Перекрестные ссылки на родственные заявки
По настоящей заявке испрашивается приоритет временной заявки на патент США № 61/288836, поданной 21 декабря 2009 года. Эта временная заявка полностью включена в настоящее описание путем ссылки.
Область техники
Настоящее изобретение в первую очередь относится к вакцинам, которые могут использоваться для иммунизации пациентов против инфекций вирусом простого герпеса типа 2 (Herpex Simplex Virus type 2, HSV-2), ассоциированными с хроническими генитальными язвами. Для вакцины используется дефектный по репликации вирус HSV-2, который в результате генноинженерных изменений экспрессирует антиген гликопротеин D HSV-2 (gD2) на высоком уровне. В предпочтительных вариантах осуществления вирус HSV-2 также экспрессирует один или более иммуномодулирующих генов, таких как IL15 и/или основные антигены HSV-1 или HSV-2, такие как gB или gC.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Вирусы простого герпеса (herpex simplex viruses, HSV) и инфекции HSV
Вирус простого герпеса типа 2 (HSV-2) является основной причиной генитальных язв. Он может вызывать как острую, продуктивную инфекцию, так и длительную латентную инфекцию с непредсказуемыми периодическими рецидивами (66). Инфекция HSV приводит к возникновению рецидивирующих генитальных язв на протяжении всей жизни пациента, а также представляет опасность для пациентов со СПИД. Было показано, что генитальная инфекция HSV-2 утраивает риск заражения ВИЧ половым путем (20), и в Африке этот повышенный риск может служить причиной 25-35% недавних инфекций ВИЧ (1).
Хотя силу и продолжительность наиболее симптоматических первичных инфекций HSV можно снизить с помощью перорального или внутримышечного введения ацикловира, валацикловира или фамцикловира, антивирусная терапия не препятствует переходу первичной инфекции в латентную и не ослабляет последующие рецидивы (66). Исходя из постоянного распространения генитального герпеса в США в течение двух последних десятков лет (19), а также учащения случаев резистентности HSV к современным антивирусным препаратам, очевидно, что в области техники существует потребность в безопасных и эффективных вакцинах против инфекций HSV (31, 60). Кроме того, тот факт, что супрессивная терапия HSV приводит к значительному снижению уровня ВИЧ в слизистых половых органов и плазме крови женщин, инфицированных HSV-2 и ВИЧ (52) позволяет предположить, что эффективная вакцина против HSV может иметь большое значение для контроля инфекции ВИЧ (1, 31).
Гликопротеин D HSV-2 (gD2)
Гликопротеин D HSV (gD) представляет собой один из основных вирусных антигенов, экспрессируемых на поверхности инфицированных клеток (21) и на оболочке вируса (24). gD необходим для проникновения вируса в клетку. Он представляет собой мишень для нейтрализующих антител против инфекции HSV (12, 49, 53). Кроме того, gD является основной вирусной мишенью для CD4+ T-клеток, включая цитотоксичность, опосредованную CD4+ Т-клетками, и CD8+ Т-клеток в человеческих и мышиных моделях инфекции HSV (27, 28, 30, 34, 47, 65, 75). В силу вышеизложенных причин, разработка вакцин против субъединиц HSV сосредоточена на gD (32, 60).
В фазе 3 клинических исследований Stanberry et al. показали, что эффективность защиты в результате вакцинации рекомбинантным gD HSV-2 (gD2) в комбинации с адъювантом AS04 от развития генитального герпеса у HSV-серонегативных женщин достигала 73-74% (62). Для мужчин и пациентов, сероположительных по HSV-1, значительной эффективности показано не было. Несмотря на то, что у иммунизированных хозяев были обнаружены gD2-специфичные гуморальный и CD4+ Т-клеточный ответы, остается неясным, может ли gD2/AS04 эффективно индуцировать CD8+ T-клеточный ответ (31, 32). Исходя из этого исследования, можно сделать вывод, что существует потребность в вакцине против HSV, индуцирующей более широкий гуморальный, а также CD4 и CD8 T-клеточный ответы на gD2 и другие вирусные антигены HSV (29, 31, 32).
Вирусные вакцины
Хорошо известно, что живые вирусные вакцины, способные к синтезу иммуногенов de novo в теле хозяина, индуцируют более широкий и более длительный иммунный ответ, чем вакцины, состоящие только из пептидов или белков. Были получены и протестированы на потенциальную пригодность в качестве вакцин против инфекции HSV различные формы HSV, дефектных по репликации и нейро-аттенуированные мутанты, компетентные по репликации (патент США № 7223411; (18)).
Поскольку вирусы, дефектные по репликации, а также нейро-аттенуированные мутанты способны в ходе ко-репликации с вирусом дикого типа или становиться компетентными по репликации в контексте вируса дикого типа, использование их в качестве человеческих вакцин несет определенный риск, в особенности для пациентов, несущих латентную инфекцию HSV (33). Поскольку было показано, что у мутантов HSV-1, дефектных по репликации, может реактивироваться предранний промотор HSV-1 в мозгу грызунов, существует дополнительный риск индукции такими мутантами продуктивной вирусной инфекции у латентно инфицированных пациентов (63). Таким образом, требуемая рекомбинантная вакцина против HSV, дефектная по репликации, должна быть способной синтезировать широкий спектр антигенов, кодируемых вирусом, а также должна кодировать уникальную функцию, предотвращающую литическую инфекцию HSV дикого типа при нахождении в одних и тех же клетках. Такой защитный механизм сведет к минимуму вероятность возникновения инфекции вирусом вакцины в результате рекомбинации вектора вакцины и вируса дикого типа, присутствующего в теле хозяина.
Сущность изобретения
В общем, настоящее изобретения основано на использовании технологии индуцированного тетрациклином включения и выключения генов (T-REx производства Invitrogen) (73) и доминантно-негативной мутантной формы полипептида UL9 HSV-1, например, UL9-C535C, для разработки безопасной и эффективной вирусной вакцины против инфекции HSV-2.
В своем первом аспекте изобретение направлено на дефектный по репликации доминантно-негативный рекомбинантный вирус Herpes simplex 2 (HSV-2). В геноме этого вируса содержится по меньшей мере первая последовательность, кодирующая первый гликопротеин D HSV-2 (gD2), функционально связанная (operably linked) с первым промотором и предпочтительно вторая последовательность, кодирующая второй HSV-2 gD2, функционально связанная со вторым промотором. Промотор (промоторы) функционально связан с последовательностью тетрациклинового оператора (tet-O) и второй последовательностью tet-O, соответственно, каждая из которых разрешает транскрипцию в отсутствие связанного тетрациклинового репрессора, но блокирует транскрипцию при связывании репрессора. Геном также включает третью последовательность, кодирующую по меньшей мере первую доминантно-негативную мутантную форму белка UL9 HSV-1 или HSV-2, под контролем третьего промотора, и предпочтительно четвертую последовательность, кодирующую вторую доминантно-негативную форму белка UL9 HSV-1 или HSV-2, под контролем четвертого промотора. Как и первый и второй промоторы, третий и четвертый промоторы оба функционально связаны с последовательностями tet-O, блокирующими транскрипцию при связывании tet-репрессора. Кроме того, в геноме вируса отсутствует последовательность, кодирующая функциональный белок ICP0. Для усиления антигенности геном также предпочтительно экспрессирует иммуномодулирующие гены, такие как IL12 или IL15 и/или основные антигены HSV-1 или HSV-2, такие как gB или gC.
Термин “функционально связаны” означает, что геномные элементы соединены вместе таким образом, что они могут выполнять свои нормальные функции. Например, ген функционально связан с промотором, если его транскрипция находится под контролем этого промотора и если в результате транскрипции получается продукт, в норме кодируемый геном. Последовательность tet-оператор функционально связана с промотором, если оператор блокирует транскрипцию с промотора в присутствии связанного tet-репрессора, но разрешает транскрипцию в отсутствие репрессора. Термин “рекомбинантный” означает вирус, чьи нуклеотидные последовательности были получены путем рекомбинации нуклеотидных последовательностей и элементов последовательностей и введения этих рекомбинированных последовательностей в вирус или в вирус-предшественник.
Предпочтительно, используемые промоторы являются такими, которые содержат TATA-элемент и последовательности tet-оператора, связанные с промоторами, содержащими два сайта связывания репрессора op2, соединенные связывающими нуклеотидами, число которых может варьировать от 2 до 20. Положение операторной последовательности важно для достижения эффективного контроля промотора. В частности, первый нуклеотид операторной последовательности должен быть расположен на расстоянии от 6 до 24 нуклеотидов к 3'-концу от последнего нуклеотида ТАТА-элемента. Структурные последовательности, кодирующие, например, gD или доминантно-негативный мутантный полипептид UL9, должны находиться к 3'-концу от оператора. В особенности предпочтительные промоторы включают предранний промотор hCMV и предранние промоторы HSV-1 и HSV-2. В особенности предпочтительны промоторы ICP4 HSV-1 или HSV-2.
В другом аспекте изобретение направлено на вакцину, которую можно использовать профилактически или терапевтически против экспрессии HSV и которая включает один или более вышеописанных рекомбинантных вирусов в стандартной дозировке. Термин “стандартная дозировка” означает однократное введение лекарственного препарата, такое как таблетка или капсула. Предпочтительно, “стандартная дозировка” представляет собой раствор лекарственного препарата с концентрацией, вызывающей терапевтический или профилактический эффект при введении выбранного объема (стандартной дозировки) пациенту путем инъекции, который находится во флаконе для инъекций. На основании эффективной дозы для мышей (2×106 БОЕ), можно сделать вывод, что минимальная эффективная доза для человека должна составлять примерно 1×107 БОЕ. Таким образом, стандартная дозировка должна содержать по меньшей мере такое количество вируса, типичная стандартная дозировка должна содержать 1×107-1×109 БОЕ. Вакцины можно хранить в лиофилизированной форме и восстанавливать в фармацевтически приемлемом носителе перед употреблением. Альтернативно, препарат может храниться непосредственно в носителе. Объем одной дозы вакцины может варьировать, но в общем должен находиться в интервале приблизительно от 0,1 мл до 10 мл и более типично приблизительно от 0,2 мл до 5 мл.
Изобретение также включает способы иммунизации пациентов против инфекции HSV-1 или HSV-2 и состояний, к которым приводят такие инфекции (например, герпетические генитальные язвы) путем введения пациентам вышеописанных вакцин. Вакцины могут вводиться инфицированным пациентам для предотвращения или подавления вспышки вируса. Любой способ введения вакцины пациенту, не приводящий к разрушению вируса, совместим с настоящим изобретением. В общем случае, препарат вводят парентеральным способом, таким как внутримышечная или внутривенная инъекция. Дозировку и график введения вакцин можно определить с помощью способов, стандартных для уровня техники. Препараты можно вводить путем однократных или многократных инъекций.
Краткое описание чертежей
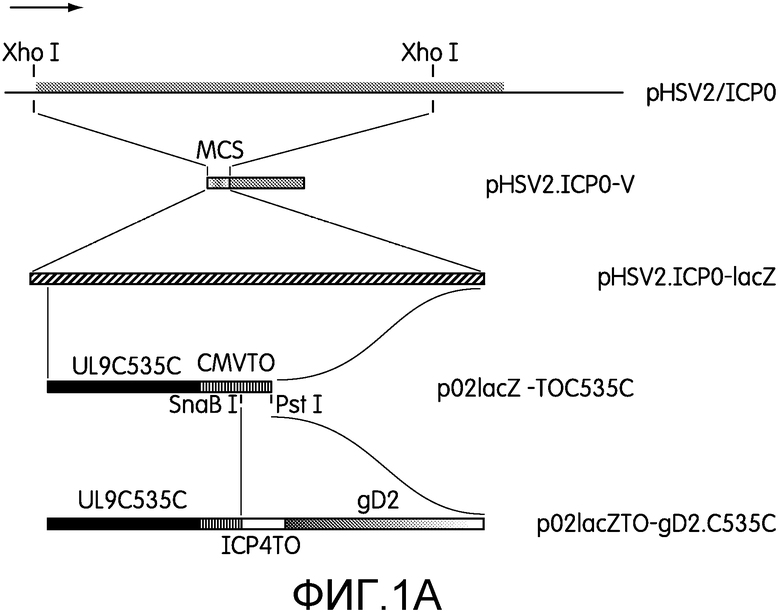
Фиг.1А и 1В: На фиг.1А представлены схематические изображения плазмид, использованных для конструирования доминантно-негативного и дефектного по репликации рекомбинантов HSV-2: N2-C535C и CJ2-gD2. Плазмида pHSV-2/ICP0 представляет собой плазмиду, включающую последовательности HSV-2 ICP0 от 268 п.н. раньше открытой рамки считывания HSV-2 ICP0 (серый прямоугольник) до 40 п.н. после сигнала полиаденилирования последовательностей, кодирующих ICP0. pHSV-2.ICP0-V конструировали путем замещения фрагмента ДНК Xho I-ICP0, содержащего последовательности с множественной клонирующей последовательностью (MCS), включающей Xho I. pHSV-2.ICP0-lacZ получали путем вставки гена LacZ (диагональная штриховка) в область MCS pHSV-2.ICP0-V. p02lacZ-TOC535C конструировали путем замены указанного фрагмента, включающего lacZ, pHSV-2.ICP0-lacZ на последовательности ДНК, кодирующие UL9-C535C (черный прямоугольник) под контролем основного предраннего промотора hCMV, включающего tetO (вертикальная штриховка, CMVTO). p02lacZTO-gD2.C535C конструировали путем замены фрагмента SnaB I/Pst I p02lacZTO-C535C на последовательности ДНК, кодирующие ген gD2 (прямоугольник с градиентной заливкой) под контролем предраннего промотора ICP4 HSV-1, включающего tetO (белый прямоугольник, ICP4TO).
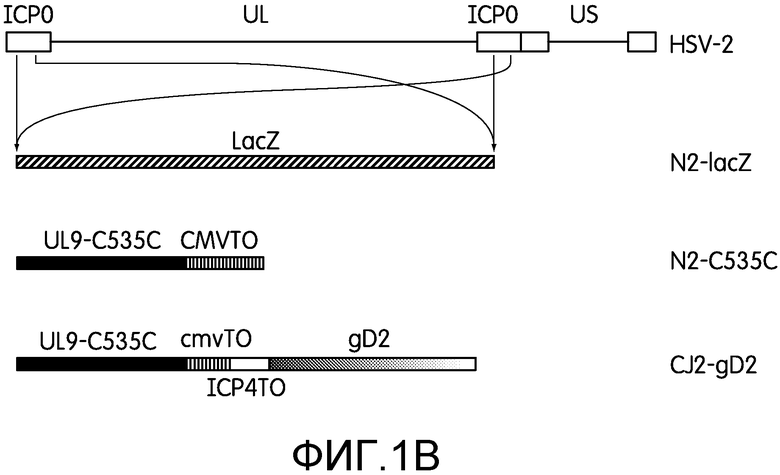
На фиг.1В приведены схематические изображения геномов HSV-2 дикого типа, мутанта HSV-2 ICP0 null (N2-lacZ), N2-C535C и CJ2-gD2. UL и US обозначают уникальную длинную и уникальную короткую область генома HSV-2 соответственно. Эти области фланкированы соответствующими областями обратных повторов (белые прямоугольники). Под развернутыми последовательностями, кодирующими ICP0 генома HSV-2, показана замена обеих копий последовательностей, кодирующих ICP0, на ген lacZ в N2-lacZ и на последовательности ДНК (1) кодирующие UL9-C535C под контролем основного предраннего промотора hCMV, несущего tetO, в N2-535C и (2) кодирующие UL9-C535C и gD2 под контролем указанных промоторов, несущих tetO, в противоположных ориентациях.
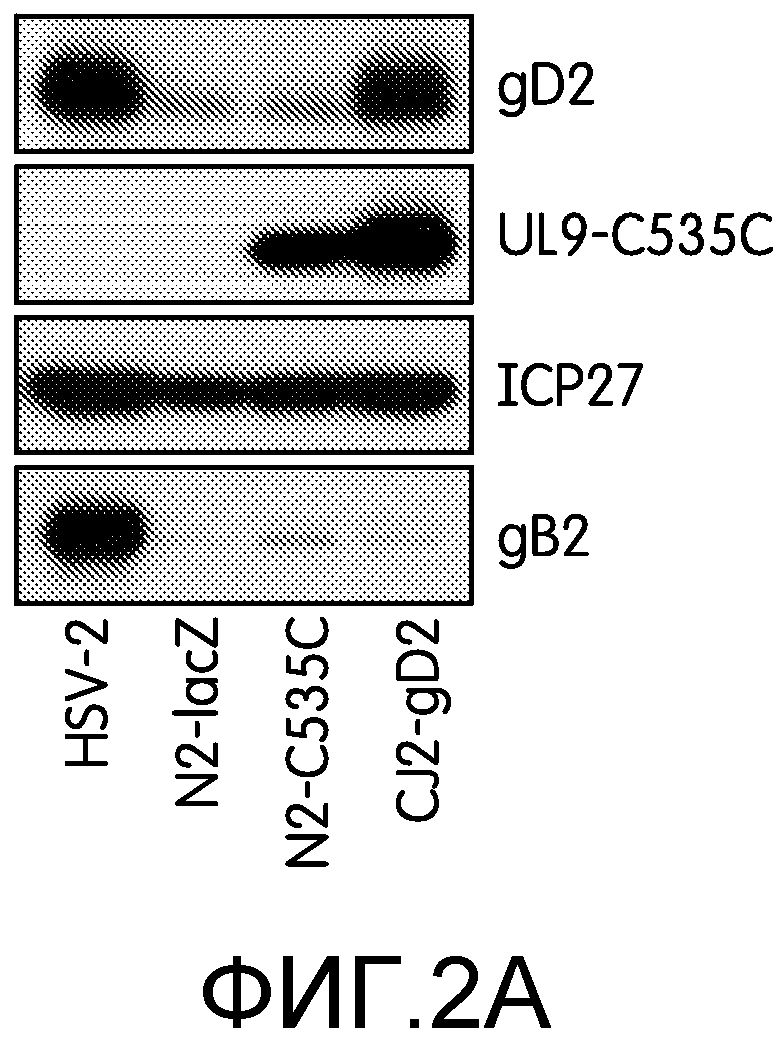
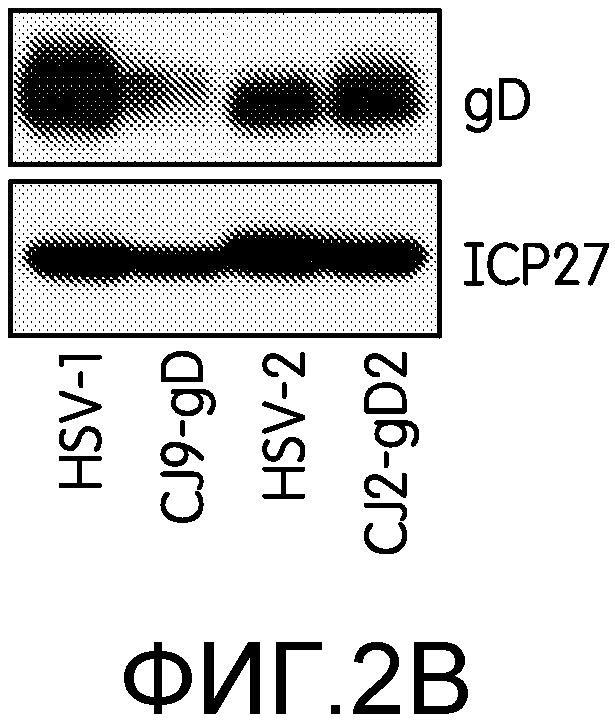
Фиг.2А и 2В: На этих фигурах показан высокий уровень экспрессии gD2 и UL9-C535C после инфекции клеток Vero CJ2-gD2. На фиг.2А клетки Vero не инфицировали или инфицировали HSV-2 дикого типа, N2-lacZ, N2-C535C или CJ2-gD2 со множественностью заражения 10 БОЕ на клетку. Эксперимент повторяли дважды. Экстракты инфицированных клеток готовили через 9 часов после инфекции. На фиг.2В клетки Vero инфицировали штаммом KOS HSV-1 дикого типа, CJ9-gD, HSV-2 дикого типа или CJ2-gD2 со множественностью инфекции 10 БОЕ на клетку. Экстракты инфицированных клеток готовили через 9 часов после инфекции. Белки, содержащиеся в экстрактах инфицированных клеток, разделяли с помощью SDS-PAGE и далее проводили иммуноблоттинг с использованием поликлональных антител к gD HSV-1 (R45), UL9, или моноклональных антител, специфичных к ICP27 и gB (Santa Cruz).
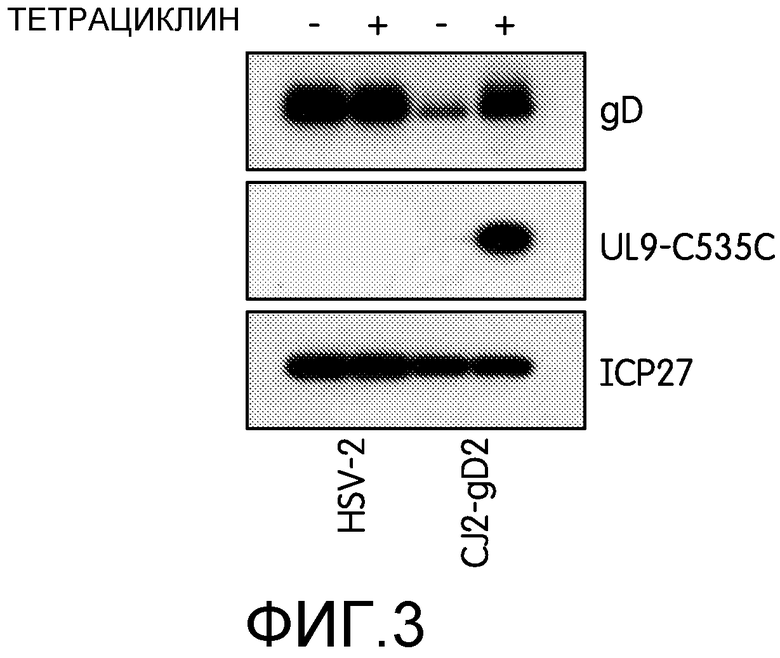
Фиг.3: На фиг.3 показана регуляция экспрессии gD2 и UL9-C535C с помощью tetR в клетках VCEP4R-28, инфицированных CJ2-gD2. Клетки VCEP4R-28 рассаживали в количестве 5×105 клеток на чашку диаметром 60 мм. Через 40 ч после рассадки клетки не инфицировали или инфицировали HSV-2 дикого типа или CJ2-gD2 при множественности инфекции 10 БОЕ на клетку в присутствии или в отсутствие тетрациклина. Эксперимент повторяли дважды. Через 9 часов после инфекции готовили экстракты инфицированных клеток. Экстракты исследовали с помощью иммуноблоттинга поликлональными антителами к gD и UL9 HSV и моноклональными антителами к ICP27.
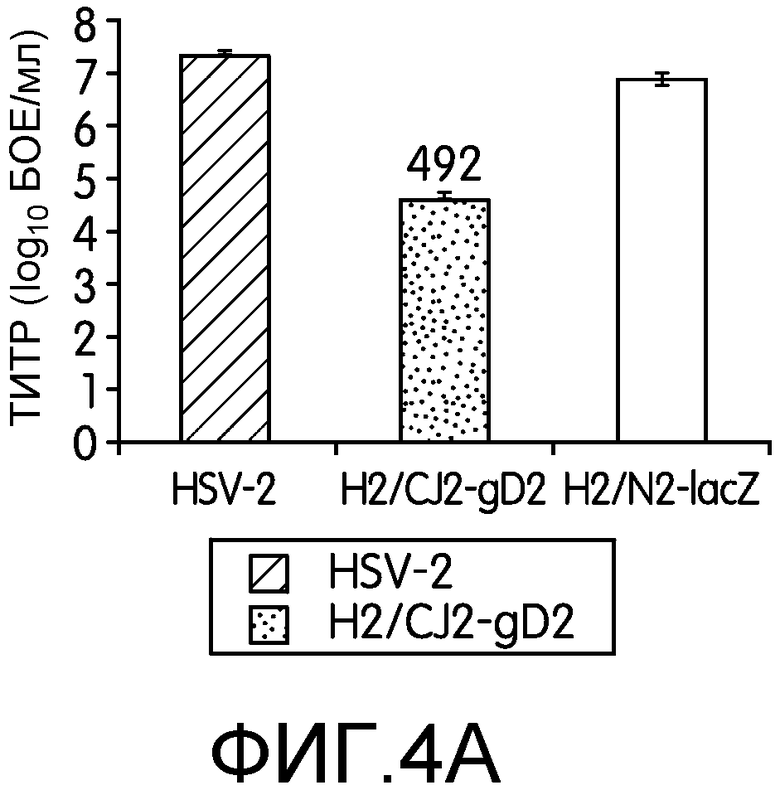
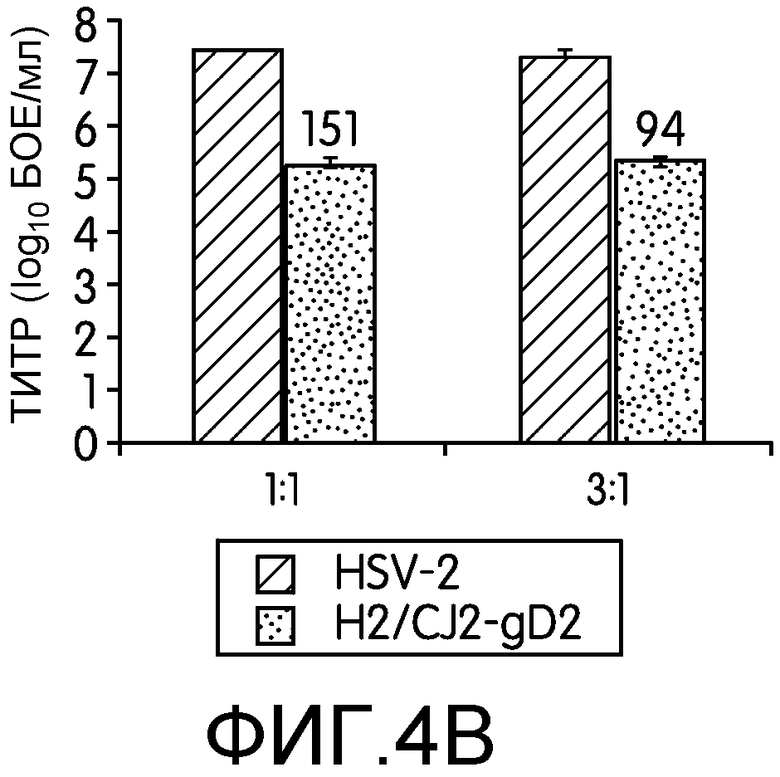
Фиг.4А и 4В: На этих фигурах показан транс-доминантно-негативный эффект CJ2-gD2 на репликацию HSV-2 дикого типа. На фиг.4А клетки Vero инфицировали штаммом 186 HSV-2 дикого типа при множественности инфекции 2 БОЕ на клетку, 186 при множественности инфекции 2 БОЕ на клетку и CJ2-gD2 при множественности инфекции 5 БОЕ на клетку или 186 при множественности инфекции 2 БОЕ на клетку и N2-lacZ при множественности инфекции 5 БОЕ на клетку. Эксперимент повторяли трижды. На фиг.4В клетки Vero инфицировали только HSV-2 дикого типа при множественности инфекции 5 БОЕ на клетку, ко-инфицировали 186 и CJ2-gD2 при множественности инфекции 5 БОЕ на клетку для обоих вирусов или инфицировали только 186 при множественности инфекции 15 БОЕ на клетку и ко-инфицировали 186 при множественности инфекции 15 БОЕ на клетку и CJ2-gD2 при множественности инфекции 5 БОЕ на клетку. Через 18 часов после инфекции собирали инфицированные клетки и определяли титры вирусов на монослоях клеток Vero. Титры вирусов представлены в виде среднее ± стандартное отклонение. Числа сверху графика означают, во сколько раз выход вируса при ко-инфекции меньше выхода вируса при одиночной инфекции.
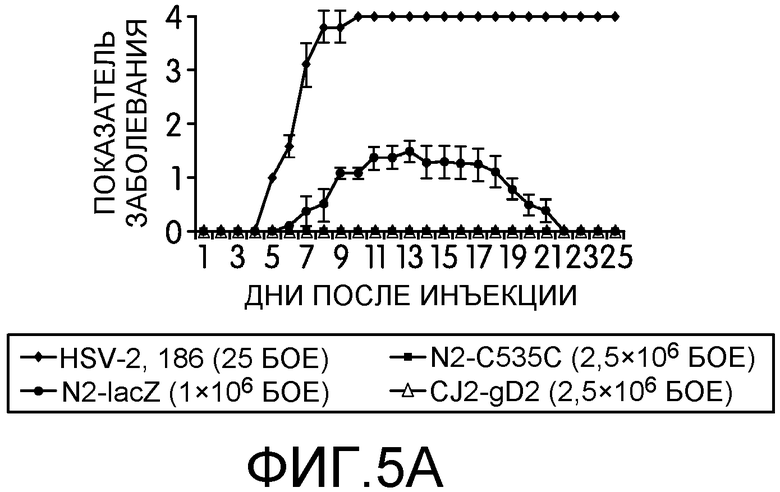
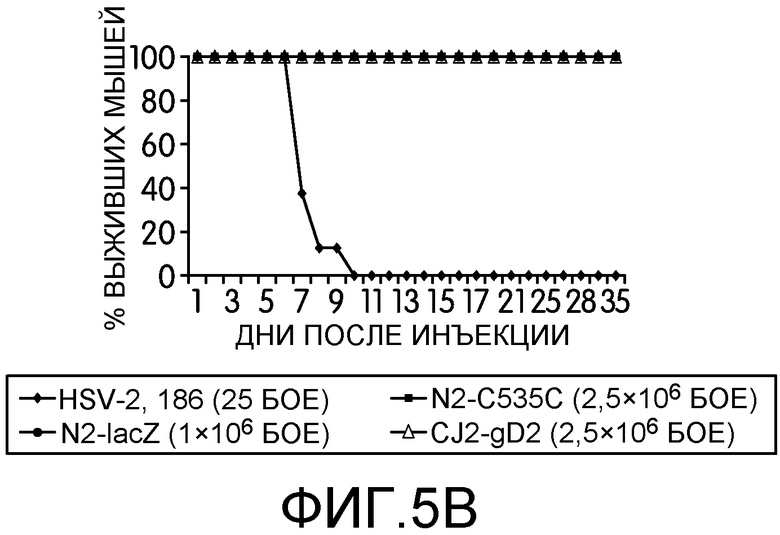
Фиг.5А и 5В: На этих фигурах показана нейровирулентность штамма 186 HSV-2 дикого типа, N2-lacZ, N2-C535C и CJ2-gD2 у мышей BALB/c после интрацеребральной прививки. Самок мышей BALB/c в возрасте 4-6 недель случайным образом разделяли на 5 групп по 8 мышей в каждой. Животных анестезировали пентобарбиталом натрия и прививали DMEM, 25 БОЕ на мышь штамма 186 HSV-2 дикого типа, 1×106 БОЕ на мышь N2-lacZ, 2,5×106 БОЕ на мышь CJ2-gD2 или N2-C535C путем интрацеребральной инъекции в левую лобную долю мозга в объеме 20 мкл на глубину 4 мм. За развитием у мышей признаков и симптомов болезни наблюдали в течение 35 дней после прививки. На фиг.5А приведены показатели заболевания в различные дни после инфекции, а на фиг.5В приведен процент выживших мышей.
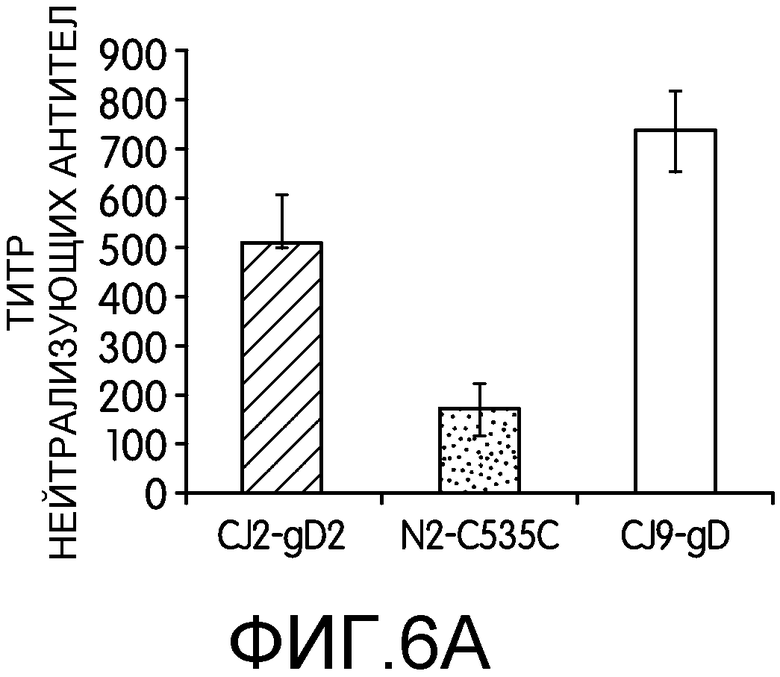
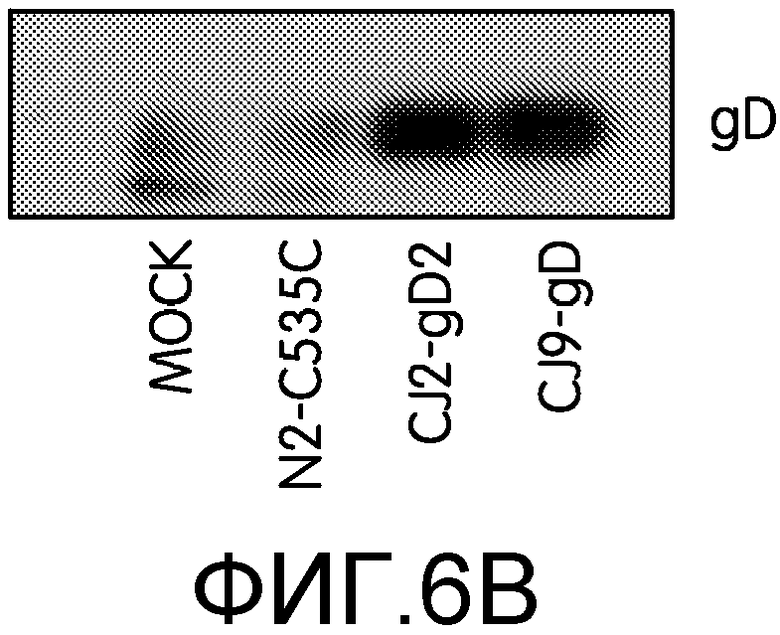
Фиг.6А и 6В: На фиг.6А и 6В показана индукция антител, специфичных к gD2, и ответов, нейтрализующих HSV-2. Самкам мышей BALB/c в возрасте 4-6 недель вводили DMEN (n=7, 6, 8, 8) или иммунизировали CJ2-gD2 (n=7, 6, 8, 8), N2-C535C (n=7, 8, 6) или CJ9-gD (n=6, 8, 6) в дозировке 2×106 БОЕ на мышь, и повторяли иммунизацию через 2 недели. Через 4-5 недель после первичной иммунизации у мышей забирали кровь из хвостовой вены. На фиг.6А сыворотку мышей из отдельной группы объединяли и инактивировали нагреванием. Определяли титры специфических антител, нейтрализующих HSV-2. Результаты приведены в виде средних титров ± стандартная ошибка среднего. На фиг.6В сыворотку контрольных мышей и мышей, иммунизированных CJ2-gD2, N2-C535C или CJ9-gD, инкубировали с клеточным экстрактом, полученным из клеток U2OS, трансфицированных плазмидой p02.4TO-gD2, экспрессирующей gD2. Комплексы gD со специфическими мышиными иммуноглобулинами преципитировали с помощью белка А, разделяли с помощью SDS-PAGE и детектировали поликлональными антителами R45, специфичными к gD.
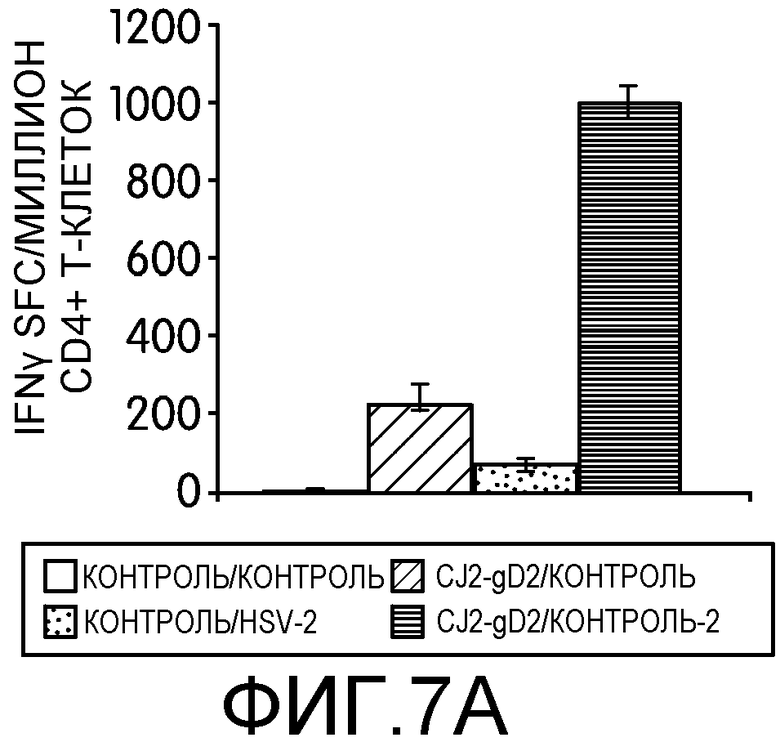
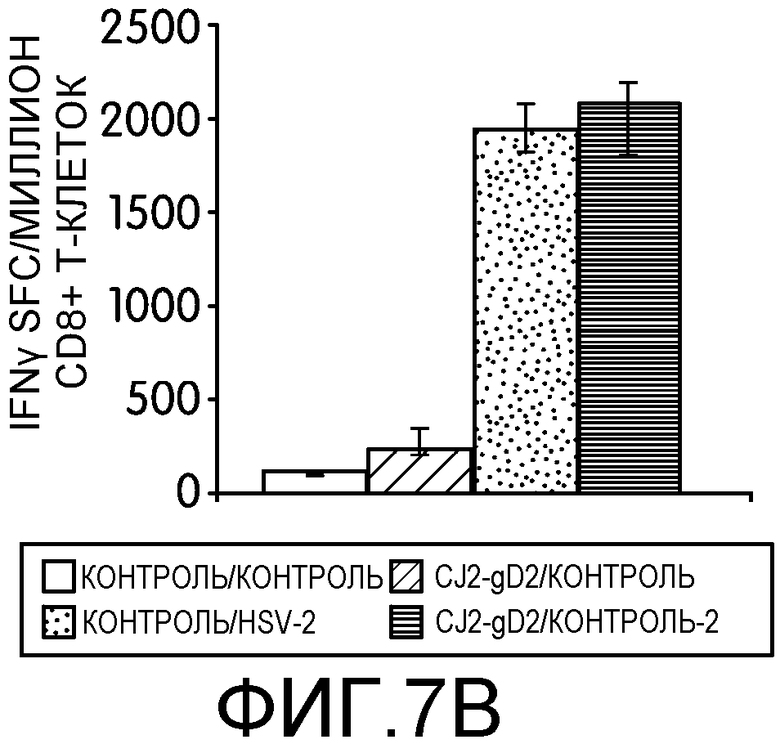
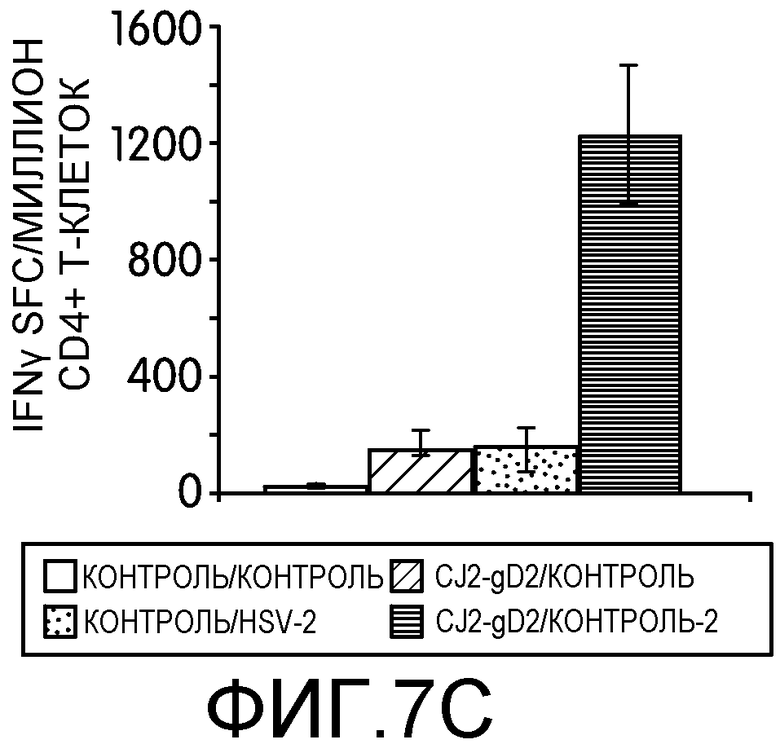
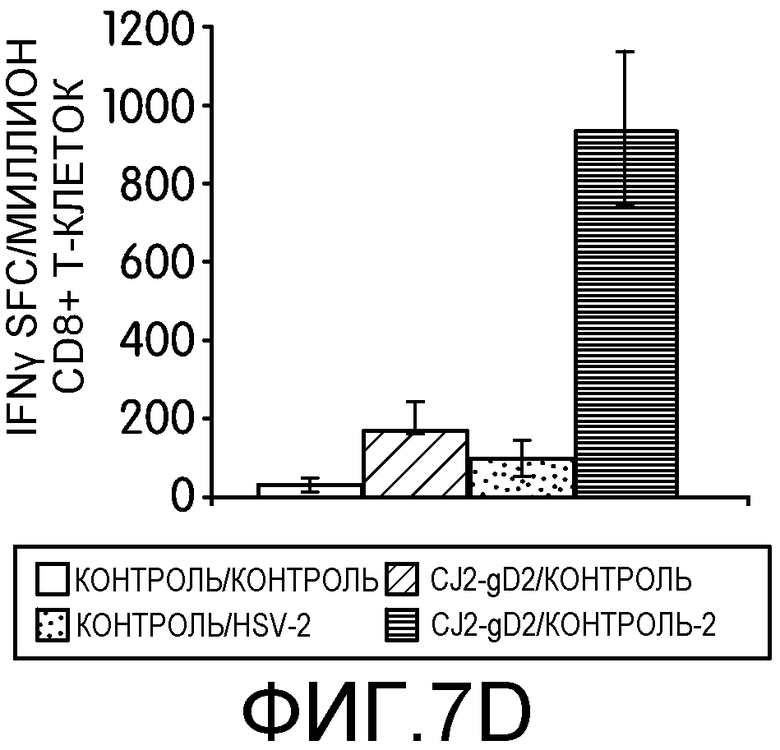
Фиг.7А-7D: На этих фигурах показана индукция CD4+ и CD8+ Т-клеточных ответов, специфичных к HSV-2, у мышей, иммунизированных CJ2-gD2. Самок мышей BALB/c не иммунизировали или иммунизировали CJ2-gD2 при множественности инфекции 2×106 БОЕ на мышь дважды с промежутком времени 2 недели. На фиг.7А и 7В контрольных и иммунизированных мышей не инфицировали или инфицировали HSV-2 дикого типа подкожно в дозировке 1×104 БОЕ на мышь через 9-10 недель после вторичной иммунизации (n=3). CD4+ и CD8+ T-клеточные ответы анализировали на 5 день после нагрузки с помощью тестов IFNγ ELISPOT на индивидуально очищенных CD4+ и CD8+ Т-клетках, выделенных из селезенки мышей, с использованием наборов реагентов Dynal для выделения мышиных CD4- и CD8- Т-клеток. На фиг.7C и 7D контрольных мышей и мышей, иммунизированных CJ2-gD2, не инфицировали или инфицировали HSV-2 дикого типа через 5-6 недель после вторичной иммунизации и далее проводили тесты IFNγ ELISPOT через 4 дня после инфекции (n=3). Число клеток, образующих пятна IFNγ (spot-forming cells, SFC) выражали в виде среднего ± стандартная ошибка среднего на миллион CD4+ или CD8+ Т-клеток.
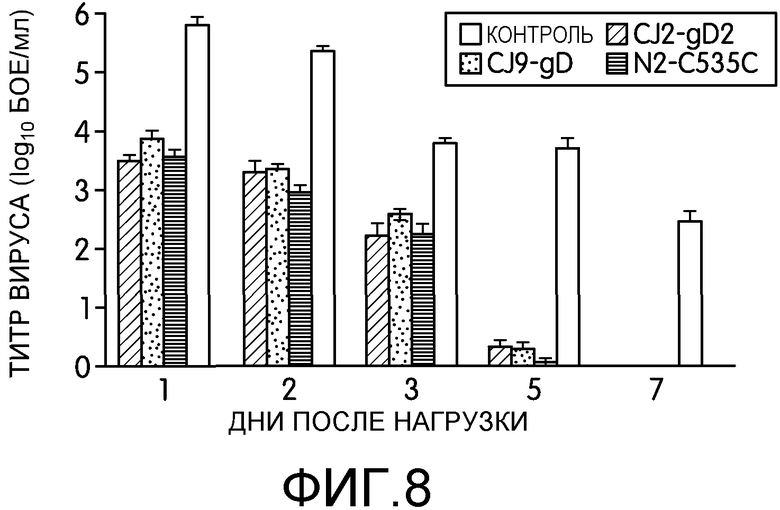
Фиг.8: На фиг.8 показано ослабление вагинальной репликации нагрузочного HSV-2 у мышей, иммунизированных CJ2-gD2. Самок мышей BALB/c в возрасте 4-6 недель случайным образом разделяли на 4 группы по 10 мышей в каждой. Мышам BALB/c в возрасте 4-6 вводили DMEM или иммунизировали CJ2-gD2, N2-C535C или CJ9-gD в дозировке 2×106 БОЕ на мышь, и вакцинацию повторяли через 2 недели. Через 5 недель мышам давали медроксипрогестерон и нагружали интравагинально 5×105 БОЕ штамма G HSV-2. Вагинальные мазки брали на 1, 2, 3, 5 и 7 день после нагрузки. Инфекционность вирусов в материале мазков оценивали по стандартному тесту образования бляшек на монослое клеток Vero. Титры вирусов приведены в виде среднего ± стандартная ошибка среднего в индивидуальных вагинальных мазках.
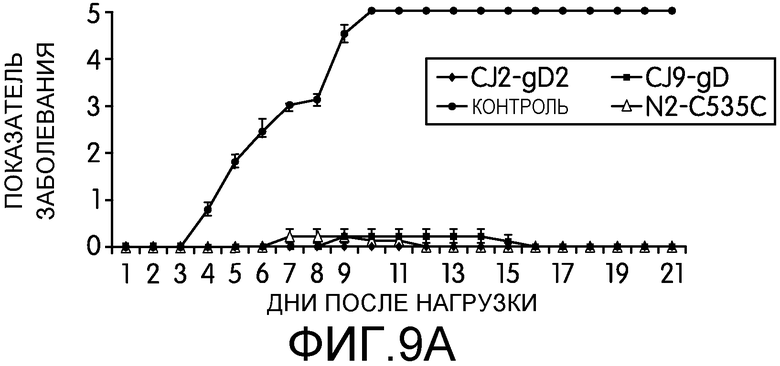
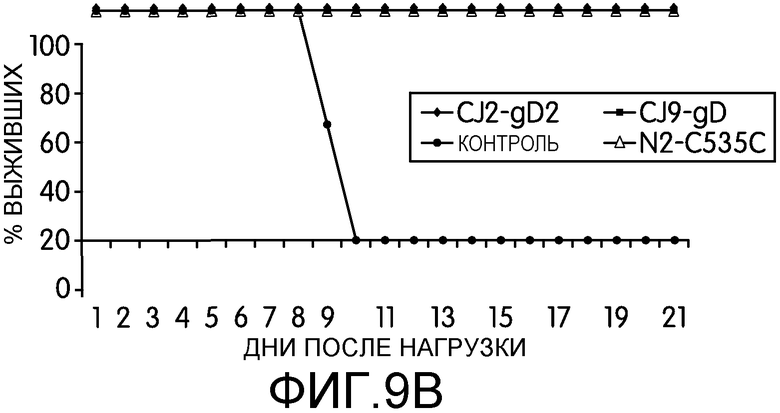
Фиг.9А и 9В: На этих фигурах показано предотвращение заболевания HSV-2 у мышей, иммунизированных CJ2-gD2. После нагрузки HSV-2 дикого типа у индивидуальных мышей, описанных в подписи к фиг.8, наблюдали в течение 21-дневного срока, наблюдали частоту генитального и системного заболевания HSV-2 (фиг.9) и выживаемость (фиг.9В), используя следующую шкалу: 0 = признаки не выражены, 1 = легкое покраснение и отек гениталий, 2 = умеренное воспаление гениталий, 3 = гнойные язвы гениталий и/или системное заболевание, 4 = паралич задних конечностей, 5 = гибель.
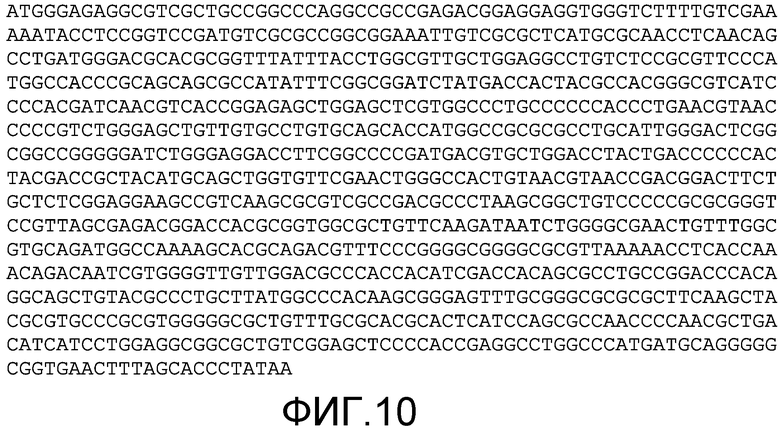
Фиг.10: На фиг.10 показана кодирующая последовательность UL9-C535C HSV-1 (SEQ ID NO:2). UL9-C535C состоит из аминокислот 1-10 UL9, трипептида Thr-Met-Gly и аминокислот 535-851 UL9 (смотрите Yao et al. (69)).
Подробное описание изобретения
Настоящее изобретение основано на идее использования технологии включения/выключения генов с помощью тетрациклина, а также доминантно-негативного пептида UL9 HSV-1 с целью получения рекомбинантного вируса HSV, дефектного по репликации и способного ингибировать инфекции HSV дикого типа (доминантно-негативного). CJ9-gD является прототипным доминантно-негативным дефектным по репликации рекомбинантным вирусом HSV-1 с высоким уровнем экспрессии основного антигена HSV-1 - гликопротеина D (gD), независимым от репликации вирусной ДНК HSV (7). В наиболее предпочтительной форме настоящего изобретения используется доминантно-негативный дефектный по репликации рекомбинантный HSV-2 (CJ2-gD2), кодирующий 2 копии гена gD HSV-2 (gD2) под контролем основного предраннего промотора ICP4 HSV-1, несущего tetO. CJ2-gD2 экспрессирует gD2 также эффективно, как и вирус HSV-2 дикого типа, и способен оказывать сильное транс-ингибирующее воздействие на репликацию HSV-2 дикого типа при ко-инфекции в одних и тех же клетках. Иммунизация CJ2-gD2 приводит к выработке эффективных нейтрализующих антител, специфичных к HSV-2, а также индукции Т-клеточных ответов, и обеспечивает полную защиту от интравагинальной инфекции HSV-2 дикого типа у мышей.
CJ2-gD2 представляет собой вакцину, более эффективно защищающую от генитальной инфекции HSV-2 и вызванного им заболевания, чем CJ9-gD. Кроме того, интрацеребральная инъекция большой дозы CJ2-gD2 не приводит к заболеваемости или смертности у мышей. Исходя из совокупности этих наблюдений, можно предположить, что CJ2-gD2 обладает определенными преимуществами перед традиционными вакцинами на основе вируса, дефектного по репликации, и вакцинами на основе субъединиц HSV-2 в защите против генитальной инфекции HSV-2 и вызванного им заболевания у человека.
Система включения/выключения генов Tet оператор/репрессор и рекомбинантная ДНК
Настоящее изобретение направлено inter alia на вирусы, содержащие гены, экспрессию которых регулируется tet-оператором и белком-репрессором. Способы, используемые для получения рекомбинантных молекул ДНК, содержащих эти элементы и последовательности ДНК, были описаны ранее (смотрите патент США № 6444871; патент США № 6251640; и патент США № 5972650), а плазмиды, содержащие индуцируемый тетрациклином переключатель транскрипции, имеются в продаже (T-RExTM производства Invitrogen, США).
Существенным признаком ДНК согласно настоящему изобретению является присутствие генов, функционально связанных с промотором, предпочтительно включающим TATA-элемент. Последовательность tet-оператор располагается между 6 и 24 нуклеотидом к 3'-концу от последнего нуклеотида ТАТА-элемента промотора и к 5'-концу от гена. Для блокирования транскрипции генов и разрешения репликации вируса вирус можно растить в клетках, экспрессирующих tet-репрессор. Сила, с которой tet-репрессор связывается с операторной последовательностью, увеличивается при использовании формы оператора, включающей два сайта связывания рецептора op2 (каждый сайт имеет нуклеотидную последовательность TCCCTATCAGTGATAGAGA (SEQ ID NO:1)), разделенных последовательностью длиной 2-20, предпочтительно 1-3 или 10-13 нуклеотидов. При связывании репрессора с этим оператором транскрипции ассоциированного гена не происходит, или она очень слаба. Если ДНК с такими характеристиками присутствует в клетке, также экспрессирующей тетрациклиновый репрессор, транскрипция гена, который может предотвратить вирусную инфекцию, будет блокироваться репрессором, связывающимся с оператором, и таким образом будет иметь место репликация вируса.
Выбор промоторов и генов
Во время продуктивной инфекции экспрессию генов HSV разделяют на три основных класса в зависимости от времени экспрессии: различают предранние (α), ранние (β) и поздние (γ) гены, причем поздние гены далее разделяют на две группы, γ1 и γ2. Экспрессия предранних генов не требует синтеза вирусных белков de novo и активируется белком VP16, ассоциированным с вирионом, совместно с клеточными факторами транскрипции при попадании вирусной ДНК в ядро. Белковые продукты предранних генов обозначаются ICP0, ICP4, ICP22, ICP27 и ICP47 (от infected cell polypeptides, полипептиды инфицированных клеток); промоторы этих генов предпочтительно используют для контроля экспрессии рекомбинантных генов, обсуждаемых в настоящей заявке.
ICP0 играет важную роль в усилении реактивации HSV из латентного состояния и дает HSV значительное преимущество в росте при низкой множественности инфекции. ICP4 представляет собой основной белковый регулятор транскрипции HSV-1, активирующий экспрессию ранних и поздних генов. ICP27 требуется для продуктивной вирусной инфекции и необходим для эффективной репликации вирусной ДНК и оптимальной экспрессии вирусных генов γ и части вирусных генов β. Считается что функция ICP47 при инфекции HSV заключается в снижении экспрессии белков основного комплекса тканевой совместимости (MHC) I класса на поверхности инфицированных клеток.
Полноразмерная последовательность геномной последовательности HSV-1 области, кодирующей UL9-C535C HSV-1, приведена на фиг.10 (SEQ ID NO:2). Из области техники известны другие последовательности, чье использование в рекомбинантных вирусах было описано ранее. Например, полноразмерная геномная последовательность HSV-1 зарегистрирована в GenBank за номером X14112. Последовательность ICP4 HSV-1 зарегистрирована в GenBank за номером Х06461; последовательность гликопротеина D HSV-1 зарегистрирована в GenBank за номером J02217; последовательность гликопротеина D HSV-2 зарегистрирована в GenBank за номером К01408; и последовательность гена UL9 HSV-1 зарегистрирована в GenBank за номером М19120 (все последовательности полностью включены в настоящую заявку путем ссылки).
Включение tet-репрессора и создание вируса
Последовательности промоторов ICP0 и ICP4 HSV, а также последовательности генов, находящихся под их эндогенным контролем, хорошо известны из уровня техники (43, 44, 56). Процедуры создания вирусных векторов, содержащих эти элементы, также были описаны ранее (смотрите опубликованную заявку на патент США № 2005-0266564). Эти промоторы не только очень активно индуцируют экспрессию генов, но сами специфически индуцируются трансактиваторным белком VP16, высвобождающимся при инфекции клетки HSV-1 или HSV-2.
После получения необходимых конструктов ДНК их можно инкорпорировать в вирус HSV-2, используя способы, хорошо известные из уровня техники (смотрите Yao et al. (68)).
Способы иммунизации
Вирусы, описанные в настоящей заявке, используют для иммунизации пациентов, как правило, путем инъекции, в качестве вакцины. Вакцина может использоваться профилактически для предотвращения инфекции HSV-1 или HSV-2 или терапевтически для ослабления уже существующей инфекции HSV-1 или HSV-2. Для получения вакцины вирусы можно суспендировать в любом фармацевтически приемлемом растворе, включая стерильный физиологический раствор, воду, физиологический раствор на фосфатном буфере, 1,2-пропиленгликоль, полигликоли, смешанные с водой, раствор Рингера и т.д. Точное количество вводимого вируса не является решающим для изобретения, однако должно представлять собой "эффективное количество", т.е. количество, достаточное для индукции иммунологического ответа, достаточно сильного для подавления инфекции HSV. В общем, ожидается, что количество первично вводимого вируса (БОЕ) будет находиться в интервале от 1×107 до 1×1010.
Эффективность дозировки, а также общую эффективность лечения можно оценить, используя стандартные иммунологические способы тестирования на присутствие антител, эффективно борющихся с HSV. Иммунологические инъекции можно повторять требуемое количество раз.
Примеры
В настоящем примере описывается создание рекомбинантного вируса HSV-2 и тесты на определение его иммунологических эффектов.
1. Материалы и способы
Клетки
Клетки почки африканской зеленой мартышки (Vero) и клетки линии остеосаркомы U2OS культивировали в среде Игла, модифицированной Дульбекко (DMEM, производство Sigma Aldrich) с добавлением 10% эмбриональной телячьей сыворотки (FBS) в присутствии 100 ед/мл пенициллина G и 100 мкг/мл стрептомицина сульфата (GIBCO, Карлсбад, штат Калифорния, США) (71). Клетки U2OS способны к функциональной комплементации делеции ICP0 HSV-1 (71). Клетки U2CEP4R11, которые представляют собой клетки U2OS, экспрессирующие tetR, культивировали в DMEM с добавлением 10% FBS и гигромицина B в концентрации 50 мкг/мл (73). Клетки VCEP4R-28, представляющие собой клетки Vero, экспрессирующие tetR, культивировали в DMEM с добавлением 10% FBS и гигромицина B в концентрации 50 мкг/мл (73).
Плазмиды
Плазмида pHSV2/ICP0 представляет собой производную плазмиды pUC19, кодирующую последовательности ICP0 HSV-2, амплифицированные с помощью PCR, от 268 п.н. выше открытой рамки считывания (ORF) ICP0 до 40 п.н. ниже сигнала полиаденилирования последовательностей, кодирующих ICP0. pHSV2.ICP0-V представляет собой плазмиду, клонирующую ICP0 HSV-2, полученную из плазмиды pHSV-2/ICP0 путем замены фрагмента ДНК Xho I-ICP0, содержащего последовательности от 25 п.н. выше кодона инициации ICP0 до 397 п.н. выше стоп-кодона ORF ICP0, на множественную клонирующую последовательность (MCS), содержащую Xho I. Плазмида pHSV2.ICP0-lacZ была получена путем вставки фрагмента HindIII-Not I-, содержащего ген LacZ pcDNA3-lacZ, в pHSV2.ICP0-V по сайтам Hind III-Not I. Плазмида pcmvtetO-UL9C535C представляет собой плазмиду, кодирующую UL9-C535C под контролем предраннего промотора hCMV, содержащего tetO (68). Плазмиду p02lacZ-TOC535C, экспрессирующую UL9-С535С под контролем основного предраннего промотора hCMV, содержащего tetO (фиг.1А), конструировали путем замены фрагмента pHSV2.ICP0-lacZ EcoR I/Age I, содержащего lacZ, на фрагмент pcmvtetOUL9-C535C EcoR I/Hind III, содержащий hcmvtetO-UL9C535C (69).
pAzgD-HSV-2 представляет собой плазмиду, кодирующую gD2 HSV-2, любезно предоставленную д-ром Patricia Spear (Northwestern University). pICP4TO-hEGF экспрессирует человеческий эпидермальный фактор роста под контролем предраннего промотора ICP4 HSV-1, несущего tetO, состоящего из последовательности промотора ICP4 HSV-1 от -377 п.н. до -19 п.н. относительно сайта начала транскрипции гена ICP4. Также, как и основной предранний промотор hCMV, несущий tetO, в плазмиде pcmvtetO-hEGF (73), промотор ICP4, содержащий tetO, включает две тандемные копии tet-операторов на 10 п.н. выше ТАТА-элемента ICP4, TATATGA. Таким образом, как и pcmvtetO-hEGF, pICP4TO-hEGF можно жестко регулировать с помощью тетрациклина в присутствии tetR, но вставка tetO не оказывает никакого влияния на активность промотора ICP4 в отсутствие tetR. Промотор ICP4, несущий tetO, в pICP4TO-hEGF имеет еще одно уникальное свойство - у него отсутствует ДНК-связывающая последовательность ICP4 (ATCGTCCACACGGAG (SEQ ID NO:3), включающая сайт инициации транскрипции гена ICP4 (51) в промоторе ICP4 дикого типа. Таким образом, в отличие от промотора ICP4 дикого типа, авто-регулирующегося ICP4 (16, 57), промотор ICP4, несущий tetO, в pICP4TO-hEGF не супрессируется основным регуляторным белком HSV-1 ICP4.
Чтобы клонировать gD2 под контроль промотора ICP4, содержащего tetO, заявители вначале сконструировали плазмиду p02ICP4-TO путем клонирования Sma I-Bam HI промотора ICP4, содержащего tetO, из pICP4TO-hEGF в pHSV2.ICP0-V в MCS вектора. p02.4TO-gD2 представляет собой плазмиду, производную p02ICP4-TO, кодирующую ген gD2 pAzgD-HSV-2 под контролем промотора ICP4, содержащего tetO.
p02lacZTO-gD2.С535С, плазмиду, кодирующую UL9-C535C под контролем предраннего промотора hCMV, содержащего tetO, с 5'-укорочением на -236 п.н. от промотора hCMV, и ген gD2 под контролем промотора tetO-ICP4 (фиг.1А), получали путем замены фрагмента SnaB I/Pst I p02lacZTO-С535С на фрагмент Hind III/Pst I p02.4TO-gD2, содержащий gD2. В p02lacZTO-gD2.С535С транскрипция гена UL9-С535С и гена gD2 происходит в противоположных направлениях.
Вирусы
HSV-2 дикого типа штаммов 186 и G размножали и оценивали по бляшкам на клетках Vero. N2-lacZ представляет собой мутант HSV-2 ICP0 null, кодирующий ген Lac Z под контролем промотора ICP0 HSV-2, где обе копии гена ICP0 заменены на ген Lac Z в pHSV2.ICP0-lacZ путем гомологичной рекомбинации с помощью трансфекции клеток U2OS линеаризованной Nhe I pHSV2.ICP0-lacZ и последующей суперинфекцией HSV-2 как описано ранее (74). Замену гена ICP0 на ген Lac Z в локусе ICP0 подтверждали с помощью PCR-анализа ДНК вируса N2-lacZ с праймерами, фланкирующими ген ICP0, и праймерами, специфичными для гена lac Z (41, 74).
N2-C535C представляет собой производное N2-lacZ, где обе копии гена Lac Z заменены на последовательности ДНК, кодирующие UL9-C535C под контролем промотора hCMV, содержащего tetO, в плазмиде p02lacZ-TOC535C (фиг.1В). Вкратце, клетки U2CEP4R11 ко-трансфецировали линеаризованной p02lacZ-TOC535C и инфекционной ДНК вируса N2-lacZ с помощью Lipofectamine 2000. Трансфицированные клетки скринировали на рекомбинационную замену генов lacZ N2-lacZ на последовательность ДНК, содержащую cmvtetOUL9-C535C с помощью стандартных тестов на образование бляшек. Бляшки окрашивали 5-бром-4-хлор-3-индолил-b-D-галактопиранозидом (X-Gal) через 96 часов после инфекции. Выделяли белые бляшки, означающие замену обеих копий гена lacZ на последовательность ДНК, кодирующую UL9-C535C. Через четыре раунда очистки бляшек удалось получить один изолят, обозначенный N2-C535C, дающий однородные белые бляшки.
CJ2-gD2 конструировали путем замены обеих копий гена Lac Z в локусе ICP0 в N2-lacZ на последовательности ДНК, кодирующие UL9-C535C под основным предранним промотором hCMV, несущим tetO, и gD2 под контролем промотора ICP4 HSV-1, содержащим tetO (фиг.1В), который состоит из последовательности промотора ICP4 HSV-1 от -377 п.н. до -19 п.н. относительно сайта начала транскрипции гена ICP4 (71).
SDS-PAGE и Вестерн-блот-анализ
Клетки Vero, рассаженные в 60-мм чашки в количестве 7,5×105 клеток на чашку, не инфицировали или инфицировали указанными вирусами со множественностью инфекции 10 БОЕ на клетку. Экстракты клеток готовили через 9 ч или 16 после инфекции (72). Белки экстрактов клеток разделяли с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) (9% акриламида), переносили на поливинилиден-дифторидные (PVDF) мембраны и детектировали с помощью поликлональных антител к gD HSV-1 (R54, любезно предоставлены д-рами Gary H. Cohen и Roselyn J Eisenberg), UL9 (любезно предоставлены Mark Challberg) или моноклональными антителами, специфичными к ICP27 и gB (Santa Cruz Biotechnology, Санта Круз, штат Калифорния, США).
Мыши
Самок мышей BALB/c в возрасте 4-6 недель приобретали у Charles River Laboratories (Уилмингтон, штат Массачусетс, США). Мышей содержали в металлических клетках по 4 мыши на клетку в условиях 12-часового цикла свет/темнота. Мыши акклиматизировались к условиям содержания в течение 1 недели до начала эксперимента. Все эксперименты на животных проводили в соответствии с протоколом, одобренным постоянным комитетом по животным Harvard Medical Area и Американской ассоциацией ветеринарных врачей.
Иммунизация и нагрузки
Мышей BALB/c случайным образом распределяли по нескольким группам. Животным выстригали шерсть на левом боку. Мышей вакцинировали 2×106 БОЕ на мышь CJ2-gD2, N2-C535C, CJ9-gD или вводили DMEM в объеме 30 мкл под кожу левого бока сзади с использованием 1-мл шприца и иглы 27G. Мышей вторично иммунизировали через 2 недели и нагружали HSV-2 дикого типа штамма G через 3 недели после вторичной иммунизации. За 5 дней до нагрузки мышам инъецировали подкожно в загривок медроксипрогестерон (SICOR Pharmaceuticals, Inc., Ирвин, штат Калифорния, США) в количестве 3 мг на мышь в объеме 20 мкл (7, 50). Для интравагинальной нагрузки мышей всех групп анестезировали, наносили альгинат кальция с помощью стерильного уретро-генитального аппликатора с альгинатом кальция производства Puritan Medical Products company LLC, Гилфорд, штат Мэн, США) и прививали интравагинально по 20 мкл культуральной среды, содержащей 5×105 БОЕ (50 LD50) HSV-2 штамма G (50). Животные оставались под анестезией в положении на спине с приподнятым задним концом тела в течение 30-45 минут после инфекции.
Анализ острой инфекции и клинические наблюдения
На 1, 2, 3, 5 и 7 день после нагрузки брали мазок слизистой оболочки влагалища с альгинатом кальция (7). Инфекционность вирусов в материале мазков оценивали с помощью стандартного теста на образования бляшек на монослое клеток Vero. После нагрузки HSV-2 дикого типа у мышей в течение 21-дневного срока наблюдали частоту возникновения генитальных язв и системного заболевания. Для оценки тяжести заболевания использовали следующую шкалу: 0 = признаки герпетической инфекции не выражены, 1 = легкое покраснение и отек гениталий, 2 = умеренное воспаление гениталий, 3 = гнойные язвы гениталий и/или системное заболевание, 4 = паралич задних конечностей, 5 = гибель (8, 50).
Детекция нейтрализующих антител, специфичных к HSV-2
Через 4 недели после первичной иммунизации у иммунизированных и контрольных мышей забирали кровь из хвостовой вены. Титры нейтрализующих антител в сыворотке крови определяли как описано ранее в присутствии комплемента (5-7) с 250 БОЕ HSV-2 дикого типа штамма 186. Титр нейтрализующих антител выражался в виде конечного серийного разведения, достаточного для снижения БОЕ HSV на 50% относительно БОЕ HSV, полученного в среде с комплементом.
Иммунопреципитация
Клетки U2OS рассаживали в количестве 7,5×106 клеток на 100-мм чашку и трансфицировали 10 мкг p02.4TO-gD с помощью Lipofectamine 2000 через 24 часа после рассадки. Экстракты клеток готовили через 48 часов после трансфекции (72). Иммунопреципитации проводили путем смешивания 10 мкл объединенной сыворотки, забранной у контрольных и иммунизированных мышей, с 70 мкл вышеописанных экстрактов клеток. Комплексы gD со специфическим мышиными IgG преципитировали с помощью белка А (Pierce Classic IP kit производства Pierce Biotechnilogy, Рокфорд, штат Иллинойс, США), разделяли с помощью SDS-PAGE и детектировали с помощью поликлональных кроличьих антител к gD, R45 и далее с помощью реакции с козьими антителами к IgG кролика, конъюгированными с HRP (Santa Cruz Biotechnology, Santa Cruz, штат Калифорния, США).
Тест IFNγ-ELISPOT
Самок мышей BALB/c иммунизировали CJ2-gD2 в дозировке 2×106 БОЕ на мышь или вводили DMEM дважды с промежутком в 2 недели. Через 5-10 недель после второй иммунизации контрольных и иммунизированных CJ2-gD2 мышей нагружали HSV-2 дикого типа штамма 186 подкожно в дозировке 1×104 БОЕ на мышь или ставили контроль нагрузки. У отдельных групп мышей (n=3) выделяли спленоциты на 4 или 5 день после нагрузки. Реакцию ELISPOT на клетках CD4+ и CD8+ проводили как описано ранее (42). Вкратце, CD4+ и CD8+ Т-клетки выделяли из спленоцитов с использованием набора реагентов Dynal для выделения мышиных CD4- и CD8- и рассаживали в 96-луночный фильтрационный планшет, предварительно покрытый моноклональными антителами, специфичными к мышиному IFNγ (AN18) в количестве 7,5×104 или 1,5×105 на лунку. Эксперимент повторяли четыре раза. После инкубации при 37ºC в течение 20 ч планшеты промывали, добавляли биотинилированные моноклональные антитела, специфичные к IFNγ (R4-6A2 производства Mabtech) при комнатной температуре и инкубировали со стрептавидином, конъюгированным со щелочной фосфатазой (Mabtech). Клетки, образующие пятна IFNγ, детектировали путем добавления субстрата BCIP/NBT. Пятна подсчитывали под препаровальной лупой; число клеток, образующих пятна IFNγ (SFC) выражали в виде среднего ± стандартная ошибка среднего на миллион CD4+ или CD8+ Т-клеток.
Количественная PCR в реальном времени
Через 16 дней после вторичной иммунизации или через 21 день после интравагинальной нагрузки 5×105 БОЕ HSV-2 штамма G у 9 или 10 мышей, иммунизированных CJ2-gD2 или CJ9-gD, забирали нижнюю поясничную и копчиковую часть позвоночника, включая спинной мозг и дорсальные корешковые ганглии. Позвоночник разрезали на 4 части, части помещали по отдельности в 0,5 мл нормальной питательной среды и замораживали при -80ºС для дальнейшей обработки. Тотальную ДНК выделяли из каждого дорсального корешкового ганглия с использованием набора реагентов Dneasy tissue kit (Quiagen, Санта-Кларита, штат Калифорния, США) и суспендировали в 400 мкл буфера АЕ. Присутствие ДНК HSV-2 определяли с помощью количественного PCR в реальном времени (система RT-PCR Applied Biosystems 7300), с использованием 100 нг ДНК ганглиев и праймеры, специфичные для ДНК-полимеразы HSV (прямой: 5' GCT CGA GTG CGA AAA AAC GTT C (SEQ ID NO:4), обратный: 5' CGG GGC GCT CGG CTA AC (SEQ ID NO:5)), как описано ранее (8). Минимальное количество достоверно определяемых копий вирусной ДНК HSV-2 составляло 1 копию на реакцию.
Статистический анализ
Для статистического анализа применяли непарные t-тесты Стьюдента. Результаты считались статистически значимыми при значении P<0,05.
II. Результаты
Конструирование CJ2-gD2
В качестве первого шага в создании доминантно-негативного дефектного по репликации рекомбинантного вируса HSV-2, экспрессирующего gD2 и UL9-C535C, заявители сконструировали делеционный мутант HSV-2 ICP0, N2-lacZ, в котором обе копии ICP0 HSV-2 штамма 186 заменены на ген LacZ под контролем промотора HSV-2 ICP0 (фиг.1В). Заявители показали, что, как и у мутанта HSV-2 ICP0 null 7314 (11), эффективность образования бляшек N2-lacZ на клетках остеосаркомы человека U2OS в 425 раз выше чем на клетках Vero, что указывает на то, что клеточная активность в клетках U2OS может функционально замещать ICP0 HSV-2. По сравнению с HSV-2 дикого типа, эффективность репликации N2-lacZ в клетках Vero ослаблена в более, чем 600 раз при множественности инфекции 0,1 БОЕ на клетку. В соответствии с этим наблюдением, интравагинальная прививка N2-lacZ в количестве 1×105 и 5×105 БОЕ на мышь не приводила к системному или локальному заболеванию, в то время как у мышей, инфицированных 1×104 БОЕ на мышь HSV-2 дикого типа развился тяжелый генитальный герпес, приведший к гибели всех животных через 11 дней после инфекции. Кроме того, после интравагинальной инфекции N2-lacZ в дозировке 5×105 БОЕ на мышь у животных не развивается реактивируемая латентная инфекция. Эти результаты указывают на то, что так же, как и у ICP0 HSV-1 (10, 11, 37, 64), делеция ICP0 HSV-2 значительно ослабляет способность вируса вызывать острую и реактивируемую латентную инфекцию in vivo.
Для повышения уровня экспрессии gD2 доминантно-негативным дефектным по репликации рекомбинантным вирусом HSV-2 заявители сконструировали доминантно-негативный дефектный по репликации рекомбинант HSV-2 (CJ2-gD2) путем замены обеих копий гена Lac Z у N2-lacZ на последовательности ДНК, кодирующие ген gD2 под контролем основного предраннего промотора ICP4 HSV-1, несущего tetO, и UL9-C535C под контролем основного предраннего промотора hCMV, содержащего tetO, укороченного на -236 п.н. от полной длины предраннего промотора hCMV (фиг.1В). Таким образом, в отличие от CJ9-gD, кодирующего одну копию вставленного гена gD HSV-1 под контролем промотора hCMV, содержащего tetO, в локусе UL9 HSV-1 (41), CJ2-gD2 содержит 2 копии гена gD2 под контролем предраннего промотора ICP4 HSV-1, несущего tetO, состоящего из последовательности промотора ICP4 HSV-1 от -377 п.н. до -19 п.н. относительно сайта начала транскрипции гена ICP4. N2-C535C представляет собой рекомбинант HSV-2, у которого обе копии гена Lac Z у N2-lacZ заменены на UL9-C535C под контролем полноразмерного предраннего промотора hCMV, несущего tetO.
Высокий уровень экспрессии gD2 и UL9-C535C при инфекции клеток Vero CJ2-gD2
Для проверки экспрессии gD2 и UL9-C535C с предраннего промотора ICP4 HSV-1, несущего tetO, и предраннего промотора hCMV, соответственно, клетки Vero инфицировали HSV-2 дикого типа, N2-lacZ, N2-C535C и CJ2-gD2 при множественности инфекции 10 БОЕ на клетку и собирали через 9 ч после инфекции. Белки инфицированных клеток анализировали с помощью Вестерн-блоттинга с использованием моноклональных антител к ICP27 HSV-1/2, поликлональных антител к UL9 и поликлональных антител к gD1 (R45). Поскольку gB2, как и gD2, является основной мишенью нейтрализующих антител, а также Т-клеточных ответов, и представляет собой продукт γ1, белки инфицированных клеток также инкубировали с моноклональными антителами, специфичными к gB. На фиг.2А показано, что уровень экспрессии предраннего белка ICP27 HSV-2 CJ2-gD2 и N2-C535C сходен с таковым HSV-2 дикого типа и N2-lacZ. В клетках, инфицированных CJ2-gD2 и N2-C535C, обнаруживали значительное количество UL9-C535C, однако в то же время в клетках, инфицированных N2-C535C, обнаружили малое количество gD2 или gB2. В отличие от инфекции N2-C535C, инфекция клеток Vero CJ2-gD2 ведет к высокому уровню экспрессии gD2, сходному с таковым в клетках, инфицированных HSV-2 дикого типа, и экспрессия gD2 не влияла на экспрессию gB2. Из приведенных результатов также следует, что также, как и в случае мутанта HSV-1 ICP0 null 7134 (71), делеция ICP0 HSV-2 в N2-lacZ значительно ослабляет экспрессию gD2. Из-за очень низкого уровня экспрессии UL9 со своего собственного раннего промотора HSV (68), в клетках, инфицированных этими четырьмя вирусами, не удалось обнаружить UL9 дикого типа. Кроме того, заявители показали, что уровень экспрессии UL9-С535С в клетках, инфицированных CJ2-gD2, был неизменно выше, чем в клетках, инфицированных N2-C535C, что позволяет предположить, что VP16-респонсивные элементы HSV, TAATGARAT, присутствующие в промоторе ICP4 HSV-1 (71) могут приводить к повышенной экспрессии UL9-C535C с предраннего промотора hCMV в описанной промоторной системе ICP4/hCMV.
Вестерн-блот анализ с использованием поликлональных антител к gD1 (R45), результаты которого приведены на фиг.2В, показал, что в клетках, инфицированных HSV-1 дикого типа, был обнаружен гораздо более высокий уровень gD, чем в клетках, инфицированных HSV-2 дикого типа, а уровень gD, обнаруживаемый в клетках, инфицированных CJ9-gD, был гораздо ниже, чем в клетках, инфицированных CJ2-gD2. Это факты говорят о том, что CJ2-gD2 экспрессирует gD2 более эффективно, чем CJ9-gD экспрессирует gD1.
Чтобы показать, что UL9-C535C и gD2, экспрессируемые в клетках Vero, инфицированных CJ2-gD2, действительно находятся под контролем промоторов, несущих tetO, далее заявители инфицировали стабильную линию клеток Vero, экспрессирующих tetR, VCEP4R-28, HSV-2 дикого типа и CJ2-gD2 при множественности инфекции 10 БОЕ на клетку в присутствии или в отсутствие тетрациклина. Белки инфицированных клеток собирали через 9 часов после инфекции и анализировали с помощью Вестерн-блота. Как показано на фиг.3, в клетках VCEP4R-28, инфицированных HSV-2 дикого типа и CJ2-gD2, в присутствии и в отсутствие тетрациклина выявлялись сходные уровни ICP27, в то время как в клетках VCEP4R-28, инфицированных CJ2-gD2, UL9-C535C выявлялся только в присутствии тетрациклина, и также в присутствии тетрациклина выявлялись значительно более высокие уровни gD2, чем в его отсутствие.
CJ2-gD2 не реплицируется в клетках Vero
Из-за отсутствия ICP0 и высокой экспрессии UL9-C535C с основного предраннего промотора hCMV, несущего tetO, CJ2-gD2 конструировали и размножали в комплементирующей по ICP0 линии клеток U2OS, U2CEP4R11, экспрессирующей tetR (68). При исследовании на образование бляшек на монослоях клеток Vero 6,65×107 БОЕ CJ2-gD2 инфекционного вируса выявлено не было, что означает, что эффективность образования бляшек CJ2-gD2 на клетках Vero снижена по меньшей мере в 6,65×107 раз по сравнению с эффективностью на комплементирующих клетках U2CEP4R11.
Ингибирование репликации HSV-2 дикого типа CJ2-gD2
Далее с помощью ко-инфекции заявители исследовали доминатно-негативный эффект высокого уровня экспрессии UL9-C535C CJ2-gD2 на репликацию HSV-2 дикого типа (фиг.4). На фиг.4А показано, что ко-инфекция клеток Vero CJ2-gD2 при множественности инфекции 5 БОЕ на клетку и HSV-2 дикого типа при множественности инфекции 2 БОЕ на клетку приводило к почти 500-кратному снижению продукции HSV-2 по сравнению с клетками, инфицированными только HSV-2 при такой же множественности инфекции, вне зависимости от того, определяли ли титры вируса в клетках Vero или U2CEP4R11. В сходном эксперименте по ко-инфекции с N2-lacZ наблюдали только небольшое снижение выхода вируса дикого типа.
Для дальнейшего изучения степени ингибирования CJ2-gD2 репликации HSV-2 дикого типа заявители провели эксперименты по ко-инфекции HSV-2 дикого типа и CJ2-gD2 при отношении множественности инфекций 1:1 и 3:1. Из результатов, приведенных на фиг.4В, видно, что CJ2-gD2 эффективно предотвращает инфекцию HSV-2 в обоих случаях. Ко-инфекция приводит к 151- и 94-кратному снижению синтеза вируса дикого типа в случае указанных соотношений по сравнению с клетками, инфицированными только HSV-2 дикого типа при множественности инфекции 5 БОЕ на клетку и 15 БОЕ на клетку соответственно.
CJ2-gD2 авирулентен при внутрицеребральной инъекции мышам
Нейровирулентность является одним из основных свойств инфекции HSV. Чтобы изучить способность CJ2-gD2 и N2-C535C реплицироваться в ЦНС, самок мышей BALB/c возрастом 5-6 недель случайным образом делили на 5 групп по 8 мышей в каждой группе. CJ2-gD2 и N2-C535C прививали напрямую в левую лобную долю мозга каждой мыши при множественности инфекции 2,5×106 БОЕ на мышь в объеме 20 мкл инсулиновой иглой 28G на глубину 4 мм (74). Заболеваемость и смертность отслеживали на протяжении 35 дней. Поскольку LD50 HSV-2 дикого типа штамма 186 у самок мышей BALB/c составляет примерно 10 БОЕ на мышь при инъекции в камеру глаза (38), группе мышей также прививали HSV-2 дикого типа в количестве 25 БОЕ на мышь. В качестве дополнительного контроля, мышей в пятой группе прививали N2-lacZ в количестве 1×106 БОЕ на мышь. На фиг.5 показано, что как и у мышей, привитых DMEM, у мышей, внутрицеребрально привитых CJ2-gD2 и N2-C535C в дозировке 2,5×106 БОЕ не наблюдалось признаков нейровирулентности в течение 35-дневного срока наблюдения, в то время как все мыши, привитые HSV-2 дикого типа в количестве 25 БОЕ на мышь, (доза меньше в 100000 раз, чем доза CJ2-gD2), умерли на 10 день после прививки, причем у всех привитых мышей наблюдались симптомы заболевания ЦНС, обычно ассоциированного с инфекцией HSV-2, включая взъерошенный мех, сгорбенную позу, атаксию и анорексию. Несмотря на то, что 100% мышей, привитых N2-lacZ, выжили, у всех мышей наблюдались симптомы энцефалита.
Индукция нейтрализующих антител, специфичных к HSV-2, а также образование gD2-специфичных антител у мышей иммунизированных CJ2-gD2
Способность CJ2-gD2 индуцировать нейтрализующие антитела, специфичные к HSV-2, определяли у мышей, иммунизированных CJ2-gD2 в дозировке 2×106 БОЕ на мышь. В качестве контроля мышей также иммунизировали N2-C535C или CJ9-gD в той же дозировке. Как показано на фиг.6А, средний титр нейтрализующих антител, специфичных к HSV-2, у мышей, иммунизированных CJ2-gD2, составлял в среднем 500, что в три раза выше, чем у мышей, иммунизированных N2-C535C (p=0,015), и сравнимо с титром нейтрализующих антител, индуцированным у мышей, иммунизированных CJ9-gD (p=0,28). У контрольных мышей в разведении 1:10 не выявлялось титра антител, специфичных к HSV-2.
На фиг.6В показано, что у мышей, иммунизированных CJ2-gD2 и CJ9-gD, наблюдались сходные уровни образования антител, специфичных к gD, при детекции соответственных иммунопреципитированных комплексов gD2 антителами к gD1 (R45). В то же время, у мышей, иммунизированных CJ2-gD2, уровни антител, специфичных к gD, были значительно выше, чем у мышей, иммунизированных N2-C535C, и контрольных. Результаты, приведенные на фиг.6, в совокупности указывают на то, что высокий уровень экспрессии gD2 CJ2-gD2 ведет к повышенной эффективности образования антител к gD2, а также к образования нейтрализующих антител, специфичных к HSV-2, по сравнению с N2-C535C.
Индукция Т-клеточного ответа, специфичного к HSV-2, у мышей, иммунизированных CJ2-gD2
Для оценки эффективности иммунизации CJ2-gD2 в отношении индукции Т-клеточного ответа, специфичного к HSV-2, заявители исследовали ответы Т-клеток памяти у иммунизированных мышей на нагрузку HSV-2 дикого типа. Контрольных или вакцинированных CJ2-gD2 мышей не нагружали или нагружали HSV-2 дикого типа через 9-10 недель после вторичной иммунизации и исследовали с помощью теста IFNγ ELISPOT CD4+ и CD8+ Т-клетки из селезенки отдельных групп мышей (n=3) на 5 день после нагрузки (фиг.7А). У мышей, вакцинированных CJ2-gD2 и нагруженных HSV-2 дикого типа, наблюдалось 4,8-кратное увеличение количества IFNγ-положительных CD4+ Т-клеток по сравнению с контрольными мышами, иммунизированными CJ2-gD2 (p<0,0001). Более существенно то, что количество CD4+ Т-клеток, секретирующих IFNγ, обнаруженных у мышей, инфицированных HSV-2 и предварительно вакцинированных CJ2-gD2, оказалось в 18 раз больше, чем у невакцинированных мышей, инфицированных HSV-2 (p<0,0001). У контрольных невакцированных и ненагруженных мышей в тех же условиях IFNγ-положительные CD4+ Т-клетки не обнаруживались. Эти наблюдения свидетельствуют о том, что CJ2-gD2 вызывает сильный ответ CD4+ Т-клеток памяти.
У мышей, вакцинированных CJ2-gD2, наблюдалось более, чем двукратное увеличение количества CD8+ Т-клеток, секретирующих IFNγ, чем у невакцинированных контрольных мышей, но сходные количества CD8+ Т-клеток, секретирующих IFNγ, наблюдались в селезенке невакцинированных мышей, инфицированных HSV-2, и мышей, вакцинированных CJ2-gD2 и инфицированных HSV-2 (фиг.7В). Заявители провели вторую серию экспериментов, в которых мышей, невакцинированных и вакцинированных CJ2-gD2, не нагружали или нагружали HSV-2 дикого типа (n=3) через 5-6 недель после второй вакцинации. Тесты CD4+ и CD8+ ELISPOT проводили через 4 дня после инфекции (фиг.7C и 7D). У мышей, иммунизированных CJ2-gD2, после инфекции HSV-2 наблюдалось 8,6- и 5,7-кратное увеличение количества CD4+ и CD8+ Т-клеток, секретирующих IFNγ, соответственно, по сравнению с неинфицированными мышами, иммунизированными CJ2-gD2 (CD4+ Т-клетки: p=0,035; CD8+ Т-клетки: p=0,01). Кроме того, у мышей, вакцинированных CJ2-gD2, после нагрузки HSV-2 количество CD4+ и CD8+ Т-клеток, секретирующих IFNγ, было в 8 и 9,5 раз больше соответственно, чем у невакцинированных мышей (CD4+ Т-клетки: р=0,036; CD8+ Т-клетки: р=0,01). Эти данные в совокупности указывает на то, что CJ2-gD2 может вызывать мощные ответы CD4+ и CD8+ Т-клеток памяти, специфичных к HSV-2, обеспечивающие эффективный вторичный иммунный ответ при инфекции HSV-2.
Защита от генитальной инфекции и заболевания, вызванного HSV-2 у иммунизированных мышей
Через 5-6 недель после первичной иммунизации мышей нагружали интравагинально HSV-2 штамма G в количестве 50 LD50 (5×105 БОЕ на мышь). Вагинальные мазки брали на 1, 2, 3, 5 и 7 день после нагрузки. У мышей наблюдали в течение 21-дневного периода частоту возникновения генитального и системного заболевания, вызванного HSV-2. Как показано на фиг.8А, выход нагрузочного вируса был снижен более, чем в 200 раз на 1 день (p<0,001) и в 130 раз на 2 день (p<0,0001) у мышей, иммунизированных CJ2-gD2 (n=9) по сравнению с неиммунизированным контролем (n=10). Заявители не наблюдали значимой разницы в ослаблении вирусного слущивания на 1, 2 и 3 день после нагрузки между группами мышей, иммунизированных CJ2-gD2 и N2-C535C (n=10), однако, по сравнению с CJ9-gD, иммунизация CJ2-gD2 более эффективно ослабляла вирусное слущивание на 1 (р=0,03), 2 (р=0,025) и 3 (p<0,007) дни. У мышей, иммунизированных CJ2-gD2, N2-C535C или CJ9-gD, на 5 день после инфекции практически не наблюдалось нагрузочного вируса, в то время как у всех неиммунизированных мышей вирус продолжал слущиваться в количестве в среднем более 5×103 БОЕ/мл. У трех иммунизированных групп мышей в материале вагинальных мазков, взятых на 7 день после нагрузки, не обнаруживалось нагрузочного вируса. В ходе другого эксперимента заявители показали, что на 5 день после нагрузки у мышей, иммунизированных CJ2-gD2, не наблюдалось вирусного слущивания, но у 5 из 7 мышей, иммунизированных N2-C535C, и у 4 из 7 мышей, иммунизированных CJ9-gD, присутствовал HSV-2 дикого типа.
Результаты, приведенные на фиг.9, свидетельствуют о том, что иммунизация мышей CJ2-gD2 полностью предотвратила развитие локальных генитальных язв и системного заболевания после нагрузки HSV-2 дикого типа (фиг.9А). У всех неиммунизированных мышей развились тяжелые генитальные язвы; все мыши погибли от инфекции HSV-2 дикого типа на 11 день после нагрузки (фиг.9В). Несмотря на то, что иммунизация N2-C535C и CJ9-gD предохраняла мышей от летальной нагрузки HSV-2 дикого типа, у 20% и 30% мышей соответственно наблюдались преходящее легкое локальное генитальное заболевание (1 балл) (таблица 1). В сходном эксперименте (таблица 1) заявители показали, что у мышей, иммунизированных CJ9-gD (n=7) у двух мышей наблюдались преходящее локальное локальное генитальное заболевание и у 1 мыши наблюдались симптомы системного заболевания. Эта мышь умерла на 14 день после нагрузки. У 3 из 7 мышей, иммунизированных N2-C535C (43%) наблюдались преходящее легкое локальное генитальное заболевание (1 балл). У мышей, иммунизированных CJ2-gD2, опять не наблюдалось симптомов локального или системного заболевания герпесом (n=7). Совокупность этих данных указывает на то, что CJ2-gD2 представляет собой более эффективную вакцину, чем N2-C535C и CJ9-gD, для защиты мышей от генитального заболевания, вызванного интравагинальной нагрузкой HSV-2 дикого типа.
Процент защиты от заболевания герпесом у контрольных и иммунизированных мышей после интравагинальной нагрузки HSV-2 дикого типа
Список литературы
1. Abu-Raddad, et al., PLoS ONE 3:e2230 (2008).
2. Ackermann, et al., J. Virol. 52:108-18 (1984).
3. Adelson, et al., J. Clin. Virol. 55:25-34 (2005).
4. Arvin, et al., Infect. Immun. 40:184-9 (1983).
5. Augustinova, et al., J. Virol. 78:5156-65 (2004).
6. Bourne, et al., Vaccine 14:1230-4 (1996).
7. Brans, et al., J. Invest. Dermatol. 129:2470-9 (2009).
8. Brans, et al., J. Invest. Dermatol. 725:2825-32 (2008).
9. Bryson, et al., J. Infect. Dis. 167:942-6 (1993).
10. Cai, et al., J. Virol. 67:1501-12 (1993).
11. Cai, et al., J. Virol. 65:4579-89 (1989).
12. Cohen, et al., J. Virol. 49:102-8 (1984).
13. Coleman, et al., J. Clin. Microbiol. 75:287-91 (1983).
14. Cooper, et al., Cell. Immunol. 239:113-20 (2006).
15. Corey, et al., N. Engl. J. Med. 314:149-51 (1986).
16. DeLuca, et al., J. Virol. 62:132-43 (1988).
17. Dolan, et al., J. Virol. 72:2010-21 (1998).
18. Dudek, et al., Virology 344:230-9 (2006).
19. Fleming, et al., N. Engl. J. Med. 557:1105-11 (1997).
20. Freeman, et al., Aids 20:73-83 (2006).
21. Glorioso, et al., J. Virol. 50:805-12 (1984).
22. Grammer, et al., J. Immunol. 145:2249-53 (1990).
23. Gupta, et al., Lancet 370:2121-31 (2007).
24. Handler, et al., J. Virol. 70:6067-70(1996).
25. Hirsch, Herpes simplex virus, p. 1144-1153. In G. L. Mandell, R. G. J. Douglas, and J. E. Bennett (ed.), Principles and practice of infectious diseases. Churchill Livingstone Inc., New York (1990).
26. Honess, et al., J. Gen. Virol. 22:159-69 (1974).
27. Hosken, et al., J. Virol. 50:5509-15 (2006).
28. Johnson, et al., J. Immunol. 145:102-10 (1990).
29. Jones, et al., Herpes 11:12-1 (2004).
30. Kim, et al., J. Immunol. 181:6604-15 (2008).
31. Koelle, et al., Annu. Rev. Med. 5P:381-395 (2008).
32. Koelle, et al., Clin. Microbiol. Rev. 16:96-113 (2003).
33. Koelle, et al., Curr. Eye Res. 30:929-42 (2005).
34. Koelle, et al., J. Clin. Invest. 91:961-8 (1993).
35. Kousoulas, et al., Virology 166:423-31 (1988).
36. Lakeman, et al., Sex. Transm. Dis. 13:61-6 (1986).
37. Leib, et al., J. Virol. 65:759-68 (1989).
38. Lewandowski, et al., J. Neuroimmunol 81:66-15 (1998).
39. Liesegang, Cornea 20:1-13 (2001).
40. Looker, et al., Sex. Transm. Infect. 81:103-1 (2005).
41. Lu, et al., J. Invest. Dermatol. 129:1114-84 (2009).
42. McGeoch, et al., J. Gen. Virol. 69:1531-74 (1988).
43. McGeoch et al., J. Gen. Virol. 72:3057-3075 (1991).
44. McGeoch et al., Nucl. Acid Res. 14:1121-1145 (1986).
45. McGeoch, et al., Nucleic Acids Res 14:3435-48 (1986).
46. Mertz, et al., Ann. Intern. Med. 116:191-202 (1992).
47. Mikloska, et al., J. Gen. Virol. 7P:353-61 (1998).
48. Mikloska, et al., J. Immunol. 164:5161-16 (2000).
49. Minson, et al., J. Gen. Virol. 67 (Pt 6): 1001-13 (1986).
50. Morrison, et al., Virology 245:178-87 (1998).
51. Muller, J. Virol. 67:858-65 (1987).
52. Nagot, et al., N. Engl J. Med. 356:190-9 (2007).
53. Para, et al., J. Virol. 55:483-8 (1985).
54. Pereira, Dev. Biol. Stand. 52:115-31 (1982).
55. Pereira, et al., Infect. Immun. 29:124-32 (1980).
56. Perry, et al., J. Gen. Virol. 67:2365-2380 (1986).
57. Roberts, et al., J. Virol. 62:4301-20 (1988).
58. Roizman, et al., Herpes simplex viruses and their replication, p. 2399-2459. In a. P. M. H. D. M. Knipe (ed.), Fields Virology, 4rd ed. Lippincott Williams & Wilkins, Philadelphia, Pa. (2001).
59. Schmidt, et al., J. Infect. Dis. 149:645-6 (1984).
60. Stanberry, Herpes 11(Suppl 3):161A-169A (2004).
61. Stanberry, et al., Clin. Infect. Dis. 30:549-66 (2000).
62. Stanberry, et al., N. Engl. J. Med. 347:1652-61 (2002).
63. Starr, et al., Gene Ther 5:615-23 (1996).
64. Stow, et al., J. Gen. Virol. 67:2571-85 (1986).
65. Tigges, et al., J. Virol. 66:1622-34 (1992).
66. Whitley, et al., Clin Infect Dis 26:541-53; quiz 554-5 (1998).
67. Xu, et al., J. Infect. Dis. 755:1019-24 (2002).
68. Yao, et al., Hum. Gene Ther. 70:1811-8 (1999).
69. Yao, et al., Hum. Gene Ther. 10:419-27 (1999).
70. Yao, et al., Antiviral Res. 53:127-33 (2002).
71. Yao, et al., J. Virol. 69:6249-58 (1995).
72. Yao, et al., J. Virol. 65:8158-68 (1994).
73. Yao, et al., Hum. Gene Ther. 9:1939-50 (1998).
74. Yao, et al., Mol. Ther. 75:1133-41 (2006).
75. Zarling, et al., J. Immunol. 136:4669-13 (1986).
Все литературные источники, процитированные в настоящей заявке, полностью включены путем ссылки. Из развернутого описания изобретения для специалиста в области техники очевидно, что изобретение может применяться с использованием широкого эквивалентного диапазона условий, параметров и т.д., без отступления от сущности и объема настоящего изобретения или любого его воплощения.


Изобретение относится к биотехнологии. Предложены дефектный по репликации доминантно-негативный рекомбинантный вирус простого герпеса 2, вакцина и способ иммунизации пациента против инфекции HSV-1 или HSV-2. Рекомбинантный вирус содержит в своем геноме две последовательности, кодирующие гликопротеин D HSV-2, связанные с промоторами ICP4, последние связаны с последовательностями тетрациклиновых операторов, и две последовательности, кодирующие белок UL9-C535C, которые связаны с промоторами hCMV, связанными с последовательностями тетрациклиновых операторов. Причем вирус не содержит последовательность, кодирующую белок ICP0. Вакцина, содержащая указанный рекомбинантный вирус, является безопасной и эффективной против инфекций HSV-1 или HSV-2 за счет экспрессии доминантно-негативной формы полипептида UL9-C535C и использования индуцированного тетрациклином включения и выключения генов. Также предложен способ иммунизации пациентов против HSV-1 или HSV-2 с использованием предложенной вакцины. Предложенные вакцина и способ иммунизации пациентов могут быть использованы в медицине для превращения или подавления вспышки HSV-1 или HSV-2. 3 н. и 14 з.п. ф-лы, 19 ил., 1 табл., 1 пр.
1. Дефектный по репликации доминантно-негативный рекомбинантный вирус простого герпеса 2 (Herpes Simplex Virus, HSV-2) для применения в качестве вакцины, геном которого включает:
a) первую последовательность, кодирующую первый гликопротеин D HSV-2 (gD2), где указанная последовательность функционально связана с первым промотором, причем указанный первый промотор представляет собой промотор ICP4, модифицированный путем исключения ДНК-связывающей последовательности ICP4, и указанный первый промотор функционально связан с первой последовательностью тетрациклинового оператора (Tet-O);
b) вторую последовательность, кодирующую второй HSV-2 gD2, где указанная вторая последовательность функционально связана со вторым промотором, причем указанный второй промотор представляет собой промотор ICP4, модифицированный путем исключения ДНК-связывающей последовательности ICP4, и причем указанный второй промотор функционально связан со второй последовательностью Tet-O;
c) третью последовательность, кодирующую белок UL9-C535C, где указанная третья последовательность функционально связана с третьим промотором, и указанный третий промотор представляет собой предранний промотор hCMV, функционально связанный с третьей последовательностью тетрациклинового оператора (Tet-O);
d) четвертую последовательность, кодирующую белок UL9-C535C, где указанная четвертая последовательность функционально связана с четвертым промотором, и указанный четвертый промотор представляет собой предранний промотор hCMV, функционально связанный с четвертой последовательностью тетрациклинового оператора (Tet-O);
и где указанный геном не включает последовательности, кодирующей функциональный белок ICP0.
2. Вирус по п. 1, где:
a) каждый из указанных первого, второго, третьего и четвертого промоторов содержит TATA-элемент;
b) каждый из указанных первого, второго, третьего и четвертого промоторов содержит 2 сайта ор2 связывания репрессора, соединенных последовательностью длиной 2-20 нуклеотидов, где первый нуклеотид указанного tet-оператора располагается между 6 и 24 нуклеотидом к 3′-концу от последнего нуклеотида указанного TATA-элемента;
c) указанная последовательность, кодирующая gD2 HSV-2, и указанная вторая последовательность, кодирующая gD2 HSV-2, располагается к 3′-концу от указанных первой и второй последовательностей Tet-O и функционально связаны с указанными первым и вторым промотором;
d) указанная последовательность, кодирующая указанную первую доминантно-негативную мутантную форму белка UL9 HSV-1 или HSV-2, располагается к 3′-концу от указанной третьей последовательности Tet-O и функционально связана с указанным третьим промотором;
e) указанная последовательность, кодирующая указанную вторую доминантно-негативную мутантную форму белка UL9 HSV-1 или HSV-2, располагается к 3′-концу от указанной четвертой последовательности Tet-O и функционально связана с указанным четвертым промотором.
3. Вирус по п. 1, где указанный вирус также экспрессирует один или более рекомбинантных иммуномодулирующих генов.
4. Вирус по п. 1, где указанный вирус экспрессирует IL12.
5. Вирус по п. 1, где указанный вирус экспрессирует IL15.
6. Вирус по п. 1, где указанный вирус также экспрессирует gB HSV-2 под контролем предраннего промотора HSV или hCMV, несущего tetO.
7. Вирус по п. 1, где указанный вирус также экспрессирует gC HSV-2 под контролем предраннего промотора HSV или hCMV, несущего tetO.
8. Вирус по п. 1, в котором промоторы ICP4 отвечают на VP16.
9. Вирус по п. 1, в котором по меньшей мере один из указанного третьего или указанного четвертого предраннего промотора hCMV и модифицированного промотора ICP4 находится в противоположной ориентации.
10. Вакцина для иммунизации пациентов против инфекции HSV-1 или HSV-2, содержащая рекомбинантный вирус по любому из пп. 1-9 в стандартной единичной дозировке, где стандартная единичная дозировка содержит однократную вводимую дозу лекарственного препарата, раствор которого вызывает терапевтический эффект.
11. Вакцина по п. 10, где указанный вирус присутствует в количестве не менее 1×107 БОЕ на дозу.
12. Вакцина по п. 10, где указанный вирус присутствует в количестве 1×107 - 1×109 БОЕ на дозу.
13. Способ иммунизации пациента против инфекции HSV-1 или HSV-2, включающий введение указанному пациенту эффективного количества вакцины по любому из пп. 10-12.
14. Способ по п. 13, где указанный пациент сероположителен по HSV-1.
15. Способ по п. 13, где указанный пациент сероположителен по HSV-2.
16. Способ по п. 13, где указанный пациент сероположителен по HSV-1 и HSV-2.
17. Способ по п. 13, где указанный пациент серонегативен по инфекции HSV-1 и HSV-2.
| WO 1990013652 A1 15.11.1990 | |||
| US 0006027730 A1 22.02.2000 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| JOURNAL OF INVESTIGATIVE DERMATOLOGY vol | |||
| Способ применения резонанс конденсатора, подключенного известным уже образом параллельно к обмотке трансформатора, дающего напряжение на анод генераторных ламп | 1922 |
|
SU129A1 |
| BRANS RICHARD et al: "Prevention of Genital Herpes Simplex | |||

