате установлено 21000 AT-U/мл, или ≅ 1,3 г/л.
Пример 6. Конструирование вектора секреции, резистентного к тетрациклину.
Выделяют 1,1 kb NruI-фермент плазмиды pBR322 (F. Bolivar et al., Gene 2, 95-113 (1977)) и лигируют его с линеаризированной путем Nru I- расщепления плазмидой pCM7051 (фиг. 4). Лигирующей смесью трансформируют E. coli HB101. Трансформанты отбирают по их резистентности к тетрациклину.
ДНК-плазмиду выделенного клона повторно изолируют и расщепляют с Nde I и Ava I. Фрагменты ДНК разделяют электрофорезом в агарозном геле. Больший фрагмент выделяют и достраивают однонитевые участки с помощью энзима Кленова, а затем лигируют.
Получают плазмиду pCMT203.
Пример 7. Секреция гирудина с применением вектора секреции pCMT 203.
Секреторный мутант WCM 100 (пример 1) трансформируют плазмидой pCMT203. Полученный штамм культивируют в 1-литровом ферментере. Через 45 ч после добавки ИПТГ определяли 42000 AT-U/мл гирудина (≅ 2,63 г/л).

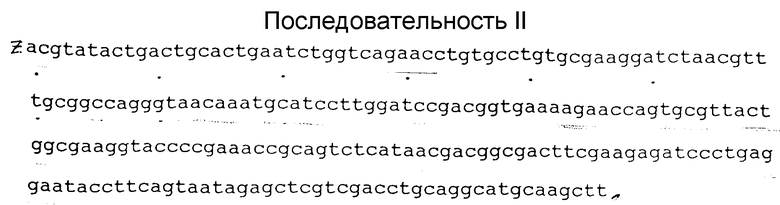

Изобретение относится к биотехнологии. Получено производное гирудина, действующее как ингибитор тромбина, обладающее специфической активностью не менее 10000 АТ ед/мг с выведенной аминокислотной последовательностью. Установлена также нуклеотидная последовательность рекомбинантной ДНК, кодирующей производное гирудина. Способ получения производного гирудина заключается в конструировании плазмиды, содержащей комбинантную ДНК, в трансформации ею секреторного мутанта Escherichia coli, культивировании трансформированных клеток в среде и в выделении целевого продукта. Производное гирудина обладает специфической активностью ингибитора тромбина. 3 с.п.ф-лы, 5 ил.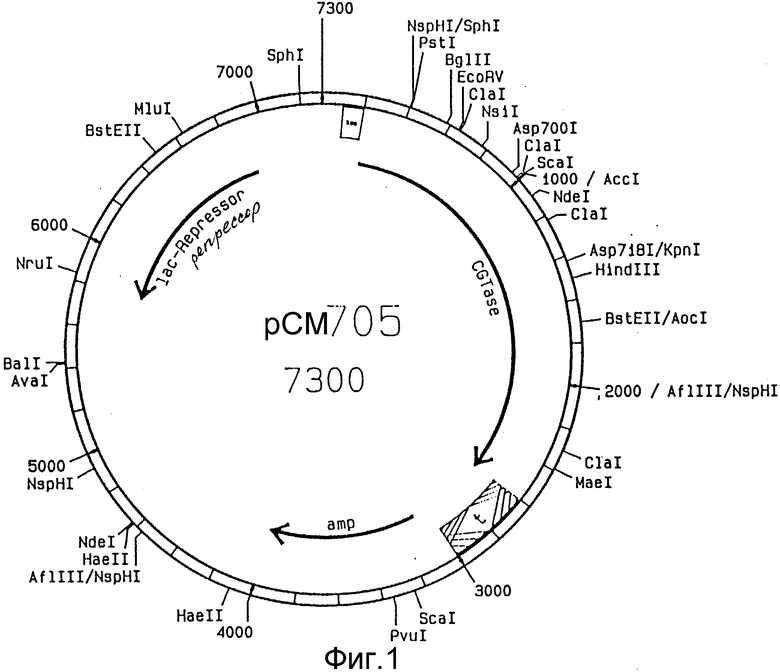
\ \\1 1. Производное гирудина, действующее как ингибитор тромбина, обладающее специфической активностью не менее 10000 АТ ед/мг, имеющее выведенную аминокислотную последовательность I, приведенную ниже, полученное культивированием секреторного мутанта Escherichia coli, трансформированного плазмидой, содержащей рекомбинантную ДНК, кодирующую упомянутое производное гирудина. \\\2 2. Рекомбинантная ДНК, кодирующая производное гирудина, действующее как ингибитор тромбина, и имеющая следующую нуклеотидную последовательность II, приведенную ниже, где Z - кодон для аминокислоты Ала. \\\2 3. Способ получения производного гирудина, действующего как ингибитор тромбина, предусматривающий конструирование плазмиды, содержащей рекомбинантную ДНК, трансформацию ею штамма Escherichia coli, культивирование трансформированных клеток в среде и выделение продукта, отличающийся тем, что трансформируют секреторный мутант Escherichia coli плазмидой, содержащей рекомбинантную ДНК по п.2.
| Пюпитр для работы на пишущих машинах | 1922 |
|
SU86A1 |
| DE 3445517, 19.06.86 | |||
| Опорное приспособление к тензометрам | 1949 |
|
SU77670A1 |
| ОДНОИМЕННОПОЛЮСНЫЙ ИНДУКТОРНЫЙ ГЕНЕРАТОР С КОМБИНИРОВАННОЙ СИСТЕМОЙ ВОЗБУЖДЕНИЯ | 0 |
|
SU200655A1 |
| ФОРМИРОВАТЕЛЬ СТУПЕНЧАТОГО СИГНАЛА | 0 |
|
SU352387A1 |
| УСТРОЙСТВО для ГЛАЗИРОВКИ ПРОДУКТОВ | 0 |
|
SU252854A1 |
| Пюпитр для работы на пишущих машинах | 1922 |
|
SU86A1 |

